p - repositories
TRANSCRIPT

EFFECT OF RED IMPORTED FIRE ANTS ON REPRODUCTION,
HEALTH, AND SURVIVAL OF NORTHERN BOBWHITES
by
JAMES MICHAEL MUELLER, B.S., M.S.
A DISSERTATION
IN
WILDLIFE SCIENCE
Submitted to the Graduate Faculty of Texas Tech University in
Partial Fulfillment of the Requirements for
the Degree of
DOCTOR OF PHILOSOPHY
Approved
August, 1999

1999 by James M. Mueller

ACKNOWLEDGMENTS
The best research is usually a product of good plaiming, adequate funding, hard
work, and both professional and personal support. I was fortunate to be the recipient of
all these factors.
This study was planned by a team of three. Chapter II was the brainchild of Steve
Demarais, who initially was my major professor for a different study. When our project
tumed sour, he guided me to the tutelage and resources of Brad Dabbert. Brad, Steve,
and I clarified our objectives, evaluated different experimental designs, and developed the
methods that led to the successful assessment of the effects of red imported fire ants on
northern bob whites. I am indebted to Steve for his support during the transition between
projects, while a committee member, and continuing after his departure to Mississippi
State University. Likewise, Brad has been the type of major professor of which most
doctoral students only dream. He has always respected my knowledge, tactfully corrected
and encouraged my shortcomings, promptly returned manuscripts, and found the money
to support this research. Chapters III, IV, and V were the result of Brad's guidance.
Steve and Brad have been great examples to me, and I hope to pass on this legacy to
students of my own.
Other faculty also contributed to the success of my research. My committee
members—Harlan Thorvilson, Danny Pence, Scott McMurry, and Rob
Mitchell—^provided helpful advice and expertise. In addition, David Wester sat down
with me at the beginning of my study to review the design and responded to my plea for
help when I could not figure out the proper statistical analyses for certain data.
I received financial support from several entities. First, the Caesar Kleberg
Endowed Fellowship in Wildlife Conservation, administered through the Department of
Range, Wildlife, and Fisheries Management, provided my stipend. The State of Texas,
through the Texas Imported Fire Ant Research and Management Plan and line item
funding for research on fire ants and noxious bmsh and weed control, funded much of the
research. Additional funding was provided by the Department of Range, Wildlife, and
II

Fisheries Management in the form of a startup grant to Brad Dabbert. The J. F. Welder
Heirs Ranch provided me field housing and free-reign over their beautiful ranch.
American Cyanamid provided the fire ant insecticide used for experimental treatments.
Implementing this project in the heat and humidity of South Texas required tough
individuals. I thank my technicians, Mike Whitson and Hunter Haucke, for their
commitment to collection of quality data under all conditions. There is only one thing
worse than having a flat tire on the way back from a long, hot day of walking after birds;
that is having a flat on the way out and the way back and having only one spare tire.
Mike and Hunter overcame situations like this regularly. When I needed to increase my
sample size, Kyle Fiedler dropped everything to come down and help catch birds. During
that brief 10-day period, we had the coldest April weather, the most rattlesnakes, and the
spiciest food that I ever care to experience, but his help was invaluable. I also thank
Andy Forbes, a fellow graduate student who worked with me throughout 1998. Andy
monitored birds for me during March 1998, so I could retum home and be with my
family. If it were not for that, my baby Abigail would not have been conceived!
Finally, I thank my family for their support of my desire to eam a doctorate. My
parents have always encouraged me to pursue my dreams. My sister, Marylee, and her
husband, Eddie, have been especially supportive of my pursuit for a doctorate. But, most
of all, I thank my beautiful wife, Kay, for all she has sacrificed by my leaving a great job
and following my ambition. Kay kept an even-keel when the project on which I started
got canceled and the new project required me to be away for 16 of 20 months. During
this time, she raised an infant, went through a second pregnancy, started her own
business, worked 2 part-time jobs, and drove to San Antonio every 4-6 weeks so I could
visit with her and Zach when the demands of the project would allow it. Her support
allowed me to successfully complete my doctoral program.
Ill

TABLE OF CONTENTS
ACKNOWLEDGMENTS ii
TABLES yi
FIGURES yii
CHAPTER
I. INTRODUCTION 1
Study Area 2
Literature Cited 3
II. NORTHERN BOB WHITE CHICK MORTALITY CAUSED BY RED
IMPORTED FIRE ANTS 4
Abstract 4
Introduction 5
Methods 6
Results 10
Discussion 13
Management Implications 16
Literature Cited 18 III. EFFECT OF RED IMPORTED FIRE ANTS ON
IMMUNOCOMPETENCE OF NORTHERN BOB WHITE CHICKS 21
Abstract 21
Introduction 21
Methods 23
Results 25
Discussion 26
Literature Cited 28
IV. PLASMA CREATINE KINASE LEVELS ASSOCIATED WITH INCREASED PROBABILITY OF MORTALITY IN WILD NORTHERN BOBWHITE 30
Abstract 30
IV

Introduction 31
Methods 32
Results 34
Discussion 37
Literature Cited 39
V. RELATIONSHIP BETWEEN PLASMA TRIGLYCERIDE, BODY
MASS, AND REPRODUCTION OF NORTHERN BOBWHITES 41
Abstract 41
Introduction 41
Methods 43
Results 44
Discussion 45
Literature Cited 51

TABLES
3.1 Masses at 22 days, sheep red blood cell antibody titers, and wing-web indexes for northern bobwhite chicks (N) from nests (n) treated with a fire ant insecticide and controls, Refugio County, Texas, 1998 26
4.1 Tests of linear relationships between the logit of survival to 6 time periods and plasma CK sampled either after transport to a laboratory or immediately in the field 35
5.1 First clutch sizes, dates of nest initiation, late winter body masses (g), and plasma triglyceride levels (mg/dL) of northem bobwhites in 1997 (« = 17) and 1998 (n = 23), Reftigio County, Texas 45
5.2 R^ and significance of late winter body mass (BM) and plasma triglycerides (TRIG) as predictors of first clutch size and date of nest initiation for northem bobwhites in Refugio County, Texas, 1997-1998 47
5.3 Comparison of northem bobwhite late winter body masses (g) and plasma triglyceride levels (mg/dL) in 1997 and 1998 between a single pasture (Willow Lake Trap [WLT], « = 13 and 4, respectively) and all locations (« = 17 and 23, respectively), Refugio County, Texas 48
VI

FIGURES
2.1 Relationship between the proportion of northem bobwhite broods surviving to 3 weeks and number of red imported fire ants in a bait cup after 30 min on the day after hatch (« = 43) 12
4.1 Relationship between survival to 16 weeks of northem bobwhite hens (n = 61) and concentration of plasma CK for birds sampled in the field 36
5.1 Lack of linear relationship between plasma triglycerides and body mass of northem bobwhites in 1997 and 1998, Refugio County, Texas (1997: R^ = 0.01, F, ,5 = 0.2, P = 0.660; 1998: R^ = 0.12, F, 2, = 2.9, P = 0.105) 46
5.2 Lack of linear relationship between plasma triglycerides of northem bobwhites and sampling date in 1998, Reftigio County, Texas (F, 2, = 0.5, P = 0.507) 49
Vll

CHAPTER I
INTRODUCTION
Northem bobwhite (Colinus virginianus) populations have declined throughout their
range during the last 30 years (Sauer et al. 1997). In a study of northem bobwhite
abundance in Texas counties, Allen et al. (1995) found that population declines were
correlated with the year the county was infested by the red imported fire ant (Solenopsis
invicta). Whether the red imported fire ant caused these declines is unknown. However,
Allen et al. (1995) found that areas treated multiple times during 2 years with a fire ant
insecticide had more northem bobwhite in the second year than untreated areas. Thus, it
appears that the red imported fire ant limits northem bobwhite densities in some way.
The red imported fire ant affects northem bobwhite by killing pipping chicks
(Mitchell 1989) and altering activity of young chicks (Pedersen et al. 1996). In addition,
the red imported fire ant may dramatically alter the abundance of invertebrates, an
important component of northem bobwhite diets, when it colonizes an area (Porter and
Savignano 1990). However, there is no evidence northem bobwhite populations are
significantly impacted by these particular effects. The red imported fire ant could affect
northem bobwhite populations through other means, such as by altering growth or
reproduction of northem bobwhite through physiological effects of envenomation.
This study evaluated potential mechanisms by which red imported fire ants impact
northem bobwhite reproduction, survival, and health. The objectives were to determine
the effects of red imported fire ants on hatching success, growth, immunocompetence,
and survival of northem bobwhite chicks.
I conducted 2 other studies simultaneously using the northem bobwhites that were
radiomarked for the study assessing the effect of red imported fire ants. The first of these
studies assessed the relationship between levels of plasma creatine kinase in trapped or
netted northem bobwhite hens and their subsequent survival. High levels of plasma
creatine kinase indicate potential tissue damage from a condition known as capture

myopathy. The second study assessed the relationship between levels of plasma
triglyceride and the date of nest initiation and reproductive effort.
Study Area
All studies were conducted on the J. F. Welder Heirs Ranch in the Texas Coastal
Prairie (Gould 1975; Reftigio County; 28°28'N, 97°11 'W; elevation = 25 m). Grasslands
were interspersed with live oak {Quercus virginiana) motts, honey mesquite (Prosopis
glandulosa), and huisache {Acacia smallii). Climate was subtropical v^th hot summers,
mild winters, and average aimual rainfall of 98 cm (Guckian 1988). Soils were primarily
fine sandy loam or clay with slopes <1% (Guckian 1988). In 1991-92, red imported fire
ants occurred on the study area at a density of 205 mounds/ha and were predominately
polygyne (Allen 1993, Allen et al. 1995).

Literature Cited
Allen, C. R. 1993. Response of wildlife to red imported fire ant population reductions in the South Texas Coastal Prairie. Thesis, Texas Tech University, Lubbock, Texas.
, R. S. Lutz, and S. Demarais. 1995. Red imported fire ant impacts on northem bobwhite populations. Ecological Applications 5:632-638.
Gould, F. W. 1975. The grasses of Texas. Texas A&M University Press, College Station, Texas.
Guckian, W. J. 1988. Soil survey of Refugio County, Texas. U.S. Department of Agriculture, Soil Conservation Service, Washington, D.C.
Mitchell, M. R. 1989. The effects of imported fire ants on nesting bobwhite quail in Texas. Texas Parks & Wildlife Department, Federal Aid in Wildlife Restoration Project W-108-R, Final Report.
Pedersen, E. K., W. E. Grant, and M. T. Longnecker. 1996. Effects of red imported fire ants on newly-hatched northem bobwhite. Journal of Wildlife Management 60:164-169.
Porter, S. D., and D. A. Savignano. 1990. Invasion of polygyne fire ants decimates native ants and dismpts arthropod community. Ecology 71:2095-2106.
Sauer, J. R., J. E. Hines, G. Gough, I. Thomas, and B. G. Peterjohn. 1997. The North American breeding bird survey results and analysis. Version 96.4. Patuxent Wildlife Research Center, Laurel, Maryland.

CHAPTER II
NORTHERN BOBWHITE CHICK MORTALITY CAUSED
BY RED IMPORTED FIRE ANTS
Abstract
Northem bobwhite (Colinus virginianus) populations have declined throughout their
range during the last 30 years. Change in land management is thought to be a primary
cause of this decline, but the invasion of northem bobwhite habitats by the red imported
fire ant (Solenopsis invicta) also may be involved. I compared hatching success and
subsequent survival of wild northem bobwhite chicks on the Coastal Prairie of Texas in
1997 and 1998 between broods that hatched under natural conditions or following fire ant
suppression treatments. In 1997, the fire ant suppression treatment resulted in a 70%
reduction in the number of red imported fire ants on baits placed in treated nests on the
day after hatch. Using 2 fire ant suppression treatments in 1998 reduced the number of
red imported fire ants on baits by >99%. No year or treatment x year interaction effects
were detected for hatching success or survival, and no treatment effect was detected for
hatching success (P > 0.10). However, the proportion of chicks surviving to 21 days was
higher (P = 0.010) for treated nests (« = 18) than control nests (n = 25; proportions of
broods surviving: 53.5 ± 8.6% [x± SE] vs. 24.7 ± 6.6%; chick survival: 60.1 ± 7.6% vs.
22.0 ± 6.2%). The probability of chick survival decreased (P < 0.001) as an index of red
imported fire ant activity in the nest increased. These results indicate northem bobwhite
chicks can suffer high levels of mortality due to red imported fire ants, which could
explain declines in some northem bobwhite populations following infestations by red
imported fire ants. Current methods for controlling red imported fire ants are expensive
and may last <3 months. Thus, strategies for mitigating the effect of red imported fire
ants to northern bobwhite populations in this area should probably focus on reducing
other mortality factors or increasing productivity.

