open research onlinelibeprints.open.ac.uk/49151/59/49151.pdfcognition and behavior accumbal...
TRANSCRIPT

Open Research OnlineThe Open University’s repository of research publicationsand other research outputs
Accumbal cholinergic interneurons differentiallyinfluence motivation related to satiety signalingJournal Item
How to cite:
Aitta-aho, Teemu; Phillips, Benjamin U.; Pappa, Elpiniki; Hay, Y. Audrey; Harnischfeger, Fiona; Heath, ChristopherJ.; Saksida, Lisa M.; Bussey, Timothy J. and Apergis-Schoute, John (2017). Accumbal cholinergic interneuronsdifferentially influence motivation related to satiety signaling. eNeuro, 4(2)
For guidance on citations see FAQs.
c© 2017 The Authors
Version: Version of Record
Link(s) to article on publisher’s website:http://dx.doi.org/doi:10.1523/ENEURO.0328-16.2017
Copyright and Moral Rights for the articles on this site are retained by the individual authors and/or other copyrightowners. For more information on Open Research Online’s data policy on reuse of materials please consult the policiespage.
oro.open.ac.uk

Cognition and Behavior
Accumbal Cholinergic Interneurons DifferentiallyInfluence Motivation Related to Satiety SignalingTeemu Aitta-aho,1,2 Benjamin U. Phillips,3 Elpiniki Pappa,1 Y. Audrey Hay,4 Fiona Harnischfeger,1
Christopher J. Heath,3,5 Lisa M. Saksida,3,6,7 Tim J. Bussey,3,6,7 and John Apergis-Schoute1,8
DOI:http://dx.doi.org/10.1523/ENEURO.0328-16.2017
1Department of Pharmacology, University of Cambridge, Cambridge CB2 1PD, UK, 2Department of Pharmacology Faculty ofMedicine, University of Helsinki, Helsinki 00014, Finland, 3Department of Psychology and Behavioural and Clinical NeuroscienceInstitute, University of Cambridge, Cambridge, CB2 3EB, UK, 4Department of Physiology, Development, and Neuroscience,University of Cambridge, Cambridge CB2 3DY, UK, 5School of Life Health and Chemical Sciences, the Open University, WaltonHall, Milton Keynes MK7 6AA, UK, 6Molecular Medicine Research Laboratories, Robarts Research Institute and Department ofPhysiology and Pharmacology, Schulich School of Medicine and Dentistry, Western University, London, ON, Canada, 7Brainand Mind Institute, Western University, London, ON, Canada, and 8Department of Neuroscience, Psychology, and Behaviour,University of Leicester, Leicester LE1 7RH, UK
AbstractSatiety, rather than all or none, can instead be viewed as a cumulative decrease in the drive to eat that developsover the course of a meal. The nucleus accumbens (NAc) is known to play a critical role in this type of valuereappraisal, but the underlying circuits that influence such processes are unclear. Although NAc cholinergicinterneurons (CINs) comprise only a small proportion of NAc neurons, their local impact on reward-basedprocesses provides a candidate cell population for investigating the neural underpinnings of satiety. The presentresearch therefore aimed to determine the role of NAc-CINs in motivation for food reinforcers in relation to satietysignaling. Through bidirectional control of CIN activity in mice, we show that when motivated by food restriction,increasing CIN activity led to a reduction in palatable food consumption while reducing CIN excitability enhancedfood intake. These activity-dependent changes developed only late in the session and were unlikely to be drivenby the innate reinforcer strength, suggesting that CIN modulation was instead impacting the cumulative changein motivation underlying satiety signaling. We propose that on a circuit level, an overall increase in inhibitory toneonto NAc output neurons played a role in the behavioral results, as activating NAc-CINs led to an inhibition ofmedium spiny neurons that was dependent on nicotinic receptor activation. Our results reveal an important rolefor NAc-CINs in controlling motivation for food intake and additionally provide a circuit-level framework forinvestigating the endogenous cholinergic circuits that signal satiety.
Received October 28, 2016; accepted March 30, 2017; First published April21, 2017.Authors report no conflict of interest.
Author Contributions: T.A.-a., B.U.P., C.J.H., L.M.S., T.B., and J.A.-S. de-signed research; T.A.-a., B.U.P., E.P., Y.A.H., F.H., C.J.H., and J.A.-S. per-formed research; T.A.-a., B.U.P., E.P., Y.A.H., F.H., and J.A.-S. analyzed data;
Significance Statement
The decrease in the drive to eat is not all or none but instead develops over the course of a meal, where witheach bite the incentive value of food is reduced, eventually resulting in the state of being sated. Suchreappraisal of value is a process that has been strongly attributed to nucleus accumbens function andprovides a motivation-based framework for investigating the neural underpinnings of satiety. Nicotine, byacting on central nicotinic acetylcholine receptors, is well know to be a potent anorectic; neverthelessprecisely how cholinergic brain circuits regulate appetite is poorly understood. The aim of this study wastherefore to determine the endogenous cholinergic circuits that underlie satiety signaling.
New Research
March/April 2017, 4(2) e0328-16.2017 1–16

IntroductionThe need for maintaining energy homeostasis has re-
sulted in the intricate wiring between specialized braincircuits for guiding food-seeking behavior. The nucleusaccumbens (NAc), located in the ventral region of thestriatum, is strategically located for integrating informa-tion relevant to such behavior, as it receives significantinputs from energy-sensing brain regions and ones thatencode for the value of environmental stimuli (Phillipsonand Griffiths, 1985; McDonald, 1991; Brog et al., 1993;Trivedi et al., 1998). By virtue of its output to motorsystems, the NAc is at an interface between these regionsand those necessary for generating complex actions im-portant for survival (Jones and Mogenson, 1980; Mogen-son et al., 1980).
Although comprising � 3% of striatal neurons (Bolamet al., 1984; Phelps et al., 1985; Contant et al., 1996; Zhouet al., 2002), cholinergic interneurons (CINs) are known toform a dense plexus of local innervation (Bolam et al.,1984; Kawaguchi, 1993; Contant et al., 1996; Descarriesand Mechawar, 2000). This small but far-reaching NAcpopulation is known to be critical for NAc function (Hoebelet al., 2007; Williams and Adinoff, 2008; Mark et al., 2011).For example, disruptions in NAc-CIN functioning havebeen linked to a number of psychiatric conditions includ-ing depression (Chau et al., 2001; Warner-Schmidt et al.,2011), addiction (Hikida et al., 2001; Itzhak and Martin,2002; Williams and Adinoff, 2008), and related withdrawalsymptomology (Hikida et al., 2003; Avena et al., 2008),highlighting the importance of this NAc neuronal popula-tion in various reward-based cognitive processes.
One theory of CIN function posits that by reducing theincentive value of rewarding stimuli, locally released NAcacetylcholine (ACh) can act as a motivational stop signal(Hoebel et al., 2007). Support for this idea comes fromdata demonstrating a behavioral link between increasedNAc cholinergic transmission and reduced seeking be-havior for drugs (Hikida et al., 2001, 2003; Zhou et al.,2007) and natural rewards (Rada et al., 2005; Avena et al.,2006). The decreased desire for food associated withsatiety is not typically an all-or-none process but insteaddevelops cumulatively over the course of a meal. Thegradual change in consumption behavior occurs through
an iterative devaluation of the incentive value of food.Such reappraisal of value is a process that has beenstrongly attributed to NAc function (Corbit et al., 2001;Cardinal et al., 2002; Singh et al., 2010; Mannella et al.,2013) and provides a motivation-based framework forinvestigating the neural underpinnings of satiety.
Support for NAc-ACh as a satiety signal comes frommicrodialysis studies demonstrating that NAc-ACh reachesmaximum levels toward the end of a meal and stronglycorrelates with a decrease in food-seeking and consump-tion behavior (Mark et al., 1992; Rada et al., 2005; Avenaet al., 2008). The physiologic impact of this local rise inACh on food intake is unclear however, as pharmacolog-ical disruptions in NAc-ACh transmission have resulted inconflicting behavioral results (Pratt and Kelley, 2004; Willet al., 2006; Perry et al., 2009; Pratt and Blackstone,2009). Moreover, selective NAc-CIN lesions both increasefood intake over the course of days and decrease theamount consumed after a 24-h fast (Hajnal et al., 2000). Inaddition, the heterogeneous pre- and postsynaptic distri-bution of cholinergic receptor subtypes and anatomicallydistinct ACh populations projecting to the NAc (Woolf andButcher, 1981; Dautan et al., 2014) have made it difficultto pinpoint the importance of NAc-CIN activity in con-trolling food intake. These challenges highlight the lim-itations of traditional pharmacological approaches inlinking functionally specialized cholinergic circuits withadaptive behavioral responses. To further characterize therelationship between NAc-CIN activity and satiety-relatedchanges in motivation, the present study used designerreceptor (DREADD) technology targeted to NAc-CINs forcontrolling their activity while mice seek out and consumefood. Our results provide further support for NAc-CINs play-ing an important role in controlling food intake and offer apotential mechanism by which NAc-CINs may act to signalsatiety through the inhibition of NAc output.
MethodsExperimental animals
All animal procedures were performed in accordancewith the UK Animals (Scientific Procedures) Act of 1986.Male heterozygous mice expressing Cre recombinase un-der the control of the ChAT promoter [ChAT::cre mice(B6N;129S6-Chattm2(cre)Lowl/J); The Jackson Laboratory]were used in the experiments. Mice were housed 2–10animals per polycarbonate cage and provided ad libitumwith water and standard lab diet (RM3, Special Diet Ser-vices, Essex, UK) in a holding room maintained under a12-h light cycle (lights off at 7 p.m.) with temperatureregulated at 22–24°C and relative humidity kept at 50–55%. Mice were genotyped using PCR from ear notchbiopsy. Before behavioral testing, the animals werehandled daily for 1 wk. One hour before each behavioralsession, the mice were transported to the respectivetesting area.
Stereotactic injectionsChAT::cre mice 2–3 mo of age were anaesthetized with
isoflurane (5% induction, 1–2% maintenance; Abbott)mixed with oxygen (flow rate 0.8–1.0 l/min) and placed in
L.M.S. and T.B. contributed unpublished reagents/analytic tools; J.A.-S. wrotethe paper.
Royal Society, Sigrid Jusélius Stiftelse, Erasmus, Herchel Smith FellowshipAcknowledgments: We thank members of the Department of Psychology for
intellectual input regarding rodent behavioral assays of motivation, AnnemiekeApergis-Schoute for assisting with analyses, Denis Burdakov for providingexperimental resources, and Bryan Roth for making designer receptors avail-able to the scientific community. We also gratefully thank the Royal Society(J.A.S.), the Sigrid Juselius Foundation (T.A.), Herchel Smith Fellowship(Y.A.H), and the Erasmus Program (E.P.) for their support.
Correspondence should be addressed to John Apergis-Schoute, Depart-ment of Neuroscience, Psychology & Behaviour, University of Leicester, Uni-versity Road, Leicester LE1 7RH. E-mail: [email protected].
DOI:http://dx.doi.org/10.1523/ENEURO.0328-16.2017Copyright © 2017 Aitta-aho et al.This is an open-access article distributed under the terms of the CreativeCommons Attribution 4.0 International license, which permits unrestricted use,distribution and reproduction in any medium provided that the original work isproperly attributed.
New Research 2 of 16
March/April 2017, 4(2) e0328-16.2017 eNeuro.org

