nitrogen form, availability, and mycorrhizal colonization affect biomass and nitrogen isotope...
TRANSCRIPT

REGULAR ARTICLE
Nitrogen form, availability, and mycorrhizal colonizationaffect biomass and nitrogen isotope patternsin Pinus sylvestris
Erik A. Hobbie & Jan V. Colpaert &Marguerite W. White & Andrew P. Ouimette &
Stephen A. Macko
Received: 7 January 2008 /Accepted: 23 April 2008 /Published online: 30 May 2008# Springer Science + Business Media B.V. 2008
Abstract Nitrogen (N) isotope patterns are useful forunderstanding carbon and nitrogen dynamics inmycorrhizal systems but questions remain about howdifferent N forms, fungal symbionts, and N availabil-ities influence δ15N signatures. Here, we studied howbiomass allocation and δ15N patterns in Pinussylvestris L. cultures were affected by nitrogen supplyrate (3% per day or 4% per day relative to thenitrogen already present), nitrogen form (ammoniumversus nitrate), and mycorrhizal colonization by fungiwith a greater (Laccaria laccata) or lesser (Suillusbovinus) ability to assimilate nitrate. Mycorrhizal(fungal) biomass was greater with ammonium thanwith nitrate nutrition for Suillus cultures but similarfor Laccaria cultures. Total biomass was less withnitrate nutrition than with ammonium nutrition for
nonmycorrhizal cultures and was less in mycorrhizalcultures than in nonmycorrhizal cultures. The seques-tration of available N by mycorrhizal fungi limitedplant N supply. This limitation and the higherenergetic cost of nitrate reduction than ammoniumassimilation appeared to control plant biomass accu-mulation. Colonization decreased foliar δ15N by 0.5to 2.2‰ (nitrate) or 1.7 to 3.5‰ (ammonium) andincreased root tip δ15N by 0 to 1‰ (nitrate) or 0.6 to2.3‰ (ammonium). Root tip δ15N and fungal biomasson root tips were positively correlated in ammoniumtreatments (r2=0.52) but not in nitrate treatments (r2=0.00). Fungal biomass on root tips was enriched in15N an estimated 6–8‰ relative to plant biomass inammonium treatments. At high nitrate availability,Suillus colonization did not reduce plant δ15N. Weconclude that: (1) transfer of 15N-depleted N frommycorrhizal fungi to plants produces low plant δ15Nsignatures and high root tip and fungal δ15N signa-tures; (2) limited nitrate reduction in fungi restrictedtransfer of 15N-depleted N to plants when nitrate issupplied and may account for many field observationsof high plant δ15N under such conditions; (3) plantscould transfer assimilated nitrogen to fungi at highnitrate supply but such transfer was without 15Nfractionation. These factors probably control plantδ15N patterns across N availability gradients and werehere incorporated into analytical equations for inter-preting nitrogen isotope patterns in mycorrhizal fungiand plants.
Plant Soil (2008) 310:121–136DOI 10.1007/s11104-008-9637-x
Responsible Editor: A. C. Borstlap.
E. A. Hobbie (*) :M. W. White :A. P. OuimetteComplex Systems Research Center,University of New Hampshire,Durham, NH 03824, USAe-mail: [email protected]
J. V. ColpaertHasselt University,Diepenbeek 3590, Belgium
S. A. MackoDepartment of Environmental Sciences,University of Virginia,Charlottesville, VA 22903, USA

Keywords Pinus . Suillus . Laccaria . Cultures .
Mycorrhizal transfer . Nitrogen isotope fractionation
Introduction
Ectomycorrhizal fungi are key symbionts of many ofthe dominant trees of tropical, temperate, and borealforests, and greatly influence carbon and nitrogen (N)cycling in these ecosystems. They are both a largesink for plant photosynthate (Hobbie and Wallander2006) and a primary source for much of the soil-derived N that will ultimately fuel plant growth. In theprevailing view, ectomycorrhizal fungi primarily increasethe access of plants to organic nitrogen and ammoniumin N-limited systems (Lipson and Näsholm 2001;Schimel and Bennett 2004) whereas the high mobilityof nitrate (Plassard et al. 1991) reduces the influenceof ectomycorrhizal fungi on uptake of this inorganicN form. However, although most ectomycorrhizalfungi prefer ammonium as an N source (Rygiewiczet al. 1984a, b; Finlay et al. 1992; Buchmann et al.1996), some can effectively use nitrate (Ho and Trappe1980; Sarjala 1990, 1991; Bailly et al. 2007), so theextent to which nitrate is supplied to plants throughtheir fungal symbionts is unclear in natural systems.
One potential tool to assess the supply of N toplants through their ectomycorrhizal symbionts is tomeasure 15N/14N ratios (expressed as δ15N values), asinternal isotopic fractionations against 15N within theectomycorrhizal mycelia decrease the δ15N of Ntransferred to the host plant and increase the δ15N offungal N (Högberg et al. 1999; Kohzu et al. 2000;Hobbie and Colpaert 2003; Brearley et al. 2003).Foliar δ15N correlated strongly and negatively withmycorrhizal colonization; conversely, Suillus, Thele-phora, and Paxillus mycorrhizae were elevated in 15Ncontent relative to available N when supplied withammonium nitrate (Hobbie and Colpaert 2003) orammonium (Högberg et al. 1999; Kohzu et al. 2000)but not with nitrate (Högberg et al. 1999). However,ectomycorrhizal fungi such as Laccaria with goodabilities to use nitrate (Finlay et al. 1992) have yet tobe used in such culture experiments, so it is stillpossible that the ectomycorrhizal role in plant supplyof nitrate could affect plant δ15N patterns.
The proportion of ammonium in the inorganic Npool and the allocation of carbon belowground and toectomycorrhizal fungi are greater at low N availability
than at high N availability (Ågren and Bosatta 1996;Aber et al. 1998; Hobbie 2006). The decrease of plantδ15N signatures with mycorrhizal colonization alsoappears greater at low than at high N availability infield studies (Hobbie et al. 2000) and in culturestudies (Hobbie and Colpaert 2003), but whether thisreflects high N retention by mycorrhizal fungi or alow importance of direct plant uptake at low Navailability is unknown. To separate the effects of Nform from those of N availability and mycorrhizalcolonization, these variables were isolated in thisstudy using appropriate culture conditions.
In this study, nonmycorrhizal and ectomycorhizalScots pine (Pinus sylvestris L.) were cultured underconditions of exponential growth at two different ratesof nutrient application, 3% per day (low N) and 4%per day (high N). Separate cultures were used withnitrate or ammonium as the main N source to determinetheir effects on the functioning of the ectomycorrhizalsymbiosis. In addition, two mycorrhizal fungi wereselected with either a greater [Laccaria laccata (Scop.)Fr.] or a lesser [Suillus bovinus (Pers.) Roussel] abilityto assimilate nitrate (Finlay et al. 1992).
After harvest, cultures were assessed for patterns inbiomass, fungal colonization, %N, and δ15N. Biomassincluded foliage, stems, roots, root tips, and fungalbiomass in extraradical hyphae. Root tips in mycor-rhizal cultures included both plant and fungal bio-mass. We hypothesized that:
1. Fungal colonization by Suillus would be greaterwith ammonium than with nitrate nutrition, fungalcolonization by Laccaria would be similar withboth N forms, and fungal colonization should begreater at low N availability than at high Navailability. Plant biomass should decline asfungal colonization increases.
We further hypothesized that fungal transfer of15N-depleted N would cause the following patterns:
2. Fungal colonization would negatively correlatewith foliar δ15N and positively correlate with roottip δ15N;
3. Relative to nonmycorrhizal cultures, foliar δ15Nin mycorrhizal cultures would decline more withammonium than with nitrate nutrition because ofgreater fungal assimilation of ammonium than ofnitrate. Foliar δ15N would decline more at low Nthan at high N supply.
122 Plant Soil (2008) 310:121–136

Materials and methods
Culture conditions
Surface-sterilized Scots pine seeds were sown in aperlite/vermiculite (2:1 by volume) mixture moistenedwith a balanced nutrient solution for Pinus sylvestris(Ingestad and Kähr 1985). The pH was adjusted to4.5. The experiment was carried out in a growthchamber with 300 µmol m−2 s−1 photosyntheticallyactive radiation, at least 70% relative air humidity,and with a day/night rhythm of 18/6 h and 22/15°C.Twenty-eight days after sowing, uniform seedlingswere selected for the experiment. A sandwichtechnique was used to inoculate 40 seedlings (VanTichelen and Colpaert 2000) either with Suillusbovinus or Laccaria laccata. Twenty nonmycorrhizalseedlings followed the same procedure in the absenceof fungal inoculum and six plants were immediatelyharvested to determine biomass (23.5±0.9 mg), Ncontent (0.368±0.010 mg), and δ15N (−0.7±0.2) atinoculation. Three days later plants were transferredto 150-cm3 containers filled with 6 g pure, acid-washed, sieved perlite (2–4 mm particles; Colpaert etal. 1999). Perlite has a low nutrient buffering capacityso plants are actually growing in a semihydroponicenvironment where it is possible to match nutrientaddition and nutrient uptake in the plants.
