neuropsychological evidence for a topographical learning...
TRANSCRIPT

NEUROPSY CHOLOGICAL EVIDENCE FOR ATOPOGRAPHICAL LEARNING MECHANISM IN
PARAHIPPOCAMPAL CORTEX
Russell EpsteinMedical Research Council Cognition and Brain Sciences Unit Cambridge UK and MIT Department of Brain and Cognitive Sciences
Cambridge USA
Edgar A DeYoeMedical College of Wisconsin Milwaukee USA
Daniel Z PressBeth Israel Deaconess Medical Center Boston USA
Allyson C RosenStanford University Stanford USA
Nancy KanwisherMIT Department of Brain and Cognitive Sciences Cambridge USA
The Parahippocampal Place Area (PPA Epstein amp Kanwisher 1998) is a region within posteriorparahippocampal cortex that responds selectively to visual stimuli that convey information about thelayout of local space Here we describe two patients who suffered damage to the PPA after vascular inci-dents Both subsequently exhibited memory problems for topographical materials and were unable tonavigate unassisted in unfamiliar environments Performance on a continuous n-back visual memorytest was significantly lower for novel scene-like stimuli than for novel object-like stimuli In contrastperformance was normal on a famous landmark recognition task and on two perceptual tasks thatrequired on-line analysis of scene geometry Both patients were able to produce accurate maps ofpremorbidly learned places but were unable to produce accurate maps of new places These results con-verge with previous neuroimaging work to demonstrate that the PPA (1) is selectively involved in pro-cessing information about the geometry of surrounding space and (2) may play a more critical role inthe encoding of this information into memory than in the initial perceptual processing recognition orrecall of this information
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 481ndash508
Oacute 2001 Psychology Press Ltdhttpwwwtandfcoukjournalspp02643294html DOI10108002643290042000215
481
Requests for reprints should be addressed to Russell Epstein MRC Cognition amp Brain Sciences Unit 15 Chaucer Road Cam-bridge CB2 2EF UK (Tel +44 (0) 1223-355-294 ext 800Fax +44 (0) 1223-359-062Email russellepsteinmrc-cbucamacuk)
We thank Damian Stanley and Alison Harris for assistance with these experiments Stephan Heckers and Facundo Manes foranatomical advice Kim Graham for comments on the manuscript Tom Hammeke for advice about neuropsychological testsMatthew Brett for assistance with MRI normalisation and Marvin Chun Frances Wang and Liz Spelke for helpful discussions Inaddition we would also like to express our thanks to GR and CO for their cheerful and enthusiastic participation in this research Thisresearch was supported by NIMH postdoctoral fellowship MH11459 to RE NEI and NIMH grants EY10244 and MH51358 toEAD and NIMH grant MH56037 to NK
supplied as Poewrpoitn files coped into document [1 2 3 4 567 9 11] Scanned amp edited [8 10 12]
INTRODUCTION
All mobile organisms must solve the fundamentalproblem of navigation One way to gain insight intothe cognitive and neural mechanisms underlyingnavigation is to investigate the functional organisa-tion of the brain areas involved Research on ani-mals and humans has suggested that a number ofregions play a role including the parietal lobes(Stark Coslett amp Saffran 1996) retrosplenial cor-tex (Chen Lin Green Barnes amp McNaughton1994)hippocampus (Maguire et al 1998OrsquoKeefeamp Nadel 1978) and parahippocampal cortex(Aguirre Detre Alsop amp DrsquoEsposito 1996Bohbot et al 1998) Here we focus on para-hippocampal cortex We describe two patients whosuffered damage to this region of cortex after vascu-lar incidents Both subsequently developed severedeficits in the topographical domain which wedefine as the domain of information relevant tospatial navigation
Previous reports in the neuropsychological liter-ature have described patients with ldquotopographicaldisorientationrdquo who are unable to successfully findtheir way through locomotor space but do notexhibit general cognitive or memory impairments(see Aguirre amp DrsquoEsposito 1999 for review) Thenature of the specific problem varies from patient topatient For example some patients show an inabil-ity to understand spatial relationships between dif-ferent locations (Levine Warach amp Farah 1985patient 2) whereas others are unable to recognisenavigationally relevant stimuli such as landmarksbut can accurately describe routes between theplaces they cannot identify (Pallis 1955) Patientsalso differ in their relative ability to handle novel vsfamiliar topographical information Some are ableto navigate through familiar environments or recallold topographical information but cannot learnnovel environments (Habib amp Sirigu 1987patients 1 and 2 Ross 1980Teng amp Squire 1999)whereas others are impaired at both learning of newmaterials and recognitionrecall of old materials(Habib amp Sirigu 1987 patients 3 and 4 Incisadella Rocchetta Cipolotti amp Warrington 1996Landis Cummings Benson amp Palmer 1986
McCarthy Evans amp Hodges 1996 Whiteley ampWarrington 1978)
Based on comprehensive review of the topo-graphical disorientation literature Aguirre andDrsquoEsposito (1999) argued that the lingual-parahippocampal region is one of several areas thatare critically involved in navigation Damage to thisregion often results in an inability to use salient top-ographical features such as landscapes and build-ings to orient oneself (ldquolandmark agnosiardquo egLandis et al 1986 Pallis 1955) or in a more gen-eral inability to learn new topographical informa-tion (ldquoanterograde disorientationrdquo eg Habib ampSirigu 1987 patients 1 and 2) In a recent study of127 patients with focal lesions Barrash DamasioAdolphs and Tranel (2000) observed that patientswith damage to this region almost always displayedsevere impairment on a real-world route learningtask Furthermore Bohbot et al (1998) found thatpatients with right-hemisphere parahippocampalcortex lesions were impaired at a human analogueof the Morris water maze task in which they wererequired to remember the location of a hidden floorplatform within a room (a deficit not found inpatients whose damage was restricted to the hippo-campus proper)
Neuroimaging studies have also implicated thelingual-parahippocampal region in navigationSeveral studies have examined brain activity in nor-mal subjects during performance of a number ofnavigational tasks including navigation through avirtual reality environment (Aguirre et al 1996)watching videotapes depicting navigation througha real environment (Maguire Frackowiak amp Frith1997) and mentally imagining navigation througha real environment (Ghaem et al 1997 Maguire etal 1998) All of these studies found greater activa-tion in the parahippocampal region (and sometimesother medial temporal regions as well Ghaem etal 1997Maguire et al 1997 1998) when the nav-igation task was compared to a less navigationallydemanding control task
What role does parahippocampal cortex play inthese navigational tasks In earlier work weaddressed this question by using fMRI to examineneural activity in parahippocampal cortex and other
EPSTEIN ET AL
482 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
brain regions while subjects viewed a wide variety ofvisual stimuli (Epstein amp Kanwisher 1998) Wefound that a region abutting the collateral sulcusnear the parahippocampal-lingual boundaryresponded significantly more strongly when sub-jects viewed navigationally relevant stimuli such asstreet scenes landscapes or buildings than whenthey viewed other kinds of visual stimuli such asfaces objects or scrambled scenes1 This responsewas found even when subjects simply watched thestimuli passively and were not required to performany navigational task We named this region theldquoparahippocampal place areardquo or PPA because itresponded strongly to depictions of places ThePPA appears to overlap with the regions activatedin previous neuroimaging studies of navigation(eg Aguirre et al 1996)
A critical determinant of PPA activationappears to be the presence in the stimulus of infor-mation about the shape or layout of the immediateenvironment The PPA responds no more stronglyto photographs of rooms filled with furniture andobjects than it does to photographs of the samerooms empty of all objects (ie just bare walls) Italso responds much more strongly to ldquoscenesrdquo madeout of Lego blocks (which have a geometric struc-ture similar to a room or street scene) than it does toobjects made out of the same Lego materials(Epstein Harris Stanley amp Kanwisher 1999)Thus the PPA is sensitive to the presence of a spe-cific kind of geometric organisation in the stimuluseven if the stimulus does not depict a real place inthe world In particular it responds more stronglyto depictions of surfaces that in some senseldquoencloserdquo the observer and define a space withinwhich one can act (ie the walls of a room or theside of the distant hill) than to depictions of surfacesthat define objects that the observer can act uponThese results are generally consistent with behav-ioural results from rats (Cheng 1986 Gallistel1990 Margules amp Gallistel 1988) and human
infants (Hermer amp Spelke 1994 1996 Hermer-Vazquez Spelke amp Katsnelson 1999) that suggestthat information about the geometry of surround-ing spacemdashthe ldquolay of the landrdquomdashplays a privilegedrole in orientation and navigation
What specific cognitive task does the PPA per-form Given the fact that landmark agnosia oftenresults from parahippocampal-lingual damage(Landis et al 1986) one might hypothesise thatthe primary role of the PPA is place recognitionidentification of familiar locations based on theirappearance andor geometric structure If so onemight expect the PPA to be sensitive to the noveltyfamiliarity of the place depicted in the stimulusHowever we found that the PPA responds just asstrongly to photographs of unfamiliar places (ieplaces the subjects had never visited) as it does tophotographs of familiar places (Epstein et al 1999Expt 1) In contrast the PPA is sensitive to thenoveltyfamiliarity of the stimulus itself respond-ing more strongly to new photographs than to pho-tographs viewed many times previously even whenall the photographs depict unfamiliar places(Epstein et al 1999 Expt 4) From these resultswe inferred that the PPA is more involved inencoding a representation of the spatial structure ofthe current scene than in relating that scene to onersquosstored cognitive map of the environment as awhole However our data did not allow us to deter-mine whether the PPA activation reflects percep-tual encoding memory encoding or both
The purpose of the present report was to moreprecisely determine the function of the PPA Tothis end we tested the ability of our two patients toperceive learn and recognise a variety of visualmaterials We predicted that the patients would bemore impaired at processing scene-like stimuli (iestimuli that depict spaces that one can navigatethrough or act within) than object-like stimuli Byusing several different kinds of tests we aimed todetermine whether any such selective impairment
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 483
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
1Although full scenes activate the PPA more strongly than do any other kind of visual stimulus we have also observed higher
activity in the PPA when subjects view buildings than when they view other kinds of objects (Epstein et al 1999) consistent withAguirre Zarahn and DrsquoEspositorsquos (1998) finding of a significant response to buildings in an adjoining and possibly overlapping regionof the anterior lingual gyrus
was in the perceptual recognition or memorydomain To anticipate we did find evidence for adeficit for visual materials that conveyed informa-tion about the shape of surrounding space Thisdeficit manifested itself during a memory encodingtest but not during perceptual tests or a recognitiontest Thus our data suggest that the PPA is neces-sary for the encoding of spatial layout informationinto memory but may not be necessary for manyaspects of the initial perceptual analysis recogni-tion or recall of this information We argue that thePPA may be a learning mechanism specifically ded-icated to encoding topographical materials intomemory
CASE DESCRIPTIONS ANDNEUROPSYCHOLOGICAL TESTING
In this section we present the case histories of thetwo patients in order to provide a general contextfor the more specific experimental tests reportedin the Experimental Investigations section Thepatients were chosen for investigation because clin-ical MRIs indicated that they had suffered damageto the parahippocampal region This section is
divided into four subsections First we present ageneral overview of the case histories Second wereport the results of some ad hoc tests that give ageneral indication of the patientrsquos navigational abil-ities Third we discuss other cognitive abilitiesincluding face recognition and episodic memoryFourth we report the results of a battery of formalneuropsychological tests performed on the patientsTable 1 summarises relevant observed and self-reported behaviour and Table 2 summarises theresults of the neuropsychological tests
Overview
Case 1 GRGR2 is a right-handed male with 18 years of educa-tion He was 60 years old at the time of testingTwo-and-a-half years prior to testing he suffered aright occipital-temporal stroke during cardiacsurgery A second stroke affecting the left occipital-temporal region occurred a week later Thesestrokes left him with a number of deficits includinglimitation of his peripheral vision (lefthemianopsia right upper quadrantanopsia)dyschromatopsia (details reported in Beauchampet al 2000) and topographical disorientation We
484 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
EPSTEIN ET AL
Table 1 Summary of observed and self-reported behaviour
GR CO
Real-world topographical Unable to recognise 714 newly learned MIT landmarks Unable to learn 3-room videogamelearning despite extensive training regime environment
Unable to find way back to lab unassisted despite Able to learn a simple route out ofextended experience with environment building after many visits
Map drawing Could not draw map of lab maps of other newly learned Map of lab was sketchy but not entirelyenvironments were inaccurate inaccurate
Drew accurate map of premorbid environment Drew accurate map of premorbid(former home) environment (currentformer home)
Scene perception Reports difficulty understanding the globality of complex Does not report a perceptual deficitscenes
Face recognition Some prosopagnosia Severe prosopagnosia
Episodic memory Largely intact though some difficulties were reported Moderate difficulties were reported
Can follow the plot of a movie or novel and report Reports some difficulty following theprevious dayrsquos activities plot of a book Accurately reported
contents of previous dayrsquos newspaper
2GRrsquos true initials are KG (cf Beauchamp Haxby Rosen amp DeYoe 2000)and COrsquos true initials are KC We use GR and CO
here to avoid confusion between the two patients
tested him over a period of 5 days in August 1998with 3 days of follow-up testing 3 weeks later and 1day of additional tests in May 1999 During thistime he was alert and cooperative Emotionalaffect and language use was normal He interactednormally in nonclinical social situations and hisconversation was appropriate and varied Heremembered the events of previous days andrecounted them accurately
In contrast to this high level of non-topographical functioning GR demonstrated adramatic inability to orient himself in space Forexample even after several days of testing he neverlearned the relative locations of the different roomsof the lab According to both GR and his wife thisinability to learn new topographical informationwas typical of his experience since his strokes as hefrequently gets lost in his daily life Soon after his
injury he moved to a neighbourhood with manysimilar-looking houses He reported that in orderto find his new house after a walk to a market six toseven blocks away he had to rely on street signs toguide him to the correct block and then examineeach house on the block in detail until he couldrecognise some feature that distinguished his homeuniquely Subsequently GR and his wife moved toa different house in a different country He reportedthat for the first 6 months after the move his newhome was like a ldquohaunted houserdquo for him insofar ashe was unable to learn his way around it For exam-ple he would repeatedly forget that the bedroomhad a balcony and he would be surprised by this factevery time he rediscovered it After several monthshowever he was able to learn the topography of thehouse sufficiently well to navigate through it (butsee below)
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 485
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 2 Summary of neuropsychological tests ( indicates percentile scores)
GR COmdashmdashmdashmdashmdashmdashmdashmdashmdashmdash mdashmdashmdashmdashmdashmdashmdashmdashmdashmdashmdashndash
IQ 122 (93 WAIS-R) 134 (99 WAIS-III)
WMS-III
Logical memory 25 immediate recall 27 immediate recall50 delayed recall 48 delayed recall(WMS-R)
Face memory 9 immediate recall 16 immediate recall16 delayed recall 9 delayed recall
Family pictures 2 immediate recall lt1 immediate recall5 delayed recall lt1 delayed recall
Rey Auditory Verbal 63 immediate recall 45 immediate recallLearning test 14 after interference list lt1 after interference list
16 delayed recall 3 delayed recall
Warrington Recognition 50 wordsMemory tests 29 scenes
lt5 faces
724 Spatial Recall 66 immediate recall2 delayed recall
Benton Face Recognition 42 lt1
Memory span Digits99 forward99 backward
Spatial37 forward50 backward
Boston Naming test Ceiling Ceiling
Case 2 COCO is a left-handed male with 20 years of educa-tion He was 60 years old at the time of testing Twoyears prior to testing he suffered a right posteriorcerebral artery stroke He first noticed on a week-end that his vision appeared blurred on the left andthat he had a tendency to bump into objects on thisside On Monday morning he was able to success-fully negotiate the route to his workplace a trip thatinvolved a bus ride followed by two subways andthen a shuttle to his office building However oncehe entered the building he found himself com-pletely disoriented unsure of its layout and wherehis office was located Interestingly the companyhe worked for had moved a few weeks earlierAlthough he had managed to arrive at this newbuilding (which was a block away from the previouslocation) he became lost upon entering it Hisneurologic exam revealed a left homonymoushemianopia but no evidence of neglect on a lettercancellation task or on sensory testing Investiga-tions revealed an ischaemic stroke with haemor-rhagic transformation in the right occipital andmesial temporal lobe On magnetic resonanceangiography he had a chronic left carotid occlusiona mild right carotid stenosis and bilateral vertebralartery stenoses Since then he has suffered fromsevere topographical disorientation primarily innew environments
CO reports that he can recognise buildings thatwere familiar to him before the stroke He can alsoform mental images of well-known landmarks suchas the Empire State Building and the BostonPublic Gardens and of the house he lived in as achild When asked to imagine standing in front ofthe Massachusetts State House (a location wellknown to him from years of living in Boston) heaccurately described the surrounding buildings andtheir locations However he reports that when helooks at a place that hersquos encountered post-strokeall he gets is a feeling of familiarity that does notallow him to determine where the place is or whenhe saw it He also reports an inability to understandhow various landmarks relate to each other inlarge-scale space Despite this difficulty he canfollow a map and frequently rides the Bostonsubway alone
Unlike GR CO was able to learn a certainamount of topographical information For exam-ple he soon learned the location of the rooms in thelab and would travel between them unassistedAfter his first visit to the lab he needed to be guidedall the way back to the subway station but after sev-eral visits he could find his way out of the buildingand to the subway on his own as long as he was firstguided to the elevators (It should be noted how-ever that the building is on the same block as thesubway station Furthermore he still needed to bedirected to the elevators even though our buildinghas a relatively simple floor plan) Thus the amountof topographical information he can learn is limitedbut not zero
COrsquos language and reasoning skills appear to benormal and his conversation is appropriate andvaried In addition to his topographical difficultieshe also has limitations of his peripheral vision onthe left side
Navigational abilities
In addition to the experiments reported in theExperimental Investigations section of this paperwe also performed a number of ad hoc tests of GRand COrsquos navigational abilities Although thesetests were informal we describe them here to pro-vide a general sketch of the nature of GR and COrsquosimpairments
Real-world topographical learningOver the course of several days we took GR and hiswife on three walks around the MIT campus whichneither had visited previously Each walk followedexactly the same path At several places in the walkwe would stop and point out different buildings andlandmarks tell them something about each one andask them to remember their appearance for latertesting Immediately after the completion of thethird walk both GR and his wife separately per-formed a forced-choice recognition test in whichthey saw photographs of 14 landmarks (mostlybuildings) from the walk and 14 novel landmarks(from another part of the MIT campus that theyhad not visited) and were required to reportwhether they had seen each one or not GRrsquos
EPSTEIN ET AL
486 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
performance was 714 correct hits for the familiarlandmarks and 1314 correct rejections for theunfamiliar landmarks In contrast his wifeobtained a perfect score on this test Given theextensive nature of the training regime GRrsquos scorestrongly suggests an impairment in learning theappearance of new real-world places Interestinglyseveral of the seven familiar landmarks that he gotright had an obvious distinctive feature that hadbeen explicitly pointed out to him For exampleone building had a distinctive clock tower andanother one was an unusual triangular shape ThusGR was occasionally able to use distinct visualfeatures to identify places even though he wasunable to recognise them holisticallymdasha commonpattern among topographical disorientationpatients
GR did not appear to have any obvious problemsin understanding the spatial structure of his imme-diate environment When given a floor plan of thebuilding at MIT in which he was being tested hewas able to successfully follow a route marked onthe plan However when the plan was taken awayhe had great difficulty finding his way back to thelab even though he had more than 30 min experi-ence with the environment and his starting positionwas less than 30 feet from the goal (Figure 2) Infact the route he took resembled a random walk Atone point he walked right past his destinationwithout recognising it and only knew for sure thathe was in the right place when he identified his wifein the room Thus his on-line appreciation of thelayout of immediately visible space appears to beintact but he cannot find his own way withoutassistance
We were unable to perform the MIT landmarkrecognition test on CO because he suffered from amedical problem that made it difficult for him towalk around campus In lieu of this we tried to seewhether he could learn a simple video game envi-ronment consisting of three connected roomsAfter about 30 min experience navigating throughthis environment (via button presses) he was askedto sketch a map of it He declined to do so statingthat it was beyond his abilities When asked howmany rooms there were he replied that he did notknow but that there were at least 10 Given the
simplicity of this artificial environment COrsquos com-plete inability to learn it is striking
Map drawingAfter several days of testing we asked both GR andhis wife to sit at different tables and independentlysketch (1) the shape of Florida (2) a bicycle (3) afloor plan of their hotel room in Boston (4) a floorplan of the first floor of the house they had been liv-ing in for the last 8 months (all post-stroke) (5) afloor plan of the main room of the lab and (6) afloor plan of the house they lived in 10 years ago(pre-stroke) GR drew the map of Florida and thebicycle perfectly His plan of the hotel room wasnotably different from the one drawn by his wifemdashfor example he reported that there was only onebed when in fact there were two and he placed thebed on the wrong side of the room (Figure 1a) Hisplan of the first floor of his current house was some-what better but was not very detailed and includedsome major errors For example he wronglyreported that there was no bathroom on this floor(Figure 1b) He was unable to produce any floorplan of the main room of the lab leaving this pageblank When his wife reminded him that this wasthe room in which he had just spent an hour readingthe newspaper and in which we had all had break-fast in that morning he replied that he didnrsquot knowwhat room she was talking about (although heappeared to remember these events) In contrast tothis near total inability to produce accurate maps ofnew environments GR produced an accurate mapof the house he lived in prior to his injury (Figure1c) He reported that his memory of this house wasldquoall thererdquo and he could imagine walking around init When asked to produce a perspective of a roomwhen he was in it he did so with ease and he wasalso able to create an accurate floor plan of theroom
In contrast CO found it very difficult to pro-duce a floor plan of the lab both when he was in itand when he was taken to another room and askedto recall it However although the maps he pro-duced were quite impoverished they were notentirely inaccurate demonstrating some ability toencode topographical information This contrastswith GRrsquos absolute inability to recall anything
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 487
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
EPSTEIN ET AL
488 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure
1F
loorplansdraw
nby
GR
andhisw
ifeof(a)
theirhotelroomin
Boston(b)theircurrenthousew
hichthey
hadm
ovedto
subsequenttoG
Rrsquosinjury(c)the
housethey
livedin
priorto
GR
rsquosinjury
Ho
telro
om
inB
osto
n
GR
His
wife
His
wife
Cu
rren
th
om
e
GR
His
wife
Fo
rme
rh
om
e
GR
about the main room of the lab once he had left itCO was also able to draw a detailed floor plan of theapartment he was living in at the time (and hadlived in for 20 years) Several months after he hadmoved from this apartment we asked him to drawsuch a floor plan again and he produced one thatwas substantially the same as the first (Figure 3)Thus their map drawing performance indicatesthat GR and CO are primarily impaired at sketch-ing places experienced after their strokes but areunimpaired at sketching places experienced beforetheir strokes This pattern is indicative of deficits inencoding but not recall
Scene perceptionAs noted earlier GR can navigate with the aid of afloor plan suggesting that his perceptual abilitiesare largely intact Interestingly however he reportsthat he does have a perceptual deficit which hedescribes as an inability to understand scenes whenthey are too complex For example he reports thathe often has trouble using maps because theyusually are too busy with details It is unclearwhether this apparent perceptual problem is a resultof his field cut some other separate perceptual
problem or a memory problem (see discussion)CO did not complain of a similar deficit GR wasan accomplished artist prior to his injury producingpaintings of striking complexity Tragically hereports that he cannot now appreciate his ownpaintings
Other cognitive abilities
Face recognitionGR also reported some difficulties with face recog-nition On his second visit we examined his abilityto learn new faces by having him perform a forced-choice recognition test on seven familiar and sevenunfamiliar faces The familiar faces were personnelfrom the lab with whom he had at that point had agood deal of experience spanning at least 3 daysAll the people in the photographs had a towelwrapped around their head in order to minimise theuse of hair as a nonfacial cue Despite the difficultyof this task he correctly identified 67 familiarfaces and made no false alarms Surprisingly theone face that he did miss was the first author of thispaper with whom he at that point had had consid-erable experience GR also performed normally ona difficult famous face recognition test (see Experi-ment 4 following) and on the Benton Face Recog-nition test (see following) Thus he has someability to recognise faces Consistent with this GRreports that he can remember a small number offaces but that he quickly ldquoloses trackrdquo if he has toremember too many
CO also appears to suffer from someprosopagnosia as evidenced by his low perfor-mance on the Benton Face Recognition test (seefollowing see also Experiment 4) Interestingly hedoes not complain of face recognition difficultiesbut rather attributes his inability to recognise peo-ple to a lack of interest in their appearance
Episodic memoryAt first glance GRrsquos episodic memory appeared tobe largely intact As mentioned he could recall theevents of the previous day accurately Furthermorehe reports that he can follow the plot of a moviewithout any difficulty However both he and hiswife report that he does have some problems with
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 489
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Figure 2 Floor plan of the building at MIT in which GR wastested Corridors are highlighted in grey GR was asked to find themain room of the lab (marked G on the map) Starting location(marked S) was a point approximately 30 feet down the hall GRrsquosroute is marked by arrows GR found this task very difficult andonly knew for certain that he had reached his goal when he caughtsight of his wife through the glass door of the lab In contrast hewas able to follow a route marked on a plan of the same floorwithout any problem or hesitation
episodic memory For example he stated that heremembered very little about a trip to Italy they hadtaken several months previously His wife reportsthat he has great difficulty keeping track of timeFor example before their trip to Boston he repeat-edly asked her what day they were supposed to leaveand seemed unable to encode this information Weobserved a similar phenomenon during testingDuring the tests that required repetition of thesame procedure several times he sometimesseemed unable to clearly remember how manytimes he had repeated the procedure Thus he doesappear to have a subtle episodic memory deficit
which may manifest itself as an inability to sepa-rately remember successive events that are not easilydistinguishable from each other
CO also reports some problems with his epi-sodic memory He relies on lists to rememberappointments stating ldquoif I didnrsquot have a list Irsquodprobably forget why I walked in the door hererdquoDespite this claim he has never missed an appoint-ment nor is he ever confused about the purpose ofthe testing appointments He can accurately reportthe day of the week He remembers the experi-menters clearly from appointment to appointmentas well as the details of the various experiments
EPSTEIN ET AL
490 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 3 Two floor plans drawn by CO of the apartment he lived in for over 20 years The first plan was drawn when CO still lived inthe apartment the second was drawn several months later when he had moved out Note that the two plans correspond very well COreported that it was quite easy to produce these floor plans
CO home Feb 99
CO (former) home June 99
when they are repeated across appointments Whenqueried about his episodic memory difficulties COreported that he will usually remember an event ifhe is reminded of it but that he has difficulty spon-taneously regenerating events Interestingly hereports that even when he is reminded of an eventand remembers it he cannot remember ldquowhere itwas or when it wasrdquo When he reads he finds it dif-ficult to follow a story if it skips around Howeverwhen queried he was able to accurately report whathe had read in the newspaper the previous day
Formal neuropsychological tests
Formal neuropsychological testing was performedon GR in September 1998and on CO in December1998 Test performances were compared to age-matched peers3
GR exhibited mild to moderate impairment inmemory relative to what would be predicted for hisintellect (WAIS-R Full Scale IQ =122 93rd per-centile superior range) and level of educationThese memory problems were more apparent onvisual than on verbal memory tests On a test ofmemory for conceptually organised text passages(Wechsler Memory Scale Logical Memory[WMS-III] Wechsler 1997b) his performancewas average for both immediate (LM I =25th per-centile standard score SS = 90) and delayed recall(LM II =50th percentile SS =100) In contrast hisscore on the WMS-III Face Memory subtest was inthe low average range for both immediate (9th per-centile SS =80) and delayed recall (16th percentileSS =85) On a test of memory for scenes (WMS-IIIFamily Pictures subtest) immediate recall was defi-cient (2nd percentile SS = 69) and delayed recallwas borderline impaired (5th percentile SS =76)GR also had difficulty learning word lists on theRey Auditory Verbal Learning Test (Spreen ampStrauss 1998) his immediate recall was average(63rd percentile SS = 105) but after the interfer-ence list his performance fell to the low averagerange for both immediate (14th percentile SS =84)
and delayed recall (16th percentile SS =85) On arecognition test of the items of the list he success-fully recognised all 15 but made 8 false alarms Per-formance on the 724 Spatial Recall Test (RaoHammeke McQuillen Khatri amp Lloyd 1984)which required him to remember seven locations ona 6times4 checkerboard was 66th percentile (SS =106)for immediate and 2nd percentile (SS = 69) fordelayed recall Performance on the Benton FaceRecognition test was normal (42nd percentile)Performance on the Boston Naming Test was atceiling indicating that language abilities wereintact Thus GR exhibits some impairment ontasks that require delayed recall of visual informa-tion (WMS-III Face Memory subtest) or verbalinformation that is not organised in terms of a storyor plot (Rey AVLT) and is clearly impaired ontasks that required delayed recall of spatial informa-tion (724 Spatial Recall Test) or stimulus location(WMS-III Family Pictures subtest) In contrasthis ability to recall organised verbal information(WMS-III Logical Memory subtest) appears to beintact
As with GR COrsquos memory performance wasgenerally below what would be expected based onintellect (WAIS-III Full Scale IQ =134 99th per-centile very superior range) and education levelAlso like GR there was some evidence that visualmemory problems were more pronounced than ver-bal memory problems Memory for text passages(WMS-R Logical Memory) was average for bothimmediate (27th percentile SS = 91) and delayedrecall (48th percentileSS =99) In contrast perfor-mance on the WMS-III Faces subtest was lowaverage for both immediate (16th percentile SS =85) and delayed recall (9th percentile SS =80) andperformance on the Family Pictures subtest wasdeficient (lt 1st percentile SSlt 62) for both imme-diate and delayed recall On the Warrington Rec-ognition Memory Tests (RMT) he scored in theaverage range for words (50th percentile SS =100)and scenes (Warrington Topographical Recogni-tion Memory Test 29th percentile SS =92) and in
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 491
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
3The formal neuropsychological testing reported in this section was performed prior to the beginning of our combined
investigation of GR and CO (ie prior to the main experimental investigations of this paper) Consequently GR and CO were nottested on exactly the same battery of tests
the deficient range (lt 5th percentile SS lt 75) forfaces The low score for faces on this test as well asthe WMS-III may reflect prosopagnosia ratherthan a memory problem as he scored in the defi-cient range (lt1st percentile SSlt62) on the BentonFace Recognition test Verbal list learning (ReyAVLT) was average on immediate recall (45th per-centile SS =98) but after hearing an interferencelist performance fell to the deficient level (015words recalled) for both immediate and delayedrecall On a recognition test for these words herecognised 1115 and made one false alarm (39thpercentile SS =96) There was a large discrepancyon tests of memory span which may indicate adecline in spatial memory digit span was superior(9+ forward and 7 backwards both 99th percentile)but spatial span was only average (5 forward and 5backward 37th and 50th percentiles respectively)Language abilities were intact (Boston NamingTest performance was at ceiling) Thus like GRCO shows some impairment at recall of visualinformation (WMS-III Face Memory subtest) andverbal information that is not organised into a nar-rative (Rey AVLT) and clear impairment at tasksthat require recall of stimulus location (Family Pic-tures subtests of WMS-III) In contrast his abilityto recall organised verbal information (WMS-RLogical Memory subtest) is more preserved
MRI RESULTS
In order to determine the extent of their lesionsstructural MRI images were taken of both subjectsDelineation of the lesions was based either onspoiled GRASS coronal images at 11mm thick-ness (GR) or anatomical T-1 weighted MPRAGEMRI sequences yielding 125 mm thick coronalimages (CO) The anterior-posterior extent of thelesions was verified on axial images
GR has bilateral occipital-temporal lesions(Figure 4) In the left hemisphere the posteriorparahippocampal gyrus (including para-hippocampal cortex) is severely damaged with onlyminimal injury to the most caudal aspect of the hip-pocampus There is also moderate damage to themedial fusiform and inferior lingual gyri In the
right hemisphere the hippocampus is spared butthere is damage to the posterior end of theparahippocampal gyrus near the parahippocampal-lingual boundary More posteriorly the lesionedarea extends along the collateral sulcus to includesignificant portions of the inferior lingual gyrus andthe medial fusiform gyrus as well as most of themedial occipital lobe below the calcarine fissure
CO has a right-hemisphere occipital-temporallesion Right posterior hippocampus and para-hippocampal gyrus are severely affected Moreposteriorly the damage covers large portions of theinferior lingual and medial fusiform gyri as well assubstantial portions of the medial occipital lobeincluding most of right calcarine cortex In addi-tion diffuse foci of increased signal on T-2weighted images was present in periventricularwhite matter in the corona radiata and centrumsemiovale with a lacunar infarct of the rightputamen There was also moderate degree of dif-fuse involutional changes with increased ventricularsize particularly notable in the right temporal hornwith ex vacuo dilitation
The structural MRIs suggested that the PPAwas severely if not completely damaged in bothhemispheres for GR and in the right hemispherefor CO In order to confirm this for GR we used ahigh-field 3-T magnet to perform a functional scanin which he viewed scenes faces and objects in thesame blocked fMRI design that we have used tolocalise the PPA in all our previous experiments(see Epstein et al 1999 for details Epstein ampKanwisher 1998) A Kolomogorov-Smirnov testfound no voxels in the parahippocampal region thatfit our previously established criterion for inclusionin the PPA (ie greater response to scenes than tofaces and objects at a uncorrected significance levelof plt 0001) This result contrasts sharply with thepattern seen in over 95 of (mostly young or mid-dle-aged) normal subjects in whom a large acti-vated region can be identified in both left and rightparahippocampal cortex using this comparison (seeFigure 5) A similar test was used to identify voxelsthat responded significantly more to faces than toobjects This comparison revealed a region of face-selective voxels in the right fusiform gyrus consis-tent with the location of the Fusiform Face Area
EPSTEIN ET AL
492 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 493
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
GR
CO
Figure 4 Structural MRIs of GR and CO showing the extent of the damaged region (marked as white) Images have been normalisedusing SPM99 and a cost function masking algorithm (provided by Matthew Brett) into standard stereotactic space (Talairach amp Tournoux1988) using the Montreal Neurological Institute template (Evans et al 1993) Numbers indicate y-Talairach coordinate for each coronalslice corresponding slice planes are also indicated on the sagittal image
(FFA) commonly seen in normal subjects(Kanwisher McDermott amp Chun 1997) Thusthe MRI was sensitive enough to detect functionaldifferences an FFA was found but no PPA Whencombined with the anatomical results this stronglysuggests that GR has no PPA
Because of safety concerns (presence of a femo-ral artery aneurysm clip) we did not perform a simi-lar high-field functional MRI of CO However asFigure 4 demonstrates CO clearly has damage tothe parahippocampal-lingual region in the righthemisphere
EXPERIMENTAL INVESTIGATIONS
In this section we report the results of four experi-ments designed to probe the consequences of dam-age to the lingual-parahippocampal region ingeneral and the PPA in particular Our primaryconcern in these experiments was to test GR andCOrsquos ability to perceive remember and recognisedifferent kinds of visual material Based on our pre-vious fMRI work we predicted that GR and CO
would have more difficulty with scene-like stimuli(ie stimuli that convey information about thegeometry of a space one can navigate through or actwithin) than with object-like stimuli Such a deficitcould account for at least some of the problems GRand CO encounter in real-world navigation
Beyond determining the kind of visual materialthat gets processed in parahippocampal cortex wealso wanted to determine the nature of the process-ing that takes place on that material In particularwe wanted to determine if parahippocampal cortexis involved in perceptual encoding memory encodingor recognition As discussed earlier (Introduction)our fMRI results provided some evidence againstplace recognition as the sole function of the PPAbut did not allow us to distinguish between percep-tual and mnemonic encoding Experiments 1 and 2examine the role of parahippocampal cortex in per-ceptual encoding Experiment 3 examines its role inmnemonic encoding and Experiment 4 examinesits role in place recognition
Experiment 1 Two-dot task
The first hypothesis we examined was thatparahippocampal cortex plays a role in perceivingthe shape of the local environment In our previousfMRI work we observed that the PPA respondsmuch more strongly during scene viewing than dur-ing object or face viewing even when subjects donothing more than watch the scenes passively with-out performing any other task This suggested thatscene perception alone may be sufficient to activatethe PPA Previous reports have indicated that dam-age to parahippocampal cortex and other medialtemporal regions leads to problems with memoryrather than perception (Bohbot et al 1998Maguire Burke Philips amp Staunton 1996 Pigottamp Milner 1993) However all of these previousresults were obtained on patients who had unilat-eral lesions GR has a bilateral occipital-temporallesion including clear damage to parahippocampalcortex in both hemispheres furthermore he reportssome kind of perceptual deficit We hypothesisedthat this deficit might involve difficulty perceivingor interpreting the geometry of scenes and otherstimuli with a similar geometric structure
EPSTEIN ET AL
494 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 5 Top row shows a single slice from a functional MRIscan performed on GR Bottom row shows results from a normalsubject (NK) for comparison Voxels that responded more to facesthan objects (left column) or more to scenes than faces and objects(right column) in a Kolomogorov-Smirnov test are highlighted bywhite arrows GR has a functioning face area (FFA) but there isno evidence of a place area (PPA)
FFA PPA
GR
NK
In this experiment subjects viewed either objectshapes or spatial layouts Stimuli consisted ofblack-and-white photographs of real-world stimuli(empty rooms vs common objects) or abstractstimuli made out of Lego blocks (Lego ldquoscenesrdquo vsLego ldquoobjectsrdquo) These stimuli have been previouslyshown to differentially activate the PPA in normalsubjects (Epstein et al 1999 Epstein ampKanwisher 1998) Two red dots were superim-posed on each photograph (Figure 6) The task wasto report which of the two red dots was over thecloser part of the object or scene This task wasdesigned to have no memory component (apartfrom remembering the instructions) but to requireaccurate 3D perception of objects and scenes
If GR or CO have difficulty perceiving or inter-preting the spatial layout of scenes then we pre-dicted they would do worse when the stimuli areempty rooms or Lego scenes than when they arereal-world or Lego objects Note that the criticalcomparison is between the Lego scenes and theLego objects as these stimuli are made of the samematerials and differ primarily in their surfacegeometry
MethodTesting for all experiments was done on aMacintosh computer The experiment consisted of80 trials (20 of each stimulus type randomly inter-mixed) Subjects began each trial by pressing thespace bar A black-and-white photograph from oneof the four stimulus categories (see earlier) thenappeared on the screen with two red dots on it Thetask was to determine which of the two red dotsindicated the closer part of the scene or objectSubjects pressed the ldquofrdquo key if the leftmost red dotwas closest and the ldquojrdquo key if the rightmost dotwas closest They were instructed to respond asquickly as possible without making errors Stimuliremained on the screen until a response was made
The stimulus set was constructed so that eachempty room was matched to a common object thathad red dots in the exact same location and eachLego layout was similarly matched to a Lego objectThus the locations of the red dots were counterbal-anced across scene and object stimuli controllingfor any effects of visual field cuts
Results and discussionResults for GR and CO and four elderly normalsubjects are shown in Table 3 Both GR and COcould do this task without difficulty and their per-formance was near ceiling There was no evidencethat they had any more trouble with the emptyrooms or Lego scenes than with the real or Legoobjects Reaction times were slightly longer for GRand CO than for normal subjects as one wouldexpect given their visual field deficits however this
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 495
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 3 Experiment 1 (two-dot) results (SD in parenthesis)
GR CO Normals
Proportion correctObject 095 080 093Lego object 100 095 098Empty room 100 100 099Lego room 095 100 096
Mean RTObject 2435 2932 2161 (980)Lego object 2052 3245 2029 (921)Empty room 2744 2291 1788 (959)Lego room 1904 2281 1630 (1127)
Figure 6 The four kinds of stimuli used in Experiment 1 Topleft lego layouts top right real-world layouts (empty rooms)bottom left lego objects bottom right real-world objects
difference did not appear to be significant Impor-tantly there was little evidence that GR or CO tooklonger to make their perceptual judgements for thereal or Lego scenes than for the real or Lego objectsThus contrary to our initial hypothesis this experi-ment found no clear evidence for selective impair-ment in the perception of scene-like stimuli
Experiment 2 Different-views matchingtask
The previous experiment demonstrated that GRand CO can extract information about the per-ceived distances between themselves and the sur-faces of an object or scene Thus in at least onesense their perception of these surfaces is pre-served However this does not mean that their abil-ity to represent the layout of a scene is unimpairedThe two-dot task requires on-line comparison oftwo surfaces in egocentric coordinates GR and COwould be able to perform this task even if they couldnot (1) represent more than two surfaces at a time(2) represent surfaces in other than strictly egocen-tric coordinates or (3) sustain surface representa-tions for longer than the time it takes to make acomparison It has been suggested that the parietallobes are involved in on-line processing of surfacegeometry in egocentric coordinates (Milner ampGoodale 1995) thus it is possible that the parietallobes (which are intact in GR and CO) may be suf-ficient to perform the two-dot task even in theabsence of more durable or complex surface repre-sentations in parahippocampal cortex
In Experiment 2 we examine GR and COrsquosability to create and sustain a representation of thegeometry of an object or scene-like layout The taskwas to view a photograph of a Lego object or Legoscene hold it in memory for 10 s and then deter-mine whether a second object or scene was the sameas the first In ldquosamerdquo trials the second photographdepicted the same scene or object depicted from adifferent viewing angle In ldquodifferentrdquo trials the sec-ond photograph depicted a different scene or objectchosen to be similar to the first one (Figure 7)Thus in order to successfully perform this tasksubjects had to form a representation of the surfacesof the object or scene sustain this representation in
memory for 10 s and then align it (presumablyusing mental rotation) to the second object orscene
MethodThe experiment consisted of two parts each with40 trials In the first part the stimuli were black andwhite photographs of objects made out of Legoblocks In the second part the stimuli were photo-graphs of ldquoscenesrdquo made out of Lego blocks whichwere designed to have a geometric structure similarto real-world places such as a roomor a city street
Subjects began each trial by pressing the spacebar A photograph of a Lego object or sceneappeared on the screen for 3 s followed by a 10-sblank interval A second photograph then appeareddepicting either (1) the same object or scene as thefirst photograph shown from a different viewingangle (offset approximately 35 degrees) or (2) a dif-ferent object or scene Subjects were instructed topress the ldquojrdquo key if the two photographs depicted thesame object or scene and the ldquof rdquo key if they did notThey were told to take as much time as they neededto make a decision but no extra time The secondphotograph remained on the screen until a responsewas made Half the trials were ldquosamerdquo trials and halfwere ldquodifferentrdquo trials
Results and discussionFigure 8 shows results for GR and CO as well as forsix elderly normal controls GRrsquos and COrsquos perfor-mance was comparable to that of the normal con-trols Furthermore GR and CO performed just aswell when the stimuli were Lego scenes as whenthey were Lego objects These results are unlikely toreflect a ceiling effect as performance for bothpatients and controls was around 80 correctThus both patients appear to be able to form a rep-resentation of the geometry of both scenes andobjects which they can sustain in short-termmemory
Experiment 3 N-back recognition memorytask
The previous experiment failed to find any impair-ment in GR and COrsquos ability to form a representa-
EPSTEIN ET AL
496 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
tion of the geometry of a scene or object andmaintain it for 10 s in the absence of the originalstimulus Not only does this result suggest that per-ception of scene-like stimuli is intact in thesepatients it further suggests that some short-termmemory functions are intact as well However we
should note that subjects in this experiment werefree to devote all their effort and attention in the 10-second blank interval to maintaining a memory ofthe stimulus Thus it is possible that GR and COcan use rehearsal mechanisms to maintain the stim-ulus in short-term memory during the blank inter-
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 497
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
(b) Lego object version
Figure 7 Stimuli and procedure for Experiment 2 (a) In the Lego layout version subjects see a Lego layout for 3 s followed by a blankscreen for 10 s A second layout then appears which is either the same layout as the first one but seen from a different angle or a differentlayout (b) The Lego object version is the same but with object stimuli
3 s 10 s OR
3 s 10 s OR
(a) Lego layout version
(b) Lego object version
val but are unable to form a more durable memorytrace for the stimulus This possibility was tested inthe current experiment which was designed toexamine GR and COrsquos ability to learn new visualinformation
We devised a continuous n-back recognitionmemory test in which subjects viewed a sequence oforiginally novel stimuli some of which wererepeated within the sequence at intervals of 1 3 or5 intervening images For each stimulus subjects
reported whether they had seen it before in theexperiment or not In different runs in the experi-ment the stimuli were black-and-white photo-graphs of (1) Lego scenes (2) Lego objects (3) real-world landscapes (4) novel objects made out ofSculpey clay (5) unfamiliar houses (6) unfamiliarfaces as well as (7) line drawings of nonsenseobjects and (8) pronounceable pseudowords (Fig-ure 9) Thus we were able to test GR and COrsquos rec-ognition memory for different kinds of materials atdifferent time intervals in a paradigm in which thepresence of intervening items between the twoappearances of each stimulus made rehearsal diffi-cult Note that because none of the specific stimuliused in this experiment were familiar in advancethis task did not require an episodic memory com-ponent and could be done on the basis of pure visualfamiliarity with the particular visual stimulus
Although many different stimulus classes weretested in this experiment the critical comparison isbetween the Lego layouts and Lego objectsbecause these are best matched with regard tomaterials and differ primarily in their geometricstructure (see Introduction) We predicted that ifGR and CO showed a memory deficit for this taskit would be greater for the Lego layouts than for the
EPSTEIN ET AL
498 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 8 Results of Experiment 2 Performance over ldquosamerdquotrials and ldquodifferentrdquo trials is averaged together Error bars show1 SD
Figure 9 Stimuli for Experiment 3 Top row Lego layouts real-world scenes houses Bottom row Lego objects clay objects nonsenseobjects faces In an eighth condition (not shown) subjects viewed pronounceable nonwords such as ldquowilchrdquo or ldquothipperrdquo
Lego objects In addition a deficit for materialsthat convey information about the layout of sur-rounding space might also be apparent when GRand CO are tested on more realistic stimuli If soone would expect them to do poorly on the land-scapes but relatively well on the clay objects linedrawings of nonsense objects faces andpseudowords
MethodFor each subject a complete experimental sessionconsisted of being run once on each of the eightstimulus classes Each run consisted of 90 trials ineach trial a stimulus was presented and subjectsreported whether or not they had seen it before inthe run (or indeed ever) by pressing the ldquojrdquo key ifthey had or the ldquof rdquo key if they had not Each stimu-lus was on the screen for either 1 2 or 3 s (see fol-lowing) and appeared promptly after the keypressreport for the previous trial Individual stimuliappeared either once (14 stimuli) twice (30 stim-uli) or four times (4 stimuli) within a given run Ofthe stimuli that were presented twice 10 wererepeated immediately (1-back) 10 were repeatedwith a lag of two intervening items (3-back) and 10were repeated with a lag of four intervening items(5-back) In order to ensure that between-subjectdifferences in performance were not due to stimulusdifferences the same stimuli were assigned to thesame conditions each time the experimentwas run
GR performed the experiment twice once inAugust 1998 and again 9 months later Because ofa computer error he was only tested on six of theeight categories (all except Lego layouts and Legoobjects) at the later date CO was also tested twiceonce in February 1999 and again 6 months laterReported results for GR and CO are the average oftheir two testing sessions Normal subjects wereonly tested once Stimulus presentation time was 1 sin GRrsquos first testing session In pilot testing COfound this too fast for the critical categories of Lego
layouts and Lego objects so presentation time wasextended to 2 s for these categories in his first test-ing session All normal subjects were tested at thesepresentation rates (2 s for Lego layouts and objects1 s for other stimuli) In COrsquos second testingsession presentation time was lengthened to 3 s forall stimulus categories to ensure that any deficitsfound were memory deficits rather than problemswith the nonmemory aspects of the task
Results and discussionResults from GR CO and 10 elderly normal con-trols are shown in Figure 10 and Table 4 Figure 10ashows results from the main comparison betweenLego layouts and Lego objects Both normal sub-jects and patients performed near ceiling for bothLego layouts and Lego objects at 1-backinterstimulus intervals verifying that GR and COcould successfully perceive the pictures as they werebeing presented In contrast there was a strikingdifference between the two patients and the normalsubjects at 5-back intervals Whereas normals didon average somewhat better on Lego layouts thanLego objects both GR and CO showed the oppo-site pattern doing much worse on Lego layoutsthan Lego objects The 3-back condition showed amixed pattern Whereas GR did better on Legoobjects than Lego layouts in this condition (similarto his performance at 5-back intervals) CO didbetter on Lego layouts than Lego objects COrsquos lowperformance on Lego objects in the 3-back condi-tion appears to be an anomaly as it is much lowerthan his performance for comparable stimuli in the1-back and 5-back conditions
Figure 10b shows the difference in performancebetween Lego layouts and Lego objects for thecritical 1-back and 5-back intervals At 5-backintervals CO was significantly impaired comparedto the normal subjects (z = ndash250) whereasGRrsquos impairment fell just short of significance (z =ndash175)4
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 499
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
4In fact these values may underestimate GR and COrsquos relative impairment for Lego layouts In the 5-back condition 4 of the 10
normal subjects had perfect performance on the Lego layouts but all scored below ceiling on the Lego objects Thus the true advantagefor Lego layouts over Lego objects for normal subjects is probably greater than that observed here in which case GR and COrsquos lowerperformance for Lego layouts compared to Lego objects would be even more abnormal Furthermore post-hoc analyses revealed thatone of the normal subjects showed a difference in performance between Lego layouts and objects that was over 2 SD below the mean ofthe other nine subjects When this outlying subject was excluded z-scores were ndash322 for CO and ndash232 for GR
Table 4 shows results for all eight stimulusclasses Although direct comparisons between sub-jects should be treated with caution since viewingtime for different items was not the same for eachsubject we do note a few interesting patterns Firstof all there is little or no evidence that GR or CO
show a memory deficit for nonsense words clayobjects or line drawings of nonsense objects GRappears to be slightly impaired on faces and housesbut CO performs normally on these categoriesStrikingly GR shows a severe deficit for the reallandscapes performing more than 40 percentagepoints lower than normals Thus even thoughthere is some evidence that GR might have someimpairment for nonscene stimuli he clearly is muchmore impaired for scene-like stimuli such as theLego scenes and landscapes CO does not show asimilar deficit for the landscapes However itshould be kept in mind that in one of his two testingsessions CO viewed each picture for 3 s which wasmuch longer than the 1 second used for all non-Lego stimuli for GR and normals Thus COrsquosapparently ldquonormalrdquo performance in these condi-tions might be overestimated
Taken together the results of this experimentindicate that GR and CO are selectively impaired atprocessing scene-like stimuli This deficit is consis-tently apparent at 5-back but not 1-back intervalsThis finding suggests that the deficit should becharacterised as a memory encoding problemHowever the present results do not allow us todetermine whether the difficulty occurs at memoryencoding or at subsequent recognition (see Epsteinamp Kanwisher 2001) If GR and CO have a generalproblem with recognition of topographical materi-als then they should not be able to recognise scenes
EPSTEIN ET AL
500 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 10a Results of Experiment 3 for GR (top) CO (middle)and for 10 age-matched normal controls Performance measure isproportion of repeated stimuli correctly identified as such correctedfor guessing using the standard formula
Figure 10b Difference between Lego layout performance andLego object performance for the 1-back and 5-back conditionsError bars indicate one 1 SD
and landmarks that were learned prior to theirinjury We tested this possibility in the nextexperiment
Experiment 4 Famous landmarksfacesrecognition task
In this experiment we examined GR and COrsquosability to recognise the visual appearances of placeslearned prior to injury Subjects saw a series offamous landmarks intermixed with nonfamouscontrols (Imaging experiments have demonstratedthat landmarks of this type can activate the PPAalmost as strongly as complete scenes Epstein etal 1999 Epstein amp Kanwisher 1998) They wererequired to say whether they knew each specificlandmark and to identify the ones they knew Sub-jects were instructed to respond affirmatively if theywere fairly certain that they recognised the land-mark but not if it only looked generally familiarFor purposes of comparison subjects also per-formed an identical test in which the stimuli werefamous and nonfamous faces In both testsdistractor items were designed to be visually andconceptually similar to the target items (see Figure11) If parahippocampal cortex is required for placerecognition then one might expect GR and CO toperform poorly compared to normals on the land-mark recognition test
MethodEach of the two experiments (famous faces famouslandmarks) consisted of 48 trials Subjects beganeach trial by pressing the space bar after which ablack-and-white photograph of a face (in thefamous faces experiment) or a landmark (in thefamous landmarks experiment) appeared on thescreen Subjects were told to press the ldquojrdquo key if theyrecognised the specific face or landmark and the ldquof rdquokey if they did not taking as much time at theyneeded to make this discrimination but no extratime Subjects were told to report that they recog-nised a face or landmark if they felt that they knewit even if they could not retrieve its name If theydid know the name they were to say it after pressingthe button Otherwise they were to say whateverthey did remember about the item
Half of the photographs were of famous and halfof nonfamous people and landmarks The famousfaces included public figures (eg George BushRichard Nixon Prince Charles) and movie stars(eg Harrison Ford Clark Gable Marlon Brando)who were prominent prior to the time of GR andCOrsquos injuries the famous landmarks were build-ings and monuments from all over the world (egEiffel Tower US Capitol Building Great Sphinx)The nonfamous stimuli were chosen to match thefamous stimuli as closely as possible Each famousstimulus had a ldquomatchrdquo For example George Bushwas matched to a man of similar age in a coat andtie and the Taj Mahal was matched to a visually
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 501
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 4 Full results of Experiment 3 Mean proportion correct (corrected for guessing) SDs in parentheses
Lego Land- Lego Clay Nonsense Non-scenes scapes Houses objects objects objects Words Faces
1-back GR 100 094 100 100 100 094 100 100CO 091 085 091 100 100 079 074 100Normals 096(008) 095(008) 093(014) 096(008) 096(006) 099(003) 098(004) 097(011)
3-back GR ndash003 068 026 047 083 083 075 052CO 066 081 037 010 094 069 095 081Normals 066(032) 077(021) 072(015) 077(020) 086(016) 083(015) 083(017) 072(020)
5-back GR 010 034 044 047 072 083 094 053CO 031 073 075 088 081 064 100 081Normals 076(026) 077(016) 068(021) 066(016) 091(014) 083(018) 093(011) 078(021)
similar Asian palace that is not famous (see Figure11)
Results and discussionResults for GR and CO as well as for five age-matched controls are shown in Figure 12 Both
GR and CO could recognise the famous landmarksas well normals In addition both GR and CO wereusually able to name the landmarks they success-fully identified
Thus GR and CO can recognize landmarkslearned prior to injury demonstrating that at leastsome of their place recognition abilities are intactWhen combined with the fact that GR and COwere able to produce detailed maps of places theylived in prior to their injury (see Case Description)these results suggest that their deficit is primarily inencoding or consolidation of new informationrather than in recognition or recall of old informa-tion Note however that these results do not provethat their place recognition abilities are entirelynormal GR and CO might suffer from a mild rec-ognition deficit that affects identification of newly-learned stimuli like the Lego layouts in Experiment3 but not overlearned stimuli like the famous land-marks Furthermore it is possible that GR and COmight be impaired at recognising premorbidlylearned places that are defined primarily by theirspatial layout even though they are unimpaired atrecognising premorbidly learned places that aredefined primarily by a single object-like landmarkor building (see General Discussion) Neverthelessthe trend of the present evidence is to suggest thatGR and CO have particular problems with noveltopographical information
In the famous-face task GRrsquos performance wasin the normal range However he sometimes haddifficulty naming the faces even though he clearly
EPSTEIN ET AL
502 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 11 Stimuli for Experiment 4 Each famous landmark(Arc de Triomphe Taj Mahal) and famous face (Barbara BushYasser Arafat) was matched to a nonfamous building or face thatalso appeared in the experiment
FA
CE
SL
AN
DM
AR
KS
Figure 12 Results of Experiment 4 Performance measure isproportion of famous stimuli reported as ldquoknownrdquo corrected forguessing using the standard formula
recognised them For example he described Ron-ald Reagan as ldquothe gipperrdquo Nancy Reagan as ldquothewife of the gipperrdquo and Clark Gable as ldquothe movieactor who said lsquoScarlett I donrsquot give a damnrsquordquo COhad a great deal of difficulty identifying the famousfaces This result is not surprising given hisextremely low performance on the Benton FaceMatching Task In addition to his topographicproblems CO appears to suffer fromprosopagnosia
GENERAL DISCUSSION
We tested two parahippocampal patients (GR andCO) on four experiments designed to measure per-formance as a function of both task (perceptionencoding and recognition) and stimulus type(scenes objects faces and words) The only taskthat revealed a clear deficit was an n-back recogni-tion memory test (Experiment 3) In order to suc-cessfully perform this task subjects had to encode adurable representation of a visual stimulus and useit for later recognition GR and CO were signifi-cantly more impaired on this task when the stimuliwere scene-like spatial layouts (ie Lego scenesand real scenes for GR) than when they wereobjects No comparable deficits were found in otherexperiments that tested layout perception (Experi-ments 1 and 2) and placelandmark recognition(Experiment 4) Taken together these results indi-cate that GR and CO are impaired in their ability toencode novel information about the geometry ofsurrounding space into memory but are lessimpaired at their ability to encode the geometry ofnovel objects
We argue that this specific topographical defi-citmdashinability to encode the spatial layout of novelscenesmdashresults from damage to the para-hippocampal place area Although the damage inboth patients extends beyond the parahippo-campal-lingual territory normally associated withthe PPA this region is clearly lesioned bilaterally inGR and in the right hemisphere in CO Further-more functional imaging demonstrated that GRhas no functioning PPA Thus the current resultsand the earlier fMRI work (Epstein et al 1999
Epstein amp Kanwisher 1998) converge to implicateparahippocampal cortex in the encoding of infor-mation about scene geometry into memory
Our assignment of the function of memoryencoding to parahippocampal cortex is consistentwith the results of earlier case reports of patientswith parahippocampal damage who have naviga-tional difficulties in novel but not familiar environ-ments (Habib amp Sirigu 1987 Ross 1980 see alsoTeng amp Squire 1999) As with GR and CO thesepatients were either less impaired or unimpaired onnontopographical memory tasks Patients have alsobeen described who exhibit the opposite pattern ofimpairment an inability to learn faces (sometimescalled ldquoprosopamnesiardquo Cipolotti Robinson Blairamp Frith 1999 Tippett Miller amp Farah 2000) orfaces and verbal materials (Maguire amp Cipolotti1998) with an unimpaired ability to learn topo-graphical materials Taken together these resultsargue for a high degree of domain-specificity in theneural systems involved in learning of new informa-tion (Gallistel 1995) The brain may have separatemechanisms for learning faces places and wordswhich can be differentially impaired after damageto specific cortical regions (Cipolotti et al 1999Incisa della Rochetta et al 1996) The presentresults suggest that the anatomical locus of themechanism dedicated to learning new topographi-cal information is in the parahippocampal cortex(Aguirre amp DrsquoEsposito 1999)
In contrast to their impaired ability to encodenew topographical information both GR and COwere able to produce detailed maps of their previousdwellings and performed normally on the famouslandmark recognition test of Experiment 4 Theseresults are generally consistent with Teng andSquirersquos (1999) report of a patient with extensivemedial temporal damage who could recallpremorbidly learned but not postmorbidly learnedtopographical information (see also Ross 1980)Thus our results provide further evidence thatmedial temporal regions such as parahippocampalcortex may be more critical for encoding or consoli-dating new information into memory than for recallof old information
One issue that our experiments do not entirelyresolve is the precise nature of the topographical
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 503
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
information encoded by parahippocampal cortexClearly this region is critical for encoding informa-tion about the geometry of the currently visiblescene even when the ldquoscenerdquo is made out of Legoblocks and therefore has no immediate navigationalrelevance However it is unclear whether this spe-cific deficit can account for all of the real-worldnavigational difficulties encountered by GR andCO GR in particular shows such a catastrophicloss of navigational ability that we suspect he is alsoimpaired at encoding the spatial relationshipsbetweenthe current scene and other locations in theworld CO probably also suffers from such difficul-ties as he reports that he has trouble rememberinghow places relate to each other in large-scale spaceWhen he was tested on a simple 3-room videogameenvironment he not only was unable to rememberthe individual rooms (claiming that there were 10rooms when there were only 3) but he also had nomemory for the spatial relationships between themThese spatial deficits may be the result of damage tothe posterior right hippocampus (in the case of CO)or of disconnection of the posterior right hippo-campus from the parahippocampal regions thatprovide much of its cortical input (in both patientsSuzuki amp Amaral 1994) In either case the abilityto form a cognitive map of new environments likethe lab or videogame space could be impaired(Maguire et al 1998 OrsquoKeefe amp Nadel 1978)However it is also possible that parahippocampalcortex itself may be critical for encoding not onlythe geometry of individual scenes but also the spa-tial relationships between the locations of thosescenes In this case the spatial deficits of GR andCO could be attributed directly to parahippo-campal damage rather than to hippocampal dam-age or disconnection
The latter scenario would be consistent withAguirre and DrsquoEspositorsquos (1999) and Barrash etalrsquos (2000) suggestion that parahippocampal cortexis critically involved in combining visual informa-tion about the appearance of scenes and landmarkswith spatial information about their locations into aunified topographical representation Howeverboth of these authors argue that the representationsof individual scenes (Barrash et al) and landmarks(Aguirre amp DrsquoEsposito) are supported not by
parahippocampal cortex but by more posterior lin-gual-fusiform regions In contrast we believe thatthe neuroimaging data indicate that parahippo-campal cortex is involved in the encoding of thegeometric structure of individual scenes althoughit may also play a role in combining these represen-tations with other spatial representations
Interestingly GR and CO were not impaired onrecognition of famous landmarks that were mostlybuildings Thus our data do not preclude the possi-bility that there may be a separate lingual orfusiform region involved in processing informationabout buildings (Aguirre Zarahn amp DrsquoEsposito1998 Chao Haxby amp Martin 1999 IshaiUngerleider Martin Schouten amp Haxby 1999)which might have been preserved in GR and COdespite extensive damage to other regions of thelingual and fusiform gyrus Alternatively GR andCOrsquos preserved building recognition abilities mightbe mediated by interactions between cortical areasthat perform shape analysis for any object-likestimulus type (Malach et al 1995Kanwisher et al1996) and anterior temporal regions in which long-term topographical and semantic memories arestored (McCarthy et al 1996)
Is perception preserved
In contrast to their inability to encode new scenesinto memory GR and CO have substantiallyretained their ability to perceive the spatial layout ofthe currently visible scene (Experiments 1 and 2)GR can follow a route marked on a floor planwhich indicates a good on-line appreciation of hiscurrent spatial surroundings Furthermore neitherGR or CO have any trouble moving through theenvironmentmdashthey move with confidence and donot bump into doors or walls Problems of the lattertype are usually associated with damage to the pari-etal lobes which are intact in GR and CO Thus atfirst glance GR and CO do not appear to haveperceptual impairments (beyond their visual fieldrestrictions)
However GR does complain of a perceptualdeficit which he describes as an inability to ldquotakeinrdquo scenes when they are too complex Similarreports of an inability to comprehend the
EPSTEIN ET AL
504 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
ldquoglobalityrdquo of a scene were made by some of theparahippocampally-damaged patients studied byHabib and Sirigu (1987) We were not able todetermine the precise nature of this deficit whichmay be partly attributable to his visual field restric-tions It appears that his deficit is more salient in thereal-world environments than in the picture-per-ception tasks of Experiments 1 and 2 perhapsbecause he finds it more difficult to organise visualinput when immersed in it than when viewing pho-tographs that subtend only part of the visual field
We suggest that GR can perceive parts of thescene when he attends to them but may be unableto integrate information over time into a coherentrepresentation of a scene or place The fact that GRand CO are impaired for Lego layouts at 5-backintervals in Experiment 3 is consistent with thishypothesis The time between the first and secondpresentations of a stimulus is very short in this case(and even shorter with 3-back intervals where GRalso shows evidence of a deficit) In both patientsthe representation of layout information formed byperception of a Lego scene must either have anabnormally short persistence be abnormally sus-ceptible to interference or both If so they mightfind it impossible to sustain a spatial frame orschema long enough to use it to organise informa-tion obtained from many different glances at a com-plex scene (Intraub 1997) The result would be aninability to understand the ldquoglobalityrdquo of a sceneeven though all the details were perfectlyperceived
Non-topographical mnemonic abilities
Although not as severe as their topographical mem-ory problems GR and CO do have some memorydifficulties in other domains For example bothperformed poorly on the Rey AVLT whichrequired them to remember a list of unrelatedwords Both also report some subtle episodic mem-ory problems (see Case Description) COrsquos prob-lems could partly result from damage to the righthippocampusmdashindeed CO appeared to have themore acute episodic memory problems HoweverGR has no hippocampal damage so the fact that hedisplays episodic memory problems suggests thatparahippocampal cortex is involved in the forma-
tion of episodic memories (cf Wagner et al1998)
In previous work (Epstein et al 1999) weargued that spatial context information encoded byparahippocampal cortex might play a role in estab-lishing event memories by encoding informationabout where events take place The inability toencode this information might cause problems intasks such as the Rey AVLT where one has to recalla list of unrelated words If spatial (and perhapstemporal) context information is not encoded itwill be harder subsequently to recall the distinctevent of hearing each word (Tulving 1983) Incontrast lack of spatial context information wouldbe less of a problem in tasks where one is onlyrequired to recognise previously encountered verbalmaterials but not to reproduce the spatiotemporalcontext in which they were experienced IndeedGR and CO performed as well as normals on thenonsense words in Experiment 4 where recogni-tion memory but not episodic memory was requiredfor successful performance Interestingly GR andCO also did relatively well on the Logical Memorysubtest of the WMS-III perhaps because the nar-rative framework of a story provides sufficient con-textual information to overcome the absence ofspatial context and mediate successful verbalretrieval
Differences between GR and CO
Although we have discussed GR and CO togetherthere are a number of relevant differences betweenthem One open question is whether these differ-ences can be attributed to the different effects ofunilateral and bilateral parahippocampal damageIn general GRrsquos topographical difficulties weremore severe than COrsquos CO did surprisingly well onthe Warrington Topographical Memory Test inwhich he was required to remember detailed real-world city scenes and also performed substantiallybetter on real scenes than Lego scenes in Experi-ment 3 Insofar as CO has intact left para-hippocampal cortex this suggests that the left PPAmight be sufficient to encode a certain amount of(perhaps semantic) information about real-worldscenes but the right PPA is necessary to encode
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 505
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)

