of fat flies and hippos, or the magic of animal size
TRANSCRIPT

NATURE STRUCTURAL & MOLECULAR BIOLOGY VOLUME 13 NUMBER 12 DECEMBER 2006 1051
the membrane interface13. It is thus intrigu-ing to find in the GlpG structure a belt of tryptophan residues that spiral around the TM5, TM2 and TM1 surfaces from the extra-cellular to the cytoplasmic end of the protein (Fig. 1a,b). These are highly conserved in bacterial GlpG proteins2. These observations raise the possibility that structurally and functionally significant lipid-protein inter-actions might be revealed by the inclusion of phospholipids during the crystallization14,15 of GlpG. But perhaps the greatest advance in our understanding of GlpG will come from inclusion of a catalytically inactive substrate in GlpG crystals.
ACKNOWLEDGMENTSThis work was supported in part by grants from the National Institute of General Medical Sciences and the National Center for Research Resources, US National Institutes of Health.
1. Wang, Y., Zhang, Y. & Ha, Y. Nature 444, 179–183 (2006).
2. Wu, Z. et al. Nat. Struct. Mol. Biol. 13, 1084–1091 (2006).
3. Ben-Shem, A., Fass, D. & Bibi, E. Proc. Natl. Acad. Sci. USA (in the press).
4. Doyle, D.A. et al. Science 280, 69–77 (1998).5. Hunte, C. et al. Nature 435, 1197–1202 (2005).6. Abramson, J. et al. Science 301, 610–615 (2003).7. Van den Berg, B. et al. Nature 427, 36–44 (2004).8. Freites, J.A., Tobias, D.J., von Heijne, G. & White,
S.H. Proc. Natl. Acad. Sci. USA 102, 15059–15064
(2005).9. Freites, J.A., Tobias, D.J. & White, S.H. Biophys. J.,
published online 29 September 2006 (doi:10.1529/biophysj.106.096065).
10. Sands, Z.A., Grottesi, A. & Sansom, M.S.P. Biophys. J. 90, 1598–1606 (2006).
11. Schmidt, D., Jiang, Q.-X. & MacKinnon, R. Nature published online 29 November 2006 (doi:10.1038/nature05416).
12. Ulmschneider, M.B., Sansom, M.S.P. & Di Nola, A. Proteins 59, 252–265 (2005).
13. Yau, W.-M., Wimley, W.C., Gawrisch, K. & White, S.H. Biochemistry 37, 14713–14718 (1998).
14. Long, S.B., Campbell, E.B. & MacKinnon, R. Science 309, 903–908 (2005).
15. Guan, L., Smirnova, I.N., Verner, G., Nagamori, S. & Kaback, H.R. Proc. Natl. Acad. Sci. USA 103, 1723–1726 (2006).
16. Humphrey, W., Dalke, W. & Schulten, K. J. Mol. Graph. 14, 33–38 (1996).
Fernando Casares, Carlos M. Luque and Maria J. Tavares are at the Centro Andaluz de Biología del Desarrollo, Consejo Superior de Investigaciones Científicas-Universidad Pablo de Olavide, 41013 Seville, Spain. e-mail: [email protected]
Of Fat flies and Hippos, or the magic of animal sizeFernando Casares, Carlos M Luque & Maria J Tavares
A stream of papers exposes the molecular nuts and bolts of a mechanism that prevents cells from becoming immortal.
During embryonic development, animals reach consistent species-specific shapes and sizes. What are the mechanisms that determine animal size and the relative dimensions of different organs and tissues? The initial phases of embryogenesis, involving cell patterning and rapid proliferation, eventually give way to cellular differentiation, with slowed cell proliferation and the commencement of widespread developmentally regulated cell death or apoptosis. During development, apoptosis not only serves to remove damaged cells but also helps sculpt tissue shape and size. Over the past few years, work primarily in Drosophila melanogaster has led to the identification of a conserved pathway, known as the Hippo (Hpo) pathway, which is key in size and shape control. As cells enter the differentiation phase of development, the Hpo pathway acts to curb proliferation of cells and increase their sensitivity to developmentally regulated apoptosis. Typically, members of the Hpo pathway behave as tumor suppressors, as their mutation results in hyperplastic growth. Loss of Hpo-pathway function in epithelial cells results in their overproliferation and resistance to apoptosis, although they retain
both their epithelial organization and cell-type specification acquired earlier during development. Flies with mutations in Hpo-pathway components develop hugely overgrown external organs, reminiscent of the loose, pleated skin of shar-pei dogs. Thus, it is clear that the Hpo pathway has a vital role in the transition from proliferation to differentiation, which is crucial for the control of size.
