phenology of paralemanea mexicana (batrachospermales, rhodophyta) in a...
TRANSCRIPT

Phenology of Paralemanea mexicana (Batrachospermales,Rhodophyta) in a high-altitude stream in central MexicoJavier Carmona Jiménez,* Miriam Guadalupe Bojorge García and Rocío Ramírez RodríguezEcology and Natural Sources, Faculty of Sciences, National Autonomous University of Mexico (UNAM), CiudadUniversitaria, Coyoacan, CP 04510, Mexico
SUMMARYThe morphology and phenology of Paralemaneamexicana (Kützing) Vis et Sheath was evaluated sea-sonally in a fifth order high-altitude stream in centralMexico. The gametophytes grew during oligotrophicand eutrophic conditions, and during particularmicrohabitat conditions: high current velocity (40–240 cm s−1), low to medium irradiance (5–973 μmolphotons m−2 s−1), and shallow depth (1–30 cm). Theabundance of gametophytes was positively correlatedwith low temperature, high current velocity and concen-trations of soluble reactive phosphorus. Interestingly,monoecious gametophytes formed two types ofbranches, true branches with sympodial pattern origi-nating from meristematic cells in variable number, andabundant false branches produced during the develop-ment of the ‘Chantransia’ stage in the surface of thegametophyte or by uniseriate filaments arising withinthe thallus lumen. These filaments generally producegametophytes and suggest that they could support thegermination of carpospores. The frequently whorledbranches are the result of a false branching patternand are exclusive to P. mexicana. These morpholo-gical and reproductive characteristics appear to bebiomechanical adaptations to avoid detachment andincrease reproductive success. Efficient reproductivestrategies of P. mexicana observed in this study can beinterpreted as adaptations to successfully colonizestreams; however, these features may not have beencommon in the study region due to restrictedmicrohabitat conditions and geographic isolation.
Key words: Batrachospermales, ecology, Paralemaneamexicana, phenology, Rhodophyta, stream.
INTRODUCTION
The red algal orders Batrachospermales is a conspicu-ous group of freshwater Rhodophyta that occurs widelyin lotic ecosystems throughout the world (Sheath &Hambrook 1990) and appear to be highly specializedto these ecosystems. Batrachospermalean taxa haveseveral morphological and reproductive adaptations
that could be linked to unidirectional current velocityFor example, morphological adaptation related withthe formation of rhizoidal filaments, or aggregation offilaments, from which an efficient attachment systemis produced (Carmona et al. 2009); abundance ofbranches or dense growth forms can reduce drag force(Sheath & Hambrook 1990); and presence of largeamounts of mucilage surrounding the thallus (Carmonaet al. 2006). As reproductive adaptations are somaticmeiosis, clonal multiplication in the ‘Chantransia’ stage(Sheath 1984; Necchi & Carmona 2002) and abun-dant spermatangia and carposporangia small in size(Carmona et al. 2009). Therefore, in the lotic ecosys-tems, water flow can be considered the environmentalparameter that is most important to the development ofthe gametophyte, reproduction, and evolutionary adap-tations of these algae.
Traditionally, the genus Paralemanea was placed inthe family Lemaneaceae (Vis & Sheath 1992); however,using molecular data, Entwisle et al. (2009) proposed amajor revision of Batrachospermales and amended thecircumscriptions of the family Batrachospermaceae toinclude Lemaneaceae and Psilosiphonaceae. The genusParalemanea genera was retained pending furtherinvestigation, noting the possibility that Paralemanea isparaphyletic in relation to Lemanea (Vis et al. 1998;Kapraun et al. 2007; Entwisle et al. 2009).