INTRODUCTION
All mobile organisms must solve the fundamentalproblem of navigation One way to gain insight intothe cognitive and neural mechanisms underlyingnavigation is to investigate the functional organisa-tion of the brain areas involved Research on ani-mals and humans has suggested that a number ofregions play a role including the parietal lobes(Stark Coslett amp Saffran 1996) retrosplenial cor-tex (Chen Lin Green Barnes amp McNaughton1994)hippocampus (Maguire et al 1998OrsquoKeefeamp Nadel 1978) and parahippocampal cortex(Aguirre Detre Alsop amp DrsquoEsposito 1996Bohbot et al 1998) Here we focus on para-hippocampal cortex We describe two patients whosuffered damage to this region of cortex after vascu-lar incidents Both subsequently developed severedeficits in the topographical domain which wedefine as the domain of information relevant tospatial navigation
Previous reports in the neuropsychological liter-ature have described patients with ldquotopographicaldisorientationrdquo who are unable to successfully findtheir way through locomotor space but do notexhibit general cognitive or memory impairments(see Aguirre amp DrsquoEsposito 1999 for review) Thenature of the specific problem varies from patient topatient For example some patients show an inabil-ity to understand spatial relationships between dif-ferent locations (Levine Warach amp Farah 1985patient 2) whereas others are unable to recognisenavigationally relevant stimuli such as landmarksbut can accurately describe routes between theplaces they cannot identify (Pallis 1955) Patientsalso differ in their relative ability to handle novel vsfamiliar topographical information Some are ableto navigate through familiar environments or recallold topographical information but cannot learnnovel environments (Habib amp Sirigu 1987patients 1 and 2 Ross 1980Teng amp Squire 1999)whereas others are impaired at both learning of newmaterials and recognitionrecall of old materials(Habib amp Sirigu 1987 patients 3 and 4 Incisadella Rocchetta Cipolotti amp Warrington 1996Landis Cummings Benson amp Palmer 1986
McCarthy Evans amp Hodges 1996 Whiteley ampWarrington 1978)
Based on comprehensive review of the topo-graphical disorientation literature Aguirre andDrsquoEsposito (1999) argued that the lingual-parahippocampal region is one of several areas thatare critically involved in navigation Damage to thisregion often results in an inability to use salient top-ographical features such as landscapes and build-ings to orient oneself (ldquolandmark agnosiardquo egLandis et al 1986 Pallis 1955) or in a more gen-eral inability to learn new topographical informa-tion (ldquoanterograde disorientationrdquo eg Habib ampSirigu 1987 patients 1 and 2) In a recent study of127 patients with focal lesions Barrash DamasioAdolphs and Tranel (2000) observed that patientswith damage to this region almost always displayedsevere impairment on a real-world route learningtask Furthermore Bohbot et al (1998) found thatpatients with right-hemisphere parahippocampalcortex lesions were impaired at a human analogueof the Morris water maze task in which they wererequired to remember the location of a hidden floorplatform within a room (a deficit not found inpatients whose damage was restricted to the hippo-campus proper)
Neuroimaging studies have also implicated thelingual-parahippocampal region in navigationSeveral studies have examined brain activity in nor-mal subjects during performance of a number ofnavigational tasks including navigation through avirtual reality environment (Aguirre et al 1996)watching videotapes depicting navigation througha real environment (Maguire Frackowiak amp Frith1997) and mentally imagining navigation througha real environment (Ghaem et al 1997 Maguire etal 1998) All of these studies found greater activa-tion in the parahippocampal region (and sometimesother medial temporal regions as well Ghaem etal 1997Maguire et al 1997 1998) when the nav-igation task was compared to a less navigationallydemanding control task
What role does parahippocampal cortex play inthese navigational tasks In earlier work weaddressed this question by using fMRI to examineneural activity in parahippocampal cortex and other
EPSTEIN ET AL
482 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
brain regions while subjects viewed a wide variety ofvisual stimuli (Epstein amp Kanwisher 1998) Wefound that a region abutting the collateral sulcusnear the parahippocampal-lingual boundaryresponded significantly more strongly when sub-jects viewed navigationally relevant stimuli such asstreet scenes landscapes or buildings than whenthey viewed other kinds of visual stimuli such asfaces objects or scrambled scenes1 This responsewas found even when subjects simply watched thestimuli passively and were not required to performany navigational task We named this region theldquoparahippocampal place areardquo or PPA because itresponded strongly to depictions of places ThePPA appears to overlap with the regions activatedin previous neuroimaging studies of navigation(eg Aguirre et al 1996)
A critical determinant of PPA activationappears to be the presence in the stimulus of infor-mation about the shape or layout of the immediateenvironment The PPA responds no more stronglyto photographs of rooms filled with furniture andobjects than it does to photographs of the samerooms empty of all objects (ie just bare walls) Italso responds much more strongly to ldquoscenesrdquo madeout of Lego blocks (which have a geometric struc-ture similar to a room or street scene) than it does toobjects made out of the same Lego materials(Epstein Harris Stanley amp Kanwisher 1999)Thus the PPA is sensitive to the presence of a spe-cific kind of geometric organisation in the stimuluseven if the stimulus does not depict a real place inthe world In particular it responds more stronglyto depictions of surfaces that in some senseldquoencloserdquo the observer and define a space withinwhich one can act (ie the walls of a room or theside of the distant hill) than to depictions of surfacesthat define objects that the observer can act uponThese results are generally consistent with behav-ioural results from rats (Cheng 1986 Gallistel1990 Margules amp Gallistel 1988) and human
infants (Hermer amp Spelke 1994 1996 Hermer-Vazquez Spelke amp Katsnelson 1999) that suggestthat information about the geometry of surround-ing spacemdashthe ldquolay of the landrdquomdashplays a privilegedrole in orientation and navigation
What specific cognitive task does the PPA per-form Given the fact that landmark agnosia oftenresults from parahippocampal-lingual damage(Landis et al 1986) one might hypothesise thatthe primary role of the PPA is place recognitionidentification of familiar locations based on theirappearance andor geometric structure If so onemight expect the PPA to be sensitive to the noveltyfamiliarity of the place depicted in the stimulusHowever we found that the PPA responds just asstrongly to photographs of unfamiliar places (ieplaces the subjects had never visited) as it does tophotographs of familiar places (Epstein et al 1999Expt 1) In contrast the PPA is sensitive to thenoveltyfamiliarity of the stimulus itself respond-ing more strongly to new photographs than to pho-tographs viewed many times previously even whenall the photographs depict unfamiliar places(Epstein et al 1999 Expt 4) From these resultswe inferred that the PPA is more involved inencoding a representation of the spatial structure ofthe current scene than in relating that scene to onersquosstored cognitive map of the environment as awhole However our data did not allow us to deter-mine whether the PPA activation reflects percep-tual encoding memory encoding or both
The purpose of the present report was to moreprecisely determine the function of the PPA Tothis end we tested the ability of our two patients toperceive learn and recognise a variety of visualmaterials We predicted that the patients would bemore impaired at processing scene-like stimuli (iestimuli that depict spaces that one can navigatethrough or act within) than object-like stimuli Byusing several different kinds of tests we aimed todetermine whether any such selective impairment
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 483
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
1Although full scenes activate the PPA more strongly than do any other kind of visual stimulus we have also observed higher
activity in the PPA when subjects view buildings than when they view other kinds of objects (Epstein et al 1999) consistent withAguirre Zarahn and DrsquoEspositorsquos (1998) finding of a significant response to buildings in an adjoining and possibly overlapping regionof the anterior lingual gyrus
was in the perceptual recognition or memorydomain To anticipate we did find evidence for adeficit for visual materials that conveyed informa-tion about the shape of surrounding space Thisdeficit manifested itself during a memory encodingtest but not during perceptual tests or a recognitiontest Thus our data suggest that the PPA is neces-sary for the encoding of spatial layout informationinto memory but may not be necessary for manyaspects of the initial perceptual analysis recogni-tion or recall of this information We argue that thePPA may be a learning mechanism specifically ded-icated to encoding topographical materials intomemory
CASE DESCRIPTIONS ANDNEUROPSYCHOLOGICAL TESTING
In this section we present the case histories of thetwo patients in order to provide a general contextfor the more specific experimental tests reportedin the Experimental Investigations section Thepatients were chosen for investigation because clin-ical MRIs indicated that they had suffered damageto the parahippocampal region This section is
divided into four subsections First we present ageneral overview of the case histories Second wereport the results of some ad hoc tests that give ageneral indication of the patientrsquos navigational abil-ities Third we discuss other cognitive abilitiesincluding face recognition and episodic memoryFourth we report the results of a battery of formalneuropsychological tests performed on the patientsTable 1 summarises relevant observed and self-reported behaviour and Table 2 summarises theresults of the neuropsychological tests
Overview
Case 1 GRGR2 is a right-handed male with 18 years of educa-tion He was 60 years old at the time of testingTwo-and-a-half years prior to testing he suffered aright occipital-temporal stroke during cardiacsurgery A second stroke affecting the left occipital-temporal region occurred a week later Thesestrokes left him with a number of deficits includinglimitation of his peripheral vision (lefthemianopsia right upper quadrantanopsia)dyschromatopsia (details reported in Beauchampet al 2000) and topographical disorientation We
484 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
EPSTEIN ET AL
Table 1 Summary of observed and self-reported behaviour
GR CO
Real-world topographical Unable to recognise 714 newly learned MIT landmarks Unable to learn 3-room videogamelearning despite extensive training regime environment
Unable to find way back to lab unassisted despite Able to learn a simple route out ofextended experience with environment building after many visits
Map drawing Could not draw map of lab maps of other newly learned Map of lab was sketchy but not entirelyenvironments were inaccurate inaccurate
Drew accurate map of premorbid environment Drew accurate map of premorbid(former home) environment (currentformer home)
Scene perception Reports difficulty understanding the globality of complex Does not report a perceptual deficitscenes
Face recognition Some prosopagnosia Severe prosopagnosia
Episodic memory Largely intact though some difficulties were reported Moderate difficulties were reported
Can follow the plot of a movie or novel and report Reports some difficulty following theprevious dayrsquos activities plot of a book Accurately reported
contents of previous dayrsquos newspaper
2GRrsquos true initials are KG (cf Beauchamp Haxby Rosen amp DeYoe 2000)and COrsquos true initials are KC We use GR and CO
here to avoid confusion between the two patients
tested him over a period of 5 days in August 1998with 3 days of follow-up testing 3 weeks later and 1day of additional tests in May 1999 During thistime he was alert and cooperative Emotionalaffect and language use was normal He interactednormally in nonclinical social situations and hisconversation was appropriate and varied Heremembered the events of previous days andrecounted them accurately
In contrast to this high level of non-topographical functioning GR demonstrated adramatic inability to orient himself in space Forexample even after several days of testing he neverlearned the relative locations of the different roomsof the lab According to both GR and his wife thisinability to learn new topographical informationwas typical of his experience since his strokes as hefrequently gets lost in his daily life Soon after his
injury he moved to a neighbourhood with manysimilar-looking houses He reported that in orderto find his new house after a walk to a market six toseven blocks away he had to rely on street signs toguide him to the correct block and then examineeach house on the block in detail until he couldrecognise some feature that distinguished his homeuniquely Subsequently GR and his wife moved toa different house in a different country He reportedthat for the first 6 months after the move his newhome was like a ldquohaunted houserdquo for him insofar ashe was unable to learn his way around it For exam-ple he would repeatedly forget that the bedroomhad a balcony and he would be surprised by this factevery time he rediscovered it After several monthshowever he was able to learn the topography of thehouse sufficiently well to navigate through it (butsee below)
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 485
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 2 Summary of neuropsychological tests ( indicates percentile scores)
GR COmdashmdashmdashmdashmdashmdashmdashmdashmdashmdash mdashmdashmdashmdashmdashmdashmdashmdashmdashmdashmdashndash
IQ 122 (93 WAIS-R) 134 (99 WAIS-III)
WMS-III
Logical memory 25 immediate recall 27 immediate recall50 delayed recall 48 delayed recall(WMS-R)
Face memory 9 immediate recall 16 immediate recall16 delayed recall 9 delayed recall
Family pictures 2 immediate recall lt1 immediate recall5 delayed recall lt1 delayed recall
Rey Auditory Verbal 63 immediate recall 45 immediate recallLearning test 14 after interference list lt1 after interference list
16 delayed recall 3 delayed recall
Warrington Recognition 50 wordsMemory tests 29 scenes
lt5 faces
724 Spatial Recall 66 immediate recall2 delayed recall
Benton Face Recognition 42 lt1
Memory span Digits99 forward99 backward
Spatial37 forward50 backward
Boston Naming test Ceiling Ceiling
Case 2 COCO is a left-handed male with 20 years of educa-tion He was 60 years old at the time of testing Twoyears prior to testing he suffered a right posteriorcerebral artery stroke He first noticed on a week-end that his vision appeared blurred on the left andthat he had a tendency to bump into objects on thisside On Monday morning he was able to success-fully negotiate the route to his workplace a trip thatinvolved a bus ride followed by two subways andthen a shuttle to his office building However oncehe entered the building he found himself com-pletely disoriented unsure of its layout and wherehis office was located Interestingly the companyhe worked for had moved a few weeks earlierAlthough he had managed to arrive at this newbuilding (which was a block away from the previouslocation) he became lost upon entering it Hisneurologic exam revealed a left homonymoushemianopia but no evidence of neglect on a lettercancellation task or on sensory testing Investiga-tions revealed an ischaemic stroke with haemor-rhagic transformation in the right occipital andmesial temporal lobe On magnetic resonanceangiography he had a chronic left carotid occlusiona mild right carotid stenosis and bilateral vertebralartery stenoses Since then he has suffered fromsevere topographical disorientation primarily innew environments
CO reports that he can recognise buildings thatwere familiar to him before the stroke He can alsoform mental images of well-known landmarks suchas the Empire State Building and the BostonPublic Gardens and of the house he lived in as achild When asked to imagine standing in front ofthe Massachusetts State House (a location wellknown to him from years of living in Boston) heaccurately described the surrounding buildings andtheir locations However he reports that when helooks at a place that hersquos encountered post-strokeall he gets is a feeling of familiarity that does notallow him to determine where the place is or whenhe saw it He also reports an inability to understandhow various landmarks relate to each other inlarge-scale space Despite this difficulty he canfollow a map and frequently rides the Bostonsubway alone
Unlike GR CO was able to learn a certainamount of topographical information For exam-ple he soon learned the location of the rooms in thelab and would travel between them unassistedAfter his first visit to the lab he needed to be guidedall the way back to the subway station but after sev-eral visits he could find his way out of the buildingand to the subway on his own as long as he was firstguided to the elevators (It should be noted how-ever that the building is on the same block as thesubway station Furthermore he still needed to bedirected to the elevators even though our buildinghas a relatively simple floor plan) Thus the amountof topographical information he can learn is limitedbut not zero
COrsquos language and reasoning skills appear to benormal and his conversation is appropriate andvaried In addition to his topographical difficultieshe also has limitations of his peripheral vision onthe left side
Navigational abilities
In addition to the experiments reported in theExperimental Investigations section of this paperwe also performed a number of ad hoc tests of GRand COrsquos navigational abilities Although thesetests were informal we describe them here to pro-vide a general sketch of the nature of GR and COrsquosimpairments
Real-world topographical learningOver the course of several days we took GR and hiswife on three walks around the MIT campus whichneither had visited previously Each walk followedexactly the same path At several places in the walkwe would stop and point out different buildings andlandmarks tell them something about each one andask them to remember their appearance for latertesting Immediately after the completion of thethird walk both GR and his wife separately per-formed a forced-choice recognition test in whichthey saw photographs of 14 landmarks (mostlybuildings) from the walk and 14 novel landmarks(from another part of the MIT campus that theyhad not visited) and were required to reportwhether they had seen each one or not GRrsquos
EPSTEIN ET AL
486 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
performance was 714 correct hits for the familiarlandmarks and 1314 correct rejections for theunfamiliar landmarks In contrast his wifeobtained a perfect score on this test Given theextensive nature of the training regime GRrsquos scorestrongly suggests an impairment in learning theappearance of new real-world places Interestinglyseveral of the seven familiar landmarks that he gotright had an obvious distinctive feature that hadbeen explicitly pointed out to him For exampleone building had a distinctive clock tower andanother one was an unusual triangular shape ThusGR was occasionally able to use distinct visualfeatures to identify places even though he wasunable to recognise them holisticallymdasha commonpattern among topographical disorientationpatients
GR did not appear to have any obvious problemsin understanding the spatial structure of his imme-diate environment When given a floor plan of thebuilding at MIT in which he was being tested hewas able to successfully follow a route marked onthe plan However when the plan was taken awayhe had great difficulty finding his way back to thelab even though he had more than 30 min experi-ence with the environment and his starting positionwas less than 30 feet from the goal (Figure 2) Infact the route he took resembled a random walk Atone point he walked right past his destinationwithout recognising it and only knew for sure thathe was in the right place when he identified his wifein the room Thus his on-line appreciation of thelayout of immediately visible space appears to beintact but he cannot find his own way withoutassistance
We were unable to perform the MIT landmarkrecognition test on CO because he suffered from amedical problem that made it difficult for him towalk around campus In lieu of this we tried to seewhether he could learn a simple video game envi-ronment consisting of three connected roomsAfter about 30 min experience navigating throughthis environment (via button presses) he was askedto sketch a map of it He declined to do so statingthat it was beyond his abilities When asked howmany rooms there were he replied that he did notknow but that there were at least 10 Given the
simplicity of this artificial environment COrsquos com-plete inability to learn it is striking
Map drawingAfter several days of testing we asked both GR andhis wife to sit at different tables and independentlysketch (1) the shape of Florida (2) a bicycle (3) afloor plan of their hotel room in Boston (4) a floorplan of the first floor of the house they had been liv-ing in for the last 8 months (all post-stroke) (5) afloor plan of the main room of the lab and (6) afloor plan of the house they lived in 10 years ago(pre-stroke) GR drew the map of Florida and thebicycle perfectly His plan of the hotel room wasnotably different from the one drawn by his wifemdashfor example he reported that there was only onebed when in fact there were two and he placed thebed on the wrong side of the room (Figure 1a) Hisplan of the first floor of his current house was some-what better but was not very detailed and includedsome major errors For example he wronglyreported that there was no bathroom on this floor(Figure 1b) He was unable to produce any floorplan of the main room of the lab leaving this pageblank When his wife reminded him that this wasthe room in which he had just spent an hour readingthe newspaper and in which we had all had break-fast in that morning he replied that he didnrsquot knowwhat room she was talking about (although heappeared to remember these events) In contrast tothis near total inability to produce accurate maps ofnew environments GR produced an accurate mapof the house he lived in prior to his injury (Figure1c) He reported that his memory of this house wasldquoall thererdquo and he could imagine walking around init When asked to produce a perspective of a roomwhen he was in it he did so with ease and he wasalso able to create an accurate floor plan of theroom
In contrast CO found it very difficult to pro-duce a floor plan of the lab both when he was in itand when he was taken to another room and askedto recall it However although the maps he pro-duced were quite impoverished they were notentirely inaccurate demonstrating some ability toencode topographical information This contrastswith GRrsquos absolute inability to recall anything
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 487
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
EPSTEIN ET AL
488 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure
1F
loorplansdraw
nby
GR
andhisw
ifeof(a)
theirhotelroomin
Boston(b)theircurrenthousew
hichthey
hadm
ovedto
subsequenttoG
Rrsquosinjury(c)the
housethey
livedin
priorto
GR
rsquosinjury
Ho
telro
om
inB
osto
n
GR
His
wife
His
wife
Cu
rren
th
om
e
GR
His
wife
Fo
rme
rh
om
e
GR
about the main room of the lab once he had left itCO was also able to draw a detailed floor plan of theapartment he was living in at the time (and hadlived in for 20 years) Several months after he hadmoved from this apartment we asked him to drawsuch a floor plan again and he produced one thatwas substantially the same as the first (Figure 3)Thus their map drawing performance indicatesthat GR and CO are primarily impaired at sketch-ing places experienced after their strokes but areunimpaired at sketching places experienced beforetheir strokes This pattern is indicative of deficits inencoding but not recall
Scene perceptionAs noted earlier GR can navigate with the aid of afloor plan suggesting that his perceptual abilitiesare largely intact Interestingly however he reportsthat he does have a perceptual deficit which hedescribes as an inability to understand scenes whenthey are too complex For example he reports thathe often has trouble using maps because theyusually are too busy with details It is unclearwhether this apparent perceptual problem is a resultof his field cut some other separate perceptual
problem or a memory problem (see discussion)CO did not complain of a similar deficit GR wasan accomplished artist prior to his injury producingpaintings of striking complexity Tragically hereports that he cannot now appreciate his ownpaintings
Other cognitive abilities
Face recognitionGR also reported some difficulties with face recog-nition On his second visit we examined his abilityto learn new faces by having him perform a forced-choice recognition test on seven familiar and sevenunfamiliar faces The familiar faces were personnelfrom the lab with whom he had at that point had agood deal of experience spanning at least 3 daysAll the people in the photographs had a towelwrapped around their head in order to minimise theuse of hair as a nonfacial cue Despite the difficultyof this task he correctly identified 67 familiarfaces and made no false alarms Surprisingly theone face that he did miss was the first author of thispaper with whom he at that point had had consid-erable experience GR also performed normally ona difficult famous face recognition test (see Experi-ment 4 following) and on the Benton Face Recog-nition test (see following) Thus he has someability to recognise faces Consistent with this GRreports that he can remember a small number offaces but that he quickly ldquoloses trackrdquo if he has toremember too many
CO also appears to suffer from someprosopagnosia as evidenced by his low perfor-mance on the Benton Face Recognition test (seefollowing see also Experiment 4) Interestingly hedoes not complain of face recognition difficultiesbut rather attributes his inability to recognise peo-ple to a lack of interest in their appearance
Episodic memoryAt first glance GRrsquos episodic memory appeared tobe largely intact As mentioned he could recall theevents of the previous day accurately Furthermorehe reports that he can follow the plot of a moviewithout any difficulty However both he and hiswife report that he does have some problems with
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 489
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Figure 2 Floor plan of the building at MIT in which GR wastested Corridors are highlighted in grey GR was asked to find themain room of the lab (marked G on the map) Starting location(marked S) was a point approximately 30 feet down the hall GRrsquosroute is marked by arrows GR found this task very difficult andonly knew for certain that he had reached his goal when he caughtsight of his wife through the glass door of the lab In contrast hewas able to follow a route marked on a plan of the same floorwithout any problem or hesitation
episodic memory For example he stated that heremembered very little about a trip to Italy they hadtaken several months previously His wife reportsthat he has great difficulty keeping track of timeFor example before their trip to Boston he repeat-edly asked her what day they were supposed to leaveand seemed unable to encode this information Weobserved a similar phenomenon during testingDuring the tests that required repetition of thesame procedure several times he sometimesseemed unable to clearly remember how manytimes he had repeated the procedure Thus he doesappear to have a subtle episodic memory deficit
which may manifest itself as an inability to sepa-rately remember successive events that are not easilydistinguishable from each other
CO also reports some problems with his epi-sodic memory He relies on lists to rememberappointments stating ldquoif I didnrsquot have a list Irsquodprobably forget why I walked in the door hererdquoDespite this claim he has never missed an appoint-ment nor is he ever confused about the purpose ofthe testing appointments He can accurately reportthe day of the week He remembers the experi-menters clearly from appointment to appointmentas well as the details of the various experiments
EPSTEIN ET AL
490 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 3 Two floor plans drawn by CO of the apartment he lived in for over 20 years The first plan was drawn when CO still lived inthe apartment the second was drawn several months later when he had moved out Note that the two plans correspond very well COreported that it was quite easy to produce these floor plans
CO home Feb 99
CO (former) home June 99
when they are repeated across appointments Whenqueried about his episodic memory difficulties COreported that he will usually remember an event ifhe is reminded of it but that he has difficulty spon-taneously regenerating events Interestingly hereports that even when he is reminded of an eventand remembers it he cannot remember ldquowhere itwas or when it wasrdquo When he reads he finds it dif-ficult to follow a story if it skips around Howeverwhen queried he was able to accurately report whathe had read in the newspaper the previous day
Formal neuropsychological tests
Formal neuropsychological testing was performedon GR in September 1998and on CO in December1998 Test performances were compared to age-matched peers3
GR exhibited mild to moderate impairment inmemory relative to what would be predicted for hisintellect (WAIS-R Full Scale IQ =122 93rd per-centile superior range) and level of educationThese memory problems were more apparent onvisual than on verbal memory tests On a test ofmemory for conceptually organised text passages(Wechsler Memory Scale Logical Memory[WMS-III] Wechsler 1997b) his performancewas average for both immediate (LM I =25th per-centile standard score SS = 90) and delayed recall(LM II =50th percentile SS =100) In contrast hisscore on the WMS-III Face Memory subtest was inthe low average range for both immediate (9th per-centile SS =80) and delayed recall (16th percentileSS =85) On a test of memory for scenes (WMS-IIIFamily Pictures subtest) immediate recall was defi-cient (2nd percentile SS = 69) and delayed recallwas borderline impaired (5th percentile SS =76)GR also had difficulty learning word lists on theRey Auditory Verbal Learning Test (Spreen ampStrauss 1998) his immediate recall was average(63rd percentile SS = 105) but after the interfer-ence list his performance fell to the low averagerange for both immediate (14th percentile SS =84)
and delayed recall (16th percentile SS =85) On arecognition test of the items of the list he success-fully recognised all 15 but made 8 false alarms Per-formance on the 724 Spatial Recall Test (RaoHammeke McQuillen Khatri amp Lloyd 1984)which required him to remember seven locations ona 6times4 checkerboard was 66th percentile (SS =106)for immediate and 2nd percentile (SS = 69) fordelayed recall Performance on the Benton FaceRecognition test was normal (42nd percentile)Performance on the Boston Naming Test was atceiling indicating that language abilities wereintact Thus GR exhibits some impairment ontasks that require delayed recall of visual informa-tion (WMS-III Face Memory subtest) or verbalinformation that is not organised in terms of a storyor plot (Rey AVLT) and is clearly impaired ontasks that required delayed recall of spatial informa-tion (724 Spatial Recall Test) or stimulus location(WMS-III Family Pictures subtest) In contrasthis ability to recall organised verbal information(WMS-III Logical Memory subtest) appears to beintact
As with GR COrsquos memory performance wasgenerally below what would be expected based onintellect (WAIS-III Full Scale IQ =134 99th per-centile very superior range) and education levelAlso like GR there was some evidence that visualmemory problems were more pronounced than ver-bal memory problems Memory for text passages(WMS-R Logical Memory) was average for bothimmediate (27th percentile SS = 91) and delayedrecall (48th percentileSS =99) In contrast perfor-mance on the WMS-III Faces subtest was lowaverage for both immediate (16th percentile SS =85) and delayed recall (9th percentile SS =80) andperformance on the Family Pictures subtest wasdeficient (lt 1st percentile SSlt 62) for both imme-diate and delayed recall On the Warrington Rec-ognition Memory Tests (RMT) he scored in theaverage range for words (50th percentile SS =100)and scenes (Warrington Topographical Recogni-tion Memory Test 29th percentile SS =92) and in
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 491
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
3The formal neuropsychological testing reported in this section was performed prior to the beginning of our combined
investigation of GR and CO (ie prior to the main experimental investigations of this paper) Consequently GR and CO were nottested on exactly the same battery of tests
the deficient range (lt 5th percentile SS lt 75) forfaces The low score for faces on this test as well asthe WMS-III may reflect prosopagnosia ratherthan a memory problem as he scored in the defi-cient range (lt1st percentile SSlt62) on the BentonFace Recognition test Verbal list learning (ReyAVLT) was average on immediate recall (45th per-centile SS =98) but after hearing an interferencelist performance fell to the deficient level (015words recalled) for both immediate and delayedrecall On a recognition test for these words herecognised 1115 and made one false alarm (39thpercentile SS =96) There was a large discrepancyon tests of memory span which may indicate adecline in spatial memory digit span was superior(9+ forward and 7 backwards both 99th percentile)but spatial span was only average (5 forward and 5backward 37th and 50th percentiles respectively)Language abilities were intact (Boston NamingTest performance was at ceiling) Thus like GRCO shows some impairment at recall of visualinformation (WMS-III Face Memory subtest) andverbal information that is not organised into a nar-rative (Rey AVLT) and clear impairment at tasksthat require recall of stimulus location (Family Pic-tures subtests of WMS-III) In contrast his abilityto recall organised verbal information (WMS-RLogical Memory subtest) is more preserved
MRI RESULTS
In order to determine the extent of their lesionsstructural MRI images were taken of both subjectsDelineation of the lesions was based either onspoiled GRASS coronal images at 11mm thick-ness (GR) or anatomical T-1 weighted MPRAGEMRI sequences yielding 125 mm thick coronalimages (CO) The anterior-posterior extent of thelesions was verified on axial images
GR has bilateral occipital-temporal lesions(Figure 4) In the left hemisphere the posteriorparahippocampal gyrus (including para-hippocampal cortex) is severely damaged with onlyminimal injury to the most caudal aspect of the hip-pocampus There is also moderate damage to themedial fusiform and inferior lingual gyri In the
right hemisphere the hippocampus is spared butthere is damage to the posterior end of theparahippocampal gyrus near the parahippocampal-lingual boundary More posteriorly the lesionedarea extends along the collateral sulcus to includesignificant portions of the inferior lingual gyrus andthe medial fusiform gyrus as well as most of themedial occipital lobe below the calcarine fissure
CO has a right-hemisphere occipital-temporallesion Right posterior hippocampus and para-hippocampal gyrus are severely affected Moreposteriorly the damage covers large portions of theinferior lingual and medial fusiform gyri as well assubstantial portions of the medial occipital lobeincluding most of right calcarine cortex In addi-tion diffuse foci of increased signal on T-2weighted images was present in periventricularwhite matter in the corona radiata and centrumsemiovale with a lacunar infarct of the rightputamen There was also moderate degree of dif-fuse involutional changes with increased ventricularsize particularly notable in the right temporal hornwith ex vacuo dilitation
The structural MRIs suggested that the PPAwas severely if not completely damaged in bothhemispheres for GR and in the right hemispherefor CO In order to confirm this for GR we used ahigh-field 3-T magnet to perform a functional scanin which he viewed scenes faces and objects in thesame blocked fMRI design that we have used tolocalise the PPA in all our previous experiments(see Epstein et al 1999 for details Epstein ampKanwisher 1998) A Kolomogorov-Smirnov testfound no voxels in the parahippocampal region thatfit our previously established criterion for inclusionin the PPA (ie greater response to scenes than tofaces and objects at a uncorrected significance levelof plt 0001) This result contrasts sharply with thepattern seen in over 95 of (mostly young or mid-dle-aged) normal subjects in whom a large acti-vated region can be identified in both left and rightparahippocampal cortex using this comparison (seeFigure 5) A similar test was used to identify voxelsthat responded significantly more to faces than toobjects This comparison revealed a region of face-selective voxels in the right fusiform gyrus consis-tent with the location of the Fusiform Face Area
EPSTEIN ET AL
492 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 493
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
GR
CO
Figure 4 Structural MRIs of GR and CO showing the extent of the damaged region (marked as white) Images have been normalisedusing SPM99 and a cost function masking algorithm (provided by Matthew Brett) into standard stereotactic space (Talairach amp Tournoux1988) using the Montreal Neurological Institute template (Evans et al 1993) Numbers indicate y-Talairach coordinate for each coronalslice corresponding slice planes are also indicated on the sagittal image
(FFA) commonly seen in normal subjects(Kanwisher McDermott amp Chun 1997) Thusthe MRI was sensitive enough to detect functionaldifferences an FFA was found but no PPA Whencombined with the anatomical results this stronglysuggests that GR has no PPA
Because of safety concerns (presence of a femo-ral artery aneurysm clip) we did not perform a simi-lar high-field functional MRI of CO However asFigure 4 demonstrates CO clearly has damage tothe parahippocampal-lingual region in the righthemisphere
EXPERIMENTAL INVESTIGATIONS
In this section we report the results of four experi-ments designed to probe the consequences of dam-age to the lingual-parahippocampal region ingeneral and the PPA in particular Our primaryconcern in these experiments was to test GR andCOrsquos ability to perceive remember and recognisedifferent kinds of visual material Based on our pre-vious fMRI work we predicted that GR and CO
would have more difficulty with scene-like stimuli(ie stimuli that convey information about thegeometry of a space one can navigate through or actwithin) than with object-like stimuli Such a deficitcould account for at least some of the problems GRand CO encounter in real-world navigation
Beyond determining the kind of visual materialthat gets processed in parahippocampal cortex wealso wanted to determine the nature of the process-ing that takes place on that material In particularwe wanted to determine if parahippocampal cortexis involved in perceptual encoding memory encodingor recognition As discussed earlier (Introduction)our fMRI results provided some evidence againstplace recognition as the sole function of the PPAbut did not allow us to distinguish between percep-tual and mnemonic encoding Experiments 1 and 2examine the role of parahippocampal cortex in per-ceptual encoding Experiment 3 examines its role inmnemonic encoding and Experiment 4 examinesits role in place recognition
Experiment 1 Two-dot task
The first hypothesis we examined was thatparahippocampal cortex plays a role in perceivingthe shape of the local environment In our previousfMRI work we observed that the PPA respondsmuch more strongly during scene viewing than dur-ing object or face viewing even when subjects donothing more than watch the scenes passively with-out performing any other task This suggested thatscene perception alone may be sufficient to activatethe PPA Previous reports have indicated that dam-age to parahippocampal cortex and other medialtemporal regions leads to problems with memoryrather than perception (Bohbot et al 1998Maguire Burke Philips amp Staunton 1996 Pigottamp Milner 1993) However all of these previousresults were obtained on patients who had unilat-eral lesions GR has a bilateral occipital-temporallesion including clear damage to parahippocampalcortex in both hemispheres furthermore he reportssome kind of perceptual deficit We hypothesisedthat this deficit might involve difficulty perceivingor interpreting the geometry of scenes and otherstimuli with a similar geometric structure
EPSTEIN ET AL
494 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 5 Top row shows a single slice from a functional MRIscan performed on GR Bottom row shows results from a normalsubject (NK) for comparison Voxels that responded more to facesthan objects (left column) or more to scenes than faces and objects(right column) in a Kolomogorov-Smirnov test are highlighted bywhite arrows GR has a functioning face area (FFA) but there isno evidence of a place area (PPA)
FFA PPA
GR
NK
In this experiment subjects viewed either objectshapes or spatial layouts Stimuli consisted ofblack-and-white photographs of real-world stimuli(empty rooms vs common objects) or abstractstimuli made out of Lego blocks (Lego ldquoscenesrdquo vsLego ldquoobjectsrdquo) These stimuli have been previouslyshown to differentially activate the PPA in normalsubjects (Epstein et al 1999 Epstein ampKanwisher 1998) Two red dots were superim-posed on each photograph (Figure 6) The task wasto report which of the two red dots was over thecloser part of the object or scene This task wasdesigned to have no memory component (apartfrom remembering the instructions) but to requireaccurate 3D perception of objects and scenes
If GR or CO have difficulty perceiving or inter-preting the spatial layout of scenes then we pre-dicted they would do worse when the stimuli areempty rooms or Lego scenes than when they arereal-world or Lego objects Note that the criticalcomparison is between the Lego scenes and theLego objects as these stimuli are made of the samematerials and differ primarily in their surfacegeometry
MethodTesting for all experiments was done on aMacintosh computer The experiment consisted of80 trials (20 of each stimulus type randomly inter-mixed) Subjects began each trial by pressing thespace bar A black-and-white photograph from oneof the four stimulus categories (see earlier) thenappeared on the screen with two red dots on it Thetask was to determine which of the two red dotsindicated the closer part of the scene or objectSubjects pressed the ldquofrdquo key if the leftmost red dotwas closest and the ldquojrdquo key if the rightmost dotwas closest They were instructed to respond asquickly as possible without making errors Stimuliremained on the screen until a response was made
The stimulus set was constructed so that eachempty room was matched to a common object thathad red dots in the exact same location and eachLego layout was similarly matched to a Lego objectThus the locations of the red dots were counterbal-anced across scene and object stimuli controllingfor any effects of visual field cuts
Results and discussionResults for GR and CO and four elderly normalsubjects are shown in Table 3 Both GR and COcould do this task without difficulty and their per-formance was near ceiling There was no evidencethat they had any more trouble with the emptyrooms or Lego scenes than with the real or Legoobjects Reaction times were slightly longer for GRand CO than for normal subjects as one wouldexpect given their visual field deficits however this
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 495
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 3 Experiment 1 (two-dot) results (SD in parenthesis)
GR CO Normals
Proportion correctObject 095 080 093Lego object 100 095 098Empty room 100 100 099Lego room 095 100 096
Mean RTObject 2435 2932 2161 (980)Lego object 2052 3245 2029 (921)Empty room 2744 2291 1788 (959)Lego room 1904 2281 1630 (1127)
Figure 6 The four kinds of stimuli used in Experiment 1 Topleft lego layouts top right real-world layouts (empty rooms)bottom left lego objects bottom right real-world objects
difference did not appear to be significant Impor-tantly there was little evidence that GR or CO tooklonger to make their perceptual judgements for thereal or Lego scenes than for the real or Lego objectsThus contrary to our initial hypothesis this experi-ment found no clear evidence for selective impair-ment in the perception of scene-like stimuli
Experiment 2 Different-views matchingtask
The previous experiment demonstrated that GRand CO can extract information about the per-ceived distances between themselves and the sur-faces of an object or scene Thus in at least onesense their perception of these surfaces is pre-served However this does not mean that their abil-ity to represent the layout of a scene is unimpairedThe two-dot task requires on-line comparison oftwo surfaces in egocentric coordinates GR and COwould be able to perform this task even if they couldnot (1) represent more than two surfaces at a time(2) represent surfaces in other than strictly egocen-tric coordinates or (3) sustain surface representa-tions for longer than the time it takes to make acomparison It has been suggested that the parietallobes are involved in on-line processing of surfacegeometry in egocentric coordinates (Milner ampGoodale 1995) thus it is possible that the parietallobes (which are intact in GR and CO) may be suf-ficient to perform the two-dot task even in theabsence of more durable or complex surface repre-sentations in parahippocampal cortex
In Experiment 2 we examine GR and COrsquosability to create and sustain a representation of thegeometry of an object or scene-like layout The taskwas to view a photograph of a Lego object or Legoscene hold it in memory for 10 s and then deter-mine whether a second object or scene was the sameas the first In ldquosamerdquo trials the second photographdepicted the same scene or object depicted from adifferent viewing angle In ldquodifferentrdquo trials the sec-ond photograph depicted a different scene or objectchosen to be similar to the first one (Figure 7)Thus in order to successfully perform this tasksubjects had to form a representation of the surfacesof the object or scene sustain this representation in
memory for 10 s and then align it (presumablyusing mental rotation) to the second object orscene
MethodThe experiment consisted of two parts each with40 trials In the first part the stimuli were black andwhite photographs of objects made out of Legoblocks In the second part the stimuli were photo-graphs of ldquoscenesrdquo made out of Lego blocks whichwere designed to have a geometric structure similarto real-world places such as a roomor a city street
Subjects began each trial by pressing the spacebar A photograph of a Lego object or sceneappeared on the screen for 3 s followed by a 10-sblank interval A second photograph then appeareddepicting either (1) the same object or scene as thefirst photograph shown from a different viewingangle (offset approximately 35 degrees) or (2) a dif-ferent object or scene Subjects were instructed topress the ldquojrdquo key if the two photographs depicted thesame object or scene and the ldquof rdquo key if they did notThey were told to take as much time as they neededto make a decision but no extra time The secondphotograph remained on the screen until a responsewas made Half the trials were ldquosamerdquo trials and halfwere ldquodifferentrdquo trials
Results and discussionFigure 8 shows results for GR and CO as well as forsix elderly normal controls GRrsquos and COrsquos perfor-mance was comparable to that of the normal con-trols Furthermore GR and CO performed just aswell when the stimuli were Lego scenes as whenthey were Lego objects These results are unlikely toreflect a ceiling effect as performance for bothpatients and controls was around 80 correctThus both patients appear to be able to form a rep-resentation of the geometry of both scenes andobjects which they can sustain in short-termmemory
Experiment 3 N-back recognition memorytask
The previous experiment failed to find any impair-ment in GR and COrsquos ability to form a representa-
EPSTEIN ET AL
496 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
tion of the geometry of a scene or object andmaintain it for 10 s in the absence of the originalstimulus Not only does this result suggest that per-ception of scene-like stimuli is intact in thesepatients it further suggests that some short-termmemory functions are intact as well However we
should note that subjects in this experiment werefree to devote all their effort and attention in the 10-second blank interval to maintaining a memory ofthe stimulus Thus it is possible that GR and COcan use rehearsal mechanisms to maintain the stim-ulus in short-term memory during the blank inter-
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 497
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
(b) Lego object version
Figure 7 Stimuli and procedure for Experiment 2 (a) In the Lego layout version subjects see a Lego layout for 3 s followed by a blankscreen for 10 s A second layout then appears which is either the same layout as the first one but seen from a different angle or a differentlayout (b) The Lego object version is the same but with object stimuli
3 s 10 s OR
3 s 10 s OR
(a) Lego layout version
(b) Lego object version
val but are unable to form a more durable memorytrace for the stimulus This possibility was tested inthe current experiment which was designed toexamine GR and COrsquos ability to learn new visualinformation
We devised a continuous n-back recognitionmemory test in which subjects viewed a sequence oforiginally novel stimuli some of which wererepeated within the sequence at intervals of 1 3 or5 intervening images For each stimulus subjects
reported whether they had seen it before in theexperiment or not In different runs in the experi-ment the stimuli were black-and-white photo-graphs of (1) Lego scenes (2) Lego objects (3) real-world landscapes (4) novel objects made out ofSculpey clay (5) unfamiliar houses (6) unfamiliarfaces as well as (7) line drawings of nonsenseobjects and (8) pronounceable pseudowords (Fig-ure 9) Thus we were able to test GR and COrsquos rec-ognition memory for different kinds of materials atdifferent time intervals in a paradigm in which thepresence of intervening items between the twoappearances of each stimulus made rehearsal diffi-cult Note that because none of the specific stimuliused in this experiment were familiar in advancethis task did not require an episodic memory com-ponent and could be done on the basis of pure visualfamiliarity with the particular visual stimulus
Although many different stimulus classes weretested in this experiment the critical comparison isbetween the Lego layouts and Lego objectsbecause these are best matched with regard tomaterials and differ primarily in their geometricstructure (see Introduction) We predicted that ifGR and CO showed a memory deficit for this taskit would be greater for the Lego layouts than for the
EPSTEIN ET AL
498 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 8 Results of Experiment 2 Performance over ldquosamerdquotrials and ldquodifferentrdquo trials is averaged together Error bars show1 SD
Figure 9 Stimuli for Experiment 3 Top row Lego layouts real-world scenes houses Bottom row Lego objects clay objects nonsenseobjects faces In an eighth condition (not shown) subjects viewed pronounceable nonwords such as ldquowilchrdquo or ldquothipperrdquo
Lego objects In addition a deficit for materialsthat convey information about the layout of sur-rounding space might also be apparent when GRand CO are tested on more realistic stimuli If soone would expect them to do poorly on the land-scapes but relatively well on the clay objects linedrawings of nonsense objects faces andpseudowords
MethodFor each subject a complete experimental sessionconsisted of being run once on each of the eightstimulus classes Each run consisted of 90 trials ineach trial a stimulus was presented and subjectsreported whether or not they had seen it before inthe run (or indeed ever) by pressing the ldquojrdquo key ifthey had or the ldquof rdquo key if they had not Each stimu-lus was on the screen for either 1 2 or 3 s (see fol-lowing) and appeared promptly after the keypressreport for the previous trial Individual stimuliappeared either once (14 stimuli) twice (30 stim-uli) or four times (4 stimuli) within a given run Ofthe stimuli that were presented twice 10 wererepeated immediately (1-back) 10 were repeatedwith a lag of two intervening items (3-back) and 10were repeated with a lag of four intervening items(5-back) In order to ensure that between-subjectdifferences in performance were not due to stimulusdifferences the same stimuli were assigned to thesame conditions each time the experimentwas run
GR performed the experiment twice once inAugust 1998 and again 9 months later Because ofa computer error he was only tested on six of theeight categories (all except Lego layouts and Legoobjects) at the later date CO was also tested twiceonce in February 1999 and again 6 months laterReported results for GR and CO are the average oftheir two testing sessions Normal subjects wereonly tested once Stimulus presentation time was 1 sin GRrsquos first testing session In pilot testing COfound this too fast for the critical categories of Lego
layouts and Lego objects so presentation time wasextended to 2 s for these categories in his first test-ing session All normal subjects were tested at thesepresentation rates (2 s for Lego layouts and objects1 s for other stimuli) In COrsquos second testingsession presentation time was lengthened to 3 s forall stimulus categories to ensure that any deficitsfound were memory deficits rather than problemswith the nonmemory aspects of the task
Results and discussionResults from GR CO and 10 elderly normal con-trols are shown in Figure 10 and Table 4 Figure 10ashows results from the main comparison betweenLego layouts and Lego objects Both normal sub-jects and patients performed near ceiling for bothLego layouts and Lego objects at 1-backinterstimulus intervals verifying that GR and COcould successfully perceive the pictures as they werebeing presented In contrast there was a strikingdifference between the two patients and the normalsubjects at 5-back intervals Whereas normals didon average somewhat better on Lego layouts thanLego objects both GR and CO showed the oppo-site pattern doing much worse on Lego layoutsthan Lego objects The 3-back condition showed amixed pattern Whereas GR did better on Legoobjects than Lego layouts in this condition (similarto his performance at 5-back intervals) CO didbetter on Lego layouts than Lego objects COrsquos lowperformance on Lego objects in the 3-back condi-tion appears to be an anomaly as it is much lowerthan his performance for comparable stimuli in the1-back and 5-back conditions
Figure 10b shows the difference in performancebetween Lego layouts and Lego objects for thecritical 1-back and 5-back intervals At 5-backintervals CO was significantly impaired comparedto the normal subjects (z = ndash250) whereasGRrsquos impairment fell just short of significance (z =ndash175)4
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 499
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
4In fact these values may underestimate GR and COrsquos relative impairment for Lego layouts In the 5-back condition 4 of the 10
normal subjects had perfect performance on the Lego layouts but all scored below ceiling on the Lego objects Thus the true advantagefor Lego layouts over Lego objects for normal subjects is probably greater than that observed here in which case GR and COrsquos lowerperformance for Lego layouts compared to Lego objects would be even more abnormal Furthermore post-hoc analyses revealed thatone of the normal subjects showed a difference in performance between Lego layouts and objects that was over 2 SD below the mean ofthe other nine subjects When this outlying subject was excluded z-scores were ndash322 for CO and ndash232 for GR
Table 4 shows results for all eight stimulusclasses Although direct comparisons between sub-jects should be treated with caution since viewingtime for different items was not the same for eachsubject we do note a few interesting patterns Firstof all there is little or no evidence that GR or CO
show a memory deficit for nonsense words clayobjects or line drawings of nonsense objects GRappears to be slightly impaired on faces and housesbut CO performs normally on these categoriesStrikingly GR shows a severe deficit for the reallandscapes performing more than 40 percentagepoints lower than normals Thus even thoughthere is some evidence that GR might have someimpairment for nonscene stimuli he clearly is muchmore impaired for scene-like stimuli such as theLego scenes and landscapes CO does not show asimilar deficit for the landscapes However itshould be kept in mind that in one of his two testingsessions CO viewed each picture for 3 s which wasmuch longer than the 1 second used for all non-Lego stimuli for GR and normals Thus COrsquosapparently ldquonormalrdquo performance in these condi-tions might be overestimated
Taken together the results of this experimentindicate that GR and CO are selectively impaired atprocessing scene-like stimuli This deficit is consis-tently apparent at 5-back but not 1-back intervalsThis finding suggests that the deficit should becharacterised as a memory encoding problemHowever the present results do not allow us todetermine whether the difficulty occurs at memoryencoding or at subsequent recognition (see Epsteinamp Kanwisher 2001) If GR and CO have a generalproblem with recognition of topographical materi-als then they should not be able to recognise scenes
EPSTEIN ET AL
500 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 10a Results of Experiment 3 for GR (top) CO (middle)and for 10 age-matched normal controls Performance measure isproportion of repeated stimuli correctly identified as such correctedfor guessing using the standard formula
Figure 10b Difference between Lego layout performance andLego object performance for the 1-back and 5-back conditionsError bars indicate one 1 SD
and landmarks that were learned prior to theirinjury We tested this possibility in the nextexperiment
Experiment 4 Famous landmarksfacesrecognition task
In this experiment we examined GR and COrsquosability to recognise the visual appearances of placeslearned prior to injury Subjects saw a series offamous landmarks intermixed with nonfamouscontrols (Imaging experiments have demonstratedthat landmarks of this type can activate the PPAalmost as strongly as complete scenes Epstein etal 1999 Epstein amp Kanwisher 1998) They wererequired to say whether they knew each specificlandmark and to identify the ones they knew Sub-jects were instructed to respond affirmatively if theywere fairly certain that they recognised the land-mark but not if it only looked generally familiarFor purposes of comparison subjects also per-formed an identical test in which the stimuli werefamous and nonfamous faces In both testsdistractor items were designed to be visually andconceptually similar to the target items (see Figure11) If parahippocampal cortex is required for placerecognition then one might expect GR and CO toperform poorly compared to normals on the land-mark recognition test
MethodEach of the two experiments (famous faces famouslandmarks) consisted of 48 trials Subjects beganeach trial by pressing the space bar after which ablack-and-white photograph of a face (in thefamous faces experiment) or a landmark (in thefamous landmarks experiment) appeared on thescreen Subjects were told to press the ldquojrdquo key if theyrecognised the specific face or landmark and the ldquof rdquokey if they did not taking as much time at theyneeded to make this discrimination but no extratime Subjects were told to report that they recog-nised a face or landmark if they felt that they knewit even if they could not retrieve its name If theydid know the name they were to say it after pressingthe button Otherwise they were to say whateverthey did remember about the item
Half of the photographs were of famous and halfof nonfamous people and landmarks The famousfaces included public figures (eg George BushRichard Nixon Prince Charles) and movie stars(eg Harrison Ford Clark Gable Marlon Brando)who were prominent prior to the time of GR andCOrsquos injuries the famous landmarks were build-ings and monuments from all over the world (egEiffel Tower US Capitol Building Great Sphinx)The nonfamous stimuli were chosen to match thefamous stimuli as closely as possible Each famousstimulus had a ldquomatchrdquo For example George Bushwas matched to a man of similar age in a coat andtie and the Taj Mahal was matched to a visually
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 501
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 4 Full results of Experiment 3 Mean proportion correct (corrected for guessing) SDs in parentheses
Lego Land- Lego Clay Nonsense Non-scenes scapes Houses objects objects objects Words Faces
1-back GR 100 094 100 100 100 094 100 100CO 091 085 091 100 100 079 074 100Normals 096(008) 095(008) 093(014) 096(008) 096(006) 099(003) 098(004) 097(011)
3-back GR ndash003 068 026 047 083 083 075 052CO 066 081 037 010 094 069 095 081Normals 066(032) 077(021) 072(015) 077(020) 086(016) 083(015) 083(017) 072(020)
5-back GR 010 034 044 047 072 083 094 053CO 031 073 075 088 081 064 100 081Normals 076(026) 077(016) 068(021) 066(016) 091(014) 083(018) 093(011) 078(021)
similar Asian palace that is not famous (see Figure11)
Results and discussionResults for GR and CO as well as for five age-matched controls are shown in Figure 12 Both
GR and CO could recognise the famous landmarksas well normals In addition both GR and CO wereusually able to name the landmarks they success-fully identified
Thus GR and CO can recognize landmarkslearned prior to injury demonstrating that at leastsome of their place recognition abilities are intactWhen combined with the fact that GR and COwere able to produce detailed maps of places theylived in prior to their injury (see Case Description)these results suggest that their deficit is primarily inencoding or consolidation of new informationrather than in recognition or recall of old informa-tion Note however that these results do not provethat their place recognition abilities are entirelynormal GR and CO might suffer from a mild rec-ognition deficit that affects identification of newly-learned stimuli like the Lego layouts in Experiment3 but not overlearned stimuli like the famous land-marks Furthermore it is possible that GR and COmight be impaired at recognising premorbidlylearned places that are defined primarily by theirspatial layout even though they are unimpaired atrecognising premorbidly learned places that aredefined primarily by a single object-like landmarkor building (see General Discussion) Neverthelessthe trend of the present evidence is to suggest thatGR and CO have particular problems with noveltopographical information
In the famous-face task GRrsquos performance wasin the normal range However he sometimes haddifficulty naming the faces even though he clearly
EPSTEIN ET AL
502 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 11 Stimuli for Experiment 4 Each famous landmark(Arc de Triomphe Taj Mahal) and famous face (Barbara BushYasser Arafat) was matched to a nonfamous building or face thatalso appeared in the experiment
FA
CE
SL
AN
DM
AR
KS
Figure 12 Results of Experiment 4 Performance measure isproportion of famous stimuli reported as ldquoknownrdquo corrected forguessing using the standard formula
recognised them For example he described Ron-ald Reagan as ldquothe gipperrdquo Nancy Reagan as ldquothewife of the gipperrdquo and Clark Gable as ldquothe movieactor who said lsquoScarlett I donrsquot give a damnrsquordquo COhad a great deal of difficulty identifying the famousfaces This result is not surprising given hisextremely low performance on the Benton FaceMatching Task In addition to his topographicproblems CO appears to suffer fromprosopagnosia
GENERAL DISCUSSION
We tested two parahippocampal patients (GR andCO) on four experiments designed to measure per-formance as a function of both task (perceptionencoding and recognition) and stimulus type(scenes objects faces and words) The only taskthat revealed a clear deficit was an n-back recogni-tion memory test (Experiment 3) In order to suc-cessfully perform this task subjects had to encode adurable representation of a visual stimulus and useit for later recognition GR and CO were signifi-cantly more impaired on this task when the stimuliwere scene-like spatial layouts (ie Lego scenesand real scenes for GR) than when they wereobjects No comparable deficits were found in otherexperiments that tested layout perception (Experi-ments 1 and 2) and placelandmark recognition(Experiment 4) Taken together these results indi-cate that GR and CO are impaired in their ability toencode novel information about the geometry ofsurrounding space into memory but are lessimpaired at their ability to encode the geometry ofnovel objects
We argue that this specific topographical defi-citmdashinability to encode the spatial layout of novelscenesmdashresults from damage to the para-hippocampal place area Although the damage inboth patients extends beyond the parahippo-campal-lingual territory normally associated withthe PPA this region is clearly lesioned bilaterally inGR and in the right hemisphere in CO Further-more functional imaging demonstrated that GRhas no functioning PPA Thus the current resultsand the earlier fMRI work (Epstein et al 1999
Epstein amp Kanwisher 1998) converge to implicateparahippocampal cortex in the encoding of infor-mation about scene geometry into memory
Our assignment of the function of memoryencoding to parahippocampal cortex is consistentwith the results of earlier case reports of patientswith parahippocampal damage who have naviga-tional difficulties in novel but not familiar environ-ments (Habib amp Sirigu 1987 Ross 1980 see alsoTeng amp Squire 1999) As with GR and CO thesepatients were either less impaired or unimpaired onnontopographical memory tasks Patients have alsobeen described who exhibit the opposite pattern ofimpairment an inability to learn faces (sometimescalled ldquoprosopamnesiardquo Cipolotti Robinson Blairamp Frith 1999 Tippett Miller amp Farah 2000) orfaces and verbal materials (Maguire amp Cipolotti1998) with an unimpaired ability to learn topo-graphical materials Taken together these resultsargue for a high degree of domain-specificity in theneural systems involved in learning of new informa-tion (Gallistel 1995) The brain may have separatemechanisms for learning faces places and wordswhich can be differentially impaired after damageto specific cortical regions (Cipolotti et al 1999Incisa della Rochetta et al 1996) The presentresults suggest that the anatomical locus of themechanism dedicated to learning new topographi-cal information is in the parahippocampal cortex(Aguirre amp DrsquoEsposito 1999)
In contrast to their impaired ability to encodenew topographical information both GR and COwere able to produce detailed maps of their previousdwellings and performed normally on the famouslandmark recognition test of Experiment 4 Theseresults are generally consistent with Teng andSquirersquos (1999) report of a patient with extensivemedial temporal damage who could recallpremorbidly learned but not postmorbidly learnedtopographical information (see also Ross 1980)Thus our results provide further evidence thatmedial temporal regions such as parahippocampalcortex may be more critical for encoding or consoli-dating new information into memory than for recallof old information
One issue that our experiments do not entirelyresolve is the precise nature of the topographical
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 503
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
information encoded by parahippocampal cortexClearly this region is critical for encoding informa-tion about the geometry of the currently visiblescene even when the ldquoscenerdquo is made out of Legoblocks and therefore has no immediate navigationalrelevance However it is unclear whether this spe-cific deficit can account for all of the real-worldnavigational difficulties encountered by GR andCO GR in particular shows such a catastrophicloss of navigational ability that we suspect he is alsoimpaired at encoding the spatial relationshipsbetweenthe current scene and other locations in theworld CO probably also suffers from such difficul-ties as he reports that he has trouble rememberinghow places relate to each other in large-scale spaceWhen he was tested on a simple 3-room videogameenvironment he not only was unable to rememberthe individual rooms (claiming that there were 10rooms when there were only 3) but he also had nomemory for the spatial relationships between themThese spatial deficits may be the result of damage tothe posterior right hippocampus (in the case of CO)or of disconnection of the posterior right hippo-campus from the parahippocampal regions thatprovide much of its cortical input (in both patientsSuzuki amp Amaral 1994) In either case the abilityto form a cognitive map of new environments likethe lab or videogame space could be impaired(Maguire et al 1998 OrsquoKeefe amp Nadel 1978)However it is also possible that parahippocampalcortex itself may be critical for encoding not onlythe geometry of individual scenes but also the spa-tial relationships between the locations of thosescenes In this case the spatial deficits of GR andCO could be attributed directly to parahippo-campal damage rather than to hippocampal dam-age or disconnection
The latter scenario would be consistent withAguirre and DrsquoEspositorsquos (1999) and Barrash etalrsquos (2000) suggestion that parahippocampal cortexis critically involved in combining visual informa-tion about the appearance of scenes and landmarkswith spatial information about their locations into aunified topographical representation Howeverboth of these authors argue that the representationsof individual scenes (Barrash et al) and landmarks(Aguirre amp DrsquoEsposito) are supported not by
parahippocampal cortex but by more posterior lin-gual-fusiform regions In contrast we believe thatthe neuroimaging data indicate that parahippo-campal cortex is involved in the encoding of thegeometric structure of individual scenes althoughit may also play a role in combining these represen-tations with other spatial representations
Interestingly GR and CO were not impaired onrecognition of famous landmarks that were mostlybuildings Thus our data do not preclude the possi-bility that there may be a separate lingual orfusiform region involved in processing informationabout buildings (Aguirre Zarahn amp DrsquoEsposito1998 Chao Haxby amp Martin 1999 IshaiUngerleider Martin Schouten amp Haxby 1999)which might have been preserved in GR and COdespite extensive damage to other regions of thelingual and fusiform gyrus Alternatively GR andCOrsquos preserved building recognition abilities mightbe mediated by interactions between cortical areasthat perform shape analysis for any object-likestimulus type (Malach et al 1995Kanwisher et al1996) and anterior temporal regions in which long-term topographical and semantic memories arestored (McCarthy et al 1996)
Is perception preserved
In contrast to their inability to encode new scenesinto memory GR and CO have substantiallyretained their ability to perceive the spatial layout ofthe currently visible scene (Experiments 1 and 2)GR can follow a route marked on a floor planwhich indicates a good on-line appreciation of hiscurrent spatial surroundings Furthermore neitherGR or CO have any trouble moving through theenvironmentmdashthey move with confidence and donot bump into doors or walls Problems of the lattertype are usually associated with damage to the pari-etal lobes which are intact in GR and CO Thus atfirst glance GR and CO do not appear to haveperceptual impairments (beyond their visual fieldrestrictions)
However GR does complain of a perceptualdeficit which he describes as an inability to ldquotakeinrdquo scenes when they are too complex Similarreports of an inability to comprehend the
EPSTEIN ET AL
504 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
ldquoglobalityrdquo of a scene were made by some of theparahippocampally-damaged patients studied byHabib and Sirigu (1987) We were not able todetermine the precise nature of this deficit whichmay be partly attributable to his visual field restric-tions It appears that his deficit is more salient in thereal-world environments than in the picture-per-ception tasks of Experiments 1 and 2 perhapsbecause he finds it more difficult to organise visualinput when immersed in it than when viewing pho-tographs that subtend only part of the visual field
We suggest that GR can perceive parts of thescene when he attends to them but may be unableto integrate information over time into a coherentrepresentation of a scene or place The fact that GRand CO are impaired for Lego layouts at 5-backintervals in Experiment 3 is consistent with thishypothesis The time between the first and secondpresentations of a stimulus is very short in this case(and even shorter with 3-back intervals where GRalso shows evidence of a deficit) In both patientsthe representation of layout information formed byperception of a Lego scene must either have anabnormally short persistence be abnormally sus-ceptible to interference or both If so they mightfind it impossible to sustain a spatial frame orschema long enough to use it to organise informa-tion obtained from many different glances at a com-plex scene (Intraub 1997) The result would be aninability to understand the ldquoglobalityrdquo of a sceneeven though all the details were perfectlyperceived
Non-topographical mnemonic abilities
Although not as severe as their topographical mem-ory problems GR and CO do have some memorydifficulties in other domains For example bothperformed poorly on the Rey AVLT whichrequired them to remember a list of unrelatedwords Both also report some subtle episodic mem-ory problems (see Case Description) COrsquos prob-lems could partly result from damage to the righthippocampusmdashindeed CO appeared to have themore acute episodic memory problems HoweverGR has no hippocampal damage so the fact that hedisplays episodic memory problems suggests thatparahippocampal cortex is involved in the forma-
tion of episodic memories (cf Wagner et al1998)
In previous work (Epstein et al 1999) weargued that spatial context information encoded byparahippocampal cortex might play a role in estab-lishing event memories by encoding informationabout where events take place The inability toencode this information might cause problems intasks such as the Rey AVLT where one has to recalla list of unrelated words If spatial (and perhapstemporal) context information is not encoded itwill be harder subsequently to recall the distinctevent of hearing each word (Tulving 1983) Incontrast lack of spatial context information wouldbe less of a problem in tasks where one is onlyrequired to recognise previously encountered verbalmaterials but not to reproduce the spatiotemporalcontext in which they were experienced IndeedGR and CO performed as well as normals on thenonsense words in Experiment 4 where recogni-tion memory but not episodic memory was requiredfor successful performance Interestingly GR andCO also did relatively well on the Logical Memorysubtest of the WMS-III perhaps because the nar-rative framework of a story provides sufficient con-textual information to overcome the absence ofspatial context and mediate successful verbalretrieval
Differences between GR and CO
Although we have discussed GR and CO togetherthere are a number of relevant differences betweenthem One open question is whether these differ-ences can be attributed to the different effects ofunilateral and bilateral parahippocampal damageIn general GRrsquos topographical difficulties weremore severe than COrsquos CO did surprisingly well onthe Warrington Topographical Memory Test inwhich he was required to remember detailed real-world city scenes and also performed substantiallybetter on real scenes than Lego scenes in Experi-ment 3 Insofar as CO has intact left para-hippocampal cortex this suggests that the left PPAmight be sufficient to encode a certain amount of(perhaps semantic) information about real-worldscenes but the right PPA is necessary to encode
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 505
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)

brain regions while subjects viewed a wide variety ofvisual stimuli (Epstein amp Kanwisher 1998) Wefound that a region abutting the collateral sulcusnear the parahippocampal-lingual boundaryresponded significantly more strongly when sub-jects viewed navigationally relevant stimuli such asstreet scenes landscapes or buildings than whenthey viewed other kinds of visual stimuli such asfaces objects or scrambled scenes1 This responsewas found even when subjects simply watched thestimuli passively and were not required to performany navigational task We named this region theldquoparahippocampal place areardquo or PPA because itresponded strongly to depictions of places ThePPA appears to overlap with the regions activatedin previous neuroimaging studies of navigation(eg Aguirre et al 1996)
A critical determinant of PPA activationappears to be the presence in the stimulus of infor-mation about the shape or layout of the immediateenvironment The PPA responds no more stronglyto photographs of rooms filled with furniture andobjects than it does to photographs of the samerooms empty of all objects (ie just bare walls) Italso responds much more strongly to ldquoscenesrdquo madeout of Lego blocks (which have a geometric struc-ture similar to a room or street scene) than it does toobjects made out of the same Lego materials(Epstein Harris Stanley amp Kanwisher 1999)Thus the PPA is sensitive to the presence of a spe-cific kind of geometric organisation in the stimuluseven if the stimulus does not depict a real place inthe world In particular it responds more stronglyto depictions of surfaces that in some senseldquoencloserdquo the observer and define a space withinwhich one can act (ie the walls of a room or theside of the distant hill) than to depictions of surfacesthat define objects that the observer can act uponThese results are generally consistent with behav-ioural results from rats (Cheng 1986 Gallistel1990 Margules amp Gallistel 1988) and human
infants (Hermer amp Spelke 1994 1996 Hermer-Vazquez Spelke amp Katsnelson 1999) that suggestthat information about the geometry of surround-ing spacemdashthe ldquolay of the landrdquomdashplays a privilegedrole in orientation and navigation
What specific cognitive task does the PPA per-form Given the fact that landmark agnosia oftenresults from parahippocampal-lingual damage(Landis et al 1986) one might hypothesise thatthe primary role of the PPA is place recognitionidentification of familiar locations based on theirappearance andor geometric structure If so onemight expect the PPA to be sensitive to the noveltyfamiliarity of the place depicted in the stimulusHowever we found that the PPA responds just asstrongly to photographs of unfamiliar places (ieplaces the subjects had never visited) as it does tophotographs of familiar places (Epstein et al 1999Expt 1) In contrast the PPA is sensitive to thenoveltyfamiliarity of the stimulus itself respond-ing more strongly to new photographs than to pho-tographs viewed many times previously even whenall the photographs depict unfamiliar places(Epstein et al 1999 Expt 4) From these resultswe inferred that the PPA is more involved inencoding a representation of the spatial structure ofthe current scene than in relating that scene to onersquosstored cognitive map of the environment as awhole However our data did not allow us to deter-mine whether the PPA activation reflects percep-tual encoding memory encoding or both
The purpose of the present report was to moreprecisely determine the function of the PPA Tothis end we tested the ability of our two patients toperceive learn and recognise a variety of visualmaterials We predicted that the patients would bemore impaired at processing scene-like stimuli (iestimuli that depict spaces that one can navigatethrough or act within) than object-like stimuli Byusing several different kinds of tests we aimed todetermine whether any such selective impairment
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 483
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
1Although full scenes activate the PPA more strongly than do any other kind of visual stimulus we have also observed higher
activity in the PPA when subjects view buildings than when they view other kinds of objects (Epstein et al 1999) consistent withAguirre Zarahn and DrsquoEspositorsquos (1998) finding of a significant response to buildings in an adjoining and possibly overlapping regionof the anterior lingual gyrus
was in the perceptual recognition or memorydomain To anticipate we did find evidence for adeficit for visual materials that conveyed informa-tion about the shape of surrounding space Thisdeficit manifested itself during a memory encodingtest but not during perceptual tests or a recognitiontest Thus our data suggest that the PPA is neces-sary for the encoding of spatial layout informationinto memory but may not be necessary for manyaspects of the initial perceptual analysis recogni-tion or recall of this information We argue that thePPA may be a learning mechanism specifically ded-icated to encoding topographical materials intomemory
CASE DESCRIPTIONS ANDNEUROPSYCHOLOGICAL TESTING
In this section we present the case histories of thetwo patients in order to provide a general contextfor the more specific experimental tests reportedin the Experimental Investigations section Thepatients were chosen for investigation because clin-ical MRIs indicated that they had suffered damageto the parahippocampal region This section is
divided into four subsections First we present ageneral overview of the case histories Second wereport the results of some ad hoc tests that give ageneral indication of the patientrsquos navigational abil-ities Third we discuss other cognitive abilitiesincluding face recognition and episodic memoryFourth we report the results of a battery of formalneuropsychological tests performed on the patientsTable 1 summarises relevant observed and self-reported behaviour and Table 2 summarises theresults of the neuropsychological tests
Overview
Case 1 GRGR2 is a right-handed male with 18 years of educa-tion He was 60 years old at the time of testingTwo-and-a-half years prior to testing he suffered aright occipital-temporal stroke during cardiacsurgery A second stroke affecting the left occipital-temporal region occurred a week later Thesestrokes left him with a number of deficits includinglimitation of his peripheral vision (lefthemianopsia right upper quadrantanopsia)dyschromatopsia (details reported in Beauchampet al 2000) and topographical disorientation We
484 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
EPSTEIN ET AL
Table 1 Summary of observed and self-reported behaviour
GR CO
Real-world topographical Unable to recognise 714 newly learned MIT landmarks Unable to learn 3-room videogamelearning despite extensive training regime environment
Unable to find way back to lab unassisted despite Able to learn a simple route out ofextended experience with environment building after many visits
Map drawing Could not draw map of lab maps of other newly learned Map of lab was sketchy but not entirelyenvironments were inaccurate inaccurate
Drew accurate map of premorbid environment Drew accurate map of premorbid(former home) environment (currentformer home)
Scene perception Reports difficulty understanding the globality of complex Does not report a perceptual deficitscenes
Face recognition Some prosopagnosia Severe prosopagnosia
Episodic memory Largely intact though some difficulties were reported Moderate difficulties were reported
Can follow the plot of a movie or novel and report Reports some difficulty following theprevious dayrsquos activities plot of a book Accurately reported
contents of previous dayrsquos newspaper
2GRrsquos true initials are KG (cf Beauchamp Haxby Rosen amp DeYoe 2000)and COrsquos true initials are KC We use GR and CO
here to avoid confusion between the two patients
tested him over a period of 5 days in August 1998with 3 days of follow-up testing 3 weeks later and 1day of additional tests in May 1999 During thistime he was alert and cooperative Emotionalaffect and language use was normal He interactednormally in nonclinical social situations and hisconversation was appropriate and varied Heremembered the events of previous days andrecounted them accurately
In contrast to this high level of non-topographical functioning GR demonstrated adramatic inability to orient himself in space Forexample even after several days of testing he neverlearned the relative locations of the different roomsof the lab According to both GR and his wife thisinability to learn new topographical informationwas typical of his experience since his strokes as hefrequently gets lost in his daily life Soon after his
injury he moved to a neighbourhood with manysimilar-looking houses He reported that in orderto find his new house after a walk to a market six toseven blocks away he had to rely on street signs toguide him to the correct block and then examineeach house on the block in detail until he couldrecognise some feature that distinguished his homeuniquely Subsequently GR and his wife moved toa different house in a different country He reportedthat for the first 6 months after the move his newhome was like a ldquohaunted houserdquo for him insofar ashe was unable to learn his way around it For exam-ple he would repeatedly forget that the bedroomhad a balcony and he would be surprised by this factevery time he rediscovered it After several monthshowever he was able to learn the topography of thehouse sufficiently well to navigate through it (butsee below)
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 485
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 2 Summary of neuropsychological tests ( indicates percentile scores)
GR COmdashmdashmdashmdashmdashmdashmdashmdashmdashmdash mdashmdashmdashmdashmdashmdashmdashmdashmdashmdashmdashndash
IQ 122 (93 WAIS-R) 134 (99 WAIS-III)
WMS-III
Logical memory 25 immediate recall 27 immediate recall50 delayed recall 48 delayed recall(WMS-R)
Face memory 9 immediate recall 16 immediate recall16 delayed recall 9 delayed recall
Family pictures 2 immediate recall lt1 immediate recall5 delayed recall lt1 delayed recall
Rey Auditory Verbal 63 immediate recall 45 immediate recallLearning test 14 after interference list lt1 after interference list
16 delayed recall 3 delayed recall
Warrington Recognition 50 wordsMemory tests 29 scenes
lt5 faces
724 Spatial Recall 66 immediate recall2 delayed recall
Benton Face Recognition 42 lt1
Memory span Digits99 forward99 backward
Spatial37 forward50 backward
Boston Naming test Ceiling Ceiling
Case 2 COCO is a left-handed male with 20 years of educa-tion He was 60 years old at the time of testing Twoyears prior to testing he suffered a right posteriorcerebral artery stroke He first noticed on a week-end that his vision appeared blurred on the left andthat he had a tendency to bump into objects on thisside On Monday morning he was able to success-fully negotiate the route to his workplace a trip thatinvolved a bus ride followed by two subways andthen a shuttle to his office building However oncehe entered the building he found himself com-pletely disoriented unsure of its layout and wherehis office was located Interestingly the companyhe worked for had moved a few weeks earlierAlthough he had managed to arrive at this newbuilding (which was a block away from the previouslocation) he became lost upon entering it Hisneurologic exam revealed a left homonymoushemianopia but no evidence of neglect on a lettercancellation task or on sensory testing Investiga-tions revealed an ischaemic stroke with haemor-rhagic transformation in the right occipital andmesial temporal lobe On magnetic resonanceangiography he had a chronic left carotid occlusiona mild right carotid stenosis and bilateral vertebralartery stenoses Since then he has suffered fromsevere topographical disorientation primarily innew environments
CO reports that he can recognise buildings thatwere familiar to him before the stroke He can alsoform mental images of well-known landmarks suchas the Empire State Building and the BostonPublic Gardens and of the house he lived in as achild When asked to imagine standing in front ofthe Massachusetts State House (a location wellknown to him from years of living in Boston) heaccurately described the surrounding buildings andtheir locations However he reports that when helooks at a place that hersquos encountered post-strokeall he gets is a feeling of familiarity that does notallow him to determine where the place is or whenhe saw it He also reports an inability to understandhow various landmarks relate to each other inlarge-scale space Despite this difficulty he canfollow a map and frequently rides the Bostonsubway alone
Unlike GR CO was able to learn a certainamount of topographical information For exam-ple he soon learned the location of the rooms in thelab and would travel between them unassistedAfter his first visit to the lab he needed to be guidedall the way back to the subway station but after sev-eral visits he could find his way out of the buildingand to the subway on his own as long as he was firstguided to the elevators (It should be noted how-ever that the building is on the same block as thesubway station Furthermore he still needed to bedirected to the elevators even though our buildinghas a relatively simple floor plan) Thus the amountof topographical information he can learn is limitedbut not zero
COrsquos language and reasoning skills appear to benormal and his conversation is appropriate andvaried In addition to his topographical difficultieshe also has limitations of his peripheral vision onthe left side
Navigational abilities
In addition to the experiments reported in theExperimental Investigations section of this paperwe also performed a number of ad hoc tests of GRand COrsquos navigational abilities Although thesetests were informal we describe them here to pro-vide a general sketch of the nature of GR and COrsquosimpairments
Real-world topographical learningOver the course of several days we took GR and hiswife on three walks around the MIT campus whichneither had visited previously Each walk followedexactly the same path At several places in the walkwe would stop and point out different buildings andlandmarks tell them something about each one andask them to remember their appearance for latertesting Immediately after the completion of thethird walk both GR and his wife separately per-formed a forced-choice recognition test in whichthey saw photographs of 14 landmarks (mostlybuildings) from the walk and 14 novel landmarks(from another part of the MIT campus that theyhad not visited) and were required to reportwhether they had seen each one or not GRrsquos
EPSTEIN ET AL
486 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
performance was 714 correct hits for the familiarlandmarks and 1314 correct rejections for theunfamiliar landmarks In contrast his wifeobtained a perfect score on this test Given theextensive nature of the training regime GRrsquos scorestrongly suggests an impairment in learning theappearance of new real-world places Interestinglyseveral of the seven familiar landmarks that he gotright had an obvious distinctive feature that hadbeen explicitly pointed out to him For exampleone building had a distinctive clock tower andanother one was an unusual triangular shape ThusGR was occasionally able to use distinct visualfeatures to identify places even though he wasunable to recognise them holisticallymdasha commonpattern among topographical disorientationpatients
GR did not appear to have any obvious problemsin understanding the spatial structure of his imme-diate environment When given a floor plan of thebuilding at MIT in which he was being tested hewas able to successfully follow a route marked onthe plan However when the plan was taken awayhe had great difficulty finding his way back to thelab even though he had more than 30 min experi-ence with the environment and his starting positionwas less than 30 feet from the goal (Figure 2) Infact the route he took resembled a random walk Atone point he walked right past his destinationwithout recognising it and only knew for sure thathe was in the right place when he identified his wifein the room Thus his on-line appreciation of thelayout of immediately visible space appears to beintact but he cannot find his own way withoutassistance
We were unable to perform the MIT landmarkrecognition test on CO because he suffered from amedical problem that made it difficult for him towalk around campus In lieu of this we tried to seewhether he could learn a simple video game envi-ronment consisting of three connected roomsAfter about 30 min experience navigating throughthis environment (via button presses) he was askedto sketch a map of it He declined to do so statingthat it was beyond his abilities When asked howmany rooms there were he replied that he did notknow but that there were at least 10 Given the
simplicity of this artificial environment COrsquos com-plete inability to learn it is striking
Map drawingAfter several days of testing we asked both GR andhis wife to sit at different tables and independentlysketch (1) the shape of Florida (2) a bicycle (3) afloor plan of their hotel room in Boston (4) a floorplan of the first floor of the house they had been liv-ing in for the last 8 months (all post-stroke) (5) afloor plan of the main room of the lab and (6) afloor plan of the house they lived in 10 years ago(pre-stroke) GR drew the map of Florida and thebicycle perfectly His plan of the hotel room wasnotably different from the one drawn by his wifemdashfor example he reported that there was only onebed when in fact there were two and he placed thebed on the wrong side of the room (Figure 1a) Hisplan of the first floor of his current house was some-what better but was not very detailed and includedsome major errors For example he wronglyreported that there was no bathroom on this floor(Figure 1b) He was unable to produce any floorplan of the main room of the lab leaving this pageblank When his wife reminded him that this wasthe room in which he had just spent an hour readingthe newspaper and in which we had all had break-fast in that morning he replied that he didnrsquot knowwhat room she was talking about (although heappeared to remember these events) In contrast tothis near total inability to produce accurate maps ofnew environments GR produced an accurate mapof the house he lived in prior to his injury (Figure1c) He reported that his memory of this house wasldquoall thererdquo and he could imagine walking around init When asked to produce a perspective of a roomwhen he was in it he did so with ease and he wasalso able to create an accurate floor plan of theroom
In contrast CO found it very difficult to pro-duce a floor plan of the lab both when he was in itand when he was taken to another room and askedto recall it However although the maps he pro-duced were quite impoverished they were notentirely inaccurate demonstrating some ability toencode topographical information This contrastswith GRrsquos absolute inability to recall anything
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 487
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
EPSTEIN ET AL
488 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure
1F
loorplansdraw
nby
GR
andhisw
ifeof(a)
theirhotelroomin
Boston(b)theircurrenthousew
hichthey
hadm
ovedto
subsequenttoG
Rrsquosinjury(c)the
housethey
livedin
priorto
GR
rsquosinjury
Ho
telro
om
inB
osto
n
GR
His
wife
His
wife
Cu
rren
th
om
e
GR
His
wife
Fo
rme
rh
om
e
GR
about the main room of the lab once he had left itCO was also able to draw a detailed floor plan of theapartment he was living in at the time (and hadlived in for 20 years) Several months after he hadmoved from this apartment we asked him to drawsuch a floor plan again and he produced one thatwas substantially the same as the first (Figure 3)Thus their map drawing performance indicatesthat GR and CO are primarily impaired at sketch-ing places experienced after their strokes but areunimpaired at sketching places experienced beforetheir strokes This pattern is indicative of deficits inencoding but not recall
Scene perceptionAs noted earlier GR can navigate with the aid of afloor plan suggesting that his perceptual abilitiesare largely intact Interestingly however he reportsthat he does have a perceptual deficit which hedescribes as an inability to understand scenes whenthey are too complex For example he reports thathe often has trouble using maps because theyusually are too busy with details It is unclearwhether this apparent perceptual problem is a resultof his field cut some other separate perceptual
problem or a memory problem (see discussion)CO did not complain of a similar deficit GR wasan accomplished artist prior to his injury producingpaintings of striking complexity Tragically hereports that he cannot now appreciate his ownpaintings
Other cognitive abilities
Face recognitionGR also reported some difficulties with face recog-nition On his second visit we examined his abilityto learn new faces by having him perform a forced-choice recognition test on seven familiar and sevenunfamiliar faces The familiar faces were personnelfrom the lab with whom he had at that point had agood deal of experience spanning at least 3 daysAll the people in the photographs had a towelwrapped around their head in order to minimise theuse of hair as a nonfacial cue Despite the difficultyof this task he correctly identified 67 familiarfaces and made no false alarms Surprisingly theone face that he did miss was the first author of thispaper with whom he at that point had had consid-erable experience GR also performed normally ona difficult famous face recognition test (see Experi-ment 4 following) and on the Benton Face Recog-nition test (see following) Thus he has someability to recognise faces Consistent with this GRreports that he can remember a small number offaces but that he quickly ldquoloses trackrdquo if he has toremember too many
CO also appears to suffer from someprosopagnosia as evidenced by his low perfor-mance on the Benton Face Recognition test (seefollowing see also Experiment 4) Interestingly hedoes not complain of face recognition difficultiesbut rather attributes his inability to recognise peo-ple to a lack of interest in their appearance
Episodic memoryAt first glance GRrsquos episodic memory appeared tobe largely intact As mentioned he could recall theevents of the previous day accurately Furthermorehe reports that he can follow the plot of a moviewithout any difficulty However both he and hiswife report that he does have some problems with
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 489
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Figure 2 Floor plan of the building at MIT in which GR wastested Corridors are highlighted in grey GR was asked to find themain room of the lab (marked G on the map) Starting location(marked S) was a point approximately 30 feet down the hall GRrsquosroute is marked by arrows GR found this task very difficult andonly knew for certain that he had reached his goal when he caughtsight of his wife through the glass door of the lab In contrast hewas able to follow a route marked on a plan of the same floorwithout any problem or hesitation
episodic memory For example he stated that heremembered very little about a trip to Italy they hadtaken several months previously His wife reportsthat he has great difficulty keeping track of timeFor example before their trip to Boston he repeat-edly asked her what day they were supposed to leaveand seemed unable to encode this information Weobserved a similar phenomenon during testingDuring the tests that required repetition of thesame procedure several times he sometimesseemed unable to clearly remember how manytimes he had repeated the procedure Thus he doesappear to have a subtle episodic memory deficit
which may manifest itself as an inability to sepa-rately remember successive events that are not easilydistinguishable from each other
CO also reports some problems with his epi-sodic memory He relies on lists to rememberappointments stating ldquoif I didnrsquot have a list Irsquodprobably forget why I walked in the door hererdquoDespite this claim he has never missed an appoint-ment nor is he ever confused about the purpose ofthe testing appointments He can accurately reportthe day of the week He remembers the experi-menters clearly from appointment to appointmentas well as the details of the various experiments
EPSTEIN ET AL
490 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 3 Two floor plans drawn by CO of the apartment he lived in for over 20 years The first plan was drawn when CO still lived inthe apartment the second was drawn several months later when he had moved out Note that the two plans correspond very well COreported that it was quite easy to produce these floor plans
CO home Feb 99
CO (former) home June 99
when they are repeated across appointments Whenqueried about his episodic memory difficulties COreported that he will usually remember an event ifhe is reminded of it but that he has difficulty spon-taneously regenerating events Interestingly hereports that even when he is reminded of an eventand remembers it he cannot remember ldquowhere itwas or when it wasrdquo When he reads he finds it dif-ficult to follow a story if it skips around Howeverwhen queried he was able to accurately report whathe had read in the newspaper the previous day
Formal neuropsychological tests
Formal neuropsychological testing was performedon GR in September 1998and on CO in December1998 Test performances were compared to age-matched peers3
GR exhibited mild to moderate impairment inmemory relative to what would be predicted for hisintellect (WAIS-R Full Scale IQ =122 93rd per-centile superior range) and level of educationThese memory problems were more apparent onvisual than on verbal memory tests On a test ofmemory for conceptually organised text passages(Wechsler Memory Scale Logical Memory[WMS-III] Wechsler 1997b) his performancewas average for both immediate (LM I =25th per-centile standard score SS = 90) and delayed recall(LM II =50th percentile SS =100) In contrast hisscore on the WMS-III Face Memory subtest was inthe low average range for both immediate (9th per-centile SS =80) and delayed recall (16th percentileSS =85) On a test of memory for scenes (WMS-IIIFamily Pictures subtest) immediate recall was defi-cient (2nd percentile SS = 69) and delayed recallwas borderline impaired (5th percentile SS =76)GR also had difficulty learning word lists on theRey Auditory Verbal Learning Test (Spreen ampStrauss 1998) his immediate recall was average(63rd percentile SS = 105) but after the interfer-ence list his performance fell to the low averagerange for both immediate (14th percentile SS =84)
and delayed recall (16th percentile SS =85) On arecognition test of the items of the list he success-fully recognised all 15 but made 8 false alarms Per-formance on the 724 Spatial Recall Test (RaoHammeke McQuillen Khatri amp Lloyd 1984)which required him to remember seven locations ona 6times4 checkerboard was 66th percentile (SS =106)for immediate and 2nd percentile (SS = 69) fordelayed recall Performance on the Benton FaceRecognition test was normal (42nd percentile)Performance on the Boston Naming Test was atceiling indicating that language abilities wereintact Thus GR exhibits some impairment ontasks that require delayed recall of visual informa-tion (WMS-III Face Memory subtest) or verbalinformation that is not organised in terms of a storyor plot (Rey AVLT) and is clearly impaired ontasks that required delayed recall of spatial informa-tion (724 Spatial Recall Test) or stimulus location(WMS-III Family Pictures subtest) In contrasthis ability to recall organised verbal information(WMS-III Logical Memory subtest) appears to beintact
As with GR COrsquos memory performance wasgenerally below what would be expected based onintellect (WAIS-III Full Scale IQ =134 99th per-centile very superior range) and education levelAlso like GR there was some evidence that visualmemory problems were more pronounced than ver-bal memory problems Memory for text passages(WMS-R Logical Memory) was average for bothimmediate (27th percentile SS = 91) and delayedrecall (48th percentileSS =99) In contrast perfor-mance on the WMS-III Faces subtest was lowaverage for both immediate (16th percentile SS =85) and delayed recall (9th percentile SS =80) andperformance on the Family Pictures subtest wasdeficient (lt 1st percentile SSlt 62) for both imme-diate and delayed recall On the Warrington Rec-ognition Memory Tests (RMT) he scored in theaverage range for words (50th percentile SS =100)and scenes (Warrington Topographical Recogni-tion Memory Test 29th percentile SS =92) and in
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 491
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
3The formal neuropsychological testing reported in this section was performed prior to the beginning of our combined
investigation of GR and CO (ie prior to the main experimental investigations of this paper) Consequently GR and CO were nottested on exactly the same battery of tests
the deficient range (lt 5th percentile SS lt 75) forfaces The low score for faces on this test as well asthe WMS-III may reflect prosopagnosia ratherthan a memory problem as he scored in the defi-cient range (lt1st percentile SSlt62) on the BentonFace Recognition test Verbal list learning (ReyAVLT) was average on immediate recall (45th per-centile SS =98) but after hearing an interferencelist performance fell to the deficient level (015words recalled) for both immediate and delayedrecall On a recognition test for these words herecognised 1115 and made one false alarm (39thpercentile SS =96) There was a large discrepancyon tests of memory span which may indicate adecline in spatial memory digit span was superior(9+ forward and 7 backwards both 99th percentile)but spatial span was only average (5 forward and 5backward 37th and 50th percentiles respectively)Language abilities were intact (Boston NamingTest performance was at ceiling) Thus like GRCO shows some impairment at recall of visualinformation (WMS-III Face Memory subtest) andverbal information that is not organised into a nar-rative (Rey AVLT) and clear impairment at tasksthat require recall of stimulus location (Family Pic-tures subtests of WMS-III) In contrast his abilityto recall organised verbal information (WMS-RLogical Memory subtest) is more preserved
MRI RESULTS
In order to determine the extent of their lesionsstructural MRI images were taken of both subjectsDelineation of the lesions was based either onspoiled GRASS coronal images at 11mm thick-ness (GR) or anatomical T-1 weighted MPRAGEMRI sequences yielding 125 mm thick coronalimages (CO) The anterior-posterior extent of thelesions was verified on axial images
GR has bilateral occipital-temporal lesions(Figure 4) In the left hemisphere the posteriorparahippocampal gyrus (including para-hippocampal cortex) is severely damaged with onlyminimal injury to the most caudal aspect of the hip-pocampus There is also moderate damage to themedial fusiform and inferior lingual gyri In the
right hemisphere the hippocampus is spared butthere is damage to the posterior end of theparahippocampal gyrus near the parahippocampal-lingual boundary More posteriorly the lesionedarea extends along the collateral sulcus to includesignificant portions of the inferior lingual gyrus andthe medial fusiform gyrus as well as most of themedial occipital lobe below the calcarine fissure
CO has a right-hemisphere occipital-temporallesion Right posterior hippocampus and para-hippocampal gyrus are severely affected Moreposteriorly the damage covers large portions of theinferior lingual and medial fusiform gyri as well assubstantial portions of the medial occipital lobeincluding most of right calcarine cortex In addi-tion diffuse foci of increased signal on T-2weighted images was present in periventricularwhite matter in the corona radiata and centrumsemiovale with a lacunar infarct of the rightputamen There was also moderate degree of dif-fuse involutional changes with increased ventricularsize particularly notable in the right temporal hornwith ex vacuo dilitation
The structural MRIs suggested that the PPAwas severely if not completely damaged in bothhemispheres for GR and in the right hemispherefor CO In order to confirm this for GR we used ahigh-field 3-T magnet to perform a functional scanin which he viewed scenes faces and objects in thesame blocked fMRI design that we have used tolocalise the PPA in all our previous experiments(see Epstein et al 1999 for details Epstein ampKanwisher 1998) A Kolomogorov-Smirnov testfound no voxels in the parahippocampal region thatfit our previously established criterion for inclusionin the PPA (ie greater response to scenes than tofaces and objects at a uncorrected significance levelof plt 0001) This result contrasts sharply with thepattern seen in over 95 of (mostly young or mid-dle-aged) normal subjects in whom a large acti-vated region can be identified in both left and rightparahippocampal cortex using this comparison (seeFigure 5) A similar test was used to identify voxelsthat responded significantly more to faces than toobjects This comparison revealed a region of face-selective voxels in the right fusiform gyrus consis-tent with the location of the Fusiform Face Area
EPSTEIN ET AL
492 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 493
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
GR
CO
Figure 4 Structural MRIs of GR and CO showing the extent of the damaged region (marked as white) Images have been normalisedusing SPM99 and a cost function masking algorithm (provided by Matthew Brett) into standard stereotactic space (Talairach amp Tournoux1988) using the Montreal Neurological Institute template (Evans et al 1993) Numbers indicate y-Talairach coordinate for each coronalslice corresponding slice planes are also indicated on the sagittal image
(FFA) commonly seen in normal subjects(Kanwisher McDermott amp Chun 1997) Thusthe MRI was sensitive enough to detect functionaldifferences an FFA was found but no PPA Whencombined with the anatomical results this stronglysuggests that GR has no PPA
Because of safety concerns (presence of a femo-ral artery aneurysm clip) we did not perform a simi-lar high-field functional MRI of CO However asFigure 4 demonstrates CO clearly has damage tothe parahippocampal-lingual region in the righthemisphere
EXPERIMENTAL INVESTIGATIONS
In this section we report the results of four experi-ments designed to probe the consequences of dam-age to the lingual-parahippocampal region ingeneral and the PPA in particular Our primaryconcern in these experiments was to test GR andCOrsquos ability to perceive remember and recognisedifferent kinds of visual material Based on our pre-vious fMRI work we predicted that GR and CO
would have more difficulty with scene-like stimuli(ie stimuli that convey information about thegeometry of a space one can navigate through or actwithin) than with object-like stimuli Such a deficitcould account for at least some of the problems GRand CO encounter in real-world navigation
Beyond determining the kind of visual materialthat gets processed in parahippocampal cortex wealso wanted to determine the nature of the process-ing that takes place on that material In particularwe wanted to determine if parahippocampal cortexis involved in perceptual encoding memory encodingor recognition As discussed earlier (Introduction)our fMRI results provided some evidence againstplace recognition as the sole function of the PPAbut did not allow us to distinguish between percep-tual and mnemonic encoding Experiments 1 and 2examine the role of parahippocampal cortex in per-ceptual encoding Experiment 3 examines its role inmnemonic encoding and Experiment 4 examinesits role in place recognition
Experiment 1 Two-dot task
The first hypothesis we examined was thatparahippocampal cortex plays a role in perceivingthe shape of the local environment In our previousfMRI work we observed that the PPA respondsmuch more strongly during scene viewing than dur-ing object or face viewing even when subjects donothing more than watch the scenes passively with-out performing any other task This suggested thatscene perception alone may be sufficient to activatethe PPA Previous reports have indicated that dam-age to parahippocampal cortex and other medialtemporal regions leads to problems with memoryrather than perception (Bohbot et al 1998Maguire Burke Philips amp Staunton 1996 Pigottamp Milner 1993) However all of these previousresults were obtained on patients who had unilat-eral lesions GR has a bilateral occipital-temporallesion including clear damage to parahippocampalcortex in both hemispheres furthermore he reportssome kind of perceptual deficit We hypothesisedthat this deficit might involve difficulty perceivingor interpreting the geometry of scenes and otherstimuli with a similar geometric structure
EPSTEIN ET AL
494 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 5 Top row shows a single slice from a functional MRIscan performed on GR Bottom row shows results from a normalsubject (NK) for comparison Voxels that responded more to facesthan objects (left column) or more to scenes than faces and objects(right column) in a Kolomogorov-Smirnov test are highlighted bywhite arrows GR has a functioning face area (FFA) but there isno evidence of a place area (PPA)
FFA PPA
GR
NK
In this experiment subjects viewed either objectshapes or spatial layouts Stimuli consisted ofblack-and-white photographs of real-world stimuli(empty rooms vs common objects) or abstractstimuli made out of Lego blocks (Lego ldquoscenesrdquo vsLego ldquoobjectsrdquo) These stimuli have been previouslyshown to differentially activate the PPA in normalsubjects (Epstein et al 1999 Epstein ampKanwisher 1998) Two red dots were superim-posed on each photograph (Figure 6) The task wasto report which of the two red dots was over thecloser part of the object or scene This task wasdesigned to have no memory component (apartfrom remembering the instructions) but to requireaccurate 3D perception of objects and scenes
If GR or CO have difficulty perceiving or inter-preting the spatial layout of scenes then we pre-dicted they would do worse when the stimuli areempty rooms or Lego scenes than when they arereal-world or Lego objects Note that the criticalcomparison is between the Lego scenes and theLego objects as these stimuli are made of the samematerials and differ primarily in their surfacegeometry
MethodTesting for all experiments was done on aMacintosh computer The experiment consisted of80 trials (20 of each stimulus type randomly inter-mixed) Subjects began each trial by pressing thespace bar A black-and-white photograph from oneof the four stimulus categories (see earlier) thenappeared on the screen with two red dots on it Thetask was to determine which of the two red dotsindicated the closer part of the scene or objectSubjects pressed the ldquofrdquo key if the leftmost red dotwas closest and the ldquojrdquo key if the rightmost dotwas closest They were instructed to respond asquickly as possible without making errors Stimuliremained on the screen until a response was made
The stimulus set was constructed so that eachempty room was matched to a common object thathad red dots in the exact same location and eachLego layout was similarly matched to a Lego objectThus the locations of the red dots were counterbal-anced across scene and object stimuli controllingfor any effects of visual field cuts
Results and discussionResults for GR and CO and four elderly normalsubjects are shown in Table 3 Both GR and COcould do this task without difficulty and their per-formance was near ceiling There was no evidencethat they had any more trouble with the emptyrooms or Lego scenes than with the real or Legoobjects Reaction times were slightly longer for GRand CO than for normal subjects as one wouldexpect given their visual field deficits however this
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 495
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 3 Experiment 1 (two-dot) results (SD in parenthesis)
GR CO Normals
Proportion correctObject 095 080 093Lego object 100 095 098Empty room 100 100 099Lego room 095 100 096
Mean RTObject 2435 2932 2161 (980)Lego object 2052 3245 2029 (921)Empty room 2744 2291 1788 (959)Lego room 1904 2281 1630 (1127)
Figure 6 The four kinds of stimuli used in Experiment 1 Topleft lego layouts top right real-world layouts (empty rooms)bottom left lego objects bottom right real-world objects
difference did not appear to be significant Impor-tantly there was little evidence that GR or CO tooklonger to make their perceptual judgements for thereal or Lego scenes than for the real or Lego objectsThus contrary to our initial hypothesis this experi-ment found no clear evidence for selective impair-ment in the perception of scene-like stimuli
Experiment 2 Different-views matchingtask
The previous experiment demonstrated that GRand CO can extract information about the per-ceived distances between themselves and the sur-faces of an object or scene Thus in at least onesense their perception of these surfaces is pre-served However this does not mean that their abil-ity to represent the layout of a scene is unimpairedThe two-dot task requires on-line comparison oftwo surfaces in egocentric coordinates GR and COwould be able to perform this task even if they couldnot (1) represent more than two surfaces at a time(2) represent surfaces in other than strictly egocen-tric coordinates or (3) sustain surface representa-tions for longer than the time it takes to make acomparison It has been suggested that the parietallobes are involved in on-line processing of surfacegeometry in egocentric coordinates (Milner ampGoodale 1995) thus it is possible that the parietallobes (which are intact in GR and CO) may be suf-ficient to perform the two-dot task even in theabsence of more durable or complex surface repre-sentations in parahippocampal cortex
In Experiment 2 we examine GR and COrsquosability to create and sustain a representation of thegeometry of an object or scene-like layout The taskwas to view a photograph of a Lego object or Legoscene hold it in memory for 10 s and then deter-mine whether a second object or scene was the sameas the first In ldquosamerdquo trials the second photographdepicted the same scene or object depicted from adifferent viewing angle In ldquodifferentrdquo trials the sec-ond photograph depicted a different scene or objectchosen to be similar to the first one (Figure 7)Thus in order to successfully perform this tasksubjects had to form a representation of the surfacesof the object or scene sustain this representation in
memory for 10 s and then align it (presumablyusing mental rotation) to the second object orscene
MethodThe experiment consisted of two parts each with40 trials In the first part the stimuli were black andwhite photographs of objects made out of Legoblocks In the second part the stimuli were photo-graphs of ldquoscenesrdquo made out of Lego blocks whichwere designed to have a geometric structure similarto real-world places such as a roomor a city street
Subjects began each trial by pressing the spacebar A photograph of a Lego object or sceneappeared on the screen for 3 s followed by a 10-sblank interval A second photograph then appeareddepicting either (1) the same object or scene as thefirst photograph shown from a different viewingangle (offset approximately 35 degrees) or (2) a dif-ferent object or scene Subjects were instructed topress the ldquojrdquo key if the two photographs depicted thesame object or scene and the ldquof rdquo key if they did notThey were told to take as much time as they neededto make a decision but no extra time The secondphotograph remained on the screen until a responsewas made Half the trials were ldquosamerdquo trials and halfwere ldquodifferentrdquo trials
Results and discussionFigure 8 shows results for GR and CO as well as forsix elderly normal controls GRrsquos and COrsquos perfor-mance was comparable to that of the normal con-trols Furthermore GR and CO performed just aswell when the stimuli were Lego scenes as whenthey were Lego objects These results are unlikely toreflect a ceiling effect as performance for bothpatients and controls was around 80 correctThus both patients appear to be able to form a rep-resentation of the geometry of both scenes andobjects which they can sustain in short-termmemory
Experiment 3 N-back recognition memorytask
The previous experiment failed to find any impair-ment in GR and COrsquos ability to form a representa-
EPSTEIN ET AL
496 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
tion of the geometry of a scene or object andmaintain it for 10 s in the absence of the originalstimulus Not only does this result suggest that per-ception of scene-like stimuli is intact in thesepatients it further suggests that some short-termmemory functions are intact as well However we
should note that subjects in this experiment werefree to devote all their effort and attention in the 10-second blank interval to maintaining a memory ofthe stimulus Thus it is possible that GR and COcan use rehearsal mechanisms to maintain the stim-ulus in short-term memory during the blank inter-
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 497
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
(b) Lego object version
Figure 7 Stimuli and procedure for Experiment 2 (a) In the Lego layout version subjects see a Lego layout for 3 s followed by a blankscreen for 10 s A second layout then appears which is either the same layout as the first one but seen from a different angle or a differentlayout (b) The Lego object version is the same but with object stimuli
3 s 10 s OR
3 s 10 s OR
(a) Lego layout version
(b) Lego object version
val but are unable to form a more durable memorytrace for the stimulus This possibility was tested inthe current experiment which was designed toexamine GR and COrsquos ability to learn new visualinformation
We devised a continuous n-back recognitionmemory test in which subjects viewed a sequence oforiginally novel stimuli some of which wererepeated within the sequence at intervals of 1 3 or5 intervening images For each stimulus subjects
reported whether they had seen it before in theexperiment or not In different runs in the experi-ment the stimuli were black-and-white photo-graphs of (1) Lego scenes (2) Lego objects (3) real-world landscapes (4) novel objects made out ofSculpey clay (5) unfamiliar houses (6) unfamiliarfaces as well as (7) line drawings of nonsenseobjects and (8) pronounceable pseudowords (Fig-ure 9) Thus we were able to test GR and COrsquos rec-ognition memory for different kinds of materials atdifferent time intervals in a paradigm in which thepresence of intervening items between the twoappearances of each stimulus made rehearsal diffi-cult Note that because none of the specific stimuliused in this experiment were familiar in advancethis task did not require an episodic memory com-ponent and could be done on the basis of pure visualfamiliarity with the particular visual stimulus
Although many different stimulus classes weretested in this experiment the critical comparison isbetween the Lego layouts and Lego objectsbecause these are best matched with regard tomaterials and differ primarily in their geometricstructure (see Introduction) We predicted that ifGR and CO showed a memory deficit for this taskit would be greater for the Lego layouts than for the
EPSTEIN ET AL
498 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 8 Results of Experiment 2 Performance over ldquosamerdquotrials and ldquodifferentrdquo trials is averaged together Error bars show1 SD
Figure 9 Stimuli for Experiment 3 Top row Lego layouts real-world scenes houses Bottom row Lego objects clay objects nonsenseobjects faces In an eighth condition (not shown) subjects viewed pronounceable nonwords such as ldquowilchrdquo or ldquothipperrdquo
Lego objects In addition a deficit for materialsthat convey information about the layout of sur-rounding space might also be apparent when GRand CO are tested on more realistic stimuli If soone would expect them to do poorly on the land-scapes but relatively well on the clay objects linedrawings of nonsense objects faces andpseudowords
MethodFor each subject a complete experimental sessionconsisted of being run once on each of the eightstimulus classes Each run consisted of 90 trials ineach trial a stimulus was presented and subjectsreported whether or not they had seen it before inthe run (or indeed ever) by pressing the ldquojrdquo key ifthey had or the ldquof rdquo key if they had not Each stimu-lus was on the screen for either 1 2 or 3 s (see fol-lowing) and appeared promptly after the keypressreport for the previous trial Individual stimuliappeared either once (14 stimuli) twice (30 stim-uli) or four times (4 stimuli) within a given run Ofthe stimuli that were presented twice 10 wererepeated immediately (1-back) 10 were repeatedwith a lag of two intervening items (3-back) and 10were repeated with a lag of four intervening items(5-back) In order to ensure that between-subjectdifferences in performance were not due to stimulusdifferences the same stimuli were assigned to thesame conditions each time the experimentwas run
GR performed the experiment twice once inAugust 1998 and again 9 months later Because ofa computer error he was only tested on six of theeight categories (all except Lego layouts and Legoobjects) at the later date CO was also tested twiceonce in February 1999 and again 6 months laterReported results for GR and CO are the average oftheir two testing sessions Normal subjects wereonly tested once Stimulus presentation time was 1 sin GRrsquos first testing session In pilot testing COfound this too fast for the critical categories of Lego
layouts and Lego objects so presentation time wasextended to 2 s for these categories in his first test-ing session All normal subjects were tested at thesepresentation rates (2 s for Lego layouts and objects1 s for other stimuli) In COrsquos second testingsession presentation time was lengthened to 3 s forall stimulus categories to ensure that any deficitsfound were memory deficits rather than problemswith the nonmemory aspects of the task
Results and discussionResults from GR CO and 10 elderly normal con-trols are shown in Figure 10 and Table 4 Figure 10ashows results from the main comparison betweenLego layouts and Lego objects Both normal sub-jects and patients performed near ceiling for bothLego layouts and Lego objects at 1-backinterstimulus intervals verifying that GR and COcould successfully perceive the pictures as they werebeing presented In contrast there was a strikingdifference between the two patients and the normalsubjects at 5-back intervals Whereas normals didon average somewhat better on Lego layouts thanLego objects both GR and CO showed the oppo-site pattern doing much worse on Lego layoutsthan Lego objects The 3-back condition showed amixed pattern Whereas GR did better on Legoobjects than Lego layouts in this condition (similarto his performance at 5-back intervals) CO didbetter on Lego layouts than Lego objects COrsquos lowperformance on Lego objects in the 3-back condi-tion appears to be an anomaly as it is much lowerthan his performance for comparable stimuli in the1-back and 5-back conditions
Figure 10b shows the difference in performancebetween Lego layouts and Lego objects for thecritical 1-back and 5-back intervals At 5-backintervals CO was significantly impaired comparedto the normal subjects (z = ndash250) whereasGRrsquos impairment fell just short of significance (z =ndash175)4
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 499
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
4In fact these values may underestimate GR and COrsquos relative impairment for Lego layouts In the 5-back condition 4 of the 10
normal subjects had perfect performance on the Lego layouts but all scored below ceiling on the Lego objects Thus the true advantagefor Lego layouts over Lego objects for normal subjects is probably greater than that observed here in which case GR and COrsquos lowerperformance for Lego layouts compared to Lego objects would be even more abnormal Furthermore post-hoc analyses revealed thatone of the normal subjects showed a difference in performance between Lego layouts and objects that was over 2 SD below the mean ofthe other nine subjects When this outlying subject was excluded z-scores were ndash322 for CO and ndash232 for GR
Table 4 shows results for all eight stimulusclasses Although direct comparisons between sub-jects should be treated with caution since viewingtime for different items was not the same for eachsubject we do note a few interesting patterns Firstof all there is little or no evidence that GR or CO
show a memory deficit for nonsense words clayobjects or line drawings of nonsense objects GRappears to be slightly impaired on faces and housesbut CO performs normally on these categoriesStrikingly GR shows a severe deficit for the reallandscapes performing more than 40 percentagepoints lower than normals Thus even thoughthere is some evidence that GR might have someimpairment for nonscene stimuli he clearly is muchmore impaired for scene-like stimuli such as theLego scenes and landscapes CO does not show asimilar deficit for the landscapes However itshould be kept in mind that in one of his two testingsessions CO viewed each picture for 3 s which wasmuch longer than the 1 second used for all non-Lego stimuli for GR and normals Thus COrsquosapparently ldquonormalrdquo performance in these condi-tions might be overestimated
Taken together the results of this experimentindicate that GR and CO are selectively impaired atprocessing scene-like stimuli This deficit is consis-tently apparent at 5-back but not 1-back intervalsThis finding suggests that the deficit should becharacterised as a memory encoding problemHowever the present results do not allow us todetermine whether the difficulty occurs at memoryencoding or at subsequent recognition (see Epsteinamp Kanwisher 2001) If GR and CO have a generalproblem with recognition of topographical materi-als then they should not be able to recognise scenes
EPSTEIN ET AL
500 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 10a Results of Experiment 3 for GR (top) CO (middle)and for 10 age-matched normal controls Performance measure isproportion of repeated stimuli correctly identified as such correctedfor guessing using the standard formula
Figure 10b Difference between Lego layout performance andLego object performance for the 1-back and 5-back conditionsError bars indicate one 1 SD
and landmarks that were learned prior to theirinjury We tested this possibility in the nextexperiment
Experiment 4 Famous landmarksfacesrecognition task
In this experiment we examined GR and COrsquosability to recognise the visual appearances of placeslearned prior to injury Subjects saw a series offamous landmarks intermixed with nonfamouscontrols (Imaging experiments have demonstratedthat landmarks of this type can activate the PPAalmost as strongly as complete scenes Epstein etal 1999 Epstein amp Kanwisher 1998) They wererequired to say whether they knew each specificlandmark and to identify the ones they knew Sub-jects were instructed to respond affirmatively if theywere fairly certain that they recognised the land-mark but not if it only looked generally familiarFor purposes of comparison subjects also per-formed an identical test in which the stimuli werefamous and nonfamous faces In both testsdistractor items were designed to be visually andconceptually similar to the target items (see Figure11) If parahippocampal cortex is required for placerecognition then one might expect GR and CO toperform poorly compared to normals on the land-mark recognition test
MethodEach of the two experiments (famous faces famouslandmarks) consisted of 48 trials Subjects beganeach trial by pressing the space bar after which ablack-and-white photograph of a face (in thefamous faces experiment) or a landmark (in thefamous landmarks experiment) appeared on thescreen Subjects were told to press the ldquojrdquo key if theyrecognised the specific face or landmark and the ldquof rdquokey if they did not taking as much time at theyneeded to make this discrimination but no extratime Subjects were told to report that they recog-nised a face or landmark if they felt that they knewit even if they could not retrieve its name If theydid know the name they were to say it after pressingthe button Otherwise they were to say whateverthey did remember about the item
Half of the photographs were of famous and halfof nonfamous people and landmarks The famousfaces included public figures (eg George BushRichard Nixon Prince Charles) and movie stars(eg Harrison Ford Clark Gable Marlon Brando)who were prominent prior to the time of GR andCOrsquos injuries the famous landmarks were build-ings and monuments from all over the world (egEiffel Tower US Capitol Building Great Sphinx)The nonfamous stimuli were chosen to match thefamous stimuli as closely as possible Each famousstimulus had a ldquomatchrdquo For example George Bushwas matched to a man of similar age in a coat andtie and the Taj Mahal was matched to a visually
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 501
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 4 Full results of Experiment 3 Mean proportion correct (corrected for guessing) SDs in parentheses
Lego Land- Lego Clay Nonsense Non-scenes scapes Houses objects objects objects Words Faces
1-back GR 100 094 100 100 100 094 100 100CO 091 085 091 100 100 079 074 100Normals 096(008) 095(008) 093(014) 096(008) 096(006) 099(003) 098(004) 097(011)
3-back GR ndash003 068 026 047 083 083 075 052CO 066 081 037 010 094 069 095 081Normals 066(032) 077(021) 072(015) 077(020) 086(016) 083(015) 083(017) 072(020)
5-back GR 010 034 044 047 072 083 094 053CO 031 073 075 088 081 064 100 081Normals 076(026) 077(016) 068(021) 066(016) 091(014) 083(018) 093(011) 078(021)
similar Asian palace that is not famous (see Figure11)
Results and discussionResults for GR and CO as well as for five age-matched controls are shown in Figure 12 Both
GR and CO could recognise the famous landmarksas well normals In addition both GR and CO wereusually able to name the landmarks they success-fully identified
Thus GR and CO can recognize landmarkslearned prior to injury demonstrating that at leastsome of their place recognition abilities are intactWhen combined with the fact that GR and COwere able to produce detailed maps of places theylived in prior to their injury (see Case Description)these results suggest that their deficit is primarily inencoding or consolidation of new informationrather than in recognition or recall of old informa-tion Note however that these results do not provethat their place recognition abilities are entirelynormal GR and CO might suffer from a mild rec-ognition deficit that affects identification of newly-learned stimuli like the Lego layouts in Experiment3 but not overlearned stimuli like the famous land-marks Furthermore it is possible that GR and COmight be impaired at recognising premorbidlylearned places that are defined primarily by theirspatial layout even though they are unimpaired atrecognising premorbidly learned places that aredefined primarily by a single object-like landmarkor building (see General Discussion) Neverthelessthe trend of the present evidence is to suggest thatGR and CO have particular problems with noveltopographical information
In the famous-face task GRrsquos performance wasin the normal range However he sometimes haddifficulty naming the faces even though he clearly
EPSTEIN ET AL
502 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 11 Stimuli for Experiment 4 Each famous landmark(Arc de Triomphe Taj Mahal) and famous face (Barbara BushYasser Arafat) was matched to a nonfamous building or face thatalso appeared in the experiment
FA
CE
SL
AN
DM
AR
KS
Figure 12 Results of Experiment 4 Performance measure isproportion of famous stimuli reported as ldquoknownrdquo corrected forguessing using the standard formula
recognised them For example he described Ron-ald Reagan as ldquothe gipperrdquo Nancy Reagan as ldquothewife of the gipperrdquo and Clark Gable as ldquothe movieactor who said lsquoScarlett I donrsquot give a damnrsquordquo COhad a great deal of difficulty identifying the famousfaces This result is not surprising given hisextremely low performance on the Benton FaceMatching Task In addition to his topographicproblems CO appears to suffer fromprosopagnosia
GENERAL DISCUSSION
We tested two parahippocampal patients (GR andCO) on four experiments designed to measure per-formance as a function of both task (perceptionencoding and recognition) and stimulus type(scenes objects faces and words) The only taskthat revealed a clear deficit was an n-back recogni-tion memory test (Experiment 3) In order to suc-cessfully perform this task subjects had to encode adurable representation of a visual stimulus and useit for later recognition GR and CO were signifi-cantly more impaired on this task when the stimuliwere scene-like spatial layouts (ie Lego scenesand real scenes for GR) than when they wereobjects No comparable deficits were found in otherexperiments that tested layout perception (Experi-ments 1 and 2) and placelandmark recognition(Experiment 4) Taken together these results indi-cate that GR and CO are impaired in their ability toencode novel information about the geometry ofsurrounding space into memory but are lessimpaired at their ability to encode the geometry ofnovel objects
We argue that this specific topographical defi-citmdashinability to encode the spatial layout of novelscenesmdashresults from damage to the para-hippocampal place area Although the damage inboth patients extends beyond the parahippo-campal-lingual territory normally associated withthe PPA this region is clearly lesioned bilaterally inGR and in the right hemisphere in CO Further-more functional imaging demonstrated that GRhas no functioning PPA Thus the current resultsand the earlier fMRI work (Epstein et al 1999
Epstein amp Kanwisher 1998) converge to implicateparahippocampal cortex in the encoding of infor-mation about scene geometry into memory
Our assignment of the function of memoryencoding to parahippocampal cortex is consistentwith the results of earlier case reports of patientswith parahippocampal damage who have naviga-tional difficulties in novel but not familiar environ-ments (Habib amp Sirigu 1987 Ross 1980 see alsoTeng amp Squire 1999) As with GR and CO thesepatients were either less impaired or unimpaired onnontopographical memory tasks Patients have alsobeen described who exhibit the opposite pattern ofimpairment an inability to learn faces (sometimescalled ldquoprosopamnesiardquo Cipolotti Robinson Blairamp Frith 1999 Tippett Miller amp Farah 2000) orfaces and verbal materials (Maguire amp Cipolotti1998) with an unimpaired ability to learn topo-graphical materials Taken together these resultsargue for a high degree of domain-specificity in theneural systems involved in learning of new informa-tion (Gallistel 1995) The brain may have separatemechanisms for learning faces places and wordswhich can be differentially impaired after damageto specific cortical regions (Cipolotti et al 1999Incisa della Rochetta et al 1996) The presentresults suggest that the anatomical locus of themechanism dedicated to learning new topographi-cal information is in the parahippocampal cortex(Aguirre amp DrsquoEsposito 1999)
In contrast to their impaired ability to encodenew topographical information both GR and COwere able to produce detailed maps of their previousdwellings and performed normally on the famouslandmark recognition test of Experiment 4 Theseresults are generally consistent with Teng andSquirersquos (1999) report of a patient with extensivemedial temporal damage who could recallpremorbidly learned but not postmorbidly learnedtopographical information (see also Ross 1980)Thus our results provide further evidence thatmedial temporal regions such as parahippocampalcortex may be more critical for encoding or consoli-dating new information into memory than for recallof old information
One issue that our experiments do not entirelyresolve is the precise nature of the topographical
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 503
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
information encoded by parahippocampal cortexClearly this region is critical for encoding informa-tion about the geometry of the currently visiblescene even when the ldquoscenerdquo is made out of Legoblocks and therefore has no immediate navigationalrelevance However it is unclear whether this spe-cific deficit can account for all of the real-worldnavigational difficulties encountered by GR andCO GR in particular shows such a catastrophicloss of navigational ability that we suspect he is alsoimpaired at encoding the spatial relationshipsbetweenthe current scene and other locations in theworld CO probably also suffers from such difficul-ties as he reports that he has trouble rememberinghow places relate to each other in large-scale spaceWhen he was tested on a simple 3-room videogameenvironment he not only was unable to rememberthe individual rooms (claiming that there were 10rooms when there were only 3) but he also had nomemory for the spatial relationships between themThese spatial deficits may be the result of damage tothe posterior right hippocampus (in the case of CO)or of disconnection of the posterior right hippo-campus from the parahippocampal regions thatprovide much of its cortical input (in both patientsSuzuki amp Amaral 1994) In either case the abilityto form a cognitive map of new environments likethe lab or videogame space could be impaired(Maguire et al 1998 OrsquoKeefe amp Nadel 1978)However it is also possible that parahippocampalcortex itself may be critical for encoding not onlythe geometry of individual scenes but also the spa-tial relationships between the locations of thosescenes In this case the spatial deficits of GR andCO could be attributed directly to parahippo-campal damage rather than to hippocampal dam-age or disconnection
The latter scenario would be consistent withAguirre and DrsquoEspositorsquos (1999) and Barrash etalrsquos (2000) suggestion that parahippocampal cortexis critically involved in combining visual informa-tion about the appearance of scenes and landmarkswith spatial information about their locations into aunified topographical representation Howeverboth of these authors argue that the representationsof individual scenes (Barrash et al) and landmarks(Aguirre amp DrsquoEsposito) are supported not by
parahippocampal cortex but by more posterior lin-gual-fusiform regions In contrast we believe thatthe neuroimaging data indicate that parahippo-campal cortex is involved in the encoding of thegeometric structure of individual scenes althoughit may also play a role in combining these represen-tations with other spatial representations
Interestingly GR and CO were not impaired onrecognition of famous landmarks that were mostlybuildings Thus our data do not preclude the possi-bility that there may be a separate lingual orfusiform region involved in processing informationabout buildings (Aguirre Zarahn amp DrsquoEsposito1998 Chao Haxby amp Martin 1999 IshaiUngerleider Martin Schouten amp Haxby 1999)which might have been preserved in GR and COdespite extensive damage to other regions of thelingual and fusiform gyrus Alternatively GR andCOrsquos preserved building recognition abilities mightbe mediated by interactions between cortical areasthat perform shape analysis for any object-likestimulus type (Malach et al 1995Kanwisher et al1996) and anterior temporal regions in which long-term topographical and semantic memories arestored (McCarthy et al 1996)
Is perception preserved
In contrast to their inability to encode new scenesinto memory GR and CO have substantiallyretained their ability to perceive the spatial layout ofthe currently visible scene (Experiments 1 and 2)GR can follow a route marked on a floor planwhich indicates a good on-line appreciation of hiscurrent spatial surroundings Furthermore neitherGR or CO have any trouble moving through theenvironmentmdashthey move with confidence and donot bump into doors or walls Problems of the lattertype are usually associated with damage to the pari-etal lobes which are intact in GR and CO Thus atfirst glance GR and CO do not appear to haveperceptual impairments (beyond their visual fieldrestrictions)
However GR does complain of a perceptualdeficit which he describes as an inability to ldquotakeinrdquo scenes when they are too complex Similarreports of an inability to comprehend the
EPSTEIN ET AL
504 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
ldquoglobalityrdquo of a scene were made by some of theparahippocampally-damaged patients studied byHabib and Sirigu (1987) We were not able todetermine the precise nature of this deficit whichmay be partly attributable to his visual field restric-tions It appears that his deficit is more salient in thereal-world environments than in the picture-per-ception tasks of Experiments 1 and 2 perhapsbecause he finds it more difficult to organise visualinput when immersed in it than when viewing pho-tographs that subtend only part of the visual field
We suggest that GR can perceive parts of thescene when he attends to them but may be unableto integrate information over time into a coherentrepresentation of a scene or place The fact that GRand CO are impaired for Lego layouts at 5-backintervals in Experiment 3 is consistent with thishypothesis The time between the first and secondpresentations of a stimulus is very short in this case(and even shorter with 3-back intervals where GRalso shows evidence of a deficit) In both patientsthe representation of layout information formed byperception of a Lego scene must either have anabnormally short persistence be abnormally sus-ceptible to interference or both If so they mightfind it impossible to sustain a spatial frame orschema long enough to use it to organise informa-tion obtained from many different glances at a com-plex scene (Intraub 1997) The result would be aninability to understand the ldquoglobalityrdquo of a sceneeven though all the details were perfectlyperceived
Non-topographical mnemonic abilities
Although not as severe as their topographical mem-ory problems GR and CO do have some memorydifficulties in other domains For example bothperformed poorly on the Rey AVLT whichrequired them to remember a list of unrelatedwords Both also report some subtle episodic mem-ory problems (see Case Description) COrsquos prob-lems could partly result from damage to the righthippocampusmdashindeed CO appeared to have themore acute episodic memory problems HoweverGR has no hippocampal damage so the fact that hedisplays episodic memory problems suggests thatparahippocampal cortex is involved in the forma-
tion of episodic memories (cf Wagner et al1998)
In previous work (Epstein et al 1999) weargued that spatial context information encoded byparahippocampal cortex might play a role in estab-lishing event memories by encoding informationabout where events take place The inability toencode this information might cause problems intasks such as the Rey AVLT where one has to recalla list of unrelated words If spatial (and perhapstemporal) context information is not encoded itwill be harder subsequently to recall the distinctevent of hearing each word (Tulving 1983) Incontrast lack of spatial context information wouldbe less of a problem in tasks where one is onlyrequired to recognise previously encountered verbalmaterials but not to reproduce the spatiotemporalcontext in which they were experienced IndeedGR and CO performed as well as normals on thenonsense words in Experiment 4 where recogni-tion memory but not episodic memory was requiredfor successful performance Interestingly GR andCO also did relatively well on the Logical Memorysubtest of the WMS-III perhaps because the nar-rative framework of a story provides sufficient con-textual information to overcome the absence ofspatial context and mediate successful verbalretrieval
Differences between GR and CO
Although we have discussed GR and CO togetherthere are a number of relevant differences betweenthem One open question is whether these differ-ences can be attributed to the different effects ofunilateral and bilateral parahippocampal damageIn general GRrsquos topographical difficulties weremore severe than COrsquos CO did surprisingly well onthe Warrington Topographical Memory Test inwhich he was required to remember detailed real-world city scenes and also performed substantiallybetter on real scenes than Lego scenes in Experi-ment 3 Insofar as CO has intact left para-hippocampal cortex this suggests that the left PPAmight be sufficient to encode a certain amount of(perhaps semantic) information about real-worldscenes but the right PPA is necessary to encode
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 505
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)

