patterns and variations in natural succession following massive ice-scour of a rocky intertidal...
TRANSCRIPT

ELSEVIER Journal of Experimental Marine Biology and Ecology,
214 (1997) 121-147
JOURNAL OF EXPERIMENTAL MARINE BIOLOGY AND ECOLOGY
Patterns and variations in natural succession following massive ice-scour of a rocky intertidal seashore
L.J. McCook”, A.R.O. Chapman
Department of Biology, Dalhousie University, Hallfax, Nova Scotia. Canada
Received I 1 April 1996; revised I2 August 1996; accepted 7 October 1996
Abstract
In April of 1987, unusual sea ice severely scoured exposed rocky seashores near Halifax, Nova Scotia. Mid-tide levels of these shores are commonly dominated by a lush canopy of perennial
fucoid rockweeds, with an understory of mussels, barnacles and crustose algae. Patches of cleared space in these canopies are often occupied by ephemeral algae. After the scour, emergent rock in
mid-shore areas was occupied by a succession of diatoms, green and blue-green ephemeral algae, and then Fucus canopy. Mussels reappeared in the understory, but rarely displaced the rockweed
canopy. These events provided an exceptional opportunity to study recovery dynamics in a
perennial intertidal community, at a scale ( > 40 km) much larger than any in previous studies. This paper describes large-scale patterns of natural succession at a variety of locations and
disturbance intensities, and then reviews the community structure in the light of several experimental tests of processes and species interactions during the succession. Formation of a Fucus canopy was ubiquitous, and variations in the recruitment and regeneration of Fucus were
major influences on the rate of succession and the abundance of other species. A survey of Fucus
and mussel dominance over about I.5 km of coastline supports this view. Most areas in this survey
were not dominated by mussels, but by the small-scale patch dynamics of ephemeral algae and Fucus, in succession. Previous work suggested that emergent rock on these shores is normally
dominated by fucoid rockweeds because predatory whelks control competition with the mussels. and because herbivorous littorinid snails control competition with ephemeral algae. However, OUI
observational and experimental results show that these interactions were not generally important during this succession, and that the importance of different species and interactions varies. Out
results emphasize the importance of several properties of the rockweeds, as the most abundant and structurally important species. These conclusions suggest caution in the extrapolation of specific, small-scale experiments to general hypotheses about community structure and recovery from disturbance. 0 1997 Elsevier Science B.V.
Ke.vwrjrds: Fucus; Ice-scour; Rocky intertidal succession; Mussel; Nova Scotia; Large scale
*Corresponding author. Present address for correspondence: Australian Institute of Marine Science & CRC:
Reef Research, PMB#3, Cape Ferguson, Townsville M.C., Queensland 4810, Australia. E-mail:
0022.0981/97/$17.00 0 1997 Elsevier Science B.V. All rights reserved
P/I SOO22-098 I (96)0275 I-7

122 L.J. McCaok, A.R.O. Chapman I J. Exp. Mar. Bid. Ed. 214 (1997) 121-147
1. Introduction
On exposed rocky seashores in the Halifax region of Nova Scotia, Canada, emergent rock in the mid-shore is largely dominated by lush canopies of the rockweeds Fucus
spiralis, Fucus vesiculosus and Fucus evanescens, at progressively lower zones. Much of the substrata under these canopies is occupied by filter feeding barnacles
(Semibalanus balanoides) and mussels (Mytilus edulis and M. trossulus), and by crustose
algae, especially Hildenbrundia rubru. Cleared space in these fucoid canopies is often
occupied by ephemeral algae, especially in winter and spring. Other common species include grazing periwinkle snails, particularly Littorinu obtusata and Littorinu rudis,
grazing gammarid amphipods, and carnivorous dogwhelks (Nucella lqillus) that eat the
barnacles and mussels (summarised in Fig. 1 in McCook and Chapman, 1991).
Extrapolating the classic studies of Menge and Lubchenco in New England suggests that Fucus rockweeds compete for primary space with both ephemeral algae and with
mussels and barnacles. Where the rockweeds persist, it is due to predator control of the invertebrates by whelks and herbivore control of ephemeral algae by littorinid snails
(especially Littorina littorea) (Menge, 1976, 1978a,b, 1991a,b; Lubchenco, 1978, 1982,
1983, 1986; Lubchenco and Menge, 1978; Petraitis, 1987, 1990; Chapman and Johnson, 1990). On exposed shores, wave action may inhibit whelk predation, leading to mussel
dominance and exclusion of rockweeds (Menge, 1976, 1978a,b; Menge and Sutherland, 1976). However, a series of recent experimental studies during a large-scale succession
in Nova Scotia have emphasised several properties of the rockweed canopy, rather than
predator/herbivore control of competition (McCook and Chapman, 1991, 1992, 1993).
The present paper places these experimental studies in the context of large-scale patterns during this succession.
In late March and early April of 1987, extensive pack-ice from the Gulf of St. Lawrence was blown onshore in the Halifax region by unusual wind conditions, and
large areas of exposed shore were severely scoured (Fig. 1). This icepack was
approximately 30 km (up to 300 km) in length, 2-20 km wide, 1-3 m thick, and included blocks as large as 10 tonnes (Whitehead, 1987 and pers. obs.). The action of
winds, tides and swells on the ice removed most biotic material from large areas of
shore, from the extreme supralittoral to more than 1 m below mean low water. Areas
that had previously been, and would later be dominated by the perennial rockweed canopy were left virtually bare of plants or animals (pers. obs.). Aerial survey showed
that over 50 km of shoreline were affected (Fig. 2). Ice-scour of such extent and intensity is extremely rare in this region, and has not
occurred for at least 25 years (Bedford Institute of Oceanography, 1987; Dinsmore, 1972; Markham, 1980). The more frequent scour reported for New England is much less extensive (Markham, 1980) and severe (since it does not remove fucoid canopies, see Wethey, 1985).
The large scale of this disturbance provided an opportunity to examine interactions and variations in succession over an unusually large scale. This contrasts with most previous studies of intertidal successions, which used patches created in established communities (e.g. Lubchenco and Menge, 1978; Murray and Littler, 1978; Sousa, 1979, 1984a; Lubchenco, 1983; Turner, 1983; Van Tamelen, 1987; Buschmann, 1990; Farrell,
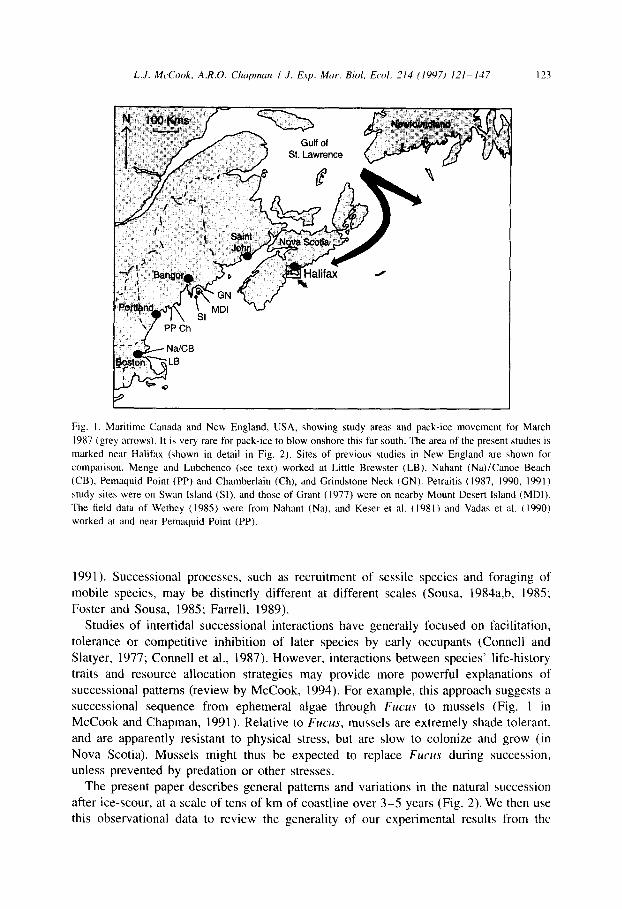
L.J. McCook, A.R.O. Chupman I J. Exp. Mar. Bid. Ed. 214 (1907) 121-147 123
b
Fig. I. Maritime Canada and New England, USA, showing study areas and pack-ice movement for March
1987 (&rey arrows). It is very rare for pack-ice to blow onshore this far south. The area of the present studies is
marked near Halifax (shown in detail in Fig. 2). Sites of previous studies in New England are shown for
comparison. Menge and Lubchenco (see text) worked at Little Brewster (LB), Nahant (Na)/Canoe Beach
(CB), Pemaquid Point (PP) and Chamberlain (Ch), and Grindstone Neck (GN). Petraitis (1987, 1990. 1991)
study sites were on Swan Island (SI), and those of Grant (1977) were on nearby Mount Desert Island (MD]).
The field data of Wethey (1985) were from Nahant (Na), and Keser et al. (1981) and Vadaa et al. ( 1990)
worked at and near Pemaquid Point (PP).
1991). Successional processes, such as recruitment of sessile species and foraging of
mobile species, may be distinctly different at different scales (Sousa, 1984a,b. 1985;
Foster and Sousa, 1985; Farrell, 1989). Studies of intertidal successional interactions have generally focused on facilitation,
tolerance or competitive inhibition of later species by early occupants (Connell and Slatyer, 1977; Connell et al., 1987). However, interactions between species’ life-history
traits and resource allocation strategies may provide more powerful explanations of successional patterns (review by McCook, 1994). For example, this approach suggests a
successional sequence from ephemeral algae through FUCU~ to mussels (Fig. 1 in McCook and Chapman, 1991). Relative to Fucus, mussels are extremely shade tolerant,
and are apparently resistant to physical stress, but are slow to colonize and grow (in
Nova Scotia). Mussels might thus be expected to replace FUCU~ during succession, unless prevented by predation or other stresses.
The present paper describes general patterns and variations in the natural succession after ice-scour, at a scale of tens of km of coastline over 3-5 years (Fig. 2). We then use this observational data to review the generality of our experimental results from the
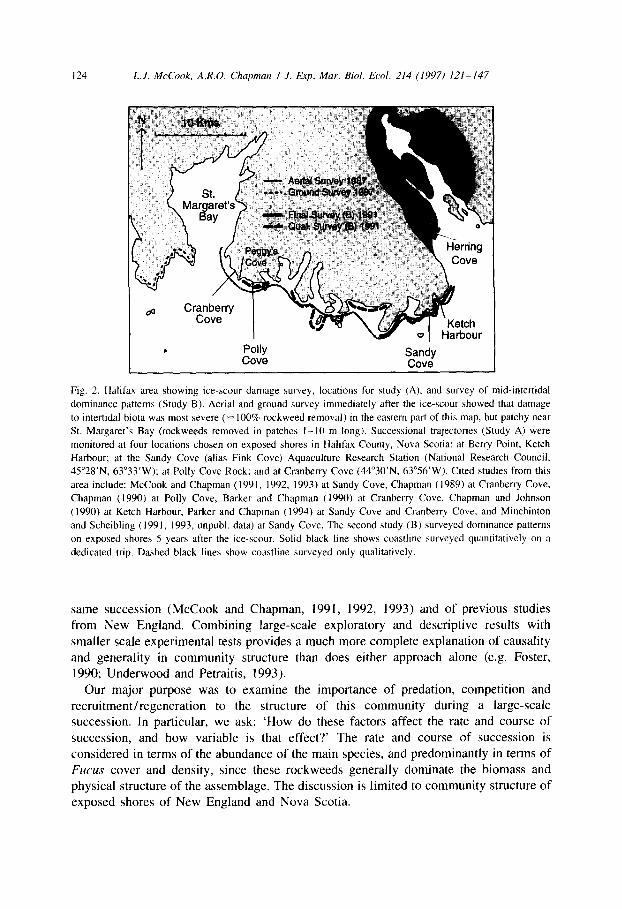
124 L.J. McCook, A.R.O. Chapman I J. Exp. Mar. Bid. Ed. 214 (1997) IZI-147
l Polly Cove
Fig. 2. Halifax area showing ice-scour damage survey, locations for study (A), and survey of mid-intertidal
dominance patterns (Study B). Aerial and ground survey immediately after the ice-scour showed that damage
to intertidal biota was most severe (= 100% rockweed removal) in the eastern part of this map, but patchy near
St. Margaret’s Bay (rockweeds removed in patches I-IO m long). Successional trajectories (Study A) were
monitored at four locations chosen on exposed shores in Halifax County, Nova Scotia: at Berry Point, Ketch
Harbour; at the Sandy Cove (alias Fink Cove) Aquaculture Research Station (National Research Council,
45”28’N, 63”33’W); at Polly Cove Rock; and at Cranberry Cove (44”30’N, 63”56’W). Cited studies from this
area include: McCook and Chapman (1991, 1992, 1993) at Sandy Cove, Chapman (1989) at Cranberry Cove,
Chapman (1990) at Polly Cove, Barker and Chapman ( 1990) at Cranberry Cove, Chapman and Johnson
(1990) at Ketch Harbour, Parker and Chapman (1994) at Sandy Cove and Cranberry Cove, and Minchinton
and Scheibling (1991, 1993, unpubl. data) at Sandy Cove. The second study (B) surveyed dominance patterns
on exposed shores 5 years after the ice-scour. Solid black line shows coastline surveyed quantitatively on a
dedicated trip. Dashed black lines show coastline surveyed only qualitatively.
same succession (McCook and Chapman, 1991, 1992, 1993) and of previous studies from New England. Combining large-scale exploratory and descriptive results with
smaller scale experimental tests provides a much more complete explanation of causality and generality in community structure than does either approach alone (e.g. Foster, 1990; Underwood and Petraitis, 1993).
Our major purpose was to examine the importance of predation, competition and recruitment/regeneration to the structure of this community during a large-scale succession. In particular, we ask: ‘How do these factors affect the rate and course of
succession, and how variable is that effect?’ The rate and course of succession is considered in terms of the abundance of the main species, and predominantly in terms of Fucus cover and density, since these rockweeds generally dominate the biomass and physical structure of the assemblage. The discussion is limited to community structure of exposed shores of New England and Nova Scotia.