Introduction
Northem bobwhite populations have declined throughout their range during the last
30 years (Sauer et al. 1997). The major cause of these declines is thought to be loss of
habitat due to changes in land use (Brerman 1991). Recently, Allen et al. (1995)
presented associative and experimental results implicating the red imported fire ant as
another factor that may be causing declines in northem bobwhite populations in areas
where their ranges overlap. They found that the begirmings of population declines of
northem bobwhites in Texas counties were correlated with the years the counties became
infested with red imported fire ants; also, multiple treatments with a fire ant insecticide
during 2 years resulted in areas having more northem bobwhites in the second year than
untreated areas (Allen et al. 1995). Thus, it appears that red imported fire ants can limit
northem bobwhite densities in some way.
Researchers have expressed strongly divergent opinions on the role red imported fire
ants may play in northem bobwhite demographics (Brerman 1991, Allen et al. 1993,
Brennan 1993). Some of these differences may stem from confusion about the different
species of fire ants (Solenopsis spp.; Allen et al. 1993). Older studies (e.g., Stoddard
1931, Travis 1938, Johnson 1961, Lehmann 1984) that reported negligible effects of fire
ants on northem bobwhites were conducted before colonization by red imported fire ants
(Allen et al. 1993). However, recent studies that experimentally addressed the effect of
red imported fire ants on northem bobwhites reported adverse effects (Allen et al. 1995,
Giuliano et al. 1996, Pedersen et al. 1996). Red imported fire ants can directly affect
individual northem bobwhites by killing chicks (Mitchell 1989, Giuliano et al. 1996) and
altering their activity (Pedersen et al. 1996). In addition, red imported fire ant
colonization may decrease the abundance of other invertebrates (Porter and Savignano
1990), an important component of northem bobwhite diets (Hurst 1972, Lehmann
1984:170-172). However, previous studies have not provided evidence that these
particular factors affect northem bobwhite populations.
I hypothesized that the time of hatching was a period in the life history of the
northem bobwhite when it was particularly vulnerable to red imported fire ants. I based

my hypothesis on 2 facts. First, other species of fire ants commonly depredate or
scavenge northem bobwhite chicks in the nest (Stoddard 1931, Travis 1938, Johnson
1961, Lehmarm 1984). Second, in a laboratory study with red imported fire ants, survival
of northem bobwhite chicks decreased followdng exposure to ^50 ants for 60 sec or 200
ants for 15 sec (Giuliano et al. 1996). In a wild northem bobwhite nest, it probably takes
^2 hr from the time the first egg in a clutch begins to hatch until the entire brood has
hatched (Stoddard 1931:36-37), and occasionally much longer (Lehmann 1984:89).
During this period, the chicks may be restricted to the nest and vulnerable to stings by red
imported fire ants. I designed this study to measure the effect of red imported fire ants on
hatching success and subsequent survival of northem bobwhite chicks. I tested the null
hypothesis that decreasing the number of red imported fire ants around northem bobwhite
nests on the Texas Coastal Prairie would not affect northem bobwhite hatching success or
subsequent survival.
Methods
Study Design
Experimental units for the study were clutches of eggs in nests I found by tracking
radiomarked birds. I assigned treatments to each clutch completely at random by using a
coin toss as each nest was found. The treatment for each nest was either suppression of
fire ants or no suppression. The model for the experiment was
};., = H + a, + P; + (ap) . + e ,.
The response variable (Yy^) was either the proportion of eggs that hatched successfully or
the proportion of chicks that survived to 3 weeks of age. Two main effects, year and
treatment, and their interaction were assessed. The 2 years of the study were 1997 and
1998.

Field Techniques
I capmred female northem bobwhites from 1 Febmary through 23 April in 1997 and
1998 primarily with walk-in ftmnel traps (Smith et al. 1981) baited with milo. Each
captured hen was radiomarked with a 6.5-g necklace-style radio transmitter without a
body loop (American Wildlife Enterprises, Montacello, Florida).
From late April until mid-July, I located most radiomarked hens at least once every
other day to find nests. I approached each bird on foot until it flushed or moved away, I
observed it, or I had circled it and determined that it might be on a nest. I never
intentionally flushed hens from nests and rarely did so inadvertently. When each nest was
found, I randomly assigned it a treatment. For the fire ant suppression treatment, I used a
handheld broadcast spreader to apply Amdro® (American Cyanamid Company, Wayne,
New Jersey), a fire ant insecticide bait, at a rate of 1.7 kg/ha. I treated a 60- x 60-m area
centered on the nest. The "no suppression" treatment included the same activities as the
fire ant suppression treatment, but without application of the fire ant insecticide bait. Any
nest found <100 m from another nest during the same year was constrained to receive the
same treatment; this constraint affected <15% of nests. Application of Amdro® at rates
used in this experiment typically results in a 90% decrease in red imported fire ant
abundance (Harlan et al. 1981). Initial measurements of effectiveness of the fire ant
suppression treatment (described below) found many red imported fire ants in several
treated nests. To more closely attain my goal of no ants in treated nests, I broadcast
Amdro® directly over the nest for several seconds during the initial application, and I
applied a second treatment of Amdro® to a 20- x 20-m area centered on the nest 1 week
before the expected hatch date. These modifications affected treatment of 3 nests that
hatched in 1997 and all treated nests in 1998.
I measured the effect of red imported fire ant reduction on hatching success and
survival of chicks to 3 weeks of age; in addition, I obtained limited data on survival of
chicks to 1 day of age. Hatching success was estimated on the day of hatch by examining
eggs and eggshells in the nest. Successfully hatched eggs were easy to distinguish by the
even line of pipping that separated the eggshell cap from the remainder of the eggshell.

Unsuccessfully hatched eggs were either unpipped or partially pipped with dead chicks
inside. In addition, any dead chicks found in or around the nest were classified as an
unsuccessfully hatched egg. On the day after hatch, I often approached hens and used the
stereotypical behavior of hens with chicks to determine the number of broods that
suffered 100% mortality within 1 day. This technique could not provide information
beyond presence or absence of chicks; broods in which nearly all chicks died could not be
distinguished from those that had 100% survival. Three weeks after the hatch, I flushed
and counted the hen and chicks to measure chick survival. I repeated these counts vsdth 1
or 2 observers and up to twice per day until I was confident of brood size.
I used a 30-mL plastic cup baited with a slice of hot dog (approx 1 g; Porter and
Tschinkel 1987) to determine efficacy of the fire ant insecticide treatment and assess red
imported fire ant foraging activity in the control nests. On the day of hatch, I removed all
unhatched eggs and eggshells from the nest to eliminate residual yolk in the eggshells to
which the ants might be attracted. On the morning after the eggs hatched, I placed a bait
cup on its side in the nest. After 30 min, I capped the bait cup and placed it in a freezer
(-20 °C) to kill the ants. Bait-cup sampling was conducted within the thermal range of
maximal foraging by red imported fire ants (22-36°C at depth of 2 cm; Porter and
Tschinkel 1987). I counted the number of red imported fire ants in each cup and stored
all ants in 70%) ethanol for later verification of species identification.
I measured the density of active red imported fire ant mounds on the study area
during August 1997. I counted the number of red imported fire ant mounds in 18 20- x
20-m areas centered on northem bobwhite nests that had received the placebo treatment.
To assess presence of red imported fire ants in all potential mounds, I used a shovel to dig
down to visible soil moisture where ants would be if present.
Data Analysis
I compared the proportions of eggs that hatched successftiUy and chicks that
survived to 3 weeks of age between treatments and years using analysis of variance
(ANOVA). The data were analyzed as a 2 x 2 factorial with years and treatments as main
8

effects and yearxtreatment interaction. The year effect was analyzed first as a fixed effect
(Model I ANOVA; Zar 1996:244) to draw inference to treatment effects during the 2
years of this study, and then analyzed as a random effect (Model lU ANOVA or mixed
model; Zar 1996:247) to draw inference to the probability of treatment effects during any
year. Thus, the denominator for the F-test assessing differences in treatment means when
the year effect was fixed was the mean square error of the model, and the denominator for
this test when the year effect was random was the mean of the sum of squares for the
interaction term. I followed the recommendations of Cochran (1943) and determined that
each nest or brood should receive equal weighting. The analysis was repeated using the
ranks of the proportions due to heteroscedasticity and nonnormality (Zar 1996:269-270).
Calculations were made with SPSS for Windows (Nomsis 1993). I report means of the
proportions of eggs that hatched successfully and chicks that survived to 3 weeks of age;
these means were used for experimental comparisons. I also report mean chick survival
to 3 weeks of age for each treatment as calculated via an equation for cluster sampling
(Cochran 1963:65). I estimated the percentage of northem bobwhite mortality at 21 days
attributed to red imported fire ants (m J at hatching by using the equation
m m^ = ( 1 - - ^ ) X 100,
where m^ = the percent mortality at 21 days for the fire ant suppression group, and m^^^ =
the percent mortality at 21 days for the control group. Means are reported ± 1 standard
error.
I used logistic regression to assess the effect of red imported fire ant foraging
activity on chick survival (PROC LOGISTIC; SAS Institute 1989). My model was
(number of chicks from nest /alive at 3 weeks)/(number of chicks from nest /that
hatched) = (number of red imported fire ants in the bait cup in nest I). I tested for a linear
relation between the logit of chick survival and the number of red imported fire ants in

the bait cup in the nest on the day after the hatch by using a full-reduced model likelihood
ratio chi-square test (SAS Institute 1989).
Results
Precipitation differed tremendously during the 2 years of the study. Rainfall from
March through July 1997 was 165% of the long-term mean; during the same period in
1998, rainfall was only 23%) of the long-term mean (National Oceanic and Atmospheric
Administration 1997, 1998).
I radiomarked 77 northem bobwhite hens in 1997 and 71 in 1998. Of 79 nests found
in 1997, 37 were depredated by vertebrates, 35 hatched, 6 were abandoned, and 1 was
trampled by cattle. In 1998,1 found 87 nests: 44 were depredated by vertebrates, 27
hatched, 13 were abandoned, 1 was lost when the hen died, and 2 were still being
incubated when the study ended. Four of the 62 successful nests in 1997-98 were
disturbed by predators before the number of eggs that hatched could be counted, and 1
nest could not be assigned to a treatment due to its proximity to both treated and untreated
nests. These 5 nests were censored. Of the remaining 57 broods, 43 were successfully
counted at 3-weeks of age to measure survival.
Effects of Red Imported Fire Ants
Hatching success did not differ by treatment (F^^2 ~ 0-98, P = 0.327) or year
(F, 53 = 1.89, P = 0.175), and no treatmentxyear interaction was detected (F, 53 = 0.53,
P = 0.472). In the 35 control nests that were successful, 457 of 483 eggs (94.6%)) hatched
successfully. I found 10 of these unsuccessful eggs pipped or hatched, and observed
dozens of red imported fire ants feeding on each dead chick. In the 22 nests treated with
insecticide, 265 of 289 eggs (91.7%)) hatched successfully. I found 4 of these
unsuccessful eggs pipped and 1 cracked, but observed no red imported fire ants on these
chicks.
Chick survival differed greatly between treatments. The proportion of chicks
surviving to 21 days was higher for treated nests (n= 18) than control nests (n = 25;
10

proportions of broods surviving: 53.5 ± 8.6%) vs. 24.7 ± 6.6%), year analyzed as a fixed
effect, F, 39 = 7.35, P = 0.010; chick survival: 60.1 ± 7.6% vs. 22.0 ± 6.2%). Death of all
chicks occurred in 52%) of broods from control nests and 22%) of broods from treated
nests. The percentage of mortality at 21 days attributed to red imported fire ants at
hatching (mj was 48.8%). For this calculation, I did not include the 2%o of chicks from
control nests that were found dead and covered with red imported fire ants because these
chicks may have died of other causes and inclusion of these mortalities had little effect on
the resultant value. No year effect (F, 39 = 0.26, P = 0.612) or treatmentxyear interaction
(F, 39 = 0.18, P = 0.670) were detected. Results using ranked data were similar. When
the year effect was analyzed as a random effect, results were less conclusive (F, , =
39.91, P = 0.100). The power of this test was low because the study was conducted for
only 2 years, which resulted in only 1 degree of freedom for the denominator in the F-test.
I monitored 12 broods on the day after hatch that were later found to have no chicks alive
at 21 days. Six of 10 control broods and both treated broods in this category had lost all
of their chicks within the first day.
The number of red imported fire ants in bait cups in control nests averaged 89 ± 59
more ants in 1997 (x= 358 ± 43 ants in 1997 and 269 ± 39 ants in 1998). The logit of
chick survival decreased as the number of red imported fire ants in a bait cup increased
(X^i = 95.0, P < 0.001; Figure 2.1). The probability of a chick surviving decreased by
0.5%) with each additional red imported fire ant. Mean red imported fire ant mound
density in August 1997 was 290 ± 60 mounds/ha (range = 50-1,000).
In 1998, 1 nest appeared to have been abandoned due to red imported fire ants. The
nest had been constmcted next to a clump of grass that contained an abandoned red
imported fire ant mound. The ants reoccupied the mound after a 3.5-cm rainfall preceded
by 2 months of drought, and they covered the eggs in the nearby northem bobwhite nest
with soil litter.
11

