phÂn lẬp, tuyỂn chỌn vÀ ĐÁnh giÁ khẢ nĂng cỐ ĐỊnh...
TRANSCRIPT

Kỷ yếu kỷ niệm 35 năm thành lập Trường ĐH ng nghiệp Th c ph m T h Minh 98 -2017)
38
PHÂN LẬP, TUYỂN CHỌN VÀ ĐÁNH GIÁ KHẢ NĂNG CỐ ĐỊNH ĐẠM
CỦA MỘT SỐ CHỦNG VI KHUẨN NỐT SẦN Ở RỄ CÂY ĐẬU PHỘNG
(ARACHIS HYPOGAEA. L)
Trần Ánh Nguyệt1, *
, Trần Minh Trí1, Hà Thanh Đạt
1,
Trần Thu Thảo1, Hồ Viết Thế
2
1Viện Lúa Đ ng Bằng S ng ửu Long
2Trường Đại học ng nghiệp Th c ph m Thành phố h Minh
*Email: [email protected]
TÓM TẮT
Vi khuẩn nốt sần được phân lập từ rễ cây đậu phộng (Arachis hypogaea. L) thông qua việc định
danh về đặc điểm, hình thái, sinh lý, sinh hóa thì chỉ có hai chủng đạt yêu cầu. Vi khuẩn phân lập được
phát triển tốt trên môi trường YEMA, gram âm, hình que, hiếu khí, có khả năng di động, không bắt màu
trên môi trường có bổ sung Congo red, không phát triển trên môi trường Glucose-Peptone-Agar, có khả
năng chịu được nồng độ muối 2% và phát triển rất tốt trên môi trường kiềm pH từ 7-10. Về khả năng tạo
gum (polysaccharide được chiết xuất từ vỏ màng của tế bào vi khuẩn) cả hai đều tạo gum cao 0,91 mg và
0,89 mg. Hai chủng vi khuẩn cho khả năng cố định đạm cao thông qua phân tích khả năng hình thành nốt
sần và đạm tổng số của thí nghiệm trong nhà lưới. Nghiệm thức sử dụng vi khuẩn nhiễm hạt trước khi
gieo đều có số lượng và khối lượng nốt sần, trọng lượng 100 hạt, chiều cao cây và trọng lượng tươi, phần
trăm của đạm tổng số của cây được chủng cao hơn một cách rõ rệt so với đối chứng. Kết quả là tuyển
chọn được hai chủng vi khuẩn nốt sần cộng sinh có khả năng cố định đạm cao góp phần nâng cao năng
suất cây trồng và giảm chi phí sản xuất.
Từ khóa: vi khuẩn nốt sần, đậu phộng, YEMA.
1. GIỚI THIỆU
Đậu phộng (Arachis hypogaea. L) là cây thực phẩm, cây công nghiệp ngắn ngày dùng để lấy dầu
trong nhóm cây trồng cạn. Nó được biết đến mang lại nhiều nguồn lợi về kinh tế, dinh dưỡng, đồng thời
cũng là cây cải thiện môi trường nhờ vào khả năng cộng sinh với các vi khuẩn cố định đạm. Mặc dù, diện
tích trồng và sản lượng thu được từ cây đậu phộng ít hơn lúa và cây đậu nành nhưng để đưa vào cơ cấu
chuyển dịch luân canh nhằm giảm áp lực sản xuất cây lúa liên tục 3 vụ như hiện nay bồi dưỡng đất giúp
cải thiện độ phì nhiêu, cắt đứt nguồn lây lan dịch bệnh của cây lúa. Mặt khác, những vùng đất không có
khả năng canh tác lúa thì việc nghiên cứu nâng cao diện tích canh tác cây trồng cạn nói chung và cây đậu
phộng nói riêng cũng là điều cần thiết trong tình hình sản xuất nông nghiệp như hiện nay.
Sử dụng phân bón sinh học là một trong những biện pháp cũng góp phần nâng cao năng suất, phẩm
chất cây trồng. Mục đích khảo sát đánh giá, khắc phục những tác hại do lạm dụng quá nhiều phân đạm
hóa học thì việc sử dụng phân bón sinh học có chứa các chủng vi khuẩn có khả năng cố định đạm, là một
trong những biện pháp có hiệu quả mà không gây ô nhiễm môi trường, tiết kiệm chi phí, cân bằng hệ sinh
thái nhưng vẫn đảm bảo góp phần vào việc gia tăng năng suất, chất lượng nông sản và đồng thời góp phần
xây dựng một nền nông nghiệp bền vững và ổn định.
Hiện nay vi khuẩn cố định đạm trên rễ cây họ đậu như đậu nành, đậu xanh, bắp đã được phát triển và
áp dụng. Bên cạnh đó cũng có những chủng vi khuẩn cố định đạm khác như: vi khuẩn Azospirillum có
khả năng cố định đạm, tổng hợp IAA, hòa tan lân và các chất dinh dưỡng khác (Bilal và ctv, 1990;
Somers ctv, 2005; Dậu và ctv, 2007). Một số chế phẩm vi khuẩn nốt sần được sử dụng cho cây họ đậu để

hân lập, tuyển chọn và đánh giá khả năng cố định đạm của một số chủng vi khu n nốt sần ở...
39
tăng cường khả năng cố định đạm đã được tập trung nghiên cứu nhiều trên cây đậu nành, bắp (Sơn và ctv,
2006). Tuy nhiên, vẫn chưa có nhiều công trình nghiên cứu về khả năng của vi khuẩn cố định đạm trên
cây đậu phộng. Chính vì những lý do trên mục tiêu của nghiên cứu này là bước đầu phân lập, tuyển chọn
và đánh giá khả năng cố định đạm một số chủng vi khuẩn nốt sần ở rễ cây đậu phộng nhằm tìm ra những
chủng vi khuẩn có khả năng cố định đạm làm nguồn nguyên liệu cho việc tạo ra phân đạm sinh học, giúp
bảo vệ môi trường và góp phần bảo đảm cho sự phát triển nông nghiệp bền vững trong khu vực.
2. VẬT LIỆU VÀ PHƢƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu
Vật liệu nghiên cứu sử dụng trong đề tài là các chủng (nòi 1-6) vi khuẩn nốt sần được phân lập từ rễ
cây đậu phộng OMĐP13 ở giai đoạn 50 ngày tuổi. Ngoài ra, còn có sử dụng 6 giống đậu phộng (OMĐP1,
OMĐP13, OMĐP15, OMĐP16, OMĐP18, OMĐP23). Các thí nghiệm được tiến hành trong phòng thí
nghiệm, nhà lưới, sử dụng các trang thiết bị, hóa chất, giống đậu phộng của bộ môn Di Truyền và Chọn
Giống-Viện Lúa Đồng Bằng Sông Cửu Long.