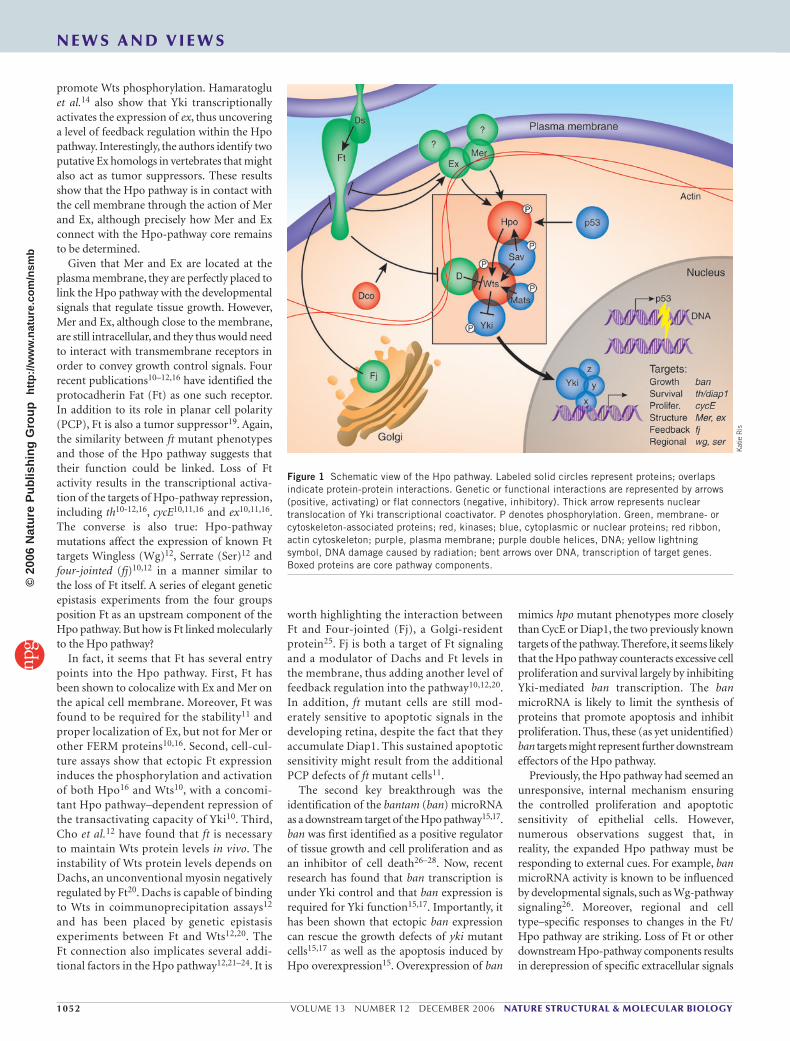
Until recently, understanding of the Hpo pathway was somewhat rudimentary. The pathway depends on the action of two serine/threonine kinases, Hpo and Warts (Wts), in combination with two adaptor proteins, Salvador (Sav) and ‘Mob as tumor suppressor’ (Mats), to phosphorylate and inhibit the transcriptional coactivator Yorkie (Yki) (Fig. 1). Eliminating Hpo, Wts or either of their partners abrogates this repression and unleashes Yki, resulting in unchecked hyperplastic cell proliferation1–7; for a recent review, see ref. 8. Two genes whose expression is activated by Yki, cyclin E (cycE) and thread (th), which encodes Drosophila inhibitor of apoptosis 1 (Diap1), have long been regarded as the main targets of the Hpo pathway in tumor suppression. Although these two genes are clearly implicated in proliferation and resistance to apoptosis, they are unlikely to be the only targets of the Hpo pathway, given that CycE overexpression combined with the inhibition of apoptosis is insufficient to trigger tissue overgrowth9. Moreover, the components of the ‘core’ Hpo pathway are intracellular proteins. Given the importance of the Hpo
pathway in controlling cell proliferation and apoptosis, it seems necessary that its function be linked to the extracellular world and be able to respond to developmental cues. In the past year, several new and exciting papers have started to shed light on such issues10–17.
