opposite regulation of calbindin and calretinin expression by brain-derived neurotrophic factor in...
TRANSCRIPT

Opposite Regulation of Calbindin and Calretinin Expression byBrain-Derived Neurotrophic Factor in Cortical Neurons
Hubert Fiumelli, Maria Kiraly, Andrea Ambrus, Pierre J. Magistretti, and Jean-Luc Martin
Institut de Physiologie, Faculte´ de Medecine, Universite´ de Lausanne, Lausanne, Switzerland
Abstract: Regulation of calbindin and calretinin expres-sion by brain-derived neurotrophic factor (BDNF) wasexamined in primary cultures of cortical neurons usingimmunocytochemistry and northern blot analysis. Herewe report that regulation of calretinin expression byBDNF is in marked contrast to that of calbindin. Indeed,chronic exposure of cultured cortical neurons for 5 daysto increasing concentrations of BDNF (0.1–10 ng/ml) re-sulted in a concentration-dependent decrease in thenumber of calretinin-positive neurons and a concentra-tion-dependent increase in the number of calbindin-im-munoreactive neurons. Consistent with the immunocyto-chemical analysis, BDNF reduced calretinin mRNA levelsand up-regulated calbindin mRNA expression, providingevidence that modifications in gene expression ac-counted for the changes in the number of calretinin- andcalbindin-containing neurons. Among other members ofthe neurotrophin family, neurotrophin-4 (NT-4), whichalso acts by activating tyrosine kinase TrkB receptors,exerted effects comparable to those of BDNF, whereasnerve growth factor (NGF) was ineffective. As for BDNFand NT-4, incubation of cortical neurons with neurotro-phin-3 (NT-3) also led to a decrease in calretinin expres-sion. However, in contrast to BDNF and NT-4, NT-3 didnot affect calbindin expression. Double-labeling experi-ments evidenced that calretinin- and calbindin-contain-ing neurons belong to distinct neuronal subpopulations,suggesting that BDNF and NT-4 exert opposite effectsaccording to the neurochemical phenotype of the targetcell. Key Words: Brain-derived neurotrophic factor—Neurotrophin-3—Neurotrophin-4 —Calretinin—Calbin-din—Cerebral cortex.J. Neurochem. 74, 1870–1877 (2000).
Brain-derived neurotrophic factor (BDNF) is a mem-ber of the neurotrophin family, which also includes nervegrowth factor (NGF), neurotrophin-3 (NT-3), and neu-rotrophin-4/5 (NT-4/5) (Lewin and Barde, 1996). BDNFbinds to and activates the low-affinity p75 receptor aswell as the specific tyrosine kinase receptor (Trk) Bthrough which the neurotrophin exerts its biologicalfunctions (Barbacid, 1994; Bothwell, 1995). There isabundant in vitro and in vivo evidence showing thatBDNF promotes the survival and differentiation of se-lective populations of neurons (Davies, 1994; Lewin and
Barde, 1996). For instance, BDNF acts as a peptidergicdifferentiation factor for GABA-containing neurons as itstimulates the expression of specific neuropeptides suchas somatostatin, substance P, neuropeptide Y, and cho-lecystokinin both in vitro and in vivo (Nawa et al., 1993,1994; Croll et al., 1994). Consistent with these observa-tions, expression of neuropeptide Y is reduced in thecerebral cortex and hippocampus of BDNF knockoutmice (Jones et al., 1994). In addition to its role as apeptidergic differentiation factor, BDNF has been re-ported to regulate the expression of calcium-bindingproteins (Ip et al., 1993; Widmer and Hefti, 1994; Martyet al., 1996; Pappas and Parnavelas, 1997). Thus, BDNFstimulates calbindin expression in cultured hippocampaland cortical neurons (Ip et al., 1993; Widmer and Hefti,1994). Along with this evidence, the expression of cal-bindin as well as of parvalbumin is reduced in the cere-bral cortex and hippocampus of BDNF knockout mice(Jones et al., 1994). In addition to calbindin and parval-bumin, calretinin is the third member of the EF-handfamily of calcium-binding proteins that is expressedthroughout the brain. Although little is known regardingtheir precise functional properties, these three calcium-binding proteins are thought to buffer intracellular freeCa21 (Baimbridge et al., 1992). In the adult rat cerebralcortex, calbindin, parvalbumin, and calretinin are local-ized in three distinct subpopulations of GABAergic neu-rons (Celio, 1990; Resibois and Rogers, 1992), whereasthey are expressed transiently in non-GABAergic neu-rons during development (Fonseca et al., 1995).
Received September 17, 1999; revised manuscript received Decem-ber 31, 1999; accepted January 5, 2000.
Address correspondence and reprint requests to Dr. J.-L. Martin atInstitut de Physiologie, Faculte´ de Medecine, 7 rue du Bugnon, CH-1005 Lausanne, Switzerland. E-mail: [email protected]
Dr. A. Ambrus is on leave from Department of Physiology, AlbertSzent-Gyo¨rgyi Medical University, H-6720 Szeged, Hungary.
Abbreviations used:BDNF, brain-derived neurotrophic factor; NGF,nerve growth factor; NT-3, neurotrophin-3; NT-4/5, neurotrophin-4/5;PBS, phosphate-buffered saline; PBS-T, phosphate-buffered salinewith 0.2% Triton X-100; SSC, saline–sodium citrate; Trk, tyrosinekinase receptor.
1870
Journal of NeurochemistryLippincott Williams & Wilkins, Inc., Philadelphia© 2000 International Society for Neurochemistry

Although regulation of calbindin and parvalbumin ex-pression by BDNF has been examined in the cerebralcortex, there has been no report on the regulation ofcalretinin expression by BDNF. Thus, the aim of thisstudy was to investigate the regulation of calretinin ex-pression by BDNF in cultures of dissociated corticalneurons and to compare this effect with that observed onthe expression of calbindin and parvalbumin. To exam-ine whether the actions of other members of the neuro-trophin family were similar to those of BDNF, we havealso tested the effects of NGF, NT-3, and NT-4 on theexpression of calbindin and calretinin.
