paper no. 11426 - ohio universitycheserver.ent.ohiou.edu/paper-gu/nace/11426 - bioenergetics... ·...
TRANSCRIPT

©2011 by NACE International. Requests for permission to publish this manuscript in any form, in part or
in whole, must be in writing to NACE International, Publications Division, 1440 South Creek Drive,
Houston, Texas 77084. The material presented and the views expressed in this paper are solely those of the
author(s) and are not necessarily endorsed by the Association.
Bioenergetics Explains When and Why More Severe MIC Pitting by SRB Can Occur
Dake Xu and Tingyue Gu (Speaker) Institute for Corrosion and Multiphase Technology
Department of Chemical and Biomolecular Engineering Ohio University
Athens, Ohio 45701 Phone: 740-593-1499; Fax: 740-593-0873; [email protected]
ABSTRACT The detection and prediction of microbiologically influence corrosion (MIC) have long been plagued by a lack of basic understanding of MIC mechanisms. In this work, bioenergetics was used to explain why and when sulfate reducing bacteria (SRB) become aggressive toward iron. The electrochemical standard reduction potentials of Fe2+/Fe and acetate + CO2/lactate are very close. This means that bioenergetically Fe oxidation is as favorable as lactate oxidation, except that the former does not provide organic carbons for cell growth. In the stationary cell growth phase, Fe oxidation when coupled with dissimilatory sulfate reduction generates energy in the form of ATPs that are used by SRB as maintenance energy. In this work experiments were carried out by subjecting established Desulfovibrio vulgaris (ATCC 7757) biofilms on carbon steel coupons to various degrees of carbon source starvation. It was found that when sessile SRB cells were starved of organic carbons, they turned to Fe oxidation and thus increased pitting weight loss. Moderately reduced carbon source caused deeper pits and a larger weight loss compared with full strength medium. Severely reduced carbon source caused wider but shallower pits and a larger weigh loss compared with moderately reduced carbon source. When the carbon source was dropped from the medium, weight loss decreased compared to that in full strength medium, probably due to a lack of a minimum amount of carbon needed for sufficient enzyme activities that are required for MIC. Biofilm morphology plays a critical
Paper No. 11426

©2011 by NACE International. Requests for permission to publish this manuscript in any form, in part or
in whole, must be in writing to NACE International, Publications Division, 1440 South Creek Drive,
Houston, Texas 77084. The material presented and the views expressed in this paper are solely those of the
author(s) and are not necessarily endorsed by the Association.
role in MIC pitting. Loose biofilms allow better mass transfer of organic carbon to the iron surface, while dense biofilms restrict the mass transfer and thus can result in organic carbon starvation in the sessile cells that are close to the iron surface. Many NRB form very loose biofilms while SRB biofilms typically have a very dense morphology. This is probably why SRB tend to be aggressive while NRB usually are not. Our bioenergetics theory for MIC also explains in some cases why the same biofilm can be “docile” and all of sudden can become aggressive.
Keywords: MIC, Mechanism, SRB, NRB, Sulfate reduction, Bioenergetics
INTRODUCTION Microbiologically Influenced Corrosion (MIC) was first recognized by Garrett about a century ago1. In the past decade, MIC has become a major problem to the oil and gas industry as well as water utility and other industries. MIC turned out to be a major factor for several major pipeline failures. They include the gas pipeline rupture and fire near Carlsbad, New Mexico on August 19, 20002 and in San Bruno, CA on September 9, 20103, both resulting in significant human casualties. MIC was also a prime suspect together with underdeposit attack in the Alaskan pipeline leak on March 2, 20064 that caused a spike in world oil prices. The awareness of MIC is growing rapidly in recent years. However, the detection and treatment of MIC are hampered by a considerable lack of basic understanding of MIC mechanisms. Sulfate Reducing Bacteria (SRB) are the most often mentioned microbes in MIC. They often prefer volatile fatty acids as the organic carbon source. Some SRB can even grow autotrophically on CO2 and H2
5, much like methanogens, and some SRB readily utilize saturated and unsaturated hydrocarbons that are readily available in oil pipelines6. It is well known that redox reactions provide energy in biological systems. MIC pitting of iron involves iron oxidation, an electrochemical reaction. Iron can be used as an electron donor by microbes in their metabolism when other donors are not readily available. Many anaerobes that respire organic carbons can potentially be aggressive if they switch from organic carbon oxidation to iron oxidation provided that such a switch is beneficial to their survival and they are able to use iron as an electron donor. Apart from SRB, microbes such as acid producing bacteria and even some nitrate reducing bacteria7, 8 are also fount to be aggressive. Bioenergetically, even methanogens may become aggressive because hydrogen oxidation may be replaced by iron oxidation with the latter be energetically more favorable due to its more negative reduction potential under biological conditions. Thus, it is reasonable to propose a more general term of XRB to replace SRB, where X represents non-oxygen oxidants such as sulfate, nitrate, nitrite, CO2, etc.