The initial breakthrough was linking the Hpo pathway to the plasma membrane, the communication interface of the cell. Merlin (Mer), and the related protein Expanded (Ex), are members of the ‘4.1, Ezrin, Radixin, Moesin’ (FERM) domain superfamily. Both are membrane- associated cytoplasmic proteins that may function as adapters linking transmembrane proteins with multiprotein complexes, the cytoskeleton or both. Mutations in neurofibromatosis type 2 (NF-2), the human ortholog of Mer, cause tumors in the central nervous system. In flies, Mer and Ex function together to regulate cell proliferation, interacting both genetically and physically18, although the molecular basis for their tumor- suppressor activity had not been clear. Hamaratoglu et al.14 noticed that the phenotypes caused by the loss of Mer and Ex were highly similar to those of Hpo-pathway mutants. Mer or ex single mutants show slight overgrowths, but double-mutant cells proliferate dramatically, indicating that the two proteins function in a partially redundant manner. mer;ex double-mutant cells upregulate Diap1 and avoid cell-cycle arrest. Genetic epistasis experiments in conjunction with biochemical and cell- culture assays show that Mer and Ex activate the pathway upstream of Hpo and cooperate to
N E W S A N D V I E W S©
2006
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
smb
1052 VOLUME 13 NUMBER 12 DECEMBER 2006 NATURE STRUCTURAL & MOLECULAR BIOLOGY
promote Wts phosphorylation. Hamaratoglu et al.14 also show that Yki transcriptionally activates the expression of ex, thus uncovering a level of feedback regulation within the Hpo pathway. Interestingly, the authors identify two putative Ex homologs in vertebrates that might also act as tumor suppressors. These results show that the Hpo pathway is in contact with the cell membrane through the action of Mer and Ex, although precisely how Mer and Ex connect with the Hpo-pathway core remains to be determined.
Given that Mer and Ex are located at the plasma membrane, they are perfectly placed to link the Hpo pathway with the developmental signals that regulate tissue growth. However, Mer and Ex, although close to the membrane, are still intracellular, and they thus would need to interact with transmembrane receptors in order to convey growth control signals. Four recent publications10–12,16 have identified the protocadherin Fat (Ft) as one such receptor. In addition to its role in planar cell polarity (PCP), Ft is also a tumor suppressor19. Again, the similarity between ft mutant phenotypes and those of the Hpo pathway suggests that their function could be linked. Loss of Ft activity results in the transcriptional activa-tion of the targets of Hpo-pathway repression, including th10-12,16, cycE10,11,16 and ex10,11,16. The converse is also true: Hpo-pathway mutations affect the expression of known Ft targets Wingless (Wg)12, Serrate (Ser)12 and four-jointed (fj)10,12 in a manner similar to the loss of Ft itself. A series of elegant genetic epistasis experiments from the four groups position Ft as an upstream component of the Hpo pathway. But how is Ft linked molecularly to the Hpo pathway?
In fact, it seems that Ft has several entry points into the Hpo pathway. First, Ft has been shown to colocalize with Ex and Mer on the apical cell membrane. Moreover, Ft was found to be required for the stability11 and proper localization of Ex, but not for Mer or other FERM proteins10,16. Second, cell-cul-ture assays show that ectopic Ft expression induces the phosphorylation and activation of both Hpo16 and Wts10, with a concomi-tant Hpo pathway–dependent repression of the transactivating capacity of Yki10. Third, Cho et al.12 have found that ft is necessary to maintain Wts protein levels in vivo. The instability of Wts protein levels depends on Dachs, an unconventional myosin negatively regulated by Ft20. Dachs is capable of binding to Wts in coimmunoprecipitation assays12 and has been placed by genetic epistasis experiments between Ft and Wts12,20. The Ft connection also implicates several addi-tional factors in the Hpo pathway12,21–24. It is
worth highlighting the interaction between Ft and Four-jointed (Fj), a Golgi-resident protein25. Fj is both a target of Ft signaling and a modulator of Dachs and Ft levels in the membrane, thus adding another level of feedback regulation into the pathway10,12,20. In addition, ft mutant cells are still mod-erately sensitive to apoptotic signals in the developing retina, despite the fact that they accumulate Diap1. This sustained apoptotic sensitivity might result from the additional PCP defects of ft mutant cells11.