Paralemanea has a pseudoparenchymatous tubeconstruction, with interwoven medullary filamentsabsent (Sheath et al. 1996a). Three species ofParalemanea have been recognized for North America,P. annulata (Kützing) M.L. Vis et Sheath, P. catenata(Kützing) M.L. Vis et Sheath and P. mexicana (Kützing)M.L.Vis et Sheath (Vis et al. 1992). Species delineationis based on the thallus length and frequency ofbranches, and P. mexicana is characterized by awhorled branch and rebranching pattern. This speciesis localized in high mountain streams in central Mexico,
*To whom correspondence should be addressed.Email: [email protected] editor: J. H. Kim.Received 5 July 2012; accepted 14 September 2013.doi: 10.1111/pre.12042
Phycological Research 2014; 62: 86–93
bs_bs_banner
© 2013 Japanese Society of Phycology

geographically isolated from the other species of thegenus (Vis et al. 1992). Some studies of the reproduc-tive success and life-history of Paralemanea specieshave been conducted in North America (Filkin & Vis2004) and the Iberian Peninsula (Carmona et al.2011). Paralemanea annulata gametophytes start togrow in winter, reproduce in spring, and releasecarpospores in early summer. The gametophytes disap-pear during low water and increased temperature con-ditions; environmental parameters that influence thephenological characteristics include current velocity,water depth, light intensity, water temperature and thepresence of epiphytic Audouinella sp. (Filkin & Vis2004). The gametophyte of P. catenata is presentthroughout the year, with the highest population coverin winter and the release of carpospores in the springand summer (Carmona et al. 2011). Similar ecologicalrequirements, such as water temperature and currentvelocity, were noted for both populations and seemed toplay a role in the phenology of these Paralemaneapopulations; however, no one environmental parameterwas observed to be most influential (Carmona et al.2011; Filkin & Vis 2004). Several factors were hypoth-esized to interact during the trigger events of thephenology of P. annulata (Filkin & Vis 2004).
Paralemanea mexicana has been collected in severalstreams from high mountain and temperate waters fromcentral Mexico (Carmona & Necchi 2002; Bojorge et al.2010). However, the ecology is poorly known and therehas been no characterization of gametophytes season-ally. The present investigation, based on seasonalobservations of a population of P. mexicana in a highaltitude stream from central Mexico, was conducted todescribe morphological and reproductive characteris-tics, as well as the environmental conditions that influ-ence gametophytes occurrence.
MATERIALS AND METHODS
Sample collection
The material used in the present study was collectedfrom the Amanalco river; a fifth-order stream segmentin a mountainous region of central Mexico (elevation1890 m; 19°13′N, 100°07′W). Field work was con-ducted from October 2007 to November 2008, includ-ing the contrasting rainy and dry seasons. Threesamplings were conducted during the early (December),middle (February) and end of the dry season (May;termed ‘dry’); December and February were the coldestmonths (termed; ‘cool dry’). Three more samplings wereconducted during the middle of the rainy season (Sep-tember, October and November; termed ‘rainy’).
Observations were made on natural substrata (boul-ders) directly in the stream bed. Water temperature andspecific conductance were measured with a Conductronic
(Puebla, Mexico) PC-18 conductivity meter. Temporalvariations were monitored by the quadrat technique(Necchi et al. 1995), which evaluates the influence ofvariables at the microhabitat level (current velocity, depthand underwater irradiance) on the vegetative and repro-ductive characteristics of the population. Each samplingdate consisted of a stream segment of 10 m long. Thesampling size consisted of five quadrats, each separatedby 2 m. Each sampling quadrat was a circle of 10 cmradius; random numbers between 0 and 180° determinedtheir locations. The individuals were collected from themiddle of the sampling unit.
Chemical analyses
Dissolved nutrients were sampled by filtering 30 mL ofstream water through 0.45 and 0.22 μm pore diametermembranes (in situ). Samples were preserved withchloroform and frozen until measured in the laboratory,with a multichannel analyzer following standard titra-tion. Samples for dissolved inorganic nitrogen (DIN)and soluble reactive phosphorous (SRP) were kept incold conditions until analyses were completed (APHAet al. 1980). Water samples for determination of anionsand pH were preserved frozen in the dark; samples forcations were preserved with 40% nitric acid (pH 2–3).Determination of carbonates was performed usingthe titration method, chlorides by the selectiveelectrode method, hardness by the titration methodwith ethylenediaminetetraacetic acid, and Na+ and K+
by the spectrophotometric atomic absorption method(Greenberg et al. 1985).
Microhabitat characterization
Microhabitat characteristics were recorded on all sam-pling dates. Microhabitat variables were measured insitu at the center of five replicate quadrats of eachsampling unit: current velocity and irradiance weremeasured as close as possible to the algae using aSwoffer 3100 current velocity meter (Swoffer Instru-ments, Inc., Seattle, WA) and a Li-Cor LI-1000quantum meter (LI-COR Corporate, Lincoln, NE) with aflat subaquatic sensor of photosynthetically activeradiation, respectively. The coefficient of variation(VC = SE/a·100, where SE = standard error anda = average) was used to evaluate the variability ofenvironmental parameters. A VC value < 10% indicatedrelatively stable parameters; a value >10% indicatedthose that changed over time and space, usually aresult of dilution/evaporation processes and biologicalactivity (sensu Margalef 1983).