•OX A A
0.0
X Fire Ant Suppression
A Control
0 X
100 200 M v x / ^ i A /y\
300 400 500
NUMBER OF ANTS
600 700 800
Figure 2.1 Relationship between the proportion of northem bobwhite broods surviving to 3 weeks and number of red imported fire ants in a bait cup after 30 min on the day after hatch (« = 43). Broods from nests treated to suppress red imported fire ants are displayed separately from broods from control nests. The logit of chick survival decreased with increasing numbers of red imported fire ants (P < 0.001). The equation depicted by the line is y = e<o55-o.oo54x)/(i + g(o.55-o.oo54x)) Arrows and associated
numbers denote the number of overlapping values at those points.
12

Discussion
Time of Mortality
The benefit of fire ant suppression in my study probably was experienced by
northem bobwhite chicks only while hatching and for a few hours afterward. Chicks
were led away from the nest by 1 or both parents, were always outside the treated area 24
hr following hatching, and were never relocated in a treated area. Thus, chicks from
control and treated nests used the same environment after leaving the nest. Few chicks in
my study were killed in the nest. After 3 weeks, however, survival of broods from
control nests was less than one-half that of broods from treated nests. Much of the
mortality may have occurred soon after the chicks hatched; this possibility is supported by
my data showing that 60%) of the control broods that suffered 100%) mortality by 21 days
had already experienced this mortality by the day after hatch. The following field
observation also supports this possibility and illustrates that deaths may have resulted
from indirect effects of red imported fire ant stings.
At a nest site with a red imported fire ant mound density of 1,000 mounds/ha, I
found 3 dead chicks (2 still in their eggs) in the nest covered by red imported fire ants. I
observed the hen flush 15m from the nest and captured 1 chick that attempted to hide at
my feet. This chick had 1 eye swollen shut from red imported fire ant stings, yellow
marks on its feet that may have been sting sites, and 1 red imported fire ant still stinging
its foot. Pedersen et al. (1996) reported that red imported fire ants often attacked and
stung newly hatched and 1-day-old northem bobwhite chicks on the eyelids, legs, and
toes; stings to the eyelids caused swelling that obscured vision, and stings to the legs and
feet sometimes caused swelling that prevented normal movement. Undoubtedly, the
probability of a chick surviving decreases if its vision and movement are hindered by red
imported fire ant stings.
Chick Survival
Past studies that estimated northem bobwhite chick survival from repeated counts of
chicks in broods associated with radiomarked adults found survival to 2 weeks of 13%) (n
13

= 5 broods; Cantu and Everett 1982) and 38% (n = 22 broods; DeVos and Mueller 1993)
and survival to 3 weeks of 38%) (n = 59 broods; DeMaso et al. 1997). The study area of
DeVos and Mueller (1993) was within the range of the red imported fire ant. When I
compared my results with these studies, survival of chicks that were protected from red
imported fire ants (60%) was higher, and survival of chicks from control nests (22%) was
within the range of previously reported estimates.
When I assumed survival rates of chicks in my 2 treatment groups were identical
after 21 days, additive mortality of northem bobwhite due to stings from red imported fire
ants at the time of hatching was 38.1 ± 7.7%. My estimate of additive mortality was the
difference in chick survival for the 2 treatments. This difference would have been greater
if indirect mortality from red imported fire ant stings did not occur within 21 days.
Giuliano et al. (1996) observed that northem bobwhite chicks that survived the most
severe exposures to red imported fire ants weighed less than controls after 9 days (the
duration of their experiment), and they speculated that depressed growth could reduce
subsequent survival because chick size may be related to survival. However, the
difference in survival rates could have decreased subsequently due to density-dependent
factors. My data could not resolve if the difference in mortality between the 2 treatment
groups changed after 21 days.
Northem bobwhite frequently attempt >1 nest/season (Rosene 1969:73, Roseberry
and Klimstra 1984:83) and may occasionally raise >1 brood in a season (reviewed by
Guthery and Kuvlesky 1998). These attributes allow the possibility that northem
bobwhite could exhibit compensatory reproduction in response to losing broods rapidly
due to red imported fire ant stings at hatching. However, when an entire brood is killed
by red imported fire ants, it occurs after an investment of >37 days by the hen (average
clutch size of 14 eggs [this study], lay rate of 1 egg/day, 23-day incubation period, and
time to build the nest and lay the first egg that Rosene [1969] estimated at 6-12 days).
Burger et al. (1995) found the interval between successfiil brood rearing and renesting
was about 14 days longer than the interval between nest failure and renesting. Thus, if
red imported fire ants kill only some chicks in a brood, even more time will pass before
14

the hen will attempt to renest. In either case, mortality by red imported fire ants at
hatching is more severe than an equivalent proportion of mortality from nest destmction
because of the greater loss of time for reproduction.
I observed no renesting after the loss of an entire brood of chicks, but I only
monitored hens through July. Guthery et al. (1988) found that nearly half the northem
bobwhite hens in my region of Texas were in laying condition in August; thus, hens had
ample time to produce additional clutches. Hens also may have produced clutches that
were incubated by males (Burger et al. 1995, DeMaso et al. 1997). Further investigation
is needed to determine how time constraints resulting from a limited reproductive season
and a large investment of time in chicks that are killed by red imported fire ants affect
potential compensatory reproduction.
The scope of inference from this study is limited to 2 years at 1 site. Here I consider
the reasonableness of extrapolating these results to other times and areas. The statistical
test of treatment effects in the context of all possible years (i.e., years as a random
variable) was not conclusive (P - 0.100). Because precipitation affects northem
bobwhite populations (Rice et al. 1993) and activity of red imported fire ants (Rhoades
and Davis 1967), it was probably the most important variable in the "year" effect in this
study. Fortuitously, precipitation during this study represented periods of flooding one
year and drought the next. Treatment effects were consistent between these conditions.
Thus, I expect effects of red imported fire ants on hatching northem bobwhites to vary
little under the extremes of precipitation that occur on the Texas Coastal Prairie.
However, many other variables could cause differences in the treatment effect, and my
data were not adequate for supporting the effects I observed over the universality of time.
Expanding the inference from this study to other regions is even more problematic.
Because characteristics of red imported fire ants and northem bobwhite may vary among
locations, replication of this study in other regions is needed to understand how well the
fiinctional relation described in Figure 2.1 applies to other regions.
15

Effectiveness of Treatment
The number of red imported fire ants in bait cups exceeded 200 ants at 3 of the first
9 treated nests in 1997 (Figure 2.1); thus, I added a second treatment to attain better ant
suppression. Changing my methodology probably did not confound results. First, chick
survival for these 3 nests did not differ (P = 0.6) from that of other nests treated in 1997.
Second, chick survival across years was not different (P = 0.4) between nests treated
either once or twice. Thus, while the additional treatment was valuable for suppressing
red imported fire ants at some nests, it did not cause a detectable change in chick survival.
I recommend using 2 applications of fire ant insecticide to suppress red imported fire ants
in research applications to ensure distinct differences in numbers of red imported fire ants
between treatments.
Interpreting Evidence at Nests
I observed the outcome of eggs hatching in 3 nests abandoned by the hen (1 due to a
western diamondback rattlesnake [Crotalus atrox] in the nest, 1 due to the initiation of
brooding of the first chick that hatched days before the others, and 1 due to disturbance).
The death of chicks in these nests would have been attributed to red imported fire ants if
the circumstances had not been understood. These nests were tallied as abandoned nests
in my study and not included in any subsequent analysis. Most of the chicks in 2 of these
broods were found dead near their nests and appeared to have been killed by red imported
fire ants. The third brood would probably have suffered the same fate, but that nest had
been treated with fire ant insecticide and the chicks were seen alive around the nest
without a parent for 2 days. These observations confirm that proving red imported fire
ants are responsible for killing northem bobwhite chicks is difficult when researchers
simply observe hatches (Johnson 1961).
Management Implications
Red imported fire ants have probably degraded suitability of habitat for northem
bobwhites throughout the Texas Coastal Prairie due to the mortality they inflict on
16

northem bobwhite chicks. Current methods of controlling red imported fire ants on large
areas using insecticides are expensive ($20.00 ha"' treatment''; Drees et al. 1996) and may
last <3 months (Apperson et al. 1984). Allen (1993) found that insecticidal control of red
imported fire ants to increase northem bobwhite and white-tailed deer (Odocoileus
virginianus) densities for lease hunting was not warranted economically. However,
insecticidal treatments may play a valuable role in management of threatened and
endangered species (e.g., Lockley 1995). One endangered species that occurs near the
study area that might benefit from insecticidal control of red imported fire ants is the
endangered Attwater's prairie chicken (Tympanuchus cupido attwateri).
Because of the high cost of reducing red imported fire ants directly through
insecticide, I think the best current management for northem bobwhite populations is
limited to mitigating red imported fire ant mortality. Thus, reduction of other mortality
factors and increasing productivity via traditional means has become more critical with
the invasion of northem bobwhite habitat by red imported fire ants. The most cost-
effective solution presently is to manage land so that it provides usable space (Guthery
1997) for northem bobwhites in the form of an interspersion of nesting, brood-rearing,
feeding, loafing, and roosting cover demonstrated for decades to benefit northem
bobwhites (Stoddard 1931, Rosene 1969).
17

Literature Cited
Allen, C. R. 1993. Response of wildlife to red imported fire ant population reductions in the South Texas Coastal Prairie. Thesis, Texas Tech University, Lubbock, Texas.
, R. S. Lutz, and S. Demarais. 1993. What about fire ants and northem bobwhites. Wildlife Society Bulletin 21: 349-351.
_ , , and . 1995. Red imported fire ant impacts on northem bobwhite populations. Ecological Applications 5:632-638.
Apperson, C. S., R. B. Leidy, and E, E. Powell. 1984, Effects of Amdro on the red imported fire ant (Hymenoptera: Formicidae) and some nontarget ant species and persistence of Amdro on a pasture habitat in North Carolina. Joumal of Economic Entomology 77:1012-1018.
Brerman, L. A. 1991. How can we reverse the northem bobwhite population decline? Wildlife Society Bulletin 19:544-555.
. 1993. Fire ants and northem bobwhites: a real problem or a red herring? Wildlife Society Bulletin 21:351-355.
Burger, L. W., Jr., M. R. Ryan, T. V. Dailey, and E. W. Kurzejeski. 1995. Reproductive strategies, success, and mating systems of northem bobwhite in Missouri. Joumal of Wildlife Management 59:417-426.
Cantu, R., and D. D. Everett. 1982. Reproductive success and brood survival of bobwhite quail as affected by grazing practices. National Bobwhite Quail Symposium 2:79-83.
Cochran, W. G. 1943. Analysis of variance for percentages based on unequal numbers. Joumal of the American Statistical Association 38:287-301.
. 1963. Sampling techniques. Second edition. John Wiley & Sons, New York, New York.
DeMaso, S. J., A. D. Peoples, S. A. Cox, and E. S. Parry. 1997. Survival of northem bobwhite chicks in western Oklahoma. Joumal of Wildlife Management 61:846-853.
DeVos, T., and B. S. Mueller. 1993. Reproductive ecology of northem bobwhite in north Florida. National Bobwhite Quail Symposium 3:83-90.
Drees, B. M., C. L. Barr, S. B. Vinson, R. E. Gold, M. E. Merchant, and D. Kostroun. 1996. Managing red imported fire ants in urban areas. Texas Agricultural Extension Service Publication B-6043.
Giuliano, W. M., C. R. Allen, R. S. Lutz, and S. Demarais. 1996. Effects of red imported fire ants on northem bobwhite chicks. Joumal of Wildlife Management 60:309-313.
18