©2011 by NACE International. Requests for permission to publish this manuscript in any form, in part or
in whole, must be in writing to NACE International, Publications Division, 1440 South Creek Drive,
Houston, Texas 77084. The material presented and the views expressed in this paper are solely those of the
author(s) and are not necessarily endorsed by the Association.
because microbes reducing any of these oxidants can potentially couple it with iron oxidation that has a very similar reduction potential (Eo´= -447 mV) as an often favored organic carbon lactate (Eo´= -430 mV) under biological conditions5. However, not all such microbes are found to be aggressive at all times. Certain conditions play critical roles, among which biofilm morphology and local nutritional and electrochemical environment. Investigations into these factors will prove critical to the understanding of MIC mechanisms.
BIOENERGETICS THEORY FOR MIC There has been no clear answer to why and when SRB attacks iron. It is well known that SRB cells do not “eat” iron1 because the tiny amount Fe ions needed as enzyme co-factors for cellular metabolism are readily available in an aqueous environment. The classical theory form MIC by SRB is the so-called Cathodic Depolarization Theory (CDT). It was initiated by von Wolzogen Kuhr and Vlugt vander in 19349 and was widely adopted by numerous publications on MIC10-12. A new MIC theory called biocatalytic cathodic sulfate reduction (BCSR) theory was introduced by Gu et al.13. The BCSR theory uses the following reactions to simplify SRB corrosion. Anodic: 4Fe 4Fe2+ + 8e- (Iron dissolution) Eo´= -447 mV (1) Cathodic: SO4
2- + 9H+ + 8e- HS- + 4H2O (BCSR) Eo´= -200 mV (2) Sulfate reduction goes forward due to biocatalysis by enzymes produced by SRB. Only the sessile SRB on the iron surface (in some cases a monolayer of sessile cells) are directly responsible for the iron dissolution if other factors such as galvanic and acid attacks are not considered. Reaction (2) actually summarizes the net result of several reactions in the enzyme catalyzed sulfate reduction steps such as those described by the CDT involving hydrogenase14. A mechanistic model-based MIC pitting prediction software program was created using the BCSR theory. It assumes that an aggressive SRB biofilm is already present on iron surface13. SRB are anaerobic prokaryotes that can use sulphate as the terminal electron acceptor in their respiration. When lactate is used as the organic carbon source, the following oxidation reaction occurs,5 CH3CHOHCOO- +H2O CH3COO- + CO2 + 4H+ + 4e- Eo´= -430 mV (3) When Reactions (2) and (3) are coupled together with biocatalysis by SRB, energy is released by the redox reaction because the Gibb‟s free energy is negative,5

©2011 by NACE International. Requests for permission to publish this manuscript in any form, in part or
in whole, must be in writing to NACE International, Publications Division, 1440 South Creek Drive,
Houston, Texas 77084. The material presented and the views expressed in this paper are solely those of the
author(s) and are not necessarily endorsed by the Association.
SO4
2- + 2CH3CHOHCOO- 2CH3COO- + 2CO2 + HS- + OH- +H2O ΔGo´= -196.4kJ/mol (4) where
ΔGo´= -nFΔEo´ <0 (5) because when concentrations are considered in the Nernst equation, ΔEo´=254 mV > 0. In Equation (5), ΔGo´ is the free energy change under standard conditions for pH=7 biological systems15, n the number of electrons, F the Faraday constant, and ΔEo´ the difference in reduction potentials between the oxidant and the reductant. A positive ΔEo´ value will yield a negative ΔGo´ indicating an exergonic reaction that means it is thermodynamically favorable and can occur spontaneously. However, favorable thermodynamics does not mean the reaction will actually proceed at a significant rate. For example, no noticeable reaction will happen when lactate is mixed with sulfate without catalysis. The kinetically retarded redox reaction is catalyzed by the enzymes in SRB. The hydrogenase enzyme in hydrogenase-positive SRB is capable of helping push the reactions forward. The redox reaction in (4) releases 196.4 kJ of energy for each mole of sulfate reduced. This is very high. It is reported that a ΔEo´ of +25mV, equivalent to a free energy change ΔGo´ of -20kJ/mol (n=8) is sufficient to support growth of SRB5. Such energy is needed by SRB metabolic activities such as active transport, signal amplification and organic synthesis. Even when cells are not growing in the stationary phase, maintenance energy is still needed12, 16. Bioenergetics plays a critical role in the understanding of MIC mechanisms. MIC pitting occurs under an SRB biofilm. When planktonic SRB cells metabolize lactate and sulfate, Reactions (2) and (3) happen in the cytoplasm17 and the electrons from iron dissolution are not needed. In fact these electrons are not available any way to the planktonic SRB cells because electrons cannot “swim” in an aqueous solution. This is why planktonic SRB do not directly cause iron dissolution through its metabolism. Because planktonic cell counts have no direct impact on the sessile SRB in a biofilm that are very close to an iron surface, they are known to have no direct correlation with MIC pitting severity. An SRB biofilm on an iron surface is typically very dense and this causes a significant mass transfer barrier to the diffusion of organic carbon. This means when the bulk organic carbon concentration is low, the small mass transfer driving force (i.e., concentration difference) may not be sufficient to supply the organic carbon to the sessile SRB cells deep in the biofilm, especially the monolayer of SRB cells right on the iron surface. In order to extract maintenance energy for their survival, the SRB cells may switch to iron oxidation to replace organic carbon oxidation to provide electrons for sulfate reduction. It turns out that the reduction potential for

