pharmacological prevention of acute lead poisoning in paramecium
TRANSCRIPT

ELSEVIER
Pharmacological
Toxicology 108 (1996) 165-173
prevention of acute lead poisoning in Paramecium
Juan Bernal*, Sergio Ruvalcaba
Deporromenta de Medicino. Cenrro BiomPdico, Universidod Autdnomo de Aguoscolientq C. P. 20100, Aguascolienres. Ags., MPxico
Received IS November 1994; accepted 31 October 1995
Abstract
To understand how lead (Pb”) and other heavy metals and chelating agents affect living cells, behavioral experi- ments in the marine ciliate Paramecium calkinsi were carried out. The duration of Backward Swimming Behavior (BSB) of Paramecium was partially reduced when celis were exposed to 100 FM of Ni2+, Cd*’ and Co2+. In contrast, Pb*’ increased Paramecium BSB in a dose-dependent manner. Thus, I, 10, 20, 50 and 100 PM of Pb” increased the dura- tion of BSB by 20.4,83.9, 143.2, 163.2 and 185.2%, respectively. The naphthalenesulphonamide W-7, a calcium channel blocker in lower organisms, abolished the increase of Paramecium BSB initially produced by PbZ’. Paramecium,
poisoned with 10 PM of Pb”, were also treated with putative Pb2+ chelating agents, such as meso-2-3-
dimercaptosuccinic acid (DMSA), Ca-Na,-EDTA and ascorbic acid. These compounds inhibited the increase of the duration of BSB initially produced by Pb2+ in a dose-dependent manner. The potency of these antidotes in blocking the effects of Pb2+ was as follows: DMSA > Ca-Na,-EDTA > ascorbic acid. These results provide evidence for a membrane-based mechanism of lead poisoning and support the use of DMSA as a lead antidote,
Keyworcis: Lead toxicity; Calcium channels; Paramecium; Pb”; DMSA; Ca-Na,-EDTA; meso-2,3-
Dimercaptosuccinic acid; Ascorbic acid
1. Introduction
Lead and other heavy metals are ubiquitous ele- ments that dramatically modify the environment (Nriagu, 1990). Many heavy metals do not have functional properties on living organisms and their toxic effects can be observed at different levels from protozoa to mammalia. Thus, it has been
* Corresponding author, Tel.: 52-49-123345 ext. 342; Fax: 52-49-143222 or 145591.
proposed that heavy metals such as Ni’+, Co*+, and Cd2+ may interfere with calcium permeability through calcium channels (Eckert and Brehm, 1979; Hagiwara and Byerly, 1981; Tsien et al., 1987; Byerly and Hagiwara, 1988; Bean 1989; Hess, 1990; Hille, 1992; Bernal and Ehrlich, 1993). In fact, the selectivity of divalent cations to block calcium channels has been considered as a criteria for calcium channel classification. Thereby, the L- type calcium channels are more sensitive to Cd2+, whereas the T-type calcium channels are more sen-
0300-483)(/96/$15.00 0 1996 Elsevier Science Ireland Ltd. All rights reserved SSDI 0300-483X(95)03272-H

166 J. Bemal, S. Ruvalcaba / Toxicology 108 (19%) 165-173
sitive to Ni2+ (Nowycky et al, 1985; Fox et al., 1987; Tsien et al., 1987, 1988; Tang et al., 1988; Bean, 1989; Chow, 1991; Hille, 1992).
There are controversial results about the effects of Pb2+ on ionic membrane permeability. In some cases. Pb2+ blocks the calcium membrane perme- ability through voltage-dependent calcium chan- nels (Audesirk and Audesirk, 1991; Busselberg et al, 1991; Evans et al., 1991; Reuveny and Narahashi, 1991). In other cases, Pb2+ permeates voltage-dependent calcium channels (Tomsig and Suszkiw, 1991) or enhances current flow through voltage-dependent calcium channels (Audesirk and Audesirk, 1991). The effect of Pb2+ on the calcium permeability of lower organisms such as protozoa is unknown. Because Paramecium is a protozoan that has only one class of voltage- dependent calcium channel characterized as a ‘T- type’-like calcium channel (Eckert and Brehm, 1979; Ehrlich et al., 1988; Bernal and Ehrlich, 1993), in this work we studied whether Pb2+ modifies the ionic calcium permeability of Paramecium.
