patterns of neuronal differentiation in neural tube mutant mice: curly tail and pax3 splotch-delayed
TRANSCRIPT

THE JOURNAL OF COMPARATIVE NEUROLOGY 368:516-526 (1996)
Patterns of Neuronal Differentiation in Neural Tube Mutant Mice:
Curly Tail and Pax3 Splotch-Delayed
CYNTHIA R. KELLER-PECK AND RICHARD J. MULLEN Department of Neurobiology and Anatomy, University of Utah School of Medicine,
Salt Lake City, Utah 84132
ABSTRACT A battery of antibodies was used to assess development of the spinal cord and its neurons in
mouse embryos with neural tube defects (NTDs). The two mutant strains examined, curly tail (ct) and splotch-delayed ( P U X ~ ~ P - ~ ) , develop an open neural tube for unrelated reasons, and thus provided for a complementary analysis. Five percent of embryos homozygous for the ct gene and 89% of embryos homozygous for the P U X ~ ~ P - ~ gene develop spina bifida in the lumbosacral region of the neuraxis. Expression of several neuronal antigens, including Islet-li2, polysialylated neural cell adhesion molecule (NCAM), neurofilaments, and a neuronal-specific nuclear protein (NeuN) recognized by monoclonal antibody A60, were used as indicators of the level of differentiation of neuronal tissue. Immunohistochemical labeling suggests that early (embryonic days 12-15) neuronal differentiation in the dorsal and ventral region of the dysraphic neural tube o a r s remarkably normally in both of the mutants. Similarly, labeling with antibodies to NCAM and neuroafdaments indicate that axonal development during early neurogenesis is unperturbed. Later stages of neuronal maturation, however, do not occur in the usual manner. Instead, the neuronal tissue begins a prodigious degeneration at embryonic day 17 (E17), so that by E l 8 only a rudimentary tissue remains. These results suggest that the aberrant morphology of the neural tube does not affect neuronal differentiation. However, the anomalous morphological and chemical environment may contribute to the neuronal degeneration observed at later stages. G 1996 Wiley-Liss, Inc.
Indexing terms: spina bifida cystica, monoclonal antibodies, neural tube defects, spinal cord, spinal dysraphism
The differentiation of cells in the central nervous system involves an ongoing process of proliferation, migration, and maturation. These events in the normal mammalian ner- vous system have been exceptionally well documented (Nornes and Carry, 1978; McConnell, 1981; Altman and Bayer, 1984); much less is known about these events in neural tube mutants. In general, the process of maturation in the spinal cord follows a ventrodorsal and rostrocaudal temporal gradient (Nornes and Carry, 1978; McConnell 1981; Altman and Bayer, 1984). In the mouse, the neural tube is just beginning to close at embryonic day 8 (ES), and consists of radially oriented undifferentiated columnar epithelial cells (Sauer, 1935; Sidman et al., 1959). By embryonic day 9, motoneurons, interneurons, and dorsal root ganglion cells are beginning to differentiate in the cervical spinal cord (Wentworth, 1984a,b). The posterior neuropore closes at E 10, completing neurulation, and thora- columbar motoneurons begin to form shortly thereafter. As a result of this process, neuronal differentiation, particu- larly in the dorsal region of the lumbosacral spinal cord,
occurs after neural tube closure is complete. In this paper, we examine this maturation process in two neural tube mutants, curly tail and splotch-delayed, to determine if neural tube closure is essential for neuronal differentiation.
The phenotype of the curly tail (c t ) mutant, first de- scribed by Gruneburg (19541, is characterized as an autoso- ma1 recessive trait with incomplete penetrance. Manifesta- tion of the c t l c t genotype is variable, with 40% of embryos exhibiting neural tube defects (NTDs), and the remaining 60% being phenotypically normal. Of the 40% with NTDs, 5% have lumbosacral myeloschisis, and the rest have curly or kinked tails. The obvious multifactorial inheritance of NTDs in ct make it one of the best animal models of the human condition (Embury et al., 1979; Neumann et al., 1994).
Accepted September 18, 1995. Address reprint requests to Dr. Richard J Mullen, 421 Wintrobe Bldg.,
Dept. Neurobiology & Anatomy, Univ. of Utah School of Medicine, Salt Lake City, UT 84132. E-mail:mullen@ msscc.med.utah.edu
8 1996 WILEY-LISS, INC.

NEURON DIFFERENTIATION IN NEURAL TUBE MUTANTS 517
The exact cause of NTDs in curly tail is unknown. The gene has been localized to distal chromosome 4 in mice (Neumann et al., 19941, although it has not been cloned. A number of experiments have been aimed at determining the morphological and cellular abnormalities leading to caudal NTDs in these mutants. I t is currently thought that the gross abnormality leading to tail defects and lumbosacral myeloschisis in curly tail mutants is a delayed closure of the posterior neuropore (PNP; Copp et al., 1982; Copp, 1985; van Straaten et al., 1992) which arises initially as a defect in primary neurulation (Copp and Brook, 1989). The underly- ing cause of the delayed closure appears to be a reduced rate of cell proliferation in gut endoderm and notochord, while the overlying neuroepithelium grows normally (Copp et al., 1988). The result of these abnormal cell proliferation rates is an enhanced ventral curvature of the embryo (Brook et al., 1991). The increased curvature places a mechanical stress on the neural folds, and they fail to fuse over the dorsal midline. Experimental prevention of ventral curva- ture in ctlct embryos by either separating the caudal neuroepithelium from all adjacent non-neuronal tissues and culturing in vitro (van Straaten et al., 1993), or by insertion of an eyelash tip longitudinally into the hindgut lumen to prevent curvature (Brook et al., 1991) alleviates the delay in PNP closure.
The splotch mutant was first described in 1947 (Russell and Roscoe, 1947). There are several allelic variants of the initial mutation; only one of which, splotch-delayed ( P a 3 ~ 3 ~ p - ~ ; Dickie, 1964) was used in these experiments. The mutation is classified as semidominant lethal; the colony is maintained by heterozygous brother-sister mating. Homo- zygous splotch-delayed embryos develop posterior NTDs and manifest defects in neural crest derived structures (Moase and Trasler, 1989). Heterozygous animals exhibit only pigmentary defects of the tail, belly, and feet.
In contrast to the curly tail mutant, the gene for the splotch-delayed mutant has been cloned (Vogan et al., 1993). It is located on the proximal portion of chromosome 1 and encodes the DNA binding protein Pax-3. I t is for this reason that the gene symbol was recently changed from Spd to P U X ~ ~ P - ~ (Peters et al., 1995). The gene is expressed during early neurogenesis in the ventricular zone of the spinal cord, particularly in the alar plate region, including the roof plate and neural crest cells. It is currently un- known how the expression of the Pax-3 protein product is related to the cascade of events resulting in normal neurula- tion. The experiments of Moase and Trasler (1991) and of Trasler and Morriss-Kay (1991) suggest that Pax-3 may be regulating extracellular matrix molecules and cell adhesion molecules, and that this might be the biochemical basis for the observed morphological aberrancies.
Although the gross abnormality produced by the curly tail or splotch-delayed gene is similar, the underlying mechanisms are not. For example, an increased ventral curvature is thought to be the primary cause of NTDs in curly tail (Brook et al., 19911, while splotch mutants have a decreased ventral curvature compared to controls (Estibi- ero et al., 1993). That different morphological characteris- tics exist between splotch and curly tail mutants is to be expected, since the splotch gene appears to affect neuroepi- thelium and the curly tail gene presumably affects ventral structures. The use of these etiologically distinct mutants in this study allowed us to ascertain whether differentiation and observed neuropathological changes of neuronal tissue are related to the gene involved, or are the result of the
abnormal morphology and environment of the open neural tube.