was in the perceptual recognition or memorydomain To anticipate we did find evidence for adeficit for visual materials that conveyed informa-tion about the shape of surrounding space Thisdeficit manifested itself during a memory encodingtest but not during perceptual tests or a recognitiontest Thus our data suggest that the PPA is neces-sary for the encoding of spatial layout informationinto memory but may not be necessary for manyaspects of the initial perceptual analysis recogni-tion or recall of this information We argue that thePPA may be a learning mechanism specifically ded-icated to encoding topographical materials intomemory
CASE DESCRIPTIONS ANDNEUROPSYCHOLOGICAL TESTING
In this section we present the case histories of thetwo patients in order to provide a general contextfor the more specific experimental tests reportedin the Experimental Investigations section Thepatients were chosen for investigation because clin-ical MRIs indicated that they had suffered damageto the parahippocampal region This section is
divided into four subsections First we present ageneral overview of the case histories Second wereport the results of some ad hoc tests that give ageneral indication of the patientrsquos navigational abil-ities Third we discuss other cognitive abilitiesincluding face recognition and episodic memoryFourth we report the results of a battery of formalneuropsychological tests performed on the patientsTable 1 summarises relevant observed and self-reported behaviour and Table 2 summarises theresults of the neuropsychological tests
Overview
Case 1 GRGR2 is a right-handed male with 18 years of educa-tion He was 60 years old at the time of testingTwo-and-a-half years prior to testing he suffered aright occipital-temporal stroke during cardiacsurgery A second stroke affecting the left occipital-temporal region occurred a week later Thesestrokes left him with a number of deficits includinglimitation of his peripheral vision (lefthemianopsia right upper quadrantanopsia)dyschromatopsia (details reported in Beauchampet al 2000) and topographical disorientation We
484 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
EPSTEIN ET AL
Table 1 Summary of observed and self-reported behaviour
GR CO
Real-world topographical Unable to recognise 714 newly learned MIT landmarks Unable to learn 3-room videogamelearning despite extensive training regime environment
Unable to find way back to lab unassisted despite Able to learn a simple route out ofextended experience with environment building after many visits
Map drawing Could not draw map of lab maps of other newly learned Map of lab was sketchy but not entirelyenvironments were inaccurate inaccurate
Drew accurate map of premorbid environment Drew accurate map of premorbid(former home) environment (currentformer home)
Scene perception Reports difficulty understanding the globality of complex Does not report a perceptual deficitscenes
Face recognition Some prosopagnosia Severe prosopagnosia
Episodic memory Largely intact though some difficulties were reported Moderate difficulties were reported
Can follow the plot of a movie or novel and report Reports some difficulty following theprevious dayrsquos activities plot of a book Accurately reported
contents of previous dayrsquos newspaper
2GRrsquos true initials are KG (cf Beauchamp Haxby Rosen amp DeYoe 2000)and COrsquos true initials are KC We use GR and CO
here to avoid confusion between the two patients
tested him over a period of 5 days in August 1998with 3 days of follow-up testing 3 weeks later and 1day of additional tests in May 1999 During thistime he was alert and cooperative Emotionalaffect and language use was normal He interactednormally in nonclinical social situations and hisconversation was appropriate and varied Heremembered the events of previous days andrecounted them accurately
In contrast to this high level of non-topographical functioning GR demonstrated adramatic inability to orient himself in space Forexample even after several days of testing he neverlearned the relative locations of the different roomsof the lab According to both GR and his wife thisinability to learn new topographical informationwas typical of his experience since his strokes as hefrequently gets lost in his daily life Soon after his
injury he moved to a neighbourhood with manysimilar-looking houses He reported that in orderto find his new house after a walk to a market six toseven blocks away he had to rely on street signs toguide him to the correct block and then examineeach house on the block in detail until he couldrecognise some feature that distinguished his homeuniquely Subsequently GR and his wife moved toa different house in a different country He reportedthat for the first 6 months after the move his newhome was like a ldquohaunted houserdquo for him insofar ashe was unable to learn his way around it For exam-ple he would repeatedly forget that the bedroomhad a balcony and he would be surprised by this factevery time he rediscovered it After several monthshowever he was able to learn the topography of thehouse sufficiently well to navigate through it (butsee below)
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 485
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 2 Summary of neuropsychological tests ( indicates percentile scores)
GR COmdashmdashmdashmdashmdashmdashmdashmdashmdashmdash mdashmdashmdashmdashmdashmdashmdashmdashmdashmdashmdashndash
IQ 122 (93 WAIS-R) 134 (99 WAIS-III)
WMS-III
Logical memory 25 immediate recall 27 immediate recall50 delayed recall 48 delayed recall(WMS-R)
Face memory 9 immediate recall 16 immediate recall16 delayed recall 9 delayed recall
Family pictures 2 immediate recall lt1 immediate recall5 delayed recall lt1 delayed recall
Rey Auditory Verbal 63 immediate recall 45 immediate recallLearning test 14 after interference list lt1 after interference list
16 delayed recall 3 delayed recall
Warrington Recognition 50 wordsMemory tests 29 scenes
lt5 faces
724 Spatial Recall 66 immediate recall2 delayed recall
Benton Face Recognition 42 lt1
Memory span Digits99 forward99 backward
Spatial37 forward50 backward
Boston Naming test Ceiling Ceiling
Case 2 COCO is a left-handed male with 20 years of educa-tion He was 60 years old at the time of testing Twoyears prior to testing he suffered a right posteriorcerebral artery stroke He first noticed on a week-end that his vision appeared blurred on the left andthat he had a tendency to bump into objects on thisside On Monday morning he was able to success-fully negotiate the route to his workplace a trip thatinvolved a bus ride followed by two subways andthen a shuttle to his office building However oncehe entered the building he found himself com-pletely disoriented unsure of its layout and wherehis office was located Interestingly the companyhe worked for had moved a few weeks earlierAlthough he had managed to arrive at this newbuilding (which was a block away from the previouslocation) he became lost upon entering it Hisneurologic exam revealed a left homonymoushemianopia but no evidence of neglect on a lettercancellation task or on sensory testing Investiga-tions revealed an ischaemic stroke with haemor-rhagic transformation in the right occipital andmesial temporal lobe On magnetic resonanceangiography he had a chronic left carotid occlusiona mild right carotid stenosis and bilateral vertebralartery stenoses Since then he has suffered fromsevere topographical disorientation primarily innew environments
CO reports that he can recognise buildings thatwere familiar to him before the stroke He can alsoform mental images of well-known landmarks suchas the Empire State Building and the BostonPublic Gardens and of the house he lived in as achild When asked to imagine standing in front ofthe Massachusetts State House (a location wellknown to him from years of living in Boston) heaccurately described the surrounding buildings andtheir locations However he reports that when helooks at a place that hersquos encountered post-strokeall he gets is a feeling of familiarity that does notallow him to determine where the place is or whenhe saw it He also reports an inability to understandhow various landmarks relate to each other inlarge-scale space Despite this difficulty he canfollow a map and frequently rides the Bostonsubway alone
Unlike GR CO was able to learn a certainamount of topographical information For exam-ple he soon learned the location of the rooms in thelab and would travel between them unassistedAfter his first visit to the lab he needed to be guidedall the way back to the subway station but after sev-eral visits he could find his way out of the buildingand to the subway on his own as long as he was firstguided to the elevators (It should be noted how-ever that the building is on the same block as thesubway station Furthermore he still needed to bedirected to the elevators even though our buildinghas a relatively simple floor plan) Thus the amountof topographical information he can learn is limitedbut not zero
COrsquos language and reasoning skills appear to benormal and his conversation is appropriate andvaried In addition to his topographical difficultieshe also has limitations of his peripheral vision onthe left side
Navigational abilities
In addition to the experiments reported in theExperimental Investigations section of this paperwe also performed a number of ad hoc tests of GRand COrsquos navigational abilities Although thesetests were informal we describe them here to pro-vide a general sketch of the nature of GR and COrsquosimpairments
Real-world topographical learningOver the course of several days we took GR and hiswife on three walks around the MIT campus whichneither had visited previously Each walk followedexactly the same path At several places in the walkwe would stop and point out different buildings andlandmarks tell them something about each one andask them to remember their appearance for latertesting Immediately after the completion of thethird walk both GR and his wife separately per-formed a forced-choice recognition test in whichthey saw photographs of 14 landmarks (mostlybuildings) from the walk and 14 novel landmarks(from another part of the MIT campus that theyhad not visited) and were required to reportwhether they had seen each one or not GRrsquos
EPSTEIN ET AL
486 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
performance was 714 correct hits for the familiarlandmarks and 1314 correct rejections for theunfamiliar landmarks In contrast his wifeobtained a perfect score on this test Given theextensive nature of the training regime GRrsquos scorestrongly suggests an impairment in learning theappearance of new real-world places Interestinglyseveral of the seven familiar landmarks that he gotright had an obvious distinctive feature that hadbeen explicitly pointed out to him For exampleone building had a distinctive clock tower andanother one was an unusual triangular shape ThusGR was occasionally able to use distinct visualfeatures to identify places even though he wasunable to recognise them holisticallymdasha commonpattern among topographical disorientationpatients
GR did not appear to have any obvious problemsin understanding the spatial structure of his imme-diate environment When given a floor plan of thebuilding at MIT in which he was being tested hewas able to successfully follow a route marked onthe plan However when the plan was taken awayhe had great difficulty finding his way back to thelab even though he had more than 30 min experi-ence with the environment and his starting positionwas less than 30 feet from the goal (Figure 2) Infact the route he took resembled a random walk Atone point he walked right past his destinationwithout recognising it and only knew for sure thathe was in the right place when he identified his wifein the room Thus his on-line appreciation of thelayout of immediately visible space appears to beintact but he cannot find his own way withoutassistance
We were unable to perform the MIT landmarkrecognition test on CO because he suffered from amedical problem that made it difficult for him towalk around campus In lieu of this we tried to seewhether he could learn a simple video game envi-ronment consisting of three connected roomsAfter about 30 min experience navigating throughthis environment (via button presses) he was askedto sketch a map of it He declined to do so statingthat it was beyond his abilities When asked howmany rooms there were he replied that he did notknow but that there were at least 10 Given the
simplicity of this artificial environment COrsquos com-plete inability to learn it is striking
Map drawingAfter several days of testing we asked both GR andhis wife to sit at different tables and independentlysketch (1) the shape of Florida (2) a bicycle (3) afloor plan of their hotel room in Boston (4) a floorplan of the first floor of the house they had been liv-ing in for the last 8 months (all post-stroke) (5) afloor plan of the main room of the lab and (6) afloor plan of the house they lived in 10 years ago(pre-stroke) GR drew the map of Florida and thebicycle perfectly His plan of the hotel room wasnotably different from the one drawn by his wifemdashfor example he reported that there was only onebed when in fact there were two and he placed thebed on the wrong side of the room (Figure 1a) Hisplan of the first floor of his current house was some-what better but was not very detailed and includedsome major errors For example he wronglyreported that there was no bathroom on this floor(Figure 1b) He was unable to produce any floorplan of the main room of the lab leaving this pageblank When his wife reminded him that this wasthe room in which he had just spent an hour readingthe newspaper and in which we had all had break-fast in that morning he replied that he didnrsquot knowwhat room she was talking about (although heappeared to remember these events) In contrast tothis near total inability to produce accurate maps ofnew environments GR produced an accurate mapof the house he lived in prior to his injury (Figure1c) He reported that his memory of this house wasldquoall thererdquo and he could imagine walking around init When asked to produce a perspective of a roomwhen he was in it he did so with ease and he wasalso able to create an accurate floor plan of theroom
In contrast CO found it very difficult to pro-duce a floor plan of the lab both when he was in itand when he was taken to another room and askedto recall it However although the maps he pro-duced were quite impoverished they were notentirely inaccurate demonstrating some ability toencode topographical information This contrastswith GRrsquos absolute inability to recall anything
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 487
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
EPSTEIN ET AL
488 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure
1F
loorplansdraw
nby
GR
andhisw
ifeof(a)
theirhotelroomin
Boston(b)theircurrenthousew
hichthey
hadm
ovedto
subsequenttoG
Rrsquosinjury(c)the
housethey
livedin
priorto
GR
rsquosinjury
Ho
telro
om
inB
osto
n
GR
His
wife
His
wife
Cu
rren
th
om
e
GR
His
wife
Fo
rme
rh
om
e
GR
about the main room of the lab once he had left itCO was also able to draw a detailed floor plan of theapartment he was living in at the time (and hadlived in for 20 years) Several months after he hadmoved from this apartment we asked him to drawsuch a floor plan again and he produced one thatwas substantially the same as the first (Figure 3)Thus their map drawing performance indicatesthat GR and CO are primarily impaired at sketch-ing places experienced after their strokes but areunimpaired at sketching places experienced beforetheir strokes This pattern is indicative of deficits inencoding but not recall
Scene perceptionAs noted earlier GR can navigate with the aid of afloor plan suggesting that his perceptual abilitiesare largely intact Interestingly however he reportsthat he does have a perceptual deficit which hedescribes as an inability to understand scenes whenthey are too complex For example he reports thathe often has trouble using maps because theyusually are too busy with details It is unclearwhether this apparent perceptual problem is a resultof his field cut some other separate perceptual
problem or a memory problem (see discussion)CO did not complain of a similar deficit GR wasan accomplished artist prior to his injury producingpaintings of striking complexity Tragically hereports that he cannot now appreciate his ownpaintings
Other cognitive abilities
Face recognitionGR also reported some difficulties with face recog-nition On his second visit we examined his abilityto learn new faces by having him perform a forced-choice recognition test on seven familiar and sevenunfamiliar faces The familiar faces were personnelfrom the lab with whom he had at that point had agood deal of experience spanning at least 3 daysAll the people in the photographs had a towelwrapped around their head in order to minimise theuse of hair as a nonfacial cue Despite the difficultyof this task he correctly identified 67 familiarfaces and made no false alarms Surprisingly theone face that he did miss was the first author of thispaper with whom he at that point had had consid-erable experience GR also performed normally ona difficult famous face recognition test (see Experi-ment 4 following) and on the Benton Face Recog-nition test (see following) Thus he has someability to recognise faces Consistent with this GRreports that he can remember a small number offaces but that he quickly ldquoloses trackrdquo if he has toremember too many
CO also appears to suffer from someprosopagnosia as evidenced by his low perfor-mance on the Benton Face Recognition test (seefollowing see also Experiment 4) Interestingly hedoes not complain of face recognition difficultiesbut rather attributes his inability to recognise peo-ple to a lack of interest in their appearance
Episodic memoryAt first glance GRrsquos episodic memory appeared tobe largely intact As mentioned he could recall theevents of the previous day accurately Furthermorehe reports that he can follow the plot of a moviewithout any difficulty However both he and hiswife report that he does have some problems with
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 489
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Figure 2 Floor plan of the building at MIT in which GR wastested Corridors are highlighted in grey GR was asked to find themain room of the lab (marked G on the map) Starting location(marked S) was a point approximately 30 feet down the hall GRrsquosroute is marked by arrows GR found this task very difficult andonly knew for certain that he had reached his goal when he caughtsight of his wife through the glass door of the lab In contrast hewas able to follow a route marked on a plan of the same floorwithout any problem or hesitation
episodic memory For example he stated that heremembered very little about a trip to Italy they hadtaken several months previously His wife reportsthat he has great difficulty keeping track of timeFor example before their trip to Boston he repeat-edly asked her what day they were supposed to leaveand seemed unable to encode this information Weobserved a similar phenomenon during testingDuring the tests that required repetition of thesame procedure several times he sometimesseemed unable to clearly remember how manytimes he had repeated the procedure Thus he doesappear to have a subtle episodic memory deficit
which may manifest itself as an inability to sepa-rately remember successive events that are not easilydistinguishable from each other
CO also reports some problems with his epi-sodic memory He relies on lists to rememberappointments stating ldquoif I didnrsquot have a list Irsquodprobably forget why I walked in the door hererdquoDespite this claim he has never missed an appoint-ment nor is he ever confused about the purpose ofthe testing appointments He can accurately reportthe day of the week He remembers the experi-menters clearly from appointment to appointmentas well as the details of the various experiments
EPSTEIN ET AL
490 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 3 Two floor plans drawn by CO of the apartment he lived in for over 20 years The first plan was drawn when CO still lived inthe apartment the second was drawn several months later when he had moved out Note that the two plans correspond very well COreported that it was quite easy to produce these floor plans
CO home Feb 99
CO (former) home June 99
when they are repeated across appointments Whenqueried about his episodic memory difficulties COreported that he will usually remember an event ifhe is reminded of it but that he has difficulty spon-taneously regenerating events Interestingly hereports that even when he is reminded of an eventand remembers it he cannot remember ldquowhere itwas or when it wasrdquo When he reads he finds it dif-ficult to follow a story if it skips around Howeverwhen queried he was able to accurately report whathe had read in the newspaper the previous day
Formal neuropsychological tests
Formal neuropsychological testing was performedon GR in September 1998and on CO in December1998 Test performances were compared to age-matched peers3
GR exhibited mild to moderate impairment inmemory relative to what would be predicted for hisintellect (WAIS-R Full Scale IQ =122 93rd per-centile superior range) and level of educationThese memory problems were more apparent onvisual than on verbal memory tests On a test ofmemory for conceptually organised text passages(Wechsler Memory Scale Logical Memory[WMS-III] Wechsler 1997b) his performancewas average for both immediate (LM I =25th per-centile standard score SS = 90) and delayed recall(LM II =50th percentile SS =100) In contrast hisscore on the WMS-III Face Memory subtest was inthe low average range for both immediate (9th per-centile SS =80) and delayed recall (16th percentileSS =85) On a test of memory for scenes (WMS-IIIFamily Pictures subtest) immediate recall was defi-cient (2nd percentile SS = 69) and delayed recallwas borderline impaired (5th percentile SS =76)GR also had difficulty learning word lists on theRey Auditory Verbal Learning Test (Spreen ampStrauss 1998) his immediate recall was average(63rd percentile SS = 105) but after the interfer-ence list his performance fell to the low averagerange for both immediate (14th percentile SS =84)
and delayed recall (16th percentile SS =85) On arecognition test of the items of the list he success-fully recognised all 15 but made 8 false alarms Per-formance on the 724 Spatial Recall Test (RaoHammeke McQuillen Khatri amp Lloyd 1984)which required him to remember seven locations ona 6times4 checkerboard was 66th percentile (SS =106)for immediate and 2nd percentile (SS = 69) fordelayed recall Performance on the Benton FaceRecognition test was normal (42nd percentile)Performance on the Boston Naming Test was atceiling indicating that language abilities wereintact Thus GR exhibits some impairment ontasks that require delayed recall of visual informa-tion (WMS-III Face Memory subtest) or verbalinformation that is not organised in terms of a storyor plot (Rey AVLT) and is clearly impaired ontasks that required delayed recall of spatial informa-tion (724 Spatial Recall Test) or stimulus location(WMS-III Family Pictures subtest) In contrasthis ability to recall organised verbal information(WMS-III Logical Memory subtest) appears to beintact
As with GR COrsquos memory performance wasgenerally below what would be expected based onintellect (WAIS-III Full Scale IQ =134 99th per-centile very superior range) and education levelAlso like GR there was some evidence that visualmemory problems were more pronounced than ver-bal memory problems Memory for text passages(WMS-R Logical Memory) was average for bothimmediate (27th percentile SS = 91) and delayedrecall (48th percentileSS =99) In contrast perfor-mance on the WMS-III Faces subtest was lowaverage for both immediate (16th percentile SS =85) and delayed recall (9th percentile SS =80) andperformance on the Family Pictures subtest wasdeficient (lt 1st percentile SSlt 62) for both imme-diate and delayed recall On the Warrington Rec-ognition Memory Tests (RMT) he scored in theaverage range for words (50th percentile SS =100)and scenes (Warrington Topographical Recogni-tion Memory Test 29th percentile SS =92) and in
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 491
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
3The formal neuropsychological testing reported in this section was performed prior to the beginning of our combined
investigation of GR and CO (ie prior to the main experimental investigations of this paper) Consequently GR and CO were nottested on exactly the same battery of tests
the deficient range (lt 5th percentile SS lt 75) forfaces The low score for faces on this test as well asthe WMS-III may reflect prosopagnosia ratherthan a memory problem as he scored in the defi-cient range (lt1st percentile SSlt62) on the BentonFace Recognition test Verbal list learning (ReyAVLT) was average on immediate recall (45th per-centile SS =98) but after hearing an interferencelist performance fell to the deficient level (015words recalled) for both immediate and delayedrecall On a recognition test for these words herecognised 1115 and made one false alarm (39thpercentile SS =96) There was a large discrepancyon tests of memory span which may indicate adecline in spatial memory digit span was superior(9+ forward and 7 backwards both 99th percentile)but spatial span was only average (5 forward and 5backward 37th and 50th percentiles respectively)Language abilities were intact (Boston NamingTest performance was at ceiling) Thus like GRCO shows some impairment at recall of visualinformation (WMS-III Face Memory subtest) andverbal information that is not organised into a nar-rative (Rey AVLT) and clear impairment at tasksthat require recall of stimulus location (Family Pic-tures subtests of WMS-III) In contrast his abilityto recall organised verbal information (WMS-RLogical Memory subtest) is more preserved
MRI RESULTS
In order to determine the extent of their lesionsstructural MRI images were taken of both subjectsDelineation of the lesions was based either onspoiled GRASS coronal images at 11mm thick-ness (GR) or anatomical T-1 weighted MPRAGEMRI sequences yielding 125 mm thick coronalimages (CO) The anterior-posterior extent of thelesions was verified on axial images
GR has bilateral occipital-temporal lesions(Figure 4) In the left hemisphere the posteriorparahippocampal gyrus (including para-hippocampal cortex) is severely damaged with onlyminimal injury to the most caudal aspect of the hip-pocampus There is also moderate damage to themedial fusiform and inferior lingual gyri In the
right hemisphere the hippocampus is spared butthere is damage to the posterior end of theparahippocampal gyrus near the parahippocampal-lingual boundary More posteriorly the lesionedarea extends along the collateral sulcus to includesignificant portions of the inferior lingual gyrus andthe medial fusiform gyrus as well as most of themedial occipital lobe below the calcarine fissure
CO has a right-hemisphere occipital-temporallesion Right posterior hippocampus and para-hippocampal gyrus are severely affected Moreposteriorly the damage covers large portions of theinferior lingual and medial fusiform gyri as well assubstantial portions of the medial occipital lobeincluding most of right calcarine cortex In addi-tion diffuse foci of increased signal on T-2weighted images was present in periventricularwhite matter in the corona radiata and centrumsemiovale with a lacunar infarct of the rightputamen There was also moderate degree of dif-fuse involutional changes with increased ventricularsize particularly notable in the right temporal hornwith ex vacuo dilitation
The structural MRIs suggested that the PPAwas severely if not completely damaged in bothhemispheres for GR and in the right hemispherefor CO In order to confirm this for GR we used ahigh-field 3-T magnet to perform a functional scanin which he viewed scenes faces and objects in thesame blocked fMRI design that we have used tolocalise the PPA in all our previous experiments(see Epstein et al 1999 for details Epstein ampKanwisher 1998) A Kolomogorov-Smirnov testfound no voxels in the parahippocampal region thatfit our previously established criterion for inclusionin the PPA (ie greater response to scenes than tofaces and objects at a uncorrected significance levelof plt 0001) This result contrasts sharply with thepattern seen in over 95 of (mostly young or mid-dle-aged) normal subjects in whom a large acti-vated region can be identified in both left and rightparahippocampal cortex using this comparison (seeFigure 5) A similar test was used to identify voxelsthat responded significantly more to faces than toobjects This comparison revealed a region of face-selective voxels in the right fusiform gyrus consis-tent with the location of the Fusiform Face Area
EPSTEIN ET AL
492 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 493
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
GR
CO
Figure 4 Structural MRIs of GR and CO showing the extent of the damaged region (marked as white) Images have been normalisedusing SPM99 and a cost function masking algorithm (provided by Matthew Brett) into standard stereotactic space (Talairach amp Tournoux1988) using the Montreal Neurological Institute template (Evans et al 1993) Numbers indicate y-Talairach coordinate for each coronalslice corresponding slice planes are also indicated on the sagittal image
(FFA) commonly seen in normal subjects(Kanwisher McDermott amp Chun 1997) Thusthe MRI was sensitive enough to detect functionaldifferences an FFA was found but no PPA Whencombined with the anatomical results this stronglysuggests that GR has no PPA
Because of safety concerns (presence of a femo-ral artery aneurysm clip) we did not perform a simi-lar high-field functional MRI of CO However asFigure 4 demonstrates CO clearly has damage tothe parahippocampal-lingual region in the righthemisphere
EXPERIMENTAL INVESTIGATIONS
In this section we report the results of four experi-ments designed to probe the consequences of dam-age to the lingual-parahippocampal region ingeneral and the PPA in particular Our primaryconcern in these experiments was to test GR andCOrsquos ability to perceive remember and recognisedifferent kinds of visual material Based on our pre-vious fMRI work we predicted that GR and CO
would have more difficulty with scene-like stimuli(ie stimuli that convey information about thegeometry of a space one can navigate through or actwithin) than with object-like stimuli Such a deficitcould account for at least some of the problems GRand CO encounter in real-world navigation
Beyond determining the kind of visual materialthat gets processed in parahippocampal cortex wealso wanted to determine the nature of the process-ing that takes place on that material In particularwe wanted to determine if parahippocampal cortexis involved in perceptual encoding memory encodingor recognition As discussed earlier (Introduction)our fMRI results provided some evidence againstplace recognition as the sole function of the PPAbut did not allow us to distinguish between percep-tual and mnemonic encoding Experiments 1 and 2examine the role of parahippocampal cortex in per-ceptual encoding Experiment 3 examines its role inmnemonic encoding and Experiment 4 examinesits role in place recognition
Experiment 1 Two-dot task
The first hypothesis we examined was thatparahippocampal cortex plays a role in perceivingthe shape of the local environment In our previousfMRI work we observed that the PPA respondsmuch more strongly during scene viewing than dur-ing object or face viewing even when subjects donothing more than watch the scenes passively with-out performing any other task This suggested thatscene perception alone may be sufficient to activatethe PPA Previous reports have indicated that dam-age to parahippocampal cortex and other medialtemporal regions leads to problems with memoryrather than perception (Bohbot et al 1998Maguire Burke Philips amp Staunton 1996 Pigottamp Milner 1993) However all of these previousresults were obtained on patients who had unilat-eral lesions GR has a bilateral occipital-temporallesion including clear damage to parahippocampalcortex in both hemispheres furthermore he reportssome kind of perceptual deficit We hypothesisedthat this deficit might involve difficulty perceivingor interpreting the geometry of scenes and otherstimuli with a similar geometric structure
EPSTEIN ET AL
494 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 5 Top row shows a single slice from a functional MRIscan performed on GR Bottom row shows results from a normalsubject (NK) for comparison Voxels that responded more to facesthan objects (left column) or more to scenes than faces and objects(right column) in a Kolomogorov-Smirnov test are highlighted bywhite arrows GR has a functioning face area (FFA) but there isno evidence of a place area (PPA)
FFA PPA
GR
NK
In this experiment subjects viewed either objectshapes or spatial layouts Stimuli consisted ofblack-and-white photographs of real-world stimuli(empty rooms vs common objects) or abstractstimuli made out of Lego blocks (Lego ldquoscenesrdquo vsLego ldquoobjectsrdquo) These stimuli have been previouslyshown to differentially activate the PPA in normalsubjects (Epstein et al 1999 Epstein ampKanwisher 1998) Two red dots were superim-posed on each photograph (Figure 6) The task wasto report which of the two red dots was over thecloser part of the object or scene This task wasdesigned to have no memory component (apartfrom remembering the instructions) but to requireaccurate 3D perception of objects and scenes
If GR or CO have difficulty perceiving or inter-preting the spatial layout of scenes then we pre-dicted they would do worse when the stimuli areempty rooms or Lego scenes than when they arereal-world or Lego objects Note that the criticalcomparison is between the Lego scenes and theLego objects as these stimuli are made of the samematerials and differ primarily in their surfacegeometry
MethodTesting for all experiments was done on aMacintosh computer The experiment consisted of80 trials (20 of each stimulus type randomly inter-mixed) Subjects began each trial by pressing thespace bar A black-and-white photograph from oneof the four stimulus categories (see earlier) thenappeared on the screen with two red dots on it Thetask was to determine which of the two red dotsindicated the closer part of the scene or objectSubjects pressed the ldquofrdquo key if the leftmost red dotwas closest and the ldquojrdquo key if the rightmost dotwas closest They were instructed to respond asquickly as possible without making errors Stimuliremained on the screen until a response was made
The stimulus set was constructed so that eachempty room was matched to a common object thathad red dots in the exact same location and eachLego layout was similarly matched to a Lego objectThus the locations of the red dots were counterbal-anced across scene and object stimuli controllingfor any effects of visual field cuts
Results and discussionResults for GR and CO and four elderly normalsubjects are shown in Table 3 Both GR and COcould do this task without difficulty and their per-formance was near ceiling There was no evidencethat they had any more trouble with the emptyrooms or Lego scenes than with the real or Legoobjects Reaction times were slightly longer for GRand CO than for normal subjects as one wouldexpect given their visual field deficits however this
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 495
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 3 Experiment 1 (two-dot) results (SD in parenthesis)
GR CO Normals
Proportion correctObject 095 080 093Lego object 100 095 098Empty room 100 100 099Lego room 095 100 096
Mean RTObject 2435 2932 2161 (980)Lego object 2052 3245 2029 (921)Empty room 2744 2291 1788 (959)Lego room 1904 2281 1630 (1127)
Figure 6 The four kinds of stimuli used in Experiment 1 Topleft lego layouts top right real-world layouts (empty rooms)bottom left lego objects bottom right real-world objects
difference did not appear to be significant Impor-tantly there was little evidence that GR or CO tooklonger to make their perceptual judgements for thereal or Lego scenes than for the real or Lego objectsThus contrary to our initial hypothesis this experi-ment found no clear evidence for selective impair-ment in the perception of scene-like stimuli
Experiment 2 Different-views matchingtask
The previous experiment demonstrated that GRand CO can extract information about the per-ceived distances between themselves and the sur-faces of an object or scene Thus in at least onesense their perception of these surfaces is pre-served However this does not mean that their abil-ity to represent the layout of a scene is unimpairedThe two-dot task requires on-line comparison oftwo surfaces in egocentric coordinates GR and COwould be able to perform this task even if they couldnot (1) represent more than two surfaces at a time(2) represent surfaces in other than strictly egocen-tric coordinates or (3) sustain surface representa-tions for longer than the time it takes to make acomparison It has been suggested that the parietallobes are involved in on-line processing of surfacegeometry in egocentric coordinates (Milner ampGoodale 1995) thus it is possible that the parietallobes (which are intact in GR and CO) may be suf-ficient to perform the two-dot task even in theabsence of more durable or complex surface repre-sentations in parahippocampal cortex
In Experiment 2 we examine GR and COrsquosability to create and sustain a representation of thegeometry of an object or scene-like layout The taskwas to view a photograph of a Lego object or Legoscene hold it in memory for 10 s and then deter-mine whether a second object or scene was the sameas the first In ldquosamerdquo trials the second photographdepicted the same scene or object depicted from adifferent viewing angle In ldquodifferentrdquo trials the sec-ond photograph depicted a different scene or objectchosen to be similar to the first one (Figure 7)Thus in order to successfully perform this tasksubjects had to form a representation of the surfacesof the object or scene sustain this representation in
memory for 10 s and then align it (presumablyusing mental rotation) to the second object orscene
MethodThe experiment consisted of two parts each with40 trials In the first part the stimuli were black andwhite photographs of objects made out of Legoblocks In the second part the stimuli were photo-graphs of ldquoscenesrdquo made out of Lego blocks whichwere designed to have a geometric structure similarto real-world places such as a roomor a city street
Subjects began each trial by pressing the spacebar A photograph of a Lego object or sceneappeared on the screen for 3 s followed by a 10-sblank interval A second photograph then appeareddepicting either (1) the same object or scene as thefirst photograph shown from a different viewingangle (offset approximately 35 degrees) or (2) a dif-ferent object or scene Subjects were instructed topress the ldquojrdquo key if the two photographs depicted thesame object or scene and the ldquof rdquo key if they did notThey were told to take as much time as they neededto make a decision but no extra time The secondphotograph remained on the screen until a responsewas made Half the trials were ldquosamerdquo trials and halfwere ldquodifferentrdquo trials
Results and discussionFigure 8 shows results for GR and CO as well as forsix elderly normal controls GRrsquos and COrsquos perfor-mance was comparable to that of the normal con-trols Furthermore GR and CO performed just aswell when the stimuli were Lego scenes as whenthey were Lego objects These results are unlikely toreflect a ceiling effect as performance for bothpatients and controls was around 80 correctThus both patients appear to be able to form a rep-resentation of the geometry of both scenes andobjects which they can sustain in short-termmemory
Experiment 3 N-back recognition memorytask
The previous experiment failed to find any impair-ment in GR and COrsquos ability to form a representa-
EPSTEIN ET AL
496 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
tion of the geometry of a scene or object andmaintain it for 10 s in the absence of the originalstimulus Not only does this result suggest that per-ception of scene-like stimuli is intact in thesepatients it further suggests that some short-termmemory functions are intact as well However we
should note that subjects in this experiment werefree to devote all their effort and attention in the 10-second blank interval to maintaining a memory ofthe stimulus Thus it is possible that GR and COcan use rehearsal mechanisms to maintain the stim-ulus in short-term memory during the blank inter-
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 497
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
(b) Lego object version
Figure 7 Stimuli and procedure for Experiment 2 (a) In the Lego layout version subjects see a Lego layout for 3 s followed by a blankscreen for 10 s A second layout then appears which is either the same layout as the first one but seen from a different angle or a differentlayout (b) The Lego object version is the same but with object stimuli
3 s 10 s OR
3 s 10 s OR
(a) Lego layout version
(b) Lego object version
val but are unable to form a more durable memorytrace for the stimulus This possibility was tested inthe current experiment which was designed toexamine GR and COrsquos ability to learn new visualinformation
We devised a continuous n-back recognitionmemory test in which subjects viewed a sequence oforiginally novel stimuli some of which wererepeated within the sequence at intervals of 1 3 or5 intervening images For each stimulus subjects
reported whether they had seen it before in theexperiment or not In different runs in the experi-ment the stimuli were black-and-white photo-graphs of (1) Lego scenes (2) Lego objects (3) real-world landscapes (4) novel objects made out ofSculpey clay (5) unfamiliar houses (6) unfamiliarfaces as well as (7) line drawings of nonsenseobjects and (8) pronounceable pseudowords (Fig-ure 9) Thus we were able to test GR and COrsquos rec-ognition memory for different kinds of materials atdifferent time intervals in a paradigm in which thepresence of intervening items between the twoappearances of each stimulus made rehearsal diffi-cult Note that because none of the specific stimuliused in this experiment were familiar in advancethis task did not require an episodic memory com-ponent and could be done on the basis of pure visualfamiliarity with the particular visual stimulus
Although many different stimulus classes weretested in this experiment the critical comparison isbetween the Lego layouts and Lego objectsbecause these are best matched with regard tomaterials and differ primarily in their geometricstructure (see Introduction) We predicted that ifGR and CO showed a memory deficit for this taskit would be greater for the Lego layouts than for the
EPSTEIN ET AL
498 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 8 Results of Experiment 2 Performance over ldquosamerdquotrials and ldquodifferentrdquo trials is averaged together Error bars show1 SD
Figure 9 Stimuli for Experiment 3 Top row Lego layouts real-world scenes houses Bottom row Lego objects clay objects nonsenseobjects faces In an eighth condition (not shown) subjects viewed pronounceable nonwords such as ldquowilchrdquo or ldquothipperrdquo
Lego objects In addition a deficit for materialsthat convey information about the layout of sur-rounding space might also be apparent when GRand CO are tested on more realistic stimuli If soone would expect them to do poorly on the land-scapes but relatively well on the clay objects linedrawings of nonsense objects faces andpseudowords
MethodFor each subject a complete experimental sessionconsisted of being run once on each of the eightstimulus classes Each run consisted of 90 trials ineach trial a stimulus was presented and subjectsreported whether or not they had seen it before inthe run (or indeed ever) by pressing the ldquojrdquo key ifthey had or the ldquof rdquo key if they had not Each stimu-lus was on the screen for either 1 2 or 3 s (see fol-lowing) and appeared promptly after the keypressreport for the previous trial Individual stimuliappeared either once (14 stimuli) twice (30 stim-uli) or four times (4 stimuli) within a given run Ofthe stimuli that were presented twice 10 wererepeated immediately (1-back) 10 were repeatedwith a lag of two intervening items (3-back) and 10were repeated with a lag of four intervening items(5-back) In order to ensure that between-subjectdifferences in performance were not due to stimulusdifferences the same stimuli were assigned to thesame conditions each time the experimentwas run
GR performed the experiment twice once inAugust 1998 and again 9 months later Because ofa computer error he was only tested on six of theeight categories (all except Lego layouts and Legoobjects) at the later date CO was also tested twiceonce in February 1999 and again 6 months laterReported results for GR and CO are the average oftheir two testing sessions Normal subjects wereonly tested once Stimulus presentation time was 1 sin GRrsquos first testing session In pilot testing COfound this too fast for the critical categories of Lego
layouts and Lego objects so presentation time wasextended to 2 s for these categories in his first test-ing session All normal subjects were tested at thesepresentation rates (2 s for Lego layouts and objects1 s for other stimuli) In COrsquos second testingsession presentation time was lengthened to 3 s forall stimulus categories to ensure that any deficitsfound were memory deficits rather than problemswith the nonmemory aspects of the task
Results and discussionResults from GR CO and 10 elderly normal con-trols are shown in Figure 10 and Table 4 Figure 10ashows results from the main comparison betweenLego layouts and Lego objects Both normal sub-jects and patients performed near ceiling for bothLego layouts and Lego objects at 1-backinterstimulus intervals verifying that GR and COcould successfully perceive the pictures as they werebeing presented In contrast there was a strikingdifference between the two patients and the normalsubjects at 5-back intervals Whereas normals didon average somewhat better on Lego layouts thanLego objects both GR and CO showed the oppo-site pattern doing much worse on Lego layoutsthan Lego objects The 3-back condition showed amixed pattern Whereas GR did better on Legoobjects than Lego layouts in this condition (similarto his performance at 5-back intervals) CO didbetter on Lego layouts than Lego objects COrsquos lowperformance on Lego objects in the 3-back condi-tion appears to be an anomaly as it is much lowerthan his performance for comparable stimuli in the1-back and 5-back conditions
Figure 10b shows the difference in performancebetween Lego layouts and Lego objects for thecritical 1-back and 5-back intervals At 5-backintervals CO was significantly impaired comparedto the normal subjects (z = ndash250) whereasGRrsquos impairment fell just short of significance (z =ndash175)4
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 499
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
4In fact these values may underestimate GR and COrsquos relative impairment for Lego layouts In the 5-back condition 4 of the 10
normal subjects had perfect performance on the Lego layouts but all scored below ceiling on the Lego objects Thus the true advantagefor Lego layouts over Lego objects for normal subjects is probably greater than that observed here in which case GR and COrsquos lowerperformance for Lego layouts compared to Lego objects would be even more abnormal Furthermore post-hoc analyses revealed thatone of the normal subjects showed a difference in performance between Lego layouts and objects that was over 2 SD below the mean ofthe other nine subjects When this outlying subject was excluded z-scores were ndash322 for CO and ndash232 for GR
Table 4 shows results for all eight stimulusclasses Although direct comparisons between sub-jects should be treated with caution since viewingtime for different items was not the same for eachsubject we do note a few interesting patterns Firstof all there is little or no evidence that GR or CO
show a memory deficit for nonsense words clayobjects or line drawings of nonsense objects GRappears to be slightly impaired on faces and housesbut CO performs normally on these categoriesStrikingly GR shows a severe deficit for the reallandscapes performing more than 40 percentagepoints lower than normals Thus even thoughthere is some evidence that GR might have someimpairment for nonscene stimuli he clearly is muchmore impaired for scene-like stimuli such as theLego scenes and landscapes CO does not show asimilar deficit for the landscapes However itshould be kept in mind that in one of his two testingsessions CO viewed each picture for 3 s which wasmuch longer than the 1 second used for all non-Lego stimuli for GR and normals Thus COrsquosapparently ldquonormalrdquo performance in these condi-tions might be overestimated
Taken together the results of this experimentindicate that GR and CO are selectively impaired atprocessing scene-like stimuli This deficit is consis-tently apparent at 5-back but not 1-back intervalsThis finding suggests that the deficit should becharacterised as a memory encoding problemHowever the present results do not allow us todetermine whether the difficulty occurs at memoryencoding or at subsequent recognition (see Epsteinamp Kanwisher 2001) If GR and CO have a generalproblem with recognition of topographical materi-als then they should not be able to recognise scenes
EPSTEIN ET AL
500 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 10a Results of Experiment 3 for GR (top) CO (middle)and for 10 age-matched normal controls Performance measure isproportion of repeated stimuli correctly identified as such correctedfor guessing using the standard formula
Figure 10b Difference between Lego layout performance andLego object performance for the 1-back and 5-back conditionsError bars indicate one 1 SD
and landmarks that were learned prior to theirinjury We tested this possibility in the nextexperiment
Experiment 4 Famous landmarksfacesrecognition task
In this experiment we examined GR and COrsquosability to recognise the visual appearances of placeslearned prior to injury Subjects saw a series offamous landmarks intermixed with nonfamouscontrols (Imaging experiments have demonstratedthat landmarks of this type can activate the PPAalmost as strongly as complete scenes Epstein etal 1999 Epstein amp Kanwisher 1998) They wererequired to say whether they knew each specificlandmark and to identify the ones they knew Sub-jects were instructed to respond affirmatively if theywere fairly certain that they recognised the land-mark but not if it only looked generally familiarFor purposes of comparison subjects also per-formed an identical test in which the stimuli werefamous and nonfamous faces In both testsdistractor items were designed to be visually andconceptually similar to the target items (see Figure11) If parahippocampal cortex is required for placerecognition then one might expect GR and CO toperform poorly compared to normals on the land-mark recognition test
MethodEach of the two experiments (famous faces famouslandmarks) consisted of 48 trials Subjects beganeach trial by pressing the space bar after which ablack-and-white photograph of a face (in thefamous faces experiment) or a landmark (in thefamous landmarks experiment) appeared on thescreen Subjects were told to press the ldquojrdquo key if theyrecognised the specific face or landmark and the ldquof rdquokey if they did not taking as much time at theyneeded to make this discrimination but no extratime Subjects were told to report that they recog-nised a face or landmark if they felt that they knewit even if they could not retrieve its name If theydid know the name they were to say it after pressingthe button Otherwise they were to say whateverthey did remember about the item
Half of the photographs were of famous and halfof nonfamous people and landmarks The famousfaces included public figures (eg George BushRichard Nixon Prince Charles) and movie stars(eg Harrison Ford Clark Gable Marlon Brando)who were prominent prior to the time of GR andCOrsquos injuries the famous landmarks were build-ings and monuments from all over the world (egEiffel Tower US Capitol Building Great Sphinx)The nonfamous stimuli were chosen to match thefamous stimuli as closely as possible Each famousstimulus had a ldquomatchrdquo For example George Bushwas matched to a man of similar age in a coat andtie and the Taj Mahal was matched to a visually
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 501
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 4 Full results of Experiment 3 Mean proportion correct (corrected for guessing) SDs in parentheses
Lego Land- Lego Clay Nonsense Non-scenes scapes Houses objects objects objects Words Faces
1-back GR 100 094 100 100 100 094 100 100CO 091 085 091 100 100 079 074 100Normals 096(008) 095(008) 093(014) 096(008) 096(006) 099(003) 098(004) 097(011)
3-back GR ndash003 068 026 047 083 083 075 052CO 066 081 037 010 094 069 095 081Normals 066(032) 077(021) 072(015) 077(020) 086(016) 083(015) 083(017) 072(020)
5-back GR 010 034 044 047 072 083 094 053CO 031 073 075 088 081 064 100 081Normals 076(026) 077(016) 068(021) 066(016) 091(014) 083(018) 093(011) 078(021)
similar Asian palace that is not famous (see Figure11)
Results and discussionResults for GR and CO as well as for five age-matched controls are shown in Figure 12 Both
GR and CO could recognise the famous landmarksas well normals In addition both GR and CO wereusually able to name the landmarks they success-fully identified
Thus GR and CO can recognize landmarkslearned prior to injury demonstrating that at leastsome of their place recognition abilities are intactWhen combined with the fact that GR and COwere able to produce detailed maps of places theylived in prior to their injury (see Case Description)these results suggest that their deficit is primarily inencoding or consolidation of new informationrather than in recognition or recall of old informa-tion Note however that these results do not provethat their place recognition abilities are entirelynormal GR and CO might suffer from a mild rec-ognition deficit that affects identification of newly-learned stimuli like the Lego layouts in Experiment3 but not overlearned stimuli like the famous land-marks Furthermore it is possible that GR and COmight be impaired at recognising premorbidlylearned places that are defined primarily by theirspatial layout even though they are unimpaired atrecognising premorbidly learned places that aredefined primarily by a single object-like landmarkor building (see General Discussion) Neverthelessthe trend of the present evidence is to suggest thatGR and CO have particular problems with noveltopographical information
In the famous-face task GRrsquos performance wasin the normal range However he sometimes haddifficulty naming the faces even though he clearly
EPSTEIN ET AL
502 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 11 Stimuli for Experiment 4 Each famous landmark(Arc de Triomphe Taj Mahal) and famous face (Barbara BushYasser Arafat) was matched to a nonfamous building or face thatalso appeared in the experiment
FA
CE
SL
AN
DM
AR
KS
Figure 12 Results of Experiment 4 Performance measure isproportion of famous stimuli reported as ldquoknownrdquo corrected forguessing using the standard formula
recognised them For example he described Ron-ald Reagan as ldquothe gipperrdquo Nancy Reagan as ldquothewife of the gipperrdquo and Clark Gable as ldquothe movieactor who said lsquoScarlett I donrsquot give a damnrsquordquo COhad a great deal of difficulty identifying the famousfaces This result is not surprising given hisextremely low performance on the Benton FaceMatching Task In addition to his topographicproblems CO appears to suffer fromprosopagnosia
GENERAL DISCUSSION
We tested two parahippocampal patients (GR andCO) on four experiments designed to measure per-formance as a function of both task (perceptionencoding and recognition) and stimulus type(scenes objects faces and words) The only taskthat revealed a clear deficit was an n-back recogni-tion memory test (Experiment 3) In order to suc-cessfully perform this task subjects had to encode adurable representation of a visual stimulus and useit for later recognition GR and CO were signifi-cantly more impaired on this task when the stimuliwere scene-like spatial layouts (ie Lego scenesand real scenes for GR) than when they wereobjects No comparable deficits were found in otherexperiments that tested layout perception (Experi-ments 1 and 2) and placelandmark recognition(Experiment 4) Taken together these results indi-cate that GR and CO are impaired in their ability toencode novel information about the geometry ofsurrounding space into memory but are lessimpaired at their ability to encode the geometry ofnovel objects
We argue that this specific topographical defi-citmdashinability to encode the spatial layout of novelscenesmdashresults from damage to the para-hippocampal place area Although the damage inboth patients extends beyond the parahippo-campal-lingual territory normally associated withthe PPA this region is clearly lesioned bilaterally inGR and in the right hemisphere in CO Further-more functional imaging demonstrated that GRhas no functioning PPA Thus the current resultsand the earlier fMRI work (Epstein et al 1999
Epstein amp Kanwisher 1998) converge to implicateparahippocampal cortex in the encoding of infor-mation about scene geometry into memory
Our assignment of the function of memoryencoding to parahippocampal cortex is consistentwith the results of earlier case reports of patientswith parahippocampal damage who have naviga-tional difficulties in novel but not familiar environ-ments (Habib amp Sirigu 1987 Ross 1980 see alsoTeng amp Squire 1999) As with GR and CO thesepatients were either less impaired or unimpaired onnontopographical memory tasks Patients have alsobeen described who exhibit the opposite pattern ofimpairment an inability to learn faces (sometimescalled ldquoprosopamnesiardquo Cipolotti Robinson Blairamp Frith 1999 Tippett Miller amp Farah 2000) orfaces and verbal materials (Maguire amp Cipolotti1998) with an unimpaired ability to learn topo-graphical materials Taken together these resultsargue for a high degree of domain-specificity in theneural systems involved in learning of new informa-tion (Gallistel 1995) The brain may have separatemechanisms for learning faces places and wordswhich can be differentially impaired after damageto specific cortical regions (Cipolotti et al 1999Incisa della Rochetta et al 1996) The presentresults suggest that the anatomical locus of themechanism dedicated to learning new topographi-cal information is in the parahippocampal cortex(Aguirre amp DrsquoEsposito 1999)
In contrast to their impaired ability to encodenew topographical information both GR and COwere able to produce detailed maps of their previousdwellings and performed normally on the famouslandmark recognition test of Experiment 4 Theseresults are generally consistent with Teng andSquirersquos (1999) report of a patient with extensivemedial temporal damage who could recallpremorbidly learned but not postmorbidly learnedtopographical information (see also Ross 1980)Thus our results provide further evidence thatmedial temporal regions such as parahippocampalcortex may be more critical for encoding or consoli-dating new information into memory than for recallof old information
One issue that our experiments do not entirelyresolve is the precise nature of the topographical
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 503
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
information encoded by parahippocampal cortexClearly this region is critical for encoding informa-tion about the geometry of the currently visiblescene even when the ldquoscenerdquo is made out of Legoblocks and therefore has no immediate navigationalrelevance However it is unclear whether this spe-cific deficit can account for all of the real-worldnavigational difficulties encountered by GR andCO GR in particular shows such a catastrophicloss of navigational ability that we suspect he is alsoimpaired at encoding the spatial relationshipsbetweenthe current scene and other locations in theworld CO probably also suffers from such difficul-ties as he reports that he has trouble rememberinghow places relate to each other in large-scale spaceWhen he was tested on a simple 3-room videogameenvironment he not only was unable to rememberthe individual rooms (claiming that there were 10rooms when there were only 3) but he also had nomemory for the spatial relationships between themThese spatial deficits may be the result of damage tothe posterior right hippocampus (in the case of CO)or of disconnection of the posterior right hippo-campus from the parahippocampal regions thatprovide much of its cortical input (in both patientsSuzuki amp Amaral 1994) In either case the abilityto form a cognitive map of new environments likethe lab or videogame space could be impaired(Maguire et al 1998 OrsquoKeefe amp Nadel 1978)However it is also possible that parahippocampalcortex itself may be critical for encoding not onlythe geometry of individual scenes but also the spa-tial relationships between the locations of thosescenes In this case the spatial deficits of GR andCO could be attributed directly to parahippo-campal damage rather than to hippocampal dam-age or disconnection
The latter scenario would be consistent withAguirre and DrsquoEspositorsquos (1999) and Barrash etalrsquos (2000) suggestion that parahippocampal cortexis critically involved in combining visual informa-tion about the appearance of scenes and landmarkswith spatial information about their locations into aunified topographical representation Howeverboth of these authors argue that the representationsof individual scenes (Barrash et al) and landmarks(Aguirre amp DrsquoEsposito) are supported not by
parahippocampal cortex but by more posterior lin-gual-fusiform regions In contrast we believe thatthe neuroimaging data indicate that parahippo-campal cortex is involved in the encoding of thegeometric structure of individual scenes althoughit may also play a role in combining these represen-tations with other spatial representations
Interestingly GR and CO were not impaired onrecognition of famous landmarks that were mostlybuildings Thus our data do not preclude the possi-bility that there may be a separate lingual orfusiform region involved in processing informationabout buildings (Aguirre Zarahn amp DrsquoEsposito1998 Chao Haxby amp Martin 1999 IshaiUngerleider Martin Schouten amp Haxby 1999)which might have been preserved in GR and COdespite extensive damage to other regions of thelingual and fusiform gyrus Alternatively GR andCOrsquos preserved building recognition abilities mightbe mediated by interactions between cortical areasthat perform shape analysis for any object-likestimulus type (Malach et al 1995Kanwisher et al1996) and anterior temporal regions in which long-term topographical and semantic memories arestored (McCarthy et al 1996)
Is perception preserved
In contrast to their inability to encode new scenesinto memory GR and CO have substantiallyretained their ability to perceive the spatial layout ofthe currently visible scene (Experiments 1 and 2)GR can follow a route marked on a floor planwhich indicates a good on-line appreciation of hiscurrent spatial surroundings Furthermore neitherGR or CO have any trouble moving through theenvironmentmdashthey move with confidence and donot bump into doors or walls Problems of the lattertype are usually associated with damage to the pari-etal lobes which are intact in GR and CO Thus atfirst glance GR and CO do not appear to haveperceptual impairments (beyond their visual fieldrestrictions)
However GR does complain of a perceptualdeficit which he describes as an inability to ldquotakeinrdquo scenes when they are too complex Similarreports of an inability to comprehend the
EPSTEIN ET AL
504 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
ldquoglobalityrdquo of a scene were made by some of theparahippocampally-damaged patients studied byHabib and Sirigu (1987) We were not able todetermine the precise nature of this deficit whichmay be partly attributable to his visual field restric-tions It appears that his deficit is more salient in thereal-world environments than in the picture-per-ception tasks of Experiments 1 and 2 perhapsbecause he finds it more difficult to organise visualinput when immersed in it than when viewing pho-tographs that subtend only part of the visual field
We suggest that GR can perceive parts of thescene when he attends to them but may be unableto integrate information over time into a coherentrepresentation of a scene or place The fact that GRand CO are impaired for Lego layouts at 5-backintervals in Experiment 3 is consistent with thishypothesis The time between the first and secondpresentations of a stimulus is very short in this case(and even shorter with 3-back intervals where GRalso shows evidence of a deficit) In both patientsthe representation of layout information formed byperception of a Lego scene must either have anabnormally short persistence be abnormally sus-ceptible to interference or both If so they mightfind it impossible to sustain a spatial frame orschema long enough to use it to organise informa-tion obtained from many different glances at a com-plex scene (Intraub 1997) The result would be aninability to understand the ldquoglobalityrdquo of a sceneeven though all the details were perfectlyperceived
Non-topographical mnemonic abilities
Although not as severe as their topographical mem-ory problems GR and CO do have some memorydifficulties in other domains For example bothperformed poorly on the Rey AVLT whichrequired them to remember a list of unrelatedwords Both also report some subtle episodic mem-ory problems (see Case Description) COrsquos prob-lems could partly result from damage to the righthippocampusmdashindeed CO appeared to have themore acute episodic memory problems HoweverGR has no hippocampal damage so the fact that hedisplays episodic memory problems suggests thatparahippocampal cortex is involved in the forma-
tion of episodic memories (cf Wagner et al1998)
In previous work (Epstein et al 1999) weargued that spatial context information encoded byparahippocampal cortex might play a role in estab-lishing event memories by encoding informationabout where events take place The inability toencode this information might cause problems intasks such as the Rey AVLT where one has to recalla list of unrelated words If spatial (and perhapstemporal) context information is not encoded itwill be harder subsequently to recall the distinctevent of hearing each word (Tulving 1983) Incontrast lack of spatial context information wouldbe less of a problem in tasks where one is onlyrequired to recognise previously encountered verbalmaterials but not to reproduce the spatiotemporalcontext in which they were experienced IndeedGR and CO performed as well as normals on thenonsense words in Experiment 4 where recogni-tion memory but not episodic memory was requiredfor successful performance Interestingly GR andCO also did relatively well on the Logical Memorysubtest of the WMS-III perhaps because the nar-rative framework of a story provides sufficient con-textual information to overcome the absence ofspatial context and mediate successful verbalretrieval
Differences between GR and CO
Although we have discussed GR and CO togetherthere are a number of relevant differences betweenthem One open question is whether these differ-ences can be attributed to the different effects ofunilateral and bilateral parahippocampal damageIn general GRrsquos topographical difficulties weremore severe than COrsquos CO did surprisingly well onthe Warrington Topographical Memory Test inwhich he was required to remember detailed real-world city scenes and also performed substantiallybetter on real scenes than Lego scenes in Experi-ment 3 Insofar as CO has intact left para-hippocampal cortex this suggests that the left PPAmight be sufficient to encode a certain amount of(perhaps semantic) information about real-worldscenes but the right PPA is necessary to encode
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 505
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)

tested him over a period of 5 days in August 1998with 3 days of follow-up testing 3 weeks later and 1day of additional tests in May 1999 During thistime he was alert and cooperative Emotionalaffect and language use was normal He interactednormally in nonclinical social situations and hisconversation was appropriate and varied Heremembered the events of previous days andrecounted them accurately
In contrast to this high level of non-topographical functioning GR demonstrated adramatic inability to orient himself in space Forexample even after several days of testing he neverlearned the relative locations of the different roomsof the lab According to both GR and his wife thisinability to learn new topographical informationwas typical of his experience since his strokes as hefrequently gets lost in his daily life Soon after his
injury he moved to a neighbourhood with manysimilar-looking houses He reported that in orderto find his new house after a walk to a market six toseven blocks away he had to rely on street signs toguide him to the correct block and then examineeach house on the block in detail until he couldrecognise some feature that distinguished his homeuniquely Subsequently GR and his wife moved toa different house in a different country He reportedthat for the first 6 months after the move his newhome was like a ldquohaunted houserdquo for him insofar ashe was unable to learn his way around it For exam-ple he would repeatedly forget that the bedroomhad a balcony and he would be surprised by this factevery time he rediscovered it After several monthshowever he was able to learn the topography of thehouse sufficiently well to navigate through it (butsee below)
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 485
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 2 Summary of neuropsychological tests ( indicates percentile scores)
GR COmdashmdashmdashmdashmdashmdashmdashmdashmdashmdash mdashmdashmdashmdashmdashmdashmdashmdashmdashmdashmdashndash
IQ 122 (93 WAIS-R) 134 (99 WAIS-III)
WMS-III
Logical memory 25 immediate recall 27 immediate recall50 delayed recall 48 delayed recall(WMS-R)
Face memory 9 immediate recall 16 immediate recall16 delayed recall 9 delayed recall
Family pictures 2 immediate recall lt1 immediate recall5 delayed recall lt1 delayed recall
Rey Auditory Verbal 63 immediate recall 45 immediate recallLearning test 14 after interference list lt1 after interference list
16 delayed recall 3 delayed recall
Warrington Recognition 50 wordsMemory tests 29 scenes
lt5 faces
724 Spatial Recall 66 immediate recall2 delayed recall
Benton Face Recognition 42 lt1
Memory span Digits99 forward99 backward
Spatial37 forward50 backward
Boston Naming test Ceiling Ceiling
Case 2 COCO is a left-handed male with 20 years of educa-tion He was 60 years old at the time of testing Twoyears prior to testing he suffered a right posteriorcerebral artery stroke He first noticed on a week-end that his vision appeared blurred on the left andthat he had a tendency to bump into objects on thisside On Monday morning he was able to success-fully negotiate the route to his workplace a trip thatinvolved a bus ride followed by two subways andthen a shuttle to his office building However oncehe entered the building he found himself com-pletely disoriented unsure of its layout and wherehis office was located Interestingly the companyhe worked for had moved a few weeks earlierAlthough he had managed to arrive at this newbuilding (which was a block away from the previouslocation) he became lost upon entering it Hisneurologic exam revealed a left homonymoushemianopia but no evidence of neglect on a lettercancellation task or on sensory testing Investiga-tions revealed an ischaemic stroke with haemor-rhagic transformation in the right occipital andmesial temporal lobe On magnetic resonanceangiography he had a chronic left carotid occlusiona mild right carotid stenosis and bilateral vertebralartery stenoses Since then he has suffered fromsevere topographical disorientation primarily innew environments
CO reports that he can recognise buildings thatwere familiar to him before the stroke He can alsoform mental images of well-known landmarks suchas the Empire State Building and the BostonPublic Gardens and of the house he lived in as achild When asked to imagine standing in front ofthe Massachusetts State House (a location wellknown to him from years of living in Boston) heaccurately described the surrounding buildings andtheir locations However he reports that when helooks at a place that hersquos encountered post-strokeall he gets is a feeling of familiarity that does notallow him to determine where the place is or whenhe saw it He also reports an inability to understandhow various landmarks relate to each other inlarge-scale space Despite this difficulty he canfollow a map and frequently rides the Bostonsubway alone
Unlike GR CO was able to learn a certainamount of topographical information For exam-ple he soon learned the location of the rooms in thelab and would travel between them unassistedAfter his first visit to the lab he needed to be guidedall the way back to the subway station but after sev-eral visits he could find his way out of the buildingand to the subway on his own as long as he was firstguided to the elevators (It should be noted how-ever that the building is on the same block as thesubway station Furthermore he still needed to bedirected to the elevators even though our buildinghas a relatively simple floor plan) Thus the amountof topographical information he can learn is limitedbut not zero
COrsquos language and reasoning skills appear to benormal and his conversation is appropriate andvaried In addition to his topographical difficultieshe also has limitations of his peripheral vision onthe left side
Navigational abilities
In addition to the experiments reported in theExperimental Investigations section of this paperwe also performed a number of ad hoc tests of GRand COrsquos navigational abilities Although thesetests were informal we describe them here to pro-vide a general sketch of the nature of GR and COrsquosimpairments
Real-world topographical learningOver the course of several days we took GR and hiswife on three walks around the MIT campus whichneither had visited previously Each walk followedexactly the same path At several places in the walkwe would stop and point out different buildings andlandmarks tell them something about each one andask them to remember their appearance for latertesting Immediately after the completion of thethird walk both GR and his wife separately per-formed a forced-choice recognition test in whichthey saw photographs of 14 landmarks (mostlybuildings) from the walk and 14 novel landmarks(from another part of the MIT campus that theyhad not visited) and were required to reportwhether they had seen each one or not GRrsquos
EPSTEIN ET AL
486 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
performance was 714 correct hits for the familiarlandmarks and 1314 correct rejections for theunfamiliar landmarks In contrast his wifeobtained a perfect score on this test Given theextensive nature of the training regime GRrsquos scorestrongly suggests an impairment in learning theappearance of new real-world places Interestinglyseveral of the seven familiar landmarks that he gotright had an obvious distinctive feature that hadbeen explicitly pointed out to him For exampleone building had a distinctive clock tower andanother one was an unusual triangular shape ThusGR was occasionally able to use distinct visualfeatures to identify places even though he wasunable to recognise them holisticallymdasha commonpattern among topographical disorientationpatients
GR did not appear to have any obvious problemsin understanding the spatial structure of his imme-diate environment When given a floor plan of thebuilding at MIT in which he was being tested hewas able to successfully follow a route marked onthe plan However when the plan was taken awayhe had great difficulty finding his way back to thelab even though he had more than 30 min experi-ence with the environment and his starting positionwas less than 30 feet from the goal (Figure 2) Infact the route he took resembled a random walk Atone point he walked right past his destinationwithout recognising it and only knew for sure thathe was in the right place when he identified his wifein the room Thus his on-line appreciation of thelayout of immediately visible space appears to beintact but he cannot find his own way withoutassistance
We were unable to perform the MIT landmarkrecognition test on CO because he suffered from amedical problem that made it difficult for him towalk around campus In lieu of this we tried to seewhether he could learn a simple video game envi-ronment consisting of three connected roomsAfter about 30 min experience navigating throughthis environment (via button presses) he was askedto sketch a map of it He declined to do so statingthat it was beyond his abilities When asked howmany rooms there were he replied that he did notknow but that there were at least 10 Given the
simplicity of this artificial environment COrsquos com-plete inability to learn it is striking
Map drawingAfter several days of testing we asked both GR andhis wife to sit at different tables and independentlysketch (1) the shape of Florida (2) a bicycle (3) afloor plan of their hotel room in Boston (4) a floorplan of the first floor of the house they had been liv-ing in for the last 8 months (all post-stroke) (5) afloor plan of the main room of the lab and (6) afloor plan of the house they lived in 10 years ago(pre-stroke) GR drew the map of Florida and thebicycle perfectly His plan of the hotel room wasnotably different from the one drawn by his wifemdashfor example he reported that there was only onebed when in fact there were two and he placed thebed on the wrong side of the room (Figure 1a) Hisplan of the first floor of his current house was some-what better but was not very detailed and includedsome major errors For example he wronglyreported that there was no bathroom on this floor(Figure 1b) He was unable to produce any floorplan of the main room of the lab leaving this pageblank When his wife reminded him that this wasthe room in which he had just spent an hour readingthe newspaper and in which we had all had break-fast in that morning he replied that he didnrsquot knowwhat room she was talking about (although heappeared to remember these events) In contrast tothis near total inability to produce accurate maps ofnew environments GR produced an accurate mapof the house he lived in prior to his injury (Figure1c) He reported that his memory of this house wasldquoall thererdquo and he could imagine walking around init When asked to produce a perspective of a roomwhen he was in it he did so with ease and he wasalso able to create an accurate floor plan of theroom
In contrast CO found it very difficult to pro-duce a floor plan of the lab both when he was in itand when he was taken to another room and askedto recall it However although the maps he pro-duced were quite impoverished they were notentirely inaccurate demonstrating some ability toencode topographical information This contrastswith GRrsquos absolute inability to recall anything
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 487
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
EPSTEIN ET AL
488 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure
1F
loorplansdraw
nby
GR
andhisw
ifeof(a)
theirhotelroomin
Boston(b)theircurrenthousew
hichthey
hadm
ovedto
subsequenttoG
Rrsquosinjury(c)the
housethey
livedin
priorto
GR
rsquosinjury
Ho
telro
om
inB
osto
n
GR
His
wife
His
wife
Cu
rren
th
om
e
GR
His
wife
Fo
rme
rh
om
e
GR
about the main room of the lab once he had left itCO was also able to draw a detailed floor plan of theapartment he was living in at the time (and hadlived in for 20 years) Several months after he hadmoved from this apartment we asked him to drawsuch a floor plan again and he produced one thatwas substantially the same as the first (Figure 3)Thus their map drawing performance indicatesthat GR and CO are primarily impaired at sketch-ing places experienced after their strokes but areunimpaired at sketching places experienced beforetheir strokes This pattern is indicative of deficits inencoding but not recall
Scene perceptionAs noted earlier GR can navigate with the aid of afloor plan suggesting that his perceptual abilitiesare largely intact Interestingly however he reportsthat he does have a perceptual deficit which hedescribes as an inability to understand scenes whenthey are too complex For example he reports thathe often has trouble using maps because theyusually are too busy with details It is unclearwhether this apparent perceptual problem is a resultof his field cut some other separate perceptual
problem or a memory problem (see discussion)CO did not complain of a similar deficit GR wasan accomplished artist prior to his injury producingpaintings of striking complexity Tragically hereports that he cannot now appreciate his ownpaintings
Other cognitive abilities
Face recognitionGR also reported some difficulties with face recog-nition On his second visit we examined his abilityto learn new faces by having him perform a forced-choice recognition test on seven familiar and sevenunfamiliar faces The familiar faces were personnelfrom the lab with whom he had at that point had agood deal of experience spanning at least 3 daysAll the people in the photographs had a towelwrapped around their head in order to minimise theuse of hair as a nonfacial cue Despite the difficultyof this task he correctly identified 67 familiarfaces and made no false alarms Surprisingly theone face that he did miss was the first author of thispaper with whom he at that point had had consid-erable experience GR also performed normally ona difficult famous face recognition test (see Experi-ment 4 following) and on the Benton Face Recog-nition test (see following) Thus he has someability to recognise faces Consistent with this GRreports that he can remember a small number offaces but that he quickly ldquoloses trackrdquo if he has toremember too many
CO also appears to suffer from someprosopagnosia as evidenced by his low perfor-mance on the Benton Face Recognition test (seefollowing see also Experiment 4) Interestingly hedoes not complain of face recognition difficultiesbut rather attributes his inability to recognise peo-ple to a lack of interest in their appearance
Episodic memoryAt first glance GRrsquos episodic memory appeared tobe largely intact As mentioned he could recall theevents of the previous day accurately Furthermorehe reports that he can follow the plot of a moviewithout any difficulty However both he and hiswife report that he does have some problems with
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 489
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Figure 2 Floor plan of the building at MIT in which GR wastested Corridors are highlighted in grey GR was asked to find themain room of the lab (marked G on the map) Starting location(marked S) was a point approximately 30 feet down the hall GRrsquosroute is marked by arrows GR found this task very difficult andonly knew for certain that he had reached his goal when he caughtsight of his wife through the glass door of the lab In contrast hewas able to follow a route marked on a plan of the same floorwithout any problem or hesitation
episodic memory For example he stated that heremembered very little about a trip to Italy they hadtaken several months previously His wife reportsthat he has great difficulty keeping track of timeFor example before their trip to Boston he repeat-edly asked her what day they were supposed to leaveand seemed unable to encode this information Weobserved a similar phenomenon during testingDuring the tests that required repetition of thesame procedure several times he sometimesseemed unable to clearly remember how manytimes he had repeated the procedure Thus he doesappear to have a subtle episodic memory deficit
which may manifest itself as an inability to sepa-rately remember successive events that are not easilydistinguishable from each other
CO also reports some problems with his epi-sodic memory He relies on lists to rememberappointments stating ldquoif I didnrsquot have a list Irsquodprobably forget why I walked in the door hererdquoDespite this claim he has never missed an appoint-ment nor is he ever confused about the purpose ofthe testing appointments He can accurately reportthe day of the week He remembers the experi-menters clearly from appointment to appointmentas well as the details of the various experiments
EPSTEIN ET AL
490 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 3 Two floor plans drawn by CO of the apartment he lived in for over 20 years The first plan was drawn when CO still lived inthe apartment the second was drawn several months later when he had moved out Note that the two plans correspond very well COreported that it was quite easy to produce these floor plans
CO home Feb 99
CO (former) home June 99
when they are repeated across appointments Whenqueried about his episodic memory difficulties COreported that he will usually remember an event ifhe is reminded of it but that he has difficulty spon-taneously regenerating events Interestingly hereports that even when he is reminded of an eventand remembers it he cannot remember ldquowhere itwas or when it wasrdquo When he reads he finds it dif-ficult to follow a story if it skips around Howeverwhen queried he was able to accurately report whathe had read in the newspaper the previous day
Formal neuropsychological tests
Formal neuropsychological testing was performedon GR in September 1998and on CO in December1998 Test performances were compared to age-matched peers3
GR exhibited mild to moderate impairment inmemory relative to what would be predicted for hisintellect (WAIS-R Full Scale IQ =122 93rd per-centile superior range) and level of educationThese memory problems were more apparent onvisual than on verbal memory tests On a test ofmemory for conceptually organised text passages(Wechsler Memory Scale Logical Memory[WMS-III] Wechsler 1997b) his performancewas average for both immediate (LM I =25th per-centile standard score SS = 90) and delayed recall(LM II =50th percentile SS =100) In contrast hisscore on the WMS-III Face Memory subtest was inthe low average range for both immediate (9th per-centile SS =80) and delayed recall (16th percentileSS =85) On a test of memory for scenes (WMS-IIIFamily Pictures subtest) immediate recall was defi-cient (2nd percentile SS = 69) and delayed recallwas borderline impaired (5th percentile SS =76)GR also had difficulty learning word lists on theRey Auditory Verbal Learning Test (Spreen ampStrauss 1998) his immediate recall was average(63rd percentile SS = 105) but after the interfer-ence list his performance fell to the low averagerange for both immediate (14th percentile SS =84)
and delayed recall (16th percentile SS =85) On arecognition test of the items of the list he success-fully recognised all 15 but made 8 false alarms Per-formance on the 724 Spatial Recall Test (RaoHammeke McQuillen Khatri amp Lloyd 1984)which required him to remember seven locations ona 6times4 checkerboard was 66th percentile (SS =106)for immediate and 2nd percentile (SS = 69) fordelayed recall Performance on the Benton FaceRecognition test was normal (42nd percentile)Performance on the Boston Naming Test was atceiling indicating that language abilities wereintact Thus GR exhibits some impairment ontasks that require delayed recall of visual informa-tion (WMS-III Face Memory subtest) or verbalinformation that is not organised in terms of a storyor plot (Rey AVLT) and is clearly impaired ontasks that required delayed recall of spatial informa-tion (724 Spatial Recall Test) or stimulus location(WMS-III Family Pictures subtest) In contrasthis ability to recall organised verbal information(WMS-III Logical Memory subtest) appears to beintact
As with GR COrsquos memory performance wasgenerally below what would be expected based onintellect (WAIS-III Full Scale IQ =134 99th per-centile very superior range) and education levelAlso like GR there was some evidence that visualmemory problems were more pronounced than ver-bal memory problems Memory for text passages(WMS-R Logical Memory) was average for bothimmediate (27th percentile SS = 91) and delayedrecall (48th percentileSS =99) In contrast perfor-mance on the WMS-III Faces subtest was lowaverage for both immediate (16th percentile SS =85) and delayed recall (9th percentile SS =80) andperformance on the Family Pictures subtest wasdeficient (lt 1st percentile SSlt 62) for both imme-diate and delayed recall On the Warrington Rec-ognition Memory Tests (RMT) he scored in theaverage range for words (50th percentile SS =100)and scenes (Warrington Topographical Recogni-tion Memory Test 29th percentile SS =92) and in
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 491
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
3The formal neuropsychological testing reported in this section was performed prior to the beginning of our combined
investigation of GR and CO (ie prior to the main experimental investigations of this paper) Consequently GR and CO were nottested on exactly the same battery of tests
the deficient range (lt 5th percentile SS lt 75) forfaces The low score for faces on this test as well asthe WMS-III may reflect prosopagnosia ratherthan a memory problem as he scored in the defi-cient range (lt1st percentile SSlt62) on the BentonFace Recognition test Verbal list learning (ReyAVLT) was average on immediate recall (45th per-centile SS =98) but after hearing an interferencelist performance fell to the deficient level (015words recalled) for both immediate and delayedrecall On a recognition test for these words herecognised 1115 and made one false alarm (39thpercentile SS =96) There was a large discrepancyon tests of memory span which may indicate adecline in spatial memory digit span was superior(9+ forward and 7 backwards both 99th percentile)but spatial span was only average (5 forward and 5backward 37th and 50th percentiles respectively)Language abilities were intact (Boston NamingTest performance was at ceiling) Thus like GRCO shows some impairment at recall of visualinformation (WMS-III Face Memory subtest) andverbal information that is not organised into a nar-rative (Rey AVLT) and clear impairment at tasksthat require recall of stimulus location (Family Pic-tures subtests of WMS-III) In contrast his abilityto recall organised verbal information (WMS-RLogical Memory subtest) is more preserved
MRI RESULTS
In order to determine the extent of their lesionsstructural MRI images were taken of both subjectsDelineation of the lesions was based either onspoiled GRASS coronal images at 11mm thick-ness (GR) or anatomical T-1 weighted MPRAGEMRI sequences yielding 125 mm thick coronalimages (CO) The anterior-posterior extent of thelesions was verified on axial images
GR has bilateral occipital-temporal lesions(Figure 4) In the left hemisphere the posteriorparahippocampal gyrus (including para-hippocampal cortex) is severely damaged with onlyminimal injury to the most caudal aspect of the hip-pocampus There is also moderate damage to themedial fusiform and inferior lingual gyri In the
right hemisphere the hippocampus is spared butthere is damage to the posterior end of theparahippocampal gyrus near the parahippocampal-lingual boundary More posteriorly the lesionedarea extends along the collateral sulcus to includesignificant portions of the inferior lingual gyrus andthe medial fusiform gyrus as well as most of themedial occipital lobe below the calcarine fissure
CO has a right-hemisphere occipital-temporallesion Right posterior hippocampus and para-hippocampal gyrus are severely affected Moreposteriorly the damage covers large portions of theinferior lingual and medial fusiform gyri as well assubstantial portions of the medial occipital lobeincluding most of right calcarine cortex In addi-tion diffuse foci of increased signal on T-2weighted images was present in periventricularwhite matter in the corona radiata and centrumsemiovale with a lacunar infarct of the rightputamen There was also moderate degree of dif-fuse involutional changes with increased ventricularsize particularly notable in the right temporal hornwith ex vacuo dilitation
The structural MRIs suggested that the PPAwas severely if not completely damaged in bothhemispheres for GR and in the right hemispherefor CO In order to confirm this for GR we used ahigh-field 3-T magnet to perform a functional scanin which he viewed scenes faces and objects in thesame blocked fMRI design that we have used tolocalise the PPA in all our previous experiments(see Epstein et al 1999 for details Epstein ampKanwisher 1998) A Kolomogorov-Smirnov testfound no voxels in the parahippocampal region thatfit our previously established criterion for inclusionin the PPA (ie greater response to scenes than tofaces and objects at a uncorrected significance levelof plt 0001) This result contrasts sharply with thepattern seen in over 95 of (mostly young or mid-dle-aged) normal subjects in whom a large acti-vated region can be identified in both left and rightparahippocampal cortex using this comparison (seeFigure 5) A similar test was used to identify voxelsthat responded significantly more to faces than toobjects This comparison revealed a region of face-selective voxels in the right fusiform gyrus consis-tent with the location of the Fusiform Face Area
EPSTEIN ET AL
492 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 493
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
GR
CO
Figure 4 Structural MRIs of GR and CO showing the extent of the damaged region (marked as white) Images have been normalisedusing SPM99 and a cost function masking algorithm (provided by Matthew Brett) into standard stereotactic space (Talairach amp Tournoux1988) using the Montreal Neurological Institute template (Evans et al 1993) Numbers indicate y-Talairach coordinate for each coronalslice corresponding slice planes are also indicated on the sagittal image
(FFA) commonly seen in normal subjects(Kanwisher McDermott amp Chun 1997) Thusthe MRI was sensitive enough to detect functionaldifferences an FFA was found but no PPA Whencombined with the anatomical results this stronglysuggests that GR has no PPA
Because of safety concerns (presence of a femo-ral artery aneurysm clip) we did not perform a simi-lar high-field functional MRI of CO However asFigure 4 demonstrates CO clearly has damage tothe parahippocampal-lingual region in the righthemisphere
EXPERIMENTAL INVESTIGATIONS
In this section we report the results of four experi-ments designed to probe the consequences of dam-age to the lingual-parahippocampal region ingeneral and the PPA in particular Our primaryconcern in these experiments was to test GR andCOrsquos ability to perceive remember and recognisedifferent kinds of visual material Based on our pre-vious fMRI work we predicted that GR and CO
would have more difficulty with scene-like stimuli(ie stimuli that convey information about thegeometry of a space one can navigate through or actwithin) than with object-like stimuli Such a deficitcould account for at least some of the problems GRand CO encounter in real-world navigation
Beyond determining the kind of visual materialthat gets processed in parahippocampal cortex wealso wanted to determine the nature of the process-ing that takes place on that material In particularwe wanted to determine if parahippocampal cortexis involved in perceptual encoding memory encodingor recognition As discussed earlier (Introduction)our fMRI results provided some evidence againstplace recognition as the sole function of the PPAbut did not allow us to distinguish between percep-tual and mnemonic encoding Experiments 1 and 2examine the role of parahippocampal cortex in per-ceptual encoding Experiment 3 examines its role inmnemonic encoding and Experiment 4 examinesits role in place recognition
Experiment 1 Two-dot task
The first hypothesis we examined was thatparahippocampal cortex plays a role in perceivingthe shape of the local environment In our previousfMRI work we observed that the PPA respondsmuch more strongly during scene viewing than dur-ing object or face viewing even when subjects donothing more than watch the scenes passively with-out performing any other task This suggested thatscene perception alone may be sufficient to activatethe PPA Previous reports have indicated that dam-age to parahippocampal cortex and other medialtemporal regions leads to problems with memoryrather than perception (Bohbot et al 1998Maguire Burke Philips amp Staunton 1996 Pigottamp Milner 1993) However all of these previousresults were obtained on patients who had unilat-eral lesions GR has a bilateral occipital-temporallesion including clear damage to parahippocampalcortex in both hemispheres furthermore he reportssome kind of perceptual deficit We hypothesisedthat this deficit might involve difficulty perceivingor interpreting the geometry of scenes and otherstimuli with a similar geometric structure
EPSTEIN ET AL
494 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 5 Top row shows a single slice from a functional MRIscan performed on GR Bottom row shows results from a normalsubject (NK) for comparison Voxels that responded more to facesthan objects (left column) or more to scenes than faces and objects(right column) in a Kolomogorov-Smirnov test are highlighted bywhite arrows GR has a functioning face area (FFA) but there isno evidence of a place area (PPA)
FFA PPA
GR
NK
In this experiment subjects viewed either objectshapes or spatial layouts Stimuli consisted ofblack-and-white photographs of real-world stimuli(empty rooms vs common objects) or abstractstimuli made out of Lego blocks (Lego ldquoscenesrdquo vsLego ldquoobjectsrdquo) These stimuli have been previouslyshown to differentially activate the PPA in normalsubjects (Epstein et al 1999 Epstein ampKanwisher 1998) Two red dots were superim-posed on each photograph (Figure 6) The task wasto report which of the two red dots was over thecloser part of the object or scene This task wasdesigned to have no memory component (apartfrom remembering the instructions) but to requireaccurate 3D perception of objects and scenes
If GR or CO have difficulty perceiving or inter-preting the spatial layout of scenes then we pre-dicted they would do worse when the stimuli areempty rooms or Lego scenes than when they arereal-world or Lego objects Note that the criticalcomparison is between the Lego scenes and theLego objects as these stimuli are made of the samematerials and differ primarily in their surfacegeometry
MethodTesting for all experiments was done on aMacintosh computer The experiment consisted of80 trials (20 of each stimulus type randomly inter-mixed) Subjects began each trial by pressing thespace bar A black-and-white photograph from oneof the four stimulus categories (see earlier) thenappeared on the screen with two red dots on it Thetask was to determine which of the two red dotsindicated the closer part of the scene or objectSubjects pressed the ldquofrdquo key if the leftmost red dotwas closest and the ldquojrdquo key if the rightmost dotwas closest They were instructed to respond asquickly as possible without making errors Stimuliremained on the screen until a response was made
The stimulus set was constructed so that eachempty room was matched to a common object thathad red dots in the exact same location and eachLego layout was similarly matched to a Lego objectThus the locations of the red dots were counterbal-anced across scene and object stimuli controllingfor any effects of visual field cuts
Results and discussionResults for GR and CO and four elderly normalsubjects are shown in Table 3 Both GR and COcould do this task without difficulty and their per-formance was near ceiling There was no evidencethat they had any more trouble with the emptyrooms or Lego scenes than with the real or Legoobjects Reaction times were slightly longer for GRand CO than for normal subjects as one wouldexpect given their visual field deficits however this
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 495
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 3 Experiment 1 (two-dot) results (SD in parenthesis)
GR CO Normals
Proportion correctObject 095 080 093Lego object 100 095 098Empty room 100 100 099Lego room 095 100 096
Mean RTObject 2435 2932 2161 (980)Lego object 2052 3245 2029 (921)Empty room 2744 2291 1788 (959)Lego room 1904 2281 1630 (1127)
Figure 6 The four kinds of stimuli used in Experiment 1 Topleft lego layouts top right real-world layouts (empty rooms)bottom left lego objects bottom right real-world objects
difference did not appear to be significant Impor-tantly there was little evidence that GR or CO tooklonger to make their perceptual judgements for thereal or Lego scenes than for the real or Lego objectsThus contrary to our initial hypothesis this experi-ment found no clear evidence for selective impair-ment in the perception of scene-like stimuli
Experiment 2 Different-views matchingtask
The previous experiment demonstrated that GRand CO can extract information about the per-ceived distances between themselves and the sur-faces of an object or scene Thus in at least onesense their perception of these surfaces is pre-served However this does not mean that their abil-ity to represent the layout of a scene is unimpairedThe two-dot task requires on-line comparison oftwo surfaces in egocentric coordinates GR and COwould be able to perform this task even if they couldnot (1) represent more than two surfaces at a time(2) represent surfaces in other than strictly egocen-tric coordinates or (3) sustain surface representa-tions for longer than the time it takes to make acomparison It has been suggested that the parietallobes are involved in on-line processing of surfacegeometry in egocentric coordinates (Milner ampGoodale 1995) thus it is possible that the parietallobes (which are intact in GR and CO) may be suf-ficient to perform the two-dot task even in theabsence of more durable or complex surface repre-sentations in parahippocampal cortex
In Experiment 2 we examine GR and COrsquosability to create and sustain a representation of thegeometry of an object or scene-like layout The taskwas to view a photograph of a Lego object or Legoscene hold it in memory for 10 s and then deter-mine whether a second object or scene was the sameas the first In ldquosamerdquo trials the second photographdepicted the same scene or object depicted from adifferent viewing angle In ldquodifferentrdquo trials the sec-ond photograph depicted a different scene or objectchosen to be similar to the first one (Figure 7)Thus in order to successfully perform this tasksubjects had to form a representation of the surfacesof the object or scene sustain this representation in
memory for 10 s and then align it (presumablyusing mental rotation) to the second object orscene
MethodThe experiment consisted of two parts each with40 trials In the first part the stimuli were black andwhite photographs of objects made out of Legoblocks In the second part the stimuli were photo-graphs of ldquoscenesrdquo made out of Lego blocks whichwere designed to have a geometric structure similarto real-world places such as a roomor a city street
Subjects began each trial by pressing the spacebar A photograph of a Lego object or sceneappeared on the screen for 3 s followed by a 10-sblank interval A second photograph then appeareddepicting either (1) the same object or scene as thefirst photograph shown from a different viewingangle (offset approximately 35 degrees) or (2) a dif-ferent object or scene Subjects were instructed topress the ldquojrdquo key if the two photographs depicted thesame object or scene and the ldquof rdquo key if they did notThey were told to take as much time as they neededto make a decision but no extra time The secondphotograph remained on the screen until a responsewas made Half the trials were ldquosamerdquo trials and halfwere ldquodifferentrdquo trials
Results and discussionFigure 8 shows results for GR and CO as well as forsix elderly normal controls GRrsquos and COrsquos perfor-mance was comparable to that of the normal con-trols Furthermore GR and CO performed just aswell when the stimuli were Lego scenes as whenthey were Lego objects These results are unlikely toreflect a ceiling effect as performance for bothpatients and controls was around 80 correctThus both patients appear to be able to form a rep-resentation of the geometry of both scenes andobjects which they can sustain in short-termmemory
Experiment 3 N-back recognition memorytask
The previous experiment failed to find any impair-ment in GR and COrsquos ability to form a representa-
EPSTEIN ET AL
496 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
tion of the geometry of a scene or object andmaintain it for 10 s in the absence of the originalstimulus Not only does this result suggest that per-ception of scene-like stimuli is intact in thesepatients it further suggests that some short-termmemory functions are intact as well However we
should note that subjects in this experiment werefree to devote all their effort and attention in the 10-second blank interval to maintaining a memory ofthe stimulus Thus it is possible that GR and COcan use rehearsal mechanisms to maintain the stim-ulus in short-term memory during the blank inter-
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 497
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
(b) Lego object version
Figure 7 Stimuli and procedure for Experiment 2 (a) In the Lego layout version subjects see a Lego layout for 3 s followed by a blankscreen for 10 s A second layout then appears which is either the same layout as the first one but seen from a different angle or a differentlayout (b) The Lego object version is the same but with object stimuli
3 s 10 s OR
3 s 10 s OR
(a) Lego layout version
(b) Lego object version
val but are unable to form a more durable memorytrace for the stimulus This possibility was tested inthe current experiment which was designed toexamine GR and COrsquos ability to learn new visualinformation
We devised a continuous n-back recognitionmemory test in which subjects viewed a sequence oforiginally novel stimuli some of which wererepeated within the sequence at intervals of 1 3 or5 intervening images For each stimulus subjects
reported whether they had seen it before in theexperiment or not In different runs in the experi-ment the stimuli were black-and-white photo-graphs of (1) Lego scenes (2) Lego objects (3) real-world landscapes (4) novel objects made out ofSculpey clay (5) unfamiliar houses (6) unfamiliarfaces as well as (7) line drawings of nonsenseobjects and (8) pronounceable pseudowords (Fig-ure 9) Thus we were able to test GR and COrsquos rec-ognition memory for different kinds of materials atdifferent time intervals in a paradigm in which thepresence of intervening items between the twoappearances of each stimulus made rehearsal diffi-cult Note that because none of the specific stimuliused in this experiment were familiar in advancethis task did not require an episodic memory com-ponent and could be done on the basis of pure visualfamiliarity with the particular visual stimulus
Although many different stimulus classes weretested in this experiment the critical comparison isbetween the Lego layouts and Lego objectsbecause these are best matched with regard tomaterials and differ primarily in their geometricstructure (see Introduction) We predicted that ifGR and CO showed a memory deficit for this taskit would be greater for the Lego layouts than for the
EPSTEIN ET AL
498 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 8 Results of Experiment 2 Performance over ldquosamerdquotrials and ldquodifferentrdquo trials is averaged together Error bars show1 SD
Figure 9 Stimuli for Experiment 3 Top row Lego layouts real-world scenes houses Bottom row Lego objects clay objects nonsenseobjects faces In an eighth condition (not shown) subjects viewed pronounceable nonwords such as ldquowilchrdquo or ldquothipperrdquo
Lego objects In addition a deficit for materialsthat convey information about the layout of sur-rounding space might also be apparent when GRand CO are tested on more realistic stimuli If soone would expect them to do poorly on the land-scapes but relatively well on the clay objects linedrawings of nonsense objects faces andpseudowords
MethodFor each subject a complete experimental sessionconsisted of being run once on each of the eightstimulus classes Each run consisted of 90 trials ineach trial a stimulus was presented and subjectsreported whether or not they had seen it before inthe run (or indeed ever) by pressing the ldquojrdquo key ifthey had or the ldquof rdquo key if they had not Each stimu-lus was on the screen for either 1 2 or 3 s (see fol-lowing) and appeared promptly after the keypressreport for the previous trial Individual stimuliappeared either once (14 stimuli) twice (30 stim-uli) or four times (4 stimuli) within a given run Ofthe stimuli that were presented twice 10 wererepeated immediately (1-back) 10 were repeatedwith a lag of two intervening items (3-back) and 10were repeated with a lag of four intervening items(5-back) In order to ensure that between-subjectdifferences in performance were not due to stimulusdifferences the same stimuli were assigned to thesame conditions each time the experimentwas run
GR performed the experiment twice once inAugust 1998 and again 9 months later Because ofa computer error he was only tested on six of theeight categories (all except Lego layouts and Legoobjects) at the later date CO was also tested twiceonce in February 1999 and again 6 months laterReported results for GR and CO are the average oftheir two testing sessions Normal subjects wereonly tested once Stimulus presentation time was 1 sin GRrsquos first testing session In pilot testing COfound this too fast for the critical categories of Lego
layouts and Lego objects so presentation time wasextended to 2 s for these categories in his first test-ing session All normal subjects were tested at thesepresentation rates (2 s for Lego layouts and objects1 s for other stimuli) In COrsquos second testingsession presentation time was lengthened to 3 s forall stimulus categories to ensure that any deficitsfound were memory deficits rather than problemswith the nonmemory aspects of the task
Results and discussionResults from GR CO and 10 elderly normal con-trols are shown in Figure 10 and Table 4 Figure 10ashows results from the main comparison betweenLego layouts and Lego objects Both normal sub-jects and patients performed near ceiling for bothLego layouts and Lego objects at 1-backinterstimulus intervals verifying that GR and COcould successfully perceive the pictures as they werebeing presented In contrast there was a strikingdifference between the two patients and the normalsubjects at 5-back intervals Whereas normals didon average somewhat better on Lego layouts thanLego objects both GR and CO showed the oppo-site pattern doing much worse on Lego layoutsthan Lego objects The 3-back condition showed amixed pattern Whereas GR did better on Legoobjects than Lego layouts in this condition (similarto his performance at 5-back intervals) CO didbetter on Lego layouts than Lego objects COrsquos lowperformance on Lego objects in the 3-back condi-tion appears to be an anomaly as it is much lowerthan his performance for comparable stimuli in the1-back and 5-back conditions
Figure 10b shows the difference in performancebetween Lego layouts and Lego objects for thecritical 1-back and 5-back intervals At 5-backintervals CO was significantly impaired comparedto the normal subjects (z = ndash250) whereasGRrsquos impairment fell just short of significance (z =ndash175)4
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 499
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
4In fact these values may underestimate GR and COrsquos relative impairment for Lego layouts In the 5-back condition 4 of the 10
normal subjects had perfect performance on the Lego layouts but all scored below ceiling on the Lego objects Thus the true advantagefor Lego layouts over Lego objects for normal subjects is probably greater than that observed here in which case GR and COrsquos lowerperformance for Lego layouts compared to Lego objects would be even more abnormal Furthermore post-hoc analyses revealed thatone of the normal subjects showed a difference in performance between Lego layouts and objects that was over 2 SD below the mean ofthe other nine subjects When this outlying subject was excluded z-scores were ndash322 for CO and ndash232 for GR
Table 4 shows results for all eight stimulusclasses Although direct comparisons between sub-jects should be treated with caution since viewingtime for different items was not the same for eachsubject we do note a few interesting patterns Firstof all there is little or no evidence that GR or CO
show a memory deficit for nonsense words clayobjects or line drawings of nonsense objects GRappears to be slightly impaired on faces and housesbut CO performs normally on these categoriesStrikingly GR shows a severe deficit for the reallandscapes performing more than 40 percentagepoints lower than normals Thus even thoughthere is some evidence that GR might have someimpairment for nonscene stimuli he clearly is muchmore impaired for scene-like stimuli such as theLego scenes and landscapes CO does not show asimilar deficit for the landscapes However itshould be kept in mind that in one of his two testingsessions CO viewed each picture for 3 s which wasmuch longer than the 1 second used for all non-Lego stimuli for GR and normals Thus COrsquosapparently ldquonormalrdquo performance in these condi-tions might be overestimated
Taken together the results of this experimentindicate that GR and CO are selectively impaired atprocessing scene-like stimuli This deficit is consis-tently apparent at 5-back but not 1-back intervalsThis finding suggests that the deficit should becharacterised as a memory encoding problemHowever the present results do not allow us todetermine whether the difficulty occurs at memoryencoding or at subsequent recognition (see Epsteinamp Kanwisher 2001) If GR and CO have a generalproblem with recognition of topographical materi-als then they should not be able to recognise scenes
EPSTEIN ET AL
500 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 10a Results of Experiment 3 for GR (top) CO (middle)and for 10 age-matched normal controls Performance measure isproportion of repeated stimuli correctly identified as such correctedfor guessing using the standard formula
Figure 10b Difference between Lego layout performance andLego object performance for the 1-back and 5-back conditionsError bars indicate one 1 SD
and landmarks that were learned prior to theirinjury We tested this possibility in the nextexperiment
Experiment 4 Famous landmarksfacesrecognition task
In this experiment we examined GR and COrsquosability to recognise the visual appearances of placeslearned prior to injury Subjects saw a series offamous landmarks intermixed with nonfamouscontrols (Imaging experiments have demonstratedthat landmarks of this type can activate the PPAalmost as strongly as complete scenes Epstein etal 1999 Epstein amp Kanwisher 1998) They wererequired to say whether they knew each specificlandmark and to identify the ones they knew Sub-jects were instructed to respond affirmatively if theywere fairly certain that they recognised the land-mark but not if it only looked generally familiarFor purposes of comparison subjects also per-formed an identical test in which the stimuli werefamous and nonfamous faces In both testsdistractor items were designed to be visually andconceptually similar to the target items (see Figure11) If parahippocampal cortex is required for placerecognition then one might expect GR and CO toperform poorly compared to normals on the land-mark recognition test
MethodEach of the two experiments (famous faces famouslandmarks) consisted of 48 trials Subjects beganeach trial by pressing the space bar after which ablack-and-white photograph of a face (in thefamous faces experiment) or a landmark (in thefamous landmarks experiment) appeared on thescreen Subjects were told to press the ldquojrdquo key if theyrecognised the specific face or landmark and the ldquof rdquokey if they did not taking as much time at theyneeded to make this discrimination but no extratime Subjects were told to report that they recog-nised a face or landmark if they felt that they knewit even if they could not retrieve its name If theydid know the name they were to say it after pressingthe button Otherwise they were to say whateverthey did remember about the item
Half of the photographs were of famous and halfof nonfamous people and landmarks The famousfaces included public figures (eg George BushRichard Nixon Prince Charles) and movie stars(eg Harrison Ford Clark Gable Marlon Brando)who were prominent prior to the time of GR andCOrsquos injuries the famous landmarks were build-ings and monuments from all over the world (egEiffel Tower US Capitol Building Great Sphinx)The nonfamous stimuli were chosen to match thefamous stimuli as closely as possible Each famousstimulus had a ldquomatchrdquo For example George Bushwas matched to a man of similar age in a coat andtie and the Taj Mahal was matched to a visually
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 501
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 4 Full results of Experiment 3 Mean proportion correct (corrected for guessing) SDs in parentheses
Lego Land- Lego Clay Nonsense Non-scenes scapes Houses objects objects objects Words Faces
1-back GR 100 094 100 100 100 094 100 100CO 091 085 091 100 100 079 074 100Normals 096(008) 095(008) 093(014) 096(008) 096(006) 099(003) 098(004) 097(011)
3-back GR ndash003 068 026 047 083 083 075 052CO 066 081 037 010 094 069 095 081Normals 066(032) 077(021) 072(015) 077(020) 086(016) 083(015) 083(017) 072(020)
5-back GR 010 034 044 047 072 083 094 053CO 031 073 075 088 081 064 100 081Normals 076(026) 077(016) 068(021) 066(016) 091(014) 083(018) 093(011) 078(021)
similar Asian palace that is not famous (see Figure11)
Results and discussionResults for GR and CO as well as for five age-matched controls are shown in Figure 12 Both
GR and CO could recognise the famous landmarksas well normals In addition both GR and CO wereusually able to name the landmarks they success-fully identified
Thus GR and CO can recognize landmarkslearned prior to injury demonstrating that at leastsome of their place recognition abilities are intactWhen combined with the fact that GR and COwere able to produce detailed maps of places theylived in prior to their injury (see Case Description)these results suggest that their deficit is primarily inencoding or consolidation of new informationrather than in recognition or recall of old informa-tion Note however that these results do not provethat their place recognition abilities are entirelynormal GR and CO might suffer from a mild rec-ognition deficit that affects identification of newly-learned stimuli like the Lego layouts in Experiment3 but not overlearned stimuli like the famous land-marks Furthermore it is possible that GR and COmight be impaired at recognising premorbidlylearned places that are defined primarily by theirspatial layout even though they are unimpaired atrecognising premorbidly learned places that aredefined primarily by a single object-like landmarkor building (see General Discussion) Neverthelessthe trend of the present evidence is to suggest thatGR and CO have particular problems with noveltopographical information
In the famous-face task GRrsquos performance wasin the normal range However he sometimes haddifficulty naming the faces even though he clearly
EPSTEIN ET AL
502 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 11 Stimuli for Experiment 4 Each famous landmark(Arc de Triomphe Taj Mahal) and famous face (Barbara BushYasser Arafat) was matched to a nonfamous building or face thatalso appeared in the experiment
FA
CE
SL
AN
DM
AR
KS
Figure 12 Results of Experiment 4 Performance measure isproportion of famous stimuli reported as ldquoknownrdquo corrected forguessing using the standard formula
recognised them For example he described Ron-ald Reagan as ldquothe gipperrdquo Nancy Reagan as ldquothewife of the gipperrdquo and Clark Gable as ldquothe movieactor who said lsquoScarlett I donrsquot give a damnrsquordquo COhad a great deal of difficulty identifying the famousfaces This result is not surprising given hisextremely low performance on the Benton FaceMatching Task In addition to his topographicproblems CO appears to suffer fromprosopagnosia
GENERAL DISCUSSION
We tested two parahippocampal patients (GR andCO) on four experiments designed to measure per-formance as a function of both task (perceptionencoding and recognition) and stimulus type(scenes objects faces and words) The only taskthat revealed a clear deficit was an n-back recogni-tion memory test (Experiment 3) In order to suc-cessfully perform this task subjects had to encode adurable representation of a visual stimulus and useit for later recognition GR and CO were signifi-cantly more impaired on this task when the stimuliwere scene-like spatial layouts (ie Lego scenesand real scenes for GR) than when they wereobjects No comparable deficits were found in otherexperiments that tested layout perception (Experi-ments 1 and 2) and placelandmark recognition(Experiment 4) Taken together these results indi-cate that GR and CO are impaired in their ability toencode novel information about the geometry ofsurrounding space into memory but are lessimpaired at their ability to encode the geometry ofnovel objects
We argue that this specific topographical defi-citmdashinability to encode the spatial layout of novelscenesmdashresults from damage to the para-hippocampal place area Although the damage inboth patients extends beyond the parahippo-campal-lingual territory normally associated withthe PPA this region is clearly lesioned bilaterally inGR and in the right hemisphere in CO Further-more functional imaging demonstrated that GRhas no functioning PPA Thus the current resultsand the earlier fMRI work (Epstein et al 1999
Epstein amp Kanwisher 1998) converge to implicateparahippocampal cortex in the encoding of infor-mation about scene geometry into memory
Our assignment of the function of memoryencoding to parahippocampal cortex is consistentwith the results of earlier case reports of patientswith parahippocampal damage who have naviga-tional difficulties in novel but not familiar environ-ments (Habib amp Sirigu 1987 Ross 1980 see alsoTeng amp Squire 1999) As with GR and CO thesepatients were either less impaired or unimpaired onnontopographical memory tasks Patients have alsobeen described who exhibit the opposite pattern ofimpairment an inability to learn faces (sometimescalled ldquoprosopamnesiardquo Cipolotti Robinson Blairamp Frith 1999 Tippett Miller amp Farah 2000) orfaces and verbal materials (Maguire amp Cipolotti1998) with an unimpaired ability to learn topo-graphical materials Taken together these resultsargue for a high degree of domain-specificity in theneural systems involved in learning of new informa-tion (Gallistel 1995) The brain may have separatemechanisms for learning faces places and wordswhich can be differentially impaired after damageto specific cortical regions (Cipolotti et al 1999Incisa della Rochetta et al 1996) The presentresults suggest that the anatomical locus of themechanism dedicated to learning new topographi-cal information is in the parahippocampal cortex(Aguirre amp DrsquoEsposito 1999)
In contrast to their impaired ability to encodenew topographical information both GR and COwere able to produce detailed maps of their previousdwellings and performed normally on the famouslandmark recognition test of Experiment 4 Theseresults are generally consistent with Teng andSquirersquos (1999) report of a patient with extensivemedial temporal damage who could recallpremorbidly learned but not postmorbidly learnedtopographical information (see also Ross 1980)Thus our results provide further evidence thatmedial temporal regions such as parahippocampalcortex may be more critical for encoding or consoli-dating new information into memory than for recallof old information
One issue that our experiments do not entirelyresolve is the precise nature of the topographical
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 503
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
information encoded by parahippocampal cortexClearly this region is critical for encoding informa-tion about the geometry of the currently visiblescene even when the ldquoscenerdquo is made out of Legoblocks and therefore has no immediate navigationalrelevance However it is unclear whether this spe-cific deficit can account for all of the real-worldnavigational difficulties encountered by GR andCO GR in particular shows such a catastrophicloss of navigational ability that we suspect he is alsoimpaired at encoding the spatial relationshipsbetweenthe current scene and other locations in theworld CO probably also suffers from such difficul-ties as he reports that he has trouble rememberinghow places relate to each other in large-scale spaceWhen he was tested on a simple 3-room videogameenvironment he not only was unable to rememberthe individual rooms (claiming that there were 10rooms when there were only 3) but he also had nomemory for the spatial relationships between themThese spatial deficits may be the result of damage tothe posterior right hippocampus (in the case of CO)or of disconnection of the posterior right hippo-campus from the parahippocampal regions thatprovide much of its cortical input (in both patientsSuzuki amp Amaral 1994) In either case the abilityto form a cognitive map of new environments likethe lab or videogame space could be impaired(Maguire et al 1998 OrsquoKeefe amp Nadel 1978)However it is also possible that parahippocampalcortex itself may be critical for encoding not onlythe geometry of individual scenes but also the spa-tial relationships between the locations of thosescenes In this case the spatial deficits of GR andCO could be attributed directly to parahippo-campal damage rather than to hippocampal dam-age or disconnection
The latter scenario would be consistent withAguirre and DrsquoEspositorsquos (1999) and Barrash etalrsquos (2000) suggestion that parahippocampal cortexis critically involved in combining visual informa-tion about the appearance of scenes and landmarkswith spatial information about their locations into aunified topographical representation Howeverboth of these authors argue that the representationsof individual scenes (Barrash et al) and landmarks(Aguirre amp DrsquoEsposito) are supported not by
parahippocampal cortex but by more posterior lin-gual-fusiform regions In contrast we believe thatthe neuroimaging data indicate that parahippo-campal cortex is involved in the encoding of thegeometric structure of individual scenes althoughit may also play a role in combining these represen-tations with other spatial representations
Interestingly GR and CO were not impaired onrecognition of famous landmarks that were mostlybuildings Thus our data do not preclude the possi-bility that there may be a separate lingual orfusiform region involved in processing informationabout buildings (Aguirre Zarahn amp DrsquoEsposito1998 Chao Haxby amp Martin 1999 IshaiUngerleider Martin Schouten amp Haxby 1999)which might have been preserved in GR and COdespite extensive damage to other regions of thelingual and fusiform gyrus Alternatively GR andCOrsquos preserved building recognition abilities mightbe mediated by interactions between cortical areasthat perform shape analysis for any object-likestimulus type (Malach et al 1995Kanwisher et al1996) and anterior temporal regions in which long-term topographical and semantic memories arestored (McCarthy et al 1996)
Is perception preserved
In contrast to their inability to encode new scenesinto memory GR and CO have substantiallyretained their ability to perceive the spatial layout ofthe currently visible scene (Experiments 1 and 2)GR can follow a route marked on a floor planwhich indicates a good on-line appreciation of hiscurrent spatial surroundings Furthermore neitherGR or CO have any trouble moving through theenvironmentmdashthey move with confidence and donot bump into doors or walls Problems of the lattertype are usually associated with damage to the pari-etal lobes which are intact in GR and CO Thus atfirst glance GR and CO do not appear to haveperceptual impairments (beyond their visual fieldrestrictions)
However GR does complain of a perceptualdeficit which he describes as an inability to ldquotakeinrdquo scenes when they are too complex Similarreports of an inability to comprehend the
EPSTEIN ET AL
504 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
ldquoglobalityrdquo of a scene were made by some of theparahippocampally-damaged patients studied byHabib and Sirigu (1987) We were not able todetermine the precise nature of this deficit whichmay be partly attributable to his visual field restric-tions It appears that his deficit is more salient in thereal-world environments than in the picture-per-ception tasks of Experiments 1 and 2 perhapsbecause he finds it more difficult to organise visualinput when immersed in it than when viewing pho-tographs that subtend only part of the visual field
We suggest that GR can perceive parts of thescene when he attends to them but may be unableto integrate information over time into a coherentrepresentation of a scene or place The fact that GRand CO are impaired for Lego layouts at 5-backintervals in Experiment 3 is consistent with thishypothesis The time between the first and secondpresentations of a stimulus is very short in this case(and even shorter with 3-back intervals where GRalso shows evidence of a deficit) In both patientsthe representation of layout information formed byperception of a Lego scene must either have anabnormally short persistence be abnormally sus-ceptible to interference or both If so they mightfind it impossible to sustain a spatial frame orschema long enough to use it to organise informa-tion obtained from many different glances at a com-plex scene (Intraub 1997) The result would be aninability to understand the ldquoglobalityrdquo of a sceneeven though all the details were perfectlyperceived
Non-topographical mnemonic abilities
Although not as severe as their topographical mem-ory problems GR and CO do have some memorydifficulties in other domains For example bothperformed poorly on the Rey AVLT whichrequired them to remember a list of unrelatedwords Both also report some subtle episodic mem-ory problems (see Case Description) COrsquos prob-lems could partly result from damage to the righthippocampusmdashindeed CO appeared to have themore acute episodic memory problems HoweverGR has no hippocampal damage so the fact that hedisplays episodic memory problems suggests thatparahippocampal cortex is involved in the forma-
tion of episodic memories (cf Wagner et al1998)
In previous work (Epstein et al 1999) weargued that spatial context information encoded byparahippocampal cortex might play a role in estab-lishing event memories by encoding informationabout where events take place The inability toencode this information might cause problems intasks such as the Rey AVLT where one has to recalla list of unrelated words If spatial (and perhapstemporal) context information is not encoded itwill be harder subsequently to recall the distinctevent of hearing each word (Tulving 1983) Incontrast lack of spatial context information wouldbe less of a problem in tasks where one is onlyrequired to recognise previously encountered verbalmaterials but not to reproduce the spatiotemporalcontext in which they were experienced IndeedGR and CO performed as well as normals on thenonsense words in Experiment 4 where recogni-tion memory but not episodic memory was requiredfor successful performance Interestingly GR andCO also did relatively well on the Logical Memorysubtest of the WMS-III perhaps because the nar-rative framework of a story provides sufficient con-textual information to overcome the absence ofspatial context and mediate successful verbalretrieval
Differences between GR and CO
Although we have discussed GR and CO togetherthere are a number of relevant differences betweenthem One open question is whether these differ-ences can be attributed to the different effects ofunilateral and bilateral parahippocampal damageIn general GRrsquos topographical difficulties weremore severe than COrsquos CO did surprisingly well onthe Warrington Topographical Memory Test inwhich he was required to remember detailed real-world city scenes and also performed substantiallybetter on real scenes than Lego scenes in Experi-ment 3 Insofar as CO has intact left para-hippocampal cortex this suggests that the left PPAmight be sufficient to encode a certain amount of(perhaps semantic) information about real-worldscenes but the right PPA is necessary to encode
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 505
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)

