original article role of cxcl13-cxcr5 crosstalk between...
TRANSCRIPT

1
ORIGINAL ARTICLE
Role of CXCL13-CXCR5 crosstalk between malignant neuroblastoma cells and
Schwannian stromal cells in neuroblastic tumors
Federica Del Grosso1, Simona Coco1, Paola Scaruffi1°, Sara Stigliani1, Francesca Valdora2, Roberto
Benelli3, Sandra Salvi4, Simona Boccardo4, Mauro Truini4, Michela Croce5, Silvano Ferrini5, Luca
Longo1,6, Gian Paolo Tonini1*
1Translational Oncopathology, National Cancer Research Institute (IST), Genoa, Italy.
1°Translational Oncopathology, National Cancer Research Institute (IST), Genoa, Italy. (present
address Center of Physiopathology of Human Reproduction, Dept. Obstetrics and Gynecology,
"San Martino" Hospital, Genoa, 16132, Italy).
2Department of Oncology and Genetics (DOBIG), University of Genoa, Genoa, Italy.
3Department of Immunology, National Cancer Research Institute (IST), Genoa, Italy.
4Department of Diagnostic Technologies, National Cancer Research Institute (IST), Genoa, Italy.
5Laboratory of Immunological Therapy, National Cancer Research Institute (IST), Genoa, Italy.
6Italian Neuroblastoma Foundation, National Cancer Research Institute (IST), Genoa, Italy
Running title: CXCL13-CXCR5 cross-talk in neuroblastoma
Keywords: neuroblastoma, metastasis, CXCL13, CXCR5, chemokine
*Corresponding author: Gian Paolo Tonini, Translational Oncopathology, National Cancer
Research Institute (IST), L.go R. Benzi, 10 - 16132 - Genoa, Italy. Phone: +39-010-5737487/381;
Fax: +39-010-5737487; e-mail: [email protected]
on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

2
Abstract
Neuroblastoma (NB) is a stroma-poor (SP) aggressive pediatric cancer belonging to Neuroblastic
Tumors (NTs), also including ganglioneuroblastoma and ganglioneuroma, two stroma-rich (SR)
less aggressive tumors. Our previous gene expression profiling analysis showed a different CXCL13
mRNA expression between SP and SR tumors. Therefore, we studied 13 SP and 13 SR tumors by
RT-qPCR and we found that CXCR5b was more expressed in SP than in SR and CXCL13 was
predominantly expressed in SR tumors. Then, we isolated neuroblastic and Schwannian stromal
cells by laser capture microdissection and we found that malignant neuroblasts express CXCR5b
mRNA, whereas Schwannian stromal cells express CXCL13. Immunohistochemistry confirmed that
stroma expresses CXCL13 but not CXCR5. To better understand the role of CXCL13 and CXCR5
in NTs we studied 11 NB cell lines and we detected a heterogeneous expression of CXCL13 and
CXCR5b. Interestingly, we found that only CXCR5b splice variant was expressed in both tumors
and NB lines, whereas CXCR5a was never detected. Moreover, we found that NB cells expressing
CXCR5 receptor migrate towards a source of recombinant CXCL13. Lastly, NB cells induced to
glial cell differentiation expressed CXCL13 mRNA and protein. The chemokine released in the
culture medium was able to stimulate chemotaxis of LA1-5S NB cells. Collectively, our data
suggest that CXCL13 produced by stromal cells may contribute to the generation of an environment
in which the malignant neuroblasts are retained, thus limiting the possible development of
metastases in patients with stroma-rich tumor.
on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

3
Introduction
Neuroblastic Tumors (NTs) are a group of pediatric cancers that onset as localized or disseminated
disease (1). Patients with localized tumor have good prognosis with a 3-years overall survival (OS)
of 90-95%, whereas patients older than one year with a disseminated disease have a worse
prognosis with a 3-years OS ranging between 30% and 35%. This dramatic behavior of the
metastatic disease is mainly due to the capacity of neuroblastoma (NB) cells to metastasize at bone,
bone marrow, liver and lymph nodes. Particularly, in patients with disseminated disease, the normal
bone marrow cell population is almost completely substituted by malignant neuroblasts that find a
favorable environment for their proliferation.
NTs show a quite heterogeneous histology with different tumor histotypes including: neuroblastoma
predominantly composed of small round undifferentiated or poorly differentiated neuroblastic cells,
with few or absent Schwannian stromal cells, ganglioneuroblastoma intermixed (GNB) and
ganglioneuroma (GN) mostly composed of Schwannian stromal cells in which some nests or very
few neuroblastic cells are present (2). The absence of stromal cells makes the tumor more
aggressive and patients have a worse outcome. On the contrary, the abundance of Schwannian
stromal cells, as observed in GNB and GN, is associated with a less aggressive tumor and a
localized disease with a more favorable prognosis (1, 3).
Several evidences support a non-malignant origin of Schwannian stromal cells (4, 5), which are
supposed to control tumor growth by secreting soluble factors that influence cell proliferation,
differentiation and angiogenesis. Indeed, various molecules have been found expressed by
Schwannian stromal cells (6, 7), although their role in controlling neuroblastic cells growth and
on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

