noxe nadh oxidase and the electron transport chain are ... · the electron transport chain (etc)...
TRANSCRIPT

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Mar. 2010, p. 1311–1319 Vol. 76, No. 50099-2240/10/$12.00 doi:10.1128/AEM.02120-09Copyright © 2010, American Society for Microbiology. All Rights Reserved.
NoxE NADH Oxidase and the Electron Transport Chain AreResponsible for the Ability of Lactococcus lactis To
Decrease the Redox Potential of Milk�†Sybille Tachon,1 Johannes Bernhard Brandsma,2 and Mireille Yvon1*
INRA, UMR 1319 Micalis, Domaine de Vilvert, F-78352 Jouy-en-Josas, France,1 and CSK Food Enrichment, Pallasweg 1,8901 BA Leeuwarden, the Netherlands2
Received 2 September 2009/Accepted 15 December 2009
The redox potential plays a major role in the microbial and sensorial quality of fermented dairy products.The redox potential of milk (around 400 mV) is mainly due to the presence of oxygen and many other oxidizingcompounds. Lactococcus lactis has a strong ability to decrease the redox potential of milk to a negative value(�220 mV), but the molecular mechanisms of milk reduction have never been addressed. In this study, weinvestigated the impact of inactivation of genes encoding NADH oxidases (noxE and ahpF) and components ofthe electron transport chain (ETC) (menC and noxAB) on the ability of L. lactis to decrease the redox potentialof ultrahigh-temperature (UHT) skim milk during growth under aerobic and anaerobic conditions. Our resultsrevealed that elimination of oxygen is required for milk reduction and that NoxE is mainly responsible for therapid removal of oxygen from milk before the exponential growth phase. The ETC also contributes slightly tooxygen consumption, especially during the stationary growth phase. We also demonstrated that the ETC isresponsible for the decrease in the milk redox potential from 300 mV to �220 mV when the oxygen concen-tration reaches zero or under anaerobic conditions. This suggests that the ETC is responsible for the reductionof oxidizing compounds other than oxygen. Moreover, we found great diversity in the reducing activities ofnatural L. lactis strains originating from the dairy environment. This diversity allows selection of specificstrains that can be used to modulate the redox potential of fermented dairy products to optimize theirmicrobial and sensorial qualities.
Lactococcus lactis is a lactic acid bacterium that is widelyused as a starter in the dairy industry. Its major role is to acidifymilk, but it is also one of the most reducing lactic acid bacteriaand produces a very negative redox potential (Eh) value in milk(5, 8). The Eh is indicative of the ability of a system to donateor accept electrons and is strongly dependent on levels ofdissolved oxygen (39). The Eh affects the microbiota of manydairy products. For instance, it affects the development andactivity of the secondary flora in cheese (especially nonstarterlactic acid bacteria) (4, 33) and the viability of probiotic bac-teria, such as Lactobacillus acidophilus or bifidobacteria, inyogurt or milk (3, 10–12, 20, 27). Moreover, the Eh affects theproduction and stability of aroma compounds (16, 23, 24).Oxygen is also responsible for formation of the brown discol-oration that affects some cheeses and results from the oxida-tion of a tyrosine catabolite produced by certain yeasts (9).Finally, the oxygen in the medium and/or the Eh affects thedevelopment of spoilage microorganisms (7) and the expres-sion of virulence factors in some pathogens, such as Bacteroidesfragilis or Staphylococcus aureus (6, 14, 37). The Eh and oxygenthus play a major role in the microbial and sensorial qualitiesof fermented dairy products.
The ability of L. lactis to decrease the Eh of milk is thought
to be related mainly to the reduction of O2 (21). Despite thefact that L. lactis has the genes encoding the electron transportchain (ETC), it does not respire unless heme is added to thegrowth medium, because it lacks heme biosynthesis genes (15).In the absence of heme, the reduction of O2 to H2O is likelydue to the NADH oxidase (NOX) NoxE (21, 26). L. lactis alsocontains the alkyl hydroperoxide reductase (AhpR) system,which is composed of an H2O2-forming NADH oxidase(AhpF) and a peroxyredoxine (AhpC) that reduces H2O2 toH2O (17). The AhpR system may also participate in reductionof oxygen to H2O. However, the Eh of oxygen-free milk (sat-urated with nitrogen) is still positive (21), while the Eh of milkfermented with L. lactis is close to �220 mV (8, 21). Thissuggests that, in addition to oxygen, L. lactis reduces otheroxidizing compounds present in milk. We recently showed thatthe functional part of the electron transport chain (ETC) of L.lactis, which is essentially composed of menaquinones andmembrane NADH dehydrogenases (NoxA and NoxB), is re-sponsible for the reduction of tetrazolium violet (TV) to form-azan when TV is added to milk (38). NoxA and/or NoxBtransfers electrons from intracellular NADH to cell mem-brane-associated menaquinones, which reduce TV at the cellsurface or in the cell membrane. Rezaïki et al. (35) also showedthat menaquinones are involved in the reduction of O2, Fe3�,and, with considerable efficiency, Cu2�. These results suggestthat the ETC may be involved in the reduction of milk, whichcontains metal cations (USDA National Nutrient Database forStandard Reference).
Despite the importance of the Eh for the quality of dairyproducts, the molecular mechanisms involved in milk reduc-
* Corresponding author. Mailing address: Micalis Institute, INRAUMR 1319, Domaine de Vilvert, 78352 Jouy-en-Josas cedex, France.Phone: 33 1 34 65 21 59. Fax: 33 1 34 65 21 63. E-mail: [email protected].
† Supplemental material for this article may be found at http://aem.asm.org/.
� Published ahead of print on 28 December 2009.
1311
on May 23, 2020 by guest
http://aem.asm
.org/D
ownloaded from

tion by L. lactis have never been addressed. In the presentstudy, we investigated the impact of inactivation of genes en-coding NADH oxidases (noxE and ahpF) and components ofthe ETC (menC and noxAB) on the reduction of ultrahigh-temperature (UHT) skim milk by L. lactis. Our results showedthat two distinct and complementary mechanisms are involvedin milk reduction. The first mechanism is essential and relieson the elimination of dissolved oxygen, principally by the NoxENADH oxidase. The second mechanism is oxygen independentand is due to the ETC, which probably reduces oxidizing com-pounds other than oxygen. Moreover, we found great diversityin the reducing activities of natural L. lactis strains, whichmakes control of the Eh in fermented dairy products throughselection and use of strains that reduce more or less conceiv-able.