Case 2 COCO is a left-handed male with 20 years of educa-tion He was 60 years old at the time of testing Twoyears prior to testing he suffered a right posteriorcerebral artery stroke He first noticed on a week-end that his vision appeared blurred on the left andthat he had a tendency to bump into objects on thisside On Monday morning he was able to success-fully negotiate the route to his workplace a trip thatinvolved a bus ride followed by two subways andthen a shuttle to his office building However oncehe entered the building he found himself com-pletely disoriented unsure of its layout and wherehis office was located Interestingly the companyhe worked for had moved a few weeks earlierAlthough he had managed to arrive at this newbuilding (which was a block away from the previouslocation) he became lost upon entering it Hisneurologic exam revealed a left homonymoushemianopia but no evidence of neglect on a lettercancellation task or on sensory testing Investiga-tions revealed an ischaemic stroke with haemor-rhagic transformation in the right occipital andmesial temporal lobe On magnetic resonanceangiography he had a chronic left carotid occlusiona mild right carotid stenosis and bilateral vertebralartery stenoses Since then he has suffered fromsevere topographical disorientation primarily innew environments
CO reports that he can recognise buildings thatwere familiar to him before the stroke He can alsoform mental images of well-known landmarks suchas the Empire State Building and the BostonPublic Gardens and of the house he lived in as achild When asked to imagine standing in front ofthe Massachusetts State House (a location wellknown to him from years of living in Boston) heaccurately described the surrounding buildings andtheir locations However he reports that when helooks at a place that hersquos encountered post-strokeall he gets is a feeling of familiarity that does notallow him to determine where the place is or whenhe saw it He also reports an inability to understandhow various landmarks relate to each other inlarge-scale space Despite this difficulty he canfollow a map and frequently rides the Bostonsubway alone
Unlike GR CO was able to learn a certainamount of topographical information For exam-ple he soon learned the location of the rooms in thelab and would travel between them unassistedAfter his first visit to the lab he needed to be guidedall the way back to the subway station but after sev-eral visits he could find his way out of the buildingand to the subway on his own as long as he was firstguided to the elevators (It should be noted how-ever that the building is on the same block as thesubway station Furthermore he still needed to bedirected to the elevators even though our buildinghas a relatively simple floor plan) Thus the amountof topographical information he can learn is limitedbut not zero
COrsquos language and reasoning skills appear to benormal and his conversation is appropriate andvaried In addition to his topographical difficultieshe also has limitations of his peripheral vision onthe left side
Navigational abilities
In addition to the experiments reported in theExperimental Investigations section of this paperwe also performed a number of ad hoc tests of GRand COrsquos navigational abilities Although thesetests were informal we describe them here to pro-vide a general sketch of the nature of GR and COrsquosimpairments
Real-world topographical learningOver the course of several days we took GR and hiswife on three walks around the MIT campus whichneither had visited previously Each walk followedexactly the same path At several places in the walkwe would stop and point out different buildings andlandmarks tell them something about each one andask them to remember their appearance for latertesting Immediately after the completion of thethird walk both GR and his wife separately per-formed a forced-choice recognition test in whichthey saw photographs of 14 landmarks (mostlybuildings) from the walk and 14 novel landmarks(from another part of the MIT campus that theyhad not visited) and were required to reportwhether they had seen each one or not GRrsquos
EPSTEIN ET AL
486 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
performance was 714 correct hits for the familiarlandmarks and 1314 correct rejections for theunfamiliar landmarks In contrast his wifeobtained a perfect score on this test Given theextensive nature of the training regime GRrsquos scorestrongly suggests an impairment in learning theappearance of new real-world places Interestinglyseveral of the seven familiar landmarks that he gotright had an obvious distinctive feature that hadbeen explicitly pointed out to him For exampleone building had a distinctive clock tower andanother one was an unusual triangular shape ThusGR was occasionally able to use distinct visualfeatures to identify places even though he wasunable to recognise them holisticallymdasha commonpattern among topographical disorientationpatients
GR did not appear to have any obvious problemsin understanding the spatial structure of his imme-diate environment When given a floor plan of thebuilding at MIT in which he was being tested hewas able to successfully follow a route marked onthe plan However when the plan was taken awayhe had great difficulty finding his way back to thelab even though he had more than 30 min experi-ence with the environment and his starting positionwas less than 30 feet from the goal (Figure 2) Infact the route he took resembled a random walk Atone point he walked right past his destinationwithout recognising it and only knew for sure thathe was in the right place when he identified his wifein the room Thus his on-line appreciation of thelayout of immediately visible space appears to beintact but he cannot find his own way withoutassistance
We were unable to perform the MIT landmarkrecognition test on CO because he suffered from amedical problem that made it difficult for him towalk around campus In lieu of this we tried to seewhether he could learn a simple video game envi-ronment consisting of three connected roomsAfter about 30 min experience navigating throughthis environment (via button presses) he was askedto sketch a map of it He declined to do so statingthat it was beyond his abilities When asked howmany rooms there were he replied that he did notknow but that there were at least 10 Given the
simplicity of this artificial environment COrsquos com-plete inability to learn it is striking
Map drawingAfter several days of testing we asked both GR andhis wife to sit at different tables and independentlysketch (1) the shape of Florida (2) a bicycle (3) afloor plan of their hotel room in Boston (4) a floorplan of the first floor of the house they had been liv-ing in for the last 8 months (all post-stroke) (5) afloor plan of the main room of the lab and (6) afloor plan of the house they lived in 10 years ago(pre-stroke) GR drew the map of Florida and thebicycle perfectly His plan of the hotel room wasnotably different from the one drawn by his wifemdashfor example he reported that there was only onebed when in fact there were two and he placed thebed on the wrong side of the room (Figure 1a) Hisplan of the first floor of his current house was some-what better but was not very detailed and includedsome major errors For example he wronglyreported that there was no bathroom on this floor(Figure 1b) He was unable to produce any floorplan of the main room of the lab leaving this pageblank When his wife reminded him that this wasthe room in which he had just spent an hour readingthe newspaper and in which we had all had break-fast in that morning he replied that he didnrsquot knowwhat room she was talking about (although heappeared to remember these events) In contrast tothis near total inability to produce accurate maps ofnew environments GR produced an accurate mapof the house he lived in prior to his injury (Figure1c) He reported that his memory of this house wasldquoall thererdquo and he could imagine walking around init When asked to produce a perspective of a roomwhen he was in it he did so with ease and he wasalso able to create an accurate floor plan of theroom
In contrast CO found it very difficult to pro-duce a floor plan of the lab both when he was in itand when he was taken to another room and askedto recall it However although the maps he pro-duced were quite impoverished they were notentirely inaccurate demonstrating some ability toencode topographical information This contrastswith GRrsquos absolute inability to recall anything
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 487
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
EPSTEIN ET AL
488 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure
1F
loorplansdraw
nby
GR
andhisw
ifeof(a)
theirhotelroomin
Boston(b)theircurrenthousew
hichthey
hadm
ovedto
subsequenttoG
Rrsquosinjury(c)the
housethey
livedin
priorto
GR
rsquosinjury
Ho
telro
om
inB
osto
n
GR
His
wife
His
wife
Cu
rren
th
om
e
GR
His
wife
Fo
rme
rh
om
e
GR
about the main room of the lab once he had left itCO was also able to draw a detailed floor plan of theapartment he was living in at the time (and hadlived in for 20 years) Several months after he hadmoved from this apartment we asked him to drawsuch a floor plan again and he produced one thatwas substantially the same as the first (Figure 3)Thus their map drawing performance indicatesthat GR and CO are primarily impaired at sketch-ing places experienced after their strokes but areunimpaired at sketching places experienced beforetheir strokes This pattern is indicative of deficits inencoding but not recall
Scene perceptionAs noted earlier GR can navigate with the aid of afloor plan suggesting that his perceptual abilitiesare largely intact Interestingly however he reportsthat he does have a perceptual deficit which hedescribes as an inability to understand scenes whenthey are too complex For example he reports thathe often has trouble using maps because theyusually are too busy with details It is unclearwhether this apparent perceptual problem is a resultof his field cut some other separate perceptual
problem or a memory problem (see discussion)CO did not complain of a similar deficit GR wasan accomplished artist prior to his injury producingpaintings of striking complexity Tragically hereports that he cannot now appreciate his ownpaintings
Other cognitive abilities
Face recognitionGR also reported some difficulties with face recog-nition On his second visit we examined his abilityto learn new faces by having him perform a forced-choice recognition test on seven familiar and sevenunfamiliar faces The familiar faces were personnelfrom the lab with whom he had at that point had agood deal of experience spanning at least 3 daysAll the people in the photographs had a towelwrapped around their head in order to minimise theuse of hair as a nonfacial cue Despite the difficultyof this task he correctly identified 67 familiarfaces and made no false alarms Surprisingly theone face that he did miss was the first author of thispaper with whom he at that point had had consid-erable experience GR also performed normally ona difficult famous face recognition test (see Experi-ment 4 following) and on the Benton Face Recog-nition test (see following) Thus he has someability to recognise faces Consistent with this GRreports that he can remember a small number offaces but that he quickly ldquoloses trackrdquo if he has toremember too many
CO also appears to suffer from someprosopagnosia as evidenced by his low perfor-mance on the Benton Face Recognition test (seefollowing see also Experiment 4) Interestingly hedoes not complain of face recognition difficultiesbut rather attributes his inability to recognise peo-ple to a lack of interest in their appearance
Episodic memoryAt first glance GRrsquos episodic memory appeared tobe largely intact As mentioned he could recall theevents of the previous day accurately Furthermorehe reports that he can follow the plot of a moviewithout any difficulty However both he and hiswife report that he does have some problems with
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 489
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Figure 2 Floor plan of the building at MIT in which GR wastested Corridors are highlighted in grey GR was asked to find themain room of the lab (marked G on the map) Starting location(marked S) was a point approximately 30 feet down the hall GRrsquosroute is marked by arrows GR found this task very difficult andonly knew for certain that he had reached his goal when he caughtsight of his wife through the glass door of the lab In contrast hewas able to follow a route marked on a plan of the same floorwithout any problem or hesitation
episodic memory For example he stated that heremembered very little about a trip to Italy they hadtaken several months previously His wife reportsthat he has great difficulty keeping track of timeFor example before their trip to Boston he repeat-edly asked her what day they were supposed to leaveand seemed unable to encode this information Weobserved a similar phenomenon during testingDuring the tests that required repetition of thesame procedure several times he sometimesseemed unable to clearly remember how manytimes he had repeated the procedure Thus he doesappear to have a subtle episodic memory deficit
which may manifest itself as an inability to sepa-rately remember successive events that are not easilydistinguishable from each other
CO also reports some problems with his epi-sodic memory He relies on lists to rememberappointments stating ldquoif I didnrsquot have a list Irsquodprobably forget why I walked in the door hererdquoDespite this claim he has never missed an appoint-ment nor is he ever confused about the purpose ofthe testing appointments He can accurately reportthe day of the week He remembers the experi-menters clearly from appointment to appointmentas well as the details of the various experiments
EPSTEIN ET AL
490 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 3 Two floor plans drawn by CO of the apartment he lived in for over 20 years The first plan was drawn when CO still lived inthe apartment the second was drawn several months later when he had moved out Note that the two plans correspond very well COreported that it was quite easy to produce these floor plans
CO home Feb 99
CO (former) home June 99
when they are repeated across appointments Whenqueried about his episodic memory difficulties COreported that he will usually remember an event ifhe is reminded of it but that he has difficulty spon-taneously regenerating events Interestingly hereports that even when he is reminded of an eventand remembers it he cannot remember ldquowhere itwas or when it wasrdquo When he reads he finds it dif-ficult to follow a story if it skips around Howeverwhen queried he was able to accurately report whathe had read in the newspaper the previous day
Formal neuropsychological tests
Formal neuropsychological testing was performedon GR in September 1998and on CO in December1998 Test performances were compared to age-matched peers3
GR exhibited mild to moderate impairment inmemory relative to what would be predicted for hisintellect (WAIS-R Full Scale IQ =122 93rd per-centile superior range) and level of educationThese memory problems were more apparent onvisual than on verbal memory tests On a test ofmemory for conceptually organised text passages(Wechsler Memory Scale Logical Memory[WMS-III] Wechsler 1997b) his performancewas average for both immediate (LM I =25th per-centile standard score SS = 90) and delayed recall(LM II =50th percentile SS =100) In contrast hisscore on the WMS-III Face Memory subtest was inthe low average range for both immediate (9th per-centile SS =80) and delayed recall (16th percentileSS =85) On a test of memory for scenes (WMS-IIIFamily Pictures subtest) immediate recall was defi-cient (2nd percentile SS = 69) and delayed recallwas borderline impaired (5th percentile SS =76)GR also had difficulty learning word lists on theRey Auditory Verbal Learning Test (Spreen ampStrauss 1998) his immediate recall was average(63rd percentile SS = 105) but after the interfer-ence list his performance fell to the low averagerange for both immediate (14th percentile SS =84)
and delayed recall (16th percentile SS =85) On arecognition test of the items of the list he success-fully recognised all 15 but made 8 false alarms Per-formance on the 724 Spatial Recall Test (RaoHammeke McQuillen Khatri amp Lloyd 1984)which required him to remember seven locations ona 6times4 checkerboard was 66th percentile (SS =106)for immediate and 2nd percentile (SS = 69) fordelayed recall Performance on the Benton FaceRecognition test was normal (42nd percentile)Performance on the Boston Naming Test was atceiling indicating that language abilities wereintact Thus GR exhibits some impairment ontasks that require delayed recall of visual informa-tion (WMS-III Face Memory subtest) or verbalinformation that is not organised in terms of a storyor plot (Rey AVLT) and is clearly impaired ontasks that required delayed recall of spatial informa-tion (724 Spatial Recall Test) or stimulus location(WMS-III Family Pictures subtest) In contrasthis ability to recall organised verbal information(WMS-III Logical Memory subtest) appears to beintact
As with GR COrsquos memory performance wasgenerally below what would be expected based onintellect (WAIS-III Full Scale IQ =134 99th per-centile very superior range) and education levelAlso like GR there was some evidence that visualmemory problems were more pronounced than ver-bal memory problems Memory for text passages(WMS-R Logical Memory) was average for bothimmediate (27th percentile SS = 91) and delayedrecall (48th percentileSS =99) In contrast perfor-mance on the WMS-III Faces subtest was lowaverage for both immediate (16th percentile SS =85) and delayed recall (9th percentile SS =80) andperformance on the Family Pictures subtest wasdeficient (lt 1st percentile SSlt 62) for both imme-diate and delayed recall On the Warrington Rec-ognition Memory Tests (RMT) he scored in theaverage range for words (50th percentile SS =100)and scenes (Warrington Topographical Recogni-tion Memory Test 29th percentile SS =92) and in
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 491
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
3The formal neuropsychological testing reported in this section was performed prior to the beginning of our combined
investigation of GR and CO (ie prior to the main experimental investigations of this paper) Consequently GR and CO were nottested on exactly the same battery of tests
the deficient range (lt 5th percentile SS lt 75) forfaces The low score for faces on this test as well asthe WMS-III may reflect prosopagnosia ratherthan a memory problem as he scored in the defi-cient range (lt1st percentile SSlt62) on the BentonFace Recognition test Verbal list learning (ReyAVLT) was average on immediate recall (45th per-centile SS =98) but after hearing an interferencelist performance fell to the deficient level (015words recalled) for both immediate and delayedrecall On a recognition test for these words herecognised 1115 and made one false alarm (39thpercentile SS =96) There was a large discrepancyon tests of memory span which may indicate adecline in spatial memory digit span was superior(9+ forward and 7 backwards both 99th percentile)but spatial span was only average (5 forward and 5backward 37th and 50th percentiles respectively)Language abilities were intact (Boston NamingTest performance was at ceiling) Thus like GRCO shows some impairment at recall of visualinformation (WMS-III Face Memory subtest) andverbal information that is not organised into a nar-rative (Rey AVLT) and clear impairment at tasksthat require recall of stimulus location (Family Pic-tures subtests of WMS-III) In contrast his abilityto recall organised verbal information (WMS-RLogical Memory subtest) is more preserved
MRI RESULTS
In order to determine the extent of their lesionsstructural MRI images were taken of both subjectsDelineation of the lesions was based either onspoiled GRASS coronal images at 11mm thick-ness (GR) or anatomical T-1 weighted MPRAGEMRI sequences yielding 125 mm thick coronalimages (CO) The anterior-posterior extent of thelesions was verified on axial images
GR has bilateral occipital-temporal lesions(Figure 4) In the left hemisphere the posteriorparahippocampal gyrus (including para-hippocampal cortex) is severely damaged with onlyminimal injury to the most caudal aspect of the hip-pocampus There is also moderate damage to themedial fusiform and inferior lingual gyri In the
right hemisphere the hippocampus is spared butthere is damage to the posterior end of theparahippocampal gyrus near the parahippocampal-lingual boundary More posteriorly the lesionedarea extends along the collateral sulcus to includesignificant portions of the inferior lingual gyrus andthe medial fusiform gyrus as well as most of themedial occipital lobe below the calcarine fissure
CO has a right-hemisphere occipital-temporallesion Right posterior hippocampus and para-hippocampal gyrus are severely affected Moreposteriorly the damage covers large portions of theinferior lingual and medial fusiform gyri as well assubstantial portions of the medial occipital lobeincluding most of right calcarine cortex In addi-tion diffuse foci of increased signal on T-2weighted images was present in periventricularwhite matter in the corona radiata and centrumsemiovale with a lacunar infarct of the rightputamen There was also moderate degree of dif-fuse involutional changes with increased ventricularsize particularly notable in the right temporal hornwith ex vacuo dilitation
The structural MRIs suggested that the PPAwas severely if not completely damaged in bothhemispheres for GR and in the right hemispherefor CO In order to confirm this for GR we used ahigh-field 3-T magnet to perform a functional scanin which he viewed scenes faces and objects in thesame blocked fMRI design that we have used tolocalise the PPA in all our previous experiments(see Epstein et al 1999 for details Epstein ampKanwisher 1998) A Kolomogorov-Smirnov testfound no voxels in the parahippocampal region thatfit our previously established criterion for inclusionin the PPA (ie greater response to scenes than tofaces and objects at a uncorrected significance levelof plt 0001) This result contrasts sharply with thepattern seen in over 95 of (mostly young or mid-dle-aged) normal subjects in whom a large acti-vated region can be identified in both left and rightparahippocampal cortex using this comparison (seeFigure 5) A similar test was used to identify voxelsthat responded significantly more to faces than toobjects This comparison revealed a region of face-selective voxels in the right fusiform gyrus consis-tent with the location of the Fusiform Face Area
EPSTEIN ET AL
492 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 493
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
GR
CO
Figure 4 Structural MRIs of GR and CO showing the extent of the damaged region (marked as white) Images have been normalisedusing SPM99 and a cost function masking algorithm (provided by Matthew Brett) into standard stereotactic space (Talairach amp Tournoux1988) using the Montreal Neurological Institute template (Evans et al 1993) Numbers indicate y-Talairach coordinate for each coronalslice corresponding slice planes are also indicated on the sagittal image
(FFA) commonly seen in normal subjects(Kanwisher McDermott amp Chun 1997) Thusthe MRI was sensitive enough to detect functionaldifferences an FFA was found but no PPA Whencombined with the anatomical results this stronglysuggests that GR has no PPA
Because of safety concerns (presence of a femo-ral artery aneurysm clip) we did not perform a simi-lar high-field functional MRI of CO However asFigure 4 demonstrates CO clearly has damage tothe parahippocampal-lingual region in the righthemisphere
EXPERIMENTAL INVESTIGATIONS
In this section we report the results of four experi-ments designed to probe the consequences of dam-age to the lingual-parahippocampal region ingeneral and the PPA in particular Our primaryconcern in these experiments was to test GR andCOrsquos ability to perceive remember and recognisedifferent kinds of visual material Based on our pre-vious fMRI work we predicted that GR and CO
would have more difficulty with scene-like stimuli(ie stimuli that convey information about thegeometry of a space one can navigate through or actwithin) than with object-like stimuli Such a deficitcould account for at least some of the problems GRand CO encounter in real-world navigation
Beyond determining the kind of visual materialthat gets processed in parahippocampal cortex wealso wanted to determine the nature of the process-ing that takes place on that material In particularwe wanted to determine if parahippocampal cortexis involved in perceptual encoding memory encodingor recognition As discussed earlier (Introduction)our fMRI results provided some evidence againstplace recognition as the sole function of the PPAbut did not allow us to distinguish between percep-tual and mnemonic encoding Experiments 1 and 2examine the role of parahippocampal cortex in per-ceptual encoding Experiment 3 examines its role inmnemonic encoding and Experiment 4 examinesits role in place recognition
Experiment 1 Two-dot task
The first hypothesis we examined was thatparahippocampal cortex plays a role in perceivingthe shape of the local environment In our previousfMRI work we observed that the PPA respondsmuch more strongly during scene viewing than dur-ing object or face viewing even when subjects donothing more than watch the scenes passively with-out performing any other task This suggested thatscene perception alone may be sufficient to activatethe PPA Previous reports have indicated that dam-age to parahippocampal cortex and other medialtemporal regions leads to problems with memoryrather than perception (Bohbot et al 1998Maguire Burke Philips amp Staunton 1996 Pigottamp Milner 1993) However all of these previousresults were obtained on patients who had unilat-eral lesions GR has a bilateral occipital-temporallesion including clear damage to parahippocampalcortex in both hemispheres furthermore he reportssome kind of perceptual deficit We hypothesisedthat this deficit might involve difficulty perceivingor interpreting the geometry of scenes and otherstimuli with a similar geometric structure
EPSTEIN ET AL
494 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 5 Top row shows a single slice from a functional MRIscan performed on GR Bottom row shows results from a normalsubject (NK) for comparison Voxels that responded more to facesthan objects (left column) or more to scenes than faces and objects(right column) in a Kolomogorov-Smirnov test are highlighted bywhite arrows GR has a functioning face area (FFA) but there isno evidence of a place area (PPA)
FFA PPA
GR
NK
In this experiment subjects viewed either objectshapes or spatial layouts Stimuli consisted ofblack-and-white photographs of real-world stimuli(empty rooms vs common objects) or abstractstimuli made out of Lego blocks (Lego ldquoscenesrdquo vsLego ldquoobjectsrdquo) These stimuli have been previouslyshown to differentially activate the PPA in normalsubjects (Epstein et al 1999 Epstein ampKanwisher 1998) Two red dots were superim-posed on each photograph (Figure 6) The task wasto report which of the two red dots was over thecloser part of the object or scene This task wasdesigned to have no memory component (apartfrom remembering the instructions) but to requireaccurate 3D perception of objects and scenes
If GR or CO have difficulty perceiving or inter-preting the spatial layout of scenes then we pre-dicted they would do worse when the stimuli areempty rooms or Lego scenes than when they arereal-world or Lego objects Note that the criticalcomparison is between the Lego scenes and theLego objects as these stimuli are made of the samematerials and differ primarily in their surfacegeometry
MethodTesting for all experiments was done on aMacintosh computer The experiment consisted of80 trials (20 of each stimulus type randomly inter-mixed) Subjects began each trial by pressing thespace bar A black-and-white photograph from oneof the four stimulus categories (see earlier) thenappeared on the screen with two red dots on it Thetask was to determine which of the two red dotsindicated the closer part of the scene or objectSubjects pressed the ldquofrdquo key if the leftmost red dotwas closest and the ldquojrdquo key if the rightmost dotwas closest They were instructed to respond asquickly as possible without making errors Stimuliremained on the screen until a response was made
The stimulus set was constructed so that eachempty room was matched to a common object thathad red dots in the exact same location and eachLego layout was similarly matched to a Lego objectThus the locations of the red dots were counterbal-anced across scene and object stimuli controllingfor any effects of visual field cuts
Results and discussionResults for GR and CO and four elderly normalsubjects are shown in Table 3 Both GR and COcould do this task without difficulty and their per-formance was near ceiling There was no evidencethat they had any more trouble with the emptyrooms or Lego scenes than with the real or Legoobjects Reaction times were slightly longer for GRand CO than for normal subjects as one wouldexpect given their visual field deficits however this
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 495
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 3 Experiment 1 (two-dot) results (SD in parenthesis)
GR CO Normals
Proportion correctObject 095 080 093Lego object 100 095 098Empty room 100 100 099Lego room 095 100 096
Mean RTObject 2435 2932 2161 (980)Lego object 2052 3245 2029 (921)Empty room 2744 2291 1788 (959)Lego room 1904 2281 1630 (1127)
Figure 6 The four kinds of stimuli used in Experiment 1 Topleft lego layouts top right real-world layouts (empty rooms)bottom left lego objects bottom right real-world objects
difference did not appear to be significant Impor-tantly there was little evidence that GR or CO tooklonger to make their perceptual judgements for thereal or Lego scenes than for the real or Lego objectsThus contrary to our initial hypothesis this experi-ment found no clear evidence for selective impair-ment in the perception of scene-like stimuli
Experiment 2 Different-views matchingtask
The previous experiment demonstrated that GRand CO can extract information about the per-ceived distances between themselves and the sur-faces of an object or scene Thus in at least onesense their perception of these surfaces is pre-served However this does not mean that their abil-ity to represent the layout of a scene is unimpairedThe two-dot task requires on-line comparison oftwo surfaces in egocentric coordinates GR and COwould be able to perform this task even if they couldnot (1) represent more than two surfaces at a time(2) represent surfaces in other than strictly egocen-tric coordinates or (3) sustain surface representa-tions for longer than the time it takes to make acomparison It has been suggested that the parietallobes are involved in on-line processing of surfacegeometry in egocentric coordinates (Milner ampGoodale 1995) thus it is possible that the parietallobes (which are intact in GR and CO) may be suf-ficient to perform the two-dot task even in theabsence of more durable or complex surface repre-sentations in parahippocampal cortex
In Experiment 2 we examine GR and COrsquosability to create and sustain a representation of thegeometry of an object or scene-like layout The taskwas to view a photograph of a Lego object or Legoscene hold it in memory for 10 s and then deter-mine whether a second object or scene was the sameas the first In ldquosamerdquo trials the second photographdepicted the same scene or object depicted from adifferent viewing angle In ldquodifferentrdquo trials the sec-ond photograph depicted a different scene or objectchosen to be similar to the first one (Figure 7)Thus in order to successfully perform this tasksubjects had to form a representation of the surfacesof the object or scene sustain this representation in
memory for 10 s and then align it (presumablyusing mental rotation) to the second object orscene
MethodThe experiment consisted of two parts each with40 trials In the first part the stimuli were black andwhite photographs of objects made out of Legoblocks In the second part the stimuli were photo-graphs of ldquoscenesrdquo made out of Lego blocks whichwere designed to have a geometric structure similarto real-world places such as a roomor a city street
Subjects began each trial by pressing the spacebar A photograph of a Lego object or sceneappeared on the screen for 3 s followed by a 10-sblank interval A second photograph then appeareddepicting either (1) the same object or scene as thefirst photograph shown from a different viewingangle (offset approximately 35 degrees) or (2) a dif-ferent object or scene Subjects were instructed topress the ldquojrdquo key if the two photographs depicted thesame object or scene and the ldquof rdquo key if they did notThey were told to take as much time as they neededto make a decision but no extra time The secondphotograph remained on the screen until a responsewas made Half the trials were ldquosamerdquo trials and halfwere ldquodifferentrdquo trials
Results and discussionFigure 8 shows results for GR and CO as well as forsix elderly normal controls GRrsquos and COrsquos perfor-mance was comparable to that of the normal con-trols Furthermore GR and CO performed just aswell when the stimuli were Lego scenes as whenthey were Lego objects These results are unlikely toreflect a ceiling effect as performance for bothpatients and controls was around 80 correctThus both patients appear to be able to form a rep-resentation of the geometry of both scenes andobjects which they can sustain in short-termmemory
Experiment 3 N-back recognition memorytask
The previous experiment failed to find any impair-ment in GR and COrsquos ability to form a representa-
EPSTEIN ET AL
496 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
tion of the geometry of a scene or object andmaintain it for 10 s in the absence of the originalstimulus Not only does this result suggest that per-ception of scene-like stimuli is intact in thesepatients it further suggests that some short-termmemory functions are intact as well However we
should note that subjects in this experiment werefree to devote all their effort and attention in the 10-second blank interval to maintaining a memory ofthe stimulus Thus it is possible that GR and COcan use rehearsal mechanisms to maintain the stim-ulus in short-term memory during the blank inter-
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 497
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
(b) Lego object version
Figure 7 Stimuli and procedure for Experiment 2 (a) In the Lego layout version subjects see a Lego layout for 3 s followed by a blankscreen for 10 s A second layout then appears which is either the same layout as the first one but seen from a different angle or a differentlayout (b) The Lego object version is the same but with object stimuli
3 s 10 s OR
3 s 10 s OR
(a) Lego layout version
(b) Lego object version
val but are unable to form a more durable memorytrace for the stimulus This possibility was tested inthe current experiment which was designed toexamine GR and COrsquos ability to learn new visualinformation
We devised a continuous n-back recognitionmemory test in which subjects viewed a sequence oforiginally novel stimuli some of which wererepeated within the sequence at intervals of 1 3 or5 intervening images For each stimulus subjects
reported whether they had seen it before in theexperiment or not In different runs in the experi-ment the stimuli were black-and-white photo-graphs of (1) Lego scenes (2) Lego objects (3) real-world landscapes (4) novel objects made out ofSculpey clay (5) unfamiliar houses (6) unfamiliarfaces as well as (7) line drawings of nonsenseobjects and (8) pronounceable pseudowords (Fig-ure 9) Thus we were able to test GR and COrsquos rec-ognition memory for different kinds of materials atdifferent time intervals in a paradigm in which thepresence of intervening items between the twoappearances of each stimulus made rehearsal diffi-cult Note that because none of the specific stimuliused in this experiment were familiar in advancethis task did not require an episodic memory com-ponent and could be done on the basis of pure visualfamiliarity with the particular visual stimulus
Although many different stimulus classes weretested in this experiment the critical comparison isbetween the Lego layouts and Lego objectsbecause these are best matched with regard tomaterials and differ primarily in their geometricstructure (see Introduction) We predicted that ifGR and CO showed a memory deficit for this taskit would be greater for the Lego layouts than for the
EPSTEIN ET AL
498 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 8 Results of Experiment 2 Performance over ldquosamerdquotrials and ldquodifferentrdquo trials is averaged together Error bars show1 SD
Figure 9 Stimuli for Experiment 3 Top row Lego layouts real-world scenes houses Bottom row Lego objects clay objects nonsenseobjects faces In an eighth condition (not shown) subjects viewed pronounceable nonwords such as ldquowilchrdquo or ldquothipperrdquo
Lego objects In addition a deficit for materialsthat convey information about the layout of sur-rounding space might also be apparent when GRand CO are tested on more realistic stimuli If soone would expect them to do poorly on the land-scapes but relatively well on the clay objects linedrawings of nonsense objects faces andpseudowords
MethodFor each subject a complete experimental sessionconsisted of being run once on each of the eightstimulus classes Each run consisted of 90 trials ineach trial a stimulus was presented and subjectsreported whether or not they had seen it before inthe run (or indeed ever) by pressing the ldquojrdquo key ifthey had or the ldquof rdquo key if they had not Each stimu-lus was on the screen for either 1 2 or 3 s (see fol-lowing) and appeared promptly after the keypressreport for the previous trial Individual stimuliappeared either once (14 stimuli) twice (30 stim-uli) or four times (4 stimuli) within a given run Ofthe stimuli that were presented twice 10 wererepeated immediately (1-back) 10 were repeatedwith a lag of two intervening items (3-back) and 10were repeated with a lag of four intervening items(5-back) In order to ensure that between-subjectdifferences in performance were not due to stimulusdifferences the same stimuli were assigned to thesame conditions each time the experimentwas run
GR performed the experiment twice once inAugust 1998 and again 9 months later Because ofa computer error he was only tested on six of theeight categories (all except Lego layouts and Legoobjects) at the later date CO was also tested twiceonce in February 1999 and again 6 months laterReported results for GR and CO are the average oftheir two testing sessions Normal subjects wereonly tested once Stimulus presentation time was 1 sin GRrsquos first testing session In pilot testing COfound this too fast for the critical categories of Lego
layouts and Lego objects so presentation time wasextended to 2 s for these categories in his first test-ing session All normal subjects were tested at thesepresentation rates (2 s for Lego layouts and objects1 s for other stimuli) In COrsquos second testingsession presentation time was lengthened to 3 s forall stimulus categories to ensure that any deficitsfound were memory deficits rather than problemswith the nonmemory aspects of the task
Results and discussionResults from GR CO and 10 elderly normal con-trols are shown in Figure 10 and Table 4 Figure 10ashows results from the main comparison betweenLego layouts and Lego objects Both normal sub-jects and patients performed near ceiling for bothLego layouts and Lego objects at 1-backinterstimulus intervals verifying that GR and COcould successfully perceive the pictures as they werebeing presented In contrast there was a strikingdifference between the two patients and the normalsubjects at 5-back intervals Whereas normals didon average somewhat better on Lego layouts thanLego objects both GR and CO showed the oppo-site pattern doing much worse on Lego layoutsthan Lego objects The 3-back condition showed amixed pattern Whereas GR did better on Legoobjects than Lego layouts in this condition (similarto his performance at 5-back intervals) CO didbetter on Lego layouts than Lego objects COrsquos lowperformance on Lego objects in the 3-back condi-tion appears to be an anomaly as it is much lowerthan his performance for comparable stimuli in the1-back and 5-back conditions
Figure 10b shows the difference in performancebetween Lego layouts and Lego objects for thecritical 1-back and 5-back intervals At 5-backintervals CO was significantly impaired comparedto the normal subjects (z = ndash250) whereasGRrsquos impairment fell just short of significance (z =ndash175)4
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 499
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
4In fact these values may underestimate GR and COrsquos relative impairment for Lego layouts In the 5-back condition 4 of the 10
normal subjects had perfect performance on the Lego layouts but all scored below ceiling on the Lego objects Thus the true advantagefor Lego layouts over Lego objects for normal subjects is probably greater than that observed here in which case GR and COrsquos lowerperformance for Lego layouts compared to Lego objects would be even more abnormal Furthermore post-hoc analyses revealed thatone of the normal subjects showed a difference in performance between Lego layouts and objects that was over 2 SD below the mean ofthe other nine subjects When this outlying subject was excluded z-scores were ndash322 for CO and ndash232 for GR
Table 4 shows results for all eight stimulusclasses Although direct comparisons between sub-jects should be treated with caution since viewingtime for different items was not the same for eachsubject we do note a few interesting patterns Firstof all there is little or no evidence that GR or CO
show a memory deficit for nonsense words clayobjects or line drawings of nonsense objects GRappears to be slightly impaired on faces and housesbut CO performs normally on these categoriesStrikingly GR shows a severe deficit for the reallandscapes performing more than 40 percentagepoints lower than normals Thus even thoughthere is some evidence that GR might have someimpairment for nonscene stimuli he clearly is muchmore impaired for scene-like stimuli such as theLego scenes and landscapes CO does not show asimilar deficit for the landscapes However itshould be kept in mind that in one of his two testingsessions CO viewed each picture for 3 s which wasmuch longer than the 1 second used for all non-Lego stimuli for GR and normals Thus COrsquosapparently ldquonormalrdquo performance in these condi-tions might be overestimated
Taken together the results of this experimentindicate that GR and CO are selectively impaired atprocessing scene-like stimuli This deficit is consis-tently apparent at 5-back but not 1-back intervalsThis finding suggests that the deficit should becharacterised as a memory encoding problemHowever the present results do not allow us todetermine whether the difficulty occurs at memoryencoding or at subsequent recognition (see Epsteinamp Kanwisher 2001) If GR and CO have a generalproblem with recognition of topographical materi-als then they should not be able to recognise scenes
EPSTEIN ET AL
500 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 10a Results of Experiment 3 for GR (top) CO (middle)and for 10 age-matched normal controls Performance measure isproportion of repeated stimuli correctly identified as such correctedfor guessing using the standard formula
Figure 10b Difference between Lego layout performance andLego object performance for the 1-back and 5-back conditionsError bars indicate one 1 SD
and landmarks that were learned prior to theirinjury We tested this possibility in the nextexperiment
Experiment 4 Famous landmarksfacesrecognition task
In this experiment we examined GR and COrsquosability to recognise the visual appearances of placeslearned prior to injury Subjects saw a series offamous landmarks intermixed with nonfamouscontrols (Imaging experiments have demonstratedthat landmarks of this type can activate the PPAalmost as strongly as complete scenes Epstein etal 1999 Epstein amp Kanwisher 1998) They wererequired to say whether they knew each specificlandmark and to identify the ones they knew Sub-jects were instructed to respond affirmatively if theywere fairly certain that they recognised the land-mark but not if it only looked generally familiarFor purposes of comparison subjects also per-formed an identical test in which the stimuli werefamous and nonfamous faces In both testsdistractor items were designed to be visually andconceptually similar to the target items (see Figure11) If parahippocampal cortex is required for placerecognition then one might expect GR and CO toperform poorly compared to normals on the land-mark recognition test
MethodEach of the two experiments (famous faces famouslandmarks) consisted of 48 trials Subjects beganeach trial by pressing the space bar after which ablack-and-white photograph of a face (in thefamous faces experiment) or a landmark (in thefamous landmarks experiment) appeared on thescreen Subjects were told to press the ldquojrdquo key if theyrecognised the specific face or landmark and the ldquof rdquokey if they did not taking as much time at theyneeded to make this discrimination but no extratime Subjects were told to report that they recog-nised a face or landmark if they felt that they knewit even if they could not retrieve its name If theydid know the name they were to say it after pressingthe button Otherwise they were to say whateverthey did remember about the item
Half of the photographs were of famous and halfof nonfamous people and landmarks The famousfaces included public figures (eg George BushRichard Nixon Prince Charles) and movie stars(eg Harrison Ford Clark Gable Marlon Brando)who were prominent prior to the time of GR andCOrsquos injuries the famous landmarks were build-ings and monuments from all over the world (egEiffel Tower US Capitol Building Great Sphinx)The nonfamous stimuli were chosen to match thefamous stimuli as closely as possible Each famousstimulus had a ldquomatchrdquo For example George Bushwas matched to a man of similar age in a coat andtie and the Taj Mahal was matched to a visually
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 501
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 4 Full results of Experiment 3 Mean proportion correct (corrected for guessing) SDs in parentheses
Lego Land- Lego Clay Nonsense Non-scenes scapes Houses objects objects objects Words Faces
1-back GR 100 094 100 100 100 094 100 100CO 091 085 091 100 100 079 074 100Normals 096(008) 095(008) 093(014) 096(008) 096(006) 099(003) 098(004) 097(011)
3-back GR ndash003 068 026 047 083 083 075 052CO 066 081 037 010 094 069 095 081Normals 066(032) 077(021) 072(015) 077(020) 086(016) 083(015) 083(017) 072(020)
5-back GR 010 034 044 047 072 083 094 053CO 031 073 075 088 081 064 100 081Normals 076(026) 077(016) 068(021) 066(016) 091(014) 083(018) 093(011) 078(021)
similar Asian palace that is not famous (see Figure11)
Results and discussionResults for GR and CO as well as for five age-matched controls are shown in Figure 12 Both
GR and CO could recognise the famous landmarksas well normals In addition both GR and CO wereusually able to name the landmarks they success-fully identified
Thus GR and CO can recognize landmarkslearned prior to injury demonstrating that at leastsome of their place recognition abilities are intactWhen combined with the fact that GR and COwere able to produce detailed maps of places theylived in prior to their injury (see Case Description)these results suggest that their deficit is primarily inencoding or consolidation of new informationrather than in recognition or recall of old informa-tion Note however that these results do not provethat their place recognition abilities are entirelynormal GR and CO might suffer from a mild rec-ognition deficit that affects identification of newly-learned stimuli like the Lego layouts in Experiment3 but not overlearned stimuli like the famous land-marks Furthermore it is possible that GR and COmight be impaired at recognising premorbidlylearned places that are defined primarily by theirspatial layout even though they are unimpaired atrecognising premorbidly learned places that aredefined primarily by a single object-like landmarkor building (see General Discussion) Neverthelessthe trend of the present evidence is to suggest thatGR and CO have particular problems with noveltopographical information
In the famous-face task GRrsquos performance wasin the normal range However he sometimes haddifficulty naming the faces even though he clearly
EPSTEIN ET AL
502 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 11 Stimuli for Experiment 4 Each famous landmark(Arc de Triomphe Taj Mahal) and famous face (Barbara BushYasser Arafat) was matched to a nonfamous building or face thatalso appeared in the experiment
FA
CE
SL
AN
DM
AR
KS
Figure 12 Results of Experiment 4 Performance measure isproportion of famous stimuli reported as ldquoknownrdquo corrected forguessing using the standard formula
recognised them For example he described Ron-ald Reagan as ldquothe gipperrdquo Nancy Reagan as ldquothewife of the gipperrdquo and Clark Gable as ldquothe movieactor who said lsquoScarlett I donrsquot give a damnrsquordquo COhad a great deal of difficulty identifying the famousfaces This result is not surprising given hisextremely low performance on the Benton FaceMatching Task In addition to his topographicproblems CO appears to suffer fromprosopagnosia
GENERAL DISCUSSION
We tested two parahippocampal patients (GR andCO) on four experiments designed to measure per-formance as a function of both task (perceptionencoding and recognition) and stimulus type(scenes objects faces and words) The only taskthat revealed a clear deficit was an n-back recogni-tion memory test (Experiment 3) In order to suc-cessfully perform this task subjects had to encode adurable representation of a visual stimulus and useit for later recognition GR and CO were signifi-cantly more impaired on this task when the stimuliwere scene-like spatial layouts (ie Lego scenesand real scenes for GR) than when they wereobjects No comparable deficits were found in otherexperiments that tested layout perception (Experi-ments 1 and 2) and placelandmark recognition(Experiment 4) Taken together these results indi-cate that GR and CO are impaired in their ability toencode novel information about the geometry ofsurrounding space into memory but are lessimpaired at their ability to encode the geometry ofnovel objects
We argue that this specific topographical defi-citmdashinability to encode the spatial layout of novelscenesmdashresults from damage to the para-hippocampal place area Although the damage inboth patients extends beyond the parahippo-campal-lingual territory normally associated withthe PPA this region is clearly lesioned bilaterally inGR and in the right hemisphere in CO Further-more functional imaging demonstrated that GRhas no functioning PPA Thus the current resultsand the earlier fMRI work (Epstein et al 1999
Epstein amp Kanwisher 1998) converge to implicateparahippocampal cortex in the encoding of infor-mation about scene geometry into memory
Our assignment of the function of memoryencoding to parahippocampal cortex is consistentwith the results of earlier case reports of patientswith parahippocampal damage who have naviga-tional difficulties in novel but not familiar environ-ments (Habib amp Sirigu 1987 Ross 1980 see alsoTeng amp Squire 1999) As with GR and CO thesepatients were either less impaired or unimpaired onnontopographical memory tasks Patients have alsobeen described who exhibit the opposite pattern ofimpairment an inability to learn faces (sometimescalled ldquoprosopamnesiardquo Cipolotti Robinson Blairamp Frith 1999 Tippett Miller amp Farah 2000) orfaces and verbal materials (Maguire amp Cipolotti1998) with an unimpaired ability to learn topo-graphical materials Taken together these resultsargue for a high degree of domain-specificity in theneural systems involved in learning of new informa-tion (Gallistel 1995) The brain may have separatemechanisms for learning faces places and wordswhich can be differentially impaired after damageto specific cortical regions (Cipolotti et al 1999Incisa della Rochetta et al 1996) The presentresults suggest that the anatomical locus of themechanism dedicated to learning new topographi-cal information is in the parahippocampal cortex(Aguirre amp DrsquoEsposito 1999)
In contrast to their impaired ability to encodenew topographical information both GR and COwere able to produce detailed maps of their previousdwellings and performed normally on the famouslandmark recognition test of Experiment 4 Theseresults are generally consistent with Teng andSquirersquos (1999) report of a patient with extensivemedial temporal damage who could recallpremorbidly learned but not postmorbidly learnedtopographical information (see also Ross 1980)Thus our results provide further evidence thatmedial temporal regions such as parahippocampalcortex may be more critical for encoding or consoli-dating new information into memory than for recallof old information
One issue that our experiments do not entirelyresolve is the precise nature of the topographical
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 503
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
information encoded by parahippocampal cortexClearly this region is critical for encoding informa-tion about the geometry of the currently visiblescene even when the ldquoscenerdquo is made out of Legoblocks and therefore has no immediate navigationalrelevance However it is unclear whether this spe-cific deficit can account for all of the real-worldnavigational difficulties encountered by GR andCO GR in particular shows such a catastrophicloss of navigational ability that we suspect he is alsoimpaired at encoding the spatial relationshipsbetweenthe current scene and other locations in theworld CO probably also suffers from such difficul-ties as he reports that he has trouble rememberinghow places relate to each other in large-scale spaceWhen he was tested on a simple 3-room videogameenvironment he not only was unable to rememberthe individual rooms (claiming that there were 10rooms when there were only 3) but he also had nomemory for the spatial relationships between themThese spatial deficits may be the result of damage tothe posterior right hippocampus (in the case of CO)or of disconnection of the posterior right hippo-campus from the parahippocampal regions thatprovide much of its cortical input (in both patientsSuzuki amp Amaral 1994) In either case the abilityto form a cognitive map of new environments likethe lab or videogame space could be impaired(Maguire et al 1998 OrsquoKeefe amp Nadel 1978)However it is also possible that parahippocampalcortex itself may be critical for encoding not onlythe geometry of individual scenes but also the spa-tial relationships between the locations of thosescenes In this case the spatial deficits of GR andCO could be attributed directly to parahippo-campal damage rather than to hippocampal dam-age or disconnection
The latter scenario would be consistent withAguirre and DrsquoEspositorsquos (1999) and Barrash etalrsquos (2000) suggestion that parahippocampal cortexis critically involved in combining visual informa-tion about the appearance of scenes and landmarkswith spatial information about their locations into aunified topographical representation Howeverboth of these authors argue that the representationsof individual scenes (Barrash et al) and landmarks(Aguirre amp DrsquoEsposito) are supported not by
parahippocampal cortex but by more posterior lin-gual-fusiform regions In contrast we believe thatthe neuroimaging data indicate that parahippo-campal cortex is involved in the encoding of thegeometric structure of individual scenes althoughit may also play a role in combining these represen-tations with other spatial representations
Interestingly GR and CO were not impaired onrecognition of famous landmarks that were mostlybuildings Thus our data do not preclude the possi-bility that there may be a separate lingual orfusiform region involved in processing informationabout buildings (Aguirre Zarahn amp DrsquoEsposito1998 Chao Haxby amp Martin 1999 IshaiUngerleider Martin Schouten amp Haxby 1999)which might have been preserved in GR and COdespite extensive damage to other regions of thelingual and fusiform gyrus Alternatively GR andCOrsquos preserved building recognition abilities mightbe mediated by interactions between cortical areasthat perform shape analysis for any object-likestimulus type (Malach et al 1995Kanwisher et al1996) and anterior temporal regions in which long-term topographical and semantic memories arestored (McCarthy et al 1996)
Is perception preserved
In contrast to their inability to encode new scenesinto memory GR and CO have substantiallyretained their ability to perceive the spatial layout ofthe currently visible scene (Experiments 1 and 2)GR can follow a route marked on a floor planwhich indicates a good on-line appreciation of hiscurrent spatial surroundings Furthermore neitherGR or CO have any trouble moving through theenvironmentmdashthey move with confidence and donot bump into doors or walls Problems of the lattertype are usually associated with damage to the pari-etal lobes which are intact in GR and CO Thus atfirst glance GR and CO do not appear to haveperceptual impairments (beyond their visual fieldrestrictions)
However GR does complain of a perceptualdeficit which he describes as an inability to ldquotakeinrdquo scenes when they are too complex Similarreports of an inability to comprehend the
EPSTEIN ET AL
504 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
ldquoglobalityrdquo of a scene were made by some of theparahippocampally-damaged patients studied byHabib and Sirigu (1987) We were not able todetermine the precise nature of this deficit whichmay be partly attributable to his visual field restric-tions It appears that his deficit is more salient in thereal-world environments than in the picture-per-ception tasks of Experiments 1 and 2 perhapsbecause he finds it more difficult to organise visualinput when immersed in it than when viewing pho-tographs that subtend only part of the visual field
We suggest that GR can perceive parts of thescene when he attends to them but may be unableto integrate information over time into a coherentrepresentation of a scene or place The fact that GRand CO are impaired for Lego layouts at 5-backintervals in Experiment 3 is consistent with thishypothesis The time between the first and secondpresentations of a stimulus is very short in this case(and even shorter with 3-back intervals where GRalso shows evidence of a deficit) In both patientsthe representation of layout information formed byperception of a Lego scene must either have anabnormally short persistence be abnormally sus-ceptible to interference or both If so they mightfind it impossible to sustain a spatial frame orschema long enough to use it to organise informa-tion obtained from many different glances at a com-plex scene (Intraub 1997) The result would be aninability to understand the ldquoglobalityrdquo of a sceneeven though all the details were perfectlyperceived
Non-topographical mnemonic abilities
Although not as severe as their topographical mem-ory problems GR and CO do have some memorydifficulties in other domains For example bothperformed poorly on the Rey AVLT whichrequired them to remember a list of unrelatedwords Both also report some subtle episodic mem-ory problems (see Case Description) COrsquos prob-lems could partly result from damage to the righthippocampusmdashindeed CO appeared to have themore acute episodic memory problems HoweverGR has no hippocampal damage so the fact that hedisplays episodic memory problems suggests thatparahippocampal cortex is involved in the forma-
tion of episodic memories (cf Wagner et al1998)
In previous work (Epstein et al 1999) weargued that spatial context information encoded byparahippocampal cortex might play a role in estab-lishing event memories by encoding informationabout where events take place The inability toencode this information might cause problems intasks such as the Rey AVLT where one has to recalla list of unrelated words If spatial (and perhapstemporal) context information is not encoded itwill be harder subsequently to recall the distinctevent of hearing each word (Tulving 1983) Incontrast lack of spatial context information wouldbe less of a problem in tasks where one is onlyrequired to recognise previously encountered verbalmaterials but not to reproduce the spatiotemporalcontext in which they were experienced IndeedGR and CO performed as well as normals on thenonsense words in Experiment 4 where recogni-tion memory but not episodic memory was requiredfor successful performance Interestingly GR andCO also did relatively well on the Logical Memorysubtest of the WMS-III perhaps because the nar-rative framework of a story provides sufficient con-textual information to overcome the absence ofspatial context and mediate successful verbalretrieval
Differences between GR and CO
Although we have discussed GR and CO togetherthere are a number of relevant differences betweenthem One open question is whether these differ-ences can be attributed to the different effects ofunilateral and bilateral parahippocampal damageIn general GRrsquos topographical difficulties weremore severe than COrsquos CO did surprisingly well onthe Warrington Topographical Memory Test inwhich he was required to remember detailed real-world city scenes and also performed substantiallybetter on real scenes than Lego scenes in Experi-ment 3 Insofar as CO has intact left para-hippocampal cortex this suggests that the left PPAmight be sufficient to encode a certain amount of(perhaps semantic) information about real-worldscenes but the right PPA is necessary to encode
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 505
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)

