observations on the histochemical distribution of alcohol
TRANSCRIPT

289
Observations on the histochemical distribution of alcoholdehydrogenase
By M. M. FERGUSON
(From the Department of Anatomy, The University, Glasgow, W.2)
With 1 plate
SummaryAlcohol dehydrogenase capable of utilizing primary alcohols (straight and branchedchain, unsaturated and cellosolves), secondary alcohols, polyhydric alcohols but nottertiary alcohols is described in the rat brain, lung, heart, liver, kidney, gut, spleen,pancreas, uterus, and seminal vesicle. With the straight chain primary alcohols theextent to which the alcohols were used increased relative to increasing chain lengthuntil the optimum length of six to seven carbon atoms had been reached, whereafteractivity decreased, probably due to the insolubility of the higher members of theseries in water. Side groups on the chain, double bonds and ethereal oxygen atoms, allhad the effect of hindering the ease of dehydrogenation of the alcohol. Secondaryalcohols were suitable as substrates; however, tertiary alcohols did not seem to besatisfactory for the demonstration of alcohol dehydrogenase. Polyhydric alcohols weresuitable substrates although they were not as well used as were the monohydricalcohols. Furfuryl alcohol dehydrogenase was found in all tissues and is apparentlya separate enzyme.
Introduction
HISTOCHEMICAL literature contains relatively little information on thedistribution of alcohol dehydrogenase in the body (Pearse, i960; Burstone,1962). Studies have been mainly confined to the retina (Kuwabara andCogan, i960; Pearse, 1961; Niemi and Merenmies, 1961), although Pearse(i960) also noted strong activity in rat parietal cells and salivary duct cellswhich were stated as being exceptional relative to other tissues. The substratenormally employed in these studies is ethanol.
The present histochemical investigation into alcohol dehydrogenase wasundertaken to acquire further information regarding the location and activityof the enzyme(s) in tissues.
Material and methodsRats of the Royal Wistar albino strain were killed by decapitation; samples
of about 5 mm3 of brain, lung, heart, liver, spleen, kidney, pancreas, gut,seminal vesicle, and uterus were removed and immediately frozen on solidcarbon dioxide. They were sectioned at 12/x in a cryostat maintained at—25° C and the sections attached to clean, dry glass coverslips by momentarythawing. Each section was incubated for 1 h at 380 C in a phosphate buffer(pH 7-4) medium containing nicotinamide adenine dinucleotide (NAD),nicotinamide, magnesium sulphate, and Nitro B.T. The reduced NAD
[Quart. J. micr. Sci., Vol. 106, pt. 4, pp. 289-97, 1965.]

290 Ferguson—Distribution of alcohol dehydrogenase
formed was coupled to the soluble Nitro B.T. which on reduction formed aninsoluble precipitate of formazan at the site of enzymic activity in the tissue.Pink monoformazan was considered to indicate weak activity and blue di-formazan stronger activity. The density of the formazan deposit was judgedvisually and slight variations could not be measured. The following alcoholswere added to give a concentration in the mixture of 5%: methanol, ethanol,n-propanol, n-butanol, n-pentanol, n-hexanol, n-heptanol, n-octanol,n-nonanol, n-decanol, n-undecanol, n-dodecanol, n-tridecanol, isopropanol,secondary butanol, pentan-2-ol, octan-2-ol, pentan-3-ol, isobutanol, tertiarybutanol, 2-methyl butan-i-ol, 3-methyl butan-i-ol, tertiary butyl carbinol,nonyl alcohol, pinacol, phenol, hydroquinone, allyl alcohol, cyclohexanol, 2-ethoxyethanol, furfuryl alcohol, ethylene glycol, propylene glycol, glycerol,butan-i :3-diol, butan-i :4-diol. Control sections were incubated in thebuffered medium which (a) contained no alcohol, or (b) contained alcoholbut no NAD. To investigate inhibition, hydroxylamine and p-chloro-mercuribenzoic acid were added to all the media in a concurrent set ofincubations.