Lead poisoning in humans is a public health problem in several countries. The group of people most affected by this problem is children in which the major impact produced by lead is in the brain (Aposhian and Aposhian, 1990; Chisolm, 1990; Goldstein, 1990; Needleman and Gatsonic, 1990; Bressler and Goldstein, 199 1; Needleman and Bell- inger, 1991; Beck, 1992; Rosen, 1992; Shannon and Graef, 1992). Much effort has been focused on improving the treatment of lead intoxications. The appropriate drugs and the correct dose for humans are still controversial. Drugs such as Ca-Naz- EDTA (Flora et al., 1986; Cory-Slechta et al., 1987; Llobet et al., 1990; Rosen et al., 1991; Chisolm, 1992), ascorbic acid (Dalley et al., 1990; Llobet et al., 1990) and meso-2,3- dimercaptosuccinic acid (DMSA) (Aposhian, 1983; Graziano, et al., 1985; Cory-Slechta, 1988; Fournier et al., 1988; Aposhian and Aposhian, 1990; Aposhian et al., 1989, 1992; Chisolm, 1990, 1992) have been used for treating lead poisoning. Although DMSA and Ca-Na,-EDTA, as well as ascorbic acid, are compounds that may eliminate lead from the body in intact animals and humans, these drugs have not been tested before as to the
protective effect on living cells, such as protozoa, from lead poisoning.
In this paper, behavioral experiments in the pro- tozoan Paramecium calkinsi were done to address the hypothesis that acute lead poisoning indeed acts on the cellular membrane of the cilia modulating the calcium permeability of Paramecium. In addition, we report that among several compounds tested, DMSA prevented Pb” poisoning in Paramecium most effectively. Prelimi- nary results of this work have already been published in abstract form (Ruvalcaba et al., 1992).
2. Metbods
2.1. Biological preparation In the present work, the marine ciliate
Paramecium calkinsi was used. Paramecium was maintained in 25% artificial sea water culture media and fed with Enterobacter aurogenes 48 h before being used for the experiments (see Bernal et al., 1991).
2.2. Chemicals Calcium disodium ethylenediaminetetraacetic
acid (Ca-Na2-EDTA), ascorbic acid, meso-2,3- dimercaptosuccinic acid (DMSA), the naphthal- enesulphonamide W-7 and dimethylsulfoxide (DMSO) as well as all salts and buffers used for preparing solutions were obtained from Sigma Chemical Co. Lead acetate (99.99%) and nitric acid were obtained from Aldrich. W-7 and DMSA were dissolved in 1% DMSO (in didistillated water) for preparing stock solutions of 10 mM. Lead acetate was dissolved in a 0.1% nitric acid solution (in didistillated water). The pH of all solu- tions containing heavy metals, chelating agents or W-7 was maintained at 7.2.
2.3. Behavioral experiments Paramecium is the most primitive organism that
has voltage-dependent calcium channels (Ehrlich et al., 1988; Ehrlich and Forte, 1988). These chan- nels are predominantly located at the ciliary cellular membrane (Dunlap, 1977; Eckert and Brehm, 1979; Machemer and Ogura, 1979).
The activation of calcium channels in
J. Bemal, S. Ruvalcaba / Toxicology 108 (1996) 165-173 167
Paramecium modulates the backward swimming behavior (BSB) of the ciliate (Eckert and Brehm, 1979; Hennessey and Kung, 1984; Ehrlich et al., 1988; Bemal et al., 1991). Because in its normal media Paramecium swims backward for about 1 to 3 s, in the experiments described below the dura- tion of BSB of Paramecium was magnified ex- perimentally by using high potassium solution. It was done by transferring Paramecium from a chamber containing normal ‘resting’ solution (NaCl, 125 mM; CaC12, 1 mM; 3-[N- Morpholinolpropane sulfonic acid (MOPS), 5 mM; pH 7.2) to a chamber containing high potas- sium ‘testing’ solution (NaCl, 62.5 mM; KCl, 62.5 mM; CaCl,, 1 mM; MOPS, 5 mM; pH 7.2). After Paramecium is placed in the high potassium ‘test- ing’ solution, the cellular membrane of the ciliate is depolarized, and Paramecium starts swimming backward immediately. In approximately 150 s, Paramecium become adapted, recover and start swimming forward again. Cells were exposed and incubated to several compounds in normal ‘resting’ solution for 5 min. After this period of in- cubation, cells were transferred to the high potas- sium ‘testing’ solution. Similar concentrations of drugs were maintained in both chambers contain- ing normal ‘resting’ and high potassium ‘testing’ solutions.