MATERIALS AND METHODS
Bacterial strains and culture conditions. All L. lactis strains used in this studyare L. lactis subsp. cremoris strains. L. lactis mutant strains were derived fromlaboratory strain TIL46 and are listed in Table 1. The noxAB mutant wasconstructed previously (38), and the menC mutant was obtained previously byinsertion mutagenesis and selection for an inability to reduce tetrazolium violet(38). Other wild-type (WT) L. lactis subsp. cremoris strains were supplied by CSKFood Enrichment (Ede, the Netherlands). Lactococcus strains were grown at30°C either in M17 broth (Difco, Detroit, MI) supplemented with glucose at afinal concentration of 0.5% (wt/vol) (GM17) or in UHT skim milk (Lescure,Surgeres, France). When required, an antibiotic (tetracycline, erythromycin, orchloramphenicol) was added at a final concentration of 5 �g ml�1. Bacterialgrowth was estimated by measuring the optical density at 480 nm (OD480), afterdilution of 0.1 ml of a culture with 0.9 ml EDTA (5 g liter�1, pH 12.0) in the caseof milk cultures. Escherichia coli strains were grown at 37°C with agitation at 200rpm in Luria-Bertani broth (LB) (Difco, Detroit, MI). When required, erythro-mycin (final concentration, 150 �g ml�1), kanamycin (final concentration, 50 �gml�1), or chloramphenicol (final concentration, 50 �g ml�1) was added.
DNA manipulations. DNA extraction, PCR, plasmid preparation, and se-quencing were performed as described by Sambrook and Russell (36). DNArestriction and modification enzymes were purchased from New England Biolabs
(Ipswich, MA) and used as recommended by the supplier. The primers used inthis study were synthesized by Eurogentec (Seraing, Belgium) and are listed inTable 2. L. lactis electrocompetent cells were prepared and transformed asdescribed previously by Holo and Nes (18). Plasmid extraction was performed asdescribed by O’Sullivan and Klaenhammer (32). PCR amplification was carriedout with an Applied Biosystems 2720 DNA thermal cycler (Courtaboeuf,France), using the Taq DNA polymerase (MP Biomedicals, Illkirch, France) orPhusion DNA polymerase (high-fidelity PCR master mixture; Finnzymes, Fin-land).
Construction of the ahpF null mutant. The ahpF mutant was constructed bydeleting 435 bp of the ahpF gene using double-crossover gene replacement. A1,741-bp DNA fragment that included the ahpF gene (1,527 bp) was PCRamplified with primers ahpF-up and ahpF-down containing EcoRI and SacIIrestriction sites, respectively (Table 2). A 435-bp deletion inside the PCR prod-uct was obtained by digestion with AvaII and ligation of the two outside frag-ments (649 and 657 bp). The truncated fragment was then PCR amplified withprimers ahpF-up and ahpF-down and subcloned into the pGEM-T Easy vector(Promega, Madison, WI). After digestion of the subcloning vector with EcoRIand SacII, the resulting truncated fragment (1,289 bp) was cloned into pG�host9(28), and the recombinant vector was introduced by electroporation into L. lactis
TABLE 1. Strains and plasmids used in this study
Strain or plasmid Relevant characteristics Source or reference
L. lactis strainsTIL46 Wild-type (WT) strain derived from L. lactis NCDO763 cured of its 2-kb plasmid NCIMB (Aberdeen, UK)noxE mutant TIL46 noxE mutant obtained by single-crossover integration of pORI::noxE; Eryr This studynoxE-pJIM mutant noxE mutant complemented with pJIM; Eryr Cmr This studynoxE-pJIM::noxE
mutantnoxE mutant complemented with pJIM::noxE; Eryr Cmr This study
ahpF mutant TIL46 ahpF deletion mutant obtained by double-crossover gene replacementwith pG�host9::�ahpF
This study
noxAB mutant TIL46 noxAB deletion mutant obtained by double-crossover gene replacement 38menC mutant TIL46 menC integration mutant obtained by double-crossover gene replacement
with pG�host9::ISS138
E. coli TG1 repA� TG1 derivative with repA gene integrated into the chromosome, allowingreplication of L. lactis plasmids
P. Renault (INRA, GénétiqueMicrobienne, Jouy-en-Josas,France)
PlasmidspGEM-T Easy 3.0-kb linear T-overhang vector for PCR fragment cloning; nonreplicative in
Gram-positive bacteria; AprPromega, Madison, WI
pGEM-TnoxE pGEM-T Easy containing a 644-bp internal fragment of the noxE gene This studypORInewlux 4.3-kb pJIM2374-derived vector; nonreplicative in Gram-positive bacteria; Eryr 13pORInoxE pORInewlux containing a 644-bp internal fragment of the noxE gene This studypG�host9 3.8-kb pWV01 carrying a thermosensitive replicon; Eryr 28pG�host9::�ahpF pG�host9 containing truncated ahpF; Eryr This studypJIM pJIM2246, 6.4-kb multicopy plasmid vector; Cmr 34pJIM::noxE pJIM2246 containing noxE of TIL46; Cmr This study
TABLE 2. Primers used in this study
Primer Sequence (5� 3 3�)a
ahpF-up ..........................................ATAAATGAATTCGTCTTGACTCACTTTATTTG (EcoRI)
ahpF-down .....................................AATTTGCCGCGGAACATTTTACCACATTTATG(SacII)
ahpF-up2 ........................................GTGGAAAGAAGATGGGGAAAahpF-down2 ...................................CTGACCGACTTTATTGCGCTnoxF ................................................CTAATCTACCAGGCAAAGnoxR................................................TGAAATACCATTAGACCCnox2F ..............................................GACCGTAATAGCAACATGAGnox2R..............................................CTTGAATCGCTAAACTGAACnox3F ..............................................ACCCGATGCTCCCGATGTTCnox3R..............................................GGTGCGTTTGTTAAATGACCpOriEry...........................................GGTCAATCGAGAATATCGluxAinv............................................CCCAGATTAACCAATCGC
a Underlining indicates restriction sites of the enzymes indicated in parentheses.
1312 TACHON ET AL. APPL. ENVIRON. MICROBIOL.
on May 23, 2020 by guest
http://aem.asm
.org/D
ownloaded from

TIL46. The double crossovers leading to the expected gene replacement werescreened and obtained as described by Biswas et al. (2). Correct chromosomaldeletion of the ahpF gene was verified using PCR.
Construction of the noxE null mutant. The noxE gene, encoding the water-forming NADH oxidase NoxE, was disrupted in L. lactis TIL46 using a singlecrossover, resulting in the noxE mutant (Table 1). An internal 644-bp DNAfragment of noxE was PCR amplified with the noxF and noxR primers (Table 2)and subcloned into the pGEM-T Easy vector (Promega, Madison, WI). Theresulting plasmid, pGEM-TnoxE (Table 1), was produced in E. coli TG1 (Table1) and digested by SacI and AatII. The digestion product was then cloned intothe nonreplicative pORInewlux vector (13), leading to pORInoxE (Table 1),which was also produced in E. coli and transformed into TIL46 electrocompetentcells. Transformants were selected with erythromycin (5 �g ml�1). The disrup-tion of noxE by pORInoxE was confirmed by PCR using the external primerspORIEry, luxAinv, Nox2F, and Nox2R (Table 2).
