a subset of cholinergic mushroom body neurons...
TRANSCRIPT

SLEEP, Vol. 36, No. 12, 2013 1809 Cholinergic MB Neurons Regulate Sleep—Yi et al
INTRODUCTIONPeople suffer from various sleep disorders such as insomnia
and hypersomnia,1 leading to a clear need to identify the neurophysiology and neurochemistry of sleep and wakeful-ness. Drosophila has emerged as a new animal model for sleep research, because its rest behavior shares cardinal features with mammalian sleep characteristics such as prolonged revers-ible immobility, increased arousal thresholds,2,3 altered brain electrical activity,4,5� ���� �������� � ����� �6 The neuro- ������� ���� ��� ������ ��� ��� ��� ��� ������ ��� ��functionally conserved.1,7 As in mammals, dopamine (DA) and octopamine (OA, the insect equivalent of norepinephrine) exert wake-promoting function, whereas serotonin (5-HT) and ������������� � � ��� ������� ������� ������� ��� ��� �����������������������1,8-13 The convenient genetic tools in fruit ���������������!���������������������� �� ����������with dopamine, octopamine, and 5-HT systems in sleep regula-tion, providing further insights into the understanding of sleep neurophysiology.14-16 Nevertheless, the function of the cholin-��� �������������������������������������������
"����������������#��$��������������� ��������������the brain, has been highlighted as a sleep control center in fruit
CHOLINERGIC MB NEURONS AND REGULATION OF SLEEP IN DROSOPHILAhttp://dx.doi.org/10.5665/sleep.3206
A Subset of Cholinergic Mushroom Body Neurons Requires Go Signaling to Regulate Sleep in DrosophilaWei Yi, PhD1,2; Yunpeng Zhang, PhD1,2; Yinjun Tian, MS1,2; Jing Guo, BS1; Yan Li, PhD1; Aike Guo, PhD1,3
1State Key Laboratory of Brain and Cognitive Science, Institute of Biophysics, Chinese Academy of Sciences, Beijing, China; 2University of Chinese Academy of Sciences, Beijing, China; 3Institute of Neuroscience, State Key Laboratory of Neuroscience, Shanghai Institutes for Biological Sciences, Shanghai, China
Submitted for publication January, 2013������������ ��������������������Accepted for publication June, 2013Address correspondence to: Aike Guo and Yan Li, Institute of Biophysics, 15 Datun Road, Chaoyang District, Beijing, 100101, China; Tel: 86-21-54921785 and 86-10-64881260; E-mail: [email protected] and [email protected]
��������������� Identifying the neurochemistry and neural circuitry of sleep regulation is critical for understanding sleep and various sleep �������������� ������������������������������������������������������������������������������������� �������������������������������!"#$��������������������������������%������������������������������������������������������������������������������still a topic of controversy.����!��&��������������������������������������������������������������������������������'����(������������������������regionally manipulated by heat induction and the Gal4/UAS system."� ��������� �#������� We found that expressing pertussis toxin (PTX) in the MB by c309-Gal4 to block Go activity led to unique sleep ���������������������������������������������������������������������&�����������������+<=�'��>�(�������������������������"#?FH�����������������������������������'�����������������������K����������������������������������������O�'��Q<���V[\�������������������������������������������������������������������������"#?FH������������������������������������������������������������������'�������������������������������������������������������]�����"#?FH����������������������������������������������������������'����������������^������$����������_��O������������� �������������`��������������������������������������������������������������������������w��������������������������������������������'���������������������������������%��&���� Mushroom body, Go signaling, cholinergic neurons, sleep$�� ����� Yi W; Zhang Y; Tian Y; Guo J; Li Y; Guo A. A subset of cholinergic mushroom body neurons requires go signaling to regulate sleep in Drosophila. SLEEP 2013;36(12):1809-1821.
��$17,18 and mediates many of the effects of DA and 5-HT on sleep.11,19,20�"%���������������%����������������������� -����� ��� ���#�������� ���������&� ����������� ���' ���������� ����%��� ������� %�*����$17,18� %���� � ��������� ��$���������� ��� ��� ����������� ��+�� ��������$� ������� ��$21 ����������������#�����������!����������������/�%��$����������������������������� *�������#�����������������sleep.18�"�� ���� ��� ��� #�� ��� ��� ������� �� ���� ����'����� ������ ������������ :�� ��������$� ��������� ��� #�� ���been found to be modulated by various types of neurons, the ���� �������������������#��������������������poorly understood.22
/�$� %� ��%� ����� ����������� ��� � ������� ��� �� ���� ��������� ��� #�� �����$� ��� #�� ;+<� ��� �����$� ���� ����� ��������=�������������������������#��������are cholinergic and their acetylcholine (Ach) output is required ���� ��� ����������� >������$� %� ?��� ����� ��� ��@� ��� #��neurons play opposite roles in sleep regulation and also require �����������������B��������������������� �����
METHODS
Drosophila��� ���>�������� ������%��*�������������� �����%&�C�E!���"F
���G�������%�"�������� �B��������C��������I�C�E!F"J���� G��� ����� K����� �C�������� ��� /������I� C�E!� L� ���G���M��!Q����R����C�����������B�������������E���>��� � ��I�#�!���SU����G���E ����=�������C�����������#�� �����#�� ���E �����I�B��3.3kb!���SU����G���M��K����F*����C����-����I���GV!���SU����G���"�*�����E�*����"�*���#�����������C��������I� WUX!���Y� ��� G��� ������ E����� �C�������� ���

SLEEP, Vol. 36, No. 12, 2013 1810 Cholinergic MB Neurons Regulate Sleep—Yi et al
F����������I�GZ[/!���Y����G���M��\������"�������C����-������C�E!��B�"KQ�� (#40918) was obtained from the Vienna Drosophila KQ��� B���I� C�E!B��"KQ��� �V[WYZK!Y�$� �109-
68!���Y� ���� QF� ���� ����� ��� Drosophila� ���� � K��� �B����� C�E!�BGS&&�>F� �_ZVW`�$� KVZjUV!���Y$� ���� ���!���SUts��_`UVS�����_`UVX����������������������E�� *�B�����Flies were raised on standard cornmeal, sugar, yeast, and agar �����$������V[��+V[��������+���*��{G�� ����������>���%���������������[Y|B$�' �����������������!���� �����'��-����������%���������������VS|B�
����' �$��� � �#(��(�)�� ��E��� ������� %�� �������� �� � ����� ���������21
}�����������Y!����~!����!�������%����� ������������������65 mm × 5 mm monitor tubes with food containing 5% sucrose in 2% agar and entrained for 24-36 h before sleep recording. {��������%�����\"�U�� �� ����������US&UU�������������������\"�V[�� �� ������ ����[U&UU���{� �������� �������%�����������������{G� � ������������������23,24�:�������{jG�������were used to give constant illumination for video recording. Web cameras with a 640 × 480 resolution were pretreated to allow reception of only infrared light. Images were acquired every 5 sec and further processed by Pysolo24 to determine the �� ����������������E���%���?���������� ��������-ment for 5 min or longer.3� E��� �������� %�� �� ������using Matlab (MathWorks, Natick, MA). Waking activity was calculated as the total walking distance divided by the total time awake.
For experiments without heat induction, sleep was assayed ���� ���� {G� � �� ��� [Y|B�� :�� ���� ���� ����� '������$���� %�� ��������� ��� �%�� {G� � �� ��� VS|B� �� �� ������ ������� ��� ����� ��� ������ ��$� ���� ��� %�� ����'���������V[!��WV|B������� *������\"�V[����\"�U���������V[!��� ����������$������������������'���%��{G� � ��%�� ��� ������VS|B��������������������
>�����!�������� �� �����������������������$����%��treated and recorded in the same condition as mentioned previ-ously, except that the experiments were performed in constant ���*���{� �������� �������%����������������� ���������Z��������� �� ���������B�� *������ ������ $�=�����$�:{����������������������������� �� ��������������"����%���������������%�� �� ��������� ��!���������������� ��?-dence level of 0.01.
� � )� ��������� ������ ��� ������ %�� ���� ��� ����� EFEE� �EFEE�
:� �$� B�� ���� :{���"��=��*!E������� ��� %�� ��� ��� ���-mine the normality of the data. Preheat and postheat records of ��������������%�� �������%�����%�!������������E������t!����"�� ���� ����? ���� ������ � �� ��� �{EG�� %�� ���as a post hoc test after one-way analysis of variance. Data were presented as mean behavioral responses and error bars ����������������������������������Ej#���G����� ���%��������%�� �����������? �������������������������error was less than 0.05 (P < 0.05).
*�����(�����(�������"��'�������������������Y�����%�����������������
������� ������� ���� ���� ��� ������� ��BGS&&�>F$�
��>F�� '������� :�� ���� ���������� ��� '������� ' ����������E����������>�����EY$���>F�'������%������ �����WV|B�����W�����������`!X�������� �������:��>�����EY$���>F�'������%�� ��������������� ������������������!������� �>����� EY�$� EYB�$� ��� ���� �� ���� W� ���� ����� �������>�����EY����"�������������������������������%���� ������ �������������������������F�E������?'�����Y�� ��������������� ��� F�E� ��� ����� ��������� �K"�� ����40 min. After three 20-min washes in phosphate buffered saline "������J!VUU��F�E"$�U�W��"������J!VUU����F�E�$������%����� *��%������� *�����������Z���������������������F�E"������V������K"��E�����%�������� ������%����������������-��������������!>�::����V&VUU������G����������E�����/������������*$�:�%�$����������������!�>F�������������������:�������V&SUU������:��������$�B������$�B��������� *�������������Y|B���������������� ����[U!����%��� ���F�E"$������� %�� �� ������ ��� Y|B� ��������� %���� ������� ����-�����B�W����������!����:����V&ZUUI�:����������������'��>�����YSS����������!�������:����V&VUUUI�:���������������� *��������������������[U!����%������F�E"$�������%��������������} �����������������������} ����{��$�B��������������� ������%������{� ��EFZ::� ���� ����� �� ����\��� *�images were scanned at 1-μm section intervals with a resolution ���V$U[Y���V$U[Y���'���B���� ����� *�%����� �������:���R��Q:/$������$�#������ ����������@ �������%�
*������+������ ��$( ��#� ���������� � GQ�� ��� WUX!���Y� ��� �109-68!���Y� ��� %��
'��� �������������%����/��::��Q%�j��������������:� �$�Ipswich). DNA fragments were then self-ligated overnight at V~|B� ����� "Y� GQ�� ����� �:����������� {������ �������� �������������Z��������F���%����������*��������� �GQ��%�� �����?�� ��� �������� ����� �� ����� �FBK�� %������� ������ ����$� Z�!B"BB�B��""BB�""���""!W�� ����Z�!B"�"B��B����BB�BB""�!W���"��FBK������ ��%����� ���������{�E"��� �����������*����GQ����� �against the Drosophila genome sequence (Flybase.org) was ������������� ���������������������������������
RESULTS
-���.��!/�)������������45/ �67��� �������'��:����For sleep research in Drosophila, a video recording system
has been newly developed, which could detect subtle move-�������������U�WS���$������'�������V+VU���������������length.23 Thus, we set up this system in our laboratory and ��������� ��?���� ����� ��� ����� ����������� ���� ��� ��� ���-����� �>����� EV��� "�� ������ ��������� ����������� �� �$�we adopted the temporal and regional gene expression targeting �"�K�j"������� :�� ���� ���$��� ��������������Y+C�E�������������!� �? �'�������������� ��������������Y!��������������"�����Y����� ����������� ������� ���be inhibited by a ubiquitously expressed and temperature-sensi-�������SU������$����SUts$����VS|B$�%��������������������Y�����������������������������������WV|B$������������expression of this effecter gene.25 The effecter gene expression was only induced by a 12-h heat-shock in 7- to 9-day-old adult ��������������'�������' �������%��������>�����V����Thus, the effect of temporal gene expression on sleep could be