Using the ct and Pa3~3~P-~mutants as a model, we have analyzed neuronal differentiation in affected regions of the spinal cord. We have found that neurogenesis proceeds relatively normally in the dysraphic neural tube through E16. Despite the abnormal proliferation of notochord cells in ct mutants reported by Copp et al. (1988), the floor plate is clearly evident, and commissural axons cross the ventral midline as expected. Similarly, both dorsal and ventral roots and the dorsal root ganglion (DRG) are well-formed prior to E l 6 in ct mutants, though they are displaced dorsally. However, the roots and DRG are small or absent from the affected region of P C L X ~ ~ P - ~ embryos, most likely due to involvement of the neural crest in these mutants. In both mutants, neurons within the NTD have begun to degenerate at E17, so that by E l 8 only a rudimentary tissue remains. These observations suggest that neural matura- tion within the open neural tube proceeds relatively nor- mally through E16. Thereafter, however, a rapid deteriora- tion of the spinal cord and rootlets occurs. A portion of these results has been published in abstract form (Keller- Peck and Mullen, 1993).
MATERIALS AND METHODS Animals and tissue preparation
Both the curly tail (c t lc t stock) and splotch-delayed ( C ~ ~ B L I G J - P U X ~ ~ P - ~ ) strains were obtained from The Jack- son Laboratory (Bar Harbor, ME), and are currently main- tained in our animal facility. The animal facility complies with NIH and USDA guidelines. The research was ap- proved by the IACUC. Animals are given food and water ad libitum and maintained on a 12 hour light/dark cycle, with the lights on at 7 A.M. and off at 7 P.M. Females homozygous for the ct gene (phenotypically either curly-tailed or straight- tailed) were mated with homozygous males (with a pheno- typically curled or kinked tail) in the late afternoon and checked for vaginal plugs the following three mornings. Heterozygous P ~ X ~ ~ P - ~ females were mated with heterozy- gous males in a similar manner. The day of finding a plug was considered day 0 (EO) of gestation.
Pregnant females were euthanized by overexposure to C02 and embryos recovered at embryonic days E l 2 to E18. The whole uterus was removed and the embryos were dissected out and examined for spina bifida. Embryos with spina bifida, and at least two phenotypically normal litter- mate controls were decapitated and immersed in fixative overnight.
Histology Embryos to be examined with antibodies A60,5A5,2H3,
and 3A10 and 40.2D6 were fixed with paraformaldehyde- lysine-periodate (PLP) fixative (McLean and Nakane, 1974). The following day they were rinsed a minimum of 10 hours in phosphate-buffered sucrose (5%), then dehydrated and embedded for either transverse or sagittal sections in polyester wax (Polyethylene glycol 400 distearate, Ruger Chemical, Irvington, NJ). The wax had been prepared as detailed by Feder (1976).
Serial sections were cut at 8 km through the entire region of spina bifida (which ranged from 1-5 mm in length depending on age and severity of the defect) and through control embryos of the same axial level. For the antibodies A60, 5A5,2H3, and 3A10, lumbar cord was compared with

518 C.R. KELLER-PECK AND RJ. MULLEN
lumbar cord within 2-3 segments. However, for the anti- body 40.2D6 mutant lumbar cord was compared with normal cord at the same segmental level. Sections were mounted on slides subbed in gelatin with chromium potas- sium sulfate. After air-drying, the sections were rehydrated and processed for immunohistochemistry. The average slide had approximately five sections.
We examined 3 E12, 3 E13, 4 E14, 2 E15, 4 E16, 7 E17, and 5 E l 8 ct /c t embryos with spina bifida in transverse section and 2 E14, l E15,3 E16,5 E17, and 3 E l 8 embryos with spina bifida in sagittal section. We examined 3 E12, 2 E13, l E14,2 E16,5 E17, and 4 E l 8 Pax3sP-d embryos with spina bifida in transverse section, and 3 E l 3 embryos with spina bifida in sagittal section. Approximately every fifth slide from every transversely sectioned embryo was stained with mAb A60. Roughly every fourth slide of E l 2 and E l 3 embryos in transverse section were labeled with either mAb 40.2D6 or 5A5. All sagittal sections were stained with the neurofilament antibodies 3A10 and 2H3; a few transverse sections were also stained with these antibodies. Since all of these embryos were collected around midday, the actual developmental ages might be more accurately described as E12.5, E13.5, etc. For simplicity we will refer to them as E12, E l 3 etc.
Antibodies The following monoclonal antibodies (mAb) were used on
polyester wax sections. Monoclonal antibody A60, a mouse IgG produced in our laboratory, identifies NeuN, a neuron- specific nuclear protein (Mullen et al., 1992). 5A5, a mouse IgM labels a polysialylated form of neural cell adhesion molecule (PSA-NCAM; Dodd et al., 1988). 2H3, a mouse IgG, recognizes a 155 kd neurofilament subunit (Dodd et al., 1988). 3A10, a mouse IgG, identifies a neurofilament-associated protein. Fi- nally, 40.2D6, a mouse I*, binds to the C-terminal portion of the Islet-1 protein (Ericson et al., 1992). However, this antibody may also recognize the Islet-2 protein (Tsuchida et al., 1994) and thus the antigen will be referred to as Islet-l/2 in this paper. The Islet-l/2 proteins are expressed in a subset of neurons, including motor neurons and dorsal root ganglion cells. 5A5, 2H3, 3A10, and 40.2D6 were obtained from the Developmental Studies Hybridoma Bank (Iowa City, IA).
Immunohistochemistry Monoclonal antibody A60 from ascites was biotinylated
according to manufacturer's directions (biotin; Vector Labs, Burlingame, CA) and used at a dilution of 1:1,500. Hybri- doma supernatants for antibodies 5A5, 2H3, and 3A10 and 40.2D6 were not biotinylated, and were diluted 1 : l O unless otherwise indicated. Sections were blocked in 3% normal serum (horse or goat depending on secondary antibody) and 0.1% HzOz for 30 minutes. Slides were then incubated with primary antibodies diluted in Tris-buffered saline (TBS) and 3% normal serum at 4°C overnight. The following day sections were rinsed in TBS and incubated with a 1:150 dilution of biotinylated secondary antibody (Vector Labs) for one hour at room temperature. The exception was the biotinylated A60 slides, which were rinsed and directly incubated with horseradish peroxidase (HRP)-labeled strep- tavidin from the Vecta ABC Elite Kit (Vector Labs) for 30 minutes. Slides labeled with secondary antibodies were sub- sequently incubated with HRP-labeled streptavidin. Pri- mary antibody location was visualized with 3-3'-diaminoben- zidine as the HRP substrate. Slides were then rinsed, dehydrated through graded alcohols, and coverslipped.
Photography Photographs were taken on a Nikon Diaphot inverted
microscope with Kodak black-and-white Technical Pan film. Negatives were electronically scanned onto a Power Macintosh with a Leafscan 45 digital camera. Images were adjusted for brightness and contrast using the Adobe Photoshop program, Additionally, a uniform gray back- ground was added around tissue sections for aesthetic purposes. No other alterations to the images were per- formed. Images were printed with a Fujix Pictrography 3000.