a stereotactic frame (David Kopf Instruments), the skullwas exposed via a small incision, and a small bilateralcraniotomy was performed to allow intracranial injections.A stainless steel beveled microinjector was lowered to acoordinate aimed at the NAc (anteroposterior �1.30 mmin relation to bregma, laterally �0.70 mm in relation tomidline, and –4.2 mm and –4.5 mm deep from the skulllevel). The microinjector was connected to a 1-�l Hamil-ton glass syringe via polyethylene tubing, and an injectionrate of 0.1 �l/min was regulated by a microprocessor-controlled programmable syringe pump (KD Scientific).Starting from the ventral site, each injection site received150-nl volume of one of the following viruses: AAV2-hSyn-DIO-mCherry, AAV2-hSyn-DIO-rM3D(Gs)-mCherry, AAV2-hSyn-DIO-hM3D(Gq)-mCherry, or AAV2-hSyn-DIO-hM4D(Gi)-mCherry (titer 1–5 � 1012 vg/mL, Gene Therapy Center,University of North Carolina School of Medicine, Chapel Hill,NC) followed by a 2-min wait, except for the dorsal site, forwhich the waiting time was extended to 4 min. For postop-erative care, mice received meloxicam (1 mg/kg s.c.; Boehr-inger Ingelheim), and a recovery period of 5 wks was allowedbefore behavioral testing.
ImmunohistochemistryTo determine the specificity of receptors expression in
CINs, mice were anaesthetized with pentobarbital (500mg/kg, intra-peritonally (i.p.); Vetoquinol) and transcardi-ally perfused first with 0.1 M PBS followed by 10% neutral-buffered formalin (Sigma-Aldrich). Brains were removed,postfixed overnight at 4°C, and cryoprotected at 4°C with30% w/v sucrose in PBS until the brains sank and werecompletely submerged. Coronal sections (30 �m) werecut on a freezing sliding microtome (model 860; AmericanOptical Company). For both mCherry and VAChT immu-nostaining, sections were washed at room temperature(RT) in 0.1 M PBS, blocked with 1% BSA (Thermo FisherScientific) supplemented with 0.3% Triton X-100 (ThermoFisher Scientific) in 0.1 M PBS. Sections were then incu-bated overnight at RT in primary antibodies diluted inblocking buffer, washed in PBS, incubated in secondaryantibodies for 2 h at RT, washed in PBS, mounted onmicroscope slides, and coverslipped. Primary antibodieswere rabbit anti-mCherry (1:1000, ab167453; Abcam) andguinea pig anti-VAChT (1:500, AB1588; EMD Millipore).Secondary antibodies were donkey anti-rabbit Alexa Fluor594 (1:1000; Abcam) and goat anti–guinea-pig AlexaFluor 488 (1:1000; Abcam). Digital images were capturedwith a Zeiss Axioskop 2 microscope (Zeiss) and QImagingQICAM Fast digital camera (QImaging). Images weremerged using ImageJ (National Institutes of Health).
ElectrophysiologyCoronal slices were made �9 wks postinjection. 250-
mm-thick slices were cut with a Leica VT 1200S vi-bratome in ice-cold artificial CSF (ACSF; see below) andallowed to recover for 1 h at 35°C in ACSF before record-ings. Patch pipettes were manufactured from borosilicateglass, and their tip resistances were 4–6 M� when filledwith K-gluconate solution (see below). Whole-cell record-ings were conducted at 37°C using an EPC-10 amplifierand Patch-Master software (HEKA Elektronik). Only cells
with access resistances of �20 M� were used for anal-ysis. Current signals were low-pass filtered at 3 kHz anddigitized at 10 kHz. Data were analyzed using Axograph,Patch-Master, and Igor Pro software. Whole-cell record-ings were performed at 35°C using an EPC-10 amplifierand Patch-Master software (HEKA Elektronik). ChAT-containing cells were visualized in acute living brain slicesusing a GFP filter set (Chroma). Clozapine-N-oxide (CNO;Sequoia Research Products) was prepared in ACSF andbath-applied at a concentration of 10 �M.
Chemicals and solutionsSlice-cutting and recording ACSF was gassed with
95% O2 and 5% CO2, and contained the following (in mM):NaCl 125, NaHCO3 25, KCl 3, NaH2PO4 1.25, CaCl2 1(cutting)/2 (recording), MgCl2 6 (cutting)/1 (recording), so-dium pyruvate 3, and glucose 25 (cutting)/5 (recording).Pipettes were filled with (in mM): potassium gluconate135, NaCl 7, Hepes 10, Na2-ATP 2, Na-GTP 0.3, andMgCl2 2; pH was adjusted to 7.3 with KOH. All chemicalswere from Sigma-Aldrich, Tocris, and Abcam.
Whole-cell recording protocolFor determining the responsiveness of transduced CINs
to CNO, whole-cell recordings were made from identifiedCINs. Immediately after whole-cell access, current pulseswere delivered for identifying signature CIN currents. Typ-ically, CINs were spontaneously active at rest, but inalmost all cases, neuronal firing decreased with time. In afew examples, constant positive current was injected fortriggering action potentials (Fig. 2B). To control for pre-synaptic changes in excitability, in all cases synaptic an-tagonists (in �M: CNQX 20, AP5 100, CGP-52432 10, andgabazine 10) were bath-perfused before CNO application.To confirm recording stability, �5 min elapsed betweenwhole-cell access and testing for CNO-mediated respon-siveness. After 2 min of additional baseline recording,CNO (10 �M) was bath-applied for 3 min, and recordingscontinued CNO-free for no less than 8 min. A voltage-clamp ramp was delivered before and immediately afterCNO termination for determining the change in restingmembrane potential (VmR; Fig. 2B). Current–voltage (I-V)relationships were obtained by performing voltage-clampramps from 0 to 120 mV in 1.5 s.
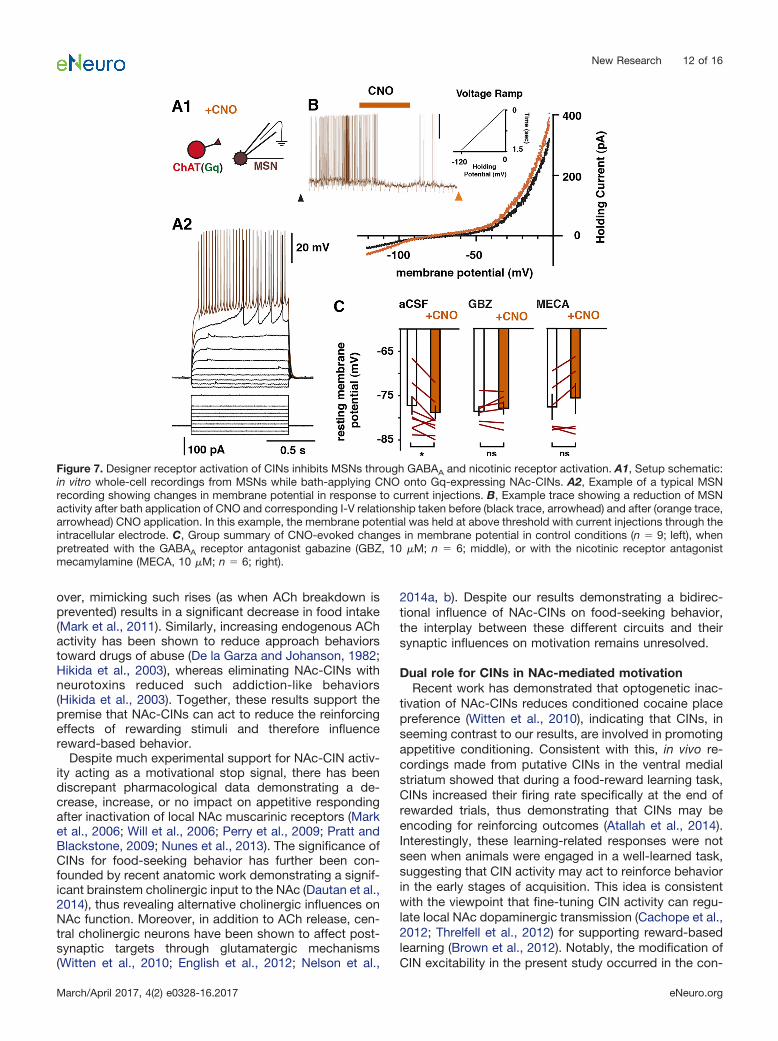
For determining the impact of CIN activation on me-dium spiny neuron (MSN) responses, whole-cell record-ings were made from putative NAc-MSNs. Immediatelyafter whole-cell access, current steps were delivered foridentifying signature MSNs currents. As was the case forwhole-cell CIN recordings, �5 min elapsed betweenwhole-cell access and testing for CNO-mediated respon-siveness. I-V relationships were obtained by performingvoltage-clamp ramps from 0 to 120 mV in 1.5 s involtage clamp immediately before recording baseline ac-tivity. Next, while in current-clamp recording mode, cur-rent was injected to bring the membrane potential ofMSNs close to threshold. After a 2-min baseline period,10 �M CNO was bath-applied for 3 min to activate Gq-expressing CINs. Changes in VmR were determined bycomparing a MSNs-injected current ramp before (Fig. 7B,black trace) recording and one 2 min after (Fig. 7B, brown
New Research 3 of 16
March/April 2017, 4(2) e0328-16.2017 eNeuro.org