3. Results
3.1. DivaIent cations modulate backward swimming in Paramecium
It was found that Ni*+, Cd2+ and Co*+ at 100 PM, reduced the duration of BSB by 25.9, 18.4 and 8.8%, respectively (Table 1). The relative potency
Table 1 Effect of divalent cations (100 pm) on the duration of Paramecium backward swimming behavior (seconds f S.D.; n = number of cells)
Ni2+ Cd2+ co2+ Pb2+
Control conditions Experimental conditions
158 f 15 (n = 36) 117.2 zt I7 (n = 28) I52 f 9.1 (n = 18) 124 zt 9.5 (n = 18) I61 ziz IO (n=22) I46 * 9.3 (n = 22) I56 zt 14.7 (n = 30) 447 zt 13.3 (n = 28)
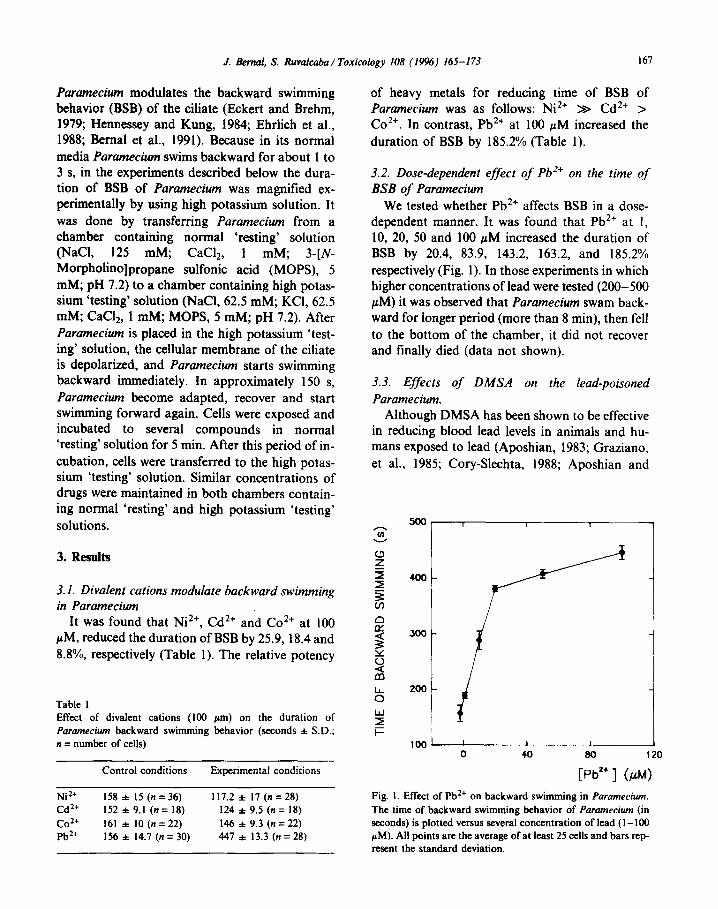
of heavy metals for reducing time of BSB of Paramecium was as follows: Ni*+ >> Cd*+ > Co*+. In contrast, Pb*+ at 100 PM increased the duration of BSB by 185.2% (Table 1).
3.2. Dose-dependent effect of Pb’+ on the time of BSB of Paramecium
We tested whether Pb*+ affects BSB in a dose- dependent manner. It was found that Pb*+ at I, 10, 20, 50 and 100 PM increased the duration of BSB by 20.4, 83.9, 143.2, 163.2, and 185.2% respectively (Fig. 1). In those experiments in which higher concentrations of lead were tested (200-500 PM) it was observed that Paramecium swam back- ward for longer period (more than 8 min), then fell to the bottom of the chamber, it did not recover and finally died (data not shown).
3.3. Effects of DMSA on the lead-poisoned Paramecium.
Although DMSA has been shown to be effective in reducing blood lead levels in animals and hu- mans exposed to lead (Aposhian, 1983; Graziano, et al., 1985; Cory-Slechta, 1988; Aposhian and
Y------7
Fig. I. Effect of Pb2+ on backward swimming in Paramecium.
The time of backward swimming behavior of Paramecium (in seconds) is plotted versus several concentration of lead (I - 100 PM). All points are the average of at least 25 cells and bars rep- resent the standard deviation.

168
-350 03
g 300
r
? 250
L? 200
s 150 is
3 100
k
w s0 I i= 0
J. Bernal, S. Ruvalcaba / Toxicology 108 (19%) 165-173
A
1 1 0
i
]
e Pb2+ 1OpM-i
-r
71 a _I
! L
I - 10 0 50 0 1 10 20 50
[DMSAI (AN [EDT4 OLM)
C
I I 0 lot I
k Pb2+
[ASCAC.] (@I)
Fig. 2. Phannaco logical prevention of acute lead poisoning on the duration of backward swimming behavior of Paramecium. The time of backward swimming behavior of Paramecium (in seconds) is plotted in control conditions (black columns) and when cells were treated with Pbz+ (10 PM) alone and with several concentrations of DMSA (A), Ca-Na,-EDTA (B) and ascorbic acid (C). Col- umns are the average of at least 25 cells and bars represent the standard deviation.
Aposhian, 1990), the capability of DMSA to pro- tions was 144.1 f 9.1 s (mean and SD); n = 15 tect cells from lead toxicity at cellular level is still (Fig. 2A, first column). In the presence of DMSA relatively unknown. Therefore, we decided to test at 50 PM, the duration of BSB was 156.1 f 8.1 s; DMSA on Paramecium in normal conditions and n = 21 (Fig. 2A, second column). when cells were exposed to lead. In these experi- When Pb*+ at 10 PM was administered alone, it ments the duration of the BSB in control condi- increased the duration of BSB by about 98% (Fig.