performance was 714 correct hits for the familiarlandmarks and 1314 correct rejections for theunfamiliar landmarks In contrast his wifeobtained a perfect score on this test Given theextensive nature of the training regime GRrsquos scorestrongly suggests an impairment in learning theappearance of new real-world places Interestinglyseveral of the seven familiar landmarks that he gotright had an obvious distinctive feature that hadbeen explicitly pointed out to him For exampleone building had a distinctive clock tower andanother one was an unusual triangular shape ThusGR was occasionally able to use distinct visualfeatures to identify places even though he wasunable to recognise them holisticallymdasha commonpattern among topographical disorientationpatients
GR did not appear to have any obvious problemsin understanding the spatial structure of his imme-diate environment When given a floor plan of thebuilding at MIT in which he was being tested hewas able to successfully follow a route marked onthe plan However when the plan was taken awayhe had great difficulty finding his way back to thelab even though he had more than 30 min experi-ence with the environment and his starting positionwas less than 30 feet from the goal (Figure 2) Infact the route he took resembled a random walk Atone point he walked right past his destinationwithout recognising it and only knew for sure thathe was in the right place when he identified his wifein the room Thus his on-line appreciation of thelayout of immediately visible space appears to beintact but he cannot find his own way withoutassistance
We were unable to perform the MIT landmarkrecognition test on CO because he suffered from amedical problem that made it difficult for him towalk around campus In lieu of this we tried to seewhether he could learn a simple video game envi-ronment consisting of three connected roomsAfter about 30 min experience navigating throughthis environment (via button presses) he was askedto sketch a map of it He declined to do so statingthat it was beyond his abilities When asked howmany rooms there were he replied that he did notknow but that there were at least 10 Given the
simplicity of this artificial environment COrsquos com-plete inability to learn it is striking
Map drawingAfter several days of testing we asked both GR andhis wife to sit at different tables and independentlysketch (1) the shape of Florida (2) a bicycle (3) afloor plan of their hotel room in Boston (4) a floorplan of the first floor of the house they had been liv-ing in for the last 8 months (all post-stroke) (5) afloor plan of the main room of the lab and (6) afloor plan of the house they lived in 10 years ago(pre-stroke) GR drew the map of Florida and thebicycle perfectly His plan of the hotel room wasnotably different from the one drawn by his wifemdashfor example he reported that there was only onebed when in fact there were two and he placed thebed on the wrong side of the room (Figure 1a) Hisplan of the first floor of his current house was some-what better but was not very detailed and includedsome major errors For example he wronglyreported that there was no bathroom on this floor(Figure 1b) He was unable to produce any floorplan of the main room of the lab leaving this pageblank When his wife reminded him that this wasthe room in which he had just spent an hour readingthe newspaper and in which we had all had break-fast in that morning he replied that he didnrsquot knowwhat room she was talking about (although heappeared to remember these events) In contrast tothis near total inability to produce accurate maps ofnew environments GR produced an accurate mapof the house he lived in prior to his injury (Figure1c) He reported that his memory of this house wasldquoall thererdquo and he could imagine walking around init When asked to produce a perspective of a roomwhen he was in it he did so with ease and he wasalso able to create an accurate floor plan of theroom
In contrast CO found it very difficult to pro-duce a floor plan of the lab both when he was in itand when he was taken to another room and askedto recall it However although the maps he pro-duced were quite impoverished they were notentirely inaccurate demonstrating some ability toencode topographical information This contrastswith GRrsquos absolute inability to recall anything
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 487
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
EPSTEIN ET AL
488 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure
1F
loorplansdraw
nby
GR
andhisw
ifeof(a)
theirhotelroomin
Boston(b)theircurrenthousew
hichthey
hadm
ovedto
subsequenttoG
Rrsquosinjury(c)the
housethey
livedin
priorto
GR
rsquosinjury
Ho
telro
om
inB
osto
n
GR
His
wife
His
wife
Cu
rren
th
om
e
GR
His
wife
Fo
rme
rh
om
e
GR
about the main room of the lab once he had left itCO was also able to draw a detailed floor plan of theapartment he was living in at the time (and hadlived in for 20 years) Several months after he hadmoved from this apartment we asked him to drawsuch a floor plan again and he produced one thatwas substantially the same as the first (Figure 3)Thus their map drawing performance indicatesthat GR and CO are primarily impaired at sketch-ing places experienced after their strokes but areunimpaired at sketching places experienced beforetheir strokes This pattern is indicative of deficits inencoding but not recall
Scene perceptionAs noted earlier GR can navigate with the aid of afloor plan suggesting that his perceptual abilitiesare largely intact Interestingly however he reportsthat he does have a perceptual deficit which hedescribes as an inability to understand scenes whenthey are too complex For example he reports thathe often has trouble using maps because theyusually are too busy with details It is unclearwhether this apparent perceptual problem is a resultof his field cut some other separate perceptual
problem or a memory problem (see discussion)CO did not complain of a similar deficit GR wasan accomplished artist prior to his injury producingpaintings of striking complexity Tragically hereports that he cannot now appreciate his ownpaintings
Other cognitive abilities
Face recognitionGR also reported some difficulties with face recog-nition On his second visit we examined his abilityto learn new faces by having him perform a forced-choice recognition test on seven familiar and sevenunfamiliar faces The familiar faces were personnelfrom the lab with whom he had at that point had agood deal of experience spanning at least 3 daysAll the people in the photographs had a towelwrapped around their head in order to minimise theuse of hair as a nonfacial cue Despite the difficultyof this task he correctly identified 67 familiarfaces and made no false alarms Surprisingly theone face that he did miss was the first author of thispaper with whom he at that point had had consid-erable experience GR also performed normally ona difficult famous face recognition test (see Experi-ment 4 following) and on the Benton Face Recog-nition test (see following) Thus he has someability to recognise faces Consistent with this GRreports that he can remember a small number offaces but that he quickly ldquoloses trackrdquo if he has toremember too many
CO also appears to suffer from someprosopagnosia as evidenced by his low perfor-mance on the Benton Face Recognition test (seefollowing see also Experiment 4) Interestingly hedoes not complain of face recognition difficultiesbut rather attributes his inability to recognise peo-ple to a lack of interest in their appearance
Episodic memoryAt first glance GRrsquos episodic memory appeared tobe largely intact As mentioned he could recall theevents of the previous day accurately Furthermorehe reports that he can follow the plot of a moviewithout any difficulty However both he and hiswife report that he does have some problems with
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 489
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Figure 2 Floor plan of the building at MIT in which GR wastested Corridors are highlighted in grey GR was asked to find themain room of the lab (marked G on the map) Starting location(marked S) was a point approximately 30 feet down the hall GRrsquosroute is marked by arrows GR found this task very difficult andonly knew for certain that he had reached his goal when he caughtsight of his wife through the glass door of the lab In contrast hewas able to follow a route marked on a plan of the same floorwithout any problem or hesitation
episodic memory For example he stated that heremembered very little about a trip to Italy they hadtaken several months previously His wife reportsthat he has great difficulty keeping track of timeFor example before their trip to Boston he repeat-edly asked her what day they were supposed to leaveand seemed unable to encode this information Weobserved a similar phenomenon during testingDuring the tests that required repetition of thesame procedure several times he sometimesseemed unable to clearly remember how manytimes he had repeated the procedure Thus he doesappear to have a subtle episodic memory deficit
which may manifest itself as an inability to sepa-rately remember successive events that are not easilydistinguishable from each other
CO also reports some problems with his epi-sodic memory He relies on lists to rememberappointments stating ldquoif I didnrsquot have a list Irsquodprobably forget why I walked in the door hererdquoDespite this claim he has never missed an appoint-ment nor is he ever confused about the purpose ofthe testing appointments He can accurately reportthe day of the week He remembers the experi-menters clearly from appointment to appointmentas well as the details of the various experiments
EPSTEIN ET AL
490 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 3 Two floor plans drawn by CO of the apartment he lived in for over 20 years The first plan was drawn when CO still lived inthe apartment the second was drawn several months later when he had moved out Note that the two plans correspond very well COreported that it was quite easy to produce these floor plans
CO home Feb 99
CO (former) home June 99
when they are repeated across appointments Whenqueried about his episodic memory difficulties COreported that he will usually remember an event ifhe is reminded of it but that he has difficulty spon-taneously regenerating events Interestingly hereports that even when he is reminded of an eventand remembers it he cannot remember ldquowhere itwas or when it wasrdquo When he reads he finds it dif-ficult to follow a story if it skips around Howeverwhen queried he was able to accurately report whathe had read in the newspaper the previous day
Formal neuropsychological tests
Formal neuropsychological testing was performedon GR in September 1998and on CO in December1998 Test performances were compared to age-matched peers3
GR exhibited mild to moderate impairment inmemory relative to what would be predicted for hisintellect (WAIS-R Full Scale IQ =122 93rd per-centile superior range) and level of educationThese memory problems were more apparent onvisual than on verbal memory tests On a test ofmemory for conceptually organised text passages(Wechsler Memory Scale Logical Memory[WMS-III] Wechsler 1997b) his performancewas average for both immediate (LM I =25th per-centile standard score SS = 90) and delayed recall(LM II =50th percentile SS =100) In contrast hisscore on the WMS-III Face Memory subtest was inthe low average range for both immediate (9th per-centile SS =80) and delayed recall (16th percentileSS =85) On a test of memory for scenes (WMS-IIIFamily Pictures subtest) immediate recall was defi-cient (2nd percentile SS = 69) and delayed recallwas borderline impaired (5th percentile SS =76)GR also had difficulty learning word lists on theRey Auditory Verbal Learning Test (Spreen ampStrauss 1998) his immediate recall was average(63rd percentile SS = 105) but after the interfer-ence list his performance fell to the low averagerange for both immediate (14th percentile SS =84)
and delayed recall (16th percentile SS =85) On arecognition test of the items of the list he success-fully recognised all 15 but made 8 false alarms Per-formance on the 724 Spatial Recall Test (RaoHammeke McQuillen Khatri amp Lloyd 1984)which required him to remember seven locations ona 6times4 checkerboard was 66th percentile (SS =106)for immediate and 2nd percentile (SS = 69) fordelayed recall Performance on the Benton FaceRecognition test was normal (42nd percentile)Performance on the Boston Naming Test was atceiling indicating that language abilities wereintact Thus GR exhibits some impairment ontasks that require delayed recall of visual informa-tion (WMS-III Face Memory subtest) or verbalinformation that is not organised in terms of a storyor plot (Rey AVLT) and is clearly impaired ontasks that required delayed recall of spatial informa-tion (724 Spatial Recall Test) or stimulus location(WMS-III Family Pictures subtest) In contrasthis ability to recall organised verbal information(WMS-III Logical Memory subtest) appears to beintact
As with GR COrsquos memory performance wasgenerally below what would be expected based onintellect (WAIS-III Full Scale IQ =134 99th per-centile very superior range) and education levelAlso like GR there was some evidence that visualmemory problems were more pronounced than ver-bal memory problems Memory for text passages(WMS-R Logical Memory) was average for bothimmediate (27th percentile SS = 91) and delayedrecall (48th percentileSS =99) In contrast perfor-mance on the WMS-III Faces subtest was lowaverage for both immediate (16th percentile SS =85) and delayed recall (9th percentile SS =80) andperformance on the Family Pictures subtest wasdeficient (lt 1st percentile SSlt 62) for both imme-diate and delayed recall On the Warrington Rec-ognition Memory Tests (RMT) he scored in theaverage range for words (50th percentile SS =100)and scenes (Warrington Topographical Recogni-tion Memory Test 29th percentile SS =92) and in
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 491
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
3The formal neuropsychological testing reported in this section was performed prior to the beginning of our combined
investigation of GR and CO (ie prior to the main experimental investigations of this paper) Consequently GR and CO were nottested on exactly the same battery of tests
the deficient range (lt 5th percentile SS lt 75) forfaces The low score for faces on this test as well asthe WMS-III may reflect prosopagnosia ratherthan a memory problem as he scored in the defi-cient range (lt1st percentile SSlt62) on the BentonFace Recognition test Verbal list learning (ReyAVLT) was average on immediate recall (45th per-centile SS =98) but after hearing an interferencelist performance fell to the deficient level (015words recalled) for both immediate and delayedrecall On a recognition test for these words herecognised 1115 and made one false alarm (39thpercentile SS =96) There was a large discrepancyon tests of memory span which may indicate adecline in spatial memory digit span was superior(9+ forward and 7 backwards both 99th percentile)but spatial span was only average (5 forward and 5backward 37th and 50th percentiles respectively)Language abilities were intact (Boston NamingTest performance was at ceiling) Thus like GRCO shows some impairment at recall of visualinformation (WMS-III Face Memory subtest) andverbal information that is not organised into a nar-rative (Rey AVLT) and clear impairment at tasksthat require recall of stimulus location (Family Pic-tures subtests of WMS-III) In contrast his abilityto recall organised verbal information (WMS-RLogical Memory subtest) is more preserved
MRI RESULTS
In order to determine the extent of their lesionsstructural MRI images were taken of both subjectsDelineation of the lesions was based either onspoiled GRASS coronal images at 11mm thick-ness (GR) or anatomical T-1 weighted MPRAGEMRI sequences yielding 125 mm thick coronalimages (CO) The anterior-posterior extent of thelesions was verified on axial images
GR has bilateral occipital-temporal lesions(Figure 4) In the left hemisphere the posteriorparahippocampal gyrus (including para-hippocampal cortex) is severely damaged with onlyminimal injury to the most caudal aspect of the hip-pocampus There is also moderate damage to themedial fusiform and inferior lingual gyri In the
right hemisphere the hippocampus is spared butthere is damage to the posterior end of theparahippocampal gyrus near the parahippocampal-lingual boundary More posteriorly the lesionedarea extends along the collateral sulcus to includesignificant portions of the inferior lingual gyrus andthe medial fusiform gyrus as well as most of themedial occipital lobe below the calcarine fissure
CO has a right-hemisphere occipital-temporallesion Right posterior hippocampus and para-hippocampal gyrus are severely affected Moreposteriorly the damage covers large portions of theinferior lingual and medial fusiform gyri as well assubstantial portions of the medial occipital lobeincluding most of right calcarine cortex In addi-tion diffuse foci of increased signal on T-2weighted images was present in periventricularwhite matter in the corona radiata and centrumsemiovale with a lacunar infarct of the rightputamen There was also moderate degree of dif-fuse involutional changes with increased ventricularsize particularly notable in the right temporal hornwith ex vacuo dilitation
The structural MRIs suggested that the PPAwas severely if not completely damaged in bothhemispheres for GR and in the right hemispherefor CO In order to confirm this for GR we used ahigh-field 3-T magnet to perform a functional scanin which he viewed scenes faces and objects in thesame blocked fMRI design that we have used tolocalise the PPA in all our previous experiments(see Epstein et al 1999 for details Epstein ampKanwisher 1998) A Kolomogorov-Smirnov testfound no voxels in the parahippocampal region thatfit our previously established criterion for inclusionin the PPA (ie greater response to scenes than tofaces and objects at a uncorrected significance levelof plt 0001) This result contrasts sharply with thepattern seen in over 95 of (mostly young or mid-dle-aged) normal subjects in whom a large acti-vated region can be identified in both left and rightparahippocampal cortex using this comparison (seeFigure 5) A similar test was used to identify voxelsthat responded significantly more to faces than toobjects This comparison revealed a region of face-selective voxels in the right fusiform gyrus consis-tent with the location of the Fusiform Face Area
EPSTEIN ET AL
492 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 493
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
GR
CO
Figure 4 Structural MRIs of GR and CO showing the extent of the damaged region (marked as white) Images have been normalisedusing SPM99 and a cost function masking algorithm (provided by Matthew Brett) into standard stereotactic space (Talairach amp Tournoux1988) using the Montreal Neurological Institute template (Evans et al 1993) Numbers indicate y-Talairach coordinate for each coronalslice corresponding slice planes are also indicated on the sagittal image
(FFA) commonly seen in normal subjects(Kanwisher McDermott amp Chun 1997) Thusthe MRI was sensitive enough to detect functionaldifferences an FFA was found but no PPA Whencombined with the anatomical results this stronglysuggests that GR has no PPA
Because of safety concerns (presence of a femo-ral artery aneurysm clip) we did not perform a simi-lar high-field functional MRI of CO However asFigure 4 demonstrates CO clearly has damage tothe parahippocampal-lingual region in the righthemisphere
EXPERIMENTAL INVESTIGATIONS
In this section we report the results of four experi-ments designed to probe the consequences of dam-age to the lingual-parahippocampal region ingeneral and the PPA in particular Our primaryconcern in these experiments was to test GR andCOrsquos ability to perceive remember and recognisedifferent kinds of visual material Based on our pre-vious fMRI work we predicted that GR and CO
would have more difficulty with scene-like stimuli(ie stimuli that convey information about thegeometry of a space one can navigate through or actwithin) than with object-like stimuli Such a deficitcould account for at least some of the problems GRand CO encounter in real-world navigation
Beyond determining the kind of visual materialthat gets processed in parahippocampal cortex wealso wanted to determine the nature of the process-ing that takes place on that material In particularwe wanted to determine if parahippocampal cortexis involved in perceptual encoding memory encodingor recognition As discussed earlier (Introduction)our fMRI results provided some evidence againstplace recognition as the sole function of the PPAbut did not allow us to distinguish between percep-tual and mnemonic encoding Experiments 1 and 2examine the role of parahippocampal cortex in per-ceptual encoding Experiment 3 examines its role inmnemonic encoding and Experiment 4 examinesits role in place recognition
Experiment 1 Two-dot task
The first hypothesis we examined was thatparahippocampal cortex plays a role in perceivingthe shape of the local environment In our previousfMRI work we observed that the PPA respondsmuch more strongly during scene viewing than dur-ing object or face viewing even when subjects donothing more than watch the scenes passively with-out performing any other task This suggested thatscene perception alone may be sufficient to activatethe PPA Previous reports have indicated that dam-age to parahippocampal cortex and other medialtemporal regions leads to problems with memoryrather than perception (Bohbot et al 1998Maguire Burke Philips amp Staunton 1996 Pigottamp Milner 1993) However all of these previousresults were obtained on patients who had unilat-eral lesions GR has a bilateral occipital-temporallesion including clear damage to parahippocampalcortex in both hemispheres furthermore he reportssome kind of perceptual deficit We hypothesisedthat this deficit might involve difficulty perceivingor interpreting the geometry of scenes and otherstimuli with a similar geometric structure
EPSTEIN ET AL
494 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 5 Top row shows a single slice from a functional MRIscan performed on GR Bottom row shows results from a normalsubject (NK) for comparison Voxels that responded more to facesthan objects (left column) or more to scenes than faces and objects(right column) in a Kolomogorov-Smirnov test are highlighted bywhite arrows GR has a functioning face area (FFA) but there isno evidence of a place area (PPA)
FFA PPA
GR
NK
In this experiment subjects viewed either objectshapes or spatial layouts Stimuli consisted ofblack-and-white photographs of real-world stimuli(empty rooms vs common objects) or abstractstimuli made out of Lego blocks (Lego ldquoscenesrdquo vsLego ldquoobjectsrdquo) These stimuli have been previouslyshown to differentially activate the PPA in normalsubjects (Epstein et al 1999 Epstein ampKanwisher 1998) Two red dots were superim-posed on each photograph (Figure 6) The task wasto report which of the two red dots was over thecloser part of the object or scene This task wasdesigned to have no memory component (apartfrom remembering the instructions) but to requireaccurate 3D perception of objects and scenes
If GR or CO have difficulty perceiving or inter-preting the spatial layout of scenes then we pre-dicted they would do worse when the stimuli areempty rooms or Lego scenes than when they arereal-world or Lego objects Note that the criticalcomparison is between the Lego scenes and theLego objects as these stimuli are made of the samematerials and differ primarily in their surfacegeometry
MethodTesting for all experiments was done on aMacintosh computer The experiment consisted of80 trials (20 of each stimulus type randomly inter-mixed) Subjects began each trial by pressing thespace bar A black-and-white photograph from oneof the four stimulus categories (see earlier) thenappeared on the screen with two red dots on it Thetask was to determine which of the two red dotsindicated the closer part of the scene or objectSubjects pressed the ldquofrdquo key if the leftmost red dotwas closest and the ldquojrdquo key if the rightmost dotwas closest They were instructed to respond asquickly as possible without making errors Stimuliremained on the screen until a response was made
The stimulus set was constructed so that eachempty room was matched to a common object thathad red dots in the exact same location and eachLego layout was similarly matched to a Lego objectThus the locations of the red dots were counterbal-anced across scene and object stimuli controllingfor any effects of visual field cuts
Results and discussionResults for GR and CO and four elderly normalsubjects are shown in Table 3 Both GR and COcould do this task without difficulty and their per-formance was near ceiling There was no evidencethat they had any more trouble with the emptyrooms or Lego scenes than with the real or Legoobjects Reaction times were slightly longer for GRand CO than for normal subjects as one wouldexpect given their visual field deficits however this
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 495
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 3 Experiment 1 (two-dot) results (SD in parenthesis)
GR CO Normals
Proportion correctObject 095 080 093Lego object 100 095 098Empty room 100 100 099Lego room 095 100 096
Mean RTObject 2435 2932 2161 (980)Lego object 2052 3245 2029 (921)Empty room 2744 2291 1788 (959)Lego room 1904 2281 1630 (1127)
Figure 6 The four kinds of stimuli used in Experiment 1 Topleft lego layouts top right real-world layouts (empty rooms)bottom left lego objects bottom right real-world objects
difference did not appear to be significant Impor-tantly there was little evidence that GR or CO tooklonger to make their perceptual judgements for thereal or Lego scenes than for the real or Lego objectsThus contrary to our initial hypothesis this experi-ment found no clear evidence for selective impair-ment in the perception of scene-like stimuli
Experiment 2 Different-views matchingtask
The previous experiment demonstrated that GRand CO can extract information about the per-ceived distances between themselves and the sur-faces of an object or scene Thus in at least onesense their perception of these surfaces is pre-served However this does not mean that their abil-ity to represent the layout of a scene is unimpairedThe two-dot task requires on-line comparison oftwo surfaces in egocentric coordinates GR and COwould be able to perform this task even if they couldnot (1) represent more than two surfaces at a time(2) represent surfaces in other than strictly egocen-tric coordinates or (3) sustain surface representa-tions for longer than the time it takes to make acomparison It has been suggested that the parietallobes are involved in on-line processing of surfacegeometry in egocentric coordinates (Milner ampGoodale 1995) thus it is possible that the parietallobes (which are intact in GR and CO) may be suf-ficient to perform the two-dot task even in theabsence of more durable or complex surface repre-sentations in parahippocampal cortex
In Experiment 2 we examine GR and COrsquosability to create and sustain a representation of thegeometry of an object or scene-like layout The taskwas to view a photograph of a Lego object or Legoscene hold it in memory for 10 s and then deter-mine whether a second object or scene was the sameas the first In ldquosamerdquo trials the second photographdepicted the same scene or object depicted from adifferent viewing angle In ldquodifferentrdquo trials the sec-ond photograph depicted a different scene or objectchosen to be similar to the first one (Figure 7)Thus in order to successfully perform this tasksubjects had to form a representation of the surfacesof the object or scene sustain this representation in
memory for 10 s and then align it (presumablyusing mental rotation) to the second object orscene
MethodThe experiment consisted of two parts each with40 trials In the first part the stimuli were black andwhite photographs of objects made out of Legoblocks In the second part the stimuli were photo-graphs of ldquoscenesrdquo made out of Lego blocks whichwere designed to have a geometric structure similarto real-world places such as a roomor a city street
Subjects began each trial by pressing the spacebar A photograph of a Lego object or sceneappeared on the screen for 3 s followed by a 10-sblank interval A second photograph then appeareddepicting either (1) the same object or scene as thefirst photograph shown from a different viewingangle (offset approximately 35 degrees) or (2) a dif-ferent object or scene Subjects were instructed topress the ldquojrdquo key if the two photographs depicted thesame object or scene and the ldquof rdquo key if they did notThey were told to take as much time as they neededto make a decision but no extra time The secondphotograph remained on the screen until a responsewas made Half the trials were ldquosamerdquo trials and halfwere ldquodifferentrdquo trials
Results and discussionFigure 8 shows results for GR and CO as well as forsix elderly normal controls GRrsquos and COrsquos perfor-mance was comparable to that of the normal con-trols Furthermore GR and CO performed just aswell when the stimuli were Lego scenes as whenthey were Lego objects These results are unlikely toreflect a ceiling effect as performance for bothpatients and controls was around 80 correctThus both patients appear to be able to form a rep-resentation of the geometry of both scenes andobjects which they can sustain in short-termmemory
Experiment 3 N-back recognition memorytask
The previous experiment failed to find any impair-ment in GR and COrsquos ability to form a representa-
EPSTEIN ET AL
496 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
tion of the geometry of a scene or object andmaintain it for 10 s in the absence of the originalstimulus Not only does this result suggest that per-ception of scene-like stimuli is intact in thesepatients it further suggests that some short-termmemory functions are intact as well However we
should note that subjects in this experiment werefree to devote all their effort and attention in the 10-second blank interval to maintaining a memory ofthe stimulus Thus it is possible that GR and COcan use rehearsal mechanisms to maintain the stim-ulus in short-term memory during the blank inter-
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 497
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
(b) Lego object version
Figure 7 Stimuli and procedure for Experiment 2 (a) In the Lego layout version subjects see a Lego layout for 3 s followed by a blankscreen for 10 s A second layout then appears which is either the same layout as the first one but seen from a different angle or a differentlayout (b) The Lego object version is the same but with object stimuli
3 s 10 s OR
3 s 10 s OR
(a) Lego layout version
(b) Lego object version
val but are unable to form a more durable memorytrace for the stimulus This possibility was tested inthe current experiment which was designed toexamine GR and COrsquos ability to learn new visualinformation
We devised a continuous n-back recognitionmemory test in which subjects viewed a sequence oforiginally novel stimuli some of which wererepeated within the sequence at intervals of 1 3 or5 intervening images For each stimulus subjects
reported whether they had seen it before in theexperiment or not In different runs in the experi-ment the stimuli were black-and-white photo-graphs of (1) Lego scenes (2) Lego objects (3) real-world landscapes (4) novel objects made out ofSculpey clay (5) unfamiliar houses (6) unfamiliarfaces as well as (7) line drawings of nonsenseobjects and (8) pronounceable pseudowords (Fig-ure 9) Thus we were able to test GR and COrsquos rec-ognition memory for different kinds of materials atdifferent time intervals in a paradigm in which thepresence of intervening items between the twoappearances of each stimulus made rehearsal diffi-cult Note that because none of the specific stimuliused in this experiment were familiar in advancethis task did not require an episodic memory com-ponent and could be done on the basis of pure visualfamiliarity with the particular visual stimulus
Although many different stimulus classes weretested in this experiment the critical comparison isbetween the Lego layouts and Lego objectsbecause these are best matched with regard tomaterials and differ primarily in their geometricstructure (see Introduction) We predicted that ifGR and CO showed a memory deficit for this taskit would be greater for the Lego layouts than for the
EPSTEIN ET AL
498 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 8 Results of Experiment 2 Performance over ldquosamerdquotrials and ldquodifferentrdquo trials is averaged together Error bars show1 SD
Figure 9 Stimuli for Experiment 3 Top row Lego layouts real-world scenes houses Bottom row Lego objects clay objects nonsenseobjects faces In an eighth condition (not shown) subjects viewed pronounceable nonwords such as ldquowilchrdquo or ldquothipperrdquo
Lego objects In addition a deficit for materialsthat convey information about the layout of sur-rounding space might also be apparent when GRand CO are tested on more realistic stimuli If soone would expect them to do poorly on the land-scapes but relatively well on the clay objects linedrawings of nonsense objects faces andpseudowords
MethodFor each subject a complete experimental sessionconsisted of being run once on each of the eightstimulus classes Each run consisted of 90 trials ineach trial a stimulus was presented and subjectsreported whether or not they had seen it before inthe run (or indeed ever) by pressing the ldquojrdquo key ifthey had or the ldquof rdquo key if they had not Each stimu-lus was on the screen for either 1 2 or 3 s (see fol-lowing) and appeared promptly after the keypressreport for the previous trial Individual stimuliappeared either once (14 stimuli) twice (30 stim-uli) or four times (4 stimuli) within a given run Ofthe stimuli that were presented twice 10 wererepeated immediately (1-back) 10 were repeatedwith a lag of two intervening items (3-back) and 10were repeated with a lag of four intervening items(5-back) In order to ensure that between-subjectdifferences in performance were not due to stimulusdifferences the same stimuli were assigned to thesame conditions each time the experimentwas run
GR performed the experiment twice once inAugust 1998 and again 9 months later Because ofa computer error he was only tested on six of theeight categories (all except Lego layouts and Legoobjects) at the later date CO was also tested twiceonce in February 1999 and again 6 months laterReported results for GR and CO are the average oftheir two testing sessions Normal subjects wereonly tested once Stimulus presentation time was 1 sin GRrsquos first testing session In pilot testing COfound this too fast for the critical categories of Lego
layouts and Lego objects so presentation time wasextended to 2 s for these categories in his first test-ing session All normal subjects were tested at thesepresentation rates (2 s for Lego layouts and objects1 s for other stimuli) In COrsquos second testingsession presentation time was lengthened to 3 s forall stimulus categories to ensure that any deficitsfound were memory deficits rather than problemswith the nonmemory aspects of the task
Results and discussionResults from GR CO and 10 elderly normal con-trols are shown in Figure 10 and Table 4 Figure 10ashows results from the main comparison betweenLego layouts and Lego objects Both normal sub-jects and patients performed near ceiling for bothLego layouts and Lego objects at 1-backinterstimulus intervals verifying that GR and COcould successfully perceive the pictures as they werebeing presented In contrast there was a strikingdifference between the two patients and the normalsubjects at 5-back intervals Whereas normals didon average somewhat better on Lego layouts thanLego objects both GR and CO showed the oppo-site pattern doing much worse on Lego layoutsthan Lego objects The 3-back condition showed amixed pattern Whereas GR did better on Legoobjects than Lego layouts in this condition (similarto his performance at 5-back intervals) CO didbetter on Lego layouts than Lego objects COrsquos lowperformance on Lego objects in the 3-back condi-tion appears to be an anomaly as it is much lowerthan his performance for comparable stimuli in the1-back and 5-back conditions
Figure 10b shows the difference in performancebetween Lego layouts and Lego objects for thecritical 1-back and 5-back intervals At 5-backintervals CO was significantly impaired comparedto the normal subjects (z = ndash250) whereasGRrsquos impairment fell just short of significance (z =ndash175)4
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 499
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
4In fact these values may underestimate GR and COrsquos relative impairment for Lego layouts In the 5-back condition 4 of the 10
normal subjects had perfect performance on the Lego layouts but all scored below ceiling on the Lego objects Thus the true advantagefor Lego layouts over Lego objects for normal subjects is probably greater than that observed here in which case GR and COrsquos lowerperformance for Lego layouts compared to Lego objects would be even more abnormal Furthermore post-hoc analyses revealed thatone of the normal subjects showed a difference in performance between Lego layouts and objects that was over 2 SD below the mean ofthe other nine subjects When this outlying subject was excluded z-scores were ndash322 for CO and ndash232 for GR
Table 4 shows results for all eight stimulusclasses Although direct comparisons between sub-jects should be treated with caution since viewingtime for different items was not the same for eachsubject we do note a few interesting patterns Firstof all there is little or no evidence that GR or CO
show a memory deficit for nonsense words clayobjects or line drawings of nonsense objects GRappears to be slightly impaired on faces and housesbut CO performs normally on these categoriesStrikingly GR shows a severe deficit for the reallandscapes performing more than 40 percentagepoints lower than normals Thus even thoughthere is some evidence that GR might have someimpairment for nonscene stimuli he clearly is muchmore impaired for scene-like stimuli such as theLego scenes and landscapes CO does not show asimilar deficit for the landscapes However itshould be kept in mind that in one of his two testingsessions CO viewed each picture for 3 s which wasmuch longer than the 1 second used for all non-Lego stimuli for GR and normals Thus COrsquosapparently ldquonormalrdquo performance in these condi-tions might be overestimated
Taken together the results of this experimentindicate that GR and CO are selectively impaired atprocessing scene-like stimuli This deficit is consis-tently apparent at 5-back but not 1-back intervalsThis finding suggests that the deficit should becharacterised as a memory encoding problemHowever the present results do not allow us todetermine whether the difficulty occurs at memoryencoding or at subsequent recognition (see Epsteinamp Kanwisher 2001) If GR and CO have a generalproblem with recognition of topographical materi-als then they should not be able to recognise scenes
EPSTEIN ET AL
500 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 10a Results of Experiment 3 for GR (top) CO (middle)and for 10 age-matched normal controls Performance measure isproportion of repeated stimuli correctly identified as such correctedfor guessing using the standard formula
Figure 10b Difference between Lego layout performance andLego object performance for the 1-back and 5-back conditionsError bars indicate one 1 SD
and landmarks that were learned prior to theirinjury We tested this possibility in the nextexperiment
Experiment 4 Famous landmarksfacesrecognition task
In this experiment we examined GR and COrsquosability to recognise the visual appearances of placeslearned prior to injury Subjects saw a series offamous landmarks intermixed with nonfamouscontrols (Imaging experiments have demonstratedthat landmarks of this type can activate the PPAalmost as strongly as complete scenes Epstein etal 1999 Epstein amp Kanwisher 1998) They wererequired to say whether they knew each specificlandmark and to identify the ones they knew Sub-jects were instructed to respond affirmatively if theywere fairly certain that they recognised the land-mark but not if it only looked generally familiarFor purposes of comparison subjects also per-formed an identical test in which the stimuli werefamous and nonfamous faces In both testsdistractor items were designed to be visually andconceptually similar to the target items (see Figure11) If parahippocampal cortex is required for placerecognition then one might expect GR and CO toperform poorly compared to normals on the land-mark recognition test
MethodEach of the two experiments (famous faces famouslandmarks) consisted of 48 trials Subjects beganeach trial by pressing the space bar after which ablack-and-white photograph of a face (in thefamous faces experiment) or a landmark (in thefamous landmarks experiment) appeared on thescreen Subjects were told to press the ldquojrdquo key if theyrecognised the specific face or landmark and the ldquof rdquokey if they did not taking as much time at theyneeded to make this discrimination but no extratime Subjects were told to report that they recog-nised a face or landmark if they felt that they knewit even if they could not retrieve its name If theydid know the name they were to say it after pressingthe button Otherwise they were to say whateverthey did remember about the item
Half of the photographs were of famous and halfof nonfamous people and landmarks The famousfaces included public figures (eg George BushRichard Nixon Prince Charles) and movie stars(eg Harrison Ford Clark Gable Marlon Brando)who were prominent prior to the time of GR andCOrsquos injuries the famous landmarks were build-ings and monuments from all over the world (egEiffel Tower US Capitol Building Great Sphinx)The nonfamous stimuli were chosen to match thefamous stimuli as closely as possible Each famousstimulus had a ldquomatchrdquo For example George Bushwas matched to a man of similar age in a coat andtie and the Taj Mahal was matched to a visually
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 501
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 4 Full results of Experiment 3 Mean proportion correct (corrected for guessing) SDs in parentheses
Lego Land- Lego Clay Nonsense Non-scenes scapes Houses objects objects objects Words Faces
1-back GR 100 094 100 100 100 094 100 100CO 091 085 091 100 100 079 074 100Normals 096(008) 095(008) 093(014) 096(008) 096(006) 099(003) 098(004) 097(011)
3-back GR ndash003 068 026 047 083 083 075 052CO 066 081 037 010 094 069 095 081Normals 066(032) 077(021) 072(015) 077(020) 086(016) 083(015) 083(017) 072(020)
5-back GR 010 034 044 047 072 083 094 053CO 031 073 075 088 081 064 100 081Normals 076(026) 077(016) 068(021) 066(016) 091(014) 083(018) 093(011) 078(021)
similar Asian palace that is not famous (see Figure11)
Results and discussionResults for GR and CO as well as for five age-matched controls are shown in Figure 12 Both
GR and CO could recognise the famous landmarksas well normals In addition both GR and CO wereusually able to name the landmarks they success-fully identified
Thus GR and CO can recognize landmarkslearned prior to injury demonstrating that at leastsome of their place recognition abilities are intactWhen combined with the fact that GR and COwere able to produce detailed maps of places theylived in prior to their injury (see Case Description)these results suggest that their deficit is primarily inencoding or consolidation of new informationrather than in recognition or recall of old informa-tion Note however that these results do not provethat their place recognition abilities are entirelynormal GR and CO might suffer from a mild rec-ognition deficit that affects identification of newly-learned stimuli like the Lego layouts in Experiment3 but not overlearned stimuli like the famous land-marks Furthermore it is possible that GR and COmight be impaired at recognising premorbidlylearned places that are defined primarily by theirspatial layout even though they are unimpaired atrecognising premorbidly learned places that aredefined primarily by a single object-like landmarkor building (see General Discussion) Neverthelessthe trend of the present evidence is to suggest thatGR and CO have particular problems with noveltopographical information
In the famous-face task GRrsquos performance wasin the normal range However he sometimes haddifficulty naming the faces even though he clearly
EPSTEIN ET AL
502 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 11 Stimuli for Experiment 4 Each famous landmark(Arc de Triomphe Taj Mahal) and famous face (Barbara BushYasser Arafat) was matched to a nonfamous building or face thatalso appeared in the experiment
FA
CE
SL
AN
DM
AR
KS
Figure 12 Results of Experiment 4 Performance measure isproportion of famous stimuli reported as ldquoknownrdquo corrected forguessing using the standard formula
recognised them For example he described Ron-ald Reagan as ldquothe gipperrdquo Nancy Reagan as ldquothewife of the gipperrdquo and Clark Gable as ldquothe movieactor who said lsquoScarlett I donrsquot give a damnrsquordquo COhad a great deal of difficulty identifying the famousfaces This result is not surprising given hisextremely low performance on the Benton FaceMatching Task In addition to his topographicproblems CO appears to suffer fromprosopagnosia
GENERAL DISCUSSION
We tested two parahippocampal patients (GR andCO) on four experiments designed to measure per-formance as a function of both task (perceptionencoding and recognition) and stimulus type(scenes objects faces and words) The only taskthat revealed a clear deficit was an n-back recogni-tion memory test (Experiment 3) In order to suc-cessfully perform this task subjects had to encode adurable representation of a visual stimulus and useit for later recognition GR and CO were signifi-cantly more impaired on this task when the stimuliwere scene-like spatial layouts (ie Lego scenesand real scenes for GR) than when they wereobjects No comparable deficits were found in otherexperiments that tested layout perception (Experi-ments 1 and 2) and placelandmark recognition(Experiment 4) Taken together these results indi-cate that GR and CO are impaired in their ability toencode novel information about the geometry ofsurrounding space into memory but are lessimpaired at their ability to encode the geometry ofnovel objects
We argue that this specific topographical defi-citmdashinability to encode the spatial layout of novelscenesmdashresults from damage to the para-hippocampal place area Although the damage inboth patients extends beyond the parahippo-campal-lingual territory normally associated withthe PPA this region is clearly lesioned bilaterally inGR and in the right hemisphere in CO Further-more functional imaging demonstrated that GRhas no functioning PPA Thus the current resultsand the earlier fMRI work (Epstein et al 1999
Epstein amp Kanwisher 1998) converge to implicateparahippocampal cortex in the encoding of infor-mation about scene geometry into memory
Our assignment of the function of memoryencoding to parahippocampal cortex is consistentwith the results of earlier case reports of patientswith parahippocampal damage who have naviga-tional difficulties in novel but not familiar environ-ments (Habib amp Sirigu 1987 Ross 1980 see alsoTeng amp Squire 1999) As with GR and CO thesepatients were either less impaired or unimpaired onnontopographical memory tasks Patients have alsobeen described who exhibit the opposite pattern ofimpairment an inability to learn faces (sometimescalled ldquoprosopamnesiardquo Cipolotti Robinson Blairamp Frith 1999 Tippett Miller amp Farah 2000) orfaces and verbal materials (Maguire amp Cipolotti1998) with an unimpaired ability to learn topo-graphical materials Taken together these resultsargue for a high degree of domain-specificity in theneural systems involved in learning of new informa-tion (Gallistel 1995) The brain may have separatemechanisms for learning faces places and wordswhich can be differentially impaired after damageto specific cortical regions (Cipolotti et al 1999Incisa della Rochetta et al 1996) The presentresults suggest that the anatomical locus of themechanism dedicated to learning new topographi-cal information is in the parahippocampal cortex(Aguirre amp DrsquoEsposito 1999)
In contrast to their impaired ability to encodenew topographical information both GR and COwere able to produce detailed maps of their previousdwellings and performed normally on the famouslandmark recognition test of Experiment 4 Theseresults are generally consistent with Teng andSquirersquos (1999) report of a patient with extensivemedial temporal damage who could recallpremorbidly learned but not postmorbidly learnedtopographical information (see also Ross 1980)Thus our results provide further evidence thatmedial temporal regions such as parahippocampalcortex may be more critical for encoding or consoli-dating new information into memory than for recallof old information
One issue that our experiments do not entirelyresolve is the precise nature of the topographical
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 503
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
information encoded by parahippocampal cortexClearly this region is critical for encoding informa-tion about the geometry of the currently visiblescene even when the ldquoscenerdquo is made out of Legoblocks and therefore has no immediate navigationalrelevance However it is unclear whether this spe-cific deficit can account for all of the real-worldnavigational difficulties encountered by GR andCO GR in particular shows such a catastrophicloss of navigational ability that we suspect he is alsoimpaired at encoding the spatial relationshipsbetweenthe current scene and other locations in theworld CO probably also suffers from such difficul-ties as he reports that he has trouble rememberinghow places relate to each other in large-scale spaceWhen he was tested on a simple 3-room videogameenvironment he not only was unable to rememberthe individual rooms (claiming that there were 10rooms when there were only 3) but he also had nomemory for the spatial relationships between themThese spatial deficits may be the result of damage tothe posterior right hippocampus (in the case of CO)or of disconnection of the posterior right hippo-campus from the parahippocampal regions thatprovide much of its cortical input (in both patientsSuzuki amp Amaral 1994) In either case the abilityto form a cognitive map of new environments likethe lab or videogame space could be impaired(Maguire et al 1998 OrsquoKeefe amp Nadel 1978)However it is also possible that parahippocampalcortex itself may be critical for encoding not onlythe geometry of individual scenes but also the spa-tial relationships between the locations of thosescenes In this case the spatial deficits of GR andCO could be attributed directly to parahippo-campal damage rather than to hippocampal dam-age or disconnection
The latter scenario would be consistent withAguirre and DrsquoEspositorsquos (1999) and Barrash etalrsquos (2000) suggestion that parahippocampal cortexis critically involved in combining visual informa-tion about the appearance of scenes and landmarkswith spatial information about their locations into aunified topographical representation Howeverboth of these authors argue that the representationsof individual scenes (Barrash et al) and landmarks(Aguirre amp DrsquoEsposito) are supported not by
parahippocampal cortex but by more posterior lin-gual-fusiform regions In contrast we believe thatthe neuroimaging data indicate that parahippo-campal cortex is involved in the encoding of thegeometric structure of individual scenes althoughit may also play a role in combining these represen-tations with other spatial representations
Interestingly GR and CO were not impaired onrecognition of famous landmarks that were mostlybuildings Thus our data do not preclude the possi-bility that there may be a separate lingual orfusiform region involved in processing informationabout buildings (Aguirre Zarahn amp DrsquoEsposito1998 Chao Haxby amp Martin 1999 IshaiUngerleider Martin Schouten amp Haxby 1999)which might have been preserved in GR and COdespite extensive damage to other regions of thelingual and fusiform gyrus Alternatively GR andCOrsquos preserved building recognition abilities mightbe mediated by interactions between cortical areasthat perform shape analysis for any object-likestimulus type (Malach et al 1995Kanwisher et al1996) and anterior temporal regions in which long-term topographical and semantic memories arestored (McCarthy et al 1996)
Is perception preserved
In contrast to their inability to encode new scenesinto memory GR and CO have substantiallyretained their ability to perceive the spatial layout ofthe currently visible scene (Experiments 1 and 2)GR can follow a route marked on a floor planwhich indicates a good on-line appreciation of hiscurrent spatial surroundings Furthermore neitherGR or CO have any trouble moving through theenvironmentmdashthey move with confidence and donot bump into doors or walls Problems of the lattertype are usually associated with damage to the pari-etal lobes which are intact in GR and CO Thus atfirst glance GR and CO do not appear to haveperceptual impairments (beyond their visual fieldrestrictions)
However GR does complain of a perceptualdeficit which he describes as an inability to ldquotakeinrdquo scenes when they are too complex Similarreports of an inability to comprehend the
EPSTEIN ET AL
504 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
ldquoglobalityrdquo of a scene were made by some of theparahippocampally-damaged patients studied byHabib and Sirigu (1987) We were not able todetermine the precise nature of this deficit whichmay be partly attributable to his visual field restric-tions It appears that his deficit is more salient in thereal-world environments than in the picture-per-ception tasks of Experiments 1 and 2 perhapsbecause he finds it more difficult to organise visualinput when immersed in it than when viewing pho-tographs that subtend only part of the visual field
We suggest that GR can perceive parts of thescene when he attends to them but may be unableto integrate information over time into a coherentrepresentation of a scene or place The fact that GRand CO are impaired for Lego layouts at 5-backintervals in Experiment 3 is consistent with thishypothesis The time between the first and secondpresentations of a stimulus is very short in this case(and even shorter with 3-back intervals where GRalso shows evidence of a deficit) In both patientsthe representation of layout information formed byperception of a Lego scene must either have anabnormally short persistence be abnormally sus-ceptible to interference or both If so they mightfind it impossible to sustain a spatial frame orschema long enough to use it to organise informa-tion obtained from many different glances at a com-plex scene (Intraub 1997) The result would be aninability to understand the ldquoglobalityrdquo of a sceneeven though all the details were perfectlyperceived
Non-topographical mnemonic abilities
Although not as severe as their topographical mem-ory problems GR and CO do have some memorydifficulties in other domains For example bothperformed poorly on the Rey AVLT whichrequired them to remember a list of unrelatedwords Both also report some subtle episodic mem-ory problems (see Case Description) COrsquos prob-lems could partly result from damage to the righthippocampusmdashindeed CO appeared to have themore acute episodic memory problems HoweverGR has no hippocampal damage so the fact that hedisplays episodic memory problems suggests thatparahippocampal cortex is involved in the forma-
tion of episodic memories (cf Wagner et al1998)
In previous work (Epstein et al 1999) weargued that spatial context information encoded byparahippocampal cortex might play a role in estab-lishing event memories by encoding informationabout where events take place The inability toencode this information might cause problems intasks such as the Rey AVLT where one has to recalla list of unrelated words If spatial (and perhapstemporal) context information is not encoded itwill be harder subsequently to recall the distinctevent of hearing each word (Tulving 1983) Incontrast lack of spatial context information wouldbe less of a problem in tasks where one is onlyrequired to recognise previously encountered verbalmaterials but not to reproduce the spatiotemporalcontext in which they were experienced IndeedGR and CO performed as well as normals on thenonsense words in Experiment 4 where recogni-tion memory but not episodic memory was requiredfor successful performance Interestingly GR andCO also did relatively well on the Logical Memorysubtest of the WMS-III perhaps because the nar-rative framework of a story provides sufficient con-textual information to overcome the absence ofspatial context and mediate successful verbalretrieval
Differences between GR and CO
Although we have discussed GR and CO togetherthere are a number of relevant differences betweenthem One open question is whether these differ-ences can be attributed to the different effects ofunilateral and bilateral parahippocampal damageIn general GRrsquos topographical difficulties weremore severe than COrsquos CO did surprisingly well onthe Warrington Topographical Memory Test inwhich he was required to remember detailed real-world city scenes and also performed substantiallybetter on real scenes than Lego scenes in Experi-ment 3 Insofar as CO has intact left para-hippocampal cortex this suggests that the left PPAmight be sufficient to encode a certain amount of(perhaps semantic) information about real-worldscenes but the right PPA is necessary to encode
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 505
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)

EPSTEIN ET AL
488 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure
1F
loorplansdraw
nby
GR
andhisw
ifeof(a)
theirhotelroomin
Boston(b)theircurrenthousew
hichthey
hadm
ovedto
subsequenttoG
Rrsquosinjury(c)the
housethey
livedin
priorto
GR
rsquosinjury
Ho
telro
om
inB
osto
n
GR
His
wife
His
wife
Cu
rren
th
om
e
GR
His
wife
Fo
rme
rh
om
e
GR
about the main room of the lab once he had left itCO was also able to draw a detailed floor plan of theapartment he was living in at the time (and hadlived in for 20 years) Several months after he hadmoved from this apartment we asked him to drawsuch a floor plan again and he produced one thatwas substantially the same as the first (Figure 3)Thus their map drawing performance indicatesthat GR and CO are primarily impaired at sketch-ing places experienced after their strokes but areunimpaired at sketching places experienced beforetheir strokes This pattern is indicative of deficits inencoding but not recall
Scene perceptionAs noted earlier GR can navigate with the aid of afloor plan suggesting that his perceptual abilitiesare largely intact Interestingly however he reportsthat he does have a perceptual deficit which hedescribes as an inability to understand scenes whenthey are too complex For example he reports thathe often has trouble using maps because theyusually are too busy with details It is unclearwhether this apparent perceptual problem is a resultof his field cut some other separate perceptual
problem or a memory problem (see discussion)CO did not complain of a similar deficit GR wasan accomplished artist prior to his injury producingpaintings of striking complexity Tragically hereports that he cannot now appreciate his ownpaintings
Other cognitive abilities
Face recognitionGR also reported some difficulties with face recog-nition On his second visit we examined his abilityto learn new faces by having him perform a forced-choice recognition test on seven familiar and sevenunfamiliar faces The familiar faces were personnelfrom the lab with whom he had at that point had agood deal of experience spanning at least 3 daysAll the people in the photographs had a towelwrapped around their head in order to minimise theuse of hair as a nonfacial cue Despite the difficultyof this task he correctly identified 67 familiarfaces and made no false alarms Surprisingly theone face that he did miss was the first author of thispaper with whom he at that point had had consid-erable experience GR also performed normally ona difficult famous face recognition test (see Experi-ment 4 following) and on the Benton Face Recog-nition test (see following) Thus he has someability to recognise faces Consistent with this GRreports that he can remember a small number offaces but that he quickly ldquoloses trackrdquo if he has toremember too many
CO also appears to suffer from someprosopagnosia as evidenced by his low perfor-mance on the Benton Face Recognition test (seefollowing see also Experiment 4) Interestingly hedoes not complain of face recognition difficultiesbut rather attributes his inability to recognise peo-ple to a lack of interest in their appearance
Episodic memoryAt first glance GRrsquos episodic memory appeared tobe largely intact As mentioned he could recall theevents of the previous day accurately Furthermorehe reports that he can follow the plot of a moviewithout any difficulty However both he and hiswife report that he does have some problems with
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 489
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Figure 2 Floor plan of the building at MIT in which GR wastested Corridors are highlighted in grey GR was asked to find themain room of the lab (marked G on the map) Starting location(marked S) was a point approximately 30 feet down the hall GRrsquosroute is marked by arrows GR found this task very difficult andonly knew for certain that he had reached his goal when he caughtsight of his wife through the glass door of the lab In contrast hewas able to follow a route marked on a plan of the same floorwithout any problem or hesitation
episodic memory For example he stated that heremembered very little about a trip to Italy they hadtaken several months previously His wife reportsthat he has great difficulty keeping track of timeFor example before their trip to Boston he repeat-edly asked her what day they were supposed to leaveand seemed unable to encode this information Weobserved a similar phenomenon during testingDuring the tests that required repetition of thesame procedure several times he sometimesseemed unable to clearly remember how manytimes he had repeated the procedure Thus he doesappear to have a subtle episodic memory deficit
which may manifest itself as an inability to sepa-rately remember successive events that are not easilydistinguishable from each other
CO also reports some problems with his epi-sodic memory He relies on lists to rememberappointments stating ldquoif I didnrsquot have a list Irsquodprobably forget why I walked in the door hererdquoDespite this claim he has never missed an appoint-ment nor is he ever confused about the purpose ofthe testing appointments He can accurately reportthe day of the week He remembers the experi-menters clearly from appointment to appointmentas well as the details of the various experiments
EPSTEIN ET AL
490 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 3 Two floor plans drawn by CO of the apartment he lived in for over 20 years The first plan was drawn when CO still lived inthe apartment the second was drawn several months later when he had moved out Note that the two plans correspond very well COreported that it was quite easy to produce these floor plans
CO home Feb 99
CO (former) home June 99
when they are repeated across appointments Whenqueried about his episodic memory difficulties COreported that he will usually remember an event ifhe is reminded of it but that he has difficulty spon-taneously regenerating events Interestingly hereports that even when he is reminded of an eventand remembers it he cannot remember ldquowhere itwas or when it wasrdquo When he reads he finds it dif-ficult to follow a story if it skips around Howeverwhen queried he was able to accurately report whathe had read in the newspaper the previous day
Formal neuropsychological tests
Formal neuropsychological testing was performedon GR in September 1998and on CO in December1998 Test performances were compared to age-matched peers3
GR exhibited mild to moderate impairment inmemory relative to what would be predicted for hisintellect (WAIS-R Full Scale IQ =122 93rd per-centile superior range) and level of educationThese memory problems were more apparent onvisual than on verbal memory tests On a test ofmemory for conceptually organised text passages(Wechsler Memory Scale Logical Memory[WMS-III] Wechsler 1997b) his performancewas average for both immediate (LM I =25th per-centile standard score SS = 90) and delayed recall(LM II =50th percentile SS =100) In contrast hisscore on the WMS-III Face Memory subtest was inthe low average range for both immediate (9th per-centile SS =80) and delayed recall (16th percentileSS =85) On a test of memory for scenes (WMS-IIIFamily Pictures subtest) immediate recall was defi-cient (2nd percentile SS = 69) and delayed recallwas borderline impaired (5th percentile SS =76)GR also had difficulty learning word lists on theRey Auditory Verbal Learning Test (Spreen ampStrauss 1998) his immediate recall was average(63rd percentile SS = 105) but after the interfer-ence list his performance fell to the low averagerange for both immediate (14th percentile SS =84)
and delayed recall (16th percentile SS =85) On arecognition test of the items of the list he success-fully recognised all 15 but made 8 false alarms Per-formance on the 724 Spatial Recall Test (RaoHammeke McQuillen Khatri amp Lloyd 1984)which required him to remember seven locations ona 6times4 checkerboard was 66th percentile (SS =106)for immediate and 2nd percentile (SS = 69) fordelayed recall Performance on the Benton FaceRecognition test was normal (42nd percentile)Performance on the Boston Naming Test was atceiling indicating that language abilities wereintact Thus GR exhibits some impairment ontasks that require delayed recall of visual informa-tion (WMS-III Face Memory subtest) or verbalinformation that is not organised in terms of a storyor plot (Rey AVLT) and is clearly impaired ontasks that required delayed recall of spatial informa-tion (724 Spatial Recall Test) or stimulus location(WMS-III Family Pictures subtest) In contrasthis ability to recall organised verbal information(WMS-III Logical Memory subtest) appears to beintact
As with GR COrsquos memory performance wasgenerally below what would be expected based onintellect (WAIS-III Full Scale IQ =134 99th per-centile very superior range) and education levelAlso like GR there was some evidence that visualmemory problems were more pronounced than ver-bal memory problems Memory for text passages(WMS-R Logical Memory) was average for bothimmediate (27th percentile SS = 91) and delayedrecall (48th percentileSS =99) In contrast perfor-mance on the WMS-III Faces subtest was lowaverage for both immediate (16th percentile SS =85) and delayed recall (9th percentile SS =80) andperformance on the Family Pictures subtest wasdeficient (lt 1st percentile SSlt 62) for both imme-diate and delayed recall On the Warrington Rec-ognition Memory Tests (RMT) he scored in theaverage range for words (50th percentile SS =100)and scenes (Warrington Topographical Recogni-tion Memory Test 29th percentile SS =92) and in
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 491
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
3The formal neuropsychological testing reported in this section was performed prior to the beginning of our combined
investigation of GR and CO (ie prior to the main experimental investigations of this paper) Consequently GR and CO were nottested on exactly the same battery of tests
the deficient range (lt 5th percentile SS lt 75) forfaces The low score for faces on this test as well asthe WMS-III may reflect prosopagnosia ratherthan a memory problem as he scored in the defi-cient range (lt1st percentile SSlt62) on the BentonFace Recognition test Verbal list learning (ReyAVLT) was average on immediate recall (45th per-centile SS =98) but after hearing an interferencelist performance fell to the deficient level (015words recalled) for both immediate and delayedrecall On a recognition test for these words herecognised 1115 and made one false alarm (39thpercentile SS =96) There was a large discrepancyon tests of memory span which may indicate adecline in spatial memory digit span was superior(9+ forward and 7 backwards both 99th percentile)but spatial span was only average (5 forward and 5backward 37th and 50th percentiles respectively)Language abilities were intact (Boston NamingTest performance was at ceiling) Thus like GRCO shows some impairment at recall of visualinformation (WMS-III Face Memory subtest) andverbal information that is not organised into a nar-rative (Rey AVLT) and clear impairment at tasksthat require recall of stimulus location (Family Pic-tures subtests of WMS-III) In contrast his abilityto recall organised verbal information (WMS-RLogical Memory subtest) is more preserved
MRI RESULTS
In order to determine the extent of their lesionsstructural MRI images were taken of both subjectsDelineation of the lesions was based either onspoiled GRASS coronal images at 11mm thick-ness (GR) or anatomical T-1 weighted MPRAGEMRI sequences yielding 125 mm thick coronalimages (CO) The anterior-posterior extent of thelesions was verified on axial images
GR has bilateral occipital-temporal lesions(Figure 4) In the left hemisphere the posteriorparahippocampal gyrus (including para-hippocampal cortex) is severely damaged with onlyminimal injury to the most caudal aspect of the hip-pocampus There is also moderate damage to themedial fusiform and inferior lingual gyri In the
right hemisphere the hippocampus is spared butthere is damage to the posterior end of theparahippocampal gyrus near the parahippocampal-lingual boundary More posteriorly the lesionedarea extends along the collateral sulcus to includesignificant portions of the inferior lingual gyrus andthe medial fusiform gyrus as well as most of themedial occipital lobe below the calcarine fissure
CO has a right-hemisphere occipital-temporallesion Right posterior hippocampus and para-hippocampal gyrus are severely affected Moreposteriorly the damage covers large portions of theinferior lingual and medial fusiform gyri as well assubstantial portions of the medial occipital lobeincluding most of right calcarine cortex In addi-tion diffuse foci of increased signal on T-2weighted images was present in periventricularwhite matter in the corona radiata and centrumsemiovale with a lacunar infarct of the rightputamen There was also moderate degree of dif-fuse involutional changes with increased ventricularsize particularly notable in the right temporal hornwith ex vacuo dilitation
The structural MRIs suggested that the PPAwas severely if not completely damaged in bothhemispheres for GR and in the right hemispherefor CO In order to confirm this for GR we used ahigh-field 3-T magnet to perform a functional scanin which he viewed scenes faces and objects in thesame blocked fMRI design that we have used tolocalise the PPA in all our previous experiments(see Epstein et al 1999 for details Epstein ampKanwisher 1998) A Kolomogorov-Smirnov testfound no voxels in the parahippocampal region thatfit our previously established criterion for inclusionin the PPA (ie greater response to scenes than tofaces and objects at a uncorrected significance levelof plt 0001) This result contrasts sharply with thepattern seen in over 95 of (mostly young or mid-dle-aged) normal subjects in whom a large acti-vated region can be identified in both left and rightparahippocampal cortex using this comparison (seeFigure 5) A similar test was used to identify voxelsthat responded significantly more to faces than toobjects This comparison revealed a region of face-selective voxels in the right fusiform gyrus consis-tent with the location of the Fusiform Face Area
EPSTEIN ET AL
492 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 493
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
GR
CO
Figure 4 Structural MRIs of GR and CO showing the extent of the damaged region (marked as white) Images have been normalisedusing SPM99 and a cost function masking algorithm (provided by Matthew Brett) into standard stereotactic space (Talairach amp Tournoux1988) using the Montreal Neurological Institute template (Evans et al 1993) Numbers indicate y-Talairach coordinate for each coronalslice corresponding slice planes are also indicated on the sagittal image
(FFA) commonly seen in normal subjects(Kanwisher McDermott amp Chun 1997) Thusthe MRI was sensitive enough to detect functionaldifferences an FFA was found but no PPA Whencombined with the anatomical results this stronglysuggests that GR has no PPA
Because of safety concerns (presence of a femo-ral artery aneurysm clip) we did not perform a simi-lar high-field functional MRI of CO However asFigure 4 demonstrates CO clearly has damage tothe parahippocampal-lingual region in the righthemisphere
EXPERIMENTAL INVESTIGATIONS
In this section we report the results of four experi-ments designed to probe the consequences of dam-age to the lingual-parahippocampal region ingeneral and the PPA in particular Our primaryconcern in these experiments was to test GR andCOrsquos ability to perceive remember and recognisedifferent kinds of visual material Based on our pre-vious fMRI work we predicted that GR and CO
would have more difficulty with scene-like stimuli(ie stimuli that convey information about thegeometry of a space one can navigate through or actwithin) than with object-like stimuli Such a deficitcould account for at least some of the problems GRand CO encounter in real-world navigation
Beyond determining the kind of visual materialthat gets processed in parahippocampal cortex wealso wanted to determine the nature of the process-ing that takes place on that material In particularwe wanted to determine if parahippocampal cortexis involved in perceptual encoding memory encodingor recognition As discussed earlier (Introduction)our fMRI results provided some evidence againstplace recognition as the sole function of the PPAbut did not allow us to distinguish between percep-tual and mnemonic encoding Experiments 1 and 2examine the role of parahippocampal cortex in per-ceptual encoding Experiment 3 examines its role inmnemonic encoding and Experiment 4 examinesits role in place recognition
Experiment 1 Two-dot task
The first hypothesis we examined was thatparahippocampal cortex plays a role in perceivingthe shape of the local environment In our previousfMRI work we observed that the PPA respondsmuch more strongly during scene viewing than dur-ing object or face viewing even when subjects donothing more than watch the scenes passively with-out performing any other task This suggested thatscene perception alone may be sufficient to activatethe PPA Previous reports have indicated that dam-age to parahippocampal cortex and other medialtemporal regions leads to problems with memoryrather than perception (Bohbot et al 1998Maguire Burke Philips amp Staunton 1996 Pigottamp Milner 1993) However all of these previousresults were obtained on patients who had unilat-eral lesions GR has a bilateral occipital-temporallesion including clear damage to parahippocampalcortex in both hemispheres furthermore he reportssome kind of perceptual deficit We hypothesisedthat this deficit might involve difficulty perceivingor interpreting the geometry of scenes and otherstimuli with a similar geometric structure
EPSTEIN ET AL
494 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 5 Top row shows a single slice from a functional MRIscan performed on GR Bottom row shows results from a normalsubject (NK) for comparison Voxels that responded more to facesthan objects (left column) or more to scenes than faces and objects(right column) in a Kolomogorov-Smirnov test are highlighted bywhite arrows GR has a functioning face area (FFA) but there isno evidence of a place area (PPA)
FFA PPA
GR
NK
In this experiment subjects viewed either objectshapes or spatial layouts Stimuli consisted ofblack-and-white photographs of real-world stimuli(empty rooms vs common objects) or abstractstimuli made out of Lego blocks (Lego ldquoscenesrdquo vsLego ldquoobjectsrdquo) These stimuli have been previouslyshown to differentially activate the PPA in normalsubjects (Epstein et al 1999 Epstein ampKanwisher 1998) Two red dots were superim-posed on each photograph (Figure 6) The task wasto report which of the two red dots was over thecloser part of the object or scene This task wasdesigned to have no memory component (apartfrom remembering the instructions) but to requireaccurate 3D perception of objects and scenes
If GR or CO have difficulty perceiving or inter-preting the spatial layout of scenes then we pre-dicted they would do worse when the stimuli areempty rooms or Lego scenes than when they arereal-world or Lego objects Note that the criticalcomparison is between the Lego scenes and theLego objects as these stimuli are made of the samematerials and differ primarily in their surfacegeometry
MethodTesting for all experiments was done on aMacintosh computer The experiment consisted of80 trials (20 of each stimulus type randomly inter-mixed) Subjects began each trial by pressing thespace bar A black-and-white photograph from oneof the four stimulus categories (see earlier) thenappeared on the screen with two red dots on it Thetask was to determine which of the two red dotsindicated the closer part of the scene or objectSubjects pressed the ldquofrdquo key if the leftmost red dotwas closest and the ldquojrdquo key if the rightmost dotwas closest They were instructed to respond asquickly as possible without making errors Stimuliremained on the screen until a response was made
The stimulus set was constructed so that eachempty room was matched to a common object thathad red dots in the exact same location and eachLego layout was similarly matched to a Lego objectThus the locations of the red dots were counterbal-anced across scene and object stimuli controllingfor any effects of visual field cuts
Results and discussionResults for GR and CO and four elderly normalsubjects are shown in Table 3 Both GR and COcould do this task without difficulty and their per-formance was near ceiling There was no evidencethat they had any more trouble with the emptyrooms or Lego scenes than with the real or Legoobjects Reaction times were slightly longer for GRand CO than for normal subjects as one wouldexpect given their visual field deficits however this
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 495
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 3 Experiment 1 (two-dot) results (SD in parenthesis)
GR CO Normals
Proportion correctObject 095 080 093Lego object 100 095 098Empty room 100 100 099Lego room 095 100 096
Mean RTObject 2435 2932 2161 (980)Lego object 2052 3245 2029 (921)Empty room 2744 2291 1788 (959)Lego room 1904 2281 1630 (1127)
Figure 6 The four kinds of stimuli used in Experiment 1 Topleft lego layouts top right real-world layouts (empty rooms)bottom left lego objects bottom right real-world objects
difference did not appear to be significant Impor-tantly there was little evidence that GR or CO tooklonger to make their perceptual judgements for thereal or Lego scenes than for the real or Lego objectsThus contrary to our initial hypothesis this experi-ment found no clear evidence for selective impair-ment in the perception of scene-like stimuli
Experiment 2 Different-views matchingtask
The previous experiment demonstrated that GRand CO can extract information about the per-ceived distances between themselves and the sur-faces of an object or scene Thus in at least onesense their perception of these surfaces is pre-served However this does not mean that their abil-ity to represent the layout of a scene is unimpairedThe two-dot task requires on-line comparison oftwo surfaces in egocentric coordinates GR and COwould be able to perform this task even if they couldnot (1) represent more than two surfaces at a time(2) represent surfaces in other than strictly egocen-tric coordinates or (3) sustain surface representa-tions for longer than the time it takes to make acomparison It has been suggested that the parietallobes are involved in on-line processing of surfacegeometry in egocentric coordinates (Milner ampGoodale 1995) thus it is possible that the parietallobes (which are intact in GR and CO) may be suf-ficient to perform the two-dot task even in theabsence of more durable or complex surface repre-sentations in parahippocampal cortex
In Experiment 2 we examine GR and COrsquosability to create and sustain a representation of thegeometry of an object or scene-like layout The taskwas to view a photograph of a Lego object or Legoscene hold it in memory for 10 s and then deter-mine whether a second object or scene was the sameas the first In ldquosamerdquo trials the second photographdepicted the same scene or object depicted from adifferent viewing angle In ldquodifferentrdquo trials the sec-ond photograph depicted a different scene or objectchosen to be similar to the first one (Figure 7)Thus in order to successfully perform this tasksubjects had to form a representation of the surfacesof the object or scene sustain this representation in
memory for 10 s and then align it (presumablyusing mental rotation) to the second object orscene
MethodThe experiment consisted of two parts each with40 trials In the first part the stimuli were black andwhite photographs of objects made out of Legoblocks In the second part the stimuli were photo-graphs of ldquoscenesrdquo made out of Lego blocks whichwere designed to have a geometric structure similarto real-world places such as a roomor a city street
Subjects began each trial by pressing the spacebar A photograph of a Lego object or sceneappeared on the screen for 3 s followed by a 10-sblank interval A second photograph then appeareddepicting either (1) the same object or scene as thefirst photograph shown from a different viewingangle (offset approximately 35 degrees) or (2) a dif-ferent object or scene Subjects were instructed topress the ldquojrdquo key if the two photographs depicted thesame object or scene and the ldquof rdquo key if they did notThey were told to take as much time as they neededto make a decision but no extra time The secondphotograph remained on the screen until a responsewas made Half the trials were ldquosamerdquo trials and halfwere ldquodifferentrdquo trials
Results and discussionFigure 8 shows results for GR and CO as well as forsix elderly normal controls GRrsquos and COrsquos perfor-mance was comparable to that of the normal con-trols Furthermore GR and CO performed just aswell when the stimuli were Lego scenes as whenthey were Lego objects These results are unlikely toreflect a ceiling effect as performance for bothpatients and controls was around 80 correctThus both patients appear to be able to form a rep-resentation of the geometry of both scenes andobjects which they can sustain in short-termmemory
Experiment 3 N-back recognition memorytask
The previous experiment failed to find any impair-ment in GR and COrsquos ability to form a representa-
EPSTEIN ET AL
496 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
tion of the geometry of a scene or object andmaintain it for 10 s in the absence of the originalstimulus Not only does this result suggest that per-ception of scene-like stimuli is intact in thesepatients it further suggests that some short-termmemory functions are intact as well However we
should note that subjects in this experiment werefree to devote all their effort and attention in the 10-second blank interval to maintaining a memory ofthe stimulus Thus it is possible that GR and COcan use rehearsal mechanisms to maintain the stim-ulus in short-term memory during the blank inter-
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 497
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
(b) Lego object version
Figure 7 Stimuli and procedure for Experiment 2 (a) In the Lego layout version subjects see a Lego layout for 3 s followed by a blankscreen for 10 s A second layout then appears which is either the same layout as the first one but seen from a different angle or a differentlayout (b) The Lego object version is the same but with object stimuli
3 s 10 s OR
3 s 10 s OR
(a) Lego layout version
(b) Lego object version
val but are unable to form a more durable memorytrace for the stimulus This possibility was tested inthe current experiment which was designed toexamine GR and COrsquos ability to learn new visualinformation
We devised a continuous n-back recognitionmemory test in which subjects viewed a sequence oforiginally novel stimuli some of which wererepeated within the sequence at intervals of 1 3 or5 intervening images For each stimulus subjects
reported whether they had seen it before in theexperiment or not In different runs in the experi-ment the stimuli were black-and-white photo-graphs of (1) Lego scenes (2) Lego objects (3) real-world landscapes (4) novel objects made out ofSculpey clay (5) unfamiliar houses (6) unfamiliarfaces as well as (7) line drawings of nonsenseobjects and (8) pronounceable pseudowords (Fig-ure 9) Thus we were able to test GR and COrsquos rec-ognition memory for different kinds of materials atdifferent time intervals in a paradigm in which thepresence of intervening items between the twoappearances of each stimulus made rehearsal diffi-cult Note that because none of the specific stimuliused in this experiment were familiar in advancethis task did not require an episodic memory com-ponent and could be done on the basis of pure visualfamiliarity with the particular visual stimulus
Although many different stimulus classes weretested in this experiment the critical comparison isbetween the Lego layouts and Lego objectsbecause these are best matched with regard tomaterials and differ primarily in their geometricstructure (see Introduction) We predicted that ifGR and CO showed a memory deficit for this taskit would be greater for the Lego layouts than for the
EPSTEIN ET AL
498 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 8 Results of Experiment 2 Performance over ldquosamerdquotrials and ldquodifferentrdquo trials is averaged together Error bars show1 SD
Figure 9 Stimuli for Experiment 3 Top row Lego layouts real-world scenes houses Bottom row Lego objects clay objects nonsenseobjects faces In an eighth condition (not shown) subjects viewed pronounceable nonwords such as ldquowilchrdquo or ldquothipperrdquo
Lego objects In addition a deficit for materialsthat convey information about the layout of sur-rounding space might also be apparent when GRand CO are tested on more realistic stimuli If soone would expect them to do poorly on the land-scapes but relatively well on the clay objects linedrawings of nonsense objects faces andpseudowords
MethodFor each subject a complete experimental sessionconsisted of being run once on each of the eightstimulus classes Each run consisted of 90 trials ineach trial a stimulus was presented and subjectsreported whether or not they had seen it before inthe run (or indeed ever) by pressing the ldquojrdquo key ifthey had or the ldquof rdquo key if they had not Each stimu-lus was on the screen for either 1 2 or 3 s (see fol-lowing) and appeared promptly after the keypressreport for the previous trial Individual stimuliappeared either once (14 stimuli) twice (30 stim-uli) or four times (4 stimuli) within a given run Ofthe stimuli that were presented twice 10 wererepeated immediately (1-back) 10 were repeatedwith a lag of two intervening items (3-back) and 10were repeated with a lag of four intervening items(5-back) In order to ensure that between-subjectdifferences in performance were not due to stimulusdifferences the same stimuli were assigned to thesame conditions each time the experimentwas run
GR performed the experiment twice once inAugust 1998 and again 9 months later Because ofa computer error he was only tested on six of theeight categories (all except Lego layouts and Legoobjects) at the later date CO was also tested twiceonce in February 1999 and again 6 months laterReported results for GR and CO are the average oftheir two testing sessions Normal subjects wereonly tested once Stimulus presentation time was 1 sin GRrsquos first testing session In pilot testing COfound this too fast for the critical categories of Lego
layouts and Lego objects so presentation time wasextended to 2 s for these categories in his first test-ing session All normal subjects were tested at thesepresentation rates (2 s for Lego layouts and objects1 s for other stimuli) In COrsquos second testingsession presentation time was lengthened to 3 s forall stimulus categories to ensure that any deficitsfound were memory deficits rather than problemswith the nonmemory aspects of the task
Results and discussionResults from GR CO and 10 elderly normal con-trols are shown in Figure 10 and Table 4 Figure 10ashows results from the main comparison betweenLego layouts and Lego objects Both normal sub-jects and patients performed near ceiling for bothLego layouts and Lego objects at 1-backinterstimulus intervals verifying that GR and COcould successfully perceive the pictures as they werebeing presented In contrast there was a strikingdifference between the two patients and the normalsubjects at 5-back intervals Whereas normals didon average somewhat better on Lego layouts thanLego objects both GR and CO showed the oppo-site pattern doing much worse on Lego layoutsthan Lego objects The 3-back condition showed amixed pattern Whereas GR did better on Legoobjects than Lego layouts in this condition (similarto his performance at 5-back intervals) CO didbetter on Lego layouts than Lego objects COrsquos lowperformance on Lego objects in the 3-back condi-tion appears to be an anomaly as it is much lowerthan his performance for comparable stimuli in the1-back and 5-back conditions
Figure 10b shows the difference in performancebetween Lego layouts and Lego objects for thecritical 1-back and 5-back intervals At 5-backintervals CO was significantly impaired comparedto the normal subjects (z = ndash250) whereasGRrsquos impairment fell just short of significance (z =ndash175)4
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 499
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
4In fact these values may underestimate GR and COrsquos relative impairment for Lego layouts In the 5-back condition 4 of the 10
normal subjects had perfect performance on the Lego layouts but all scored below ceiling on the Lego objects Thus the true advantagefor Lego layouts over Lego objects for normal subjects is probably greater than that observed here in which case GR and COrsquos lowerperformance for Lego layouts compared to Lego objects would be even more abnormal Furthermore post-hoc analyses revealed thatone of the normal subjects showed a difference in performance between Lego layouts and objects that was over 2 SD below the mean ofthe other nine subjects When this outlying subject was excluded z-scores were ndash322 for CO and ndash232 for GR
Table 4 shows results for all eight stimulusclasses Although direct comparisons between sub-jects should be treated with caution since viewingtime for different items was not the same for eachsubject we do note a few interesting patterns Firstof all there is little or no evidence that GR or CO
show a memory deficit for nonsense words clayobjects or line drawings of nonsense objects GRappears to be slightly impaired on faces and housesbut CO performs normally on these categoriesStrikingly GR shows a severe deficit for the reallandscapes performing more than 40 percentagepoints lower than normals Thus even thoughthere is some evidence that GR might have someimpairment for nonscene stimuli he clearly is muchmore impaired for scene-like stimuli such as theLego scenes and landscapes CO does not show asimilar deficit for the landscapes However itshould be kept in mind that in one of his two testingsessions CO viewed each picture for 3 s which wasmuch longer than the 1 second used for all non-Lego stimuli for GR and normals Thus COrsquosapparently ldquonormalrdquo performance in these condi-tions might be overestimated
Taken together the results of this experimentindicate that GR and CO are selectively impaired atprocessing scene-like stimuli This deficit is consis-tently apparent at 5-back but not 1-back intervalsThis finding suggests that the deficit should becharacterised as a memory encoding problemHowever the present results do not allow us todetermine whether the difficulty occurs at memoryencoding or at subsequent recognition (see Epsteinamp Kanwisher 2001) If GR and CO have a generalproblem with recognition of topographical materi-als then they should not be able to recognise scenes
EPSTEIN ET AL
500 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 10a Results of Experiment 3 for GR (top) CO (middle)and for 10 age-matched normal controls Performance measure isproportion of repeated stimuli correctly identified as such correctedfor guessing using the standard formula
Figure 10b Difference between Lego layout performance andLego object performance for the 1-back and 5-back conditionsError bars indicate one 1 SD
and landmarks that were learned prior to theirinjury We tested this possibility in the nextexperiment
Experiment 4 Famous landmarksfacesrecognition task
In this experiment we examined GR and COrsquosability to recognise the visual appearances of placeslearned prior to injury Subjects saw a series offamous landmarks intermixed with nonfamouscontrols (Imaging experiments have demonstratedthat landmarks of this type can activate the PPAalmost as strongly as complete scenes Epstein etal 1999 Epstein amp Kanwisher 1998) They wererequired to say whether they knew each specificlandmark and to identify the ones they knew Sub-jects were instructed to respond affirmatively if theywere fairly certain that they recognised the land-mark but not if it only looked generally familiarFor purposes of comparison subjects also per-formed an identical test in which the stimuli werefamous and nonfamous faces In both testsdistractor items were designed to be visually andconceptually similar to the target items (see Figure11) If parahippocampal cortex is required for placerecognition then one might expect GR and CO toperform poorly compared to normals on the land-mark recognition test
MethodEach of the two experiments (famous faces famouslandmarks) consisted of 48 trials Subjects beganeach trial by pressing the space bar after which ablack-and-white photograph of a face (in thefamous faces experiment) or a landmark (in thefamous landmarks experiment) appeared on thescreen Subjects were told to press the ldquojrdquo key if theyrecognised the specific face or landmark and the ldquof rdquokey if they did not taking as much time at theyneeded to make this discrimination but no extratime Subjects were told to report that they recog-nised a face or landmark if they felt that they knewit even if they could not retrieve its name If theydid know the name they were to say it after pressingthe button Otherwise they were to say whateverthey did remember about the item
Half of the photographs were of famous and halfof nonfamous people and landmarks The famousfaces included public figures (eg George BushRichard Nixon Prince Charles) and movie stars(eg Harrison Ford Clark Gable Marlon Brando)who were prominent prior to the time of GR andCOrsquos injuries the famous landmarks were build-ings and monuments from all over the world (egEiffel Tower US Capitol Building Great Sphinx)The nonfamous stimuli were chosen to match thefamous stimuli as closely as possible Each famousstimulus had a ldquomatchrdquo For example George Bushwas matched to a man of similar age in a coat andtie and the Taj Mahal was matched to a visually
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 501
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 4 Full results of Experiment 3 Mean proportion correct (corrected for guessing) SDs in parentheses
Lego Land- Lego Clay Nonsense Non-scenes scapes Houses objects objects objects Words Faces
1-back GR 100 094 100 100 100 094 100 100CO 091 085 091 100 100 079 074 100Normals 096(008) 095(008) 093(014) 096(008) 096(006) 099(003) 098(004) 097(011)
3-back GR ndash003 068 026 047 083 083 075 052CO 066 081 037 010 094 069 095 081Normals 066(032) 077(021) 072(015) 077(020) 086(016) 083(015) 083(017) 072(020)
5-back GR 010 034 044 047 072 083 094 053CO 031 073 075 088 081 064 100 081Normals 076(026) 077(016) 068(021) 066(016) 091(014) 083(018) 093(011) 078(021)
similar Asian palace that is not famous (see Figure11)
Results and discussionResults for GR and CO as well as for five age-matched controls are shown in Figure 12 Both
GR and CO could recognise the famous landmarksas well normals In addition both GR and CO wereusually able to name the landmarks they success-fully identified
Thus GR and CO can recognize landmarkslearned prior to injury demonstrating that at leastsome of their place recognition abilities are intactWhen combined with the fact that GR and COwere able to produce detailed maps of places theylived in prior to their injury (see Case Description)these results suggest that their deficit is primarily inencoding or consolidation of new informationrather than in recognition or recall of old informa-tion Note however that these results do not provethat their place recognition abilities are entirelynormal GR and CO might suffer from a mild rec-ognition deficit that affects identification of newly-learned stimuli like the Lego layouts in Experiment3 but not overlearned stimuli like the famous land-marks Furthermore it is possible that GR and COmight be impaired at recognising premorbidlylearned places that are defined primarily by theirspatial layout even though they are unimpaired atrecognising premorbidly learned places that aredefined primarily by a single object-like landmarkor building (see General Discussion) Neverthelessthe trend of the present evidence is to suggest thatGR and CO have particular problems with noveltopographical information
In the famous-face task GRrsquos performance wasin the normal range However he sometimes haddifficulty naming the faces even though he clearly
EPSTEIN ET AL
502 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 11 Stimuli for Experiment 4 Each famous landmark(Arc de Triomphe Taj Mahal) and famous face (Barbara BushYasser Arafat) was matched to a nonfamous building or face thatalso appeared in the experiment
FA
CE
SL
AN
DM
AR
KS
Figure 12 Results of Experiment 4 Performance measure isproportion of famous stimuli reported as ldquoknownrdquo corrected forguessing using the standard formula
recognised them For example he described Ron-ald Reagan as ldquothe gipperrdquo Nancy Reagan as ldquothewife of the gipperrdquo and Clark Gable as ldquothe movieactor who said lsquoScarlett I donrsquot give a damnrsquordquo COhad a great deal of difficulty identifying the famousfaces This result is not surprising given hisextremely low performance on the Benton FaceMatching Task In addition to his topographicproblems CO appears to suffer fromprosopagnosia
GENERAL DISCUSSION
We tested two parahippocampal patients (GR andCO) on four experiments designed to measure per-formance as a function of both task (perceptionencoding and recognition) and stimulus type(scenes objects faces and words) The only taskthat revealed a clear deficit was an n-back recogni-tion memory test (Experiment 3) In order to suc-cessfully perform this task subjects had to encode adurable representation of a visual stimulus and useit for later recognition GR and CO were signifi-cantly more impaired on this task when the stimuliwere scene-like spatial layouts (ie Lego scenesand real scenes for GR) than when they wereobjects No comparable deficits were found in otherexperiments that tested layout perception (Experi-ments 1 and 2) and placelandmark recognition(Experiment 4) Taken together these results indi-cate that GR and CO are impaired in their ability toencode novel information about the geometry ofsurrounding space into memory but are lessimpaired at their ability to encode the geometry ofnovel objects
We argue that this specific topographical defi-citmdashinability to encode the spatial layout of novelscenesmdashresults from damage to the para-hippocampal place area Although the damage inboth patients extends beyond the parahippo-campal-lingual territory normally associated withthe PPA this region is clearly lesioned bilaterally inGR and in the right hemisphere in CO Further-more functional imaging demonstrated that GRhas no functioning PPA Thus the current resultsand the earlier fMRI work (Epstein et al 1999
Epstein amp Kanwisher 1998) converge to implicateparahippocampal cortex in the encoding of infor-mation about scene geometry into memory
Our assignment of the function of memoryencoding to parahippocampal cortex is consistentwith the results of earlier case reports of patientswith parahippocampal damage who have naviga-tional difficulties in novel but not familiar environ-ments (Habib amp Sirigu 1987 Ross 1980 see alsoTeng amp Squire 1999) As with GR and CO thesepatients were either less impaired or unimpaired onnontopographical memory tasks Patients have alsobeen described who exhibit the opposite pattern ofimpairment an inability to learn faces (sometimescalled ldquoprosopamnesiardquo Cipolotti Robinson Blairamp Frith 1999 Tippett Miller amp Farah 2000) orfaces and verbal materials (Maguire amp Cipolotti1998) with an unimpaired ability to learn topo-graphical materials Taken together these resultsargue for a high degree of domain-specificity in theneural systems involved in learning of new informa-tion (Gallistel 1995) The brain may have separatemechanisms for learning faces places and wordswhich can be differentially impaired after damageto specific cortical regions (Cipolotti et al 1999Incisa della Rochetta et al 1996) The presentresults suggest that the anatomical locus of themechanism dedicated to learning new topographi-cal information is in the parahippocampal cortex(Aguirre amp DrsquoEsposito 1999)
In contrast to their impaired ability to encodenew topographical information both GR and COwere able to produce detailed maps of their previousdwellings and performed normally on the famouslandmark recognition test of Experiment 4 Theseresults are generally consistent with Teng andSquirersquos (1999) report of a patient with extensivemedial temporal damage who could recallpremorbidly learned but not postmorbidly learnedtopographical information (see also Ross 1980)Thus our results provide further evidence thatmedial temporal regions such as parahippocampalcortex may be more critical for encoding or consoli-dating new information into memory than for recallof old information
One issue that our experiments do not entirelyresolve is the precise nature of the topographical
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 503
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
information encoded by parahippocampal cortexClearly this region is critical for encoding informa-tion about the geometry of the currently visiblescene even when the ldquoscenerdquo is made out of Legoblocks and therefore has no immediate navigationalrelevance However it is unclear whether this spe-cific deficit can account for all of the real-worldnavigational difficulties encountered by GR andCO GR in particular shows such a catastrophicloss of navigational ability that we suspect he is alsoimpaired at encoding the spatial relationshipsbetweenthe current scene and other locations in theworld CO probably also suffers from such difficul-ties as he reports that he has trouble rememberinghow places relate to each other in large-scale spaceWhen he was tested on a simple 3-room videogameenvironment he not only was unable to rememberthe individual rooms (claiming that there were 10rooms when there were only 3) but he also had nomemory for the spatial relationships between themThese spatial deficits may be the result of damage tothe posterior right hippocampus (in the case of CO)or of disconnection of the posterior right hippo-campus from the parahippocampal regions thatprovide much of its cortical input (in both patientsSuzuki amp Amaral 1994) In either case the abilityto form a cognitive map of new environments likethe lab or videogame space could be impaired(Maguire et al 1998 OrsquoKeefe amp Nadel 1978)However it is also possible that parahippocampalcortex itself may be critical for encoding not onlythe geometry of individual scenes but also the spa-tial relationships between the locations of thosescenes In this case the spatial deficits of GR andCO could be attributed directly to parahippo-campal damage rather than to hippocampal dam-age or disconnection
The latter scenario would be consistent withAguirre and DrsquoEspositorsquos (1999) and Barrash etalrsquos (2000) suggestion that parahippocampal cortexis critically involved in combining visual informa-tion about the appearance of scenes and landmarkswith spatial information about their locations into aunified topographical representation Howeverboth of these authors argue that the representationsof individual scenes (Barrash et al) and landmarks(Aguirre amp DrsquoEsposito) are supported not by
parahippocampal cortex but by more posterior lin-gual-fusiform regions In contrast we believe thatthe neuroimaging data indicate that parahippo-campal cortex is involved in the encoding of thegeometric structure of individual scenes althoughit may also play a role in combining these represen-tations with other spatial representations
Interestingly GR and CO were not impaired onrecognition of famous landmarks that were mostlybuildings Thus our data do not preclude the possi-bility that there may be a separate lingual orfusiform region involved in processing informationabout buildings (Aguirre Zarahn amp DrsquoEsposito1998 Chao Haxby amp Martin 1999 IshaiUngerleider Martin Schouten amp Haxby 1999)which might have been preserved in GR and COdespite extensive damage to other regions of thelingual and fusiform gyrus Alternatively GR andCOrsquos preserved building recognition abilities mightbe mediated by interactions between cortical areasthat perform shape analysis for any object-likestimulus type (Malach et al 1995Kanwisher et al1996) and anterior temporal regions in which long-term topographical and semantic memories arestored (McCarthy et al 1996)
Is perception preserved
In contrast to their inability to encode new scenesinto memory GR and CO have substantiallyretained their ability to perceive the spatial layout ofthe currently visible scene (Experiments 1 and 2)GR can follow a route marked on a floor planwhich indicates a good on-line appreciation of hiscurrent spatial surroundings Furthermore neitherGR or CO have any trouble moving through theenvironmentmdashthey move with confidence and donot bump into doors or walls Problems of the lattertype are usually associated with damage to the pari-etal lobes which are intact in GR and CO Thus atfirst glance GR and CO do not appear to haveperceptual impairments (beyond their visual fieldrestrictions)
However GR does complain of a perceptualdeficit which he describes as an inability to ldquotakeinrdquo scenes when they are too complex Similarreports of an inability to comprehend the
EPSTEIN ET AL
504 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
ldquoglobalityrdquo of a scene were made by some of theparahippocampally-damaged patients studied byHabib and Sirigu (1987) We were not able todetermine the precise nature of this deficit whichmay be partly attributable to his visual field restric-tions It appears that his deficit is more salient in thereal-world environments than in the picture-per-ception tasks of Experiments 1 and 2 perhapsbecause he finds it more difficult to organise visualinput when immersed in it than when viewing pho-tographs that subtend only part of the visual field
We suggest that GR can perceive parts of thescene when he attends to them but may be unableto integrate information over time into a coherentrepresentation of a scene or place The fact that GRand CO are impaired for Lego layouts at 5-backintervals in Experiment 3 is consistent with thishypothesis The time between the first and secondpresentations of a stimulus is very short in this case(and even shorter with 3-back intervals where GRalso shows evidence of a deficit) In both patientsthe representation of layout information formed byperception of a Lego scene must either have anabnormally short persistence be abnormally sus-ceptible to interference or both If so they mightfind it impossible to sustain a spatial frame orschema long enough to use it to organise informa-tion obtained from many different glances at a com-plex scene (Intraub 1997) The result would be aninability to understand the ldquoglobalityrdquo of a sceneeven though all the details were perfectlyperceived
Non-topographical mnemonic abilities
Although not as severe as their topographical mem-ory problems GR and CO do have some memorydifficulties in other domains For example bothperformed poorly on the Rey AVLT whichrequired them to remember a list of unrelatedwords Both also report some subtle episodic mem-ory problems (see Case Description) COrsquos prob-lems could partly result from damage to the righthippocampusmdashindeed CO appeared to have themore acute episodic memory problems HoweverGR has no hippocampal damage so the fact that hedisplays episodic memory problems suggests thatparahippocampal cortex is involved in the forma-
tion of episodic memories (cf Wagner et al1998)
In previous work (Epstein et al 1999) weargued that spatial context information encoded byparahippocampal cortex might play a role in estab-lishing event memories by encoding informationabout where events take place The inability toencode this information might cause problems intasks such as the Rey AVLT where one has to recalla list of unrelated words If spatial (and perhapstemporal) context information is not encoded itwill be harder subsequently to recall the distinctevent of hearing each word (Tulving 1983) Incontrast lack of spatial context information wouldbe less of a problem in tasks where one is onlyrequired to recognise previously encountered verbalmaterials but not to reproduce the spatiotemporalcontext in which they were experienced IndeedGR and CO performed as well as normals on thenonsense words in Experiment 4 where recogni-tion memory but not episodic memory was requiredfor successful performance Interestingly GR andCO also did relatively well on the Logical Memorysubtest of the WMS-III perhaps because the nar-rative framework of a story provides sufficient con-textual information to overcome the absence ofspatial context and mediate successful verbalretrieval
Differences between GR and CO
Although we have discussed GR and CO togetherthere are a number of relevant differences betweenthem One open question is whether these differ-ences can be attributed to the different effects ofunilateral and bilateral parahippocampal damageIn general GRrsquos topographical difficulties weremore severe than COrsquos CO did surprisingly well onthe Warrington Topographical Memory Test inwhich he was required to remember detailed real-world city scenes and also performed substantiallybetter on real scenes than Lego scenes in Experi-ment 3 Insofar as CO has intact left para-hippocampal cortex this suggests that the left PPAmight be sufficient to encode a certain amount of(perhaps semantic) information about real-worldscenes but the right PPA is necessary to encode
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 505
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)