2.2. Phƣơng pháp nghiên cứu
Thí nghiệm hoàn thiện quy trình phân lập vi khuẩn cố định đạm từ nốt sần của rễ cây đậu phộng đã
được trồng 50 ngày, tiến hành phân lập trên môi trường YEMA, làm thuần, quan sát hình dạng, thử
nghiệm sinh hóa, kiểm tra sự hình thành nốt sần trong nhà lưới, xác định khả năng cố định đạm của vi
khuẩn với cây đậu phộng, đánh giá hiệu quả cố định đạm.
2.2.1. Chu n bị m i trường phân lập, quan sát hình dạng, định danh, thử nghiệm sinh hóa
Trong các thí nghiệm chúng tôi thử nghiệm các môi trường YEMA (Yeast Extract Mannitol Agar)
dùng để phân lập vi khuẩn (Fred và ctv, 1932), YEMA cũng sử dụng môi trường trên thạch đứng để quan
sát khả năng tăng trưởng yếm hay kỵ khí, hoặc bổ sung sung từ 0 đến 4% muối NaCl thử nghiệm khả
năng tăng trưởng. Môi trường chọn lọc cho vi khuẩn nốt sần như: YEMA có bổ sung Congo red (Hahn và
ctv, 1966), và môi trường (GPA) glucose-peptone-agar (Kleczkowska và ctv, 1968). Chuẩn bị môi trường
dinh dưỡng có chứa 0,2% tinh bột (Xu và ctv,1995) và môi trường Hofer đánh giá sự tăng trưởng trên
môi trường có pH = 4-12 (Hofer và ctv, 1935). Đối với các thí nghiệm tăng trưởng trên môi trường
YEMA có bổ sung 0-4% muối và môi trường kiềm Hofer có pH = 4-12, mỗi nghiệm thức lập lại 3 ống,
mỗi thí nghiệm lập lại hai lần độc lập.
2.2.2. Thu mẫu và các bước phân lập vi khu n cố định đạm
Đào nhẹ nhàng rễ giống cây đậu phộng OMĐP13 được 50 ngày tuổi và chọn rễ có chứa nốt sần có
màu hồng, khỏe mạnh. Dùng kéo cắt nốt sần từ khối u, sau đó đem đi khử trùng bề mặt của nốt sần bằng
dung dịch CaOCl2 (4%) trong 3 phút. Tiếp theo các nốt sần được rửa lại nhiều lần với nước cất vô trùng
mục đích là để loại bỏ CaOCl2 còn bám trên bề mặt. Sau đó đặt chúng vào dung dịch cồn 70% trong vòng
3 phút, tiếp tục lặp lại các bước rữa với nước cất vô trùng. Lần lượt các nốt sần sạch được nghiền trong
eppendort có chứa nước cất vô trùng tạo thành dịch huyền phù. Dung dịch huyền phù trên được pha loãng
theo bậc 10 và nồng độ từ 10-2
đến 10-6
được cấy vào 3 đĩa petri môi trường YEMA. Các đĩa sau khi cấy
được ủ ở nhiệt độ phòng từ 2-3 ngày, quan sát thấy khuẩn lạc mọc ghi nhận kết quả và tiếp tục cấy
chuyền và làm thuần.
2.2.3. ác phương pháp sử dụng để phân biệt, định danh và quan sát s tăng trưởng
Các phương pháp được dùng để phân biệt, quan sát các đặc điểm nuôi cấy và các hình thái của vi
khuẩn cố định đạm như nhuộm Gram (Vincent và ctv, 1970) và quan sát gram, hình dạng dưới kính hiển
vi, cũng như khả năng tăng trưởng trên môi trường thạch đứng. Khảo sát sự tăng trưởng trên môi trường
có bổ sung glucose, manitol và laclose.
2.2.4. S sản xuất gum
Chuẩn bị môi trường YEMA lỏng, sau đó cho vào bình arlen mỗi arlen tương ứng 100 mL môi
trường. Trước đó đã chuẩn bị môi trường nhân sinh khối của vi khuẩn như sau: Cho 10 mL môi trường

Trần Thị nh Ngu ệt, Trần Minh Tr , à Thanh Đạt, Trần Thu Thảo, Viết Thế
40
YEMA lỏng vào 2 ống nghiệm tương ứng hai nòi, cấy vi khuẩn vào để qua đêm lắc trong 24h ở nhiệt độ
phòng. Sau đó hút 1mL dung dịch vi khuẩn cho vào arlen tương ứng đã chuẩn bị trước còn lại đối chứng.
Ủ liên tục 15 ngày trên máy lắc. Kế tiếp cho 250 µl cồn 95% và 100 mL aceton vào để yên trong 24 h,
tránh sự di chuyển. Tiếp tục lọc dung dịch bằng giấy lọc, sau đó sấy khô ở 37 ºC từ 2-3 ngày. Quan sát,
cân số lượng gum được tạo ra của chủng vi khuẩn và ghi nhận kết quả.
2.2.5. Thử nghiệm catalase bằng phản ứng với oxi già 3%)
Thử nghiệm này nhằm xác định khả năng hiếu khí hay kỵ khí tùy nghi của vi khuẩn phân lập được.
Chuẩn bị thành phần môi trường YEMA có bổ sung Congo red. Cấy mỗi chủng vi khuẩn đã phân lập vào
3 đĩa môi trường YEMA, ủ từ 2-3 ngày nuôi cấy. Khi khuẩn lạc đã mọc hoàn chỉnh, thử với H2O2 (3%),
quan sát và ghi nhận có sự sủi bọt hay không sủi bọt của 3 khuẩn lạc trên 3 đĩa môi trường lập lại của
từng dòng vi khuẩn.
2.2.6. S phân hủ m i trường tinh bột theo Xu và ctv 995)
Chuẩn bị môi trường dinh dưỡng có chứa 0,2% tinh bột, thành phần 1l môi trường như sau: Peptone
(5 g), Yeast extract (3 g), NaCl (5,0 g), bột gạo (2 g), agar (15 g). Cấy lần lượt mỗi chủng vi khuẩn vào 3
đĩa petri môi trường, ủ ở nhiệt độ phòng 1-2 ngày. Sau đó nhỏ dung dịch nhuộm gram Iod vào 3 khuẩn
lạc/mỗi đĩa petri. Quan sát, đánh giá sự hiện diện quầng sáng xung quanh khuẩn lạc.
2.2.7. Kiểm tra s hình thành nốt sần sau khi chủng vi khu n cố định đạm bằng phương pháp chậu
plastic trong nhà lưới
Mục đích kiểm tra vi khuẩn phân lập được có khả năng hình thành nốt sần, cố định đạm ở quy mô
nhà lưới. Quy trình được tiến hành như sau: Lấy đất ngoài ruộng cộng thêm cát, phơi khô, đập nhỏ đem
thanh trùng ở 121 ºC trong 30 phút. Rửa, lau cồn và phơi khô các chậu plastic, cho đất vào và tiến hành
gieo đậu phộng. Các giống đậu phộng trước khi gieo khử trùng bề mặt bằng cồn 70%, rửa thật sạch lại
với nước cất vô trùng, thấm khô. Tất cả các quá trình đều thực hiện trong điều kiện vô trùng tránh sự xâm
nhiễm của các vi sinh vật khác.
