patch structure in a tropical rocky shore community in brazil: a mosaic of successional states?
TRANSCRIPT

This article was downloaded by: [University of Glasgow]On: 05 May 2013, At: 19:56Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
OpheliaPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/smar19
Patch Structure in a Tropical Rocky Shore Communityin Brazil: A Mosaic of Successional States?K. R. Sgrott Sauer Machado a c , A. R. O. Chapman a d & R. Coutinho ba Department of Biology, Dalhousie University, Halifax, Nova Scotia, B3H 4Jl, Canadab IEAPM, Rua Kioto 253, 28910 Arraial do Cabo, Rio de Janeiro, Brazilc IEAPM, Rua Kioto 253, 28910, Arraial do Cabo, Brazild Institut für Meereskunde, Dusternbrooker Weg 20, 2300 Kiel 1, GermanyPublished online: 20 Feb 2012.
To cite this article: K. R. Sgrott Sauer Machado , A. R. O. Chapman & R. Coutinho (1992): Patch Structure in a Tropical RockyShore Community in Brazil: A Mosaic of Successional States?, Ophelia, 35:3, 187-195
To link to this article: http://dx.doi.org/10.1080/00785326.1992.10429926
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form toanyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug doses shouldbe independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims,proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly or indirectly inconnection with or arising out of the use of this material.

OPHELIA 35 (3): 187-195 Guly 1992)
PATCH STRUCTURE IN A TROPICALROCKY SHORE COMMUNITY IN BRAZIL:
A MOSAIC OF SUCCESSIONAL STATES?
K. R. Sgrott Sauer Machado!' a, A. R. 0. Chapman'. b & R. Coutinho2
'Departrnent of Biology, Dalhousie University, Halifax, Nova Scotia. B3H 4Jl, Canada2IEAPM, Rua Kioto 253,28910 Arraial do Cabo, Rio de Janeiro, Brazil
-Present address: IEAPM, Rua Kioto 253, 28910 Arraial do Cabo, BrazilbPresent address and address for reprints: Institut fUr Meereskunde,
Dusternbrooker Weg 20, 2300 Kiel 1, Germany
ABSTRACT
The community of benthic organisms in the lower zone of a rocky shore at Praia Rasa, Brazil is distinctly patchy in structure. We examined the hypothesis that two types of patch (a. red algal turfandb. colonial coelenterate) represent different temporal stages in the same successional trajectory. Wemade observations on a) the stability of patch boundaries, b) successional sequence in experimentalclearings within both patch types and c) species composition through time in undisturbed patches.In most cases, the coelenterate overgrew the algae at the patch boundaries, thus showing a competitive asymmetry. In the successional sequence, diatoms were followed by ulvoid species. Thereafter,within clearings in the algal patches, a red algal turf was once more established over one year. Incontrast, the mid successional period in clearings within the coelenterate patch was characterizedby very low cover of sessile organisms. Colonization by red algal turfs and by the coelenterate colonyis largely dependent on vegetative spread. Thus later succession is not predictable on the basis oftime alone. Supply-side processes are clearly of equal importance. Therefore it cannot be statedwithout qualification that patches of red algal turfs and colonial coelenterate at Praia Rasa are partof the same successional sequence.
INTRODUCTION
Community structure is described in terms of patterns in time and space of species abundances and distributions. Spatial distributions occur at many scales ofresolution. The smallest is the space occupied by a single organism or in which amobile organism spends its life; a patch is occupied by many individuals; a regionincludes more than one patch or population linked by dispersal. Finally, there isa biogeographic scale of resolution (Wiens et al. 1986). Our work is concernedwith patch scale structure on a rocky intertidal shore occurring in Brazil. Thecontrast between this shore in the south Atlantic Ocean and those in the cold/temperate waters of the north Atlantic is striking. In the cold waters of Nova Scotia
Dow
nloa
ded
by [
Uni
vers
ity o
f G
lasg
ow]
at 1
9:56
05
May
201
3

188 SCROTT SAUER MACHADO ET AL.
the vegetation is structured into clear belts across the tidal gradient. Each belt hasconsiderable homogeneity of canopy structure and, on similarly exposed shores,there is little variation in structure on scales of kilometres (pers. obs., A. Chapman). In contrast, within the recognizable universal belts on Brazilian shoresaround Cabo Frio (Rio de Janeiro state), there is considerable heterogeneity incanopy structure so that distinct patches are readily observable (Yoneshigue1985). These patches occur on a variety of spatial scales from centimetres tometres. We chose to work on a shore at Praia Rasa where patches are at the largestscale and are therefore tractable for experimental analysis. We should point outthat there is large regional variation in community organization and that there isno basis to generalize from our findings to other shores of Cabo Frio region andespecially not to tropical shores in general.