Immediately after inoculation four different nutri-ent supply regimes were applied. Half of the plantsreceived ammonium as the N source, the other halfnitrate. The stock solutions for the ammoniumtreatment contained (in µM): K2SO4 (292), K2HPO4
(116), (NH4)2SO4 (1777), Ca(H2PO4)2•H2O (73),MgSO4•6H2O (123), H3BO3 (9.3), MnSO4•H2O(3.7), Fe(NO3)3•9H2O (6.3), ZnSO4•7H2O (0.2),CuCl2•2H2O (0.2), Na2MoO4•2H2O (0.04). Nutrientsolution supplied to the plants contained less than25 mg N l−1. The stock solution for the nitratetreatment contained (in µM): K2SO4 (141), K2HPO4
(116), KH2PO4 (126), KNO3 (2967), HNO3 (184), Ca(NO3)2•4H2O (73), Mg(NO3)2•6H2O (124), H3BO3
(9.3), Mn(NO3)2•4H2O (3.7), Fe(NO3)3•9H2O (6.3),Zn(NO3)2•4H2O (0.2), CuCl2•2H2O (0.2), andNa2MoO4•2H2O (0.04). For both N source treat-ments, two supply levels were used. Nutrients wereeither added at a constant relative addition rate of 3%per day (low N treatment) or 4% per day (high Ntreatment), where the percent of nutrient added daily
is relative to that already present in the culture.Nitrogen was the limiting nutrient in all treatments.These nutrient regimes are suboptimal so that theseedlings will adjust their relative growth rate to thenutrient addition rates (Ingestad and Kähr 1985).
Analyses
Plants were harvested after 80 days, at which time thecumulative N added to each plant during theexperimental period was 6.93 mg for the high Ntreatments or 1.97 mg for the low N treatments. Non-absorbed N was washed from the perlite with 200 mlof nitrogen-free nutrient solution. The containers werethen centrifuged at 35 g for 30 seconds to remove thesolution retained in the perlite. The entire plant andsoil was pulled from the containers; the roots werethen separated from the perlite (Colpaert et al. 1999).Root tips were detached from roots, mycorrhizaebeing included in the former fraction. Subsamples ofperlite were frozen in liquid nitrogen for ergosteroldeterminations. Ergosterol was analysed by HPLCand results were converted to fungal biomass usingconcentrations of 4.2 and 7.1 mg ergosterol per gbiomass for Laccaria laccata and Suillus bovinus,respectively (Nylund and Wallander 1992). Aftersample drying at 60°C, weights of foliage, stems,roots, and root tips were determined. Dried plant andperlite material was ground with a ball mill at 200 Hzfor 2 minutes. Samples were stored at room temper-ature prior to stable isotope analyses.
Nitrogen isotope and concentration values weremeasured using a GV Micromass Optima (Man-chester, UK) linked through continuous-flow to aCarlo Erba elemental analyser. Samples werereferenced against concurrently run acetanilide andpine needle (NIST 1575) standards that had beencalibrated against ammonium sulfate (N1 and N2)from the International Atomic Energy Administra-tion. Nitrogen isotope values were expressed as δ15N(‰)=(Rsample/Rstandard−1) 1,000‰, with R=15N/14Nand the standard is atmospheric N2. The precision ofstandards was typically ±0.2‰.
Total biomass was calculated as the sum of themasses of needles, stems, roots, root tips, andmycorhizal hyphae in perlite, whereas mycorrhizalbiomass was calculated as the sum of mycorrhizalfungi on root tips and mycorrhizal hyphae in perlite.Relative belowground allocation was the sum of
Plant Soil (2008) 310:121–136 123
roots, root tips, and mycorrhizal hyphae in perlitedivided by total biomass. Relative growth rates(RGR) in percent per day were calculated by theequation, final biomass/initial biomass=yx, with y=1+RGR and x = growth period in days between initialand final biomass estimates.
The data were analyzed using the statisticalpackage Statview (Abacus Concepts) with a three-way ANOVA at a significance level of 0.05, withnitrogen source (ammonium or nitrate), supply rate(high versus low), and mycorrhizal associate (non-mycorrhizal, Suillus, or Laccaria) as the independentvariables. Correlations among variables and t-testswere also analyzed at a significance level of 0.05.
Results
Biomass, fungal colonization, and nitrogen retention
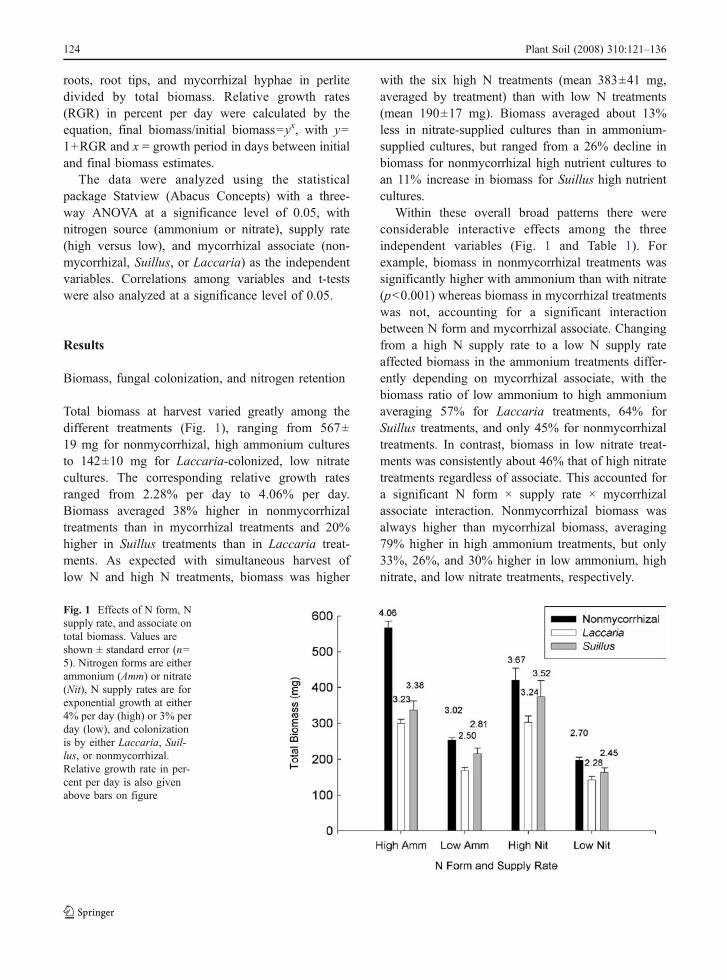
Total biomass at harvest varied greatly among thedifferent treatments (Fig. 1), ranging from 567±19 mg for nonmycorrhizal, high ammonium culturesto 142±10 mg for Laccaria-colonized, low nitratecultures. The corresponding relative growth ratesranged from 2.28% per day to 4.06% per day.Biomass averaged 38% higher in nonmycorrhizaltreatments than in mycorrhizal treatments and 20%higher in Suillus treatments than in Laccaria treat-ments. As expected with simultaneous harvest oflow N and high N treatments, biomass was higher
with the six high N treatments (mean 383±41 mg,averaged by treatment) than with low N treatments(mean 190±17 mg). Biomass averaged about 13%less in nitrate-supplied cultures than in ammonium-supplied cultures, but ranged from a 26% decline inbiomass for nonmycorrhizal high nutrient cultures toan 11% increase in biomass for Suillus high nutrientcultures.
Within these overall broad patterns there wereconsiderable interactive effects among the threeindependent variables (Fig. 1 and Table 1). Forexample, biomass in nonmycorrhizal treatments wassignificantly higher with ammonium than with nitrate(p<0.001) whereas biomass in mycorrhizal treatmentswas not, accounting for a significant interactionbetween N form and mycorrhizal associate. Changingfrom a high N supply rate to a low N supply rateaffected biomass in the ammonium treatments differ-ently depending on mycorrhizal associate, with thebiomass ratio of low ammonium to high ammoniumaveraging 57% for Laccaria treatments, 64% forSuillus treatments, and only 45% for nonmycorrhizaltreatments. In contrast, biomass in low nitrate treat-ments was consistently about 46% that of high nitratetreatments regardless of associate. This accounted fora significant N form × supply rate × mycorrhizalassociate interaction. Nonmycorrhizal biomass wasalways higher than mycorrhizal biomass, averaging79% higher in high ammonium treatments, but only33%, 26%, and 30% higher in low ammonium, highnitrate, and low nitrate treatments, respectively.