J. Bend, S. Ruvalcaba / Toxicology IO8 (19%) 165-173 169
2A, third column). In the presence of PbL+ (10 PM), DMSA at 1, 10, 20 and 50 pM, reduced Pb2+ effects on BSB by 6.9, 23.3, 49.8 and 98.7% respectively (Fig. 2A). According to these data, the ED% found for DMSA to inhibit the effects in- itially produced by Pb2+ on the duration of BSB of Paramecium was 20.5 PM.
3.4. Effect of Ca-NarEDTA on the lead-poisoned Paramecium
Ca-Na2-EDTA has been used as the most com- mon compound to treat lead intoxication in hu- mans and other mammals (Cory-Slechta et al., 1987; Aposhian and Aposhian, 1990; Llobet et al., 1990; Chisolm, 1990, 1992; Rosen et al., 1991). We decided to evaluate if this drug may protect Paramecium from acute lead poisoning. In these series of experiments the duration of BSB in con- trol conditions was 141.2 f 6.1 s; n = 15 cells. In the presence of 10 and 100 NM of Ca-Na,-EDTA alone, the duration of BSB was 137.2 f 6.9 s; n = 16 cells and 138.6 f 7.3 s; n = 18 cells, respec- tively (Fig. 2B). When cells were treated with Pb2+ at 10 PM, it increased the duration of BSB up to 281.3 & 6.7 s; n = 15 cells. Pb2+ at 10 PM tested along with Ca-Na2-EDTA at 10 and 100 PM, reduced the time of BSB, initially increased by lead, to 180.4 f 4.6 s; n = 10 cells and 143.5 f 5.6 s; n = 20 cells, respectively. The per- cent of inhibition of Ca-Na2-EDTA at 10 and 100 PM to inhibit lead effects on BSB was 72.1 and 98.3%, respectively (Fig. 2-B).
3.5. Effect of ascorbic acid on the lead-poisoned Paramecium
Some reports suggest that vitamins may be useful for lead poisoning treatment (Flora et al, 1986; Dalley et al., 1990; Llobet et al., 1990). We studied whether ascorbic acid protects Paramecium from lead poisoning. In control experiments, the duration of BSB was 164.1 f 4.1 s; n = 21 cells (black column, Fig. 2C). In the presence of ascorbic acid alone (100 FM), the duration of BSB was 166.3 f 5.9 s; n = 25 cells (second column, Fig. 2C). When cells were expos- ed to lead at 10 PM, it increased the duration of BSB up to 326.1 * 7.6 s; n = 20 cells, producing an increment of 98.7% on the duration of BSB
Fig. 3. Effect of W-7 on lead-poisoned Paramecium. The time of backward swimming behavior of Paramecium (in seconds) is plotted in control conditions and when cells were treated with W-7 and/or Pb’+. Columns are the average of at least 15 cells and bars represent the standard deviation.
(fourth column, Fig. 2C). When cells were treated with lead (at 10 PM) along with ascorbic acid at the concentrations of 10, 50, 100 and 200 PM, it was found that ascorbic acid reduced lead’s effect on the duration of BSB by 8.6, 21.6, 37.0 and 46.9%, respectively, (Fig. 2C). The ECSo of ascor- bic acid to prevent lead poisoning in Paramecium was found to be >200 PM.
3.6. Lead effect on the duration of Paramecium backward swimming behavior is modulated by W-7
The naphthalenesulfonamide W-7 (Hidaka et al., 1979), has shown to be effective in blocking the calcium channel in Paramecium (Hennessey and Kung, 1984; Ehrlich et al., 1988; Bernal and Ehrlich, 1993; Barry and Bernal, 1993). To test the hypothesis that the effect of lead on BSB of Paramecium actually affects calcium permeability of the ciliate, the action of W-7 on the BSB of Paramecium in the presence or absence of lead (10 PM) was studied. In these experiments, the dura- tion of BSB of Paramecium in control conditions was 158.35 f 6.1 s; n = 22 cells. When cells were treated with W-7 at 100 PM, the time of BSB was reduced to 17.5 i 3.0 s; n = 20 cells (Fig. 3, sec- ond column). When Pb2+ at 10 JLM was tested

170 J. Bemal. S. Ruvalcaba / Toxicology 108 (1996) 165-173
alone, the duration of BSB was increased to 285.1 f 7.4 s; n = 21 cells (Fig. 3, third column). The effect of W-7 on the lead- poisoned Paramecium was tested as follows: cells were ex- posed to Pb2+ at 10 FM, then W-7 at 100 PM was added 3 min after Paramecium started swimming backward. Immediately after W-7 was added, Paramecium began swimming forward again (Fig. 3, fourth column).
