paleoenvironmental evolution of the east carpathian foredeep
TRANSCRIPT

Global and Planetary Change 103 (2013) 135–148
Contents lists available at SciVerse ScienceDirect
Global and Planetary Change
j ourna l homepage: www.e lsev ie r .com/ locate /g lop lacha
Paleoenvironmental evolution of the East Carpathian foredeep during the lateMiocene–early Pliocene (Dacian Basin; Romania)
M. Stoica a,⁎, I. Lazăr a, W. Krijgsman b, I. Vasiliev b, D. Jipa c, A. Floroiu a
a Department of Geology, Faculty of Geology and Geophysics, Bucharest University, Bălcescu Bd. 1, 010041, Romaniab Paleomagnetic Laboratory ‘Fort Hoofddijk’, Utrecht University, Budapestlaan 17, 3584 CD Utrecht, The Netherlandsc National Institute of Marine Geology and Geoecology, GeoEcoMar, Dimitrie Onciul Street 23-25, Bucharest, RO-70318, Romania
⁎ Corresponding author. Department of Geology, FacuBucharest University, Bălcescu Bd. 1, 010041, Romania.
E-mail address: [email protected] (M. Stoica
0921-8181/$ – see front matter © 2012 Elsevier B.V. Alldoi:10.1016/j.gloplacha.2012.04.004
a b s t r a c t
a r t i c l e i n f oArticle history:Received 27 May 2011Accepted 12 April 2012Available online 21 April 2012
Keywords:ParatethysCarpathian foredeepbiostratigraphypaleoenvironmentMessinianostracodsDacian Basin
The thick and continuous Mio-Pliocene sedimentary successions of the Focşani Depression in the Dacian Basinof Romania provide an excellent opportunity to study the paleoecological changes in the Eastern Paratethysduring the time when the Mediterranean and Black Sea experienced major sea level fluctuations related tothe closure and re-opening of the marine connection to the Atlantic Ocean during the Messinian Salinity Crisis.These successions form the basis of high-resolution magneto-biostratigraphic studies that allow a detailedcorrelation to the standard Geological Time Scale. Here, we analyze the paleoenvironmental evolution of theEast Carpathian foredeep by integrating micro- and macropaleontological data and sedimentological analyses.The ostracod andmollusc fossil associations from the Râmnicu Sărat river section indicate that the late Maeotiandepositional environment was characterized by shallow waters and littoral to fluvio-deltaic sediments. TheMaeotian–Pontian boundary (6.04 Ma) is marked by a marine ingression, comprising benthic (agglutinatedand calcareous) and planktonic (Streptochilus spp.) foraminifera and nanofossils. Following this marine ingres-sion, the Lower Pontian (Odessian; 6.04–5.8 Ma) fauna shows an increased bathymetry of the basin. Thepresence of ostracod species with eye tubercles indicates depositional environments within the photic zone(b100 m). The Middle Pontian (Portaferrian; 5.8–5.5 Ma) is marked by a widespread sea level loweringresulting in dominant fluvio-deltaic conditions. This ecostratigraphy demonstrates that the main Messiniansea-level draw down (at 5.6–5.5 Ma) occurred in mid-Portaferrian times. Paleoenvironmental indicatorsshow that the water level in the Foçsani Depression dropped less than 100 m during Mediterranean desiccation.The Dacian Basin remained filled with water, suggesting a positive hydrological balance for the region. This iscompatible with the presence of a shallow barrier at Dobrogea (the Galati passage), separating the DacianBasin from the Black Sea Basin during the lateMiocene. The Portaferrian–Bosphorian boundary (5.5 Ma) ismarkedby a second transgressive event, but this time without marine foraminifera. We conclude that the Dacian Basinformed a semi-isolated entity during the Portaferrian and experienced connectivity to the Black Seadomain during the Odessian and Bosphorian.
© 2012 Elsevier B.V. All rights reserved.
1. Introduction
The Miocene paleoenvironmental evolution of the Mediterraneanand Paratethys regions is largely influenced by the opening and clo-sure of the marine connections that strongly regulated the water ex-change between the two domains and with the Indian and Atlanticoceans (e.g. Harzhauser and Kowalke, 2002; Popov et al., 2006;Harzhauser et al., 2007; De Leeuw et al., 2010). This progressivelyresulted in restricted conditions, significant water level variations, de-velopment of endemic faunas and changes of salinities, culminating inthe deposition of massive evaporites during the Badenian in the
lty of Geology and Geophysics,Tel.: +40 21 3143508.).
rights reserved.
Paratethys (Peryt, 2006) and the Messinian in the Mediterranean(Hsü et al., 1973).
The Paratethys–Mediterranean water connection during theMessinian is still heavily debated and the location of such a marinegateway has not been proven so far (Popov et al., 2006; Krijgsmanet al., 2010). The present-day connection through the Bosphorus isbelieved to have originated much later in the Pliocene (e.g. Çagatayet al., 2006), but the presence of many fossils of Paratethyan affinityin the latest Messinian Mediterranean sequences (Esu, 2007) supportthe hypothesis that the Paratethys Sea drained into the Mediterraneanafter its desiccation phase, refilling it progressively with brackishwater and resulting in the establishment of Lago Mare facies all overthe Mediterranean Basin (Cita et al., 1978; Cosentino et al., 2006;Orszag-Sperber, 2006). It has also been shown that the water level inthe Black Sea dropped because of Mediterranean desiccation (Dinu etal., 2005; Gillet et al., 2007; Krijgsman et al., 2010; Vasiliev et al.,

136 M. Stoica et al. / Global and Planetary Change 103 (2013) 135–148
2011), indicating that a marine Paratethys–Mediterranean connectionmust have been present at Messinian times.
In this paper we focus on the thick and continuous sedimentarysuccessions of the Focşani Depression, one of the main late Miocenedepocenters in the Dacian Basin of Romania, to investigate thepaleoenvironmental changes in the Eastern Paratethys during theMessinian. We integrate paleoecological information from ostracods,foraminifera andmolluscs with sedimentological analyses and a recent-ly developed chronology (Krijgsman et al., 2010). This allows making adetailed reconstruction of the paleogeographic and paleoenvironmentalevolutions of the East Carpathian foredeep during the late Miocene andearly Pliocene.
2. Geological background
2.1. The Dacian Basin of the Eastern Paratethys
The Paratethys domain (Laskarev, 1924) developed as the northernbranch of the former Tethys Ocean and became progressively separatedfrom the southern Mediterranean branch by ongoing tectonic move-ments in the African–Eurasian collision process (Fig. 1). The initiationof Paratethys took place 35 millions of years ago, from where it subse-quently evolved as a large intra- and intercontinental marine domainthat extended from the Alps to the Aral Sea (Báldi, 1980; Rusu, 1988;Schultz et al., 2005). Until the Middle Miocene, Paratethys was in com-munication with the normal marine environments of the Mediterra-nean Basin and Indian Ocean. The different Paratethys basins, mainlythe Alpine–Carpathian Basin, the Black Sea Depression and the CaspianDepression, were intercommunicating, with periodical phases of isola-tion. By the end of the Middle Miocene (late Badenian–Sarmatian),
a
b
Late Eocene
Miocene
Tethys Ocean
Mediterranean
Paratethys
Fig. 1. Schematic paleogeographic maps of the a) late Eocene and b) late Miocene indicatingarea in the East Carpathian foredeep. Time scale for the Dacian Basin (after Vasiliev et al., 2
Paratethys communications with the Mediterranean and Indian Oceanwere restricted and eventually closed (Rögl, 1998). Subsequently, Para-tethys itself subdivided into several brackish (Lake Pannon) and low sa-linity restricted marine basins (Eastern Paratethys) during the lateMiocene, all of them developing their own endemic fauna.
The Carpathian orogenwas tectonically uplifted to become a barrierin Sarmatian times (~11–12 Ma), separating the Pannonian and Tran-sylvanian Basins of the Central Paratethys from the Dacian and EuxinianBasins of the Eastern Paratethys (Sanders et al., 1999; Cloetingh et al.,2004; Vasiliev et al., 2009). In the Central Paratethys, this resulted in amajor environmental change from marine (Sarmatian) to mainly freshwater environments (Pannonian), recently magnetostratigraphicallydated to take place between 11.6 and 11.3 Ma (Vasiliev et al., 2010;Paulissen et al., 2011; Ter Borgh et al., 2013-this volume; De Leeuw etal., 2013). The Dacian Basin evolved as a separate paleogeographic enti-ty where marine conditions prevailed much longer than in the CentralParatethys (Saulea et al., 1969). This resulted in the confusingnomenclature of a Sarmatian s.l. stage, commonly used in Eastern Para-tethys time scales, which was recently solved by using Volhynian,Bessarabian and Khersonain stages instead (Fig. 1c).
2.2. The Râmnicu Sărat section in the East Carpathian foredeep
The Dacian Basin developed as a part of the Eastern Paratethys,located in between the Southern Carpathians, Pre-Balkan area andthe Dobrogean High (Matenco and Bertotti, 2000). The eastern partof the Dacian Basin (called Focşani Depression because of its high subsi-dence character) comprises the thickest Neogene sedimentary coverof the Dacian Basin, approaching ~13 km (Tarapoanca et al., 2003).Excellent exposures are on the western flank of the basin along the
c
Paratethys retreat (after Blakey, 2011). The star locates the relative position of the study004 and Krijgsman et al., 2010) and its relation with the standard GTS.