RESULTS Neuronal differentiation during early
(E12-El5) development Serial transverse sections through the length of the open
neural tube were examined immunohistochemically to as- sess the level of differentiation of neuronal tissue. Sections from both ct and P U X ~ ~ P - ~ homozygous embryos were then compared to control sections taken from the same axial level. We examined spinal cord maturation using several criteria. First, the cells should express the neuron-specific antigen of mAb A60, a general indicator of neuronal differentiation that is expressed shortly after the final mitotic division (Mullen et al., 1992). Second, there should be present a subset of cells in the ventral horn, motor neurons, that express Islet-li2. Third, commissural axons should be able to reach the ventral midline as indicated by the antibody 5A5 to the polysialylated form of NCAM. Finally, there should be demonstrable formation of spinal roots, indicated by neurofilament-specific antibodies 2H3 and 3A10. The combined expression of these antigens suggests a high level of differentiation of the spinal cord and its neurons.
The E l 2 normal spinal cord appears essentially the same as an adult cord despite the presence of a large central canal. Control sections stained with mAb A60 reveal numer- ous post-mitotic neurons throughout the basal plate and a moderate amount of these cells in the alar plate (Fig. lA,E). Furthermore, a thin outer layer of white matter has begun to develop. In embryos with spina bifida, the spinal cord is everted, producing a characteristic wing-like appearance. Despite this gross morphological insult, cells within the open neural tube of E l 2 ct and P U X ~ ~ P - ~ embryos stain with mAb A60 uniformly throughout the basal plate (Fig. lB,F). At the magnification shown in Figure 1, differentiation of the alar plate appears retarded in the open neural tube, but at higher magnification the number of A60 positive cells appears comparable to controls. At this stage the ventricu- lar zone and floor plate are clearly evident. Neurogenesis and histogenesis appear to have initiated quite normally within the dysraphic neural tube.
At E l 4 the number of A60 positive cells in the spinal cord of control embryos continues to increase, particularly in the dorsal region (Fig. lC,G). The ventral and lateral funiculi are also well-defined at this stage. This trend is similar in embryos with spina bifida. In affected embryos from the curly tail strain (Fig. 1D) the dorsal root ganglia are distinct and the cells are immunopositive for the antigen recognized by A60. In the splotch-delayed strain (Fig. lH), the dorsal root ganglia are often absent or severely reduced in size. However, those cells that are present within the DRG also express the A60 antigen.
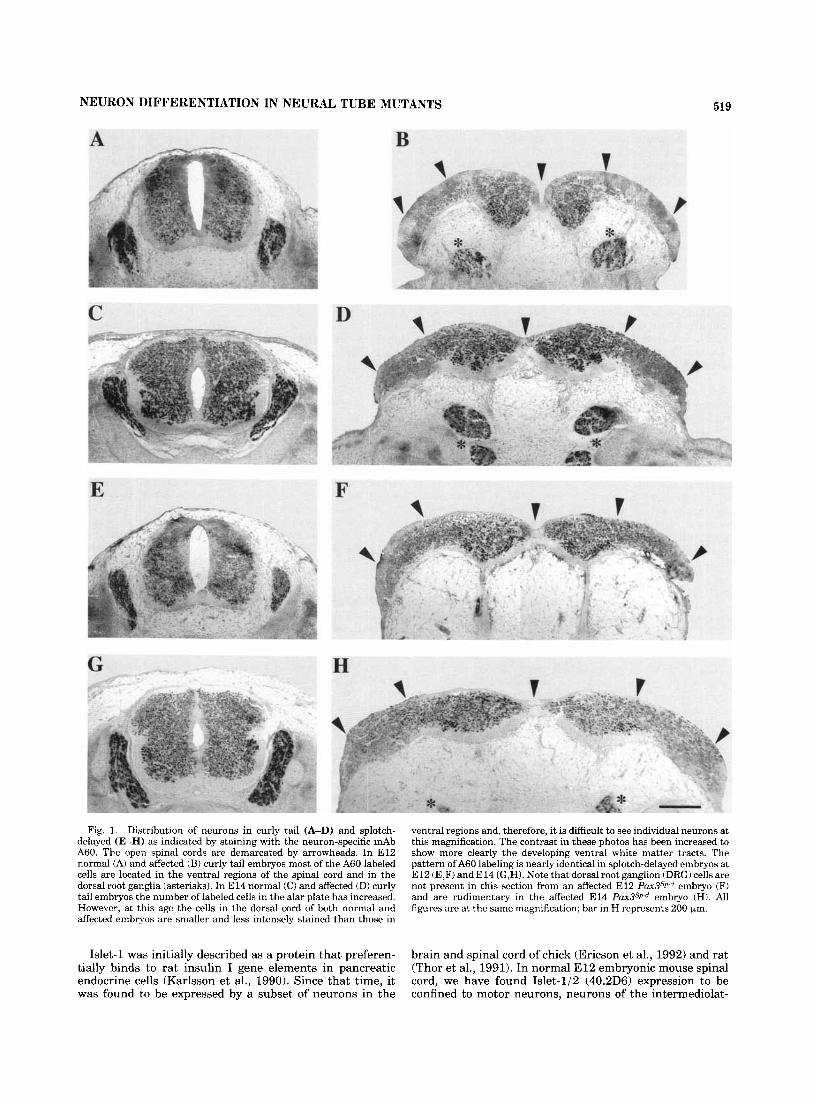
NEURON DIFFERENTIATION IN NEURAL TUBE MUTANTS 519
Fig. 1. Distribution of neurons in curly tail (A-D) and splotch- delayed (E-H) as indicated by staining with the neuron-specific mAb A60. The open spinal cords are demarcated by arrowheads. In E l 2 normal (A) and affected (B) curly tail embryos most of the A60 labeled cells are located in the ventral regions of the spinal cord and in the dorsal root ganglia (asterisks). In E l 4 normal (C) and affected (D) curly tail embryos the number of labeled cells in the alar plate has increased. However, at this age the cells in the dorsal cord of both normal and affected embryos are smaller and less intensely stained than those in
ventral regions and, therefore, it is difficult to see individual neurons at this magnification. The contrast in these photos has been increased to show more clearly the developing ventral white matter tracts. The pattern of A60 labeling is nearly identical in splotch-delayed embryos at E l 2 (E,F) and E l 4 (G,H). Note that dorsal root ganglion (DRG) cells are not present in this section from an affected E l 2 P U X ~ ~ P - ~ embryo (F) and are rudimentary in the affected E l 4 P U X ~ ~ P - ~ embryo (H). All figures are at the same magnification; bar in H represents 200 km.
Islet-1 was initially described as a protein that preferen- tially binds to rat insulin I gene elements in pancreatic endocrine cells (Karlsson et al., 1990). Since that time, it was found to be expressed by a subset of neurons in the
brain and spinal cord of chick (Ericson et al., 1992) and rat (Thor et al., 1991). In normal E l 2 embryonic mouse spinal cord, we have found Islet-1/2 (40.2D6) expression to be confined to motor neurons, neurons of the intermediolat-

520 C.R. KELLER-PECK AND R.J. MULLEN
eral nucleus and occasional cells of the alar plate. It is also expressed within the dorsal root ganglion. Thus, it appears to be a useful marker to identify subtypes of neurons present within the spinal cord of embryos with neural tube defects .
No conspicuous alterations in Islet-l/Z expression were observed between E l 2 embryos with NTDs and their normal littermate controls (Fig. 2A-D). The pattern of Islet-l/2 positive cells in the ventral and intermediate horn of affected embryos appeared to repeat the pattern observed in ct and P U X ~ ~ P - ~ normal embryos. Moreover, this trend continued through E13. However, in embryos several days older we were unable to detect any Islet-l/2 positive cells in the spinal cord of normal or affected embryos, although immunopositive cells were still present in the dorsal root ganglia (not shown).