ResultsThe results obtained with each tissue and substrate are summarized in
table 1. Some typical photomicrographs are shown in figs. 1 to 4.Brain
The brain was poorly reactive with only traces of diformazan beingdeposited after incubation with octan-2-ol and 2-ethoxyethanol. However,furfuryl alcohol was relatively well used. Enzymic activity was evidentthroughout the sections and nerve cells could not be differentiated fromneuroglia.Lung
No activity could be demonstrated when sections were incubated withmethanol, but ethanol, propanol, and butanol were utilized to a slight extentwith light deposits of formazan in the alveolar walls. Pentanol, hexanol, andheptanol were all well used by the alveolar walls where diformazan was heavilydeposited. Octanol produced only a slight deposition of formazan and non-anol, decanol, undecanol, dodecanol, and tridecanol were not used. Octan-2-ol, 2-methyl butan-i-ol, and 3-methyl butan-i-ol were suitable substrateswhereas only a trace of diformazan was deposited when sections of lung were
FIG. I. Bronchial wall showing intense alcohol dehydrogenase activity in the respiratoryepithelium with hexanol as substrate. Diformazan also deposited in the adjacent alveolartissue. X 375.
FIG. 2. With heptanol as substrate, renal tubules containing fairly heavy deposition ofdiformazan. No significant deposit in the glomeruli. x 120.
FIG. 3. Diformazan deposited in the intestinal epithelium with pentanol as substrate.Less activity is demonstrated in submucosa, and the muscular coats have no significantdeposits of diformazan. x 90.
FIG. 4. Seminal vesicle incubated with furfuryl alcohol showing intense activity, especiallyin the epithelium. X 120.
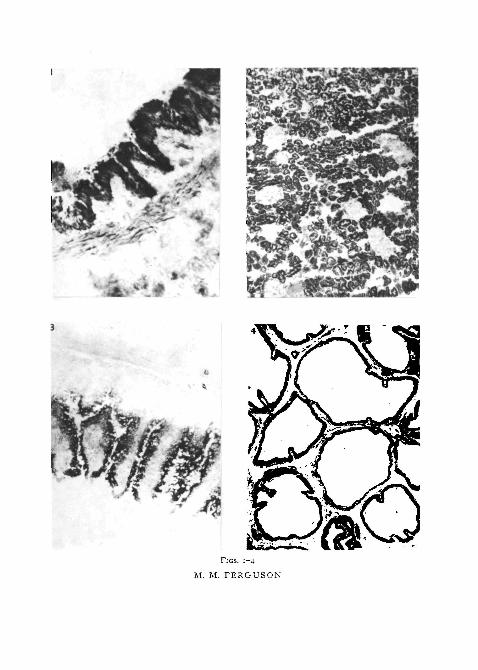
FIGS. 1-4
M. M. FERGUSON

Ferguson—Distribution of alcohol dehydrogenase 291
incubated with isopropanol, allyl alcohol, 2-ethoxyethanol, and propyleneglycol. Furfuryl alcohol was extremely well used, as indicated by a densediformazan deposition in the alveolar walls. The remaining alcohols did notseem to be utilized to any significant extent.
Heart
Methanol was not used by any of the cardiac tissue. Ethanol and propanolwere poorly used but butanol, pentanol, hexanol, heptanol, and octanol wereall well utilized, especially hexanol, with a heavy deposition of diformazanin the endocardium and myocardium. Activity was also demonstrable withoctan-2-ol and 3-methyl butan-i-ol, and trace amounts of diformazan weredeposited in the heart wall when isopropanol, pentan-2-ol, pentan-3-ol,2-methyl butan-i-ol, nonyl alcohol, allyl alcohol, propylene glycol, and2-ethoxyethanol were used as substrates. Intense deposits of diformazan wereseen in all layers of the heart wall on incubation with furfuryl alcohol.
Liver
The liver did not appear to utilize methanol. Ethanol, propanol, and octanolwere moderately well utilized by the hepatic cells whereas butanol, pentanol,hexanol, and heptanol were extremely well used with an even, dense depo-sition of diformazan throughout the substance of the liver. Moderatedeposits of diformazan were observed when sections of liver were incubatedwith isopropanol, pentan-2-ol, octan-2-ol, pentan-3-ol, isobutanol, 2-methylbutan-i-ol, 3-methyl butan-i-ol, nonyl alcohol, allyl alcohol, propyleneglycol, and furfuryl alcohol. Trace activity was further demonstrable withcyclohexanol, ethylene glycol, butan-i: 3-diol, and butan-i :4-diol as sub-strates.