Fig. 1 Effects of N form, Nsupply rate, and associate ontotal biomass. Values areshown ± standard error (n=5). Nitrogen forms are eitherammonium (Amm) or nitrate(Nit), N supply rates are forexponential growth at either4% per day (high) or 3% perday (low), and colonizationis by either Laccaria, Suil-lus, or nonmycorrhizal.Relative growth rate in per-cent per day is also givenabove bars on figure
124 Plant Soil (2008) 310:121–136
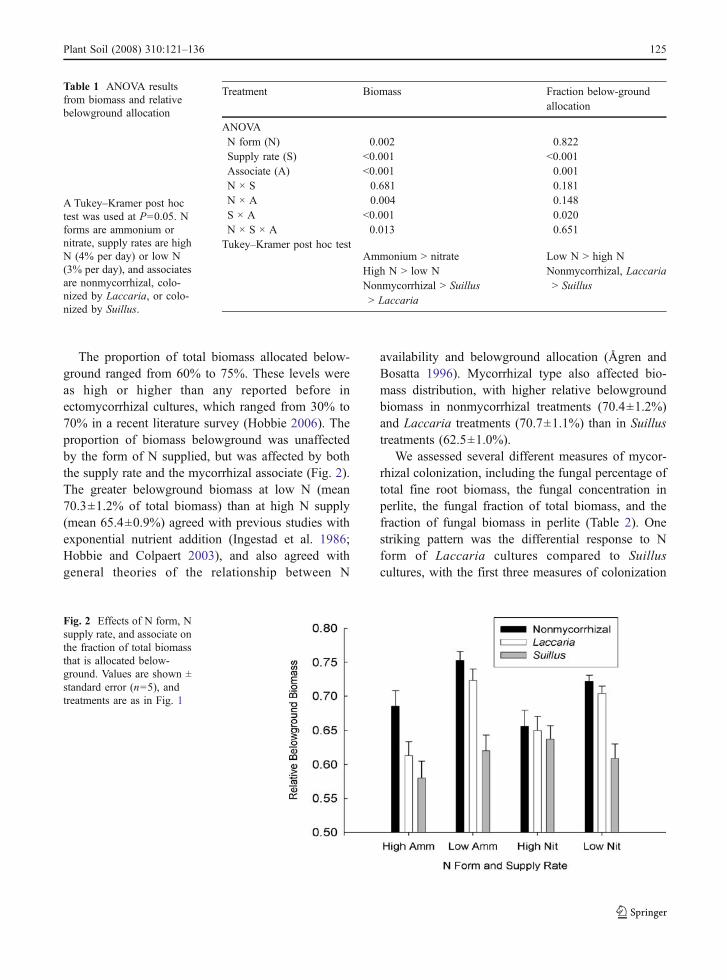
The proportion of total biomass allocated below-ground ranged from 60% to 75%. These levels wereas high or higher than any reported before inectomycorrhizal cultures, which ranged from 30% to70% in a recent literature survey (Hobbie 2006). Theproportion of biomass belowground was unaffectedby the form of N supplied, but was affected by boththe supply rate and the mycorrhizal associate (Fig. 2).The greater belowground biomass at low N (mean70.3±1.2% of total biomass) than at high N supply(mean 65.4±0.9%) agreed with previous studies withexponential nutrient addition (Ingestad et al. 1986;Hobbie and Colpaert 2003), and also agreed withgeneral theories of the relationship between N
availability and belowground allocation (Ågren andBosatta 1996). Mycorrhizal type also affected bio-mass distribution, with higher relative belowgroundbiomass in nonmycorrhizal treatments (70.4±1.2%)and Laccaria treatments (70.7±1.1%) than in Suillustreatments (62.5±1.0%).
We assessed several different measures of mycor-rhizal colonization, including the fungal percentage oftotal fine root biomass, the fungal concentration inperlite, the fungal fraction of total biomass, and thefraction of fungal biomass in perlite (Table 2). Onestriking pattern was the differential response to Nform of Laccaria cultures compared to Suilluscultures, with the first three measures of colonization
Treatment Biomass Fraction below-groundallocation
ANOVAN form (N) 0.002 0.822Supply rate (S) <0.001 <0.001Associate (A) <0.001 0.001N × S 0.681 0.181N × A 0.004 0.148S × A <0.001 0.020N × S × A 0.013 0.651Tukey–Kramer post hoc test
Ammonium > nitrate Low N > high NHigh N > low N Nonmycorrhizal, Laccaria
> SuillusNonmycorrhizal > Suillus> Laccaria
Table 1 ANOVA resultsfrom biomass and relativebelowground allocation
A Tukey–Kramer post hoctest was used at P=0.05. Nforms are ammonium ornitrate, supply rates are highN (4% per day) or low N(3% per day), and associatesare nonmycorrhizal, colo-nized by Laccaria, or colo-nized by Suillus.
Fig. 2 Effects of N form, Nsupply rate, and associate onthe fraction of total biomassthat is allocated below-ground. Values are shown ±standard error (n=5), andtreatments are as in Fig. 1
Plant Soil (2008) 310:121–136 125

having a highly significant interaction between Nform and associate (p<0.01 for all, Table 2). If theanalysis is done separately by mycorrhizal associate,all three colonization measures for Suillus cultures aresignificantly higher for ammonium supply than fornitrate supply (p<0.001 for all three) whereas forLaccaria cultures none of these measures of coloni-zation differed significantly by N form (p=0.080,0.461, and 0.148, respectively).
Mycorrhizal colonization on roots varied with Nform, N availability, and fungal symbiont. As apercentage of total fine root biomass, mycorrhizalcolonization on fine roots ranged from 15.2±2.0% fornitrate-supplied Suillus cultures at high N to 29.3±1.6% for ammonium-supplied Suillus cultures at highN (Table 2). Mycorrhizal colonization was generallyhigher on roots with ammonium-supplied culturesthan with nitrate-supplied cultures (p=0.025). How-ever, colonization on roots in Laccaria treatments wassimilar for nitrate-supplied cultures and for ammoni-um-supplied cultures (p=0.081), whereas coloniza-tion on Suillus treatments was much higher with
ammonium-supplied cultures (p<0.001), accountingfor a highly significant interaction between N formand fungal associate (p<0.001). Mycorrhizal coloni-zation on roots was actually higher at high N (23% ofroot biomass) than at low N (20% of root biomass, p=0.003), in contrast to previous results with pinescultured with ammonium nitrate and colonized withSuillus luteus and Thelephora terrestris (Hobbie andColpaert 2003). Colonization increased with increas-ing N supply for all associate × N form combinationsexcept for nitrate-supplied Suillus cultures, whichwere similar for both supply rates.
In mycorrhizal treatments, fungal biomass (on fineroots plus in perlite) ranged from 11% to 26% of totalbiomass (Table 2). Regardless of N supply rate ormycorrhizal symbiont, fungal biomass varied littleunder ammonium nutrition, ranging from 19% to 23%of total biomass. In contrast, fungal biomass undernitrate nutrition was much higher in Laccaria treat-ments (~25% of total biomass) than in Suillus treat-ments (11–15% of total biomass). Total biomass wasnegatively correlated with the fungal proportion of
Table 2 Fungal colonization of fine roots and perlite and percentage of total biomass from fungi (designated %fungal biomass)
N form Ammonium supply Nitrate supply
Treatment Fungi onroots (%)
Fungi inperlite (mg/g)
Percent fungalbiomass
Fungi inperlite (%)
Fungi onroots (%)
Fungi inperlite (mg/g)
Percent fungalbiomass
Fungi inperlite (%)
High NNM 3.0±0.1 – – – 2.4±0.1 – – –Laccaria 21.2±2.2 5.5±0.2 22.7±3.0 49.6±2.4 27.7±2.2 5.0±0.3 25.7±2.3 39.2±3.8Suillus 29.3±1.6 2.8±0.2 21.2±1.6 24.4±2.3 15.2±2.0 1.2±0.1 11.4±1.2 18.0±2.8Low NNM 2.6±0.2 – – – 1.5±0.2 – – –Laccaria 17.9±0.8 2.9±0.2 22.1±1.1 47.3±3.9 19.8±2.2 3.1±0.2 25.4±2.1 53.2±1.2Suillus 22.7±2.6 1.6±0.1 15.0±3.4 29.1±3.4 15.8±1.4 1.6±0.2 14.8±0.9 39.2±3.9ANOVAN form (N) 0.025 <0.001 0.475 0.930Supply rate (S) 0.003 <0.001 0.484 <0.001Associate (A) 0.505 <0.001 <0.001 <0.001N × S 0.624 <0.001 0.062 <0.001N × A <0.001 0.009 0.003 0.367S × A 0.330 <0.001 0.715 0.120N × S × A 0.037 0.144 0.074 0.985
Fungal colonization was determined by ergosterol measurements, with ergosterol: fungal biomass conversions given in methods.Fungal colonization of roots is given as a percentage of fine root biomass, fungal colonization of perlite is given in mg per g perlite.Fungi in perlite (%) is the percentage of total fungal biomass (roots plus perlite) that was detected in perlite. Statistical analyses onlyincluded mycorrhizal treatments. Tukey–Kramer post hoc test, fungi on roots, fungi in perlite: ammonium > nitrate, high N > low N;% fungal biomass: Laccaria > Suillus; fungi in perlite (%): high N < low N, Laccaria > Suillus
NM nonmycorrhizal
126 Plant Soil (2008) 310:121–136
total biomass at both low N supply (r2=0.27, p=0.025) and high N supply (r2=0.28, p=0.019).