Guthery, F. S. 1997. A philosophy of habitat management for northem bobwhites. Joumal of Wildlife Management 61:291-301.
, N. E. Koerth, and D. S. Smith. 1988. Reproduction of northem bobwhites in semiarid environments. Joumal of Wildlife Management 52:144-149.
, and W. P. Kuvlesky, Jr. 1998. The effect of multiple-brooding on age ratios of quail. Joumal of Wildlife Management 62:540-549.
Harlan, D. P., W. A. Banks, H. L. Collins, and C. E. Stringer. 1981. Large area tests of AC-217,300 bait for control of imported fire ants in Alabama, Louisiana, and Texas. Southwestern Entomologist 6:150-157.
Hurst, G. A. 1972. Insects and bobwhite quail brood habitat management. National Bobwhite Quail Symposium 1:65-82.
Johnson, A. S. 1961. Antagonistic relationships between ants and wildlife with special reference to imported fire ants and bobwhite quail in the Southeast. Proceedings of the Aimual Conference of the Southeastem Association of Game and Fish Commissioners 15:88-107.
Lehmann, V. W. 1984. Bobwhites in the Rio Grande Plain of Texas. Texas A&M University Press, College Station, Texas.
Lockley, T. C. 1995. Effect of imported fire ant predation on a population of the least tern—an endangered species. Southwestem Entomologist 20:517-519.
Mitchell, M. R. 1989. The effects of imported fire ants on nesting bobwhite quail in Texas. Texas Parks & Wildlife Department, Federal Aid in Wildlife Restoration Project W-108-R, Final Report.
National Oceanic and Atmospheric Administration. 1997. Climatological data annual summary: Texas. National Climatic Data Center, Asheville, North Carolina.
. 1998. Climatological data annual surrunary: Texas. National Climatic Data Center, Asheville, North Carolina.
Nomsis, M. J. 1993. SPSS® for Windows: base system user's guide. Release 6.0. SPSS, Chicago, Illinois.
Pedersen, E. K., W. E. Grant, and M. T. Longnecker. 1996. Effects of red imported fire ants on newly-hatched northem bobwhite. Joumal of Wildlife Management 60:164-169.
Porter, S. D., and D. A. Savignano. 1990. Invasion of polygyne fire ants decimates native ants and dismpts arthropod community. Ecology 71:2095-2106.
, and W. R. Tschinkel. 1987. Foraging in Solenopsis invicta (Hymenoptera: Formicidae): effects of weather and season. Envirormiental Entomology 16:802-808.
19

Rhoades, W. C , and D. R. Davis. 1967. Effects of meteorological factors on the biology and control of the imported fire ant. Joumal of Economic Entomology 60:554-558.
Rice, S. M., F. S. Guthery, G. S. Spears, S. J. DeMaso, and B. H. Koerth. 1993. A precipitation-habitat model for northem bobwhites on semiarid rangeland. Joumal of Wildlife Management 57:92-102.
Roseberry, J. L., and W. D. Klimstra. 1984. Population ecology of the bobwhite. Southern Illinois University Press, Carbondale, Illinois.
Rosene, W. 1969. The bobwhite quail: its life and management. Rutgers University Press, New Bmnswick, New Jersey.
SAS Institute. 1989. SAS/STAT user's guide. Version 6. Fourth edition. Volume 2. SAS Institute, Gary, North Carolina.
Sauer, J. R., J. E. Hines, G. Gough, I. Thomas, and B. G. Peterjohn. 1997. The North American breeding bird survey results and analysis. Version 96.4. Patuxent Wildlife Research Center, Laurel, Maryland.
Smith, H. D., F. A. Stormer, and R. D. Godfrey, Jr. 1981. A collapsible quail trap. U.S. Forest Service Research Note RM-400.
Stoddard, H. L. 1931. The bobwhite quail: its habits, preservation and increase. Charles Scribner's Sons, New York, New York.
Travis, B. V. 1938. The fire ant (Solenopsis spp.) as a pest of quail. Joumal of Economic Entomology 31:649-652.
Zar, J. H. 1996. Biostatistical analysis. Third edition. Prentice Hall, Upper Saddle River, New Jersey.
20

CHAPTER III
EFFECT OF RED IMPORTED FIRE ANTS ON IMMUNOCOMPETENCE
OF NORTHERN BOBWHITE CHICKS
Abstract
Northem bobwhites can suffer high mortality due to stings from red imported fire
ants while hatching. I conducted a study on wild northem bobwhites to determine if
chicks that survived natural exposure to red imported fire ants at hatching suffered
reduced growth or development of immunocompetence. I compared 22-day chick masses
and measures of humoral and cell-mediated immunity between chicks exposed to natural
levels of red imported fire ants at hatching and chicks from nests where ants were
exterminated. No variables differed between treatments. These results suggest that
growth and immunocompetence of northem bobwhite chicks are probably not affected
greatly by red imported fire ant stings at hatching. Sublethal effects such as reduced
growth can be demonstrated under laboratory conditions, but may occur only rarely under
natural conditions due to the rapid recmitment of red imported fire ants to food sources
and subsequent rapid accumulation of stings by chicks.
Introduction
The red imported fire ant (Solenopsis invicta) is a pest species accidentally
introduced into the United States from South America between 1933 and 1941 (Buren
1972, Rhoades 1977). Its range in the United States rapidly expanded and now
encompasses more than 1 million km^ (Williams 1994). The primary diet of these ants is
insects and other small invertebrates (reviewed in Vinson and Greenberg 1986); however,
they frequently attack and kill vertebrates (reviewed in Allen et al. 1994), including
nestiing wood ducks (Aix sponsa) (Ridlehuber 1982), cliff swallows (Hirundo
pyrrhonota) (Sikes and Amold 1986), least tems (Sterna antillarum) (Lockley 1995), and
northem bobwhites (Chapter II).
21

Before the invasion and spread of red imported fire ants, researchers found that
native fire ants (genus Solenopsis), such as S. geminata and S. richteri, killed 4-9%) of
hatching northem bobwhite (Stoddard 1931, Travis 1938, Johnson 1961). However, the
impact of red imported fire ants to northem bobwhite is more severe than that of native
fire ants. In Chapter II, I estimated that 38.1%) of all northem bobwhites that hatched in
1997-98 died as a result of red imported fire ant stings inflicted at the time of hatching.
This estimate of the proportion of northem bobwhites killed may have been negatively
biased if an assumption, that all mortalities caused by ant stings at hatching occurred
within 21 days, was violated. This assumption was required because chick survival was
measured 21 days after each brood hatched.
Could red imported fire ant stings to chicks at hatching cause mortality >21 days
after hatching? Giuliano et al. (1996) found northem bobwhite chicks that survived the
most severe exposures to red imported fire ants in a laboratory study weighed 12% less
than controls after 9 days (the duration of the experiment). However, 77%) of the chicks
at this level of exposure died (Giuliano et al. 1996). Thus, under laboratory conditions,
most chicks died when exposed to a level of red imported fire ant stings that reduced
growth. Wild northem bobwhites, however, face greater difficulty in acquiring optimal
nutrition, and the effect of red imported fire ant stings may be more severe. For example,
chicks in lab studies might be capable of obtaining optimal nutrition after being partially
blinded by red imported fire ant stings because feed is easily found and provided ad
libitum. Thus, levels of red imported fire ant exposure equivalent to the study by
Giuliano et al. (1996) would probably cause greater mortality and reductions in growth in
wild northem bobwhites; in addition, reductions in growth might occur over a broader
range of red imported fire ant exposure.
Giuliano et al. (1996) did not describe the mechanism by which red imported fire ant
stings reduced growth of northem bobwhite chicks. However, Pedersen et al. (1996)
reported that red imported fire ants often stung the eyelids, legs, and toes of northem
bobwhite chicks. Stings to the eyelid usually caused the eye to swell closed, and stings to
the leg or foot sometimes caused swelling that prevented normal movement. If stings
22

result in reduced intake of protein, chick growth, development of the bursa of Fabricius
and spleen, and cell-mediated immunity might be suppressed (Lochmiller et al. 1993).
Thus, chicks that survive stings from red imported fire ants while hatching may be unable
to acquire an adequate ration of high-protein foods, leading to slower growth, and
resulting in greater susceptibility to pathogens for weeks afterward.
To test whether red imported fire ant stings at hatching could potentially influence
wild northem bobwhite survival >21 days after hatching, I compared 22-day body mass,
cell-mediated immunity, and humoral immunity between northem bobwhite chicks that
hatched from nests with natural levels of red imported fire ants and those from nests
where ants were exterminated. These chicks left the experimentally treated areas around
each nest soon after hatching and thereafter occupied the same natural environment with
uimianipulated red imported fire ant populations.
Methods
Field Techniques
Chapter II provided a detailed description of the experimental treatments. Briefly, I
found nests by monitoring radiomarked northem bobwhite hens from late April until mid
July 1998. When a nest was found, it was randomly assigned to either receive a fire ant
suppression treatment or be a control. For the fire ant suppression treatment, a fire ant
insecticide was applied (1.7 kilograms/ha of Amdro®, American Cyanamid Company,
Wayne, New Jersey) using a handheld broadcast spreader. Treatments were applied to a
60- x 60-m area centered on the nest 3 weeks before the expected hatch date followed by
a retreatment 2 weeks later to the 20- x 20-m core area aroimd the nest. Five variables
were compared between chicks from treated and control nests: hatching success, 21-day
survival, 22-day body mass, humoral immunity, and cell-mediated immunity. Methods
and results for comparisons of hatching success and 21-day survival were reported in
Chapter II. The following describes my methods for capturing chicks and measuring 22-
day body mass, humoral immunity, and cell-mediated immunity.
23

Chick Capture
Broods were captured 22 days after hatching by netting at night. Coveys were found
by tracking the signal from the hen's radio transmitter and illuminating the brood with a
handheld 500,000 candlepower spotlight. Broods were captured using a handheld net
(Mighty Net, Animal Management, Inc., Heafford Junction, Wisconsin; netting was
replaced with 1/4 inch mesh). Chicks were weighed (± 0.1 g), banded with aluminum
legbands, and radiomarked with transmitters weighing 0.7 g (American Wildlife
Enterprises, Montacello, Florida). To assess chick immunocompetence, chicks were
recaptured after 5 and 6 days by locating them using telemetry and capturing them by
hand or net.
Humoral Immunity
Humoral immunity was assessed by measuring antibody production in response to
an injection of sheep red blood cells. Chicks were inoculated intramuscularly with
0.5 mL of 5% sheep red blood cells (Colorado Semm Company, Denver, Colorado) in
phosphate buffered saline (Lochmiller et al. 1993) when initially captured and
radiomarked. Antibody production was assessed after 6 days by pricking the ulnar vein
and collecting blood into capillary tubes. Blood was kept shaded at ambient temperature
for up to 4 hr and then centrifuged for 10 min. Serum was pipetted from the capillary
tubes, placed into cryovials, stored at -20 °C for <9 weeks, shipped on dry ice, and then
stored at -84 °C for 6 months until assayed.
Antibody responses to the sheep red blood cell inoculation was measured using a
microhemagglutination assay (Wegmann and Smithies 1966). As described by
Lochmiller et al. (1993), serial 2-fold dilutions of heat-inactivated semm (56°C for 30
min) in phosphate-buffered saline were mixed with an equal volume of \% sheep red
blood cell solution in phosphate-buffered saline and incubated at 40°C for 1 hr. Titers
were expressed as the logj of the reciprocal of the highest dilution of semm showing a
positive hemagglutination.
24