Complementation of the noxE mutant. Complementation of the noxE mutantwas achieved by cloning the noxE gene into the pJIM2246 multicopy plasmidvector. A 1,728-bp fragment encoding noxE with its putative promoter andterminator was amplified by PCR from TIL46 total DNA with primers nox3F andnox3R (Table 2) using the Phusion high-fidelity DNA polymerase. This fragmentwas cloned into pGEM-T Easy, and the resulting plasmid was produced in E. coliTG1 and digested with SpeI and SacII. The fragment was then cloned into theSpeI/SacII-linearized vector pJIM2246. The resulting plasmid, pJIM2246::noxE(Table 1), was produced in E. coli TG1 repA� and used to transform the noxEmutant. The resulting strain was designated noxE-pJIM::noxE. A control strainwas constructed by introducing the empty vector pJIM2246 into the noxE mutant(noxE-pJIM).
Cell extract preparation. Cells were harvested from L. lactis cultures by cen-trifugation (4,000 � g, 15 min, 4°C). The cell pellets were washed twice with 5 ml50 mM potassium phosphate buffer (pH 7.0) and stored at �20°C until they wereused. The cells were resuspended in 50 mM potassium phosphate buffer (pH 7.0)containing 0.6 g of glass beads and disrupted twice for 45 s at 180 V and 4.5 m/s(Fast Prep orbital mixer; MP Biomedicals, Illkirch, France). Each cell extract wasrecovered by centrifugation (17,400 � g, 20 min, 4°C). The protein concentrationwas determined using a Coomassie (Bradford) protein assay kit (Pierce, Rock-ford, IL), and bovine serum albumin (BSA) was used as the standard.
NOX assay. NADH oxidase (NOX) activity was determined by using a 1-mlreaction mixture consisting of 50 mM potassium phosphate buffer (pH 7.0), 0.3mM EDTA, and 0.2 mM NADH. The reaction was initiated by addition of anappropriate quantity of freshly prepared cell extract. NADH oxidation was mon-itored by spectrophotometry at 340 nm and 25°C (Uvikon XL; Biotek Instru-ments, Colmar, France). NOX activity was calculated using the molar extinctioncoefficient of NADH (6,220 M�1 cm�1) and was expressed in units per milligramof protein. One unit of NADH oxidase activity is defined as the amount ofenzyme which catalyzes the oxidation of 1 �mol NADH per min at 25°C. Controlexperiments without the cell extract were performed to verify the absence ofspontaneous NADH oxidation.
Milk fermentation and measurement of pH, dissolved oxygen, and redoxpotential. All milk fermentations were performed at 30°C in 1-liter fermentors(BioStat Q plus; Sartorius) containing 500 ml UHT skim milk (Lescure, Surgeres,France). Milk was inoculated (1%, vol/vol) with a milk culture at an opticaldensity at 480 nm (OD480) of 1.8. The cultures were kept homogeneous by gentlestirring (100 rpm). Aerobic incubation was performed with air in the headspace,which remained in contact with the atmosphere via a sterile filter so that oxygencould diffuse into the medium during fermentation (low-oxygen conditions).Anaerobic cultures were grown in nitrogen-saturated milk with the headspacecontinuously sparged with nitrogen. During L. lactis growth, acidification and theoxygen concentration were monitored using pH sensors (EASYFERM plus K8;Hamilton) and dissolved oxygen (pO2) sensors (OXYFERM FDA 160; Hamil-ton). The pH sensors were calibrated using standard solutions with pH values of4.0 and 7.0. The oxygen sensors were calibrated using water sparged with air(21% oxygen) and nitrogen (0% oxygen). The redox potential was measuredusing redox electrodes (EASYFERM PLUS K8 Rx; Hamilton) that werecleaned, polished, and verified as described by Jeanson et al. (21). Data wereacquired using MFCS/win 2.0 software (B. Braun Biotech International). Acid-ification data were expressed as the �pH, which is the difference (in pH units)between the initial pH (pH 6.6) and the pH measured at each time. As describedby Cachon et al. (8), the measured redox potential (Em) was converted to Eh,with reference to the hydrogen electrode, using the following formula: Eh �Em � Er, where Er is 204 mV at 30°C for our Ag/AgCl electrodes. In addition,because redox potential values are pH dependent (19), Eh was transformed tothe redox potential at pH 7 (Eh7) with the Leistner-Mirna equation: Eh7 � Eh �� � (7 � pH) (25). The � constant determined for milk experimentally at 30°C
was 53. Bacterial growth was estimated each hour by measuring the opticaldensity at 480 nm (OD480) after dilution of 0.1 ml of a milk culture with 0.9 mlEDTA (5 g liter�1, pH 12.0). Fermentation experiments with L. lactis TIL46 andthe derived mutants were performed at least three times independently, and datawere compared using Student’s t test. Experiments with CSK strains were per-formed only once or twice.
RESULTS
Analysis of milk reduction mechanism using knockout mu-tants. (i) NADH oxidase activity. The NADH oxidase (NOX)activities of TIL46 (WT) and the derived noxE and ahpF mu-tants grown in GM17 were determined using extracts of cellsharvested at the end of the exponential growth phase, whenNOX activity was optimal (Table 3). Disruption of the noxEgene suppressed 95% of the NOX activity. Conversely, thenoxE mutant complemented with the pJIM expression vectorharboring noxE exhibited an 8-fold increase in activity com-pared with the WT strain. Finally, inactivation of the ahpFgene did not affect NOX activity.
(ii) Aerobic growth and acidification in milk. The abilities ofthe different mutant strains to grow in milk and acidify underlow-oxygen conditions (gentle stirring at 100 rpm) were exam-ined and compared with the abilities of the WT parent (Fig. 1Aand B). The growth of the noxE mutant did not include anexponential phase, and both the growth rate and the finalbiomass were significantly (P 0.001) reduced compared withthe growth rate and the final biomass of the WT strain. Theacidification rate was also significantly affected (P 0.001) butless than the growth rate, suggesting that the noxE mutantproduced more lactic acid than the wild-type strain. Thegrowth and acidification kinetics were restored when a noxEexpression vector (pJIM::noxE) was introduced into the noxEmutant (see Fig. S1 in the supplemental material), indicatingthat the growth defect was actually due to the absence of NOXactivity. The growth and acidification rates of the ahpF mutantwere also strongly reduced (P 0.001) compared with thegrowth and acidification rates of the WT (Fig. 1), while themenC and noxAB mutants grew and acidified slightly slowerthan the WT strain but the final biomasses and pH reachedabout the same levels as those of the WT strain (Fig. 1).