©2011 by NACE International. Requests for permission to publish this manuscript in any form, in part or
in whole, must be in writing to NACE International, Publications Division, 1440 South Creek Drive,
Houston, Texas 77084. The material presented and the views expressed in this paper are solely those of the
author(s) and are not necessarily endorsed by the Association.
Fe2+/Fe (-447mV) is slightly more favorable than CO2+acetate/lactate (-430mV), which means iron oxidation actually provides slightly more energy than lactate oxidation when coupled with sulfate reduction. The ATP molecules generated from the redox reaction also benefits other cells nearby because they can be passed around. Lactate is a common organic carbon for SRB growth in laboratory investigations. It is more energetic than other organic carbons such as formate and acetate because it has a more negative standard reduction potential as shown in Table 1. Our new bioenergetics theory for MIC suggests that an established SRB biofilm may be more aggressive under the carbon starvation condition. Experimental verification is desired.
EXPERIMENTAL CONDITIONS
Table 2 shows test conditions used in this work. C1018 carbon steel disk coupons (1.2cm in diameter with top surface uncoated) were put into 120ml anaerobic vials containing SRB (Desulfovibrio vulgaris, ATCC 7757) and 100 ml ATCC1249 medium (Table 3). Each vial contained 2 coupons. Each test condition was duplicated using 3 vials. After 7 days the coupons coated with established SRB biofilms were transferred to new 120 ml vials containing fresh ATCC 1249 medium or ATCC 1249 medium with diluted (sodium citrate and sodium lactate) carbon source concentrations to test carbon source starvation effects. After 7 days the coupons were placed in the fresh media with various carbon source amounts, coupons were examined using SEM and sessile cells were counted. To count the sessile cells, coupons were first rinsed with distilled water and sonicated for 30 seconds and vortexed. Residual cells were removed from the coupon surface using a scraper. SRB Test Kit (Sani-Check® Product #100) was then used to test the cells in the cell suspension obtained from removed sessile cells. The MPN (Most Probable Number) method was only occasionally used to double-check the test kit results because it was labor-intensive. The culture medium was deoxygenated using nitrogen sparging before inoculation. To observe the biofilm on the coupon surface, the sessile cells were first fixed in 4% (wt) glutaraldehyde for 4 hours and then rinsed with a graded series (25%, 50%, 75%, 100% v/v) of ethanol to dehydrate. It was subsequently critical point dried and coated with Au. To view the pits on a carbon steel coupon, a duplicate coupon in the same vial had to be used. The coupon surface was cleaned with Clark‟s solution before weight loss analysis and SEM observation. The same coupon was again used for pit depth profile analysis under an Infinite Focus Microscope (IFM). The entire coupon surface was first scanned under IFM at a

