past and present initiatives on the biological control of
TRANSCRIPT

Past and present initiatives on the biologicalcontrol of Lantana camara (Verbenaceae) in
South Africa
J-R. Baars & S. NeserARC - Plant Protection Research Institute, Private Bag X134, Pretoria, 0001 South Africa
Lantana camara, a highly invasive weed in many countries, has been targeted for biological
control in South Africa since the early 1960s. An earlier review in 1991 indicated that, despite the
establishment of several natural enemy species, the programme has largely been unsuccessful.
In this paper we review initiatives undertaken during the 1990s and discuss (i) the status of the
natural enemies established on the weed, (ii) factors that have limited the impact of these
agents, (iii) the potential of eleven new biocontrol candidates currently under evaluation for re-
lease and (iv) the problem of expanded host ranges of imported natural enemies under labora-
tory conditions. Ultimately, the success of the programme will depend on the establishment of a
suite of natural enemies, attacking several parts of the weed, which are able to cope with the ex-
treme variability and wide distribution of L. camara in South Africa. Despite the problems associ-
ated with the programme, L. camara remains a candidate for biological control in South Africa.
Key words: Lantana camara, varieties, biological weed control, natural enemies, host-specificity
testing, Lippia.
Lantana camara sensu lato (Verbenaceae; Fig. 1), afloriferous, prickly, thicket-forming shrub, whichis commonly known as lantana, originates fromtropical and subtropical South and Central Amer-ica (Stirton 1977). As a popular ornamental plant,several varieties (cultivars) of lantana have beenwidely distributed throughout the tropics, sub-tropics and warm temperate regions of the world.Following its worldwide introduction, lantana hasbecome naturalized in some 50 countries and israted as one of the world’s worst weeds (Holmet al. 1977).
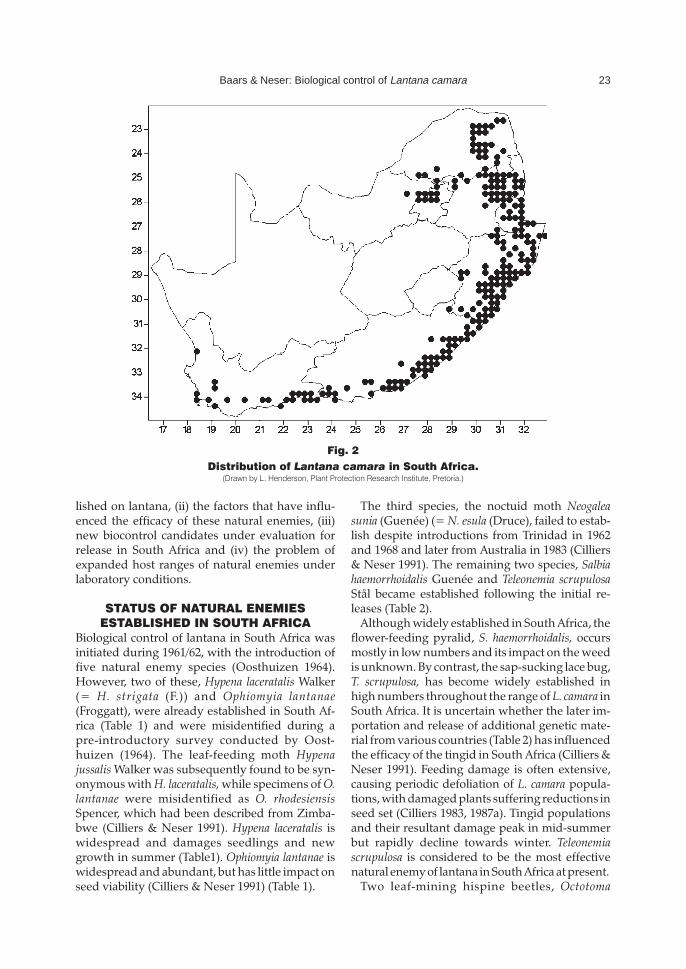
Lantana is an aggressive, vigorously growingweed that tolerates a wide variety of environmen-tal conditions, but thrives better in humid than indry regions. In South Africa it is presently natural-ized in the warm, moist subtropical and temperateregions (Oosthuizen 1964; Stirton 1977) of theNorthern Province, Gauteng, Mpumalanga andKwaZulu-Natal, as well as the southern coastal re-gions of the Eastern and Western Cape Provinces(Fig. 2). Lantana invades river-banks, mountainslopes and valleys, pastures and commercialforests where it forms impenetrable stands thatobstruct access and utilization. Through allelo-pathic suppression of indigenous plant species,lantana invasions also interrupt regenerationprocesses (Gentle & Duggin 1997a) and reduce thebiodiversity of natural ecosystems.
Lantana leaves, stems and fruits contain toxic
compounds, notably the pentacyclic triterpenes(Kellerman et al. 1996), lantadene A and B (Morton1994), which if consumed can cause photosensiti-zation, liver and kidney damage, paralysis of thegall bladder, intestinal haemorrhage and deathwithin 1–4 days in cattle, sheep and horses. Live-stock previously exposed to lantana are less likelyto suffer from the acute symptoms caused byingestion, while those with no previous exposureare likely to be severely affected. The expectedannual impact of cattle mortalities from lantanapoisoning in South Africa was estimated to be inexcess of R1.7 million (Kellerman et al. 1996).
Lantana is an extremely variable entity that pres-ents a complex taxonomic problem (Munir 1996).Its conspicuous variability in general morphology(Howard 1970; Smith & Smith 1982; Cilliers 1987a;Munir 1996), physiology and genetic composition(Spies & Stirton 1982a,b,c; Spies 1984; Spies & DuPlessis 1987), has led to the recognition of hun-dreds of cultivars or varieties, all of which are clas-sified as L. camara. The weed is a polyploidcomplex, presumably derived from the deliberatehybridization of species in the genus Lantana(Stirton 1977) and, subsequent to naturalization inSouth Africa, hybridization was shown to con-tinue in the field (Spies & Stirton 1982a,b,c). Spies(1984) argued that L. camara is in an active phase ofevolution, with intermediates in a transitionalstage of speciation. Although the species is consid-

ered to be unstable, certain varieties dominate inlocalized areas and remain relatively stable inspace and time. Following ecological disturbances,lantana stands may recolonize as a mixture of vari-eties, but soon revert to the former dominant con-dition (Spies & Du Plessis 1987).