(iii) Dissolved oxygen consumption. The abilities of the WTstrain and mutants to consume dissolved oxygen were alsoexamined during milk fermentation under low-oxygen condi-tions (Fig. 2). The WT strain very rapidly consumed dissolved
TABLE 3. NADH oxidase activities in cell extracts of TIL46 (WT),the noxE mutant, the noxE mutant containing empty pJIM2246
(noxE-pJIM mutant), the noxE mutant complementedwith pJIM-noxE (noxE-pJIM::noxE mutant),
and the ahpF mutanta
Strain NADH oxidase activity(U mg�1)
WT.................................................................................... 0.30 0.05noxE mutant ....................................................................0.014 0.003noxE-pJIM mutant..........................................................0.016 0.005noxE-pJIM::noxE mutant............................................... 2.5 0.8ahpF mutant .................................................................... 0.32 0.15
a Cell extracts were prepared from GM17 cultures at the end of the exponen-tial growth phase. The data are means standard deviations of at least fourdeterminations.
VOL. 76, 2010 MECHANISMS OF MILK REDUCTION BY L. LACTIS 1313
on May 23, 2020 by guest
http://aem.asm
.org/D
ownloaded from
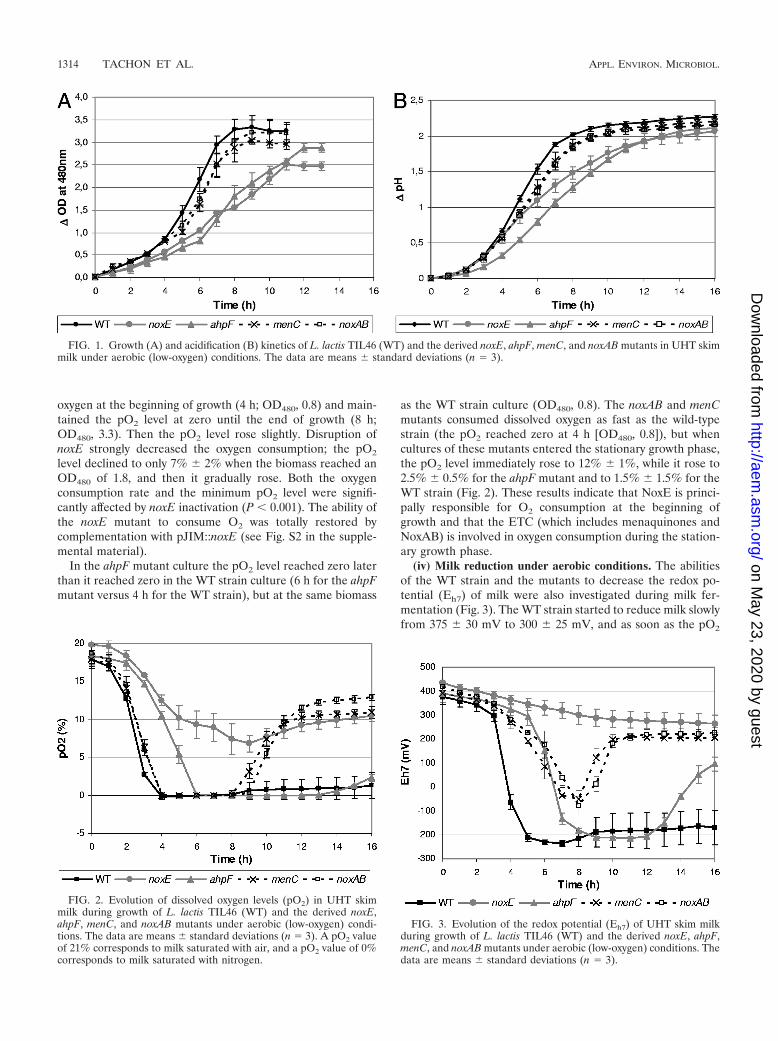
oxygen at the beginning of growth (4 h; OD480, 0.8) and main-tained the pO2 level at zero until the end of growth (8 h;OD480, 3.3). Then the pO2 level rose slightly. Disruption ofnoxE strongly decreased the oxygen consumption; the pO2
level declined to only 7% 2% when the biomass reached anOD480 of 1.8, and then it gradually rose. Both the oxygenconsumption rate and the minimum pO2 level were signifi-cantly affected by noxE inactivation (P 0.001). The ability ofthe noxE mutant to consume O2 was totally restored bycomplementation with pJIM::noxE (see Fig. S2 in the supple-mental material).
In the ahpF mutant culture the pO2 level reached zero laterthan it reached zero in the WT strain culture (6 h for the ahpFmutant versus 4 h for the WT strain), but at the same biomass
as the WT strain culture (OD480, 0.8). The noxAB and menCmutants consumed dissolved oxygen as fast as the wild-typestrain (the pO2 reached zero at 4 h [OD480, 0.8]), but whencultures of these mutants entered the stationary growth phase,the pO2 level immediately rose to 12% 1%, while it rose to2.5% 0.5% for the ahpF mutant and to 1.5% 1.5% for theWT strain (Fig. 2). These results indicate that NoxE is princi-pally responsible for O2 consumption at the beginning ofgrowth and that the ETC (which includes menaquinones andNoxAB) is involved in oxygen consumption during the station-ary growth phase.
(iv) Milk reduction under aerobic conditions. The abilitiesof the WT strain and the mutants to decrease the redox po-tential (Eh7) of milk were also investigated during milk fer-mentation (Fig. 3). The WT strain started to reduce milk slowlyfrom 375 30 mV to 300 25 mV, and as soon as the pO2
FIG. 1. Growth (A) and acidification (B) kinetics of L. lactis TIL46 (WT) and the derived noxE, ahpF, menC, and noxAB mutants in UHT skimmilk under aerobic (low-oxygen) conditions. The data are means standard deviations (n � 3).
FIG. 2. Evolution of dissolved oxygen levels (pO2) in UHT skimmilk during growth of L. lactis TIL46 (WT) and the derived noxE,ahpF, menC, and noxAB mutants under aerobic (low-oxygen) condi-tions. The data are means standard deviations (n � 3). A pO2 valueof 21% corresponds to milk saturated with air, and a pO2 value of 0%corresponds to milk saturated with nitrogen.
FIG. 3. Evolution of the redox potential (Eh7) of UHT skim milkduring growth of L. lactis TIL46 (WT) and the derived noxE, ahpF,menC, and noxAB mutants under aerobic (low-oxygen) conditions. Thedata are means standard deviations (n � 3).