SLEEP, Vol. 36, No. 12, 2013 1811 Cholinergic MB Neurons Regulate Sleep—Yi et al
:�!����—Blocking Go signaling in c309 neurons leads to sleep increase. (A)z������������������������������������������&�������������������������F����!{F|$������������������������������}����������!���������$��������!��������$�������������������(B)~�������O�������+<=�'��>Ftub-Gal80ts%K\~�zw�F� ������������������� �������'��Q<ts/+;UAS-PTX/+ during the sleep recording procedure. Diagonal crossing lines represent heat shock period. (C)~�������O�������+<=�'��>F����'��Q<ts;UAS-PTX/+ are averaged for the preheat or the post-heat recording days. =�5>? Sleep parameters ��������`��������������������������������������������������������+<=�'��>F����'��Q<ts%K\~�zw�F� ����������������������������+<=�'��>F�and tub-Gal80tsF�%K\~�zw�F� ��������������������(D), change in sleep amount calculated by subtracting sleep before heat induction from sleep after heat induction (E), number of sleep bouts (F), average duration of sleep bout during the day or night (G)��������������������������������������������night (H). n�+<�+�������������|������������`�������������������������������(E) and paired Student t-test in other panels. Data are presented ������������������������������������z�<�<<�%�����������O���������������z�<�<��
A
C D
E
G H
L18°C
31°C
elcosion Pre-heat recordadaption Post-heat record4-6 days 24-36h D L D L D L D L D
F
0
40
80
120Day
***
Aver
age
dura
tion
of s
leep
bou
ts (m
in)
***
Nightpre-heatpost-heat
0
40
80
120Day
***
Aver
age
dura
tion
of w
ake
bout
s (m
in)
***
Nightpre-heatpost-heat
0 12 24 36 48 60 72 84 96 108 120(h)0
20
40
60
Sleep
am
ount
per
60
min
(mm
)
c309/tub-Gal80ts;UAS-PTX/+
tub-Gal80ts/+;UAS-PTX/+
0
20
40
60
M
inut
es o
f sle
ep
per
60
min
(min
)
pre-heat post-heat
c309/tub-Gal80ts;UAS-PTX/+
0 12 24
HS
B
0
400
800
1200
pre-heat recordpost-heat record
Slee
p am
ount
(min
) ***
c309
/tubG
al80ts
;UAS-P
TX/+ c309
/+
tubGal8
0ts /+
;UAS-P
TX/+
0
5
10
15
20Day
***
Num
ber o
f sle
ep b
outs ***
Nightpre-heatpost-heat
c309
/tubG
al80ts
;U
AS-PTX/+
c309
/+
tubGal8
0ts /+
;UAS-P
TX/+
c309
/tubG
al80ts
;U
AS-PTX/+
c309
/+
tubGal8
0ts /+
;UAS-P
TX/+
c309
/tubG
al80ts
;U
AS-PTX/+
c309
/+
tubGal8
0ts /+
;UAS-P
TX/+
c309
/tubG
al80ts
;U
AS-PTX/+
c309
/+
tubGal8
0ts /+
;UAS-P
TX/+
c309
/tubG
al80ts
;U
AS-PTX/+
c309
/+
tubGal8
0ts /+
;UAS-P
TX/+
c309
/tubG
al80ts
;U
AS-PTX/+
c309
/+
tubGal8
0ts /+
;UAS-P
TX/+
0
100
200
300
400
c309/tubGal80ts;PTX/+
c309/+
Day
***
Chan
ge in
sle
ep a
mou
nt (m
in)
Night
n.s.
tubGal80ts/+;PTX/+

SLEEP, Vol. 36, No. 12, 2013 1812 Cholinergic MB Neurons Regulate Sleep—Yi et al
�������� ��� ��������� ��� ���?�� ��%�� ��� ������and post-heat recording period (Figure 1A).
=����������������� ����� ��������������� ����������F"J���� � �? � ����������������� ������� �����26,27�� ��� ���#�� ������ � ���� ��� ���� ��� ���21� E����������$� ����������'������F"J�%���� WUX!���Y$��������#�!'���������Y$�������������� ��� �����������������>�����V����������1D). We observed temporally reduced sleep amount during ��������� *����������������� WUX���F"J����� �����������>�����V����"��' ������������������������������ �������������� *!���� ��F"J�'������������������������to heat-induced sleep loss, we performed the heat shock induc-tion during the daytime, during which the sleep amount was ����� ��� �>����� E[���� =� ���� ������ �� ����*������ ���������������������'������F"J���� WUX��������>����� E[���� "��$� ��� ����� ����� ����� '������ ���F"J���� WUX�������������������� ��$��������� ��������this increase is not a result of heat-induced sleep loss.
���������� ��� ��� ���?�� ��� �����$� %� ���� �� ����� ���excessive sleep occurred predominantly during the daytime, whereas the amount of nighttime sleep was unchanged �>����� Vj��� "�� �� ���� ������� ��� %�� ������������to both prolonged sleep bout duration and more sleep bouts �>�����V>�����V��$��������������������������' -sive daytime sleepiness. Meanwhile, the wake bout duration ������� ��� ������� %�� ����? ������ � ���� �>����� V/�$��������� ����� WUX� �� F"J� ��� ���� ���? ����� ����������������� ������ ��� %�*����� %��� ��� ��� ������ ��� ��mainly wakeful. In contrast, nighttime sleep was fragmented ����� F"J� '�����$� ���� ���� ��� �� ����? ������ �� ����number of sleep bouts, as well as a decreased average duration �����������������������>�����V>�����V����=�*����������-��������������%������������� �>�����V/�$� �� ������������������+%�*�%�� �������������������������������and wake states at night.
"�������������+%�*��������������%�*��� �� �-dian rhythm, and we thus tested this possibility under constant dark (DD) condition. Our results showed that temporally '������ F"J� ��� WUX!���Y� '������ ������ ������ WUX��������������������������������� ����������������rhythmicity, but only decreased the relative rhythmic power �"����EV���=�������������������������������������GG� ������������������������������������� *�������� �������in c309 neurons also induced dramatic sleep increment and �� ������+%�*�����������$�������������������{G� ����-������>�����EW���"�������������������������� ����� WUX���F"J����%������ ����������� �� �������������problem. We noticed that heat shock also induces moderate �� ��� ��� ��� ����� ������ ��� ���!���SUts+�IC�E!F"J+�� �����������>�����EWB���=��� ������������������������!���SUts����+���C�E!F"J����������� ��������������������heat to some, albeit subtle, extent, which may only be observed under DD conditions.
7��� ���!/�)������������45/ �6����� �������')�����"������������������������ ������������ ������������$�%�
����������"F$��� ������������� ������������������������$����������������������� ���������� WUX�����������������������(Figure 1A). In contrast to hypersomnia resulting from blocking
���� ������$������������� ���������� WUX�������������������?- ����� �����������������>�����[������[����������? �-�������� ����������� ������ ����� ������������ ������� ���c309 neurons induced sleep decrease during both the daytime ���� ��������� �>����� [B��� G������ ��� ������$� ��� �����number decreased, but sleep duration was comparable to control ����>�����[G�����[j������ ��� �������$���������������during the nighttime was unchanged, whereas sleep duration was ������� ������ ���� �>�����[G�����[j���=�*����������-tion was increased during both daytime and nighttime, indicating more consolidated wakefulness (Figure 2F). Taken together with ������������� WUX���F"J���$�%����������������� �����in c309 neurons to reinforce wakefulness, and that the signals �����������������������%�*!����������������
/�:�������������45� ����"-@KQ$���V��������#�!�� ��Sleep
We next examined which of the neurons labeled by WUX!���Y� %�� �������� ���� ��� �����!������� ����� ��� =��� �BGS&&�>F� ���>F�� %�� �����������'������� WUX!���Y$�����������#��;+<�����������������some neurons scattered through the central brain were strongly ������ �>����� EY��$� ������� %���� �� ������� ������28 /�%��$� %��� %� ���� �� �>F� '������ ���� W� ���� ���`!� ���X!���!���������������� *��>�����W��$�%����������� �%�� '������ ������ ����� %�� ������ ��� WUX!���Y��>����� W���� :�� ������ �� ����� ����� ��� ��� ��� ��� ������ ���������������������������%�� �������� *�%��administered in our behavioral studies (Figures 1A and 3A). In ����� ����$�'������������#��%������ ���������;+<� ���neurons, whose axons run exclusively into the core area of the ;+<����22 (Figures 3D through 3F). When heat induction was performed for 3 days before eclosion, stronger expression in all ;+<�������������%���������>�����EY����"��$������������� ����������������� ������� WUX!���Y�'�������������are due to different developmental stages.
"�� ������� %����� ��� ���� ��� #�� ������ ������ ������ ������ ������ ��� #�� ������ �� ���!#��neurons) were responsible for the sleep defects, we intro-�� �� #�!���SU� ��� � �? ����� �������� ���Y� � ������� ������ #�� �����29� �>����� WB��� B��������� #�!���SU� %���� WUX!���Y� �������� ��� *�� ��� F"J� '������ ���� ����� �� ������������������ ������"F expression induced ������ ������>�����Y������Y����:����������$� ������������ ��+%�*������ ���� �������� ��������� ��� �������������������������%���� ��������������F"J��������"F experi-���� �>����� YB� �������� Yj��� "�� ����� �������������������������������� ���������� WUX!���������!#��������had no effect on either total sleep amount or sleep structure, �������� ����� ��� WUX!#�� ������ ��� �������� ���� �������� ����� WUX���F"J����� WUX������"F����
=�����'��������������������������%��$�������%��$�%���� ��� ��� ���� ���� ����� ���������� B� �� !�#F!���-�����������*�����F����������%��������� ���������������%��$����� �����������'������� L$��� ������������� ��������F��� ������� �������$���� WUX!���Y���������17 As expected, we observed severely reduced and fragmented ��� %��� � L� %�� ���������� '���� ��� WUX� �����$�however, sleep was still severely reduced and fragmented when