Kidney
With each of the substrates utilized by the kidney the sites of alcoholdehydrogenase activity seemed to be similar; the glomeruli contained little ornone, but the surrounding tubules in the cortex appeared to possess moderateactivity with heaviest deposition of diformazan in the inner zone of cortexadjacent to the medulla, whereas the medulla itself usually had only slightactivity. Methanol was not used by the kidney to any significant extent.Ethanol was moderately utilized; however, propanol, butanol, pentanol,heptanol, and octanol were more satisfactory substrates, producing a fairlyheavy deposition of diformazan. Pentanol was the only alcohol observed tocause a trace of diformazan to be deposited in the glomeruli. Octan-2-ol, nonylalcohol, and propylene glycol were moderately well used but pentan-2-ol,3-methyl butan-i-ol, and ethylene glycol were only poorly utilized. Furfurylalcohol was well used throughout cortical tissue, and to a lesser extent inglomeruli and medulla, producing an intense deposition of diformazan.

292 Ferguson—Distribution of alcohol dehydrogenase
Gut
Sections of gut incubated in methanol produced no deposit of diformazan.Alcohol dehydrogenase activity was demonstrable with ethanol, propanol,butanol, pentanol, hexanol, heptanol, and octanol as substrates. Diformazanwas heavily deposited in the intestinal epithelium with moderate deposits inthe lamina propria and submucosa. Brunner's glands in duodenal sectionsalso possessed alcohol dehydrogenase activity. The muscular coats onlycontained light deposits of diformazan with these alcohols. Isopropanol,octan-2-ol, pentan-3-ol, isobutanol, 2-methyl butan-i-ol, 3-methyl butan-i-ol, propylene glycol, and 2-ethoxyethanol were all moderately well usedin the same sites; also, there was a trace of activity demonstrable on incubatingsections in nonyl alcohol, ethylene glycol, allyl alcohol, butan-i ^-diol, andbutan-i :4-diol. Furfuryl alcohol was well used by the intestines and thedeposition of diformazan in the muscular coats appeared to be slightly moreintense than with the other alcohols.
Spleen
The spleen did not use methanol as a substrate and ethanol only to a slightextent. Propanol and octanol were only poorly utilized but butanol, pentanol,hexanol, and heptanol were satisfactory as substrates for the demonstrationof alcohol dehydrogenase in the spleen. The capsule was not observed tocontain significant formazan deposits; the red and white pulp seemed to beequally active. Octan-2-ol and 3-methyl butan-i-ol were also fairly wellutilized by the splenic pulp, whereas with isopropanol, pentan-2-ol, pentan-3-ol, allyl alcohol, 2-methyl butan-i-ol, propylene glycol, and nonyl alcoholas substrates only traces of diformazan were seen. The spleen used furfurylalcohol extremely well, with heavy deposits of diformazan throughout.
Pancreas
The control sections of pancreas developed a colour within 15 min ofincubation, thus rendering it very difficult to record observations in sectionswhich contained anything less than an intense deposition of diformazan.Diformazan was much more heavily deposited than in control sections whensections were incubated with propanol, butanol, pentanol, hexanol, heptanol,and isopropanol. Activity appeared to be more intense in the exocrine tissuerather than in the islets. Furfuryl alcohol was also well utilized.
UterusThe endometrium was observed to contain a heavier deposit of diformazan
than did the myometrium when incubated with the substrates which wereused. Ethanol and octanol were poorly utilized in comparison to propanol,butanol, pentanol, heptanol, and especially hexanol, which were all wellused. A light deposit of diformazan was further seen when uterine sectionswere incubated in 2-methyl butan-i-ol, 3-methyl butan-i-ol, allyl alcohol,

Ferguson—Distribution of alcohol dehydrogenase 293
propylene glycol, 2-ethoxyethanol, and butan-i :4-diol. Furfuryl alcohol wasalso well used.