Results reported in this study clearly show that regu-lation of calretinin expression by BDNF is in markedcontrast to that of calbindin. Indeed, BDNF reduces theexpression of calretinin mRNA and the number of cal-retinin-positive neurons, although it stimulates the ex-pression of calbindin mRNA and the number of calbin-din-immunoreactive neurons. NT-4 exhibited effectssimilar to those of BDNF, whereas NGF did not affectthe expression of these two calcium-binding proteins. Itis interesting that NT-3 decreased calretinin expression,whereas it did not regulate calbindin expression.
EXPERIMENTAL PROCEDURES
Preparation of primary cultures of mouse corticalneurons
All experiments were carried out in accordance to the Eu-ropean Communities Council Directive regarding care and useof animals for experimental procedures.
Primary cultures of cerebral cortical neurons were preparedfrom 17-day-old Swiss mouse embryos as previously described(Cardinaux et al., 1997; Pellegri et al., 1998; Fiumelli et al.,1999). Cells were plated at a density of 23 105/cm2 on 90-315-mm culture plates (northern blots) or on glass coverslips (12mm in diameter; immunocytochemistry) in Dulbecco’s modi-fied Eagle’s medium supplemented with 1 mM glutamine, 7.5mM sodium bicarbonate, 5 mM HEPES buffer (pH 7.0), 0.1mg/ml streptomycin, and 0.06 mg/ml penicillin. A mixture ofhormones and salt containing 25mg/ml insulin, 100mg/mltransferrin, 60mM putrescine, 20 nM progesterone, and 30 nMsodium selenate was added to the culture medium. Cells weremaintained for 6 days at 37°C in a humidified atmosphere of95% air and 5% CO2. As previously reported (Stella et al.,1995), immunostaining of 6 days in vitro cortical cultures withcell-specific antibodies yields 94% neuron-specific enolase-immunoreactive cells and 99% neurofilament-immunoreactivecells.
Chronic treatment of cultured cortical neurons withneurotrophins
For northern blot experiments, cells were exposed daily to0.1, 1, 3, 5, or 10 ng/ml BDNF, 5 ng/ml NGF, 5 ng/ml NT-3,or 5 ng/ml NT-4 for 5 days starting 1 day after plating. On day6, total RNA was extracted and electrophoresed as describedbelow.
For immunocytochemistry, cultures in separate wells wereexposed daily to 0.1, 0.5, 1, 5, or 10 ng/ml BDNF, 5 ng/mlNGF, 5 ng/ml NT-3, or 5 ng/ml NT-4 for 5 days starting 1 dayafter plating. On day 6, cultures were fixed and subjected toimmunocytochemistry.
Northern blot analysisAt the end of the stimulation, cells were washed twice with
ice-cold phosphate-buffered saline (PBS; pH 7.4), and totalRNA was extracted from cultured cortical neurons using theCsCl centrifugation procedure (Chirgwin et al., 1979). TotalRNA (10 mg) was electrophoresed on a 1.3% agarose/2Mformaldehyde gel and transferred onto a GeneScreen nylonmembrane (NEN Life Science, Zaventem, Belgium). Hybrid-ization was performed overnight at 65°C in 50% formamide,53 saline–sodium citrate (SSC), 50 mM Tris-HCl (pH 7.5),0.1% sodium pyrophosphate, 1% sodium dodecyl sulfate, 0.2%polyvinylpyrrolidone, 0.2% Ficoll, 5 mM EDTA, 0.2% bovineserum albumin, and 150mg/ml salmon sperm DNA with a32P-antisense calbindin, calretinin, or parvalbumin riboprobe.Filters were then washed under high-stringency conditions(twice with 23 SSC containing 0.1% sodium dodecyl sulfate at65°C for 15 min and twice with 0.13 SSC containing 0.1%sodium dodecyl sulfate at 65°C for 30 min) and apposed toautoradiography films (Hyperfilms; Amersham Pharmacia Bio-tech, Dubendorf, Switzerland) at270°C with an intensifyingscreen. Differences in RNA gel loading and blotting wereassessed by rehybridizing the filters with a32P-antisense actinriboprobe (Martin et al., 1995; Pellegri et al., 1998). Hybrid-ization and washing conditions for actin were identical to thosedescribed above. After autoradiography, films were densito-metrically scanned, and results were quantified using theMacintosh-based National Institutes of Health Image program(version 1.61). Densitometric values for calretinin and calbin-din mRNAs were normalized to corresponding actin mRNAvalues. Results are mean6 SEM percentages of control fromthree independent experiments.
32P-antisense riboprobes were generated using either T7 orSP6 RNA polymerase and [a-32P]UTP from a linearizedpGEM-T vector (Promega, Zu¨rich, Switzerland) containing theappropriate cDNA fragment. The 749-bp calbindin cDNA frag-ment was obtained by RT and PCR amplification of total RNAfrom primary cultures of mouse cortical neurons with a setof oligonucleotide primers (59-ATGGCAGAATCCCACCT-GCA-39 and 59-GTTCGGTACAGCTTCCCTCC-39) located at286–305 and 1,015–1,034 bp, respectively, in the coding re-gion of the rat calbindin cDNA sequence (Hunziker andSchrickel, 1988). The 490-bp calretinin cDNA fragment wasobtained by RT and PCR amplification of total RNA fromcultured cortical neurons with a set of oligonucleotide primers(59-TGCTTCAGGCAGCACGTGGG-39 and 59-CAATCTC-CAGGTCCTTTCTG-39) located at 59–78 and 529–548 bp,respectively, in the coding region of the mouse calretinincDNA sequence (Ellis and Rogers, 1993). The 333-bp parval-bumin cDNA fragment was obtained by RT and PCR amplifi-cation of total RNA from adult mouse brain with a set ofoligonucleotide primers (59-ATGTCGATGACAGACGTGCT-39and 59-TTACGTTTCAGCCACCAGAG-39) located at 1–20and 314–333 bp, respectively, in the coding region of themouse parvalbumin cDNA sequence (Zuhlke et al., 1989).
The identity of the amplified calbindin, calretinin, and parv-albumin cDNA fragments was confirmed by sequencing usingan automated DNA sequencer (ALF DNA Analysis System;Pharmacia, Uppsala, Sweden).