Gametophyte characterization
The number of thalli (gametophytes) within each sam-pling unit was recorded by visual estimation using a
© 2013 Japanese Society of Phycology
87Phenology of Paralemanea mexicana

175 cm2 viewfinder. The ‘Chantransia’ stage was notobserved in natural conditions. Twenty-five thalli wererandomly selected (five in each sampling unit) andpreserved in 3% formaldehyde for subsequent analysisin the laboratory. The following morphological andphenological characteristics were defined during pre-liminary tests and previous research: thallus height;number of true and false branches; number of nodeswith spermatangia; number of visible carposporophytes;number of gametophytes with open tips; and number of‘Chantransia’ stages associated with the gametophytesurface (Vis et al. 1992; Sheath et al. 1996b; Carmona& Necchi 2002; Filkin & Vis 2004). In addition, epi-phytic and aquatic invertebrates were identified fromeach thallus to examine species associations (Merrit& Cummins 1996; Wiggins 1996). Olympus BX51and SZX7 microscopes with a DP12 and E-330microphotography system were used for microscopicanalyses.
Statistics
Kruskal–Wallis test were performed to assess significantdifferences in environmental and morphometric meas-urements among six samplings dates. When tests weresignificant, the Mann–Whitney test was conductedamong all dates to detect which differed significantly.Associations among morphometric, reproductive data,and microhabitat variables during the six samplingdates were assessed using the Spearman correlationcoefficient (Gotelli & Ellison 2004). The analyses wereconducted using SPSS 18 software (Levesque 2006).
RESULTS
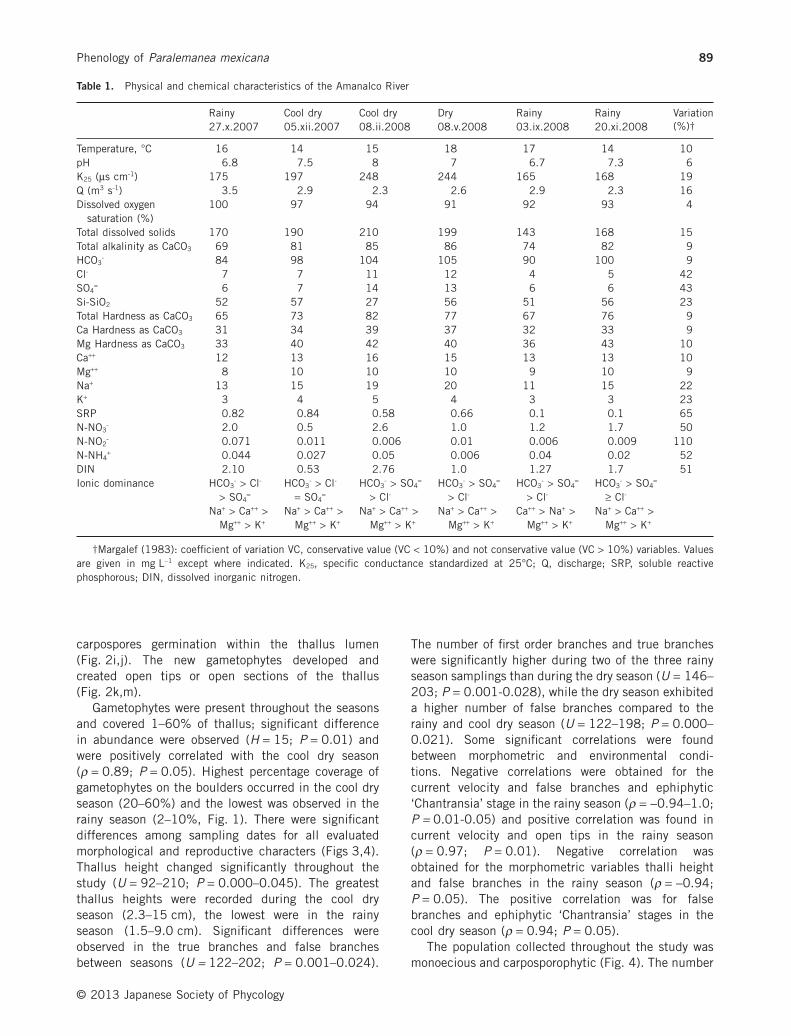
During the six collection periods of the year, the riverwater exhibited relatively low ionic content (143–210 mg L−1 of total dissolved solids; specific conduct-ance 165–248 μS cm–1), with a near neutral pH (6.7–8), mild temperature (14–18°C), high dissolved oxygensaturation (91–100%), low total alkalinity (74–86 mgCaCO3 L−1) and a dominance of sodium and bicarbonate(Table 1). The physical characteristics were consistentduring the six sampling periods (VC < 10%). Theremaining physical and chemical variables (discharge,specific conductance, total dissolved solids, sodium,potassium, chloride, sulfate, nitrate, nitrite, ammo-nium, dissolved silica and SRP) exhibited larger vari-ations (VC 15–110%), with increased values duringcool dry and dry seasons. Changes in DIN were great,probably by the contribution of upstream agriculturalactivities. Gametophytes occurred predominantly inmicrohabitat conditions that exhibited fast currentvelocity (40–240 cm s−1), low to medium irradiance(5–973 μmol photons m−2 s−1), shallow depth(1–30 cm), and boulders as a substratum (Fig. 1). The
Kruskal–Wallis test revealed significant differences forcurrent velocity (H = 12; P = 0.04) and irradiance(H = 24; P < 0.00) among sampling dates.