about the main room of the lab once he had left itCO was also able to draw a detailed floor plan of theapartment he was living in at the time (and hadlived in for 20 years) Several months after he hadmoved from this apartment we asked him to drawsuch a floor plan again and he produced one thatwas substantially the same as the first (Figure 3)Thus their map drawing performance indicatesthat GR and CO are primarily impaired at sketch-ing places experienced after their strokes but areunimpaired at sketching places experienced beforetheir strokes This pattern is indicative of deficits inencoding but not recall
Scene perceptionAs noted earlier GR can navigate with the aid of afloor plan suggesting that his perceptual abilitiesare largely intact Interestingly however he reportsthat he does have a perceptual deficit which hedescribes as an inability to understand scenes whenthey are too complex For example he reports thathe often has trouble using maps because theyusually are too busy with details It is unclearwhether this apparent perceptual problem is a resultof his field cut some other separate perceptual
problem or a memory problem (see discussion)CO did not complain of a similar deficit GR wasan accomplished artist prior to his injury producingpaintings of striking complexity Tragically hereports that he cannot now appreciate his ownpaintings
Other cognitive abilities
Face recognitionGR also reported some difficulties with face recog-nition On his second visit we examined his abilityto learn new faces by having him perform a forced-choice recognition test on seven familiar and sevenunfamiliar faces The familiar faces were personnelfrom the lab with whom he had at that point had agood deal of experience spanning at least 3 daysAll the people in the photographs had a towelwrapped around their head in order to minimise theuse of hair as a nonfacial cue Despite the difficultyof this task he correctly identified 67 familiarfaces and made no false alarms Surprisingly theone face that he did miss was the first author of thispaper with whom he at that point had had consid-erable experience GR also performed normally ona difficult famous face recognition test (see Experi-ment 4 following) and on the Benton Face Recog-nition test (see following) Thus he has someability to recognise faces Consistent with this GRreports that he can remember a small number offaces but that he quickly ldquoloses trackrdquo if he has toremember too many
CO also appears to suffer from someprosopagnosia as evidenced by his low perfor-mance on the Benton Face Recognition test (seefollowing see also Experiment 4) Interestingly hedoes not complain of face recognition difficultiesbut rather attributes his inability to recognise peo-ple to a lack of interest in their appearance
Episodic memoryAt first glance GRrsquos episodic memory appeared tobe largely intact As mentioned he could recall theevents of the previous day accurately Furthermorehe reports that he can follow the plot of a moviewithout any difficulty However both he and hiswife report that he does have some problems with
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 489
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Figure 2 Floor plan of the building at MIT in which GR wastested Corridors are highlighted in grey GR was asked to find themain room of the lab (marked G on the map) Starting location(marked S) was a point approximately 30 feet down the hall GRrsquosroute is marked by arrows GR found this task very difficult andonly knew for certain that he had reached his goal when he caughtsight of his wife through the glass door of the lab In contrast hewas able to follow a route marked on a plan of the same floorwithout any problem or hesitation
episodic memory For example he stated that heremembered very little about a trip to Italy they hadtaken several months previously His wife reportsthat he has great difficulty keeping track of timeFor example before their trip to Boston he repeat-edly asked her what day they were supposed to leaveand seemed unable to encode this information Weobserved a similar phenomenon during testingDuring the tests that required repetition of thesame procedure several times he sometimesseemed unable to clearly remember how manytimes he had repeated the procedure Thus he doesappear to have a subtle episodic memory deficit
which may manifest itself as an inability to sepa-rately remember successive events that are not easilydistinguishable from each other
CO also reports some problems with his epi-sodic memory He relies on lists to rememberappointments stating ldquoif I didnrsquot have a list Irsquodprobably forget why I walked in the door hererdquoDespite this claim he has never missed an appoint-ment nor is he ever confused about the purpose ofthe testing appointments He can accurately reportthe day of the week He remembers the experi-menters clearly from appointment to appointmentas well as the details of the various experiments
EPSTEIN ET AL
490 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 3 Two floor plans drawn by CO of the apartment he lived in for over 20 years The first plan was drawn when CO still lived inthe apartment the second was drawn several months later when he had moved out Note that the two plans correspond very well COreported that it was quite easy to produce these floor plans
CO home Feb 99
CO (former) home June 99
when they are repeated across appointments Whenqueried about his episodic memory difficulties COreported that he will usually remember an event ifhe is reminded of it but that he has difficulty spon-taneously regenerating events Interestingly hereports that even when he is reminded of an eventand remembers it he cannot remember ldquowhere itwas or when it wasrdquo When he reads he finds it dif-ficult to follow a story if it skips around Howeverwhen queried he was able to accurately report whathe had read in the newspaper the previous day
Formal neuropsychological tests
Formal neuropsychological testing was performedon GR in September 1998and on CO in December1998 Test performances were compared to age-matched peers3
GR exhibited mild to moderate impairment inmemory relative to what would be predicted for hisintellect (WAIS-R Full Scale IQ =122 93rd per-centile superior range) and level of educationThese memory problems were more apparent onvisual than on verbal memory tests On a test ofmemory for conceptually organised text passages(Wechsler Memory Scale Logical Memory[WMS-III] Wechsler 1997b) his performancewas average for both immediate (LM I =25th per-centile standard score SS = 90) and delayed recall(LM II =50th percentile SS =100) In contrast hisscore on the WMS-III Face Memory subtest was inthe low average range for both immediate (9th per-centile SS =80) and delayed recall (16th percentileSS =85) On a test of memory for scenes (WMS-IIIFamily Pictures subtest) immediate recall was defi-cient (2nd percentile SS = 69) and delayed recallwas borderline impaired (5th percentile SS =76)GR also had difficulty learning word lists on theRey Auditory Verbal Learning Test (Spreen ampStrauss 1998) his immediate recall was average(63rd percentile SS = 105) but after the interfer-ence list his performance fell to the low averagerange for both immediate (14th percentile SS =84)
and delayed recall (16th percentile SS =85) On arecognition test of the items of the list he success-fully recognised all 15 but made 8 false alarms Per-formance on the 724 Spatial Recall Test (RaoHammeke McQuillen Khatri amp Lloyd 1984)which required him to remember seven locations ona 6times4 checkerboard was 66th percentile (SS =106)for immediate and 2nd percentile (SS = 69) fordelayed recall Performance on the Benton FaceRecognition test was normal (42nd percentile)Performance on the Boston Naming Test was atceiling indicating that language abilities wereintact Thus GR exhibits some impairment ontasks that require delayed recall of visual informa-tion (WMS-III Face Memory subtest) or verbalinformation that is not organised in terms of a storyor plot (Rey AVLT) and is clearly impaired ontasks that required delayed recall of spatial informa-tion (724 Spatial Recall Test) or stimulus location(WMS-III Family Pictures subtest) In contrasthis ability to recall organised verbal information(WMS-III Logical Memory subtest) appears to beintact
As with GR COrsquos memory performance wasgenerally below what would be expected based onintellect (WAIS-III Full Scale IQ =134 99th per-centile very superior range) and education levelAlso like GR there was some evidence that visualmemory problems were more pronounced than ver-bal memory problems Memory for text passages(WMS-R Logical Memory) was average for bothimmediate (27th percentile SS = 91) and delayedrecall (48th percentileSS =99) In contrast perfor-mance on the WMS-III Faces subtest was lowaverage for both immediate (16th percentile SS =85) and delayed recall (9th percentile SS =80) andperformance on the Family Pictures subtest wasdeficient (lt 1st percentile SSlt 62) for both imme-diate and delayed recall On the Warrington Rec-ognition Memory Tests (RMT) he scored in theaverage range for words (50th percentile SS =100)and scenes (Warrington Topographical Recogni-tion Memory Test 29th percentile SS =92) and in
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 491
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
3The formal neuropsychological testing reported in this section was performed prior to the beginning of our combined
investigation of GR and CO (ie prior to the main experimental investigations of this paper) Consequently GR and CO were nottested on exactly the same battery of tests
the deficient range (lt 5th percentile SS lt 75) forfaces The low score for faces on this test as well asthe WMS-III may reflect prosopagnosia ratherthan a memory problem as he scored in the defi-cient range (lt1st percentile SSlt62) on the BentonFace Recognition test Verbal list learning (ReyAVLT) was average on immediate recall (45th per-centile SS =98) but after hearing an interferencelist performance fell to the deficient level (015words recalled) for both immediate and delayedrecall On a recognition test for these words herecognised 1115 and made one false alarm (39thpercentile SS =96) There was a large discrepancyon tests of memory span which may indicate adecline in spatial memory digit span was superior(9+ forward and 7 backwards both 99th percentile)but spatial span was only average (5 forward and 5backward 37th and 50th percentiles respectively)Language abilities were intact (Boston NamingTest performance was at ceiling) Thus like GRCO shows some impairment at recall of visualinformation (WMS-III Face Memory subtest) andverbal information that is not organised into a nar-rative (Rey AVLT) and clear impairment at tasksthat require recall of stimulus location (Family Pic-tures subtests of WMS-III) In contrast his abilityto recall organised verbal information (WMS-RLogical Memory subtest) is more preserved
MRI RESULTS
In order to determine the extent of their lesionsstructural MRI images were taken of both subjectsDelineation of the lesions was based either onspoiled GRASS coronal images at 11mm thick-ness (GR) or anatomical T-1 weighted MPRAGEMRI sequences yielding 125 mm thick coronalimages (CO) The anterior-posterior extent of thelesions was verified on axial images
GR has bilateral occipital-temporal lesions(Figure 4) In the left hemisphere the posteriorparahippocampal gyrus (including para-hippocampal cortex) is severely damaged with onlyminimal injury to the most caudal aspect of the hip-pocampus There is also moderate damage to themedial fusiform and inferior lingual gyri In the
right hemisphere the hippocampus is spared butthere is damage to the posterior end of theparahippocampal gyrus near the parahippocampal-lingual boundary More posteriorly the lesionedarea extends along the collateral sulcus to includesignificant portions of the inferior lingual gyrus andthe medial fusiform gyrus as well as most of themedial occipital lobe below the calcarine fissure
CO has a right-hemisphere occipital-temporallesion Right posterior hippocampus and para-hippocampal gyrus are severely affected Moreposteriorly the damage covers large portions of theinferior lingual and medial fusiform gyri as well assubstantial portions of the medial occipital lobeincluding most of right calcarine cortex In addi-tion diffuse foci of increased signal on T-2weighted images was present in periventricularwhite matter in the corona radiata and centrumsemiovale with a lacunar infarct of the rightputamen There was also moderate degree of dif-fuse involutional changes with increased ventricularsize particularly notable in the right temporal hornwith ex vacuo dilitation
The structural MRIs suggested that the PPAwas severely if not completely damaged in bothhemispheres for GR and in the right hemispherefor CO In order to confirm this for GR we used ahigh-field 3-T magnet to perform a functional scanin which he viewed scenes faces and objects in thesame blocked fMRI design that we have used tolocalise the PPA in all our previous experiments(see Epstein et al 1999 for details Epstein ampKanwisher 1998) A Kolomogorov-Smirnov testfound no voxels in the parahippocampal region thatfit our previously established criterion for inclusionin the PPA (ie greater response to scenes than tofaces and objects at a uncorrected significance levelof plt 0001) This result contrasts sharply with thepattern seen in over 95 of (mostly young or mid-dle-aged) normal subjects in whom a large acti-vated region can be identified in both left and rightparahippocampal cortex using this comparison (seeFigure 5) A similar test was used to identify voxelsthat responded significantly more to faces than toobjects This comparison revealed a region of face-selective voxels in the right fusiform gyrus consis-tent with the location of the Fusiform Face Area
EPSTEIN ET AL
492 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 493
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
GR
CO
Figure 4 Structural MRIs of GR and CO showing the extent of the damaged region (marked as white) Images have been normalisedusing SPM99 and a cost function masking algorithm (provided by Matthew Brett) into standard stereotactic space (Talairach amp Tournoux1988) using the Montreal Neurological Institute template (Evans et al 1993) Numbers indicate y-Talairach coordinate for each coronalslice corresponding slice planes are also indicated on the sagittal image
(FFA) commonly seen in normal subjects(Kanwisher McDermott amp Chun 1997) Thusthe MRI was sensitive enough to detect functionaldifferences an FFA was found but no PPA Whencombined with the anatomical results this stronglysuggests that GR has no PPA
Because of safety concerns (presence of a femo-ral artery aneurysm clip) we did not perform a simi-lar high-field functional MRI of CO However asFigure 4 demonstrates CO clearly has damage tothe parahippocampal-lingual region in the righthemisphere
EXPERIMENTAL INVESTIGATIONS
In this section we report the results of four experi-ments designed to probe the consequences of dam-age to the lingual-parahippocampal region ingeneral and the PPA in particular Our primaryconcern in these experiments was to test GR andCOrsquos ability to perceive remember and recognisedifferent kinds of visual material Based on our pre-vious fMRI work we predicted that GR and CO
would have more difficulty with scene-like stimuli(ie stimuli that convey information about thegeometry of a space one can navigate through or actwithin) than with object-like stimuli Such a deficitcould account for at least some of the problems GRand CO encounter in real-world navigation
Beyond determining the kind of visual materialthat gets processed in parahippocampal cortex wealso wanted to determine the nature of the process-ing that takes place on that material In particularwe wanted to determine if parahippocampal cortexis involved in perceptual encoding memory encodingor recognition As discussed earlier (Introduction)our fMRI results provided some evidence againstplace recognition as the sole function of the PPAbut did not allow us to distinguish between percep-tual and mnemonic encoding Experiments 1 and 2examine the role of parahippocampal cortex in per-ceptual encoding Experiment 3 examines its role inmnemonic encoding and Experiment 4 examinesits role in place recognition
Experiment 1 Two-dot task
The first hypothesis we examined was thatparahippocampal cortex plays a role in perceivingthe shape of the local environment In our previousfMRI work we observed that the PPA respondsmuch more strongly during scene viewing than dur-ing object or face viewing even when subjects donothing more than watch the scenes passively with-out performing any other task This suggested thatscene perception alone may be sufficient to activatethe PPA Previous reports have indicated that dam-age to parahippocampal cortex and other medialtemporal regions leads to problems with memoryrather than perception (Bohbot et al 1998Maguire Burke Philips amp Staunton 1996 Pigottamp Milner 1993) However all of these previousresults were obtained on patients who had unilat-eral lesions GR has a bilateral occipital-temporallesion including clear damage to parahippocampalcortex in both hemispheres furthermore he reportssome kind of perceptual deficit We hypothesisedthat this deficit might involve difficulty perceivingor interpreting the geometry of scenes and otherstimuli with a similar geometric structure
EPSTEIN ET AL
494 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 5 Top row shows a single slice from a functional MRIscan performed on GR Bottom row shows results from a normalsubject (NK) for comparison Voxels that responded more to facesthan objects (left column) or more to scenes than faces and objects(right column) in a Kolomogorov-Smirnov test are highlighted bywhite arrows GR has a functioning face area (FFA) but there isno evidence of a place area (PPA)
FFA PPA
GR
NK
In this experiment subjects viewed either objectshapes or spatial layouts Stimuli consisted ofblack-and-white photographs of real-world stimuli(empty rooms vs common objects) or abstractstimuli made out of Lego blocks (Lego ldquoscenesrdquo vsLego ldquoobjectsrdquo) These stimuli have been previouslyshown to differentially activate the PPA in normalsubjects (Epstein et al 1999 Epstein ampKanwisher 1998) Two red dots were superim-posed on each photograph (Figure 6) The task wasto report which of the two red dots was over thecloser part of the object or scene This task wasdesigned to have no memory component (apartfrom remembering the instructions) but to requireaccurate 3D perception of objects and scenes
If GR or CO have difficulty perceiving or inter-preting the spatial layout of scenes then we pre-dicted they would do worse when the stimuli areempty rooms or Lego scenes than when they arereal-world or Lego objects Note that the criticalcomparison is between the Lego scenes and theLego objects as these stimuli are made of the samematerials and differ primarily in their surfacegeometry
MethodTesting for all experiments was done on aMacintosh computer The experiment consisted of80 trials (20 of each stimulus type randomly inter-mixed) Subjects began each trial by pressing thespace bar A black-and-white photograph from oneof the four stimulus categories (see earlier) thenappeared on the screen with two red dots on it Thetask was to determine which of the two red dotsindicated the closer part of the scene or objectSubjects pressed the ldquofrdquo key if the leftmost red dotwas closest and the ldquojrdquo key if the rightmost dotwas closest They were instructed to respond asquickly as possible without making errors Stimuliremained on the screen until a response was made
The stimulus set was constructed so that eachempty room was matched to a common object thathad red dots in the exact same location and eachLego layout was similarly matched to a Lego objectThus the locations of the red dots were counterbal-anced across scene and object stimuli controllingfor any effects of visual field cuts
Results and discussionResults for GR and CO and four elderly normalsubjects are shown in Table 3 Both GR and COcould do this task without difficulty and their per-formance was near ceiling There was no evidencethat they had any more trouble with the emptyrooms or Lego scenes than with the real or Legoobjects Reaction times were slightly longer for GRand CO than for normal subjects as one wouldexpect given their visual field deficits however this
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 495
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 3 Experiment 1 (two-dot) results (SD in parenthesis)
GR CO Normals
Proportion correctObject 095 080 093Lego object 100 095 098Empty room 100 100 099Lego room 095 100 096
Mean RTObject 2435 2932 2161 (980)Lego object 2052 3245 2029 (921)Empty room 2744 2291 1788 (959)Lego room 1904 2281 1630 (1127)
Figure 6 The four kinds of stimuli used in Experiment 1 Topleft lego layouts top right real-world layouts (empty rooms)bottom left lego objects bottom right real-world objects
difference did not appear to be significant Impor-tantly there was little evidence that GR or CO tooklonger to make their perceptual judgements for thereal or Lego scenes than for the real or Lego objectsThus contrary to our initial hypothesis this experi-ment found no clear evidence for selective impair-ment in the perception of scene-like stimuli
Experiment 2 Different-views matchingtask
The previous experiment demonstrated that GRand CO can extract information about the per-ceived distances between themselves and the sur-faces of an object or scene Thus in at least onesense their perception of these surfaces is pre-served However this does not mean that their abil-ity to represent the layout of a scene is unimpairedThe two-dot task requires on-line comparison oftwo surfaces in egocentric coordinates GR and COwould be able to perform this task even if they couldnot (1) represent more than two surfaces at a time(2) represent surfaces in other than strictly egocen-tric coordinates or (3) sustain surface representa-tions for longer than the time it takes to make acomparison It has been suggested that the parietallobes are involved in on-line processing of surfacegeometry in egocentric coordinates (Milner ampGoodale 1995) thus it is possible that the parietallobes (which are intact in GR and CO) may be suf-ficient to perform the two-dot task even in theabsence of more durable or complex surface repre-sentations in parahippocampal cortex
In Experiment 2 we examine GR and COrsquosability to create and sustain a representation of thegeometry of an object or scene-like layout The taskwas to view a photograph of a Lego object or Legoscene hold it in memory for 10 s and then deter-mine whether a second object or scene was the sameas the first In ldquosamerdquo trials the second photographdepicted the same scene or object depicted from adifferent viewing angle In ldquodifferentrdquo trials the sec-ond photograph depicted a different scene or objectchosen to be similar to the first one (Figure 7)Thus in order to successfully perform this tasksubjects had to form a representation of the surfacesof the object or scene sustain this representation in
memory for 10 s and then align it (presumablyusing mental rotation) to the second object orscene
MethodThe experiment consisted of two parts each with40 trials In the first part the stimuli were black andwhite photographs of objects made out of Legoblocks In the second part the stimuli were photo-graphs of ldquoscenesrdquo made out of Lego blocks whichwere designed to have a geometric structure similarto real-world places such as a roomor a city street
Subjects began each trial by pressing the spacebar A photograph of a Lego object or sceneappeared on the screen for 3 s followed by a 10-sblank interval A second photograph then appeareddepicting either (1) the same object or scene as thefirst photograph shown from a different viewingangle (offset approximately 35 degrees) or (2) a dif-ferent object or scene Subjects were instructed topress the ldquojrdquo key if the two photographs depicted thesame object or scene and the ldquof rdquo key if they did notThey were told to take as much time as they neededto make a decision but no extra time The secondphotograph remained on the screen until a responsewas made Half the trials were ldquosamerdquo trials and halfwere ldquodifferentrdquo trials
Results and discussionFigure 8 shows results for GR and CO as well as forsix elderly normal controls GRrsquos and COrsquos perfor-mance was comparable to that of the normal con-trols Furthermore GR and CO performed just aswell when the stimuli were Lego scenes as whenthey were Lego objects These results are unlikely toreflect a ceiling effect as performance for bothpatients and controls was around 80 correctThus both patients appear to be able to form a rep-resentation of the geometry of both scenes andobjects which they can sustain in short-termmemory
Experiment 3 N-back recognition memorytask
The previous experiment failed to find any impair-ment in GR and COrsquos ability to form a representa-
EPSTEIN ET AL
496 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
tion of the geometry of a scene or object andmaintain it for 10 s in the absence of the originalstimulus Not only does this result suggest that per-ception of scene-like stimuli is intact in thesepatients it further suggests that some short-termmemory functions are intact as well However we
should note that subjects in this experiment werefree to devote all their effort and attention in the 10-second blank interval to maintaining a memory ofthe stimulus Thus it is possible that GR and COcan use rehearsal mechanisms to maintain the stim-ulus in short-term memory during the blank inter-
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 497
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
(b) Lego object version
Figure 7 Stimuli and procedure for Experiment 2 (a) In the Lego layout version subjects see a Lego layout for 3 s followed by a blankscreen for 10 s A second layout then appears which is either the same layout as the first one but seen from a different angle or a differentlayout (b) The Lego object version is the same but with object stimuli
3 s 10 s OR
3 s 10 s OR
(a) Lego layout version
(b) Lego object version
val but are unable to form a more durable memorytrace for the stimulus This possibility was tested inthe current experiment which was designed toexamine GR and COrsquos ability to learn new visualinformation
We devised a continuous n-back recognitionmemory test in which subjects viewed a sequence oforiginally novel stimuli some of which wererepeated within the sequence at intervals of 1 3 or5 intervening images For each stimulus subjects
reported whether they had seen it before in theexperiment or not In different runs in the experi-ment the stimuli were black-and-white photo-graphs of (1) Lego scenes (2) Lego objects (3) real-world landscapes (4) novel objects made out ofSculpey clay (5) unfamiliar houses (6) unfamiliarfaces as well as (7) line drawings of nonsenseobjects and (8) pronounceable pseudowords (Fig-ure 9) Thus we were able to test GR and COrsquos rec-ognition memory for different kinds of materials atdifferent time intervals in a paradigm in which thepresence of intervening items between the twoappearances of each stimulus made rehearsal diffi-cult Note that because none of the specific stimuliused in this experiment were familiar in advancethis task did not require an episodic memory com-ponent and could be done on the basis of pure visualfamiliarity with the particular visual stimulus
Although many different stimulus classes weretested in this experiment the critical comparison isbetween the Lego layouts and Lego objectsbecause these are best matched with regard tomaterials and differ primarily in their geometricstructure (see Introduction) We predicted that ifGR and CO showed a memory deficit for this taskit would be greater for the Lego layouts than for the
EPSTEIN ET AL
498 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 8 Results of Experiment 2 Performance over ldquosamerdquotrials and ldquodifferentrdquo trials is averaged together Error bars show1 SD
Figure 9 Stimuli for Experiment 3 Top row Lego layouts real-world scenes houses Bottom row Lego objects clay objects nonsenseobjects faces In an eighth condition (not shown) subjects viewed pronounceable nonwords such as ldquowilchrdquo or ldquothipperrdquo
Lego objects In addition a deficit for materialsthat convey information about the layout of sur-rounding space might also be apparent when GRand CO are tested on more realistic stimuli If soone would expect them to do poorly on the land-scapes but relatively well on the clay objects linedrawings of nonsense objects faces andpseudowords
MethodFor each subject a complete experimental sessionconsisted of being run once on each of the eightstimulus classes Each run consisted of 90 trials ineach trial a stimulus was presented and subjectsreported whether or not they had seen it before inthe run (or indeed ever) by pressing the ldquojrdquo key ifthey had or the ldquof rdquo key if they had not Each stimu-lus was on the screen for either 1 2 or 3 s (see fol-lowing) and appeared promptly after the keypressreport for the previous trial Individual stimuliappeared either once (14 stimuli) twice (30 stim-uli) or four times (4 stimuli) within a given run Ofthe stimuli that were presented twice 10 wererepeated immediately (1-back) 10 were repeatedwith a lag of two intervening items (3-back) and 10were repeated with a lag of four intervening items(5-back) In order to ensure that between-subjectdifferences in performance were not due to stimulusdifferences the same stimuli were assigned to thesame conditions each time the experimentwas run
GR performed the experiment twice once inAugust 1998 and again 9 months later Because ofa computer error he was only tested on six of theeight categories (all except Lego layouts and Legoobjects) at the later date CO was also tested twiceonce in February 1999 and again 6 months laterReported results for GR and CO are the average oftheir two testing sessions Normal subjects wereonly tested once Stimulus presentation time was 1 sin GRrsquos first testing session In pilot testing COfound this too fast for the critical categories of Lego
layouts and Lego objects so presentation time wasextended to 2 s for these categories in his first test-ing session All normal subjects were tested at thesepresentation rates (2 s for Lego layouts and objects1 s for other stimuli) In COrsquos second testingsession presentation time was lengthened to 3 s forall stimulus categories to ensure that any deficitsfound were memory deficits rather than problemswith the nonmemory aspects of the task
Results and discussionResults from GR CO and 10 elderly normal con-trols are shown in Figure 10 and Table 4 Figure 10ashows results from the main comparison betweenLego layouts and Lego objects Both normal sub-jects and patients performed near ceiling for bothLego layouts and Lego objects at 1-backinterstimulus intervals verifying that GR and COcould successfully perceive the pictures as they werebeing presented In contrast there was a strikingdifference between the two patients and the normalsubjects at 5-back intervals Whereas normals didon average somewhat better on Lego layouts thanLego objects both GR and CO showed the oppo-site pattern doing much worse on Lego layoutsthan Lego objects The 3-back condition showed amixed pattern Whereas GR did better on Legoobjects than Lego layouts in this condition (similarto his performance at 5-back intervals) CO didbetter on Lego layouts than Lego objects COrsquos lowperformance on Lego objects in the 3-back condi-tion appears to be an anomaly as it is much lowerthan his performance for comparable stimuli in the1-back and 5-back conditions
Figure 10b shows the difference in performancebetween Lego layouts and Lego objects for thecritical 1-back and 5-back intervals At 5-backintervals CO was significantly impaired comparedto the normal subjects (z = ndash250) whereasGRrsquos impairment fell just short of significance (z =ndash175)4
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 499
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
4In fact these values may underestimate GR and COrsquos relative impairment for Lego layouts In the 5-back condition 4 of the 10
normal subjects had perfect performance on the Lego layouts but all scored below ceiling on the Lego objects Thus the true advantagefor Lego layouts over Lego objects for normal subjects is probably greater than that observed here in which case GR and COrsquos lowerperformance for Lego layouts compared to Lego objects would be even more abnormal Furthermore post-hoc analyses revealed thatone of the normal subjects showed a difference in performance between Lego layouts and objects that was over 2 SD below the mean ofthe other nine subjects When this outlying subject was excluded z-scores were ndash322 for CO and ndash232 for GR
Table 4 shows results for all eight stimulusclasses Although direct comparisons between sub-jects should be treated with caution since viewingtime for different items was not the same for eachsubject we do note a few interesting patterns Firstof all there is little or no evidence that GR or CO
show a memory deficit for nonsense words clayobjects or line drawings of nonsense objects GRappears to be slightly impaired on faces and housesbut CO performs normally on these categoriesStrikingly GR shows a severe deficit for the reallandscapes performing more than 40 percentagepoints lower than normals Thus even thoughthere is some evidence that GR might have someimpairment for nonscene stimuli he clearly is muchmore impaired for scene-like stimuli such as theLego scenes and landscapes CO does not show asimilar deficit for the landscapes However itshould be kept in mind that in one of his two testingsessions CO viewed each picture for 3 s which wasmuch longer than the 1 second used for all non-Lego stimuli for GR and normals Thus COrsquosapparently ldquonormalrdquo performance in these condi-tions might be overestimated
Taken together the results of this experimentindicate that GR and CO are selectively impaired atprocessing scene-like stimuli This deficit is consis-tently apparent at 5-back but not 1-back intervalsThis finding suggests that the deficit should becharacterised as a memory encoding problemHowever the present results do not allow us todetermine whether the difficulty occurs at memoryencoding or at subsequent recognition (see Epsteinamp Kanwisher 2001) If GR and CO have a generalproblem with recognition of topographical materi-als then they should not be able to recognise scenes
EPSTEIN ET AL
500 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 10a Results of Experiment 3 for GR (top) CO (middle)and for 10 age-matched normal controls Performance measure isproportion of repeated stimuli correctly identified as such correctedfor guessing using the standard formula
Figure 10b Difference between Lego layout performance andLego object performance for the 1-back and 5-back conditionsError bars indicate one 1 SD
and landmarks that were learned prior to theirinjury We tested this possibility in the nextexperiment
Experiment 4 Famous landmarksfacesrecognition task
In this experiment we examined GR and COrsquosability to recognise the visual appearances of placeslearned prior to injury Subjects saw a series offamous landmarks intermixed with nonfamouscontrols (Imaging experiments have demonstratedthat landmarks of this type can activate the PPAalmost as strongly as complete scenes Epstein etal 1999 Epstein amp Kanwisher 1998) They wererequired to say whether they knew each specificlandmark and to identify the ones they knew Sub-jects were instructed to respond affirmatively if theywere fairly certain that they recognised the land-mark but not if it only looked generally familiarFor purposes of comparison subjects also per-formed an identical test in which the stimuli werefamous and nonfamous faces In both testsdistractor items were designed to be visually andconceptually similar to the target items (see Figure11) If parahippocampal cortex is required for placerecognition then one might expect GR and CO toperform poorly compared to normals on the land-mark recognition test
MethodEach of the two experiments (famous faces famouslandmarks) consisted of 48 trials Subjects beganeach trial by pressing the space bar after which ablack-and-white photograph of a face (in thefamous faces experiment) or a landmark (in thefamous landmarks experiment) appeared on thescreen Subjects were told to press the ldquojrdquo key if theyrecognised the specific face or landmark and the ldquof rdquokey if they did not taking as much time at theyneeded to make this discrimination but no extratime Subjects were told to report that they recog-nised a face or landmark if they felt that they knewit even if they could not retrieve its name If theydid know the name they were to say it after pressingthe button Otherwise they were to say whateverthey did remember about the item
Half of the photographs were of famous and halfof nonfamous people and landmarks The famousfaces included public figures (eg George BushRichard Nixon Prince Charles) and movie stars(eg Harrison Ford Clark Gable Marlon Brando)who were prominent prior to the time of GR andCOrsquos injuries the famous landmarks were build-ings and monuments from all over the world (egEiffel Tower US Capitol Building Great Sphinx)The nonfamous stimuli were chosen to match thefamous stimuli as closely as possible Each famousstimulus had a ldquomatchrdquo For example George Bushwas matched to a man of similar age in a coat andtie and the Taj Mahal was matched to a visually
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 501
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 4 Full results of Experiment 3 Mean proportion correct (corrected for guessing) SDs in parentheses
Lego Land- Lego Clay Nonsense Non-scenes scapes Houses objects objects objects Words Faces
1-back GR 100 094 100 100 100 094 100 100CO 091 085 091 100 100 079 074 100Normals 096(008) 095(008) 093(014) 096(008) 096(006) 099(003) 098(004) 097(011)
3-back GR ndash003 068 026 047 083 083 075 052CO 066 081 037 010 094 069 095 081Normals 066(032) 077(021) 072(015) 077(020) 086(016) 083(015) 083(017) 072(020)
5-back GR 010 034 044 047 072 083 094 053CO 031 073 075 088 081 064 100 081Normals 076(026) 077(016) 068(021) 066(016) 091(014) 083(018) 093(011) 078(021)
similar Asian palace that is not famous (see Figure11)
Results and discussionResults for GR and CO as well as for five age-matched controls are shown in Figure 12 Both
GR and CO could recognise the famous landmarksas well normals In addition both GR and CO wereusually able to name the landmarks they success-fully identified
Thus GR and CO can recognize landmarkslearned prior to injury demonstrating that at leastsome of their place recognition abilities are intactWhen combined with the fact that GR and COwere able to produce detailed maps of places theylived in prior to their injury (see Case Description)these results suggest that their deficit is primarily inencoding or consolidation of new informationrather than in recognition or recall of old informa-tion Note however that these results do not provethat their place recognition abilities are entirelynormal GR and CO might suffer from a mild rec-ognition deficit that affects identification of newly-learned stimuli like the Lego layouts in Experiment3 but not overlearned stimuli like the famous land-marks Furthermore it is possible that GR and COmight be impaired at recognising premorbidlylearned places that are defined primarily by theirspatial layout even though they are unimpaired atrecognising premorbidly learned places that aredefined primarily by a single object-like landmarkor building (see General Discussion) Neverthelessthe trend of the present evidence is to suggest thatGR and CO have particular problems with noveltopographical information
In the famous-face task GRrsquos performance wasin the normal range However he sometimes haddifficulty naming the faces even though he clearly
EPSTEIN ET AL
502 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 11 Stimuli for Experiment 4 Each famous landmark(Arc de Triomphe Taj Mahal) and famous face (Barbara BushYasser Arafat) was matched to a nonfamous building or face thatalso appeared in the experiment
FA
CE
SL
AN
DM
AR
KS
Figure 12 Results of Experiment 4 Performance measure isproportion of famous stimuli reported as ldquoknownrdquo corrected forguessing using the standard formula
recognised them For example he described Ron-ald Reagan as ldquothe gipperrdquo Nancy Reagan as ldquothewife of the gipperrdquo and Clark Gable as ldquothe movieactor who said lsquoScarlett I donrsquot give a damnrsquordquo COhad a great deal of difficulty identifying the famousfaces This result is not surprising given hisextremely low performance on the Benton FaceMatching Task In addition to his topographicproblems CO appears to suffer fromprosopagnosia
GENERAL DISCUSSION
We tested two parahippocampal patients (GR andCO) on four experiments designed to measure per-formance as a function of both task (perceptionencoding and recognition) and stimulus type(scenes objects faces and words) The only taskthat revealed a clear deficit was an n-back recogni-tion memory test (Experiment 3) In order to suc-cessfully perform this task subjects had to encode adurable representation of a visual stimulus and useit for later recognition GR and CO were signifi-cantly more impaired on this task when the stimuliwere scene-like spatial layouts (ie Lego scenesand real scenes for GR) than when they wereobjects No comparable deficits were found in otherexperiments that tested layout perception (Experi-ments 1 and 2) and placelandmark recognition(Experiment 4) Taken together these results indi-cate that GR and CO are impaired in their ability toencode novel information about the geometry ofsurrounding space into memory but are lessimpaired at their ability to encode the geometry ofnovel objects
We argue that this specific topographical defi-citmdashinability to encode the spatial layout of novelscenesmdashresults from damage to the para-hippocampal place area Although the damage inboth patients extends beyond the parahippo-campal-lingual territory normally associated withthe PPA this region is clearly lesioned bilaterally inGR and in the right hemisphere in CO Further-more functional imaging demonstrated that GRhas no functioning PPA Thus the current resultsand the earlier fMRI work (Epstein et al 1999
Epstein amp Kanwisher 1998) converge to implicateparahippocampal cortex in the encoding of infor-mation about scene geometry into memory
Our assignment of the function of memoryencoding to parahippocampal cortex is consistentwith the results of earlier case reports of patientswith parahippocampal damage who have naviga-tional difficulties in novel but not familiar environ-ments (Habib amp Sirigu 1987 Ross 1980 see alsoTeng amp Squire 1999) As with GR and CO thesepatients were either less impaired or unimpaired onnontopographical memory tasks Patients have alsobeen described who exhibit the opposite pattern ofimpairment an inability to learn faces (sometimescalled ldquoprosopamnesiardquo Cipolotti Robinson Blairamp Frith 1999 Tippett Miller amp Farah 2000) orfaces and verbal materials (Maguire amp Cipolotti1998) with an unimpaired ability to learn topo-graphical materials Taken together these resultsargue for a high degree of domain-specificity in theneural systems involved in learning of new informa-tion (Gallistel 1995) The brain may have separatemechanisms for learning faces places and wordswhich can be differentially impaired after damageto specific cortical regions (Cipolotti et al 1999Incisa della Rochetta et al 1996) The presentresults suggest that the anatomical locus of themechanism dedicated to learning new topographi-cal information is in the parahippocampal cortex(Aguirre amp DrsquoEsposito 1999)
In contrast to their impaired ability to encodenew topographical information both GR and COwere able to produce detailed maps of their previousdwellings and performed normally on the famouslandmark recognition test of Experiment 4 Theseresults are generally consistent with Teng andSquirersquos (1999) report of a patient with extensivemedial temporal damage who could recallpremorbidly learned but not postmorbidly learnedtopographical information (see also Ross 1980)Thus our results provide further evidence thatmedial temporal regions such as parahippocampalcortex may be more critical for encoding or consoli-dating new information into memory than for recallof old information
One issue that our experiments do not entirelyresolve is the precise nature of the topographical
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 503
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
information encoded by parahippocampal cortexClearly this region is critical for encoding informa-tion about the geometry of the currently visiblescene even when the ldquoscenerdquo is made out of Legoblocks and therefore has no immediate navigationalrelevance However it is unclear whether this spe-cific deficit can account for all of the real-worldnavigational difficulties encountered by GR andCO GR in particular shows such a catastrophicloss of navigational ability that we suspect he is alsoimpaired at encoding the spatial relationshipsbetweenthe current scene and other locations in theworld CO probably also suffers from such difficul-ties as he reports that he has trouble rememberinghow places relate to each other in large-scale spaceWhen he was tested on a simple 3-room videogameenvironment he not only was unable to rememberthe individual rooms (claiming that there were 10rooms when there were only 3) but he also had nomemory for the spatial relationships between themThese spatial deficits may be the result of damage tothe posterior right hippocampus (in the case of CO)or of disconnection of the posterior right hippo-campus from the parahippocampal regions thatprovide much of its cortical input (in both patientsSuzuki amp Amaral 1994) In either case the abilityto form a cognitive map of new environments likethe lab or videogame space could be impaired(Maguire et al 1998 OrsquoKeefe amp Nadel 1978)However it is also possible that parahippocampalcortex itself may be critical for encoding not onlythe geometry of individual scenes but also the spa-tial relationships between the locations of thosescenes In this case the spatial deficits of GR andCO could be attributed directly to parahippo-campal damage rather than to hippocampal dam-age or disconnection
The latter scenario would be consistent withAguirre and DrsquoEspositorsquos (1999) and Barrash etalrsquos (2000) suggestion that parahippocampal cortexis critically involved in combining visual informa-tion about the appearance of scenes and landmarkswith spatial information about their locations into aunified topographical representation Howeverboth of these authors argue that the representationsof individual scenes (Barrash et al) and landmarks(Aguirre amp DrsquoEsposito) are supported not by
parahippocampal cortex but by more posterior lin-gual-fusiform regions In contrast we believe thatthe neuroimaging data indicate that parahippo-campal cortex is involved in the encoding of thegeometric structure of individual scenes althoughit may also play a role in combining these represen-tations with other spatial representations
Interestingly GR and CO were not impaired onrecognition of famous landmarks that were mostlybuildings Thus our data do not preclude the possi-bility that there may be a separate lingual orfusiform region involved in processing informationabout buildings (Aguirre Zarahn amp DrsquoEsposito1998 Chao Haxby amp Martin 1999 IshaiUngerleider Martin Schouten amp Haxby 1999)which might have been preserved in GR and COdespite extensive damage to other regions of thelingual and fusiform gyrus Alternatively GR andCOrsquos preserved building recognition abilities mightbe mediated by interactions between cortical areasthat perform shape analysis for any object-likestimulus type (Malach et al 1995Kanwisher et al1996) and anterior temporal regions in which long-term topographical and semantic memories arestored (McCarthy et al 1996)
Is perception preserved
In contrast to their inability to encode new scenesinto memory GR and CO have substantiallyretained their ability to perceive the spatial layout ofthe currently visible scene (Experiments 1 and 2)GR can follow a route marked on a floor planwhich indicates a good on-line appreciation of hiscurrent spatial surroundings Furthermore neitherGR or CO have any trouble moving through theenvironmentmdashthey move with confidence and donot bump into doors or walls Problems of the lattertype are usually associated with damage to the pari-etal lobes which are intact in GR and CO Thus atfirst glance GR and CO do not appear to haveperceptual impairments (beyond their visual fieldrestrictions)
However GR does complain of a perceptualdeficit which he describes as an inability to ldquotakeinrdquo scenes when they are too complex Similarreports of an inability to comprehend the
EPSTEIN ET AL
504 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
ldquoglobalityrdquo of a scene were made by some of theparahippocampally-damaged patients studied byHabib and Sirigu (1987) We were not able todetermine the precise nature of this deficit whichmay be partly attributable to his visual field restric-tions It appears that his deficit is more salient in thereal-world environments than in the picture-per-ception tasks of Experiments 1 and 2 perhapsbecause he finds it more difficult to organise visualinput when immersed in it than when viewing pho-tographs that subtend only part of the visual field
We suggest that GR can perceive parts of thescene when he attends to them but may be unableto integrate information over time into a coherentrepresentation of a scene or place The fact that GRand CO are impaired for Lego layouts at 5-backintervals in Experiment 3 is consistent with thishypothesis The time between the first and secondpresentations of a stimulus is very short in this case(and even shorter with 3-back intervals where GRalso shows evidence of a deficit) In both patientsthe representation of layout information formed byperception of a Lego scene must either have anabnormally short persistence be abnormally sus-ceptible to interference or both If so they mightfind it impossible to sustain a spatial frame orschema long enough to use it to organise informa-tion obtained from many different glances at a com-plex scene (Intraub 1997) The result would be aninability to understand the ldquoglobalityrdquo of a sceneeven though all the details were perfectlyperceived
Non-topographical mnemonic abilities
Although not as severe as their topographical mem-ory problems GR and CO do have some memorydifficulties in other domains For example bothperformed poorly on the Rey AVLT whichrequired them to remember a list of unrelatedwords Both also report some subtle episodic mem-ory problems (see Case Description) COrsquos prob-lems could partly result from damage to the righthippocampusmdashindeed CO appeared to have themore acute episodic memory problems HoweverGR has no hippocampal damage so the fact that hedisplays episodic memory problems suggests thatparahippocampal cortex is involved in the forma-
tion of episodic memories (cf Wagner et al1998)
In previous work (Epstein et al 1999) weargued that spatial context information encoded byparahippocampal cortex might play a role in estab-lishing event memories by encoding informationabout where events take place The inability toencode this information might cause problems intasks such as the Rey AVLT where one has to recalla list of unrelated words If spatial (and perhapstemporal) context information is not encoded itwill be harder subsequently to recall the distinctevent of hearing each word (Tulving 1983) Incontrast lack of spatial context information wouldbe less of a problem in tasks where one is onlyrequired to recognise previously encountered verbalmaterials but not to reproduce the spatiotemporalcontext in which they were experienced IndeedGR and CO performed as well as normals on thenonsense words in Experiment 4 where recogni-tion memory but not episodic memory was requiredfor successful performance Interestingly GR andCO also did relatively well on the Logical Memorysubtest of the WMS-III perhaps because the nar-rative framework of a story provides sufficient con-textual information to overcome the absence ofspatial context and mediate successful verbalretrieval
Differences between GR and CO
Although we have discussed GR and CO togetherthere are a number of relevant differences betweenthem One open question is whether these differ-ences can be attributed to the different effects ofunilateral and bilateral parahippocampal damageIn general GRrsquos topographical difficulties weremore severe than COrsquos CO did surprisingly well onthe Warrington Topographical Memory Test inwhich he was required to remember detailed real-world city scenes and also performed substantiallybetter on real scenes than Lego scenes in Experi-ment 3 Insofar as CO has intact left para-hippocampal cortex this suggests that the left PPAmight be sufficient to encode a certain amount of(perhaps semantic) information about real-worldscenes but the right PPA is necessary to encode
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 505
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)

episodic memory For example he stated that heremembered very little about a trip to Italy they hadtaken several months previously His wife reportsthat he has great difficulty keeping track of timeFor example before their trip to Boston he repeat-edly asked her what day they were supposed to leaveand seemed unable to encode this information Weobserved a similar phenomenon during testingDuring the tests that required repetition of thesame procedure several times he sometimesseemed unable to clearly remember how manytimes he had repeated the procedure Thus he doesappear to have a subtle episodic memory deficit
which may manifest itself as an inability to sepa-rately remember successive events that are not easilydistinguishable from each other
CO also reports some problems with his epi-sodic memory He relies on lists to rememberappointments stating ldquoif I didnrsquot have a list Irsquodprobably forget why I walked in the door hererdquoDespite this claim he has never missed an appoint-ment nor is he ever confused about the purpose ofthe testing appointments He can accurately reportthe day of the week He remembers the experi-menters clearly from appointment to appointmentas well as the details of the various experiments
EPSTEIN ET AL
490 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 3 Two floor plans drawn by CO of the apartment he lived in for over 20 years The first plan was drawn when CO still lived inthe apartment the second was drawn several months later when he had moved out Note that the two plans correspond very well COreported that it was quite easy to produce these floor plans
CO home Feb 99
CO (former) home June 99
when they are repeated across appointments Whenqueried about his episodic memory difficulties COreported that he will usually remember an event ifhe is reminded of it but that he has difficulty spon-taneously regenerating events Interestingly hereports that even when he is reminded of an eventand remembers it he cannot remember ldquowhere itwas or when it wasrdquo When he reads he finds it dif-ficult to follow a story if it skips around Howeverwhen queried he was able to accurately report whathe had read in the newspaper the previous day
Formal neuropsychological tests
Formal neuropsychological testing was performedon GR in September 1998and on CO in December1998 Test performances were compared to age-matched peers3
GR exhibited mild to moderate impairment inmemory relative to what would be predicted for hisintellect (WAIS-R Full Scale IQ =122 93rd per-centile superior range) and level of educationThese memory problems were more apparent onvisual than on verbal memory tests On a test ofmemory for conceptually organised text passages(Wechsler Memory Scale Logical Memory[WMS-III] Wechsler 1997b) his performancewas average for both immediate (LM I =25th per-centile standard score SS = 90) and delayed recall(LM II =50th percentile SS =100) In contrast hisscore on the WMS-III Face Memory subtest was inthe low average range for both immediate (9th per-centile SS =80) and delayed recall (16th percentileSS =85) On a test of memory for scenes (WMS-IIIFamily Pictures subtest) immediate recall was defi-cient (2nd percentile SS = 69) and delayed recallwas borderline impaired (5th percentile SS =76)GR also had difficulty learning word lists on theRey Auditory Verbal Learning Test (Spreen ampStrauss 1998) his immediate recall was average(63rd percentile SS = 105) but after the interfer-ence list his performance fell to the low averagerange for both immediate (14th percentile SS =84)
and delayed recall (16th percentile SS =85) On arecognition test of the items of the list he success-fully recognised all 15 but made 8 false alarms Per-formance on the 724 Spatial Recall Test (RaoHammeke McQuillen Khatri amp Lloyd 1984)which required him to remember seven locations ona 6times4 checkerboard was 66th percentile (SS =106)for immediate and 2nd percentile (SS = 69) fordelayed recall Performance on the Benton FaceRecognition test was normal (42nd percentile)Performance on the Boston Naming Test was atceiling indicating that language abilities wereintact Thus GR exhibits some impairment ontasks that require delayed recall of visual informa-tion (WMS-III Face Memory subtest) or verbalinformation that is not organised in terms of a storyor plot (Rey AVLT) and is clearly impaired ontasks that required delayed recall of spatial informa-tion (724 Spatial Recall Test) or stimulus location(WMS-III Family Pictures subtest) In contrasthis ability to recall organised verbal information(WMS-III Logical Memory subtest) appears to beintact
As with GR COrsquos memory performance wasgenerally below what would be expected based onintellect (WAIS-III Full Scale IQ =134 99th per-centile very superior range) and education levelAlso like GR there was some evidence that visualmemory problems were more pronounced than ver-bal memory problems Memory for text passages(WMS-R Logical Memory) was average for bothimmediate (27th percentile SS = 91) and delayedrecall (48th percentileSS =99) In contrast perfor-mance on the WMS-III Faces subtest was lowaverage for both immediate (16th percentile SS =85) and delayed recall (9th percentile SS =80) andperformance on the Family Pictures subtest wasdeficient (lt 1st percentile SSlt 62) for both imme-diate and delayed recall On the Warrington Rec-ognition Memory Tests (RMT) he scored in theaverage range for words (50th percentile SS =100)and scenes (Warrington Topographical Recogni-tion Memory Test 29th percentile SS =92) and in
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 491
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
3The formal neuropsychological testing reported in this section was performed prior to the beginning of our combined
investigation of GR and CO (ie prior to the main experimental investigations of this paper) Consequently GR and CO were nottested on exactly the same battery of tests
the deficient range (lt 5th percentile SS lt 75) forfaces The low score for faces on this test as well asthe WMS-III may reflect prosopagnosia ratherthan a memory problem as he scored in the defi-cient range (lt1st percentile SSlt62) on the BentonFace Recognition test Verbal list learning (ReyAVLT) was average on immediate recall (45th per-centile SS =98) but after hearing an interferencelist performance fell to the deficient level (015words recalled) for both immediate and delayedrecall On a recognition test for these words herecognised 1115 and made one false alarm (39thpercentile SS =96) There was a large discrepancyon tests of memory span which may indicate adecline in spatial memory digit span was superior(9+ forward and 7 backwards both 99th percentile)but spatial span was only average (5 forward and 5backward 37th and 50th percentiles respectively)Language abilities were intact (Boston NamingTest performance was at ceiling) Thus like GRCO shows some impairment at recall of visualinformation (WMS-III Face Memory subtest) andverbal information that is not organised into a nar-rative (Rey AVLT) and clear impairment at tasksthat require recall of stimulus location (Family Pic-tures subtests of WMS-III) In contrast his abilityto recall organised verbal information (WMS-RLogical Memory subtest) is more preserved
MRI RESULTS
In order to determine the extent of their lesionsstructural MRI images were taken of both subjectsDelineation of the lesions was based either onspoiled GRASS coronal images at 11mm thick-ness (GR) or anatomical T-1 weighted MPRAGEMRI sequences yielding 125 mm thick coronalimages (CO) The anterior-posterior extent of thelesions was verified on axial images
GR has bilateral occipital-temporal lesions(Figure 4) In the left hemisphere the posteriorparahippocampal gyrus (including para-hippocampal cortex) is severely damaged with onlyminimal injury to the most caudal aspect of the hip-pocampus There is also moderate damage to themedial fusiform and inferior lingual gyri In the
right hemisphere the hippocampus is spared butthere is damage to the posterior end of theparahippocampal gyrus near the parahippocampal-lingual boundary More posteriorly the lesionedarea extends along the collateral sulcus to includesignificant portions of the inferior lingual gyrus andthe medial fusiform gyrus as well as most of themedial occipital lobe below the calcarine fissure
CO has a right-hemisphere occipital-temporallesion Right posterior hippocampus and para-hippocampal gyrus are severely affected Moreposteriorly the damage covers large portions of theinferior lingual and medial fusiform gyri as well assubstantial portions of the medial occipital lobeincluding most of right calcarine cortex In addi-tion diffuse foci of increased signal on T-2weighted images was present in periventricularwhite matter in the corona radiata and centrumsemiovale with a lacunar infarct of the rightputamen There was also moderate degree of dif-fuse involutional changes with increased ventricularsize particularly notable in the right temporal hornwith ex vacuo dilitation
The structural MRIs suggested that the PPAwas severely if not completely damaged in bothhemispheres for GR and in the right hemispherefor CO In order to confirm this for GR we used ahigh-field 3-T magnet to perform a functional scanin which he viewed scenes faces and objects in thesame blocked fMRI design that we have used tolocalise the PPA in all our previous experiments(see Epstein et al 1999 for details Epstein ampKanwisher 1998) A Kolomogorov-Smirnov testfound no voxels in the parahippocampal region thatfit our previously established criterion for inclusionin the PPA (ie greater response to scenes than tofaces and objects at a uncorrected significance levelof plt 0001) This result contrasts sharply with thepattern seen in over 95 of (mostly young or mid-dle-aged) normal subjects in whom a large acti-vated region can be identified in both left and rightparahippocampal cortex using this comparison (seeFigure 5) A similar test was used to identify voxelsthat responded significantly more to faces than toobjects This comparison revealed a region of face-selective voxels in the right fusiform gyrus consis-tent with the location of the Fusiform Face Area
EPSTEIN ET AL
492 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 493
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
GR
CO
Figure 4 Structural MRIs of GR and CO showing the extent of the damaged region (marked as white) Images have been normalisedusing SPM99 and a cost function masking algorithm (provided by Matthew Brett) into standard stereotactic space (Talairach amp Tournoux1988) using the Montreal Neurological Institute template (Evans et al 1993) Numbers indicate y-Talairach coordinate for each coronalslice corresponding slice planes are also indicated on the sagittal image
(FFA) commonly seen in normal subjects(Kanwisher McDermott amp Chun 1997) Thusthe MRI was sensitive enough to detect functionaldifferences an FFA was found but no PPA Whencombined with the anatomical results this stronglysuggests that GR has no PPA
Because of safety concerns (presence of a femo-ral artery aneurysm clip) we did not perform a simi-lar high-field functional MRI of CO However asFigure 4 demonstrates CO clearly has damage tothe parahippocampal-lingual region in the righthemisphere
EXPERIMENTAL INVESTIGATIONS
In this section we report the results of four experi-ments designed to probe the consequences of dam-age to the lingual-parahippocampal region ingeneral and the PPA in particular Our primaryconcern in these experiments was to test GR andCOrsquos ability to perceive remember and recognisedifferent kinds of visual material Based on our pre-vious fMRI work we predicted that GR and CO
would have more difficulty with scene-like stimuli(ie stimuli that convey information about thegeometry of a space one can navigate through or actwithin) than with object-like stimuli Such a deficitcould account for at least some of the problems GRand CO encounter in real-world navigation
Beyond determining the kind of visual materialthat gets processed in parahippocampal cortex wealso wanted to determine the nature of the process-ing that takes place on that material In particularwe wanted to determine if parahippocampal cortexis involved in perceptual encoding memory encodingor recognition As discussed earlier (Introduction)our fMRI results provided some evidence againstplace recognition as the sole function of the PPAbut did not allow us to distinguish between percep-tual and mnemonic encoding Experiments 1 and 2examine the role of parahippocampal cortex in per-ceptual encoding Experiment 3 examines its role inmnemonic encoding and Experiment 4 examinesits role in place recognition
Experiment 1 Two-dot task
The first hypothesis we examined was thatparahippocampal cortex plays a role in perceivingthe shape of the local environment In our previousfMRI work we observed that the PPA respondsmuch more strongly during scene viewing than dur-ing object or face viewing even when subjects donothing more than watch the scenes passively with-out performing any other task This suggested thatscene perception alone may be sufficient to activatethe PPA Previous reports have indicated that dam-age to parahippocampal cortex and other medialtemporal regions leads to problems with memoryrather than perception (Bohbot et al 1998Maguire Burke Philips amp Staunton 1996 Pigottamp Milner 1993) However all of these previousresults were obtained on patients who had unilat-eral lesions GR has a bilateral occipital-temporallesion including clear damage to parahippocampalcortex in both hemispheres furthermore he reportssome kind of perceptual deficit We hypothesisedthat this deficit might involve difficulty perceivingor interpreting the geometry of scenes and otherstimuli with a similar geometric structure
EPSTEIN ET AL
494 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 5 Top row shows a single slice from a functional MRIscan performed on GR Bottom row shows results from a normalsubject (NK) for comparison Voxels that responded more to facesthan objects (left column) or more to scenes than faces and objects(right column) in a Kolomogorov-Smirnov test are highlighted bywhite arrows GR has a functioning face area (FFA) but there isno evidence of a place area (PPA)
FFA PPA
GR
NK
In this experiment subjects viewed either objectshapes or spatial layouts Stimuli consisted ofblack-and-white photographs of real-world stimuli(empty rooms vs common objects) or abstractstimuli made out of Lego blocks (Lego ldquoscenesrdquo vsLego ldquoobjectsrdquo) These stimuli have been previouslyshown to differentially activate the PPA in normalsubjects (Epstein et al 1999 Epstein ampKanwisher 1998) Two red dots were superim-posed on each photograph (Figure 6) The task wasto report which of the two red dots was over thecloser part of the object or scene This task wasdesigned to have no memory component (apartfrom remembering the instructions) but to requireaccurate 3D perception of objects and scenes
If GR or CO have difficulty perceiving or inter-preting the spatial layout of scenes then we pre-dicted they would do worse when the stimuli areempty rooms or Lego scenes than when they arereal-world or Lego objects Note that the criticalcomparison is between the Lego scenes and theLego objects as these stimuli are made of the samematerials and differ primarily in their surfacegeometry
MethodTesting for all experiments was done on aMacintosh computer The experiment consisted of80 trials (20 of each stimulus type randomly inter-mixed) Subjects began each trial by pressing thespace bar A black-and-white photograph from oneof the four stimulus categories (see earlier) thenappeared on the screen with two red dots on it Thetask was to determine which of the two red dotsindicated the closer part of the scene or objectSubjects pressed the ldquofrdquo key if the leftmost red dotwas closest and the ldquojrdquo key if the rightmost dotwas closest They were instructed to respond asquickly as possible without making errors Stimuliremained on the screen until a response was made
The stimulus set was constructed so that eachempty room was matched to a common object thathad red dots in the exact same location and eachLego layout was similarly matched to a Lego objectThus the locations of the red dots were counterbal-anced across scene and object stimuli controllingfor any effects of visual field cuts
Results and discussionResults for GR and CO and four elderly normalsubjects are shown in Table 3 Both GR and COcould do this task without difficulty and their per-formance was near ceiling There was no evidencethat they had any more trouble with the emptyrooms or Lego scenes than with the real or Legoobjects Reaction times were slightly longer for GRand CO than for normal subjects as one wouldexpect given their visual field deficits however this
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 495
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 3 Experiment 1 (two-dot) results (SD in parenthesis)
GR CO Normals
Proportion correctObject 095 080 093Lego object 100 095 098Empty room 100 100 099Lego room 095 100 096
Mean RTObject 2435 2932 2161 (980)Lego object 2052 3245 2029 (921)Empty room 2744 2291 1788 (959)Lego room 1904 2281 1630 (1127)
Figure 6 The four kinds of stimuli used in Experiment 1 Topleft lego layouts top right real-world layouts (empty rooms)bottom left lego objects bottom right real-world objects
difference did not appear to be significant Impor-tantly there was little evidence that GR or CO tooklonger to make their perceptual judgements for thereal or Lego scenes than for the real or Lego objectsThus contrary to our initial hypothesis this experi-ment found no clear evidence for selective impair-ment in the perception of scene-like stimuli
Experiment 2 Different-views matchingtask
The previous experiment demonstrated that GRand CO can extract information about the per-ceived distances between themselves and the sur-faces of an object or scene Thus in at least onesense their perception of these surfaces is pre-served However this does not mean that their abil-ity to represent the layout of a scene is unimpairedThe two-dot task requires on-line comparison oftwo surfaces in egocentric coordinates GR and COwould be able to perform this task even if they couldnot (1) represent more than two surfaces at a time(2) represent surfaces in other than strictly egocen-tric coordinates or (3) sustain surface representa-tions for longer than the time it takes to make acomparison It has been suggested that the parietallobes are involved in on-line processing of surfacegeometry in egocentric coordinates (Milner ampGoodale 1995) thus it is possible that the parietallobes (which are intact in GR and CO) may be suf-ficient to perform the two-dot task even in theabsence of more durable or complex surface repre-sentations in parahippocampal cortex
In Experiment 2 we examine GR and COrsquosability to create and sustain a representation of thegeometry of an object or scene-like layout The taskwas to view a photograph of a Lego object or Legoscene hold it in memory for 10 s and then deter-mine whether a second object or scene was the sameas the first In ldquosamerdquo trials the second photographdepicted the same scene or object depicted from adifferent viewing angle In ldquodifferentrdquo trials the sec-ond photograph depicted a different scene or objectchosen to be similar to the first one (Figure 7)Thus in order to successfully perform this tasksubjects had to form a representation of the surfacesof the object or scene sustain this representation in
memory for 10 s and then align it (presumablyusing mental rotation) to the second object orscene
MethodThe experiment consisted of two parts each with40 trials In the first part the stimuli were black andwhite photographs of objects made out of Legoblocks In the second part the stimuli were photo-graphs of ldquoscenesrdquo made out of Lego blocks whichwere designed to have a geometric structure similarto real-world places such as a roomor a city street
Subjects began each trial by pressing the spacebar A photograph of a Lego object or sceneappeared on the screen for 3 s followed by a 10-sblank interval A second photograph then appeareddepicting either (1) the same object or scene as thefirst photograph shown from a different viewingangle (offset approximately 35 degrees) or (2) a dif-ferent object or scene Subjects were instructed topress the ldquojrdquo key if the two photographs depicted thesame object or scene and the ldquof rdquo key if they did notThey were told to take as much time as they neededto make a decision but no extra time The secondphotograph remained on the screen until a responsewas made Half the trials were ldquosamerdquo trials and halfwere ldquodifferentrdquo trials
Results and discussionFigure 8 shows results for GR and CO as well as forsix elderly normal controls GRrsquos and COrsquos perfor-mance was comparable to that of the normal con-trols Furthermore GR and CO performed just aswell when the stimuli were Lego scenes as whenthey were Lego objects These results are unlikely toreflect a ceiling effect as performance for bothpatients and controls was around 80 correctThus both patients appear to be able to form a rep-resentation of the geometry of both scenes andobjects which they can sustain in short-termmemory
Experiment 3 N-back recognition memorytask
The previous experiment failed to find any impair-ment in GR and COrsquos ability to form a representa-
EPSTEIN ET AL
496 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
tion of the geometry of a scene or object andmaintain it for 10 s in the absence of the originalstimulus Not only does this result suggest that per-ception of scene-like stimuli is intact in thesepatients it further suggests that some short-termmemory functions are intact as well However we
should note that subjects in this experiment werefree to devote all their effort and attention in the 10-second blank interval to maintaining a memory ofthe stimulus Thus it is possible that GR and COcan use rehearsal mechanisms to maintain the stim-ulus in short-term memory during the blank inter-
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 497
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
(b) Lego object version
Figure 7 Stimuli and procedure for Experiment 2 (a) In the Lego layout version subjects see a Lego layout for 3 s followed by a blankscreen for 10 s A second layout then appears which is either the same layout as the first one but seen from a different angle or a differentlayout (b) The Lego object version is the same but with object stimuli
3 s 10 s OR
3 s 10 s OR
(a) Lego layout version
(b) Lego object version
val but are unable to form a more durable memorytrace for the stimulus This possibility was tested inthe current experiment which was designed toexamine GR and COrsquos ability to learn new visualinformation
We devised a continuous n-back recognitionmemory test in which subjects viewed a sequence oforiginally novel stimuli some of which wererepeated within the sequence at intervals of 1 3 or5 intervening images For each stimulus subjects
reported whether they had seen it before in theexperiment or not In different runs in the experi-ment the stimuli were black-and-white photo-graphs of (1) Lego scenes (2) Lego objects (3) real-world landscapes (4) novel objects made out ofSculpey clay (5) unfamiliar houses (6) unfamiliarfaces as well as (7) line drawings of nonsenseobjects and (8) pronounceable pseudowords (Fig-ure 9) Thus we were able to test GR and COrsquos rec-ognition memory for different kinds of materials atdifferent time intervals in a paradigm in which thepresence of intervening items between the twoappearances of each stimulus made rehearsal diffi-cult Note that because none of the specific stimuliused in this experiment were familiar in advancethis task did not require an episodic memory com-ponent and could be done on the basis of pure visualfamiliarity with the particular visual stimulus
Although many different stimulus classes weretested in this experiment the critical comparison isbetween the Lego layouts and Lego objectsbecause these are best matched with regard tomaterials and differ primarily in their geometricstructure (see Introduction) We predicted that ifGR and CO showed a memory deficit for this taskit would be greater for the Lego layouts than for the
EPSTEIN ET AL
498 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 8 Results of Experiment 2 Performance over ldquosamerdquotrials and ldquodifferentrdquo trials is averaged together Error bars show1 SD
Figure 9 Stimuli for Experiment 3 Top row Lego layouts real-world scenes houses Bottom row Lego objects clay objects nonsenseobjects faces In an eighth condition (not shown) subjects viewed pronounceable nonwords such as ldquowilchrdquo or ldquothipperrdquo
Lego objects In addition a deficit for materialsthat convey information about the layout of sur-rounding space might also be apparent when GRand CO are tested on more realistic stimuli If soone would expect them to do poorly on the land-scapes but relatively well on the clay objects linedrawings of nonsense objects faces andpseudowords
MethodFor each subject a complete experimental sessionconsisted of being run once on each of the eightstimulus classes Each run consisted of 90 trials ineach trial a stimulus was presented and subjectsreported whether or not they had seen it before inthe run (or indeed ever) by pressing the ldquojrdquo key ifthey had or the ldquof rdquo key if they had not Each stimu-lus was on the screen for either 1 2 or 3 s (see fol-lowing) and appeared promptly after the keypressreport for the previous trial Individual stimuliappeared either once (14 stimuli) twice (30 stim-uli) or four times (4 stimuli) within a given run Ofthe stimuli that were presented twice 10 wererepeated immediately (1-back) 10 were repeatedwith a lag of two intervening items (3-back) and 10were repeated with a lag of four intervening items(5-back) In order to ensure that between-subjectdifferences in performance were not due to stimulusdifferences the same stimuli were assigned to thesame conditions each time the experimentwas run
GR performed the experiment twice once inAugust 1998 and again 9 months later Because ofa computer error he was only tested on six of theeight categories (all except Lego layouts and Legoobjects) at the later date CO was also tested twiceonce in February 1999 and again 6 months laterReported results for GR and CO are the average oftheir two testing sessions Normal subjects wereonly tested once Stimulus presentation time was 1 sin GRrsquos first testing session In pilot testing COfound this too fast for the critical categories of Lego
layouts and Lego objects so presentation time wasextended to 2 s for these categories in his first test-ing session All normal subjects were tested at thesepresentation rates (2 s for Lego layouts and objects1 s for other stimuli) In COrsquos second testingsession presentation time was lengthened to 3 s forall stimulus categories to ensure that any deficitsfound were memory deficits rather than problemswith the nonmemory aspects of the task
Results and discussionResults from GR CO and 10 elderly normal con-trols are shown in Figure 10 and Table 4 Figure 10ashows results from the main comparison betweenLego layouts and Lego objects Both normal sub-jects and patients performed near ceiling for bothLego layouts and Lego objects at 1-backinterstimulus intervals verifying that GR and COcould successfully perceive the pictures as they werebeing presented In contrast there was a strikingdifference between the two patients and the normalsubjects at 5-back intervals Whereas normals didon average somewhat better on Lego layouts thanLego objects both GR and CO showed the oppo-site pattern doing much worse on Lego layoutsthan Lego objects The 3-back condition showed amixed pattern Whereas GR did better on Legoobjects than Lego layouts in this condition (similarto his performance at 5-back intervals) CO didbetter on Lego layouts than Lego objects COrsquos lowperformance on Lego objects in the 3-back condi-tion appears to be an anomaly as it is much lowerthan his performance for comparable stimuli in the1-back and 5-back conditions
Figure 10b shows the difference in performancebetween Lego layouts and Lego objects for thecritical 1-back and 5-back intervals At 5-backintervals CO was significantly impaired comparedto the normal subjects (z = ndash250) whereasGRrsquos impairment fell just short of significance (z =ndash175)4
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 499
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
4In fact these values may underestimate GR and COrsquos relative impairment for Lego layouts In the 5-back condition 4 of the 10
normal subjects had perfect performance on the Lego layouts but all scored below ceiling on the Lego objects Thus the true advantagefor Lego layouts over Lego objects for normal subjects is probably greater than that observed here in which case GR and COrsquos lowerperformance for Lego layouts compared to Lego objects would be even more abnormal Furthermore post-hoc analyses revealed thatone of the normal subjects showed a difference in performance between Lego layouts and objects that was over 2 SD below the mean ofthe other nine subjects When this outlying subject was excluded z-scores were ndash322 for CO and ndash232 for GR
Table 4 shows results for all eight stimulusclasses Although direct comparisons between sub-jects should be treated with caution since viewingtime for different items was not the same for eachsubject we do note a few interesting patterns Firstof all there is little or no evidence that GR or CO
show a memory deficit for nonsense words clayobjects or line drawings of nonsense objects GRappears to be slightly impaired on faces and housesbut CO performs normally on these categoriesStrikingly GR shows a severe deficit for the reallandscapes performing more than 40 percentagepoints lower than normals Thus even thoughthere is some evidence that GR might have someimpairment for nonscene stimuli he clearly is muchmore impaired for scene-like stimuli such as theLego scenes and landscapes CO does not show asimilar deficit for the landscapes However itshould be kept in mind that in one of his two testingsessions CO viewed each picture for 3 s which wasmuch longer than the 1 second used for all non-Lego stimuli for GR and normals Thus COrsquosapparently ldquonormalrdquo performance in these condi-tions might be overestimated
Taken together the results of this experimentindicate that GR and CO are selectively impaired atprocessing scene-like stimuli This deficit is consis-tently apparent at 5-back but not 1-back intervalsThis finding suggests that the deficit should becharacterised as a memory encoding problemHowever the present results do not allow us todetermine whether the difficulty occurs at memoryencoding or at subsequent recognition (see Epsteinamp Kanwisher 2001) If GR and CO have a generalproblem with recognition of topographical materi-als then they should not be able to recognise scenes
EPSTEIN ET AL
500 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 10a Results of Experiment 3 for GR (top) CO (middle)and for 10 age-matched normal controls Performance measure isproportion of repeated stimuli correctly identified as such correctedfor guessing using the standard formula
Figure 10b Difference between Lego layout performance andLego object performance for the 1-back and 5-back conditionsError bars indicate one 1 SD
and landmarks that were learned prior to theirinjury We tested this possibility in the nextexperiment
Experiment 4 Famous landmarksfacesrecognition task
In this experiment we examined GR and COrsquosability to recognise the visual appearances of placeslearned prior to injury Subjects saw a series offamous landmarks intermixed with nonfamouscontrols (Imaging experiments have demonstratedthat landmarks of this type can activate the PPAalmost as strongly as complete scenes Epstein etal 1999 Epstein amp Kanwisher 1998) They wererequired to say whether they knew each specificlandmark and to identify the ones they knew Sub-jects were instructed to respond affirmatively if theywere fairly certain that they recognised the land-mark but not if it only looked generally familiarFor purposes of comparison subjects also per-formed an identical test in which the stimuli werefamous and nonfamous faces In both testsdistractor items were designed to be visually andconceptually similar to the target items (see Figure11) If parahippocampal cortex is required for placerecognition then one might expect GR and CO toperform poorly compared to normals on the land-mark recognition test
MethodEach of the two experiments (famous faces famouslandmarks) consisted of 48 trials Subjects beganeach trial by pressing the space bar after which ablack-and-white photograph of a face (in thefamous faces experiment) or a landmark (in thefamous landmarks experiment) appeared on thescreen Subjects were told to press the ldquojrdquo key if theyrecognised the specific face or landmark and the ldquof rdquokey if they did not taking as much time at theyneeded to make this discrimination but no extratime Subjects were told to report that they recog-nised a face or landmark if they felt that they knewit even if they could not retrieve its name If theydid know the name they were to say it after pressingthe button Otherwise they were to say whateverthey did remember about the item
Half of the photographs were of famous and halfof nonfamous people and landmarks The famousfaces included public figures (eg George BushRichard Nixon Prince Charles) and movie stars(eg Harrison Ford Clark Gable Marlon Brando)who were prominent prior to the time of GR andCOrsquos injuries the famous landmarks were build-ings and monuments from all over the world (egEiffel Tower US Capitol Building Great Sphinx)The nonfamous stimuli were chosen to match thefamous stimuli as closely as possible Each famousstimulus had a ldquomatchrdquo For example George Bushwas matched to a man of similar age in a coat andtie and the Taj Mahal was matched to a visually
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 501
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 4 Full results of Experiment 3 Mean proportion correct (corrected for guessing) SDs in parentheses
Lego Land- Lego Clay Nonsense Non-scenes scapes Houses objects objects objects Words Faces
1-back GR 100 094 100 100 100 094 100 100CO 091 085 091 100 100 079 074 100Normals 096(008) 095(008) 093(014) 096(008) 096(006) 099(003) 098(004) 097(011)
3-back GR ndash003 068 026 047 083 083 075 052CO 066 081 037 010 094 069 095 081Normals 066(032) 077(021) 072(015) 077(020) 086(016) 083(015) 083(017) 072(020)
5-back GR 010 034 044 047 072 083 094 053CO 031 073 075 088 081 064 100 081Normals 076(026) 077(016) 068(021) 066(016) 091(014) 083(018) 093(011) 078(021)
similar Asian palace that is not famous (see Figure11)
Results and discussionResults for GR and CO as well as for five age-matched controls are shown in Figure 12 Both
GR and CO could recognise the famous landmarksas well normals In addition both GR and CO wereusually able to name the landmarks they success-fully identified
Thus GR and CO can recognize landmarkslearned prior to injury demonstrating that at leastsome of their place recognition abilities are intactWhen combined with the fact that GR and COwere able to produce detailed maps of places theylived in prior to their injury (see Case Description)these results suggest that their deficit is primarily inencoding or consolidation of new informationrather than in recognition or recall of old informa-tion Note however that these results do not provethat their place recognition abilities are entirelynormal GR and CO might suffer from a mild rec-ognition deficit that affects identification of newly-learned stimuli like the Lego layouts in Experiment3 but not overlearned stimuli like the famous land-marks Furthermore it is possible that GR and COmight be impaired at recognising premorbidlylearned places that are defined primarily by theirspatial layout even though they are unimpaired atrecognising premorbidly learned places that aredefined primarily by a single object-like landmarkor building (see General Discussion) Neverthelessthe trend of the present evidence is to suggest thatGR and CO have particular problems with noveltopographical information
In the famous-face task GRrsquos performance wasin the normal range However he sometimes haddifficulty naming the faces even though he clearly
EPSTEIN ET AL
502 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 11 Stimuli for Experiment 4 Each famous landmark(Arc de Triomphe Taj Mahal) and famous face (Barbara BushYasser Arafat) was matched to a nonfamous building or face thatalso appeared in the experiment
FA
CE
SL
AN
DM
AR
KS
Figure 12 Results of Experiment 4 Performance measure isproportion of famous stimuli reported as ldquoknownrdquo corrected forguessing using the standard formula
recognised them For example he described Ron-ald Reagan as ldquothe gipperrdquo Nancy Reagan as ldquothewife of the gipperrdquo and Clark Gable as ldquothe movieactor who said lsquoScarlett I donrsquot give a damnrsquordquo COhad a great deal of difficulty identifying the famousfaces This result is not surprising given hisextremely low performance on the Benton FaceMatching Task In addition to his topographicproblems CO appears to suffer fromprosopagnosia
GENERAL DISCUSSION
We tested two parahippocampal patients (GR andCO) on four experiments designed to measure per-formance as a function of both task (perceptionencoding and recognition) and stimulus type(scenes objects faces and words) The only taskthat revealed a clear deficit was an n-back recogni-tion memory test (Experiment 3) In order to suc-cessfully perform this task subjects had to encode adurable representation of a visual stimulus and useit for later recognition GR and CO were signifi-cantly more impaired on this task when the stimuliwere scene-like spatial layouts (ie Lego scenesand real scenes for GR) than when they wereobjects No comparable deficits were found in otherexperiments that tested layout perception (Experi-ments 1 and 2) and placelandmark recognition(Experiment 4) Taken together these results indi-cate that GR and CO are impaired in their ability toencode novel information about the geometry ofsurrounding space into memory but are lessimpaired at their ability to encode the geometry ofnovel objects
We argue that this specific topographical defi-citmdashinability to encode the spatial layout of novelscenesmdashresults from damage to the para-hippocampal place area Although the damage inboth patients extends beyond the parahippo-campal-lingual territory normally associated withthe PPA this region is clearly lesioned bilaterally inGR and in the right hemisphere in CO Further-more functional imaging demonstrated that GRhas no functioning PPA Thus the current resultsand the earlier fMRI work (Epstein et al 1999
Epstein amp Kanwisher 1998) converge to implicateparahippocampal cortex in the encoding of infor-mation about scene geometry into memory
Our assignment of the function of memoryencoding to parahippocampal cortex is consistentwith the results of earlier case reports of patientswith parahippocampal damage who have naviga-tional difficulties in novel but not familiar environ-ments (Habib amp Sirigu 1987 Ross 1980 see alsoTeng amp Squire 1999) As with GR and CO thesepatients were either less impaired or unimpaired onnontopographical memory tasks Patients have alsobeen described who exhibit the opposite pattern ofimpairment an inability to learn faces (sometimescalled ldquoprosopamnesiardquo Cipolotti Robinson Blairamp Frith 1999 Tippett Miller amp Farah 2000) orfaces and verbal materials (Maguire amp Cipolotti1998) with an unimpaired ability to learn topo-graphical materials Taken together these resultsargue for a high degree of domain-specificity in theneural systems involved in learning of new informa-tion (Gallistel 1995) The brain may have separatemechanisms for learning faces places and wordswhich can be differentially impaired after damageto specific cortical regions (Cipolotti et al 1999Incisa della Rochetta et al 1996) The presentresults suggest that the anatomical locus of themechanism dedicated to learning new topographi-cal information is in the parahippocampal cortex(Aguirre amp DrsquoEsposito 1999)
In contrast to their impaired ability to encodenew topographical information both GR and COwere able to produce detailed maps of their previousdwellings and performed normally on the famouslandmark recognition test of Experiment 4 Theseresults are generally consistent with Teng andSquirersquos (1999) report of a patient with extensivemedial temporal damage who could recallpremorbidly learned but not postmorbidly learnedtopographical information (see also Ross 1980)Thus our results provide further evidence thatmedial temporal regions such as parahippocampalcortex may be more critical for encoding or consoli-dating new information into memory than for recallof old information
One issue that our experiments do not entirelyresolve is the precise nature of the topographical
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 503
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
information encoded by parahippocampal cortexClearly this region is critical for encoding informa-tion about the geometry of the currently visiblescene even when the ldquoscenerdquo is made out of Legoblocks and therefore has no immediate navigationalrelevance However it is unclear whether this spe-cific deficit can account for all of the real-worldnavigational difficulties encountered by GR andCO GR in particular shows such a catastrophicloss of navigational ability that we suspect he is alsoimpaired at encoding the spatial relationshipsbetweenthe current scene and other locations in theworld CO probably also suffers from such difficul-ties as he reports that he has trouble rememberinghow places relate to each other in large-scale spaceWhen he was tested on a simple 3-room videogameenvironment he not only was unable to rememberthe individual rooms (claiming that there were 10rooms when there were only 3) but he also had nomemory for the spatial relationships between themThese spatial deficits may be the result of damage tothe posterior right hippocampus (in the case of CO)or of disconnection of the posterior right hippo-campus from the parahippocampal regions thatprovide much of its cortical input (in both patientsSuzuki amp Amaral 1994) In either case the abilityto form a cognitive map of new environments likethe lab or videogame space could be impaired(Maguire et al 1998 OrsquoKeefe amp Nadel 1978)However it is also possible that parahippocampalcortex itself may be critical for encoding not onlythe geometry of individual scenes but also the spa-tial relationships between the locations of thosescenes In this case the spatial deficits of GR andCO could be attributed directly to parahippo-campal damage rather than to hippocampal dam-age or disconnection
The latter scenario would be consistent withAguirre and DrsquoEspositorsquos (1999) and Barrash etalrsquos (2000) suggestion that parahippocampal cortexis critically involved in combining visual informa-tion about the appearance of scenes and landmarkswith spatial information about their locations into aunified topographical representation Howeverboth of these authors argue that the representationsof individual scenes (Barrash et al) and landmarks(Aguirre amp DrsquoEsposito) are supported not by
parahippocampal cortex but by more posterior lin-gual-fusiform regions In contrast we believe thatthe neuroimaging data indicate that parahippo-campal cortex is involved in the encoding of thegeometric structure of individual scenes althoughit may also play a role in combining these represen-tations with other spatial representations
Interestingly GR and CO were not impaired onrecognition of famous landmarks that were mostlybuildings Thus our data do not preclude the possi-bility that there may be a separate lingual orfusiform region involved in processing informationabout buildings (Aguirre Zarahn amp DrsquoEsposito1998 Chao Haxby amp Martin 1999 IshaiUngerleider Martin Schouten amp Haxby 1999)which might have been preserved in GR and COdespite extensive damage to other regions of thelingual and fusiform gyrus Alternatively GR andCOrsquos preserved building recognition abilities mightbe mediated by interactions between cortical areasthat perform shape analysis for any object-likestimulus type (Malach et al 1995Kanwisher et al1996) and anterior temporal regions in which long-term topographical and semantic memories arestored (McCarthy et al 1996)
Is perception preserved
In contrast to their inability to encode new scenesinto memory GR and CO have substantiallyretained their ability to perceive the spatial layout ofthe currently visible scene (Experiments 1 and 2)GR can follow a route marked on a floor planwhich indicates a good on-line appreciation of hiscurrent spatial surroundings Furthermore neitherGR or CO have any trouble moving through theenvironmentmdashthey move with confidence and donot bump into doors or walls Problems of the lattertype are usually associated with damage to the pari-etal lobes which are intact in GR and CO Thus atfirst glance GR and CO do not appear to haveperceptual impairments (beyond their visual fieldrestrictions)
However GR does complain of a perceptualdeficit which he describes as an inability to ldquotakeinrdquo scenes when they are too complex Similarreports of an inability to comprehend the
EPSTEIN ET AL
504 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
ldquoglobalityrdquo of a scene were made by some of theparahippocampally-damaged patients studied byHabib and Sirigu (1987) We were not able todetermine the precise nature of this deficit whichmay be partly attributable to his visual field restric-tions It appears that his deficit is more salient in thereal-world environments than in the picture-per-ception tasks of Experiments 1 and 2 perhapsbecause he finds it more difficult to organise visualinput when immersed in it than when viewing pho-tographs that subtend only part of the visual field
We suggest that GR can perceive parts of thescene when he attends to them but may be unableto integrate information over time into a coherentrepresentation of a scene or place The fact that GRand CO are impaired for Lego layouts at 5-backintervals in Experiment 3 is consistent with thishypothesis The time between the first and secondpresentations of a stimulus is very short in this case(and even shorter with 3-back intervals where GRalso shows evidence of a deficit) In both patientsthe representation of layout information formed byperception of a Lego scene must either have anabnormally short persistence be abnormally sus-ceptible to interference or both If so they mightfind it impossible to sustain a spatial frame orschema long enough to use it to organise informa-tion obtained from many different glances at a com-plex scene (Intraub 1997) The result would be aninability to understand the ldquoglobalityrdquo of a sceneeven though all the details were perfectlyperceived
Non-topographical mnemonic abilities
Although not as severe as their topographical mem-ory problems GR and CO do have some memorydifficulties in other domains For example bothperformed poorly on the Rey AVLT whichrequired them to remember a list of unrelatedwords Both also report some subtle episodic mem-ory problems (see Case Description) COrsquos prob-lems could partly result from damage to the righthippocampusmdashindeed CO appeared to have themore acute episodic memory problems HoweverGR has no hippocampal damage so the fact that hedisplays episodic memory problems suggests thatparahippocampal cortex is involved in the forma-
tion of episodic memories (cf Wagner et al1998)
In previous work (Epstein et al 1999) weargued that spatial context information encoded byparahippocampal cortex might play a role in estab-lishing event memories by encoding informationabout where events take place The inability toencode this information might cause problems intasks such as the Rey AVLT where one has to recalla list of unrelated words If spatial (and perhapstemporal) context information is not encoded itwill be harder subsequently to recall the distinctevent of hearing each word (Tulving 1983) Incontrast lack of spatial context information wouldbe less of a problem in tasks where one is onlyrequired to recognise previously encountered verbalmaterials but not to reproduce the spatiotemporalcontext in which they were experienced IndeedGR and CO performed as well as normals on thenonsense words in Experiment 4 where recogni-tion memory but not episodic memory was requiredfor successful performance Interestingly GR andCO also did relatively well on the Logical Memorysubtest of the WMS-III perhaps because the nar-rative framework of a story provides sufficient con-textual information to overcome the absence ofspatial context and mediate successful verbalretrieval
Differences between GR and CO
Although we have discussed GR and CO togetherthere are a number of relevant differences betweenthem One open question is whether these differ-ences can be attributed to the different effects ofunilateral and bilateral parahippocampal damageIn general GRrsquos topographical difficulties weremore severe than COrsquos CO did surprisingly well onthe Warrington Topographical Memory Test inwhich he was required to remember detailed real-world city scenes and also performed substantiallybetter on real scenes than Lego scenes in Experi-ment 3 Insofar as CO has intact left para-hippocampal cortex this suggests that the left PPAmight be sufficient to encode a certain amount of(perhaps semantic) information about real-worldscenes but the right PPA is necessary to encode
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 505
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)

when they are repeated across appointments Whenqueried about his episodic memory difficulties COreported that he will usually remember an event ifhe is reminded of it but that he has difficulty spon-taneously regenerating events Interestingly hereports that even when he is reminded of an eventand remembers it he cannot remember ldquowhere itwas or when it wasrdquo When he reads he finds it dif-ficult to follow a story if it skips around Howeverwhen queried he was able to accurately report whathe had read in the newspaper the previous day
Formal neuropsychological tests
Formal neuropsychological testing was performedon GR in September 1998and on CO in December1998 Test performances were compared to age-matched peers3
GR exhibited mild to moderate impairment inmemory relative to what would be predicted for hisintellect (WAIS-R Full Scale IQ =122 93rd per-centile superior range) and level of educationThese memory problems were more apparent onvisual than on verbal memory tests On a test ofmemory for conceptually organised text passages(Wechsler Memory Scale Logical Memory[WMS-III] Wechsler 1997b) his performancewas average for both immediate (LM I =25th per-centile standard score SS = 90) and delayed recall(LM II =50th percentile SS =100) In contrast hisscore on the WMS-III Face Memory subtest was inthe low average range for both immediate (9th per-centile SS =80) and delayed recall (16th percentileSS =85) On a test of memory for scenes (WMS-IIIFamily Pictures subtest) immediate recall was defi-cient (2nd percentile SS = 69) and delayed recallwas borderline impaired (5th percentile SS =76)GR also had difficulty learning word lists on theRey Auditory Verbal Learning Test (Spreen ampStrauss 1998) his immediate recall was average(63rd percentile SS = 105) but after the interfer-ence list his performance fell to the low averagerange for both immediate (14th percentile SS =84)
and delayed recall (16th percentile SS =85) On arecognition test of the items of the list he success-fully recognised all 15 but made 8 false alarms Per-formance on the 724 Spatial Recall Test (RaoHammeke McQuillen Khatri amp Lloyd 1984)which required him to remember seven locations ona 6times4 checkerboard was 66th percentile (SS =106)for immediate and 2nd percentile (SS = 69) fordelayed recall Performance on the Benton FaceRecognition test was normal (42nd percentile)Performance on the Boston Naming Test was atceiling indicating that language abilities wereintact Thus GR exhibits some impairment ontasks that require delayed recall of visual informa-tion (WMS-III Face Memory subtest) or verbalinformation that is not organised in terms of a storyor plot (Rey AVLT) and is clearly impaired ontasks that required delayed recall of spatial informa-tion (724 Spatial Recall Test) or stimulus location(WMS-III Family Pictures subtest) In contrasthis ability to recall organised verbal information(WMS-III Logical Memory subtest) appears to beintact
As with GR COrsquos memory performance wasgenerally below what would be expected based onintellect (WAIS-III Full Scale IQ =134 99th per-centile very superior range) and education levelAlso like GR there was some evidence that visualmemory problems were more pronounced than ver-bal memory problems Memory for text passages(WMS-R Logical Memory) was average for bothimmediate (27th percentile SS = 91) and delayedrecall (48th percentileSS =99) In contrast perfor-mance on the WMS-III Faces subtest was lowaverage for both immediate (16th percentile SS =85) and delayed recall (9th percentile SS =80) andperformance on the Family Pictures subtest wasdeficient (lt 1st percentile SSlt 62) for both imme-diate and delayed recall On the Warrington Rec-ognition Memory Tests (RMT) he scored in theaverage range for words (50th percentile SS =100)and scenes (Warrington Topographical Recogni-tion Memory Test 29th percentile SS =92) and in
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 491
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
3The formal neuropsychological testing reported in this section was performed prior to the beginning of our combined
investigation of GR and CO (ie prior to the main experimental investigations of this paper) Consequently GR and CO were nottested on exactly the same battery of tests
the deficient range (lt 5th percentile SS lt 75) forfaces The low score for faces on this test as well asthe WMS-III may reflect prosopagnosia ratherthan a memory problem as he scored in the defi-cient range (lt1st percentile SSlt62) on the BentonFace Recognition test Verbal list learning (ReyAVLT) was average on immediate recall (45th per-centile SS =98) but after hearing an interferencelist performance fell to the deficient level (015words recalled) for both immediate and delayedrecall On a recognition test for these words herecognised 1115 and made one false alarm (39thpercentile SS =96) There was a large discrepancyon tests of memory span which may indicate adecline in spatial memory digit span was superior(9+ forward and 7 backwards both 99th percentile)but spatial span was only average (5 forward and 5backward 37th and 50th percentiles respectively)Language abilities were intact (Boston NamingTest performance was at ceiling) Thus like GRCO shows some impairment at recall of visualinformation (WMS-III Face Memory subtest) andverbal information that is not organised into a nar-rative (Rey AVLT) and clear impairment at tasksthat require recall of stimulus location (Family Pic-tures subtests of WMS-III) In contrast his abilityto recall organised verbal information (WMS-RLogical Memory subtest) is more preserved
MRI RESULTS
In order to determine the extent of their lesionsstructural MRI images were taken of both subjectsDelineation of the lesions was based either onspoiled GRASS coronal images at 11mm thick-ness (GR) or anatomical T-1 weighted MPRAGEMRI sequences yielding 125 mm thick coronalimages (CO) The anterior-posterior extent of thelesions was verified on axial images
GR has bilateral occipital-temporal lesions(Figure 4) In the left hemisphere the posteriorparahippocampal gyrus (including para-hippocampal cortex) is severely damaged with onlyminimal injury to the most caudal aspect of the hip-pocampus There is also moderate damage to themedial fusiform and inferior lingual gyri In the
right hemisphere the hippocampus is spared butthere is damage to the posterior end of theparahippocampal gyrus near the parahippocampal-lingual boundary More posteriorly the lesionedarea extends along the collateral sulcus to includesignificant portions of the inferior lingual gyrus andthe medial fusiform gyrus as well as most of themedial occipital lobe below the calcarine fissure
CO has a right-hemisphere occipital-temporallesion Right posterior hippocampus and para-hippocampal gyrus are severely affected Moreposteriorly the damage covers large portions of theinferior lingual and medial fusiform gyri as well assubstantial portions of the medial occipital lobeincluding most of right calcarine cortex In addi-tion diffuse foci of increased signal on T-2weighted images was present in periventricularwhite matter in the corona radiata and centrumsemiovale with a lacunar infarct of the rightputamen There was also moderate degree of dif-fuse involutional changes with increased ventricularsize particularly notable in the right temporal hornwith ex vacuo dilitation
The structural MRIs suggested that the PPAwas severely if not completely damaged in bothhemispheres for GR and in the right hemispherefor CO In order to confirm this for GR we used ahigh-field 3-T magnet to perform a functional scanin which he viewed scenes faces and objects in thesame blocked fMRI design that we have used tolocalise the PPA in all our previous experiments(see Epstein et al 1999 for details Epstein ampKanwisher 1998) A Kolomogorov-Smirnov testfound no voxels in the parahippocampal region thatfit our previously established criterion for inclusionin the PPA (ie greater response to scenes than tofaces and objects at a uncorrected significance levelof plt 0001) This result contrasts sharply with thepattern seen in over 95 of (mostly young or mid-dle-aged) normal subjects in whom a large acti-vated region can be identified in both left and rightparahippocampal cortex using this comparison (seeFigure 5) A similar test was used to identify voxelsthat responded significantly more to faces than toobjects This comparison revealed a region of face-selective voxels in the right fusiform gyrus consis-tent with the location of the Fusiform Face Area
EPSTEIN ET AL
492 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 493
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
GR
CO
Figure 4 Structural MRIs of GR and CO showing the extent of the damaged region (marked as white) Images have been normalisedusing SPM99 and a cost function masking algorithm (provided by Matthew Brett) into standard stereotactic space (Talairach amp Tournoux1988) using the Montreal Neurological Institute template (Evans et al 1993) Numbers indicate y-Talairach coordinate for each coronalslice corresponding slice planes are also indicated on the sagittal image
(FFA) commonly seen in normal subjects(Kanwisher McDermott amp Chun 1997) Thusthe MRI was sensitive enough to detect functionaldifferences an FFA was found but no PPA Whencombined with the anatomical results this stronglysuggests that GR has no PPA
Because of safety concerns (presence of a femo-ral artery aneurysm clip) we did not perform a simi-lar high-field functional MRI of CO However asFigure 4 demonstrates CO clearly has damage tothe parahippocampal-lingual region in the righthemisphere
EXPERIMENTAL INVESTIGATIONS
In this section we report the results of four experi-ments designed to probe the consequences of dam-age to the lingual-parahippocampal region ingeneral and the PPA in particular Our primaryconcern in these experiments was to test GR andCOrsquos ability to perceive remember and recognisedifferent kinds of visual material Based on our pre-vious fMRI work we predicted that GR and CO
would have more difficulty with scene-like stimuli(ie stimuli that convey information about thegeometry of a space one can navigate through or actwithin) than with object-like stimuli Such a deficitcould account for at least some of the problems GRand CO encounter in real-world navigation
Beyond determining the kind of visual materialthat gets processed in parahippocampal cortex wealso wanted to determine the nature of the process-ing that takes place on that material In particularwe wanted to determine if parahippocampal cortexis involved in perceptual encoding memory encodingor recognition As discussed earlier (Introduction)our fMRI results provided some evidence againstplace recognition as the sole function of the PPAbut did not allow us to distinguish between percep-tual and mnemonic encoding Experiments 1 and 2examine the role of parahippocampal cortex in per-ceptual encoding Experiment 3 examines its role inmnemonic encoding and Experiment 4 examinesits role in place recognition
Experiment 1 Two-dot task
The first hypothesis we examined was thatparahippocampal cortex plays a role in perceivingthe shape of the local environment In our previousfMRI work we observed that the PPA respondsmuch more strongly during scene viewing than dur-ing object or face viewing even when subjects donothing more than watch the scenes passively with-out performing any other task This suggested thatscene perception alone may be sufficient to activatethe PPA Previous reports have indicated that dam-age to parahippocampal cortex and other medialtemporal regions leads to problems with memoryrather than perception (Bohbot et al 1998Maguire Burke Philips amp Staunton 1996 Pigottamp Milner 1993) However all of these previousresults were obtained on patients who had unilat-eral lesions GR has a bilateral occipital-temporallesion including clear damage to parahippocampalcortex in both hemispheres furthermore he reportssome kind of perceptual deficit We hypothesisedthat this deficit might involve difficulty perceivingor interpreting the geometry of scenes and otherstimuli with a similar geometric structure
EPSTEIN ET AL
494 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 5 Top row shows a single slice from a functional MRIscan performed on GR Bottom row shows results from a normalsubject (NK) for comparison Voxels that responded more to facesthan objects (left column) or more to scenes than faces and objects(right column) in a Kolomogorov-Smirnov test are highlighted bywhite arrows GR has a functioning face area (FFA) but there isno evidence of a place area (PPA)
FFA PPA
GR
NK
In this experiment subjects viewed either objectshapes or spatial layouts Stimuli consisted ofblack-and-white photographs of real-world stimuli(empty rooms vs common objects) or abstractstimuli made out of Lego blocks (Lego ldquoscenesrdquo vsLego ldquoobjectsrdquo) These stimuli have been previouslyshown to differentially activate the PPA in normalsubjects (Epstein et al 1999 Epstein ampKanwisher 1998) Two red dots were superim-posed on each photograph (Figure 6) The task wasto report which of the two red dots was over thecloser part of the object or scene This task wasdesigned to have no memory component (apartfrom remembering the instructions) but to requireaccurate 3D perception of objects and scenes
If GR or CO have difficulty perceiving or inter-preting the spatial layout of scenes then we pre-dicted they would do worse when the stimuli areempty rooms or Lego scenes than when they arereal-world or Lego objects Note that the criticalcomparison is between the Lego scenes and theLego objects as these stimuli are made of the samematerials and differ primarily in their surfacegeometry
MethodTesting for all experiments was done on aMacintosh computer The experiment consisted of80 trials (20 of each stimulus type randomly inter-mixed) Subjects began each trial by pressing thespace bar A black-and-white photograph from oneof the four stimulus categories (see earlier) thenappeared on the screen with two red dots on it Thetask was to determine which of the two red dotsindicated the closer part of the scene or objectSubjects pressed the ldquofrdquo key if the leftmost red dotwas closest and the ldquojrdquo key if the rightmost dotwas closest They were instructed to respond asquickly as possible without making errors Stimuliremained on the screen until a response was made
The stimulus set was constructed so that eachempty room was matched to a common object thathad red dots in the exact same location and eachLego layout was similarly matched to a Lego objectThus the locations of the red dots were counterbal-anced across scene and object stimuli controllingfor any effects of visual field cuts
Results and discussionResults for GR and CO and four elderly normalsubjects are shown in Table 3 Both GR and COcould do this task without difficulty and their per-formance was near ceiling There was no evidencethat they had any more trouble with the emptyrooms or Lego scenes than with the real or Legoobjects Reaction times were slightly longer for GRand CO than for normal subjects as one wouldexpect given their visual field deficits however this
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 495
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 3 Experiment 1 (two-dot) results (SD in parenthesis)
GR CO Normals
Proportion correctObject 095 080 093Lego object 100 095 098Empty room 100 100 099Lego room 095 100 096
Mean RTObject 2435 2932 2161 (980)Lego object 2052 3245 2029 (921)Empty room 2744 2291 1788 (959)Lego room 1904 2281 1630 (1127)
Figure 6 The four kinds of stimuli used in Experiment 1 Topleft lego layouts top right real-world layouts (empty rooms)bottom left lego objects bottom right real-world objects
difference did not appear to be significant Impor-tantly there was little evidence that GR or CO tooklonger to make their perceptual judgements for thereal or Lego scenes than for the real or Lego objectsThus contrary to our initial hypothesis this experi-ment found no clear evidence for selective impair-ment in the perception of scene-like stimuli
Experiment 2 Different-views matchingtask
The previous experiment demonstrated that GRand CO can extract information about the per-ceived distances between themselves and the sur-faces of an object or scene Thus in at least onesense their perception of these surfaces is pre-served However this does not mean that their abil-ity to represent the layout of a scene is unimpairedThe two-dot task requires on-line comparison oftwo surfaces in egocentric coordinates GR and COwould be able to perform this task even if they couldnot (1) represent more than two surfaces at a time(2) represent surfaces in other than strictly egocen-tric coordinates or (3) sustain surface representa-tions for longer than the time it takes to make acomparison It has been suggested that the parietallobes are involved in on-line processing of surfacegeometry in egocentric coordinates (Milner ampGoodale 1995) thus it is possible that the parietallobes (which are intact in GR and CO) may be suf-ficient to perform the two-dot task even in theabsence of more durable or complex surface repre-sentations in parahippocampal cortex
In Experiment 2 we examine GR and COrsquosability to create and sustain a representation of thegeometry of an object or scene-like layout The taskwas to view a photograph of a Lego object or Legoscene hold it in memory for 10 s and then deter-mine whether a second object or scene was the sameas the first In ldquosamerdquo trials the second photographdepicted the same scene or object depicted from adifferent viewing angle In ldquodifferentrdquo trials the sec-ond photograph depicted a different scene or objectchosen to be similar to the first one (Figure 7)Thus in order to successfully perform this tasksubjects had to form a representation of the surfacesof the object or scene sustain this representation in
memory for 10 s and then align it (presumablyusing mental rotation) to the second object orscene
MethodThe experiment consisted of two parts each with40 trials In the first part the stimuli were black andwhite photographs of objects made out of Legoblocks In the second part the stimuli were photo-graphs of ldquoscenesrdquo made out of Lego blocks whichwere designed to have a geometric structure similarto real-world places such as a roomor a city street
Subjects began each trial by pressing the spacebar A photograph of a Lego object or sceneappeared on the screen for 3 s followed by a 10-sblank interval A second photograph then appeareddepicting either (1) the same object or scene as thefirst photograph shown from a different viewingangle (offset approximately 35 degrees) or (2) a dif-ferent object or scene Subjects were instructed topress the ldquojrdquo key if the two photographs depicted thesame object or scene and the ldquof rdquo key if they did notThey were told to take as much time as they neededto make a decision but no extra time The secondphotograph remained on the screen until a responsewas made Half the trials were ldquosamerdquo trials and halfwere ldquodifferentrdquo trials
Results and discussionFigure 8 shows results for GR and CO as well as forsix elderly normal controls GRrsquos and COrsquos perfor-mance was comparable to that of the normal con-trols Furthermore GR and CO performed just aswell when the stimuli were Lego scenes as whenthey were Lego objects These results are unlikely toreflect a ceiling effect as performance for bothpatients and controls was around 80 correctThus both patients appear to be able to form a rep-resentation of the geometry of both scenes andobjects which they can sustain in short-termmemory
Experiment 3 N-back recognition memorytask
The previous experiment failed to find any impair-ment in GR and COrsquos ability to form a representa-
EPSTEIN ET AL
496 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
tion of the geometry of a scene or object andmaintain it for 10 s in the absence of the originalstimulus Not only does this result suggest that per-ception of scene-like stimuli is intact in thesepatients it further suggests that some short-termmemory functions are intact as well However we
should note that subjects in this experiment werefree to devote all their effort and attention in the 10-second blank interval to maintaining a memory ofthe stimulus Thus it is possible that GR and COcan use rehearsal mechanisms to maintain the stim-ulus in short-term memory during the blank inter-
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 497
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
(b) Lego object version
Figure 7 Stimuli and procedure for Experiment 2 (a) In the Lego layout version subjects see a Lego layout for 3 s followed by a blankscreen for 10 s A second layout then appears which is either the same layout as the first one but seen from a different angle or a differentlayout (b) The Lego object version is the same but with object stimuli
3 s 10 s OR
3 s 10 s OR
(a) Lego layout version
(b) Lego object version
val but are unable to form a more durable memorytrace for the stimulus This possibility was tested inthe current experiment which was designed toexamine GR and COrsquos ability to learn new visualinformation
We devised a continuous n-back recognitionmemory test in which subjects viewed a sequence oforiginally novel stimuli some of which wererepeated within the sequence at intervals of 1 3 or5 intervening images For each stimulus subjects
reported whether they had seen it before in theexperiment or not In different runs in the experi-ment the stimuli were black-and-white photo-graphs of (1) Lego scenes (2) Lego objects (3) real-world landscapes (4) novel objects made out ofSculpey clay (5) unfamiliar houses (6) unfamiliarfaces as well as (7) line drawings of nonsenseobjects and (8) pronounceable pseudowords (Fig-ure 9) Thus we were able to test GR and COrsquos rec-ognition memory for different kinds of materials atdifferent time intervals in a paradigm in which thepresence of intervening items between the twoappearances of each stimulus made rehearsal diffi-cult Note that because none of the specific stimuliused in this experiment were familiar in advancethis task did not require an episodic memory com-ponent and could be done on the basis of pure visualfamiliarity with the particular visual stimulus
Although many different stimulus classes weretested in this experiment the critical comparison isbetween the Lego layouts and Lego objectsbecause these are best matched with regard tomaterials and differ primarily in their geometricstructure (see Introduction) We predicted that ifGR and CO showed a memory deficit for this taskit would be greater for the Lego layouts than for the
EPSTEIN ET AL
498 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 8 Results of Experiment 2 Performance over ldquosamerdquotrials and ldquodifferentrdquo trials is averaged together Error bars show1 SD
Figure 9 Stimuli for Experiment 3 Top row Lego layouts real-world scenes houses Bottom row Lego objects clay objects nonsenseobjects faces In an eighth condition (not shown) subjects viewed pronounceable nonwords such as ldquowilchrdquo or ldquothipperrdquo
Lego objects In addition a deficit for materialsthat convey information about the layout of sur-rounding space might also be apparent when GRand CO are tested on more realistic stimuli If soone would expect them to do poorly on the land-scapes but relatively well on the clay objects linedrawings of nonsense objects faces andpseudowords
MethodFor each subject a complete experimental sessionconsisted of being run once on each of the eightstimulus classes Each run consisted of 90 trials ineach trial a stimulus was presented and subjectsreported whether or not they had seen it before inthe run (or indeed ever) by pressing the ldquojrdquo key ifthey had or the ldquof rdquo key if they had not Each stimu-lus was on the screen for either 1 2 or 3 s (see fol-lowing) and appeared promptly after the keypressreport for the previous trial Individual stimuliappeared either once (14 stimuli) twice (30 stim-uli) or four times (4 stimuli) within a given run Ofthe stimuli that were presented twice 10 wererepeated immediately (1-back) 10 were repeatedwith a lag of two intervening items (3-back) and 10were repeated with a lag of four intervening items(5-back) In order to ensure that between-subjectdifferences in performance were not due to stimulusdifferences the same stimuli were assigned to thesame conditions each time the experimentwas run
GR performed the experiment twice once inAugust 1998 and again 9 months later Because ofa computer error he was only tested on six of theeight categories (all except Lego layouts and Legoobjects) at the later date CO was also tested twiceonce in February 1999 and again 6 months laterReported results for GR and CO are the average oftheir two testing sessions Normal subjects wereonly tested once Stimulus presentation time was 1 sin GRrsquos first testing session In pilot testing COfound this too fast for the critical categories of Lego
layouts and Lego objects so presentation time wasextended to 2 s for these categories in his first test-ing session All normal subjects were tested at thesepresentation rates (2 s for Lego layouts and objects1 s for other stimuli) In COrsquos second testingsession presentation time was lengthened to 3 s forall stimulus categories to ensure that any deficitsfound were memory deficits rather than problemswith the nonmemory aspects of the task
Results and discussionResults from GR CO and 10 elderly normal con-trols are shown in Figure 10 and Table 4 Figure 10ashows results from the main comparison betweenLego layouts and Lego objects Both normal sub-jects and patients performed near ceiling for bothLego layouts and Lego objects at 1-backinterstimulus intervals verifying that GR and COcould successfully perceive the pictures as they werebeing presented In contrast there was a strikingdifference between the two patients and the normalsubjects at 5-back intervals Whereas normals didon average somewhat better on Lego layouts thanLego objects both GR and CO showed the oppo-site pattern doing much worse on Lego layoutsthan Lego objects The 3-back condition showed amixed pattern Whereas GR did better on Legoobjects than Lego layouts in this condition (similarto his performance at 5-back intervals) CO didbetter on Lego layouts than Lego objects COrsquos lowperformance on Lego objects in the 3-back condi-tion appears to be an anomaly as it is much lowerthan his performance for comparable stimuli in the1-back and 5-back conditions
Figure 10b shows the difference in performancebetween Lego layouts and Lego objects for thecritical 1-back and 5-back intervals At 5-backintervals CO was significantly impaired comparedto the normal subjects (z = ndash250) whereasGRrsquos impairment fell just short of significance (z =ndash175)4
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 499
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
4In fact these values may underestimate GR and COrsquos relative impairment for Lego layouts In the 5-back condition 4 of the 10
normal subjects had perfect performance on the Lego layouts but all scored below ceiling on the Lego objects Thus the true advantagefor Lego layouts over Lego objects for normal subjects is probably greater than that observed here in which case GR and COrsquos lowerperformance for Lego layouts compared to Lego objects would be even more abnormal Furthermore post-hoc analyses revealed thatone of the normal subjects showed a difference in performance between Lego layouts and objects that was over 2 SD below the mean ofthe other nine subjects When this outlying subject was excluded z-scores were ndash322 for CO and ndash232 for GR
Table 4 shows results for all eight stimulusclasses Although direct comparisons between sub-jects should be treated with caution since viewingtime for different items was not the same for eachsubject we do note a few interesting patterns Firstof all there is little or no evidence that GR or CO
show a memory deficit for nonsense words clayobjects or line drawings of nonsense objects GRappears to be slightly impaired on faces and housesbut CO performs normally on these categoriesStrikingly GR shows a severe deficit for the reallandscapes performing more than 40 percentagepoints lower than normals Thus even thoughthere is some evidence that GR might have someimpairment for nonscene stimuli he clearly is muchmore impaired for scene-like stimuli such as theLego scenes and landscapes CO does not show asimilar deficit for the landscapes However itshould be kept in mind that in one of his two testingsessions CO viewed each picture for 3 s which wasmuch longer than the 1 second used for all non-Lego stimuli for GR and normals Thus COrsquosapparently ldquonormalrdquo performance in these condi-tions might be overestimated
Taken together the results of this experimentindicate that GR and CO are selectively impaired atprocessing scene-like stimuli This deficit is consis-tently apparent at 5-back but not 1-back intervalsThis finding suggests that the deficit should becharacterised as a memory encoding problemHowever the present results do not allow us todetermine whether the difficulty occurs at memoryencoding or at subsequent recognition (see Epsteinamp Kanwisher 2001) If GR and CO have a generalproblem with recognition of topographical materi-als then they should not be able to recognise scenes
EPSTEIN ET AL
500 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 10a Results of Experiment 3 for GR (top) CO (middle)and for 10 age-matched normal controls Performance measure isproportion of repeated stimuli correctly identified as such correctedfor guessing using the standard formula
Figure 10b Difference between Lego layout performance andLego object performance for the 1-back and 5-back conditionsError bars indicate one 1 SD
and landmarks that were learned prior to theirinjury We tested this possibility in the nextexperiment
Experiment 4 Famous landmarksfacesrecognition task
In this experiment we examined GR and COrsquosability to recognise the visual appearances of placeslearned prior to injury Subjects saw a series offamous landmarks intermixed with nonfamouscontrols (Imaging experiments have demonstratedthat landmarks of this type can activate the PPAalmost as strongly as complete scenes Epstein etal 1999 Epstein amp Kanwisher 1998) They wererequired to say whether they knew each specificlandmark and to identify the ones they knew Sub-jects were instructed to respond affirmatively if theywere fairly certain that they recognised the land-mark but not if it only looked generally familiarFor purposes of comparison subjects also per-formed an identical test in which the stimuli werefamous and nonfamous faces In both testsdistractor items were designed to be visually andconceptually similar to the target items (see Figure11) If parahippocampal cortex is required for placerecognition then one might expect GR and CO toperform poorly compared to normals on the land-mark recognition test
MethodEach of the two experiments (famous faces famouslandmarks) consisted of 48 trials Subjects beganeach trial by pressing the space bar after which ablack-and-white photograph of a face (in thefamous faces experiment) or a landmark (in thefamous landmarks experiment) appeared on thescreen Subjects were told to press the ldquojrdquo key if theyrecognised the specific face or landmark and the ldquof rdquokey if they did not taking as much time at theyneeded to make this discrimination but no extratime Subjects were told to report that they recog-nised a face or landmark if they felt that they knewit even if they could not retrieve its name If theydid know the name they were to say it after pressingthe button Otherwise they were to say whateverthey did remember about the item
Half of the photographs were of famous and halfof nonfamous people and landmarks The famousfaces included public figures (eg George BushRichard Nixon Prince Charles) and movie stars(eg Harrison Ford Clark Gable Marlon Brando)who were prominent prior to the time of GR andCOrsquos injuries the famous landmarks were build-ings and monuments from all over the world (egEiffel Tower US Capitol Building Great Sphinx)The nonfamous stimuli were chosen to match thefamous stimuli as closely as possible Each famousstimulus had a ldquomatchrdquo For example George Bushwas matched to a man of similar age in a coat andtie and the Taj Mahal was matched to a visually
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 501
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 4 Full results of Experiment 3 Mean proportion correct (corrected for guessing) SDs in parentheses
Lego Land- Lego Clay Nonsense Non-scenes scapes Houses objects objects objects Words Faces
1-back GR 100 094 100 100 100 094 100 100CO 091 085 091 100 100 079 074 100Normals 096(008) 095(008) 093(014) 096(008) 096(006) 099(003) 098(004) 097(011)
3-back GR ndash003 068 026 047 083 083 075 052CO 066 081 037 010 094 069 095 081Normals 066(032) 077(021) 072(015) 077(020) 086(016) 083(015) 083(017) 072(020)
5-back GR 010 034 044 047 072 083 094 053CO 031 073 075 088 081 064 100 081Normals 076(026) 077(016) 068(021) 066(016) 091(014) 083(018) 093(011) 078(021)
similar Asian palace that is not famous (see Figure11)
Results and discussionResults for GR and CO as well as for five age-matched controls are shown in Figure 12 Both
GR and CO could recognise the famous landmarksas well normals In addition both GR and CO wereusually able to name the landmarks they success-fully identified
Thus GR and CO can recognize landmarkslearned prior to injury demonstrating that at leastsome of their place recognition abilities are intactWhen combined with the fact that GR and COwere able to produce detailed maps of places theylived in prior to their injury (see Case Description)these results suggest that their deficit is primarily inencoding or consolidation of new informationrather than in recognition or recall of old informa-tion Note however that these results do not provethat their place recognition abilities are entirelynormal GR and CO might suffer from a mild rec-ognition deficit that affects identification of newly-learned stimuli like the Lego layouts in Experiment3 but not overlearned stimuli like the famous land-marks Furthermore it is possible that GR and COmight be impaired at recognising premorbidlylearned places that are defined primarily by theirspatial layout even though they are unimpaired atrecognising premorbidly learned places that aredefined primarily by a single object-like landmarkor building (see General Discussion) Neverthelessthe trend of the present evidence is to suggest thatGR and CO have particular problems with noveltopographical information
In the famous-face task GRrsquos performance wasin the normal range However he sometimes haddifficulty naming the faces even though he clearly
EPSTEIN ET AL
502 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 11 Stimuli for Experiment 4 Each famous landmark(Arc de Triomphe Taj Mahal) and famous face (Barbara BushYasser Arafat) was matched to a nonfamous building or face thatalso appeared in the experiment
FA
CE
SL
AN
DM
AR
KS
Figure 12 Results of Experiment 4 Performance measure isproportion of famous stimuli reported as ldquoknownrdquo corrected forguessing using the standard formula
recognised them For example he described Ron-ald Reagan as ldquothe gipperrdquo Nancy Reagan as ldquothewife of the gipperrdquo and Clark Gable as ldquothe movieactor who said lsquoScarlett I donrsquot give a damnrsquordquo COhad a great deal of difficulty identifying the famousfaces This result is not surprising given hisextremely low performance on the Benton FaceMatching Task In addition to his topographicproblems CO appears to suffer fromprosopagnosia
GENERAL DISCUSSION
We tested two parahippocampal patients (GR andCO) on four experiments designed to measure per-formance as a function of both task (perceptionencoding and recognition) and stimulus type(scenes objects faces and words) The only taskthat revealed a clear deficit was an n-back recogni-tion memory test (Experiment 3) In order to suc-cessfully perform this task subjects had to encode adurable representation of a visual stimulus and useit for later recognition GR and CO were signifi-cantly more impaired on this task when the stimuliwere scene-like spatial layouts (ie Lego scenesand real scenes for GR) than when they wereobjects No comparable deficits were found in otherexperiments that tested layout perception (Experi-ments 1 and 2) and placelandmark recognition(Experiment 4) Taken together these results indi-cate that GR and CO are impaired in their ability toencode novel information about the geometry ofsurrounding space into memory but are lessimpaired at their ability to encode the geometry ofnovel objects
We argue that this specific topographical defi-citmdashinability to encode the spatial layout of novelscenesmdashresults from damage to the para-hippocampal place area Although the damage inboth patients extends beyond the parahippo-campal-lingual territory normally associated withthe PPA this region is clearly lesioned bilaterally inGR and in the right hemisphere in CO Further-more functional imaging demonstrated that GRhas no functioning PPA Thus the current resultsand the earlier fMRI work (Epstein et al 1999
Epstein amp Kanwisher 1998) converge to implicateparahippocampal cortex in the encoding of infor-mation about scene geometry into memory
Our assignment of the function of memoryencoding to parahippocampal cortex is consistentwith the results of earlier case reports of patientswith parahippocampal damage who have naviga-tional difficulties in novel but not familiar environ-ments (Habib amp Sirigu 1987 Ross 1980 see alsoTeng amp Squire 1999) As with GR and CO thesepatients were either less impaired or unimpaired onnontopographical memory tasks Patients have alsobeen described who exhibit the opposite pattern ofimpairment an inability to learn faces (sometimescalled ldquoprosopamnesiardquo Cipolotti Robinson Blairamp Frith 1999 Tippett Miller amp Farah 2000) orfaces and verbal materials (Maguire amp Cipolotti1998) with an unimpaired ability to learn topo-graphical materials Taken together these resultsargue for a high degree of domain-specificity in theneural systems involved in learning of new informa-tion (Gallistel 1995) The brain may have separatemechanisms for learning faces places and wordswhich can be differentially impaired after damageto specific cortical regions (Cipolotti et al 1999Incisa della Rochetta et al 1996) The presentresults suggest that the anatomical locus of themechanism dedicated to learning new topographi-cal information is in the parahippocampal cortex(Aguirre amp DrsquoEsposito 1999)
In contrast to their impaired ability to encodenew topographical information both GR and COwere able to produce detailed maps of their previousdwellings and performed normally on the famouslandmark recognition test of Experiment 4 Theseresults are generally consistent with Teng andSquirersquos (1999) report of a patient with extensivemedial temporal damage who could recallpremorbidly learned but not postmorbidly learnedtopographical information (see also Ross 1980)Thus our results provide further evidence thatmedial temporal regions such as parahippocampalcortex may be more critical for encoding or consoli-dating new information into memory than for recallof old information
One issue that our experiments do not entirelyresolve is the precise nature of the topographical
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 503
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
information encoded by parahippocampal cortexClearly this region is critical for encoding informa-tion about the geometry of the currently visiblescene even when the ldquoscenerdquo is made out of Legoblocks and therefore has no immediate navigationalrelevance However it is unclear whether this spe-cific deficit can account for all of the real-worldnavigational difficulties encountered by GR andCO GR in particular shows such a catastrophicloss of navigational ability that we suspect he is alsoimpaired at encoding the spatial relationshipsbetweenthe current scene and other locations in theworld CO probably also suffers from such difficul-ties as he reports that he has trouble rememberinghow places relate to each other in large-scale spaceWhen he was tested on a simple 3-room videogameenvironment he not only was unable to rememberthe individual rooms (claiming that there were 10rooms when there were only 3) but he also had nomemory for the spatial relationships between themThese spatial deficits may be the result of damage tothe posterior right hippocampus (in the case of CO)or of disconnection of the posterior right hippo-campus from the parahippocampal regions thatprovide much of its cortical input (in both patientsSuzuki amp Amaral 1994) In either case the abilityto form a cognitive map of new environments likethe lab or videogame space could be impaired(Maguire et al 1998 OrsquoKeefe amp Nadel 1978)However it is also possible that parahippocampalcortex itself may be critical for encoding not onlythe geometry of individual scenes but also the spa-tial relationships between the locations of thosescenes In this case the spatial deficits of GR andCO could be attributed directly to parahippo-campal damage rather than to hippocampal dam-age or disconnection
The latter scenario would be consistent withAguirre and DrsquoEspositorsquos (1999) and Barrash etalrsquos (2000) suggestion that parahippocampal cortexis critically involved in combining visual informa-tion about the appearance of scenes and landmarkswith spatial information about their locations into aunified topographical representation Howeverboth of these authors argue that the representationsof individual scenes (Barrash et al) and landmarks(Aguirre amp DrsquoEsposito) are supported not by
parahippocampal cortex but by more posterior lin-gual-fusiform regions In contrast we believe thatthe neuroimaging data indicate that parahippo-campal cortex is involved in the encoding of thegeometric structure of individual scenes althoughit may also play a role in combining these represen-tations with other spatial representations
Interestingly GR and CO were not impaired onrecognition of famous landmarks that were mostlybuildings Thus our data do not preclude the possi-bility that there may be a separate lingual orfusiform region involved in processing informationabout buildings (Aguirre Zarahn amp DrsquoEsposito1998 Chao Haxby amp Martin 1999 IshaiUngerleider Martin Schouten amp Haxby 1999)which might have been preserved in GR and COdespite extensive damage to other regions of thelingual and fusiform gyrus Alternatively GR andCOrsquos preserved building recognition abilities mightbe mediated by interactions between cortical areasthat perform shape analysis for any object-likestimulus type (Malach et al 1995Kanwisher et al1996) and anterior temporal regions in which long-term topographical and semantic memories arestored (McCarthy et al 1996)
Is perception preserved
In contrast to their inability to encode new scenesinto memory GR and CO have substantiallyretained their ability to perceive the spatial layout ofthe currently visible scene (Experiments 1 and 2)GR can follow a route marked on a floor planwhich indicates a good on-line appreciation of hiscurrent spatial surroundings Furthermore neitherGR or CO have any trouble moving through theenvironmentmdashthey move with confidence and donot bump into doors or walls Problems of the lattertype are usually associated with damage to the pari-etal lobes which are intact in GR and CO Thus atfirst glance GR and CO do not appear to haveperceptual impairments (beyond their visual fieldrestrictions)
However GR does complain of a perceptualdeficit which he describes as an inability to ldquotakeinrdquo scenes when they are too complex Similarreports of an inability to comprehend the
EPSTEIN ET AL
504 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
ldquoglobalityrdquo of a scene were made by some of theparahippocampally-damaged patients studied byHabib and Sirigu (1987) We were not able todetermine the precise nature of this deficit whichmay be partly attributable to his visual field restric-tions It appears that his deficit is more salient in thereal-world environments than in the picture-per-ception tasks of Experiments 1 and 2 perhapsbecause he finds it more difficult to organise visualinput when immersed in it than when viewing pho-tographs that subtend only part of the visual field
We suggest that GR can perceive parts of thescene when he attends to them but may be unableto integrate information over time into a coherentrepresentation of a scene or place The fact that GRand CO are impaired for Lego layouts at 5-backintervals in Experiment 3 is consistent with thishypothesis The time between the first and secondpresentations of a stimulus is very short in this case(and even shorter with 3-back intervals where GRalso shows evidence of a deficit) In both patientsthe representation of layout information formed byperception of a Lego scene must either have anabnormally short persistence be abnormally sus-ceptible to interference or both If so they mightfind it impossible to sustain a spatial frame orschema long enough to use it to organise informa-tion obtained from many different glances at a com-plex scene (Intraub 1997) The result would be aninability to understand the ldquoglobalityrdquo of a sceneeven though all the details were perfectlyperceived
Non-topographical mnemonic abilities
Although not as severe as their topographical mem-ory problems GR and CO do have some memorydifficulties in other domains For example bothperformed poorly on the Rey AVLT whichrequired them to remember a list of unrelatedwords Both also report some subtle episodic mem-ory problems (see Case Description) COrsquos prob-lems could partly result from damage to the righthippocampusmdashindeed CO appeared to have themore acute episodic memory problems HoweverGR has no hippocampal damage so the fact that hedisplays episodic memory problems suggests thatparahippocampal cortex is involved in the forma-
tion of episodic memories (cf Wagner et al1998)
In previous work (Epstein et al 1999) weargued that spatial context information encoded byparahippocampal cortex might play a role in estab-lishing event memories by encoding informationabout where events take place The inability toencode this information might cause problems intasks such as the Rey AVLT where one has to recalla list of unrelated words If spatial (and perhapstemporal) context information is not encoded itwill be harder subsequently to recall the distinctevent of hearing each word (Tulving 1983) Incontrast lack of spatial context information wouldbe less of a problem in tasks where one is onlyrequired to recognise previously encountered verbalmaterials but not to reproduce the spatiotemporalcontext in which they were experienced IndeedGR and CO performed as well as normals on thenonsense words in Experiment 4 where recogni-tion memory but not episodic memory was requiredfor successful performance Interestingly GR andCO also did relatively well on the Logical Memorysubtest of the WMS-III perhaps because the nar-rative framework of a story provides sufficient con-textual information to overcome the absence ofspatial context and mediate successful verbalretrieval
Differences between GR and CO
Although we have discussed GR and CO togetherthere are a number of relevant differences betweenthem One open question is whether these differ-ences can be attributed to the different effects ofunilateral and bilateral parahippocampal damageIn general GRrsquos topographical difficulties weremore severe than COrsquos CO did surprisingly well onthe Warrington Topographical Memory Test inwhich he was required to remember detailed real-world city scenes and also performed substantiallybetter on real scenes than Lego scenes in Experi-ment 3 Insofar as CO has intact left para-hippocampal cortex this suggests that the left PPAmight be sufficient to encode a certain amount of(perhaps semantic) information about real-worldscenes but the right PPA is necessary to encode
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 505
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)