ImmunocytochemistryDiaminobenzidine labeling.At the end of the incubation
period, cultures were fixed for 25 min in 4% paraformaldehyde(except for GABA immunostaining, where a mixture of 4%paraformaldehyde and 1% glutaraldehyde was used). Depend-ing on the primary antibody tested, cultures were preincubated
J. Neurochem., Vol. 74, No. 5, 2000
1871REGULATION OF CALBINDIN AND CALRETININ BY BDNF

for 1 h in PBSwith 0.2% Triton X-100 (PBS-T) containingeither 4% normal goat serum (for polyclonal antibodies) or 4%normal horse serum (for monoclonal antibodies). Cultures werethen incubated overnight at room temperature in a solutioncontaining the primary antibody diluted in PBS-T containing4% normal goat or horse serum. The antibodies used were thefollowing: rabbit anti-GABA (Sigma), 1:10,000; rabbit anti-calretinin (Swant, Switzerland), 1:5,000; mouse monoclonalanti-calbindin (Swant), 1:2,000; and mouse monoclonal parv-albumin (Swant), 1:2,000. The next day, cultures were rinsed inPBS-T and incubated for 30 min in the presence of biotinylatedgoat anti-rabbit IgG or biotinylated horse anti-mouse IgG (Vec-tor Laboratories, Burlingame, CA, U.S.A.), both diluted at1:200 in PBS-T. Cultures were then washed in PBS, andstaining was completed using an avidin–biotin–peroxidasecomplex (ABC; Vectastain Elite ABC kits; Vector Laborato-ries). After rinsing in PBS, cultures were incubated for 5–10min in PBS containing 0.05% 3,39-diaminobenzidine and0.002% H2O2. Then, the coverslips were extensively rinsed,dehydrated, and mounted on glass slides with Eukitt. Culturestreated identically, except for replacing the primary antibody bynonimmune serum from the adequate animal, served as controlfor antibody specificity. This treatment yielded nondetectablestaining.
Microscopic examination for quantification was performedunder bright-field illumination using a quadratic counting gridplaced in the ocular of the microscope (Olympus model BH-2).Counts of labeled cells were made using a 253 objective lensin a total population of at least 3,000 cells. The percentage ofimmunoreactive neurons was determined by counting the num-ber of labeled cells and by reporting this value relative to thetotal number of cells. Data represent at least two independentexperiments, each comprising triplicate culture preparations.Results are expressed as percentages of control. Statisticalanalysis was performed using Student’st test.
Double-labeling experiments.For colocalization studies ofGABA with calretinin or calbindin, cells on glass coverslipswere fixed in a mixture of 4% paraformaldehyde and 0.1%glutaraldehyde at room temperature for 25 min. After washingin PBS-T, coverslips were incubated in 4% normal donkeyserum diluted in PBS-T for 50 min. This step was followed byan overnight incubation at room temperature in a cocktail ofprimary antibodies, diluted in PBS-T as follows: cocktail 1,rabbit anti-GABA (Sigma; 1:5,000) and mouse monoclonalanti-calbindin (Swant; 1:1,000); cocktail 2, mouse monoclonalanti-GABA (Chemicon; 1:1,000) and rabbit anti-calretinin(Swant; 1:5,000); and cocktail 3, rabbit anti-calretinin (Swant;1:5,000) and mouse monoclonal anti-calbindin (Swant;1:1,000). The next day, after two 7-min rinses in PBS-T andone 7-min rinse in PBS, coverslips were incubated in thefollowing mixtures of secondary antibodies for 20 min: mixture1, fluorescein isothiocyanate-conjugated anti-mouse IgG andCY3-conjugated anti-rabbit IgG (1:1,000 final concentration ofeach); and mixture 2, fluorescein isothiocyanate-conjugatedanti-rabbit IgG and CY3-conjugated anti-mouse IgG (1:1,000).All secondary antibodies were raised in donkeys (JacksonImmunoResearch Laboratories, West Grove, PA, U.S.A.).Eventually, coverslips were washed and mounted on glassslides with Vectashield (Vector Laboratories).
Stained cells were visualized under a Leitz (Wetzlar, Ger-many) fluorescence microscope. The quantitative analysis ofdouble-labeled neurons was performed by checking at least 100cells on each coverslip. Experiments were performed in tripli-cates.
RESULTS
Effects of BDNF on number of calbindin- andcalretinin-immunoreactive neurons
Regulation of calbindin and calretinin expression byBDNF was examined in primary cultures of mouse cor-
FIG. 1. BDNF increases the number of calbindin-immunoreac-tive neurons. Cultures of cortical neurons were treated for 5 daysin the presence of increasing concentrations of BDNF. A: Immu-nocytochemical detection of calbindin-positive neurons in con-trol (Ctrl) cultures and in cultures treated with 5 ng/ml BDNF. Bar5 20 mm. B: Quantitative analysis of calbindin-immunoreactiveneurons was performed as described in Experimental Proce-dures. Data are mean 6 SEM (bars) percentages of Ctrl valuesfrom three to eight determinations from three independent ex-periments. In Ctrl cultures, the percentage of calbindin-positiveneurons was found to represent 5.85 6 0.4% (n 5 16) of the totalcell population. *p , 0.05, ***p , 0.001 by Student’s t testbetween Ctrl and BDNF-treated cultures.
FIG. 2. BDNF decreases the number of calretinin-immunoreac-tive neurons. Cultures of cortical neurons were treated for 5 daysin the presence of increasing concentrations of BDNF. A: Immu-nocytochemical detection of calretinin-positive neurons in con-trol (Ctrl) cultures and in cultures treated with 5 ng/ml BDNF. Bar5 20 mm. B: Quantitative analysis of calretinin-immunoreactiveneurons was performed as described in Experimental Proce-dures. Data are mean 6 SEM (bars) percentages of Ctrl valuesfrom six to 18 determinations from four independent experi-ments. In Ctrl cultures, the percentage of calretinin-positive neu-rons was found to represent 6.94 6 0.25% (n 5 18) of the totalcell population. ***p , 0.001 by Student’s t test between Ctrl andBDNF-treated cultures.