Based on taxonomic characteristics, the specimenswere ascribed to P. mexicana (Fig 2a–m). The thalliwere generally branched, and branches of first andsecond order were usually whorled (Fig. 2a). Two typesof branches were identified. The first type, truebranches, originated from meristematic cells thatdivide in the apical region of thallus in variable number(Fig. 2b,c). The meristematic cells produce severalapical or intercalary branches along the thallus(Fig. 2d,e). The second type, false branches, were pro-duced by: (i) the development of the ‘Chantransia’stage at the surface of the gametophyte and the originof abundant new gametophytes (Fig. 2f–h), and (ii)uniseriate filaments arising from the supposed
Fig. 1. Current velocity, irradiance, depth and percentage cover
in quadrats with Paralemanea mexicana. Sampling dates indi-
cated with the same letter do not significantly differ (Mann–
Whitney test, α = 0.05). n = 25 (average ± 1 SE).
© 2013 Japanese Society of Phycology
88 J. C. Jiménez et al.
carpospores germination within the thallus lumen(Fig. 2i,j). The new gametophytes developed andcreated open tips or open sections of the thallus(Fig. 2k,m).
Gametophytes were present throughout the seasonsand covered 1–60% of thallus; significant differencein abundance were observed (H = 15; P = 0.01) andwere positively correlated with the cool dry season(ρ = 0.89; P = 0.05). Highest percentage coverage ofgametophytes on the boulders occurred in the cool dryseason (20–60%) and the lowest was observed in therainy season (2–10%, Fig. 1). There were significantdifferences among sampling dates for all evaluatedmorphological and reproductive characters (Figs 3,4).Thallus height changed significantly throughout thestudy (U = 92–210; P = 0.000–0.045). The greatestthallus heights were recorded during the cool dryseason (2.3–15 cm), the lowest were in the rainyseason (1.5–9.0 cm). Significant differences wereobserved in the true branches and false branchesbetween seasons (U = 122–202; P = 0.001–0.024).
The number of first order branches and true brancheswere significantly higher during two of the three rainyseason samplings than during the dry season (U = 146–203; P = 0.001-0.028), while the dry season exhibiteda higher number of false branches compared to therainy and cool dry season (U = 122–198; P = 0.000–0.021). Some significant correlations were foundbetween morphometric and environmental condi-tions. Negative correlations were obtained for thecurrent velocity and false branches and ephiphytic‘Chantransia’ stage in the rainy season (ρ = −0.94–1.0;P = 0.01-0.05) and positive correlation was found incurrent velocity and open tips in the rainy season(ρ = 0.97; P = 0.01). Negative correlation wasobtained for the morphometric variables thalli heightand false branches in the rainy season (ρ = −0.94;P = 0.05). The positive correlation was for falsebranches and ephiphytic ‘Chantransia’ stages in thecool dry season (ρ = 0.94; P = 0.05).