©2011 by NACE International. Requests for permission to publish this manuscript in any form, in part or
in whole, must be in writing to NACE International, Publications Division, 1440 South Creek Drive,
Houston, Texas 77084. The material presented and the views expressed in this paper are solely those of the
author(s) and are not necessarily endorsed by the Association.
low resolution to find a region with deepest pits. This region was then zoomed in at a high resolution to produce a detailed pit depth profile for reporting.
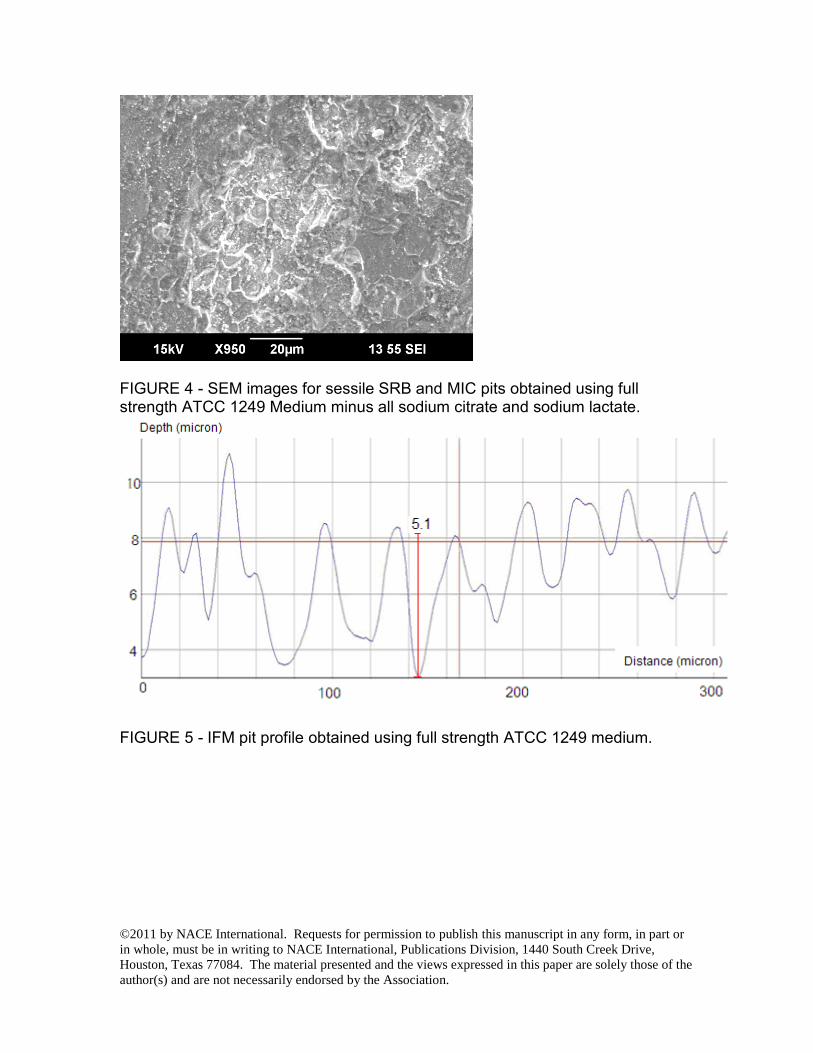
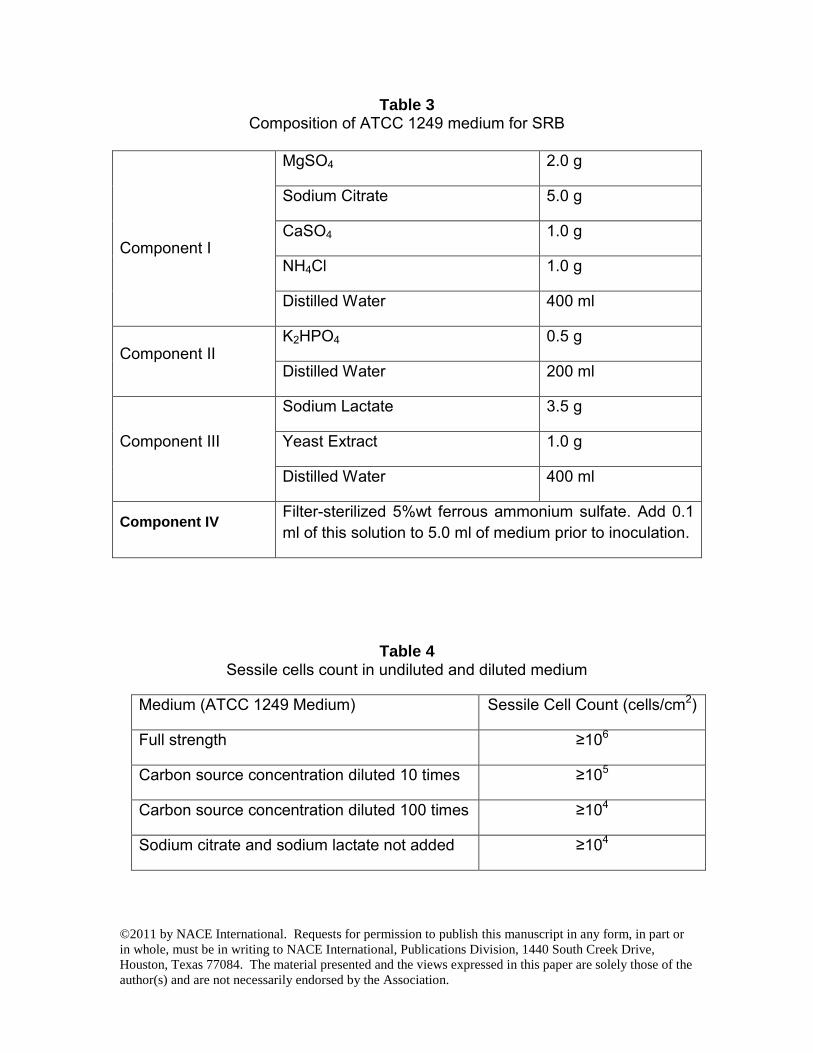
RESULTS AND DISCUSSION In this work, the biofilms were pre-grown on disk coupons in a full strength culture medium before the medium was changed to start the carbon starvation tests. The initial sodium citrate and sodium lactate concentrations were reduced by 10X, 100X and also completely eliminated in different vials to simulate various degree of carbon source starvation. The sessile cell counts in the biofilms on the coupon surfaces for 10X medium dilution and 100X dilution and no carbon source were 101, 102 and 102 times lower than that for the full strength medium as shown in Table 4. Table 4 indicates that the biofilms with 100X less carbon source and no carbon source lost a large amount of sessile cells. The sessile cell counts in Table 4 are consistent with biofilm images shown in Figs. 1 to 4. The sessile SRB images in the figures indicate the general health of the biofilms. They do not represent the hidden SRB cells underneath near the iron surface that were actually responsible for BCSR. It should be pointed out that yeast extract in the ATCC1249 medium also contained a small amount of carbon. It was not diluted in the carbon starvation tests in this work. Microbial cells also have the ability to scavenge carbon sources from dead cells. Fig. 1 shows the SEM images of pits for the regular strength medium. When the carbon source was diluted by 10X (Fig. 2), and 100X (Fig. 3), pits appeared to be larger in width compared to the pits for full strength medium. The largest pit in Fig. 2 (moderate carbon starvation) is considerably larger than that in Fig. 1 (full medium). The pits in Fig. 3 (severe carbon starvation) appear to be relatively shallow compared with Fig. 2. This was probably because the biofilm could not grow into the newly created pit bottom under severe carbon starvation. When sodium citrate and sodium lactate were completely removed from the medium, the coupon surface was rough under magnification but it did not show any well-defined MIC pits (Fig. 4). The pit depth profiles from IFM in Figs. 5 to 7 are consistent with observations from Figs. 1 to 3. No pit depth profile was presented for Fig. 4 conditions because the pits were very shallow and not well-defined. The SEM images (Figs. 1 to 4) and Table 4 data came from one experiment (Test #1). This carbon source starvation experiment was repeated twice (Test #2 and Test #3). Among the three separate tests, pit image analysis was done for Test #1 and Test #3. Both of them showed the same qualitative trend as presented in Figs. 1 to 4. Fig. 8 shows the weight loss data for the three tests. They are consistent with the pit images shown in Figs. 1 to 4. Fig. 8 shows that