Sau 5 ngày đậu phộng bắt đầu nảy mầm khỏi mặt đất, tổng số 22 chậu của mỗi giống OMĐP được
gieo, 6 chậu đối chứng không chủng (K), 8 chậu chủng 1 (C1), 8 chậu chủng 2 (C2). Một mL dịch vi
khuẩn nuôi cấy sau 24 h có nồng độ 2x107 được xác định bằng phương pháp hộp đĩa bơm đều vào các
gốc đậu phộng, sau đó dán bề mặt chậu với màng phủ plastic trong khoảng 1-2 ngày. Tiếp theo, chờ đậu
phộng lớn lên ta đục lỗ cho cây ra khỏi bên ngoài. Thí nghiệm được tưới nước cất vô trùng và quan sát sự
hình thành nốt sần sau 60 ngày gieo.
Rễ cây từ mỗi chậu được đào một cách cẩn thận, rửa sạch với nước và đếm số lượng nốt sần được
hình thành từ mỗi chậu. Các chỉ tiêu khác như trọng lượng tươi của nốt sần, trọng lượng khô, đường kính
của nốt sần, màu sắc nốt sần, số quả trên cây, trọng lượng 100 hạt, sự sinh trưởng và phát triển của cây
cũng được ghi nhận.
2.2.8. Xác định nitơ tổng số bằng phương pháp Kjeldahl theo Bremner, 960)
Thân và lá của sáu giống đậu phộng 60 ngày tuổi, trên hai nghiệm thức chủng và không chủng được
cắt và chuyển mẫu phân tích đạm tổng số ở bộ môn Khoa Học Đất - Viện lúa Đồng Bằng Sông Cửu Long
theo phương pháp Kjeldahl.
2.3. Xử lý kết quả và tính toán số liệu
Số liệu được thu thập xử lý trên máy tính bằng chương trình Excel và Cropstat 7.2 (IRRI, 1996).
Mỗi nhóm thí nghiệm khác nhau tính toán hệ số biến thiên (CV%) và LSD (5%).
3. KẾT QUẢ VÀ THẢO LUẬN
3.1. Quá trình phân lập vi khuẩn cố định đạm từ nốt sần của rễ cây đậu phộng
Sau khi chọn được nốt sần khỏe có màu hồng từ rễ cây họ đậu chúng tôi đã tiến hành phân lập và
làm thuần được 6 chủng vi khuẩn. Qua kết quả sơ tuyển cho thấy 6 chủng đều sinh trưởng nhanh, khuẩn
lạc mọc sau 2 ngày nuôi cấy, và kích thước khuẩn lạc đạt được 5,5-6,5 mm sau 7 ngày nuôi cấy. Màu sắc

hân lập, tuyển chọn và đánh giá khả năng cố định đạm của một số chủng vi khu n nốt sần ở...
41
của khuẩn lạc từ trắng nhạt đến trắng trong, hình dạng khuẩn lạc từ tròn đến bầu dục, bề mặt có chủng thì
lồi, dẹt nhầy nhớt, màu sắc khuẩn lạc trắng trong, có đường viền trong suốt, hoặc trắng sữa quanh khuẩn
lạc, kích thước tăng nhanh sau 72 giờ nuôi cấy. Khi quan sát dưới kính hiển vi, vi khuẩn có khả năng di
động, hiếu khí, hình que ngắn, là vi khuẩn gram âm bắt màu hồng khi nhuộm.
Để loại bỏ những chủng không thuộc nhóm cộng sinh nốt sần, 6 chủng tiếp tục thử nghiệm trên môi
trường YEMA có bổ sung Congo red bởi vì đây là chất chị thị đặc biệt hữu hiệu và là môi trường chọn
lọc tốt nhất để phân biệt hay kiểm tra độ thuần của vi khuẩn nốt sần (Hình 1). Kết quả ở là khuẩn lạc vi
khuẩn có hình dạng trắng tròn, mờ sáng lấp lánh, màu trắng hoặc màu hơi đỏ nguyên nhân là do sự hấp
thụ của loài này với thuốc nhuộm rất yếu, tiếp tục loại bỏ những chủng có khả năng hấp thụ mạnh với
thuốc nhuộm Congo red (Hình 1).
ình . Hình dạng của khuẩn lạc trên môi trường YEMA bổ sung Congo red chủng hấp thụ yếu (B),
hấp thụ mạnh (A)
Dựa trên nguyên tắc vi khuẩn cố định đạm không sử dụng peptone cho quá trình sinh trưởng. Do
vậy, môi trường chọn lọc GPA dùng để phân biệt vi khuẩn phân lập từ nốt sần sau khi ủ từ 2-3 ngày. Kết
quả cho thấy rằng các chủng phân lập này không phát triển hay phát triển rất yếu trên môi trường này vì
đơn giản các nòi vi khuẩn này không thể sử dụng peptone làm nguồn nguyên liệu cho sự nhân đôi tế bào,
cũng như phát triển (Hình 2).
Qua hai thử nghiệm trên môi trường chọn lọc trên chúng tôi lựa chọn được 2 chủng đáp ứng hai điều
kiện không tăng trưởng trên môi trường GPA, và hấp thụ yếu đối với Congo red. Hai chủng C1 và C2 này
tiếp tục các thử nghiệm sinh lý sinh hóa và khả năng hình thành nốt sần và cố định đạm.
ình . Quan sát sự mọc của khuẩn lạc trên môi trường thử nghiệm GPA
(glucose- peptone-agar).
3.2. Quan sát đặc điểm sinh lý và sinh hóa của chủng vi khuẩn nốt sần
Khi quan sát khả năng tăng trưởng trên môi trường YEMA thạch đứng của hai chủng chúng tôi ghi
nhận rằng vết cấy mọc tốt từ đáy ống nghiệm đến bề mặt thạch sâu sau hai ngày cấy đều đó chúng tỏ rằng
hai chủng có khả năng di động. Ngoài ra, xác định khả năng hiếu khí bằng các nhỏ dung dịch H2O2 (3%)
vào các khuẩn lạc quan sát thấy hầu hết đều có hiện tượng sủi bọt xảy ra chứng tỏ cả hai chủng trên đều
hiếu khí.