J. Neurochem., Vol. 74, No. 5, 2000
1872 H. FIUMELLI ET AL.
tical neurons using immunocytochemical staining andnorthern blot analysis. In control cultures, the percentageof calbindin- and calretinin-positive neurons was 5.856 0.4 (n5 16) and 6.946 0.25% (n5 18) of the totalcell population, respectively. Treatment of cultured cor-tical neurons with increasing concentrations of BDNFresulted in a concentration-dependent increase in thenumber of calbindin-positive neurons compared withcontrol cultures (Fig. 1). For instance, the number ofcalbindin-containing neurons almost doubled after stim-ulation with 5 ng/ml BDNF reaching 10.886 0.71% (n5 8) of the total neuronal population.
In contrast to its effect on calbindin expression, BDNFinduced a marked decrease in the number of calretinin-positive neurons (Fig. 2). For instance, daily treatmentwith 5 ng/ml BDNF for 5 days reduced by 60% thenumber of cortical neurons expressing calretinin as thenumber of calretinin-positive cells dropped to 2.686 0.31% (n5 12).
Parvalbumin-positive neurons were detected neither incontrol nor in BDNF-treated cultures.
Considering the striking opposite effects of BDNF onthe expression of these two cytosolic calcium-bindingproteins, it was of importance to determine whether ornot these proteins were localized in the same neuronalpopulation. Double-labeling experiments were carriedout to investigate the degree of colocalization of calbin-din and calretinin. In agreement with previous studies(Celio, 1990; Resibois and Rogers, 1992; Gonchar andBurkhalter, 1997), our results revealed that only 6.26 0.9% (n5 4) of calbindin-containing neurons coex-pressed calretinin. This percentage of colocalization wasnot altered by treatment with BDNF (data not shown),indicating that the opposite regulation of these two cal-cium-binding proteins does not occur within the sameneurons.
Although calbindin and calretinin are distributed inmainly nonoverlapping neuronal populations, they areknown to be colocalized, at least in part, with the inhib-itory neurotransmitter GABA (Rogers, 1992; Kubotaet al., 1994; Gonchar and Burkhalter, 1997; Miettinenet al., 1997). We therefore examined their degree ofcolocalization with GABA with particular emphasis onthe effects exerted by BDNF. Double-labeling analysisrevealed that calbindin was highly colocalized withGABA because 92.16 3.6% (n 5 4) of calbindin-positive neurons expressed GABA, whereas only 37.86 1.5% (n 5 5) of calretinin-positive neurons wereGABA-immunoreactive. After treatment with 5 ng/mlBDNF, the percentage of colocalization of calbindin withGABA was not changed (94.86 1.1%; n5 4), whereasthe percentage of colocalization of GABA with calretininwas significantly increased from 37.86 1.5 to 50.16 1.6% (n5 5, p , 0.001).
Calbindin up-regulation by BDNF was not associatedwith an increase in the number of GABAergic neurons.Indeed, the percentage of GABAergic neurons (11.36 0.6%, n5 6) remained unchanged after exposure to 5ng/ml BDNF (11.86 0.5%, n5 6), which is consistent
with the unaffected expression of GABA in the cerebralcortex and hippocampus of BDNF knockout mice (Joneset al., 1994).
Effects of BDNF on levels of calbindin andcalretinin mRNAs
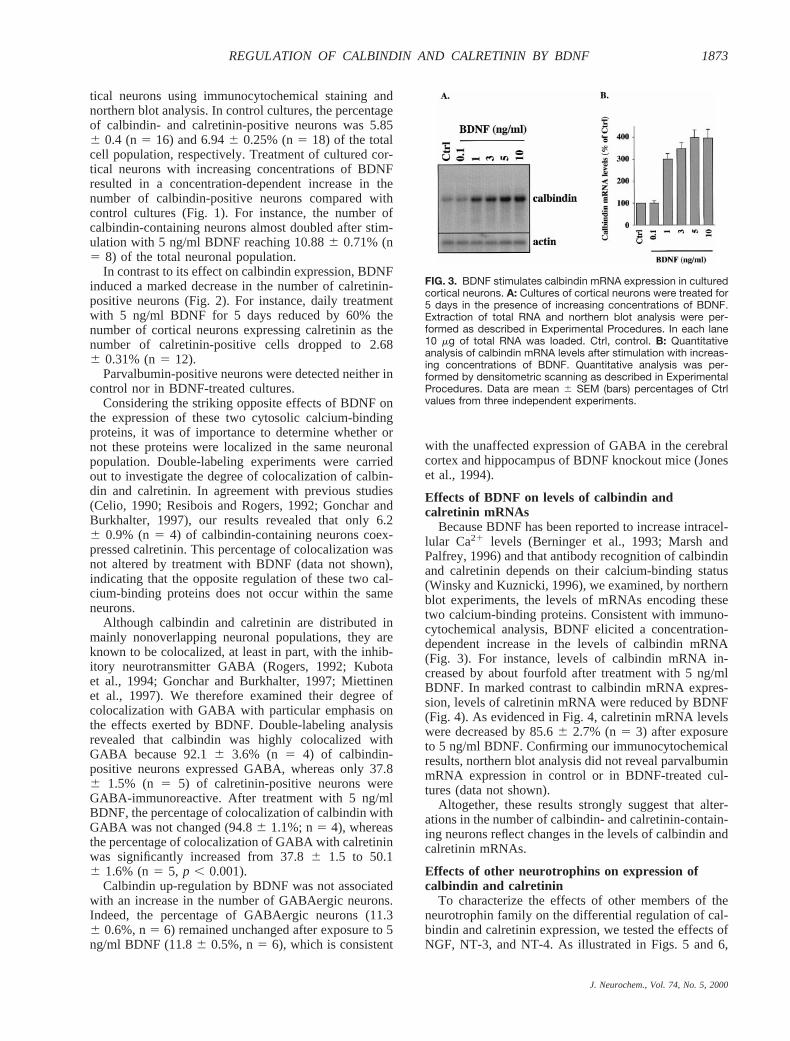
Because BDNF has been reported to increase intracel-lular Ca21 levels (Berninger et al., 1993; Marsh andPalfrey, 1996) and that antibody recognition of calbindinand calretinin depends on their calcium-binding status(Winsky and Kuznicki, 1996), we examined, by northernblot experiments, the levels of mRNAs encoding thesetwo calcium-binding proteins. Consistent with immuno-cytochemical analysis, BDNF elicited a concentration-dependent increase in the levels of calbindin mRNA(Fig. 3). For instance, levels of calbindin mRNA in-creased by about fourfold after treatment with 5 ng/mlBDNF. In marked contrast to calbindin mRNA expres-sion, levels of calretinin mRNA were reduced by BDNF(Fig. 4). As evidenced in Fig. 4, calretinin mRNA levelswere decreased by 85.66 2.7% (n5 3) after exposureto 5 ng/ml BDNF. Confirming our immunocytochemicalresults, northern blot analysis did not reveal parvalbuminmRNA expression in control or in BDNF-treated cul-tures (data not shown).