Seminal vesicle
Alcohol dehydrogenase was readily demonstrable in the epithelium of theseminal vesicle and to a much lesser extent in the lamina propria and muscularcoat. Ethanol and octanol were poorly utilized by the seminal vesicle whereas
TABLE I
Alcohol dehydrogenase activity of various tissues
MethanolEthanolPropanolButanolPentanolHexanolHeptanolOctanolNonanolDecanolUndecanolDodecanolTridecanolIsopropanolSec. butanolPentan-2-olOclan-2-olPentan-3-olIsobutanolTen. butanol3-Methyl
butan-i-ol3-Methyl
butan-i-olTert. butyl
carbinolNonyl
alcoholPinacolPhenolHydroquinoneAtlyl alcoholCyclohexanoli-ethoxy-
ethanolFurfuryl
alcoholEthylene
glycolPropylene
glycolClycerolButan-i :3-
diolSutan-T :4-
dial
Brain
0
a0
0
0
0
0
0
0
0
0
0
0
0
00
trace0
0
0
0
0
0
0
0
0
0
0
0
trace
+0
0
0
0
0
Lung
o
tracetracetrace+++
trace0
O
O
0
O
trace0
0
+0
0
0
+
+
0
0
0
0
0
trace0
trace
+ +0
trace
0
0
0
Heart
0
tracetrace++
+ +++0
0
0
0
0
trace0
trace+
trace0
0
trace
+0
trace
0
0
0
trace0
trace
+ +0
trace
0
0
0
Liver
o
++
+ ++ ++ +++
trace000
0
+0
++++0
+
+
0
+0
0
0
+trace
++
trace
+
otrace
trace
Kidney
o
trace
++++++0
0
0
0
0
0
0
trace+0
0
0
trace
+
+0
0
0
0
0
0
+ +trace
trace
0
0
0
Cut
0
++++
+ +++
trace0
0
0
0
+0
0
+++0
+
+0
trace
0
0
0
trace0
++ +trace
+
+trace
trace
Spleen
0
tracetrace
++++
trace0
0
0
0
0
trace0
trace+
trace0
0
trace
+0
trace
0
0
0
trace0
0
+ +a
trace
0
0
0
Pancreas
++++
+
+ +
Uterus
o
trace
+++
+ ++
trace0
0
0
0
0
0
0
0
0
0
0
0
trace
+
o
trace
0
0
0
trace0
trace
+0
trace
0
0
trace
Seminalvesicle
o
+++++
tracetrace
0
0
0
O
0
0
0
0
trace0
trace0
trace
trace
0
0
0
0
0
+•
trace
+0
+0
0
0
moderate activity, indicated by a heavier deposition of diformazan, was shownwhen propanol, butanol, pentanol, hexanol, and heptanol were used assubstrates. Allyl alcohol and furfuryl alcohol were also quite well utilizedbut when sections were incubated with propylene glycol, octan-2-ol, 2-methyl

294 Ferguson—Distribution of alcohol dehydrogenase
butan-i-ol, 3-methyl butan-i-ol, isobutanol, and 2-ethoxyethanol only atrace of diformazan was deposited.
DiscussionThe enzyme alcohol dehydrogenase catalyses the inter-conversion of an
alcohol with its corresponding aldehyde,ADH
R.CH2OH+NAD+ ^ R.CHO+NADH+H+and the mechanism of the reaction has been extensively described (Theorelland Bonnichsen, 1951; Theorell and Chance, 1951; Theorell, Nygaard, andBonnichsen, 1955; Theorell, 1958; Winer and Theorell, i960; Theorell, 1961;Theorell and McKinley-McKee, 1961; Dalziel, 1961; Plane and Theorell,1961; Snyder, Vogel, and Schulman, 1963; Mahler, Baker, and Sherer, 1962;Dalziel, 1962; Yonetani, 1963; Plane and Long, 1963; and Dalziel, 1963).