Cell-mediated Immunitv
Cell-mediated immunity was assessed by measuring wing web inflammation caused
by an intradermal injection of the T-cell mitogen phytohemagglutinin (PHA-P; Sigma, St.
Louis, Missouri). The thickness of each wing web (± 0.01 mm) of 27-day-old chicks was
measured using a digital thickness gauge (Digimatic Thickness Gage, model 547-520,
Mitutoyo Corporation, Takatsu-ku, Kawasaki, Kanagawa, Japan). The right wing web
was injected with 0.5 mg of PHA-P in 0.1 mL of phosphate buffered saline to stimulate
inflammation and the left wing web with 0.1 mL of phosphate buffered saline to serve as
a control (Lochmiller et al. 1993). After 24 hr, the chicks were recaptured and wdng-web
thickness was remeasured. Wing-web swelling was calculated as the difference in wing-
web thickness before and 1 day after injection. The cell-mediated immune response,
termed the wing-web index, was the difference in swelling between the right and left
wing webs (Lochmiller et al. 1993).
Statistical Analysis
Brood means for 22-day chick masses, antibody titers, and wing-web indexes were
compared between treatments using t-tests. Brood means rather than means of all chicks
were used because experimental treatments were applied to nests, not eggs or chicks.
Thus, variables for chicks within a brood were probably correlated. The assumptions of
normality and homoscedasticity were assessed where appropriate using the Shapiro and
Wilk (1965) and Levene (1960) tests, respectively. Analyses were conducted using SPSS
for Windows, release 6.0 (Nomsis 1993) and GPOWER, version 2.0f (Erdfelder et al.
1996). Statistical significance was determined with a = 0.05.
Results
I captured and radiomarked 37 chicks from 11 nests. Of these, 9 lost their radio
transmitters, 6 died, 5 could not be found and were presumed to either have a faulty radio
transmitter or have been depredated, and 17 were successftiUy monitored until they were
28 days old.
25

No differences were detected between chicks that hatched from treated and control
nests for 22-day mass, humoral immunity, or cell-mediated immunity (Table 3.1). Chick
masses ranged from 18.7 to 54.8 g. The heaviest chick failed to express a primary
antibody response to sheep red blood cell immunization and had the third lowest wing-
web index. Excluding this chick, hemagglutinating antibody titers (log2) ranged from 2 to
16 and wing-web indexes ranged from 0.21 to 1.24 mm. Mass was not linearly related to
individual chick antibody titers (F, 9 = 0.13, F = 0.726) or wing-web indexes (F, ,4 =
0.08, F = 0.788).
Table 3.1. Masses at 22 days, sheep red blood cell antibody titers, and wing-web indexes for northem bobwhite chicks (N) from nests (n) treated with a fire ant insecticide and controls, Refugio County, Texas, 1998.
Variable
A
Treated
n X SE N
Control
n X SE / P Power
Mass (g)
Antibody titer (logj)
Wing-web index (mm)
22 6 38.1 2.6
6 4 8.5 2.9
10 5 0.74 0.15
15 5 35.4 3.1 0.69 0.51 0.09
5 3 13.7 2.3 1.32 0.25 0.18
6 3 0.91 0.13 0.77 0.47 0.10
Discussion
Body mass at 22 days and immunocompetence of northem bobwhite chicks were not
affected by exposure to natural levels of red imported fire ants at hatching. Few
comparable data were available to determine the reasonableness of the observed values,
so I calculated an optimal 22-day body mass using an equation derived by Blem and Zara
(1980) for northem bobwhites fed ad libitum on a 30%) protein diet. Blem and Zara
(1980) stated that their asymptotic weight of 205 g was undoubtedly greater than reported
previously for northem bobwhites due to the genetic strain used. For a more realistic
asymptotic weight, I used the average mass of 175 northem bobwhite captured on my
study area in Febmary 1997 and 1998 (x = 169 g). This mean should be a reasonable
estimate of asymptotic weight because body mass of northem bobwhites peaks in
26

Febmary when averaged by gender (Roseberry and Klimstra 1971). Using the other
growth parameters of Blem and Zara (1980) and the site-specific estimate of asymptotic
size, the predicted 22-day body mass of northem bobwhite was 37.9 g. Chicks from
treated nests and control nests attained 100.5%) and 93.4%) of this predicted 22-day body
mass, respectively.
The probability of falsely finding no difference between treatments in body mass and
immunocompetence was high due to the small sample size. For example, the power for
detecting the observed difference in body masses between treatments was 0.09 (Table
3.1). The effect size for this test was 0.41, which is generally considered a small- to
medium-sized effect (Cohen 1977). For a more biologically relevant assessment of
power, I calculated the power of detecting a difference between body mass treatment
means of 10 g. Nestler et al. (1942) reported that 21-day body mass of northem
bobwhites fed either a 22%) or 30%) protein diet differed by about 10 g. The power of
detecting a 10-g difference with the sample sizes (n = 5 and 6), standard deviation (6.7),
and alpha (0.05) of this study was 0.60. Thus, this study had a 60% probability of
detecting an effect of the same magnitude as a 22% versus 30%) protein diet. These
results should be used in context with previous studies to focus future research at the
specific mechanisms by which red imported fire ants impact northem bobwhites. While
my sample size was inadequate for detecting small differences between treatments, my
observed treatment means and previous studies (Giuliano et al. 1996) suggest that growth
and immunocompetence of northem bobwhite chicks are probably not affected greatly by
red imported fire ant stings at hatching. However, northem bobwhite chick survival is
greatly reduced by red imported fire ant stings at hatching (Chapter II).
Under laboratory conditions, the effect of red imported fire ant stings on growth of
northem bobwhite chicks was small and only detectable under conditions that resulted in
77% mortality (Giuliano et al. 1996). Only a narrow range of red imported fire ant
exposure appears to reduce growth without causing death. Thus, I speculate that, under
natural conditions, few hatching northem bobwhite chicks are exposed to red imported
fire ants at a level that is sub-lethal but biologically detrimental.
27

Literature Cited
Allen, C. R., S. Demarais, and R. S. Lutz. 1994. Red imported fire ant impact on wildlife: an overview. Texas Joumal of Science 46:51-59.
Blem, C. R., and J. Zara. 1980. The energetics of young bobwhite (Colinus virginianus). Comparative Biochemistry and Physiology 67A:611-615.
Buren, W. F. 1972. Revisionary studies on the taxonomy of the imported fire ants. Joumal of the Georgia Entomological Society 7:1-26.
Cohen, J. 1977. Statistical power analysis for the behavioral sciences. Revised edition. Academic Press, New York, New York.
Erdfelder, E., Faul, F., and A. Buchner. 1996. GPOWER: A general power analysis program. Behavior Research Methods, Instmments, & Computers 28:1-11.
GiuHano, W. M., C. R. Allen, R. S. Lutz, and S. Demarais. 1996. Effects of red imported fire ants on northem bobwhite chicks. Joumal of Wildlife Management 60:309-313.
Johnson, A. S. 1961. Antagonistic relationships between ants and wildlife with special reference to imported fire ants and bobwhite quail in the Southeast. Proceedings of the Aimual Conference of the Southeastem Association of Game and Fish Commissioners 15:88-107.
Levene, H. 1960. Robust tests for equality of variances. Pages 278-292 in I. Olkin, editor. Contributions to probability and statistics. Stanford University Press, Palo Alto, California.
Lochmiller, R. L., M. R. Vestey, and J. C. Boren. 1993. Relationship between protein nutritional status and immunocompetence in northem bobwhite chicks. Auk 110:503-510.
Lockley, T. C. 1995. Effect of imported fire ant predation on a population of the least tern—an endangered species. Southwestem Entomologist 20:517-519.
Nestler, R. B., W. W. Bailey, and H. E. McCIure. 1942. Protein requirements of bobwhite quail chicks for survival, growth, and efficiency of feed utilization. Journal of Wildlife Management 6:185-193.
Nomsis, M. J. 1993. SPSS® for Windows: base system user's guide. Release 6.0. SPSS, Chicago, Illinois.
Pedersen, E. K., W. E. Grant, and M. T. Longnecker. 1996. Effects of red imported fire ants on newly-hatched northem bobwhite. Joumal of Wildlife Management 60:164-169.
Rhoades, R. B. 1977. Medical aspects of the imported fire ant. University Presses of Florida, Gainesville, Florida.
28

Ridlehuber, K. T. 1982. Fire ant predation on wood duck ducklings and pipped eggs. Southwestem Naturalist 27:222.
Roseberry, J. L., and W. D. Klimstra. 1971. Annual weight cycles in male and female bobwhite quail. Auk 88:116-123.
Shapiro, S. S., and M. B. Wilk. 1965. An analysis of variance test for normality (complete samples). Biometrika 52:591-611.
Sikes, P. J., and K. A. Amold. 1986. Red imported fire ant (Solenopsis invicta) predation on cliff swallows (Hirundo pyrrhonota) nestlings in east-central Texas. Southwestem Naturalist 31:105-106.
Stoddard, H. L. 1931. The bobwhite quail: its habits, preservation and increase. Charles Scribner's Sons, New York, New York.
Travis, B. V. 1938. The fire ant (Solenopsis spp.) as a pest of quail. Joumal of Economic Entomology 31:649-652.
Vinson, S. B., and L. Greenberg. 1986. The biology, physiology, and ecology of imported fire ants. Pages 193-226 in S. B. Vinson, editor. Economic impact and control of social insects. Praeger Publishers, Westport, Connecticut.
Wegmann, T. G., and O. Smithies. 1966. A simple hemagglutination system requiring small amoimts of cells and antibodies. Transfusion 6:67-73.
Williams, D. F. 1994. Control of the introduced pest Solenopsis invicta in the United States. Pages 282-292 in D. F. Williams, editor. Exotic ants: biology, impact, and control of introduced species. Westview Press, Boulder, Colorado.
29

CHAPTER IV
PLASMA CREATINE KINASE LEVELS ASSOCIATED WITH INCREASED
PROBABILITY OF MORTALITY IN WILD NORTHERN BOBWHITE
Abstract
I assessed the effect of capture myopathy on survival of northem bobwhites (Colinus
virginianus) during January through August of 1997 (n = 48) and 1998 (n = 56) in Texas.
I captured bobwhites by trapping and netting and measured plasma creatine kinase (CK)
concentrations at capture. Elevated plasma CK levels are an indication of muscular
damage associated with capture myopathy. My hypothesis was that northem bobwhite
plasma CK measured at capture would be negatively associated with subsequent survival.
Plasma CK of birds differed due to sampling time/location (P = 0.002), but not capture
method (P = 0.870) or year (P = 0.222). Average plasma CK of groups of birds
processed in the lab (9,290 ± 1,000 u/L [x± SE], « = 13) was almost twice that of groups
processed in the field (5,240 ±510 u/L, n = 38). For birds sampled in the field, plasma
CK was negatively associated with probability of survival to 8, 12, 16, and 20 weeks (P <
0.1), but not with probability of survival to 4 weeks (P = 0.773). The probability of a bird
surviving decreased by 14%) with each 1,000 u/L increase of plasma CK. Plasma CK was
not associated v^th probability of survival to any time period for birds sampled after
transport to a field lab (P > 0.5). Survival estimates differed between birds having high
versus low plasma CK (Z= 1.58, P = 0.057; 54.8 ± 8.2% vs. 72.4 ± 7.5%). If plasma CK
accurately indicated severity of capture myopathy, then capture myopathy resulted in
^ 12.2%o negative bias of the survival estimate (63.6 ± 5.7%o) calculated using all of the
data. These results suggest that demographic estimates based on captured animals are
biased. Plasma CK may be a useftal tool to identify individual animals whose probability
of survival has been influenced by capture-related trauma, and should thus be removed
from analyses.
30

Introduction
Much of our knowledge of wildlife has been gained through studies that capture
animals, mark or radiomark them, and then monitor their fate. Biologists have long
realized that the capture event could alter the fate of animals and have attempted to
account for this effect. For example, mark-recapture models have been developed for
situations where the marking event reduces short-term survival of the animal (Pollock et
al. 1990). In radiotelemetry studies, data from animals that die within some specified
interval after capture are often censored (Cox and Afton 1998). For animals that survive
this interval, the assumption is made that subsequent survival of the radiomarked
population is representative of the population of interest (Pollock et al. 1989). However,
current methods of incorporating potential negative capture effects in demographic
analyses are problematic because they typically assume a short-term effect that is
constant for all animals.
A common injury to wdld animals that results from capture is a condition referred to
as capture myopathy. The condition is characterized by acute degeneration of muscle
tissue resulting from intense muscular exertion and trauma caused by a chase, stmggle, or
transport (Hulland 1993). Low pH vsdthin muscle tissue resulting from the buildup of
lactic acid from anaerobic glycolysis results in increased cell membrane permeability and
cell lysis (Wobeser 1997:218). This releases cellular enzymes, such as CK to the blood,
and elevated levels of these enzymes in serum or plasma are indicative of skeletal or
cardiac muscle damage (Bollinger et al. 1989). Scarring of skeletal and cardiac muscles
may occur, which can contribute to death weeks or months after the causative event
(Hulland 1993). The degree of capture myopathy among individuals captured under
similar conditions, as indicated by semm or plasma concentrations of cellular enzymes,
can vary greatly (Bollinger et al. 1989). Thus, assumptions that subsequent survival of
captured animals is representative of the population of interest or that capture effects are
short-term and constant for all capture animals may be violated if capture myopathy
occurs.
31