Our objective was to analyse the causes of patch structure that we observed.There are several hypotheses which should not be regarded as mutually exclusive.According to Sousa (1984), localized disturbance occurs on a variety of temporaland spatial scales. Following a disturbance, which clears space for colonization,a successional process ensues. During this process, the species composition of apatch changes greatly. It is possible that patch structure on the shore simplyrepresents spatial variability in the time since local disturbance occurred. If thisis the only process producing mosaic structure, we can expect that patches clearedsimultaneously will develop similar community structures at similar times. Conversely, clearances produced at different times should result in patches withdifferent structural configurations. It is this notion that we examine here.
It is also necessary to point out immediately that other factors can producepatchiness. For example, there may be spatial variability in the supply of larvalstages (Gaines & Roughgarden 1985). Or patches may be maintained by inertialcompetition (Dayton et al. 1984). The role of competition in patch organizationat Praia Rasa is treated separately (Sgrott Sauer Machado 1991).
The low intertidal zone at Praia Rasa is dominated by three types of perennialpatches occurring on meter scales. Patches of algal turf are dominated by Hypneacervicornis, H. musciformis and Acanthophora spicifera (hereafter Hypnea). The remaining patch types are dominated by sessile animals. One of these (Zoanthus sp.,a colonial coelenterate) was included in this study. In addition, there are largepatches at Praia Rasa covered by the casts of a sessile polychaete, Phragmatopoma(not included in this study). In our work we tested the hypothesis that patches ofHypnea and Zoanthus represent different parts of the same successional trajectory.That is to say, if the successional sequence is allowed to proceed through its fullcourse on a given area ofsubstrate, then this area will be colonized by Hypnea andZoanthus at different times.
The field work was carried out from the Instituto de Estudos do Mar Almirante Paulo Moreira inArraial do Cabo and we thank Commandant Emmanuel de Almeida Gama and his staff for their
Dow
nloa
ded
by [
Uni
vers
ity o
f G
lasg
ow]
at 1
9:56
05
May
201
3

PATCH ORGANIZATION ON A BRAZILIAN SHORE 189
assistance. We are particularly grateful to Drs. Y. Yoneshigue and J. Valentin without whom wecould not have completed the study. Marcelo Machado was extremely helpful in the field, in the organization of the data and in the statistical analysis. We thank him for his work. Data analysis wasdone in Halifax and there Laurence McCook was invaluable for his expert advice - again we arevery grateful to him.
Much of the funding was provided through grant A6497 from the Natural Sciences and Environmental Research Council of Canada. Additional funds came from the Comissao Interministerealde Recursos do Mar (CIRM) in Brazil. Scholarship money for KRSSM came in part fromCoordenacao de Aperfeir;oamento de Pessoal de Nivel Superior (CAPES) and Conselho Nacionalde Desenvolvimento Cientlfico e Tecnol6gico (CNPq). RC was supported by a grant from CNPq.We thank these funding agencies.
MATERIAL AND METHODS
Study site
Praia Rasa (22°44'S, 41°57'W) is ca. 150 km east of Rio de Janeiro. Most of thesubstratum in the intertidal zone is Precambrian gneiss and is exposed to the eastand southeast. The shore has a slight incline and the tidal amplitude varies between 0.8 and 1.0 m. The site is moderately sheltered. Water temperature rangesfrom 19-27°C and the area is clearly out of the cold water upwelling system of thenearby Cabo Frio region. Mean daily air temperatures range from to 16-28°C(Yoneshigue 1985).
Patch boundary stability
Patches of Hypnea and Zoanthus have sharp mutual boundaries. We checked thestability of these boundaries over one year. Eleven steel bolts were inserted intoholes drilled into the rock exactly at patch boundaries in November 1989. A Ryobi™gasoline powered hammer drill was used to make the holes. Over one yearwe made 3 sets ofobservations to determine whether either species had retreatedor advanced from the inserted bolts.