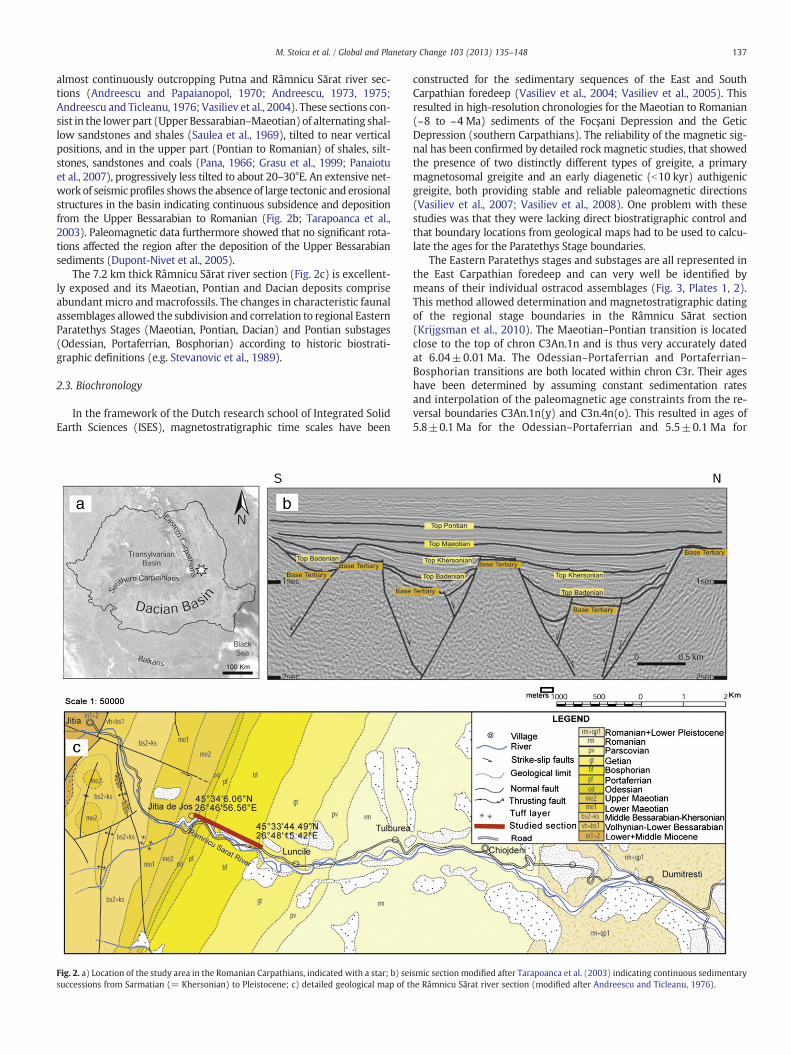
137M. Stoica et al. / Global and Planetary Change 103 (2013) 135–148
almost continuously outcropping Putna and Râmnicu Sărat river sec-tions (Andreescu and Papaianopol, 1970; Andreescu, 1973, 1975;Andreescu and Ticleanu, 1976; Vasiliev et al., 2004). These sections con-sist in the lower part (Upper Bessarabian–Maeotian) of alternating shal-low sandstones and shales (Saulea et al., 1969), tilted to near verticalpositions, and in the upper part (Pontian to Romanian) of shales, silt-stones, sandstones and coals (Pana, 1966; Grasu et al., 1999; Panaiotuet al., 2007), progressively less tilted to about 20–30°E. An extensive net-work of seismic profiles shows the absence of large tectonic and erosionalstructures in the basin indicating continuous subsidence and depositionfrom the Upper Bessarabian to Romanian (Fig. 2b; Tarapoanca et al.,2003). Paleomagnetic data furthermore showed that no significant rota-tions affected the region after the deposition of the Upper Bessarabiansediments (Dupont-Nivet et al., 2005).
The 7.2 km thick Râmnicu Sărat river section (Fig. 2c) is excellent-ly exposed and its Maeotian, Pontian and Dacian deposits compriseabundant micro andmacrofossils. The changes in characteristic faunalassemblages allowed the subdivision and correlation to regional EasternParatethys Stages (Maeotian, Pontian, Dacian) and Pontian substages(Odessian, Portaferrian, Bosphorian) according to historic biostrati-graphic definitions (e.g. Stevanovic et al., 1989).
2.3. Biochronology
In the framework of the Dutch research school of Integrated SolidEarth Sciences (ISES), magnetostratigraphic time scales have been
Fig. 2. a) Location of the study area in the Romanian Carpathians, indicated with a star; b) seisuccessions from Sarmatian (= Khersonian) to Pleistocene; c) detailed geological map of t
constructed for the sedimentary sequences of the East and SouthCarpathian foredeep (Vasiliev et al., 2004; Vasiliev et al., 2005). Thisresulted in high-resolution chronologies for the Maeotian to Romanian(~8 to ~4 Ma) sediments of the Focşani Depression and the GeticDepression (southern Carpathians). The reliability of the magnetic sig-nal has been confirmed by detailed rock magnetic studies, that showedthe presence of two distinctly different types of greigite, a primarymagnetosomal greigite and an early diagenetic (b10 kyr) authigenicgreigite, both providing stable and reliable paleomagnetic directions(Vasiliev et al., 2007; Vasiliev et al., 2008). One problem with thesestudies was that they were lacking direct biostratigraphic control andthat boundary locations from geological maps had to be used to calcu-late the ages for the Paratethys Stage boundaries.
The Eastern Paratethys stages and substages are all represented inthe East Carpathian foredeep and can very well be identified bymeans of their individual ostracod assemblages (Fig. 3, Plates 1, 2).This method allowed determination and magnetostratigraphic datingof the regional stage boundaries in the Râmnicu Sărat section(Krijgsman et al., 2010). The Maeotian–Pontian transition is locatedclose to the top of chron C3An.1n and is thus very accurately datedat 6.04±0.01 Ma. The Odessian–Portaferrian and Portaferrian–Bosphorian transitions are both located within chron C3r. Their ageshave been determined by assuming constant sedimentation ratesand interpolation of the paleomagnetic age constraints from the re-versal boundaries C3An.1n(y) and C3n.4n(o). This resulted in ages of5.8±0.1 Ma for the Odessian–Portaferrian and 5.5±0.1 Ma for
smic section modified after Tarapoanca et al. (2003) indicating continuous sedimentaryhe Râmnicu Sărat river section (modified after Andreescu and Ticleanu, 1976).

Fig. 3. Upper Miocene to Lower Pliocene ostracod assemblages and the main paleoenvironmental settings and events from the Râmnicu Sărat valley section of the Dacian Basin(modified after Krijgsman et al., 2010). Salinity and water level curves are schematic interpretations of the various paleoenvironmental indicators.
138 M. Stoica et al. / Global and Planetary Change 103 (2013) 135–148
the Portaferrian–Bosphorian boundary, respectively. The Pontian–Dacian boundary, and thus also the Bosphorian–Getian substageboundary, is located within the reversed chron C3n.2r at an age of4.70±0.05 Ma, in excellent agreement with earlier results fromthe south Carpathian foredeep (Vasiliev et al., 2005; Krijgsman etal., 2010).