Chemical and mechanical control of lantana wasreviewed by Cilliers & Neser (1991) who con-cluded that, although very effective, these meth-ods are often labour-intensive and expensive.These control measures usually offer only tempo-rary relief unless continual follow-up treatmentsare used. Controlled low- to moderate-intensityfires appear to reduce invasions by lantana, andcan be an effective, preventative management
strategy (Gentle & Duggin 1997b). However, theuse of fires is not always a suitable option as infes-tations are often close to, or in, indigenous forests,grazing lands and plantations. The difficulties andexpenses incurred by conventional control mea-sures have fostered the hope that biological con-trol may provide a solution.
The biological control programme, includingpreviously introduced natural enemies on L.camara in South Africa, was reviewed by Cilliers &Neser (1991), while other aspects of the weed werereviewed by Swarbrick et al. (1995) and Munir(1996). In this paper we mainly review biocontrolinitiatives undertaken during the 1990s, notably (i)the status of the natural enemies previously estab-
22 African Entomology Memoir No. 1 (1999): 21–33
Lantana camara.(Drawn by R. Weber, National Botanical Institute, Pretoria.)
50mm
lished on lantana, (ii) the factors that have influ-enced the efficacy of these natural enemies, (iii)new biocontrol candidates under evaluation forrelease in South Africa and (iv) the problem ofexpanded host ranges of natural enemies underlaboratory conditions.
STATUS OF NATURAL ENEMIESESTABLISHED IN SOUTH AFRICA
Biological control of lantana in South Africa wasinitiated during 1961/62, with the introduction offive natural enemy species (Oosthuizen 1964).However, two of these, Hypena laceratalis Walker(= H. strigata (F.)) and Ophiomyia lantanae(Froggatt), were already established in South Af-rica (Table 1) and were misidentified during apre-introductory survey conducted by Oost-huizen (1964). The leaf-feeding moth Hypenajussalis Walker was subsequently found to be syn-onymous with H. laceratalis, while specimens of O.lantanae were misidentified as O. rhodesiensisSpencer, which had been described from Zimba-bwe (Cilliers & Neser 1991). Hypena laceratalis iswidespread and damages seedlings and newgrowth in summer (Table1). Ophiomyia lantanae iswidespread and abundant, but has little impact onseed viability (Cilliers & Neser 1991) (Table 1).
The third species, the noctuid moth Neogaleasunia (Guenée) (= N. esula (Druce), failed to estab-lish despite introductions from Trinidad in 1962and 1968 and later from Australia in 1983 (Cilliers& Neser 1991). The remaining two species, Salbiahaemorrhoidalis Guenée and Teleonemia scrupulosaStål became established following the initial re-leases (Table 2).
Although widely established in South Africa, theflower-feeding pyralid, S. haemorrhoidalis, occursmostly in low numbers and its impact on the weedis unknown. By contrast, the sap-sucking lace bug,T. scrupulosa, has become widely established inhigh numbers throughout the range of L. camara inSouth Africa. It is uncertain whether the later im-portation and release of additional genetic mate-rial from various countries (Table 2) has influencedthe efficacy of the tingid in South Africa (Cilliers &Neser 1991). Feeding damage is often extensive,causing periodic defoliation of L. camara popula-tions, with damaged plants suffering reductions inseed set (Cilliers 1983, 1987a). Tingid populationsand their resultant damage peak in mid-summerbut rapidly decline towards winter. Teleonemiascrupulosa is considered to be the most effectivenatural enemy of lantana in South Africa at present.
Two leaf-mining hispine beetles, Octotoma
Baars & Neser: Biological control of Lantana camara 23
Distribution of Lantana camara in South Africa.(Drawn by L. Henderson, Plant Protection Research Institute, Pretoria.)

scabripennis Guérin-Méneville and Uroplata girardiPic, released in the early 1970s (Cilliers & Neser1991), have become established and coexist in themoist, subtropical areas of the country (Table 2).Uroplata girardi, although present in Mpumalangaand the Northern Province, is most successful inthe coastal regions of KwaZulu-Natal Province,where it is widely established and reaches highpopulation densities. By contrast, O. scabripennis ismost abundant at inland sites, where populationspeak infrequently in localized areas. Populationsof both species reach damaging levels duringmid-summer and often cause the defoliation oflantana stands (Cilliers 1987b), but the plants mayrecover rapidly. Although a third leaf-miningbeetle, Octotoma championi Baly, failed to establishafter releases in 1978 (Cilliers & Neser 1991), fur-ther releases were made at an inland, low altitudesite near Nelspruit (Mpumalanga Province) in1995. The population persisted in low numbers forthe following two seasons, but its present status isunknown (Table 2).
The leaf-mining fly, Calycomyza lantanae (Frick),released in 1982 (Cilliers & Neser 1991) has also be-come widespread (Table 2). Populations initiallyestablished in the warm, subtropical regions andsubsequently became widely established in thetemperate regions. The blotch mines initiated bythe flies appear more abundant in the subtropicalregions, with most of the damage visible on ac-tively-growing plants, seedlings and coppicegrowth. Additional material was released in 1989
(Table 2) but its effect on the established popula-tions is unknown. The impact of C. lantanaepopulations on lantana infestations is unknown,but is reduced by the extensive larval parasitismobserved throughout the fly’s range.
Besides the three species already mentioned, afurther two moths were released in South Africabut were later found to have already been estab-lished (Table 1). Both were introduced via Hawaiiin 1984 (Cilliers & Neser 1991; Julien & Griffiths1998) and neither has inflicted appreciabledamage on the weed. Larvae of the plume mothLantanophaga pusillidactyla (Walker) feed on thebases of the flowers, binding them together. Feed-ing damage causes a typical streak of aborted flow-ers among the cluster, leaving the other flowers todevelop normally (Perkins & Swezey 1924).Larvae of the tortricid moth Epinotia lantana(Busck) attack the flowers, hollowing out thereceptacle, and also burrow into the young shoottips (Perkins & Swezey 1924).
Although never deliberately introduced, thetineid moth Cremastobombycia lantanella Busck iswidely established in South Africa (Table 1). Thelarvae initiate serpentine leaf mines, which laterdevelop into ‘blister ’mines, and later pupate inspindle-shaped, ribbed cocoons, which are sus-pended within the ‘blisters’. Populations areusually low, suffer heavy parasitism, and alsocause negligible damage to lantana. Moths be-lieved to be C. lantanella and L. pusillidactyla havebeen reared from different indigenous Lippia
24 African Entomology Memoir No. 1 (1999): 21–33
Status of natural enemies present on Lantana camara in South Africa prior to the initiation of thebiological control programme (‘generalists’ not included).