SLEEP, Vol. 36, No. 12, 2013 1813 Cholinergic MB Neurons Regulate Sleep—Yi et al
WUX!���Y� %�� ������� %���� #�!���SU� �>����� EZ��� =�����������������������������������������������������������#���������!#�������$�%����������������������������������#��������%��������� WUX�������
:�������� ������� ����������������� �� �������� ��� �� �-motor activity control.30 Although we noticed decreased
waking activity (the walking distance divided by waking time) ��� ����� WUX� �� F"J� ���� WUX� �� ���"F� ��� �>����� E~�$� ������������������� �� ������� WUX���F"J������ ������� WUX������"F�����>�����VB�����[B���#�����$� WUX$#�!���SU� �� F"J� ��� ���� '������� ������� ��� �� %�*����� ��������>�����E~�$�%����������'�����������������
:�!����—Elevating Go signaling in c309 neurons decreases sleep. (A)~�������O���������������������������(��������'�GTP������+<=�'��>����tub-Gal80ts. =-5:?~��������������������!������������$��������������������!�����������$���������`������+<=�'��>FK\~�'�GTP;tub-Gal80tsF� ���and their parental controls c309-Gal4/+ and UAS-GoGTP/+;tub-Gal80tsF� ��������������������(B), change in sleep amount during daytime and nighttime (C), number of sleep bouts (D), average duration of sleep bouts during daytime and nighttime (E)����������������������������������������������nighttime (F). n�+<�+�������������|������������`�������������������������������(C)�������������~������t-test in other panels. Data are �������������������������������������������z�<�<<�%�����������O���������������z�<�<��
A B
C D
F
-600
-400
-200
0
200
400c309/GoGTP;tubGal80ts/+
c309/+GoGTP/+;tubGal80ts/+
***
***
Day
***
Chan
ge in
sle
ep a
mou
nt (m
in)
Night
0
20
40
60
Min
utes
of
sleep
p
er 6
0 m
in (m
in)
pre-heat post-heat
c309/UAS-GoGTP
;tub-Gal80ts/+
0 12 240
400
800
1200
pre-heat recordpost-heat record
Slee
p am
ount
(min
)
***
c309
/UAS-G
oGTP
;tu
bGal8
0ts /+ c3
09/+
UAS-GoG
TP /+;
tubGal8
0ts /+
0
5
10
15
20Day
Num
ber o
f sle
ep b
outs **
Nightpre-heatpost-heat
c309
/UAS-G
oGTP
;tu
bGal8
0ts /+
c309
/+
UAS-GoG
TP /+;
tubGal8
0ts /+
c309
/UAS-G
oGTP
;tu
bGal8
0ts /+
c309
/+
UAS-GoG
TP /+;
tubGal8
0ts /+
E
0
40
80
120Day
***
Aver
age
dura
tion
of s
leep
bou
ts (m
in)
Nightpre-heatpost-heat
c309
/UAS-G
oGTP
;tu
bGal8
0ts /+
c309
/+
UAS-GoG
TP /+;
tubGal8
0ts /+
c309
/UAS-G
oGTP
;tu
bGal8
0ts /+
c309
/+
UAS-GoG
TP /+;
tubGal8
0ts /+
40
80
120
160Day ***
Aver
age
dura
tion
of w
ake
bout
s (m
in)
***
Nightpre-heatpost-heat
c309
/UAS-G
oGTP
;tu
bGal8
0ts /+
c309
/+
UAS-GoG
TP /+;
tubGal8
0ts /+
c309
/UAS-G
oGTP
;tu
bGal8
0ts /+
c309
/+
UAS-GoG
TP /+;
tubGal8
0ts /+

SLEEP, Vol. 36, No. 12, 2013 �W�6 Cholinergic MB Neurons Regulate Sleep—Yi et al
������� �>����� Y����"�� ����� ����� ����� ����������������� ���������� WUX����!#����������������� ��� �������� ������I���%��$����������� ���������������������� ������������� ���������� ��� ����� ��� ��������������locomotor activity defects. The sleep defects we observed ����������������������������? ������ �������� ��������������=��������>�::������������ � *���������������������� ���#��;+<������� �����=������� �����'���������F"J�������"F over a 12-h period did not alter the global structures ��� ��� #�� %��� '������ ��� ����� ��� ��� ��� ��� ���!����� ��������>�����E`�$�����[U����������'�������>�����ES���Thus, we suggest that the behavioral changes are not due to �������������!������� ������ �����������#��
/���#�X���������������"-���!���'�Y( �> ���''�����:��������������'#�!�� ����
j���� �� ����!���� ���Y� ���� %���� ��@� ��� ��������sites in the genome may share similar expression patterns. =������� ��� �������� ������ WUX!���Y� ���S$~~`$XWW����the second chromosome, upstream of the gene scabrous. We � *�����'�����������������������@� ������Y������>�����Z��������������!���� �������� ����������������(Figure 3A), and unexpectedly their expression patterns were
���������B�������%���� WUX!���Y$� �109-68!���Y���%�������� %�*� ������ ��� ��� #�� ���� ������� '������ ��������� #�� �>����� Z�� ���� ZB�$� %���� QF[V[[!���Y� ����QFY[SX!���Y� ��%�� ������ '������ ��� #�� ;+<� ��������� ���� �� �� �� '������ ������ ��� #�� �>�����ZG� ���� Zj��� "�� ���Y� ���� ����%� �� ��� ������� '�������� �� �� ��� ��� ��� ����� ��� ��� ����������� ��� *���� ���� ��������������������'������F"J�%����QF[V[[!���Y����QFY[SX!���Y$��������� �109-68!���Y$�������������? ������ ���������������$���%����� ����� WUX���F"J�����>�����Z>��� "�� ����� ������� ������� ���� �������� ����� ���� ������� ��� ���#��;+<� ��������� �� �������� ��������� ����� WUX���F"J����
=� ��������� ������� ����� ��� *���� ��� � ������� ��� F"J���� ��� #�� ��� �� ��� ������$21 and thus we wondered %�� �����������#��������%��������������������������������������=��������#�����Y����$�%�� ����%������ ���'������������#������������������������������������%������� ����������� *��������������>�����W����GZ[/!���Y���%��������'���������;+<����� ������$�moderate expression in posterior neurons, but no expression in ��� ������$�%����KVZjUV!���Y�����QFWU~V!���Y����������� ;+<� ��$� ���� $� ���� �������� ������ ��� �������
:�!������+<=�'��>���(���������"#?FH����������������������� ���� (A) Procedure of heat induction of mCD8::GFP for examining the expression pattern. =-5$?�����"#��������+<=�'��>��������������(���������������������������������"#�'��Q<�=�5:?z��]���������������"#��������(D) ��������������`�������������������������?����=75:?�����������!���$�������������������"#��������'z������!�����$��������������+<=�'��>�(���������������������������������������������"#?FH�����~������������������<����#���������<�����5:.
3 days
18°C 31°C
7-9 dayseclosion dissection
c309/UAS-mGFP;tubGal80ts/+
A
B C
D
FasII
� tipE F�
�
�
�
c309,MBGal80/UAS-mGFP;tubGal80ts/+
c309/UAS-mGFP;tubGal80ts/+
GFP+FasII