Altogether, these results strongly suggest that alter-ations in the number of calbindin- and calretinin-contain-ing neurons reflect changes in the levels of calbindin andcalretinin mRNAs.
Effects of other neurotrophins on expression ofcalbindin and calretinin
To characterize the effects of other members of theneurotrophin family on the differential regulation of cal-bindin and calretinin expression, we tested the effects ofNGF, NT-3, and NT-4. As illustrated in Figs. 5 and 6,
FIG. 3. BDNF stimulates calbindin mRNA expression in culturedcortical neurons. A: Cultures of cortical neurons were treated for5 days in the presence of increasing concentrations of BDNF.Extraction of total RNA and northern blot analysis were per-formed as described in Experimental Procedures. In each lane10 mg of total RNA was loaded. Ctrl, control. B: Quantitativeanalysis of calbindin mRNA levels after stimulation with increas-ing concentrations of BDNF. Quantitative analysis was per-formed by densitometric scanning as described in ExperimentalProcedures. Data are mean 6 SEM (bars) percentages of Ctrlvalues from three independent experiments.
J. Neurochem., Vol. 74, No. 5, 2000
1873REGULATION OF CALBINDIN AND CALRETININ BY BDNF

NT-4, which also acts by activating TrkB receptors(Klein et al., 1992), exerted effects comparable to thoseof BDNF. Indeed, NT-4 decreased calretinin mRNAlevels and the number of calretinin-expressing neurons(Fig. 5), whereas it increased the expression of calbindinmRNA and the number of calbindin-immunoreactiveneurons (Fig. 6). In contrast, NGF did not regulate theexpression of these two calcium-binding proteins (Figs. 5and 6). Similarly to BDNF and NT-4, treatment of cul-tured cortical neurons with NT-3 also led to a reductionin calretinin mRNA expression and in the number ofcalretinin-positive neurons (Fig. 5). However, unlikeBDNF and NT-4, NT-3 did not increase calbindinmRNA expression or the number of calbindin-immuno-reactive neurons (Fig. 6).
DISCUSSION
The purpose of the present study was to investigate theregulation of the expression of three calcium-bindingproteins, i.e., calretinin, calbindin, and parvalbumin, byBDNF and by other members of the neurotrophin family.
The main result reported here is an opposite regulationby BDNF and NT-4 of calretinin and calbindin expres-sion in cultured cortical neurons. Indeed, in response tochronic treatment by BDNF or NT-4, the expression ofcalbindin mRNA and the number of calbindin-immuno-reactive neurons are strongly up-regulated (Figs. 1, 3,and 6), whereas the expression of calretinin mRNA andthe number of calretinin-immunoreactive neurons areremarkably decreased (Figs. 2, 4, and 5). Similar treat-ment of cortical neurons with NT-3 also leads to amarked reduction in the levels of calretinin mRNA andin the number of calretinin-positive neurons (Fig. 5).However, in contrast to BDNF and NT-4, NT-3 does notincrease calbindin expression (Fig. 6). The lack of parv-
albumin, the third calcium-binding protein investigatedin this study, may be ascribed to the delayed appearanceof parvalbumin as compared with calretinin and calbin-din in rodent cerebral cortex in vivo. Indeed, the latterappear in the rat cortical anlage at embryonic day 14(Enderlin et al., 1987; Sanchez et al., 1992; Fonsecaet al., 1995), whereas parvalbumin is first detected afterpostnatal week 1 (Solbach and Celio, 1991; Sanchezet al., 1992).
Neurotrophins regulate cell functions by binding toand activating specific Trks. NGF is the specific ligandfor TrkA, whereas BDNF and NT-4 preferentially acti-vate TrkB (Bothwell, 1995). NT-3 binds to TrkC and toa lesser extent to TrkB (Soppet et al., 1991). TrkB andTrkC receptors have been reported to be abundant during
FIG. 5. Effects of neurotrophins on number of calretinin-immu-noreactive neurons and on level of calretinin mRNA. Cultures ofcortical neurons were treated for 5 days in the presence of NGF,BDNF, NT-3, or NT-4 added at a concentration of 5 ng/ml. A:Immunocytochemical detection of calretinin-immunoreactiveneurons and cell counting were performed as described in Ex-perimental Procedures. Data are mean 6 SEM (bars) percent-ages of control (Ctrl) values from six determinations from twoindependent experiments. In Ctrl cultures, the percentage ofcalretinin-positive neurons was found to represent 7.52 6 0.92%(n 5 6) of the total cell population. **p , 0.01 by Student’s t testbetween Ctrl and treated cultures. B: Northern blot analysis ofcalretinin mRNA expression after treatment with NGF, BDNF,NT-3, or NT-4. Extraction of total RNA and northern blot analysiswere performed as described in Experimental Procedures. Ineach lane 10 mg of total RNA was loaded. C: Quantitative anal-ysis of calretinin mRNA levels after stimulation with NGF, BDNF,NT-3, or NT-4. Quantitative analysis was performed by densito-metric scanning as described in Experimental Procedures. Dataare mean 6 SEM (bars) percentages of control from three inde-pendent experiments.
FIG. 4. BDNF inhibits calretinin mRNA expression in culturedcortical neurons. A: Cultures of cortical neurons were treated for5 days in the presence of increasing concentrations of BDNF.Extraction of total RNA and northern blot analysis were per-formed as described in Experimental Procedures. In each lane10 mg of total RNA was loaded. Ctrl, control. B: Quantitativeanalysis of calretinin mRNA levels after stimulation with increas-ing concentrations of BDNF. Quantitative analysis was per-formed by densitometric scanning as described in ExperimentalProcedures. Data are mean 6 SEM (bars) percentages of Ctrlvalues from three independent experiments.