4
dissemination is not completely understood. In several tumors apart NTs, the presence of stromal
cells is associated with a better disease outcome and the contiguity between stroma and tumor cells
has an important role in cancer growth and invasion.
Recently, we studied a series of NTs by gene expression profiling analysis and we observed
different levels of CXCL13 transcripts in microdissected neuroblastic cells with respect to
Schwannian stromal cells (5, 8). Our results showed that this chemokine was more expressed in
stromal cells than in malignant neuroblasts, suggesting a functional role of CXCL13 in the
interaction between stroma and neuroblastic cells.
Chemokines are a large family of molecules that, together with their receptors, are involved in the
regulation of chemotaxis, cell proliferation, cell migration and several other crucial cell-cell
interactions (9). Moreover, chemokines play a major role in tumor growth, angiogenesis and
invasion (10, 11) in several cancers and neoplastic cells may express chemokine receptors (12).
Over the last years, CCR1, CCR5, CCR6, CCR9, CXCR1, CXCR2, CXCR4, CXCR5, and CXCR6
were found expressed in NB and it has been shown that CXCR4 is virtually expressed in all human
NB cell lines (13-16).
Starting from our observation we further investigated the expression of CXCL13 and its receptor
CXCR5 in neuroblastic primary tumors and human NB cell lines. CXCL13 and CXCR5 mRNAs
were differently expressed between Schwannian stromal cells and neuroblastic cells suggesting a
cell-cell crosstalk via CXCL13-CXCR5 axis. This cross-talk may contribute to retain neuroblastic
cells within stroma-rich tumors and possibly to inhibit the malignant cells dissemination. Our
on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

5
results add new information about the CXCL13-CXCR5 axis in NTs and in the cross-talk between
neuroblastic and stromal cells.
Materials and Methods
Tumor samples, NB cell lines and total RNA isolation. Tumor specimens were collected from 26
patients at onset of disease, who were diagnosed with a primary NT and referred to the Gaslini
Children’s Hospital, Genoa, Italy. The study was approved by Ethics committee of the Gaslini
Children’s Hospital, and informed consent was obtained by all children’s legal guardians.
According to the International Neuroblastoma Pathology Committee (2), 13 samples were from SP
tumors with at least 80% of neuroblastic cells, and 13 were from SR with Schwannian stromal cells
ranging from 80% to 90% (Supplementary Table 1). Eleven NB cell lines (ACN, GI-CA-N, GI-LI-
N, GI-ME-N, IMR-5, IMR-32, LA1-5S, SH-SY5Y, SK-N-BE(2), SK-N-BE(2)c, SK-N-SH) and the
Raji B-lymphocytic cell line were cultured at 37°C and 5% CO2 in RPMI 1640, supplemented with
L-glutamine, penicilline/streptomycin, non-essential amino acids, and 10% Foetal Bovine Serum
(FBS) (Lonza, Basel, Switzerland). Cells were removed from substrate with PBS/EDTA. All cell
lines were tested for mycoplasma contamination and authenticated (Supplementary Table 2). Total
RNA was isolated from NB cell lines by PerfectPureTM RNA Cell Kit (5Prime, Hamburg,
Germany), and from tumor tissues by PerfectPureTM Tissue RNA Cell Kit (5Prime, Hamburg,
Germany), and treated with RNase-free DNase I. RNA integrity and quantification were checked by
RNA 6000 Nano LabChip kit and 2100 BioAnalyzer instrument (Agilent Technologies, Santa
Clara, CA, USA).
on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

6
Isolation of neuroblastic and Schwannian stromal cells by laser capture microdissection and
total RNA extraction. About 800 cells were laser microdissected from 3 SP and 3 SR frozen
tumors in order to obtain pure cell populations of neuroblastic cells and Schwannian stromal cells,
as previously described (8). Total RNA from laser capture microdissection-derived material was
extracted by PicoPureTM RNA isolation kit (Arcturus Engineering, CA, USA), including a DNase
treatment, RNA quality control and quantification wereas performed by RNA 6000 Pico LabChip
kit and the 2100 BioAnalyzer instrument (Agilent Technologies).
Reverse transcription quantitative real-time PCR (RT-qPCR). RNA (1 μg) from NB cell lines
and tumors was reverse transcribed using 20 pmoles of random hexamers (Eppendorf, Hamburg,
Germany) and 200 U of SuperScript II enzyme (Invitrogen Life Technologies, Carlsbad, CA, USA).
RNA (50 ng) from microdissected cells was amplified and reverse transcribed by WT-OvationTM
RNA Amplification System kit (NuGEN Technologies, San Carlos, CA, USA). CXCL13, CXCR5,
CXCR5a, S100A6 and 18S rRNA transcripts were quantified by TaqMan® Gene Expression Assays
(Applied Biosystems, Foster City, CA, USA). An assay specific for CXCR5b isoform was designed
by PrimerDesign Ltd (Hants, UK). 18S rRNA was used as reference gene. Any amplified product
with a quantification cycle (Cq) higher than 36 cycle was considered undetectable. According to the
Minimum Information for Publication of Quantitative Real-Time PCR Experiments (MIQE) (17,
18), a checklist including technical details is submitted as Supplementary data (MIQE checklist).
The relative amount of each transcript was determined using the equation 2-dCq, where dCq =
(Cqtarget gene – Cq18S rRNA).
on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