Order/Family Natural enemy species Mode of attack Status Limitations
Diptera
Agromyzidae Ophiomyia lantanae (Froggatt) Fruit miner Widely established and abundant Low impact on seed viability;heavy parasitism
Lepidoptera
Noctuidae Hypena laceratalis1 Walker Leaf chewer Widely established with considerable Larvae are only active during latedamage to seedlings and new growth. summer and autumn and are oftenAlso attacks native Lippia and Priva parasitizedspecies
Pterophoridae Lantanophaga pusillidactyla1 Flower, fruit and Widely established, but present in low Low abundance and possible(Walker) seed chewer numbers. Also attacks native Lippia high levels of parasitism
species
Tortricidae Epinotia lantana1 (Busck) Flower-peduncle Widely established Unknownand shoot-tip borer
Gracillariidae Cremastobombycia lantanella1 Leaf miner Widely established, but present in low Heavy parasitismBusck numbers. Attacks native Lippia species
1: scrutiny of identifications, also of specimens from indigenous plants, required.

species. It is possible that these two species andperhaps even H. laceratalis and E. lantana were in-advertently introduced into South Africa togetherwith lantana plants, although they may be indige-nous. The geographic range, correct identities andstatus of the different lepidopterans associated withlantana in South Africa are in need of revision.
FACTORS AFFECTING THE IMPACT OFESTABLISHED AGENTS
Despite the establishment of several natural ene-mies on L. camara in South Africa (Oosthuizen1964; Cilliers & Neser 1991), the biological controlprogramme in South Africa has had limitedsuccess. This has largely been attributed to thegenetic diversity of lantana, which has made it anextremely variable target weed. The diversity ofvarieties presents the natural enemies with severalmorphological and physiological barriers to utili-zation (Cilliers 1983; Neser & Cilliers 1990; Cilliers& Neser 1991). Indeed, the introduced complex ofL. camara is a man-made polyploid entity (Stirton1977), which is far removed from the parentspecies occurring in its native range. Introducednatural enemies are therefore poorly adapted tocope with the diversity of ‘new’ entities natural-
ized in the countries of introduction.Several natural enemies (e.g. T. scrupulosa and C.
lantanae) were reported to display preferences forcertain varieties of lantana (Radunz 1971; Harley& Kassulke 1974; Harley et al. 1979; Cilliers 1987b;Cilliers & Neser 1991). In some cases (e.g. E. xantho-chaeta), agents have reportedly failed to establishbecause of the phenomenon of varietal resistanceor reduced compatibility (Cilliers & Neser 1991).Agents from different isolated varieties of L.camara, or from other species such as L. tiliaefolia(Winder & Harley 1983), may thus all be consid-ered as ‘new’ associations when deployed againstthe plants in South Africa. Consequently, the inter-actions between the various natural enemies andthe different lantana varieties are complex anddifficult to predict.
Harley & Kassulke (1974) and Cilliers & Neser(1991), amongst others, have thus emphasized theneed to import different ‘strains’ of the naturalenemies already released, so as to increase thenumber of lantana varieties attacked. However,the distinction must be made between agentspreviously established and those that failed toestablish. Since little is known about the extent towhich established agents have realized their
Baars & Neser: Biological control of Lantana camara 25
Status of natural enemies released and established on Lantana camara in South Africa.
Order/ Natural enemy species Origin Main Mode of Status Damage inflictedFamily releases attack
Coleoptera
Chrysomelidae Octotoma scabripennis Mexico via 1971 Leaf miner Established in the warm, moist Extensive defoliation,Guérin-Méneville Hawaii via eastern range of lantana. Abun- but localized
Australia dant in localized inland areas
Octotoma championi Costa Rica via 1978 Leaf miner Persisted in low numbers for two UnknownBaly Australia 1995 seasons after the last release.
Establishment unconfirmed
Uroplata girardi Pic Paraguay via 1974 Leaf miner Abundant in KwaZulu-Natal Extensive defoliation inHawaii via 1983 coastal regions. Present in low coastal regionsAustralia numbers in the warm, moist
inland range of lantanaDiptera
Agromyzidae Calycomyza lantanae Trinidad via 1982 Leaf miner Widely established in low Unknown(Frick) Australia numbers. Heavily parasitized
Florida, USA 1989
Hemiptera
Tingidae Teleonemia scrupulosa Mexico via Flower and Widely established in large numbers Complete defoliation andStål Hawaii 1961 leaf sucker across the entire range of lantana; abortion of flowers in
via Australia 1971 severe damage sporadic subtropical regionsvia Mauritius 1984Florida, USA 1989
Lepidoptera
Pyralidae Salbia haemorrhoidalis Cuba via Hawaii 1962 Flower and Widely established in low numbers. UnknownGuenée fruit feeder Reared from native Lippiaspecies

potential, importations of new genetic material(e.g. T. scrupulosa) may not necessarily alter anagent’s status in South Africa. Unless the impactsof established natural enemies are well quantified,it seems inappropriate to introduce new material.Neser & Cilliers (1990) highlighted the difficultiesof monitoring new genetic material in large,well-established populations, which may result ininconclusive evidence unless suitable techniquesare used. By contrast, the re-importation of new‘strains’ of species that failed to establish, collectedfrom different species in the lantana complex andfrom climates that match target release sites, isexpected to improve the chances of establishment(Neser & Cilliers 1990; Cilliers & Neser 1991).
To increase the number of lantana varietiesattacked in South Africa, Neser & Cilliers (1990)suggested that local varieties should be exposed tothe natural enemies within the native range of L.camara. This would afford several advantages butwould depend on detailed comparative perfor-mance studies on the South African varieties. Thesewould indicate the natural enemies that can copewith the different varieties as well as the degree ofsuitability of their ‘new’ hosts.
High rates of parasitism in the field have beenobserved for C. lantanae, C. lantanella, O. lantanaeand H. laceratalis and appear to significantly re-duce the efficacy of these natural enemies in SouthAfrica. Eutreta xanthochaeta is also known to beheavily parasitized in other countries where it hasestablished (Daun & Messing 1996), suggestingthat the recruitment of native generalistparasitoids may have contributed to its failure toestablish in South Africa. However, the possibilityof heavy parasitism by native parasitoids shouldnot deter releases of new species or ‘strains’ of nat-ural enemies. Parasitized populations could stillinflict significant damage, provided that the popu-lations are able to proliferate and survive seasonaland other adversities.
NEW BIOCONTROL CANDIDATESUNDER EVALUATION
The biology, host specificity and potential ofseveral species that are being evaluated for re-lease in South Africa (Table 3) are summarizedbelow.