L.J. McCook, A.R.O. Chapman I J. Exp. Mar. Bid. Ed. 214 (1997) 121-147 125
The specific questions asked are:
1. What was the general pattern of species abundances during this succession?
2. How did that pattern vary: between sites, locations and successional stages; and,
between studies, including earlier work in New England. 3. Are the patterns seen in our experimental studies representative?
4. Did areas with different patch size and severity of disturbance differ in rate or course of succession? In particular: What is the relationship between distance to mature
rockweeds and time to recovery of canopy, or recruitment density?
5. Did vegetative regeneration from damaged holdfasts contribute to canopy recovery’?
6. How are species abundances related for Fucus, ephemeral algae and herbivores; or
for Fucus, filter feeders and whelks?
The paper includes two observational studies. The first (A) describes and interprets
species abundances over 3 years in permanent quadrats at four locations between Halifax
Harbour and St. Margaret’s Bay (Fig. 2; a ‘natural trajectory experiment’ sensu
Diamond, 1986). The second study (B) is a simple survey to identify broad patterns of
community composition and dominance, nearly 5 years after the ice-scour (a ‘natural snapshot experiment’ sensu Diamond, 1986). Both studies are restricted to shores
exposed to direct wave action, and to the FUCUS vesiculosus L. and Fucus evanescens C.
Ag. zones. The emphasis in both studies is on exploratory description of patterns, and the
ability of different hypotheses to explain those patterns. In particular, the ‘dominance’ survey (B) assessed the generality of explanations of
dominance by mussels or rockweeds on exposed shores. If mussel exclusion of rockweeds due to decreased predation on exposed shores (Menge, 1976, 1978a,b) is
general, then a large proportion of exposed midshores should be dominated by mussels,
to the exclusion of Fucus (see also Foster, 1990; Underwood and Petraitis, 1993). However, there has been little or no quantitative assessment of this dominance, and
many rocky shores in this area and in New England are seemingly dominated by
rockweeds, including most exposed shores (pers. obs.; Keser et al., 1981; Cousens,
1985; Petraitis, 1987; Chapman and Johnson, 1990; Vadas et al., 1990). By estimating
the relative dominance of mussels and Fucus, we could assess the generality of different
explanations of community structure.
2. Study sites and methods
2.1. Study A: Natural successional trajectories
2.1.1. Locations and study sites
To obtain descriptions of successional trajectories, we monitored the abundance of all macroscopic species in permanent quadrats at four locations (Fig. 2) for over 3 years. The four locations were chosen as apparently representative of exposed shores over a geographic range of about 40 km, and covering a range of disturbance intensities (Table

126 L.J. McCook, A.R.O. Chapman I J. Exp. Mar. Biol. Ecol. 214 (1997) 121-147
Table I Study sites
Location scour
severity
Patch Size Site Zones
(m) numbers
Ketch Harbour Severe 400 3
Sandy Cove Severe 1000
I
3 6
4
Polly Cove Moderate IO I Cranberry Cove Moderate l-O.2 6
Undamaged 0 I
Series of heights, F. evanescens+F. vesiculosus Series of heights, F. evanescens+F. vesicufosus
(overlapping I site with:)
Series of distance from fertile Fucus F. evane.wens Series along shore, F. evanesces and F. Vesiculosus F. Vesiculosus
Details given in text; ‘Site’ refers to a group of 5 quadrat
I). The damage at the two eastern locations, Ketch Harbour and Sandy Cove, was extreme. Most or all of the biota had been removed from nearly all of the shore for
distances of many hundreds of metres, leaving only small refuges of a few metres in
length in some, unusually protected locations. Further west (Cranberry and Polly Coves), however, the damage was less severe, and more patchy. Patches of tens of metres of
damaged shore were interspersed with apparently undamaged vegetation. Observations
of the ice pack suggested that the eastern sites had been scoured by hard packed ice, whereas the western areas had been exposed to large (several m3), free-floating blocks of
brash ice, which caused localised damage to the shore communities.
Sites were chosen to represent a range of biological and physical conditions and
disturbance severities, but all four locations are directly exposed to severe wave action
from the open Atlantic Ocean (see Figs. 1 and 2, waves > 10 m). There is little public
access to these four locations, ensuring natural succession with minimal human interference. Throughout this paper, ‘location’ refers to the four areas in Fig. 2, separated at the scale of 10s of km. ‘Site’ refers to a group of (5) quadrats, groups
separated at scales of l-100 m. Locations included between 1 and 6 sites each (Table 1). At each site, 5 permanent, marked quadrats were randomly chosen from 10 plots. All sites were on gently sloping granite rock with fairly similar texture.
At Ketch Harbour (K), three sites (i.e., 3 groups of 5 quadrats) were located at a range of heights through the F. vesiculosus and F. evanescens zones on a gently shelving rock
promontory. At Sandy Cove (SC), three sites were located at different heights in the F.
evanescens and F. vesiculosus zones. One of these sites also served as the fourth in a series of four sites located at distances of 0.1-30 m from a small ( = 25 m’) refuge of fertile Fucus (both spp.). These sites were on a gently sloping shelf, which was also used
for the experiment reported in McCook and Chapman (1991). At Polly Cove Rock (PC), an exposed rock island, one site was placed on exposed, gently sloping rock in the F. evanescens zone. At Cranberry Cove (CC), six sites were located over several hundred metres of shore, in patches of partially damaged F. vesiculosus and F. evanescens
canopy. A seventh site, in intact canopy immediately adjacent to a partially damaged site, provided an undisturbed site for comparison.

L.J. McCook, A.R.O. Chapman I .I. Exp. Mar. Biol. Ecol. 214 (1997) 121-147 121
2.1.2. Sampling and analyses
Abundance of all macroscopic species was initially measured every few months, and
then at longer intervals. Abundances were measured as density and/or percent cover, in
10 X 10 cm quadrats. Percent cover of sessile species was estimated by identifying all
species present at 30 random points on a 100 point strung grid. Density of Fucus was counted as shoots or stipes, using jeweller’s magnifying glasses when necessary. Juvenile FLICKS morphologies inter-graded (see McLachlan et al., 1971) and hence could
not generally be identified to species. Description of the succession is also based on extensive qualitative field notes and photographs taken during early stages of the
succession. Data analyses include estimates of several environmental parameters. (i) Patch size
for each site was estimated as the order of magnitude of the distance to intact rockweed
beds of similar size to the disturbed area. (ii) The severity of the scour was estimated by
the cover of crustose algae remaining (assumes similar pre-disturbance covers), and by
the number of holdfasts left in each quadrat (also assumes similar pre-disturbance
densities). ‘Severity’ here measures the damage to the biota (sensu Sousa, 1984b). (iii)
Dispersal distance was estimated by the distance to the nearest mature Fucus plants for
each quadrat. Five summary variables are used to compare successional trajectories. (i) Rate of
canopy recovery was (inversely) estimated as the number of months before the first date
at which estimated canopy cover was 100%. Canopy closure (100% cover) of Fucus is an important stage in recovery, although it does not represent ‘mature’ canopy, since
Fucus shoots were often = 10 cm long at this point. (ii) Maximum density of Fucus
was used to indicate Fucus abundance before self-thinning. (iii) Initial Fucus recruit- ment was estimated by the maximum Fucus density for the first six months, based on the
recruitment period measured in Fig. 2E of McCook and Chapman (1993). To contrast with these variables, which measure earlier succession, (iv) final cover, and/or (v) final
density are used to identify lasting effects.
Statistical analyses are only used to quantify the relationships, not as tests. Data were inspected for homogeneity of variance (Cochran’s Test for anova) and independence 01
residuals where appropriate, but some heteroscedasticity, and unbalanced design in the
anova for effects of location (5 5 n < 20), were tolerated, especially since the P values were very small. Note that %SST, used to estimate magnitude of effects, is not reliable
for comparisons between studies (Underwood and Petraitis, 1993). Note also that, as with any such observational study, locations and sites differ in numerous aspects as well as the factors of interest. Causality is only attributed where patterns are supported by
experimental results. Data were analyzed using SystatTM (Wilkinson, 1989).
2.2. Study B: Survey of mid-intertidal community dominance patterns
2.2.1. Methods and survey locations
To assess the broad-scale importance of processes leading to dominance by mussels or rockweeds, we surveyed dominance patterns on exposed rocky shores in the study areas, nearly 5 years after the ice-scour disturbance. The proportions of shore dominated by Fucus or mussels were estimated for approximately 15 km of exposed coastline (Fig. 2),