J. Neurochem., Vol. 74, No. 5, 2000
1874 H. FIUMELLI ET AL.

mouse brain embryonic and postnatal development(Klein et al., 1990; Escandon et al., 1994; Lamballeet al., 1994), whereas levels of TrkA were found to below (Yan and Johnson, 1988; Koh and Loy, 1989; Mar-tin-Zanca et al., 1990). Consistent with these in vivostudies, northern blot analysis of Trk receptors in our 6days in vitro cultures has revealed the presence of TrkBand TrkC but not TrkA mRNAs (data not shown), pro-viding an explanation for the lack of response of thispreparation to NGF (Figs. 5 and 6). The similarity be-tween the effects of BDNF and NT-4 on the regulation ofcalbindin and calretinin expression is consistent withtheir actions via TrkB receptors. As to the absence ofcalbindin regulation by NT-3, it may be ascribed to thefact that in our cultures calbindin is largely confined to
GABAergic neurons (92.16 3.5%, n5 4), which do notcolocalize TrkC mRNA, as shown in the rat visual cortex(Gorba and Wahle, 1999).
There is a general agreement that many calbindin- andcalretinin-containing neurons coexpress GABA in ratcerebral cortex. Although the colocalization of calretininwith GABA differs between studies (Rogers, 1992; Gon-char and Burkhalter, 1997; Miettinen et al., 1997), fewerdiscrepancies exist in the degree of overlap betweencalbindin and GABA, which have been repeatedly ob-served to be highly colocalized (Rogers, 1992; Kubotaet al., 1994; Gonchar and Burkhalter, 1997). Our double-labeling experiments reveal a high percentage of cellscoexpressing calbindin and GABA (92.16 3.5%, n5 4). The still important coexpression of GABA withcalbindin after treatment with BDNF (94.86 1.1%, n5 4) indicates that up-regulation of calbindin expressionby BDNF (Figs. 1, 3, and 6) takes place in GABAergicneurons. Thus, after exposure to BDNF, the vast majorityof GABAergic neurons (;90%) also contain calbindin.In marked contrast to calbindin, the ratio of calretinin-positive neurons coexpressing GABA is only 37.86 1.5% (n5 4), suggesting that the majority of calreti-nin-labeled cells correspond to non-GABAergic neurons.In agreement with these observations, calretinin has beenshown to be transiently expressed in certain GABA-negative populations during cortical (Fonseca et al.,1995; Yan et al., 1995) and hippocampal (Jiang andSwann, 1997) ontogenesis. Although TrkB receptors areexpressed on both pyramidal and GABAergic cells (Ca-belli et al., 1996; Cellerino et al., 1996; Gorba andWahle, 1999), the significant increase in the colocaliza-tion of GABA with calretinin after treatment with BDNF(50.16 1.6%; n5 6) indicates that down-regulation ofcalretinin expression by BDNF (Figs. 2, 4, and 5) pref-erentially occurs in non-GABAergic neurons. Thus, ac-cording to the neurochemical phenotype of the targetcell, TrkB receptor activation by BDNF or NT-4 leads toan opposite regulation of calbindin and calretinin viadistinct transduction pathways.
Although calbindin has been previously shown to beup-regulated by BDNF in vivo and in vitro (Ip et al.,1993; Jones et al., 1994; Widmer and Hefti, 1994), this isthe first time that inhibition of calretinin expression byBDNF has been observed. This down-regulation of cal-retinin expression by BDNF may have a physiologicalrelevance during development. Indeed, it has been re-ported that, during the second and third postnatal week,the expression of calretinin decreases sharply to reachadult levels in the rat cerebral cortex (Schierle et al.,1997). It is interesting that studies in the rat visual cortexhave shown that the expression of BDNF mRNA ismarkedly increased during the same period (Castrenet al., 1992; Bozzi et al., 1995; Schoups et al., 1995).Similar decreases in calretinin levels have been reportedin the rat hippocampus after the second postnatal week(Villa et al., 1994; Jiang and Swann, 1997), where BDNFexpression has been shown to reach high levels relativeto earlier stages of development (Maisonpierre et al.,
FIG. 6. Effects of neurotrophins on number of calbindin-immu-noreactive neurons and on level of calbindin mRNA. Cultures ofcortical neurons were treated for 5 days in the presence of NGF,BDNF, NT-3, or NT-4 added at a concentration of 5 ng/ml. A:Immunocytochemical detection of calbindin-immunoreactiveneurons and cell counting were performed as described in Ex-perimental Procedures. Data are mean 6 SEM (bars) percent-ages of control (Ctrl) values from six determinations from twoindependent experiments. In Ctrl cultures the percentage ofcalbindin-positive neurons was found to represent 6.49 6 0.72%(n 5 6) of the total cell population. **p , 0.01 by Student’s t testbetween Ctrl and treated cultures. B: Northern blot analysis ofcalbindin mRNA expression after treatment with NGF, BDNF,NT-3, or NT-4. Extraction of total RNA and northern blot analysiswere performed as described in Experimental Procedures. Ineach lane 10 mg of total RNA was loaded. C: Quantitative anal-ysis of calbindin mRNA levels after stimulation with NGF, BDNF,NT-3, or NT-4. Quantitative analysis was performed by densito-metric scanning as described in Experimental Procedures. Dataare mean 6 SEM (bars) percentages of Ctrl values from threeindependent experiments.
J. Neurochem., Vol. 74, No. 5, 2000
1875REGULATION OF CALBINDIN AND CALRETININ BY BDNF

1990; Friedman et al., 1991). The temporal correlationbetween the appearance of BDNF and the disappearanceof calretinin, together with our observations, stronglysuggests that the ontogenetic down-regulation of calreti-nin observed in the cerebral cortex in vivo may resultfrom a BDNF-mediated mechanism. However, a definitelink between BDNF and calretinin down-regulation invivo awaits further experiments using BDNF knockoutmice.