Aceria lantanae (Acari: Eriophyidae)The eriophyid mite, Aceria lantanae (Cook), is
known to cause two characteristic kinds ofdamage to Lantana species in South and CentralAmerica (Flechtmann & Harley 1974). In South
American countries it causes ‘crinkle’ galls on theleaf surface (Flechtmann 1973). In Central Americait distorts flower buds into inflorescence gallscomprising a mass of tiny green leaves (Keifer &Denmark 1976). These two symptoms are geo-graphically distinct and seldom coexist, withreported incidences of both kinds of symptomsfrom Florida (Cromroy 1983), Mexico (Palmer &Pullen 1995) and Venezuela. Although regardedas morphologically identical, mites inducing thetwo symptoms appear to be distinct and mayconstitute separate biotypes or species.
The lantana inflorescence mite was first importedfrom Florida, USA (Craemer 1989). Althoughattempts to rear the mite on local lantana varietiesin quarantine were largely unsuccessful, develop-ing colonies induced limited symptoms. In 1996, acolony was initiated from rooted material im-ported from Florida, but later died out as succes-sive daughter plants were severely damaged byexcessive gall formation. Mite-induced gallsappear to act as metabolic sinks, but also form inplace of flowers and thereby primarily reduce seedset. Heavy infestations of the inflorescence miteobserved in Florida, USA, after an exceptionallywet season in 1998, resulted in large stands oflantana being devoid of mature flowers. Followingintroductions from several countries (Table 3) in1997, a mixture of flower gall mite colonies devel-oped characteristic symptoms on several SouthAfrican and Australian lantana varieties in quaran-tine (Urban, unpubl.). Successful gall inductionappears to require a combination of suitablephysiological (differentiating flower buds on anactively-growing plant) and environmental condi-tions (high humidity) during infestation. Host-specificity screening is underway. Additionalmaterial was imported from Mexico and Vene-zuela in 1998 (Table 3), in an attempt to boost thegenetic base of the laboratory culture.
Falconia intermedia (Hemiptera: Miridae)Two separate laboratory cultures of Falconia
intermedia (Distant) were collected in Jamaica in1994 and in Guatemala in 1997 (Neser, unpubl.).This species is endemic to Central America whereit is associated with several species in the L. camaracomplex (Palmer & Pullen 1995, 1998).
Adults and nymphs are highly active andmobile, and feed on the undersides of leaves.Feeding damage causes chlorosis, characterizedby the stippling of the upper leaf surfaces, and cancause them to desiccate and abscise. Eggs aredeposited singly or in small groups, on the
26 African Entomology Memoir No. 1 (1999): 21–33

undersides of the leaves. Under laboratory condi-tions the eggs hatched after about 13 days and thenymphs (there are five instars) completed theirdevelopment in about 14 days; adults lived for upto two months and laid about four eggs per day.
Nymphs of F. intermedia completed their devel-opment on L. camara and some native species ofLippia (Verbenaceae). No survival and develop-ment occurred on the native Lantana rugosaThunb. or the introduced ornamental Lantanamontevidensis (Sprengler) Briquet, and perfor-mance was poor on the introduced Lantana trifoliaL. In adult starvation trials F. intermedia had signifi-
cantly higher rates of survival and egg productionon L. camara than on the various Lippia species.During multi-choice trials, L. camara was consis-tently selected as the preferred host plant for feed-ing and oviposition. Based on these results, F.intermedia was cleared for release, and released, inSouth Africa in April 1999.
Teleonemia vulgata (Hemiptera: Tingidae)A culture of the sap-sucking lace-bug, Teleonemia
vulgata Drake & Hambleton, was collected in 1996in Rio de Janeiro, Brazil, near the only locality fromwhich this species was previously recorded
Baars & Neser: Biological control of Lantana camara 27
Status of biocontrol candidates recently evaluated for release on Lantana camara in South Africa.
Order/ Natural enemy species Date Origin StatusFamily imported
Acari
Eriophyidae Aceria lantanae 1989 Florida, USA. Imported for culturing on local lantana varieties and undergoing(Cook) 1996 Florida, USA; biological studies
19971 Florida, USA. Jamaica; New material imported as separate ‘strains’ to determineMexico compatibility with local lantana varieties. Undergoing host-
19981 Mexico; Venezuela specificity tests
Hemiptera
Miridae Falconia intermedia 19951 Jamaica Released in April 1999(Distant) 19971,2 Guatemala. Imported to provide new genetic material for laboratory culture
Tingidae Teleonemia vulgata 19961 Brazil Undergoing host-specificity testsDrake & Hambleton
Membracidae Aconophora compressa 19951 Mexico via Australia Undergoing host-specificity testsWalker 1997 Guatemala Guatemalan culture (only) terminated
Coleoptera
Apionidae Coelocephalapion sp. 19971 Mexico, Cárdenas Undergoing biological studies and host-specificity screeningAppears acceptable for release in South Africa
19981,2 Mexico, Veracruz Imported to provide new genetic material for laboratory culture
Chrysomelidae Omophoita albicollis 1993 Jamaica Rejected because of wide host range under laboratory conditionsFabricius Culture terminated
Charidotis pygmaea 19941 Colombia via Australia Undergoing performance trials on local lantana varieties forBuzzi 19981 Colombia via Australia comparison with performance on Lantana montevidensis
Alagoasa prob. 19971 Mexico Undergoing biological studies and host-specificity screeningquadrilineata (Harold)
?Longitarsus sp. 1996 Trinidad; Florida Culture died in quarantine1997 Mexico Culture died in quarantine19981 Mexico, Veracruz Undergoing biological studies and developing culturing techniques
?L. columbicus Harold 19981,2 Venezuela Undergoing biological studies and developing culturing techniques
Diptera
Agromyzidae Ophiomyia camarae 1995 Florida, USA Culture died in quarantineSpencer 1996 Trinidad Culture died in quarantine
19971 Florida, USA; Mexico Undergoing host-specificity tests. Appears acceptable for releasein South Africa
19981,2 Florida, USA New material imported to boost the quarantine culture
Tephritidae Eutreta xanthochaeta 19981 Hawaii Undergoing biological studies and host-specificity testsAldrich
1: importations from which the current laboratory cultures originate.2: separate laboratory culture maintained.