128 L.J. MeCook, A.R.O. Chapman I J. Exp. Mar. Biol. Ed. 214 (1997) 121-147
from Duncan’s Cove (44”295’N, 63”31S’W), at the mouth of Halifax Harbour, to
Pennant Point (44”26’N, 63”39’W), near Sambro, Halifax County, using a small boat. The survey was carried out during low tide (from 2 h before low tide to 3.5 h after) on the 5th November 1991. Good visibility and calm sea conditions allowed us to cruise
between 10 and 30 m from the shore. The survey was restricted to rocky bedrock shores exposed to direct wave action from the open ocean, and excluded sandy and cobble
beaches and boulder fields. Note that the estimated length of the survey includes only
exposed shores, and is very approximate owing to the difficulty of measuring irregular
coastlines. The midshore was considered as a belt transect of varying width, and scored as
sections of homogeneous occupancy. In each section, proportions of area were assigned to three dominance categories: (i) mussel dominance (little or no canopy): (ii) closed
canopy; (iii) other. Space not occupied by Fucus or mussels had varying amounts of bare rock, crustose algae or often lush growths of ephemeral algae, often with recruiting
or regenerating Fucus shoots. Where dominance varied much at a small scale ( < 10 m),
we estimated ranges for each category. Dominance is used here to mean high
proportional biomass. Since we are interested in whether mussels may exclude Fucus
canopy (and not just persist as understory), space was only considered to be dominated
by mussels if Fucus canopy was largely absent.
Although the dominance estimates are only approximate, they are unlikely to
underestimate mussel dominance, since very small mussel patches ( < 30 cm diameter) were easily detected and scored. The size of most Fucus individuals means that areas
must be larger than this to count as closed canopy. Furthermore, the survey was carried out soon after a severe storm that appeared to have removed some rockweed. Since
mussels are probably less susceptible to wave damage than Fucus, this would reduce the
proportion of rockweed dominance and increase mussel dominance by revealing mussels
previously present as understory.
Also shown in Fig. 2 are several areas of coastline qualitatively surveyed for patches
of mussel dominance during the previous summer. These observations serve as indications of the generality of the quantitative survey.
3. Results
3.1. Study A: Natural successional trajectories
3.1. I. Patterns and variability in successional trajectory: Generul observations
A general pattern of succession was apparent, with an initial bloom of ephemeral, mainly filamentous green algae during the spring and early summer, replaced by ephemeral blue-green algae during the later summer (Fig. 3 and qualitative pers. obs.). Predominant eukaryotic ephemeral algae were the filamentous green algae Ulothrix
flacca and Urospora penicilliformis, along with Bangia atropurpurea, Porphyra species, Entermorpha intestinalis and Ulva lactuca. The blue-green algae were mainly Calothrix contarenii and Lyngbya majuscula. These ephemerals were replaced by Fucus, which
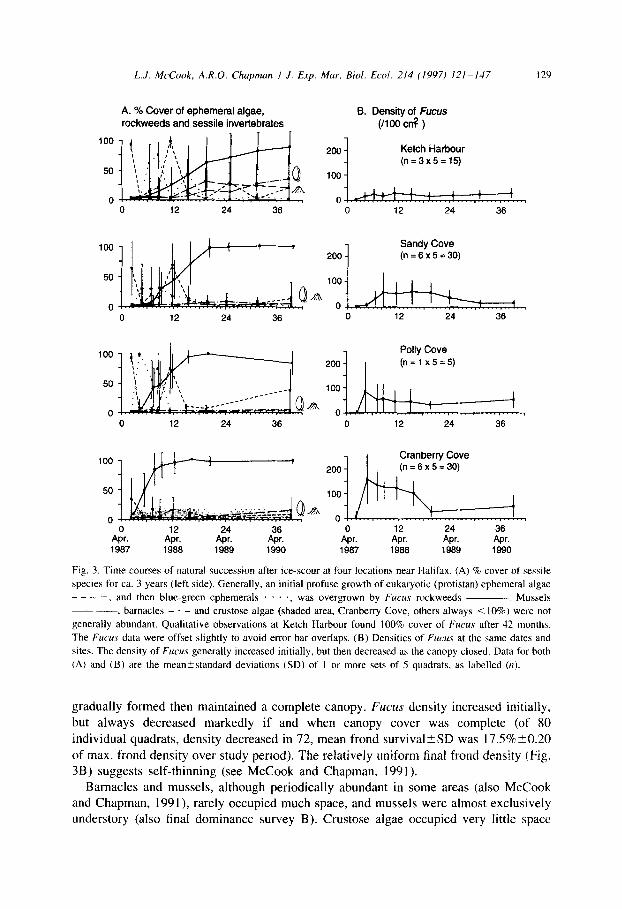
L.J. MrCook, A.R.O. Chapman I J. Exp. Mar. Bid. Ed. 214 (1997) 121-147 12’)
A. % Cover of ephemeral algae, rockweeds and sessile invertebrates
8. Density of Fucus (1100 cn3 )
0 12 24 36 0 12 24 36
0 12 24 36 0 12 24 36
Air. 24 0 12 24
Apr. Apr. Apr. Apr. $r. 1987 1988 1969 1990 1987 1988 1969 1990
Fig. 3. Time courses of natural succession after ice-scour at four locations near Halifax. (A) % cover of sesslle
species for ca. 3 years (left side). Generally, an initial profuse growth of eukaryotic (protistan) ephemeral algae
- -, and then blue-green ephemerals . . . ., was overgrown by Fucus rockweeds -. Mussels
~ --, barnacles - - and crustose algae (shaded area, Cranberry Cove, others always < 10%) were not
generally abundant. Qualitative observations at Ketch Harbour found 100% cover of Fucus after 42 months.
The Fucus data were offset slightly to avoid error bar overlaps. (B) Densities of Fucus at the same dates and
sites. The density of Fucus generally increased initially, but then decreased as the canopy closed. Data for both
(A) and (B) are the meankstandard deviations (SD) of I or more sets of 5 quadrats, as labelled (n).
gradually formed then maintained a complete canopy. Fucus density increased initially. but always decreased markedly if and when canopy cover was complete (of 80
individual quadrats, density decreased in 72, mean frond survival+-SD was 17.5%‘c-+O.20
of max. frond density over study period). The relatively uniform final frond density (Fig. 3B) suggests self-thinning (see McCook and Chapman, 1991).
Barnacles and mussels, although periodically abundant in some areas (also McCook and Chapman, 1991), rarely occupied much space, and mussels were almost exclusively understory (also final dominance survey B). Crustose algae occupied very little space

130 L.J. McCook, A.R.O. Chapman I J. Exp. Mar. Biol. Ecol. 214 (1997) 121-147
throughout the three years of observations, except where they were relatively undamaged by the scour.
After 3 years, successional dynamics at some sites were overshadowed by normal
patch dynamics such as canopy loss to wave disturbance (pers. obs., e.g. Polly Cove Fig. 3). However, total understory cover of crustose algae was still small relative to
unscoured areas, suggesting that these assemblages had not fully recovered from the
scour by this time (MeankSEM cover of crusts = 7.9%+ 1.9 for Sandy Cove, compared
to 21.8%& 1.74 for an undisturbed site at Cranberry Cove, anova and contrast, P <
0.001).
The alternating abundance of eukaryotic and prokaryotic, blue-green ephemeral algae
was a seasonal pattern rather than succession, since it also occurred in cleared patches in later years (McCook and Chapman, 1993). However, these seasonal cycles disappeared
later in the succession as Fucus cover increased (i.e. successional change).
While there was little variation in the qualitative pattern of succession during the study period, there were considerable differences in quantitative patterns and timing
(Fig. 3). Canopy regrowth was much faster and (overall) mussel abundance much
smaller at Sandy Cove than at Ketch Harbour, despite apparently similar scale and intensity of scour at the two locations. Two particular sites with similar exposure and
height, one at each location, differed by nearly 2 years in time to canopy closure. Fucus
cover and density varied considerably even within sites, especially when canopy cover was increasing. Fucus density even varied considerably within sites with similar canopy
cover.
Most of the differences in successional course were correlated with differences in timing of Fucus canopy formation or with secondary disturbances to canopy, such as
storm damage (e.g. Fig. 3, Polly Cove final date). Differences in timing of canopy
formation and consequent shading of substrate probably explain the marked differences in patterns of ephemeral algal abundance at different locations (Fig. 3, Cranberry Cove, cf. others).
Herbivore and whelk densities were temporally and spatially highly variable (at these
scales) but generally small (Fig. 4), except where crevices and later canopy provided
shelter from waves. Before canopy recovery, herbivores were only abundant near crevices and other shelter, and created bare areas in the ephemeral algae surrounding the
crevices. Density tended to be higher and more even after canopy formation, except at Ketch Harbour. The density of Littorina rudis relative to L. obtusaru was often lower
where canopy cover was higher (cf. McCook and Chapman, 1991). L. littorea was extremely rare throughout this study, being more common in sheltered sites.
The rate of Fucus canopy formation was faster where moisture availability was greater, based on relationships between canopy formation and wave exposure, shore height, drainage and shading. Although such physical factors are not the focus of this study, they were a major influence on Fucus abundance (also McCook and Chapman, 1993).
3.1.2. Effects of location, patch size, disturbance intensity and dispersal distance There were considerable differences between locations in the timing of canopy
recovery and in density of Fucus (Figs. 3 and 5A). Time to complete canopy cover was