The population collected throughout the study wasmonoecious and carposporophytic (Fig. 4). The number
Table 1. Physical and chemical characteristics of the Amanalco River
Rainy Cool dry Cool dry Dry Rainy Rainy Variation(%)†27.x.2007 05.xii.2007 08.ii.2008 08.v.2008 03.ix.2008 20.xi.2008
Temperature, °C 16 14 15 18 17 14 10pH 6.8 7.5 8 7 6.7 7.3 6K25 (μs cm-1) 175 197 248 244 165 168 19Q (m3 s-1) 3.5 2.9 2.3 2.6 2.9 2.3 16Dissolved oxygen
saturation (%)100 97 94 91 92 93 4
Total dissolved solids 170 190 210 199 143 168 15Total alkalinity as CaCO3 69 81 85 86 74 82 9HCO3
- 84 98 104 105 90 100 9Cl- 7 7 11 12 4 5 42SO4
= 6 7 14 13 6 6 43Si-SiO2 52 57 27 56 51 56 23Total Hardness as CaCO3 65 73 82 77 67 76 9Ca Hardness as CaCO3 31 34 39 37 32 33 9Mg Hardness as CaCO3 33 40 42 40 36 43 10Ca++ 12 13 16 15 13 13 10Mg++ 8 10 10 10 9 10 9Na+ 13 15 19 20 11 15 22K+ 3 4 5 4 3 3 23SRP 0.82 0.84 0.58 0.66 0.1 0.1 65N-NO3
- 2.0 0.5 2.6 1.0 1.2 1.7 50N-NO2
- 0.071 0.011 0.006 0.01 0.006 0.009 110N-NH4
+ 0.044 0.027 0.05 0.006 0.04 0.02 52DIN 2.10 0.53 2.76 1.0 1.27 1.7 51Ionic dominance HCO3
- > Cl-
> SO4=
HCO3- > Cl-
= SO4=
HCO3- > SO4
=
> Cl-HCO3
- > SO4=
> Cl-HCO3
- > SO4=
> Cl-HCO3
- > SO4=
≥ Cl-
Na+ > Ca++ >Mg++ > K+
Na+ > Ca++ >Mg++ > K+
Na+ > Ca++ >Mg++ > K+
Na+ > Ca++ >Mg++ > K+
Ca++ > Na+ >Mg++ > K+
Na+ > Ca++ >Mg++ > K+
†Margalef (1983): coefficient of variation VC, conservative value (VC < 10%) and not conservative value (VC > 10%) variables. Valuesare given in mg L−1 except where indicated. K25, specific conductance standardized at 25°C; Q, discharge; SRP, soluble reactivephosphorous; DIN, dissolved inorganic nitrogen.
© 2013 Japanese Society of Phycology
89Phenology of Paralemanea mexicana

of spermatangial rings, number of carposporophytes andopen tips exhibited significant differences betweensampling dates (U = 43–212; P = 0.000–0.04). Thespermatangial rings and carposporohytes were signifi-cantly higher during November, December and February(cooler months) than during May, October and Septem-
ber (warmer months, U = 19–196; P = 0.0–0.019). Thegreatest number of open tips (U = 140–210; P =0.001–0.04) and the number of epiphytic ‘Chantransia’stage (U = 53–207; P = 0.000–0.03) were observedduring the warm dry season. One correlation was foundbetween the thallus height and the highest number of
Fig. 2. Morphological and reproductive features of Paralemanea mexicana. (a) General habit and branches usually whorled. (b)
Meristematic cells (arrows) in the apical region. (c) Meristematic cells (arrows) along the apical region. (d) True branches in the apical
region of thallus. (e) True branches in the apical and intercalar region of thallus. (f) ‘Chantransia’ stage and rhizoidal system with the origin
of abundant new gametophytes (arrow). (g) Epiphytic ‘Chantransia’ stage in apical region of the gametophyte. (h) Cross section of
the cortex showing ‘Chantransia’ stage and new gametophytes (arrows). (i) Uniseriate filaments arising from the carpospores within the
thallus lumen. (j) Detail of the uniseriate filament arising from a carpospore. (k) Open tip section of thallus with new gametophytes. (l) Open
lateral section of thallus with new gametophytes. (m) Damaged section of thallus with new gametophytes (arrow). Scale bar: 1 cm for Figs.
a–h, k–m; 30 μm for Figs. i, j.
© 2013 Japanese Society of Phycology
90 J. C. Jiménez et al.

carposphorophytes and spermatangial rings (ρ = 0.90;P = 0.05) during the cool dry season.