trace) CNO termination. Only stable recordings were in-cluded in the dataset.
Food intake behaviorFor habituating animals to the experimental conditions,
mice were first gently handled for 1 wk in their homecages, and thereafter placed at the onset of the dark cycle(7 p.m.) on two consecutive days into individual cages fora 2-h food intake measurement. After the habituation, thead libitum diet-fed mice were administered either withCNO (1 mg/kg, i.p., Sequoia) or vehicle (10 mL/kg, i.p.,sterile 0.9% saline supplemented with 0.5% DMSO) 30min before the onset of the dark cycle in their homecages. This i.p. injection of CNO timeframe has beenshown to reliably impact virally expressed designer recep-tors in the CNS (Alexander et al., 2009). At the onset of thedark cycle, mice were transferred to the individual cageswith food (standard diet, as above) on the food hopperand water bottle installed, and food intake and bodyweight were measured. The mice were tested with bothCNO and vehicle administered in a counterbalanced orderwith a 2-d interval between sessions. After the acute foodintake experiment, the mice were fasted for 24 h in homecages, and all mice received an injection of CNO (1 mg/kg, i.p.) 30 min before the onset of the dark cycle. At theonset of the dark cycle, the mice were placed into theindividual cages with food on the hopper and water bottleinstalled, and food intake and body weight were measured.
Operant behaviorApparatus
Experiments were performed in Bussey-Saksida mouseoperant touchscreen chambers (Campden Instruments)as presented previously (Mar et al., 2013; Heath et al.,2015). The apparatus has a perforated stainless steel floorand trapezoidal walls bordering the area from a foodmagazine to a touchscreen (12.1 inches; resolution 800 �600) equipped with infrared beam arrays at �5 mm fromthe screen surface to detect nose-poke responses with-out animals having to apply pressure on the screen for aresponse to be detected. To guide responding and decreaseunintentional touches on the screen, a screen mask made ofblack acrylic with a row of five 4 � 4-cm openings 1 cmapart from each other and 1.5 cm from the floor level wasplaced in front of the screen. The white square visual stim-ulus is presented only in the central opening/location. Theapparatus is enclosed in a sound-attenuating chamber witha fan to provide ventilation and mask background noise. Thefood magazine connects to a pump delivering a reward(Yazoo Strawberry milkshake; Friesland Campina). A LEDand a speaker delivered a magazine light and a tone, re-spectively, at the reward delivery.
Touchscreen trainingAll testing was performed during the light cycle. Before
start of experiments, the mice were food restricted to85–90% of the free-feeding weight with water available adlibitum in the home cage throughout. Mice were firsthabituated to consume the milkshake reward in theirhome cages for 2 d to avoid hyponeophagia, and to theapparatus for two consecutive days, with all mice con-
suming 200 �L milkshake that was delivered into themagazine before the session start. To train the mice toassociate the stimulus with the reward delivery (20 �L), a60-min session was started with a presentation of thevisual stimulus for 30 s followed by a delivery of a tone(1000 ms, 3 kHz), a magazine light, and a reward. If thevisual stimulus was touched three times the quantity ofreward was delivered (60 �L). The magazine light wasturned off at the reward collection and followed by 5-sintertrial interval (ITI). All animals collected 30 rewards andthus reached the training criterion.
Fixed ratio trainingAfter the initial training, the mice were trained for fixed
ratio (FR) performance with the following parameters:stimulus removal for 500 ms, session length of 60 min,reward volume of 20 �L, ITI of 4.5 s, and tone of 10 ms at10 kHz. Mice were first trained to FR1 (one screen touchfor reinforcer delivery) and FR3, then continued to FR5.Animals reached the criterion when 30 trials were com-pleted during a session.
FR5 uncapped probeAnimals were administered with CNO (1 mg/kg, i.p.) or
vehicle 30 min before placing them into the chambers andtested for FR5 performance with no trial limit in 60-minsessions as in FR training. The mice were tested with bothCNO and vehicle administered in a counterbalanced orderwith a 3-d interval with a break day and a training sessionin between the test sessions.
Progressive ratioNext, mice were tested on a progressive ratio 4 (PR4)
schedule that required incremental increases in the num-ber of touches required for reinforcer delivery, as previ-ously described (Heath et al., 2015). Sessions wereterminated after either 5 min of inactivity or after 60 minhad elapsed. The mice were tested with CNO (1 mg/kg,i.p.) or vehicle administered in a counterbalanced orderwith a single baseline nondrug day between each drugtest session.
Data analysis and statisticsAll touchscreen testing data were automatically stored
in a database within ABET II touch testing software. Thetotal response time was defined as the amount of timebetween the first and last touch of a single trial and con-verted to rate for both FR and PR sessions. Individual ses-sions of FR response rate data were binned in blocks of 20trials and fitted with the parabolic function y b � x2 � a forstabilizing the variance and normalizing the distribution.Individual sessions of PR response rate data were fittedwith the negative exponential function y a–b�n as perpreviously reported analysis (Bailey et al., 2016). Thesedata were analyzed using repeated-measures ANOVA withtrial bin and CNO/vehicle condition as within-subject fac-tors. Post-reinforcement pause (PRP) was defined as thetime elapsed between the removal of the head from thereward magazine and the emission of the first touch ofthe next trial. These data were binned in blocks of 50 trialsfor FR sessions. Statistical analysis of this measure wasconducted using linear mixed models, as this class of
New Research 4 of 16
March/April 2017, 4(2) e0328-16.2017 eNeuro.org