Nitrogen concentrations
Nitrogen concentrations were examined in needles,roots, root tips (Table 3), and stems (data not shown).Concentrations were highest in root tips (mean 2.33±0.10%) and foliage (mean 1.72±0.05%), and lowestin stems (mean 1.21±0.06%) and roots (mean 1.06±0.03%). None of the treatments affected stem Nconcentrations significantly. In contrast, foliar %Nvaried with N form, supply rate, and mycorrhizaltype, with ammonium-supplied cultures higher by0.19% than nitrate-supplied cultures and high Ncultures higher by 0.30% than low N cultures. Foliar%N in Laccaria cultures was lower than in bothSuillus and nonmycorrhizal cultures, by 0.19% and0.32%, respectively. Root %N was greater by 0.13%in high N than in low N treatments, but was notsignificantly affected by N form or mycorrhizalassociate. Mycorrhizal root tips averaged higher inN content than nonmycorrhizal root tips (2.72%versus 1.77%). Suillus-colonized root tips were muchhigher in N content with ammonium supply (mean3.10%) than with nitrate supply (mean 2.15%). In
contrast, Laccaria-colonized root tips did not differsignificantly among N forms (mean 2.60% forammonium supply versus 2.30% for nitrate supply).Root tips in Laccaria, Suillus and nonmycorrhizaltreatments averaged respectively 0.91%, 0.92% and -0.04% higher in %N than the corresponding foliage,presumably reflecting the presence on mycorrhizalroot tips of fungal matter high in N (Hobbie andColpaert 2003).
Nitrogen isotope patterns
The two N sources differed somewhat in δ15N, withammonium at 1.4‰ and nitrate at −1.3‰. However,the mean δ15N of measured components in ammoni-um treatments (−1.7‰) was not significantly higherthan nitrate treatments (−1.9‰), presumably becauseof high losses of 15N-enriched compounds in ammo-nium treatments. Within a treatment, the δ15N valuesof individual components varied by up to 7‰.Relative to needle δ15N, roots averaged 1.4‰ higher,root tips 2.1‰ higher, and stems 1.1‰ lower(Table 4). Significant differences in δ15N amongneedles, roots, and root tips were confined tomycorrhizal treatments. In contrast, stems were
Table 3 %N of needles, roots and root tips under different treatments
N form Ammonium supply Nitrate supply
Treatment Needles Roots Root tips Needles Roots Root tips
High NNM 1.79±0.07 1.02±0.06 1.73±0.06 1.89±0.13 1.15±0.07 1.64±0.14Laccaria 1.79±0.18 0.97±0.10 2.83±0.27 1.70±0.12 1.26±0.06 2.31±0.10Suillus 1.96±0.07 1.24±0.11 3.51±0.17 1.71±0.21 1.10±0.10 1.91±0.39Low NNM 1.78±0.04 1.15±0.08 1.21±0.07 1.58±0.09 0.90±0.12 2.49±0.49Laccaria 1.62±0.13 0.89±0.04 2.37±0.15 1.08±0.13 1.05±0.08 2.30±0.27Suillus 1.57±0.03 0.92±0.09 2.69±0.16 1.72±0.01 1.06±0.17 2.39±0.23ANOVA Needles Roots Root tipsN form (N) 0.003 0.306 0.148Supply rate (S) <0.001 0.024 0.586Associate (A) 0.001 0.208 <0.001N × S 0.781 0.495 0.001N × A 0.230 0.741 <0.001S × A 0.387 0.318 0.517N × S × A 0.039 0.053 0.373
Statistical significance was determined by ANOVA followed by a post hoc Tukey–Kramer test at P=0.05. Tukey–Kramer post hoctest, needles: ammonium > nitrate; needles, roots: high N > low N; needles: nonmycorrhizal > Laccaria; root tips: Laccaria, Suillus >nonmycorrhizal
Plant Soil (2008) 310:121–136 127
depleted in 15N relative to other pools in all treat-ments, averaging 0.8‰ and 1.5‰ depleted in 15Nrelative to needles in mycorrhizal treatments andnonmycorrhizal treatments, respectively. Needle andstem δ15N were positively correlated (r2=0.30, p<0.001, n=60), root tip δ15N was negatively correlatedwith both needle δ15N (r2=0.19, p=0.001, n=49) andstem δ15N (r2=0.21, p<0.001, n=49), and root δ15Nwas uncorrelated with all other pools.
Needle and stem δ15N declined significantly inmycorrhizal treatments relative to nonmycorrhizaltreatments. When averaged by treatment, colonizationby mycorrhizal fungi decreased foliar δ15N in ammo-nium-supplied cultures by 2.6±0.4‰ (n=4) and innitrate-supplied cultures by 1.3±0.3‰ (Table 4).Colonization decreased stem δ15N in ammonium-supplied cultures by 1.1±0.3‰ and in nitrate-suppliedcultures by 1.5±0.5‰.
Unlike other pools, root tips increased in δ15N inmycorrhizal treatments relative to nonmycorrhizaltreatments. The average increase in nitrate treatmentswas 0.6‰ and the average increase in ammoniumtreatments was 1.6‰. Treatment means for root tipsranged from 1.9‰ for ammonium-supplied Suillus
treatments at high N to −1.9‰ for ammonium-supplied nonmycorrhizal treatments at low N. Roottips in ammonium treatments were 1.2‰ higher thanroot tips in nitrate treatments.
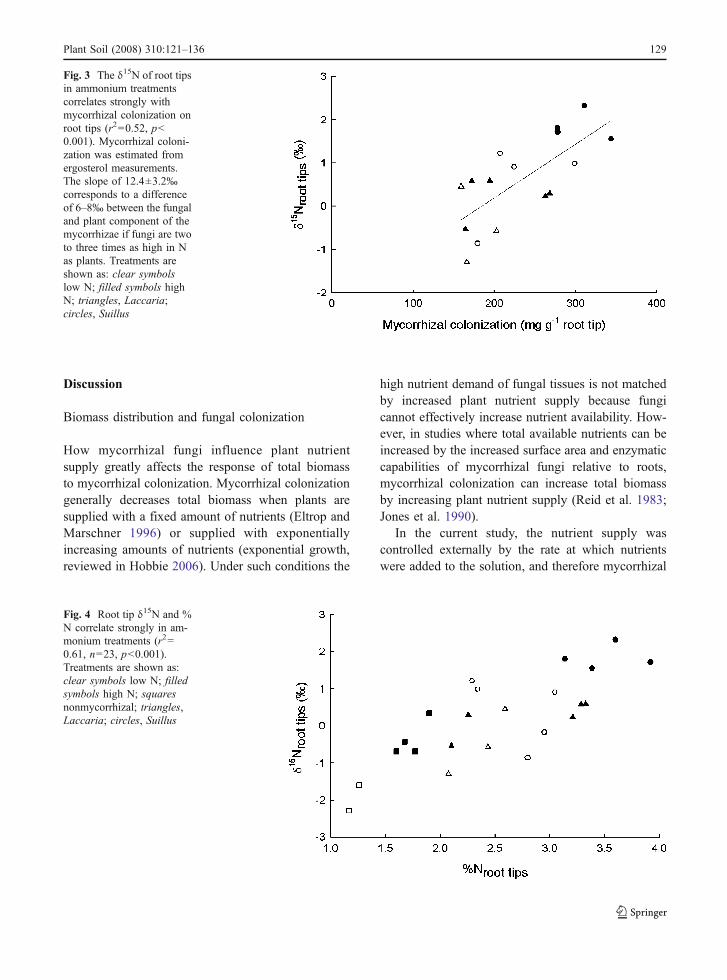
Because root tips varied in the amount of fungalbiomass and this fungal biomass differed in δ15N and%N from root biomass, comparing root tip δ15N withN concentrations and mycorrhizal colonization canprovide insights into N dynamics between mycorrhi-zal fungi and host plants. Root tip δ15N correlatedwith fungal biomass on root tips in ammoniumtreatments (r2=0.52, p=0.011, slope=12.4±3.2‰,Fig. 3) but not in nitrate treatments (r2=0.00, p=0.842). Root tip δ15N and %N were also significantlycorrelated overall (r2=0.41) but differed widelyamong the three associate treatments. Root tip δ15Nand %N were not correlated in nonmycorrhizaltreatments, weakly correlated in Laccaria treatments(r2=0.25, p=0.039), and strongly correlated in Suillustreatments (r2=0.62, p<0.001). When examinedseparately by N form, root tip δ15N and %N werehighly correlated in ammonium treatments (r2=0.61,p<0.001, Fig. 4) but uncorrelated in nitrate treatments(r2=0.13, p=0.066).