Studies of capture myopathy in wild birds have involved necropsy and examination
of muscular lesions (Spraker et al. 1987) and measurement of serum enzymes, usually
CK and aspartate aminotransferase (Bollinger et al. 1989, Dabbert and Powell 1993).
Additionally, researchers have associated particular capture and handling techniques with
subsequent survival, and speculated that the differences in survival rates were likely due
to differences in the number of individuals predated as a result of debilitating effects of
capture myopathy (Cox and Afton 1998). However, no studies have measured serum or
plasma enzymes to determine if higher levels were associated with increased mortality.
The objective of this study was to assess whether concentration of plasma CK, an
indicator of capture myopathy, was associated with survival of radiomarked northem
bobwhite.
Methods
Northem bobwhites were captured from January through March of 1997 and 1998
using walk-in funnel traps (Smith et al. 1981) baited with milo and by netting coveys at
night. Traps were checked traps twice per day, about 4 hr after sunrise and 30 min before
sunset, and coveys were netted 2-4 hr after sunset. I could not estimate how long birds
had been trapped, but trap checks were timed to coincide with the end of morning and
evening northem bobwhite activity periods. Birds were immediately removed from traps
or the net and placed them into ventilated sacks or cardboard boxes. Each group of
captured birds was processed either immediately in the field or after transport to a lab
about 11 km away. Birds processed in the field were typically handled <10 min and then
released immediately. In contrast, birds transported to the lab were held for various times
before processing, often held overnight, and then released back at the capture location.
All hens were weighed, marked with an aluminum legband (National Band and Tag
Co., Newport, Kentucky), radiomarked, bled, and released. Each radio transmitter
weighed 6.5 g, had a mortality sensor, and was a necklace style with no body loop
(American Wildlife Enterprises, Montacello, Florida). Each hen was bled by pricking the
ulnar or brachial vein with a 25-gauge needle and collecting blood into heparinized
32

capillary tubes. Capillary tubes were sealed with clay, transported to a field lab, and
centrifuged. Tubes were then scored with a diamond-tipped pen and broken immediately
above the white blood cell layer. Plasma was pipetted into cryovials, frozen initially at
-20 °C for <4 months and then -84 °C for < 18 months, and shipped on dry ice to a
commercial laboratory (Veterinary Associates Laboratory, Edmond, Oklahoma) for
measurement of CK using a Technicon RA-1000® clinical chemistry analyzer (Bayer
Corporation, Pittsburgh, Pennsylvania). Hemolytic and lipemic samples were discarded
due to potential measurement errors (Alleman 1990). Radiomarked birds were monitored
for survival 1-3 times per week from capture until July or August of each year.
Survival rates were calculated using the staggered-entry Kaplan-Meier model
(Pollock et al. 1989) of program BCAPLAN (T. Kulowiec, Missouri Department of
Conservation, Columbia, Missouri). Precision of the survival estimator is poor when
based on <20 individuals (Pollock et al. 1989), so calculations were limited to periods
when ^20 birds were radiomarked. This resulted in survival estimates for 2 165-day
periods: 10 Febmary 1997 through 24 July 1997 and 14 Febmary 1998 through 28 July
1998.
The effects of capture method (trap or net), processing location (field or lab), and
year (1997 or 1998) on levels of plasma CK were assessed using analysis of variance.
The levels of CK from each group of birds captured at a single time were probably
correlated, violating the assumption for analysis of variance of independent error terms
(Sokal and Rohlf 1995:393). Thus, each group of birds captured during a single capture
event was used as the experimental unit and average CK for each group of birds as the
response variable.
The relationship between survival and plasma CK was assessed using logistic
regression. Survival was assessed at 4, 8, 12, 16, 20, and 24 weeks. Individual birds
were used as the experimental units because survival probabilities were probably
uncorrelated with capture events. I tested for a linear relationship between the logit of
survival and plasma CK using a fiill/reduced model likelihood ratio chi-squared test (SAS
33

Institute 1989). Analyses were performed using PROC LOGISTIC of version 6.12 of
SAS (SAS Institute, Gary, North Carolina).
The assumptions of normality and homoscedasticity were assessed where
appropriate using the Shapiro-Wilk (Shapiro and Wilk 1965) and Levene (1960) tests,
respectively. Analyses were conducted using SPSS for Windows, release 6.0 (SPSS,
Chicago, Illinois), unless specified otherwise. Statistical significance was determined
using an a-Ievel of 0.1 rather than the traditional a-Ievel of 0.05 to reduce the
probability of Type II error (Skalski and Robson 1992:21-22). Means are reported ± 1
standard error.
Results
I captured, bled, radiomarked, and monitored 104 northem bobwhite hens (48 in
1997 and 56 in 1998). Survival of hens was 0.525 ± 0.079 from 10 Febmary to 24 July
1997 and 0.625 ± 0.078 from 14 Febmary to 28 July 1998.
Levels of plasma CK with respect to capture method, processing location, and year
were homoscedastic but nonnormal. Because ANOVA is robust to nonnormality (Zar
1996:187), I chose not to transform data or use nonparametric alternatives. Plasma CK
was influenced by processing location (F, 47 = 10.63, P = 0.002), but not capture method
(F, 47 = 0.03, P = 0.870) or year (F, 47 = 1.53, P = 0.222). Average plasma CK of groups
of birds processed in the lab (9,290 ± 1,000 u/L, n=\3) was almost twice that of groups
processed in the field (5,240 ±510 u/L, n = 38). Because of the observed difference in
plasma CK of birds processed in the field versus the lab, I assessed the relationship
between plasma CK and survival separately for each group. For birds that were
transported back to the lab for processing, plasma CK was not related to the logit of
survival for any interval (Table 4.1). For birds processed in the field, plasma CK was
related to the logit of survival measured to 8, 12, 16, and 20 weeks (Table 4.1). The
probability of a bird surviving to 16 weeks decreased by 14% with each 1,000 u/L
increase of plasma CK (Figure 4.1).
34

Table 4.1. Tests of linear relationships between the logit of survival to 6 time periods and plasma CK sampled either after transport to a laboratory or immediately in the field.
Sampling location Survival interval (weeks)
Laboratory
Field
4
8
12
16
20
24
4
8
12
16
20
24
n /
32
31
30
30
28
16
72
72
70
67
59
43
<0.01
0.25
0.40
0.43
0.26
0.03
0.08
3.00
2.97
3.59
3.42
0.04
0.957
0.616
0.528
0.514
0.613
0.874
0.773
0.083
0.085
0.058
0.065
0.845
35

> >
CO
1.0
0.8
0.6
0.4
0.2
0.0
• • • • • • • •
0 5,000 10,000
Plasma CK (u/L)
15,000
Figure 4.1. Relationship between survival to 16 weeks of northem bobwhite hens (n 61) and concentration of plasma CK for birds sampled in the field. The logit of survival decreased with increasing levels of plasma CK (P = 0.058). The equation depicted by the line is y = e ' 23-000013ix)/( i + d 23-0.00013 Ix)
) .
36

Discussion
Plasma CK was negatively related to survival of northem bobwhites when plasma
CK was sampled immediately after birds were removed from the traps or net. These
results suggest that the effects of capture do not impact all animals equally, an
assumption of models that attempt to incorporate capture effects. Plasma CK may be a
useful diagnostic tool for determining the degree that each animal was impacted by
capture, and the subsequent effect on its survival probability. For birds handled in the
field, I was surprised to find a relationship between plasma CK and survival measured to
every interval from 8 to 20 weeks, but not to 4 weeks (Table 4.1). This suggests that the
effect of capture myopathy on northem bobwhite survival may take >4 weeks to manifest
itself. A conditioning period >4 weeks would be required to account for these effects
using current methods for studies of radiomarked animals; few researchers will find this
feasible given the cost of field studies and the current battery life of transmitters for
avians.
To explore the effect of plasma CK on survival estimates, I tested the hypothesis that
survival of northem bobwhites with high plasma CK was lower than survival of northem
bobwhites with low plasma CK. Hill and Murray (1987) estimated the normal bounds of
plasma CK to be 90-1,590 u/L. Only 1 \% of the birds sampled in the field fell within
this range. Median plasma CK of birds sampled in the field during both years was
3,760 u/L. I used this value to divide these birds into low and high plasma CK groups. I
combined both years of data and restricted the comparison to the interval when ^20
radiomarked birds were monitored (Pollock et al. 1989); this was the 118-day period from
16 Febmary to 14 June. Survival was significantiy lower for the high CK group (1-tailed
Z-test, Pollock et al. 1989; Z= 1.58, P = 0.057; 54.8 ± 8.2% vs. 72.4 ± 7.5%). Estimated
survival during this interval using the combined data was 63.6 ± 5.7%). Assuming plasma
CK accurately indicated severity of capture myopathy and that birds in the low CK group
were unaffected by capture myopathy, then capture myopathy resulted in a -12.2% bias of
the survival estimate calculated using all of the data. Because only 8 of 36 birds in the
37

low CK group had plasma CK levels wdthin the normal range (Hill and Murray 1987), it
is doubtful that these birds were completely unaffected by capture myopathy. Thus, this
estimate of negative bias due to capture myopathy may be conservative. Also, other
factors may have caused further bias in the survival estimate. For example, Osborne et al.
(1997) found that simulated radio transmitters caused birds to lose body mass and lipids,
and thus could potentially lower survival. These other potential effects could not be
assessed in this study. Researchers must recognize that the assumption that capture and
radiomarking animals has either no effect or a constant, short-term effect may be severely
violated.
The failure to find a relationship between survival and plasma CK for birds
sampled in the lab may have been due to the smaller sample size or the lack of
consistency in sampling time for this group. The rise in plasma CK caused by physical
exercise or capture of animals is time dependent. Tripp and Schmitz (1982) found that
CK levels in domestic turkeys were unchanged immediately following 15 min of
continuous exercise, but rose about 5-fold during the next 4 hr and remained at that level
for at least another 24 hr. Dabbert and Powell (1993) found that CK levels in domestic
mallards (Anas platyrhynchos) sampled 45 min following the initiation of capture and
transport was 6-fold that of baseline levels. Thus, plasma CK probably begins to rise
yvithin 15 to 45 min of capture. The difference in plasma CK between birds handled in
the field and lab was probably largely due to the greater time that elapsed between
capture and sampling. The difference may also have partially been due to the effect of
transportation; the tme cause cannot be determined because sampling time and
transportation are confounded. This example illustrates that handling and sampling times
must be consistent for semm or plasma enzyme results to be of any value.
38

Literature Cited
Alleman, A. R. 1990. The effects of hemolysis and lipemia on serum biochemical constituents. Veterinary Medicine 85:1272-1284.
Bollinger, T., G. Wobeser, R. G. Clark, D. J. Nieman, and J. R. Smith. 1989. Concentration of creatine kinase and aspartate aminotransferase in the blood of wild mallards following capture by three methods for banding. Joumal of Wildlife Diseases 25:225-231.
Cox, R. R., Jr., and A. D. Afton. 1998. Effects of capture and handling on survival of female northem pintails. Joumal of Field Ornithology 69:276-287.
Dabbert, C. B., and K. C. Powell. 1993. Semm enzymes as indicators of capture myopathy in mallards (Anas platyrhynchos). Joumal of Wildlife Diseases 29:304-309.
Hill, E. F,, and H. C. Murray. 1987. Seasonal variation in diagnostic enzymes and biochemical constituents of captive northem bobwhites and passerines. Comparative Biochemistry and Physiology 87B:933-940.
Hulland, T.J. 1993. Muscle and tendon. Pages 183-265 m K. V. F. Jubb, P. C. Kennedy, and N. Palmer, editors. Pathology of domestic animals. Fourth edition. Volume 1. Academic Press, New York, New York.
Levene, H. 1960. Robust tests for equality of variances. Pages 278-292 in I. Olkin, editor. Contributions to probability and statistics. Stanford University Press, Palo Alto, California.
Osbome, D. A., B. J. Frawley, and H. P. Weeks, Jr. 1997. Effects of radio tags on captive northem bobwhite (Colinus virginianus) body composition and survival. American Midland Naturalist 137:213-224.
Pollock, K. H., S. R. Winterstein, C. M. Bunck, and P. D. Curtis. 1989. Survival analysis in telemetry studies: the staggered entry design. Joumal of Wildlife Management 53:7-15.
, J. D. Nichols, C. Brownie, and J. E. Hines. 1990. Statistical inference for capture-recapture experiments. Wildlife Monograph 107:1-97.
SAS Institute. 1989. SAS/STAT user's guide. Version 6. Fourth edition. Volume 2. SAS Institute, Gary, North Carolina.
Shapiro, S. S., and M. B. Wilk. 1965. An analysis of variance test for normality (complete samples). Biometrika 52:591-611.
Skalski, J. R., and D. S. Robson. 1992. Techniques for wildlife investigations: design and analysis of capmre data. Academic Press, San Diego, California.
39