Succession
We made space available in each patch type so that the successional process couldproceed. This was done in summer and in winter in two Hypnea patches where wecleared all living organisms, except crustose coralline species, from 10 randomlyplaced 400 ern" experimental areas. Five similar areas were set up (summer only)in a Zoanthus patch. Clearing was done with scrapers and wire brushes. Wedeliberately did not sterilize the surface of the rock because this phenomenonrarely, if ever, occurs naturally.
Species succession was followed by observing changes in cover of sessile organisms in each experimental area.. Cover was determined by a point intercept technique using a 10x 10 cm quadrat with 30 randomly chosen points.
Dow
nloa
ded
by [
Uni
vers
ity o
f G
lasg
ow]
at 1
9:56
05
May
201
3

190 SCROTT SAUER MACHADO ET AL.
Table 1. Competitive overgrowth patterns at 11 points on boundaries separating patches of Hypnea(H) and Zoanthus (Z). Data in each cell indicate winner (H or Z) and amount of overgrowth (em).
When patch boundary remained unchanged, this is indicated by -.
Date Boundary point number
1 2 3 4 5 6 7 8 9 10 11March 1990 Z+1 - - H+2 Z+1 H+4 - Z +2 - Z+ 1June 1990 Z+4 - Z+1 Z+2 Z+1 Z + 2 - H+l Z+1 Z+1November 1990 H+2 Z+1 Z+2 H+l Z+2 H+l Z+1 Z+4 Z+2 Z+7
Mature communities
We observed changes in species abundances in mature patches of Hypnea andZoanthus. In each patch type, we recorded changes in cover (see above) throughtime ofsessile species in 5 or 10 randomly distributed 10 x 10 em quadrats. Theseobservations allowed comparisons to be made between successional and maturecommunity states.
RESULTS
Boundary interactions
At 10 of the 11 bolts installed at patch boundaries, one ofthe species overgrew theother and increased its range over one year of observations (Table 1). In mostcases of overgrowth (seven) Zoanthus was the winner.
Succession