©2011 by NACE International. Requests for permission to publish this manuscript in any form, in part or
in whole, must be in writing to NACE International, Publications Division, 1440 South Creek Drive,
Houston, Texas 77084. The material presented and the views expressed in this paper are solely those of the
author(s) and are not necessarily endorsed by the Association.
under carbon starvation, weight loss increased. However, this trend stopped when sodium citrate and sodium lactate were completely removed. It is reasonable to argue that reason probably was that the sessile SRB cells lacked a minimum amount of carbon needed to maintain sufficient enzyme activities for MIC pitting. In the carbon starvation tests, sulfate concentration was stoichiometrically much larger than the organic carbon concentration when a 10x or higher reduction in organic carbon was used. This means sulfate deficiency should not be a problem in the SRB biofilm. This observation may help explain why some NRB are found to be aggressive7,8, while many are considered docile. The latter probably have loose biofilms as described by Larsen et al. compared to very dense SRB biofilms.17 With a fluffy biofilm, mass transfer may not be a limitation for carbon source diffusion, which means sufficient organic carbon can penetrate the biofilm. It was reported18 in one study that when the bulk nitrate concentration was 0.96 mM, the nitrate concentration very close to the steel surface was 0.86 mM. The carbon source distribution (not reported) should have a similar picture in that study when there was sufficient carbon source in the bulk fluid because typical organic carbon molecules such as lactate (a small molecule) have similar diffusivities as nitrate and sulfate. During the revision of this work, a newly accepted paper by Sherar et al.20 came to our attention. It provided experimental evidence that supported our bioenergetics theory for MIC. It reported that in the absence of organic carbon, SRB cells in their microbial consortium (dominated by SRB) isolated from an oil well formed nanowires that extended to the iron surface. Nanowires (pili) are known to channel electrons for reduction reactions in microbial metabolism. In this reported case, microbes due to the lack of organic carbon needed to harvest the maintenance energy from extracellular iron oxidation coupled with intracellular sulfate reduction. They facilitated this by growing nanowires to transport electrons donated from iron oxidation to the cytoplasm inside SRB cells where sulfate reduction takes place. The role of iron sulfate film on the iron surface form during iron oxidation and sulfate reduction has been widely reported. Both aggressive and passivating effects have been reported21, 22. This factor deserves further investigation because the film can interfere with interpretations of experimental results.
CONCLUSIONS Our new bioenergetics theory for MIC suggests that established SRB biofilms under starvation may be forced to switch from organic carbon oxidation to iron oxidation for maintenance energy. Iron oxidation is bioenergetically as favorable