Acknowledgment: The authors wish to thank Igor Allamanfor expert advice on northern blot analysis. This work wassupported by the Swiss National Science Foundation withgrants 3100-050806.97 to J.-L.M. and 3100-40565.94 to P.J.M.
REFERENCES
Baimbridge K. G., Celio M. R., and Rogers J. H. (1992) Calcium-binding proteins in the nervous system.Trends Neurosci.15,303–308.
Barbacid M. (1994) The Trk family of neurotrophin receptors.J. Neu-robiol. 25, 1386–1403.
Berninger B., Garcia D. E., Inagaki N., Hahnel C., and Lindholm D.(1993) BDNF and NT-3 induce intracellular Ca21 elevation inhippocampal neurones.Neuroreport4, 1303–1306.
Bothwell M. (1995) Functional interactions of neurotrophins and neu-rotrophin receptors.Annu. Rev. Neurosci.18, 223–253.
Bozzi Y., Pizzorusso T., Cremisi F., Rossi F. M., Barsacchi G., andMaffei L. (1995) Monocular deprivation decreases the expressionof messenger RNA for brain-derived neurotrophic factor in the ratvisual cortex.Neuroscience69, 1133–1144.
Cabelli R. J., Allendoerfer K. L., Radeke M. J., Welcher A. A.,Feinstein S. C., and Shatz C. J. (1996) Changing patterns ofexpression and subcellular localization of TrkB in the developingvisual system.J. Neurosci.16, 7965–7980.
Cardinaux J. R., Magistretti P. J., and Martin J. L. (1997) Brain-derivedneurotrophic factor stimulates phosphorylation of stathmin in cor-tical neurons.Brain Res. Mol. Brain Res.51, 220–228.
Castren E., Zafra F., Thoenen H., and Lindholm D. (1992) Lightregulates expression of brain-derived neurotrophic factor mRNAin rat visual cortex.Proc. Natl. Acad. Sci. USA89, 9444–9448.
Celio M. R. (1990) Calbindin D-28k and parvalbumin in the ratnervous system.Neuroscience35, 375–475.
Cellerino A., Maffei L., and Domenici L. (1996) The distribution ofbrain-derived neurotrophic factor and its receptor trkB in parval-bumin-containing neurons of the rat visual cortex.Eur. J. Neuro-sci. 8, 1190–1197.
Chirgwin J. M., Przybyla A. E., MacDonald R. J., and Rutter W. J.(1979) Isolation of biologically active ribonucleic acid fromsources enriched in ribonuclease.Biochemistry18, 5294–5299.
Croll S. D., Wiegand S. J., Anderson K. D., Lindsay R. M., and NawaH. (1994) Regulation of neuropeptides in adult rat forebrain by theneurotrophins BDNF and NGF.Eur. J. Neurosci.6, 1343–1353.
Davies A. M. (1994) The role of neurotrophins in the developingnervous system.J. Neurobiol.25, 1334–1348.
Ellis J. and Rogers J. (1993) Design and specificity of hammerheadribozymes against calretinin mRNA.Nucleic Acids Res.21,5171–5178.
Enderlin S., Norman A. W., and Celio M. R. (1987) Ontogeny of thecalcium binding protein calbindin D-28k in the rat nervous sys-tem.Anat. Embryol. (Berl.)177,15–28.
Escandon E., Soppet D., Rosenthal A., Mendoza-Ramirez J. L., SzonyiE., Burton L. E., Henderson C. E., Parada L. F., and Nikolics K.(1994) Regulation of neurotrophin receptor expression duringembryonic and postnatal development.J. Neurosci.14, 2054–2068.
Fiumelli H., Jabaudon D., Magistretti P. J., and Martin J. L. (1999)BDNF stimulates expression, activity and release of tissue-type
plasminogen activator in mouse cortical neurons.Eur. J. Neurosci.11, 1639–1646.
Fonseca M., del Rio J. A., Martinez A., Gomez S., and Soriano E.(1995) Development of calretinin immunoreactivity in the neo-cortex of the rat.J. Comp. Neurol.361,177–192.
Friedman W. J., Olson L., and Persson H. (1991) Cells that expressbrain-derived neurotrophic factor mRNA in the developing post-natal rat brain.Eur. J. Neurosci.3, 688–697.
Gonchar Y. and Burkhalter A. (1997) Three distinct families ofGABAergic neurons in rat visual cortex.Cereb. Cortex7, 347–358.
Gorba T. and Wahle P. (1999) Expression of TrkB and TrkC but notBDNF mRNA in neurochemically identified interneurons in ratvisual cortex in vivo and in organotypic cultures.Eur. J. Neurosci.11, 1179–1190.
Hunziker W. and Schrickel S. (1988) Rat brain calbindin D28: sixdomain structure and extensive amino acid homology withchicken calbindin D28.Mol. Endocrinol.2, 465–473.
Ip N. Y., Li Y., Yancopoulos G. D., and Lindsay R. M. (1993) Culturedhippocampal neurons show responses to BDNF, NT-3, and NT-4,but not NGF.J. Neurosci.13, 3394–3405.
Jiang M. and Swann J. W. (1997) Expression of calretinin in diverseneuronal populations during development of rat hippocampus.Neuroscience81, 1137–1154.
Jones K. R., Farinas I., Backus C., and Reichardt L. F. (1994) Targeteddisruption of the BDNF gene perturbs brain and sensory neurondevelopment but not motor neuron development.Cell 76, 989–999.
Klein R., Martin-Zanca D., Barbacid M., and Parada L. F. (1990)Expression of the tyrosine kinase receptor gene trkB is confined tothe murine embryonic and adult nervous system.Development109,845–850.
Klein R., Lamballe F., Bryant S., and Barbacid M. (1992) The trkBtyrosine protein kinase is a receptor for neurotrophin-4.Neuron8,947–956.
Koh S. and Loy R. (1989) Localization and development of nervegrowth factor-sensitive rat basal forebrain neurons and their af-ferent projections to hippocampus and neocortex.J. Neurosci.9,2999–3018.
Kubota Y., Hattori R., and Yui Y. (1994) Three distinct subpopulationsof GABAergic neurons in rat frontal agranular cortex.Brain Res.649,159–173.