7
CXCR5 detection in NB cells by flow cytometry
NB cell suspensions (3x105 cells/tube) and Raji cell line suspension (1.5x105 cells/tube) were
washed with PBS and incubated with 2.5 μg/ml monoclonal mouse anti-human CXCR5/Blr-1
primary antibody (R&D Systems, Inc., Minneapolis, MN, USA) in 2% BSA/PBS for 30 minutes on
ice. After incubation, cells were washed twice with 2% BSA/PBS and 50 μl of goat anti-mouse
IgG2b-PE secondary (Becton Dickinson, San Josè, CA, USA) antibody was added. Cell
suspensions were incubated for 30 minutes on ice. Percentage of positive cells and mean
fluorescence intensity of stained cells were analyzed using a FACScan (Becton Dickinson). The
background levels were set using cells stained with secondary antibody alone and viable cells were
discriminated by evaluating the level of propidium incorporation.
CXCR5 and CXCL13 detection in NB cells by immunofluorescence. NB cells were directly
fixed in a methanol fixing solution and were dropped on microscope glass slides and dried at room
temperature (RT). Slides were incubated 30 minutes with 10% goat serum in PBS. Primary
monoclonal anti-human CXCR5/Blr-1 mouse antibody was diluted 1:40 with PBS 1.5% normal
goat serum, applied on slides at a concentration of 12 μg/ml and incubated 1 hour at RT in a
humidified chamber. After washing, slides were incubated 40 minutes at RT in a humidified
chamber with goat anti-mouse-IgG2b-FITC conjugated secondary antibody (Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA) diluted to a concentration of 1.5 μg/ml with PBS 3%
goat normal serum. In order to identify the expression of CXCL13, acetone permeabilized cells
were incubated 1 hour at room temperature in a humidified chamber with primary anti-human
CXCL13 goat antibody diluted 1:50 with PBS 1.5% normal donkey serum, applied on slides at 2
on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

8
μg/ml. After washing, slides were incubated 45 minutes at RT in a humidified chamber with donkey
anti-goat IgG(H+L) TRITC conjugated secondary antibody (Jackson Immunoresearch Europe Ltd,
Suffolk, UK) diluted 1:50 with PBS 3% donkey normal serum. Staining without primary antibodies
was performed as negative control. Slides were mounted with mounting medium and inspected
under the microscope with a fluorescence lamp.
CXCR5 detection in primary tumor by immunofluorescence. Frozen sections of 4 �m from 5
SP and 5 SR tumors were fixed in 4% paraformaldehyde for 2 minutes. Sections were incubated
with Ultra V Block (Thermo Fisher Scientific, LabVision Inc., Waltham, MA, USA) for 8 minutes
at RT to block unspecific binding of the antibody. Primary monoclonal anti-human CXCR5/Blr-1
mouse antibody was diluted 1:40 with PBS and incubated 1 hour at RT in a humidified chamber.
After washes three-time, slides were incubated for 45 minutes at RT in a humidified chamber with
goat anti-mouse IgG (H+L) Alexa Fluor® 488 Conjugate (Molecular Probe, Invitrogen, Inc.,
Carlsbad, CA, USA) diluted 1:100 in PBS and counterstained with DAPI. Staining without primary
antibody was performed as negative control.
CXCR5 and CXCL13 detection in primary tumors and NB cells by immunohistochemistry.
Immunohistochemistry analysis was performed using polymeric complex technique and automated
immunostainer Benchmark XT (Ventana Medical System, Tucson, AR, USA). Sections of 5 μm
from the same tumor samples were air dried and fixed (10 minutes with acetone and 5-10 seconds
in methanol at RT) and methanol fixed NB cell lines (SH-SY5Y and LA1-5S) were incubated for
30 minutes at 37°C with monoclonal anti-human CXCR5/BLR1 mouse antibody (20 μg/ml for NB
cell lines; 10 μg/ml for tumor sections) and anti-human CXCL13/BLC/BCA-1 goat antibody (0.8
on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

9
μg/ml for tumor sections) (R&D Systems, Inc) and anti-human GFAP (Glial fibrillary acidic
protein) policlonal rabbit antibody (Ventana Medical System). Anti-CXCR5 antibody was detected
by UltraViewRedIM detection kit (Ventana Medical System) based on polymeric alkaline
phosphatase detection system. Staining with anti-CXCL13 antibody was detected by LSAB+System
HRP Kit (DAKO Cytomation, Glostrup, Denmark). Fixed cells and sections were counterstained
with Gill modified hematoxylin (Ventana Medical System), dehydrated, and mounted. Staining
without primary antibody was performed as negative control. Staining of Raji cells and frozen or
paraffin-embedded from lymph nodes and spleen samples was performed as positive control.
Glial differentiation assays. N-type (SH-SY5Y), I-type (SK-N-BE(2)c) and S-type (LA1-5S) NB
cell lines were grown by adding 10 mM 5’-Bromo-2’-deoxyuridine (BrdU) (Sigma Aldrich, Inc.,
San Louis, MO, USA) to cell culture medium to induce glial differentiation (19). Treatment was
maintained for 3 weeks by renewing media three times per week. During the first and the second
week of treatment, cell proliferation was not affected by the treatment and cells were split twice.
Culture media of treated cells were collected every three days for 3 weeks and then concentrated
from 40 ml to 200 μl with centrifugal filter devices Centricon Plus-70 (Millipore, Inc., Billerica,
MA, USA). Total RNA was extracted from the cells after 1, 2 and 3 weeks of treatment with BrdU.
Pictures of differentiated cells have been taken after 3 weeks of treatment.
CXCL13 detection in NB cell line conditioned medium by ELISA. CXCL13 protein
concentration was detected in NB cell line conditioned medium by Quantikine Human CXCL13
Immunoassay (R&D Systems, Inc.), following the manufacturer’s recommendations. The assay
employs the quantitative sandwich enzyme immunoassay technique. A monoclonal antibody for
on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