Paralemanea mexicana occurrence was associatedwith other freshwater Rhodophyceae: Batrachosper-mum gelatinosum (Linnaeus) De Candolle and Sirodotiasuecica Kylin and several epiphytic and metaphyticcyanobacteria: Blennothrix heterotricha (Gomont exGomont) Anagnostidis et Komarek, Phormodium inter-ruptum Kützing ex Gomont and Placoma regulare Broadyet Ingerfield. Epiphytic aquatic larvae and pupae inver-tebrates of the orders Simuliidae (Simulium sp.),Chironomidae (Rheotanytarsus sp.) and Hydroptilidae(Hydroptila sp. and Atopsyche sp.) built cases on thegametophyte. A positive correlation was observedbetween the numbers of epiphytic ‘Chantransia’ stages,the number of insect cases, and the number of associ-ated species in May (dry season, ρ = 0.90–0.97;P = 0.05–0.01).
DISCUSSION
The stream conditions observed during the currentstudy were similar to previous records in high mountainstreams in central Mexico (Carmona & Necchi 2002;Bojorge et al. 2010): mild temperature (12–16°C),slightly acid to neutral pH (5.5–7.6), moderate specificconductance (169–248 μS cm−1), rocky substrata, andmedium to fast current velocity (>35 cm s−1). Thestream segment showed both oligotrophic to eutrophicconditions during the year: DIN 0.02–2 mg L−1 andSRP 0.2–0.8 mg L−1 (Dodds 2003). According toBowman et al. (2005), enrichment of the limiting nutri-ent in mountain streams can increase the abundance ofbenthic algae, which could contribute to the increasedpercent cover recorded during the cold dry season atthis site.
The highest cover percent of gametophytes wasobserved during the cool season and indicates theirassociation with low temperature, and conversely, withlow cover percent during the rainy season with highdischarge. This result is comparable with other popu-lations of Paralemanea species from high-altitudestreams from tropical climates in Central and SouthAmerica; e.g. Paralemanea annulata (Necchi & Zucchi1995) and P. mexicana (Carmona et al. 2004; Bojorgeet al. 2010). The preference for cool temperatures(<20°C) seems to be a common feature among speciesof Batrachospermales collected in temperate regions;for example, Lemanea fluviatilis (Thirb & Benson-Evans1984, 1985; Carmona et al. 2011), P. annulata (Filkin& Vis 2004), P. catenata (Kucera & Marvan 2004;Carmona et al. 2011), as well as populations analyzedunder culture conditions: P. catenata and Paralemaneasp. (Necchi & Carmona 2002).
The gametophytes of the P. mexicana population incentral Mexico exhibited several peculiar characteris-
Fig. 3. Morphometric characteristics measured for Paralemanea
mexicana. Sampling dates indicated with the same letter do not
significantly differ (Mann–Whitney test, α = 0.05). n = 25
(average ± 1 SE).
Fig. 4. Reproductive characteristics measured for Paralemanea
mexicana. Sampling dates indicated with the same letter do not
significantly differ (Mann–Whitney test, α = 0.05). n = 25
(average ± 1 SE).
© 2013 Japanese Society of Phycology
91Phenology of Paralemanea mexicana

tics. They contained true branches with a sympodialbranching pattern. This branching pattern is importantto taxonomically differentiate specimens of P. mexicana(with branches) and P. catanata and P. annulata(without branches; Vis et al. 1992). The frequentlywhorled branches are the result of a false branchedpattern and are apparently exclusive to P. mexicana.Considering that true and false branches are distin-guishing taxonomic features, we concluded that thenumber and branching pattern are variable and corre-lated with environmental changes, especially withchanges in current velocity.
The correlation between the number of open tips(i.e. greater number of carpospores released) and thenumber of epiphytic ‘Chantransia’ stages can result inthe highest number of false branches in the dry season.In P. annulata, the increase of open tips is thought tooccur earlier in the areas with higher current velocity,due to abrasion from sand and detritus (Filkin & Vis2004).
Gametophytic thalli of P. mexicana populationsappeared to prefer specific microhabitat conditions,such low irradiance (average 235 μmol photonsm−2 s−1), boulder substrate and particularly high currentvelocity (average 70 cm s−1). Several characteristicsseem to provide adaptations to high current velocity: (i)abundant spermatangial rings increase fertilizationsuccess, a feature that can be related to the presence ofcarposporophytes in all seasons; (ii) aggregations ofgametophytes and abundant whorled branches alongthe surface of the substrate provide anchorage for newgametophytes. In our field study, the ‘Chantransia’stages of P. mexicana were microscopic and formed asystem of rhizoidal filaments, or a cluster of cells,which produced new gametophytes (clonal reproduc-tion), similar to what has been observed in severalBatrachospermales species in culture (Necchi &Carmona 2002); (iii) germination of carpospores in thethalli lumen gives the gametophytes the possibility ofdeveloping protected by the cortex of their parentalthalli.