L.J. McCook, A.R.O. Chapman I J. Exp. Mar. Biol. Ed. 214 (1997) 121 147 131
A. Herbivore Densities B. Whellk Densities (/lOocm2) (11 oocrnq
12 24 36
Sandy Cove
I;:. , , *r, V .
vx 441. t 4 _
0 12 24 36
0.1
0 0 12 24 36
0.1
0 0 12 24 96
0 12 24 36
10
5 1
Cranberry Cove
. T .V ’
0 I_____ I ‘.‘--.... -
0 12 24 96
0.5
i .
l * .
4 0.11 .
.&.l ow-... ... s
&. :;r . gr . 36 Apr.
1967 1966 1969 1990
0- Apr. 1967
Fig. 4. Abundance of herbivores and carnivorous whelks during succession. (A) Densities of herbivores
Lifrorino rudis, Litrorina obrusata and amphipods for the same sites and dates as Fig. 3; (B) Densities of
carnivorous whelks. Data for herbivores and whelks were transformed to log( I -density/ 100 cm’). Data are
means of numbers of plots indicated in Fig. 3; standard deviations, not shown for clarity, were generally large
(=mean).
generally shorter and maximum density higher at the western locations, but differences
in Fucus cover and density were small by the end of the study. Within location/between site differences in recovery were generally minor, apparently because severity of damage
was fairly homogeneous at that smaller scale.
Patch size was correlated with recovery rate (Fig. 5B), as was scour severity, estimated by cover of crustose algae remaining after the scour (R = - 0.529, P -C 0.001). This latter effect was not significant within locations because cover of crustose algae was uniformly very low at the eastern locations (e.g. Ancova on location using initial cover of crustose algae as covariate gave Pcovar,nre = 0.34 and 0.9, %SST < 1% for F. vesiculosus and F. evanescens, respectively). Severity of damage to Fucus was
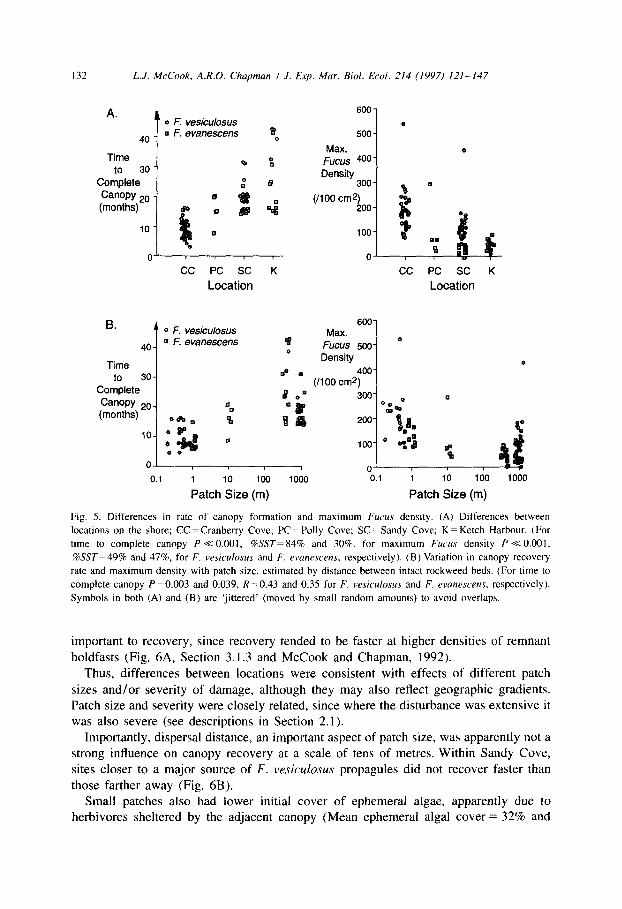
132 L.J. McCook, A.R.O. Chapman I .I. Exp. Mar. Biol. Ecol. 214 (1997) 121-147
A.
i
(0 F. vesiculosus
40 q F. evanescens fl
0
Time to 30
Complete Canopy *O (months)
0
CC PC SC K
Location
B. 0 F. vesiculosus
40 a F. evanescens 9
0
Time d
0.1
I
1 IO 100 1000
Patch Size (m)
600
500
FY:s 400
Density 300
(/loo cm%,
0 CC PC SC K
Location
600 MElX.
Fucus 500
Density 400
100 cm2) 1
o- 0.1 1 10
Patch Size
Fig. 5. Differences in rate of canopy formation and maximum Fucus density. (A) Differences between
locations on the shore; CC =Cranberry Cove; PC= Polly Cove; SC=Sandy Cove; K= Ketch Harbour. (For
time to complete canopy P +z 0.001, ‘&SST=84% and 300/c, for maximum Fucus density P +z 0.001,
?WST=49% and 47%, for F. vesiculouus and F. rvanrscens, respectively). (B) Variation in canopy recovery
rate and maximum density with patch size, estimated by distance between intact rockweed beds. (For time to
complete canopy P =0.003 and 0.039, Rz0.43 and 0.35 for F. vesiculosus and F. evanescens, respectively).
Symbols in both (A) and (B) are ‘jittered’ (moved by small random amounts) to avoid overlaps.
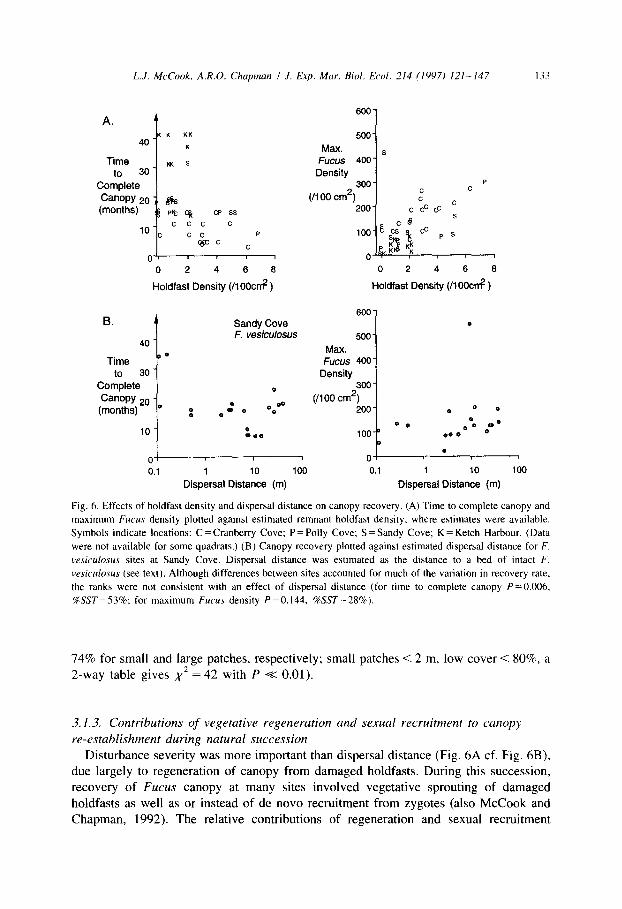
important to recovery, since recovery tended to be faster at higher densities of remnant holdfasts (Fig. 6A, Section 3.1.3 and McCook and Chapman, 1992).
Thus, differences between locations were consistent with effects of different patch
sizes and/or severity of damage, although they may also reflect geographic gradients. Patch size and severity were closely related, since where the disturbance was extensive it was also severe (see descriptions in Section 2.1).
Importantly, dispersal distance, an important aspect of patch size, was apparently not a strong influence on canopy recovery at a scale of tens of metres. Within Sandy Cove, sites closer to a major source of F. vesiculosus propagules did not recover faster than
those farther away (Fig. 6B). Small patches also had lower initial cover of ephemeral algae, apparently due to
herbivores sheltered by the adjacent canopy (Mean ephemeral algal cover = 32% and
L.J. McCook, A.R.O. Chapman I .I. Exp. Mar. Bid. Ed. 214 (1997) 121-147 133
A. t
W ss
0 ‘1 1
0 2 4 6 6
Holdfast Density (/lOOct~? )
500
Max. Fucus 400
Density 300
(/I 00 cm2) 200
100
0
0 2 4 6 6
Holdfast Density (/l OOcrr? )
B. h Sandy Cove F. vesiculosus
40 -
Time Be
to 30
Complete e Canopy 2. -o
(months) $0 e. *
:
10 - :e.
600 - a
500 -
Max.
Fucus 406-
Density 300 -
(/l 00 cm2) 200- B B 0
00 100-O *% 00 0e* B
P
0’ 0-l 0
I
0.1 1 10 100 0.1 1 10 100
Dispersal Distance (m) Dispersal Distance (m)
Fig. 6. Effects of holdfast density and dispersal distance on canopy recovery. (A) Time to complete canopy and
maximum Fucus density plotted against estimated remnant holdfast density, where estimates were available.
Symbols indicate locations: C = Cranberry Cove; P = Polly Cove; S = Sandy Cove; K = Ketch Harbour. (Data
were not available for some quadrats.) (B) Canopy recovery plotted against estimated dispersal distance for F.
wsiculosus sites at Sandy Cove. Dispersal distance was estimated as the distance to a bed of intact F.
vesiculosus (see text). Although differences between sites accounted for much of the variation in recovery rate,
the ranks were not consistent with an effect of dispersal distance (for time to complete canopy P=O.O06,
KLST=53%; for maximum Fucus density P =O.l44, %LST=28%).
74% for small and large patches, respectively; small patches < 2 m, low cover < 80%, a
2-way table gives x2 = 42 with P K 0.01).
3.1.3. Contributions of vegetative regeneration and sexual recruitment to canopy
re-establishment during natural succession
Disturbance severity was more important than dispersal distance (Fig. 6A cf. Fig. 6B), due largely to regeneration of canopy from damaged holdfasts. During this succession, recovery of Fucus canopy at many sites involved vegetative sprouting of damaged
holdfasts as well as or instead of de novo recruitment from zygotes (also McCook and Chapman, 1992). The relative contributions of regeneration and sexual recruitment