Alcohol dehydrogenase was originally isolated from yeast (Neglein andWulff, 1937; Racker, 1950) and later from horse liver (Bonnichsen, 1951).This enzyme has also been described in species of Pseudomonas (Baptist andCoon, 1959; Azoulay and Heydeman, 1963). Although related in action andbasic structure, alcohol dehydrogenases from these various sources exhibitdifferent characteristics. Enzymes from each source contain zinc (Vallee andHoch, 1957) and also have active sulphydryl groups of which liver alcoholdehydrogenase is thought to have twenty-eight (Witter, i960). The liveralcohol dehydrogenase molecule is now believed to consist of two sub-unitswith a compound molecular weight of 84,000 (Dalziel, 1963; Wratten andCleland, 1963) whereas yeast alcohol dehydrogenase, consisting of four sub-units has a combined molecular weight in the region of 150,000 (Wratten andCleland, 1963). Both enzymes are soluble and can be inhibited by generalsulphydryl inhibitors such as hydroxylamine or p-chloromercuribenzoicacid (Theorell and Bonnichsen, 1951; Vallee and Hoch, 1957; Li, Ulmer, andVallee, 1962) or by 1 :io phenanthroline which binds with the zinc in themolecule (Snyder, Vogel, and Schulman, 1963). A further point of com-parison is that the yeast alcohol dehydrogenase tends to favour the conversionof aldehyde to alcohol whereas liver alcohol dehydrogenase favours a shiftin the opposite direction (Wratten and Cleland, 1963). Alcohol dehydrogenasehas also been demonstrated biochemically in kidney, gut, testis, and lung inaddition to liver (Deuel, 1957; Zackman and Olson, 1961).
Wald and Hubbard (1948-9) described a retinene reductase in the retinacapable of converting retinene to vitamin A and this was later found to bevery similar to liver alcohol dehydrogenase (Hubbard and Wald, 1951) in itsaction, although yeast alcohol dehydrogenase was incapable of catalysing thisreaction. Glover, Goodwin, and Morton (1948) described an alcohol de-hydrogenase in gut wall capable of converting the retinene formed fromcarotene into vitamin A. However, Zackman and Olson (1961) on comparingretinene reductase and alcohol dehydrogenase from various organs came tothe conclusion that these were two similar but nevertheless different enzymes.

Ferguson—Distribution of alcohol dehydrogenase 295
Primary alcohols are oxidized by most aerobic cells with increasing easeas the chain length increases (Schueler, i960), resulting in the eventualliberation of water and carbon dioxide. Secondary alcohols tend to beoxidized to the corresponding ketones (Merrit and Tomkins, 1959), e.g. con-version of isopropanol to acetone, but tertiary alcohols were not well oxidizedand are usually excreted conjugated with glucuronic acid. Different groupsof alcohols were investigated in this series :
Straight chain alcohols (methanol to tridecanol). In the present seriesutilization of the straight chain alcohols improved as chain length increased,reaching an optimum about hexanol. Methanol was not utilized in any tissue.After this, activity dropped but as the incubations were carried out in anaqueous medium and the longer chain alcohols became progressively lesssoluble, it is probable that solubility factors influence the results. This histo-chemical technique is less sensitive in that very slight activity which may berecorded spectrophotometrically may fail to produce a significant depositionof diformazan. It is perplexing to find that the longer chain alcohols are moretoxic than ethanol but yet utilized much better; there is an increase in toxicityto nerve cells as the chain length increases (Jenkins, Hartung, Hamlin, andData, 1957).
Aromatic alcohols (phenol, hydroquinone). Neither of these substances wasused by the tissues, and this would be expected due to the relationship of thehydroxyl group to the rest of the molecule. Instead of oxidation they areusually excreted as glucuronides.
Cellosolves (2-ethoxyethanol). This alcohol was only moderately well usedby a variety of tissues but the ethereal oxygen atom obviously impedes itsdehydrogenation, when compared to butanol and pentanol, with similar chainlengths.
Unsaturated alcohols (allyl alcohol). The double bond in this moleculeappeared to impede dehydrogenation.
Branched alcohols (isobutanol, 2-methyl butan-i-ol, 3-methyl butan-i-ol,nonyl alcohol, tertiary butanol, tertiary butyl carbinol). The two tertiaryalcohols were not used to any significant extent and the remainder were onlypoorly used in comparison to the corresponding straight chain alcohol.It would appear from this that the altered configuration of the moleculeaffects substrate-enzyme binding. It is relevant to note in this connexion thatthe better utilization of 3-methyl butan-i-ol compared to 2-methyl butan-i-ol suggests that the nearer the branched methyl group is to the hydroxylgroup the greater is the degree of steric hindrance.