Commissural axon trajectory in embryos with spina bifida. We next examined whether commissural axons reach their ventral midline target in E l 2 embryos with spina bifida. These axons were labeled using mAb 5A5 to a polysialylated form of NCAM (PSA-NCAM), which is ex- pressed by motor neurons and the commissural axons that cross the ventral floor plate. In normal embryos, the cell bodies of commissural neurons differentiate in the dorsal spinal cord and extend their axons ventrally (Fig. 2E,G). These axons initially project in the transverse plane until they cross the floor plate. After this point, they turn into the longitudinal plane and project rostrally in the contralat- eral ventral funiculus (Holley, 1982; Wentworth, 1984b; Bovolenta and Dodd, 1990). The ventral trajectory of commissural axons in E l 2 embryos with an open neural tube occurs with remarkable fidelity; a dense band of stained axons can be seen running beneath the floor plate (Fig. 2F,H). The sections shown were examined at a higher magnification to confirm that the axons were running transversely beneath the floor plate (Fig. 2H inset). The presence of the ventral funiculus in affected embryos attests to the fact that axonal processes appear to be developing normally. However, we have not determined whether rostral connections have been made appropriately.
Dorsal and ventral root development in embryos with spina bifida. Serial sections in both sagittal and trans- verse planes of embryos with spina bifida were analyzed immunohistochemically and histologically to assess develop- ment of the spinal nerves. Axons both entering and leaving the spinal cord contain neurofilaments that are recognized by the antibodies 2H3 and 3A10. The purpose of using these antibodies was twofold. First, they identify the spinal nerves, and second, positive expression of their antigen further indicates differentiation of ventral horn and DRG neurons.
In sagittal section of curly tail embryos, axons are clearly observed exiting the dysraphic neural tissue and traversing an enlarged subarachnoid space (Fig. 3A). The DRG are also suspended in the expanded subarachnoid space, sur- rounded by a loose meshwork of tissue (Fig. 3B). Both the DRG and the spinal nerves are pulled dorsally as the neural tissue rises above the epidermis. In E l 3 transverse section, it is evident that axons from the DRG have penetrated the dorsal horn despite the splayed-out morphology of the open neural tube (Fig. 3 0 . Moreover, the ventral roots exit the spinal cord, form distinct nerves and head toward the periphery. By E l 5 however, the nerves begin to fragment and degenerate. By E17, neither dorsal nor ventral roots can be detected (not shown).
In contrast to the curly tail mutants, we were unable to detect dorsal root axons in affected splotch-delayed embryos
even at early stages of development. Presumably, this is because of the decreased migration of neural crest cells and hence rudimentary DRGs present in these embryos. How- ever, ventral roots did exit the affected spinal cord (Fig. 3D), as in the curly tail embryos.
Neuronal differentiation during late (E16-El8) development
At slightly later stages of development, E16, the number of A60 positive cells both in the normal (Fig. 4A,E) and the dysraphic neural tissue (Fig. 4B,F) of ct and P U X ~ ~ P - ~ embryos continues to increase. In the normal embryo, the dorsal, ventral, and lateral funiculi are well developed. In contrast, the dorsal funiculus and the dorsal portion of the lateral funiculus have not developed in spina bifida em- bryos, as evidenced by the A60 positive cells continuing to the lateral edge of the dorsal neural tube. Thus, during early development, the fact that the neural folds have failed to fuse does not appear to affect neuronal differentiation as judged by the expression of the A60 antigen. It does, however, severely affect the formation of dorsal white matter tracts.
Maturation of the open spinal cord does not continue after E16. Beginning at E17, the neural tissue starts to fragment and degenerate and the ventral funiculi disap- pear. Most of the cells within the affected region of the spinal cord maintain their expression of the A60 antigen, nonetheless, by E l 8 only rudimentary tissue remains (Fig. 4D,H). Some variability in the extent of degeneration of these embryos was observed, however, all embryos exam- ined had lost the majority of their neurons by E18. Al- though the DRG neurons also degenerate in curly tail spina bifida embryos, the rate appears to be substantially less than in the spinal cord. The abnormal morphology of the spinal cord combined with the abnormal environment is associated with a severe neuronal loss in late development.
In order to more fully describe the extent of degeneration of neurons in El8 c t embryos, an analysis of the spinal cord beginning 5 mm rostral to the lesion and continuing to the tail was under- taken. Sections greater than 4 mm rostral to the lesion reveal a spinal cord similar in morphology and dimension to control sections. Perhaps somewhat surprisingly, however, 2 mm rostral to the exposed neural tissue (Fig. 5A) the spinal cord completely lacks any organization and is roughly one quarter the diameter (one fifteenth the area) of a normal section. However, the DRGs rostral to the lesion appear relatively normal (not shown). It is interesting to note that the disorganized tissue observed slightly rostral to the exposed neural tissue is not found in embryos younger than E16. Figure 5B depicts the spinal cord just as it becomes exposed at the dorsal surface. The dimensions are severely reduced compared to controls and only a few A60 positive cells are evident. As sections continue caudally through the lesion (Fig. 5C,D) the neural tissue takes on the characteristic wing-like appearance, increases in area, and A60 labeling becomes more obvious. In the curly tail strain, a normal E l 8 spinal cord has an area of roughly 0.43 mm2 (Fig. 4C); the area of the abnormal cord in Figure 4D is approximately 0.044 mm2, indicating the enormous num- ber of cells lost within the last days of development.
Neurons degenerate in El8 embryos.
DISCUSSION Despite the decreasing incidence of spina bifida among
newborn infants in the United States, the Birth Defects
NEURON DIFFERENTIATION IN NEURAL TUBE MUTANTS 521
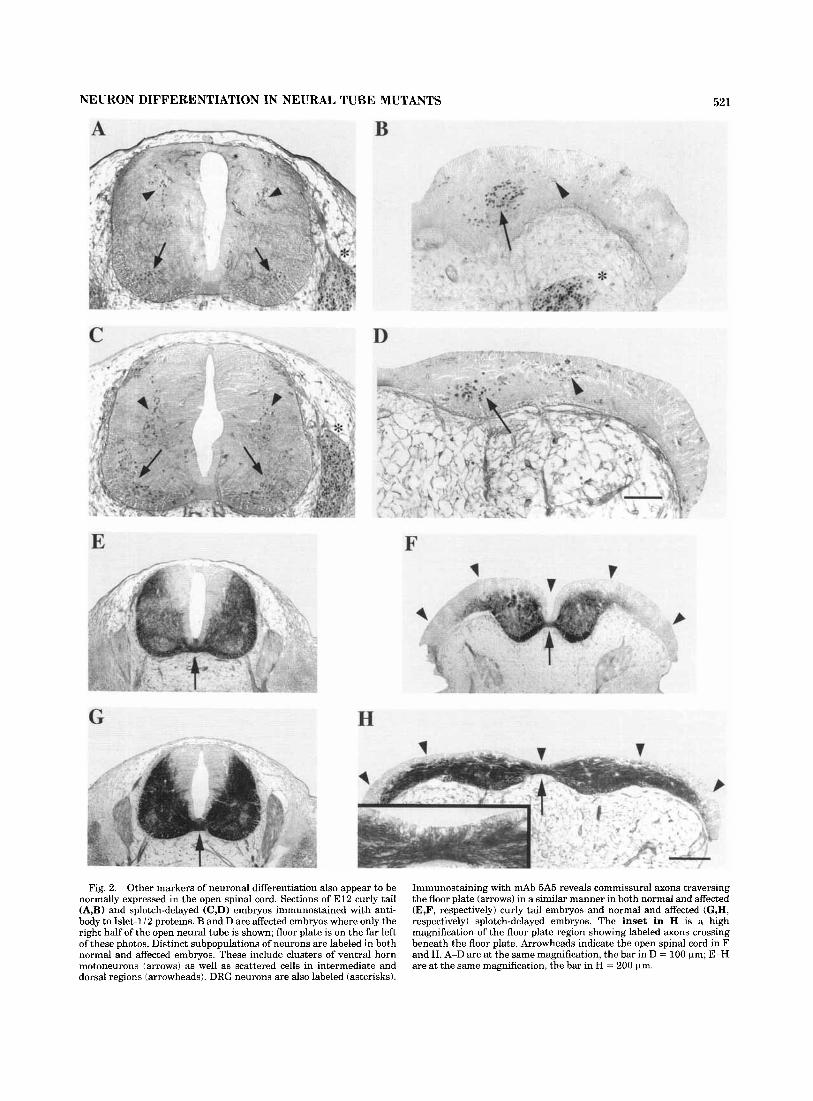
Fig. 2. Other markers of neuronal differentiation also appear to be normally expressed in the open spinal cord. Sections of El2 curly tail (A,B) and splotch-delayed (C,D) embryos immunostained with anti- body to Islet-li2 proteins. B and D are affected embryos where only the right half of the open neural tube is shown; floor plate is on the far left of these photos. Distinct subpopulations of neurons are labeled in both normal and affected embryos. These include clusters of ventral horn motoneurons (arrows) as well as scattered cells in intermediate and dorsal regions (arrowheads). DRG neurons are also labeled (asterisks).