134 L.J. McCook, A.R.O. Chapman I J. Exp. Mar. Biol. Ecol. 214 (1997) 121-147
varied considerably between locations, but are difficult to determine precisely, since the importance of regeneration only became apparent during the succession.
At the less severely scoured Cranberry Cove and Polly Cove sites (Fig. 5B Fig. 6A),
vegetative sprouting from original holdfasts was directly observed to contribute to
canopy recovery, particularly of F. vesiculosus. Tiny shoots ( I 2 mm) appeared on
holdfasts within 3 months (June 1987), and gradually increased in size over that summer. The establishing canopy at these sites was patchy at a scale typical for
regenerating holdfasts. Canopy growing from regenerating holdfasts generally forms
distinct clumps 5 cm or more apart (see McCook and Chapman, 1992, Fig. 2B), reflecting the density of original mature holdfasts, whereas new recruitment (when
abundant) was usually more evenly distributed, at a scale of l-10 cm. This patchiness could be detected as bimodal distributions of Fucus density, since quadrat size was
similar to patch size.
Both regeneration and recruitment contributed to recovery of F. evanescens canopy at Cranberry and Polly Coves. Recruitment of F. evanescens to sites lower on the shore
was prolific (= 100 shoots/100 cm*) during the first 6 months of the succession, but
thick clumps of larger fronds from single holdfasts were visible among the recruits. Isolated plants of F. evanescens were common subtidally in these areas, and probably
acted as sources of propagules.
Regeneration from holdfasts appeared less important among the more severely scoured sites at Sandy Cove and Ketch Harbour. Recruitment seemed to be the major
contributor to F. evanescens canopy, since shoot density was generally high, and fairly even. Density of shoots was never as high as at the western sites, and canopy cover
increased more slowly. F. evanescens shoots first appeared through a thick cover of ephemerals, making it difficult to determine their origin, and regeneration from holdfasts
cannot be discounted at these sites. Among F. vesiculosus at Sandy Cove and Ketch Harbour, vegetative regeneration
appears to have contributed to canopy recovery, but the overall contribution was
probably small. Some holdfasts were visible after the scour, but these were severely
damaged and recovered more slowly (see also McCook and Chapman, 1992; Minchinton et al., unpubl. data, found similar patterns elsewhere at Sandy Cove). Although densities
were patchy (bimodal frequencies), recovery was generally slower, and comparable to that seen in the absence of regeneration (McCook and Chapman, 1992).
Overall, it seems that more severe damage resulted in less vegetative regeneration,
and slower recovery of canopy at Sandy Cove and Ketch Harbour, compared to Cranberry and Polly Coves (Fig. 5A Fig. 6A). Vegetative regeneration was clearly
important at the western locations, but apparently made only a small contribution to canopy recovery at the two eastern locations.
3.1.4. Correlations between species abundances
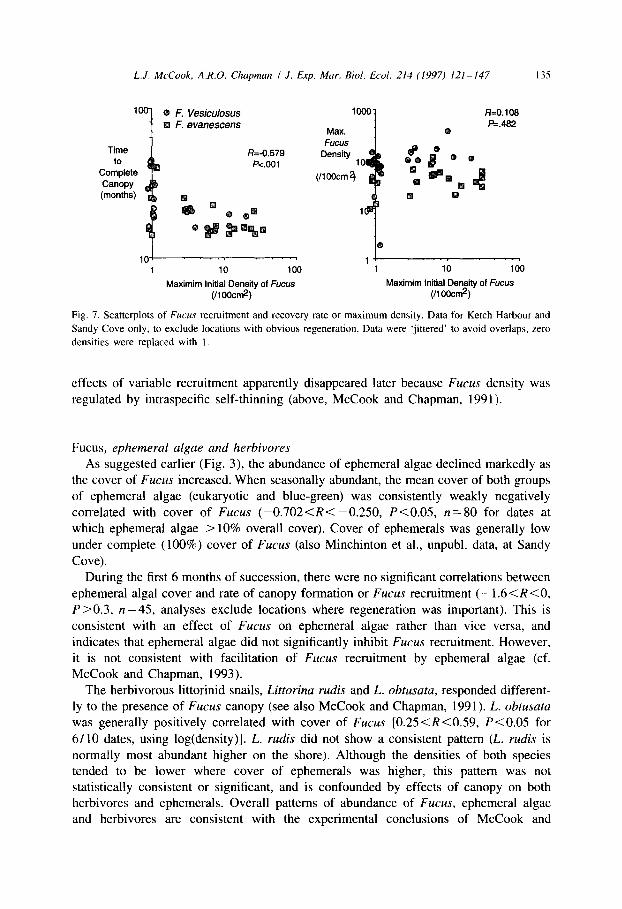
Effects of Fucus recruitment
Initial recruitment of Fucus was significantly correlated with rate of canopy formation (on a log/log scale) but had no significant effects on maximum density (Fig. 7), or final cover or density of Fucus (data not shown; density refers to fronds not plants). The
L.J. McCook, A.R.O. Chapman I J. Exp. Mar. Biol. Ecol. 214 (1997) 121-147 135
1 @ F. Vesiculosus F. evanescens
R=0.109 /=+I82
Time l0
Complete Canopy (months)
FUCUS
R-O.579 Density PC.001 1
(/l OOcm 3
1 10 100 1 10 100
Maximim Initial Density of Fucus Maximim Initial Density of Fucus (/lOOCmq (H OOcn?)
Fig. 7. Scatterplots of Fucus recruitment and recovery rate or maximum density. Data for Ketch Harbour and
Sandy Cove only, to exclude locations with obvious regeneration. Data were ‘jittered’ to avoid overlaps, zero
densities were replaced with I.
effects of variable recruitment apparently disappeared later because Fucus density was regulated by intraspecific self-thinning (above, McCook and Chapman, 1991).
Fucus, ephemeral algae and herbivores
As suggested earlier (Fig. 3), the abundance of ephemeral algae declined markedly as
the cover of Fucus increased. When seasonally abundant, the mean cover of both groups
of ephemeral algae (eukaryotic and blue-green) was consistently weakly negatively correlated with cover of Fucus (-0.702<R< -0.250, P<O.O5, n=80 for dates at
which ephemeral algae > 10% overall cover). Cover of ephemerals was generally low
under complete (100%) cover of Fucus (also Minchinton et al., unpubl. data, at Sandy
Cove). During the first 6 months of succession, there were no significant correlations between
ephemeral algal cover and rate of canopy formation or Fucus recruitment (- 1.6<R<O,
P>O.3, n =45, analyses exclude locations where regeneration was important). This is
consistent with an effect of Fucus on ephemeral algae rather than vice versa, and
indicates that ephemeral algae did not significantly inhibit Fucus recruitment. However, it is not consistent with facilitation of Fucus recruitment by ephemeral algae (cf. McCook and Chapman, 1993).
The herbivorous littorinid snails, Littorina rudis and L. obtusata, responded different-
ly to the presence of Fucus canopy (see also McCook and Chapman, 1991). L. obtusata
was generally positively correlated with cover of Fucus [0.25<R<0.59, PcO.05 for
6/10 dates, using log(density)]. L. rudis did not show a consistent pattern (L. rudis is
normally most abundant higher on the shore). Although the densities of both species tended to be lower where cover of ephemerals was higher. this pattern was not statistically consistent or significant, and is confounded by effects of canopy on both herbivores and ephemerals. Overall patterns of abundance of Fucus, ephemeral algae and herbivores are consistent with the experimental conclusions of McCook and
136 L.J. McCook, A.R.O. Chapman I J. Exp. Mar. Biol. Ecol. 214 (1997) 121-147
l F. vesiculosus I F. evanescens
% Cover of Fm~s
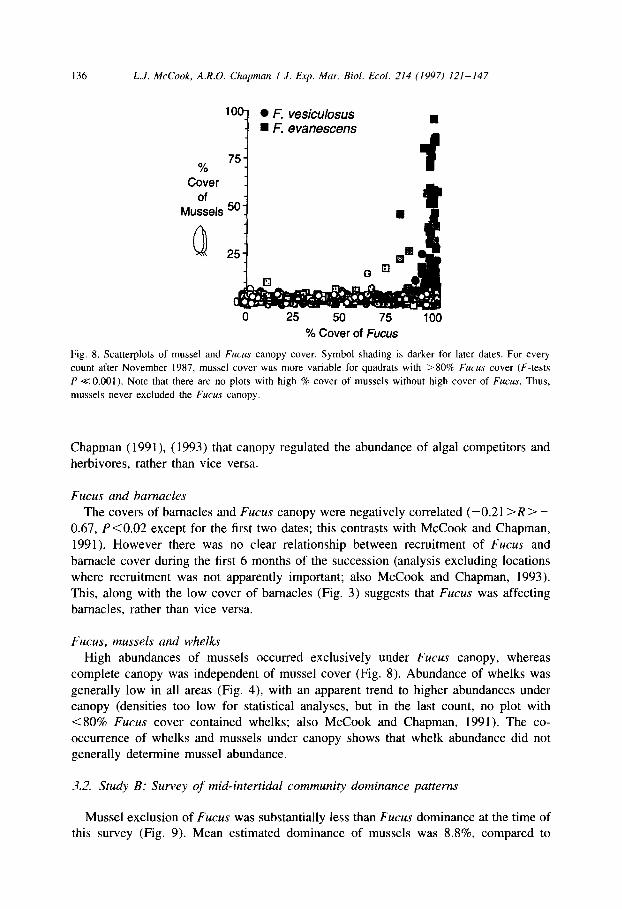
Fig. 8. Scatterplots of mussel and Fucus canopy cover. Symbol shading is darker for later dates. For every
count after November 1987, mussel cover was more variable for quadrats with >80% Fucus cover (F-tests
P i< 0.001). Note that there are no plots with high % cover of mussels without high cover of Fucus. Thus,
mussels never excluded the Fucus canopy.
Chapman ( 1991), (1993) that canopy regulated the abundance of algal competitors and herbivores, rather than vice versa.
Fucus and barnacles
The covers of barnacles and Fucus canopy were negatively correlated (-0.21 >R> -
0.67, PcO.02 except for the first two dates; this contrasts with McCook and Chapman,
1991). However there was no clear relationship between recruitment of Fucus and
barnacle cover during the first 6 months of the succession (analysis excluding locations
where recruitment was not apparently important; also McCook and Chapman, 1993). This, along with the low cover of barnacles (Fig. 3) suggests that Fucus was affecting
barnacles, rather than vice versa.
Fucus, mussels and whelks
High abundances of mussels occurred exclusively under Fucus canopy, whereas
complete canopy was independent of mussel cover (Fig. 8). Abundance of whelks was
generally low in all areas (Fig. 4), with an apparent trend to higher abundances under canopy (densities too low for statistical analyses, but in the last count, no plot with < 80% Fucus cover contained whelks; also McCook and Chapman, 1991). The co- occurrence of whelks and mussels under canopy shows that whelk abundance did not generally determine mussel abundance.
3.2. Study B: Survey of mid-intertidal community dominance patterns
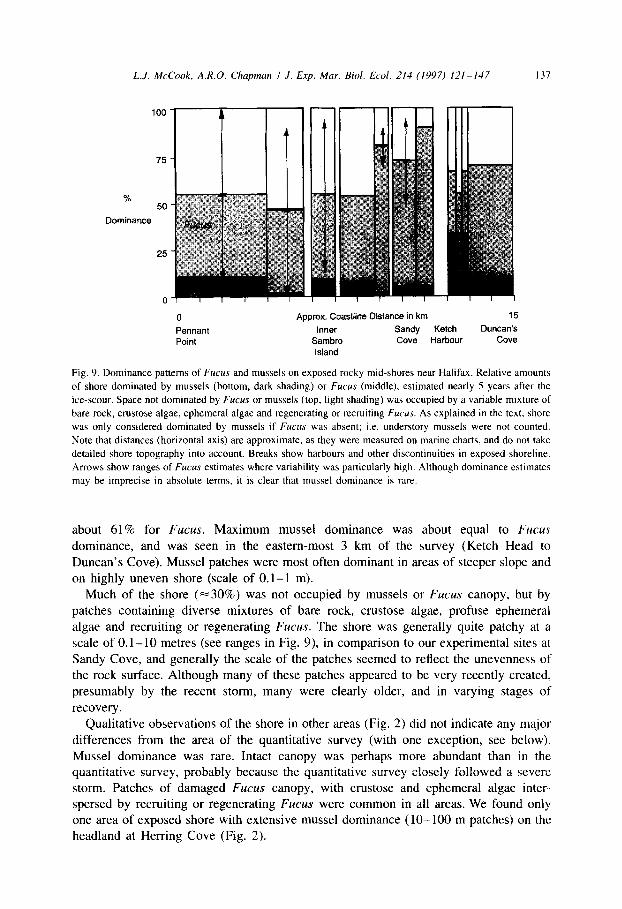
Mussel exclusion of Fucus was substantially less than Fucus dominance at the time of this survey (Fig. 9). Mean estimated dominance of mussels was 8.8%, compared to
L.J. McCook, A.R.O. Chapman I J. Exp. Mar. Biol. Ecol. 214 (1997) 121-147 137
%
Dominance
100
75
50
25
0
0
Pennant Point
Approx. Coastline Distance in km
Inner Sandy Ketch Sambro Cove Harbour Island
15
Duncan’s Cove
Fig. 9. Dominance patterns of Fucus and mussels on exposed rocky mid-shores near Halifax. Relative amounts
of shore dominated by mussels (bottom, dark shading) or Fucus (middle), estimated nearly 5 years after the
ice-scour. Space not dominated by Fucus or mussels (top, light shading) was occupied by a variable mixture of
bare rock, crustose algae, ephemeral algae and regenerating or recruiting Focus. As explained in the text, shore
was only considered dominated by mussels if Fucus was absent; i.e. understory mussels were not counted.
Note that distances (horizontal axis) are approximate, as they were measured on marine charts, and do not take
detailed shore topography into account. Breaks show harbours and other discontinuities in exposed shoreline.
Arrows show ranges of Fucus estimates where variability was particularly high. Although dominance estimates
may be imprecise in absolute terms, it is clear that mussel dominance is rare.
about 61% for Fucus. Maximum mussel dominance was about equal to Fucus dominance, and was seen in the eastern-most 3 km of the survey (Ketch Head to
Duncan’s Cove). Mussel patches were most often dominant in areas of steeper slope and
on highly uneven shore (scale of 0. l-l m). Much of the shore (~30%) was not occupied by mussels or Fucus canopy, but by
patches containing diverse mixtures of bare rock, crustose algae, profuse ephemeral algae and recruiting or regenerating Fucus. The shore was generally quite patchy at a
scale of 0.1-10 metres (see ranges in Fig. 9), in comparison to our experimental sites at
Sandy Cove, and generally the scale of the patches seemed to reflect the unevenness of the rock surface. Although many of these patches appeared to be very recently created,
presumably by the recent storm, many were clearly older, and in varying stages of
recovery. Qualitative observations of the shore in other areas (Fig. 2) did not indicate any major
differences from the area of the quantitative survey (with one exception, see below). Mussel dominance was rare. Intact canopy was perhaps more abundant than in the quantitative survey, probably because the quantitative survey closely followed a severe
storm. Patches of damaged Fucus canopy, with crustose and ephemeral algae inter- spersed by recruiting or regenerating Fucus were common in all areas. We found only one area of exposed shore with extensive mussel dominance (lo-100 m patches) on the headland at Herring Cove (Fig. 2).