Để quan sát tính tăng trưởng của 2 chủng vi khuẩn cố định đạm chúng tôi cũng quan sát khả năng
tăng trưởng trên môi trường YEMA có bổ sung muối NaCl 0,5-4% và môi trường Hofer pH=4-12. Sau
hai ngày cấy hai chủng vi khuẩn vào môi trường lỏng nhận thấy rằng có sự khác biệt giữa các nghiệm
thức: ở nồng độ muối đối chứng là không cấy vi khuẩn thì không có sự phát triển môi trường vẫn trong. Ở
các nồng độ muối 0,5-2% 2 chủng phát triển rất tốt làm đục cả môi trường, bắt đầu phát triển yếu ở nồng
độ 2,5% và nồng độ từ 3-4% không phát triển (Hình 3). Kết quả quan sát trên 2 lần thiết kế thí nghiệm
độc lập và cho kết quả tương tự là chủng phân lập được có thể chống chịu mặn được ở 2% và hy vọng
rằng chủng này có thể đáp ứng với những loại đất có các nồng độ cation và anion cao. Quan sát sự sinh
A B

Trần Thị nh Ngu ệt, Trần Minh Tr , à Thanh Đạt, Trần Thu Thảo, Viết Thế
42
trưởng và độ đục của hai chủng vi khuẩn trên môi trường YEM lỏng có sự khác biệt về pH từ 4-12. Ở các
ống nghiệm có pH từ 4-5 không phát triển được, pH = 6 phát triển nhưng rất yếu, còn ở các ống nghiệm
có pH từ 7-10 phát triển rất tốt, riêng ở pH bằng 11 có sự phát triển nhưng yếu và pH = 12 là không phát
triển (Bảng 1).
ình 3 Sự sinh trưởng và phát triển của chủng vi khuẩn số 2 ở các nồng độ muối NaCl từ 0,5 - 4%.
Bảng 1. Sự tăng trưởng của 2 chủng vi khuẩn nốt sần ở các độ pH 4-12
Độ pH
Chủng 4 5 6 7 8 9 10 11 12
Chủng 1 - - + + + + + + -
Chủng 2 - - + + + + + + -
Ngoài ra, chúng tôi cũng tập trung quan sát sự sản xuất gum (polysaccharide được chiết xuất từ vỏ
màng của tế bào vi khuẩn) và tiêu thụ các nguồn đường carbon. Khi cấy vi khuẩn trong môi trường lỏng,
ủ ở nhiệt độ 37 ºC trong 15 ngày, kết tủa rồi sấy khô cho thấy chủng thứ nhất tạo được khối lượng gum
nhiều nhất đạt 0,91 mg/100 mL và chủng thứ hai thấp hơn chỉ đạt 0,89 mg/100 mL trong khi đó thì đối
chứng chỉ đạt 0,3 mg/100 mL (Bảng 2). Đều đó cho thấy rằng cả hai nòi đều phát triển và cho kết quả cao
hơn so với đối chứng không có hiện diện của vi khuẩn. Thí nghiệm sử dụng các nguồn đường carbon cho
thấy rằng chỉ có manitol cho sự phát triển mạnh đục hoàn toàn môi trường sau hai ngày nuôi cấy trong khi
đó đối với các nguồn glucose và lactose thì phát triển rất yếu đều này được thể hiện bởi mật độ quang của
môi trường có chứa manitol cao hơn so với glucose và lactose (Bảng 2). Kết quả này cũng phù hợp bởi vì
môi trường dùng để phân lập ban đầu YEMA có chứa nhiều đường manitol.
Bảng 2. Sự sản xuất gum và tiêu thụ các nguồn đường carbon
Chủng Manitol Glucose Lactose Sản xuất Gum (mg/100 mL)
Chủng 1 Rất mạnh Rất yếu Rất yếu 0,91
Chủng 2 Rất mạnh Rất yếu Rất yếu 0,89
Do mỗi loại vi khuẩn có khả năng tăng trưởng và phát triển trên môi trường nhất định bằng cách sử
dụng các nguồn đường làm vật liệu. Với mục đích xem hai chủng vi khuẩn phân lập được có khả năng sử
dụng tinh bột như một đường phức được cấu tạo từ glucose như là nguồn năng lượng của carbon và năng
lượng cho sự phát triển. Đây là môi trường quyết định vi khuẩn có khả năng tạo ra enzyme amylase, vì
chính enzyme này sẽ phân cắt các phân tử glucose có trong tinh bột.
hân lập, tuyển chọn và đánh giá khả năng cố định đạm của một số chủng vi khu n nốt sần ở...
43
Giống
Số lượng nốt sần
(Nốt)
Trọng lượng tươi
nốt sần
(g)
Trọng lượng khô
nốt sần
(g)
C1 C2 K C1 C2 K C1 C2 K
OMĐP1 10,0 11,5 1,8 0,21 0,36 0,009 0,06 0,06 0,004
OMĐP13 14,6 14,1 1,0 0,51 0,47 0,004 0,13 0,11 0,0003
OMĐP15 13,0 16,1 1,0 0,54 0,35 0,01 0,12 0,08 0.004
OMĐP16 8,0 13,8 2,9 0,3 0,77 0,02 0,08 0,06 0,005
OMĐP18 12,1 13,9 1,0 0,36 0,42 0,001 0.05 0,09 0,0001
OMĐP23 13,3 16,1 2,8 0.5 0,54 0,02 0,11 0,12 0.005
Cv (%) 70,1 100,9 103,4
LSD (5%) 6,4 0,3 0.07
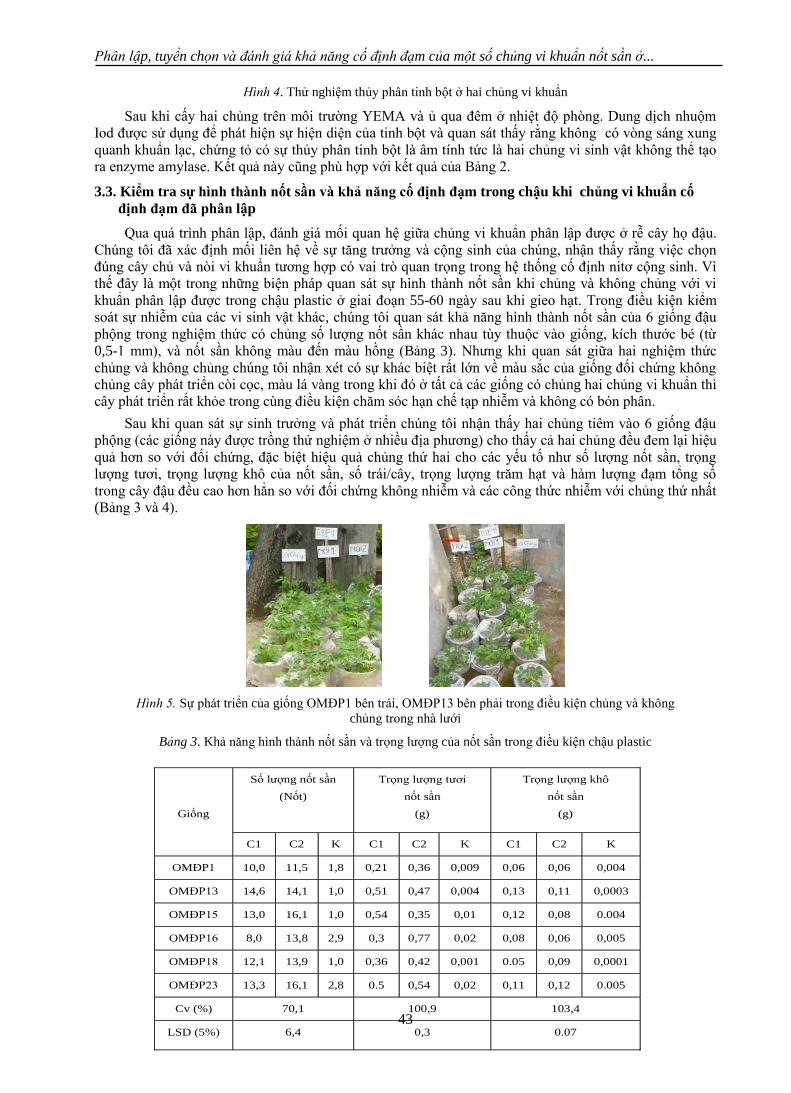
ình 4. Thử nghiệm thủy phân tinh bột ở hai chủng vi khuẩn
Sau khi cấy hai chủng trên môi trường YEMA và ủ qua đêm ở nhiệt độ phòng. Dung dịch nhuộm
Iod được sử dụng để phát hiện sự hiện diện của tinh bột và quan sát thấy rằng không có vòng sáng xung
quanh khuẩn lạc, chứng tỏ có sự thủy phân tinh bột là âm tính tức là hai chủng vi sinh vật không thể tạo
ra enzyme amylase. Kết quả này cũng phù hợp với kết quả của Bảng 2.