Table 4 Effects of N form, N availability, and fungal associate on δ15N (‰) of needles, stems, roots, and root tips by ANOVA
N form Ammonium supply Nitrate supply
Treatment Needles Stems Roots Root tips Needles Stems Roots Root tips
High NNM −0.7±0.3 −3.9±0.5 −1.3±0.4 −0.4±0.2 −1.1±0.3 −2.6±0.8 −1.7±0.2 −1.5±0.3Laccaria −3.0±0.5 −4.3±0.4 −1.5±0.4 0.2±0.2 −3.3±0.4 −3.9±0.2 −1.5±0.2 −1.0±0.4Suillus −4.2±0.9 −4.9±0.5 −0.7±0.7 −1.9±0.2 −1.6±0.1 −2.8±0.5 −1.6±0.4 −1.6±0.3Low NNM −1.6±0.5 −2.7±1.4 −0.8±0.3 −1.9±0.3 −1.9±0.6 −2.3±0.5 −1.3±0.5 −1.7±0.3Laccaria −4.6±0.4 −4.7±0.3 −0.5±0.5 −0.5±0.5 −3.3±0.4 −4.5±0.7 −1.6±0.3 −0.9±0.7Suillus −3.3±0.4 −3.8±0.6 −1.0±0.1 −0.4±0.4 −3.2±0.4 −4.7±0.5 −1.5±0.3 0.7±0.2ANOVAN form (N) 0.028 0.118 0.014 <0.001Supply rate (S) 0.042 0.848 0.227 0.031Associate (A) <0.001 0.005 0.946 <0.001N × S 0.482 0.071 0.530 0.001N × A 0.440 0.821 0.892 0.003S × A 0.761 0.338 0.412 0.451N × S × A 0.003 0.275 0.407 0.333
A post hoc Tukey–Kramer test was used at P=0.05. The source signatures were 1.4‰ for ammonium and −1.7‰ for nitrate. Valueswere not corrected for the contribution of initial N. Tukey–Kramer post hoc test, roots: nitrate > ammonium; root tips: ammonium >nitrate; foliage: high N > Low N; foliage, stems: nonmycorrhizal > Laccaria, Suillus; root tips: Laccaria, Suillus > nonmycorrhizal
NM nonmycorrhizal
128 Plant Soil (2008) 310:121–136
Discussion
Biomass distribution and fungal colonization
How mycorrhizal fungi influence plant nutrientsupply greatly affects the response of total biomassto mycorrhizal colonization. Mycorrhizal colonizationgenerally decreases total biomass when plants aresupplied with a fixed amount of nutrients (Eltrop andMarschner 1996) or supplied with exponentiallyincreasing amounts of nutrients (exponential growth,reviewed in Hobbie 2006). Under such conditions the
high nutrient demand of fungal tissues is not matchedby increased plant nutrient supply because fungicannot effectively increase nutrient availability. How-ever, in studies where total available nutrients can beincreased by the increased surface area and enzymaticcapabilities of mycorrhizal fungi relative to roots,mycorrhizal colonization can increase total biomassby increasing plant nutrient supply (Reid et al. 1983;Jones et al. 1990).
In the current study, the nutrient supply wascontrolled externally by the rate at which nutrientswere added to the solution, and therefore mycorrhizal
Fig. 3 The δ15N of root tipsin ammonium treatmentscorrelates strongly withmycorrhizal colonization onroot tips (r2=0.52, p<0.001). Mycorrhizal coloni-zation was estimated fromergosterol measurements.The slope of 12.4±3.2‰corresponds to a differenceof 6–8‰ between the fungaland plant component of themycorrhizae if fungi are twoto three times as high in Nas plants. Treatments areshown as: clear symbolslow N; filled symbols highN; triangles, Laccaria;circles, Suillus
Fig. 4 Root tip δ15N and %N correlate strongly in am-monium treatments (r2=0.61, n=23, p<0.001).Treatments are shown as:clear symbols low N; filledsymbols high N; squaresnonmycorrhizal; triangles,Laccaria; circles, Suillus
Plant Soil (2008) 310:121–136 129

colonization could not improve nutrient supply.Accordingly, the smaller biomass recorded here formycorrhizal treatments probably reflects the highsequestration of N in fungal tissues for a given carbonallocation belowground (low fungal C/N). This Nsequestration should also restrict the N supply toshoots for growing new photosynthetic machinery,and should therefore limit plant biomass accumula-tion. An additional factor hypothesized to limitbiomass accumulation is the high respiration rates ofmycorrhizal fungi relative to plant tissues (Andersenand Rygiewicz 1995; Eltrop and Marschner 1996).However, the metabolic efficiency with which fungiconvert sugars to biomass does not appear lower thanthat of heterotrophic tissues in plants (e.g., Henn andChapela 2000), and the ratio of respiration to biomassappears generally similar in mycorrhizal and non-mycorrhizal root systems in Pinus culture (Colpaertet al. 1996). Accordingly, N retention appears to bethe primary mechanism accounting for biomassdifferences between mycorrhizal and nonmycorrhizalcultures.
In general, higher biomass is expected withammonium nutrition than with nitrate nutrition be-cause of the extra energy required to reduce nitrateto ammonium prior to incorporation into biomass(Raven et al. 1992). Here, biomass in nitrate treat-ments averaged 87% of that in ammonium treatments,with the average of the two nonmycorrhizal treat-ments at 76% and the two mycorrhizal low Ntreatments also low (75% for Suillus and 84% forLaccaria). In contrast, the two mycorrhizal treatmentsat high N had similar biomass with nitrate nutritionthan with ammonium nutrition. This unusual patternpresumably arose because the fungal ability to reduceand assimilate nitrate at high nitrate supply was lessthan the fungal ability to assimilate ammonium. As aconsequence, exudates of amino acids from pine roots(e.g., Laheurte et al. 1990) may have been animportant N source for fungi in high nitrate treatmentsfor their N supply and the fungal functioning as acarbon sink (particularly the extraradical hyphae) mayhave been impaired as a result. For example, theproportion of total Laccaria biomass in perliteaveraged 39% for high nitrate treatments versus50% for other treatments, whereas Suillus in perliteaveraged 18% for high nitrate treatments versus 31%for other treatments (Table 2), accounting for a highlysignificant N form × supply rate interaction (p<
0.001). Patterns of root tip %N also suggest that highnitrate limited fungal N retention, with root tip %Nhigher in high ammonium than in low ammoniumtreatments but equivalent in high nitrate and lownitrate treatments (Table 3), accounting for a highlysignificant N form × supply rate interaction (p=0.001). We conclude that the ability of fungi toassimilate N and the high N concentration of fungaltissues limited N supply to host plants under low Navailability and under ammonium nutrition. Theproportion of available N assimilated by mycorrhizalfungi was therefore less at high nitrate availabilitythan at other N form × N availability combinations.his higher plant N availability would presumably leadto greater plant biomass production under theseconditions.
In contrast to biomass patterns in nitrate treat-ments, the large reduction of biomass in mycorrhizaltreatments compared to nonmycorrhizal treatments athigh ammonium supply (44% reduction versus anaverage reduction of 22% in other treatments)indicated that N retention by mycorrhizal fungi wasvery high under these conditions. This is furthersupported by high values at high ammonium supplyfor the fungal proportion of total biomass (22%,Table 2) and the high N concentrations in mycorrhizalroot tips (2.8% for Laccaria, 3.5% for Suillus,Table 3). We note that these patterns are driven bythe high colonization in high ammonium treatmentsby Suillus, as the fungal proportion of total biomassdid not vary significantly across the four Laccariatreatments (Table 2). Separate ANOVAs for Suillustreatments and Laccaria treatments show that thefungal proportion of total biomass for Suillus wassignificantly higher for ammonium than for nitratetreatments (p<0.001) but was unaffected by N formin Laccaria treatments (p=0.079). The high propor-tion of total biomass attributed to Suillus at highammonium supply and the low fungal proportion oftotal biomass attributed to Suillus at high nitratesupply resulted in a significant N form × supplyinteraction (p=0.009); the N form × supply interac-tion was not significant for Laccaria treatments (p=0.950).
These biomass values correspond to exponentialgrowth rates (Fig. 1). By making some simpleassumptions, we can calculate the effect of the extraenergetic cost of nitrate nutrition versus ammoniumnutrition on plant growth rates. Ek (1997) reported for
130 Plant Soil (2008) 310:121–136

Betula pendula–Paxillus involutus symbioses thatoverall respiration for nitrate nutrition was twice thatwith ammonium nutrition, at 9.2 mol versus 4 molCO2 per mole N assimilated. Assuming that nitratenutrition requires an additional 5 mol of carbon permole of assimilated N relative to ammonium nutri-tion, metabolic efficiency is 50% for growth withammonium, and an average molar C:N is 40, thenammonium nutrition will require 80 mol of carbon togrow 40 mol and nitrate nutrition will require 85 molof carbon to grow 40 mol. Therefore, the growth rateof nitrate nutrition will be theoretically 80/85 (94%)that of ammonium nutrition. The growth rate at low Nsupply with nitrate relative to ammonium was close tothis (averaging 89%), whereas the growth rates athigh nitrate supply with Suillus, Laccaria, and non-mycorrhizal cultures were 104%, 100%, and 90% ofthat with high ammonium supply, respectively. Thehigh growth rates (and accordingly, high biomass) inmycorrhizal treatments at high nitrate supply arepresumably explained by the relatively low mycor-rhizal ability for nitrate assimilation and sequestrationin these treatments, as previously discussed.