138 L.J. Me&ok, A.R.O. Chapman I J. Exp. Mar. Bid. Ed. 214 (1997) 121-147
4. Discussion
Integrating the large-scale observations in this paper with smaller scale experimental
studies on the same succession (McCook and Chapman, 1991, 1992, 1993 and Minchinton et al., unpubl. data) offers several insights into the structure of exposed rocky seashores in the North-west Atlantic. This section includes specific discussion of
the dominance survey (B, Section 4.1) and then a general review of three aspects of the
community structure: the rarity and scale of the disturbance; the generality and
variability of patterns and processes; and alternative interpretations of community
organization in terms of rockweed life-history traits instead of predator control of
competition.
4.1. Study B: Survey of mid-intertidal community dominance patterns
The results of this survey show that, 5 years after a major disturbance, mussels rarely excluded rockweeds on exposed, mid-intertidal seashores in this area. Our survey was
unlikely to underestimate mussel dominance, but even allowing a several-fold margin of
error would not indicate overall mussel dominance. Much of the shore is occupied by rockweed beds of varying size. These beds appear to be dynamic patches in various
stages of recovery from small-scale disturbances, such as storm waves. At Sandy Cove,
Fucus cover at experimental sites was more patchy than at the time of the experiments
(see McCook and Chapman, 1991, 1992, 1993, this paper Study A). Extensive,
uninterrupted Fucus beds may be somewhat limited in successional stage and geog-
raphy.
Greater abundance of mussels on steep and irregular shores may arise because the
irregularities provide attachment foci, or because greater wave impact and turbulence
restrict predators. The apparent relationship between heterogeneous shore topography, and patchiness in the rockweed beds may be due to increased turbulence on steep and uneven shores. Flat areas of shore may dissipate the force of waves more gradually,
making them less likely to remove canopy. It is significant that the only extensive
mussel beds found (Herring Cove) were near a large sewage outfall and a village which relies on septic systems, and are within Halifax Harbour.
Clearly the composition of this community is highly variable in space (scale of l-10
m or more; Fig. 9) and in time (scale of seasons to years, Fig. 3), and disturbance is an
important influence on community structure. Mussel exclusion of rockweeds is excep-
tional, and does not represent a general successional ‘climax’ on exposed shores in this
area. This survey also demonstrates the value of quantifying, even approximately, how well experimental sites serve as representative samples of general patterns.
4.2. General discussion: Community structure during natural succession after ice-
scour
4.2.1. Implications of the rarity, severity and large scale of the disturbance
The rarity, severity and spatial scale of this disturbance both heighten the value and limit the context of the descriptions. Our studies provide rare comparisons of processes

L.J. McCook, A.R.O. Chapman I .I. Exp. Mar. Bid. Ed. 214 (1997) 121-147 139
occurring on rocky shores over scales of 1- lo4 m. The succession served as a valuable
test of the application of previous explanations of community structure (Section 1) to
large-scale and unusual circumstances. Furthermore, such catastrophic disturbances may
have considerable consequences for long-term community composition (e.g. Wethey, 1985).
Nonetheless, our studies remain anecdotal accounts of an exceptional succession, involving a particular set of circumstances and environmental conditions. Similar results
should not be assumed for other apparently similar successions, since effects of factors such as extent, severity, patchiness, seasonality and timing of the disturbance, weather,
and species availability can all vary, often stochastically (Sousa, 1984b, 1985; McCook,
1994). Indeed, our studies demonstrate that the organization of this assemblage often
varies with specific circumstances.
The scale of the scour demonstrated both variations in small-scale processes (between
sites) and scale-dependent effects. Variations in small-scale patterns include the differ-
ences in rate of succession between sites (Results A), even under very similar
circumstances (e.g. Fig. 3, Ketch Harbour, cf. Sandy Cove; also variations in Fig. 9). Of
the scale-dependent effects, many can be explained by the effects of patch size and proximity of canopy on physical conditions, although some are confounded by the
correlation between severity and extent of disturbance (also Sousa, 1985). For example, growth of perennial algae was faster in smaller patches (Results A), probably due both to
the less severe damage in small patches, and to enhanced moisture from the adjacent
canopy; moisture availability was critical to Fucus growth (Results A). In very small
patches (< 1 m) canopy recovery may involve overgrowth by surrounding plants (pets.
obs.). Disturbance scale did not appear important to Fucus dispersal (Results A),
apparently because canopy recovery commonly involved regeneration (Results A;
McCook and Chapman, 1992). For ephemeral algae, barnacles and mussels, abundance was often reduced in smaller patches, apparently due to foraging by littorinids or whelks
from nearby canopy areas (Results A; McCook and Chapman, 1991). Generally low
herbivore abundance during the large-scale succession would also limit the importance of refuges from herbivores during Fucus recruitment (cf. Lubchenco, 1983).
4.2.2. Generality and variability in succession and community organization
The large scale of this succession also allows assessment of variations in small-scale
processes between locations and under varying biological and environmental conditions.
Importantly, the large-scale patterns of species abundances, and relationships between
them (Results A and Results B), demonstrate that many of our experimental conclusions
from Sandy Cove (McCook and Chapman, 1991, 1992, 1993) applied generally over a broad range of locations and disturbance intensities.
Both the descriptive and experimental studies found an essentially similar pattern of
succession over a wide variety of conditions. At every site there was a general pattern of seasonal ephemeral algae, replaced by self-thinning rockweeds with an understory of mussels, barnacles and crustose algae, whether canopy recovery involved recruitment or regeneration. Rockweed dominance, although dynamic, appears general and mussel dominance is rare. Importantly, many of the relationships and experimental effects noted during these studies disappeared after longer periods (- l-5 years, this paper, McCook

140 L.J. McCook, A.R.O. Chapman I J. Exp. Mar. Biol. Ecol. 214 (19973 121-147
and Chapman, 1991, 1992), suggesting that much of the variation in the system is temporary.
Nonetheless, both patterns and processes varied considerably in time and space during this succession, and between studies. Many of the experimental effects and species
interactions in our own and previous studies varied with scale of disturbance (see Section 4.2.1) and between guilds, circumstances and studies (Results A and Results B;
and Menge, 1976, 1991a,b; Lubchenco, 1983, 1986; Parker and Chapman, 1994;
Petraitis, 1987, 1990, 1991; Barker and Chapman, 1990; McCook and Chapman, 1991, 1992, 1993). This variability demands caution in the interpretation of general patterns of
succession (McCook, 1994) or community structure (e.g. Menge, 1976, 1991a,b;
Lubchenco, 1983, 1986) based on specific experiments.
4.2.3. The trees in the forest: Fucus life-history traits and rockweed canopy as major
influences on community structure
Our discussions of community structure have focused on interactions between
particular species’ properties and life-history traits under specific circumstances, since this perspective may be more useful in understanding variable species interactions than
the more general terms of competition/predation or inhibition/facilitation (Sousa, 1980;
McCook and Chapman, 1991; McCook, 1994; Underwood and Anderson, 1994; e.g.
ephemeral algae to Fucus transition, see Section 4.2.5).
We consider that three specific traits or properties of the rockweeds are especially
valuable to understanding Fucus’ role as a structuring influence on the whole
assemblage: (i) Fucus as a physical canopy; (ii) Fucus’ ‘supply-side’ ecology: recruitment levels and the ability of Fucus to regenerate vegetatively from damaged
tissues; and (iii) self-thinning during canopy formation (intraspecific competition).
The Fucus canopy dramatically alters the physical environment in the understory, shading competitors (intra- and inter-specific), providing shelter from dehydration, wave
battering and perhaps from thermal stress, and increasing the dimensionality of the physical space (McCook and Chapman, 1991; Reed and Foster, 1984; Duggins et al.,
1990). Many of the observed differences in successional patterns and interspecific
interactions can be explained by these physical properties of the canopy, and influences on its recovery. During early succession, proximity to canopy (patch size) influenced
abundance of most guilds (McCook and Chapman, 1991, 1993 this paper). During later
succession, shelter by the Fucus canopy appeared to generally influence the abundance of filter feeders, herbivores and carnivores; and shading by the canopy seemed to affect
ephemeral algae, and recruitment and regeneration and self-thinning of Fucus itself
(McCook and Chapman, 1991, 1992 this paper; Minchinton and Scheibling, 1993; also Menge, 1976; Grant, 1977).
The general importance of the fucoid canopy to community structure is not surprising. As canopy vegetation. the rockweeds dominate the biomass, productivity and physical structure of these communities, yet there have been few previous tests of the rockweed’s own role (cf. studies of subtidal or terrestrial forests).
4.2.4. The roles of FUCUS recruitment, regeneration and intraspec$c competition
Given the importance of the fucoid canopy to the organization of the community as a whole, the mechanism of Fucus recovery from disturbance is important to the processes

L.J. McCook, A.R.O. Chapman I J. Exp. Mar. Bid. Ed. Z/4 (1997) 121-147 141
of succession. Sources and consequences of variations in Fucus recruitment, discussed
in detail in McCook and Chapman (1993), also Section 4.2.5), may generally account
for some variation in natural community composition (this paper). Variations in
recruitment may be important even when propagule supply is plentiful (Vadas et al., 1990; McCook and Chapman, 1993; cf. Menge, 1991a).
The impact of variations in Fucus recruitment also varies because Fucus recovery from disturbance often involves vegetative regeneration. During the natural succession,
vegetative regeneration was the major mechanism of recovery for F. vesiculosus where
damage was not extreme, and is probably important after more common, wave-induced
canopy damage (Studies A and B of this paper, McCook and Chapman, 1992;
Minchinton et al., unpubl. data). Variations in extent of damage to rockweeds can explain
much of the variation in rate of canopy formation (see Results A and McCook and
Chapman, 1992). However, the significance of variations in recruitment or regeneration appears to
decline with time since the disturbance (Results A, McCook and Chapman, 1992)
probably because intraspecific competition and self-thinning play a major role in regulating Fucus abundance (Results A, McCook and Chapman, 1991; Menge and
Lubchenco, pers. comm.). Abundance of Fucus almost universally increased when canopy was incomplete or thinned, but density is apparently tightly regulated once the
canopy is complete (Results A, McCook and Chapman, 1991). Intraspecific regulation of canopy biomass seems a major influence on abundance of Fucus, and hence on the
rest of the assemblage.
4.2.5. Interspecific interactions and community organization
Herbivory and plant-plunt competition with ephemeral algae
Interactions with ephemeral algae did not appear to significantly affect Fucus during this succession (cf. Lubchenco, 1986; McCook and Chapman, 1993), although the
seasonal abundance of ephemeral algae was strongly inhibited by Fucus canopy (Results
A, McCook and Chapman, 1991, 1993; also Menge, 1976; Lubchenco, 1986; Chapman, 1989, 1990). There was no evidence that Fucus recovery, whether by recruitment or
regeneration, was inhibited by ephemeral algae, even in the absence of herbivores.
Interestingly, Fucus recruitment and regeneration, as well as ephemeral algae, were also
inhibited by Fucus canopy, apparently due to a combination of shading and herbivore protection by canopy. This suggests first that early success of the ephemeral algae was a
consequence of rapid colonization and growth alone, and not of inhibition of Fucus; and
second, that Fucus’ later success was due to some combination of facilitation of recruitment and/or Fucus’ greater longevity and height (which leads to shading).
In the terminology of Connell and Slatyer (1977), the ephemeral-Fucus transition
clearly varies between facilitation (McCook and Chapman, 1991), tolerance (Results A) and inhibition (e.g. Lubchenco, 1983, 1986). We interpret this variation as the interaction between life-history traits of Fucus and of the different ephemeral algae. For example, different groups and abundances of ephemeral algae would have different effects on the Fucus recruits’ vulnerability to wave exposure, dehydration and shading, and the importance of these effects will depend on circumstances such as location, topography,