10
CXCL13 was pre-coated onto a microplate. Human recombinant CXCL13 (hrCXCL13) standards,
saliva, untreated and BrdU differentiated NB cells conditioned media were pipetted into the wells
and CXCL13 was bound by the immobilized antibody. After washing, an enzyme-linked
monoclonal antibody specific for CXCL13 was added. After incubation and extensive washing, a
substrate solution was added and the amount of CXCL13 was quantified by a colorimetric
measurement on a standard curve of serial dilution of hrCXCL13.
NB cell migration assay. Cell migration assays were performed on SH-SY5Y, ACN and LA1-5S
cells in microchambers (NeuroProbes Gaithersburg, MD, USA) as previously described (20). Cells
were extensively washed with PBS, re-suspended in serum-free medium, and placed in the upper
compartment. The two compartments of the chamber were separated by 5 μm pore size
polycarbonate filters coated with 0.01% gelatine. The hrCXCL13 (100 or 200 ng/ml) and
concentrated serum-free conditioned medium from BrdU-differentiated NB cells were used as
chemoattractants in the lower chamber. To obtain specific neutralization of hrCXCL13 bioactivity a
blocking anti-human CXCL13/BLC/BCA-1 goat antibody (R&D Systems, Inc) was added to
hrCXCL13 and to BrdU differentiated NB cells conditioned medium in the lower chamber at 3.5
μg/ml and 0.5 μg/ml, respectively. In order to specifically neutralize CXCR5 receptor on NB cell
surface, cells were previously incubated 30 minutes with 15 μg/ml blocking monoclonal anti-human
CXCR5/BLR1 mouse antibody (R&D Systems, Inc). After 6 hours of incubation at 37°C in 5%
CO2, the filters were recovered, the cells on the upper surface were mechanically removed, and the
cells on the lower surface were fixed and stained. The migration was measured by counting the
on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

11
remained cells on the filter lower surface. Experiments were done in six replicates and repeated
three times.
Results
CXCL13 and CXCR5b mRNA expression in whole tumor, isolated neuroblastic and
Schwannian stromal cells and in NB cell lines. We performed RT-qPCR of 26 tumor samples to
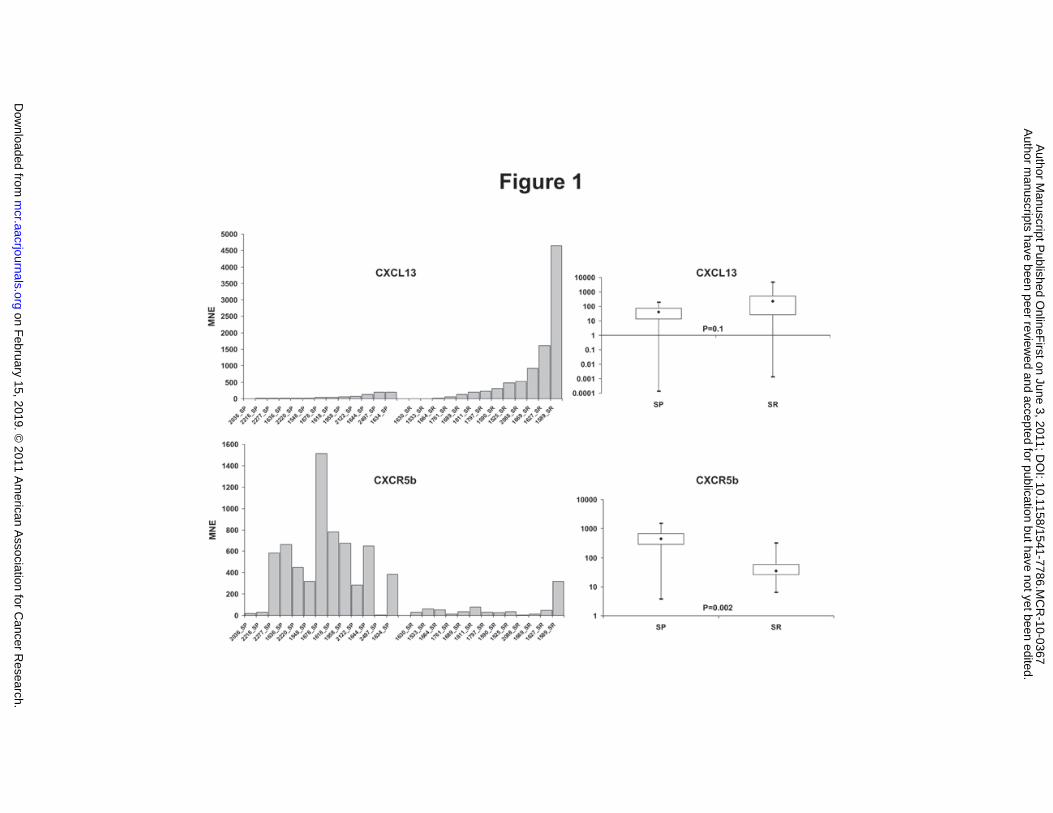
assess the expression of CXCL13 and CXCR5b mRNA in NTs. We firstly observed that SP tumors
showed higher levels of CXCR5b mRNA than SR tumors. Conversely, CXCL13 was more
expressed in SR tumors (Figure 1). Noteworthy, RT-qPCR analysis showed CXCR5b transcript
variant (accession number 032966) (21) in NTs. In our analysis we used primers for both CXCR5a
and CXCR5b variants but only the CXCRb transcript was detected whereas CXCR5a was not
detectable in any samples (data not shown).
Since NTs show tissue heterogeneity and non-malignant cells may infiltrate the tumor, we isolated
either neuroblastic or Schwannian stromal cells from 6 NTs (3 SP and 3 SR) by laser capture
microdissection. Next, we extracted total RNA from both purified cell populations and we
performed RT-qPCR for CXCL13 and CXCR5b. We observed expression of CXCL13 in the stromal
cell components, as opposed to very low or undetectable levels observed in isolated neuroblastic
cells (Figure 2), supporting our results obtained by whole tumors analysis. On the contrary, we
definitely detected CXCR5b expression in the isolated neuroblastic cells but not in stromal cells
(Figure 2). Afterwards, we investigated the expression of CXCL13 and CXCR5b mRNA in 11 NB
cell lines and we detected a variable expression of CXCL13 and CXCR5b. Among NB cell lines,
3/11 (27%) express CXCL13 (GI-LI-N, IMR5, SK-N-SH) and 9/11 (82%) express CXCR5b (GI-LI-
on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