3. Paleoenvironmental evolution of the East Carpathian foredeep
3.1. Methods
The Râmnicu Sărat river section was investigated in detail for mi-cropaleontological analyses primarily focusing on foraminifera andostracods. In addition, sedimentological and stratonomic investiga-tions have been performed on the Maeotian–Pontian successions.The sedimentological data coverage is discontinuous, as detailed study lo-cations have been chosen according to outcrop conditions, but additionalobservations have been made on smaller-sized outcrops located in
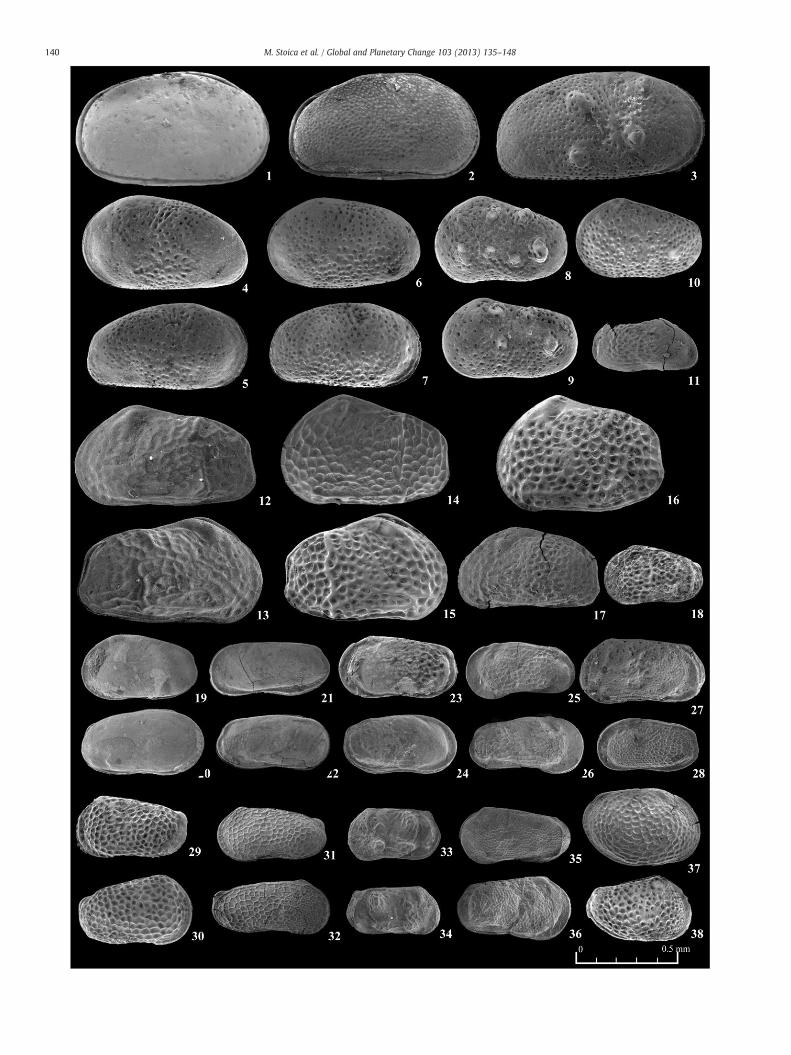
Plate 1.Most relevant ostracod species (Superfamily Cypridoidea) from the Upper Maeotianindividuals, external lateral views, LV = left valve, RV = right valve, RM=micropaleontolo2) Candona (Caspiocypris) pontica Sokač, carapace, view from RV, RM 207, Lower Pontian; 34) Candona (Typhlocyprella) ankae Krstić, RV, RM 215, Middle Pontian; 5) Candona (Typhltocypria) ossoinaensis Krstić. carapace, view from RV, RM 88, Upper Pontian; 7) Candona negsp., carapace, view from RV, RM 89, Upper Pontian; 9), 10) Candoniella sp., 9) carapace, view(Zalányi), carapace, view from RV, RM 207, Lower Pontian; 12) Candona (Hastacandona) hytoniella acuminata (Zalányi), RV, RM 94, Upper Pontian; 14) Pontoniella striata (MandelstamRV, RM 220, Upper Pontian; 16) Zonocypris membranae Stancheva, fragmented LV, RM 215from LV; 18) carapace, view from RV; all specimens from RM 207, Lower Pontian; 19), 20)all specimens from RM 88, Upper Pontian; 21), 22) Cypria sp.; 21) carapace, view from RV;niles), LV, RM 201, Upper Maeotian; 25) Ilyocypris sp., carapace, view from LV, RM 79, UpperRV; RM 88, Upper Pontian.
between the main exposures. Samples were processed by standard mi-cropaleontological methods, sieved over sieves of 63 μm and hand-picked under a microscope. Macropaleontological analyses on mol-lusks have been performed in less detail. Taxonomic identificationsand ecological inferences were based on Stancheva (1968, 1990);Sokač (1972, 1990a, 1990b); Hanganu (1974); Hanganu andPapaianopol (1977; 1982); Krstić and Stancheva (1989); Olteanu(1989, 1995); Papaianopol (1989); Meisch (2000); Gliozzi et al.(2005, 2008); Stoica et al. (2007); Harzhauser et al. (2008); andCziczer et al. (2009).
3.2. The Upper Maeotian
The Upper Maeotian of the Râmnicu Sărat river valley is well ex-posed on the outcrops downstream of Jitia village (Fig. 2c). In general,the Upper Maeotian is represented by alternations of sandstone andsilt. Many sandy units display channel structures, and wave ripples arefrequently observed on the upper part of the sandstone beds. Internal
and Pontian of the Râmnicu Sărat valley section (all valves of ostracods belong to adultgical samples no.); 1. Amplocypris ex. gr. dorsobrevis Sokač, LV, RM 215, Middle Pontian;) Candona (Caspiocypris) alta (Zalányi), carapace, view from RV, RM 88, Upper Pontian;ocypris) sp., carapace, view from the RV, RM 207, Lower Pontian; 6) Candona (Camp-lecta G.O. Sars, carapace, view from RV, RM 87, Upper Pontian; 8) Fabaeoformiscandonafrom RV, 10) LV; all from RM 207, Lower Pontian; 11) Candona (Hastacandona) lotzyisterica Kristić & Stancheva, carapace, view from RV, RM 207, Lower Pontian; 13) Pon-), RV, RM 220, Upper Pontian; 15) Pontoniella quadrata (Krstić), carapace, view from
, Middle Pontian; 17), 18) Candona (Zalanyiella) venusta (Zalányi); 17) carapace, viewCypria tocorjescui Hanganu; 19) carapace, view from LV; 20) carapace, view from RV;22) carapace, view from LV; RM 207, Lower Pontian; 23), 24) Pseudocandona sp. (juve-Pontian; and 26), 27) Bakunella dorsoarcuata (Zalányi); 26) LV; 27) carapace, view from


a
c
d
b e
1 cm
1 cm
1 mm
1 m
Sandstone
Clay
Wave ripples
Biostratigraphical sample
Microgastropods (Hydrobia spp., Theodoxus spp.)
Unionids
Legend:
Stra
tigra
phic
leve
l (m
)
Log
of
Upp
er M
aeot
ian
depo
sits
Subs
tage
Age
(M
a)
Pola
rity
Sy
nthe
tic lo
g
1 cm
Fig. 4. Stratigraphy, lithology and paleontology of the Upper Maeotian of the Râmnicu Sărat valley section: a) wave ripples on the upper bedding plane of a sand layer; b) ostracodassemblage dominated by Cyprideis sp.; c) accumulation of microgastropods (Hydrobia spp.); d) oolite sandstones with abundant gastropods (Theodoxus spp.); e) Psilunio (Psilunio)ex. gr. subrecurvus (Sinzow).
141M. Stoica et al. / Global and Planetary Change 103 (2013) 135–148
sedimentary structures consist of small scale cross-lamination, whichare occasionally disturbed, probably due to the elimination of waterduring sediment compaction. The most valuable paleoenvironmentalsignal comes from the presence of wave rippled sediments, pointingto deposition in shallow water, probably on the lower shoreface.Based on these structures the thick bedded sandy layers are interpretedas shallow water deposits that probably accumulated in the proximityof the river mouth under strong fluvial control.
The ostracods are highly abundant but very poor in species number(Figs. 3, 4, Plate 1). The ostracod assemblages consist of several speciesof Cyprideis (Cyprideis pannonica and C. ex. gr. torosa) and some rarecandonids like Pseudocandona sp. (juveniles) and Candoniella sp. Spo-radically, Leptocythere blanda is present.
Plate 2. Most relevant ostracod species (Superfamily Cytheroidea) from the Upper Maeotianbelong to adult individuals, external lateral views, LV = left valve, RV = right valve, RM = mRV, RM 116, Upper Maeotian; 2), 3) Cyprideis ex. gr. torosa Sars; 2) smooth specimen, carapPontian; 4), 5) Cyprideis sp. 1; 4) LV; 5) RV; RM 54, Lower Dacian; 6), 7) Cyprideis sp. 2; 6) LLV, male; RM 5, Lower Dacian; 10) Cytherissa sp., LV, RM 79, Upper Pontian; 11) Pontoleberismotasi Olteanu; 12) LV, male; 13) carapace, view from RV, female; RM 220, Upper Pontian; 194, Upper Pontian; 16) Tyrrhenocythere filipescui (Hanganu), LV, RM 88, Upper Pontian; 17)ex. gr. bosqueti (Livental), LV, RM 88, Upper Pontian; 19), 20) Amnicythere cymbula (Liventalgr. cymbula (Livental); 21) LV; 22) RV; RM 207, Lower Pontian; 23), 24) Amnicythere costSchneider; 25) LV; 26) RV; RM 207, Lower Pontian; 27) Leptocythere blanda, LV, RM 215, Up29), 30) Amnicythere andrusovi (Livental); 29) LV; 30) RV; RM 94, Upper Pontian; 31), 32)Maeotocythere bacuana (Livental); 33) LV; 34) RV; RM 220, Upper Pontian; 35), 36) MaeotoLivental, LV, RM 220, Upper Pontian; and 38) Loxoconcha babazananica Livental, RV, RM 96
The dominance of fresh–brackish water and littoral conditions isalso shown by the presence of abundant microgastropods likeHydrobiasp. and Teodoxus sp. and Unionidae bivalves— Psilunio (Psilunio) ex. gr.subrecurvus (Fig. 4).
The observed macro and micro faunas indicate low salinity(0–5‰) environments, typical of littoral setting or within lakesof short life time. In the upper part of the Maeotian, intercalationsof oolitic sandstones with micro-gastropods are common (Fig. 4c,d).