statistical modeling tolerates incompleteness, and ani-mals cumulatively stopped responding throughout thesession. All statistical testing was conducted with a sig-nificance level of p � 0.05.
ResultsDesigner receptors are expressed in NAc-CINS andin sufficient quantities for modulating theirexcitability
For effectively manipulating NAc-CIN activity in vivo, weused a Cre-recombinase approach to express DREADDsin ChAT-cre� transgenic mice. Viral constructs that con-tained mCherry only (MCY), Gq, Gs, or Gi-coupled DREADDswere bilaterally injected in the medial division of the NAccore and shell. Immunocytochemical analysis revealedthat viral expression in CINs was specific, since MCY-expressing neurons were seen to be immunopositive forthe vesicular ACh transporter VAChT with high specificity(Fig. 1A, B). Consistent with previous studies (Bolamet al., 1984; Phelps et al., 1985; Contant et al., 1996; Zhouet al., 2002), the extent of their dendritic and axonalprocesses was limited to the NAc core and shell and didnot extend outside the NAc (Fig. 1A, C). To determinewhether the DREADD-specific ligand CNO was indeedsufficient to modulate CIN activity, we performed whole-cell recordings from identified transduced cells in acutebrain slices. Recorded neurons were regular spiking,mostly spontaneously active at rest, and showed signa-ture CIN intrinsic currents (Fig. 2A). As expected, in the
presence of 10 �M CNO, Gq-expressing CINs consis-tently showed a depolarization (Fig. 2B1, C; VmR: n 10;control, –60.7 � 3.1 mV; CNO, –49.9 � 3.4 mV; paired ttest, p 0.0007), whereas Gi-expressing CINs were hy-perpolarized (VmR: n 9; control, –63.8 � 3.8 mV; CNO,–67.2 � 3.9 mV; paired t test, p 0.02) with bath appli-cation of CNO (Fig. 2B2, C). CNO-evoked responses in GsCINs were significantly depolarizing (VmR: n 11; control,–55.4 � 2.7 mV; CNO, –52.8 � 2.6 mV; paired t test, p 0.04), but the responses were smaller in magnitude thanthose seen in Gq-expressing CINs (change in mV: Gq 10, 10.8 � 1.0 mV; Gs 11, 2.5 � 1.5 mV; unpaired t test,p � 0.0001; Fig. 2C). In contrast, there was no change inthe membrane potential of MCY-expressing CINs afterCNO application (VmR: n 6; control, –62.3 � 4.8 mV;CNO, –63.8 � 4.8 mV; paired t test, p 0.13). Compar-isons of CIN input resistance before and after CNO deliv-ery revealed a significant decrease in Gi-expressing CINs(n 9: Control, 329 � 28 M�; CNO, 282 � 24 M�; pairedt test, p 0.02) whereas neither Gq- nor Gs-expressingCINs showed any change in response to CNO (Gq, n 10; control, 341 � 22 M�; CNO, 359 � 43 M�; paired ttest, p 0.56: Gs, n 9; control, 327 � 62 M�; CNO,318 � 55 M�; paired t test, p 0.564). Together, theseresults provide evidence that Gq-, Gi-, and Gs-linkeddesigner receptors were selectively expressed in NAc-CINs and in sufficient quantities for modulating their ex-citability.
Figure 1. Selective expression of viral constructs in NAc cholinergic interneurons. Viral targeting of NAc-CINs in ChAT-cre miceresulted in selective expression of MCY (B2) in neurons expressing the vesicular ACh transporter VAChT (B1, B3) in the medial coreand shell region of the NAc (n 10; A, C). C, Darker shading corresponds to the qualitative greater density of the dendritic andsomatic extent of ACh-CIN labeling. ac, anterior commissure; ms, medial septum; NAcC, nucleus accumbens core; NAcS, nucleusaccumbens shell. White arrows, VAChT-expressing neurons; yellow arrowheads, neurons expressing both VAChT and MCY. Scalebars: A, 100 �m; B, 0.5 mm.
New Research 5 of 16
March/April 2017, 4(2) e0328-16.2017 eNeuro.org

CIN activity modulation does not change normalfeeding behavior
Previous pharmacological research investigating thecholinergic regulation of NAc-dependent feeding behaviorhas resulted in conflicting results, with reports of bothACh receptor-specific increases and decreases in foodintake that have been attributed to NAc-CINs (Hoebel
et al., 2007; Baldo et al., 2013). However, CIN communi-cation is not restricted to ACh release, as CINs are knownto excite postsynaptic targets by releasing glutamate (Hi-gley et al., 2011; Nelson et al., 2014a, b). Moreover, recentanatomic data has shown that mesopontine brainstemACh neurons also project to the NAc (Dautan et al., 2014),indicating that cholinergic regulation of NAc processes is
Figure 2. Designer receptor expression was sufficient for modulating CIN excitability in vitro. A, Recorded neurons were regularspiking and mostly spontaneously active at rest and showed signature CIN intrinsic currents. B, Bath application of 10 �MCNO reliably depolarized the membrane potential of neurons expression Gq-coupled (n 10; B1, C) and Gs-coupled (n 11;B3, C) designer receptors, whereas neurons expressing Gi-coupled (n 9) receptors were inhibited (B2). Black arrowhead(left), baseline current ramp injection; colored arrow (right), post-CNO current ramp injection. C, Results summary forCNO-mediated response from left to right in Gq-expressing (green), Gi-expressing (red), Gs-expressing (blue), and MCY-only(black; n 6) CINs plotted as the change in membrane potential. Synaptic antagonists (in �M) CNQX 20, AP5 100, CGP-5243210, gabazine 10.
New Research 6 of 16
March/April 2017, 4(2) e0328-16.2017 eNeuro.org

not restricted to CIN activity. As such, the relationshipbetween NAc-CIN activity and food intake remains unre-solved. Using a designer-receptor approach in vivo, weinvestigated how modifying the excitability of NAc-CINsaffected feeding behavior. To determine the impact of CINactivity on daily home-cage feeding bouts, mice express-ing Gq-linked (n 10), Gs-linked (n 14), and Gi-linked(n 10) linked DREADDs and MCY only (n 15) inNAc-CINs were injected with either CNO or vehicle (VEH)30 min before the start of the active phase, when micenormally show a significant increase in food intake. For allconditions, VEH and CNO injections were counterbal-anced over two testing sessions that were separated by 2days. Regardless of whether they received CNO or VEHinjection, mice ate an equal amount of home-cage diet2 hours after the start of the active phase (two-wayANOVA: group effect, F(3,90) 1.00, p 0.40; drugeffect, F(1,90) 0.003, p 0.96; interaction, F(3,90) 1.28, p 0.29; Fig. 3A). In addition to demonstratingthat CNO in control conditions (MCY group) has noimpact on normal eating behavior, these results provideevidence that changes in CIN activity do not adverselyaffect food intake by inducing taste aversion, as in-creases in NAc-ACh activity have been previouslyshown to do (Taylor et al., 2011).
Under states of high motivation, CIN modulationdifferentially affects food intake
In the preceding experiment, the motivation to con-sume food was relatively low, since animals had freeaccess to homecage food in their normal feeding environ-ment. Previous studies have shown CINs to be importantin reward-based behavior when the stimuli used arestrongly reinforcing, such as drugs of abuse and naturalrewards (Hajnal et al., 2000; Hikida et al., 2001, 2003). Totest for the involvement of NAc-CIN under heightenedlevels of motivation, we increased the incentive value ofthe home-cage food by fasting mice for 24 hours beforereintroducing food 30 min after CNO injections. Previousstudies have demonstrated that when fasted on multipleoccasions, mice increase their activity in anticipation forfood (Mieda et al., 2004; Storch and Weitz, 2009). Toavoid potential confounds from altered behavior inducedby multiple fasting events, a single fasting event was im-posed, therefore allowing for a between-subject compari-son. At 1 hour after the reintroduction of home-cage food,differences in food consumption between MCY and Gq-expressing mice began to emerge but were not statisticallydifferent (two-way ANOVA; between groups, F(3,95) 1.44, p� 0.05; within-group, F(1,95) 3.08, p � 0.05; Fig. 3B). At2 hours, however, Gq mice consumed significantly less thanMCY and Gi mice (MCY, 5.7 � 0.2; Gq, 4.8 � 0.6; Gi, 5.9 �
Figure 3. Impact of CIN activity modulation on food intake after free food access and a 24-h fast. A, MCY, Gs, Gq, and GiNAc-CIN–expressing mice with ad libitum food access were given an injection of either VEH or CNO 30 min before the dark cycle(active phase), and food consumption was measured 150 min later. B, CNO injections 30 min before the active phase in mice fastedfor 24 h resulted in a significant reduction in food intake for Gq-expressing compared with both MCY- and Gi-expressing mice at 2h after reintroducing home-cage food. Gq, n 10; Gs, n 14; Gi, n 10; MCY, n 15. �, p � 0.05.
New Research 7 of 16
March/April 2017, 4(2) e0328-16.2017 eNeuro.org