12
N, GI-ME-N, IMR-32, IMR5, LA-1-5S, SH-SY5Y, SK-N-BE(2), SK-N-BE(2)c, SK-N-SH). GI-LI-
N, IMR5, SK-N-SH cells express both CXCL13 and CXCR5b and ACN and GI-CA-N cells do not
express either CXCL13 or CXCR5b (data not shown).
CXCR5 and CXCL13 protein expression in NTs and in NB cell lines. To demonstrate that
Schwannian stromal cells of SR tumors express CXCL13 but not CXCR5 we carried out
immunohistochemistry analysis on tumor tissue. As shown in Figure 3C, Schwannian stromal cells
strongly express CXCL13, whereas CXCR5 is not expressed in the same tissue (Figure 3D). Since
CXCR5b mRNA was found variable expressed we employed immunofluorescence technique to
have a better sensibility to detect CXCR5 expression. CXCR5 protein was observed in neuroblastic
cells (Figure 3E and 3F), but not in SR tumors (data not shown).
Next, we analyzed SH-SY5Y and LA1-5S NB cell lines by either immunofluorescence (Figure 3G
and 3H) or immunohistochemistry (Figure 3I and 3L) and we observed a mild surface expression of
CXCR5 receptor in about 20% of SH-SY5Y and 30% of LA1-5S NB cells, respectively (Figure 3G
and 3H). Immunofluorescence analysis also showed some cytoplasmic staining, possibly related to
a CXCR5 internalization. Immunofluorescence analysis was also performed in the other NB cell
lines with similar results and we confirmed CXCR5 expression in SK-N-BE(2), SK-N-BE(2)c and
SK-N-SH cells by FACS analysis (data not shown). Although IMR-5, SK-N-SH and GI-LI-N,
expressed CXCL13 mRNA, only IMR-5 showed CXCL13 cytoplasmatic expression but the
secreted protein was not detected (data not shown).
Induction of NB cells to glial differentiation and CXCL13 production. Human NB cell lines
retain a bi-lineage potential and may differentiate towards a neuroblast or stromal/glial phenotype
on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

13
upon different conditions (19). Since secreted CXCL13 was not detectable by ELISA in the
conditionated medium of any NB cell line (data not shown), we treated SH-SY5Y, SK-N-BE(2)c
and LA1-5S cells with BrdU for 3 weeks to evaluate whether NB cells produce CXCL13 after glial
differentiation induction. Following BrdU treatment most of SH-SY5Y and SK-N-BE(2)c cells
acquired an evident flat, large and more adherent cell morphology resembling Schwannian stromal
cells (Figure 4A). A morphological change was also observed in LA1-5S cells at a less extent. The
induction towards glial cell lineage was shown by increase of S100A6 mRNA expression in all three
cell lines (Figure 4A). In addition, a variable de novo CXCL13 mRNA induction was detected in
NB cell lines reaching a maximum at 3 weeks in BrdU-differentiated SK-N-BE(2)c cells (Figure
4B). The secretion of CXCL13 protein in the culture medium was remarkable in differentiated SK-
N-BE(2)c and LA1-5S (Figure 4C).
Triggering of NB cells migration by CXCL13. We tested whether the expression of CXCR5
could be sufficient to trigger NB cell migration in response to CXCL13 stimulation by chemotaxis
assays. Figure 5A shows that the CXCR5-positive SH-SY5Y and LA1-5S cells significantly
migrated in presence of hrCXCL13 chemokine, while the CXR5-negative ACN cells failed to
migrate (p = not significant, data not shown). LA1-5S cell migration was inhibited by either
blocking the receptor or the chemokine activity, following addition of specific antibodies against
CXCR5 or hrCXCL13 (Figure 5B). LA1-5S cell migration was also observed when the conditioned
medium of NB cells differentiated to the glial cell lineage were used as chemoattractants, thus
further substantiating the possibility that cell-released CXCL13 induces NB cell migration (Figure
5C). This result shows that NB cells induced to glial cell lineage and acquiring a Schwannian-like
on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

14
morphology secretes CXCL13 that stimulates chemotaxis in neuroblasts expressing CXCR5.
Moreover, migration of LA1-5S NB cells was significantly suppressed by the addition of a
neutralizing anti-CXCL13 antibody. On the contrary, CXCR5-negative ACN cells showed a more
limited migration than LA1-5S in response to the conditioned medium of Schwannian-like cells and
their migration was not inhibited by anti-CXCL13 antibody (Figure 5C).
Discussion
Chemokines play several roles in both physiological and pathological conditions. Hereby, we show
that CXCL13 and its receptor CXCR5 are involved in the relation between neuroblastic and
Schwannian stromal cells of NTs, a heterogeneous group of pediatric cancers. Several chemokines
have been found expressed in NB; in particular, the role of the CXCR4-CXCL12 axis in NB cells
proliferation, survival and in bone marrow dissemination is still controversial (13, 14, 22). Airoldi
et al. (15) showed that NB cells express CXCR5 and they hypothesized that malignant cells are able
to migrate even in the bone marrow in response to CXCL13 stimulation.
We confirm that NB cells express CXCR5 receptor and we add a detailed analysis of CXCL13 and
CXCR5 expression in NTs. Our results demonstrate that only malignant neuroblasts express
CXCR5 protein and we show, for the first time, that CXCL13 is also secreted by Schwannian
stromal cells of neuroblastic tumors.
NTs stroma-rich tumors are usually composed of abundant stroma and less amount of neuroblastic
cells; these tumors onset as a localized mass and rarely show metastatic disease. Patients with
stroma-rich tumor are at low risk of relapse after complete surgical tumor resection and they usually
have a good outcome. Since we observed that CXCL13 is expressed by Schwannian stromal cells,
on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