3.3. The Maeotian–Pontian transition
An important transgressive event takes place at the Maeotian–Pontian boundary, characterized by a sudden lithological change to
, Pontian and Lower Dacian of the Râmnicu Sărat valley section (all valves of ostracodsicropaleontological sample no.); 1) Cyprideis pannonica (Méhes), carapace, view from
ace, view from RV, RM 114, Upper Maeotian; 3) nodded specimen, RV, RM 226, UpperV; 7) RV; RM 65, Upper Pontian; 8), 9) Cytherissa bogatschovi Livental; 8) LV, female; 9)pontica (Stancheva), fragmented RV, RM 209, Lower Pontian; 12), 13) Tyrrhenocythere4), 15) Tyrrhenocythere ex gr. motasi Olteanu; 14) LV; 15) carapace, view from RV; RMTyrrhenocythere pannonicum (Olteanu), LV, RM 209, Lower Pontian; 18) Maeotocythere); 19) LV; 20) carapace, view from RV; RM 220, Upper Pontian; 21), 22) Amnicythere ex.ata Olteanu; 23) LV; 24) RV; RM 220, Upper Pontian; 25), 26) Amnicythere ex gr. lataper Maeotian; 28) Amnicythere ex. gr. polymorpha Olteanu, LV, RM 219, Upper Pontian;Amnicythere palimpsesta (Livental); 31) LV; 32) RV; RM 207, Lower Pontian; 33), 34)cythere incusa Olteanu; 35) LV; 36) RV; RM 203, Lower Pontian; 37) Loxoconcha petasa, Upper Pontian. (see on page 140)

142 M. Stoica et al. / Global and Planetary Change 103 (2013) 135–148
dominantly silty clays. This event coincides with a short-time replace-ment of the fresherwater fauna by a fauna of significantly higher salinity.These assemblages are dominated by the occurrence of benthonic calcar-eous foraminifers (species of Ammonia and Porosononion) and especiallyof agglutinated foraminifers (species of Ammotium). The enigmaticplanktonic foraminifera genus Streptochilus has also been observed inlarge numbers at the same level (Fig. 5). The biserial planktonic forami-nifera were earlier described from the Upper Maeotian deposits of theWestern Caucasus as belonging to the genus Bolivina, and some Bolivinaspecies were also reported from the Taman peninsula (Maissuradze,1988). Several morphological species of Streptochilus (Foraminifera)were described in the Miocene of the western Indian Ocean and theeastern Atlantic. They had previously been assigned to the benthicgenus Bolivina, but evidence on their apertural morphology, togetherwith accumulation rate data and isotopic composition shows that theylived as plankton, and should be assigned to the planktic genusStreptochilus (Smart and Thomas, 2006, 2007). It was suggested thatStreptochilus may have evolved polyphyletically, either from biserialplanktic or benthic ancestors, possibly in response to the occurrenceof relatively eutrophic environmental conditions caused by intermittentupwelling, leading to high algal growth rates but low transport efficien-cy of organic matter to the sea floor (Smart and Thomas, 2007). Recentinvestigations of the Upper Badenian and Sarmatian of the Transylva-nian Basin revealed particular, small sized planktonic foraminiferal as-semblages that include also the biserial foraminifera Streptochilusoccurring in relation to a transgressive event close to the end ofBadenian and provide evidence for a paleogeographic connection tothe Indo-Pacific area at that time (Filipescu and Silye, 2008).
Subs
tage
Age
Po
lari
ty
Synt
hetic
log
Stra
tigra
phic
leve
l (m
)
Log
of
Upp
er M
aeot
ian
/ L
ower
Pon
tian
inte
rval
1 cm
1 mm
Sandstone
Clay
Wave
Biostsamp
Legend:
Marls
Infl
ux o
f m
arin
e w
ater
Fig. 5. Stratigraphy, lithology and paleontology of the Maeotian–Pontian transition intervalagglutinated and calcareous benthic foraminifers; b), c) Ammotium sp.; d), e) Ammonia beccCongeria (Andrusoviconca) amygadaloides novorossica Sinzow rich layers.
Starting with the Pontian, a large diversification of the ostracod faunatook place, marked by a sudden increase of species of Pontoniella andCandona: Pontoniella acuminata, Candona (Hastacandona) lotzy,Candona (Hastacandona) hysterica, Candona (Zalanyiella) venusta,Fabaeformiscandona sp., Candona (Caspiocypris) alta, Candona(Caspiocypris) pontica (Fig. 3, Plate 1). Cyprideis species are still veryfrequent.
The transitional interval is further characterized by shell accumu-lations with the bivalve Congeria (Andrusoviconca) amygadaloidesnovorossica, a biostratigraphic marker for the Maeotian–Pontianboundary. The association of C. (A) amiygdaloides novorossica, micro-gastropods and unionids (Psilunio sp.) macrofauna is abruptly rep-laced by an association of limnocardiids (Fig. 5), marking the baseof the Pontian according to its original definition.
The Maeotian–Pontian boundary as defined here, is thus markedby an influx of marine water into the Paratethys, probably comingfrom the Mediterranean or alternatively from the Indian ocean(Krijgsman et al., 2010; Ruban, 2010). After this very rapid marinewater influx, the environment became less saline, as testified by theabundance and diversity of Candoninae in the Lower Pontian.
3.4. The Lower Pontian (Odessian)
As a consequence of the Maeotian–Pontian transgression, theLower Pontian starts with fine pelitic successions, deposited in deeperwaters (~100–150 m). The littoral and fluvial facies from the latestMaeotian are replaced by more distal ones; gray marls and clayswith frequent intercalations of ferruginous marls and silts (Fig. 6).
a
b
c
d
e
f
gh i
j k l1 cm 1 cm
1mm 0.5mm
ripples
ratigraphical le
Pseudoprosodacna sp
Congeria (Andrusoviconca) amygadaloides novorossica
of the Râmnicu Sărat valley section: a) micropaleontological assemblage dominated byarii (Linné); f), g) Porosononion ex. gr. subgranosus (Egger); h), i) Streptochilus sp.; j)–l)

143M. Stoica et al. / Global and Planetary Change 103 (2013) 135–148
An important rejuvenation of the ostracod fauna is observed in theLower Pontian, resulting in colonization by a large number of species:C. (Caspiocypris) alta, C. (Caspiocypris) pontica, Candona (Camptocypria)ossoinaensis, C. (Zalanyiella) venusta, C. (Hastacandona) hysterica,C. (Hastacandona) lotzyi, P. acuminata, Pontoniella quadrata, Pontoniellastriata, Fabaeformiscandona sp., Candona (Typhlocyprella) ankae, Cypriatocorjescui, Cypria sp., Bakunella dorsoarcuata, Cytherissa sp., C. pannonica,Tyrrhenocythere pannonicum, Amnicythere cymbula, Amnicythere ex.gr.,cymbula, Amnicythere costata, Amnicythere andrusovi, L. blanda (Figs. 3,6; Plates 1,2). The presence of ostracods with eye tubercles indicatesthe need of the photic zone for their development, in agreement withthe presence of green charophyta algae in the same samples. At thebase of the Lower Pontian there is a level with pyrite filled ostracods(Fig. 6), which is similarly observed in the Taman Peninsula of Russia(Krijgsman et al., 2010), pointing to large-scale dissoxic conditionsthroughout the Eastern Paratethys.
The Upper Maeotian mollusc fauna is replaced by limnocardiidsbivalves: Paradacna abichi, Pseudoprosodacna littoralis littoralis,Prosodacna sturi, Caladacna steindachneri, Didacna subcarinata,Limnocardium (Tauricardium) subsquamulosum, Monodacna (Pseu-docatilus) pseudocattillus and Congeria zagrabiensis (Fig. 6).
These faunal assemblages indicate that immediately after theshort marine influx, the salinity of the Dacian Basin waters decreased
Stra
tigr
aphi
c le
vel
Log
of
Low
er
Pont
ian
depo
sits
c
1 cm
1 mm
Subs
tage
Age
(M
a)
Pol
arity
Sy
nthe
tic
log
Sandstone
Clay
Biostratsample
Pseudop
Congeria amygad
Legend:
Marls
Fig. 6. Stratigraphy, lithology and paleontology of the Lower Pontian of the Râmnicu Sărat vtions rich in Limnocardiidae shells; c) micropaleontological assemblage from the basal partmarls rich in Paradacna abichi (R. Hoernes); f) Limnocardiids sandstone (Didacna sp., PseudPseudoprosodacna littoralis littoralis (Eichwald).
again because of the high influx of continental waters. The LowerPontian paleoenvironmentwas generally fresh to brackishwith salinitiesof 7–8‰ (Fig. 3).
3.5. The Middle Pontian (Portaferrian)
The Middle Pontian represents a regressive phase in the DacianBasin. There are a large number of fluvial sandstones (Fig. 7), fossilsoils, coaly clays and even thin coal levels. Rare occurrences of sand-stones with limnocardiids bivalves are found. The sedimentary recordbecomes scarce in fossils, although ostracods and molluscs typicalof shallow water environment (littoral, fluvial and even subaeriallyexposed) have been found.
The ostracod fauna is rather scarce if compared with the LowerPontian one (Fig. 3). The Middle Pontian species are represented by:Amplocypris ex. gr. dorsobrevis (a species with robust shell capableof living in littoral environment where sands are deposited in hydrody-namically active regimes), Candoniella sp., Zonocypris membranae(species able to quickly colonize temporary, short-living lakes),Cyprideis ex.gr. torosa (littoral species), Tyrrhenocythere motasi (aspecieswith robust shell capable of living in sandy–silty environments).These species are known to adapt rapidly in changing environmentsfrom the lakes located close to rivers, to marshes and lagoons (Plate 2).
a
d e
f
h
1 cm 1 cm
1 cm
1 cm
igraphical
rosodacna sp
(Andrusoviconca) aloides novorossica
Paradacna abichi
Congeria rhomboidea
Dreissena spp.
Didacna sp
b
g
alley section: a), b) dominant marly sediments with thin ferruginous sandy intercala-of Pontian dominated by Candonidae ostracods with carapaces filled with pyrite; d), e)ocatillus sp., Pseudoprosodacna sp.); and g), h) ferruginous sandy layer with abundant

Subs
tage
Age
(M
a)
Pola
rity
Sy
nthe
tic
log
Stra
tigr
aphi
c le
vel
Log
of
Mid
dle
Pont
ian
depo
sits
b
c e
fd 1cm
1cm
1cm 1cm
1cm
Sandstone
Clay
Biostratigraphical sample
Terrestrial gastropods
Legend:
Marls
Coaly clay
Lignite Viviparus spp.