Smith, H. D., F. A. Stormer, and R. D. Godfrey, Jr. 1981. A collapsible quail trap. U.S. Forest Service Research Note RM-400.
Sokal, R. R., and F. J. Rohlf 1995. Biometry. Third edition. W. H. Freeman and Company, New York, New York.
Spraker, T. R., W. J. Adrian, and W. R. Lance. 1987. Capture myopathy in wild mrkeys (Meleagris gallopavo) following trapping, handling, and transportation in Colorado. Joumal of Wildlife Diseases 23:447-453.
Tripp, M. J., and J. A. Schmitz. 1982. Influence of physical exercise on plasma creatine kinase activity in healthy and dystrophic turkeys and sheep. American Joumal of Veterinary Research 43:2220-2223.
Wobeser, G. A. 1997. Diseasesof wild waterfowl. Second edition. Plenum Press, New York, New York.
Zar, J. H. 1996. Biostatistical analysis. Third edition. Prentice Hall, Upper Saddle River, New Jersey.
40

CHAPTER V
RELATIONSHIP BETWEEN PLASMA TRIGLYCERIDE, BODY MASS,
AND REPRODUCTION OF NORTHERN BOBWHITES
Abstract
The earliest potential initiation of northem bobwhite (Colinus virginianus)
reproduction is limited by photoperiod. Secondary factors such as lipid reserves, diet, and
stress often limit the beginning of northem bobwhite reproduction, potentially reducing
reproductive success and causing a shorter reproductive season. I measured late winter
body masses and plasma triglycerides of wild northem bobwhites and subsequent
reproductive timing and effort in 1997 and 1998 on the Coastal Prairie of Texas. Using
body mass and plasma triglyceride level as indices of body fat, I tested the hypothesis that
the onset of reproduction and first clutch size was influenced by late winter lipid reserves.
Northem bobwhite plasma triglycerides were higher (P < 0.001) and more variable (P =
0.019) in 1998, and nesting began 15 ± 1.6 days (x± SE) earlier than in 1997. However,
within each year no combination of body mass and triglycerides was associated with
timing of nesting or size of first clutch (P > 0.1). In addition, body masses were not
correlated with plasma triglycerides (P > 0.1), My findings suggest that individual
plasma triglyceride level and body mass are unsuitable variables for assessing within-
population differences in reproductive timing. However, mean plasma triglycerides for a
population may be useful for assessing differences in reproductive timing among years
and locations. The relationship between triglycerides and hormones directly affecting
gonadal recmdescence, such as luteinizing hormone (LH) and prolactin, is uncertain for
wild northem bobwhites. Thus, future studies should assess causes and patterns of
change in these hormones.
Introduction
The initiation of reproduction in birds is controlled by the endocrine system (Bahr
and Johnson 1991). Photoperiod has an overriding influence on reproduction in many
41

species, including northem bobwhite (Kirkpatrick and Leopold 1952), by regulating
hypothalamic secretions of follicle stimulating hormone (FSH) and LH (Famer and
Follett 1979, Carey 1996). After minimum photoperiod is available to birds, secondary
factors may influence hormone levels and determine when individuals begin to reproduce.
Physiological condition, diet, and stress are 3 interrelated secondary factors influencing
northem bobwhite reproduction. For example, water deprivation in northem bobwhites
decreases semm progesterone, delays or prevents egg production, decreases clutch size,
and causes smaller eggs (Cain and Lien 1985, Koerth and Guthery 1991, Giuliano et al.
1995). Insufficient dietary energy increases semm corticosterone and decreases northem
bobwhite egg production (Giuliano et al. 1996). And, lower body mass in late winter may
delay the onset of egg laying in pheasants, probably due to levels of stress and
reproductive hormones (Gates and Woehler 1968). Physiological condition, diet, and
stress are probably strongly correlated among wild birds at any particular time and place
because these factors are largely dependent on environmental conditions. However,
differences in condition, diet, and stress within a population may explain within-year
variability in the timing of reproduction and reproductive effort.
Late winter lipid reserves probably influence future reproduction (Gates and
Woehler 1968), although lipid reserves are less important to reproduction of grouses and
presumably other galliforms as compared to waterfowl (Thomas 1988). Lipid reserves
could increase size of the first clutch by allowing a female to maintain and recmit more
follicles for ovulation, largely through the action of FSH (Carey 1996). Lipid reserves are
correlated with northem bobwhite body masses during winter (Rebel 1972, Frawley et al.
1999), and with both body masses and plasma triglycerides when measured in other
avians (Bacon et al. 1989, Dabbert et al. 1997). Thus, late winter body masses and
plasma triglycerides may explain within-year variability of reproductive timing and first
clutch sizes. I tested the hypothesis that the onset of reproduction and first clutch size of
northem bobwhites was influenced by late winter lipid reserves, using body masses and
plasma triglyceride levels as indices of body fat.
42

Methods
Methods for capturing and monitoring northem bobwhites were described in Chapter
II. Briefly, female northem bobwhites were captured from 1 Febmary through 23 April in
1997 and 1998 primarily with walk-in funnel traps (Smith et al. 1981) baited with milo.
All hens were weighed, marked with an aluminum legband (National Band and Tag Co.,
Newport, Kentucky), radiomarked (American Wildlife Enterprises, Montacello, Florida),
bled, and released. Each hen was bled by pricking the ulnar or brachial vein with a 25-
gauge needle and collecting blood into heparinized capillary tubes. Capillary tubes were
sealed with clay, transported to a field lab, and centrifuged. Tubes were then scored with
a diamond-tipped pen and broken immediately above the white blood cell layer. Plasma
was pipetted into cryovials, frozen initially at -20°C and then -84°C, and shipped on dry
ice to a commercial laboratory (Veterinary Associates Laboratory, Edmond, Oklahoma)
for measurement of triglycerides using a Technicon RA-1000® clinical chemistry
analyzer (Bayer Corporation, Pittsburgh, Pennsylvania). Hemolytic and lipemic samples
were discarded due to potential measurement errors (Alleman 1990).
From late April until mid-July, most radiomarked hens were located at least once
every other day to find nests. Each bird was approached on foot until it flushed or moved
away, was observed, or had been circled by the researcher, indicating that it might be on a
nest. Two sets of stake flags were placed at a distance of 5 and 10 m from the nest
oriented so that each set was aligned with the nest. The position of the nest could be
determined on subsequent visits by sighting down both sets of stake flags and
determining the point where the 2 lines intersected. This marking method was used so
that predators investigating the stake flags were not drawn directly to the nest. Hens were
never intentionally flushed from nests and rarely flushed inadvertently. To confirm the
nest location and clutch size, the nest site was checked at times when the hen might be
away from the nest feeding. When the nest was unoccupied, the number of eggs was
recorded. The date that eggs in a nest began to be incubated was estimated by averaging
the last date that a hen was not found on a nest and the first date that a hen was found
43

incubating eggs on a nest. The date that nesting began was estimated by subtracting the
number of eggs in a nest from the estimated date that incubation of eggs began.
Data Analysis
The value of late-winter body mass and plasma triglycerides as predictors of first
clutch size and date of first nest initiation was assessed using multiple linear regression.
Body masses and plasma triglycerides were recorded from birds captured 6-16 Febmary
1997 and 4 Febmary-7 March 1998. Records for 5 nests initiated >30 days after the
earliest nests each year were censored because it was suspected that the initial nests for
these birds were not detected. First nests would probably not have been detected if they
were depredated before incubation began or if the male incubated the eggs.
Residuals were tested for normality using the Shapiro-Wilk test (Shapiro and Wilk
1965). Homoscedasticity and linearity were assessed by examining plots of residual and
predicted values. Analyses were conducted using SPSS for Windows, release 6.0
(Nomsis 1993). Statistical significance was determined with a = 0.05. Means are
reported ± 1 SE.
Results
Body masses, plasma triglyceride levels, dates of nest initiation, and sizes of first
clutches were obtained for 17 northem bobwhite hens in 1997 and 23 in 1998 (Table 5.1).
Northem bobwhites began nesting 15 ± 1.6 days earlier in 1998 ( 35 = 6.70, P < 0.001).
Clutch sizes did not differ between years (t-^^ = 0.86, P = 0.394). The mean and variance
of plasma triglycerides were higher in 1998 (mean: t-test for unequal variances, 30 =
-5.53, P < 0.001; variance: Levene's [1960] test, F, 33 = 5.99, P = 0.019). Body masses
were lower in 1998 (t-test for equal variances, 35 = 2.09, P = 0.043). Plasma triglycerides
were not correlated with body mass either year (1997: R^ = 0.01, F, ,5 = 0.20, P = 0.660;
1998: R^ = 0.12, F, 2, = 2.87, P = 0.105; Figure 5.1).
44

Table 5.1. First clutch sizes, dates of nest initiation, late winter body masses (g), and plasma triglyceride levels (mg/dL) of northem bobwhites in 1997 (n= \1) and 1998 (n = 23), Refugio County, Texas.
First clutch size
Date of nest initiation
Body mass
Plasma triglyceride
1997
X
15.7
3 May
167.7
122.2
SE
0.3
1.9
2.6
9.6
1998
X
15.2
18 April
161.1
250.1
SE
0.4
1.4
1.9
21.0
Tests for relationships among variables were conducted separately for each year
because birds began reproducing earlier in 1998. No combination of body mass and
triglycerides successfully predicted date of nest initiation or first clutch size (Table 5.2).
Assumptions of normality and homoscedasticity were violated for analyses of clutch size
in 1998 due to a clutch of 22 eggs. The analysis was repeated with this record removed
with identical results.
Discussion
Late winter body mass and plasma triglycerides were not related to date of nest
initiation or first clutch size either year. This contradicts patterns found in turkeys
(Meleagris gallopavo; Badyaev et al, 1996), where body mass in Febmary-March and
nest initiation date were the best predictors of clutch size of first nests. However, in the
turkey study (Badyaev et al. 1996), the partial r for female body mass was 0,10. Thus,
turkey body mass accounted for only an additional !%> of the variation in clutch sizes
given the other variable in their model. If body mass influenced clutch size this weakly in
my study, I could not have detected it. Both studies support earlier conclusions that
galliform reproduction relies very little on lipid reserves (Thomas 1988),
45

^ 500 _ i 5 P 400 • ^^
(0 ^ 300 '&—
0 ^ 200 D)
^ 100
n u
•
• •
^
•
•
•
•
•
1997
•
• • •
1
3- 500
I' 400 (/)
^ 300
^ 200
• 100 0
-
-
•
•
-
• •
1
•
•
• •
• •
•
• ••
•
•
•
• •
• •
• •
1
1998
•
1
140 150 160 170 180 190
Body Mass (g)
Figure 5,1. Lack of linear relation between plasma triglycerides and body mass of northem bobwhites in 1997 and 1998, Reftigio County, Texas (1997: R^ 0.01, F , ,5 = 0.20, P = 0.660; 1998: R^ = 0.12, F , 2, = 2.87, P = 0.105).
46