(Drake & Ruhoff 1965). Damage in the fieldappears similar to that caused by T. scrupulosa.Several species of Teleonemia are associated withthe Lantana species complex in South and CentralAmerica, and some have been released as bio-control agents in various countries (Julien &Griffiths 1998). With the exception of T. validicornisStål (Harley & Kassulke 1971), all six Teleonemiaspecies that were studied proved suitable forrelease. Teleonemia vulgata has not been previouslyevaluated as a biocontrol agent against L. camara.
Adults and nymphs prefer to feed on flowers,but readily feed on leaves and vegetative buds.Eggs are inserted singly or in small clusters into theventral veins and periphery of leaves, and into theflower peduncle. Eggs take about 14 days to hatchand the nymphs pass through five instars, takingabout 15 days to complete their development.Feeding damage results in the abortion of floretsand the desiccation and abscission of leaves.
During no-choice tests, nymphs completed nor-mal development on L. camara, L. rugosa, L. trifolia,L. montevidensis, and different indigenous Lippiaspecies. Survival and development on the nativePriva cordifolia Druce (Verbenaceae) was low, indi-cating its unsuitability as a host. Preliminary adultstarvation trials on these plant species indicatedthat ovariole development was better on L. camara,resulting in a higher deposition of fat reserves,well-developed ovaries and eggs. During choicetests, gravid females preferred to feed and ovipositon L. camara, with comparatively fewer eggs de-posited on the other plant species. Additionalmulti-choice trials are to be completed before T.vulgata may be considered for release.
Aconophora compressa (Hemiptera:Membracidae)
A culture of the stem-sucking membracid,Aconophora compressa Walker, from Cuernavaca,Mexico, was obtained from the Queensland De-partment of Natural Resources, Australia, in 1995(Table 3). This species is endemic to the Neotropics(Palmer et al. 1996). Specimens collected in 1997(Neser, unpubl.) in Guatemala appeared some-what different from the Mexican material and aseparate quarantine culture was maintained, butlater destroyed when it was confirmed to be thesame species.
Adults feed on the stems of lantana and remaingregarious during the pre-oviposition period.Females then disperse to insert the eggs intoactively-growing stems, forming egg cases ofabout 70 eggs that are covered by a protective
froth. The females tend the egg cases and emerg-ing nymphs, until these complete their develop-ment (Palmer et al. 1996). Nymphs undergo fiveinstars and display a variable rate of development,ranging from 28–108 days in the greenhouse.Feeding damage causes flowers to abort and shoottips to wilt and die back, often causing long sec-tions of thick stems to die.
Host-specificity tests indicated that among thedifferent species of Lantana, A. compressa wasspecific to L. camara, confirming the results ofPalmer et al. (1996) in Mexico and Australia. Starva-tion tests showed that feeding on a number ofindigenous Lippia species allowed ovariole devel-opment, oviposition and high rates of nymphalemergence, similar to that on L. camara. However,preliminary multi-choice trials with inexperi-enced adults revealed preferences for L. camara.Species of Lippia may thus be able to serve asmarginal hosts under field conditions and furtherperformance and choice trials will be conductedbefore A. compressa is considered for release inSouth Africa.
Coelocephalapion sp. (Coleoptera:Apionidae)
The petiole-boring apionid, an undescribed spe-cies of Coelocephalapion (Kissinger, pers. comm.),was first collected in Cardenas, Tabasco Province,Mexico, during a survey in 1997, and subsequentlyat a number of sites in Veracruz, Mexico, in 1998.
Adults feed on the leaves of lantana, and eggs areinserted into the leaf petiole and flower peduncle.Larvae burrow for a short distance into the planttissue and induce a small gall. The later instars re-main relatively sedentary and feed on callus tissuewithin the gall. Larvae pass through four instarsand pupate inside the gall, and adults emergefrom a small exit hole chewed in the gall. Thelife cycle from egg to adult is approximately30–35 days in the laboratory and the adults live for4–5 months. Galls induced on the leaf petioledisrupt the supply of liquids and may cause smallleaves to desiccate and abscise. Flower gallsappear to prevent the development of seeds in thelaboratory.
Preliminary tests suggested that the weevils arehost specific and perform best on L. camara.During starvation trials, adults fed and ovipositedon the two introduced species, L. montevidensisand L. trifolia, and on five indigenous species,namely Lantana rugosa, Lippia javanica Spreng.,Lippia rehmannii Pearson, Lippia wilmsii Pearsonand Lippia scaberrima Sond. Although a number of
28 African Entomology Memoir No. 1 (1999): 21–33

eggs were deposited on these plants during thesetrials, far fewer larvae completed their develop-ment on them than on L. camara. During multi-choice tests, adults consistently preferred to feedand oviposit on lantana. The specific ovipositionbehaviour of the apionid is thought to limit thenumber of suitable hosts. The results so far indi-cate that the apionid may be acceptable for releasein South Africa.
Charidotis pygmaea (Coleoptera:Chrysomelidae)
The leaf-feeding tortoise beetle, Charidotispygmaea Buzzi, was imported into South Africafrom Brazil, via Australia, in 1994 (Table 3). Adultsand larvae feed on the leaves, causing characteris-tic longitudinal feeding scars on the upper leaf sur-faces. Eggs are deposited singly on leaves, semi-embedded into the lower epidermis and coveredby a hardened froth excretion. Protective frothextends from the posterior end of the egg andforms a raised covering over the egg, most likelyfunctioning as protection against parasitoids. Thelarvae undergo five instars, lasting about 38 daysto pupation, which occurs on the leaf surface.
Although originally collected on Lantanatiliaefolia (Swarbrick et al. 1995), one of the closelyrelated species in the L. camara complex and verysimilar to the weedy forms, C. pygmaea wasthought to be potentially specific to L. camara.However, adults performed better on the intro-duced L. montevidensis (creeping lantana) than onvarious L. camara varieties. Survival and develop-ment also occurred on the indigenous L. rugosaand various Lippia species, but performance waspoor relative to that on L. montevidensis and L.camara (Sparks, unpubl.). Various sterile forms of L.montevidensis, and hybrids, have been introducedinto South Africa via the ornamental trade andare popular garden plants. The fertile form inAustralia is regarded as invasive (Munir 1996) andwas also targeted for biological control with therelease of C. pygmaea (Julien & Griffiths 1998).Since C. pygmaea appears to be better suited to L.montevidensis than to local South African lantanavarieties, it seems unlikely that it will be furtherconsidered for release in South Africa.