SLEEP, Vol. 36, No. 12, 2013 1815 Cholinergic MB Neurons Regulate Sleep—Yi et al
GZ[/!���Y�����KVZjUV!���Y����������'������ ��� ������������>�����~����������~B��
"������������� ����F"J�'���������GZ[/!���Y�����������? ������ ��� �� ��� ������� �>����� ~G���=��� �����-�� ���� #�!���SU� ��� ������� GZ[/!���Y� � ������� ��� ��� #��expression, sleep amount was statistically indistinguishable ����� ���� ����� ���� �� *� ���� ����$� �������� ����� ���� �������������;+<����� +�������������������������������������������>�����EX������EXG���:���������$���� *�������� ����������KVZjUV!���Y����QFWU~V!���Y����������������� ����? ������ � ���� ��� �>����� ~j��� #��%���$�these sleep effects could be abolished by the presence of #�!���SU$����������������������������#��������'������
%�� ������� �>����� EX�� �������� EXG��� "�� '������������� ��� ��� ���� ���Y� ���� ������� ��� ��� ;+<� ���� +posterior neurons. Therefore, our results demonstrate that in ���������������;+<� ��������$�������������������������������������������#�������$�� �������;+<����� +���-rior neurons, and their effects on sleep regulation are opposite ������;+<� ���������
"-@KQ$���V������ ��$(������!�� ���&�5��!�� ���!)$(��!� �*�(���������'
"��'�����%�� ������������������������������#��;+<� ��������$�%�������SU�����������������'���������� �? ��������������B��3.3kb!���SU�������GV!���SU����
:�!���6—MB-Gal80 eliminates PTX- and GoGTP- induced sleep effects in c309-Gal4. (A)~�������O�����������������������������������������������MB-Gal80 is introduced. =-57? Temporally expression of PTX or GoGTP�����+<=�'��>�"#�'��Q<��������������O�����������������������(B), number of sleep bouts (C), sleep bout duration in daytime (D) or sleep bout duration in nighttime (E). Light gray bars and dark gray bars indicate sleep parameters for }����������!{|$���������������������������������������������|������������`����������~������t-tests; n = 30-36 in each group. Data are presented as means ± standard error of the mean. ***P < 0.001, paired Student t-test.
0
20
40
60
Min
utes
of
sleep
p
er 6
0 m
in (m
in)
pre-heat recordpost-heat record
0 12 24
A
B
c309,MB-Gal80/tub-Gal80ts;UAS-PTX/+
0
20
40
60
pre-heat recordpost-heat record
0 12 24
c309,MB-Gal80/UAS-GoGTP;tub-Gal80ts/+
D E
C
0
400
800
1200 pre-heat recordpost-heat record
Slee
p am
ount
(min
)
c309
,MBGal8
0/
GoGTP ;tu
bGal8
0ts /+
c309
,MBGal8
0/
tubGal8
0ts ;P
TX/+
c309
,MBGal8
0/+
GoGTP /+;
tubGal8
0ts /+
tubGal8
0ts /+;
PTX/+
0
40
80
120
Aver
age
dura
tion
of
slee
p bo
uts
in d
ay (m
in) pre-heat record
post-heat record
c309
,MBGal8
0/
GoGTP ;tu
bGal8
0ts /+
c309
,MBGal8
0/
tubGal8
0ts ;P
TX/+
c309
,MBGal8
0/+
GoGTP /+;
tubGal8
0ts /+
tubGal8
0ts /+;
PTX/+0
40
80
120
Aver
age
dura
tion
of
sleep
bou
ts in
nig
ht (m
in)
pre-heat recordpost-heat record
c309
,MBGal8
0/
GoGTP ;tu
bGal8
0ts /+
c309
,MBGal8
0/
tubGal8
0ts ;P
TX/+
c309
,MBGal8
0/+
GoGTP /+;
tubGal8
0ts /+
tubGal8
0ts /+;
PTX/+
0
10
20
30
40
Num
ber o
f s
leep
bou
ts
pre-heat recordpost-heat record
c309
,MBGal8
0/
GoGTP ;tu
bGal8
0ts /+
c309
,MBGal8
0/
tubGal8
0ts ;P
TX/+
c309
,MBGal8
0/+
GoGTP /+;
tubGal8
0ts /+
tubGal8
0ts /+;
PTX/+

SLEEP, Vol. 36, No. 12, 2013 1816 Cholinergic MB Neurons Regulate Sleep—Yi et al
������� ��� ����� ���Y� � ������� ��� �������� � �����31
���� ������� � �����$32 respectively. We found that WUX!���Y� '������ ��� ��� #�� %�� ���� ��� ��� ��� ������ � ��� ��GV!���SU$� ���� %�� �������� ��������� ���B��!���SU� �>����� `�� ���� `��$� �������� ����� WUX!#�������� ��� �������� � ���� ���� ������� �� =� ���� '��!���� ��� ��� ����� ��� ��� ���� ���� ������ ����� ���������B��!���SU$�����������GV!���SU$�%���� WUX!���Y� �������� *���� ��������� ���������������������������%����F"J������������� �%�������"F������ ������� �>�����`B���"�� ���������������� WUX!#���������������� ����������� ����������cholinergic.
}� ������ ��� �������������������B�"���������������acetylcholine transport and release.33� E��� %�� ���� ��� ���
when we temporally knocked down vAChT����KQ��������� ��in the c309 neurons at the adult stage for up to 5 days (data ������%��$������������������?� ����*�� *��%������������B�"�����������������������=���vAChT was continu-����� *�� *�� ��%�� ��� WUX� �����$� ���� ��%�� ����?�!cantly reduced amounts of sleep compared with their parental ��������>�����̀ G���B���������B��!���SU����#�!���SU�%���� WUX!���Y� ����������� *������������ ������>�����̀ G���=��������� ������������� ����#�!���SU��������������!nated this sleep reduction, but also caused an increase of sleep amount (Figure 7D). We suggest that some c309 neurons ���������#����������������������B��������$�%������� ��������� %�� ���� � ������ ��� ��� ������ ���� �������������� B������ � ����������� �B��"�� �� '����
:�!���\�����������'����������+<=��������'��>����������������������������������� (A) Schematic diagram of the P-element insertion sites of several Gal4 lines in the scabrous ���������������\���������������������������������������������������w����������������������������������������������the second chromosome. =-5�?�������������(���������������������������'��>�������������(�������������'��Q<ts. c309 (B), NP2122-Gal4 (D) and NP4289-Gal4 (E)������������(�������������"#?FH����������������~��109-68-Gal4 (C)�����������(�������������"#�~������������������<���(F)w��������(����������zw�����[z}�}}���[z>}Q=�'��>�����������~��109-68-Gal4, leads to increases in sleep amount. Numbers on bars indicate ����������� ���������������|������������`����������~������t-test. ***P < 0.001.
B C D
A
F
8667603 8667928 8667933 8667936 8668049
NP4289 sca109-68 c309 NP2122scabrous
c309>mGFP;tubGal80ts sca109-68>mGFP;tubGal80ts NP2122>mGFP;tubGal80ts NP4289>mGFP;tubGal80ts
E
55 34 17 37 16 27 330
400
800
1200
Slee
p am
ount
(min
)
pre-heat recordpost-heat record******
NP2122
/
tubGal8
0ts ;
UAS-PTX/+
NP2122
/+
tubGal8
0ts /+;
UAS-PTX/+
NP4289
/
tubGal8
0ts ;
UAS-PTX/+
sca1
09-68 /
tubGal8
0ts ;
UAS-PTX/+
Sca1
09-68 /+
NP4289
/+
SLEEP, Vol. 36, No. 12, 2013 1817 Cholinergic MB Neurons Regulate Sleep—Yi et al
only in cholinergic neurons and is critical for the synthesis of acetylcholine.34����*�� *������%��ChAT�%���� WUX!���Y$�%�observed similar sleep amount decrease, which could also be ������������#�!���SU��>�����EVU���B���������*�� *����down of vAChT or ChAT� ��� ���#��������� ��#�����-opment, thus causing the sleep defects. We then checked the
�����������������#��������������������>�::����������"���������������%������������ ����������������������'������ �>����� EVV�� �������� EVVj��� "��$� ��� ���-������� ����� ����� WUX!#�� ������ ��� �������� $� ��������� ����B�� ������ ��� ��� ������ ����� ��� ������ ����in sleep regulation.
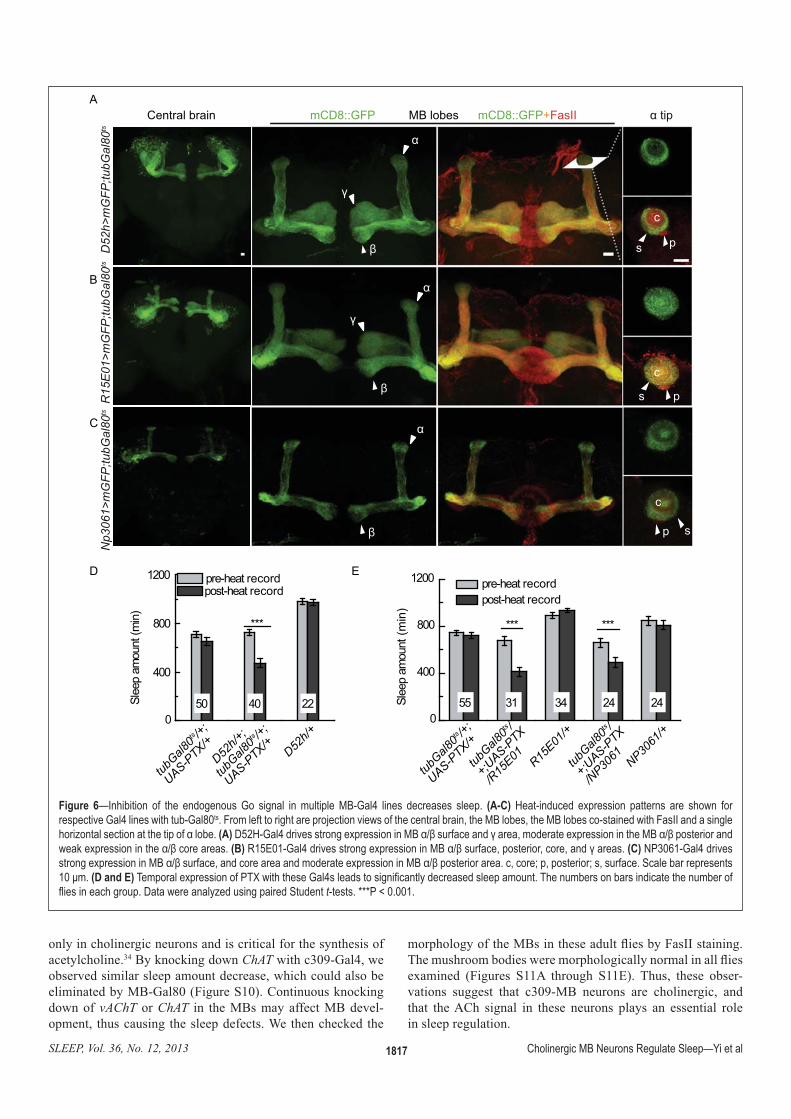
:�!��� ^—Inhibition of the endogenous Go signal in multiple MB-Gal4 lines decreases sleep. =)5$? ������������ �(�������� �������� ��� ����� �������������'��>�������������'��Q<ts���������������������]��������������������������������"#���������"#�������������������������������������`����������������������?�����(A)|�}��'��>�������������(����������"#?FH�������������������������(�������������"#?FH�����������������(�������������?FH����������(B)V���<��'��>�������������(����������"#?FH���������������������������������(C) NP3061-Gal4 drives �������(����������"#?FH�������������������������������(����������"#?FH��������������������%�����������%����������~������������������<���(D and E)w��������(����������zw����������'��>�������������O���������������������������w���������������������������������� ���������������|������������`�������������~������t-tests. ***P < 0.001.
AmCD8::GFP
B
C
D E
50 40 220
400
800
1200
Slee
p am
ount
(min
)
pre-heat recordpost-heat record
***
mCD8::GFP+FasII ��tip
55 31 34 24 240
400
800
1200
***
pre-heat recordpost-heat record
S
leep
am
ount
(min
)
***
�
�
�
�
�
�
�
�
ps
c
ps
c
p s
c
Central brain MB lobes
D52
h/+;
tubGal8
0ts /+;
UAS-PTX/+
D52h/+
tubGal8
0ts /+;
UAS-PTX/+
tubGal8
0ts /
+;UAS-P
TX
/R15
E01 R15E01
/+
tubGal8
0ts /+;
UAS-PTX/+
tubGal8
0ts /
+;UAS-P
TX
/NP30
61 NP3061
/+
R15
E01
>mG
FP;tu
bGal
80ts
D52
h>m
GFP
;tubG
al80
tsN
p306
1>m
GFP
;tubG
al80
ts