4. Diion
In this paper we have found that acute lead poi- soning in Paramecium increased its duration of BSB, whereas other heavy metals decreased the time of BSB. Also we found that the effect initially produced by lead on Paramecium BSB was prevented when cells were treated with putative heavy metal chelating agents, such as DMSA, Ca- Na2-EDTA or ascorbic acid.
In the experiments we report here, the potency of divalent cations in reducing the duration of BSB was as follows: Ni2+ >> Cd2+ > Co2+. These divalent cations have shown different specificity in blocking the calcium influx into the cell through voltage-dependent calcium channels in several preparations (Hagiwara and Byerly, 1981; Nowycky et al, 1985; Tsien et al., 1987, 1988; Bean, 1989; Hess, 1990; Bernal and Ehrlich, 1993). It can be expected that Ni2+ is the most effective in blocking BSB of Paramecium for several reasons: first, because different calcium channel types have been found in several preparations (Fedulova et al, 1985; Armstrong and Matteson, 1985; Nowycky et al, 1985), it has been suggested that Ni2+ is more specific as a ‘T type’ calcium channel blocker (Tsien et al., 1988; Tang et al., 1988; Bean, 1989; Hille, 1992). Second, the type of calcium channel that has been proposed to be pres- ent in Paramecium is the ‘T-type’ because this channel has small conductance, is inactivating and is not sensitive to dihydropyridines (Ehrlich et a1.,1988; Bemal and Ehrlich, 1993). Third, assum- ing that BSB is an indirect estimation of the voltage-dependent calcium channel in Paramecium (Eckert and Brehm, 1979; Haga et al., 1982; Hen- nessey and Kung, 1984; Bemal et al., 1991), the ac- tion of an inorganic calcium channel, blocker such
as Ni2+, should inhibit the duration of BSB. Similar results were obtained by Eckert (1972).
In contrast with the findings described above, an interesting result of the behavioral experiments with Paramecium is that the acute exposure of Paramecium to Pb2+ increased in a dose- dependent manner the duration of BSB (see Fig. 1). Different possibilities could explain these re- sults: First, because BSB in Paramecium is an indi- rect estimation of calcium channel activation (Hennessey and Kung, 1984; Ehrlich et al, 1988; Bemal et al., 1991), there is a chance that Pb2+ modifies the kinetics of calcium channels and in- duces Paramecium backwards swimming for a longer time. It can be explained by two mechanisms: (1) increasing the time activation of the channel and (2) by blocking the inactivation process of the calcium channel (Brehm et al., 1980; Eckert and Chad, 1984: Chad and Eckert, 1986). In both situations the channel will be open for a long period of time, promoting the massive influx of calcium, increasing the duration of BSB and eventually killing the cell, depending upon lead concentration used. Second, it is possible that Pb2+ may permeate the cellular membrane prob- ably through calcium channels, as suggested in other cells (Tomsig and Suszkiw, 1991), Thus, Pb2+ may activate a calcium-calmodulin- dependent process (Cheung, 1984) or a protein kainase C-dependent process (Markovak and Goldstein, 1988) that could be associated with an increase of the duration of Paramecium BSB. This is plausible because it has been proposed that Pb2+ activates calmodulin with higher affinity than calcium by itself (Cheung, 1984) and that pro- tein kinase C can be activated by Pb2+ at picomolar concentrations (Markovac and Goldstein, 1988). Pb2+ may also increases the cur- rent flow through voltage-dependent calcium channels in Paramecium as found in neuroblastoma cells (Audesirk and Audesirk, 1991). Thus, membrane mechanisms involved in BSB may be modulated by a calmodulin- dependent or protein kinase C-dependent process, triggered either by Ca2+ or Pb2+, that increases the time of Paramecium BSB. Finally, the last pos- sible explanation is that Pb2+ may interact with the binding sites of the calcium channel or with the

J. Bema~. S. Ruvalcaba/ Toxicology 108 (1996) 165-173 171
sulphydryl groups (SH-) (Chow, 1991; Rosenberg and Xio-Hua 1991) producing conformational changes of the channel and promoting the massive influx of calcium into the cell. To answer these possibilities, further electrophysiological experi- ments such as two microelectrode voltage clamp and single calcium channel recording have to be done. These experiments are in progress in our lab- oratory. Despite the possible mechanism by which Pb*+ promotes the increase of the duration of BSB in Paramecium, it was observed that the cal- cium permeability of Paramecium was modified by Pb*+ because a calcium channel blocker such as a W-7 (Hennessey and Kung, 1984; Ehrlich et al., 1988; Barry and Bemal, 1993) inhibited Pb*+ ef- fects on BSB of Paramecium (see Fig. 3).