©2011 by NACE International. Requests for permission to publish this manuscript in any form, in part or
in whole, must be in writing to NACE International, Publications Division, 1440 South Creek Drive,
Houston, Texas 77084. The material presented and the views expressed in this paper are solely those of the
author(s) and are not necessarily endorsed by the Association.
as lactate oxidation based on their standard reduction potentials. Our experimental data using D. vulgaris were consistent with this theory. When the amount of carbon source in the medium was reduced by 10X and 100X, coupon weight loss increased as expected and pits appeared to be larger in width. However, shallower pits were observed for 100X reduction in carbon source. This was probably because severely starved SRB cells could not readily expand into the newly created pits causing deeper pits. It will be interesting to see the effect of a cyclic carbon source variation on MIC pitting, because it provides both the starvation phase and the biofilm growth phase. It should be noted that when the medium did not contain sodium citrate and sodium lactate, no well-defined MIC pits were found and weight loss was lower than that for full strength medium indicating lower ability for the SRB cells to attack iron likely due to weakened enzyme activities. More in-depth investigations including using different microbes (SRB and non-SRB anaerobes) and sulfate-free media (to eliminate the effects of iron sulfate film) are needed to further explore the validity of the bioenergetics theory presented in this work.

©2011 by NACE International. Requests for permission to publish this manuscript in any form, in part or
in whole, must be in writing to NACE International, Publications Division, 1440 South Creek Drive,
Houston, Texas 77084. The material presented and the views expressed in this paper are solely those of the
author(s) and are not necessarily endorsed by the Association.
FIGURE 1 - SEM images for sessile SRB and MIC pits obtained using full strength ATCC 1249 Medium.

©2011 by NACE International. Requests for permission to publish this manuscript in any form, in part or
in whole, must be in writing to NACE International, Publications Division, 1440 South Creek Drive,
Houston, Texas 77084. The material presented and the views expressed in this paper are solely those of the
author(s) and are not necessarily endorsed by the Association.

©2011 by NACE International. Requests for permission to publish this manuscript in any form, in part or
in whole, must be in writing to NACE International, Publications Division, 1440 South Creek Drive,
Houston, Texas 77084. The material presented and the views expressed in this paper are solely those of the
author(s) and are not necessarily endorsed by the Association.
FIGURE 2 - SEM images for sessile SRB and MIC pits obtained using full strength ATCC 1249 Medium minus 90% carbon source.

©2011 by NACE International. Requests for permission to publish this manuscript in any form, in part or
in whole, must be in writing to NACE International, Publications Division, 1440 South Creek Drive,
Houston, Texas 77084. The material presented and the views expressed in this paper are solely those of the
author(s) and are not necessarily endorsed by the Association.

©2011 by NACE International. Requests for permission to publish this manuscript in any form, in part or
in whole, must be in writing to NACE International, Publications Division, 1440 South Creek Drive,
Houston, Texas 77084. The material presented and the views expressed in this paper are solely those of the
author(s) and are not necessarily endorsed by the Association.
FIGURE 3 - SEM images for sessile SRB and MIC pits obtained using full strength ATCC 1249 Medium minus 99% carbon source.
©2011 by NACE International. Requests for permission to publish this manuscript in any form, in part or
in whole, must be in writing to NACE International, Publications Division, 1440 South Creek Drive,
Houston, Texas 77084. The material presented and the views expressed in this paper are solely those of the
author(s) and are not necessarily endorsed by the Association.
FIGURE 4 - SEM images for sessile SRB and MIC pits obtained using full strength ATCC 1249 Medium minus all sodium citrate and sodium lactate.
FIGURE 5 - IFM pit profile obtained using full strength ATCC 1249 medium.