Plots cleared in H y Pneapatches. The changes in species compositions in experimental plots cleared in summer and in winter are shown in Fig. 1. Data on diatomcover are not included, but these organisms were the first invaders in all plots,declining rapidly in cover as Enteromorpha species became dominant. It the plotscleared in summer, Enteromorpha declined after 4 weeks when Ulua invaded theunits (Fig. 1). After about 13 weeks, the cover of Ulua fell and perennial seaweedsincreased, predominantly Acanthophora, Hypnea and Laurencia. By the end of oneyear, these species had assumed complete cover.
The plots cleared in winter were observed for 18 weeks. The successional sequence diatoms - Enteromorpha - Uloa was common to plots cleared in summerand winter (Fig. 1). The rate of succession was somewhat slower in winter andperennial algal species achieved little cover.
Plots cleared in Z {Jant h us patch. Five plots were cleared in the summer and followedfor 40 weeks. The results were strikingly different from plots cleared in the Hypnea patches (Fig. 2). The early invasion of diatoms and Enteromorpha was similar.However, Giffordia sp. was much more successful than Enteromorphaduring weeks
Dow
nloa
ded
by [
Uni
vers
ity o
f G
lasg
ow]
at 1
9:56
05
May
201
3
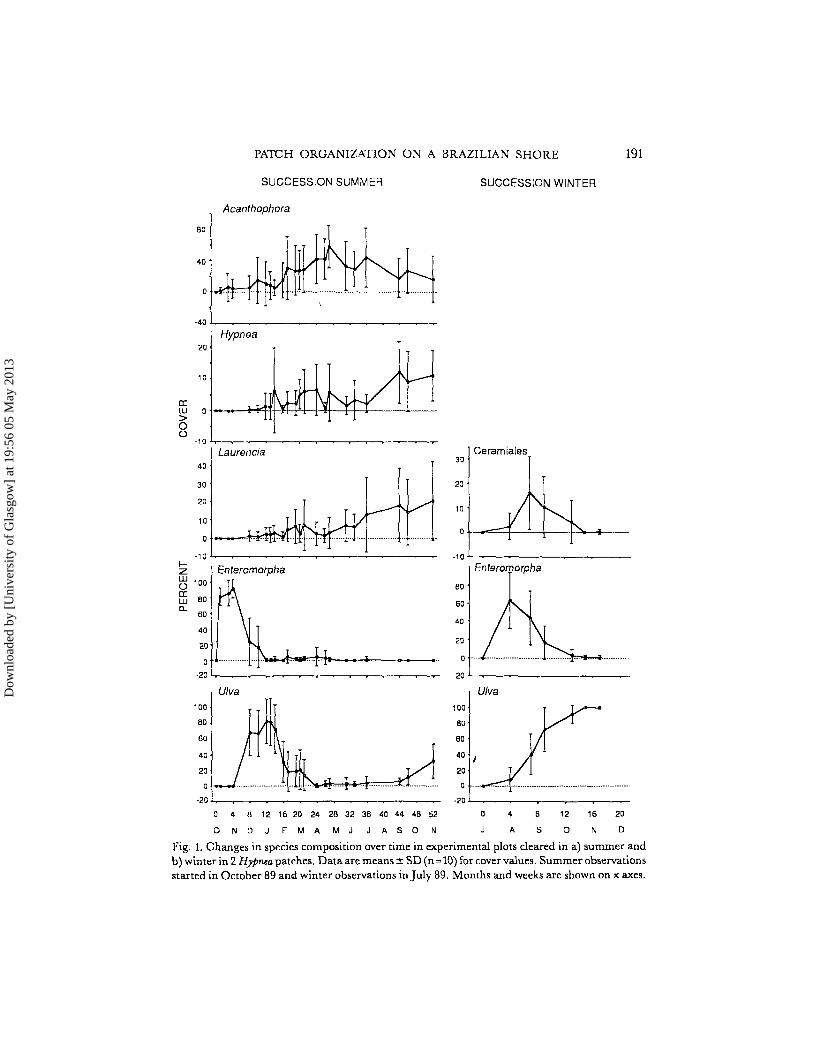
PATCH ORGANIZATION ON A BRAZILIAN SHORE 191
80
40
SUCCESSION SUMMER
Acanthophora
SUCCESSION WINTER
ONOJFMAMJJASON J AS 0 NO
Fig. 1. Changes in species composition over time in experimental plots cleared in a) summer andb) winter in 2 Hypnea patches. Data are means ± SD (n= 10) for cover values. Summer observationsstarted in October 89 and winter observations in July 89. Months and weeks are shown on x axes.
PATCH ORGANIZATION ON A BRAZILIAN SHORE 191
80
40
SUCCESSION SUMMER
Acanthophora
SUCCESSION WINTER
-40
Hypnea20
10
a:W>00
-10
Laurencia Ceramiales30
4[
3C 20
2C10
10a
-10 -10t-
Enreromorpha I Enteromorohazw lOa0 60a:
60w 60C-60
4040
2020
a-20 -20
'Ilv« UlvalOa 100
60 60
60 60
40 40
20 20
a a-20 -20
a 4 B 12 16 20 24 26 32 36 40 44 46 52 0 4 6 12 16 20
ONOJFMAMJJASON AS 0 NO
Fig. 1. Changes in species composition over time in experimental plots cleared in a) summer andb) winter in 2 Hypnea patches. Data are means ± SD (n= 10) for cover values. Summer observationsstarted in October 89 and winter observations in July 89. Months and weeks are shown on x axes.