Immunostaining with mAb 5A5 reveals commissural axons traversing the floor plate (arrows) in a similar manner in both normal and affected (E,F, respectively) curly tail embryos and normal and affected (G,H, respectively) splotch-delayed embryos. The inset in H is a high magnification of the floor plate region showing labeled axons crossing beneath the floor plate. Arrowheads indicate the open spinal cord in F and H. A-D are at the same magnification, the bar in D = 100 pm; E-H are at the same magnification, the bar in H = 200 pm.
522 C.R. KELLER-PECK AND R J . MULLEN
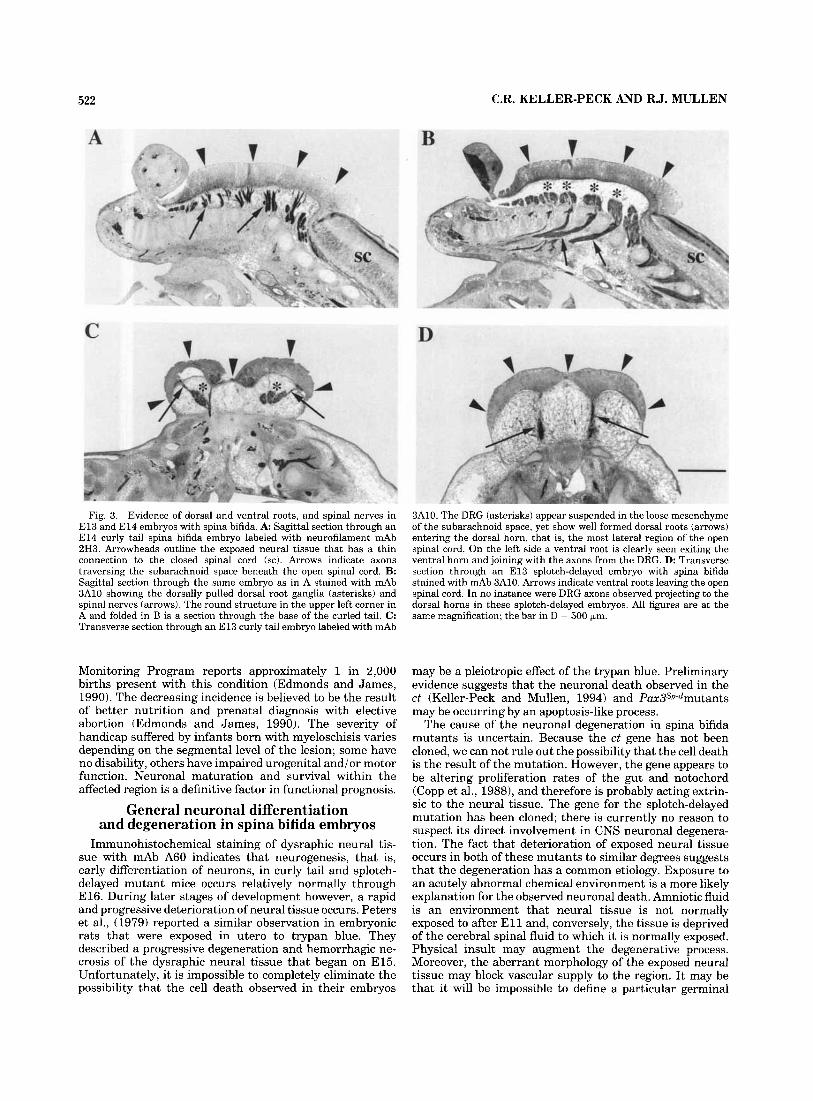
Fig. 3. Evidence of dorsal and ventral roots, and spinal nerves in E l 3 and E l 4 embryos with spina bifida. A: Sagittal section through an El4 curly tail spina bifida embryo labeled with neurofilament mAb 2H3. Arrowheads outline the exposed neural tissue that has a thin connection to the closed spinal cord (sc). Arrows indicate axons traversing the subarachnoid space beneath the open spinal cord. B: Sagittal section through the same embryo as in A stained with mAb 3A10 showing the dorsally pulled dorsal root ganglia (asterisks) and spinal nerves (arrows). The round structure in the upper left corner in A and folded in B is a section through the base of the curled tail. C : Transverse section through an E l 3 curly tail embryo labeled with mAb
Monitoring Program reports approximately 1 in 2,000 births present with this condition (Edmonds and James, 1990). The decreasing incidence is believed to be the result of better nutrition and prenatal diagnosis with elective abortion (Edmonds and James, 1990). The severity of handicap suffered by infants born with myeloschisis varies depending on the segmental level of the lesion; some have no disability, others have impaired urogenital and/or motor function. Neuronal maturation and survival within the affected region is a definitive factor in functional prognosis.
General neuronal differentiation and degeneration in spina bifida embryos
Immunohistochemical staining of dysraphic neural tis- sue with mAb A60 indicates that neurogenesis, that is, early differentiation of neurons, in curly tail and splotch- delayed mutant mice occurs relatively normally through E16. During later stages of development however, a rapid and progressive deterioration of neural tissue occurs. Peters et al., (1979) reported a similar observation in embryonic rats that were exposed in utero to trypan blue. They described a progressive degeneration and hemorrhagic ne- crosis of the dysraphic neural tissue that began on E l 5 Unfortunately, it is impossible to completely eliminate the possibility that the cell death observed in their embryos
3A10. The DRG (asterisks) appear suspended in the loose mesenchyme of the subarachnoid space, yet show well formed dorsal roots (arrows) entering the dorsal horn, that is, the most lateral region of the open spinal cord. On the left side a ventral root is clearly seen exiting the ventral horn and joining with the axons from the DRG. D: Transverse section through an E l 3 splotch-delayed embryo with spina bifida stained with mAb 3A10. Arrows indicate ventral roots leaving the open spinal cord. In no instance were DRG axons observed projecting to the dorsal horns in these splotch-delayed embryos. All figures are at the same magnification; the bar in D = 500 km.
may be a pleiotropic effect of the trypan blue. Preliminary evidence suggests that the neuronal death observed in the ct (Keller-Peck and Mullen, 1994) and Pax3sP-dmutants may be occurring by an apoptosis-like process.