Secondary alcohols (isopropanol, secondary butanol, pentan-2-ol, pentan-3-01, octan-2-ol). Again with these alcohol substrates the longer-chainedoctan-2-ol is better utilized than the shorter pentan-2-ol, but this in turnis better used than secondary butanol. Although in the present study noobvious differences were found in the tissue location of alcohol dehydrogenasecapable of utilizing primary or secondary alcohols, differences have beenobserved in the rodent ovary (unpublished observations), and in the light of

296 Ferguson—Distribution of alcohol dehydrogenase
these observations it seems fair to postulate the existence of more than onealcohol dehydrogenase. Retinene reductase, if it is indeed a different enzymefrom primary alcohol dehydrogenase as stated by Zackman and Olson (1961),may be another member of this group of enzymes.
Cyclohexanol. Cyclohexanol has been recorded as being utilized by Pseudo-monas (Jacoby and Frederiks, 1962) and liver alcohol dehydrogenase (Merritand Tomkins, 1959), but apart from a trace of diformazan deposited in liversections, a dehydrogenase capable of this action could not be demonstratedhistochemically. The possibility of this being yet another alcohol dehydro-genase requiring slightly different conditions needs more detailed investi-gation; on the other hand, the inactivity could be ascribed to the relativeinsolubility of cyclohexanol in water.
Polyhydric alcohols (ethylene glycol, propylene glycol, glycerol, butan-i :3-diol, butan-i :4-diol, pinacol). These polyhydric alcohols in general do notseem to be well used by any tissue examined.
Furfuryl alcohol. Furfuryl alcohol is structurally unrelated to the otheralcohols examined and is considered to be a by-product of pentose meta-bolism. This step of dehydrogenation appeared to occur in all tissues and itseems probable that this enzyme is connected with normal cellular meta-bolism.
The straight-chain alcohols which were well used by many tissues are alsotoxic and it is interesting to note that fatty acids and alcohols compete forbinding sites in the formation of dissociable ternary complexes with thealcohol dehydrogenase and NAD and further, there is also an affinity forlonger-chain fatty acids (Winer and Theorell, i960; Yonetani and Theorell,1962). Fatty acid amides and aldehydes also similarly compete.
Hydroxy fatty acids are dehydrogenated by a similar NAD-dependentdehydrogenase (Mintz and Heinrikson, 1961) in connexion with omega oxi-dation. Therefore, one method by which longer-chain alcohols could exerta toxic effect is by complexing with these enzymes due to the similarity ofmolecular structure, thus interfering with normal fatty acid metabolism.However, there is no supporting evidence for this.
Control sections of some tissues, notably pancreas, developed a depositof diformazan after a while even though no substrate was added. The possi-bility exists that the tissue could contain intrinsic substrates capable of beingdehydrogenated and thus producing a deposit of diformazan.
Of all the tissues examined, brain was the least active except with furfurylalcohol, the significance of which has already been discussed. NAD lipo-amide oxidoreductase, succinic dehydrogenase (Nachlas, Tsou, De Souza,Cheng, and Seligman, 1957), malate, lactate, and a-glycerophosphate de-hydrogenases (Chason and Pearse, 1961) have all been described in brain butChason and Pearse (1961) point out that a high concentration of nicotinamideis required to protect the NAD against the diphosphopyridine nucleotidasefound in high concentrations in the brain and this may be a causative factorin obtaining the poor results.

Ferguson—Distribution of alcohol dehydrogenase 297
The fact that the brain is not capable of metabolizing the various alcoholsto any significant extent may possibly be a reason why their toxicity manifestsitself primarily in nervous tissues.
The author acknowledges research facilities provided by the Anatomy Department,Glasgow University.
ReferencesAZOULAY, E., and HEYDEMAN, M. T., 1963. Biochim. biophys. Acta, 73, 1.BAPTIST, J. N., and COON, M. J., 1959. Amer. chem. Soc. Boston Meetings., Div. Biol.