The generally lower belowground allocation inSuillus (63±1%) than in Laccaria treatments (71±1%) may stem from the relatively high proportion ofammonium-derived N transferred from suilloid fungi,including Suillus, to host pines (Bidartondo et al.2001). A high N supply from roots to shoots willgenerally decrease the proportion of fixed carbonallocated belowground (Ågren and Bosatta 1996).Similarly, if Laccaria sequesters more of the N ittakes up than Suillus, then this should restrict plant Nsupply and increase belowground carbon allocationby the host plants. Here, the higher biomass ofmycorrhizal fungi in Laccaria than in Suillus culturesshould lead to a high fungal N sequestration inLaccaria cultures, restricting plant N supply andtherefore increasing plant carbon allocation to mycor-rhizal fungi.
Nitrogen fluxes and nitrogen isotope patterns
Foliar δ15N values generally decreased with mycor-rhizal colonization, in agreement with previousculture studies (Högberg et al. 1999; Kohzu et al.2000; Hobbie and Colpaert 2003). Previous studieswith Pinus sylvestris reported that foliar δ15N de-clined 0.5–2.8‰ with mycorrhizal colonization under
ammonium nitrate nutrition (Hobbie and Colpaert2003), declined 1.5–3.0‰ with ammonium nutrition(Högberg et al. 1999), and declined less than 1.0‰with nitrate nutrition (Högberg et al. 1999). Thesevalues can be compared to those in the present study,with a decline in foliar δ15N of 1.7–3.5‰ withammonium nutrition and 0.5–2.2‰ with nitratenutrition. The transfer of 15N-depleted N from fungito plants coupled with N retention by fungi appears tocause these patterns and also result in 15N-enrichedfungal biomass. The 15N patterns can thereforeprovide insight into N partitioning between mycor-rhizal fungi and host plants.
The hypothesis that nitrate would fractionatedifferently than ammonium was partially supportedby our results, with fractionation for the two N formsdiffering for cultures with Suillus but not forLaccaria. Suillus colonization at high nitrate avail-ability was reasonably high (about 15% of root mass)but δ15N values for all tissues were unchanged fromnonmycorrhizal cultures. Additional support for dif-ferential fractionation of nitrate versus ammoniumwas reported in a study by Högberg et al. (1999), inwhich rhizomorphs of Suillus bovinus were 4.9‰enriched in 15N relative to Pinus sylvestris shootswith ammonium nutrition but were not enriched in15N relative to Pinus sylvestris shoots with nitratenutrition. Two explanations for these patterns appearplausible: either nitrate was assimilated by plants andreduced N subsequently passed to fungi, or nitratewas assimilated separately by plants and fungi andreduced N was not exchanged between the symbionts.These two possibilities cannot be distinguished usingδ15N patterns but could be distinguished if nitratereductase activity in fungal hyphae and plant rootscould be separately determined. Because both mycor-rhizal fungi and pines secrete amino acids (Laheurteet al. 1990; Müller et al. 2007), the relative N status,secretion rates, and uptake capacities of the plant andfungal partners should determine the net transfer of Nbetween plants and fungi.
Another potential marker of the transfer of reducedN from plants to fungi is the relative fungal biomassin extraradical hyphae versus in ectomycorrhizae. Ifmycorrhizal fungi derive their N from plants ratherthan directly from the culture media, then mycorrhi-zae will obtain N before extraradical hyphae, and onemight expect to see less biomass in extraradicalhyphae as well. The small relative allocation to
Plant Soil (2008) 310:121–136 131

extraradical hyphae in the high nitrate treatment withSuillus cultures, with only ~18% of fungal biomass inextraradical hyphae (Table 2), therefore agrees withless nitrate assimilation by fungi in this treatment.
Further evidence of the different fates of nitrateversus ammonium comes from measurements ofrespiration and 15N labeling in culture studies. Basedon 15N labeling patterns in Betula and Piceasymbiotic with Paxillus involutus, Ek et al. (1994)suggested that nitrate, but not ammonium, was passedfrom fungi to plants without metabolic assimilation.Under such conditions, no isotopic fractionation isexpected. Similarly, in Picea-Pisolithus symbioses,mycorrhizal respiration rates were estimated at 3.4times higher than root respiration rates under nitratenutrition and 11.1 times higher with ammoniumnutrition (Eltrop and Marschner 1996), presumablyindicating little involvement of mycorrhizal fungi innitrate reduction.
Partial nitrate reduction in roots can theoreticallyenrich foliage in 15N relative to roots (Robinson et al.1998; Evans 2001). Although pines are generallyconsidered to have low nitrate reductase activity inneedles (Plassard et al. 1991; Downs et al. 1993), theproportion of nitrate reduction in needles (relative toroots) increases with increasing nitrate availability(Sarjala 1991). However, the lack of 15N enrichmenthere of foliage relative to roots suggests that all nitratewas assimilated by the roots and associated ectomy-corrhizal fungi. Alternatively, recycling of N betweenfoliage and roots could diminish 15N differencesbetween roots and foliage and result in root andplants behaving effectively as a single isotopic pool.
Based on our previous study using ammoniumnitrate as the N source, we hypothesized that needleδ15N should correlate strongly and negatively withmycorrhizal colonization. This did not turn out to bethe case, with needle δ15N uncorrelated with thefungal fraction of total biomass. Because not allapplied N was recovered and the recovered N inammonium treatments was depleted in 15N relative tothe supplied N, we cannot distinguish betweenpossible fractionation against 15N during N lossprocesses and fractionation against 15N during initialuptake. However, differences between root tips andfoliage in δ15N reflect mycorrhizal transfer of 15N-depleted N to plants and the consequent retention of15N-enriched N in fungal tissues. Thus, the value
δ15Nroot tips–needles should reflect fungal biomass.Interestingly, δ15Nroot tips–needles correlated stronglywith the fungal proportion of total biomass for Suillustreatments (r2=0.50, p<0.001) but not for Laccariatreatments (r2=0.04, p=0.486; data not shown).Several factors could contribute to this, including (1)whether fungal N transfer is proportional to fungalbiomass, (2) how closely total fungal biomasscorrelates with the fungal colonization of fine roots,and (3) whether 15N discrimination processes are thesame in these two fungal species. In addition, whetherfungal N can be treated as a single well-mixed pool orwhether only a portion of fungal N is labile enough toparticipate in the reactions that fractionate against 15Nwill also influence 15N partitioning between ectomy-corrhizal fungi and their host plants (Hobbie andColpaert 2003).
Relatively large increases in 15N content ofmycorrhizal root tips compared to uncolonized roottips with ammonium nutrition (mean 1.7‰) indicatedfractionation against 15N during creation of com-pounds transferred from fungi to host plants. Incontrast, small increases in 15N content of mycor-rhizally colonized root tips relative to uncolonizedroot tips with nitrate nutrition (mean 0.5‰) presum-ably indicated less fungal assimilation of nitrate intoreduced N prior to transfer to host plants. However,the ability of Laccaria to use nitrate is well-established and even Suillus appears to have assimi-lated some nitrate at low N availability.
Correlations between δ15N and mycorrhizal colo-nization of root tips with ammonium nutritionindicate that the relative proportion of 15N-enrichedfungal N and 15N-depleted plant N in root tips causesthese patterns (Fig. 3). The slope of the regression,12.4±3.2‰, can be used to estimate the 15Nenrichment between plants and fungal matter byaccounting for the different N concentrations infungal sheaths and plant root tips. If the N concen-tration in fungi is two to three times that of plant roottips, then a root tip composed of 25% fungal matterconsists of 40–50% fungal N. For a root tipenrichment in 15N of 3‰ relative to foliage, fungalN would be 6–8‰ enriched in 15N relative to plant N.
The estimated fungal enrichment of 6–8‰ issomewhat higher than in previous culture studies.For example, Hobbie and Colpaert (2003) calculatedin a culture study using ammonium nitrate that fungal
132 Plant Soil (2008) 310:121–136

sheaths of Suillus luteus and Thelephora terrestrisaveraged 5.5‰ and 4.4‰ enriched in 15N relative topine roots, Kohzu et al. (2000) reported that fungalmats of Suillus granulatus were 3.3‰ enriched in 15Nrelative to pine needles, and Högberg et al. (1996)measured fungal sheaths from Picea roots as 2.4–6.4‰ enriched relative to the root core in a fieldstudy. We believe that these differing patterns can bereconciled if ammonium is assumed to be the N formresponsible for most 15N fractionation between plantsand fungi and nitrate diminishes the overall fraction-ation between plants and fungi.
These results have several implications for inter-preting δ15N patterns in ectomycorrhizal plants andfungi. Other researchers have suggested that the lowδ15N of mycorrhizal fruiting bodies of Laccariarelative to that of fungi such as Suillus reflectsdifferent N sources for these fungi, with Suillus ableto use complex organic substrates and Laccariaspecializing on inorganic N sources (Taylor et al.2003; Hobbie and Hobbie 2006). With only a singleavailable N source, roughly equal colonization levelsof root tips by Laccaria and Suillus with ammoniumnutrition led to Laccaria-colonized root tips averaging1.3‰ depleted in 15N relative to Suillus-colonizedroot tips. This indicates two possibilities: either theintrinsic processing of N by Laccaria produces lessfractionation against 15N than for Suillus or Laccariatransfers less N than Suillus under identical growthconditions, and therefore is less enriched in 15N thanSuillus. In either case, 15N-depleted fruiting bodies inLaccaria do not necessarily reflect uptake of differentN sources than those used by 15N-enriched fungaltaxa. The positive correlation between root tip δ15Nand fungal colonization in Suillus but not in Laccaria(Fig. 3) also suggests that the processes causingdifferences between fungal and plant δ15N are weakerin Laccaria cultures than in Suillus cultures.