142 L.J. McCook, A.R.O. Chapman I J. Exp. Mar. Biol. Ecol. 214 (1997) 121-147
weather and abundance of ephemeral algae and Fucus (Lubchenco, 1978, 1982, 1983,
1986; cf. McCook and Chapman, 1993). There was no evidence that herbivore control of ephemeral algae was important to
Fucus success during the early stages of either the natural succession (Results A) or experimental primary succession (McCook and Chapman, 1993). Herbivore densities
were generally low, as a consequence of the scale of the disturbance (and ephemeral
algae did not inhibit MUCUS). The role of herbivory later in this succession was not tested,
but may be more important since herbivore density seemed to increase with canopy formation (Fig. 4A; Lubchenco, 1983, 1986; Petraitis, 1983, 1987; Chapman and
Johnson, 1990; Parker and Chapman, 1994; discussion in McCook and Chapman, 1991,
1993).
4.2.6. Plant-animal interactions with barnacles
The inverse relationship between cover of barnacles and rockweeds during the natural succession (Study A) was probably due to canopy inhibition of barnacles, rather than
vice versa. Barnacle cover was generally low and had no apparent effect on Fucus
recruitment (Results A; McCook and Chapman, 1993), in contrast to previous results in
New England and Nova Scotia (Menge, 1976; Grant, 1977; Lubchenco, 1983; Petraitis,
1987, 1990; Chapman and Johnson, 1990). Canopy may have reduced barnacle
recruitment (Menge, 1976; Grant, 1977; Minchinton and Scheibling, 1993; in contrast to
McCook and Chapman, 1991) and/or sheltered predators (Menge, 1976; Grant, 1977;
McCook and Chapman, 1991; our Results A). Importantly, the results (Survey A) for the large-scale succession show that the conclusions from removal of canopy in small
patches (McCook and Chapman, 1991) were not general, but underestimated the importance of the canopy. Foraging by whelks from adjacent canopy may have confounded the experimental study. Interactions between rockweeds and barnacles
apparently vary with disturbance scale, and with recruitment and abundance of both
species.
4.2.7. Plant-animal competition with mussels, predation and wave exposure: An
alternative view
Mussels very rarely excluded Fucus, and rarely recruited except under Fucus canopy on exposed shores during this succession. This was despite low whelk abundance. We
have suggested that, at least during the first 2-5 years of succession, competitive
dominance by mussels does not represent the normal ‘climax’ on these shores, and that Fucus dynamics are predominantly controlled by intraspecific interactions rather than
interactions with mussels (this paper, McCook and Chapman, 199 1, 1993; Minchinton et al., unpubl. data). Frequent disturbance followed by rapid recovery of canopy, by recruitment or regeneration, may maintain this situation, since extensive mussel recruitment depended on the Fucus canopy, yet that canopy shelters the mussels’ major
predators, the whelks. Nonetheless, there are exceptions to this pattern and mussel dominated patches do
occur, often on particularly exposed headlands (Fig. 9; see also Menge, 1976). Menge ( 1976, 1978a; Menge and Sutherland, 1976) felt that these patches occurred because predation intensity was low at exposed sites. This released mussels to exclude the

L.J. McCook, A.R.O. Chapman I J. Exp. Mar. Bid. Ed. 214 (1997) 121-147 143
competitively inferior rockweeds. According to this view, the predator’s vulnerability to
waves is critical to the mussel-canopy interaction, and canopy is only removed in mussel
beds. However, in our .studies, mussels generally recruited under canopy, suggesting an
alternative interpretation of this pattern: If disturbances (waves) remove the canopy and
consequently the predators (whelks), then mussels may (or may not) persist or increase (McCook and Chapman, 1991). According to this view, it is the canopy, not the
predator, which is critically vulnerable to wave battering, and it is disturbance, not
competition, which removes the canopy. Predation may still regulate mussel abundance (but see Petraitis, 1995). This interpretation does not require that mussels are competi-
tively superior (or inferior) to Fucus at low levels of predation.
Neither hypothesis can serve as a general explanation of both Menge’s (1976) results
and our own. Our interpretation assumes that mussels are generally preceded by Fucus, as in our studies, but this was not the case during Menge’s experiments. Photographs of
Menge’s site at Pemaquid Point show that abundant recruitment and dominance by mussels occurred without prior occupation by Fucus (Fig. 4 in Menge and Farrell, 1989,
and pers. comm.). Clearly mussels could pre-empt space in those circumstances, and canopy shelter of predators is not relevant.
Similarly, Menge’s model cannot explain the rarity of mussel dominance in our results
(Figs. 8, 9 McCook and Chapman, 1991). Indeed mussels seemed to depend on the
canopy during recruitment, in contrast to Menge’s studies. Wave exposure in our study
areas is at least as severe as Pemaquid Point in the Gulf of Maine, since our study sites
are fully exposed to the Atlantic Ocean (pers. obs. at both areas; Figs. 1 and 2). So why would mussel patches be rare throughout our study area and predominate at
Herring Cove (Fig. 2 Survey B), and at Pemaquid Point during Menge’s work? Petraitis
(1995) has recently shown that growth rates are critical to the impacts of recruitment and mortality, including predation, on mussel cover. Slow growing mussels may fail to
increase coverage, even at low levels of mortality, whereas mussels in highly productive
areas may maintain dominance in the face of disturbance and predation. Mussels at Pemaquid Point are filtering the waters of the highly productive Gulf of Maine and Bay
of Fundy, in contrast to the more oligotrophic waters of Atlantic Nova Scotia. Petraitis’
(1995) results suggest that, in our area, mussels are unable to grow (and recruit) fast
enough to outweigh even small losses to predation or disturbance. Presumably, the
nutrient supplement from sewage at Herring Cove (Results B) enhances mussel growth sufficiently to outweigh mortality (a ‘not-so-natural experiment’). Oceanographic
differences between the Gulf of Maine/Bay of Fundy and Atlantic may also influence larval retention and supply.
At local and small scales, neither wave exposure nor water quality alone can explain the patterns of dominance. Often adjacent, similar patches or small promontories,
separated by a few metres, are differently dominated by Fucus canopy or mussels (in this area, pers. obs.), and canopy loss often does not lead to mussel dominance (Fig. 9). These patches probably represent small-scale variation in factors such as predation, recruitment/regeneration/immigration of both competitors (Minchinton and Scheibling,
1991), the order of occupancy, growth rates, and effectiveness of canopy shelter on different shore topographies (McCook and Chapman, 199 1).

144 L.J. McCook, A.R.O. Chapman I J. Exp. Mar. Biol. Ecol. 214 (1997) 121-147
Thus, different explanations of patterns of species abundance are relevant under different circumstances, from geographic scales to differences within a few metres.
While hardly novel (e.g. Gleason, 1926), this conclusion has important implications, particularly since the earlier work in New England has provided a basis for synthetic
explanations of community structure on these shores and in natural communities in
general (e.g. Menge and Sutherland, 1976, 1987; Begon et al., 1986). Our studies
demonstrate that these are not sufficient explanations of the structure of an assemblage with the same suite of species under different circumstances. Thus, their application as
general models of community structure should be treated with caution.
4.3. Conclusions
Rocky shore community structure during an exceptional, large-scale succession after
ice-scour was studied using broad-scale observation (this paper) and experimental manipulations (McCook and Chapman, 1991, 1992, 1993). The general pattern of
succession involved a profuse growth of ephemeral algae that was overgrown by a
canopy of Fucus rockweeds. Sessile invertebrates appeared at variable but generally low
abundances, and mussels were rarely abundant except as understory. Ephemeral algae
did not inhibit Fucus significantly (Results A and B, McCook and Chapman, 1991,
1993). The rate of succession was markedly influenced by qualitative and quantitative
variations in Fucus re-establishment. Variable recruitment influenced recovery, but was often overshadowed by vegetative regeneration from surviving fragments of holdfast
tissue. This regeneration is apparently an important mechanism of recovery from a wide variety of disturbances (Results A and B, McCook and Chapman, 1992, 1993).
The Fucus canopy was a major influence on abundances of other algae, sessile
invertebrates, herbivores and whelks, and on the recruitment, regeneration and growth/
self thinning of Fucus itself. Predation, herbivory and competition with sessile
invertebrates had no significant effects on Fucus or other species during experimental
primary succession, and competition with mussels was of minor importance during later natural succession (this paper, McCook and Chapman, 1991).
Taken together, these studies show that predator control of competition, between
Fucus rockweeds and ephemeral algae or mussels, was not an adequate explanation of the patterns of species abundance seen during this succession. Our results emphasize the importance of several properties and traits of FUCUS, as the most abundant and
structurally important species.
Acknowledgments
We are indebted to J. Skeat, for careful field assistance and for commenting on and
proof-reading the manuscript. The work benefited enormously from advice and comments from A. Metaxas, C. Beauchamp, S. Walde, R. Scheibling, D. Webb, T. Minchinton and T. Done. We thank P. Petraitis, B. Menge and an anonymous reviewer for valuable discussion and C. Bird for identification of samples and advice. Field