SLEEP, Vol. 36, No. 12, 2013 1818 Cholinergic MB Neurons Regulate Sleep—Yi et al
E�� � ����� ��������� ���� ����$� WUX!���Y� ���� �����'������ ��� ��� ;+<� ���� +�������� ���� �� �����$� ���
��������� ��� #�� ;+<� ��� ������ �>�����EY���� "��� %� %������ %�� �� ��������� ��� #�� ������ ��������� ��� ������ � ��� ��� WUX� �� ��B�"KQ��� ����=� ������ ����� %��� GZ[/!���Y� %������%����� ������������������>F�'��-���$������>F��������������%����������� ��� ��� ��� #�� ;+<� ��� �����$�whereas strong expression was observed in ���#��;+<����� +�������������������$�which is identical to heat induction at adult ���� ����� �>����� EV[�� �������� EV[B���:������ ���� ��!���SU� ������������� ���'������������#������������������������� �����������������;+<����� +���-������������>�����EV[G���������EV[>�$������������������;+<����� +�������������������� ������������������ �������� ��=� ���� ��� GZ[/!���Y� ���� ������-����� '���� ��B�"KQ�� to reduce �B�� ������ ��� ��� ���� +�������� ������ ������� B��������� *�� *���� ��%��of vAChT in D52h neurons did not cause ����!���� ��� ������ �� �� ��� ��� #���>����� EVV>��� :�� ������� %���� ��� ������ �������� WUX�����B�"KQ�����$����GZ[/�����B�"KQ��������%���� ����sleep compared to their parental controls (Figure 7D), which could be eliminated by ��� ��� � ��� #�!���SU� ��� B��!���SU��>����� EV[���� "�����$� ��� ��!�������� �� �� ��� WUX� �� ��B�"KQ�� ����������������������������������� +�������� ���� �� ������� "�� ���������� ����� ����B�� ������ �� ������� ������#��;+<� ���������������������$����� ��� ���� +�������� ���� �� ������ ���promote wakefulness.
Q������$���� ��������B�����������#��neurons resulted in similar sleep defects ��� ���� ��� WUX� �� ���"F� ��$� �� �������decreased total sleep amount, fewer sleep bouts during the daytime, and compa-rable sleep bout numbers and shorter sleep bout duration during the nighttime (Figure EVW� ���� >����� [G� ���� [j��� "�� ?��-���� ����� ����� ��� ��� �������� � #�������$� ��� ��� ����� �� ��� ����������������������������B���������
DISCUSSIONC��������������$�%������������
��!����� ������������������ ����������������������������� �������#����� ����������������������=�������������%����@�- ��� ������ ��� #�� �������� � ������������ ����� ��� ��������� ����� �� �������
��� �������� ���� >��������$� ��� �%�� ������ ��� #��neurons play opposite roles in sleep regulation.
:�!���_—c309-MB neurons are cholinergic, and reducing the ACh signal in these neurons affects sleep.=)5-? c309 expression in the MB is blocked by Cha-Gal80 but not GAD1-Gal80 in adult stage ���� ~������������������<���(C) Cha-Gal80, but not GAD1-Gal80, eliminates sleep defects induced by temporally expression of PTX or GoGTP mediated by c309-Gal4. n = 25-39 in each group. (D) Continuously expressing vAChTRNAi���+<=�'��>��������������������������������������"#�'��Q<������'��Q<�~�����������������������|�}���\��wRNAi ����[���������������������������������� ������������������z�<�<<���������~������t-tests (C)���������analysis of variance (D)�������������O���������������
c309/mGFP;tubGal80ts/cha-Gal80
A B
D
c309/mGFP;tubGal80ts/GAD1-Gal80
0
400
800
1200
Slee
p am
ount
(min
)
pre-heat recordpost-heat record
***
***
C
c309
/GoG
TP ;tub-
Gal80ts /ch
aGal8
0
GoGTP /+;
tubGal8
0ts /+
tubGal8
0ts /+;
PTX/+
c309
/tubG
al80ts
;PTX/ch
aGal8
0
c309/+
;chaG
al80/+
c309
/GoG
TP ;tub-
Gal80ts /G
AD1Gal8
0
c309
/tubG
al80ts
;PTX/G
AD1Gal8
0
c309/+
;GAD1G
al80/+
33 43 22 54 52 29 25 31 330
400
800
1200
*** ***
Slee
p am
ount
(min
)
n.s. ***
c309
/+;vA
chTRNAi /+
c309
/+
c309
/MBGal8
0
;vAch
TRNAi /+
vAch
TRNAi /+
c309
/MBGal8
0
c309
/;vAch
TRNAi
/chaG
al80
c309
/+;ch
aGal8
0/+
D52
h/+;;
vAch
TRNAi /+
D52h/+