3.3. Kiểm tra sự hình thành nốt sần và khả năng cố định đạm trong chậu khi chủng vi khuẩn cố
định đạm đã phân lập
Qua quá trình phân lập, đánh giá mối quan hệ giữa chủng vi khuẩn phân lập được ở rễ cây họ đậu.
Chúng tôi đã xác định mối liên hệ về sự tăng trưởng và cộng sinh của chúng, nhận thấy rằng việc chọn
đúng cây chủ và nòi vi khuẩn tương hợp có vai trò quan trọng trong hệ thống cố định nitơ cộng sinh. Vì
thế đây là một trong những biện pháp quan sát sự hình thành nốt sần khi chủng và không chủng với vi
khuẩn phân lập được trong chậu plastic ở giai đoạn 55-60 ngày sau khi gieo hạt. Trong điều kiện kiểm
soát sự nhiễm của các vi sinh vật khác, chúng tôi quan sát khả năng hình thành nốt sần của 6 giống đậu
phộng trong nghiệm thức có chủng số lượng nốt sần khác nhau tùy thuộc vào giống, kích thước bé (từ
0,5-1 mm), và nốt sần không màu đến màu hồng (Bảng 3). Nhưng khi quan sát giữa hai nghiệm thức
chủng và không chủng chúng tôi nhận xét có sự khác biệt rất lớn về màu sắc của giống đối chứng không
chủng cây phát triển còi cọc, màu lá vàng trong khi đó ở tất cả các giống có chủng hai chủng vi khuẩn thì
cây phát triển rất khỏe trong cùng điều kiện chăm sóc hạn chế tạp nhiễm và không có bón phân.
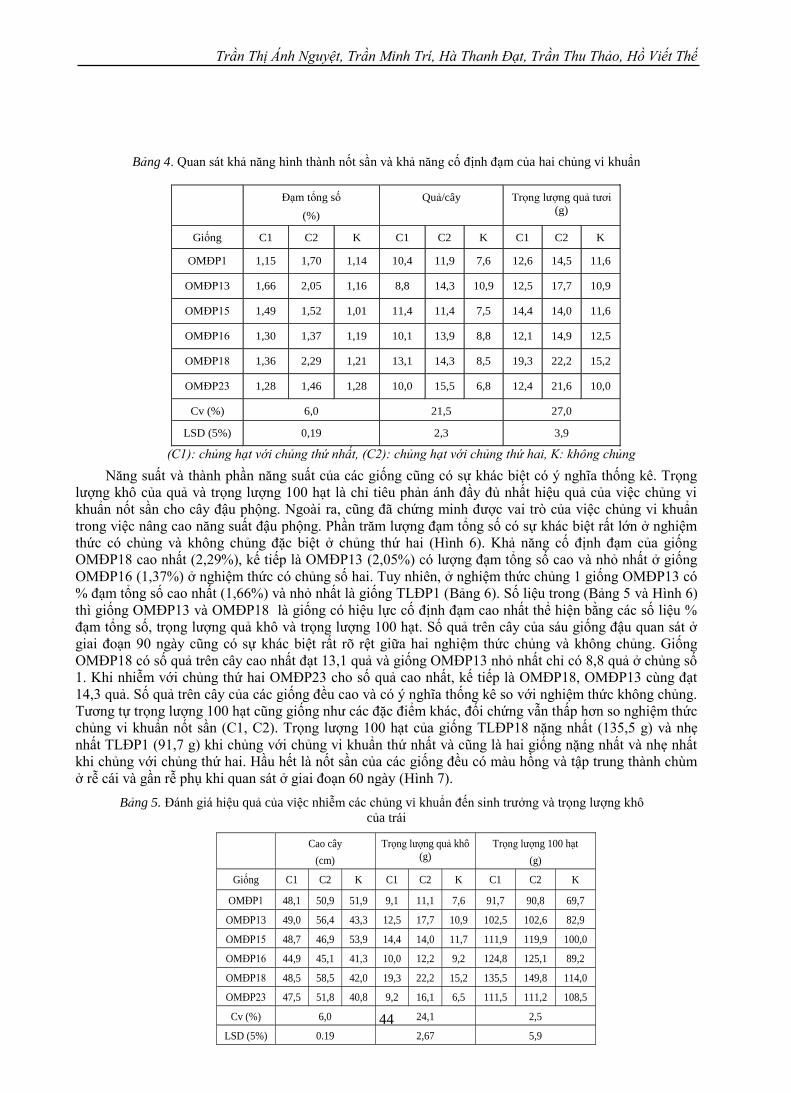
Sau khi quan sát sự sinh trưởng và phát triển chúng tôi nhận thấy hai chủng tiêm vào 6 giống đậu
phộng (các giống này được trồng thử nghiệm ở nhiều địa phương) cho thấy cả hai chủng đều đem lại hiệu
quả hơn so với đối chứng, đặc biệt hiệu quả chủng thứ hai cho các yếu tố như số lượng nốt sần, trọng
lượng tươi, trọng lượng khô của nốt sần, số trái/cây, trọng lượng trăm hạt và hàm lượng đạm tổng số
trong cây đậu đều cao hơn hẳn so với đối chứng không nhiễm và các công thức nhiễm với chủng thứ nhất
(Bảng 3 và 4).