1314 TACHON ET AL. APPL. ENVIRON. MICROBIOL.
on May 23, 2020 by guest
http://aem.asm
.org/D
ownloaded from

reached zero (at 4 h), the Eh7 immediately dropped to �200mV and then stabilized at around �230 10 mV until the endof the exponential growth phase (Fig. 3). The noxE mutantslowly reduced milk, and the Eh7 only declined to 260 40mV, which is in line with the relatively high oxygen level (pO2,�10%) in milk. The reducing ability of the noxE mutant wasrestored by complementation with pJIM::noxE (see Fig. S3 inthe supplemental material). The ahpF mutant reduced milklater than the WT, when the pO2 reached zero (after 6 h), butthen the Eh7 dropped to the same level that was observed withthe WT (�213 5 mV) (Fig. 3). Finally, the mutants whoseETC was affected (menC and noxAB mutants) reduced milkvery gradually to an Eh7 value of �65 50 mV at the end ofgrowth (8 h; OD480, �3), although the pO2 reached zero at thebeginning of growth (4 h; OD480, 0.8) (Fig. 2). For all strains,the Eh7 of milk increased after the end of growth, in line withthe increase in the pO2. All of these results indicate that NoxE
contributes markedly to the reduction of aerated milk via elim-ination of oxygen. The ETC is also strongly involved in milkreduction, probably by reducing oxidizing compounds otherthan the oxygen present in milk.
(v) Milk reduction under anaerobic conditions. The abilitiesof the WT strain and the mutants to grow, acidify, and reducemilk under anaerobiosis were also examined (Fig. 4). The anaer-obic growth rate of TIL46 in milk (Fig. 4A) was slightly reduced(P 0.05) compared with the aerobic growth rate and was similarto the aerobic growth rate of the noxE mutant (P � 0.17) (Fig. 1),indicating that NAD regeneration via NoxE (under aerobiosis)stimulates the growth of L. lactis TIL46 in milk. Of course, theimpact of noxE inactivation on anaerobic growth and acidificationwas weak because NoxE is inactive in the absence of oxygen. Incontrast, the negative impact of ahpF inactivation on anaerobicgrowth (P 0.001) was stronger than the impact on aerobicgrowth, suggesting that the impaired growth of the ahpF mutant
FIG. 4. Growth (A) and acidification kinetics (B) of L. lactis TIL46 (WT) and the derived noxE, ahpF, menC, and noxAB mutants innitrogen-saturated (anaerobic conditions) UHT skim milk and evolution of the redox potential (Eh7) of milk during growth (C). The data aremeans standard deviations (n � 3).
VOL. 76, 2010 MECHANISMS OF MILK REDUCTION BY L. LACTIS 1315
on May 23, 2020 by guest
http://aem.asm
.org/D
ownloaded from
was not due to its NADH oxidase activity. The growth rates (P 0.01) and final biomasses (P 0.01) of the menC and noxABmutants were also strongly affected (Fig. 4A).
Under anaerobiosis, the Eh7 of milk before inoculation wasessentially the same as that in the presence of O2 (350 50mV). Disruption of noxE or ahpF did not affect the ability ofTIL46 to decrease the Eh7 of milk (Fig. 4C). The Eh7 droppedimmediately (before the pH of milk started to decrease) from350 50 mV to a stable value of �265 15 mV. With themenC mutant the Eh7 dropped 1 h later than it dropped withthe WT strain, and it stabilized at a higher value (about�180 10 mV). Finally, the noxAB mutant reduced milk verygradually, and the Eh7 stabilized at the same level that it sta-bilized with the menC mutant (�180 10 mV). Thus, ourresults showed that the NoxA and/or NoxB NADH dehydro-genase and, to a lesser extent, menaquinones are involved inmilk reduction by L. lactis TIL46 under anaerobic conditions.
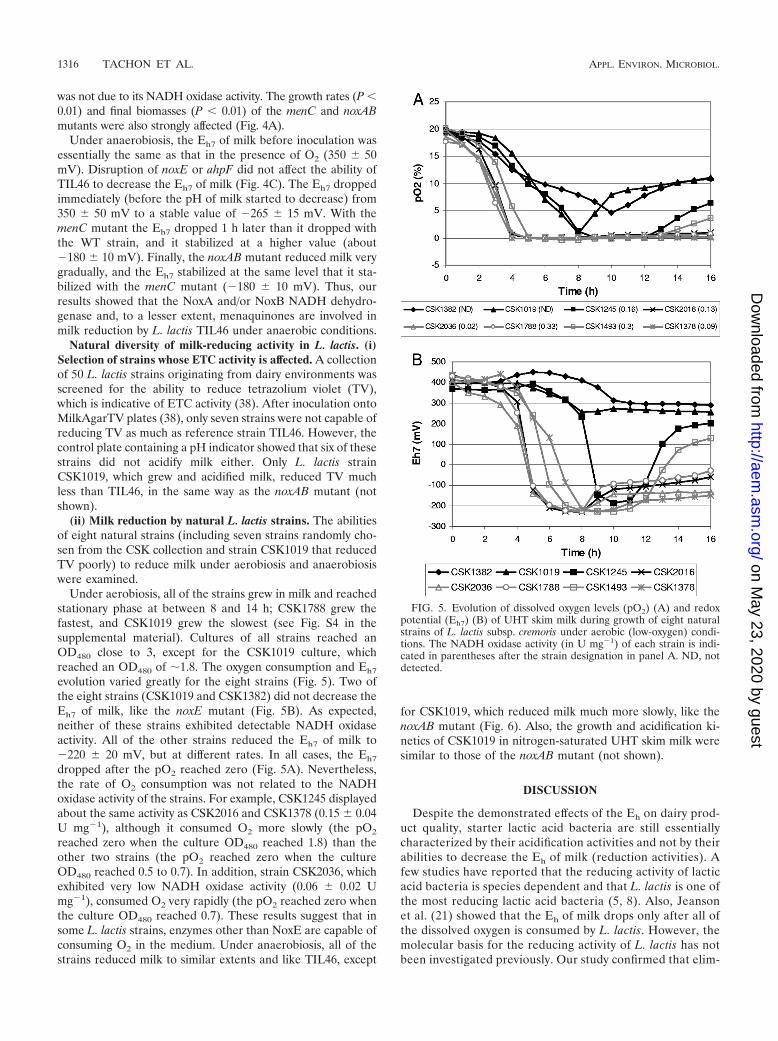
Natural diversity of milk-reducing activity in L. lactis. (i)Selection of strains whose ETC activity is affected. A collectionof 50 L. lactis strains originating from dairy environments wasscreened for the ability to reduce tetrazolium violet (TV),which is indicative of ETC activity (38). After inoculation ontoMilkAgarTV plates (38), only seven strains were not capable ofreducing TV as much as reference strain TIL46. However, thecontrol plate containing a pH indicator showed that six of thesestrains did not acidify milk either. Only L. lactis strainCSK1019, which grew and acidified milk, reduced TV muchless than TIL46, in the same way as the noxAB mutant (notshown).