Pseudoprosodacna sp.
Dreissena spp.
Unionids
a
Fig. 7. Stratigraphy, lithology and paleontology of the Middle Pontian of the Râmnicu Sărat valley section: a) sandy channel deposits separated by flood plain, lacustrine and con-tinental deposits; b)–d) layers with abundant Unionidae bivalves: b) Psilunio sp., c), d) Unio (Rumanunio) rumanus Tournouër; e) Cepaea sp.; and f) Theodoxus (Calvertia) slavonicus(Brusina).
144 M. Stoica et al. / Global and Planetary Change 103 (2013) 135–148
The molluscan fauna is also rare and represented by fresh waterlacustrine and fluvial forms: unioniids — Unio (Rumanunio) rumanusand dreiseniids — Dreissena polymorpha (Fig. 7). Frequently, planorbidgastropods typical of continental environments (temporary ponds andlakes) have been identified.
These sedimentological and faunistical changes were caused by animportant base level drop from basinal environments to flood plaindeposits. The changes in lithofacies were reflected in changes infauna that became taxonomically poorer and have visible effects inthe marginal areas of the basin (Stoica et al., 2007). The faunal assem-blages indicate fresh water environments (0–4‰) and a relativelysmall drop (~100 m) of the water level (Fig. 3).
3.6. The Upper Pontian (Bosphorian)
The Upper Pontian deposits accumulated in a more distal setting,where finer, pelitic sediments are alternated with sandy units. Thesefine deposits are represented by massive or stratified gray marlsand clays, rich in fossils, alternating with thinner sandstones andshell accumulations (Fig. 8). The Upper Pontian is marked by a gradualupward increase in littoral andfluvial deposits, evolving into deltaic andfluvial deposits toward the Pontian–Dacian boundary. This is caused bya transgressive episode that marks the base of Upper Pontian.
A new faunal bloom occurs and some of the species common inthe Lower Pontian become dominant again (Fig. 3). Next to the
Lower Pontian species, some new species appear that will continue toexist up to Dacian (Hanganu, 1974; Hanganu and Papaianopol, 1982;Olteanu, 1989, 1995; Stoica et al., 2007). The main ostracod speciesfrom the Upper Pontian of the Rîmnicu Sarat Valley are: A. ex. gr.dorsobrevis, C. (Caspiocypris) alta, C. (Camptocypria) ossoinaensis,Candona (Camptocypria) balcanica, C. (Zalanyiella) venusta, P.acuminata, P. quadrata, P. striata, Candona neglecta, Candoniella sp.,Cypria tocorjescui, Cypria sp., B. dorsoarcuata, Cytherissa boghatschovi,C. ex.gr. pannonica, Cyprideis ex. gr. torosa, Cyprideis sp.2, T. motasi, T.ex. gr.motasi, Tyrrhenocythere filipescui, A. cymbula, A. ex. gr. cymbula,A. costata, A. andrussovi, Amnicythere palimpsesta, Amnicythereex. gr. lata, Maeotocythere ex. gr. bosqueti, Maeotocythere bacuana,Maeotocythere incusa, Loxoconcha babazananica, Loxoconcha petasa(Plate 2). Two species of the Amplocypris genus are recorded in theUpper Miocene and Pliocene deposits of the Dacian Basin (Hanganuand Papaianopol, 1977;Olteanu, 1995; Floroiu et al., 2011):A. dorsobrevisSokač and Amplocypris odessaensis Ilnitzkaia. More additional morpho-metric studies (Danielopol et al., 2011a) are needed to find out whichis the real taxonomic affiliation of Amplocypris species in Paratethys.The presence of Leptocytheridae in the Paratethys Neogene is alsoquestioned. Many species formerly attributed to the genus Leptocythereare now re-evaluated and assigned to other leptocytherids generalike Amnicythere, Callistocythere, Euxinocythere, Maeotocythere,Mediocytherideis (Stancheva, 1968; Gliozzi et al., 2005; Boomer etal., 2010). There is still a lack of consensus in the literature on the

a b
c d
f g
e
h i
1cm
1cm 1cm 1cm
1cm 1cm 1mm
Subs
tage
Age
(M
a)
Pola
rity
Sy
nthe
tic
log
Stra
tigr
aphi
c le
vel
Log
of
Upp
er
Pont
ian
depo
sits
Sandstone
Clay
Biostratigraphical sample
Legend:
Marls
Coaly clay
Lignite
Viviparus spp.
Pseudoprosodacna sp. Dreissena spp.
Valenciennius anulatus
Limnocardium (Tauricardium) sp..
Pseudocatillus sp. Plagiodacana sp.
Fig. 8. Stratigraphy, lithology and paleontology of the Upper Pontian of the Râmnicu Sărat valley section; a) the transition from littoral sandy deposits of Middle Pontian to morebasinal fine grained sediments at the base of Upper Pontian; b) wave ripples on the upper bedding plane of a sandy layer from the Middle Pontian–Upper Pontian boundary interval;c) ostracod assemblage from the basal part of the Upper Pontian marked by the abundancy of Tyrrhenocythere spp.; d)–i) most abundant mollusc species from the basal part of theUpper Pontian; d) Plagiodacna sp.; e), f) Pseudocatillus sp.; g) Chartchoncha bayerni (R. Hoernes); h) Limnocardium (Tauricardium) sp.; and i) Valenciennius annulatus Rousseau.
145M. Stoica et al. / Global and Planetary Change 103 (2013) 135–148
generic assignment of leptocytherid species (Namiotko et al., 2011) anddetailed comparative morphological studies must be developed for thisgroup of ostracods (Danielopol et al., 2011b).
The bivalves are represented by numerous species that are mostlytypical for brackish waters (Papaianopol, 1989): D. polymorpha,Prosodacana (Prosodacna) mrazeci, Prosodacana (Prosodacna) savae,Prosodacana sturi, Pseudocatillus sp., Plagiodacna sp., C. steindachneri,Limnocardium nobile, Limnocardium sp., D. subcarinata, Lunadacna lunae,Phyllocardium planum giganteum, L. (Tauricardium) subsquamulosum,Limnocardium (Tauricardium) sp. Unio (Rumanunio) rumanus, Congeriabotenica, and Chartoconcha bayerni. (Fig. 8). Gastropods are also abun-dant and among them are: Viviparus acatinoides, Viviparus neumayrineumayri, Lithoglyphus sp., Bulimus (Tylopoma) speciosus, Melanopsis(Melanopsis) decolata, Zagrabica reticulata, Valenciennius annulatus, andValenciennius sp.
Paleoenvironmentally, the base of the Bosphorian corresponds toa second transgressive event in the Dacian Basin, showing a majorfaunal change in the ostracod assemblages and a lithological changeto more basinal sequences. The Portaferrian–Bosphorian transitionis marked by a second bloom of ostracod fauna that indicate salinitiesof 7–8‰ during the Upper Pontian (Fig. 3).
4. Discussion
4.1. Dacian Basin water level changes during the late Miocene–earlyPliocene
The presence of agglutinated and calcareous foraminifera of ma-rine origin at the Maeotian–Pontian boundary interval suggests thata major flooding event has taken place in the Dacian Basin by marinewaters, probably by establishing a connection to the Mediterraneanor Indian Ocean. An Odessian transgression has also been recognizedin seismic profiles from the western Dacian Basin (Getic Depression)where the Lower Pontian corresponds to a progradational–aggradationalunit developed during rising water levels (Leever et al., 2010). A LowerPontian transgression has further been documented in other Paratethysbasins and is biostratigraphically marked by a migration of faunal ele-ments from the Pannonian Basin (Hungary) into the Eastern Paratethysand by a migration of typical Aegean species into the Black Sea domain(Stevanovic et al., 1989; Popov et al., 2006). In addition, a marine nano-fossil influx, comprising assemblages that are correlative to SubzoneNN11b, has been reported from the Maeotian–Pontian boundary inter-val in the Dacian Basin (Marunteanu and Papaianopol, 1998). Fossil

146 M. Stoica et al. / Global and Planetary Change 103 (2013) 135–148
assemblages from the Taman Peninsula of Russia show similar marineassociations at the Maeotian–Pontian boundary interval indicatingthat this transgression extended at least into the eastern part of theBlack Sea Basin (Krijgsman et al., 2010).
A major water-level drop in the Dacian Basin took place duringPortaferrian times (between 5.8 and 5.5 Ma) and resulted in the re-establishment of fluvio-deltaic conditions in the Focşani Depression.Our paleontological data indicate that depositional environmentswithin the photic zone (b100 m) abruptly change to faunal assem-blages typical of shallow waters (~0–50 m) and terrestrial environ-ments during the Portaferrian. This implies that only a relativelysmall water level drop has taken place (~100 m) in the Dacian Basinduring the Portaferrian, although the exact amplitude is still a matterof debate. In seismic profiles of the western part of the Dacian Basin,the Portaferrian shows mainly progradational units, deposited duringperiods of low water levels (Leever et al., 2010). The correspondingwater level drop is here roughly estimated at a maximum of 200 m,based on the elevation difference between the shelf edges aboveand below the sequence boundary. The other regions of the DacianBasin that developed in water depths of ~100 m in the early Pontianstarted to evolve in deltaic, fluvial, lacustrine and/or littoral environ-ments at the beginning of the Middle Pontian. This indicates that thewater level in the entire Dacian Basin dropped during the Porta-ferrian, which would be compatible with the presence of a shallowbarrier at Dobrogea, separating the Dacian Basin from the Black SeaBasin.