15
and neuroblastic cells express CXCR5 we hypothesize that CXCL13-CXCR5 axis may contribute
to retain malignant CXCR5-positive neuroblasts in stroma-rich tumors limiting their metastatic
spreading. Such hypothesis is strongly supported by the ability of NB cells to migrate towards a
source of hrCXCL13. Moreover, our data indicate that the CXCR5b mRNA variant encodes for a
functionally active CXCR5b protein, which is capable to specifically respond to the CXCL13
stimulus.
The role of Schwannian stromal cells in NTs has been widely debated. Ambros et al. (4) suggested
that these cells secrete factors that limit the proliferation of malignant neuroblasts and Chienski et
al. (7) indicated that Schwannian stromal cells release other factors that modulate angiogenesis.
Here, we demonstrate that CXCL13 is produced by Schwannian stromal cells adding new
information about the factor released by stroma of NTs and we suggest a relevant role of the
CXCL13/CXCR5b axis in the cross-talk between neuroblasts and stromal cells. The novel
discovery that Schwannian stromal cells express CXCL13 supports the previous findings that cells
of neural origin are able to produce CXCL13 (24, 25).
As an in vitro model to test the possible involvement of CXCL13 in the cross-talk between
malignant neuroblasts and Schwannian stromal cells we induced glial cell differentiation of NB
cells. NB cell lines treated with BrdU induce immature neuroblasts to a more mature phenotype
with a large and sub-adherent Schwannian-like morphology (19). After BrdU treatment, NB cells
stop to proliferate and produce calcyclin (23), a glial and Schwannian cells marker. We show for the
first time, that NB cells, which possess a bi-lineage potential (19), express CXCL13 only after
induction of a glial phenotype. Furthermore, we demonstrated that LA1-5S NB cells are able to
on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

16
migrate under the effect of the conditioned medium of BrdU differentiated NB cells and that this
effect is blocked by CXCL13-neutralizing antibodies. Overall data indicate that differentiated
neuroblasts phenotypically similar to Schwannian stromal cell are able to secrete CXCL13 and to
exert a chemotactic effect on CXCR5-positive NB cells, whereas CXCR5-negative NB cells
showed limited migration possible due to a slight effect of other chemotactic factors produced by
Schwannian-like cells. Collectively our results, enforced by the finding that anti-CXCL13 antibody
significantly inhibits the migration of CXCR5-positive NB cells in response to the conditioned
medium, strongly indicate a close relation between Schwannian stromal cells and neuroblastic cells
mediated via a CXCL13-CXCR5 interaction. Hence, in addition to the other factors that exert their
effects on neuroblast survival and differentiation, and on tumor angiogenesis (4, 6, 7), our finding
that Schwannian-like cells are able to secrete a chemotactic factor acting on neuroblastic cell
migration adds insight on a broad role of Schwannian stromal cells in stroma-rich neuroblastic
tumors.
In conclusion, our model suggests that CXCL13 produced by Schwannian stroma may contribute to
limit dissemination of CXCR5b-expressing neuroblasts outside the SR tumors, whereas CXCR5-
positive neuroblastic cells of stroma-poor tumors more easily disseminate to CXCL13-producing
distal sites, such as the bone marrow. Although we cannot exclude the influence of CXCL13
released from distal bone marrow sites on neuroblastona cells, this effect appears unlikely in the
case of localized tumors. Indeed, very low concentrations of CXCL13 are usually present in the
systemic circulation, while the Schwannian stromal cells and malignant neuroblasts are in close
contact thus facilitating local CXCL13-CXCR5 interaction. Obviously, this model cannot be the
on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

17
only reason for the different biological and clinical behavior of SP and SR tumors, which is
predominantly related to the different genetic and molecular characteristics observed in localized
and metastatic tumors (26).
on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

18
Acknowledgements
We are grateful to surgeons, clinicians, pathologists of the Associazione Italiana Ematologia
Oncologia Pediatrica (AIEOP) for providing tumor samples, to Dr. Alessandro De Ambrosis for the
helpful discussions and to Silvia De Luca (Fondazione Italiana per la Lotta al Neuroblastoma, Genoa,
Italy) for language revision. MC and L L are supported by Fondazione Italiana per la Lotta al
Neuroblastoma, FV is a fellow of the University of Genoa. The work was supported by: Associazione
Italiana per la Ricerca sul Cancro (AIRC, Project IG 5331, 2008), Fondazione Italiana per la Lotta al
Neuroblastoma, Ministero dell’Università, Ricerca Scientifica e Tecnologica, Italy.
The authors have no conflicting financial interests.
on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