ình 5 Sự phát triển của giống OMĐP1 bên trái, OMĐP13 bên phải trong điều kiện chủng và không
chủng trong nhà lưới
Bảng 3. Khả năng hình thành nốt sần và trọng lượng của nốt sần trong điều kiện chậu plastic
A B
Trần Thị nh Ngu ệt, Trần Minh Tr , à Thanh Đạt, Trần Thu Thảo, Viết Thế
44
Bảng 4. Quan sát khả năng hình thành nốt sần và khả năng cố định đạm của hai chủng vi khuẩn
): chủng hạt với chủng thứ nhất, ): chủng hạt với chủng thứ hai, K: kh ng chủng
Năng suất và thành phần năng suất của các giống cũng có sự khác biệt có ý nghĩa thống kê. Trọng
lượng khô của quả và trọng lượng 100 hạt là chỉ tiêu phản ánh đầy đủ nhất hiệu quả của việc chủng vi
khuẩn nốt sần cho cây đậu phộng. Ngoài ra, cũng đã chứng minh được vai trò của việc chủng vi khuẩn
trong việc nâng cao năng suất đậu phộng. Phần trăm lượng đạm tổng số có sự khác biệt rất lớn ở nghiệm
thức có chủng và không chủng đặc biệt ở chủng thứ hai (Hình 6). Khả năng cố định đạm của giống
OMĐP18 cao nhất (2,29%), kế tiếp là OMĐP13 (2,05%) có lượng đạm tổng số cao và nhỏ nhất ở giống
OMĐP16 (1,37%) ở nghiệm thức có chủng số hai. Tuy nhiên, ở nghiệm thức chủng 1 giống OMĐP13 có
% đạm tổng số cao nhất (1,66%) và nhỏ nhất là giống TLĐP1 (Bảng 6). Số liệu trong (Bảng 5 và Hình 6)
thì giống OMĐP13 và OMĐP18 là giống có hiệu lực cố định đạm cao nhất thể hiện bằng các số liệu %
đạm tổng số, trọng lượng quả khô và trọng lượng 100 hạt. Số quả trên cây của sáu giống đậu quan sát ở
giai đoạn 90 ngày cũng có sự khác biệt rất rõ rệt giữa hai nghiệm thức chủng và không chủng. Giống
OMĐP18 có số quả trên cây cao nhất đạt 13,1 quả và giống OMĐP13 nhỏ nhất chỉ có 8,8 quả ở chủng số
1. Khi nhiễm với chủng thứ hai OMĐP23 cho số quả cao nhất, kế tiếp là OMĐP18, OMĐP13 cùng đạt
14,3 quả. Số quả trên cây của các giống đều cao và có ý nghĩa thống kê so với nghiệm thức không chủng.
Tương tự trọng lượng 100 hạt cũng giống như các đặc điểm khác, đối chứng vẫn thấp hơn so nghiệm thức
chủng vi khuẩn nốt sần (C1, C2). Trọng lượng 100 hạt của giống TLĐP18 nặng nhất (135,5 g) và nhẹ
nhất TLĐP1 (91,7 g) khi chủng với chủng vi khuẩn thứ nhất và cũng là hai giống nặng nhất và nhẹ nhất
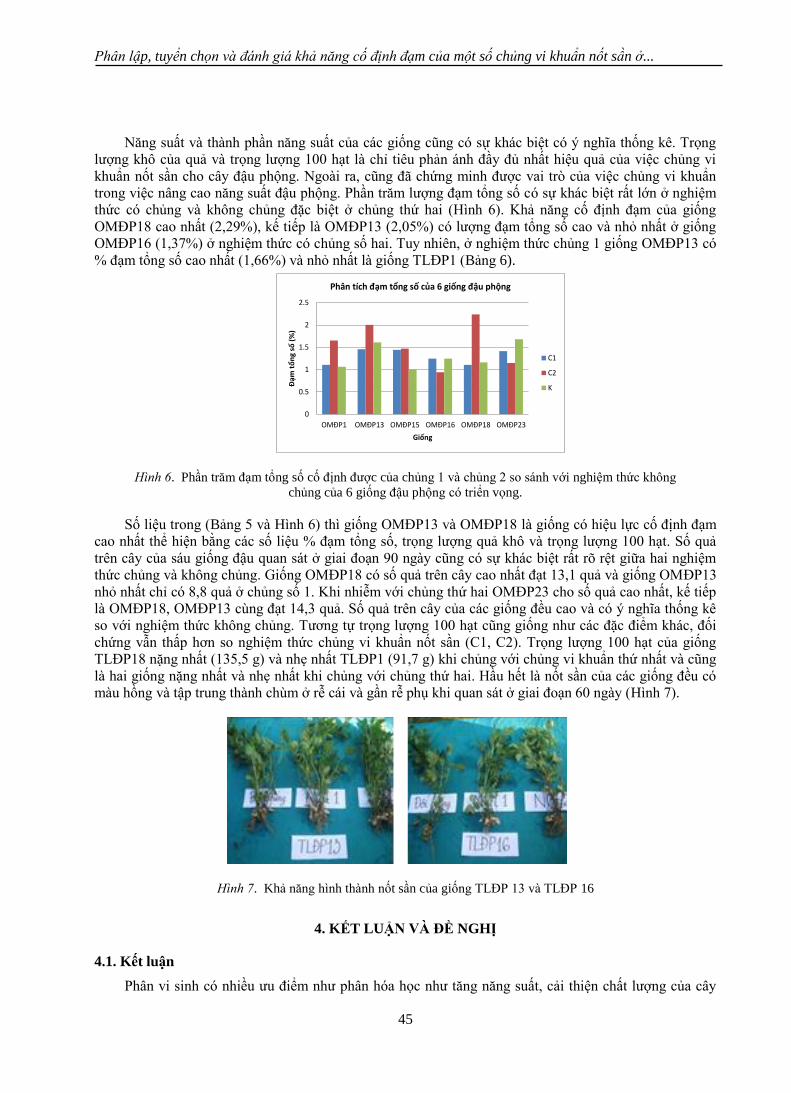
khi chủng với chủng thứ hai. Hầu hết là nốt sần của các giống đều có màu hồng và tập trung thành chùm
ở rễ cái và gần rễ phụ khi quan sát ở giai đoạn 60 ngày (Hình 7).