The Upper Pontian is marked by a second transgression in the EastCarpathian foredeep. Interpolation assuming constant sedimentationrates suggests that this Bosphorian transgression started at an age of5.5±0.1 Ma. In thewestern Dacian Basin, seismic profiles show a trans-gressive system tract at the base of the Bosphorian as well (Leever,2007). In the Topolog region of the south Carpathian foredeep, the Bos-phorian is found transgressive on Maeotian deposits while the upper-most Maeotian, Odessian and Portaferrian are missing (Stoica et al.,2007; Floroiu et al., 2011). Our detailed micropaleontological studiesshow that the base of the Bosphorian consists of fresh to brackishwater ostracods, indicating a continental origin for this transgression,probably related to a regional climatic change toward a positive hydro-logical balance for the Paratethys region (e.g. Krijgsman et al., 2010).
A Pliocene marine flooding of the Dacian Basin was previouslysuggested based on the Hinova section (western Dacian Basin),which was interpreted to represent the bottom set of a ‘Gilbert fandelta’ produced by Mediterranean inflow water (Clauzon et al., 2005).We have resampled this Hinova section of Clauzon et al. (2005); theirFig. 6 for biostratigraphic dating. Our micropaleontological analysesshow that this section corresponds to the Lower Pontian (6.0–5.8 Ma),and thus not to the Pliocene, because it comprises the Odessian ostra-cods, C. (Caspiocypris) alta, C. (Camptocypria) balcanica, P. acuminata,P. striata, C. tocorjescui, B. dorsoarcuata, A. andrusovi, Amnicythere sp.,Mediocytherideis sp. and abundant limnocardiid bivalves, especiallyP. abichi. In addition, recent micropaleontological and stratigraphicstudies of the assumed foreset beds (Clauzon et al., 2005) clearly indicatea Badenian–Sarmatian age for these coarse clastic sediments that aremost likely tectonically tiltedMiddleMiocene alluvial fan conglomerates(Marinescu, 1978; Jipa et al., 2011).
4.2. Paratethys–Mediterranean connectivity during the late Miocene andearly Pliocene
The Black Sea itself has experienced significant water levelchanges during the Portaferrian. The sedimentary succession exposedat the Black Seamargin of Russia (TamanPeninsula) shows a conspicuousenvironmental change in Portaferrian, by brackish marls with Pontianfauna indicative of the lower photic zone (50–100 m deep) abruptlychanging to condensed coastal sequences of reddish sands, attributedthere to the base of the Kimmerian (= Middle Portaferrian,
Krijgsman et al., 2010). This indicates that the Black Sea experienceda sea level drop of at least 50–100 m as well.
Drilling in the Black Sea Basin (DSDP Leg 42B) had earlier revealeda peculiar sedimentary layer at the Mio-Pliocene boundary, interpretedas shallow, supratidal and intertidal sediments in the otherwise deep-water sequence and as caused by a sudden drastic (~1600m) loweringof the Black Seawater-level (Hsü andGiovanoli, 1979). In addition, seis-mic profiles of the Black Sea Basin show evidence of deep canyon cut-ting (Dinu et al., 2005; Gillet et al., 2007). This major down dropscenario of the Black Sea implies a negative water budget for the Para-tethys. In contrast, the hydrological balance of the Black Sea basin isalso considered as positive to explain overspilling of Paratethys watersduring the Mediterranean Lago Mare facies (Cita et al., 1978; Esu,2007; Gliozzi and Grossi, 2008). In that scenario it is envisaged thatthe water level of the Black Sea only dropped to the sill height of thepaleo-Bosphorus. The magnitude and, in some cases, even the sign ofthe Paratethys sea or lake level changes during the Messinian are thusstill seriously debated and subject to ongoing controversy.
Paleogeographic data from the Aegean region indicate that no(significant) gateway existed between Mediterranean and Paratethys(Çagatay et al., 2006) and that only ephemeral marine incursions tookplace in the Eastern Paratethys during the Mio-Pliocene period(Stevanovic et al., 1989; Marunteanu and Papaianopol, 1998; Popov etal., 2006). Consequently, water level in the Paratethys could beexpected to havefluctuated in relation toMessinian paleoclimatologicalchanges.
Our biostratigraphic results suggest that a marine connectionexisted at the Maeotian–Pontian boundary at 6.05 Ma, evidenced bythe influx of foraminifera in the Dacian Basin. This is further confirmedby earlier observations of a short calcareous nanofossil influx at theMaeotian/Pontian boundary interval (Marunteanu and Papaianopol,1998). According to earlier paleogeographic reconstructions, the mostlikely marine connection is to the Mediterranean (Popov et al., 2006),but the available biostratigraphic data also do not exclude a gatewayto the Indian Ocean (Krijgsman et al., 2010). Our new biochronologyfor the Dacian Basin furthermore shows that the successively youngernanofossil influx at the Pontian–Dacian boundary, previously suggestedto correlate to the Zanclean flooding event of the Mediterranean(Marunteanu and Papaianopol, 1998), is now dated at 4.7 Ma.
5. Conclusions
New biostratigraphic data from the thick and continuous sedi-mentary successions of the Focşani Depression are incorporated intoa magnetostratigraphic time frame. The integration of paleoecologicalinformation from ostracods, foraminifera and molluscs with sedimen-tary structures enables detailed reconstructions of the paleogeo-graphic and paleoenvironmental evolutions of the East Carpathianforedeep during the Messinian.
• The Upper Maeotian is marked by sedimentary structures that indi-cate the coexistence of fluvial and littoral environments. The faunalrecord suggests low salinity (0–4‰) environments, typical ofperennial-lake or littoral setting or of temporary lakes and ponds.
• An important transgression takes place at the Maeotian–Pontianboundary, characterized by a short-time paleoenvironmentalchange to significantly higher salinities (20–30‰). The faunal assem-blages are dominated by the occurrence of benthic (agglutinated andcalcareous) and planktonic foraminifera. It is the first time that thesehave been observed at the Maeotian/Pontian interval in the DacianBasin, and they indicate an influx of marine waters at 6.05 Ma.
• The Lower Pontian starts with fine pelitic sediments, deposited indeeper water, but still within the photic zone. The salinity decreasedto 7–8‰ because of a positive water balance determined by thehigher influx of continental waters.

147M. Stoica et al. / Global and Planetary Change 103 (2013) 135–148
• The Middle Pontian shows a base level drop with visible effects inthe marginal areas of the basin. It is scarce in fossils, and mainly os-tracods and molluscs typical of shallow and fresh water (0–4‰) la-custrine environments have been found, species known to adaptrapidly to changing environments from lakes located close to rivers.
• The Upper Pontian deposits accumulated in a more distal setting,with finer pelitic sediments, alternated with sandy units. Theupper part is marked by an increase in littoral and fluvial deposits,evolving into deltaic and fluvial deposits (salinity between 7 and8‰) toward the Pontian–Dacian boundary as a consequence of pro-gressive basin filling with sediments.
Acknowledgments
We thank especially Georghe Popescu for his help with bio-stratigraphical analyses of the Râmnicu Sărat data (foraminifera). Weacknowledge the constructive comments of Oleg Mandic and twoanonymous reviewers. This work was financially supported by theNetherlands Research Centre for Integrated Solid Earth Sciences(ISES) and the Netherlands Geosciences Foundation (ALW) withsupport from the Netherlands Organization for Scientific Research(NWO). This research was also supported by CNCSIS Romaniafunding (ID 960 and 1 Euroc).
References
Andreescu, I., 1973. Precizari asupra limitelor etajului Meotian. Acad. RSR, Stud. Cercet.Geol. Geofiz.Geogr., Ser. Geol. 18/2, 541–558.
Andreescu, I., 1975. Limitele si subdiviziunile Pontianului. Acad. RSR, Stud. Cercet. Geol.Geofiz.Geogr., Ser. Geol. 20/2, 235–245.
Andreescu, I., Papaianopol, I., 1970. Biostratigrafia depozitelor sarmatiene dintre vaileMilcov si Râmnicu Sărat. Acad. RSR, Stud. Cercet. Geol. Geofiz.Geogr., Ser. Geol.15/2, 499–512.
Andreescu, I. and Ticleanu, N., 1976. Geological map 1:50 000, sheet 113b, Dumitreşti.IGR, Bucharest.
Báldi, T., 1980. The early history of the Paratethys. Földtani Közlöny, Bulletin of theHungarian Geological Society 110, 456–472.
Blakey, R., 2011. Paleogeographic Maps in Rectangular Format: Eocene and Miocene. ,http://www2.nau.edu/rcb7/rect_globe.html2011.
Boomer, I., Guichard, F., Lericolais, G., 2010. Late Pleistocene to Recent ostracod assem-blages from the western Black Sea. Journal of Micropalaeontology 29, 119–133.
Çagatay, M.N., Görür, N., Flecker, R., Sakinç, M., Tünoglu, C., Ellam, R., Krijgsman, W.,Vincent, S., Dikbas, A., 2006. Paratethyan–Mediterranean connectivity in the Seaof Marmara region (NW Turkey) during the Messinian. Sedimentary Geology188–189, 171–187.