SLEEP, Vol. 36, No. 12, 2013 1819 Cholinergic MB Neurons Regulate Sleep—Yi et al
:������� ���)�� !�������` ������"-@KQ`���������#�������$���������������������������� ���
�����������;+<����$�������� �����;+<� ���������35 They ��� ���������� ����� ������� ��� ��@� ��� ;+<� ���� � �����$����� ���������������������������� �!������������Y�����as shown in previous studies22,28,36������������������>�����W�$�ZG$� ����Zj�$� �������������������'���������?�� ����������� ������� K ����$� ��� #�� ;+<� ��� ������ ����been highlighted to play a special permissive role for long-term memory consolidation in Drosophila.37 In the sleep research, ������������#����������������������%�*!���������������������$� ��� ��� ���� ��� ������ ����� #�� ;+<� ��� ������have sleep-promoting function by manipulating PKA activity %������ ��� �� ���������17� /�%��$� ��� ���Y� ���� ��� ����'���� ��� ���!#�� �����$� � �� �� ��� ���� ���� �����(PI), in which PKA signal is also required to regulate sleep.16 "��$������������ ����%��������#�� ��������������!�����%�*!���������������$�������%������������������:������ ���������� ���"�K�j"����$� WUX����!#��������also contributed greatly to the PKA-mediated sleep regula-������>�����EZ�I���%��$���� ���������%����#�!���SU����� ���������%�������Y������������ ��������������� �109-68, NP2122 and NP4289), we demonstrated that the sleep defects %�������������������� ���������� WUX��������������������������#��;+<� ����������"�*���������%����������������� �� �� ��� ��� *������� ����*�� *���!��%���B�� ��������� ������� ���Y� �������� �������������� #�� �����$� %������������������#��;+<� ��������������������������!������������ �$�%���������@� ������������#��neurons exert wake-promoting function.
:�� ��������� ����� ������ ����������������� �#��������are spatially very close, forming the inner layer and outer layer ��� ��� #�� ;+<� ���$� �� �����$� ������� ��� ���������� �����these neurons may have direct connections. In the behavior '������$� %��� ��� *���� ��� ����������� ��� ��������� �������� �����$� ��� ��� ���?�� %�� ���� ��*� ����� %��� ���%����� *����������������� ��������>�����~j���=�����suspect that there may have been functional interaction between ����%��#����������������������������#�����������������interactions between the sleep-promoting and wake-promoting �� ��������������������� ���������+%�*�%�� �������� ����!�����%��$38 whereas the molecular pathway has not ��� �� ������ "�� ��������� ��� ����������� ��� ����������������� ��!� ����%�*!����������������� ����� ��������may play an essential role in mediating the inhibitory signals between the antagonistic neuropils.
$(������!��V ���������������"-V������������������#�����������������������������������������
functions in sleep as well as learning and memory, its neuro- ������� ��� ���� ��� %��� ���� ������� :���������������� B��"� �� �� ������ ��� ��� #�� ���$39 however, it is not �� ���������������������%�����#��������'���ChAT���������� ������@ ���������� �������� �������40 The 3.3 Kb sequence upstream of ChAT gene directs expression �������������������������������������������B��"��������41 B��3.3kb!���SU����%�� �����SU�'������%����� �������������*������� $� �������������������Y�'������������#�� ������ ��� ������� %��*42 and this study (Figure 7A),
���������������#����������� �������� ��Q������$�%���%�that knocking down vAChT and ChAT�%���� WUX!���Y��������� ��� ��� � ����$� %�� �� ��� �� ��������� ��� B��3.3kb-���SU����#�!���SU��>�����`G�����>�����EVU���"�������������� ���?��� ��� ����������� � �������� �����������������#����������� �������� �����������B����������������������������������������
In mammals, pharmacological and electrophysiological studies suggest the cholinergic nuclei in the brainstem promote rapid eye movement sleep or wake, whereas in the forebrain they correlate with wake only.43,44 Direct lesions of different putative cholinergic nuclei showed different effects in rapid �����������$��������� ����� ������� ���������B��system in sleep.45� :�� ���� ����� ��� ��$� ��� �����B�� �������in different subsets of cholinergic neurons affected sleep in different ways (Figure 7D), suggesting there are also different �������� ������� ����� ������'����������� ���� �����sleep regulation. Interestingly, the functionally opposite cholin-��� ������������������@� ������� ��������"�������������������� �� ����������%��������%���������������������cholinergic system in sleep control.
/���!� ���! �$(������!��V������������'���������� ����? ���� �����$� ��� *���� ��� � ������� ��� ��� #��
;+<� ��������� ��� ��� � ���� ������������"�����displayed greatly increased daytime sleep when they should ��%�*�����>�����Vj�$������������������������������Moreover, they also showed fragmented nighttime sleep and ���������+%�*�%�� ���>�����V>���������V/���"��phenomena are always found in narcolepsy, one of the most common hypersomnia sleep disorders, but not in other types of hypersomnia such as idiopathic hypersomnia.46 Narcolepsy �� ����� ��������%�����? �� �������'����������'����� ������� ��� � ������ �;11� ���� ��� ����������� F"J!��������+�� ������ in vitro and in vivo in mesopontine neurons, including the cholinergic nuclei.47,48 Furthermore, increased �B��������������������������� �������������$49 whereas whether and how the cholinergic system is involved in the pathology of narcolepsy are unclear. Our work with fruit �����%���������� *���������������������� ����������� �? �cholinergic neurons led to several narcolepsy-like sleep defects, ������������? ������������������� ���� �������� � �����may be directly related to this sleep disorder. Future studies ��������� �������������������B�������� ������������� �����neural circuits may provide new understanding for the patho-genesis of narcolepsy.
ABBREVIATIONS�B�$�� ��� �����B��"$�B������� ����������DA, dopamineG�#E$�Drosophila activity monitor systemDD, constant dark condition{G$������!���*� ��������#�$�#�������������>F$�������!������!�>FOA, octopaminePIs, pars intercerebralisPKA, protein kinase A

SLEEP, Vol. 36, No. 12, 2013 1820 Cholinergic MB Neurons Regulate Sleep—Yi et al
F"J$���������'��Kj#$������������������"�K�j"$���������������������������'������
targeting system.��B�"$��� ������ ��� ���������������\"$�����������
ACKNOWLEDGMENTS"�� ������� ����*�M�� Q�� R��$���� E����$���� "�������$� E��
=�����$����K����$�"��E�*��$�M��K��$�����M��\����$���%��������������������E�� *� ���$����}�����Drosophila�KQ���center and the Drosophila� ���� � K��� � ���� ���� ����� *��=���������������J��G��>�������������%�������������-%��$� ���\��F��\����$�\��/��=�$�����/��#��{�� ��������%����manuscript preparing, to J. W. Hou for technical assistance, and ������������������{������� ���� ����� ������
DISCLOSURE STATEMENTThis was not an industry supported study. This work
%�� �������� ��� ������ ����� ��� �X`W�� F��@ �� �������[UVVB��UUYUU����G�����*�����$����Q��������E �� �>�����-��������B������������WVVWUU[`����G�����*����$�WVU`UXZ~�����XVVW[`UX� ���G���M���{��$����/������"�����F��@ ����� ���B����� ����� ��� E �� � ������� �EBJ[!M=!K!VZ~� ���G���M���{��$���������E������ �F��������K�� ��F��������������B����� ��������E �� ��������JG�U[UYUVUU��
REFERENCES1. E����� �$� #������ j�� ���� � ��� ��� ���� ��� �������� B���
[UVVIVY~&VXY
SLEEP, Vol. 36, No. 12, 2013 1821 Cholinergic MB Neurons Regulate Sleep—Yi et al
42. =��B!{$�J���E$�>��"!>$�������E� �? �������������Q#G��� �����for long-term memory consolidation in Drosophila ellipsoid body. Nat Q��� ��[UU`IVU&VZ`S!S~�
43. E �%�����RK$�K����"��Q�����������������������%�*����&���� � �� ����� ���� �������� �������B����Q��������� ���[UUSI~&W~`!`S�
44. E���������FR$��������RB$�/���*��ER��� ��� ��������������������������Kj#���&���� �� ���������� ���� �������� ������������� ���������������� �����������K��F����� ���"�'� ���VXS`I[`&VW`!Z~�
45. =���� //$� R��� �j�� Q�����'� � ����� ��� ��� �����������pontomesencephalic tegmentum-cholinergic cell area in the cat. II. j�� ���������!%�*���������������K�VXSSIYZS&[SZ!WU[�
46. B�����{�� B���? ������ ��� ��� �������� :�&� {!B������ "{$� ���E��&��� ����������������*��Q%�R��&�R����=������E��$�:� $�[UUZ&SW!X�
47. #����!����������j$��������#$��������B$�K����!E�����>$�G�������:��E��!%�*������� ��������� ����@ ������������ �����V����'������ ��� ������ ����� ���� ��� ��� ��� ����� ������� ��������� j��� R�Q��� ��[UUSI[S&WWV!YV�
48. \��� M$� #�%�� M$� M�����*�� �$� �� ���� ��'��� � ����� ���!V� �����' ������� ��� ������ ��'��!�������� �!������$� %���� ��'���receptor type-2 couples to both pertussis toxin-sensitive and -insensitive �!��������R�F����� ���E ��[UUWIX[&[ZX!~~�
49. �����������#$�����*�E{$�F�����Fj$�������Q�� ����� ���'���� �����knockout mice express enhanced cholinergic properties in laterodorsal ���������������j���R�Q��� ��[UVUIW[&VWU!Y[�

SLEEP, Vol. 36, No. 12, 2013 1821A Cholinergic MB Neurons Regulate Sleep—Yi et al
SUPPLEMENTAL MATERIAL
:�!������w������� ��������������� ���������������� �� ���� (A) ������� ��������� ������� �� �� ���������� � �� � ���� ����� �����recording. (B) A representative picture captured by the video system. ����������������� ������������������"���������� ���������������`�������������������������������������������!���������$�close to the food (tube 8), in the middle (tube 3), and close to the cotton !����}$�#������������������������������������������� �������������false negatives (active but taken as inactive) and false positives (inactive ����������������$�������������������`�������������
PositionFalseNegativeZT12-13
FalseNegativeZT16
FalsePositiveZT12-13
FalsePositiveZT16
1 0.001 0 0 02 0.00833 0 0 03 0.00667 0 0 08 0 0 0 0
A
B
:�!��� ��—Temporal expression of pertussis toxin (PTX) in c309 neurons leads to sleep increment. (A) ~���� ���O��� �� �+<=�Gal4/tub-Gal80ts%K\~�zw�F� ��� !����� ������ �^������ n = 29) and ����������� ������� ����� ����'��Q<ts/+;UAS-PTX/+ (gray solid circles, n�}�$�&������������������������������������F������������(B) ~������������������O������������������+<=�'��>F����'��Q<ts;UAS-zw�F� ���������(��������zw�����}���������������������������the daytime. Data are presented as the mean ± standard error of the mean.
0 12 24 36 48 60 72 84 96 108 120h0
20
40
60
Slee
p am
ount
per
60
min
(mm
)
tubGal80ts/c309;PTX/+ tubGal80ts/+;PTX/+
0
400
800
1200
c309/+tubGal80ts/+;PTX/+
pre-heat record post-heat record
Sl
eep
amou
nt (m
in)
***
c309/tubGal80ts
;PTX/+
A
B
pre-heat record heat post-heat record
Table S1���������������������������������������� ���������}������������������������}�����������
Genotype Period ± SEM (h) Power ± SEM %R Group numberc309-Gal4/tub-Gal80ts;UAS-PTX/+ 24.0 ± 0.5 1823.6 ± 46.6 100 15C309-Gal4/+ 23.3 ± 0.1 2570.7 ± 81.3 100 16

SLEEP, Vol. 36, No. 12, 2013 1821B Cholinergic MB Neurons Regulate Sleep—Yi et al
:�!�����—Blocking Go signaling in c309 neurons leads to hypersomnia during constant dark (DD) condition. (A) ~���� ���O��� �� �+<=�'��>Ftub-Gal80ts%K\~���������� ��(�� !zw�$F� ��� !����� ������ �^������n���$��������������������� ����� ����'��Q<ts/+;UAS-PTX/+ (gray solid circles, n�Q$�{���������������������������������������subjective light/dark periods. (B and C) Sleep amount (B) and bout number (C) �� �����O������ ��������� ����� ���������� �������� zw��(�����������+<=��������|�����������`����������~������t-test. ***P < 0.001; *P < 0.05. Data are presented as the mean ± standard error of the mean. Numbers on the bars represent group number.
15 16 80
400
800
1200 ***
Slee
p am
ount
in D
D(m
in)
pre-heat record
post-heat record
c309/tub-Gal80ts;UAS-PTX/+
c309/+ tub-Gal80ts/+;UAS-PTX/+
15 16 80
10
20
30
40 pre-heat record
post-heat record
Sle
ep b
out n
umbe
r in
DD
***
*
c309/tub-Gal80ts;UAS-PTX/+
c309/+ tub-Gal80ts/+;UAS-PTX/+
A
B
0
20
40
60
Sle
ep a
mou
nt p
er 6
0 m
in(m
m)
c309/tub-Gal80ts;UAS-PTX/+ tub-Gal80ts/+;UAS-PTX/+
pre-heat record heat post-heat record
C
:�!��� �6��+<=�'��> �(�������� ������� �� ��������� ������� w�� �genotype is c309-Gal4/UAS-mCD8::GFP;tub-Gal80ts/+. (A) ���� ����maintained continually at the restrictive temperature of tub-Gal80ts (31°C) �� ����� '��> �������� ����� ����� ��������� � ���� ����� ��������� w��'z������������������(�������������������������������!"#$������������?FH����������(B)���������������Q���������������+���+�����������������������������������������������������'z�(����������������������"#���������������������?FH����������(C)����������������������������������������������������'��Q<ts (18°C) to block Gal4 activity until being dissected 7 days after eclosion. w����������'z�����������������~������������������<���
31°C 7 days
dissecteclosion
3 days18°C
31°C
eclosiondissect
c309/UAS-mGFP;tubGal80ts/+
�
�
�
�
�
�
A
B
18°C 7 days
dissecteclosion
C
c309/UAS-mGFP;tubGal80ts/+
c309/UAS-mGFP;tubGal80ts/+

SLEEP, Vol. 36, No. 12, 2013 1821C Cholinergic MB Neurons Regulate Sleep—Yi et al
:�!��� �\—Temporal expression of mc* in either all c309 neurons or c309 non-mushroom body (MB) neurons decreases sleep. (A) Change in sleep amount (calculated by subtracting sleep before heat induction from sleep after heat-induction) in c309-Gal4/UAS-mc*;tub-Gal80tsF� ���or in c309-Gal4,MB-Gal80/UAS-mc*; tub-Gal80tsF� ��� ��� ������������������������������� ��� �����O������ ��������� ���� ���� �� ����� ���������������� �+<=�'��>F� ���� �+<=�'��>�"#�'��Q<F� ��� �� K\~����F�%tub-Gal80tsF� ���� |��� ��� �����`�� �� ������� �������� �� ������������z�<�<<�%������������O���������������z�<�<��(B) Temporal expression of mc* in either all c309 neurons or c309 non-MB neurons decreases sleep bout duration. Paired Student t-tests; ***P < 0.001. Data are presented as means ± standard error of the mean. n = 30-36 in each group.
0
20
40
60
Aver
age
dura
tion
of s
leep
bou
ts (m
in)
******
A
B pre-heat recordpost-heat record
c309
/UAS-m
c*;
tubGal8
0ts /+
c309
,MBGal8
0/
UAS-mc*;
tubGal8
0ts /+
c309
,MBGal8
0/+
c309
/+
UAS-mc*/
+;
tubGal8
0ts /+
-750
-600
-450
-300
-150
0
C
hang
e in
sle
ep a
mou
nt (m
in)
n.s.***
***
c309
/UAS-m
c*;
tubGal8
0ts /+
c309
,MBGal8
0/
UAS-mc*;
tubGal8
0ts /+
c30
9,
MBGal80/+
c309
/+UAS-m
c*/+;
tubGal8
0ts /+
***
*** :�!��� �^—Modulating Go activity in c309 neurons decreases the ������ �������� �� ���� w������� �(�������� �� ��������� ��(�� !zw�$or GoGTP ���� �+<=�'��> �� �+<=�'��>�"#�'��Q< ������� �����O������������� �� ������ �������� !��� ����� ������� �������� ������� �� �������������� ����$�{��������������������������� ��������������activity for 2 light-dark days before or after heat induction, respectively. |�����������`����������~������t-tests. ***P < 0.001; n = 30-36 in each group. Data are presented as means ± standard error of the mean.
0
40
80
120
*********
GoGTP
Wak
ing
activ
ity
(m
m/m
in)
PTX c309
MB-Gal80
***
+-+-
--+-
+-++
--++
+---
-++-
-+++
-+--
pre-heat recordpost-heat record
tubGal80ts + - + - + + + +

SLEEP, Vol. 36, No. 12, 2013 1821D Cholinergic MB Neurons Regulate Sleep—Yi et al
:�!����_—Temporal expression of pertussis toxin (PTX) or GoGTP did not affect the global morphology of mushroom bodies (MBs) at the end of the postheat recording. (A) Procedure for heat induction and dissection for B through E. Expression of PTX or GoGTP��������������}������� ��������������������at about 60 h after heat shock. =-57?V����������������]��������������� �����������������������'����������������������������������������(F) Procedure for dissection for G and H�w�� ��������������Q���������zw���'�GTP expression before dissection. (G and H) Representative projection ��������� �����������������������'����������������������������������������n��<�������������������~������������������<���
L18°C
31°C
elcosion adaption4-6 days 24-36hr D L D L D L D L D D
dissect
c309/tubGal80ts;PTX/+ tubGal80ts/+;PTX/+
c309/GoGTP;tubGal80ts/+ GoGTP/+;tubGal80ts/+
L18°C
elcosion adaption4-6 days 24-36hr D L D L D L D L D D
dissect
c309/tubGal80ts;PTX/+ c309/GoGTP;tubGal80ts/+
A
B C
D E
F
G H