the deficient range (lt 5th percentile SS lt 75) forfaces The low score for faces on this test as well asthe WMS-III may reflect prosopagnosia ratherthan a memory problem as he scored in the defi-cient range (lt1st percentile SSlt62) on the BentonFace Recognition test Verbal list learning (ReyAVLT) was average on immediate recall (45th per-centile SS =98) but after hearing an interferencelist performance fell to the deficient level (015words recalled) for both immediate and delayedrecall On a recognition test for these words herecognised 1115 and made one false alarm (39thpercentile SS =96) There was a large discrepancyon tests of memory span which may indicate adecline in spatial memory digit span was superior(9+ forward and 7 backwards both 99th percentile)but spatial span was only average (5 forward and 5backward 37th and 50th percentiles respectively)Language abilities were intact (Boston NamingTest performance was at ceiling) Thus like GRCO shows some impairment at recall of visualinformation (WMS-III Face Memory subtest) andverbal information that is not organised into a nar-rative (Rey AVLT) and clear impairment at tasksthat require recall of stimulus location (Family Pic-tures subtests of WMS-III) In contrast his abilityto recall organised verbal information (WMS-RLogical Memory subtest) is more preserved
MRI RESULTS
In order to determine the extent of their lesionsstructural MRI images were taken of both subjectsDelineation of the lesions was based either onspoiled GRASS coronal images at 11mm thick-ness (GR) or anatomical T-1 weighted MPRAGEMRI sequences yielding 125 mm thick coronalimages (CO) The anterior-posterior extent of thelesions was verified on axial images
GR has bilateral occipital-temporal lesions(Figure 4) In the left hemisphere the posteriorparahippocampal gyrus (including para-hippocampal cortex) is severely damaged with onlyminimal injury to the most caudal aspect of the hip-pocampus There is also moderate damage to themedial fusiform and inferior lingual gyri In the
right hemisphere the hippocampus is spared butthere is damage to the posterior end of theparahippocampal gyrus near the parahippocampal-lingual boundary More posteriorly the lesionedarea extends along the collateral sulcus to includesignificant portions of the inferior lingual gyrus andthe medial fusiform gyrus as well as most of themedial occipital lobe below the calcarine fissure
CO has a right-hemisphere occipital-temporallesion Right posterior hippocampus and para-hippocampal gyrus are severely affected Moreposteriorly the damage covers large portions of theinferior lingual and medial fusiform gyri as well assubstantial portions of the medial occipital lobeincluding most of right calcarine cortex In addi-tion diffuse foci of increased signal on T-2weighted images was present in periventricularwhite matter in the corona radiata and centrumsemiovale with a lacunar infarct of the rightputamen There was also moderate degree of dif-fuse involutional changes with increased ventricularsize particularly notable in the right temporal hornwith ex vacuo dilitation
The structural MRIs suggested that the PPAwas severely if not completely damaged in bothhemispheres for GR and in the right hemispherefor CO In order to confirm this for GR we used ahigh-field 3-T magnet to perform a functional scanin which he viewed scenes faces and objects in thesame blocked fMRI design that we have used tolocalise the PPA in all our previous experiments(see Epstein et al 1999 for details Epstein ampKanwisher 1998) A Kolomogorov-Smirnov testfound no voxels in the parahippocampal region thatfit our previously established criterion for inclusionin the PPA (ie greater response to scenes than tofaces and objects at a uncorrected significance levelof plt 0001) This result contrasts sharply with thepattern seen in over 95 of (mostly young or mid-dle-aged) normal subjects in whom a large acti-vated region can be identified in both left and rightparahippocampal cortex using this comparison (seeFigure 5) A similar test was used to identify voxelsthat responded significantly more to faces than toobjects This comparison revealed a region of face-selective voxels in the right fusiform gyrus consis-tent with the location of the Fusiform Face Area
EPSTEIN ET AL
492 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 493
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
GR
CO
Figure 4 Structural MRIs of GR and CO showing the extent of the damaged region (marked as white) Images have been normalisedusing SPM99 and a cost function masking algorithm (provided by Matthew Brett) into standard stereotactic space (Talairach amp Tournoux1988) using the Montreal Neurological Institute template (Evans et al 1993) Numbers indicate y-Talairach coordinate for each coronalslice corresponding slice planes are also indicated on the sagittal image
(FFA) commonly seen in normal subjects(Kanwisher McDermott amp Chun 1997) Thusthe MRI was sensitive enough to detect functionaldifferences an FFA was found but no PPA Whencombined with the anatomical results this stronglysuggests that GR has no PPA
Because of safety concerns (presence of a femo-ral artery aneurysm clip) we did not perform a simi-lar high-field functional MRI of CO However asFigure 4 demonstrates CO clearly has damage tothe parahippocampal-lingual region in the righthemisphere
EXPERIMENTAL INVESTIGATIONS
In this section we report the results of four experi-ments designed to probe the consequences of dam-age to the lingual-parahippocampal region ingeneral and the PPA in particular Our primaryconcern in these experiments was to test GR andCOrsquos ability to perceive remember and recognisedifferent kinds of visual material Based on our pre-vious fMRI work we predicted that GR and CO
would have more difficulty with scene-like stimuli(ie stimuli that convey information about thegeometry of a space one can navigate through or actwithin) than with object-like stimuli Such a deficitcould account for at least some of the problems GRand CO encounter in real-world navigation
Beyond determining the kind of visual materialthat gets processed in parahippocampal cortex wealso wanted to determine the nature of the process-ing that takes place on that material In particularwe wanted to determine if parahippocampal cortexis involved in perceptual encoding memory encodingor recognition As discussed earlier (Introduction)our fMRI results provided some evidence againstplace recognition as the sole function of the PPAbut did not allow us to distinguish between percep-tual and mnemonic encoding Experiments 1 and 2examine the role of parahippocampal cortex in per-ceptual encoding Experiment 3 examines its role inmnemonic encoding and Experiment 4 examinesits role in place recognition
Experiment 1 Two-dot task
The first hypothesis we examined was thatparahippocampal cortex plays a role in perceivingthe shape of the local environment In our previousfMRI work we observed that the PPA respondsmuch more strongly during scene viewing than dur-ing object or face viewing even when subjects donothing more than watch the scenes passively with-out performing any other task This suggested thatscene perception alone may be sufficient to activatethe PPA Previous reports have indicated that dam-age to parahippocampal cortex and other medialtemporal regions leads to problems with memoryrather than perception (Bohbot et al 1998Maguire Burke Philips amp Staunton 1996 Pigottamp Milner 1993) However all of these previousresults were obtained on patients who had unilat-eral lesions GR has a bilateral occipital-temporallesion including clear damage to parahippocampalcortex in both hemispheres furthermore he reportssome kind of perceptual deficit We hypothesisedthat this deficit might involve difficulty perceivingor interpreting the geometry of scenes and otherstimuli with a similar geometric structure
EPSTEIN ET AL
494 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 5 Top row shows a single slice from a functional MRIscan performed on GR Bottom row shows results from a normalsubject (NK) for comparison Voxels that responded more to facesthan objects (left column) or more to scenes than faces and objects(right column) in a Kolomogorov-Smirnov test are highlighted bywhite arrows GR has a functioning face area (FFA) but there isno evidence of a place area (PPA)
FFA PPA
GR
NK
In this experiment subjects viewed either objectshapes or spatial layouts Stimuli consisted ofblack-and-white photographs of real-world stimuli(empty rooms vs common objects) or abstractstimuli made out of Lego blocks (Lego ldquoscenesrdquo vsLego ldquoobjectsrdquo) These stimuli have been previouslyshown to differentially activate the PPA in normalsubjects (Epstein et al 1999 Epstein ampKanwisher 1998) Two red dots were superim-posed on each photograph (Figure 6) The task wasto report which of the two red dots was over thecloser part of the object or scene This task wasdesigned to have no memory component (apartfrom remembering the instructions) but to requireaccurate 3D perception of objects and scenes
If GR or CO have difficulty perceiving or inter-preting the spatial layout of scenes then we pre-dicted they would do worse when the stimuli areempty rooms or Lego scenes than when they arereal-world or Lego objects Note that the criticalcomparison is between the Lego scenes and theLego objects as these stimuli are made of the samematerials and differ primarily in their surfacegeometry
MethodTesting for all experiments was done on aMacintosh computer The experiment consisted of80 trials (20 of each stimulus type randomly inter-mixed) Subjects began each trial by pressing thespace bar A black-and-white photograph from oneof the four stimulus categories (see earlier) thenappeared on the screen with two red dots on it Thetask was to determine which of the two red dotsindicated the closer part of the scene or objectSubjects pressed the ldquofrdquo key if the leftmost red dotwas closest and the ldquojrdquo key if the rightmost dotwas closest They were instructed to respond asquickly as possible without making errors Stimuliremained on the screen until a response was made
The stimulus set was constructed so that eachempty room was matched to a common object thathad red dots in the exact same location and eachLego layout was similarly matched to a Lego objectThus the locations of the red dots were counterbal-anced across scene and object stimuli controllingfor any effects of visual field cuts
Results and discussionResults for GR and CO and four elderly normalsubjects are shown in Table 3 Both GR and COcould do this task without difficulty and their per-formance was near ceiling There was no evidencethat they had any more trouble with the emptyrooms or Lego scenes than with the real or Legoobjects Reaction times were slightly longer for GRand CO than for normal subjects as one wouldexpect given their visual field deficits however this
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 495
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 3 Experiment 1 (two-dot) results (SD in parenthesis)
GR CO Normals
Proportion correctObject 095 080 093Lego object 100 095 098Empty room 100 100 099Lego room 095 100 096
Mean RTObject 2435 2932 2161 (980)Lego object 2052 3245 2029 (921)Empty room 2744 2291 1788 (959)Lego room 1904 2281 1630 (1127)
Figure 6 The four kinds of stimuli used in Experiment 1 Topleft lego layouts top right real-world layouts (empty rooms)bottom left lego objects bottom right real-world objects
difference did not appear to be significant Impor-tantly there was little evidence that GR or CO tooklonger to make their perceptual judgements for thereal or Lego scenes than for the real or Lego objectsThus contrary to our initial hypothesis this experi-ment found no clear evidence for selective impair-ment in the perception of scene-like stimuli
Experiment 2 Different-views matchingtask
The previous experiment demonstrated that GRand CO can extract information about the per-ceived distances between themselves and the sur-faces of an object or scene Thus in at least onesense their perception of these surfaces is pre-served However this does not mean that their abil-ity to represent the layout of a scene is unimpairedThe two-dot task requires on-line comparison oftwo surfaces in egocentric coordinates GR and COwould be able to perform this task even if they couldnot (1) represent more than two surfaces at a time(2) represent surfaces in other than strictly egocen-tric coordinates or (3) sustain surface representa-tions for longer than the time it takes to make acomparison It has been suggested that the parietallobes are involved in on-line processing of surfacegeometry in egocentric coordinates (Milner ampGoodale 1995) thus it is possible that the parietallobes (which are intact in GR and CO) may be suf-ficient to perform the two-dot task even in theabsence of more durable or complex surface repre-sentations in parahippocampal cortex
In Experiment 2 we examine GR and COrsquosability to create and sustain a representation of thegeometry of an object or scene-like layout The taskwas to view a photograph of a Lego object or Legoscene hold it in memory for 10 s and then deter-mine whether a second object or scene was the sameas the first In ldquosamerdquo trials the second photographdepicted the same scene or object depicted from adifferent viewing angle In ldquodifferentrdquo trials the sec-ond photograph depicted a different scene or objectchosen to be similar to the first one (Figure 7)Thus in order to successfully perform this tasksubjects had to form a representation of the surfacesof the object or scene sustain this representation in
memory for 10 s and then align it (presumablyusing mental rotation) to the second object orscene
MethodThe experiment consisted of two parts each with40 trials In the first part the stimuli were black andwhite photographs of objects made out of Legoblocks In the second part the stimuli were photo-graphs of ldquoscenesrdquo made out of Lego blocks whichwere designed to have a geometric structure similarto real-world places such as a roomor a city street
Subjects began each trial by pressing the spacebar A photograph of a Lego object or sceneappeared on the screen for 3 s followed by a 10-sblank interval A second photograph then appeareddepicting either (1) the same object or scene as thefirst photograph shown from a different viewingangle (offset approximately 35 degrees) or (2) a dif-ferent object or scene Subjects were instructed topress the ldquojrdquo key if the two photographs depicted thesame object or scene and the ldquof rdquo key if they did notThey were told to take as much time as they neededto make a decision but no extra time The secondphotograph remained on the screen until a responsewas made Half the trials were ldquosamerdquo trials and halfwere ldquodifferentrdquo trials
Results and discussionFigure 8 shows results for GR and CO as well as forsix elderly normal controls GRrsquos and COrsquos perfor-mance was comparable to that of the normal con-trols Furthermore GR and CO performed just aswell when the stimuli were Lego scenes as whenthey were Lego objects These results are unlikely toreflect a ceiling effect as performance for bothpatients and controls was around 80 correctThus both patients appear to be able to form a rep-resentation of the geometry of both scenes andobjects which they can sustain in short-termmemory
Experiment 3 N-back recognition memorytask
The previous experiment failed to find any impair-ment in GR and COrsquos ability to form a representa-
EPSTEIN ET AL
496 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
tion of the geometry of a scene or object andmaintain it for 10 s in the absence of the originalstimulus Not only does this result suggest that per-ception of scene-like stimuli is intact in thesepatients it further suggests that some short-termmemory functions are intact as well However we
should note that subjects in this experiment werefree to devote all their effort and attention in the 10-second blank interval to maintaining a memory ofthe stimulus Thus it is possible that GR and COcan use rehearsal mechanisms to maintain the stim-ulus in short-term memory during the blank inter-
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 497
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
(b) Lego object version
Figure 7 Stimuli and procedure for Experiment 2 (a) In the Lego layout version subjects see a Lego layout for 3 s followed by a blankscreen for 10 s A second layout then appears which is either the same layout as the first one but seen from a different angle or a differentlayout (b) The Lego object version is the same but with object stimuli
3 s 10 s OR
3 s 10 s OR
(a) Lego layout version
(b) Lego object version
val but are unable to form a more durable memorytrace for the stimulus This possibility was tested inthe current experiment which was designed toexamine GR and COrsquos ability to learn new visualinformation
We devised a continuous n-back recognitionmemory test in which subjects viewed a sequence oforiginally novel stimuli some of which wererepeated within the sequence at intervals of 1 3 or5 intervening images For each stimulus subjects
reported whether they had seen it before in theexperiment or not In different runs in the experi-ment the stimuli were black-and-white photo-graphs of (1) Lego scenes (2) Lego objects (3) real-world landscapes (4) novel objects made out ofSculpey clay (5) unfamiliar houses (6) unfamiliarfaces as well as (7) line drawings of nonsenseobjects and (8) pronounceable pseudowords (Fig-ure 9) Thus we were able to test GR and COrsquos rec-ognition memory for different kinds of materials atdifferent time intervals in a paradigm in which thepresence of intervening items between the twoappearances of each stimulus made rehearsal diffi-cult Note that because none of the specific stimuliused in this experiment were familiar in advancethis task did not require an episodic memory com-ponent and could be done on the basis of pure visualfamiliarity with the particular visual stimulus
Although many different stimulus classes weretested in this experiment the critical comparison isbetween the Lego layouts and Lego objectsbecause these are best matched with regard tomaterials and differ primarily in their geometricstructure (see Introduction) We predicted that ifGR and CO showed a memory deficit for this taskit would be greater for the Lego layouts than for the
EPSTEIN ET AL
498 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 8 Results of Experiment 2 Performance over ldquosamerdquotrials and ldquodifferentrdquo trials is averaged together Error bars show1 SD
Figure 9 Stimuli for Experiment 3 Top row Lego layouts real-world scenes houses Bottom row Lego objects clay objects nonsenseobjects faces In an eighth condition (not shown) subjects viewed pronounceable nonwords such as ldquowilchrdquo or ldquothipperrdquo
Lego objects In addition a deficit for materialsthat convey information about the layout of sur-rounding space might also be apparent when GRand CO are tested on more realistic stimuli If soone would expect them to do poorly on the land-scapes but relatively well on the clay objects linedrawings of nonsense objects faces andpseudowords
MethodFor each subject a complete experimental sessionconsisted of being run once on each of the eightstimulus classes Each run consisted of 90 trials ineach trial a stimulus was presented and subjectsreported whether or not they had seen it before inthe run (or indeed ever) by pressing the ldquojrdquo key ifthey had or the ldquof rdquo key if they had not Each stimu-lus was on the screen for either 1 2 or 3 s (see fol-lowing) and appeared promptly after the keypressreport for the previous trial Individual stimuliappeared either once (14 stimuli) twice (30 stim-uli) or four times (4 stimuli) within a given run Ofthe stimuli that were presented twice 10 wererepeated immediately (1-back) 10 were repeatedwith a lag of two intervening items (3-back) and 10were repeated with a lag of four intervening items(5-back) In order to ensure that between-subjectdifferences in performance were not due to stimulusdifferences the same stimuli were assigned to thesame conditions each time the experimentwas run
GR performed the experiment twice once inAugust 1998 and again 9 months later Because ofa computer error he was only tested on six of theeight categories (all except Lego layouts and Legoobjects) at the later date CO was also tested twiceonce in February 1999 and again 6 months laterReported results for GR and CO are the average oftheir two testing sessions Normal subjects wereonly tested once Stimulus presentation time was 1 sin GRrsquos first testing session In pilot testing COfound this too fast for the critical categories of Lego
layouts and Lego objects so presentation time wasextended to 2 s for these categories in his first test-ing session All normal subjects were tested at thesepresentation rates (2 s for Lego layouts and objects1 s for other stimuli) In COrsquos second testingsession presentation time was lengthened to 3 s forall stimulus categories to ensure that any deficitsfound were memory deficits rather than problemswith the nonmemory aspects of the task
Results and discussionResults from GR CO and 10 elderly normal con-trols are shown in Figure 10 and Table 4 Figure 10ashows results from the main comparison betweenLego layouts and Lego objects Both normal sub-jects and patients performed near ceiling for bothLego layouts and Lego objects at 1-backinterstimulus intervals verifying that GR and COcould successfully perceive the pictures as they werebeing presented In contrast there was a strikingdifference between the two patients and the normalsubjects at 5-back intervals Whereas normals didon average somewhat better on Lego layouts thanLego objects both GR and CO showed the oppo-site pattern doing much worse on Lego layoutsthan Lego objects The 3-back condition showed amixed pattern Whereas GR did better on Legoobjects than Lego layouts in this condition (similarto his performance at 5-back intervals) CO didbetter on Lego layouts than Lego objects COrsquos lowperformance on Lego objects in the 3-back condi-tion appears to be an anomaly as it is much lowerthan his performance for comparable stimuli in the1-back and 5-back conditions
Figure 10b shows the difference in performancebetween Lego layouts and Lego objects for thecritical 1-back and 5-back intervals At 5-backintervals CO was significantly impaired comparedto the normal subjects (z = ndash250) whereasGRrsquos impairment fell just short of significance (z =ndash175)4
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 499
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
4In fact these values may underestimate GR and COrsquos relative impairment for Lego layouts In the 5-back condition 4 of the 10
normal subjects had perfect performance on the Lego layouts but all scored below ceiling on the Lego objects Thus the true advantagefor Lego layouts over Lego objects for normal subjects is probably greater than that observed here in which case GR and COrsquos lowerperformance for Lego layouts compared to Lego objects would be even more abnormal Furthermore post-hoc analyses revealed thatone of the normal subjects showed a difference in performance between Lego layouts and objects that was over 2 SD below the mean ofthe other nine subjects When this outlying subject was excluded z-scores were ndash322 for CO and ndash232 for GR
Table 4 shows results for all eight stimulusclasses Although direct comparisons between sub-jects should be treated with caution since viewingtime for different items was not the same for eachsubject we do note a few interesting patterns Firstof all there is little or no evidence that GR or CO
show a memory deficit for nonsense words clayobjects or line drawings of nonsense objects GRappears to be slightly impaired on faces and housesbut CO performs normally on these categoriesStrikingly GR shows a severe deficit for the reallandscapes performing more than 40 percentagepoints lower than normals Thus even thoughthere is some evidence that GR might have someimpairment for nonscene stimuli he clearly is muchmore impaired for scene-like stimuli such as theLego scenes and landscapes CO does not show asimilar deficit for the landscapes However itshould be kept in mind that in one of his two testingsessions CO viewed each picture for 3 s which wasmuch longer than the 1 second used for all non-Lego stimuli for GR and normals Thus COrsquosapparently ldquonormalrdquo performance in these condi-tions might be overestimated
Taken together the results of this experimentindicate that GR and CO are selectively impaired atprocessing scene-like stimuli This deficit is consis-tently apparent at 5-back but not 1-back intervalsThis finding suggests that the deficit should becharacterised as a memory encoding problemHowever the present results do not allow us todetermine whether the difficulty occurs at memoryencoding or at subsequent recognition (see Epsteinamp Kanwisher 2001) If GR and CO have a generalproblem with recognition of topographical materi-als then they should not be able to recognise scenes
EPSTEIN ET AL
500 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 10a Results of Experiment 3 for GR (top) CO (middle)and for 10 age-matched normal controls Performance measure isproportion of repeated stimuli correctly identified as such correctedfor guessing using the standard formula
Figure 10b Difference between Lego layout performance andLego object performance for the 1-back and 5-back conditionsError bars indicate one 1 SD
and landmarks that were learned prior to theirinjury We tested this possibility in the nextexperiment
Experiment 4 Famous landmarksfacesrecognition task
In this experiment we examined GR and COrsquosability to recognise the visual appearances of placeslearned prior to injury Subjects saw a series offamous landmarks intermixed with nonfamouscontrols (Imaging experiments have demonstratedthat landmarks of this type can activate the PPAalmost as strongly as complete scenes Epstein etal 1999 Epstein amp Kanwisher 1998) They wererequired to say whether they knew each specificlandmark and to identify the ones they knew Sub-jects were instructed to respond affirmatively if theywere fairly certain that they recognised the land-mark but not if it only looked generally familiarFor purposes of comparison subjects also per-formed an identical test in which the stimuli werefamous and nonfamous faces In both testsdistractor items were designed to be visually andconceptually similar to the target items (see Figure11) If parahippocampal cortex is required for placerecognition then one might expect GR and CO toperform poorly compared to normals on the land-mark recognition test
MethodEach of the two experiments (famous faces famouslandmarks) consisted of 48 trials Subjects beganeach trial by pressing the space bar after which ablack-and-white photograph of a face (in thefamous faces experiment) or a landmark (in thefamous landmarks experiment) appeared on thescreen Subjects were told to press the ldquojrdquo key if theyrecognised the specific face or landmark and the ldquof rdquokey if they did not taking as much time at theyneeded to make this discrimination but no extratime Subjects were told to report that they recog-nised a face or landmark if they felt that they knewit even if they could not retrieve its name If theydid know the name they were to say it after pressingthe button Otherwise they were to say whateverthey did remember about the item
Half of the photographs were of famous and halfof nonfamous people and landmarks The famousfaces included public figures (eg George BushRichard Nixon Prince Charles) and movie stars(eg Harrison Ford Clark Gable Marlon Brando)who were prominent prior to the time of GR andCOrsquos injuries the famous landmarks were build-ings and monuments from all over the world (egEiffel Tower US Capitol Building Great Sphinx)The nonfamous stimuli were chosen to match thefamous stimuli as closely as possible Each famousstimulus had a ldquomatchrdquo For example George Bushwas matched to a man of similar age in a coat andtie and the Taj Mahal was matched to a visually
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 501
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 4 Full results of Experiment 3 Mean proportion correct (corrected for guessing) SDs in parentheses
Lego Land- Lego Clay Nonsense Non-scenes scapes Houses objects objects objects Words Faces
1-back GR 100 094 100 100 100 094 100 100CO 091 085 091 100 100 079 074 100Normals 096(008) 095(008) 093(014) 096(008) 096(006) 099(003) 098(004) 097(011)
3-back GR ndash003 068 026 047 083 083 075 052CO 066 081 037 010 094 069 095 081Normals 066(032) 077(021) 072(015) 077(020) 086(016) 083(015) 083(017) 072(020)
5-back GR 010 034 044 047 072 083 094 053CO 031 073 075 088 081 064 100 081Normals 076(026) 077(016) 068(021) 066(016) 091(014) 083(018) 093(011) 078(021)
similar Asian palace that is not famous (see Figure11)
Results and discussionResults for GR and CO as well as for five age-matched controls are shown in Figure 12 Both
GR and CO could recognise the famous landmarksas well normals In addition both GR and CO wereusually able to name the landmarks they success-fully identified
Thus GR and CO can recognize landmarkslearned prior to injury demonstrating that at leastsome of their place recognition abilities are intactWhen combined with the fact that GR and COwere able to produce detailed maps of places theylived in prior to their injury (see Case Description)these results suggest that their deficit is primarily inencoding or consolidation of new informationrather than in recognition or recall of old informa-tion Note however that these results do not provethat their place recognition abilities are entirelynormal GR and CO might suffer from a mild rec-ognition deficit that affects identification of newly-learned stimuli like the Lego layouts in Experiment3 but not overlearned stimuli like the famous land-marks Furthermore it is possible that GR and COmight be impaired at recognising premorbidlylearned places that are defined primarily by theirspatial layout even though they are unimpaired atrecognising premorbidly learned places that aredefined primarily by a single object-like landmarkor building (see General Discussion) Neverthelessthe trend of the present evidence is to suggest thatGR and CO have particular problems with noveltopographical information
In the famous-face task GRrsquos performance wasin the normal range However he sometimes haddifficulty naming the faces even though he clearly
EPSTEIN ET AL
502 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 11 Stimuli for Experiment 4 Each famous landmark(Arc de Triomphe Taj Mahal) and famous face (Barbara BushYasser Arafat) was matched to a nonfamous building or face thatalso appeared in the experiment
FA
CE
SL
AN
DM
AR
KS
Figure 12 Results of Experiment 4 Performance measure isproportion of famous stimuli reported as ldquoknownrdquo corrected forguessing using the standard formula
recognised them For example he described Ron-ald Reagan as ldquothe gipperrdquo Nancy Reagan as ldquothewife of the gipperrdquo and Clark Gable as ldquothe movieactor who said lsquoScarlett I donrsquot give a damnrsquordquo COhad a great deal of difficulty identifying the famousfaces This result is not surprising given hisextremely low performance on the Benton FaceMatching Task In addition to his topographicproblems CO appears to suffer fromprosopagnosia
GENERAL DISCUSSION
We tested two parahippocampal patients (GR andCO) on four experiments designed to measure per-formance as a function of both task (perceptionencoding and recognition) and stimulus type(scenes objects faces and words) The only taskthat revealed a clear deficit was an n-back recogni-tion memory test (Experiment 3) In order to suc-cessfully perform this task subjects had to encode adurable representation of a visual stimulus and useit for later recognition GR and CO were signifi-cantly more impaired on this task when the stimuliwere scene-like spatial layouts (ie Lego scenesand real scenes for GR) than when they wereobjects No comparable deficits were found in otherexperiments that tested layout perception (Experi-ments 1 and 2) and placelandmark recognition(Experiment 4) Taken together these results indi-cate that GR and CO are impaired in their ability toencode novel information about the geometry ofsurrounding space into memory but are lessimpaired at their ability to encode the geometry ofnovel objects
We argue that this specific topographical defi-citmdashinability to encode the spatial layout of novelscenesmdashresults from damage to the para-hippocampal place area Although the damage inboth patients extends beyond the parahippo-campal-lingual territory normally associated withthe PPA this region is clearly lesioned bilaterally inGR and in the right hemisphere in CO Further-more functional imaging demonstrated that GRhas no functioning PPA Thus the current resultsand the earlier fMRI work (Epstein et al 1999
Epstein amp Kanwisher 1998) converge to implicateparahippocampal cortex in the encoding of infor-mation about scene geometry into memory
Our assignment of the function of memoryencoding to parahippocampal cortex is consistentwith the results of earlier case reports of patientswith parahippocampal damage who have naviga-tional difficulties in novel but not familiar environ-ments (Habib amp Sirigu 1987 Ross 1980 see alsoTeng amp Squire 1999) As with GR and CO thesepatients were either less impaired or unimpaired onnontopographical memory tasks Patients have alsobeen described who exhibit the opposite pattern ofimpairment an inability to learn faces (sometimescalled ldquoprosopamnesiardquo Cipolotti Robinson Blairamp Frith 1999 Tippett Miller amp Farah 2000) orfaces and verbal materials (Maguire amp Cipolotti1998) with an unimpaired ability to learn topo-graphical materials Taken together these resultsargue for a high degree of domain-specificity in theneural systems involved in learning of new informa-tion (Gallistel 1995) The brain may have separatemechanisms for learning faces places and wordswhich can be differentially impaired after damageto specific cortical regions (Cipolotti et al 1999Incisa della Rochetta et al 1996) The presentresults suggest that the anatomical locus of themechanism dedicated to learning new topographi-cal information is in the parahippocampal cortex(Aguirre amp DrsquoEsposito 1999)
In contrast to their impaired ability to encodenew topographical information both GR and COwere able to produce detailed maps of their previousdwellings and performed normally on the famouslandmark recognition test of Experiment 4 Theseresults are generally consistent with Teng andSquirersquos (1999) report of a patient with extensivemedial temporal damage who could recallpremorbidly learned but not postmorbidly learnedtopographical information (see also Ross 1980)Thus our results provide further evidence thatmedial temporal regions such as parahippocampalcortex may be more critical for encoding or consoli-dating new information into memory than for recallof old information
One issue that our experiments do not entirelyresolve is the precise nature of the topographical
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 503
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
information encoded by parahippocampal cortexClearly this region is critical for encoding informa-tion about the geometry of the currently visiblescene even when the ldquoscenerdquo is made out of Legoblocks and therefore has no immediate navigationalrelevance However it is unclear whether this spe-cific deficit can account for all of the real-worldnavigational difficulties encountered by GR andCO GR in particular shows such a catastrophicloss of navigational ability that we suspect he is alsoimpaired at encoding the spatial relationshipsbetweenthe current scene and other locations in theworld CO probably also suffers from such difficul-ties as he reports that he has trouble rememberinghow places relate to each other in large-scale spaceWhen he was tested on a simple 3-room videogameenvironment he not only was unable to rememberthe individual rooms (claiming that there were 10rooms when there were only 3) but he also had nomemory for the spatial relationships between themThese spatial deficits may be the result of damage tothe posterior right hippocampus (in the case of CO)or of disconnection of the posterior right hippo-campus from the parahippocampal regions thatprovide much of its cortical input (in both patientsSuzuki amp Amaral 1994) In either case the abilityto form a cognitive map of new environments likethe lab or videogame space could be impaired(Maguire et al 1998 OrsquoKeefe amp Nadel 1978)However it is also possible that parahippocampalcortex itself may be critical for encoding not onlythe geometry of individual scenes but also the spa-tial relationships between the locations of thosescenes In this case the spatial deficits of GR andCO could be attributed directly to parahippo-campal damage rather than to hippocampal dam-age or disconnection
The latter scenario would be consistent withAguirre and DrsquoEspositorsquos (1999) and Barrash etalrsquos (2000) suggestion that parahippocampal cortexis critically involved in combining visual informa-tion about the appearance of scenes and landmarkswith spatial information about their locations into aunified topographical representation Howeverboth of these authors argue that the representationsof individual scenes (Barrash et al) and landmarks(Aguirre amp DrsquoEsposito) are supported not by
parahippocampal cortex but by more posterior lin-gual-fusiform regions In contrast we believe thatthe neuroimaging data indicate that parahippo-campal cortex is involved in the encoding of thegeometric structure of individual scenes althoughit may also play a role in combining these represen-tations with other spatial representations
Interestingly GR and CO were not impaired onrecognition of famous landmarks that were mostlybuildings Thus our data do not preclude the possi-bility that there may be a separate lingual orfusiform region involved in processing informationabout buildings (Aguirre Zarahn amp DrsquoEsposito1998 Chao Haxby amp Martin 1999 IshaiUngerleider Martin Schouten amp Haxby 1999)which might have been preserved in GR and COdespite extensive damage to other regions of thelingual and fusiform gyrus Alternatively GR andCOrsquos preserved building recognition abilities mightbe mediated by interactions between cortical areasthat perform shape analysis for any object-likestimulus type (Malach et al 1995Kanwisher et al1996) and anterior temporal regions in which long-term topographical and semantic memories arestored (McCarthy et al 1996)
Is perception preserved
In contrast to their inability to encode new scenesinto memory GR and CO have substantiallyretained their ability to perceive the spatial layout ofthe currently visible scene (Experiments 1 and 2)GR can follow a route marked on a floor planwhich indicates a good on-line appreciation of hiscurrent spatial surroundings Furthermore neitherGR or CO have any trouble moving through theenvironmentmdashthey move with confidence and donot bump into doors or walls Problems of the lattertype are usually associated with damage to the pari-etal lobes which are intact in GR and CO Thus atfirst glance GR and CO do not appear to haveperceptual impairments (beyond their visual fieldrestrictions)
However GR does complain of a perceptualdeficit which he describes as an inability to ldquotakeinrdquo scenes when they are too complex Similarreports of an inability to comprehend the
EPSTEIN ET AL
504 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
ldquoglobalityrdquo of a scene were made by some of theparahippocampally-damaged patients studied byHabib and Sirigu (1987) We were not able todetermine the precise nature of this deficit whichmay be partly attributable to his visual field restric-tions It appears that his deficit is more salient in thereal-world environments than in the picture-per-ception tasks of Experiments 1 and 2 perhapsbecause he finds it more difficult to organise visualinput when immersed in it than when viewing pho-tographs that subtend only part of the visual field
We suggest that GR can perceive parts of thescene when he attends to them but may be unableto integrate information over time into a coherentrepresentation of a scene or place The fact that GRand CO are impaired for Lego layouts at 5-backintervals in Experiment 3 is consistent with thishypothesis The time between the first and secondpresentations of a stimulus is very short in this case(and even shorter with 3-back intervals where GRalso shows evidence of a deficit) In both patientsthe representation of layout information formed byperception of a Lego scene must either have anabnormally short persistence be abnormally sus-ceptible to interference or both If so they mightfind it impossible to sustain a spatial frame orschema long enough to use it to organise informa-tion obtained from many different glances at a com-plex scene (Intraub 1997) The result would be aninability to understand the ldquoglobalityrdquo of a sceneeven though all the details were perfectlyperceived
Non-topographical mnemonic abilities
Although not as severe as their topographical mem-ory problems GR and CO do have some memorydifficulties in other domains For example bothperformed poorly on the Rey AVLT whichrequired them to remember a list of unrelatedwords Both also report some subtle episodic mem-ory problems (see Case Description) COrsquos prob-lems could partly result from damage to the righthippocampusmdashindeed CO appeared to have themore acute episodic memory problems HoweverGR has no hippocampal damage so the fact that hedisplays episodic memory problems suggests thatparahippocampal cortex is involved in the forma-
tion of episodic memories (cf Wagner et al1998)
In previous work (Epstein et al 1999) weargued that spatial context information encoded byparahippocampal cortex might play a role in estab-lishing event memories by encoding informationabout where events take place The inability toencode this information might cause problems intasks such as the Rey AVLT where one has to recalla list of unrelated words If spatial (and perhapstemporal) context information is not encoded itwill be harder subsequently to recall the distinctevent of hearing each word (Tulving 1983) Incontrast lack of spatial context information wouldbe less of a problem in tasks where one is onlyrequired to recognise previously encountered verbalmaterials but not to reproduce the spatiotemporalcontext in which they were experienced IndeedGR and CO performed as well as normals on thenonsense words in Experiment 4 where recogni-tion memory but not episodic memory was requiredfor successful performance Interestingly GR andCO also did relatively well on the Logical Memorysubtest of the WMS-III perhaps because the nar-rative framework of a story provides sufficient con-textual information to overcome the absence ofspatial context and mediate successful verbalretrieval
Differences between GR and CO
Although we have discussed GR and CO togetherthere are a number of relevant differences betweenthem One open question is whether these differ-ences can be attributed to the different effects ofunilateral and bilateral parahippocampal damageIn general GRrsquos topographical difficulties weremore severe than COrsquos CO did surprisingly well onthe Warrington Topographical Memory Test inwhich he was required to remember detailed real-world city scenes and also performed substantiallybetter on real scenes than Lego scenes in Experi-ment 3 Insofar as CO has intact left para-hippocampal cortex this suggests that the left PPAmight be sufficient to encode a certain amount of(perhaps semantic) information about real-worldscenes but the right PPA is necessary to encode
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 505
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
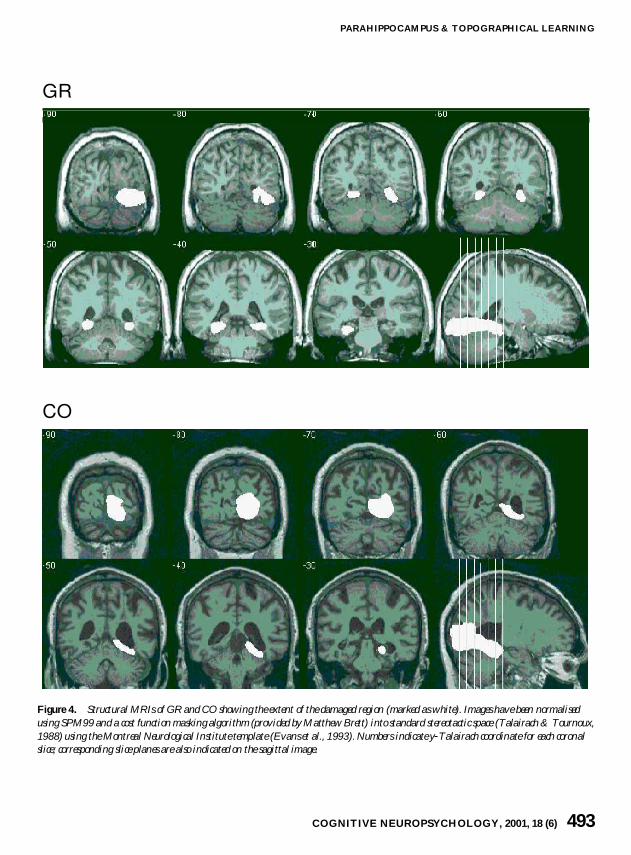
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 493
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
GR
CO
Figure 4 Structural MRIs of GR and CO showing the extent of the damaged region (marked as white) Images have been normalisedusing SPM99 and a cost function masking algorithm (provided by Matthew Brett) into standard stereotactic space (Talairach amp Tournoux1988) using the Montreal Neurological Institute template (Evans et al 1993) Numbers indicate y-Talairach coordinate for each coronalslice corresponding slice planes are also indicated on the sagittal image
(FFA) commonly seen in normal subjects(Kanwisher McDermott amp Chun 1997) Thusthe MRI was sensitive enough to detect functionaldifferences an FFA was found but no PPA Whencombined with the anatomical results this stronglysuggests that GR has no PPA
Because of safety concerns (presence of a femo-ral artery aneurysm clip) we did not perform a simi-lar high-field functional MRI of CO However asFigure 4 demonstrates CO clearly has damage tothe parahippocampal-lingual region in the righthemisphere
EXPERIMENTAL INVESTIGATIONS
In this section we report the results of four experi-ments designed to probe the consequences of dam-age to the lingual-parahippocampal region ingeneral and the PPA in particular Our primaryconcern in these experiments was to test GR andCOrsquos ability to perceive remember and recognisedifferent kinds of visual material Based on our pre-vious fMRI work we predicted that GR and CO
would have more difficulty with scene-like stimuli(ie stimuli that convey information about thegeometry of a space one can navigate through or actwithin) than with object-like stimuli Such a deficitcould account for at least some of the problems GRand CO encounter in real-world navigation
Beyond determining the kind of visual materialthat gets processed in parahippocampal cortex wealso wanted to determine the nature of the process-ing that takes place on that material In particularwe wanted to determine if parahippocampal cortexis involved in perceptual encoding memory encodingor recognition As discussed earlier (Introduction)our fMRI results provided some evidence againstplace recognition as the sole function of the PPAbut did not allow us to distinguish between percep-tual and mnemonic encoding Experiments 1 and 2examine the role of parahippocampal cortex in per-ceptual encoding Experiment 3 examines its role inmnemonic encoding and Experiment 4 examinesits role in place recognition
Experiment 1 Two-dot task
The first hypothesis we examined was thatparahippocampal cortex plays a role in perceivingthe shape of the local environment In our previousfMRI work we observed that the PPA respondsmuch more strongly during scene viewing than dur-ing object or face viewing even when subjects donothing more than watch the scenes passively with-out performing any other task This suggested thatscene perception alone may be sufficient to activatethe PPA Previous reports have indicated that dam-age to parahippocampal cortex and other medialtemporal regions leads to problems with memoryrather than perception (Bohbot et al 1998Maguire Burke Philips amp Staunton 1996 Pigottamp Milner 1993) However all of these previousresults were obtained on patients who had unilat-eral lesions GR has a bilateral occipital-temporallesion including clear damage to parahippocampalcortex in both hemispheres furthermore he reportssome kind of perceptual deficit We hypothesisedthat this deficit might involve difficulty perceivingor interpreting the geometry of scenes and otherstimuli with a similar geometric structure
EPSTEIN ET AL
494 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 5 Top row shows a single slice from a functional MRIscan performed on GR Bottom row shows results from a normalsubject (NK) for comparison Voxels that responded more to facesthan objects (left column) or more to scenes than faces and objects(right column) in a Kolomogorov-Smirnov test are highlighted bywhite arrows GR has a functioning face area (FFA) but there isno evidence of a place area (PPA)
FFA PPA
GR
NK
In this experiment subjects viewed either objectshapes or spatial layouts Stimuli consisted ofblack-and-white photographs of real-world stimuli(empty rooms vs common objects) or abstractstimuli made out of Lego blocks (Lego ldquoscenesrdquo vsLego ldquoobjectsrdquo) These stimuli have been previouslyshown to differentially activate the PPA in normalsubjects (Epstein et al 1999 Epstein ampKanwisher 1998) Two red dots were superim-posed on each photograph (Figure 6) The task wasto report which of the two red dots was over thecloser part of the object or scene This task wasdesigned to have no memory component (apartfrom remembering the instructions) but to requireaccurate 3D perception of objects and scenes
If GR or CO have difficulty perceiving or inter-preting the spatial layout of scenes then we pre-dicted they would do worse when the stimuli areempty rooms or Lego scenes than when they arereal-world or Lego objects Note that the criticalcomparison is between the Lego scenes and theLego objects as these stimuli are made of the samematerials and differ primarily in their surfacegeometry
MethodTesting for all experiments was done on aMacintosh computer The experiment consisted of80 trials (20 of each stimulus type randomly inter-mixed) Subjects began each trial by pressing thespace bar A black-and-white photograph from oneof the four stimulus categories (see earlier) thenappeared on the screen with two red dots on it Thetask was to determine which of the two red dotsindicated the closer part of the scene or objectSubjects pressed the ldquofrdquo key if the leftmost red dotwas closest and the ldquojrdquo key if the rightmost dotwas closest They were instructed to respond asquickly as possible without making errors Stimuliremained on the screen until a response was made
The stimulus set was constructed so that eachempty room was matched to a common object thathad red dots in the exact same location and eachLego layout was similarly matched to a Lego objectThus the locations of the red dots were counterbal-anced across scene and object stimuli controllingfor any effects of visual field cuts
Results and discussionResults for GR and CO and four elderly normalsubjects are shown in Table 3 Both GR and COcould do this task without difficulty and their per-formance was near ceiling There was no evidencethat they had any more trouble with the emptyrooms or Lego scenes than with the real or Legoobjects Reaction times were slightly longer for GRand CO than for normal subjects as one wouldexpect given their visual field deficits however this
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 495
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 3 Experiment 1 (two-dot) results (SD in parenthesis)
GR CO Normals
Proportion correctObject 095 080 093Lego object 100 095 098Empty room 100 100 099Lego room 095 100 096
Mean RTObject 2435 2932 2161 (980)Lego object 2052 3245 2029 (921)Empty room 2744 2291 1788 (959)Lego room 1904 2281 1630 (1127)
Figure 6 The four kinds of stimuli used in Experiment 1 Topleft lego layouts top right real-world layouts (empty rooms)bottom left lego objects bottom right real-world objects
difference did not appear to be significant Impor-tantly there was little evidence that GR or CO tooklonger to make their perceptual judgements for thereal or Lego scenes than for the real or Lego objectsThus contrary to our initial hypothesis this experi-ment found no clear evidence for selective impair-ment in the perception of scene-like stimuli
Experiment 2 Different-views matchingtask
The previous experiment demonstrated that GRand CO can extract information about the per-ceived distances between themselves and the sur-faces of an object or scene Thus in at least onesense their perception of these surfaces is pre-served However this does not mean that their abil-ity to represent the layout of a scene is unimpairedThe two-dot task requires on-line comparison oftwo surfaces in egocentric coordinates GR and COwould be able to perform this task even if they couldnot (1) represent more than two surfaces at a time(2) represent surfaces in other than strictly egocen-tric coordinates or (3) sustain surface representa-tions for longer than the time it takes to make acomparison It has been suggested that the parietallobes are involved in on-line processing of surfacegeometry in egocentric coordinates (Milner ampGoodale 1995) thus it is possible that the parietallobes (which are intact in GR and CO) may be suf-ficient to perform the two-dot task even in theabsence of more durable or complex surface repre-sentations in parahippocampal cortex
In Experiment 2 we examine GR and COrsquosability to create and sustain a representation of thegeometry of an object or scene-like layout The taskwas to view a photograph of a Lego object or Legoscene hold it in memory for 10 s and then deter-mine whether a second object or scene was the sameas the first In ldquosamerdquo trials the second photographdepicted the same scene or object depicted from adifferent viewing angle In ldquodifferentrdquo trials the sec-ond photograph depicted a different scene or objectchosen to be similar to the first one (Figure 7)Thus in order to successfully perform this tasksubjects had to form a representation of the surfacesof the object or scene sustain this representation in
memory for 10 s and then align it (presumablyusing mental rotation) to the second object orscene
MethodThe experiment consisted of two parts each with40 trials In the first part the stimuli were black andwhite photographs of objects made out of Legoblocks In the second part the stimuli were photo-graphs of ldquoscenesrdquo made out of Lego blocks whichwere designed to have a geometric structure similarto real-world places such as a roomor a city street
Subjects began each trial by pressing the spacebar A photograph of a Lego object or sceneappeared on the screen for 3 s followed by a 10-sblank interval A second photograph then appeareddepicting either (1) the same object or scene as thefirst photograph shown from a different viewingangle (offset approximately 35 degrees) or (2) a dif-ferent object or scene Subjects were instructed topress the ldquojrdquo key if the two photographs depicted thesame object or scene and the ldquof rdquo key if they did notThey were told to take as much time as they neededto make a decision but no extra time The secondphotograph remained on the screen until a responsewas made Half the trials were ldquosamerdquo trials and halfwere ldquodifferentrdquo trials
Results and discussionFigure 8 shows results for GR and CO as well as forsix elderly normal controls GRrsquos and COrsquos perfor-mance was comparable to that of the normal con-trols Furthermore GR and CO performed just aswell when the stimuli were Lego scenes as whenthey were Lego objects These results are unlikely toreflect a ceiling effect as performance for bothpatients and controls was around 80 correctThus both patients appear to be able to form a rep-resentation of the geometry of both scenes andobjects which they can sustain in short-termmemory
Experiment 3 N-back recognition memorytask
The previous experiment failed to find any impair-ment in GR and COrsquos ability to form a representa-
EPSTEIN ET AL
496 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
tion of the geometry of a scene or object andmaintain it for 10 s in the absence of the originalstimulus Not only does this result suggest that per-ception of scene-like stimuli is intact in thesepatients it further suggests that some short-termmemory functions are intact as well However we
should note that subjects in this experiment werefree to devote all their effort and attention in the 10-second blank interval to maintaining a memory ofthe stimulus Thus it is possible that GR and COcan use rehearsal mechanisms to maintain the stim-ulus in short-term memory during the blank inter-
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 497
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
(b) Lego object version
Figure 7 Stimuli and procedure for Experiment 2 (a) In the Lego layout version subjects see a Lego layout for 3 s followed by a blankscreen for 10 s A second layout then appears which is either the same layout as the first one but seen from a different angle or a differentlayout (b) The Lego object version is the same but with object stimuli
3 s 10 s OR
3 s 10 s OR
(a) Lego layout version
(b) Lego object version
val but are unable to form a more durable memorytrace for the stimulus This possibility was tested inthe current experiment which was designed toexamine GR and COrsquos ability to learn new visualinformation
We devised a continuous n-back recognitionmemory test in which subjects viewed a sequence oforiginally novel stimuli some of which wererepeated within the sequence at intervals of 1 3 or5 intervening images For each stimulus subjects
reported whether they had seen it before in theexperiment or not In different runs in the experi-ment the stimuli were black-and-white photo-graphs of (1) Lego scenes (2) Lego objects (3) real-world landscapes (4) novel objects made out ofSculpey clay (5) unfamiliar houses (6) unfamiliarfaces as well as (7) line drawings of nonsenseobjects and (8) pronounceable pseudowords (Fig-ure 9) Thus we were able to test GR and COrsquos rec-ognition memory for different kinds of materials atdifferent time intervals in a paradigm in which thepresence of intervening items between the twoappearances of each stimulus made rehearsal diffi-cult Note that because none of the specific stimuliused in this experiment were familiar in advancethis task did not require an episodic memory com-ponent and could be done on the basis of pure visualfamiliarity with the particular visual stimulus
Although many different stimulus classes weretested in this experiment the critical comparison isbetween the Lego layouts and Lego objectsbecause these are best matched with regard tomaterials and differ primarily in their geometricstructure (see Introduction) We predicted that ifGR and CO showed a memory deficit for this taskit would be greater for the Lego layouts than for the
EPSTEIN ET AL
498 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 8 Results of Experiment 2 Performance over ldquosamerdquotrials and ldquodifferentrdquo trials is averaged together Error bars show1 SD
Figure 9 Stimuli for Experiment 3 Top row Lego layouts real-world scenes houses Bottom row Lego objects clay objects nonsenseobjects faces In an eighth condition (not shown) subjects viewed pronounceable nonwords such as ldquowilchrdquo or ldquothipperrdquo
Lego objects In addition a deficit for materialsthat convey information about the layout of sur-rounding space might also be apparent when GRand CO are tested on more realistic stimuli If soone would expect them to do poorly on the land-scapes but relatively well on the clay objects linedrawings of nonsense objects faces andpseudowords
MethodFor each subject a complete experimental sessionconsisted of being run once on each of the eightstimulus classes Each run consisted of 90 trials ineach trial a stimulus was presented and subjectsreported whether or not they had seen it before inthe run (or indeed ever) by pressing the ldquojrdquo key ifthey had or the ldquof rdquo key if they had not Each stimu-lus was on the screen for either 1 2 or 3 s (see fol-lowing) and appeared promptly after the keypressreport for the previous trial Individual stimuliappeared either once (14 stimuli) twice (30 stim-uli) or four times (4 stimuli) within a given run Ofthe stimuli that were presented twice 10 wererepeated immediately (1-back) 10 were repeatedwith a lag of two intervening items (3-back) and 10were repeated with a lag of four intervening items(5-back) In order to ensure that between-subjectdifferences in performance were not due to stimulusdifferences the same stimuli were assigned to thesame conditions each time the experimentwas run
GR performed the experiment twice once inAugust 1998 and again 9 months later Because ofa computer error he was only tested on six of theeight categories (all except Lego layouts and Legoobjects) at the later date CO was also tested twiceonce in February 1999 and again 6 months laterReported results for GR and CO are the average oftheir two testing sessions Normal subjects wereonly tested once Stimulus presentation time was 1 sin GRrsquos first testing session In pilot testing COfound this too fast for the critical categories of Lego
layouts and Lego objects so presentation time wasextended to 2 s for these categories in his first test-ing session All normal subjects were tested at thesepresentation rates (2 s for Lego layouts and objects1 s for other stimuli) In COrsquos second testingsession presentation time was lengthened to 3 s forall stimulus categories to ensure that any deficitsfound were memory deficits rather than problemswith the nonmemory aspects of the task
Results and discussionResults from GR CO and 10 elderly normal con-trols are shown in Figure 10 and Table 4 Figure 10ashows results from the main comparison betweenLego layouts and Lego objects Both normal sub-jects and patients performed near ceiling for bothLego layouts and Lego objects at 1-backinterstimulus intervals verifying that GR and COcould successfully perceive the pictures as they werebeing presented In contrast there was a strikingdifference between the two patients and the normalsubjects at 5-back intervals Whereas normals didon average somewhat better on Lego layouts thanLego objects both GR and CO showed the oppo-site pattern doing much worse on Lego layoutsthan Lego objects The 3-back condition showed amixed pattern Whereas GR did better on Legoobjects than Lego layouts in this condition (similarto his performance at 5-back intervals) CO didbetter on Lego layouts than Lego objects COrsquos lowperformance on Lego objects in the 3-back condi-tion appears to be an anomaly as it is much lowerthan his performance for comparable stimuli in the1-back and 5-back conditions
Figure 10b shows the difference in performancebetween Lego layouts and Lego objects for thecritical 1-back and 5-back intervals At 5-backintervals CO was significantly impaired comparedto the normal subjects (z = ndash250) whereasGRrsquos impairment fell just short of significance (z =ndash175)4
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 499
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
4In fact these values may underestimate GR and COrsquos relative impairment for Lego layouts In the 5-back condition 4 of the 10
normal subjects had perfect performance on the Lego layouts but all scored below ceiling on the Lego objects Thus the true advantagefor Lego layouts over Lego objects for normal subjects is probably greater than that observed here in which case GR and COrsquos lowerperformance for Lego layouts compared to Lego objects would be even more abnormal Furthermore post-hoc analyses revealed thatone of the normal subjects showed a difference in performance between Lego layouts and objects that was over 2 SD below the mean ofthe other nine subjects When this outlying subject was excluded z-scores were ndash322 for CO and ndash232 for GR
Table 4 shows results for all eight stimulusclasses Although direct comparisons between sub-jects should be treated with caution since viewingtime for different items was not the same for eachsubject we do note a few interesting patterns Firstof all there is little or no evidence that GR or CO
show a memory deficit for nonsense words clayobjects or line drawings of nonsense objects GRappears to be slightly impaired on faces and housesbut CO performs normally on these categoriesStrikingly GR shows a severe deficit for the reallandscapes performing more than 40 percentagepoints lower than normals Thus even thoughthere is some evidence that GR might have someimpairment for nonscene stimuli he clearly is muchmore impaired for scene-like stimuli such as theLego scenes and landscapes CO does not show asimilar deficit for the landscapes However itshould be kept in mind that in one of his two testingsessions CO viewed each picture for 3 s which wasmuch longer than the 1 second used for all non-Lego stimuli for GR and normals Thus COrsquosapparently ldquonormalrdquo performance in these condi-tions might be overestimated
Taken together the results of this experimentindicate that GR and CO are selectively impaired atprocessing scene-like stimuli This deficit is consis-tently apparent at 5-back but not 1-back intervalsThis finding suggests that the deficit should becharacterised as a memory encoding problemHowever the present results do not allow us todetermine whether the difficulty occurs at memoryencoding or at subsequent recognition (see Epsteinamp Kanwisher 2001) If GR and CO have a generalproblem with recognition of topographical materi-als then they should not be able to recognise scenes
EPSTEIN ET AL
500 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 10a Results of Experiment 3 for GR (top) CO (middle)and for 10 age-matched normal controls Performance measure isproportion of repeated stimuli correctly identified as such correctedfor guessing using the standard formula
Figure 10b Difference between Lego layout performance andLego object performance for the 1-back and 5-back conditionsError bars indicate one 1 SD
and landmarks that were learned prior to theirinjury We tested this possibility in the nextexperiment
Experiment 4 Famous landmarksfacesrecognition task
In this experiment we examined GR and COrsquosability to recognise the visual appearances of placeslearned prior to injury Subjects saw a series offamous landmarks intermixed with nonfamouscontrols (Imaging experiments have demonstratedthat landmarks of this type can activate the PPAalmost as strongly as complete scenes Epstein etal 1999 Epstein amp Kanwisher 1998) They wererequired to say whether they knew each specificlandmark and to identify the ones they knew Sub-jects were instructed to respond affirmatively if theywere fairly certain that they recognised the land-mark but not if it only looked generally familiarFor purposes of comparison subjects also per-formed an identical test in which the stimuli werefamous and nonfamous faces In both testsdistractor items were designed to be visually andconceptually similar to the target items (see Figure11) If parahippocampal cortex is required for placerecognition then one might expect GR and CO toperform poorly compared to normals on the land-mark recognition test
MethodEach of the two experiments (famous faces famouslandmarks) consisted of 48 trials Subjects beganeach trial by pressing the space bar after which ablack-and-white photograph of a face (in thefamous faces experiment) or a landmark (in thefamous landmarks experiment) appeared on thescreen Subjects were told to press the ldquojrdquo key if theyrecognised the specific face or landmark and the ldquof rdquokey if they did not taking as much time at theyneeded to make this discrimination but no extratime Subjects were told to report that they recog-nised a face or landmark if they felt that they knewit even if they could not retrieve its name If theydid know the name they were to say it after pressingthe button Otherwise they were to say whateverthey did remember about the item
Half of the photographs were of famous and halfof nonfamous people and landmarks The famousfaces included public figures (eg George BushRichard Nixon Prince Charles) and movie stars(eg Harrison Ford Clark Gable Marlon Brando)who were prominent prior to the time of GR andCOrsquos injuries the famous landmarks were build-ings and monuments from all over the world (egEiffel Tower US Capitol Building Great Sphinx)The nonfamous stimuli were chosen to match thefamous stimuli as closely as possible Each famousstimulus had a ldquomatchrdquo For example George Bushwas matched to a man of similar age in a coat andtie and the Taj Mahal was matched to a visually
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 501
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 4 Full results of Experiment 3 Mean proportion correct (corrected for guessing) SDs in parentheses
Lego Land- Lego Clay Nonsense Non-scenes scapes Houses objects objects objects Words Faces
1-back GR 100 094 100 100 100 094 100 100CO 091 085 091 100 100 079 074 100Normals 096(008) 095(008) 093(014) 096(008) 096(006) 099(003) 098(004) 097(011)
3-back GR ndash003 068 026 047 083 083 075 052CO 066 081 037 010 094 069 095 081Normals 066(032) 077(021) 072(015) 077(020) 086(016) 083(015) 083(017) 072(020)
5-back GR 010 034 044 047 072 083 094 053CO 031 073 075 088 081 064 100 081Normals 076(026) 077(016) 068(021) 066(016) 091(014) 083(018) 093(011) 078(021)
similar Asian palace that is not famous (see Figure11)
Results and discussionResults for GR and CO as well as for five age-matched controls are shown in Figure 12 Both
GR and CO could recognise the famous landmarksas well normals In addition both GR and CO wereusually able to name the landmarks they success-fully identified
Thus GR and CO can recognize landmarkslearned prior to injury demonstrating that at leastsome of their place recognition abilities are intactWhen combined with the fact that GR and COwere able to produce detailed maps of places theylived in prior to their injury (see Case Description)these results suggest that their deficit is primarily inencoding or consolidation of new informationrather than in recognition or recall of old informa-tion Note however that these results do not provethat their place recognition abilities are entirelynormal GR and CO might suffer from a mild rec-ognition deficit that affects identification of newly-learned stimuli like the Lego layouts in Experiment3 but not overlearned stimuli like the famous land-marks Furthermore it is possible that GR and COmight be impaired at recognising premorbidlylearned places that are defined primarily by theirspatial layout even though they are unimpaired atrecognising premorbidly learned places that aredefined primarily by a single object-like landmarkor building (see General Discussion) Neverthelessthe trend of the present evidence is to suggest thatGR and CO have particular problems with noveltopographical information
In the famous-face task GRrsquos performance wasin the normal range However he sometimes haddifficulty naming the faces even though he clearly
EPSTEIN ET AL
502 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 11 Stimuli for Experiment 4 Each famous landmark(Arc de Triomphe Taj Mahal) and famous face (Barbara BushYasser Arafat) was matched to a nonfamous building or face thatalso appeared in the experiment
FA
CE
SL
AN
DM
AR
KS
Figure 12 Results of Experiment 4 Performance measure isproportion of famous stimuli reported as ldquoknownrdquo corrected forguessing using the standard formula
recognised them For example he described Ron-ald Reagan as ldquothe gipperrdquo Nancy Reagan as ldquothewife of the gipperrdquo and Clark Gable as ldquothe movieactor who said lsquoScarlett I donrsquot give a damnrsquordquo COhad a great deal of difficulty identifying the famousfaces This result is not surprising given hisextremely low performance on the Benton FaceMatching Task In addition to his topographicproblems CO appears to suffer fromprosopagnosia
GENERAL DISCUSSION
We tested two parahippocampal patients (GR andCO) on four experiments designed to measure per-formance as a function of both task (perceptionencoding and recognition) and stimulus type(scenes objects faces and words) The only taskthat revealed a clear deficit was an n-back recogni-tion memory test (Experiment 3) In order to suc-cessfully perform this task subjects had to encode adurable representation of a visual stimulus and useit for later recognition GR and CO were signifi-cantly more impaired on this task when the stimuliwere scene-like spatial layouts (ie Lego scenesand real scenes for GR) than when they wereobjects No comparable deficits were found in otherexperiments that tested layout perception (Experi-ments 1 and 2) and placelandmark recognition(Experiment 4) Taken together these results indi-cate that GR and CO are impaired in their ability toencode novel information about the geometry ofsurrounding space into memory but are lessimpaired at their ability to encode the geometry ofnovel objects
We argue that this specific topographical defi-citmdashinability to encode the spatial layout of novelscenesmdashresults from damage to the para-hippocampal place area Although the damage inboth patients extends beyond the parahippo-campal-lingual territory normally associated withthe PPA this region is clearly lesioned bilaterally inGR and in the right hemisphere in CO Further-more functional imaging demonstrated that GRhas no functioning PPA Thus the current resultsand the earlier fMRI work (Epstein et al 1999
Epstein amp Kanwisher 1998) converge to implicateparahippocampal cortex in the encoding of infor-mation about scene geometry into memory
Our assignment of the function of memoryencoding to parahippocampal cortex is consistentwith the results of earlier case reports of patientswith parahippocampal damage who have naviga-tional difficulties in novel but not familiar environ-ments (Habib amp Sirigu 1987 Ross 1980 see alsoTeng amp Squire 1999) As with GR and CO thesepatients were either less impaired or unimpaired onnontopographical memory tasks Patients have alsobeen described who exhibit the opposite pattern ofimpairment an inability to learn faces (sometimescalled ldquoprosopamnesiardquo Cipolotti Robinson Blairamp Frith 1999 Tippett Miller amp Farah 2000) orfaces and verbal materials (Maguire amp Cipolotti1998) with an unimpaired ability to learn topo-graphical materials Taken together these resultsargue for a high degree of domain-specificity in theneural systems involved in learning of new informa-tion (Gallistel 1995) The brain may have separatemechanisms for learning faces places and wordswhich can be differentially impaired after damageto specific cortical regions (Cipolotti et al 1999Incisa della Rochetta et al 1996) The presentresults suggest that the anatomical locus of themechanism dedicated to learning new topographi-cal information is in the parahippocampal cortex(Aguirre amp DrsquoEsposito 1999)
In contrast to their impaired ability to encodenew topographical information both GR and COwere able to produce detailed maps of their previousdwellings and performed normally on the famouslandmark recognition test of Experiment 4 Theseresults are generally consistent with Teng andSquirersquos (1999) report of a patient with extensivemedial temporal damage who could recallpremorbidly learned but not postmorbidly learnedtopographical information (see also Ross 1980)Thus our results provide further evidence thatmedial temporal regions such as parahippocampalcortex may be more critical for encoding or consoli-dating new information into memory than for recallof old information
One issue that our experiments do not entirelyresolve is the precise nature of the topographical
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 503
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
information encoded by parahippocampal cortexClearly this region is critical for encoding informa-tion about the geometry of the currently visiblescene even when the ldquoscenerdquo is made out of Legoblocks and therefore has no immediate navigationalrelevance However it is unclear whether this spe-cific deficit can account for all of the real-worldnavigational difficulties encountered by GR andCO GR in particular shows such a catastrophicloss of navigational ability that we suspect he is alsoimpaired at encoding the spatial relationshipsbetweenthe current scene and other locations in theworld CO probably also suffers from such difficul-ties as he reports that he has trouble rememberinghow places relate to each other in large-scale spaceWhen he was tested on a simple 3-room videogameenvironment he not only was unable to rememberthe individual rooms (claiming that there were 10rooms when there were only 3) but he also had nomemory for the spatial relationships between themThese spatial deficits may be the result of damage tothe posterior right hippocampus (in the case of CO)or of disconnection of the posterior right hippo-campus from the parahippocampal regions thatprovide much of its cortical input (in both patientsSuzuki amp Amaral 1994) In either case the abilityto form a cognitive map of new environments likethe lab or videogame space could be impaired(Maguire et al 1998 OrsquoKeefe amp Nadel 1978)However it is also possible that parahippocampalcortex itself may be critical for encoding not onlythe geometry of individual scenes but also the spa-tial relationships between the locations of thosescenes In this case the spatial deficits of GR andCO could be attributed directly to parahippo-campal damage rather than to hippocampal dam-age or disconnection
The latter scenario would be consistent withAguirre and DrsquoEspositorsquos (1999) and Barrash etalrsquos (2000) suggestion that parahippocampal cortexis critically involved in combining visual informa-tion about the appearance of scenes and landmarkswith spatial information about their locations into aunified topographical representation Howeverboth of these authors argue that the representationsof individual scenes (Barrash et al) and landmarks(Aguirre amp DrsquoEsposito) are supported not by
parahippocampal cortex but by more posterior lin-gual-fusiform regions In contrast we believe thatthe neuroimaging data indicate that parahippo-campal cortex is involved in the encoding of thegeometric structure of individual scenes althoughit may also play a role in combining these represen-tations with other spatial representations
Interestingly GR and CO were not impaired onrecognition of famous landmarks that were mostlybuildings Thus our data do not preclude the possi-bility that there may be a separate lingual orfusiform region involved in processing informationabout buildings (Aguirre Zarahn amp DrsquoEsposito1998 Chao Haxby amp Martin 1999 IshaiUngerleider Martin Schouten amp Haxby 1999)which might have been preserved in GR and COdespite extensive damage to other regions of thelingual and fusiform gyrus Alternatively GR andCOrsquos preserved building recognition abilities mightbe mediated by interactions between cortical areasthat perform shape analysis for any object-likestimulus type (Malach et al 1995Kanwisher et al1996) and anterior temporal regions in which long-term topographical and semantic memories arestored (McCarthy et al 1996)
Is perception preserved
In contrast to their inability to encode new scenesinto memory GR and CO have substantiallyretained their ability to perceive the spatial layout ofthe currently visible scene (Experiments 1 and 2)GR can follow a route marked on a floor planwhich indicates a good on-line appreciation of hiscurrent spatial surroundings Furthermore neitherGR or CO have any trouble moving through theenvironmentmdashthey move with confidence and donot bump into doors or walls Problems of the lattertype are usually associated with damage to the pari-etal lobes which are intact in GR and CO Thus atfirst glance GR and CO do not appear to haveperceptual impairments (beyond their visual fieldrestrictions)
However GR does complain of a perceptualdeficit which he describes as an inability to ldquotakeinrdquo scenes when they are too complex Similarreports of an inability to comprehend the
EPSTEIN ET AL
504 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
ldquoglobalityrdquo of a scene were made by some of theparahippocampally-damaged patients studied byHabib and Sirigu (1987) We were not able todetermine the precise nature of this deficit whichmay be partly attributable to his visual field restric-tions It appears that his deficit is more salient in thereal-world environments than in the picture-per-ception tasks of Experiments 1 and 2 perhapsbecause he finds it more difficult to organise visualinput when immersed in it than when viewing pho-tographs that subtend only part of the visual field
We suggest that GR can perceive parts of thescene when he attends to them but may be unableto integrate information over time into a coherentrepresentation of a scene or place The fact that GRand CO are impaired for Lego layouts at 5-backintervals in Experiment 3 is consistent with thishypothesis The time between the first and secondpresentations of a stimulus is very short in this case(and even shorter with 3-back intervals where GRalso shows evidence of a deficit) In both patientsthe representation of layout information formed byperception of a Lego scene must either have anabnormally short persistence be abnormally sus-ceptible to interference or both If so they mightfind it impossible to sustain a spatial frame orschema long enough to use it to organise informa-tion obtained from many different glances at a com-plex scene (Intraub 1997) The result would be aninability to understand the ldquoglobalityrdquo of a sceneeven though all the details were perfectlyperceived
Non-topographical mnemonic abilities
Although not as severe as their topographical mem-ory problems GR and CO do have some memorydifficulties in other domains For example bothperformed poorly on the Rey AVLT whichrequired them to remember a list of unrelatedwords Both also report some subtle episodic mem-ory problems (see Case Description) COrsquos prob-lems could partly result from damage to the righthippocampusmdashindeed CO appeared to have themore acute episodic memory problems HoweverGR has no hippocampal damage so the fact that hedisplays episodic memory problems suggests thatparahippocampal cortex is involved in the forma-
tion of episodic memories (cf Wagner et al1998)
In previous work (Epstein et al 1999) weargued that spatial context information encoded byparahippocampal cortex might play a role in estab-lishing event memories by encoding informationabout where events take place The inability toencode this information might cause problems intasks such as the Rey AVLT where one has to recalla list of unrelated words If spatial (and perhapstemporal) context information is not encoded itwill be harder subsequently to recall the distinctevent of hearing each word (Tulving 1983) Incontrast lack of spatial context information wouldbe less of a problem in tasks where one is onlyrequired to recognise previously encountered verbalmaterials but not to reproduce the spatiotemporalcontext in which they were experienced IndeedGR and CO performed as well as normals on thenonsense words in Experiment 4 where recogni-tion memory but not episodic memory was requiredfor successful performance Interestingly GR andCO also did relatively well on the Logical Memorysubtest of the WMS-III perhaps because the nar-rative framework of a story provides sufficient con-textual information to overcome the absence ofspatial context and mediate successful verbalretrieval
Differences between GR and CO
Although we have discussed GR and CO togetherthere are a number of relevant differences betweenthem One open question is whether these differ-ences can be attributed to the different effects ofunilateral and bilateral parahippocampal damageIn general GRrsquos topographical difficulties weremore severe than COrsquos CO did surprisingly well onthe Warrington Topographical Memory Test inwhich he was required to remember detailed real-world city scenes and also performed substantiallybetter on real scenes than Lego scenes in Experi-ment 3 Insofar as CO has intact left para-hippocampal cortex this suggests that the left PPAmight be sufficient to encode a certain amount of(perhaps semantic) information about real-worldscenes but the right PPA is necessary to encode
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 505
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)