Cita, M.B., Wright, R.C., Ryan, W.B.F., Longinelli, A., 1978. Messinian paleoenvironments.In: Hsü, K.J. (Ed.), Initial Reports of the Deep Sea Drilling Project, pp. 1003–1035.
Clauzon, G., Suc, J.P., Popescu, S.M., Marunteanu, M., Rubino, J.L., Marinescu, F., Melinte, M.C.,2005. Influence of Mediterranean sea-level changes on the Dacic Basin (Eastern Para-tethys) during the late Neogene: the Mediterranean Lago Mare facies deciphered.Basin Research 17, 437–462.
Cloetingh, S.A.P.L., Burov, E., Matenco, L., Toussaint, G., Bertotti, G., Andriessen, P.A.M.,Wortel, M.J.R., Spakman, W., 2004. Thermo-mechanical controls on the mode ofcontinental collision in the SE Carpathians (Romania). Earth and Planetary ScienceLetters 218, 57–76.
Cosentino, D., Federici, I., Cipollari, P., Gliozzi, E., 2006. Environments and tectonic in-stability in central Italy (Garigliano Basin) during the late Messinian Lago-Mare epi-sode: new data from the onshore Mondragone 1 well. Sedimentary Geology188–189, 297–317.
Cziczer, I., Magyar, I., Pipík, R., Böhme, M., Stjepan, C., Bakrač, K., Sütő-Szentai Lantos, M.,Babinszki, E., Müller, P., 2009. Life in the sublittoral zone of long-lived Lake Pannon:paleontological analysis of the Upper Miocene Szák Formation,Hungary. Internation-al Journal of Earth Sciences (Geologische Rundschau) 98, 1741–1766.
Danielopol, D.L., Gross, M., Harzhauser, M., Minati, K., Piller, W.E., 2011a. How and whyto achieve greater objectivity in taxonomy, exemplified by a fossil ostracod(Amplocypris abscissa) from the Miocene Lake Pannon. Joannea Geologie und Pala-eontologie 11, 273–326.
Danielopol, D.L., Gross, M., Namiotko, T., Minati, K., Piller, W.E., Harzhauser, M., 2011b.Comparative morphology of ostracod Leptocytheridae — a prospect for better un-derstanding the origin and evolution of selected Amnicythere taxa in long-livedLake Pannon (late Miocene). Joannea Geologie und Palaeontologie 11, 50–52.
De Leeuw, A., Bukowski, K., Krijgsman, W., Kuiper, K.F., 2010. Age of the Badenian sa-linity crisis: impact of Miocene climate variability on the Circum-Mediterraneanregion. Geology 38, 715–718.
De Leeuw, A., Filipescu, S., Matenco, L., Krijgsman, W., Kuiper, K., Stoica, M., 2013. Pa-leomagnetic and chronostratigraphic constraints on the evolution of the MiddleMiocene Transylvanian Basin (Romania) and implications for Central Paratethys
stratigraphy and emplacement of the Tisza-Dacia plate. Global and PlanetaryChange 103, 82–98.
Dinu, C., Wong, H.K., Tambrea, D., Matenco, L., 2005. Stratigraphic and structural charac-teristics of the Romanian Black Sea shelf. Tectonophysics 410, 417–435.
Dupont-Nivet, G., Vasiliev, I., Langereis, C.G., Krijgsman, W., Panaiotu, C., 2005. Neogenetectonic evolution of the southern and eastern Carpathians constrained by paleo-magnetism. Earth and Planetary Science Letters 236 (1–2), 374–387.
Esu, D., 2007. Latest Messinian “Lago-Mare” Lymnocardiinae from Italy: close relationswith the Pontian fauna from the Dacic Basin. Geobios 40 (3), 291–302.
Filipescu, S., Silye, L., 2008. New Paratethyan biozones of planktonic foraminifera de-scribed from the Middle Miocene of the Transylvanian Basin (Romania). GeologicaCarpathica 59 (6), 537–544.
Floroiu, A., Stoica, M., Vasiliev, I., Krijgsman, W., 2011. Maeotian/Pontian ostracods inthe Badislava — Topolog area (south Carpathian foredeep, Romania). Geo-Eco-Ma-rina 17, 177–184.
Gillet, H., Lericolais, G., Rehault, J.-P., 2007. Messinian event in the Black Sea: evidenceof a Messinian erosional surface. Marine Geology 244, 142–165.
Gliozzi, E., Grossi, F., 2008. Late Messinian Lago-Mare ostracod palaeoecology: a correspon-dence analysis approach. Palaeogeography, Palaeoclimatology, Palaeoecology 264(3–4), 288–295.
Gliozzi, E., Rodriguez-Lazaro, J., Nachite, D., Martin-Rubio, M., Bekkali, R., 2005. An over-view of Neogene brackish leptocytherids from Italy and Spain: biochronologicaland palaeogeographical implications. Palaeogeography, Palaeoclimatology, Palaeo-ecology 225, 283–301.
Grasu, C., Catana, C., Bobos, I., 1999. Molasa Carpatilor Orientali, Petrografie sisedimentogeneza, Editura Tehnica, Bucuresti, p. 227 (in Romanian).
Hanganu, E., 1974. Observation sur l'ostracofaune pontienne de la région comprise a lavallée du Danube et la vallée du Motru. Revista Española de Micropaleontologia VI(3), 335–346.
Hanganu, E., Papaianopol, I., 1977. Les subdivisions du Dacien fondées sur les associa-tions de malacofaune et l'ostracofaune. Bulletin de la Société belge de Géologie85, 63–88.
Hanganu, E.N., Papaianopol, I., 1982. Associations significatives du Pontien du BassinDacique (Roumanie). Bulletin de la Société belge de Géologie 91, 51–59.
Harzhauser, M., Kowalke, T., 2002. Sarmatian (Late Middle Miocene) gastropod assem-blages of the Central Paratethys. Facies (46), 57–82.
Harzhauser, M., Piller, W.E., Latal, C., 2007. Geodynamic impact on the stable isotopesignatures in a shallow epicontinental sea. Terra Nova 19 (5), 324–330.
Harzhauser, M., Kern, A., Soliman, A., Minati, K., Piller, W.E., Danielopol, D.E., Zuschin, M.,2008. Centennial- to decadal scale environmental shifts in and around Lake Pannon(Vienna Basin) related to a major late Miocene lake level rise. Palaeogeography,Palaeoclimatology, Palaeoecology 270, 102–115.
Hsü, K.J., Giovanoli, F., 1979. Messinian event in the Black Sea. Palaeogeography,Palaeoclimatology, Palaeoecology 29, 75–93.
Hsü, K.J., Ryan, W.B.F., Cita, M.B., 1973. Late Miocene desiccation of the Mediterranean.Nature 242, 240–244.
Jipa, D., Stoica, M., Andreescu, I., Floroiu, A., Maximov, G., 2011. Zanclean Gilbert-typefan deltas in the Turnu Severin area (Dacian Basin, Romania). A critical analysis.Geo-Eco-Marina 17, 177–187.
Krijgsman, W., Stoica, M., Vasiliev, I., Popov, V.V., 2010. Rise and fall of the Paratethys Seaduring the Messinian salinity crisis. Earth and Planetary Science Letters 290, 183–191.
Krstić, N., Stancheva, N., 1989. Ostracods of Eastern Serbia and Northern Bulgaria withnotice on a Northern Turkey assemblage. In: Malez, M., Stevanovic, P. (Eds.),Chronostratigraphie und Neostratotypen, pp. 753–819. Bd. VIII, Pontien Pl1, JAZUet SANU, Zagreb-Beograd.
Laskarev, V., 1924. Sur le equivalents du Sarmatien superieur en Serbie. Recueil detraveaux offert a M. Jovan Cvijic par ses amis et collaborateurs, pp. 73–85.
Leever, K.A., 2007. Foreland of the Romanian Carpathians: Controls on late OrogenicSedimentary Basin Evolution and Paratethys Paleogeography. VU University Am-sterdam, Amsterdam. 182 pp.
Leever, K., Matenco, L., Rabagia, T., Cloetingh, S., Krijgsman,W., Stoica, M., 2010. Messiniansea level fall in the Dacic Basin (Eastern Paratethys): palaeogeographical implicationsfrom seismic sequence stratigraphy. Terra Nova 22, 12–17, http://dx.doi.org/10.1111/j.1365-3121.2009.00910.x.
Maissuradze, L.S., 1988. Foraminifery meotisa zapadnoi Gruzii. Metzniereba. (73 pp., 32pls. Tbilisi. in Russian).
Marunteanu, M., Papaianopol, I., 1998. Mediterranean nannoplankton in the Dacicbasin. Rom. J. Stratigraphy 78, 115–121.
Marinescu, F., 1978. Stratigrafia Neogenului Superior din Sectorul Vestic al BazinuluiDacic. Edit. Acad. R.S.R.Marunteanu, M. and Papaianopol, I., 1998. Mediterraneannannoplankton in the Dacic basin. Romanian Journal of Stratigraphy 78, 115–121.
Matenco, L., Bertotti, G., 2000. Tertiary tectonic evolution of the external East Car-pathians (Romania). Tectonophysics 316, 255–286.
Meisch, C., 2000. Freshwater Ostracoda of Western and Central Europe. SpectrumAcademicher Verlag, Heidelberg-Berlin, p. 522.