(ii) Milk reduction by natural L. lactis strains. The abilitiesof eight natural strains (including seven strains randomly cho-sen from the CSK collection and strain CSK1019 that reducedTV poorly) to reduce milk under aerobiosis and anaerobiosiswere examined.
Under aerobiosis, all of the strains grew in milk and reachedstationary phase at between 8 and 14 h; CSK1788 grew thefastest, and CSK1019 grew the slowest (see Fig. S4 in thesupplemental material). Cultures of all strains reached anOD480 close to 3, except for the CSK1019 culture, whichreached an OD480 of �1.8. The oxygen consumption and Eh7
evolution varied greatly for the eight strains (Fig. 5). Two ofthe eight strains (CSK1019 and CSK1382) did not decrease theEh7 of milk, like the noxE mutant (Fig. 5B). As expected,neither of these strains exhibited detectable NADH oxidaseactivity. All of the other strains reduced the Eh7 of milk to�220 20 mV, but at different rates. In all cases, the Eh7
dropped after the pO2 reached zero (Fig. 5A). Nevertheless,the rate of O2 consumption was not related to the NADHoxidase activity of the strains. For example, CSK1245 displayedabout the same activity as CSK2016 and CSK1378 (0.15 0.04U mg�1), although it consumed O2 more slowly (the pO2
reached zero when the culture OD480 reached 1.8) than theother two strains (the pO2 reached zero when the cultureOD480 reached 0.5 to 0.7). In addition, strain CSK2036, whichexhibited very low NADH oxidase activity (0.06 0.02 Umg�1), consumed O2 very rapidly (the pO2 reached zero whenthe culture OD480 reached 0.7). These results suggest that insome L. lactis strains, enzymes other than NoxE are capable ofconsuming O2 in the medium. Under anaerobiosis, all of thestrains reduced milk to similar extents and like TIL46, except
for CSK1019, which reduced milk much more slowly, like thenoxAB mutant (Fig. 6). Also, the growth and acidification ki-netics of CSK1019 in nitrogen-saturated UHT skim milk weresimilar to those of the noxAB mutant (not shown).
DISCUSSION
Despite the demonstrated effects of the Eh on dairy prod-uct quality, starter lactic acid bacteria are still essentiallycharacterized by their acidification activities and not by theirabilities to decrease the Eh of milk (reduction activities). Afew studies have reported that the reducing activity of lacticacid bacteria is species dependent and that L. lactis is one ofthe most reducing lactic acid bacteria (5, 8). Also, Jeansonet al. (21) showed that the Eh of milk drops only after all ofthe dissolved oxygen is consumed by L. lactis. However, themolecular basis for the reducing activity of L. lactis has notbeen investigated previously. Our study confirmed that elim-
FIG. 5. Evolution of dissolved oxygen levels (pO2) (A) and redoxpotential (Eh7) (B) of UHT skim milk during growth of eight naturalstrains of L. lactis subsp. cremoris under aerobic (low-oxygen) condi-tions. The NADH oxidase activity (in U mg�1) of each strain is indi-cated in parentheses after the strain designation in panel A. ND, notdetected.
1316 TACHON ET AL. APPL. ENVIRON. MICROBIOL.
on May 23, 2020 by guest
http://aem.asm
.org/D
ownloaded from

ination of O2 is actually a prerequisite for milk reductionand revealed that another mechanism is necessary for theEh7 to fall to �220 mV. Indeed, we identified and charac-terized two major mechanisms involved in milk reduction byL. lactis (Fig. 7). The first mechanism relied on the elimi-nation of dissolved oxygen. We showed that NoxE activitywas essential for early O2 consumption during growth be-cause noxE inactivation hampered a decrease in the pO2
level to zero and consequently prevented a drop in the Eh
value. AhpF and the ETC did not appear to contributenotably to O2 consumption during the early growth phase.Indeed, the observed delay in the disappearance of O2 withthe ahpF mutant was mainly due to the delay in growth, andinactivation of the ETC did not affect O2 consumption sig-nificantly. However, AhpF and the ETC may be responsiblefor the slight consumption of O2 observed with the noxEmutant (Fig. 2). The minor role played by AhpF in oxygenelimination was consistent with the low specific activity ofthe pure enzyme (15 U mg�1) compared to that of NoxE (95U mg�1) (22) and with the fact that its disruption had noimpact on the NADH oxidase activity of cell extracts. Dur-ing the stationary growth phase, NoxE appeared to be muchless active, probably because it is unstable when it is sub-jected to overoxidation and is inhibited by an acidic pH (22).In this phase, the ETC contributed more markedly to O2
consumption. Indeed, inactivation of the ETC led to amarked increase in the pO2 at the end of growth. Thecontribution of the ETC to O2 consumption is in line withthe ability of menaquinones to reduce O2 to O2
�� (35) andwith the higher levels of menaquinones found in stationary-phase cells than in exponential-phase cells (35).
Nevertheless, elimination of O2 could not explain the dropin the Eh7 to �220 mV because the mutants whose ETC wasaffected (menC and noxAB mutants) consumed O2 completelyand quite rapidly but did not reduce milk as much as the WT.When the dissolved oxygen level reached zero, Eh7 was stillpositive and then gradually declined to �65 mV. These results
suggest that the ETC, which is made up of menaquinones andNoxA and/or NoxB, reduced oxidizing compounds other thanO2. These compounds may be metal cations or oxidized sulfurcompounds, which can be reduced to free thiols that have avery negative standard potential. Indeed, menaquinones arecapable of reducing metal cations and other oxidized com-pounds, such as TV (35, 38). The compounds reduced bymenaquinones seemed to be present in milk only under aero-biosis, because under anaerobiosis the menC mutant reducedmilk as rapidly as the WT. However, the final Eh7 value washigher with the menC mutant than with the WT. Surprisingly,the milk-reducing activity of the noxAB mutant was affectedmore than the milk-reducing activity of the menC mutant un-der anaerobiosis, suggesting that NoxA and/or NoxB reducedsome compounds directly and without the intervention ofmenaquinones, probably in the membrane where NoxA andNoxB are localized.
Our conclusion that menaquinones are necessary for reduc-tion of the Eh of milk to a very negative value is in agreementwith the fact that the most reducing lactic acid bacteria, such asL. lactis and Enterococcus faecalis, produce menaquinones,while Streptococcus thermophilus, which reduces the Eh of milkto a much lesser extent (to 8 mV), does not produce thesecompounds (5, 8, 31). Moreover, we did not find any mena-quinone biosynthesis genes in the sequenced genomes of Lac-tobacillus helveticus and Lactobacillus delbrueckii subsp. bul-garicus, which reduce milk in the same way as S. thermophilus(5), suggesting that they do not produce menaquinones either.However, several Lactobacillus plantarum strains reduce the Eh
of milk to a negative value (�154 mV) (5), while the se-quenced strain (WCF1) appears to lack several genes involvedin menaquinone biosynthesis. Other mechanisms may be in-volved in the reducing activity of L. plantarum, or menaqui-none production may be strain dependent.