Omophoita albicollis (Coleoptera:Chrysomelidae)
A culture of the leaf-feeding flea beetle, Omo-phoita albicollis Fabricius, was collected in Jamaicain 1993. Adults feed voraciously on flowers andleaves and deposit eggs under leaf litter on the soil
surface. Larvae emerge from the leaf litter andfeed on recently abscised leaves, or on leaves onbranches near the ground. Larvae pass throughthree instars, taking 21–25 days to complete theirdevelopment, and then burrow into the soil topupate within a hardened chamber. In the labora-tory adults emerge from the soil after about 10days (Sparks, unpubl.).
During starvation trials, larvae survived anddeveloped on several species of Verbenaceae,including L. trifolia, several indigenous Lippiaspecies and Phyla nodiflora Greene. Larvae alsodeveloped on several indigenous and economi-cally important species in the Lamiaceae. Duringmulti-choice trials, adults fed on several species ofVerbenaceae and Lamiaceae and caused damagesimilar to that on L. camara. The unacceptably widehost range of O. albicollis resulted in its rejection asa biocontrol candidate for release in South Africa(Sparks, unpubl.) and the laboratory culture wasterminated.
Alagoasa prob. quadrilineata (Coleoptera:Chrysomelidae)
A supply of the leaf-feeding flea beetle, Alagoasaprob. quadrilineata (Harold), was collected inCatemaco, Mexico in 1997 (Neser, unpubl.). Dur-ing a survey in Mexico in 1998 (Baars & Sparksunpubl.), similar adults were collected on mem-bers of the L. camara complex in the Yucatán,Campeche, Tabasco and Veracruz provinces.
Adults and larvae feed on the leaves, and larvaepupate in the soil. The adults have three distinctcolour and pattern forms (uniform, blotched andstriped). Preliminary host-specificity tests indi-cated that survival and development occurred onclosely related plants within the Verbenaceae(Sparks, unpubl.). Alagoasa prob. quadrilineata is atan early stage of evaluation and further studies onits biology and host-specificity will be conductedbefore it is considered for release in South Africa.
?Longitarsus sp. (Coleoptera:Chrysomelidae)
Adults of the flea beetles believed to belong tothe genus Longitarsus were collected from Trini-dad and Florida in 1996 (Table 3); they failed toestablish in the laboratory. A further culture of a?Longitarsus sp., collected in Mexico in 1997 per-sisted for several generations in the laboratory butthe adults did not reappear after the first winter.Another culture was collected from Veracruz inMexico in 1998 and still persists in the laboratory. Aculture of a flea beetle, provisionally identified as
Baars & Neser: Biological control of Lantana camara 29

Longitarsus columbicus columbicus Harold, was col-lected in Maracay, Venezuela in 1998. This specieswas previously recorded on Lantana spp. in Vene-zuela (Savini 1997). The identity and possibleconspecificity of the specimens collected in thevarious countries they were not determined andwere maintained as separate cultures.
Adults of ?Longitarsus columbicus columbicus feedon the leaves of lantana, and deposit their eggs inthe leaf litter. Emerging larvae burrow into the soiland feed externally on secondary rootlets. Larvaepupate in hardened cases near the soil surface.The immature stages require about 2 months fordevelopment in the laboratory, and the adultslived in excess of 2 months. Reliable culture tech-niques are being developed before host-specificitytests can be conducted.
Ophiomyia camarae (Diptera:Agromyzidae)
The herringbone leaf miner, Ophiomyia camaraeSpencer, originates from South and CentralAmerica. Adult flies are small and very active,usually inserting a single egg into the epidermaltissue on the abaxial aspect of leaves. The emerg-ing larva burrows into the mesophyll tissue, typi-cally towards the midrib of the leaf. The larva thenburrows laterally, in a characteristic ‘herring-bone’pattern, into the leaf lamina, and also into thepetiole. The late instar larva usually burrows to-wards the tip of the leaf, or elsewhere near theedge of the leaf, where pupation occurs. Leavesdamaged by the larvae may abscise prematurely,and larval development is completed in abscisedleaves. High populations were recorded inFlorida, USA (Stegmaier 1966), and were similarlyobserved there during 1987 and 1998.
Characteristic leaf mines have been recorded onL. tiliaefolia in Brazil (Winder & Harley 1983), on L.camara in Mexico (Palmer & Pullen 1995), Trinidadand Florida (Stegmaier 1966), on L. trifolia in Vene-zuela and on unidentified Lantana species in Ja-maica, Guatemala and Brazil. Although theidentities of the flies were not all confirmed, itappears that O. camarae is associated with variousspecies of the genus Lantana and is able to utilizethem as hosts across its native geographical range.Preliminary host-specificity trials confirmed thisassociation and the fly has performed well onseveral local lantana varieties. Although oviposi-tion and larval development occurred on someindigenous Lippia species, performance was farbetter on L. camara (Simelane, unpubl.) and the flywill be considered for release.
Eutreta xanthochaeta (Diptera:Tephritidae)
The stem-galling tephritid, Eutreta xanthochaetaAldrich, was one of the first natural enemiesreleased against L. camara in Hawaii in 1902(Perkins & Swezey 1924). The fly was first im-ported into South Africa in 1983 (Cilliers & Neser1991) (Table 3) and was considered to be safe forrelease. It was reared in quarantine for one genera-tion and released in very low numbers at onesite near Hartbeespoort (North-West Province).Although galls were initially recovered, the flyfailed to establish, probably because too few werereleased, and on lantana varieties with which theymay also have been incompatible (Cilliers & Neser1991). A new culture was imported from Hawaii in1998 and although previously released, is under-going biology studies and host-range tests in quar-antine. These tests will incorporate indigenousplant species that were not tested previously,because of the observed tendency of other naturalenemies of lantana to have expanded host ranges,and in the light of present environmental legisla-tion pertaining to biocontrol agents in SouthAfrica.
Adult females insert eggs into actively-growingshoot tips. Larvae burrow into the stem, causingthe shoot to swell and form an elongated or roundgall of up to 15 mm in diameter. Rapid growth ofthe apical stem above the gall is initially stunted,but growth may resume after the flies emerge ifthe shoot tip has not died (Perkins & Swezey 1924).Larvae feed on the proliferated gall tissue, takingup to 45 days to complete development in thegreenhouse. Pupation occurs within the gall, andadults emerge through an epidermal window pre-pared by the larva.
Preliminary quarantine tests indicate that E.xanthochaeta oviposits on several lantana varieties,all of which appear suitable for larval develop-ment. Oviposition and development also occurs onindigenous Lippia species (Klein, unpubl.) andperformance and choice trials will be conductedbefore further releases in South Africa are consid-ered.