©2011 by NACE International. Requests for permission to publish this manuscript in any form, in part or
in whole, must be in writing to NACE International, Publications Division, 1440 South Creek Drive,
Houston, Texas 77084. The material presented and the views expressed in this paper are solely those of the
author(s) and are not necessarily endorsed by the Association.
FIGURE 6 - IFM pit profile obtained using full strength ATCC 1249 medium minus 90% carbon source.
FIGURE 7 - IFM pit profile obtained using full strength ATCC 1249 medium minus 99% carbon source.

©2011 by NACE International. Requests for permission to publish this manuscript in any form, in part or
in whole, must be in writing to NACE International, Publications Division, 1440 South Creek Drive,
Houston, Texas 77084. The material presented and the views expressed in this paper are solely those of the
author(s) and are not necessarily endorsed by the Association.
FIGURE 8 - Weight loss 7 days after starvation test started.
Table 1 Standard potentials of redox couples in biological systems at pH 75
Redox couple n Eo´(mV)
Fe2+/Fe 2 -447
CO2 + acetate/lactate 4 -430
CO2/formate 2 -432
2H+/H2 2 -414
2CO2/acetate 8 -290
4CO2/butyrate 20 -280
3CO2/propionate 7 -280

©2011 by NACE International. Requests for permission to publish this manuscript in any form, in part or
in whole, must be in writing to NACE International, Publications Division, 1440 South Creek Drive,
Houston, Texas 77084. The material presented and the views expressed in this paper are solely those of the
author(s) and are not necessarily endorsed by the Association.
SO42-/HS- 8 -200
NO3-/NH3 8 +360
Table 2 Experimental conditions
Parameters Conditions
SRB strain Desulfovibrio desulfuricans (ATCC 7757)
Temperature 37ºC
Culture medium ATCC 1249 medium
Carbon source reduction 0%, 10%, 99%, 100%
Initial pH 7.0±0.1
Test duration 7 days
Material C1018 steel
©2011 by NACE International. Requests for permission to publish this manuscript in any form, in part or
in whole, must be in writing to NACE International, Publications Division, 1440 South Creek Drive,
Houston, Texas 77084. The material presented and the views expressed in this paper are solely those of the
author(s) and are not necessarily endorsed by the Association.
Table 3 Composition of ATCC 1249 medium for SRB
Component I
MgSO4 2.0 g
Sodium Citrate 5.0 g
CaSO4 1.0 g
NH4Cl 1.0 g
Distilled Water 400 ml
Component II K2HPO4 0.5 g
Distilled Water 200 ml
Component III
Sodium Lactate 3.5 g
Yeast Extract 1.0 g
Distilled Water 400 ml
Component IV Filter-sterilized 5%wt ferrous ammonium sulfate. Add 0.1 ml of this solution to 5.0 ml of medium prior to inoculation.
Table 4
Sessile cells count in undiluted and diluted medium
Medium (ATCC 1249 Medium) Sessile Cell Count (cells/cm2)
Full strength ≥106
Carbon source concentration diluted 10 times ≥105
Carbon source concentration diluted 100 times ≥104
Sodium citrate and sodium lactate not added ≥104

©2011 by NACE International. Requests for permission to publish this manuscript in any form, in part or
in whole, must be in writing to NACE International, Publications Division, 1440 South Creek Drive,
Houston, Texas 77084. The material presented and the views expressed in this paper are solely those of the
author(s) and are not necessarily endorsed by the Association.