Namiotko, T., Danielopol, D.L., Belmecheri, S., Gross, M., von Grafenstein, U., 2011. OnLeptocytheridae ostracods of long-lived Lake Ohrid (Albania/Macedonia). JoanneaGeologie und Palaeontologie 11, 151–153.
Olteanu, R., 1989. La faune d'ostracodes pontiens du Bassin Dacique. Chronostratigraphieund Neostratotypen Pliozän Neogen der Zentrale Paratethys, Bd. VIII Pontien, Jugosl.Akad. Wissen und Künste, pp. 722–726.
Olteanu, R., 1995. Dacian ostracods. In: Marinescu, F., Papaianopol, I. (Eds.), Pliozän Pl1Dazien. Chronostratigraphie und Neostratotypen. Neogen der Zentrale Paratethys.Editura Academeiei Romane, Bucharest, p. 530.
Orszag-Sperber, F., 2006. Changing perspectives in the concept of “Lago-Mare” in Medi-terranean late Miocene evolution. Sedimentary Geology 188–189, 259–277.

148 M. Stoica et al. / Global and Planetary Change 103 (2013) 135–148
Pana, I., 1966. Studiul depozitelor Pliocene din regiunea cuprinsa intre valea Buzaului sivalea Balaneasa. Studii tehnice si economice, Seria J-Stratigrafie, nr.1: Bucharest.
Panaiotu, C.E., Vasiliev, I., Panaiotu, C.G., Krijgsman, W., Langereis, C.G., 2007. Provenanceanalysis as a key to orogenic exhumation: a case study from the East Carpathians(Romania). Terra Nova 19, 120–126.
Papaianopol, I., 1989. Considération sur le mollusques pontiens de Roumanie.Chronostratigraphie und Neostratotypen Pliozän Neogen der Zentrale Paratethys,Bd. VIII Pontien, Jugosl. Akad. Wissen und Künste, pp. 582–591.
Paulissen, W., Luthi, S.M., Grunert, P., Ćorić, S. And, Harzhauser, M., 2011. Integratedhigh-resolution stratigraphy of a Middle to late Miocene sedimentary sequencein the central part of the Vienna Basin. Geol. Carp. 62, 155–169.
Peryt, T.M., 2006. The beginning, development and termination of the Middle Badeniansalinity crisis in Central Paratethys. Sedimentary Geology 188–189, 379–396.
Popov, S.V., Shcherba, I.G., Ilyina, L.B., Nevesskaya, L.A., Paramonova, N.P., Khondkarian, S.O.,Magyar, I., 2006. Late Miocene to Pliocene papaeogeography of the Paratethys and itsrelation to the Mediterranean. Palaeogeography, Palaeoclimatology, Palaeoecology238, 91–106.
Rögl, F., 1998. Palaeogeographic considerations for Mediterranean and Paratethys sea-ways (Oligocene to Miocene). Annalen des Naturhistorischen Museums in Wien99, 279–310.
Ruban, D.A., 2010. Stratigraphic evidence of a late Maeotian (late Miocene) punctuatedtransgression in the Tanais Palaeobay (northern part of the Eastern Paratethys,South-West Russia). Geologos 16 (3), 169–181.
Rusu, A., 1988. Oligocene events in Transylvania (Romania) and the first separation ofParatethys, D.S. Institute of Geological Geofiz 72–73, 207–223.
Sanders, C.A.E., Andriessen, P.A.M., Cloetingh, S.A.P.L., 1999. Life cycle of the EastCarpathian orogen: erosion history of a doubly vergent critical wedge assessedby fission track thermochronology. Journal of Geophysical Research B: SolidEarth 104 (B12), 29095–29112.
Saulea, E., Popescu, I., Sandulescu, M., 1969. Lithofacies Atlas, VI Neogene, Sheets 4–8and 10–13. Institutul de Geologie si Geofizica.
Schultz, H.-M., Bechtel, A., Sachsenhofer, R.F., 2005. The birth of the Paratethys duringthe Oligocene: from Tethys to an ancient Black Sea analogue. Global and PlanetaryChange 49, 163–176.
Smart, C.W., Thomas, E., 2006. The enigma of early Miocene biserial planktic foraminif-era. Geology 34, 1041–1044.
Smart, C.W., Thomas, E., 2007. Emendation of the genus Streptochilus Brönnimann andResig, 1971 (Foraminifera), and new species from the Lower Miocene of the Atlan-tic and Indian Oceans. Micropaleontology 53 (1–2), 73–103.
Sokač, A., 1972. Pannonian and Pontian ostracode fauna of Mt. Medvednica. Palaeontol.Jugosl., Zagreb 2, 1–140.
Sokač, A., 1990a. The Pontian in Croatia. In: Stevanovič, P., Nevesskaja, L.A., Marinescu,Fl., Sokač, A., Jámbor, Á. (Eds.), Chronostratigraphie und Neostratotypen: Neogender Westlichen (“Zentrale”) Paratethys 8. Pontien (1989), Jazu and Sanu, Zagreb–Beograd, pp. 180–194.
Sokač, A., 1990b. Pontian ostracod fauna in the Pannonian Basin. In: Stevanovič, P.,Nevesskaja, L.A., Marinescu, Fl, Sokač, A., Jámbor, Á. (Eds.), Chronostratigraphie
und Neostratotypen: Neogen der Westlichen (“Zentrale”) Paratethys 8. Pontien(1989) Jazu and Sanu, Zagreb–Beograd, pp. 672–721.
Stancheva, M., 1968. New data on the subfamily Leptocytherinae Hanai, 1957. BulgarianAcademy of Sciences, Sofia Bulletin of the Geological Institute 17, 37–48.
Stancheva, M., 1990. Upper Miocene ostracods from northern Bulgaria. GeologicaBalcanica 5, 1–116.
Stevanovic, P., Nevesskaya, L.A., Marinescu, F., Sokac, A., Jámbor, A., 1989.Chronostratigraphie und Neostratotypen: Pliozän Pl1, Pontien. JAZU & SANU,Zagreb–Beograd. 952 pp.
Stoica, M., Lazar, I., Vasiliev, I., Krijgsman, W., 2007. Mollusc assemblages of the Pontianand Dacian deposits from the Topolog— Arges area (southern Carpathian foredeep— Romania). Geobios 40, 391–405.
Tarapoanca, M., Bertotti, G., Matenco, L., Dinu, C., Cloetingh, S.A.P.L., 2003. Architectureof the Foçsani Depression: a 13 km deep basin in the Carpathians bend zone(Romania). Tectonics 22 (6), 1074, http://dx.doi.org/10.1029/2002TC001486.
Ter Borgh, M., Vasiliev, I., Stoica, M., Knežević, S., Matenco, L., Krijgsman, W., Rundić, L.,Cloetingh, S., 2013. The isolation of the Pannonian basin (Central Paratethys): newconstraints from magnetostratigraphy and biostratigraphy. Global and PlanetaryChange 103, 99–118.
Vasiliev, I., Krijgsman, W., Langereis, C.G., Panaiotu, C.E., Matenco, L., Bertotti, G., 2004.Towards an astrochronological framework for the eastern Paratethys Mio-Pliocenesedimentary sequences of the Focsani basin (Romania). Earth and Planetary ScienceLetters 227, 231–247.
Vasiliev, I., Krijgsman, W., Stoica, M., Langereis, C.G., 2005. Mio-Pliocenemagnetostratigraphy in the southern Carpathian foredeep and Mediterranean–Paratethys correlations. Terra Nova 17, 376–384.
Vasiliev, I., Dekkers, M.J., Krijgsman, W., Franke, C., Langereis, C.G., Mullender, T.A.T.,2007. Early diagenetic greigite as a recorder of the palaeomagnetic signal in Miocene–Pliocene sedimentary rocks of the Carpathian foredeep (Romania). Geophysical JournalInternational 171, 613–629.
Vasiliev, I., Franke, C., Meeldijk, J.D., Dekkers, M.J., Langereis, C.G., Krijgsman, W., 2008.Putative greigite magnetofossils from the Pliocene epoch. Nature Geoscience 1,782–786 710.1038/ngeo1335.
Vasiliev, I., Matenco, L., Krijgsman, W., 2009. The syn- and post-collisional evolution ofthe Romanian Carpathian foredeep: new constraits from anisotropy of magneticsusceptibility and paleostress analyses. Tectonophysics 473, 457–465, http://dx.doi.org/10.1016/j.tecto.2009.04.002.
Vasiliev, I., de Leeuw, A., Filipescu, S., Krijgsman, W., Kuiper, K., Stoica, M., Briceag, A.,2010. The age of the Sarmatian–Pannonian transition in the Transylvanian Basin(Central Paratethys). Palaeogeography, Palaeoclimatology, Palaeoecology 297,54–69, http://dx.doi.org/10.1016/j.palaeo.2010.07.015.
Vasiliev, I., Iosifidi, A.G., Khramov, A.N., Krijgsman, W., Kuiper, K.F., Langereis, C.G.,Popov, V.V., Stoica, M., Tomsha, V.A., Yudin, S.V., 2011. Magnetostratigraphy andradio-isotopic dating of Upper Miocene–Lower Pliocene sedimentary successions ofthe Black Sea Basin (Taman Peninsula, Russia). Palaeogeography, Palaeoclimatology,Palaeoecology 310, 163–175, http://dx.doi.org/10.1016/j.palaeo.2011.06.022.