The second key breakthrough was the identification of the bantam (ban) microRNA as a downstream target of the Hpo pathway15,17. ban was first identified as a positive regulator of tissue growth and cell proliferation and as an inhibitor of cell death26–28. Now, recent research has found that ban transcription is under Yki control and that ban expression is required for Yki function15,17. Importantly, it has been shown that ectopic ban expression can rescue the growth defects of yki mutant cells15,17 as well as the apoptosis induced by Hpo overexpression15. Overexpression of ban
mimics hpo mutant phenotypes more closely than CycE or Diap1, the two previously known targets of the pathway. Therefore, it seems likely that the Hpo pathway counteracts excessive cell proliferation and survival largely by inhibiting Yki-mediated ban transcription. The ban microRNA is likely to limit the synthesis of proteins that promote apoptosis and inhibit proliferation. Thus, these (as yet unidentified) ban targets might represent further downstream effectors of the Hpo pathway.
Previously, the Hpo pathway had seemed an unresponsive, internal mechanism ensuring the controlled proliferation and apoptotic sensitivity of epithelial cells. However, numerous observations suggest that, in reality, the expanded Hpo pathway must be responding to external cues. For example, ban microRNA activity is known to be influenced by developmental signals, such as Wg- pathway signaling26. Moreover, regional and cell type–specific responses to changes in the Ft/Hpo pathway are striking. Loss of Ft or other downstream Hpo-pathway components results in derepression of specific extracellular signals
Figure 1 Schematic view of the Hpo pathway. Labeled solid circles represent proteins; overlaps indicate protein-protein interactions. Genetic or functional interactions are represented by arrows (positive, activating) or flat connectors (negative, inhibitory). Thick arrow represents nuclear translocation of Yki transcriptional coactivator. P denotes phosphorylation. Green, membrane- or cytoskeleton-associated proteins; red, kinases; blue, cytoplasmic or nuclear proteins; red ribbon, actin cytoskeleton; purple, plasma membrane; purple double helices, DNA; yellow lightning symbol, DNA damage caused by radiation; bent arrows over DNA, transcription of target genes. Boxed proteins are core pathway components.
Kat
ie R
is
N E W S A N D V I E W S©
2006
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
smb

NATURE STRUCTURAL & MOLECULAR BIOLOGY VOLUME 13 NUMBER 12 DECEMBER 2006 1053
during development, such as Wg or the Notch ligand Serrate, but only in certain regions of the epithelium. Meanwhile, the loss of Mer and Ex, or Hpo, or Wts from the fly retina causes exces-sive proliferation of the interommatidial cells (between facets of the fly compound eye) but has no effect upon photoreceptors10,12,14. The reason for the different responses to pertur-bations in the Hpo pathway is unknown, but investigating their basis will be crucial to under-standing how the Hpo pathway helps shape organs and tissues during development.
One potential source of the apparent tissue and cell specificity of the Hpo pathway may reside in the transcription factors with which Yki interacts. These might be expressed dif-ferentially so that different combinations of Yki transcription factors generate distinct transcriptional responses. Some clues to the identity of Yki partners come from studies on the mammalian counterpart of Yki, the Yes- associated protein (YAP), which shares 31% amino acid identity with Yki6 and has previously been described as a transcriptional coactivator of distinct transcription factors such as p73, PEBP2α and the TEAD/TEF family. Yki and YAP have two WW domains, known to interact with PPXY-contain-ing polypeptides. However, the Drosophila homologs of PEBP2α and TEAD2, Runt and Scalloped, respectively, do not contain intact YAP- binding motifs, making their sta-tus as candidate Yki partners less clear-cut. Either way, the existence of several potential Yki partners is consistent with the idea that additional Hpo-pathway targets await discov-ery. This may be especially true for factors involved in cell cycle control. Increasing CycE levels results in an accelerated cell cycle in G1 but also a compensatory lengthening of S phase9. In contrast, Yki-induced proliferation is characterized by a global acceleration of the cell cycle5,6. These observations indicate that cell cycle regulators other than CycE must also be under Hpo- pathway control.
The publications reviewed here do much more than illuminate the mechanisms involved in the control of animal growth and shape: they pose many exciting new questions. How are Ex and Mer molecularly connected with the Hpo-pathway core? It seems that Mer participates in the pathway parallel to Ft and Ex10,16. Does Mer interact with a specific transmembrane receptor, as Ex interacts with Ft? Although this might be the case, an alternative possibility is that Mer functions as a constitutively active compo-nent of the pathway. In this way, Mer would confer an ever-present, albeit mild, brake on cell proliferation and an intrinsic sensitivity to apoptotic signals.