EXPANDED LABORATORY HOSTRANGES OF NATURAL ENEMIES
Recent host-specificity tests have indicated thatmost of the natural enemies currently under eval-uation accepted related native plant species tovarying degrees under laboratory conditions.Ironically, natural enemies selected for thebiocontrol of lantana are required to cope with the
30 African Entomology Memoir No. 1 (1999): 21–33

variability of the weed, but are also required to notaccept closely related species. Two species in thegenus Lantana and four in the genus Lippia consid-ered native to South Africa (Arnold & De Wet1993), are generally accepted as hosts by the re-cently imported natural enemies of lantana underlaboratory conditions. Some insect species (e.g. A.compressa and C. pygmaea), which were acceptablefor release in Australia (Julien & Griffiths 1998),may accept and marginally affect these non-targetindigenous species in South Africa.
To reconcile physiological and true host ranges,laboratory and field studies were conducted onspecies that have been established in South Africafor decades. The tingid, T. scrupulosa, displayednon-host specific behaviour in the laboratory,feeding and developing on a wide range of specieswithin the Verbenaceae. However, preliminarystudies provided no evidence of an extended hostrange in the field where T. scrupulosa was observedto display only limited feeding on some nativeLippia species. These studies provided further evi-dence of the conservative nature of laboratorytests and suggest that extended laboratory hostranges are often, as in many biocontrolprogrammes worldwide, not realized under fieldconditions.
Although some of the natural enemy speciespresently under evaluation (e.g. Coelocephalapionsp.) do not accept certain closely related nativespecies, most others appear to perform well onseveral closely related species. Aconophora compressa(Palmer et al. 1996) and T. scrupulosa (Harley &Kassulke 1971) have been recorded on Lippiaspecies in their native range, but detailed perfor-mance studies on other promising species (e.g. F.intermedia) in the laboratory have indicated thatalthough various Lippia species are acceptable,performance was poor relative to that on L. camara.One solution to determining the acceptability ofexpanded laboratory host ranges would be toaccept possible limited feeding on such non-targetspecies in the field, as an ecologically justifiable‘trade-off ’ against the benefits of releasing agentsthat have the potential to suppress such an envi-ronmentally damaging target weed. Alternatively,the number of new natural enemies that will ulti-mately be considered acceptable for release inSouth Africa will be limited, with obvious con-straints on the biocontrol programme.
CONCLUSIONSThe success of the biocontrol programme againstlantana will depend on a suite of natural enemies
that can cope with the extreme variability andwide distribution of the weed in South Africa,without compromising native Verbenaceae. It isapparent that the insect agents currently estab-lished on lantana in South Africa do not exertsufficient control and that additional naturalenemies are required. In this respect, Cilliers &Neser (1991) suggested a possible shift in the selec-tion of new natural enemies, in which stem borers,root feeders and flower feeders should take prece-dence over leaf feeders.
Natural enemies established in South Africa,notably T. scrupulosa, O. scabripennis and U. girardi,periodically defoliate lantana stands (Cilliers1987b), but fail to sustain these levels of damage tothe photosynthetic niche, and thus the resourceproduction by the plant. This niche is therefore inneed of additional herbivore pressure and newleaf feeders should also be considered. However,several niches are not affected by natural enemyattack and the suite of candidate biocontrol agentsshould comprise species that attack as many partsof the plant as possible. In pursuit of this objective,natural enemies under consideration includefeeders on leaves, stems, petioles and shoots,flowerbuds and roots. The abundance of potentialagents suggests that, despite the problems associ-ated with the programme, L. camara remains a can-didate for biological control in South Africa.
ACKNOWLEDGEMENTSWe thank A.J. Urban, H. Klein, H.E. Sparks, F.Heystek and D.O. Simelane of the Plant ProtectionResearch Institute for technical and research assis-tance and for the use of unpublished data. We arealso grateful to M.P. Hill, T. Olckers and A.J. Urban(all of PPRI) for comments on the manuscript. Weare grateful to various organizations for providingfinancial support for the research programmeagainst lantana, including the National Depart-ment of Agriculture, Department of Water Affairsand Forestry, H.L. Hall & Sons, Hans MerenskyFoundation and the Agricultural Research Coun-cil of South Africa.
REFERENCESARNOLD, T.H. & DE WET, B.C. 1993. Plants of southern
Africa: names and distribution. Memoirs of the Botani-cal Survey of South Africa 62: 596–637.
CILLIERS, C.J. 1983. The weed, Lantana camara L., andthe insect natural enemies imported for its biologicalcontrol into South Africa. Journal of the EntomologicalSociety of Southern Africa 46: 131–138.
CILLIERS, C.J. 1987a. Notes on the biology of the estab-lished insect natural enemies of Lantana camara L.(Verbenaceae) and their seasonal history in South
Baars & Neser: Biological control of Lantana camara 31

Africa. Journal of the Entomological Society of SouthernAfrica 50: 1–13.
CILLIERS, C.J. 1987b. The evaluation of three insectnatural enemies for the biological control of the weedLantana camara L. Journal of the Entomological Society ofSouthern Africa 50: 15–34.
CILLIERS, C.J. & NESER, S. 1991. Biological control ofLantana camara (Verbenaceae) in South Africa. Agri-culture, Ecosystems and Environment 37: 57–75.
CRAEMER, C. 1989. Eriophyidae (Acari) as potentialcontrol agents of South African weeds, with descrip-tions of a new species of Tegonotus Nalepa and ofParaphytoptus Nalepa. M.Sc. thesis, Rand AfrikaansUniversity, Johannesburg.
CROMROY, H.L. 1983. Potential use of mites in biologicalcontrol of terrestrial and aquatic weeds. In: Hoy,M.A., Cunningham, G.L. & Knutson, L. (Eds) Biologi-cal Control of Pests by Mites. 61–66. University ofCalifornia, Berkeley.
DAUN, J.J. & MESSING, R.H. 1996. Response of twoopiine fruit f ly parasitoids (Hymenoptera:Braconidae) to the lantana gall fly (Diptera:Tephritidae). Biological Control 25: 1428–1437.
DRAKE, C.J. & RUHOFF, F.A. 1965. Lacebugs of theworld: a catalog (Hemiptera: Tingidae). Bulletin of theUnited States National Museum 243: 1–634.
FLECHTMANN, C.H.W. 1973. On an eriophyid mite(Acari) from lantana from Brazil. Anais da SociedadeEntomológica do Brasil 2: 109–110.
FLECHTMANN, C.H.W. & HARLEY, K.L.S. 1974. Prelim-inary report on mites (Acari) associated with Lantanacamara L. in the Neotropical region. Anais de SociedadeEntomológica do Brasil 3: 69–71.