L.J. McCook, A.R.O. Chapman I J. Exp. Mar. Biol. Ecol. 214 (1997) 121-147 145
assistance by G. Croft, D. Paquet, V. Brzeski, S. Burton, A. Faucher, B. Gjetvaj, T.
Horton, P. Jones, T. Parker, M. Patterson, P. Skeat, I. Suthers, Z. Tooze and J. W. Guo is
sincerely appreciated. Thanks also go to W. Muschenheim for the loan of the toys. We
acknowledge the National Research Council Aquaculture Research Station at Sandy Cove, for access to the shore, and for their frequent warmth and assistance. The work
was funded by an NSERC grant (A.R.O.C.) and L.M. was supported by an I.W. Killam Memorial Predoctoral Fellowship and a Dalhousie University Graduate Fellowship.
Finally, L.M. thanks Cocoa, for lab assistance, field companionship and for providing perspective.
References
Barker, K.M. and A.R.O. Chapman, 1990. Feeding preferences of periwinkles among four species of Fucu,s.
Mar. Biol., Vol. 106, pp. 113-I 18.
Bedford Institute of Oceanography, 1987. State-of-the-ocean: March. Weekly Briejing, 24 April 1987.
Begon, M., J.L. Harper and C.R. Townsend, 1986. Ecology: Individuals, populations and communities,
Sinauer Associates, MA, 876 pp.
Buschmann, A.H., 1990. The role of herbivory and desiccation on early successional patterns of intertidal
macroalgae in southern Chile. J. Exp. Mar. Biol. Ecol., Vol. 139, pp. 221-230.
Chapman, A.R.O., 1989. Abundance of Fucus spiralis and ephemeral seaweeds in a high eulittoral zone:
Effects of grazers, canopy and substratum type. Mar. Biol., Vol. 102, pp. 565-572.
Chapman, A.R.O., 1990. Effects of grazing, canopy cover and substratum type on the abundances of common
specres of seaweeds inhabiting littoral fringe tide pools. Bar. Mar., Vol. 33, pp. 319-326.
Chapman, A.R.O. and C.R. Johnson, 1990. Disturbance and organization of macroalgal assemblages in the
Northwest Atlantic. Hydrobiologia, Vol. 192, pp. 77-l 21.
Connell, J.H., I.R. Noble and R.O. Slatyer, 1987. On the mechanisms producing successional change. Oikos,
Vol. 50, pp. 1366137.
Connell, J.H. and R.O. Slatyer, 1977. Mechanisms of succession in natural communities and their role in
community stability and organization. Am. Nat., Vol. 11 1, pp. I1 19-I 144.
Cousens, R., 1985. Frond size distributions and the effects of the algal canopy on the behaviour of
Ascophyllum nodosum (L.) Le Jolis. J. Exp. Mar. Biol. Ecol., Vol. 92, pp. 23 l-249.
Diamond, J., 1986. Overview: Laboratory experiments, field experiments and natural experiments. In,
Community Ecology, edited by J. Diamond and T.J. Case, Harper and Row, New York, pp. 3-22.
Dinsmore, R.P., 1972. Ice and its drift into the North Atlantic ocean. ICNAF Spec. Publ. No. 8, pp. 89-l 27.
Duggins, D.O., I.E. Eckman and Sewell, A.T. 1990. Ecology of understory kelp environments. II. Effects of
kelps on recruitment of benthic invertebrates. J. Exp. Mar. Biol. Ecol., Vol. 143, pp. 27-45.
Farrell, T., 1989. Succession in a rocky intertidal community: The importance of disturbance size and position
within a disturbed patch. J. Exp. Mar. Biol. Ecol.,Vol. 128, pp. 57-73.
Farrell, T.M., 1991. Models and mechanisms of succession: An example from a rocky intertidal community.
Ecol. Monogr., Vol. 61, pp. 95-l 13.
Foster, M.S., 1990. Organization of macroalgal assemblages in the Northeast Pacitic: The assumption of
homogeneity and the illusion of generality. Hydrobiologiu, Vol. 192, pp. 2 l-33.
Foster, MS. and W.P. Sousa, 1985. Succession. In, Handbook of Phycological Methods, Ecologiccrl Field
Methods: Macroalgae, edited by M.M. Littler and D.S. Littler, Cambridge Univ. Press, Cambridge, pp. 269-290.
Gleason, H.A., 1926. The individualistic concept of the plant association. Bull. Torrey Bot. Club, Vol. 53, pp. l-26.
Grant, W.S., 1977. High intertidal community organization on a rocky headland in Maine, USA. Mar. Biol.,
Vol. 44, pp. 15-25.

146 L.J. McCook, A.R.O. Chapman I 1. Exp. Mar. Biol. Ecol. 214 (1997) 121-147
Keser, M., R.L. Vadas and B.R. Larson, 1981. Regrowth of Ascophyllum nodosum and Fucus vesiculosus
under various harvesting regimes in Maine, USA. Bor. Mar., Vol. 24, pp. 29-38.
Lubchenco, J., 1978. Plant species diversity in a marine intertidal community: Importance of herbivore food
preference and algal competitive abilities. Am. Nar., Vol. 112, pp. 23-39.
Lubchenco, J., 1982. Effects of grazers and algal competitors on fucoid colonization in tide pools. J. Phycol.,
Vol. 18, pp. 544-550.
Lubchenco, J., 1983. Littorina and Fucus: Effects of herbivores, substratum heterogeneity and plant escapes
during succession. Ecology, Vol. 64, pp. 1 I l6- 1123.
Lubchenco, J., 1986. Relative importance of competition and predation: Early colonization by seaweeds in
New England. In, Communiry Ecology, edited by J. Diamond and T.J. Case, Harper and Row, New York,
pp. 5377555.
Lubchenco, J. and B.A. Menge, 1978. Community development and persistence in a low rocky intertidal zone.
Ecol. Monogr., Vol. 48, pp. 67-94.
Markham, W.E., 1980. Ice arias: Easrern Canadian seaboard, Canadian Government Publishing Centre, Hull,
Quebec, 96 pp.
McCook, L.J., 1994. Understanding ecological community succession: Causal models and theories, a review.
Vegetafio, Vol. 1 IO, pp. 115-147.
McCook, L.J. and A.R.O. Chapman, 1991. Community succession following massive ice-scour on an exposed
rocky shore: Effects of Fucu.s canopy algae and of mussels during late succession. J. Exp. Mar. Biol. Ecol.,
Vol. 154, pp. 137-169.
McCook, L.J. and A.R.O. Chapman, 1992,Vegetative regeneration of Fucus rockweed canopy as a mechanism
of secondary succession on an exposed rocky shore. Bar. Mar., Vol. 35, pp. 35-46.
McCook, L.J. and A.R.O. Chapman, 1993. Community succession following massive ice-scour on a rocky
intertidal shore: Recruitment, competition and predation during early, primary succession. Mar. Biof., Vol.
115, pp. 565-575.
McLachlan, J., L.C.-M. Chen and T. Edelstein, 1971. The culture of four species of Fucus under laboratory
conditions. Can. J. Bar., Vol. 49, pp. 1463-1469.
Menge, B.A., 1976. Organization of the New England rocky intertidal community: Role of predation,
competition and environmental heterogeneity. Ecol. Monogr., Vol. 46, pp. 355-393.
Menge, B.A., 1978a. Predation intensity in a rocky intertidal community: Relation between predator foraging
activity and environmental harshness. Oecologia, Vol. 34, pp. 1 - 16.
Menge, B.A., 1978b. Predation intensity in a rocky intertidal community: Effect of an algal canopy, wave
action and desiccation on predator feeding rates. Oecologia, Vol. 34, pp. 17-35.
Menge, B.A., 1991a. Relative importance of recruitment and other causes of variation in rocky intertidal
community structure. J. Exp. Mar. Biol. Ecol., Vol. 146, pp. 69-100.
Menge, B.A., 1991b. Generalizing from experiments: Is predation strong or weak in the New England rocky
intertidal. Oecologia, Vol. 88, pp. l-8.
Menge, B.A. and T.M. Farrell, 1989. Community structure and interaction webs in shallow marine hard-
bottom communities: Tests of an environmental stress model. Adv. Ecol. Res., Vol. 19, pp. 189-262.
Menge, B.A. and J.P. Sutherland, 1976. Species diversity gradients: Synthesis of the roles of predation,
competition and temporal heterogeneity. Am. Nat., Vol. 1 10, pp. 35 1-369.
Menge, B.A. and J.P. Sutherland, 1987. Community regulation: Variation in disturbance, competition and
predation in relation to environmental stress and recruitment. Am. Nat., Vol. 130, pp. 730-757.
Minchinton, T.E. and R.E. Scheibling, 1991. The influence of larval supply and settlement on the population
structure of barnacles. Ecology, Vol. 72, pp. 1867-1879.
Minchinton, T.E. and R.E. Scheibling, 1993. Free space availability and larval substratum selection as
determinants of barnacle [Semibulanus balanoides (L.)] population structure in a developing rocky intertidal
community. Mar. Ecol. Prog. Ser., Vol. 95, pp. 233-244.
Murray, S.N. and M.M. Littler, 1978. Patterns of algal succession in a perturbated marine intertidal
community. J. Phycol., Vol. 14, pp. 506-512.
Parker, T. and A.R.O. Chapman, 1994. Separating the grazing effects of periwinkles and amphipods on a
seaweed community dominated by Fucus disfichus. Ophelia, Vol. 39, pp. 75-91. Petraitis, P.S., 1983. Grazing patterns of the periwinkle and their effect on sessile intertidal organisms.
Ecology, Vol. 64, pp. 522-533.

L.J. McCook, A.R.O. Chupman I J. Exp. Mar. Biol. Ecol. 214 (1997) 121-147 147
Petraitis, P.S., 1987. Factors organizing rocky intertidal communities of New England: Herbivory and
predation in sheltered bays. J. Exp. Mar. Biol. Ecol., Vol. 109, pp. 117-I 36.
Petraitis, P.S.. 1990. Direct and indirect effects of predation, herbivory and surface rugosity on mussel
recruitment. Oecologia, Vol. 83, pp. 405-413.
Petraitis, P.S., 1991. Recruitment of the mussel Myrilus edulis L. on sheltered and exposed shores in Mame.
USA. .I. Exp. Mar. Biol. Ecol., Vol. 147, pp. 65-80.
Petraitis, P.S.. 1995. The role of growth in maintaining spatial dominance by mussels (Myrilus edulis). Ecology,
Vol. 76, pp. 1337-1346.
Reed, D.C. and MS. Foster, 1984. The effects of canopy shading on algal recruitment and growth in a giant
kelp forest. Ecology, Vol. 65, pp. 937-948.
Sousa, W.P., 1979. Experimental investigations of disturbance and ecological succession in a rocky intertidal
algal community. Ecol. Monogr., Vol. 49, pp. 227-254.
Sousa, W.P., 1980. The responses of a community to disturbance: The importance of successional age and
species life histories. Oecologia, Vol. 45, pp. 72-81.
Sousa, W.P., 1984a. Intertidal mosaics: Patch size, propagule availability and spatial variable patterns of
succession. Ecology, Vol. 65, pp. 1918-1935.
Sousa, W.P., 1984b. The role of disturbance in natural communities. Ann. Rev. Ecol. Sysr., Vol. 15. pp.
353-391.
Sousa, W.P., 1985. Disturbance and patch dynamics on rocky intertidal shores. In, The ecology of narurcrl
disturbance and parch dynamics, edited by S.T.A. Pickett and P.S. White, Academic, Orlando, pp. lOl- 124.
Turner, T., 1983. Facilitation as a successional mechanism in a rocky intertidal community. Am. Nat.,Vol. 121,
pp. 720-728.
Underwood, A.J. and M.J. Anderson, 1994. Seasonal and temporal aspects of recruitment and succession in an
intertidal estuarine fouling assemblage. J. Mar. Biol. Ass. U.K., Vol. 74, pp. 5633584.
Underwood, A.J. and P.S. Petraitis, 1993. Structure of intertidal assemblages in different locations: How can
local processes be compared? Species diversity in ecological communitirs: Historical and ,qeo,~ruphicaL
perspecfives, edited by R. Ricklefs and D. Schluter, Univ. Chicago Press, Chicago, pp. 39-S I
Vadaa, R.L., W.A. Wright and S.L. Miller, 1990. Recruitment of Ascophyllum nodosum: Wave action as it
source of mortality. Mar. Ecol. Prq. Ser.,Vol. 61, pp. 2633272.
Van Tamelen, P.G., 1987. Early successional mechanisms in the rocky intertidal: The role of direct and indirect
interactions. J. Exp. Mar. Biol. Ecol., Vol. 112, pp. 39-48.
Wethey, D.S., 1985. Catastrophe, extinction and species diversity: A rocky intertidal example. Ecology. Vol.
66, pp. 445-456.
Whitehead, P., 1987. Ice in Halifax Harbour. Unpuhl. Coasr Curd Reporr, Ref. No. 9106.8.
Wilkinson, L., 1989. Sysrat: The system for sfutistics (Macintosh version), Sysiat Inc., Evanston, IL, 638 pp.