Lamballe F., Smeyne R. J., and Barbacid M. (1994) Developmentalexpression of trkC, the neurotrophin-3 receptor, in the mammaliannervous system.J. Neurosci.14, 14–28.
Lewin G. R. and Barde Y. A. (1996) Physiology of the neurotrophins.Annu. Rev. Neurosci.19, 289–317.
Maisonpierre P. C., Belluscio L., Friedman B., Alderson R. F., Wie-gand S. J., Furth M. E., Lindsay R. M., and Yancopoulos G. D.(1990) NT-3, BDNF, and NGF in the developing rat nervoussystem: parallel as well as reciprocal patterns of expression.Neu-ron 5, 501–509.
Marsh H. N. and Palfrey H. C. (1996) Neurotrophin-3 and brain-derived neurotrophic factor activate multiple signal transductionevents but are not survival factors for hippocampal pyramidalneurons.J. Neurochem.67, 952–963.
Martin J.-L., Gasser D., and Magistretti P. J. (1995) Vasoactive intes-tinal peptide and pituitary adenylate cyclase–activating polypep-tide potentiate c-fos expression induced by glutamate in culturedcortical neurons.J. Neurochem.65, 1–9.
Martin-Zanca D., Barbacid M., and Parada L. F. (1990) Expression ofthe trk proto-oncogene is restricted to the sensory cranial andspinal ganglia of neural crest origin in mouse development.GenesDev.4, 683–694.
Marty S., Berninger B., Carroll P., and Thoenen H. (1996) GABAergicstimulation regulates the phenotype of hippocampal interneuronsthrough the regulation of brain-derived neurotrophic factor.Neu-ron 16, 565–570.
Miettinen M., Pitkanen A., and Miettinen R. (1997) Distribution ofcalretinin-immunoreactivity in the rat entorhinal cortex: coexist-ence with GABA.J. Comp. Neurol.378,363–378.
J. Neurochem., Vol. 74, No. 5, 2000
1876 H. FIUMELLI ET AL.

Nawa H., Bessho Y., Carnahan J., Nakanishi S., and Mizuno K. (1993)Regulation of neuropeptide expression in cultured cerebral corti-cal neurons by brain-derived neurotrophic factor.J. Neurochem.60, 772–775.
Nawa H., Pelleymounter M. A., and Carnahan J. (1994) Intraventric-ular administration of BDNF increases neuropeptide expression innewborn rat brain.J. Neurosci.14, 3751–3765.
Pappas I. S. and Parnavelas J. G. (1997) Neurotrophins and basicfibroblast growth factor induce the differentiation of calbindin-containing neurons in the cerebral cortex.Exp. Neurol.144,302–314.
Pellegri G., Magistretti P. J., and Martin J. L. (1998) VIP and PACAPpotentiate the action of glutamate on BDNF expression in mousecortical neurones.Eur. J. Neurosci.10, 272–280.
Resibois A. and Rogers J. H. (1992) Calretinin in rat brain: an immu-nohistochemical study.Neuroscience46, 101–134.
Rogers J. H. (1992) Immunohistochemical markers in rat cortex: co-localization of calretinin and calbindin-D28k with neuropeptidesand GABA.Brain Res.587,147–157.
Sanchez M. P., Frassoni C., Alvarez-Bolado G., Spreafico R., andFairen A. (1992) Distribution of calbindin and parvalbuminin the developing somatosensory cortex and its primordium inthe rat: an immunocytochemical study.J. Neurocytol.21, 717–736.
Schierle G. S., Gander J. C., D’Orlando C., Ceilo M. R., and VogtW. D. (1997) Calretinin-immunoreactivity during postnatal devel-opment of the rat isocortex: a qualitative and quantitative study.Cereb. Cortex7, 130–142.
Schoups A. A., Elliott R. C., Friedman W. J., and Black I. B. (1995)NGF and BDNF are differentially modulated by visual experiencein the developing geniculocortical pathway.Brain Res. Dev. BrainRes.86, 326–334.
Solbach S. and Celio M. R. (1991) Ontogeny of the calcium bindingprotein parvalbumin in the rat nervous system.Anat. Embryol.(Berl.) 184,103–124.
Soppet D., Escandon E., Maragos J., Middlemas D. S., Reid S. W.,Blair J., Burton L. E., Stanton B. R., Kaplan D. R., Hunter T.,Nikolics K., and Parada L. F. (1991) The neurotrophic factorsbrain-derived neurotrophic factor and neurotrophin-3 are ligandsfor the trkB tyrosine kinase receptor.Cell 65, 895–903.
Stella N., Pellerin L., and Magistretti P. J. (1995) Modulation of theglutamate-evoked release of arachidonic acid from mouse corticalneurons: involvement of a pH-sensitive membrane phospholipaseA2. J. Neurosci.15, 3307–3317.
Villa A., Podini P., Panzeri M. C., Racchetti G., and Meldolesi J.(1994) Cytosolic Ca21 binding proteins during rat brain ageing:loss of calbindin and calretinin in the hippocampus, with nochange in the cerebellum.Eur. J. Neurosci.6, 1491–1499.
Widmer H. R. and Hefti F. (1994) Stimulation of GABAergic neurondifferentiation by NT-4/5 in cultures of rat cerebral cortex.BrainRes. Dev. Brain Res.80, 279–284.
Winsky L. and Kuznicki J. (1996) Antibody recognition of calcium-binding proteins depends on their calcium-binding status.J. Neu-rochem.66, 764–771.
Yan Q. and Johnson E. M. J. (1988) An immunohistochemical study ofthe nerve growth factor receptor in developing rats.J. Neurosci.8,3481–3498.
Yan Y. H., van Brederode J. F., and Hendrickson A. E. (1995) Devel-opmental changes in calretinin expression in GABAergic andnonGABAergic neurons in monkey striate cortex.J. Comp. Neu-rol. 363,78–92.
Zuhlke C., Schoffl F., Jockusch H., Simon D., and Guenet J. L. (1989)cDNA sequence and chromosomal localization of the mouse parv-albumin gene, Pva.Genet. Res.54, 37–43.
J. Neurochem., Vol. 74, No. 5, 2000
1877REGULATION OF CALBINDIN AND CALRETININ BY BDNF