Table 5.2. R^ and significance of late winter body mass (BM) and plasma triglycerides (TRIG) as predictors of first clutch size and date of nest initiation for northem bobwhites in Refugio County, Texas, 1997-1998.
Dependent variable Year Independent variables R'
First clutch size
Date of nest initiation
1997 BM + TRIG
BM
TRIG
1998 BM + TRIG
BM
TRIG
1997 BM + TRIG
BM
TRIG
1998 BM + TRIG
BM
TRIG
0.19
0.12
0.09
0.03
0.00
0.03
0.03
0.02
0.02
0.08
0.03
0.03
0.235
0.172
0.249
0.752
0.970
0.466
0.794
0.582
0.634
0.414
0.463
0.435
In 1997, plasma triglyceride levels from my study were similar to winter plasma
triglyceride levels reported for northem bobwhites housed in outdoor pens and fed a
commercial gamebird feed ad libitum (Hill and Murray 1987). However, triglyceride
levels in 1998 were twice as high and twice as variable. I examined the relationship
between sampling date and plasma triglycerides in 1998 to determine if my sampling
interval extended into the beginning of the breeding season when triglycerides increase 3-
to 6-fold (Hill and Murray 1987). If this occurred, I predicted that plasma triglycerides
would increase during the 31-day sampling interval in 1998. However, I found no linear
relationship between plasma triglycerides and sampling date (F, 21 = 0.5, P = 0.507;
Figure 5.2). I suspected that differences in sampling locations also may have caused the
observed annual differences. In 1997, 13 of 17 samples were obtained from birds
captured within a 1-km radius. In 1998, samples were obtained from 8 locations
47

separated by >2 km, and the largest number of samples from any single location was 7.
To test whether the differences in the means and variability of plasma triglycerides were
due to location, I compared the samples collected in 1997 from a single location with 4
samples collected from that location in 1998, Surprisingly, the means and trends for 1997
and 1998 of plasma triglyceride levels and body masses from this single location were
nearly identical to that of all locations (Table 5.3). Thus, I do not know why triglyceride
levels were so much greater and variable in 1998.
Table 5.3. Comparison of northem bobwhite late winter body masses (g) and plasma triglyceride levels (mg/dL) in 1997 and 1998 between a single pasture (Willow Lake Trap [WLT], « = 13 and 4, respectively) and all locations (n = 17 and 23, respectively), Refugio County, Texas. Means are followed by standard deviations (SD).
1997 1998
WLT All WLT All
SD X SD X SD X SD
Body mass 166.8 11.2
Plasma triglyceride 125.8 38.8
167.7
122.2
10.8
39.5
156.3
262.0
8.9
190.5
161.1
250.1
9.3
100.9
Higher plasma triglycerides in 1998 were associated with earlier reproduction.
However, I found no relationship between individual plasma triglyceride levels and date
of reproduction. I suspect that northem bobwhites in my study were differentially
affected by numerous extrinsic factors, such as differences in diet caused by habitat
quality and disturbance by predators. In addition, some extrinsic factors probably
affected all of the birds to nearly the same extent within a given year; these effects would
include winter severity and precipitation. My resuUs suggest that plasma triglycerides
may be useful as an indicator of population trends in timing of reproduction, but not for
predicting reproductive timing of individuals.
48
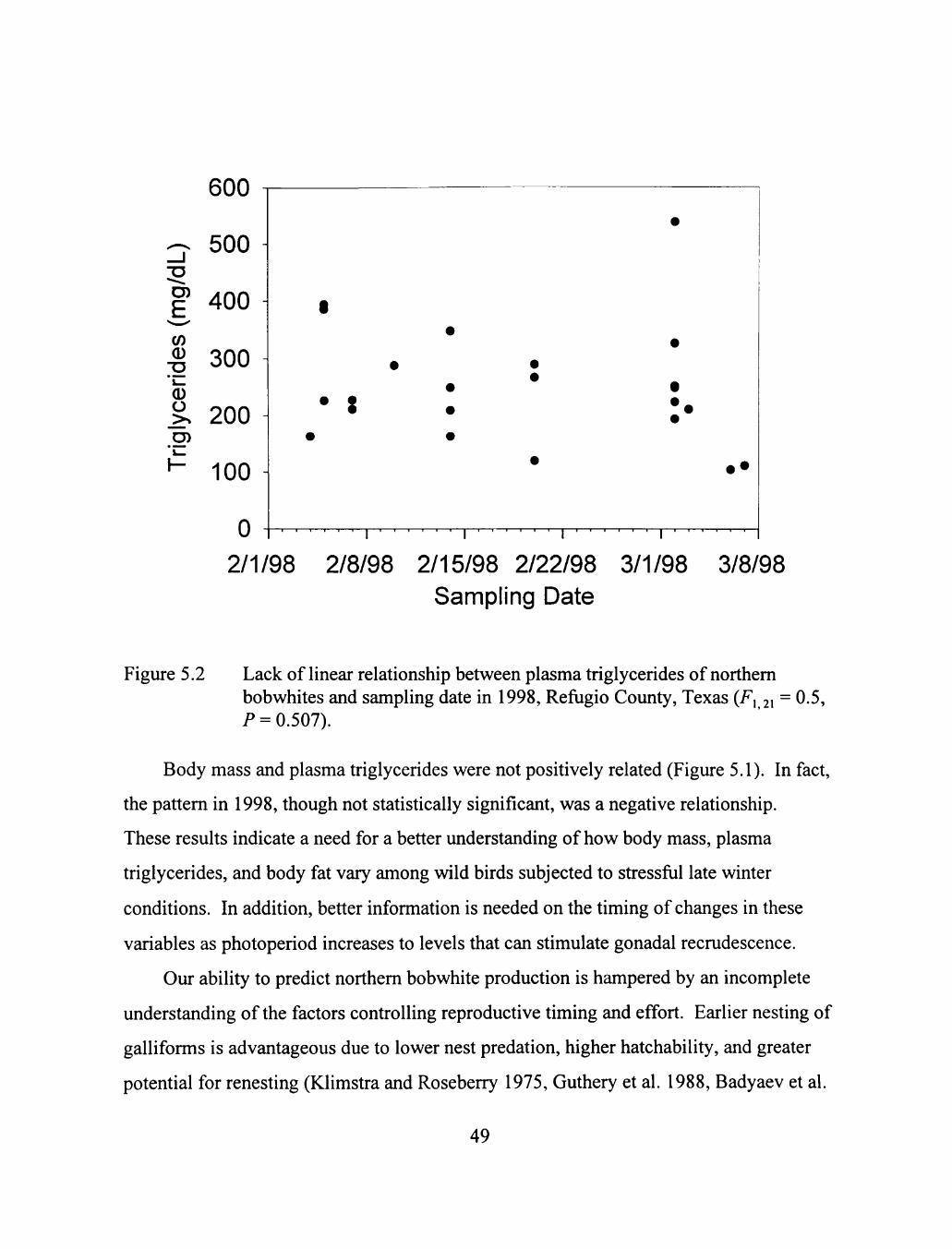
600
2/1/98 2/8/98 2/15/98 2/22/98 3/1/98 3/8/98 Sampling Date
Figure 5.2 Lack of linear relationship between plasma triglycerides of northem bobwhites and sampling date in 1998, Refugio County, Texas (F, 21 = 0.5, P = 0.507).
Body mass and plasma triglycerides were not positively related (Figure 5.1). In fact,
the pattern in 1998, though not statistically significant, was a negative relationship.
These results indicate a need for a better understanding of how body mass, plasma
triglycerides, and body fat vary among wild birds subjected to stressftjl late winter
conditions. In addition, better information is needed on the timing of changes in these
variables as photoperiod increases to levels that can stimulate gonadal recmdescence.
Our ability to predict northem bobwhite production is hampered by an incomplete
understanding of the factors controlling reproductive timing and effort. Earlier nesting of
galliforms is advantageous due to lower nest predation, higher hatchability, and greater
potential for renesting (Klimstra and Roseberry 1975, Guthery et al. 1988, Badyaev et al.
49

1996, but see Lehmann 1946), Photoperiod limits the potential reproductive period in
northem bobwhites (Kirkpatrick and Leopold 1952), but secondary factors such as lipid
reserves, diet, and stress can constrain the realized reproductive period (Gates and
Woehler 1968, Cain and Lien 1985, Koerth and Guthery 1991, Giuliano et al, 1995,
1996). My findings indicate that plasma triglycerides and body masses may be useful for
comparing populations among years and locations, but these variables do not explain
within-population variation in reproductive timing and effort. Future research on
reproduction of northem bobwhites should measure hormones more closely linked to
gonadal recrudescence and ovulation and inhibitory effects of stress hormones such as
corticosterone. Research on reproduction of wild northem bobwhite can build on the
excellent foundation of recent laboratory studies that measured reproductive and stress
hormones (Cain and Lien 1985, Giuliano et al. 1995, 1996).
50

Literature Cited
Alleman, A, R. 1990. The effects of hemolysis and lipemia on semm biochemical constituents. Veterinary Medicine 85:1272-1284.
Bacon, W. L,, K, E, Nestor, and E, C, Naber. 1989. Prediction of carcass composition of turkeys by blood lipids. Poultry Science 681282-1288.
Badyaev, A. V,, T. E. Martin, and W. J. Etges. 1996, Habitat sampling and habitat selection by female wild turkeys: ecological correlates and reproductive consequences. Auk 113:636-646.
Bahr, J. M., and P. A. Johnson. 1991. Reproduction in poultry. Pages 555-575 in P. T. Cupps, editor. Reproduction in domestic animals. Fourth edition. Harcourt Brace Jovanovich, San Diego, California.
Cain, J. R., and R. J. Lien. 1985. A model for drought inhibition of bobwhite quail (Colinus virginianus) reproductive systems. Comparative Biochemistry and Physiology 82A:925-930.
Carey, C, 1996. Female reproductive energetics. Pages 324-374 m C. Carey, editor. Avian energetics and nutritional ecology. Chapman & Hall, New York, New York.
Dabbert, C, B., T, E. Martin, and K. C. Powell. 1997. Use of body measurements and semm metabolites to estimate the nutritional status of mallards wintering in the Mississippi Alluvial Valley, USA. Joumal of Wildlife Diseases 33:57-63.
Famer, D. S., and B. K. Follett. 1979. Reproductive periodicity in birds. Pages 829-872 in E. J. W. Barrington, editor. Hormones and evolution. Volume 2. Academic Press, New York, New York.
Frawley, B. J., D. A. Osbome, H. P. Weeks, Jr., L. W. Burger, Jr., and T. V, Dailey. 1999. Use of total body electrical conductivity to predict northem bobwhite lipid mass. Joumal of Wildlife Management 63:695-704.
Gates, J. M., and E. E. Woehler, 1968. Winter weight loss related to subsequent weights and reproduction in penned pheasant hens. Joumal of Wildlife Management 32:234-247.
Giuliano, W, M,, R. S. Lutz, and R Patino. 1995. A simulated insect diet as a water source for quail: effects on body mass and reproduction. Comparative Biochemistry and Physiology 111 A:299-302.
, , and . 1996, Reproductive responses of adult female northem bobwhite and scaled quail to nutritional stress. Joumal of Wildlife Management 60:302-309.
Guthery, F. S., N. E. Koerth, and D. S. Smith. 1988. Reproduction of northem bobwhites in semiarid environments. Joumal of Wildlife Management 52:144-149,
51

Hill, E. C , and H. C. Murray. 1987. Seasonal variation in diagnostic enzymes and biochemical constituents of captive northem bobwhites and passerines. Comparative Biochemistry and Physiology 87B:933-940,
Kirkpatrick, C. M., and A. C. Leopold. 1952. The role of darkness in sexual activity of thequaiL Science 116:280-281.
Klimstra, W. D., and J. L. Roseberry. 1975. Nesting ecology of the bobwhite in southern Illinois. Wildlife Monographs 41:1-37.
Koerth, N. E,, and F. S. Guthery. 1991. Water restriction effects on northem bobwhite reproduction. Joumal of Wildlife Management 55:132-137.
Lehmann, V. W. 1946. Bobwhite quail reproduction on southwestem Texas. Joumal of Wildlife Management 10:111-123.
Levene, H. 1960. Robust tests for equality of variances. Pages 278-292 in I. Olkin, editor. Contributions to probability and statistics. Stanford University Press, Palo Alto, California.
Nomsis, M. J. 1993. SPSS® for Windows: base system user's guide. Release 6.0. SPSS, Chicago, Illinois.
Robel, R. J. 1972. Body fat content of bobwhites in relation to food plantings in Kansas. National Bobwhite Quail Symposium 1:139-149.
Shapiro, S. S., and M. B. Wilk. 1965. An analysis of variance test for normality (complete samples). Biometrika 52:591-611.
Smith, H. D., F. A. Stormer, and R. D. Godfrey, Jr. 1981. A collapsible quail trap. U.S. Forest Service Research Note RM-400.
Thomas, V. G. 1988. Body conditions, ovarian hierarchies, and their relation to egg formation in Anseriform and Galliform species. Pages 353-363 in H. Oullet, editor. Acta XIX Congressus Intemationalis Omithologici. University of Ottawa Press, Ottawa, Canada.
52