©2011 by NACE International. Requests for permission to publish this manuscript in any form, in part or
in whole, must be in writing to NACE International, Publications Division, 1440 South Creek Drive,
Houston, Texas 77084. The material presented and the views expressed in this paper are solely those of the
author(s) and are not necessarily endorsed by the Association.
REFERENCES
1. S. B. Axelsen, T. Rogne, “Do micro-organisms „eat‟ metal?” Microbiologically influenced corrosion of industrial materials, Contract No. BRRT-CT98-5084, 1998.
2. NTSB Report, “Pipeline Accident Report Natural Gas Pipeline Rupture and Fire Near Carlsbad, New Mexico,” NTSB/PAR-03/01, National Transportation Safety Board, August 19, 2000.
3. NACE International Industry News, “Microbiologically influenced corrosion considered among the possible causes of the San Bruno blast,” Industry News, NACE International, September 24, 2010.
4. G. A. Jacobson, “Corrosion at Prudhoe Bay-a lesson on the line,” Materials Performances, 46(8): 27-34, 2007.
5. R. K. Thauer, E. Stackebrandt, W. A. Hamilton, “Energy metabolism phylogenetic diversity of sulphate-reducing bacteria,” In: Sulphate-Reducing Bacteria: Environmental and Engineered Systems, L. L. Barton and W. A. Hamilton (eds.), pp. 1-37, Cambridge University Press, Cambridge, 2007. 6. D. Novelli, C. E. ZoBell, “Assimilation of petroleum hydrocarbons by sulfate-reducing bacteria,” Journal of Bacteriology, 47: 447–448, 1944.
7. B. Dunsmore, T. Whitfield, “Corrosion by Sulfate Reducing Bacteria that Utilize Nitrate,” CORROSION/2004, Paper No. 04763. (Houston, TX: NACE 2004) 8. M. J. Feio, V. Rainha, M. A. Reis, A. R. Lino, I. T. E. Fonseca, “The Influence of the Desulfovibrio desulfuricans 14 ATCC 27774 on the corrosion of mild steel,” Materials and Corrosion, 51: 691-697, 2000. 9. C. A. H. von Wolzogen Kuhr, L.S. Vlugt vander, “The graphitization, of cast iron as an electrochemical process in anaerobic soils,” Water, 18: 147-165, 1934.
10. W. S. Borenstein, Microbiologically Influenced Corrosion Handbook, Industrial Press, New York, 1994.
11. D. Thierry, W. Sand, “Microbially Influenced Corrosion,” In: Corrosion Mechanisms in Theory and Practice, P. Marcus, J. Oudar (eds.), pp. 457-499, Marcel Dekker, New York, 1995.
12. H. D. Peck, “Bioenergetic Strategies of the Sulfate-Reducing Bacteria,” In: The Sulfate-Reducing Bacteria: Contemporary Perspectives, Odom, J.M., Singleton, R., Jr. (eds.), p.57, Springer, Berlin-New York, 1993.

©2011 by NACE International. Requests for permission to publish this manuscript in any form, in part or
in whole, must be in writing to NACE International, Publications Division, 1440 South Creek Drive,
Houston, Texas 77084. The material presented and the views expressed in this paper are solely those of the
author(s) and are not necessarily endorsed by the Association.
13. T. Gu, K. Zhao, S. Nesic, “A Practical Mechanistic Model for MIC Based on a Biocatalytic Cathodic Sulfate Reduction Theory,” CORROSION/2009, Paper No. 09390. (Houston, TX: NACE 2009) 14. T. Gu, D. Xu, “Demystifying MIC Mechanisms,” CORROSION/2010, Paper No. 10213. (Houston, TX: NACE 2010) 15. W. A. Hamilton, “Bioenergetics of sulphate-reducing bacteria in relation to their environmental impact,” Biodegradation, 9: 201–212, 1998. 16. F. Chen and M. R. Johns, “Relationship between substrate inhibition and maintenance energy of Chlamydomonas reinhardtii in heterotrophic culture,” Journal of Applied Phycology, 8: 15-19, 1996. 17. I. A. C Pereira, S. A. Haveman, G. Voordouw, “Biochemical, genetic and genomic characterization of anaerobic electron transport pathways in sulphate-reducing Delta proteobacteria,” In: Sulphate-Reducing Bacteria: Environmental and Engineered Systems, L. L. Barton and W. A. Hamilton (eds.), pp. 215-240, Cambridge University Press, Cambridge, 2007. 18. J. Larsen, B. Dunsmore, C. Schwermer, “Investigation Into Halfdan Water Injection Biofilm Using Chemical Microsensors,” CORROSION/2007, Paper No. 07507. ( Houston, TX: NACE 2007) 19. C. U. Schwermer, G. Lavik, R. M. M. Abed et al., “Impact of nitrate on the structure and function of bacterial biofilm communities in pipelines used for injection of seawater into oil fields,” Applied Environmental Microbiology, 74: 2841–2851, 2008. 20. B.W.A. Sherar, I.M. Power, P.G. Keech, S. Mitlin, G. Southam, D.W. Shoesmith, “Characterizing the Effect of Carbon Steel Exposure in Sulfide Containing Solutions to Microbially Induced Corrosion,” Corrosion Science, accepted (2010), doi: 10.1016/j.corsci.2010.11.027. 21. R. A. King, J. D. A. Miller, "Corrosion by the sulfate-reducing bacteria," Nature, 233: 491-492, 1971. 22. I. B. Beech, C. C. Gaylarde, “Recent advances in the study of biocorrosion- An overview,” Revista de Microbiología, 30: 177-190, 1999.