0.4 g/100 g body weight; one-way ANOVA, group effectF(3,44) 3.70, p 0.01; Bonferroni post hoc test; MCY vs.Gq p � 0.05, Gi vs. Gq p � 0.05; Fig. 3B), suggesting thatincreases in NAc-CIN activity, when animals are motivatedby way of a 24-h fast, can act to reduce food consumption.
These results are in agreement with past research re-lating increased NAc-ACh to reduced seeking behaviorfor natural rewards (Rada et al., 2005; Avena et al., 2006)and for drugs of abuse (Hikida et al., 2001, 2003; Zhou
et al., 2007). Based on such findings, it has been pro-posed that NAc-ACh can act to promote satiety signalingby reducing the incentive value of rewarding stimuli. Toshed light on the interaction between hunger, food re-ward, and satiety signaling, we trained food-restrictedmice to perform an operant task with a FR schedule ofreinforcement for receiving a palatable food reward (Fig. 4A).The motivating nature of the stimulus and the spaced feed-ing bouts provided by a FR regimen allowed us to measure
Figure 4. Differential modulation of CIN excitability impacts food-seeking behavior in a time-dependent manner. A, Mice were trained ona FR5 schedule of reinforcement for a palatable food reward. B, Summary of trials completed after CNO injection in DREADD-expressingand MCY mice. C, Between-group comparison of response rate over time. D, Within-group comparisons (VEH vs. CNO) of response rateover time for all groups. MCY (D1), n 12; Gs (D2), n 14; Gi (D3), n 10; Gq (D4), n 10. �, p � 0.05; ��, p � 0.01.
New Research 8 of 16
March/April 2017, 4(2) e0328-16.2017 eNeuro.org

changes in response rate that may reflect the cumulativedecrease in food intake that underlies satiety signaling. An-imals were trained to touch an illuminated touchscreen lo-cation five times (FR5) to receive 20 �L of a palatable foodreward delivered to a well opposite the touchscreen. Con-sistent with our results demonstrating a relationship be-tween CIN activity and a reduction in food intake in fastedmice, activating NAc-CINs with Gq- and Gi-linked DRE-ADDs resulted in an respective decrease and increase intotal trials completed, compared with MCY animals (MCY,180 � 6; Gq, 150 � 13; Gi, 207 � 5; Gs, 188 � 11; one-wayANOVA: group effect F(3,44) 5.80, p 0.002; Bonferronipost hoc test; MCY vs. Gq p 0.04, Gs vs. Gq p � 0.03, Givs. Gq p 0.009, MCY vs. Gi p 0.02; Fig. 4B).
To further characterize the impact of CINs on food-seeking behavior, we next measured changes in responserate between CNO-injected and control mice throughoutthe entire 1-hour session (Fig. 4). Despite equal initialrates, Gq (n 10) animals responded significantly lessthan Gi (n 10) animals toward the end of the session(one-way ANOVA: group effect F(3,42) 3.39, p 0.02;Bonferroni post hoc test; Gi vs. Gq p 0.01; Fig. 4C),whereas the VEH-treated groups were not significantlydifferent from one another (one-way ANOVA: group effectF(3,42) 1.62, p 0.20; data not shown). Within-subjectcomparisons (VEH and CNO sessions) for each grouprevealed a significant interaction between trial and injec-tion type (CNO vs. VEH) where Gq activation led to areduction in responding (one-way ANOVA with repeatedmeasures, group effect F(1,9) 5.07, p � 0.05; interaction,bin � treatment, F(19,171) 1.96, p 0.01; Fig. 4D4),whereas Gi-mediated inhibition of NAc-CINs resulted inan elevated level of food seeking compared with controlconditions (one-way ANOVA with repeated measures:group effect F(1,9) 3.22, p � 0.05; interaction, bin �
treatment, F(19,171) 2.65, p � 0.001; Fig. 4D3). In con-trast, neither MCY (n 12) nor Gs (n 14) animals treatedwith CNO showed altered food-seeking behavior com-pared with treatment with VEH (one-way ANOVA withrepeated measures: Gs group effect F(1,11) 0.58, p �0.05, interaction, bin � treatment, F(19,209) 0.68, p �0.05; MCY group effect F(1,13) 0.78, p � 0.05, interac-tion, bin � treatment, F(19,247) 1.36, p � 0.05; Figs. 4D1,D2). For all groups, mice injected with either CNO or VEHwere equally active throughout the session (two-wayANOVA: group effect, F(3,84) 1.54, p 0.21; drug effect,F(1,84) 0.31, p 0.58; interaction, F(3,84) 0.15, p 0.93), suggesting that there was no overt behavioral def-icit in responding (Fig. 5A).
We next measured the time after reward consumptionbefore animals voluntarily begin the next trial (Fig. 5B1).This delay, known as the post-reinforcement pause (PRP)has been shown to increase with food consumption and isthought to reflect the decrease in motivation for food dueto satiety (Sidman and Stebbins, 1954; Patrikiou andKeehn, 1964; Felton and Lyon, 1966). Consistent with thenotion that changes in CIN activity can affect motivationfor food, Gi-expressing mice injected with CNO showed asignificant decrease in PRP magnitude compared withcontrol mice (one-way ANOVA: Gi main effect by treat-
ment F(1,67) 6.16, p 0.01; interaction, bin � treatmentF(3,63) 1.17, p � 0.05; Fig. 5B3). Gq animals treated withCNO, however, did not show an increase in PRP com-pared with control conditions (one-way ANOVA: Gq maineffect by treatment F(1,45) 3.24, p 0.08; interaction, bin� treatment F(3,45) 1.81, p � 0.05; Fig. 5B5). Similarly,CNO-treated MCY mice showed no difference comparedwith control conditions (one-way ANOVA: MCY, main effectby treatment F(1,81) 0.34, p � 0.05; interaction, bin �treatment, F(3,81) 0.66, p � 0.05; Fig. 5B2). Surprisingly,Gs mice showed a significant difference in control andCNO conditions (main effect by treatment F(1,79) 5.01, p� 0.05; interaction, bin � treatment F(3,79) 0.06, p �0.05).
Stimulus incentive value that is largely independentof satiety signaling is unaffected by CIN modulation
By demonstrating that changes in CIN excitability canhave opposing effects on food intake, these results indi-cate that CINs play an active role in regulating motivation.In is unclear, however, whether these CIN-induced differ-ences in motivation were due to changes in the innatereinforcer strength of the stimuli that were unrelated toprior food consumption. To test this, mice were trained ona PR schedule of reinforcement in which, after successfultrial completion, the correct touches necessary for fooddelivery incrementally increased by four (Fig. 6A). The PRtask is well known to gauge reinforcer strength (Hodos,1961; Eagle et al., 1999) that is mostly independent ofsatiety signaling. The increased behavioral demand im-posed by this schedule of reinforcement results in signif-icantly less food being consumed compared with a FR5regimen. In the current study, PR4 and FR5 control miceconsumed on average 14.4 � 1.1 (�288 �L) and 169.5 �7.4 (�3.4 mL) rewards, respectively. The small amount offood consumed in the PR4 task allowed us to determinewhether the time-dependent changes in response ratescould be attributed to adjustments in reinforcer strengththat is largely independent of satiety signaling. Bothbetween-subject (Fig. 6B) and within-subject (Fig. 6C)analyses revealed that modulating CIN activity while ani-mals sought food rewards in a PR4 task had no significanteffect on trials completed (one-way ANOVA: group effectF(3,44) 0.80, p � 0.05; Fig. 6B) or on response rates(within group comparisons one-way ANOVA with repeatedmeasures, group effect: MCY (n 12); F(1,13) 0.52, p �0.05; interaction, bin � treatment, F(19,247) 0.75, p � 0.05.Gs (n 14): F(1,13) 0.39, p � 0.05; interaction, bin �treatment, F(19,247) 0.71, p � 0.05. Gq (n 10): F(1,9) 0.05, p � 0.05; interaction, bin � treatment, F(19,171) 0.42,p � 0.05. Gi (n 10): F(1,13) 0.16, p � 0.05; interaction, bin� treatment, F(19,247) 0.97, p � 0.05; Fig. 6C), supportingthe viewpoint that CIN activity does not change the overallincentive value of the rewarding stimuli.
Chemo-genetic activation of CINs inhibits accumbaloutput neurons through nicotinic and GABAergicreceptor activation
It has previously been shown that NAc-CINs can effec-tively inhibit NAc output MSNs (Witten et al., 2010; Nelsonet al., 2014a). In these studies, activating NAc-CINs op-
New Research 9 of 16
March/April 2017, 4(2) e0328-16.2017 eNeuro.org

togenetically with millisecond precision while recordingresponses in MSNs showed that the CIN-evoked inhibi-tion was likely due to an increase in GABAA receptoractivation (De Rover et al., 2002; Nelson et al., 2014a).DREADD approaches lack such clear temporal relation-ships, since activity is manipulated through G-protein–coupled second messenger pathways. We thus tested invitro whether activating CINs using Gq DREADDs cansimilarly inhibit MSNs (Fig. 7A). Bath application of 10 �M
CNO in acute brain slices expressing Gq DREADDs inCINs resulted in a significant hyperpolarization of MSNs(n 9: control, –76.2 � 2.1 mV; CNO, –78.6 � 1.5 mV;paired t test, p 0.03; Fig. 7B, C, left) that was dependenton GABAA receptor activation (pretreatment with 10 �Mgabazine, n 6: control, –78.9 � 1.0 mV; CNO, –77.1 �2.1 mV; paired t test, p 0.46; Fig. 7C, middle). Inaddition, these changes in membrane potential were sim-ilarly prevented by pretreating the tissue with the nonspe-cific nicotinic receptor antagonist mecamylamine (10 �M;n 6: control, –76.6 � 3.2 mV; CNO, –74.9 � 3.9 mV;paired t test, p 0.15; Fig. 7C, right), a result consistentwith recent work showing that CIN activation in the dorsalstriatum can drive GABA release from local interneuronsthat decreases MSN activity (Nelson et al., 2014a, b).
These results suggest that Gq-mediated activation ofNAc-CINs may act to release ACh that then influenceslocal NAc networks.
DiscussionThe present study demonstrates that modulation of
NAc-CIN activity is sufficient to bring about changes infood-motivated behaviors. Through DREADD-mediatedbidirectional control of CIN activity, we show that, whenmotivated by food restriction, increasing CIN activity ledto a reduction in palatable food consumption, whereasreducing CIN excitability enhanced food intake. Theseactivity-dependent changes developed late in the sessionand were unlikely to be driven by changes in the innatereinforcer strength, since CIN manipulation did not affectoperant responses to the reinforcer when food consump-tion was minimal. These results support the idea that CINmodulation was instead affecting the cumulative changein motivation underlying satiety signaling. Consistent withpast findings, we found that activating CINs in vitro led toan inhibition of NAc output neurons that was dependenton both GABAA and nicotinic receptor activation, sug-gesting that NAc-CINs can act to dampen MSN activity
Figure 5. Modulation of CINs impacts measure of satiety. A, In the FR5 operant task, all groups in VEH and CNO conditions showedsimilar total locomotor activity. B, Within-subject comparisons for the time taken, after reward consumption, for all groups to start anew trial (PRP). MCY (B2), n 12; Gi (B3), n 10; Gs (B4), n 14; Gq (B5), n 10. �, p � 0.05.
New Research 10 of 16
March/April 2017, 4(2) e0328-16.2017 eNeuro.org