19
Figures Legends
Figure 1. CXCL13 and CXCR5b expression in tumors.
CXCL13 and CXCR5b expression by RT-qPCR in 26 NTs: 13 SP and 13 SR tumors. In the left
panels the level of CXCL13 and CXCR5 in SP and SR are shown whereas the right panels show the
box plot representation; p value is reported. Most of SR tumors express CXCL13 and very few
express CXCR5; on the contrary, SP tumors express CXCR5 at various amount. CXCL13 is very
low or undetectable expressed in SP tumors. MNE: Mean Normalized Expression.
Figure 2. CXCL13 and CXCR5b expression in isolated neuroblastic and Schwannian stromal
cells.
CXCL13 and CXCR5 expression by RT-qPCR in neuroblastic (Nb) and Schwannian stromal (SS)
cell populations isolated by Laser Capture Microdissection. Isolated SS express CXCL13 that is
almost undetectable in Nb cells. The receptor was found expressed in isolated neuroblastic cells of
2750_Nb sample and at very low levels in the remaining Nb samples but not detected in SS cells.
MNE: Mean Normalized Expression.
Figure 3. CXCL13 and CXCR5 protein expression in tumors and NB cell lines.
CXCR5 and CXCL13 expression in GNB intermixed stroma-rich and NB stroma-poor tumors and
in NB cells detected by immunohistochemistry (Panel A-D and I-L) and immunofluorescence
(Panel E-H). Panel A shows hematoxylin and eosin staining of GNB intermixed stroma-rich; in
panel B Schwannian stromal cells are markedly stained by GFAP (Glial fibrillary acidic protein)
antibody (Cell Marque Inc., Rocklin, CA, USA); the same cells are positive for CXCL13 (panel C)
but negative for CXCR5 (panel D). Detectable CXCR5 protein expression is observed in NB
on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

20
stroma-poor by immunofluorescence (Panel E and F; magnification 40x and 100x). CXCR5
expression is clearly detected in SH-SY-5Y and LA1-5S cells by immunofluorescence (panel G and
H) and immunohistochemistry (I and L). Fluorescent positive signals are evident on the cell
membrane and in the cytoplasm of some cells, indicating the internalization of CXCR5 (panel G
and H). The arrows in panel I and L indicate the CXCR5 positive edge of SH-SY-5Y and LA1-5S
cells. An AxioImager M1 microscope with light and fluorescence lamp (Zeiss Inc., Oberkochen,
Germany) was used for cells and tumor tissue observation. Images were captured by Image capture
system AxioVision Release 4.6.
Figure 4. Effects of treatment with 5’-bromo-2’-deoxyuridine (BrdU) on morphological
features and S100A6 mRNA expression (A) and CXCL13 mRNA expression (B) and detection
of secreted CXCL13 protein (C) in different NB cell lines.
SH-SY5Y, SK-N-BE(2)c and LA1-5S cells belonging to Neuroblastic, Intermediate and Substrate-
adherent NB subtypes, respectively, were grown in presence of 10 μM BrdU in order to induce glial
differentiation and obtain a model for Schwannian-like cell. Panel A shows the effect of BrdU on
morphological features after 3 weeks of treatment: large flattening and subadherent cell shape is
clearly shown in SH-SY5Y and SK-N-BE(2)c cells but less evident in LA1-5S cells. Morphological
changes correlate with the induction of S100A6 (calcyclin) by RT-qPCR in SH-SY5Y, SK-N-
BE(2)c and LA1-5S cells after treatment with 10 μM BrdU for 1, 2 or 3 weeks, indicating the
induction of cells towards glial-cell lineage. A corresponding CXCL13 mRNA expression is
observed in SK-N-BE(2)c and LA1-5S cell lines but not in SH-SY5Y (Panel B). Panel C shows the
amount of secreted CXCL13 protein in cell conditioned media collected for 3 weeks. Detectable
on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

21
secretion of the chemokine in LA1-5S and SK-N-BE(2)c conditioned media is depending on
treatment with BrdU. W: week; MNE: Mean Normalized Expression.
Figure 5. CXCL-13 mediates neuroblastoma cells migration.
Panel A shows the number of migrating SH-SY5Y and LA1-5S NB cells after 6 hours of incubation
at two different doses (100 ng/ml, 200 ng/ml) of hrCXCL13 added in the lower chamber. A low
number of cells migrate in the presence of SFM (serum free medium). Panel B shows that CXCR5
positive LA1-5S migration is specifically mediated by the CXCR5/CXCL13 interaction by
inhibiting both the receptor and the chemokine with anti-CXCR5 (15 ng/ml) and anti-CXCL13 (3.5
ng/ml) blocking antibody, respectively. Panel C shows CXCR5-positive LA1-5S and CXCR5-
negative ACN NB cells chemotaxis in the presence of concentrated serum-free conditioned-medium
of BrdU-differentiated NB cells secreting CXCL13. Migration of LA1-5S cells is significantly
reduced by anti-CXCL13 (0.5 ng/ml) blocking antibody added to the conditioned medium. ACN
cells showed limited migration that was not influenced by anti-CXCL13 antibody. Statistical
analysis was performed by unpaired two-tailed t-test. X-axis shows the different chemoattractants,
y-axis the number of cells migrated/field. ns: not significant.
on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

22
References
1. Maris JM, Hogarty MD, Bagatell R, Cohn SL. Neuroblastoma. Lancet 2007;
369(9579):2106-2120.
2. Shimada H, Ambros IM, Dehner LP, Hata J, Joshi VV, Roald B. Terminology and
morphologic criteria of neuroblastic tumors: recommendations by the International
Neuroblastoma Pathology Committee. Cancer 1999; 86:349-363.
3. Shimada H, Umehara S, Monobe Y, et al. International neuroblastoma pathology
classification for prognostic evaluation of patients with peripheral neuroblastic tumors.
Cancer 2001; 92(9):2451-2461.
4. Ambros IM, Zellner A, Roald B, et al. Role of ploidy, chromosome 1p, and Schwann cells
in the maturation of neuroblastoma. N Engl J Med 1996; 334(23):1505-1511.
5. Coco S, Defferrari R, Scaruffi P, et al. Genome analysis and gene expression profiling of
neuroblastoma and ganglioneuroblastoma reveal differences between neuroblastic and
Schwannian stromal cells. J Pathol 2005; 207(3):346-357.
6. Huang D, Rutkowski JL, Brodeur GM, et al. Schwann cell-conditioned medium inhibits
angiogenesis. Cancer Research 2000; 60:5966-5971.
7. Chienski A, Liu S, Crawford SE, et al. SPARC is a key schwannian-derived inhibitor
controlling neuroblastoma tumor angiogenesis. Cancer Research 2002; 62:7357-7363.
8. Albino D, Scaruffi P, Moretti S, et al. Identification of low intratumoral gene expression
heterogeneity in neuroblastic tumors by genome-wide expression analysis and game theory.
Cancer 2008; 113(6):1412-1422.
on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