GENTLE, C.B. & DUGGIN, J.A. 1997a. Allelopathy as acompetitive strategy in persistent thickets of Lantanacamara L. in three Australian forest communities.Plant Ecology 132: 85–95.
GENTLE, C.B. & DUGGIN, J.A. 1997b. Lantana camara L.invasions in dry rainforest-open ecotones: the role ofdisturbances associated with fire and cattle grazing.Australian Journal of Ecology 22: 298–306.
HARLEY, K.L.S. & KASSULKE, R.C. 1971. Tingidae for bi-ological control of Lantana camara [Verbenaceae].Entomophaga 16: 389–410.
HARLEY, K.L.S. & KASSULKE, R.C. 1974. The suitabilityof Phytobia lantanae Frick for biological control ofLantana camara in Australia. Journal of the AustralianEntomological Society 13: 229–233.
HARLEY, K.L.S., KERR, J.D. & KASSULKE, R.C. 1979.Effects in S.E. Queensland during 1967–72 of insectsintroduced to control Lantana camara. Entomophaga24: 65–72.
HOLM, L.G., PLUCKNETT, D.L., PANCHO, J.V. &HERBERGER, J.P. 1977. The World’s Worst Weeds. TheUniversity Press of Hawaii, Honolulu.
HOWARD, R.A. 1970. Lantana camara – a prize and a peril.American Horticultural Society Winter: 31–36.
JULIEN, M.H. & GRIFFITHS, M.W. 1998. Biological Con-trol of Weeds: A World Catalogue of Agents and theirTarget Weeds. 4th Edition. CABI Publishing, UK.
KEIFER, H.H. & DENMARK, H.A. 1976. Eriophyeslantanae Cook (Acarina: Eriophyidae) in Florida.Florida Department of Agricultural Services, Divisionof Plant Industry, Entomology Circular No. 166, 2 pp.
KELLERMAN, T.S., NAUDE, T.W. & FOURIE, N. 1996.
The distribution, diagnoses and estimated economicimpact of plant poisonings and mycotoxicoses inSouth Africa. Onderstepoort Journal of Veterinary Re-search 63: 65–90.
MORTON, J.F. 1994. Lantana, or red sage (Lantana camaraL., [Verbenaceae]), notorious weed and populargarden flower; some cases of poisoning in Florida.Economic Botany 48: 259–270.
MUNIR, A.A. 1996. A taxonomic review of Lantana camaraL. and L. montevidensis (Spreng.) Briq. (Verbenaceae)in Australia. Journal of the Adelaide Botanical Gardens 17:1–27.
NESER, S. & CILLIERS, C.J. 1990. Work towards biologi-cal control of Lantana camara: perspectives. In:Delfosse, E.S. (Ed.) Proceedings of the Seventh Interna-tional Symposium on Biological Control of Weeds.363–369. Istituto Sperimentale per la PatologiaVegetale Ministero dell’Agricoltura e delle Foreste,Rome.
OOSTHUIZEN, M.J. 1964. The biological control ofLantana camara L. in Natal. Journal of the EntomologicalSociety of Southern Africa 27: 3–16.
PALMER, W.A. & PULLEN, K.R . 1995. Thephytophagous arthropods associated with Lantanacamara, L. hirsuta, L. urticifolia and L. urticoides(Verbenaceae) in North America. Biological Control 5:54–72.
PALMER, W.A., WILLSON, B.W. & PULLEN, K.R. 1996.The host range of Aconophora compressa Walker(Homoptera: Membracidae): a potential biologicalcontrol agent for Lantana camara L. (Verbenaceae).Proceedings of the Entomological Society of Washington98: 617–624.
PALMER, W.A. & PULLEN, K.R. 1998. The host range ofFalconia intermedia (Distant) (Hemiptera: Miridae): apotential biological control agent for Lantana camaraL. (Verbenaceae). Proceedings of the EntomologicalSociety of Washington 100: 633–635.
PERKINS, R.C.L. & SWEZEY, O.H. 1924. The introduc-tion into Hawaii of insects that attack lantana. HawaiiSugar Planters Association Experimental Station Bulletin(Entomological Series) 16: 1–83.
RADUNZ, L.A.J. 1971. Some aspects of the preferentialbehaviour of Teleonemia scrupulosa Stål, towards itshost plant Lantana camara. Hons. thesis, University ofQueensland.
SAVINI, V. 1997. Evaluación de los datos sobre losphytophaga dañinos en Venezuela (Coleoptera).Boletin de Entomologia Venezolana (Serie Monografias)1: 95–96.
SMITH, L.S. & SMITH, D.A. 1982. The naturalisedLantana camara complex in eastern Australia.Queensland Botanical Bulletin No. 1. QueenslandDepartment of Primary Industry, Brisbane.
SPIES, J.J. 1984. A cytotaxonomic study of Lantana camara(Verbenaceae) from South Africa. South African Jour-nal of Botany 3: 231–250.
SPIES, J.J. & DU PLESSIS, H. 1987. Sterile Lantana camara:fact or theory. South African Journal of Plant and Soil 4:171–174.
SPIES, J.J. & STIRTON, C.H. 1982a. Chromosome num-bers of South African plants. South African Journal ofBotany 48: 21–22.
SPIES, J.J. & STIRTON, C.H. 1982b. Meiotic studies ofsome South African cultivars of Lantana camara
32 African Entomology Memoir No. 1 (1999): 21–33

(Verbenaceae). Bothalia 14: 101–111.SPIES, J.J. & STIRTON, C.H. 1982c. Embryo sac develop-
ment in some South African cultivars of Lantanacamara. Bothalia 14: 113–117.
STEGMAIER, C.E. 1966. A leaf miner on Lantana inFlorida, Ophiomyia camarae (Diptera, Agromyzidae).The Florida Entomologist 49: 151–152.
STIRTON, C.H. 1977. Some thoughts on the polyploidcomplex Lantana camara L. (Verbenaceae). Proceedings
of the Second National Weeds Conference of South Africa.321–344. A.A. Balkema, Cape Town.
SWARBRICK, J.T., WILLSON, B.W. & HANNAN-JONES,M.A. 1995. The biology of Australian weeds: Lantanacamara L. Plant Protection Quarterly 10: 82–95.
WINDER, J.A. & HARLEY, K.L.S. 1983. Phytophagousinsects on lantana in Brazil and their potential for bio-logical control in Australia. Tropical Pest Management29: 346–362.
Baars & Neser: Biological control of Lantana camara 33