Within L. lactis subsp. cremoris, we observed quite a broad
FIG. 7. Schematic representation of the mechanisms involved inmilk reduction by L. lactis. NoxAB, membrane dehydrogenases NoxAand NoxB; NoxE, NADH oxidase; AhpR, alkyl hydroperoxide reduc-tase system; OX, oxidized compounds; RED, reduced compounds;MK, oxidized menaquinones; MKH2, reduced menaquinones. Oxi-dized compounds are indicated by black type and are in boxes, whilereduced compounds are indicated by gray type and are in boxes.
FIG. 6. Reducing activity of strain CSK1019 and the TIL46 noxABmutant during growth in nitrogen-saturated UHT skim milk (anaero-bic conditions).
VOL. 76, 2010 MECHANISMS OF MILK REDUCTION BY L. LACTIS 1317
on May 23, 2020 by guest
http://aem.asm
.org/D
ownloaded from

diversity of milk-reducing activities. This diversity appeared tobe related in part to NADH oxidase activity because the twostrains without detectable NADH oxidase activity did not re-duce milk. However, some strains with low NADH oxidaseactivity eliminated O2 quite rapidly. This might have been dueto the possibility that the NADH oxidase activities of thesestrains in milk were higher than the activities determined inM17 medium, even though we found that the NADH oxidaseactivities of TIL46 in cells grown in these two media weresimilar. In fact, two other genes are predicted to encodeNADH oxidases in the genomes of L. lactis subsp. cremorisMG1363 and SK11 (30, 40). The first gene, noxC, encodes a547-residue protein in both genomes, while the second gene,noxD, is a pseudogene in MG1363 (which was derived fromTIL46) but encodes a 443-residue protein in SK11. The lattergene may be present in some natural strains and may be ex-pressed differently in M17 and milk. Moreover, certain naturalstrains may also exhibit other oxidase activities, such as pyru-vate oxidase activity, the genes for which are present in bothsequenced genomes, or a strong ETC activity, which enablethem to consume all of the oxygen present.
The diversity of the amounts of menaquinone produced byL. lactis strains in milk cultures has been examined previously(31). The amount produced ranged from 50 to 600 nmol per gof lyophilized culture. This great variation could have an im-pact on the reducing activities of different strains. However, inthe present study, the TV reduction test revealed that the ETCactivity of only one strain was affected, probably at the level ofthe NADH dehydrogenases. This result indicates that all of thestrains tested produced enough menaquinone to reduce TV.The NoxE activity of the selected strain CSK1019 was alsoaffected. However, the other natural strain without NoxE ac-tivity (CSK1382) and the noxE mutant reduced TV as well asreference strain TIL46, indicating that the TV test does notallow selection of strains on the basis of their oxygen consump-tion. To conclude, great variation in reducing activity wasfound in natural L. lactis strains originating from the dairyenvironment. The broad diversity allows selection of specificstrains that can be used to modulate the redox potential offermented dairy products.
ACKNOWLEDGMENTS
This work was supported by Eureka Research grant �!3562-LABREDOX. We are grateful to CSK Food Enrichment for financialsupport.
We thank Lucy Henno and Monique Cantonnet (INRA) for theirtechnical assistance and Veronique Monnet (INRA) for her criticalreading of the manuscript.
REFERENCES
1. Reference deleted.2. Biswas, I., A. Gruss, D. Ehrlich, and E. Maguin. 1993. High-efficiency gene
inactivation and replacement system for gram-positive bacteria. J. Bacteriol.175:3628–3635.
3. Bolduc, M.-P., Y. Raymond, P. Fustier, C. P. Champagne, and J.-C. Vuil-lemard. 2006. Sensitivity of bifidobacteria to oxygen and redox-potential innon-fermented pasteurized milk. Int. Dairy J. 16:1038–1048.
4. Boucher, B., C. Brothersen, and J. Broadbent. 2006. Influence of starter andnonstarter lactic acid bacteria on medium redox. Aust. J. Dairy Technol.61:116–118.
5. Brasca, M., S. Morandi, R. Lodi, and A. Tamburini. 2007. Redox potentialto discriminate among species of lactic acid bacteria. J. Appl. Microbiol.103:1516–1524.
6. Bronner, S., H. Monteil, and G. Prevost. 2004. Regulation of virulence
determinants in Staphylococcus aureus: complexity and applications. FEMSMicrobiol. Rev. 28:183–200.
7. Brown, M. H., and O. Emberger. 1980. Oxidation-reduction potential, p.112–125. In J. H. Silliker et al. (ed.), Microbial ecology of foods, vol. 1.Academic Press, New York, NY.
8. Cachon, R., S. Jeanson, M. Aldarf, and C. Divies. 2002. Characterization oflactic acid starters based on acidification and reduction activities. Lait 82:281–288.
9. Carreira, A., L. M. Ferreira, and V. Loureiro. 2001. Brown pigments pro-duced by Yarrowia lipolytica result from extracellular accumulation of ho-mogentisic acid. Appl. Environ. Microbiol. 67:3463–3468.
10. Dave, R. I., and N. P. Shah. 1997. Effect of cysteine on the viability of yogurtand probiotic bacteria in yogurts made with commercial starter cultures. Int.Dairy J. 7:535–545.
11. Dave, R. I., and N. P. Shah. 1997. Effectiveness of ascorbic acid as an oxygenscavenger in improving viability of probiotic bacteria in yoghurts made withcommercial starter cultures. Int. Dairy J. 7:435–443.
12. Dave, R. I., and N. P. Shah. 1998. Ingredient supplementation effects onviability of probiotic bacteria in yogurt. J. Dairy Sci. 81:2804–2816.
13. Delorme, C., D. S. Ehrlich, and P. Renault. 1999. Regulation of expressionof the Lactococcus lactis histidine operon. J. Bacteriol. 18:2026–2037.
14. de Oliveira Ferreira, E., E. A. Yates, M. Goldner, R. C. Vommaro, F. Costae Silva Filho, D. Barreiros Petropolis, and R. Pilotto Domingues. 2008. Theredox potential interferes with the expression of lamin binding molecules inBacteroides fragilis. Mem. Inst. Oswaldo Cruz 103:683–689.
15. Duwat, P., S. Sourice, B. Cesselin, G. Lamberet, K. Vido, P. Gaudu, Y. LeLoir, F. Violet, P. Loubiere, and A. Gruss. 2001. Respiration capacity of thefermenting bacterium Lactococcus lactis and its positive effects on growthand survival. J. Bacteriol. 183:4509–4516.
16. Green, M. L., and D. J. Manning. 1982. Development of texture and flavourin cheese and other fermented products. J. Dairy Res. 49:737–748.