23
9. Bonecchi R, Galliera E, Borroni EM, Corsi MM, Locati M, Mantovani A. Chemokines and
chemokine receptors: an overview [review]. Front Biosci 2009; 14:540-551.
10. Balkwill F. Cancer and the chemokine network. Nat Rev Cancer 2004; 4(7):540-550.
11. Mantovani A, Savino , Locati M, Zammataro L, Allavena P, Bonecchi R. The chemokine
system in cancer biology and therapy [review]. Cytokine Growth Factor Rev 2010;21(1):27-
39.
12. Slettenaar VIF, Wilson JL. The chemokine network: a target in cancer biology? Adv Drug
Deliv Rev 2006;58(8):962-74.
13. Gross N, Meier R. Chemokines in neuroectodermal cancers: The crucial growth signal from
the soil. Semin Cancer Biol 2009; 19:103-110.
14. Raffaghello L, Cocco C, Corrias MV, Airoldi I, Pistoia V. Chemokines in neuroectodermal
tumour progression and metastasis. Semin Cancer Biol 2009; 19:97-102.
15. Airoldi I, Cocco C, Morandi F, Prigione I, Pistoia V. CXCR5 may be involved in the
attraction of human metastatic neuroblastoma cells to the bone marrow. Cancer Immunol
Immunother 2008; 57(4):541-548.
16. Meier R, Mühlethaler-Mottet A, Flahaut M, et al. The chemokine receptor CXCR4 strongly
promotes neuroblastoma primary tumour and metastatic growth, but not invasion. PLoS
ONE 2007; 10;2(10):e1016.
17. Lefever S, Hellemans J, Pattyn F, et al. RDML: structured language and reporting guidelines
for real-time quantitative PCR data. Nucleic Acids Res 2009; 37(7):2065-9
on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

24
18. Bustin SA, Benes V, Garson JA, et al. The MIQE Guidelines: Minimum Information for
publication of Quantitative Real-Time PCR experiments. Clin Chem 2009; 55(4):611-622
19. Acosta S, Lavarino C, Paris R, et al. Comprehensive characterization of neuroblastoma cell
lines subtypes reveals bilineage potential similar to neural crest stem cells. BMC Dev Biol.
2009; 9:12.
20. Albini A, Benelli R, Giunciuglio D, et al. Identification of a novel domain of HIV Tat
involved in monocyte chemotaxis. Biol Chem 1998; 273:15895-15900.
21. Barella L, Loetscher M, Tobler A, Baggiolini M, Moser B. Sequence variation of a novel
heptahelical leucocyte receptor through alternative transcript formation. Biochem J 1995;
309:773-779.
22. Airoldi I, Raffaghello L, Piovan E, et al. CXCL12 does not attract CXCR4+ human
metastatic neuroblastoma cells: clinical implications. Clin Cancer Res 2006;12(1):77-82.
23. Tonini GP, Casalaro A, Cara A, Di Martino D. Inducible expression of calcyclin, a gene
with strong homology to S-100 protein, during neuroblastoma cell differentiation and its
prevalent expression in Schwann-like cell lines. Cancer Res. 1991;51(6):1733-1737.
24. Van de Pavert SA, Olivier BJ, Goverse G, Vondenhoff MF, et al. Chemokine CXCL13 is
essential for lymph node initiation and is induced by retinoic acid and neuronal stimulation.
Nat Immunol. 2009;10(11):1193-9.
25. Ramesh G, Borda JT, Gill A, et al. Possible role of glial cells in the onset and progression of
Lyme neuroborreliosis. J Neuroinflammation. 2009;6:23.
26. Maris JM. Recent Advances in Neuroblastoma. N Engl J Med 2010; 362:2202-2211.
on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367
on February 15, 2019. ©
2011 Am
erican Association for C
ancer Research.
mcr.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on June 3, 2011; D
OI: 10.1158/1541-7786.M
CR
-10-0367

on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

on February 15, 2019. ©
2011 Am
erican Association for C
ancer Research.
mcr.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on June 3, 2011; D
OI: 10.1158/1541-7786.M
CR
-10-0367

on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367

Published OnlineFirst June 3, 2011.Mol Cancer Res Federica Del Grosso, Simona Coco, Paola Scaruffi, et al. neuroblastic tumorsneuroblastoma cells and Schwannian stromal cells in Role of CXCL13-CXCR5 crosstalk between malignant
Updated version
10.1158/1541-7786.MCR-10-0367doi:
Access the most recent version of this article at:
Material
Supplementary
http://mcr.aacrjournals.org/content/suppl/2011/06/03/1541-7786.MCR-10-0367.DC1
Access the most recent supplemental material at:
Manuscript
Authoredited. Author manuscripts have been peer reviewed and accepted for publication but have not yet been
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://mcr.aacrjournals.org/content/early/2011/06/03/1541-7786.MCR-10-0367To request permission to re-use all or part of this article, use this link
on February 15, 2019. © 2011 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on June 3, 2011; DOI: 10.1158/1541-7786.MCR-10-0367