Three different chelating agents were tested on lead-poisoned Paramecium. These three com- pounds prevented, in a dose-dependent manner, the effects initially produced by lead with different potencies: DMSA >> Ca-Na2-EDTA > ascorbic acid (see Fig. 2). An interesting finding of these re- sults is the fact that compounds suggested as lead antidotes in humans are effective in giving protec- tion from lead poisoning at unicellular level; DMSA being the most effective. There is experi- mental evidence that supports the hypothesis that DMSA is the most promising chelating agent available for lead poisoning treatment in humans (Cory-Slechta, 1988; Aposhian and Aposhian, 1990; Chisolm, 1990, 1992; Aposhian et al., 1992). DMSA removes Pb*+ from the body without clinical side-effects. It can be administrated orally without reducing plasma concentration of Zn*+, Mn*+ and Fe*+. A major advantage of meso-2,3- DMSA is that it decreases brain Pb*+. Although the pharmacokinetics and biotransformation of DMSA are well known when this compound is
given orally to humans (Aposhian et al., 1989; Aposhian and Aposhian, 1990), the efficiency of this compound to treat acute lead poisoning ‘in vitro’ at unicellular level has not been evaluated. In our results, DMSA by itself did not modify the time of BSB in Paramecium (see Fig. 2A). In con- trast, when DMSA at different concentrations was tested along with Pb*+ (10 FM), acute lead poi- soning was prevented by DMSA in a dose- dependent manner (see Fig. 2A). Several explana-
tions can be advanced to explain this phenome- non: (1) if we assume that DMSA chelates two molecules of Pb*+ with one of the sulfur and one of the oxygen atoms of DMSA (Aposhian and Aposhian, 1990), this compound protects living cells from lead poisoning by chelating the total free Pb*+ added to the extracellular media in combina- tion with DMSA (see Fig. 2A); (2) because DMSA shows highly charged carboxyl groups, it cannot enter the cells. Therefore, DMSA is distributed in the extracellular media and it has to act at this level to protect living cells from lead poisoning; and (3) it has been proposed that a thiol-disulfide exchange or a direct reaction between DMSA and L-cysteine takes place and is necessary for DMSA activation. Thus, there is a chance that the DMSA, present in the extracellular media, interacts with cysteine groups of the cellular membrane, and pro- duces a chelate potentiation effect of DMSA to avoid Pb*+ effects on the cellular membrane. This last possibility could be possible if we assume that the interaction between DMSA and Pb*+ occurs at the cellular membrane.
In conclusion, in this paper we have presented experimental results which suggest that lead modifies the calcium ionic permeability of Paramecium and that DMSA can be used as a lead antidote at unicellular level.
Acknowledgments
The authors are very thankful to Dr. Arnulfo
Albores, Jose Carlos Escalera and Barbara DiGregorio for criticism and to Teresa Villalobos for secretarial assistance.
This work was supported in part by the grant 005 l-N9106 from CONACYT, Mixico and by the grant PIBB91-5 from Universidad Aut6noma de Aguascalientes, Mtxico.
References
Aposhian, H.V. (1983) DMSA and DMSA-water soluble anti- dotes for heavy metal poisoning. Annu. Rev. Pharmacol. Toxicol. 23, 193-215.
Aposhian, H.V. and Aposhian, M.M. (1990) Meso-2,3- dimercaptosuccinic acid: chemical, pharmacological and toxicological properties of an orally effective metal chelating agent.. Annu. Rev. Pharmacol. Toxicol. 30. 279-306.

172 J. Bema/, S. Ruvalcaba/ Toxicology 108 (19%) 165-173
Aposhian, H.V., Maiorino, R.M., Dart, R.C. and Perry, D.F. (1989) Urinary excretion of meso-2,3-dimercaptosuccinic acid in human subjects. Clin. Pharmacol. Ther. 45. 520-526.
Aposhian, H.V., Maiorino, R.M., Rivera, M., Bruce, DC., Dart, R.C. et al. (1992) Human studies with chelating agents, DMPS and DMSA. Clin. Toxicol. 3q4), 505-528.
Armstrong, C.M. and Matteson, D.R. (1985) Two distinct populations of calcium channels in a clonal line of pituitary cells. Science 2, 65-67.
Audesirk, G. and Audesirk, T. (1991) Effects of inorganic lead on voltage-sensitive calcium channels in N 1 E- 115 neuroblastoma cell. Neurotoxicology 12, 519-528.
Barry, S. and Bemal. J. (1993) Antimalarial drugs modulate calcium currents in Paramecium. J. Gen. Comp. Physiol. 172, 457-466.
Bean, B.P. (1989) Classes of calcium channels in vertebrate cells. Annu. Rev. Physiol. 51, 367-384.
Beck, B. (1992) An update on exposure and effects of lead. Fundam. Appl. Toxicol. 18, I- 16.