Differences between Laccaria and Suillus in Nretention may influence the cycling of carbon and Nin the plant-fungal symbiosis. If Laccaria transfers asmaller fraction of the N it assimilates to host plantsthan Suillus, this would lead to generally smallerfungal-plant differences in 15N content than withSuillus. By assimilating for its own use much of theavailable nitrate, Laccaria can remain a strong sinkfor plant carbohydrates. With nitrate nutrition, fungalbiomass is therefore considerably higher in Laccaria
than in Suillus. Strong N retention by Laccaria at lownitrate supply presumably accounts for the very lowfoliar N (1.05%) in this treatment (Table 2).
In cultures at low nitrate availability, the relativedepletion in 15N of foliage relative to root tips inmycorrhizal cultures of 2.4–2.5‰ and the depletion in15N of foliage relative to nonmycorrhizal foliage of1.3–1.4‰ suggest that both Laccaria and Suillus arereducing nitrate to ammonium that is then assimilatedby the fungi (Table 3). In contrast, similar δ15N valuesin nonmycorrhizal cultures and Suillus cultures athigh nitrate availability suggest that nitrate must bereduced within plant roots and then transferred fromplants to fungi. Thus, under these conditions Suillusdepends at least in part on plant-derived N whereasLaccaria can still obtain its own nitrogen.
Nitrogen dynamics in ectomycorrhizal symbioseswith ammonium and nitrate nutrition are shownschematically in Fig. 5. In this figure, ammonium isprimarily assimilated by fungi whereas nitrate isassimilated by both fungi and plants, depending onfungal species and fungal N status. Nitrate nutrition isshown separately into Suillus and Laccaria, sincebehavior of those two symbionts was quite different,particularly at high nitrate supply rates. Nitrate that isnot assimilated by fungi is taken up by plants withoutfractionation, whereupon it is reduced and assimilat-ed. A portion of plant-derived amino acids may besecreted from plant root tips into the intracellularspace where it is available for either uptake by fungior reuptake by plants. Such uptake by ectomycor-rhizal fungi of plant-derived amino acids (specifically,of glutamate) has long been hypothesized as part of aglutamine-glutamate shuttle between fungi and plants(Martin et al. 1986; Smith and Smith 1990). Such netN transfers from plants to fungi are probably morecommon at high plant N status than at low plant Nstatus, and would reduce 15N differences betweenfungi and plants.
The pattern of N dynamics shown in Fig. 5 isfurther supported by a study of the effects ofammonium versus nitrate nutrition in ectomycorrhizalsymbioses using 15N-labeled ammonium nitrate(He et al. 2005). In this study, 15NH4
14NO3 and14NH4
15NO3 were supplied to common mycorrhizalnetworks of Pisolithus connected to Eucalyptus andCasuarina plants. The N flux from the donorcompartment to the receiver plant through the
Plant Soil (2008) 310:121–136 133

mycorrhizal network was about twice as great forammonium than for nitrate, presumably becauseammonium assimilation by mycorrhizal fungi couldbe followed directly by transfer of organic N to eitherplant, whereas transfer of nitrate-derived N fromdonor compartments to receiver plants would requirenitrate reduction and assimilation by donor plants,subsequent transfer of N from plant to fungus, andthen N transfer from fungus to receiver plant. InPinus–Hebeloma symbioses, transfer of reduced Ncompounds from fungi to plants was suggested torepress plant nitrite reductase (Bailly et al. 2007), anda similar repression of fungal nitrite reductase inmycorrhizae but not in extraradical hyphae (Bailly etal. 2007) may very well have been caused by fluxesof reduced N from plants to fungi.
The primary step creating differences in plant δ15Nacross different culture conditions appears to be therelative importance of synthesis of 15N-depletedcompounds by fungi prior to transfer to host plants.Because nitrate is only partially reduced by fungi,particularly at high N availabilities, fractionationbetween plants and fungi diminishes with nitratenutrition. In addition, transfer of N from plants tofungi further diminishes the isotopic differencebetween these two pools. These mechanisms canplausibly explain much of the reported variability inplant δ15N signatures across gradients in N availabil-
ity of field studies (Garten 1993; Emmett et al. 1998;Hobbie et al. 2000).
Analytical solutions to δ15N patterns in mycorrhizalfungi and plants
General equations for plant and fungal δ15N thatincorporate the supply of N from plants to fungi canbe established. In Hobbie et al. (2000), the authorsproposed a series of analytical equations to accountfor the influence of mycorrhizal fungi and Npartitioning on plant and fungal δ15N. The mostgeneral expression for plant δ15N was given as:
δ15Nplant ¼ δ15NavailableN � f 1� Trð Þ$ ð1Þwith f the proportion of plant N derived frommycorrhizal fungi, Tr the proportion of N taken upby mycorrhizal fungi that was subsequently passed onto plants, and $ the isotopic discrimination against 15Nduring the synthesis of fungal transfer compounds.
The equivalent expression for fungal δ15N was
δ15Nfungi ¼ δ15NavailableN þ Tr$ ð2Þ
These expressions did not account for the possiblesupply of N from plants to mycorrhizal fungi. If oneassumes that a portion of fungal N can be derivedfrom plants (fp) and a portion from fungal uptake (ff),
Fig. 5 Schematic of N and C dynamics in mycorrhizal systemsunder ammonium and nitrate nutrition. Nitrate nutrition isseparated into Suillus and Laccaria systems, since behavior ofthose two symbionts was quite different, particularly at highnitrate supply rates. Thickness of arrows represents the relativeflux and size of boxes represents the relative biomass within a
pool. Net fluxes are shown, although some fluxes areundoubtedly bidirectional, such as fluxes of assimilated N asamino acids between plants and mycorrhizal fungi. Arrowsfrom inorganic N represent probable sites of assimilation intoorganic N
134 Plant Soil (2008) 310:121–136

with ff+fp=1, then Eq. 2 can be transformed into thefollowing equation:
δ15Nfungi ¼ δ15NavailableN þ $ f fTr þ f pf Tr � 1ð Þ� � ð3ÞThe quantity (Tr−1) is always negative. Therefore,δ15Nfungi will decrease as the fraction of fungal Nderived from plants increases, and the isotopicdifference between fungi and plants will also decrease.
The 15N difference between plants and fungi canbe calculated as:
δ15Nfungi � δ15Nplants ¼ $ff Tr þ f � fTrð Þ ð4ÞThese equations provide an analytical framework forinterpreting plant and fungal δ15N patterns undervariable N availability. For example, similar δ15Nvalues of needles and root tips (both −1.6‰) in theSuillus treatment at high nitrate supply suggest thatmost of the fungal N in root tips must be derived fromplants, and ff must be quite low for this specifictreatment.
This treatment can be contrasted with calculatedpatterns in the Laccaria treatment at low ammoniumsupply where the 15N enrichment in root tips relativeto foliage was highest (4.1‰). In this treatment, roottip %N is about twice that in the equivalent non-mycorrhizal treatment, and fungal colonization of roottips is about 20%. Accordingly, root tip N is about60% fungal, and the 4.1‰ enrichment in 15N of roottips relative to foliage in this treatment corresponds toa value of δ15Nfungi − δ15Nplants of 6.8‰ (4.1‰/0.6).Since the maximum value of the quantity Tr+ f− fTr inEq. 4 is 1, then for a $ value of 9‰, ff is at least 6.8/9,or 76%. These formulations do not depend on thespecific compound that is transferred from fungi toplants, and can readily accommodate recent sugges-tions that ammonia may be the transfer compound inectomycorrhizal symbioses (Chalot et al. 2006).
Acknowledgment This work was supported by NSF DEB-0235727 and by the Fund for Scientific Research-Flanders(Belgium; project G. 0033.03). Assistance with labwork byCarine Put and Julia Peters and comments on the manuscript byJohn Hobbie, Joe Craine, and several anonymous reviewers aregratefully acknowledged.