The cause of the neuronal degeneration in spina bifida mutants is uncertain. Because the ct gene has not been cloned, we can not rule out the possibility that the cell death is the result of the mutation. However, the gene appears to be altering proliferation rates of the gut and notochord (Copp et al., 19881, and therefore is probably acting extrin- sic to the neural tissue. The gene for the splotch-delayed mutation has been cloned; there is currently no reason to suspect its direct involvement in CNS neuronal degenera- tion. The fact that deterioration of exposed neural tissue occurs in both of these mutants to similar degrees suggests that the degeneration has a common etiology. Exposure to an acutely abnormal chemical environment is a more likely explanation for the observed neuronal death. Amniotic fluid is an environment that neural tissue is not normally exposed to after E l l and, conversely, the tissue is deprived of the cerebral spinal fluid to which it is normally exposed. Physical insult may augment the degenerative process. Moreover, the aberrant morphology of the exposed neural tissue may block vascular supply to the region. It may be that it will be impossible to define a particular germinal
NEURON DIFFERENTIATION IN NEURAL TUBE MUTANTS 523
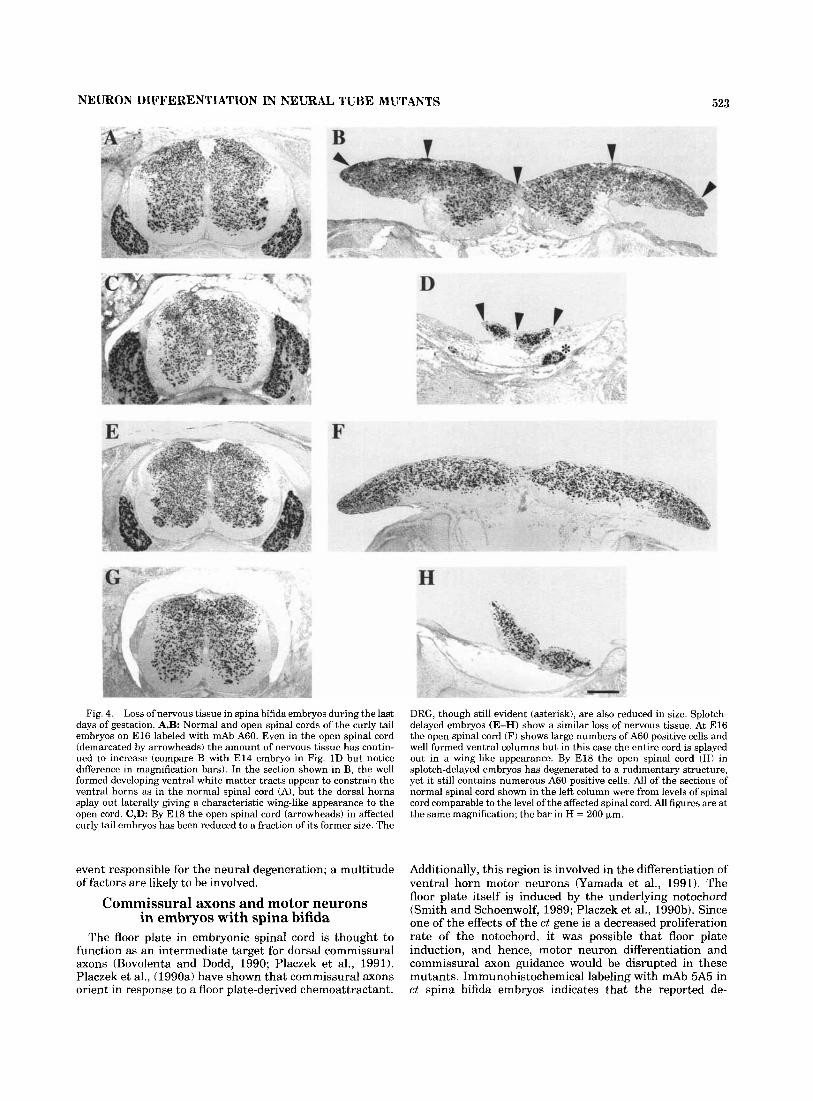
Fig. 4. Loss of nervous tissue in spina bifida embryos during the last days of gestation. A,B: Normal and open spinal cords of the curly tail embryos on E l 6 labeled with mAb A60. Even in the open spinal cord (demarcated by arrowheads) the amount of nervous tissue has contin- ued to increase (compare B with E l 4 embryo in Fig. 1D but notice difference in magnification bars). In the section shown in B, the well formed developing ventral white matter tracts appear to constrain the ventral horns as in the normal spinal cord (A), but the dorsal horns splay out laterally giving a characteristic wing-like appearance to the open cord. C,D: By E l 8 the open spinal cord (arrowheads) in affected curly tail embryos has been reduced to a fraction of its former size. The
event responsible for the neural degeneration; a multitude of factors are likely to be involved.
Commissural axons and motor neurons in embryos with spina bifida
The floor plate in embryonic spinal cord is thought to function as an intermediate target for dorsal commissural s o n s (Bovolenta and Dodd, 1990; Placzek et al., 1991). Placzek et al., (1990a) have shown that commissural s o n s orient in response to a floor plate-derived chemoattractant.
DRG, though still evident (asterisk), are also reduced in size. Splotch- delayed embryos (E-H) show a similar loss of nervous tissue. At E l 6 the open spinal cord (F) shows large numbers of A60 positive cells and well formed ventral columns but in this case the entire cord is splayed out in a wing-like appearance. By E l8 the open spinal cord (HI in splotch-delayed embryos has degenerated to a rudimentary structure, yet it still contains numerous A60 positive cells. All of the sections of normal spinal cord shown in the left column were from levels of spinal cord comparable to the level of the affected spinal cord. All figures are at the same magnification; the bar in H = 200 pm.
Additionally, this region is involved in the differentiation of ventral horn motor neurons (Yamada et al., 1991). The floor plate itself is induced by the underlying notochord (Smith and Schoenwolf, 1989; Placzek et al., 1990b). Since one of the effects of the ct gene is a decreased proliferation rate of the notochord, it was possible that floor plate induction, and hence, motor neuron differentiation and commissural axon guidance would be disrupted in these mutants. Immunohistochemical labeling with mAb 5A5 in ct spina bifida embryos indicates that the reported de-

524 C.R. KELLER-PECK AND R.J. MULLEN
creased proliferation of notochord cells does not affect commissural axon migration. This can also be taken as the first evidence that dorsal neural cell types are differentiat- ing normally in neural tube mutants.
Although there are no serious abnormalities in notochord or floor plate formation in splotch-delayed embryos, it has been documented that E9 embryos with NTDs exhibit abnormal post-translational processing of NCAM (Moase and Trader, 1991; Neale and Trader, 1994). In particular, the authors found that heterozygous and homozygous
C
D
splotch embryos express polysialylated NCAM at an abnor- mally early developmental stage. Variations in post- translational modification, especially with respect to polysi- alic acid content, could modify adhesiveness of axons and thus alter axonal trajectories. Immunohistochemical label- ing with mAb 5A5 to PSA-NCAM clearly demonstrates that dorsal commissural axons develop normally in affected Pa3~3sp-~ embryos. However, further studies are required to determine whether these axons are able to make their proper diencephalic connections.