It is likely that we have only just begun to appreciate the full complexity of this system of growth control. The initial pic-ture of a core Hpo pathway responsible for controlling epithelial growth is now elaborated with feedback loops, input from developmental cues and environmental challenges. All these elaborations ensure that brakes and accelerators of prolifera-tion and apoptosis are in a developmentally regulated equilibrium within the cell. Recent work suggests that the Hpo pathway may not be strictly limited to growth-control func-tions. For example, it has been shown that DNA damage results in the activation of the Hpo pathway through p53-dependent Hpo phosphorylation, making damaged cells more susceptible to elimination by apoptosis13. Ft has a role in PCP and patterning along the axis of fly appendages23,24. Wts participates in the choice of which rhodopsin is expressed during photoreceptor maturation29. Ex also has a role in PCP30 and, together with Mer, may have a general role in membrane recycling and thus affect several signaling pathways in addition to Hpo31. In addition, genes such as hpo, wts and yki also participate in cell-shape control2,6. Doubtless, future research will uncover many more magic
tricks played by nature in shaping organisms, from fruitflies to hippos to humans.
1. Xu, T., Wang, W., Zhang, S., Stewart, R.A. & Yu, W. Development 121, 1053–1063 (1995).
2. Justice, R.W., Zilian, O., Woods, D.F., Noll, M. & Bryant, P.J. Genes Dev. 9, 534–546 (1995).
3. Tapon, N. et al. Cell 110, 467–478 (2002).4. Harvey, K.F., Pfleger, C.M. & Hariharan, I.K. Cell 114,
457–467 (2003).5. Wu, S., Huang, J., Dong, J. & Pan, D. Cell 114, 445–
456 (2003).6. Huang, J., Wu, S., Barrera, J., Matthews, K. & Pan, D.
Cell 122, 421–434 (2005).7. Lai, Z.C. et al. Cell 120, 675–685 (2005).8. Edgar, B.A. Cell 124, 267–273 (2006).9. Neufeld, T.P., de la Cruz, A.F., Johnston, L.A. & Edgar,
B.A. Cell 93, 1183–1193 (1998).10. Willecke, M. et al. Curr. Biol. 16, 2090–2100
(2006).11. Bennett, F.C. & Harvey, K.F. Curr. Biol. 16, 2101–2110
(2006).12. Cho, E. et al. Nat. Genet. 38, 1142–1150 (2006).13. Colombani, J., Polesello, C., Josue, F. & Tapon, N. Curr.
Biol. 16, 1453–1458 (2006).14. Hamaratoglu, F. et al. Nat. Cell Biol. 8, 27–36
(2006).15. Nolo, R., Morrison, C.M., Tao, C., Zhang, X. & Halder, G.
Curr. Biol. 16, 1895–1904 (2006).16. Silva, E., Tsatskis, Y., Gardano, L., Tapon, N. & McNeill, H.
Curr. Biol. 16, 2081–2089 (2006).17. Thompson, B.J. & Cohen, S.M. Cell 126, 767–774
(2006).18. McCartney, B.M., Kulikauskas, R.M., LaJeunesse,
D.R. & Fehon, R.G. Development 127, 1315–1324 (2000).
19. Bryant, P.J., Huettner, B., Held, L.I., Jr. Ryerse, J. & Szidonya, J. Dev. Biol. 129, 541–554 (1988).
20. Mao, Y. et al. Development 133, 2539–2551 (2006). 21. Zilian, O. et al. Development 126, 5409–5420
(1999).22. Matakatsu, H. & Blair, S.S. Development 133, 2315–
2324 (2006).23. Matakatsu, H. & Blair, S.S. Development 131, 3785–
3794 (2004).24. Cho, E. & Irvine, K.D. Development 131, 4489–4500
(2004).25. Strutt, H., Mundy, J., Hofstra, K. & Strutt, D.
Development 131, 881–890 (2004).26. Brennecke, J., Hipfner, D.R., Stark, A., Russell, R.B.
& Cohen, S.M. Cell 113, 25–36 (2003).27. Hipfner, D.R., Weigmann, K. & Cohen, S.M. Genetics
161, 1527–1537 (2002).28. Raisin, S., Pantalacci, S., Breittmayer, J.P. & Leopold, P.
Genetics 164, 1015–1025 (2003).29. Mikeladze-Dvali, T. et al. Cell 122, 775–787 (2005).30. Blaumueller, C.M. & Mlodzik, M. Mech. Dev. 92, 251–
262 (2000).31. Maitra, S., Kulikauskas, R.M., Gavilan, H. & Fehon,
R.G. Curr. Biol. 16, 702–709 (2006).
N E W S A N D V I E W S©
2006
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
smb