Bảng 5. Đánh giá hiệu quả của việc nhiễm các chủng vi khuẩn đến sinh trưởng và trọng lượng khô
của trái
Đạm tổng số
(%)
Quả/cây Trọng lượng quả tươi
(g)
Giống C1 C2 K C1 C2 K C1 C2 K
OMĐP1 1,15 1,70 1,14 10,4 11,9 7,6 12,6 14,5 11,6
OMĐP13 1,66 2,05 1,16 8,8 14,3 10,9 12,5 17,7 10,9
OMĐP15 1,49 1,52 1,01 11,4 11,4 7,5 14,4 14,0 11,6
OMĐP16 1,30 1,37 1,19 10,1 13,9 8,8 12,1 14,9 12,5
OMĐP18 1,36 2,29 1,21 13,1 14,3 8,5 19,3 22,2 15,2
OMĐP23 1,28 1,46 1,28 10,0 15,5 6,8 12,4 21,6 10,0
Cv (%) 6,0 21,5 27,0
LSD (5%) 0,19 2,3 3,9
Cao cây
(cm)
Trọng lượng quả khô
(g)
Trọng lượng 100 hạt
(g)
Giống C1 C2 K C1 C2 K C1 C2 K
OMĐP1 48,1 50,9 51,9 9,1 11,1 7,6 91,7 90,8 69,7
OMĐP13 49,0 56,4 43,3 12,5 17,7 10,9 102,5 102,6 82,9
OMĐP15 48,7 46,9 53,9 14,4 14,0 11,7 111,9 119,9 100,0
OMĐP16 44,9 45,1 41,3 10,0 12,2 9,2 124,8 125,1 89,2
OMĐP18 48,5 58,5 42,0 19,3 22,2 15,2 135,5 149,8 114,0
OMĐP23 47,5 51,8 40,8 9,2 16,1 6,5 111,5 111,2 108,5
Cv (%) 6,0 24,1 2,5
LSD (5%) 0.19 2,67 5,9
hân lập, tuyển chọn và đánh giá khả năng cố định đạm của một số chủng vi khu n nốt sần ở...
45
Năng suất và thành phần năng suất của các giống cũng có sự khác biệt có ý nghĩa thống kê. Trọng
lượng khô của quả và trọng lượng 100 hạt là chỉ tiêu phản ánh đầy đủ nhất hiệu quả của việc chủng vi
khuẩn nốt sần cho cây đậu phộng. Ngoài ra, cũng đã chứng minh được vai trò của việc chủng vi khuẩn
trong việc nâng cao năng suất đậu phộng. Phần trăm lượng đạm tổng số có sự khác biệt rất lớn ở nghiệm
thức có chủng và không chủng đặc biệt ở chủng thứ hai (Hình 6). Khả năng cố định đạm của giống
OMĐP18 cao nhất (2,29%), kế tiếp là OMĐP13 (2,05%) có lượng đạm tổng số cao và nhỏ nhất ở giống
OMĐP16 (1,37%) ở nghiệm thức có chủng số hai. Tuy nhiên, ở nghiệm thức chủng 1 giống OMĐP13 có
% đạm tổng số cao nhất (1,66%) và nhỏ nhất là giống TLĐP1 (Bảng 6).
ình 6. Phần trăm đạm tổng số cố định được của chủng 1 và chủng 2 so sánh với nghiệm thức không
chủng của 6 giống đậu phộng có triển vọng.
Số liệu trong (Bảng 5 và Hình 6) thì giống OMĐP13 và OMĐP18 là giống có hiệu lực cố định đạm
cao nhất thể hiện bằng các số liệu % đạm tổng số, trọng lượng quả khô và trọng lượng 100 hạt. Số quả
trên cây của sáu giống đậu quan sát ở giai đoạn 90 ngày cũng có sự khác biệt rất rõ rệt giữa hai nghiệm
thức chủng và không chủng. Giống OMĐP18 có số quả trên cây cao nhất đạt 13,1 quả và giống OMĐP13
nhỏ nhất chỉ có 8,8 quả ở chủng số 1. Khi nhiễm với chủng thứ hai OMĐP23 cho số quả cao nhất, kế tiếp
là OMĐP18, OMĐP13 cùng đạt 14,3 quả. Số quả trên cây của các giống đều cao và có ý nghĩa thống kê
so với nghiệm thức không chủng. Tương tự trọng lượng 100 hạt cũng giống như các đặc điểm khác, đối
chứng vẫn thấp hơn so nghiệm thức chủng vi khuẩn nốt sần (C1, C2). Trọng lượng 100 hạt của giống
TLĐP18 nặng nhất (135,5 g) và nhẹ nhất TLĐP1 (91,7 g) khi chủng với chủng vi khuẩn thứ nhất và cũng
là hai giống nặng nhất và nhẹ nhất khi chủng với chủng thứ hai. Hầu hết là nốt sần của các giống đều có
màu hồng và tập trung thành chùm ở rễ cái và gần rễ phụ khi quan sát ở giai đoạn 60 ngày (Hình 7).
ình 7. Khả năng hình thành nốt sần của giống TLĐP 13 và TLĐP 16
4. KẾT LUẬN VÀ ĐỀ NGHỊ
4.1. Kết luận
Phân vi sinh có nhiều ưu điểm như phân hóa học như tăng năng suất, cải thiện chất lượng của cây
0
0.5
1
1.5
2
2.5
OMĐP1 OMĐP13 OMĐP15 OMĐP16 OMĐP18 OMĐP23
Đạ
m t
ổn
g số
(%
)
Giống
Phân tích đạm tổng số của 6 giống đậu phộng
C1
C2
K

Trần Thị nh Ngu ệt, Trần Minh Tr , à Thanh Đạt, Trần Thu Thảo, Viết Thế
46
trồng, giảm chi phí sản suất, ngoài ra còn góp phần bảo vệ sinh thái môi trường và nông nghiệp bền vững.
Tuy nhiên, tình hình sản xuất phân vi sinh ở nước ta vẫn chưa đáp ứng của nhu cầu thực tiển do nhiều
nguyên nhân, trong đó người tiêu dùng chưa mạnh dạn sử dụng là do chất lượng sản phẩm vẫn chưa ổn
định. Để hoàn thiện nâng cao chất lượng chế phẩm phân vi sinh cố định đạm thì việc phân lập, tuyển chọn
chính xác là khâu quan trọng nhất thì nghiên cứu này đã cung cấp được ý nghĩa khoa học cơ bản.
Đã hoàn thiện được các bước phân lập, tuyển chọn và xác định nhanh vi khuẩn nốt sần trong rễ cây
đậu phộng. Trong số các chủng phân lập được bước đầu xác định được 6 chủng (NS1-NS6) có đặc điểm
và hình dạng khuẩn lạc tương đối điển hình. Qua quá trình sàng lọc bằng các phản ứng sinh lý, sinh hóa,
đã sơ tuyển chỉ được hai chủng có khả năng phát triển mạnh, thuộc nhóm mọc nhanh, hiếu khí, có khả
năng chịu được pH cao, chịu mặn. Khuẩn lạc này không bắt màu với thuốc nhuộm Congo red, không phát
triển trên môi trường peptone và kháng sinh. Khi quan sát dưới kính hiển vi có khả năng di động, hình
que và là vi khuẩn gram âm.