The Islet-1/2 labeling pattern in both mutants is similar to that seen in controls. However, cautious interpretation is necessary when examining sections stained with Islet anti- bodies. Many of the previously described studies of Islet-1 expression were done using rabbit polyclonal antibodies (Ericson et al., 1992; Thor et al., 1991). In retrospect, these antibodies are probably recognizing both Islet-1 and Islet-2 (Tsuchida et al., 1994). The same is probably also true of the monoclonal antibody (40.2D6) used in our experiments. We have found that our ability to detect Islet-l/2 immuno- reactivity in the spinal cord of normal and affected embryos decreases significantly after E14. Nonetheless, staining in the dorsal root ganglion at this stage and later continues to be robust. This may indicate that 40.2D6 has a higher affinity for Islet-1 since this protein stops being expressed in the spinal cord before Islet-2 (Tsuchida et al., 1994).
The role of the roof plate in dorsoventral patterning
There are two main lines of evidence that suggest that specification of dorsal phenotype occurs normally in spina bifida mutants. First, dorsal horn neurons are competent to respond to signals that guide their axons toward the ventral floor plate. Second, the expression pattern of Islet-1 /2 remains restricted to primarily the ventral horn. These patterns indicate that the general organization of the mutant cord is, in all likelihood, normal. However, despite the normal general organization, there are indications that when the neural tube fails to close, roof plate features do not develop. For example, using sections from the same embryos employed in this study, Hamre et al. (1996) found that the roof plate marker annexin IV (Hamre et al., 1995) is not expressed in the dorsal region of embryos with spina bifida, and that the characteristic morphology of roof plate cells is also absent.
These results suggest that specification of dorsal pheno- type is not dependent on roof plate formation. This is in contrast to specification of ventral fate, which is directly dependent on floor plate formation (Placzek et al., 1991). There are several means by which dorsal fate could be
Fig. 5. Rostrocaudal analysis of an E l8 curly tail embryo with spina bifida laheled with mAb A60. A: Transverse section of spinal cord rostra1 to the region of exposed neural tissue. Although the spinal cord in this region appears to be closed, note the absence of well-defined gray and white matter and the paucity A60 labeled cells. Also, compare size of this spinal cord with those in Figure 4A,C (note the slight difference in magnification). B: Region slightly caudal to A but in a region where the spinal cord is open. The spinal cord has either degenerated or been stretched to an extent that only a small amount of cord with a few A60 labeled cells can be seen. C: Region caudal to B. Neural tissue is completely exposed but appears to have regained a somewhat more defined morphology. D: Section through the most caudal region of the open spinal cord. Note at all levels (B-D) there are distinct DRGs (asterisks) but reduced in size compared to normal (compare with Fig. 4). The bar represents 200 pm.

NEURON DIFFERENTIATION IN NEURAL TUBE MUTANTS 525
acquired. It may be a default pathway, dorsal fate being specified by gradients set up by the floor plate. Alterna- tively, molecules such as dorsalin-1 (Basler et al., 1993) which is expressed in the dorsal neural tube shortly after neural tube closure, and has been found to prevent differen- tiation of motor neurons in spinal cord explants, may be intricately involved in setting up neuronal patterns. Addi- tionally, interactions between the dorsal neural tube and the overlying ectoderm could influence dorsal phenotype. Despite the abnormal morphology of the neural tube in spina bifida embryos, interactions between the ectoderm and dorsal neural tube are still possible; however, the ectoderm would be in contact with the open neural tube at the lateral regions of the presumptive dorsal horn.
SUMMARY We have analyzed spinal cord maturation in two neural
tube mutants: curly tail and splotch-delayed. Our data indicate that neuronal differentiation, motor neuron speci- fication and dorsal commissural axon development in spina bifida embryos occurs normally during early embryogen- esis. In ct embryos dorsal root axon development also occurs normally at this time. In contrast, P a 3 ~ 3 ~ p - ~ embryos have decreased or absent dorsal roots. Despite the seemingly normal early development in both mutants, the neural tissue undergoes a prodigious degeneration during late embryogenesis resulting in almost complete destruction of the exposed spinal cord.
In a broader sense, these data suggest that the potential clinical outcome in infants with spina bifida may be far better than previously supposed. Because initial neuronal production and differentiation occurs normally to such a great extent in affected embryos, the major cause of dis- ability may be the tremendous neuronal loss occurring at late gestational ages. Perhaps drug treatments or in utero surgeries designed to protect the neural tissue from chemi- cal and physical insult may eliminate these disabilities.
ACKNOWLEDGMENTS This work was done as part of a Multicenter Agreement
for Studying Neural Tube Defects in Mutant Mice funded by the National Institute for Child Health and Human Development, NIH, through Cooperative Agreement HD28845. This manuscript represents a portion of a disser- tation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy at the University of Utah School of Medicine, Salt Lake City, Utah.
The monoclonal antibodies 5A5, 2H3, 3A10, and 40.2D6 developed by T. Jessell and J. Dodd were obtained from the Developmental Studies Hybridoma Bank maintained by the Department of Pharmacology and Molecular Sciences, The Johns Hopkins University School of Medicine, Baltimore, MD 21205, and the Department of Biology, University of Iowa, Iowa City, IA 52242, under contract N01-HD-2-3144 from the NICHD.
We thank Drs. Sheryl Scott, Gary Schoenwolf, Dan Goldowitz, and Kristin Hamre for their helpful comments and criticisms. We also thank Christopher Krejci, Connie Shwalb, and Christopher Galloway for technical assistance and Tim Gallagher for assistance with the figures.
LITERATURE CITED Altman, J., and S.A. Bayer (1984) The development of the rat spinal cord.
Adv. Anat. Embryol. Cell Biol. 85:l-168. Basler, K., T. Edlund, T.M. Jessell, and T. Yamada (1993) Control of cell
pattern in the neural tube: Regulation of cell differentiation by dorsa- lin-l, a novel TGFb family memher. Cell 73t687-702.
Bovolenta, P., and J.Dodd (1990) Guidance of commissural growth cones at the floor plate in embryonic rat spinal cord. Development 109:435-447.
Brook, F.A., AS. Shum, H.W.M. VanStraaten, and A.J. Copp (1991) Curvature of the caudal region is responsible for failure of neural tube closure in the curly tail (ct) mouse embryo. Development 11S671-678.
Copp, A.J. (1985) Relationship between timing of posterior neuropore closure and development of spinal neural tube defects in mutant (curly tail) and normal mouse embryos in culture. J. Embryol. Exp. Morphol. 88:39-54.
Copp, A.J., and F.A. Brook (1989) Does lumbosacral spina bifida arise by failure of neural folding or by defective canalisation. J. Med. Gen. 26:160-166.
Copp, A.J., F.A. Brook, and H.J. Roberts (1988) Acell-type-speeificabnormal- ity of cell proliferation in mutant (curly tail) mouse embryos developing spinal neural tube defects. Development 104.285-295,
Copp, A.J., M.J. Seller, and P.E. Polani (1982) Neural tube development in mutant (curly-tail) and normal mouse embryos: the timing in posterior neuropore closure in vivo and in vitro. J. Embryol. Exp. Morphol. 69:151-167.
Dickie, M.M. (1964) New Splotch alleles in the mouse. J. Hered. 55:97-101. Dodd, J., S.B. Morton, D. Karagogeos, M. Yamamoto, and T.M. Jessell (1988)
Spatial regulation of axonal glycoprotein expression on subsets of embryonic spinal neurons. Neuron 1:105-116.