The occurrence of P. mexicana in this study corre-sponded with associated species of Sirodotia suecicaand Batrachospermum gelatinosum, a group of temper-ate species (Kwandrans et al. 2002). The presence ofthe epiphytic ‘Chantransia’ stage and gametophytes ofthese species on the Paralemanea surface suggestedthat the thallus serves as a refuge from the high currentvelocity. Similarly, P. mexicana was frequently colo-nized by Simuliidae, Chironomidae and Hydroptilidae;this has also been observed in previous studies in NorthAmerica (Sheath et al. 1996a). However, in the presentstudy, we found no evidence of damage to the gameto-phyte; these insect larvae appear to use the red algae asrefuge from fast current velocities during its final larvaestage (Sheath & Hambrook 1990).
Efficient reproductive strategies of P. mexicanaobserved in this study can be interpreted as adaptationsfor successful colonization in streams; however, thisadaptation may not have been common in the studyregion because it is restricted to an isolated geographicregion with particular microhabitat conditions. This factsuggests that special morphological and reproductiveadaptations promote successful competition with otherspecies in similar hydrological systems in mountainstreams.
ACKNOWLEDGMENTS
We thank S. Castillo (ICMyL-UNAM) for nutrient analy-ses, A. Aguayo Ríos, N.E. Ceniceros B., and O. Cruz R.(IGeof-UNAM) for major ion analyses, M. Cartagena andJ. Ramírez-Lynn for fieldwork assistance, ProfessorNarcís Prat for macroinvertebrate determination andScienceDocs, Inc. for English review. JCJ receivedfinancial support from Research Grant PAPIIT-UNAM(IN209107) and CONACyT (52386).
REFERENCES
APHA, AWWA and WPCF. 1980. Métodos normalizados para elanálisis de aguas potables y residuales. Ediciones Días deSantos, Madrid.
Bojorge, M., Carmona, J., Cartajena, M. and Beltrán, Y. 2010.Temporal and spatial distribution of macroalgal commu-nities of mountain streams in Valle de Bravo Basin, centralMéxico. Hydrobiologia 641: 159–69.
Bowman, M. F., Chambers, P. A. and Schindler, D. W. 2005.Epilithic algal abundance in relation to anthropogenicchanges in phosphorus bioavailability and limitation inmountain rivers. Can. J. Fish. Aquat. Sci. 62: 174–84.
Carmona, J. and Necchi, O. Jr. 2002. Taxonomy and distri-bution of Paralemanea (Lemaneaceae, Rhodophyta) inCentral Mexico. Cryptogam., Algol. 23: 39–49.
Carmona, J., Montejano, G. and Cantoral, E. 2004. The dis-tribution of Rhodophyta in streams of Central Mexico.Arch. Hydrobiol. Suppl. Algol. Stud. 114: 39–52.
Carmona, J., Montejano, G. and Necchi, O. Jr. 2006. Ecologyand morphological characterization of gametophyte and‘Chantransia’ stages of Sirodotia huillensis (Batrachos-permales, Rhodophyta) from a stream in central Mexico.Phycol. Res. 54: 108–15.
Carmona, J., Bojorge, M., Beltrán, Y. and Ramírez, R. 2009.Phenology of Sirodotia suecica (Batrachospermaceae,Rhodophyta) in a high altitude stream in central Mexico.Phycol. Res. 57: 118–26.
Carmona, J., Perona, E., Sánchez-Díaz, E. and Loza, V.2011. Morphological and ecological characterization ofBatrachospermales (Rhodophyta) in the Jarama Basin(Iberian Peninsula). Limnetica 30: 117–28.
© 2013 Japanese Society of Phycology
92 J. C. Jiménez et al.

Dodds, W. K. 2003. Misuse of inorganic N and soluble reac-tive P concentrations to indicate nutrient status of surfaceWaters. J. North Am. Benthol. Soc. 22: 171–81.
Entwisle, T. J., Vis, M. L., Chiasson, W. B., Necchi, O. Jrand Sherwood, A. R. 2009. Systematics of theBatrachospermales (Rhodophyta) – A synthesis. J. Phycol.45: 704–15.