Bemal, J. and Ehrlich, B.E. (1993) Guanine nucleotides modulate calcium currents in a marine Paramecium. J. Exp. Biol. 176, 117-133.
Bemal, J., Kelsey, A.M. and Ehrlich, B.E. (1991) GTP- Gamma-S increases the duration of backward swimming behavior and the calcium action potential in marine Paramecium, J. Exp. Biol. 155, 505-518.
Brehm, P., Eckert, R. and Tillotson, D. (1980) Calcium- mediated inactivation of calcium current in Paramecium. J. Physiol. Lond. 306, 193-203.
Bressler, J.P. and Goldstein, G.W. (1991) Mechanisms of lead neurotoxicity. B&hem. Phamtacol. 41(4), 479-484.
Busselberg, D., Evans, M. L., Rahmann, H. and Carpenter, D.O. (1991) Lead and zinc block a voltage-activated chan- nel of Aplysia neurons. J. Neurophysiol. 65(4), 786-795.
Byerly, L. and Hagiwara, S. (1988) Calcium channel diversity. In: A.D. Grinnel, D. Armstrong, M.B. Jackson (Eds), Cal- cium and Ion Channel Modulation, Plenum Press, New York, pp. 3-18.
Chad, E.J. and Eckert, R. (1986) An enzymatic mechanism for calcium current inactivation in dialyzed He/ix neurones. J. Physiol. 378, 31-51.
Cheung, W.Y. (1984) Calmodulin: its potential role in cell pro- liferation and heavy metal toxicity. Fed. Proc. 43, 2995-2999.
Chisolm, J.J. (1990) Evaluation of the potential role of chela- tion therapy in treatment of low to moderate lead ex- posures. Environ. Health Perspect. 89, 67-74.
Chisolm, J.J. (1992) BAL, EDTA, DMSA and DMPS in the treatment of lead poisoning in children. Clin. Toxicol. 30(4), 493-504.
Chow, R.H. (1991) Cadmium block of squid calcium currents. Macroscopic data and a kinetic model. J. Gen. Physiol. 98, 751-770.
Gory-Slechta, D.A. (1988) Mobilization of lead over the course of DMSA chelation therapy and long-term efficacy. J. Phar- macol. Exp. Ther. 246(l), 84-91.
Gory-Slechta, D. A., Weiss, B. and Cox, C. (1987) Mobilization and redistribution of lead over the course of calcium
disodium ethylenediamine tetracetate chelation therapy. J. Pharmacol. Exp. Ther. 243, 804-813.
Dalley, J.W., Gupta, P.K. and Hung, CT. (1990) A physiological pharmacokinetic model describing the dis- position of lead in the absence and presence of L-ascorbic acid in rats. Toxicol. Lett. 50, 337-348.
Dunlap, K. (1977) Localization of calcium channels in
Paramecium cauahm. J. Physiol., Lond. 271, 119-133. Eckert, R. (1972) Bioelectric control of ciliary activity. Science
176, 473-480. Eckert, R. and Brehm, P. (1979) Ionic mechanisms of excitation
in Paramecium. Annu. Rev. Biophys. Bioeng. 8, 353-383. Eckert, R. and Chad, E.J. (1984) Inactivation of calcium chan-
nels. Prog. Biophys. Mol. Biol. 44. 215-267. Ehrlich, B.E. and Forte, M.A. (1988) Calcium channels incor-
porated into planar lipid bilayers: phenomenology, phar- macology and phylogeny. In: A.D. Grinnell, D. Armstrong and M.B. Jackson (Eds.), Calcium and Ion Channel Modu- lation. Plenum Press, New York, pp. 33-41.
Ehrlich, B.E., Jacobson, A.R., Hinrichsen, R., Sayre, L.M. and Forte, M.A. (1988) Paramecium calcium channels are blocked by a family of calmodulin antagonists. Proc. Natl. Acad. Sci. USA 85, 5718-5722.
Evans, M.L., Busselberg, D. and Carpenter, D.O. (1991) Pb+* blocks calcium currents of cultured dorsal root ganglion cells. Neurosci. Lett. 129, 103-106.
Fedulova, S.A., Kostyuk, P.G. and Veselovsky, N.S. (1985) Two types of calcium channels in the somatic membrane of newborn rat dorsal root ganglion neurons. J. Physiol. 359, 431-446.
Flora, S.J., Singh, S. and Tandom, S.K. (1986) Chelation in metal intoxication. XVIII: Combining effects of thiamine and Ca-Naz-EDTA and versenate on lead toxicity. Life Sci. 38, 67-71.
Fournier, L., Thomas, G., Gamier, R., Buisine, A., Houze, P., Pradier F. and Daily S. (1988) 2,3-Dimercaptosuccinic acid treatment of heavy metal poisoning in humans. Med. Tox- icol. 3, 499-504.