References
Aber JD, McDowell WH, Nadelhoffer KJ, Magill AH,Berntson G, Kamekea M, McNulty SG, Currie W, RustadL, Fernandez I (1998) Nitrogen saturation in temperate
forest ecosystems: hypotheses revisited. BioScience48:921–934
Ågren GI, Bosatta E (1996) Theoretical ecosystem ecology.Cambridge University Press, Cambridge
Andersen CP, Rygiewicz PT (1995) Allocation of carbon inmycorrhizal Pinus ponderosa seedlings exposed to ozone.New Phytol 131:471–480
Bailly J, Debaud JC, Verner MC, Plassard C, Chalot M,Marmeisse R, Fraissinet-Tachet L (2007) How does asymbiotic fungus modulate expression of its host-plantnitrite reductase? New Phytol 175:155–165
Bidartondo MI, Ek H, Wallander H, Söderström B (2001) Donutrient additions alter carbon sink strength of ectomycor-rhizal fungi? New Phytol 151:543–550
Brearley FQ, Press MC, Scholes JD (2003) Nutrients obtainedfrom leaf litter can improve the growth of dipterocarpseedlings. New Phytol 160:101–110
Buchmann N, Gebauer G, Schulze ED (1996) Partitioning of15N-labeled ammonium and nitrate among soil, litter,below- and above-ground biomass of trees and understoryin a 15-year-old Picea abies plantation. Biogeochemistry33:1–23
Chalot M, Blaudez D, Brun A (2006) Ammonia: a candidatefor nitrogen transfer at the mycorrhizal interface. TrendsPlant Sci 11:263–266
Colpaert JV, Van Laere A, Van Assche JA (1996) Carbon andnitrogen allocation in ectomycorrhizal and non-mycorrhizalPinus sylvestris L. seedlings. Tree Physiol 16:787–793
Colpaert JV, Van Tichelen KK, Van Assche JA, Van Laere A(1999) Short-term phosphorus uptake rates in mycorrhizaland non-mycorrhizal roots of intact Pinus sylvestris seed-lings. New Phytol 143:589–597
Downs MR, Nadelhoffer KJ, Melillo JM, Aber JD (1993)Foliar and fine root nitrate reductase-activity in seedlingsof 4 forest tree species in relation to nitrogen availability.Trees 7:233–236
Ek H (1997) The influence of nitrogen fertilization on the carboneconomy of Paxillus involutus in ectomycorrhizal associ-ation with Betula pendula. New Phytol 135:133–142
Ek H, Andersson S, Arnebrant K, Söderström B (1994) Growthand assimilation of NH4
+ and NO3− by Paxillus involutus
in association with Betula pendula and Picea abies asaffected by substrate pH. New Phytol 128:629–637
Eltrop L, Marschner H (1996) Growth and mineral nutrition ofnon-mycorrhizal and mycorrhizal Norway spruce (Piceaabies) seedlings grown in semi-hydroponic sand cul-ture.1. Growth and mineral nutrient uptake in plantssupplied with different forms of nitrogen. New Phytol133:469–478
Emmett BA, Kjonaas OJ, Gundersen P, Koopmans C, Tietema A,Sleep D (1998) Natural abundance of 15N in forests across anitrogen deposition gradient. For Ecol Manage 101:9–18
Evans RD (2001) Physiological mechanisms influencing plantnitrogen isotope composition. Trends Plant Sci 6:121–126
Finlay RD, Frostegård A, Sonnerfeldt A-M (1992) Utilizationof organic and inorganic nitrogen sources by ectomycor-rhizal fungi in pure culture and in symbiosis with Pinuscontorta Dougl. ex Loud. New Phytol 120:105–115
Garten CT (1993) Variation in foliar 15N abundance and theavailability of soil nitrogen on Walker Branch watershed.Ecology 74:2098–2113
Plant Soil (2008) 310:121–136 135

He X, Critchley C, Ng H, Bledsoe C (2005) Nodulated N2-fixing Casuarina cunninghamiana is the sink for net Ntransfer from non-N2-fixing Eucalyptus maculata via anectomycorrhizal fungus Pisolithus sp. using 15NH4
+ or15NO3
− supplied as ammonium nitrate. New Phytol167:897–912
Henn MR, Chapela IH (2000) Differential C isotope discrim-ination by fungi during decomposition of C3- and C4-derived sucrose. Appl Environ Microbiol 66:4180–4186
Ho I, Trappe JM (1980) Nitrate reductase activity of non-mycorrhizal Douglas-fir rootlets and of some associatedmycorrhizal fungi. Plant Soil 54:395–398
Hobbie EA (2006) Carbon allocation to ectomycorrhizal fungicorrelates with total belowground allocation in culturestudies. Ecology 87:563–569
Hobbie EA, Colpaert JV (2003) Nitrogen availability andcolonization by mycorrhizal fungi correlate with nitrogenisotope patterns in plants. New Phytol 157:115–126
Hobbie JE, Hobbie EA (2006) 15N in symbiotic fungi andplants estimates nitrogen and carbon flux rates in arctictundra. Ecology 87:816–822
Hobbie EA, Wallander H (2006) Integrating ectomycorrhizalfungi into quantitative frameworks of forest carbon andnitrogen cycling. In: Gadd GM (ed) Fungi in biogeochem-ical cycles. Cambridge University Press, Cambridge, pp98–128
Hobbie EA, Macko SA, Williams MT (2000) Correlationsbetween foliar δ15N and nitrogen concentrations mayindicate plant-mycorrhizal interactions. Oecologia122:273–283
Högberg P, Högbom L, Schinkel H, Högberg M, Johannisson C,Wållmark H (1996) 15N abundance of surface soils, rootsand mycorrhizas in profiles of European forest soils.Oecologia 108:207–214
Högberg P, Högberg MN, Quist ME, Ekblad A, Näsholm T(1999) Nitrogen isotope fractionation during nitrogenuptake by ectomycorrhizal and non-mycorrhizal Pinussylvestris. New Phytol 142:569–576
Ingestad T, Kähr M (1985) Nutrition and growth of coniferousseedlings at varied relative nitrogen addition rate. PhysiolPlant 65:109–116
Ingestad T, Arveby AS, Kähr M (1986) The influence ofectomycorrhiza on nitrogen nutrition and growth of Pinussylvestris seedlings. Physiol Plant 68:575–582
Jones MD, Durall DM, Tinker PB (1990) Phosphorus relation-ships and production of extramatrical hyphae by two typesof willow ectomycorrhizas at different soil phosphoruslevels. New Phytol 115:259–267
Kohzu A, Tateishi T, Yamada A, Koba K, Wada E (2000)Nitrogen isotope fractionation during nitrogen transportfrom ectomycorrhizal fungi, Suillus granulatus, to thehost plant, Pinus densiflora. Soil Sci Plant Nutr 46:733–739
Laheurte F, Leyval C, Berthelin J (1990) Root exudates ofmaize, pine and beech seedlings influenced by mycorrhi-zal and bacterial inoculation. Symbiosis 9:111–116
Lipson D, Näsholm T (2001) The unexpected versatility ofplants: organic nitrogen use and availability in terrestrialecosystems. Oecologia 128:305–316
Martin F, Stewart GR, Genetet I, Le Tacon L (1986)Assimilation of 15NH4
+ by beech (Fagus sylvatica L.)ectomycorrhizas. New Phytol 102:85–94
Müller T, Avolio M, Olivi M, Benjdia M, Rikirsch E, Kasaras A,Fitz M, Chalot M, Wipf D (2007) Nitrogen transport in theectomycorrhiza association: the Hebeloma cylindrospo-rum-Pinus pinaster model. Phytochemistry 68:41–51
Nylund JE, Wallander H (1992) Ergosterol analysis as a meansof quantifying mycorrhizal biomass. Methods Microbiol24:77–88
Plassard C, Scheromm P, Mousain D, Salsac L (1991)Assimilation of mineral nitrogen and ion balance in thetwo partners of ectomycorrhizal symbiosis: data andhypothesis. Experientia 47:340–349
Raven JA, Wollenweber B, Handley LL (1992) A comparisonof ammonium and nitrate as nitrogen sources for photo-lithotrophs. New Phytol 121:19–32
Reid CPP, Kidd FA, Ekwebelam SA (1983) Nitrogen nutrition,photosynthesis and carbon allocation in ectomycorrhizalpine. Plant Soil 71:415–432
Robinson D, Handley LL, Scrimgeour CM (1998) A theory for15N/14N fractionation in nitrate-grown vascular plants.Planta 205:397–406
Rygiewicz PT, Bledsoe CS, Zasoski RJ (1984a) Effects ofectomycorrhizae and solution pH on 15N-ammonium uptakeby coniferous seedlings. Can J For Res 14:885–892
Rygiewicz PT, Bledsoe CS, Zasoski RJ (1984b) Effects ofectomycorrhizae and solution pH on 15N-nitrate uptake byconiferous seedlings. Can J For Res 14:893–899
Sarjala T (1990) Effect of nitrate and ammonium concentrationon nitrate reductase activity in five species of mycorrhizalfungi. Physiologia Plantarum 79:65–70
Sarjala T (1991) Effect of mycorrhiza and nitrate nutrition onnitrate reductase activity in Scots pine seedlings. PhysiolPlant 81:89–94
Schimel JP, Bennett J (2004) Nitrogen mineralization: chal-lenges of a changing paradigm. Ecology 85:591–602
Smith SE, Smith FA (1990) Structure and function of theinterfaces in biotrophic symbioses as they relate to nutrienttransport. New Phytol 114:1–38
Taylor AFS, Fransson PM, Högberg P, Högberg MN, Plam-boeck AH (2003) Species level patterns in 13C and 15Nabundance of ectomycorrhizal and saprotrophic fungalsporocarps. New Phytol 159:757–774
Van Tichelen KK, Colpaert JV (2000) Kinetics of phosphateabsorption by mycorrhizal and non-mycorrhizal Scots pineseedlings. Physiol Plant 110:96–103
136 Plant Soil (2008) 310:121–136