SLEEP, Vol. 36, No. 12, 2013 1821E Cholinergic MB Neurons Regulate Sleep—Yi et al
:�!����W—Temporal expression of pertussis toxin (PTX) or GoGTP did not affect the general morphology of mushroom bodies (MBs) at the 20th day after heat induction. (A) Procedure for heat induction and dissection for B through F. PTX or GoGTP�������������(���������}������� ��������������������20 days after heat shock. =-5:?V����������������]��������������� �����������������������'����������������������������������������n = 10-15 for �������������~������������������<���
L18°C
31°C
elcosion adaption4-6 days 24-36hr 18 daysD L D L D L D L D
dissect
c309/tubGal80ts;PTX/+ tubGal80ts/+;PTX/+
c309/GoGTP;tubGal80ts/+ GoGTP/+;tubGal80ts/+
c309/+
A
B C
D E
F
SLEEP, Vol. 36, No. 12, 2013 1821F Cholinergic MB Neurons Regulate Sleep—Yi et al
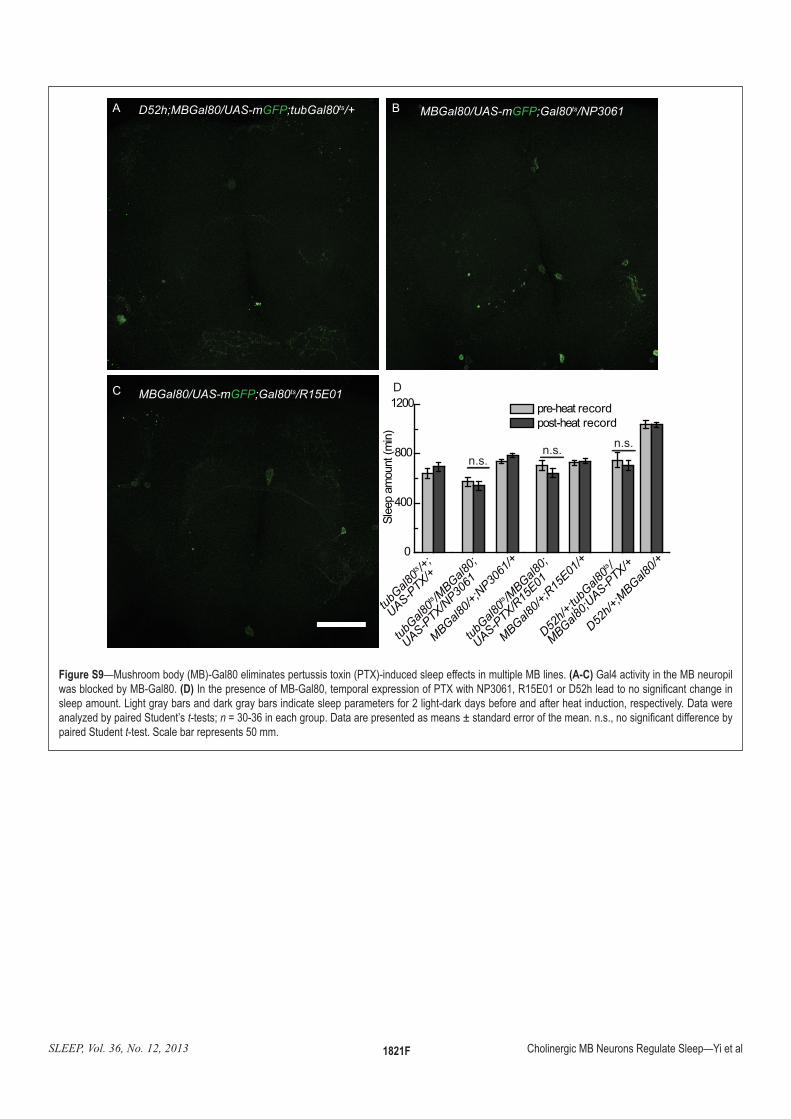
:�!����4—Mushroom body (MB)-Gal80 eliminates pertussis toxin (PTX)-induced sleep effects in multiple MB lines. =)5$? Gal4 activity in the MB neuropil ������������"#�'��Q<�(D)���������������"#�'��Q<����������(����������zw�����[z+<���V���<���|�}��������������O������������������������{�����������������������������������������������������}�������������������������������������������������������|������������`����������~��������t-tests; n�+<�+�������������|����������������������������������������������������������O����������������paired Student t-test. Scale bar represents 50 mm.
D52h;MBGal80/UAS-mGFP;tubGal80ts/+
0
400
800
1200 pre-heat record post-heat record
Slee
p am
ount
(min
) n.s.
n.s. n.s.
A B
C D
MBGal80/UAS-mGFP;Gal80ts/NP3061
MBGal80/UAS-mGFP;Gal80ts/R15E01
tubGal8
0ts /+;
UAS-PTX/+
MBGal80/+
;NP30
61/+
tubGal8
0ts /M
BGal80;
UAS-PTX/N
P3061
MBGal80/+
;R15
E01/+
tubGal8
0ts /M
BGal80;
UAS-PTX/R
15E01
D52h/+
;MBGal8
0/+
D52h/+
;tubG
al80ts /
MBGal80;U
AS-PTX/+

SLEEP, Vol. 36, No. 12, 2013 1821G Cholinergic MB Neurons Regulate Sleep—Yi et al
:�!���������������������������+<=�"#����������������������Continuously expressing chatRNAi by c309-Gal4 leads to reduced sleep �������������������������"#�'��Q<�w���������������������������������� ��� ���������������z�<�<<� �������������������variance. Data are presented as means ± standard error of the mean.
22 34 28 32 230
300
600
900
1200
***
***
Slee
p am
ount
(min
)
***
c309
/+;ch
atRNAi /+
c309
/+
c309
,MBGal8
0
/+;ch
atRNAi /+;
chat
RNAi /+
c309
,MBGal8
0/+
:�!������—Continuous expression of vAchTRNAi or chat-RNAi in c309 or D52h neurons did not affect the general morphology of MBs. =)5:? Representative ���]��������������� �����������������������'����������������������������������������n��<�������������������~������������������<���
c309/+;vAChTRNAi/+ c309/+
A B C
D E F
vAChTRNAi/+
c309/+;chatRNAi/+ chatRNAi/+ D52h/+;;vAchTRNAi/+
SLEEP, Vol. 36, No. 12, 2013 1821H Cholinergic MB Neurons Regulate Sleep—Yi et al
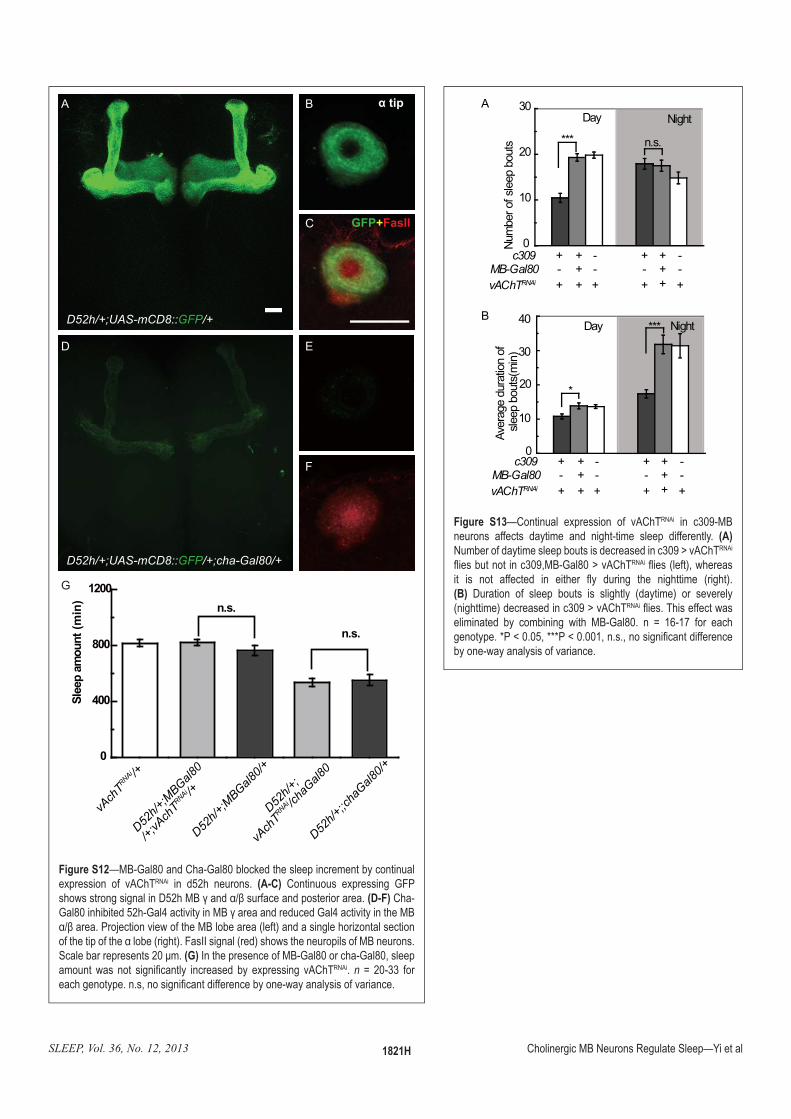
:�!������—MB-Gal80 and Cha-Gal80 blocked the sleep increment by continual expression of vAChTRNAi in d52h neurons. =)5$? Continuous expressing GFP �������������������|�}�"#����?FH������������������������=�5:? Cha-'��Q<����������}��'��>����������"#���������������'��>�������������"#?FH�����z��]���������������"#��������!����$��������������`�������������������������?����!�����$�����������!���$�������������������"#��������~�����������������}<���(G) In the presence of MB-Gal80 or cha-Gal80, sleep ������ ��� ��� �����O������ ��������� �� �(�������� �\��wRNAi. n = 20-33 for ������������������������O������������������������������������������
D52h/+;UAS-mCD8::GFP/+
GFP+FasII
� tip
D52h/+;UAS-mCD8::GFP/+;cha-Gal80/+
0
400
800
1200
Sl
eep
amou
nt (m
in) n.s.
n.s.
A B
C
D E
F
G
D52h/+
;MBGal8
0
/+;vA
chTRNAi /+
vAch
TRNAi /+
D52h/+
;MBGal8
0/+
D52
h/+;
vAch
TRNAi /ch
aGal8
0
D52h/+
;;cha
Gal80/+
:�!��� ���—Continual expression of vAChTRNAi in c309-MB neurons affects daytime and night-time sleep differently. (A) [��������������������������������������+<=��\��wRNAi ��������� ���+<=�"#�'��Q<��\��wRNAi ���!����$���������� �� ��� �������� �� ������ � ������ ��� ��������� !�����$�(B) Duration of sleep bouts is slightly (daytime) or severely !���������$������������+<=��\��wRNAi ����w���������������������� �� ��������� ���� "#�'��Q<� � � ����� ��� ��������������z�<�<�����z�<�<<��������������O������������������������������������������
0
10
20
30
n.s.
Night
vAChTRNAi MB-Gal80
Num
ber o
f sle
ep b
outs
c309
***
Day
+-+
+++
--+
+-+
+++
--+
0
10
20
30
40 Night
*
Av
erag
e du
ratio
n of
sle
ep b
outs
(min
)
***Day
vAChTRNAi MB-Gal80
c309 +-+
+++
--+
+-+
+++
--+
A
B