Chein., Comm. No. 19.BONNICHSEN, R. K., 1951. Acta chem. scand., 4, 715.BURSTONE, M. S., 1962. In Enzyme histochemistry, Ch. 12. New York (Academic Press).CHASON, J. L., and PEARSE, A. G. E., 1961. J. Neurochem., 6, 259.DALZIEL, K., 1961. Biochem. J., 80, 440.DALZIEL, K., 1962. Biochem. J., 84, 244.DALZIEL, K., 1963. J. biol. Chem., 238, 2850.DEUEL, H. J., 1957. In Lipids, Vol. iii. New York (Interscience Publishers).GLOVER, J., GOODWIN, T. W., and MORTON, R. A., 1948. Biochem. J., 43, 109 and 512.HUBBARD, R., and WALD, G., 1951. Proc. nat. Acad. Sci. U.S.A., 37, 69.JACOBY, W. B., and FREDERIKS, J., 1962. Biochim. biophys. Acta., 58, 217.JENKINS, G. L., HARTUNG, W. H., HAMLIN, K. E., and DATA, J. B., 1957. In The chemistry
of organic medicinal products. New York (John Wiley and Sons).KUWABARA, T., and COGAN, D. G., i960. J. Histochem. Cytochem., 8, 214.Li, T. K., ULMER, D. D., and VALLEE, B. L., 1962. Biochem., i, 114.MAHLER, H. R., BAKER, R. H., and SHERER, V. J., 1962. Biochem., 1, 41 and 47.MERRIT, A. D., and TOMKINS, G. M., 1959. J. biol. Chem., 234, 2778.MINTZ, M. A., and HEINRIKSON, R. L., 1961. Biochim. biophys. Acta, 46, 45.NACHLAS, M. M., TSOU, K. C, SOUZA, E. DE, CHENG, C. S., and SELIGMAN, A. M., 1957.
J. Histochem. Cytochem., 5, 420.NEGLEIN, E., and WULFF, H. J., 1937. Biochem. Z., 289, 436.NIEMI, M., and MERENMIES, E., 1961. J. Neurochem., 6, 206.PEARSE, A. G. E., i960. In Histochemistry theoretical and applied, Ch. 20. London (Churchill
Ltd.).PEARSE, A. G. E., 1961. In The structure of the eye. London (Academic Press).PLANE, R. A., and LONG, T. V., 1963. Acta chem. scand., 17, Suppl. 91.PLANE, R. A., and THEORELL, H., 10.61. Acta chem. scand., 15, 1797.RACKER, E., 1950. J. biol. Chem., 184, 313.SCHUELER, F. W., i960. In Chemodynamks and drug design, pp. 267-70. New York (McGraw-
Hill Book Co.).SNYDER, R., VOGEL, W. H., and SCHULMAN, P., 1963. Biochem. Biophys. Res. Comm., 13,
335-THEORELL, H., 1958. Advanc. Enzymol., 20, 31.THEORELL, H., 1961. Fed. Proc, 20, 967.THEORELL, H., and BONNICHSEN, R. K., 1951. Acta chem. scand., 5, 1105.THEORELL, H., and CHANCE, B., 1951. Acta chem. scand., 5, 1127.THEORELL, H., and MCKINLEY-MCKEE, J. S. M., 1961. Nature, Lond., 192, 47.THEORELL, H., NYGAARD, A. P., and BONNICHSEN, R. K., 1955. Acta chem. scand., 9, 1148.VALLEE, B. L., and HOCH, F. L., 1957. J. biol. Chem., 225, 185.WALD, G., and HUBBARD, R., 1948-9. J. gen. Physiol., 32, 367.WINER, A. D., and THEORELL, H., i960. Acta chem. scand., 14, 1729.WITTER, A., i960. Acta chem. scand., 14, 1717.WRATTEN, C. C, and CLELAND, W. W., 1963. Biochem., 2, 935.YONETANI, T., 1963. Acta chem. scand., 17, Suppl. 16.YONETANI, T., and THEORELL, H., 1962. Arch. Biochem., 99, 433.ZACKMAN, R. D., and OLSON, J. A., 1961. J. biol. Chem., 236, 2309.