Edmonds, L.D., and L.M. James (1990) Temporal trends in the prevalence of congenital malformations at birth based on the birth defects monitoring program, United States, 1979-1987. MMWR 39 (No. SS-4):19-23.
Embury, S., M.J. Seller, M. Adinolfi, and P.E. Polani (1979) Neural tube defects in curly-tail mice. Incidence, expression and similarity to the human condition. Proc. R. SOC. Lond. [Biol.] 206:85-94.
Ericson, J., S. Thor, T. Edlund, T. Jessell, and T. Yamada (1992) Early stages of motor neuron differentiation revealed by expression of homeobox gene Islet-1. Science 256:1555-1560.
Estiheiro, J.P., F.A. Brook, and A.J. Copp (1993) Interaction between splotch (Sp) and curly tail (ct) mouse mutants in the embryonic development of neural tube defects. Development 119:113-121.
Feder, N. (1976) Solitary cells and enzyme exchange in tetrapartental mice. Nature 263.67439.
Gruneberg, H. (1954) Genetical studies on the skeleton of the mouse VIII. Curly-tail. J. Genetics 5 2 5 2 4 7 .
Hamre, K.M., K.P. Chepenik, and D. Goldowitz (1995) The annexins: Specific markers of midline structures and sensory neurons in the developing murine central nervous system. J. Comp. Neurol. 352421- 435.
Hamre, K.M., C.R. Keller-Peck, R.M. Campbell, A.C. Peterson, R.J. Mullen, and D. Goldowitz (1996) Annexin IV is a marker of roof and floor plate development in the murine CNS. J. Comp. Neurol. 368:527-537.
Holley, J.A. (1982) Early development of the circumferential axonal pathway in mouse and chick spinal cord. J. Comp. Neurol. 205:371-382.
Karlsson, O., S. Thor, T. Norberg, H. Ohlsson, and T. Edlund (1990) Insulin gene enhancer binding protein Isl-1 is a member of a novel class of proteins containing both a homeo- and a Cys-His domain. Nature 344:879-882.
Keller-Peck, C., and R.J. Mullen (1993) Evidence for late neuronal degenera- tion in the open neural tube of curly-tail mutant mice. SOC. Neurosci. Abs. 19:181.
Keller-Peck, C., and R.J. Mullen (1994) Pathogenesis in the dysraphic neural tube of curly-tail mutant mice. SOC. Neurosci. Abst. 202333.
McConnell, J.A. (1981) Identification of early neurons in the brainstem and spinal cord. 11. An autoradiographic study in the mouse. J. Comp. Neurol. 200:273-288.
McLean, I.W., and P.K. Nakane (1974) Periodate-lysine-paraformaldehyde fixative, a new fixative for immunoelectron microscopy. J. Histochem. Cytochem. 22:1077-1083.
Moase, C.E., and D.G. Trasler (1989) Spinal ganglia reduction in the splotch-delayed mouse neural tube defect mutant. Teratology 40:67-75.
Moase, C.E., and D.G. Trasler (1991) N-CAM alterations in splotch neural tube defect mouse embryos. Development 113:1049-1058.

526 C.R. KELLER-PECK AND RJ. MULLEN
Mullen, R.J., C.R. Buck, and A.M. Smith (1992) NeuN, a neuronal specific nuclear protein in vertebrates. Development 116:201-211.
Neale, S.A., and D.G. Trader (1994) Early sialyation on N-CAM in splotch neural tube defect mouse embryos. Teratology 50:118-124.
Neumann, P.E., W.N. Frankel, V.A. Letts, J.M. Coffin, A.J. Copp, and M. Bernfield (1994) Multifactorial inheritance of neural tube defects: local- ization of the major gene and recognition of modifiers in ct mutant mice. Nature Genet. 6:357-362.
Nornes, H.O., and M. Carry (1978) Neurogenesis in spinal cord of mouse: an autoradiographic analysis. Brain Res. 159:1-16.
Peters, J., R. Selley, and Y. Cocking (1995) Mouse gene list. Mouse Genome 93:184-357.
Peters, P.W.J., J.A.M.A. Dormans, and J.A.G. Geelen (1979) Light micro- scopic and ultrastructural observations in advanced stages of induced exencephaly and spinal bifida. Teratology 19:3345.
Placzek, M., M. Tessier-Lavigne, T. Jessell, and J. Dodd (1990a) Orientation of commissural axon8 in uitro in response to a floor plate-derived chemoattractant. Development 11 Ot19-30.
Placzek, M., M. Tessier-Lavigne, T. Yamada, T. Jessell, and J. Dodd (1990b) Mesodermal control of neural cell identity: Floor plate induction by the notochord. Science 250t985-988.
Placzek, M., T. Yamada, M. Tessier-Lavigne, T. Jessell, and J. Dodd (1991) Control of dorsoventral pattern in vertebrate neural development: induction and polarizing properties of the floor plate. Development Supplement 2t105-122.
Russell, W.L., and B. Roscoe (1947) Splotch, a new mutation in the house mouse, Mus musculus. Genetics 32:102.
Sauer, F.C. (1935) The cellular structure of the neural tube. J. Comp. Neurol. 63: 13-23.
Sidman, R.L., I.L. Miale, and N. Feder (1959) Cell proliferation and migration in the promitive ependymal zone; an autoradiographic study of histogenesis in the nervous system. Exp. Neurol. 1:322-333.
Smith, J.L., and G.C. Schoenwolf (1989) Notochordal induction of cell wedging in the chick neural plate and its role in neural tube formation. J. Exp. Zool. 25Ot49-62.
Thor, S., J. Ericson, T. Brannstrom, and T. Edlund (1991) The homeodo- main LIM protein Isl-1 is expressed in subsets of neurons and endocrine cells in the adult rat. Neuron 7:881-889.
Trader, D.G., and G. Morriss-Kay (1991) Immunohistochemical localization of chondroitin and heparan sulfate proteoglycans in pre-spina bifida splotch mouse embryos. Teratology 44t571-579.
Tsuchida, T., M. Ensini, S.B. Morton, M. Baldassare, T. Edlund, T.M. Jessell, and S.L. Pfaff (1994) Topographic organization of embryonic motor neurons defined by expression of LIM homeobox genes. Cell 79,957-970.
van Straaten, H.W.M., J.W.M. Hekking, C. Consten, and A.J. Copp (1993) Intrinsic and extrinsic factors in the mechanism of neurulation: Effect of curvature if the body axis on closure of the posterior neuropore. Development 11 7: 1163-1 172.
van Straaten, H.W.M., J.W.M. Hekking, A.J. Copp, and M. Bernfield (1992) Deceleration and acceleration in the rate of posterior neuropore closure during neurulation in the curly tail (ct) mouse embryo. Anat. Embryol. 185.169-174.
Vogan, K.J., D.J. Epstein, D.G. Trader, and P. Gros (1993) The splotch- delayed (Spd) mouse mutant carries a point mutation within the paired box of the Pax-3 gene. Genomics 17t364-369.
Wentworth, L.E. (1984a) The development of the cervical spinal cord of the mouse embryo: I. A golgi analysis of ventral root neuron differentiation. J. Comp. Neurol. 22281-95.
Wentworth, L.E. (1984b) The development of the cervical spinal cord of the mouse embryo. 11. A golgi analysis of sensory, commissural, and associa- tion cell differentiation. J. Comp. Neurol. 222:96-115.
Yamada, T., M. Placzek, H. Tanaka, J. Dodd, and T.M. Jessell (1991) Control of cell pattern in the developing nervous system: polarizing activity of the floor plate and notochord. Cell 64t635-647.