Dow
nloa
ded
by [
Uni
vers
ity o
f G
lasg
ow]
at 1
9:56
05
May
201
3
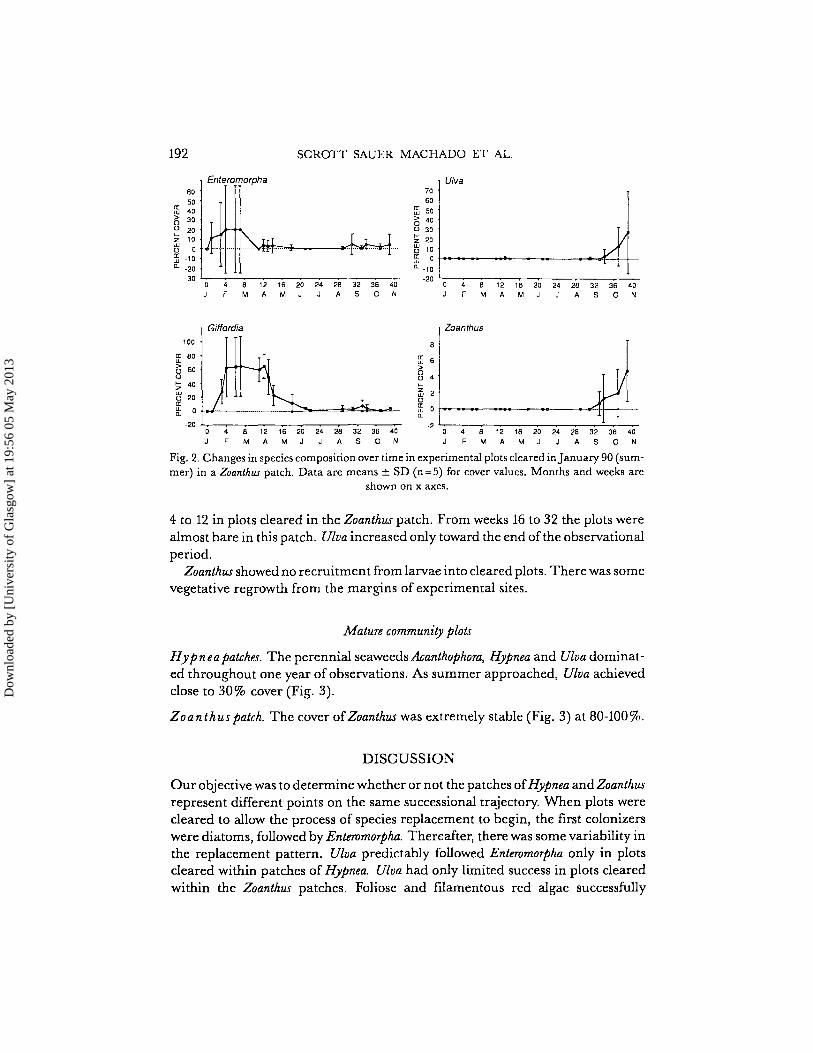
192 SCROTT SAUER MACHADO ET AL.
Enteromorpha Ulva
o 4 8 12 16 20 24 28 32 35 40J FMAMJ JASON
.. ¥t4 8 12 16 20 24 28 32 36 40F M A M J J A S 0 N
7060
ffi 50> 40
8 30
!z 20
t~ I:: :"-20 _
6050
ffi 40Ei 30o 20~ 10
~ affi -10
u, -20 I ---30
Giffordia Zoanthus
-2 ! ...
4 8 12 15 20 24 28 32 36 40
FMAMJJASON
100
c: 80wEi 50o~ 40wo 20c:
~ _2~j:1 1== i ~. A
o 4 8 12 15 20 24 28 32 36 40
J FMAMJJASON
c:w>o" 4>-~ 2
"c:wc,
Jr'V1
Fig. 2. Changes in species composition over time in experimental plots cleared in]anuary 90 (summer) in a Zoanthus patch. Data are means ± SD (n=5) for cover values. Months and weeks are
shown on x axes.
4 to 12 in plots cleared in the Zoanthus patch. From weeks 16 to 32 the plots werealmost bare in this patch. Uloa increased only toward the end ofthe observationalperiod.
Zoanthus showed no recruitment from larvae into cleared plots. There was somevegetative regrowth from the margins of experimental sites.
Mature community plots
Hyp ne a patches. The perennial seaweeds Acanthophora, Hypnea and Ulua dominated throughout one year of observations. As summer approached, Ulua achievedclose to 30% cover (Fig. 3).
Zo anth us patch. The cover of Zoanthus was extremely stable (Fig. 3) at 80-100%.