through a feedforward activation of local GABAergic cir-cuits.
These bidirectional changes in food-seeking behaviorwere induced through Gq- or Gi-linked modified musca-rinic second messenger systems. Because muscarinic M3receptors are found in low levels in the striatum (Yasudaet al., 1993), these effects were unlikely caused by dis-rupted M3 receptor signaling but instead were likelycaused by increases in CIN activity (Fig. 2B1, C). It isconceivable, however, that Gi-mediated increases in re-sponding were attributed to disruptions in M2/4 secondmessenger signaling, as activating M2/4 autoreceptors onCINs can reduce Ca2� influx (Yan and Surmeier, 1996).Occluding this signaling pathway would potentially dis-rupt this Ca2� modulation, resulting in elevated excitabil-ity of CIN axon terminals. We predict that such a scenariowould result in enhanced transmission and would there-fore resemble the end effect of Gq activation. In contrastto Gq-activated CINs, inhibiting CINs through Gi-DREADDs
increased food-seeking behavior. These results thus sup-port the interpretation that our manipulations had a bidi-rectional impact on CIN excitability that resulted inopposing influences on food-seeking behavior. Similar toprevious studies, many of these responses lasted wellafter CNO washout (Alexander et al., 2009). Notably, de-spite Gs-expressing CINs showing CNO-mediated changes inactivity in vitro, these changes in excitability were insuffi-cient to affect behavior.
NAc cholinergic control of food intakeOur results are consistent with previous studies dem-
onstrating a link between NAc-ACh transmission and de-creases in food-directed motivation. Past studies haveshown that toward the end of a meal there is a local risein NAc-ACh that strongly correlates with reductions infood intake (Mark et al., 1992), the time course of whichcan be extended in conditions where meal duration isprolonged (Rada et al., 2005; Avena et al., 2008). More-
Figure 6. Reinforcer strength largely independent of satiety signaling is unaffected by CIN modulation. A, Mice were trained on a PRschedule of reinforcement in which correct touches for a palatable food reward consecutively increased by four. B, All groupscompleted an equal amount of trials, thus consuming a similar amount of food reward. C, Within group comparisons (VEH vs. CNO)of response rate over time or all groups. MCY (C1), n 12; Gi (C2), n 10; Gs (C3), n 14; Gq (C4), n 10.
New Research 11 of 16
March/April 2017, 4(2) e0328-16.2017 eNeuro.org
over, mimicking such rises (as when ACh breakdown isprevented) results in a significant decrease in food intake(Mark et al., 2011). Similarly, increasing endogenous AChactivity has been shown to reduce approach behaviorstoward drugs of abuse (De la Garza and Johanson, 1982;Hikida et al., 2003), whereas eliminating NAc-CINs withneurotoxins reduced such addiction-like behaviors(Hikida et al., 2003). Together, these results support thepremise that NAc-CINs can act to reduce the reinforcingeffects of rewarding stimuli and therefore influencereward-based behavior.
Despite much experimental support for NAc-CIN activ-ity acting as a motivational stop signal, there has beendiscrepant pharmacological data demonstrating a de-crease, increase, or no impact on appetitive respondingafter inactivation of local NAc muscarinic receptors (Market al., 2006; Will et al., 2006; Perry et al., 2009; Pratt andBlackstone, 2009; Nunes et al., 2013). The significance ofCINs for food-seeking behavior has further been con-founded by recent anatomic work demonstrating a signif-icant brainstem cholinergic input to the NAc (Dautan et al.,2014), thus revealing alternative cholinergic influences onNAc function. Moreover, in addition to ACh release, cen-tral cholinergic neurons have been shown to affect post-synaptic targets through glutamatergic mechanisms(Witten et al., 2010; English et al., 2012; Nelson et al.,
2014a, b). Despite our results demonstrating a bidirec-tional influence of NAc-CINs on food-seeking behavior,the interplay between these different circuits and theirsynaptic influences on motivation remains unresolved.
Dual role for CINs in NAc-mediated motivationRecent work has demonstrated that optogenetic inac-
tivation of NAc-CINs reduces conditioned cocaine placepreference (Witten et al., 2010), indicating that CINs, inseeming contrast to our results, are involved in promotingappetitive conditioning. Consistent with this, in vivo re-cordings made from putative CINs in the ventral medialstriatum showed that during a food-reward learning task,CINs increased their firing rate specifically at the end ofrewarded trials, thus demonstrating that CINs may beencoding for reinforcing outcomes (Atallah et al., 2014).Interestingly, these learning-related responses were notseen when animals were engaged in a well-learned task,suggesting that CIN activity may act to reinforce behaviorin the early stages of acquisition. This idea is consistentwith the viewpoint that fine-tuning CIN activity can regu-late local NAc dopaminergic transmission (Cachope et al.,2012; Threlfell et al., 2012) for supporting reward-basedlearning (Brown et al., 2012). Notably, the modification ofCIN excitability in the present study occurred in the con-
Figure 7. Designer receptor activation of CINs inhibits MSNs through GABAA and nicotinic receptor activation. A1, Setup schematic:in vitro whole-cell recordings from MSNs while bath-applying CNO onto Gq-expressing NAc-CINs. A2, Example of a typical MSNrecording showing changes in membrane potential in response to current injections. B, Example trace showing a reduction of MSNactivity after bath application of CNO and corresponding I-V relationship taken before (black trace, arrowhead) and after (orange trace,arrowhead) CNO application. In this example, the membrane potential was held at above threshold with current injections through theintracellular electrode. C, Group summary of CNO-evoked changes in membrane potential in control conditions (n 9; left), whenpretreated with the GABAA receptor antagonist gabazine (GBZ, 10 �M; n 6; middle), or with the nicotinic receptor antagonistmecamylamine (MECA, 10 �M; n 6; right).
New Research 12 of 16
March/April 2017, 4(2) e0328-16.2017 eNeuro.org