Tuyển chọn được hai chủng vi khuẩn có khả năng tăng trưởng trên môi trường YEMA có bổ sung
thêm nồng độ muối 2% và phát triển rất tốt trên môi trường kiềm pH từ 7-10. Mặt khác cả hai nòi cho kết
quả âm tính khi thử nghiệm tinh bột, chỉ có khả năng phát triển và tiêu thụ trong môi trường có chứa
nguồn đường manitol, phát triển rất yếu trong môi trường có chứa glucose và lactose. Về khả năng tạo
gum, chủng thứ nhất có khả năng tạo gum cao nhất 0,91 mg và thấp hơn ở chủng thứ hai 0,89 mg. Khi
chủng hai dòng vi khuẩn này riêng lẻ cho từng giống đậu phộng được trồng invitro trong nhà lưới, kết
luận rằng hai chủng vi khuẩn tiếp tục cho khả năng cố định đạm cao, và khả năng hình thành nốt sần so
với nghiệm thức không chủng. Các nghiệm thức dùng vi khuẩn nhiễm hạt đều có số lượng, khối lượng
nốt sần, trọng lượng 100 hạt, trọng lượng quả tươi, trọng lượng khô lớn hơn một cách rõ rệt so với đối
chứng.
Về ý nghĩa thực tiển kết quả của đề tài là cơ sở cho việc phân lập và chọn lọc các dòng vi khuẩn cố
định đạm trong rễ của cây lạc, có hoạt tính cố định đạm cao để sản xuất chế phẩm vi sinh.
4.2. Đề nghị
Do thời gian tiến hành nghiên cứu còn ngắn và giới hạn chỉ trong một vụ, nên để có kết quả chính
xác và đầy đủ hơn, đề nghị tiếp tục tiến hành thí nghiệm ở các vụ tiếp theo, và trên các chân đất khác
nhau để có kết luận hoàn thiện hơn.
Sử dụng sự hỗ trợ của sinh học phân tử để giúp sàn lọc, định danh vi khuẩn cố định đạm nhanh
chóng, chính xác giúp rút ngắn thời gian.
Quan sát khả năng hình thành nốt sần với cây đậu phộng trồng thử nghiệm trong điều kiện tự nhiên
có bổ sung lượng phân bón thích hợp.
Dựa vào chủng vi khuẩn phân lập được tiếp tục nghiên cứu các chế phẩm phân bón vi khuẩn nốt sần
ở dạng rắn, lỏng hay bán rắn cho đậu phộng.
TÀI LIỆU THAM KHẢO
1. Bilal R; Rarul G; Queshi J. A; Malik K.A. - Characterization of Azospirillum and related
diazotrophs associated with roots of plants growing in saline soils. World J Microbiol 6 (1990) 46-
59.
2. Somers E. D; Patcek P; Gysegom M. Srinivasan and J. Vanderleyden. - Azospirillum brasilense
produces the auxin-like phenylacetic acid by using yhe key enzyme for indole-3-acetic acid
biosynthesis. App Environ Microbiol 71 (2005) 1803-1810.
3. Lăng Ngọc Dậu, Nguyễn thị Xuân My, Cao Ngọc Diệp. - Khả năng cố định đạm, hòa tan lân và
sinh tổng hợp IAA của vi khuẩn Azospirillum lipoferum. (2007) Viện công nghệ sinh hoc, Đại học
cần thơ.
4. Tran Thi Ngoc Son; Cao Ngoc Diep and Truong Thi Minh Giang. -Effect of Bradyrhizobia and
Phosphate solubilizing bacteria application on soybean in rotational system in the Mekong Delta.
Omon Rice 14 (2006) 48-57.

hân lập, tuyển chọn và đánh giá khả năng cố định đạm của một số chủng vi khu n nốt sần ở...
47
5. Fred E.B., Baldwin I.L; McMoy E. -Root nodule bacteria and leguminous plants. Univ of
Wisconsin Studies in Science 5 (1932) 343-356.
6. Hahn NJ. -The congored raction in bacteria and its use-fulness in the identification of rhizobia. Can
J Microbiol 12 (1996) 725-33.
7. Kleczkowska Nutman PS. -The identification and classification of rhizobia ‘Identification methods
for mi-crobiologists; Part B ed. B.M. Gibbs and D.A. Shapton Academic press, London and New
York, (1968) pp 51.
8. Hofer AW. - Method for distinguishing between legume bacteria amd their most common
contaminant. J Amer Soc Agron 27 (1935) 228-30.
9. Vincent J.M. A manual for the practical study of the root nodule bacteria. IPB Handbook 1970 no.
15. England; Black Well Scientific Publications Ltd., Oxford.
10. Xu L. M., Ge C., Cui Z., Li J., Fan H. Bradyrhizobium liaoningense sp. nov., isolated from the
root nodules of soybean. Int J Syst Evol Microbiol 45 (1995) 706-711.
11. Bremner J.M. - Determination of nitrogen in soil by the Kjeldahl method. The Journal of
Agricultural Science 55 (1960) 11-33.
ABSTRACT
ISOLATION AND CHARACTERIZATION OF NITROGEN FIXING BACTERIA COLLECTED
FROM ROOT NODULES OF PEANUT (Arachis hypogaea. L)
Tran Anh Nguyet1, *
, Tran Minh Tri1, Ha Thanh Đat
1, Tran Thu Thao
1, Ho Viet The
2
1Cuu Long Delta Rice Research Institute,
2HCM University of Food Industry
*Email: [email protected]
Six isolated strains of nitrogen fixing bacteria isolated from root nodules of peanut (Arachis
hypogaea. L). Through identifying of characteristics, morphology, physiology, biochemistry, only two
isolates meet the above requirements. Fixing nodules bacteria grow well on YEMA medium, gram
negative, rod shape, aerobic, mobility ability, showed good growth on Congo red yeast extract mannitol
agar (YEMA) medium and did not produce chromo genesis on Congo red medium, not formed colonies
on Glucose-Peptone-Agar medium, they able to grow in NaCl 0.5- 2% and growing Hofer’s alkaline
broth (pH 7-10). On the other hand, two isolates gave negative results for intrinsic antibiotic resistance
and starch hydrolysis. Isolate 1 produced maximum gum production 0.91 mg and minimum isolate 2
produced 0.89 mg. Glucose and lactose consumption were very weak compared with mannitol carbon
source. Two strains of bacteria continue to demonstrate a high nitrogen-fixing ability through the ability
to form nodules and percentage total nitrogen in net house condition. The treatments used infectious
bacteria showed the significance difference with without treatments the number of nodules, 100 seeds
weight, plant height and plant fresh weight significantly greater than compared to without treatment. On
the basis of result obtained in the present observation it can be concluded that isolate two strains were
found to be the best isolate of nitrogen fixing bacteria and can be exploited as bio-fertilizers for better
peanut yield production and reduced cost production.
Keywords: nitrogen fixing bacteria, peanut, root nodules, YEMA.