DISCUSSION
Our objective was to determine whether or not the patches ofHypnea and Zoanthus
represent different points on the same successional trajectory. When plots werecleared to allow the process of species replacement to begin, the first colonizerswere diatoms, followed by Enteromorpha. Thereafter, there was some variability inthe replacement pattern. Uloa predictably followed Enteromorpha only in plotscleared within patches of Hypnea. Uloa had only limited success in plots clearedwithin the Zoanthus patches. Foliose and filamentous red algae successfully
Dow
nloa
ded
by [
Uni
vers
ity o
f G
lasg
ow]
at 1
9:56
05
May
201
3

PATCH ORGANIZATION ON A BRAZILIAN SHORE
OBSERVATIONS IN MATURE HYPNEA PATCHES
193
Acanthophora
M
oc~w~~u~~
~20
u 10 !~ 0 j: : : 1 ::
-10o 4 8 12 16 20 24 28 32 36 40 44 48 52
o N 0 J F M A M J J A SON
Hypnea
80
ocw 60>ou 40~
~ 20o
~ 0 I I I • I
-20 ,--,-~~----~---~~-~
o 4 8 12 16 20 24 28 32 36 40 44 48 52o N D J F M A M J J A SON
Laurencia
30OCW>815czwu 0ocwCL
-15 ,--,~~~~~_~~~~~~ _
o 4 8 12 16 20 24 28 32 36 40 44 48 52
ONDJFMAMJJASON
Ulva
50
rr 40w>030o~ 20wo 10OC I~ JI~ Ott·············-·I-··~,
·10 ...' ~__~ ~~~ _
o 4 8 12 16 20 24 28 32 36 40 44 48 52
ONDJ FMAMJ JASON
OBSERVATIONS IN A MATURE ZOANTHUSPATCH
Zoanthus 1Ulva
OC 100 OC 15w w> >o 80 0o o 7.5~ 60 ~
z
o j. +w w~ 40 o
OC .W WCL 20 CL
I f I -7.50 4 8 1216202428323640 0 4 8 12 16 20 24 28 32 36 40J F M A M J J A S ON J F M A M J J A S ON
Fig. 3. Species composition over 40-52 mo in mature patches of a) Hypnea and b)Zoanthus. Data are means ± SD (n =5 for Zoanthus and n =10 for Hypnea) of percentcover values. Observations started in October 90 in Hypnea patches and January
90 in Zoanthus patch. Months and weeks are shown on x axes.
colonized cleared plots at various times following the fall in Enteromorpha abundance.
Our observations of succession on cleared plots over one year do not provideclear evidence that Zoanthus replaces the suite of red algal genera which we havegrouped under Hypnea. Probably, the observation period was insufficiently long.Nevertheless, our observations on the stability of patch boundaries show that thecoelenterate most frequently expands its borders by growing over the red algae.
Dow
nloa
ded
by [
Uni
vers
ity o
f G
lasg
ow]
at 1
9:56
05
May
201
3

194 SCROTT SAUER MACHADO ET AL.
This is interpreted as evidence that in the long term, Zoanthus replaces Hypnea inthe successional sequence. It must be made clear that for both types of organisms,vegetative growth from colony perimeters represents the major mechanism forinvasion. Invasion by larvae and/or spores is extremely limited. This means thatthe late successional sequence depends on the proximity of adult organisms. Apparently, Enteromorpha and Ulva are able to invade by spores. Hence, the early sequence of succession is predictable.
The weight of evidence suggests that patches of Hypnea and Zoanthusdo indeedrepresent different temporal stages in the same successional sequence. Thus themosaic structure depends on the time since disturbance, but also on the slow rateof vegetative spread in both forms. Whereas it seems almost certain that diatomsand ulvoid algae will dominate early successional stages because of their abilityto disperse widely, it is by no means predictable that either Hypnea or Zoanthuswillfollow in the sequence. This is perhaps the most important point to emerge fromthe study. It means that the temporal structuring of the community is not deterministic.
Since vegetative supply processes are so important in the patch structuring atPraia Rasa, it cannot be concluded that mosaic organization depends solely ontime since disturbance cleared space for colonization. This is an example of theimportance of supply-side mechanisms (Underwood & Denley 1984) in controlling the structure of complex communities. We would emphasize that there aremany benthic species which invade mostly through vegetative regeneration orspread. This has been shown recently for Fucus (McCook & Chapman 1992) andis well known for Ascophyllum (Baardseth 1970). These 2 fucoid genera dominatethe biomass ofnorth Atlantic shores and, for this reason alone, it seems importantto begin studying supply-side processes based on vegetative growth.
Results from clearings in the Zoanihus patch (Fig. 2) provide some evidence fora possible poisoning of the substratum by this coelenterate. This phenomenonhas been observed before (Kennelly 1987). Only limited colonization occurred,even among ephemeral species. This contrasted strongly with results from theplots cleared in the Hypneapatches (Fig. 1). We have no knowledge of the mechanisms involved here. But it is worth stressing that this little known "carry-over"poisoning effect could have great importance in benthic structuring importanceif it occurs widely.