text of a familiar home cage and on well-trained operanttasks (FR5 and PR4). Our results may support a differen-tial role for NAc-CINs between initial reinforcement learn-ing and incentive updating in well-learned conditions, asis the case with satiety signaling. Consistent with thisview, NAc-ACh transmission can act to suppress thereinstatement of heroin-seeking on a well-learned operanttask (Zhou et al., 2007). More recently, Lee et al. (2016)further implicated CINs in updating well-learned rein-forced behavior by showing that NAc-CIN manipulationcan modulate extinction learning. The wide range of mo-tivational changes observed after NAc-CIN manipulationsuggests that this cellular population likely integrates sig-nals from multiple sources to modulate at different timesmotivational output.
Inhibitory CIN impact on NAc outputFrom a circuit-level perspective, the CIN-mediated de-
crease in food-seeking behavior may result from an over-all increase in inhibitory tone onto NAc output neurons.Previous studies have shown that optogenetically activat-ing CINs can effectively inhibit MSNs (Witten et al., 2010).Consistent with the known connectivity of CINs residing inthe dorsal striatum (English et al., 2012; Nelson et al.,2014a; Faust et al., 2015), our results indicate that inhibitoryGABA responses evoked by CNO onto Gq-expressing NAc-CINs were dependent on nicotinic receptor activation and werelikely due to the feedforward activation of local interneuronsthat inhibit NAc-MSNs. Different populations of MSNs thatcan be broadly classified by their neuropeptide and do-pamine (D1 and D2) receptor expression have beenshown to differentially affect the rewarding aspects ofcocaine. Whereas D1-type MSN activation can drivecocaine-mediated learning, D2-type MSN activation hasthe opposite effect (Lobo et al., 2010). More recently, ithas been shown that in mice activating D1- as well asD2-expressing NAc-MSNs can increase motivation forfood rewards (Soares-Cunha et al., 2016). Interestingly,devaluing the food reward by allowing mice free access tofood before the behavioral task completely disrupted theenhanced motivation after D2-MSN stimulation. More-over, optical inhibition of D2-MSNs diminished operantresponding for food. These results are consistent with ourcurrent findings demonstrating that DREADD-activatedCINs can reduce food-seeking behavior while also de-creasing the excitability of NAc-MSNs. Whether or notthese synaptic effects were preferentially on D1- or D2-expressing MSNs was not investigated in the currentstudy. Dopaminergic inputs to spatially distinct CINs havebeen shown to differentially impact their activity (Chuhmaet al., 2014), but whether specialized differences in micro-circuit connectivity between NAc-CINs and MSNs existremains unclear.
Our results support a role for NAc-CINs in bidirectionallyinfluencing satiety. One potential mechanism by which CINactivity can translate into reduced food-seeking behavior isvia an indirect inhibition of NAc-MSNs. The afferents thatdrive CIN activity remain unclear, however. Potential satiety-related sources that are positioned to impact CINs includehypothalamic energy-sensing circuits (Sternson, 2013) and
amygdala neurons known to encode for reappraisingstimulus value (Morrison and Salzman, 2010). Interest-ingly, recent work has revealed that CINs express insulinreceptors whose activation excites CINs (Stouffer et al.,2015), thus demonstrating an additional influence on CINswith the appropriate time course for progressively drivingsatiety. A more complete understanding of the variouscircuits that impact CINs will help reveal the various influ-ences on CINs that contribute to their regulation over thedrive to eat.
ReferencesAlexander GM, Rogan SC, Abbas AI, Armbruster BN, Pei Y, Allen JA,
Nonneman RJ, Hartmann J, Moy SS, Nicolelis MA, McNamara JO(2009) Remote control of neuronal activity in transgenic miceexpressing evolved G protein-coupled receptors. Neuron 63:27–39. CrossRef
Atallah HE, McCool AD, Howe MW, Graybiel AM (2014) Neurons inthe ventral striatum exhibit cell-type-specific representations ofoutcome during learning. Neuron 82:1145–1156. CrossRef Med-line
Avena NM, Rada P, Moise N, Hoebel BG (2006) Sucrose shamfeeding on a binge schedule releases accumbens dopamine re-peatedly and eliminates the acetylcholine satiety response. Neu-roscience 139:813–820. CrossRef Medline
Avena NM, Rada P, Hoebel BG (2008) Underweight rats have en-hanced dopamine release and blunted acetylcholine response inthe nucleus accumbens while bingeing on sucrose. Neuroscience156:865–871. CrossRef Medline
Bailey MR, Williamson C, Mezias C, Winiger V, Silver R, Balsam PD,Simpson EH (2016) The effects of pharmacological modulation ofthe serotonin 2C receptor on goal-directed behavior in mice. Psychop-harmacology (Berl) 233:615–624. CrossRef Medline
Baldo BA, Pratt WE, Will MJ, Hanlon EC, Bakshi VP, Cador M (2013)Principles of motivation revealed by the diverse functions of neu-ropharmacological and neuroanatomical substrates underlyingfeeding behavior. Neurosci Biobehav Rev 37:1985–1998. Cross-Ref Medline
Bolam JP, Wainer BH, Smith AD (1984) Characterization of cholin-ergic neurons in the rat neostriatum. A combination of cholineacetyltransferase immunocytochemistry, Golgi-impregnation andelectron microscopy. Neuroscience 12:711–718. CrossRef
Brog JS, Salyapongse A, Deutch AY, Zahm DS (1993) The patternsof afferent innervation of the core and shell in the ‘Accumbens’part of the rat ventral striatum: immunohistochemical detection ofretrogradely transported fluoro�gold. J Comp Neurol 338:255–278.CrossRef Medline
Brown MTC, Tan KR, O’Connor EC, Nikonenko I, Muller D, LüscherC (2012) Ventral tegmental area GABA projections pause accum-bal cholinergic interneurons to enhance associative learning. Na-ture 492:452–456. CrossRef
Cachope R, Mateo Y, Mathur BN, Irving J, Wang HL, Morales M,Lovinger DM, Cheer JF (2012) Selective activation of cholinergicinterneurons enhances accumbal phasic dopamine release: set-ting the tone for reward processing. Cell Rep 2:33–41. CrossRefMedline
Cardinal RN, Parkinson JA, Hall J, Everitt BJ (2002) Emotion andmotivation: the role of the amygdala, ventral striatum, and prefron-tal cortex. Neurosci Biobehav Rev 26:321–352.
Chau DT, Rada P, Kosloff RA, Taylor JL, Hoebel BG (2001) Nucleusaccumbens muscarinic receptors in the control of behavioral de-pression: antidepressant-like effects of local M 1 antagonist in thePorsolt swim test. Neuroscience 104:791–798. CrossRef
Chuhma N, Mingote S, Moore H, Rayport S (2014) Dopamine neu-rons control striatal cholinergic neurons via regionally heteroge-neous dopamine and glutamate signaling. Neuron 81:901–912.CrossRef Medline
New Research 13 of 16
March/April 2017, 4(2) e0328-16.2017 eNeuro.org

Contant C, Umbriaco D, Garcia S, Watkins KC, Descarries L (1996)Ultrastructural characterization of the acetylcholine innervation inadult rat neostriatum. Neuroscience 71:937–947. Medline
Corbit LH, Muir JL, Balleine BW (2001) The role of the nucleusaccumbens in instrumental conditioning: evidence of a functionaldissociation between accumbens core and shell. J Neurosci 21:3251–3260.
Dautan D, Huerta-Ocampo I, Witten IB, Deisseroth K, Bolam JP,Gerdjikov T, Mena-Segovia J (2014) A major external source ofcholinergic innervation of the striatum and nucleus accumbensoriginates in the brainstem. J Neurosci 34:4509–4518. CrossRef
De la Garza R, Johanson CE (1982) Effects of haloperidol andphysostigmine on self-administration of local anesthetics. Phar-macol Biochem Behav 17:1295–1299. Medline
De Rover M, Lodder JC, Kits KS, Schoffelmeer AN, Brussaard AB(2002) Cholinergic modulation of nucleus accumbens mediumspiny neurons. Eur J Neurosci 16:2279–2290. Medline
Descarries L, Mechawar N (2000) Ultrastructural evidence for diffusetransmission by monoamine and acetylcholine neurons of thecentral nervous system. Prog. Brain Res 125:27–47. CrossRefMedline
Eagle DM, Humby T, Dunnett SB, Robbins TW (1999) Effects ofregional striatal lesions on motor, motivational, and executiveaspects of progressive-ratio performance in rats. Behav. Neurosci113:718–731. Medline
English DF, Ibanez-Sandoval O, Stark E, Tecuapetla F, Buzsáki G,Deisseroth K, Tepper JM, Koos T (2012) GABAergic circuits me-diate the reinforcement-related signals of striatal cholinergic in-terneurons. Nat Neurosci 15:123–130.
Faust TW, Assous M, Shah F, Tepper JM, Koós T (2015) Novel fastadapting interneurons mediate cholinergic-induced fast GABAAinhibitory postsynaptic currents in striatal spiny neurons. Eur JNeurosci 42:1764–1774. CrossRef Medline
Felton M, Lyon DO (1966) The post�reinforcement pause. J Exp AnalBehav 9:131–134. CrossRef Medline
Hajnal A, Székely M, Gálosi R, Lénárd L (2000) Accumbens cholin-ergic interneurons play a role in the regulation of body weight andmetabolism. Physiol Behav 70:95–103. Medline
Heath CJ, Bussey TJ, Saksida LM (2015) Motivational assessment ofmice using the touchscreen operant testing system: effects ofdopaminergic drugs. Psychopharmacology (Berl) 232:4043–4057.CrossRef Medline
Higley MJ, Gittis AH, Oldenburg IA, Balthasar N, Seal RP, EdwardsRH, Lowell BB, Kreitzer AC, Sabatini BL (2011) Cholinergic in-terneurons mediate fast VGluT3-dependent glutamatergic trans-mission in the striatum. PLoS One 6:e19155. CrossRef Medline
Hikida T, Kaneko S, Isobe T, Kitabatake Y, Watanabe D, Pastan I,Nakanishi S (2001) Increased sensitivity to cocaine by cholinergiccell ablation in nucleus accumbens. Proc Natl Acad Sci U S A98:13351–13354. CrossRef Medline
Hikida T, Kitabatake Y, Pastan I, Nakanishi S (2003) Acetylcholineenhancement in the nucleus accumbens prevents addictive be-haviors of cocaine and morphine. Proc Natl Acad Sci U S A100:6169–6173. CrossRef Medline
Hodos W (1961) Progressive ratio as a measure of reward strength.Science (NY) 134:943–944. Medline
Hoebel BG, Avena NM, Rada P (2007) Accumbens dopamine-acetylcholine balance in approach and avoidance. Curr Opin Phar-macol 7:617–627. CrossRef Medline
Itzhak Y, Martin JL (2002) Cocaine-induced conditioned place pref-erence in mice: induction, extinction and reinstatement by relatedpsychostimulants. Neuropsychopharmacology 26:130–134. Cross-Ref Medline
Jones DL, Mogenson GJ (1980) Nucleus accumbens to globuspallidus GABA projection subserving ambulatory activity. Am JPhysiol Integr Comp Physiol 238:R65–R69. Medline
Kawaguchi Y (1993) Physiological, morphological, and histochemicalcharacterization of three classes of interneurons in rat neostriatum.J Neurosci 13:4908–4923. Medline
Lee J, Finkelstein J, Choi JY, Witten IB (2016) Linking cholinergicinterneurons, synaptic plasticity, and behavior during the extinction ofa cocaine-context association. Neuron 90:1071–1085. CrossRefMedline
Lobo MK, Covington HE, Chaudhury D, Friedman AK, Sun H,Damez-Werno D, Dietz DM, Zaman S, Koo JW, Kennedy PJ (2010)Cell type–specific loss of BDNF signaling mimics optogeneticcontrol of cocaine reward. Science 330:385–390. CrossRef Med-line
Mannella F, Gurney K, Baldassarre G (2013) The nucleus accumbensas a nexus between values and goals in goal-directed behavior: areview and a new hypothesis. Front Behav Neurosci 7:e1–e29.CrossRef
Mar AC, Horner AE, Nilsson SRO, Alsiö J, Kent BA, Kim CH, HolmesA, Saksida LM, Bussey TJ (2013) The touchscreen operant plat-form for assessing executive function in rats and mice. Nat Protoc8:985–2005. CrossRef
Mark GP, Kinney AE, Grubb MC, Zhu X, Finn DA, Mader SL, BergerSP, Bechtholt AJ (2006) Injection of oxotremorine in nucleus ac-cumbens shell reduces cocaine but not food self-administration inrats. Brain Res 1123:51–59. CrossRef
Mark GP, Rada P, Pothos E, Hoebel BG (1992) Effects of feeding anddrinking on acetylcholine release in the nucleus accumbens, stria-tum, and hippocampus of freely behaving rats. J Neurochem58:2269–2274. Medline
Mark GP, Shabani S, Dobbs LK, Hansen ST (2011) Cholinergicmodulation of mesolimbic dopamine function and reward. PhysiolBehav 104:76–81. CrossRef Medline
McDonald AJ (1991) Organization of amygdaloid projections to theprefrontal cortex and associated striatum in the rat. Neuroscience44:1–14. Medline
Mieda M, Williams SC, Sinton CM, Richardson JA, Sakurai T, Yanag-isawa M (2004) Orexin neurons function in an efferent pathway ofa food-entrainable circadian oscillator in eliciting food-anticipatoryactivity and wakefulness. J Neurosci 24:10493–10501. CrossRef
Mogenson GJ, Jones DL, Yim CY (1980) From motivation to action:functional interface between the limbic system and the motorsystem. Prog Neurobiol 14:69–97. Medline
Morrison SE, Salzman CD (2010) Re-valuing the amygdala. Curr OpinNeurobiol 20:221–230. CrossRef Medline
Nelson AB, Bussert TG, Kreitzer AC, Seal RP (2014a) Striatal cho-linergic neurotransmission requires VGLUT3. J Neurosci 34:8772–8777. CrossRef Medline
Nelson AB, Hammack N, Yang CF, Shah NM, Seal RP, Kreitzer AC(2014b) Striatal cholinergic interneurons drive GABA release fromdopamine terminals. Neuron 82:63–70. CrossRef Medline
Nunes EJ, Randall PA, Podurgiel S, Correa M, Salamone JD (2013)Nucleus accumbens neurotransmission and effort-related choicebehavior in food motivation: effects of drugs acting on dopamine,adenosine, and muscarinic acetylcholine receptors. NeurosciBiobehav Rev 37:2015–2025. CrossRef Medline
Patrikiou VD, Keehn JD (1964) The effect of partial satiation onpauses after reinforcements scheduled on alternating fixed ratios.Psychon Sci 1:79–80. CrossRef
Perry ML, Baldo BA, Andrzejewski ME, Kelley AE (2009) Muscarinicreceptor antagonism causes a functional alteration in nucleusaccumbens �-opiate-mediated feeding behavior. Behav Brain Res197:225–229. CrossRef Medline
Phelps PE, Houser CR, Vaughn JE (1985) Immunocytochemicallocalization of choline acetyltransferase within the rat neostriatum:a correlated light and electron microscopic study of cholinergicneurons and synapses. J Comp Neurol 238:286–307. CrossRefMedline
Phillipson OT, Griffiths AC (1985) The topographic order of inputs tonucleus accumbens in the rat. Neuroscience 16:275–296. Medline
Pratt WE, Kelley AE (2004) Nucleus accumbens acetylcholine regu-lates appetitive learning and motivation for food via activation ofmuscarinic receptors. Behav Neurosci 118:730. CrossRef Medline
New Research 14 of 16
March/April 2017, 4(2) e0328-16.2017 eNeuro.org