Filkin, N. R. and Vis, M. L. 2004. Phenology of Paralemaneaannulata (Lemaneaceae, Rhodophyta) in a Ohio woodlandstream. Hydrobiologia 518: 159–68.
Gotelli, N. J. and Ellison, A. M. 2004. A Primer of EcologicalStatistics. Sinauer Associates, Inc, Sunderland, MA.
Greenberg, A. E., Trussell, R. R. and Clesceri, L. S. 1985.Standard Methods for the Examination of Water and WasteWater, 16th edn. American Public Health Association(APHA), Washington, DC.
Kapraun, D. F., Braly, K. S. and Freshwater, D. W. 2007.Nuclear DNA content variation in the freshwater red algalorder Batrachospermales and Thoreales (Florideophyceae,Nemaliophycidae). Phycologia 46: 54–62.
Kucera, P. and Marvan, P. 2004. Taxonomy and distribution ofLemanea and Paralemanea (Lemaneaceae, Rhodophyta) inthe Czech Republic. Preslia 76: 163–74.
Kwandrans, J., Eloranta, P. and Bengtsson, R. 2002. Sötvat-tensensrödalger Sverige – en översikt och ett nyfynd.Svensk Bot. Tidsk. 96: 274–80.
Levesque, R. 2006. SPSS Programming and Data Manege-ment, A Guide for SPSS and SAS Users, 3rd edn. RaynaldLevesque and SPSS Inc, Chicago, IL.
Margalef, R. 1983. Ecología. Omega, Barcelona.Merrit, R. and Cummins, K. 1996. An Introduction to the
Aquatic Insects of North America, 3rd edn. Kendall/Hunt,Dubuque, IA.
Necchi, O. Jr and Carmona, J. 2002. Somatic meiosis anddevelopment of the gametophyte in Batrachospermalessensu lato. Phycologia 41: 340–7.
Necchi, O. Jr and Zucchi, M. R. 1995. Record of Paralemanea(Lemaneaceae, Rhodophyta) in South America. Arch.Hydrobiol. Suppl. Algol. Stud. 78: 33–8.
Necchi, O. Jr, Branco, L. H. Z. and Branco, C. C. Z. 1995.Comparison of three techniques for estimating periphytonabundance in bedrock streams. Arch. Hydrobiol. 134:393–402.
Sheath, R. G. 1984. The biology of freshwater red algae.In Round, F. E. and Chapman, D. J. (Eds) Progressin Phycological Research, Vol. 3. Biopress, Bristol,pp. 89–157.
Sheath, R. G. and Hambrook, J. A. 1990. Freshwater ecology.In Cole, K. M. and Sheath, R. G. (Eds) Biology of the RedAlgae. Cambridge University Press, Cambridge, pp. 423–53.
Sheath, R. G., Müller, K. M., Vis, M. L. and Entwisle, T. J.1996a. A re-examination of the morphology, ultrastructureand classification of genera in the Lemaneaceae(Batrachospermales, Rhodophyta). Phycol. Res. 44: 233–46.
Sheath, R. G., Müller, K. M., Colbo, M. H. and Cole,K. M. 1996b. Incorporation of freshwater Rhodophytainto the cases of chironomid larvae (Chironomidae,Diptera) from North America. J. Phycol. 32: 949–52.
Thirb, H. H. and Benson-Evans, K. 1984. The effect of tem-perature on the growth of Lemanea thalli and carposporegermination. Arch. Hydrobiol. 103: 341–6.
Thirb, H. H. and Benson-Evans, K. 1985. The effect of watertemperature, current velocity and suspended solids on thedistribution, growth and seasonality of Lemanea fluviatilis(C. Ag.) Rhodophyta, in the River Usk and other SouthWales rivers. Hydrobiologia 127: 63–78.
Vis, M. L., and Sheath, R. G. 1992. Systematics of thefreshwater red algal family Lemaneaceae in North America.Phycologia 31: 164–79.
Vis, M. L., Saunders, G. W., Sheath, R. G., Dunse, K. andEntwisle, T. J. 1998. Phylogeny of the Batrachospermales(Rhodophyta) Inferred from rbcL and 18S ribosomal DNAgenes sequences. J. Phycol. 34: 341–50.
Wiggins, G. 1996. Larvae of the North America CaddisflyGenera (Trichoptera). University of Toronto Press, Toronto,ON.
© 2013 Japanese Society of Phycology
93Phenology of Paralemanea mexicana