Finally, it should be stressed here that, in the tropical system we chose to workin, competition for space among sessile species appears to be of considerable importance as a community structuring force. This observation contrasts stronglywith work in the Bay ofPanama where interspecific competition for space on theshore is suggested to be irrelevant (Menge & Lubchenco 1981, Menge et al. 1983,1985, 1986, Lubchenco et al. 1984). Because of the lack of concordance, we alsoundertook an experimental analysis to rank competition and predation as controlling factors in species abundances (Sgrott Sauer Machado 1991).
Dow
nloa
ded
by [
Uni
vers
ity o
f G
lasg
ow]
at 1
9:56
05
May
201
3

PATCH ORGANIZATION ON A BRAZILIAN SHORE
REFERENCES
195
Baardseth. E., 1970. A synopsis of biological data on knobbed wrack, Ascophyllum nodosum (Linn.)LeJolis. - FAa Fisheries Synopsis No. 38. 41 pp.
Dayton, P. K., V. Currie, T. Gerrodete, B. D. Keller, R. Rosenthal & D. Van Tresca, 1984. Patchdynamics and stability of some California kelp communities. - Ecol. Monogr. 54: 253-289.
Gaines, S. D. &j. R. Roughgarden, 1985. Larval settlement rate: a leading determinant of structure in an ecological community of the marine intertidal zone. - Proc. natn. Acad. Sci. USA 82:3701-3711.
Kennelly, S. j., 1987. Inhibition of kelp recruitment by turfing algae and consequences for an Australian kelp community. - j. expo mar. BioI. Ecol. 68: 257-276.
Lubchenco,j., B. A. Menge, S. D. Garrity, Pj. Lubchenco, L. R. Ashkenas, S. D. Gaines, R. Emlet,j. Lucas & S. Strauss, 1984. Structure, persistence, and role of consumers in a tropical intertidal community (Taboguilla Island, Bay of Panama). - j. expo mar. BioI. Ecol. 78: 23-73.
McCook, L. j. & A. R. O. Chapman, 1992. Mechanisms of primary and secondary succession onan exposed rocky shore: vegetative regeneration of Fucus rockweed canopy. - Bot. Mar. .35:35-46.
Menge, B. A., L. R. Ashkenas & A. Matson, 1983. Use of artificial holes in studying commuity development in cryptic marine habitats in a tropical intertidal region. - Mar. BioI. 77: 129-142.
Menge, B. A. &j. Lubchenco, 1981. Community organization in temperate and tropical rocky intertidal habitats: prey refuges in relation to consumer pressure gradients. - EcoL Monogr. 51:429-450.
Menge, B. A, j. Lubchenco & L. R. Ashkerias, 1985. Diversity, heterogeneity and consumer pressure in a tropical intertidal community. - Oecologia 65: 394-405.
Menge, B. A, j. Lubchenco & L. R. Ashkenas, 1986. Experimental separation of effects of consumers on sessile prey in the low zone of a rocky shore in the Bay of Panama: direct and indirectconsequences of food web complexity. - j. expo mar. BioI. Ecol. 100: 225-269.
Sgrott Sauer Machado, K., 1991.The role of post-settlement predation and competition in a tropical intertidal region at Praia Rasa, Buzios - Rio de Janeiro, Brazil. - M.sc. thesis, DalhousieUniv. 102 pp.
Sousa, W. P., 1984. Intertidal mosaics: patch size, propagule availability, and spatially variable patterns of succession. - Ecology 65: 1918-1935.
Underwood, A. j. & E. Denley, 1984. Marine community paradigms. -In D. R. Strong, D. S. Simberloff, L. G. Arele & A B. Thistle (eds): Ecological Communities: Conceptual Issues and theEvidence, pp. 151-180. Princeton University, Princeton.
Wiens, j. A, j. F. Addicott, T. j. Case & j. Diamond, 1986. Overview: the importance of spatialand temporal scale in ecological investigations. - In j. Diamond & T. j. Case (eds): CommunityEcology, pp. 145-153. Harper and Row, New York.
Yoneshigue, Y, 1985. Taxonomie et Ecologie des Algues Marine dans la Region de Cabo Frio (Riode Janeiro, Bresil) - Ph. D. thesis, Universite de Marseille. 466 pp.
Dow
nloa
ded
by [
Uni
vers
ity o
f G
lasg
ow]
at 1
9:56
05
May
201
3