Pratt WE, Blackstone K (2009) Nucleus accumbens acetylcholineand food intake: decreased muscarinic tone reduces feeding butnot food-seeking. Behav Brain Res 198:252–257. CrossRef
Rada P, Avena NM, Hoebel BG (2005) Daily bingeing on sugarrepeatedly releases dopamine in the accumbens shell. Neurosci-ence 134:737–744. CrossRef Medline
Sidman M, Stebbins WC (1954) Satiation effects under fixed-ratioschedules of reinforcement. J. Comp. Physiol Psychol 47:114.Medline
Singh T, McDannald M, Haney R, Cerri D, Schoenbaum G (2010)Nucleus accumbens core and shell are necessary for reinforcerdevaluation effects on pavlovian conditioned responding. FrontIntegr Neurosci 4:126. CrossRef Medline
Soares-Cunha C, Coimbra B, David-Pereira A, Borges S, Pinto L,Costa P, Sousa N, Rodrigues AJ (2016) Activation of D2 dopaminereceptor-expressing neurons in the nucleus accumbens increasesmotivation. Nat Commun 7. CrossRef
Sternson SM (2013) Hypothalamic survival circuits: blueprints forpurposive behaviors. Neuron 77:810–824. CrossRef Medline
Storch KF, Weitz CJ (2009) Daily rhythms of food-anticipatory be-havioral activity do not require the known circadian clock. PNAS106:6808–6813. CrossRef Medline
Stouffer MA, Woods CA, Patel JC, Lee CR, Witkovsky P, Bao L,Machold RP, Jones KT, de Vaca SC, Reith MEA, Carr KD, Rice ME(2015) Insulin enhances striatal dopamine release by activatingcholinergic interneurons and thereby signals reward. Nat Commun6:8543. CrossRef
Taylor KM, Mark GP, Hoebel BG (2011) Conditioned taste aversionfrom neostigmine or methyl-naloxonium in the nucleus accum-bens. Physiol Behav 104:82–86. CrossRef Medline
Threlfell S, Lalic T, Platt NJ, Jennings KA, Deisseroth K, Cragg SJ (2012)Striatal dopamine release is triggered by synchronized activity in cholin-ergic interneurons. Neuron 75:58–64. CrossRef Medline
Trivedi P, Yu H, MacNeil DJ, Van der Ploeg LHT, Guan X-M (1998)Distribution of orexin receptor mRNA in the rat brain. FEBS Lett438:71–75. Medline
Warner-Schmidt JL, Vanover KE, Chen EY, Marshall JJ, Greengard P(2011) Antidepressant effects of selective serotonin reuptake in-hibitors (SSRIs) are attenuated by antiinflammatory drugs in miceand humans. Proc Natl Acad Sci U S A 108:9262–9267. CrossRefMedline
Will MJ, Pratt WE, Kelley AE (2006) Pharmacological characterizationof high-fat feeding induced by opioid stimulation of the ventralstriatum. Physiol Behav 89:226–234. CrossRef Medline
Williams MJ, Adinoff B (2008) The role of acetylcholine in cocaineaddiction. Neuropsychopharmacology 33:1779–1797. CrossRefMedline
Witten IB, Lin S-C, Brodsky M, Prakash R, Diester I, Anikeeva P,Gradinaru V, Ramakrishnan C, Deisseroth K (2010) Cholinergicinterneurons control local circuit activity and cocaine conditioning.Science 330:1677–1681. CrossRef Medline
Woolf NJ, Butcher LL (1981) Cholinergic neurons in the caudate-putamen complex proper are intrinsically organized: a combinedEvans blue and acetylcholinesterase analysis. Brain Res Bull7:487–507. Medline
Yan Z, Surmeier DJ (1996) Muscarinic (m2/m4) receptors reduceN-and P-type Ca2� currents in rat neostriatal cholinergic interneu-rons through a fast, membrane-delimited, G-protein pathway. JNeurosci 16:2592–2604.
Yasuda RP, Ciesla WI, Flores LR, Wall SJ, Li M, Satkus SA,Weisstein JS, Spagnola BV, Wolfe BB (1993) Development ofantisera selective for m4 and m5 muscarinic cholinergic recep-tors: distribution of m4 and m5 receptors in rat brain. MolPharmacol 43:149–157.
Zhou F, Wilson CJ, Dani JA (2002) Cholinergic interneuron charac-teristics and nicotinic properties in the striatum. J Neurobiol 53:590–605. CrossRef Medline
Zhou W, Liu H, Zhang F, Tang S, Zhu H, Lai M, Kalivas PW (2007)Role of acetylcholine transmission in nucleus accumbens andventral tegmental area in heroin-seeking induced by conditionedcues. Neuroscience 144:1209–1218. CrossRef Medline
New Research 15 of 16
March/April 2017, 4(2) e0328-16.2017 eNeuro.org