Goldstein, G.W. (1990) Lead poisoning and brain cell function. Environ. Health Perspect. 89, 91-94.
Graziano, J.H., Siris, SE., Lolacono, B.S., Silvergeld, J.S. and Turgeon, L. (1985) 2,3-Dimercaptosuccinic acid as an an- tidote for lead intoxication. Clin. Pharmacol. Ther. 37, 4, 43 l-438.
Haga, N., Forte, M., Siami, Y. and Kung, C. (1982) Microin- jection of cytoplasm as a test of complementation in Paramecium. J. Cell. Biol. 96, 1072-1080.
Hagiwara, S. and Byerly, L. (1981) Calcium channels. Annu. Rev. Neurosci. 4, 69-125.
Hennessey, T. and Kung, C. (1984) An anticalmodulin drug W- 7 inhibits the voltage-dependent calcium current in Paramecium mu&rum. J. Exp. Biol. 110, 169-181.
Hess, P. (1990) Calcium channels in vertebrate cells. Annu. Rev. Neurosci. 13, 337-56.
Hidaka, H., Yamaki, T., Totsuka, T. and Asano, M. (1979) Selective inhibitor of calcium-binding modulator of phosphodiesterase produce vascular relaxation and inhibits actin-myosin alteration. Mol. Pharmacol. 15, 15-59.
Hille, B. (1992) Ionic Channels of Excitable Membranes. Scc-

1. Bemal, S. Ruvalcnba / Toxicology 108 (19%) 165-l 73 173
ond Edition, University of Washington. Sinauer Associates Inc. Publisher, Sunderland, Massachusetts.
Llobet, J.M.. Domingo, J.L., Patemain, J.L. and Corbella, J. (1990) Treatment of acute lead intoxication. A quantitative comparison of a number of chelating agents. Arch. En- viron. Contam. Toxicol. 19, 185-189.
Machemer, H. and Ogura, A. (1979) Ionic conductances of membranes in ciliated and deciliated Paramecium. J. Physiol., Lond. 296, 46-60.
Markovac, J. and Goldstein, G.W. (1988) Picomolar concen- trations of lead stimulate protein kinase C. Nature 334. 71-73.
Needleman, H.L. and Bellinger, D. (1991) The health effects of low lead exposure to lead. Annu. Rev. Health 12, II l-140.
Needleman, H.L. and Gatsonic, CA. (1990) Low level lead ex- posure and the IQ of children: a meta-analysis of modern studies. J. Am. Med. Assoc. 263, 673-678.
Nowycky, M.C.. Fox, A.P. and Tsien, R.W. (1985) Three types of neuronal calcium channels with different calcium agonist sensitivity. Nature 316, 440-443.
Nriagu, J.O. (1990) Global metal pollution. Environment 32(7), 7-32.
Reuveny, E. and Narahashi, T. (1991) Potent blocking action of lead on voltage-activated calcium channels in human neuroblastoma cells SH-SY 5Y. Brain Res. 545, 3 12-3 14.
Rosen, J.F. (1992) Health effects of lead at low exposure levels. AJDC 146, 1278-1281.
Rosen, J.F.. Markowitz, M.E., Kalef-Ezra, J.A. and Slatkin.
D.N. (1991) Sequential measurements of bone lead content by LX-ray fluorescence in Ca-Naz-EDTA-treated lead toxic children. Environ. Health Perspect. 93, 271-277.
Rosenberg, R.L. and Xiao-Hua, Ch. (1991) Characterization and localization of two ion-binding sites within the pore of cardiac L-type calcium channels. J. Gen. Phys~ol. 97, 1207-1226.
Ruvalcaba, S., Yamamoto, L., Acosta, 0. and Bemal, J. (1992) Pharmacological prevention of acute lead poisoning in Pmamerium. Biophys. J. 61(2), Al28-Al59.
Shannon, M.W. and Graef. J.W. (1992) Lead intoxication in in-
fancy. Pediatrics 89( I ), 87-90. Tang, CM., Presser, F. and Morad, M. (1988) Amiloride selec-
tively blocks the low threshold (T) calcium channel. Science 240, 213-215.
Tomsig. J.L. and Suszkiw. J.B. (1991) Permeation of Pb*+ through calcium channels: fura- measurements of voltage- and dihydropyridine-sensitive Pb*+ entry in isolated bovine chromaftin cells. Biochem. Biophys. Acta 1069, 197-200.
Tsien, R.W., Hess, P., McCleskey, E.W. and Rosenberg, R.L. (1987) Calcium channels: mechanisms of selectivity, permeation and block. Annu. Rev. Biophys., Biophys. Chem. 16, 265-290.
Tsien, R-W., Lipscombe, D., Madison, D.V., Bley, K.R. and Fox, A.P. (1988) Multiple types of neuronal calcium chan- nels and their selective modulation. Trends Neurosci. 1 I, 431-438.