17. Higuchi, M., Y. Yamamoto, L. B. Poole, M. Shimada, Y. Sato, N. Takahashi,and Y. Kamio. 1999. Functions of two types of NADH oxidases in energymetabolism and oxidative stress of Streptococcus mutans. J. Bacteriol. 181:5940–5947.
18. Holo, H., and I. F. Nes. 1989. High-frequency transformation, by electropo-ration, of Lactococcus lactis subsp. cremoris grown with glycine in osmoticallystabilized media. Appl. Environ. Microbiol. 55:3119–3123.
19. Jacob, H. E. 1970. Redox potential. Methods Microbiol. 2:91–123.20. Jayamanne, V. S., and M. R. Adams. 2009. Modelling the effects of pH,
storage temperature and redox potential (Eh) on the survival of bi-fidobacteria in fermented milk. Int. J. Food Sci. Technol. 44:1131–1138.
21. Jeanson, S., N. Hilgert, M. O. Coquillard, C. Seukpanya, M. Faiveley, P.Neveu, C. Abraham, V. Georgescu, P. Fourcassie, and E. Beuvier. 2009. Milkacidification by Lactococcus lactis is improved by decreasing the level ofdissolved oxygen rather than decreasing redox potential in the milk prior toinoculation. Int. J. Food Microbiol. 131:75–81.
22. Jiang, R., B. R. Riebel, and A. S. Bommarius. 2005. Comparison of alkylhydroperoxide reductase (AhpR) and water-forming NADH oxidase fromLactococcus lactis ATCC19435. Adv. Synth. Catal. 347:1139–1146.
23. Kieronczyk, A., R. Cachon, G. Feron, and M. Yvon. 2006. Addition ofoxidizing or reducing molecules to the reaction medium influences aminoacid conversion to aroma compounds by Lactococcus lactis. J. Appl. Micro-biol. 101:1114–1122.
24. Kristoffersen, T. 1985. Development of flavor in cheese. Milchwissenschaft40:197–199.
25. Leistner, L., and A. Mirna. 1959. Das Redoxpotential von Pokellaken. Fleis-chwirtschaft 8:659–666.
26. Lopez de Felipe, F., and J. Hugenholtz. 2001. Purification and characteriza-tion of the water forming NADH-oxidase from Lactococcus lactis. Int. DairyJ. 11:37–44.
27. Lourens-Hattingh, A., and B. C. Viljoen. 2001. Yogurt as probiotic carrierfood. Int. Dairy J. 11:1–17.
28. Maguin, E., P. Duwat, T. Hege, D. Ehrlich, and A. Gruss. 1992. New ther-mosensitive plasmid for gram-positive bacteria. J. Bacteriol. 174:5633–5638.
29. Reference deleted.30. Makarova, K., A. Slesarev, Y. Wolf, A. Sorokin, B. Mirkin, E. Koonin, A.
Pavlov, N. Pavlova, V. Karamychev, N. Polouchine, V. Shakhova, I. Grig-oriev, Y. Lou, D. Rohksar, S. Lucas, K. Huang, D. M. Goodstein, T.Hawkins, V. Plengvidhya, D. Welker, J. Hughes, Y. Goh, A. Benson, K.Baldwin, J. H. Lee, I. Diaz-Muniz, B. Dosti, V. Smeianov, W. Wechter, R.Barabote, G. Lorca, E. Altermann, R. Barrangou, B. Ganesan, Y. Xie, H.Rawsthorne, D. Tamir, C. Parker, F. Breidt, J. Broadbent, R. Hutkins, D.O’Sullivan, J. Steele, G. Unlu, M. Saier, T. Klaenhammer, P. Richardson,S. Kozyavkin, B. Weimer, and D. Mills. 2006. Comparative genomics ofthe lactic acid bacteria. Proc. Natl. Acad. Sci. U. S. A. 103:15611–15616.
31. Morishita, T., N. Tamura, T. Makino, and S. Kudo. 1999. Production ofmenaquinones by lactic acid bacteria. J. Dairy Sci. 82:1897–1903.
32. O’Sullivan, D. J., and T. R. Klaenhammer. 1993. Rapid mini-prep isolationof high-quality plasmid DNA from Lactococcus and Lactobacillus spp. Appl.Environ. Microbiol. 59:2730–2733.
33. Ouvry, A., Y. Wache, R. Tourdot-Marechal, C. Divies, and R. Cachon. 2002.
1318 TACHON ET AL. APPL. ENVIRON. MICROBIOL.
on May 23, 2020 by guest
http://aem.asm
.org/D
ownloaded from

Effects of oxidoreduction potential combined with acetic acid, NaCl andtemperature on the growth, acidification, and membrane properties of Lac-tobacillus plantarum. FEMS Microbiol. Lett. 214:257–261.
34. Renault, P., G. Corthier, N. Goupil, C. Delorme, and S. D. Ehrlich. 1996.Plasmid vectors for Gram-positive bacteria switching from high to low copynumber. Gene 183:175–182.
35. Rezaïki, L., G. Lamberet, A. Derre, A. Gruss, and P. Gaudu. 2008. Lacto-coccus lactis produces short-chain quinones that cross-feed group B Strepto-coccus to activate respiration growth. Mol. Microbiol. 67:947–957.
36. Sambrook, J., and D. W. Russell. 2001. Molecular cloning: a laboratorymanual, 3rd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor,NY.
37. Ster, C., F. B. Gilbert, T. Cochard, and B. Poutrel. 2005. Transcriptionalprofiles of regulatory and virulence factors of Staphylococcus aureus of bo-
vine origin: oxygen impact and strain-to-strain variations. Mol. Cell. Probes19:227–235.
38. Tachon, S., D. Michelon, E. Chambellon, M. Cantonnet, C. Mezange, L.Henno, R. Cachon, and M. Yvon. 2009. Experimental conditions affectwhether tetrazolium violet reduction by Lactococcus lactis occurs at mena-quinone or membrane NADH dehydrogenase sites of the electron transportchain. Microbiology 155:2941–2948.
39. Tengerdy, R. 1961. Redox potential changes in the 2-keto-L-gluconic acidfermentation. I. Correlation between redox potential and dissolved oxygenconcentration. J. Biochem. Microbiol. Technol. Eng. 3:241–253.
40. Wegmann, U., M. O’Connell-Motherway, A. Zomer, G. Buist, C. Shearman,C. Canchaya, M. Ventura, A. Goesmann, M. J. Gasson, O. P. Kuipers, D. vanSinderen, and J. Kok. 2007. Complete genome sequence of the prototypelactic acid bacterium Lactococcus lactis subsp. cremoris MG1363. J. Bacte-riol. 189:3256–3270.
VOL. 76, 2010 MECHANISMS OF MILK REDUCTION BY L. LACTIS 1319
on May 23, 2020 by guest
http://aem.asm
.org/D
ownloaded from