(FFA) commonly seen in normal subjects(Kanwisher McDermott amp Chun 1997) Thusthe MRI was sensitive enough to detect functionaldifferences an FFA was found but no PPA Whencombined with the anatomical results this stronglysuggests that GR has no PPA
Because of safety concerns (presence of a femo-ral artery aneurysm clip) we did not perform a simi-lar high-field functional MRI of CO However asFigure 4 demonstrates CO clearly has damage tothe parahippocampal-lingual region in the righthemisphere
EXPERIMENTAL INVESTIGATIONS
In this section we report the results of four experi-ments designed to probe the consequences of dam-age to the lingual-parahippocampal region ingeneral and the PPA in particular Our primaryconcern in these experiments was to test GR andCOrsquos ability to perceive remember and recognisedifferent kinds of visual material Based on our pre-vious fMRI work we predicted that GR and CO
would have more difficulty with scene-like stimuli(ie stimuli that convey information about thegeometry of a space one can navigate through or actwithin) than with object-like stimuli Such a deficitcould account for at least some of the problems GRand CO encounter in real-world navigation
Beyond determining the kind of visual materialthat gets processed in parahippocampal cortex wealso wanted to determine the nature of the process-ing that takes place on that material In particularwe wanted to determine if parahippocampal cortexis involved in perceptual encoding memory encodingor recognition As discussed earlier (Introduction)our fMRI results provided some evidence againstplace recognition as the sole function of the PPAbut did not allow us to distinguish between percep-tual and mnemonic encoding Experiments 1 and 2examine the role of parahippocampal cortex in per-ceptual encoding Experiment 3 examines its role inmnemonic encoding and Experiment 4 examinesits role in place recognition
Experiment 1 Two-dot task
The first hypothesis we examined was thatparahippocampal cortex plays a role in perceivingthe shape of the local environment In our previousfMRI work we observed that the PPA respondsmuch more strongly during scene viewing than dur-ing object or face viewing even when subjects donothing more than watch the scenes passively with-out performing any other task This suggested thatscene perception alone may be sufficient to activatethe PPA Previous reports have indicated that dam-age to parahippocampal cortex and other medialtemporal regions leads to problems with memoryrather than perception (Bohbot et al 1998Maguire Burke Philips amp Staunton 1996 Pigottamp Milner 1993) However all of these previousresults were obtained on patients who had unilat-eral lesions GR has a bilateral occipital-temporallesion including clear damage to parahippocampalcortex in both hemispheres furthermore he reportssome kind of perceptual deficit We hypothesisedthat this deficit might involve difficulty perceivingor interpreting the geometry of scenes and otherstimuli with a similar geometric structure
EPSTEIN ET AL
494 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 5 Top row shows a single slice from a functional MRIscan performed on GR Bottom row shows results from a normalsubject (NK) for comparison Voxels that responded more to facesthan objects (left column) or more to scenes than faces and objects(right column) in a Kolomogorov-Smirnov test are highlighted bywhite arrows GR has a functioning face area (FFA) but there isno evidence of a place area (PPA)
FFA PPA
GR
NK
In this experiment subjects viewed either objectshapes or spatial layouts Stimuli consisted ofblack-and-white photographs of real-world stimuli(empty rooms vs common objects) or abstractstimuli made out of Lego blocks (Lego ldquoscenesrdquo vsLego ldquoobjectsrdquo) These stimuli have been previouslyshown to differentially activate the PPA in normalsubjects (Epstein et al 1999 Epstein ampKanwisher 1998) Two red dots were superim-posed on each photograph (Figure 6) The task wasto report which of the two red dots was over thecloser part of the object or scene This task wasdesigned to have no memory component (apartfrom remembering the instructions) but to requireaccurate 3D perception of objects and scenes
If GR or CO have difficulty perceiving or inter-preting the spatial layout of scenes then we pre-dicted they would do worse when the stimuli areempty rooms or Lego scenes than when they arereal-world or Lego objects Note that the criticalcomparison is between the Lego scenes and theLego objects as these stimuli are made of the samematerials and differ primarily in their surfacegeometry
MethodTesting for all experiments was done on aMacintosh computer The experiment consisted of80 trials (20 of each stimulus type randomly inter-mixed) Subjects began each trial by pressing thespace bar A black-and-white photograph from oneof the four stimulus categories (see earlier) thenappeared on the screen with two red dots on it Thetask was to determine which of the two red dotsindicated the closer part of the scene or objectSubjects pressed the ldquofrdquo key if the leftmost red dotwas closest and the ldquojrdquo key if the rightmost dotwas closest They were instructed to respond asquickly as possible without making errors Stimuliremained on the screen until a response was made
The stimulus set was constructed so that eachempty room was matched to a common object thathad red dots in the exact same location and eachLego layout was similarly matched to a Lego objectThus the locations of the red dots were counterbal-anced across scene and object stimuli controllingfor any effects of visual field cuts
Results and discussionResults for GR and CO and four elderly normalsubjects are shown in Table 3 Both GR and COcould do this task without difficulty and their per-formance was near ceiling There was no evidencethat they had any more trouble with the emptyrooms or Lego scenes than with the real or Legoobjects Reaction times were slightly longer for GRand CO than for normal subjects as one wouldexpect given their visual field deficits however this
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 495
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 3 Experiment 1 (two-dot) results (SD in parenthesis)
GR CO Normals
Proportion correctObject 095 080 093Lego object 100 095 098Empty room 100 100 099Lego room 095 100 096
Mean RTObject 2435 2932 2161 (980)Lego object 2052 3245 2029 (921)Empty room 2744 2291 1788 (959)Lego room 1904 2281 1630 (1127)
Figure 6 The four kinds of stimuli used in Experiment 1 Topleft lego layouts top right real-world layouts (empty rooms)bottom left lego objects bottom right real-world objects
difference did not appear to be significant Impor-tantly there was little evidence that GR or CO tooklonger to make their perceptual judgements for thereal or Lego scenes than for the real or Lego objectsThus contrary to our initial hypothesis this experi-ment found no clear evidence for selective impair-ment in the perception of scene-like stimuli
Experiment 2 Different-views matchingtask
The previous experiment demonstrated that GRand CO can extract information about the per-ceived distances between themselves and the sur-faces of an object or scene Thus in at least onesense their perception of these surfaces is pre-served However this does not mean that their abil-ity to represent the layout of a scene is unimpairedThe two-dot task requires on-line comparison oftwo surfaces in egocentric coordinates GR and COwould be able to perform this task even if they couldnot (1) represent more than two surfaces at a time(2) represent surfaces in other than strictly egocen-tric coordinates or (3) sustain surface representa-tions for longer than the time it takes to make acomparison It has been suggested that the parietallobes are involved in on-line processing of surfacegeometry in egocentric coordinates (Milner ampGoodale 1995) thus it is possible that the parietallobes (which are intact in GR and CO) may be suf-ficient to perform the two-dot task even in theabsence of more durable or complex surface repre-sentations in parahippocampal cortex
In Experiment 2 we examine GR and COrsquosability to create and sustain a representation of thegeometry of an object or scene-like layout The taskwas to view a photograph of a Lego object or Legoscene hold it in memory for 10 s and then deter-mine whether a second object or scene was the sameas the first In ldquosamerdquo trials the second photographdepicted the same scene or object depicted from adifferent viewing angle In ldquodifferentrdquo trials the sec-ond photograph depicted a different scene or objectchosen to be similar to the first one (Figure 7)Thus in order to successfully perform this tasksubjects had to form a representation of the surfacesof the object or scene sustain this representation in
memory for 10 s and then align it (presumablyusing mental rotation) to the second object orscene
MethodThe experiment consisted of two parts each with40 trials In the first part the stimuli were black andwhite photographs of objects made out of Legoblocks In the second part the stimuli were photo-graphs of ldquoscenesrdquo made out of Lego blocks whichwere designed to have a geometric structure similarto real-world places such as a roomor a city street
Subjects began each trial by pressing the spacebar A photograph of a Lego object or sceneappeared on the screen for 3 s followed by a 10-sblank interval A second photograph then appeareddepicting either (1) the same object or scene as thefirst photograph shown from a different viewingangle (offset approximately 35 degrees) or (2) a dif-ferent object or scene Subjects were instructed topress the ldquojrdquo key if the two photographs depicted thesame object or scene and the ldquof rdquo key if they did notThey were told to take as much time as they neededto make a decision but no extra time The secondphotograph remained on the screen until a responsewas made Half the trials were ldquosamerdquo trials and halfwere ldquodifferentrdquo trials
Results and discussionFigure 8 shows results for GR and CO as well as forsix elderly normal controls GRrsquos and COrsquos perfor-mance was comparable to that of the normal con-trols Furthermore GR and CO performed just aswell when the stimuli were Lego scenes as whenthey were Lego objects These results are unlikely toreflect a ceiling effect as performance for bothpatients and controls was around 80 correctThus both patients appear to be able to form a rep-resentation of the geometry of both scenes andobjects which they can sustain in short-termmemory
Experiment 3 N-back recognition memorytask
The previous experiment failed to find any impair-ment in GR and COrsquos ability to form a representa-
EPSTEIN ET AL
496 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
tion of the geometry of a scene or object andmaintain it for 10 s in the absence of the originalstimulus Not only does this result suggest that per-ception of scene-like stimuli is intact in thesepatients it further suggests that some short-termmemory functions are intact as well However we
should note that subjects in this experiment werefree to devote all their effort and attention in the 10-second blank interval to maintaining a memory ofthe stimulus Thus it is possible that GR and COcan use rehearsal mechanisms to maintain the stim-ulus in short-term memory during the blank inter-
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 497
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
(b) Lego object version
Figure 7 Stimuli and procedure for Experiment 2 (a) In the Lego layout version subjects see a Lego layout for 3 s followed by a blankscreen for 10 s A second layout then appears which is either the same layout as the first one but seen from a different angle or a differentlayout (b) The Lego object version is the same but with object stimuli
3 s 10 s OR
3 s 10 s OR
(a) Lego layout version
(b) Lego object version
val but are unable to form a more durable memorytrace for the stimulus This possibility was tested inthe current experiment which was designed toexamine GR and COrsquos ability to learn new visualinformation
We devised a continuous n-back recognitionmemory test in which subjects viewed a sequence oforiginally novel stimuli some of which wererepeated within the sequence at intervals of 1 3 or5 intervening images For each stimulus subjects
reported whether they had seen it before in theexperiment or not In different runs in the experi-ment the stimuli were black-and-white photo-graphs of (1) Lego scenes (2) Lego objects (3) real-world landscapes (4) novel objects made out ofSculpey clay (5) unfamiliar houses (6) unfamiliarfaces as well as (7) line drawings of nonsenseobjects and (8) pronounceable pseudowords (Fig-ure 9) Thus we were able to test GR and COrsquos rec-ognition memory for different kinds of materials atdifferent time intervals in a paradigm in which thepresence of intervening items between the twoappearances of each stimulus made rehearsal diffi-cult Note that because none of the specific stimuliused in this experiment were familiar in advancethis task did not require an episodic memory com-ponent and could be done on the basis of pure visualfamiliarity with the particular visual stimulus
Although many different stimulus classes weretested in this experiment the critical comparison isbetween the Lego layouts and Lego objectsbecause these are best matched with regard tomaterials and differ primarily in their geometricstructure (see Introduction) We predicted that ifGR and CO showed a memory deficit for this taskit would be greater for the Lego layouts than for the
EPSTEIN ET AL
498 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 8 Results of Experiment 2 Performance over ldquosamerdquotrials and ldquodifferentrdquo trials is averaged together Error bars show1 SD
Figure 9 Stimuli for Experiment 3 Top row Lego layouts real-world scenes houses Bottom row Lego objects clay objects nonsenseobjects faces In an eighth condition (not shown) subjects viewed pronounceable nonwords such as ldquowilchrdquo or ldquothipperrdquo
Lego objects In addition a deficit for materialsthat convey information about the layout of sur-rounding space might also be apparent when GRand CO are tested on more realistic stimuli If soone would expect them to do poorly on the land-scapes but relatively well on the clay objects linedrawings of nonsense objects faces andpseudowords
MethodFor each subject a complete experimental sessionconsisted of being run once on each of the eightstimulus classes Each run consisted of 90 trials ineach trial a stimulus was presented and subjectsreported whether or not they had seen it before inthe run (or indeed ever) by pressing the ldquojrdquo key ifthey had or the ldquof rdquo key if they had not Each stimu-lus was on the screen for either 1 2 or 3 s (see fol-lowing) and appeared promptly after the keypressreport for the previous trial Individual stimuliappeared either once (14 stimuli) twice (30 stim-uli) or four times (4 stimuli) within a given run Ofthe stimuli that were presented twice 10 wererepeated immediately (1-back) 10 were repeatedwith a lag of two intervening items (3-back) and 10were repeated with a lag of four intervening items(5-back) In order to ensure that between-subjectdifferences in performance were not due to stimulusdifferences the same stimuli were assigned to thesame conditions each time the experimentwas run
GR performed the experiment twice once inAugust 1998 and again 9 months later Because ofa computer error he was only tested on six of theeight categories (all except Lego layouts and Legoobjects) at the later date CO was also tested twiceonce in February 1999 and again 6 months laterReported results for GR and CO are the average oftheir two testing sessions Normal subjects wereonly tested once Stimulus presentation time was 1 sin GRrsquos first testing session In pilot testing COfound this too fast for the critical categories of Lego
layouts and Lego objects so presentation time wasextended to 2 s for these categories in his first test-ing session All normal subjects were tested at thesepresentation rates (2 s for Lego layouts and objects1 s for other stimuli) In COrsquos second testingsession presentation time was lengthened to 3 s forall stimulus categories to ensure that any deficitsfound were memory deficits rather than problemswith the nonmemory aspects of the task
Results and discussionResults from GR CO and 10 elderly normal con-trols are shown in Figure 10 and Table 4 Figure 10ashows results from the main comparison betweenLego layouts and Lego objects Both normal sub-jects and patients performed near ceiling for bothLego layouts and Lego objects at 1-backinterstimulus intervals verifying that GR and COcould successfully perceive the pictures as they werebeing presented In contrast there was a strikingdifference between the two patients and the normalsubjects at 5-back intervals Whereas normals didon average somewhat better on Lego layouts thanLego objects both GR and CO showed the oppo-site pattern doing much worse on Lego layoutsthan Lego objects The 3-back condition showed amixed pattern Whereas GR did better on Legoobjects than Lego layouts in this condition (similarto his performance at 5-back intervals) CO didbetter on Lego layouts than Lego objects COrsquos lowperformance on Lego objects in the 3-back condi-tion appears to be an anomaly as it is much lowerthan his performance for comparable stimuli in the1-back and 5-back conditions
Figure 10b shows the difference in performancebetween Lego layouts and Lego objects for thecritical 1-back and 5-back intervals At 5-backintervals CO was significantly impaired comparedto the normal subjects (z = ndash250) whereasGRrsquos impairment fell just short of significance (z =ndash175)4
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 499
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
4In fact these values may underestimate GR and COrsquos relative impairment for Lego layouts In the 5-back condition 4 of the 10
normal subjects had perfect performance on the Lego layouts but all scored below ceiling on the Lego objects Thus the true advantagefor Lego layouts over Lego objects for normal subjects is probably greater than that observed here in which case GR and COrsquos lowerperformance for Lego layouts compared to Lego objects would be even more abnormal Furthermore post-hoc analyses revealed thatone of the normal subjects showed a difference in performance between Lego layouts and objects that was over 2 SD below the mean ofthe other nine subjects When this outlying subject was excluded z-scores were ndash322 for CO and ndash232 for GR
Table 4 shows results for all eight stimulusclasses Although direct comparisons between sub-jects should be treated with caution since viewingtime for different items was not the same for eachsubject we do note a few interesting patterns Firstof all there is little or no evidence that GR or CO
show a memory deficit for nonsense words clayobjects or line drawings of nonsense objects GRappears to be slightly impaired on faces and housesbut CO performs normally on these categoriesStrikingly GR shows a severe deficit for the reallandscapes performing more than 40 percentagepoints lower than normals Thus even thoughthere is some evidence that GR might have someimpairment for nonscene stimuli he clearly is muchmore impaired for scene-like stimuli such as theLego scenes and landscapes CO does not show asimilar deficit for the landscapes However itshould be kept in mind that in one of his two testingsessions CO viewed each picture for 3 s which wasmuch longer than the 1 second used for all non-Lego stimuli for GR and normals Thus COrsquosapparently ldquonormalrdquo performance in these condi-tions might be overestimated
Taken together the results of this experimentindicate that GR and CO are selectively impaired atprocessing scene-like stimuli This deficit is consis-tently apparent at 5-back but not 1-back intervalsThis finding suggests that the deficit should becharacterised as a memory encoding problemHowever the present results do not allow us todetermine whether the difficulty occurs at memoryencoding or at subsequent recognition (see Epsteinamp Kanwisher 2001) If GR and CO have a generalproblem with recognition of topographical materi-als then they should not be able to recognise scenes
EPSTEIN ET AL
500 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 10a Results of Experiment 3 for GR (top) CO (middle)and for 10 age-matched normal controls Performance measure isproportion of repeated stimuli correctly identified as such correctedfor guessing using the standard formula
Figure 10b Difference between Lego layout performance andLego object performance for the 1-back and 5-back conditionsError bars indicate one 1 SD
and landmarks that were learned prior to theirinjury We tested this possibility in the nextexperiment
Experiment 4 Famous landmarksfacesrecognition task
In this experiment we examined GR and COrsquosability to recognise the visual appearances of placeslearned prior to injury Subjects saw a series offamous landmarks intermixed with nonfamouscontrols (Imaging experiments have demonstratedthat landmarks of this type can activate the PPAalmost as strongly as complete scenes Epstein etal 1999 Epstein amp Kanwisher 1998) They wererequired to say whether they knew each specificlandmark and to identify the ones they knew Sub-jects were instructed to respond affirmatively if theywere fairly certain that they recognised the land-mark but not if it only looked generally familiarFor purposes of comparison subjects also per-formed an identical test in which the stimuli werefamous and nonfamous faces In both testsdistractor items were designed to be visually andconceptually similar to the target items (see Figure11) If parahippocampal cortex is required for placerecognition then one might expect GR and CO toperform poorly compared to normals on the land-mark recognition test
MethodEach of the two experiments (famous faces famouslandmarks) consisted of 48 trials Subjects beganeach trial by pressing the space bar after which ablack-and-white photograph of a face (in thefamous faces experiment) or a landmark (in thefamous landmarks experiment) appeared on thescreen Subjects were told to press the ldquojrdquo key if theyrecognised the specific face or landmark and the ldquof rdquokey if they did not taking as much time at theyneeded to make this discrimination but no extratime Subjects were told to report that they recog-nised a face or landmark if they felt that they knewit even if they could not retrieve its name If theydid know the name they were to say it after pressingthe button Otherwise they were to say whateverthey did remember about the item
Half of the photographs were of famous and halfof nonfamous people and landmarks The famousfaces included public figures (eg George BushRichard Nixon Prince Charles) and movie stars(eg Harrison Ford Clark Gable Marlon Brando)who were prominent prior to the time of GR andCOrsquos injuries the famous landmarks were build-ings and monuments from all over the world (egEiffel Tower US Capitol Building Great Sphinx)The nonfamous stimuli were chosen to match thefamous stimuli as closely as possible Each famousstimulus had a ldquomatchrdquo For example George Bushwas matched to a man of similar age in a coat andtie and the Taj Mahal was matched to a visually
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 501
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 4 Full results of Experiment 3 Mean proportion correct (corrected for guessing) SDs in parentheses
Lego Land- Lego Clay Nonsense Non-scenes scapes Houses objects objects objects Words Faces
1-back GR 100 094 100 100 100 094 100 100CO 091 085 091 100 100 079 074 100Normals 096(008) 095(008) 093(014) 096(008) 096(006) 099(003) 098(004) 097(011)
3-back GR ndash003 068 026 047 083 083 075 052CO 066 081 037 010 094 069 095 081Normals 066(032) 077(021) 072(015) 077(020) 086(016) 083(015) 083(017) 072(020)
5-back GR 010 034 044 047 072 083 094 053CO 031 073 075 088 081 064 100 081Normals 076(026) 077(016) 068(021) 066(016) 091(014) 083(018) 093(011) 078(021)
similar Asian palace that is not famous (see Figure11)
Results and discussionResults for GR and CO as well as for five age-matched controls are shown in Figure 12 Both
GR and CO could recognise the famous landmarksas well normals In addition both GR and CO wereusually able to name the landmarks they success-fully identified
Thus GR and CO can recognize landmarkslearned prior to injury demonstrating that at leastsome of their place recognition abilities are intactWhen combined with the fact that GR and COwere able to produce detailed maps of places theylived in prior to their injury (see Case Description)these results suggest that their deficit is primarily inencoding or consolidation of new informationrather than in recognition or recall of old informa-tion Note however that these results do not provethat their place recognition abilities are entirelynormal GR and CO might suffer from a mild rec-ognition deficit that affects identification of newly-learned stimuli like the Lego layouts in Experiment3 but not overlearned stimuli like the famous land-marks Furthermore it is possible that GR and COmight be impaired at recognising premorbidlylearned places that are defined primarily by theirspatial layout even though they are unimpaired atrecognising premorbidly learned places that aredefined primarily by a single object-like landmarkor building (see General Discussion) Neverthelessthe trend of the present evidence is to suggest thatGR and CO have particular problems with noveltopographical information
In the famous-face task GRrsquos performance wasin the normal range However he sometimes haddifficulty naming the faces even though he clearly
EPSTEIN ET AL
502 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 11 Stimuli for Experiment 4 Each famous landmark(Arc de Triomphe Taj Mahal) and famous face (Barbara BushYasser Arafat) was matched to a nonfamous building or face thatalso appeared in the experiment
FA
CE
SL
AN
DM
AR
KS
Figure 12 Results of Experiment 4 Performance measure isproportion of famous stimuli reported as ldquoknownrdquo corrected forguessing using the standard formula
recognised them For example he described Ron-ald Reagan as ldquothe gipperrdquo Nancy Reagan as ldquothewife of the gipperrdquo and Clark Gable as ldquothe movieactor who said lsquoScarlett I donrsquot give a damnrsquordquo COhad a great deal of difficulty identifying the famousfaces This result is not surprising given hisextremely low performance on the Benton FaceMatching Task In addition to his topographicproblems CO appears to suffer fromprosopagnosia
GENERAL DISCUSSION
We tested two parahippocampal patients (GR andCO) on four experiments designed to measure per-formance as a function of both task (perceptionencoding and recognition) and stimulus type(scenes objects faces and words) The only taskthat revealed a clear deficit was an n-back recogni-tion memory test (Experiment 3) In order to suc-cessfully perform this task subjects had to encode adurable representation of a visual stimulus and useit for later recognition GR and CO were signifi-cantly more impaired on this task when the stimuliwere scene-like spatial layouts (ie Lego scenesand real scenes for GR) than when they wereobjects No comparable deficits were found in otherexperiments that tested layout perception (Experi-ments 1 and 2) and placelandmark recognition(Experiment 4) Taken together these results indi-cate that GR and CO are impaired in their ability toencode novel information about the geometry ofsurrounding space into memory but are lessimpaired at their ability to encode the geometry ofnovel objects
We argue that this specific topographical defi-citmdashinability to encode the spatial layout of novelscenesmdashresults from damage to the para-hippocampal place area Although the damage inboth patients extends beyond the parahippo-campal-lingual territory normally associated withthe PPA this region is clearly lesioned bilaterally inGR and in the right hemisphere in CO Further-more functional imaging demonstrated that GRhas no functioning PPA Thus the current resultsand the earlier fMRI work (Epstein et al 1999
Epstein amp Kanwisher 1998) converge to implicateparahippocampal cortex in the encoding of infor-mation about scene geometry into memory
Our assignment of the function of memoryencoding to parahippocampal cortex is consistentwith the results of earlier case reports of patientswith parahippocampal damage who have naviga-tional difficulties in novel but not familiar environ-ments (Habib amp Sirigu 1987 Ross 1980 see alsoTeng amp Squire 1999) As with GR and CO thesepatients were either less impaired or unimpaired onnontopographical memory tasks Patients have alsobeen described who exhibit the opposite pattern ofimpairment an inability to learn faces (sometimescalled ldquoprosopamnesiardquo Cipolotti Robinson Blairamp Frith 1999 Tippett Miller amp Farah 2000) orfaces and verbal materials (Maguire amp Cipolotti1998) with an unimpaired ability to learn topo-graphical materials Taken together these resultsargue for a high degree of domain-specificity in theneural systems involved in learning of new informa-tion (Gallistel 1995) The brain may have separatemechanisms for learning faces places and wordswhich can be differentially impaired after damageto specific cortical regions (Cipolotti et al 1999Incisa della Rochetta et al 1996) The presentresults suggest that the anatomical locus of themechanism dedicated to learning new topographi-cal information is in the parahippocampal cortex(Aguirre amp DrsquoEsposito 1999)
In contrast to their impaired ability to encodenew topographical information both GR and COwere able to produce detailed maps of their previousdwellings and performed normally on the famouslandmark recognition test of Experiment 4 Theseresults are generally consistent with Teng andSquirersquos (1999) report of a patient with extensivemedial temporal damage who could recallpremorbidly learned but not postmorbidly learnedtopographical information (see also Ross 1980)Thus our results provide further evidence thatmedial temporal regions such as parahippocampalcortex may be more critical for encoding or consoli-dating new information into memory than for recallof old information
One issue that our experiments do not entirelyresolve is the precise nature of the topographical
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 503
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
information encoded by parahippocampal cortexClearly this region is critical for encoding informa-tion about the geometry of the currently visiblescene even when the ldquoscenerdquo is made out of Legoblocks and therefore has no immediate navigationalrelevance However it is unclear whether this spe-cific deficit can account for all of the real-worldnavigational difficulties encountered by GR andCO GR in particular shows such a catastrophicloss of navigational ability that we suspect he is alsoimpaired at encoding the spatial relationshipsbetweenthe current scene and other locations in theworld CO probably also suffers from such difficul-ties as he reports that he has trouble rememberinghow places relate to each other in large-scale spaceWhen he was tested on a simple 3-room videogameenvironment he not only was unable to rememberthe individual rooms (claiming that there were 10rooms when there were only 3) but he also had nomemory for the spatial relationships between themThese spatial deficits may be the result of damage tothe posterior right hippocampus (in the case of CO)or of disconnection of the posterior right hippo-campus from the parahippocampal regions thatprovide much of its cortical input (in both patientsSuzuki amp Amaral 1994) In either case the abilityto form a cognitive map of new environments likethe lab or videogame space could be impaired(Maguire et al 1998 OrsquoKeefe amp Nadel 1978)However it is also possible that parahippocampalcortex itself may be critical for encoding not onlythe geometry of individual scenes but also the spa-tial relationships between the locations of thosescenes In this case the spatial deficits of GR andCO could be attributed directly to parahippo-campal damage rather than to hippocampal dam-age or disconnection
The latter scenario would be consistent withAguirre and DrsquoEspositorsquos (1999) and Barrash etalrsquos (2000) suggestion that parahippocampal cortexis critically involved in combining visual informa-tion about the appearance of scenes and landmarkswith spatial information about their locations into aunified topographical representation Howeverboth of these authors argue that the representationsof individual scenes (Barrash et al) and landmarks(Aguirre amp DrsquoEsposito) are supported not by
parahippocampal cortex but by more posterior lin-gual-fusiform regions In contrast we believe thatthe neuroimaging data indicate that parahippo-campal cortex is involved in the encoding of thegeometric structure of individual scenes althoughit may also play a role in combining these represen-tations with other spatial representations
Interestingly GR and CO were not impaired onrecognition of famous landmarks that were mostlybuildings Thus our data do not preclude the possi-bility that there may be a separate lingual orfusiform region involved in processing informationabout buildings (Aguirre Zarahn amp DrsquoEsposito1998 Chao Haxby amp Martin 1999 IshaiUngerleider Martin Schouten amp Haxby 1999)which might have been preserved in GR and COdespite extensive damage to other regions of thelingual and fusiform gyrus Alternatively GR andCOrsquos preserved building recognition abilities mightbe mediated by interactions between cortical areasthat perform shape analysis for any object-likestimulus type (Malach et al 1995Kanwisher et al1996) and anterior temporal regions in which long-term topographical and semantic memories arestored (McCarthy et al 1996)
Is perception preserved
In contrast to their inability to encode new scenesinto memory GR and CO have substantiallyretained their ability to perceive the spatial layout ofthe currently visible scene (Experiments 1 and 2)GR can follow a route marked on a floor planwhich indicates a good on-line appreciation of hiscurrent spatial surroundings Furthermore neitherGR or CO have any trouble moving through theenvironmentmdashthey move with confidence and donot bump into doors or walls Problems of the lattertype are usually associated with damage to the pari-etal lobes which are intact in GR and CO Thus atfirst glance GR and CO do not appear to haveperceptual impairments (beyond their visual fieldrestrictions)
However GR does complain of a perceptualdeficit which he describes as an inability to ldquotakeinrdquo scenes when they are too complex Similarreports of an inability to comprehend the
EPSTEIN ET AL
504 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
ldquoglobalityrdquo of a scene were made by some of theparahippocampally-damaged patients studied byHabib and Sirigu (1987) We were not able todetermine the precise nature of this deficit whichmay be partly attributable to his visual field restric-tions It appears that his deficit is more salient in thereal-world environments than in the picture-per-ception tasks of Experiments 1 and 2 perhapsbecause he finds it more difficult to organise visualinput when immersed in it than when viewing pho-tographs that subtend only part of the visual field
We suggest that GR can perceive parts of thescene when he attends to them but may be unableto integrate information over time into a coherentrepresentation of a scene or place The fact that GRand CO are impaired for Lego layouts at 5-backintervals in Experiment 3 is consistent with thishypothesis The time between the first and secondpresentations of a stimulus is very short in this case(and even shorter with 3-back intervals where GRalso shows evidence of a deficit) In both patientsthe representation of layout information formed byperception of a Lego scene must either have anabnormally short persistence be abnormally sus-ceptible to interference or both If so they mightfind it impossible to sustain a spatial frame orschema long enough to use it to organise informa-tion obtained from many different glances at a com-plex scene (Intraub 1997) The result would be aninability to understand the ldquoglobalityrdquo of a sceneeven though all the details were perfectlyperceived
Non-topographical mnemonic abilities
Although not as severe as their topographical mem-ory problems GR and CO do have some memorydifficulties in other domains For example bothperformed poorly on the Rey AVLT whichrequired them to remember a list of unrelatedwords Both also report some subtle episodic mem-ory problems (see Case Description) COrsquos prob-lems could partly result from damage to the righthippocampusmdashindeed CO appeared to have themore acute episodic memory problems HoweverGR has no hippocampal damage so the fact that hedisplays episodic memory problems suggests thatparahippocampal cortex is involved in the forma-
tion of episodic memories (cf Wagner et al1998)
In previous work (Epstein et al 1999) weargued that spatial context information encoded byparahippocampal cortex might play a role in estab-lishing event memories by encoding informationabout where events take place The inability toencode this information might cause problems intasks such as the Rey AVLT where one has to recalla list of unrelated words If spatial (and perhapstemporal) context information is not encoded itwill be harder subsequently to recall the distinctevent of hearing each word (Tulving 1983) Incontrast lack of spatial context information wouldbe less of a problem in tasks where one is onlyrequired to recognise previously encountered verbalmaterials but not to reproduce the spatiotemporalcontext in which they were experienced IndeedGR and CO performed as well as normals on thenonsense words in Experiment 4 where recogni-tion memory but not episodic memory was requiredfor successful performance Interestingly GR andCO also did relatively well on the Logical Memorysubtest of the WMS-III perhaps because the nar-rative framework of a story provides sufficient con-textual information to overcome the absence ofspatial context and mediate successful verbalretrieval
Differences between GR and CO
Although we have discussed GR and CO togetherthere are a number of relevant differences betweenthem One open question is whether these differ-ences can be attributed to the different effects ofunilateral and bilateral parahippocampal damageIn general GRrsquos topographical difficulties weremore severe than COrsquos CO did surprisingly well onthe Warrington Topographical Memory Test inwhich he was required to remember detailed real-world city scenes and also performed substantiallybetter on real scenes than Lego scenes in Experi-ment 3 Insofar as CO has intact left para-hippocampal cortex this suggests that the left PPAmight be sufficient to encode a certain amount of(perhaps semantic) information about real-worldscenes but the right PPA is necessary to encode
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 505
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)

In this experiment subjects viewed either objectshapes or spatial layouts Stimuli consisted ofblack-and-white photographs of real-world stimuli(empty rooms vs common objects) or abstractstimuli made out of Lego blocks (Lego ldquoscenesrdquo vsLego ldquoobjectsrdquo) These stimuli have been previouslyshown to differentially activate the PPA in normalsubjects (Epstein et al 1999 Epstein ampKanwisher 1998) Two red dots were superim-posed on each photograph (Figure 6) The task wasto report which of the two red dots was over thecloser part of the object or scene This task wasdesigned to have no memory component (apartfrom remembering the instructions) but to requireaccurate 3D perception of objects and scenes
If GR or CO have difficulty perceiving or inter-preting the spatial layout of scenes then we pre-dicted they would do worse when the stimuli areempty rooms or Lego scenes than when they arereal-world or Lego objects Note that the criticalcomparison is between the Lego scenes and theLego objects as these stimuli are made of the samematerials and differ primarily in their surfacegeometry
MethodTesting for all experiments was done on aMacintosh computer The experiment consisted of80 trials (20 of each stimulus type randomly inter-mixed) Subjects began each trial by pressing thespace bar A black-and-white photograph from oneof the four stimulus categories (see earlier) thenappeared on the screen with two red dots on it Thetask was to determine which of the two red dotsindicated the closer part of the scene or objectSubjects pressed the ldquofrdquo key if the leftmost red dotwas closest and the ldquojrdquo key if the rightmost dotwas closest They were instructed to respond asquickly as possible without making errors Stimuliremained on the screen until a response was made
The stimulus set was constructed so that eachempty room was matched to a common object thathad red dots in the exact same location and eachLego layout was similarly matched to a Lego objectThus the locations of the red dots were counterbal-anced across scene and object stimuli controllingfor any effects of visual field cuts
Results and discussionResults for GR and CO and four elderly normalsubjects are shown in Table 3 Both GR and COcould do this task without difficulty and their per-formance was near ceiling There was no evidencethat they had any more trouble with the emptyrooms or Lego scenes than with the real or Legoobjects Reaction times were slightly longer for GRand CO than for normal subjects as one wouldexpect given their visual field deficits however this
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 495
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 3 Experiment 1 (two-dot) results (SD in parenthesis)
GR CO Normals
Proportion correctObject 095 080 093Lego object 100 095 098Empty room 100 100 099Lego room 095 100 096
Mean RTObject 2435 2932 2161 (980)Lego object 2052 3245 2029 (921)Empty room 2744 2291 1788 (959)Lego room 1904 2281 1630 (1127)
Figure 6 The four kinds of stimuli used in Experiment 1 Topleft lego layouts top right real-world layouts (empty rooms)bottom left lego objects bottom right real-world objects
difference did not appear to be significant Impor-tantly there was little evidence that GR or CO tooklonger to make their perceptual judgements for thereal or Lego scenes than for the real or Lego objectsThus contrary to our initial hypothesis this experi-ment found no clear evidence for selective impair-ment in the perception of scene-like stimuli
Experiment 2 Different-views matchingtask
The previous experiment demonstrated that GRand CO can extract information about the per-ceived distances between themselves and the sur-faces of an object or scene Thus in at least onesense their perception of these surfaces is pre-served However this does not mean that their abil-ity to represent the layout of a scene is unimpairedThe two-dot task requires on-line comparison oftwo surfaces in egocentric coordinates GR and COwould be able to perform this task even if they couldnot (1) represent more than two surfaces at a time(2) represent surfaces in other than strictly egocen-tric coordinates or (3) sustain surface representa-tions for longer than the time it takes to make acomparison It has been suggested that the parietallobes are involved in on-line processing of surfacegeometry in egocentric coordinates (Milner ampGoodale 1995) thus it is possible that the parietallobes (which are intact in GR and CO) may be suf-ficient to perform the two-dot task even in theabsence of more durable or complex surface repre-sentations in parahippocampal cortex
In Experiment 2 we examine GR and COrsquosability to create and sustain a representation of thegeometry of an object or scene-like layout The taskwas to view a photograph of a Lego object or Legoscene hold it in memory for 10 s and then deter-mine whether a second object or scene was the sameas the first In ldquosamerdquo trials the second photographdepicted the same scene or object depicted from adifferent viewing angle In ldquodifferentrdquo trials the sec-ond photograph depicted a different scene or objectchosen to be similar to the first one (Figure 7)Thus in order to successfully perform this tasksubjects had to form a representation of the surfacesof the object or scene sustain this representation in
memory for 10 s and then align it (presumablyusing mental rotation) to the second object orscene
MethodThe experiment consisted of two parts each with40 trials In the first part the stimuli were black andwhite photographs of objects made out of Legoblocks In the second part the stimuli were photo-graphs of ldquoscenesrdquo made out of Lego blocks whichwere designed to have a geometric structure similarto real-world places such as a roomor a city street
Subjects began each trial by pressing the spacebar A photograph of a Lego object or sceneappeared on the screen for 3 s followed by a 10-sblank interval A second photograph then appeareddepicting either (1) the same object or scene as thefirst photograph shown from a different viewingangle (offset approximately 35 degrees) or (2) a dif-ferent object or scene Subjects were instructed topress the ldquojrdquo key if the two photographs depicted thesame object or scene and the ldquof rdquo key if they did notThey were told to take as much time as they neededto make a decision but no extra time The secondphotograph remained on the screen until a responsewas made Half the trials were ldquosamerdquo trials and halfwere ldquodifferentrdquo trials
Results and discussionFigure 8 shows results for GR and CO as well as forsix elderly normal controls GRrsquos and COrsquos perfor-mance was comparable to that of the normal con-trols Furthermore GR and CO performed just aswell when the stimuli were Lego scenes as whenthey were Lego objects These results are unlikely toreflect a ceiling effect as performance for bothpatients and controls was around 80 correctThus both patients appear to be able to form a rep-resentation of the geometry of both scenes andobjects which they can sustain in short-termmemory
Experiment 3 N-back recognition memorytask
The previous experiment failed to find any impair-ment in GR and COrsquos ability to form a representa-
EPSTEIN ET AL
496 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
tion of the geometry of a scene or object andmaintain it for 10 s in the absence of the originalstimulus Not only does this result suggest that per-ception of scene-like stimuli is intact in thesepatients it further suggests that some short-termmemory functions are intact as well However we
should note that subjects in this experiment werefree to devote all their effort and attention in the 10-second blank interval to maintaining a memory ofthe stimulus Thus it is possible that GR and COcan use rehearsal mechanisms to maintain the stim-ulus in short-term memory during the blank inter-
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 497
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
(b) Lego object version
Figure 7 Stimuli and procedure for Experiment 2 (a) In the Lego layout version subjects see a Lego layout for 3 s followed by a blankscreen for 10 s A second layout then appears which is either the same layout as the first one but seen from a different angle or a differentlayout (b) The Lego object version is the same but with object stimuli
3 s 10 s OR
3 s 10 s OR
(a) Lego layout version
(b) Lego object version
val but are unable to form a more durable memorytrace for the stimulus This possibility was tested inthe current experiment which was designed toexamine GR and COrsquos ability to learn new visualinformation
We devised a continuous n-back recognitionmemory test in which subjects viewed a sequence oforiginally novel stimuli some of which wererepeated within the sequence at intervals of 1 3 or5 intervening images For each stimulus subjects
reported whether they had seen it before in theexperiment or not In different runs in the experi-ment the stimuli were black-and-white photo-graphs of (1) Lego scenes (2) Lego objects (3) real-world landscapes (4) novel objects made out ofSculpey clay (5) unfamiliar houses (6) unfamiliarfaces as well as (7) line drawings of nonsenseobjects and (8) pronounceable pseudowords (Fig-ure 9) Thus we were able to test GR and COrsquos rec-ognition memory for different kinds of materials atdifferent time intervals in a paradigm in which thepresence of intervening items between the twoappearances of each stimulus made rehearsal diffi-cult Note that because none of the specific stimuliused in this experiment were familiar in advancethis task did not require an episodic memory com-ponent and could be done on the basis of pure visualfamiliarity with the particular visual stimulus
Although many different stimulus classes weretested in this experiment the critical comparison isbetween the Lego layouts and Lego objectsbecause these are best matched with regard tomaterials and differ primarily in their geometricstructure (see Introduction) We predicted that ifGR and CO showed a memory deficit for this taskit would be greater for the Lego layouts than for the
EPSTEIN ET AL
498 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 8 Results of Experiment 2 Performance over ldquosamerdquotrials and ldquodifferentrdquo trials is averaged together Error bars show1 SD
Figure 9 Stimuli for Experiment 3 Top row Lego layouts real-world scenes houses Bottom row Lego objects clay objects nonsenseobjects faces In an eighth condition (not shown) subjects viewed pronounceable nonwords such as ldquowilchrdquo or ldquothipperrdquo
Lego objects In addition a deficit for materialsthat convey information about the layout of sur-rounding space might also be apparent when GRand CO are tested on more realistic stimuli If soone would expect them to do poorly on the land-scapes but relatively well on the clay objects linedrawings of nonsense objects faces andpseudowords
MethodFor each subject a complete experimental sessionconsisted of being run once on each of the eightstimulus classes Each run consisted of 90 trials ineach trial a stimulus was presented and subjectsreported whether or not they had seen it before inthe run (or indeed ever) by pressing the ldquojrdquo key ifthey had or the ldquof rdquo key if they had not Each stimu-lus was on the screen for either 1 2 or 3 s (see fol-lowing) and appeared promptly after the keypressreport for the previous trial Individual stimuliappeared either once (14 stimuli) twice (30 stim-uli) or four times (4 stimuli) within a given run Ofthe stimuli that were presented twice 10 wererepeated immediately (1-back) 10 were repeatedwith a lag of two intervening items (3-back) and 10were repeated with a lag of four intervening items(5-back) In order to ensure that between-subjectdifferences in performance were not due to stimulusdifferences the same stimuli were assigned to thesame conditions each time the experimentwas run
GR performed the experiment twice once inAugust 1998 and again 9 months later Because ofa computer error he was only tested on six of theeight categories (all except Lego layouts and Legoobjects) at the later date CO was also tested twiceonce in February 1999 and again 6 months laterReported results for GR and CO are the average oftheir two testing sessions Normal subjects wereonly tested once Stimulus presentation time was 1 sin GRrsquos first testing session In pilot testing COfound this too fast for the critical categories of Lego
layouts and Lego objects so presentation time wasextended to 2 s for these categories in his first test-ing session All normal subjects were tested at thesepresentation rates (2 s for Lego layouts and objects1 s for other stimuli) In COrsquos second testingsession presentation time was lengthened to 3 s forall stimulus categories to ensure that any deficitsfound were memory deficits rather than problemswith the nonmemory aspects of the task
Results and discussionResults from GR CO and 10 elderly normal con-trols are shown in Figure 10 and Table 4 Figure 10ashows results from the main comparison betweenLego layouts and Lego objects Both normal sub-jects and patients performed near ceiling for bothLego layouts and Lego objects at 1-backinterstimulus intervals verifying that GR and COcould successfully perceive the pictures as they werebeing presented In contrast there was a strikingdifference between the two patients and the normalsubjects at 5-back intervals Whereas normals didon average somewhat better on Lego layouts thanLego objects both GR and CO showed the oppo-site pattern doing much worse on Lego layoutsthan Lego objects The 3-back condition showed amixed pattern Whereas GR did better on Legoobjects than Lego layouts in this condition (similarto his performance at 5-back intervals) CO didbetter on Lego layouts than Lego objects COrsquos lowperformance on Lego objects in the 3-back condi-tion appears to be an anomaly as it is much lowerthan his performance for comparable stimuli in the1-back and 5-back conditions
Figure 10b shows the difference in performancebetween Lego layouts and Lego objects for thecritical 1-back and 5-back intervals At 5-backintervals CO was significantly impaired comparedto the normal subjects (z = ndash250) whereasGRrsquos impairment fell just short of significance (z =ndash175)4
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 499
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
4In fact these values may underestimate GR and COrsquos relative impairment for Lego layouts In the 5-back condition 4 of the 10
normal subjects had perfect performance on the Lego layouts but all scored below ceiling on the Lego objects Thus the true advantagefor Lego layouts over Lego objects for normal subjects is probably greater than that observed here in which case GR and COrsquos lowerperformance for Lego layouts compared to Lego objects would be even more abnormal Furthermore post-hoc analyses revealed thatone of the normal subjects showed a difference in performance between Lego layouts and objects that was over 2 SD below the mean ofthe other nine subjects When this outlying subject was excluded z-scores were ndash322 for CO and ndash232 for GR
Table 4 shows results for all eight stimulusclasses Although direct comparisons between sub-jects should be treated with caution since viewingtime for different items was not the same for eachsubject we do note a few interesting patterns Firstof all there is little or no evidence that GR or CO
show a memory deficit for nonsense words clayobjects or line drawings of nonsense objects GRappears to be slightly impaired on faces and housesbut CO performs normally on these categoriesStrikingly GR shows a severe deficit for the reallandscapes performing more than 40 percentagepoints lower than normals Thus even thoughthere is some evidence that GR might have someimpairment for nonscene stimuli he clearly is muchmore impaired for scene-like stimuli such as theLego scenes and landscapes CO does not show asimilar deficit for the landscapes However itshould be kept in mind that in one of his two testingsessions CO viewed each picture for 3 s which wasmuch longer than the 1 second used for all non-Lego stimuli for GR and normals Thus COrsquosapparently ldquonormalrdquo performance in these condi-tions might be overestimated
Taken together the results of this experimentindicate that GR and CO are selectively impaired atprocessing scene-like stimuli This deficit is consis-tently apparent at 5-back but not 1-back intervalsThis finding suggests that the deficit should becharacterised as a memory encoding problemHowever the present results do not allow us todetermine whether the difficulty occurs at memoryencoding or at subsequent recognition (see Epsteinamp Kanwisher 2001) If GR and CO have a generalproblem with recognition of topographical materi-als then they should not be able to recognise scenes
EPSTEIN ET AL
500 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 10a Results of Experiment 3 for GR (top) CO (middle)and for 10 age-matched normal controls Performance measure isproportion of repeated stimuli correctly identified as such correctedfor guessing using the standard formula
Figure 10b Difference between Lego layout performance andLego object performance for the 1-back and 5-back conditionsError bars indicate one 1 SD
and landmarks that were learned prior to theirinjury We tested this possibility in the nextexperiment
Experiment 4 Famous landmarksfacesrecognition task
In this experiment we examined GR and COrsquosability to recognise the visual appearances of placeslearned prior to injury Subjects saw a series offamous landmarks intermixed with nonfamouscontrols (Imaging experiments have demonstratedthat landmarks of this type can activate the PPAalmost as strongly as complete scenes Epstein etal 1999 Epstein amp Kanwisher 1998) They wererequired to say whether they knew each specificlandmark and to identify the ones they knew Sub-jects were instructed to respond affirmatively if theywere fairly certain that they recognised the land-mark but not if it only looked generally familiarFor purposes of comparison subjects also per-formed an identical test in which the stimuli werefamous and nonfamous faces In both testsdistractor items were designed to be visually andconceptually similar to the target items (see Figure11) If parahippocampal cortex is required for placerecognition then one might expect GR and CO toperform poorly compared to normals on the land-mark recognition test
MethodEach of the two experiments (famous faces famouslandmarks) consisted of 48 trials Subjects beganeach trial by pressing the space bar after which ablack-and-white photograph of a face (in thefamous faces experiment) or a landmark (in thefamous landmarks experiment) appeared on thescreen Subjects were told to press the ldquojrdquo key if theyrecognised the specific face or landmark and the ldquof rdquokey if they did not taking as much time at theyneeded to make this discrimination but no extratime Subjects were told to report that they recog-nised a face or landmark if they felt that they knewit even if they could not retrieve its name If theydid know the name they were to say it after pressingthe button Otherwise they were to say whateverthey did remember about the item
Half of the photographs were of famous and halfof nonfamous people and landmarks The famousfaces included public figures (eg George BushRichard Nixon Prince Charles) and movie stars(eg Harrison Ford Clark Gable Marlon Brando)who were prominent prior to the time of GR andCOrsquos injuries the famous landmarks were build-ings and monuments from all over the world (egEiffel Tower US Capitol Building Great Sphinx)The nonfamous stimuli were chosen to match thefamous stimuli as closely as possible Each famousstimulus had a ldquomatchrdquo For example George Bushwas matched to a man of similar age in a coat andtie and the Taj Mahal was matched to a visually
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 501
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 4 Full results of Experiment 3 Mean proportion correct (corrected for guessing) SDs in parentheses
Lego Land- Lego Clay Nonsense Non-scenes scapes Houses objects objects objects Words Faces
1-back GR 100 094 100 100 100 094 100 100CO 091 085 091 100 100 079 074 100Normals 096(008) 095(008) 093(014) 096(008) 096(006) 099(003) 098(004) 097(011)
3-back GR ndash003 068 026 047 083 083 075 052CO 066 081 037 010 094 069 095 081Normals 066(032) 077(021) 072(015) 077(020) 086(016) 083(015) 083(017) 072(020)
5-back GR 010 034 044 047 072 083 094 053CO 031 073 075 088 081 064 100 081Normals 076(026) 077(016) 068(021) 066(016) 091(014) 083(018) 093(011) 078(021)
similar Asian palace that is not famous (see Figure11)
Results and discussionResults for GR and CO as well as for five age-matched controls are shown in Figure 12 Both
GR and CO could recognise the famous landmarksas well normals In addition both GR and CO wereusually able to name the landmarks they success-fully identified
Thus GR and CO can recognize landmarkslearned prior to injury demonstrating that at leastsome of their place recognition abilities are intactWhen combined with the fact that GR and COwere able to produce detailed maps of places theylived in prior to their injury (see Case Description)these results suggest that their deficit is primarily inencoding or consolidation of new informationrather than in recognition or recall of old informa-tion Note however that these results do not provethat their place recognition abilities are entirelynormal GR and CO might suffer from a mild rec-ognition deficit that affects identification of newly-learned stimuli like the Lego layouts in Experiment3 but not overlearned stimuli like the famous land-marks Furthermore it is possible that GR and COmight be impaired at recognising premorbidlylearned places that are defined primarily by theirspatial layout even though they are unimpaired atrecognising premorbidly learned places that aredefined primarily by a single object-like landmarkor building (see General Discussion) Neverthelessthe trend of the present evidence is to suggest thatGR and CO have particular problems with noveltopographical information
In the famous-face task GRrsquos performance wasin the normal range However he sometimes haddifficulty naming the faces even though he clearly
EPSTEIN ET AL
502 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 11 Stimuli for Experiment 4 Each famous landmark(Arc de Triomphe Taj Mahal) and famous face (Barbara BushYasser Arafat) was matched to a nonfamous building or face thatalso appeared in the experiment
FA
CE
SL
AN
DM
AR
KS
Figure 12 Results of Experiment 4 Performance measure isproportion of famous stimuli reported as ldquoknownrdquo corrected forguessing using the standard formula
recognised them For example he described Ron-ald Reagan as ldquothe gipperrdquo Nancy Reagan as ldquothewife of the gipperrdquo and Clark Gable as ldquothe movieactor who said lsquoScarlett I donrsquot give a damnrsquordquo COhad a great deal of difficulty identifying the famousfaces This result is not surprising given hisextremely low performance on the Benton FaceMatching Task In addition to his topographicproblems CO appears to suffer fromprosopagnosia
GENERAL DISCUSSION
We tested two parahippocampal patients (GR andCO) on four experiments designed to measure per-formance as a function of both task (perceptionencoding and recognition) and stimulus type(scenes objects faces and words) The only taskthat revealed a clear deficit was an n-back recogni-tion memory test (Experiment 3) In order to suc-cessfully perform this task subjects had to encode adurable representation of a visual stimulus and useit for later recognition GR and CO were signifi-cantly more impaired on this task when the stimuliwere scene-like spatial layouts (ie Lego scenesand real scenes for GR) than when they wereobjects No comparable deficits were found in otherexperiments that tested layout perception (Experi-ments 1 and 2) and placelandmark recognition(Experiment 4) Taken together these results indi-cate that GR and CO are impaired in their ability toencode novel information about the geometry ofsurrounding space into memory but are lessimpaired at their ability to encode the geometry ofnovel objects
We argue that this specific topographical defi-citmdashinability to encode the spatial layout of novelscenesmdashresults from damage to the para-hippocampal place area Although the damage inboth patients extends beyond the parahippo-campal-lingual territory normally associated withthe PPA this region is clearly lesioned bilaterally inGR and in the right hemisphere in CO Further-more functional imaging demonstrated that GRhas no functioning PPA Thus the current resultsand the earlier fMRI work (Epstein et al 1999
Epstein amp Kanwisher 1998) converge to implicateparahippocampal cortex in the encoding of infor-mation about scene geometry into memory
Our assignment of the function of memoryencoding to parahippocampal cortex is consistentwith the results of earlier case reports of patientswith parahippocampal damage who have naviga-tional difficulties in novel but not familiar environ-ments (Habib amp Sirigu 1987 Ross 1980 see alsoTeng amp Squire 1999) As with GR and CO thesepatients were either less impaired or unimpaired onnontopographical memory tasks Patients have alsobeen described who exhibit the opposite pattern ofimpairment an inability to learn faces (sometimescalled ldquoprosopamnesiardquo Cipolotti Robinson Blairamp Frith 1999 Tippett Miller amp Farah 2000) orfaces and verbal materials (Maguire amp Cipolotti1998) with an unimpaired ability to learn topo-graphical materials Taken together these resultsargue for a high degree of domain-specificity in theneural systems involved in learning of new informa-tion (Gallistel 1995) The brain may have separatemechanisms for learning faces places and wordswhich can be differentially impaired after damageto specific cortical regions (Cipolotti et al 1999Incisa della Rochetta et al 1996) The presentresults suggest that the anatomical locus of themechanism dedicated to learning new topographi-cal information is in the parahippocampal cortex(Aguirre amp DrsquoEsposito 1999)
In contrast to their impaired ability to encodenew topographical information both GR and COwere able to produce detailed maps of their previousdwellings and performed normally on the famouslandmark recognition test of Experiment 4 Theseresults are generally consistent with Teng andSquirersquos (1999) report of a patient with extensivemedial temporal damage who could recallpremorbidly learned but not postmorbidly learnedtopographical information (see also Ross 1980)Thus our results provide further evidence thatmedial temporal regions such as parahippocampalcortex may be more critical for encoding or consoli-dating new information into memory than for recallof old information
One issue that our experiments do not entirelyresolve is the precise nature of the topographical
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 503
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
information encoded by parahippocampal cortexClearly this region is critical for encoding informa-tion about the geometry of the currently visiblescene even when the ldquoscenerdquo is made out of Legoblocks and therefore has no immediate navigationalrelevance However it is unclear whether this spe-cific deficit can account for all of the real-worldnavigational difficulties encountered by GR andCO GR in particular shows such a catastrophicloss of navigational ability that we suspect he is alsoimpaired at encoding the spatial relationshipsbetweenthe current scene and other locations in theworld CO probably also suffers from such difficul-ties as he reports that he has trouble rememberinghow places relate to each other in large-scale spaceWhen he was tested on a simple 3-room videogameenvironment he not only was unable to rememberthe individual rooms (claiming that there were 10rooms when there were only 3) but he also had nomemory for the spatial relationships between themThese spatial deficits may be the result of damage tothe posterior right hippocampus (in the case of CO)or of disconnection of the posterior right hippo-campus from the parahippocampal regions thatprovide much of its cortical input (in both patientsSuzuki amp Amaral 1994) In either case the abilityto form a cognitive map of new environments likethe lab or videogame space could be impaired(Maguire et al 1998 OrsquoKeefe amp Nadel 1978)However it is also possible that parahippocampalcortex itself may be critical for encoding not onlythe geometry of individual scenes but also the spa-tial relationships between the locations of thosescenes In this case the spatial deficits of GR andCO could be attributed directly to parahippo-campal damage rather than to hippocampal dam-age or disconnection
The latter scenario would be consistent withAguirre and DrsquoEspositorsquos (1999) and Barrash etalrsquos (2000) suggestion that parahippocampal cortexis critically involved in combining visual informa-tion about the appearance of scenes and landmarkswith spatial information about their locations into aunified topographical representation Howeverboth of these authors argue that the representationsof individual scenes (Barrash et al) and landmarks(Aguirre amp DrsquoEsposito) are supported not by
parahippocampal cortex but by more posterior lin-gual-fusiform regions In contrast we believe thatthe neuroimaging data indicate that parahippo-campal cortex is involved in the encoding of thegeometric structure of individual scenes althoughit may also play a role in combining these represen-tations with other spatial representations
Interestingly GR and CO were not impaired onrecognition of famous landmarks that were mostlybuildings Thus our data do not preclude the possi-bility that there may be a separate lingual orfusiform region involved in processing informationabout buildings (Aguirre Zarahn amp DrsquoEsposito1998 Chao Haxby amp Martin 1999 IshaiUngerleider Martin Schouten amp Haxby 1999)which might have been preserved in GR and COdespite extensive damage to other regions of thelingual and fusiform gyrus Alternatively GR andCOrsquos preserved building recognition abilities mightbe mediated by interactions between cortical areasthat perform shape analysis for any object-likestimulus type (Malach et al 1995Kanwisher et al1996) and anterior temporal regions in which long-term topographical and semantic memories arestored (McCarthy et al 1996)
Is perception preserved
In contrast to their inability to encode new scenesinto memory GR and CO have substantiallyretained their ability to perceive the spatial layout ofthe currently visible scene (Experiments 1 and 2)GR can follow a route marked on a floor planwhich indicates a good on-line appreciation of hiscurrent spatial surroundings Furthermore neitherGR or CO have any trouble moving through theenvironmentmdashthey move with confidence and donot bump into doors or walls Problems of the lattertype are usually associated with damage to the pari-etal lobes which are intact in GR and CO Thus atfirst glance GR and CO do not appear to haveperceptual impairments (beyond their visual fieldrestrictions)
However GR does complain of a perceptualdeficit which he describes as an inability to ldquotakeinrdquo scenes when they are too complex Similarreports of an inability to comprehend the
EPSTEIN ET AL
504 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
ldquoglobalityrdquo of a scene were made by some of theparahippocampally-damaged patients studied byHabib and Sirigu (1987) We were not able todetermine the precise nature of this deficit whichmay be partly attributable to his visual field restric-tions It appears that his deficit is more salient in thereal-world environments than in the picture-per-ception tasks of Experiments 1 and 2 perhapsbecause he finds it more difficult to organise visualinput when immersed in it than when viewing pho-tographs that subtend only part of the visual field
We suggest that GR can perceive parts of thescene when he attends to them but may be unableto integrate information over time into a coherentrepresentation of a scene or place The fact that GRand CO are impaired for Lego layouts at 5-backintervals in Experiment 3 is consistent with thishypothesis The time between the first and secondpresentations of a stimulus is very short in this case(and even shorter with 3-back intervals where GRalso shows evidence of a deficit) In both patientsthe representation of layout information formed byperception of a Lego scene must either have anabnormally short persistence be abnormally sus-ceptible to interference or both If so they mightfind it impossible to sustain a spatial frame orschema long enough to use it to organise informa-tion obtained from many different glances at a com-plex scene (Intraub 1997) The result would be aninability to understand the ldquoglobalityrdquo of a sceneeven though all the details were perfectlyperceived
Non-topographical mnemonic abilities
Although not as severe as their topographical mem-ory problems GR and CO do have some memorydifficulties in other domains For example bothperformed poorly on the Rey AVLT whichrequired them to remember a list of unrelatedwords Both also report some subtle episodic mem-ory problems (see Case Description) COrsquos prob-lems could partly result from damage to the righthippocampusmdashindeed CO appeared to have themore acute episodic memory problems HoweverGR has no hippocampal damage so the fact that hedisplays episodic memory problems suggests thatparahippocampal cortex is involved in the forma-
tion of episodic memories (cf Wagner et al1998)
In previous work (Epstein et al 1999) weargued that spatial context information encoded byparahippocampal cortex might play a role in estab-lishing event memories by encoding informationabout where events take place The inability toencode this information might cause problems intasks such as the Rey AVLT where one has to recalla list of unrelated words If spatial (and perhapstemporal) context information is not encoded itwill be harder subsequently to recall the distinctevent of hearing each word (Tulving 1983) Incontrast lack of spatial context information wouldbe less of a problem in tasks where one is onlyrequired to recognise previously encountered verbalmaterials but not to reproduce the spatiotemporalcontext in which they were experienced IndeedGR and CO performed as well as normals on thenonsense words in Experiment 4 where recogni-tion memory but not episodic memory was requiredfor successful performance Interestingly GR andCO also did relatively well on the Logical Memorysubtest of the WMS-III perhaps because the nar-rative framework of a story provides sufficient con-textual information to overcome the absence ofspatial context and mediate successful verbalretrieval
Differences between GR and CO
Although we have discussed GR and CO togetherthere are a number of relevant differences betweenthem One open question is whether these differ-ences can be attributed to the different effects ofunilateral and bilateral parahippocampal damageIn general GRrsquos topographical difficulties weremore severe than COrsquos CO did surprisingly well onthe Warrington Topographical Memory Test inwhich he was required to remember detailed real-world city scenes and also performed substantiallybetter on real scenes than Lego scenes in Experi-ment 3 Insofar as CO has intact left para-hippocampal cortex this suggests that the left PPAmight be sufficient to encode a certain amount of(perhaps semantic) information about real-worldscenes but the right PPA is necessary to encode
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 505
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)

difference did not appear to be significant Impor-tantly there was little evidence that GR or CO tooklonger to make their perceptual judgements for thereal or Lego scenes than for the real or Lego objectsThus contrary to our initial hypothesis this experi-ment found no clear evidence for selective impair-ment in the perception of scene-like stimuli
Experiment 2 Different-views matchingtask
The previous experiment demonstrated that GRand CO can extract information about the per-ceived distances between themselves and the sur-faces of an object or scene Thus in at least onesense their perception of these surfaces is pre-served However this does not mean that their abil-ity to represent the layout of a scene is unimpairedThe two-dot task requires on-line comparison oftwo surfaces in egocentric coordinates GR and COwould be able to perform this task even if they couldnot (1) represent more than two surfaces at a time(2) represent surfaces in other than strictly egocen-tric coordinates or (3) sustain surface representa-tions for longer than the time it takes to make acomparison It has been suggested that the parietallobes are involved in on-line processing of surfacegeometry in egocentric coordinates (Milner ampGoodale 1995) thus it is possible that the parietallobes (which are intact in GR and CO) may be suf-ficient to perform the two-dot task even in theabsence of more durable or complex surface repre-sentations in parahippocampal cortex
In Experiment 2 we examine GR and COrsquosability to create and sustain a representation of thegeometry of an object or scene-like layout The taskwas to view a photograph of a Lego object or Legoscene hold it in memory for 10 s and then deter-mine whether a second object or scene was the sameas the first In ldquosamerdquo trials the second photographdepicted the same scene or object depicted from adifferent viewing angle In ldquodifferentrdquo trials the sec-ond photograph depicted a different scene or objectchosen to be similar to the first one (Figure 7)Thus in order to successfully perform this tasksubjects had to form a representation of the surfacesof the object or scene sustain this representation in
memory for 10 s and then align it (presumablyusing mental rotation) to the second object orscene
MethodThe experiment consisted of two parts each with40 trials In the first part the stimuli were black andwhite photographs of objects made out of Legoblocks In the second part the stimuli were photo-graphs of ldquoscenesrdquo made out of Lego blocks whichwere designed to have a geometric structure similarto real-world places such as a roomor a city street
Subjects began each trial by pressing the spacebar A photograph of a Lego object or sceneappeared on the screen for 3 s followed by a 10-sblank interval A second photograph then appeareddepicting either (1) the same object or scene as thefirst photograph shown from a different viewingangle (offset approximately 35 degrees) or (2) a dif-ferent object or scene Subjects were instructed topress the ldquojrdquo key if the two photographs depicted thesame object or scene and the ldquof rdquo key if they did notThey were told to take as much time as they neededto make a decision but no extra time The secondphotograph remained on the screen until a responsewas made Half the trials were ldquosamerdquo trials and halfwere ldquodifferentrdquo trials
Results and discussionFigure 8 shows results for GR and CO as well as forsix elderly normal controls GRrsquos and COrsquos perfor-mance was comparable to that of the normal con-trols Furthermore GR and CO performed just aswell when the stimuli were Lego scenes as whenthey were Lego objects These results are unlikely toreflect a ceiling effect as performance for bothpatients and controls was around 80 correctThus both patients appear to be able to form a rep-resentation of the geometry of both scenes andobjects which they can sustain in short-termmemory
Experiment 3 N-back recognition memorytask
The previous experiment failed to find any impair-ment in GR and COrsquos ability to form a representa-
EPSTEIN ET AL
496 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
tion of the geometry of a scene or object andmaintain it for 10 s in the absence of the originalstimulus Not only does this result suggest that per-ception of scene-like stimuli is intact in thesepatients it further suggests that some short-termmemory functions are intact as well However we
should note that subjects in this experiment werefree to devote all their effort and attention in the 10-second blank interval to maintaining a memory ofthe stimulus Thus it is possible that GR and COcan use rehearsal mechanisms to maintain the stim-ulus in short-term memory during the blank inter-
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 497
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
(b) Lego object version
Figure 7 Stimuli and procedure for Experiment 2 (a) In the Lego layout version subjects see a Lego layout for 3 s followed by a blankscreen for 10 s A second layout then appears which is either the same layout as the first one but seen from a different angle or a differentlayout (b) The Lego object version is the same but with object stimuli
3 s 10 s OR
3 s 10 s OR
(a) Lego layout version
(b) Lego object version
val but are unable to form a more durable memorytrace for the stimulus This possibility was tested inthe current experiment which was designed toexamine GR and COrsquos ability to learn new visualinformation
We devised a continuous n-back recognitionmemory test in which subjects viewed a sequence oforiginally novel stimuli some of which wererepeated within the sequence at intervals of 1 3 or5 intervening images For each stimulus subjects
reported whether they had seen it before in theexperiment or not In different runs in the experi-ment the stimuli were black-and-white photo-graphs of (1) Lego scenes (2) Lego objects (3) real-world landscapes (4) novel objects made out ofSculpey clay (5) unfamiliar houses (6) unfamiliarfaces as well as (7) line drawings of nonsenseobjects and (8) pronounceable pseudowords (Fig-ure 9) Thus we were able to test GR and COrsquos rec-ognition memory for different kinds of materials atdifferent time intervals in a paradigm in which thepresence of intervening items between the twoappearances of each stimulus made rehearsal diffi-cult Note that because none of the specific stimuliused in this experiment were familiar in advancethis task did not require an episodic memory com-ponent and could be done on the basis of pure visualfamiliarity with the particular visual stimulus
Although many different stimulus classes weretested in this experiment the critical comparison isbetween the Lego layouts and Lego objectsbecause these are best matched with regard tomaterials and differ primarily in their geometricstructure (see Introduction) We predicted that ifGR and CO showed a memory deficit for this taskit would be greater for the Lego layouts than for the
EPSTEIN ET AL
498 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 8 Results of Experiment 2 Performance over ldquosamerdquotrials and ldquodifferentrdquo trials is averaged together Error bars show1 SD
Figure 9 Stimuli for Experiment 3 Top row Lego layouts real-world scenes houses Bottom row Lego objects clay objects nonsenseobjects faces In an eighth condition (not shown) subjects viewed pronounceable nonwords such as ldquowilchrdquo or ldquothipperrdquo
Lego objects In addition a deficit for materialsthat convey information about the layout of sur-rounding space might also be apparent when GRand CO are tested on more realistic stimuli If soone would expect them to do poorly on the land-scapes but relatively well on the clay objects linedrawings of nonsense objects faces andpseudowords
MethodFor each subject a complete experimental sessionconsisted of being run once on each of the eightstimulus classes Each run consisted of 90 trials ineach trial a stimulus was presented and subjectsreported whether or not they had seen it before inthe run (or indeed ever) by pressing the ldquojrdquo key ifthey had or the ldquof rdquo key if they had not Each stimu-lus was on the screen for either 1 2 or 3 s (see fol-lowing) and appeared promptly after the keypressreport for the previous trial Individual stimuliappeared either once (14 stimuli) twice (30 stim-uli) or four times (4 stimuli) within a given run Ofthe stimuli that were presented twice 10 wererepeated immediately (1-back) 10 were repeatedwith a lag of two intervening items (3-back) and 10were repeated with a lag of four intervening items(5-back) In order to ensure that between-subjectdifferences in performance were not due to stimulusdifferences the same stimuli were assigned to thesame conditions each time the experimentwas run
GR performed the experiment twice once inAugust 1998 and again 9 months later Because ofa computer error he was only tested on six of theeight categories (all except Lego layouts and Legoobjects) at the later date CO was also tested twiceonce in February 1999 and again 6 months laterReported results for GR and CO are the average oftheir two testing sessions Normal subjects wereonly tested once Stimulus presentation time was 1 sin GRrsquos first testing session In pilot testing COfound this too fast for the critical categories of Lego
layouts and Lego objects so presentation time wasextended to 2 s for these categories in his first test-ing session All normal subjects were tested at thesepresentation rates (2 s for Lego layouts and objects1 s for other stimuli) In COrsquos second testingsession presentation time was lengthened to 3 s forall stimulus categories to ensure that any deficitsfound were memory deficits rather than problemswith the nonmemory aspects of the task
Results and discussionResults from GR CO and 10 elderly normal con-trols are shown in Figure 10 and Table 4 Figure 10ashows results from the main comparison betweenLego layouts and Lego objects Both normal sub-jects and patients performed near ceiling for bothLego layouts and Lego objects at 1-backinterstimulus intervals verifying that GR and COcould successfully perceive the pictures as they werebeing presented In contrast there was a strikingdifference between the two patients and the normalsubjects at 5-back intervals Whereas normals didon average somewhat better on Lego layouts thanLego objects both GR and CO showed the oppo-site pattern doing much worse on Lego layoutsthan Lego objects The 3-back condition showed amixed pattern Whereas GR did better on Legoobjects than Lego layouts in this condition (similarto his performance at 5-back intervals) CO didbetter on Lego layouts than Lego objects COrsquos lowperformance on Lego objects in the 3-back condi-tion appears to be an anomaly as it is much lowerthan his performance for comparable stimuli in the1-back and 5-back conditions
Figure 10b shows the difference in performancebetween Lego layouts and Lego objects for thecritical 1-back and 5-back intervals At 5-backintervals CO was significantly impaired comparedto the normal subjects (z = ndash250) whereasGRrsquos impairment fell just short of significance (z =ndash175)4
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 499
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
4In fact these values may underestimate GR and COrsquos relative impairment for Lego layouts In the 5-back condition 4 of the 10
normal subjects had perfect performance on the Lego layouts but all scored below ceiling on the Lego objects Thus the true advantagefor Lego layouts over Lego objects for normal subjects is probably greater than that observed here in which case GR and COrsquos lowerperformance for Lego layouts compared to Lego objects would be even more abnormal Furthermore post-hoc analyses revealed thatone of the normal subjects showed a difference in performance between Lego layouts and objects that was over 2 SD below the mean ofthe other nine subjects When this outlying subject was excluded z-scores were ndash322 for CO and ndash232 for GR
Table 4 shows results for all eight stimulusclasses Although direct comparisons between sub-jects should be treated with caution since viewingtime for different items was not the same for eachsubject we do note a few interesting patterns Firstof all there is little or no evidence that GR or CO
show a memory deficit for nonsense words clayobjects or line drawings of nonsense objects GRappears to be slightly impaired on faces and housesbut CO performs normally on these categoriesStrikingly GR shows a severe deficit for the reallandscapes performing more than 40 percentagepoints lower than normals Thus even thoughthere is some evidence that GR might have someimpairment for nonscene stimuli he clearly is muchmore impaired for scene-like stimuli such as theLego scenes and landscapes CO does not show asimilar deficit for the landscapes However itshould be kept in mind that in one of his two testingsessions CO viewed each picture for 3 s which wasmuch longer than the 1 second used for all non-Lego stimuli for GR and normals Thus COrsquosapparently ldquonormalrdquo performance in these condi-tions might be overestimated
Taken together the results of this experimentindicate that GR and CO are selectively impaired atprocessing scene-like stimuli This deficit is consis-tently apparent at 5-back but not 1-back intervalsThis finding suggests that the deficit should becharacterised as a memory encoding problemHowever the present results do not allow us todetermine whether the difficulty occurs at memoryencoding or at subsequent recognition (see Epsteinamp Kanwisher 2001) If GR and CO have a generalproblem with recognition of topographical materi-als then they should not be able to recognise scenes
EPSTEIN ET AL
500 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 10a Results of Experiment 3 for GR (top) CO (middle)and for 10 age-matched normal controls Performance measure isproportion of repeated stimuli correctly identified as such correctedfor guessing using the standard formula
Figure 10b Difference between Lego layout performance andLego object performance for the 1-back and 5-back conditionsError bars indicate one 1 SD
and landmarks that were learned prior to theirinjury We tested this possibility in the nextexperiment
Experiment 4 Famous landmarksfacesrecognition task
In this experiment we examined GR and COrsquosability to recognise the visual appearances of placeslearned prior to injury Subjects saw a series offamous landmarks intermixed with nonfamouscontrols (Imaging experiments have demonstratedthat landmarks of this type can activate the PPAalmost as strongly as complete scenes Epstein etal 1999 Epstein amp Kanwisher 1998) They wererequired to say whether they knew each specificlandmark and to identify the ones they knew Sub-jects were instructed to respond affirmatively if theywere fairly certain that they recognised the land-mark but not if it only looked generally familiarFor purposes of comparison subjects also per-formed an identical test in which the stimuli werefamous and nonfamous faces In both testsdistractor items were designed to be visually andconceptually similar to the target items (see Figure11) If parahippocampal cortex is required for placerecognition then one might expect GR and CO toperform poorly compared to normals on the land-mark recognition test
MethodEach of the two experiments (famous faces famouslandmarks) consisted of 48 trials Subjects beganeach trial by pressing the space bar after which ablack-and-white photograph of a face (in thefamous faces experiment) or a landmark (in thefamous landmarks experiment) appeared on thescreen Subjects were told to press the ldquojrdquo key if theyrecognised the specific face or landmark and the ldquof rdquokey if they did not taking as much time at theyneeded to make this discrimination but no extratime Subjects were told to report that they recog-nised a face or landmark if they felt that they knewit even if they could not retrieve its name If theydid know the name they were to say it after pressingthe button Otherwise they were to say whateverthey did remember about the item
Half of the photographs were of famous and halfof nonfamous people and landmarks The famousfaces included public figures (eg George BushRichard Nixon Prince Charles) and movie stars(eg Harrison Ford Clark Gable Marlon Brando)who were prominent prior to the time of GR andCOrsquos injuries the famous landmarks were build-ings and monuments from all over the world (egEiffel Tower US Capitol Building Great Sphinx)The nonfamous stimuli were chosen to match thefamous stimuli as closely as possible Each famousstimulus had a ldquomatchrdquo For example George Bushwas matched to a man of similar age in a coat andtie and the Taj Mahal was matched to a visually
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 501
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 4 Full results of Experiment 3 Mean proportion correct (corrected for guessing) SDs in parentheses
Lego Land- Lego Clay Nonsense Non-scenes scapes Houses objects objects objects Words Faces
1-back GR 100 094 100 100 100 094 100 100CO 091 085 091 100 100 079 074 100Normals 096(008) 095(008) 093(014) 096(008) 096(006) 099(003) 098(004) 097(011)
3-back GR ndash003 068 026 047 083 083 075 052CO 066 081 037 010 094 069 095 081Normals 066(032) 077(021) 072(015) 077(020) 086(016) 083(015) 083(017) 072(020)
5-back GR 010 034 044 047 072 083 094 053CO 031 073 075 088 081 064 100 081Normals 076(026) 077(016) 068(021) 066(016) 091(014) 083(018) 093(011) 078(021)
similar Asian palace that is not famous (see Figure11)
Results and discussionResults for GR and CO as well as for five age-matched controls are shown in Figure 12 Both
GR and CO could recognise the famous landmarksas well normals In addition both GR and CO wereusually able to name the landmarks they success-fully identified
Thus GR and CO can recognize landmarkslearned prior to injury demonstrating that at leastsome of their place recognition abilities are intactWhen combined with the fact that GR and COwere able to produce detailed maps of places theylived in prior to their injury (see Case Description)these results suggest that their deficit is primarily inencoding or consolidation of new informationrather than in recognition or recall of old informa-tion Note however that these results do not provethat their place recognition abilities are entirelynormal GR and CO might suffer from a mild rec-ognition deficit that affects identification of newly-learned stimuli like the Lego layouts in Experiment3 but not overlearned stimuli like the famous land-marks Furthermore it is possible that GR and COmight be impaired at recognising premorbidlylearned places that are defined primarily by theirspatial layout even though they are unimpaired atrecognising premorbidly learned places that aredefined primarily by a single object-like landmarkor building (see General Discussion) Neverthelessthe trend of the present evidence is to suggest thatGR and CO have particular problems with noveltopographical information
In the famous-face task GRrsquos performance wasin the normal range However he sometimes haddifficulty naming the faces even though he clearly
EPSTEIN ET AL
502 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 11 Stimuli for Experiment 4 Each famous landmark(Arc de Triomphe Taj Mahal) and famous face (Barbara BushYasser Arafat) was matched to a nonfamous building or face thatalso appeared in the experiment
FA
CE
SL
AN
DM
AR
KS
Figure 12 Results of Experiment 4 Performance measure isproportion of famous stimuli reported as ldquoknownrdquo corrected forguessing using the standard formula
recognised them For example he described Ron-ald Reagan as ldquothe gipperrdquo Nancy Reagan as ldquothewife of the gipperrdquo and Clark Gable as ldquothe movieactor who said lsquoScarlett I donrsquot give a damnrsquordquo COhad a great deal of difficulty identifying the famousfaces This result is not surprising given hisextremely low performance on the Benton FaceMatching Task In addition to his topographicproblems CO appears to suffer fromprosopagnosia
GENERAL DISCUSSION
We tested two parahippocampal patients (GR andCO) on four experiments designed to measure per-formance as a function of both task (perceptionencoding and recognition) and stimulus type(scenes objects faces and words) The only taskthat revealed a clear deficit was an n-back recogni-tion memory test (Experiment 3) In order to suc-cessfully perform this task subjects had to encode adurable representation of a visual stimulus and useit for later recognition GR and CO were signifi-cantly more impaired on this task when the stimuliwere scene-like spatial layouts (ie Lego scenesand real scenes for GR) than when they wereobjects No comparable deficits were found in otherexperiments that tested layout perception (Experi-ments 1 and 2) and placelandmark recognition(Experiment 4) Taken together these results indi-cate that GR and CO are impaired in their ability toencode novel information about the geometry ofsurrounding space into memory but are lessimpaired at their ability to encode the geometry ofnovel objects
We argue that this specific topographical defi-citmdashinability to encode the spatial layout of novelscenesmdashresults from damage to the para-hippocampal place area Although the damage inboth patients extends beyond the parahippo-campal-lingual territory normally associated withthe PPA this region is clearly lesioned bilaterally inGR and in the right hemisphere in CO Further-more functional imaging demonstrated that GRhas no functioning PPA Thus the current resultsand the earlier fMRI work (Epstein et al 1999
Epstein amp Kanwisher 1998) converge to implicateparahippocampal cortex in the encoding of infor-mation about scene geometry into memory
Our assignment of the function of memoryencoding to parahippocampal cortex is consistentwith the results of earlier case reports of patientswith parahippocampal damage who have naviga-tional difficulties in novel but not familiar environ-ments (Habib amp Sirigu 1987 Ross 1980 see alsoTeng amp Squire 1999) As with GR and CO thesepatients were either less impaired or unimpaired onnontopographical memory tasks Patients have alsobeen described who exhibit the opposite pattern ofimpairment an inability to learn faces (sometimescalled ldquoprosopamnesiardquo Cipolotti Robinson Blairamp Frith 1999 Tippett Miller amp Farah 2000) orfaces and verbal materials (Maguire amp Cipolotti1998) with an unimpaired ability to learn topo-graphical materials Taken together these resultsargue for a high degree of domain-specificity in theneural systems involved in learning of new informa-tion (Gallistel 1995) The brain may have separatemechanisms for learning faces places and wordswhich can be differentially impaired after damageto specific cortical regions (Cipolotti et al 1999Incisa della Rochetta et al 1996) The presentresults suggest that the anatomical locus of themechanism dedicated to learning new topographi-cal information is in the parahippocampal cortex(Aguirre amp DrsquoEsposito 1999)
In contrast to their impaired ability to encodenew topographical information both GR and COwere able to produce detailed maps of their previousdwellings and performed normally on the famouslandmark recognition test of Experiment 4 Theseresults are generally consistent with Teng andSquirersquos (1999) report of a patient with extensivemedial temporal damage who could recallpremorbidly learned but not postmorbidly learnedtopographical information (see also Ross 1980)Thus our results provide further evidence thatmedial temporal regions such as parahippocampalcortex may be more critical for encoding or consoli-dating new information into memory than for recallof old information
One issue that our experiments do not entirelyresolve is the precise nature of the topographical
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 503
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
information encoded by parahippocampal cortexClearly this region is critical for encoding informa-tion about the geometry of the currently visiblescene even when the ldquoscenerdquo is made out of Legoblocks and therefore has no immediate navigationalrelevance However it is unclear whether this spe-cific deficit can account for all of the real-worldnavigational difficulties encountered by GR andCO GR in particular shows such a catastrophicloss of navigational ability that we suspect he is alsoimpaired at encoding the spatial relationshipsbetweenthe current scene and other locations in theworld CO probably also suffers from such difficul-ties as he reports that he has trouble rememberinghow places relate to each other in large-scale spaceWhen he was tested on a simple 3-room videogameenvironment he not only was unable to rememberthe individual rooms (claiming that there were 10rooms when there were only 3) but he also had nomemory for the spatial relationships between themThese spatial deficits may be the result of damage tothe posterior right hippocampus (in the case of CO)or of disconnection of the posterior right hippo-campus from the parahippocampal regions thatprovide much of its cortical input (in both patientsSuzuki amp Amaral 1994) In either case the abilityto form a cognitive map of new environments likethe lab or videogame space could be impaired(Maguire et al 1998 OrsquoKeefe amp Nadel 1978)However it is also possible that parahippocampalcortex itself may be critical for encoding not onlythe geometry of individual scenes but also the spa-tial relationships between the locations of thosescenes In this case the spatial deficits of GR andCO could be attributed directly to parahippo-campal damage rather than to hippocampal dam-age or disconnection
The latter scenario would be consistent withAguirre and DrsquoEspositorsquos (1999) and Barrash etalrsquos (2000) suggestion that parahippocampal cortexis critically involved in combining visual informa-tion about the appearance of scenes and landmarkswith spatial information about their locations into aunified topographical representation Howeverboth of these authors argue that the representationsof individual scenes (Barrash et al) and landmarks(Aguirre amp DrsquoEsposito) are supported not by
parahippocampal cortex but by more posterior lin-gual-fusiform regions In contrast we believe thatthe neuroimaging data indicate that parahippo-campal cortex is involved in the encoding of thegeometric structure of individual scenes althoughit may also play a role in combining these represen-tations with other spatial representations
Interestingly GR and CO were not impaired onrecognition of famous landmarks that were mostlybuildings Thus our data do not preclude the possi-bility that there may be a separate lingual orfusiform region involved in processing informationabout buildings (Aguirre Zarahn amp DrsquoEsposito1998 Chao Haxby amp Martin 1999 IshaiUngerleider Martin Schouten amp Haxby 1999)which might have been preserved in GR and COdespite extensive damage to other regions of thelingual and fusiform gyrus Alternatively GR andCOrsquos preserved building recognition abilities mightbe mediated by interactions between cortical areasthat perform shape analysis for any object-likestimulus type (Malach et al 1995Kanwisher et al1996) and anterior temporal regions in which long-term topographical and semantic memories arestored (McCarthy et al 1996)
Is perception preserved
In contrast to their inability to encode new scenesinto memory GR and CO have substantiallyretained their ability to perceive the spatial layout ofthe currently visible scene (Experiments 1 and 2)GR can follow a route marked on a floor planwhich indicates a good on-line appreciation of hiscurrent spatial surroundings Furthermore neitherGR or CO have any trouble moving through theenvironmentmdashthey move with confidence and donot bump into doors or walls Problems of the lattertype are usually associated with damage to the pari-etal lobes which are intact in GR and CO Thus atfirst glance GR and CO do not appear to haveperceptual impairments (beyond their visual fieldrestrictions)
However GR does complain of a perceptualdeficit which he describes as an inability to ldquotakeinrdquo scenes when they are too complex Similarreports of an inability to comprehend the
EPSTEIN ET AL
504 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
ldquoglobalityrdquo of a scene were made by some of theparahippocampally-damaged patients studied byHabib and Sirigu (1987) We were not able todetermine the precise nature of this deficit whichmay be partly attributable to his visual field restric-tions It appears that his deficit is more salient in thereal-world environments than in the picture-per-ception tasks of Experiments 1 and 2 perhapsbecause he finds it more difficult to organise visualinput when immersed in it than when viewing pho-tographs that subtend only part of the visual field
We suggest that GR can perceive parts of thescene when he attends to them but may be unableto integrate information over time into a coherentrepresentation of a scene or place The fact that GRand CO are impaired for Lego layouts at 5-backintervals in Experiment 3 is consistent with thishypothesis The time between the first and secondpresentations of a stimulus is very short in this case(and even shorter with 3-back intervals where GRalso shows evidence of a deficit) In both patientsthe representation of layout information formed byperception of a Lego scene must either have anabnormally short persistence be abnormally sus-ceptible to interference or both If so they mightfind it impossible to sustain a spatial frame orschema long enough to use it to organise informa-tion obtained from many different glances at a com-plex scene (Intraub 1997) The result would be aninability to understand the ldquoglobalityrdquo of a sceneeven though all the details were perfectlyperceived
Non-topographical mnemonic abilities
Although not as severe as their topographical mem-ory problems GR and CO do have some memorydifficulties in other domains For example bothperformed poorly on the Rey AVLT whichrequired them to remember a list of unrelatedwords Both also report some subtle episodic mem-ory problems (see Case Description) COrsquos prob-lems could partly result from damage to the righthippocampusmdashindeed CO appeared to have themore acute episodic memory problems HoweverGR has no hippocampal damage so the fact that hedisplays episodic memory problems suggests thatparahippocampal cortex is involved in the forma-
tion of episodic memories (cf Wagner et al1998)
In previous work (Epstein et al 1999) weargued that spatial context information encoded byparahippocampal cortex might play a role in estab-lishing event memories by encoding informationabout where events take place The inability toencode this information might cause problems intasks such as the Rey AVLT where one has to recalla list of unrelated words If spatial (and perhapstemporal) context information is not encoded itwill be harder subsequently to recall the distinctevent of hearing each word (Tulving 1983) Incontrast lack of spatial context information wouldbe less of a problem in tasks where one is onlyrequired to recognise previously encountered verbalmaterials but not to reproduce the spatiotemporalcontext in which they were experienced IndeedGR and CO performed as well as normals on thenonsense words in Experiment 4 where recogni-tion memory but not episodic memory was requiredfor successful performance Interestingly GR andCO also did relatively well on the Logical Memorysubtest of the WMS-III perhaps because the nar-rative framework of a story provides sufficient con-textual information to overcome the absence ofspatial context and mediate successful verbalretrieval
Differences between GR and CO
Although we have discussed GR and CO togetherthere are a number of relevant differences betweenthem One open question is whether these differ-ences can be attributed to the different effects ofunilateral and bilateral parahippocampal damageIn general GRrsquos topographical difficulties weremore severe than COrsquos CO did surprisingly well onthe Warrington Topographical Memory Test inwhich he was required to remember detailed real-world city scenes and also performed substantiallybetter on real scenes than Lego scenes in Experi-ment 3 Insofar as CO has intact left para-hippocampal cortex this suggests that the left PPAmight be sufficient to encode a certain amount of(perhaps semantic) information about real-worldscenes but the right PPA is necessary to encode
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 505
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)

tion of the geometry of a scene or object andmaintain it for 10 s in the absence of the originalstimulus Not only does this result suggest that per-ception of scene-like stimuli is intact in thesepatients it further suggests that some short-termmemory functions are intact as well However we
should note that subjects in this experiment werefree to devote all their effort and attention in the 10-second blank interval to maintaining a memory ofthe stimulus Thus it is possible that GR and COcan use rehearsal mechanisms to maintain the stim-ulus in short-term memory during the blank inter-
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 497
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
(b) Lego object version
Figure 7 Stimuli and procedure for Experiment 2 (a) In the Lego layout version subjects see a Lego layout for 3 s followed by a blankscreen for 10 s A second layout then appears which is either the same layout as the first one but seen from a different angle or a differentlayout (b) The Lego object version is the same but with object stimuli
3 s 10 s OR
3 s 10 s OR
(a) Lego layout version
(b) Lego object version
val but are unable to form a more durable memorytrace for the stimulus This possibility was tested inthe current experiment which was designed toexamine GR and COrsquos ability to learn new visualinformation
We devised a continuous n-back recognitionmemory test in which subjects viewed a sequence oforiginally novel stimuli some of which wererepeated within the sequence at intervals of 1 3 or5 intervening images For each stimulus subjects
reported whether they had seen it before in theexperiment or not In different runs in the experi-ment the stimuli were black-and-white photo-graphs of (1) Lego scenes (2) Lego objects (3) real-world landscapes (4) novel objects made out ofSculpey clay (5) unfamiliar houses (6) unfamiliarfaces as well as (7) line drawings of nonsenseobjects and (8) pronounceable pseudowords (Fig-ure 9) Thus we were able to test GR and COrsquos rec-ognition memory for different kinds of materials atdifferent time intervals in a paradigm in which thepresence of intervening items between the twoappearances of each stimulus made rehearsal diffi-cult Note that because none of the specific stimuliused in this experiment were familiar in advancethis task did not require an episodic memory com-ponent and could be done on the basis of pure visualfamiliarity with the particular visual stimulus
Although many different stimulus classes weretested in this experiment the critical comparison isbetween the Lego layouts and Lego objectsbecause these are best matched with regard tomaterials and differ primarily in their geometricstructure (see Introduction) We predicted that ifGR and CO showed a memory deficit for this taskit would be greater for the Lego layouts than for the
EPSTEIN ET AL
498 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 8 Results of Experiment 2 Performance over ldquosamerdquotrials and ldquodifferentrdquo trials is averaged together Error bars show1 SD
Figure 9 Stimuli for Experiment 3 Top row Lego layouts real-world scenes houses Bottom row Lego objects clay objects nonsenseobjects faces In an eighth condition (not shown) subjects viewed pronounceable nonwords such as ldquowilchrdquo or ldquothipperrdquo
Lego objects In addition a deficit for materialsthat convey information about the layout of sur-rounding space might also be apparent when GRand CO are tested on more realistic stimuli If soone would expect them to do poorly on the land-scapes but relatively well on the clay objects linedrawings of nonsense objects faces andpseudowords
MethodFor each subject a complete experimental sessionconsisted of being run once on each of the eightstimulus classes Each run consisted of 90 trials ineach trial a stimulus was presented and subjectsreported whether or not they had seen it before inthe run (or indeed ever) by pressing the ldquojrdquo key ifthey had or the ldquof rdquo key if they had not Each stimu-lus was on the screen for either 1 2 or 3 s (see fol-lowing) and appeared promptly after the keypressreport for the previous trial Individual stimuliappeared either once (14 stimuli) twice (30 stim-uli) or four times (4 stimuli) within a given run Ofthe stimuli that were presented twice 10 wererepeated immediately (1-back) 10 were repeatedwith a lag of two intervening items (3-back) and 10were repeated with a lag of four intervening items(5-back) In order to ensure that between-subjectdifferences in performance were not due to stimulusdifferences the same stimuli were assigned to thesame conditions each time the experimentwas run
GR performed the experiment twice once inAugust 1998 and again 9 months later Because ofa computer error he was only tested on six of theeight categories (all except Lego layouts and Legoobjects) at the later date CO was also tested twiceonce in February 1999 and again 6 months laterReported results for GR and CO are the average oftheir two testing sessions Normal subjects wereonly tested once Stimulus presentation time was 1 sin GRrsquos first testing session In pilot testing COfound this too fast for the critical categories of Lego
layouts and Lego objects so presentation time wasextended to 2 s for these categories in his first test-ing session All normal subjects were tested at thesepresentation rates (2 s for Lego layouts and objects1 s for other stimuli) In COrsquos second testingsession presentation time was lengthened to 3 s forall stimulus categories to ensure that any deficitsfound were memory deficits rather than problemswith the nonmemory aspects of the task
Results and discussionResults from GR CO and 10 elderly normal con-trols are shown in Figure 10 and Table 4 Figure 10ashows results from the main comparison betweenLego layouts and Lego objects Both normal sub-jects and patients performed near ceiling for bothLego layouts and Lego objects at 1-backinterstimulus intervals verifying that GR and COcould successfully perceive the pictures as they werebeing presented In contrast there was a strikingdifference between the two patients and the normalsubjects at 5-back intervals Whereas normals didon average somewhat better on Lego layouts thanLego objects both GR and CO showed the oppo-site pattern doing much worse on Lego layoutsthan Lego objects The 3-back condition showed amixed pattern Whereas GR did better on Legoobjects than Lego layouts in this condition (similarto his performance at 5-back intervals) CO didbetter on Lego layouts than Lego objects COrsquos lowperformance on Lego objects in the 3-back condi-tion appears to be an anomaly as it is much lowerthan his performance for comparable stimuli in the1-back and 5-back conditions
Figure 10b shows the difference in performancebetween Lego layouts and Lego objects for thecritical 1-back and 5-back intervals At 5-backintervals CO was significantly impaired comparedto the normal subjects (z = ndash250) whereasGRrsquos impairment fell just short of significance (z =ndash175)4
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 499
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
4In fact these values may underestimate GR and COrsquos relative impairment for Lego layouts In the 5-back condition 4 of the 10
normal subjects had perfect performance on the Lego layouts but all scored below ceiling on the Lego objects Thus the true advantagefor Lego layouts over Lego objects for normal subjects is probably greater than that observed here in which case GR and COrsquos lowerperformance for Lego layouts compared to Lego objects would be even more abnormal Furthermore post-hoc analyses revealed thatone of the normal subjects showed a difference in performance between Lego layouts and objects that was over 2 SD below the mean ofthe other nine subjects When this outlying subject was excluded z-scores were ndash322 for CO and ndash232 for GR
Table 4 shows results for all eight stimulusclasses Although direct comparisons between sub-jects should be treated with caution since viewingtime for different items was not the same for eachsubject we do note a few interesting patterns Firstof all there is little or no evidence that GR or CO
show a memory deficit for nonsense words clayobjects or line drawings of nonsense objects GRappears to be slightly impaired on faces and housesbut CO performs normally on these categoriesStrikingly GR shows a severe deficit for the reallandscapes performing more than 40 percentagepoints lower than normals Thus even thoughthere is some evidence that GR might have someimpairment for nonscene stimuli he clearly is muchmore impaired for scene-like stimuli such as theLego scenes and landscapes CO does not show asimilar deficit for the landscapes However itshould be kept in mind that in one of his two testingsessions CO viewed each picture for 3 s which wasmuch longer than the 1 second used for all non-Lego stimuli for GR and normals Thus COrsquosapparently ldquonormalrdquo performance in these condi-tions might be overestimated
Taken together the results of this experimentindicate that GR and CO are selectively impaired atprocessing scene-like stimuli This deficit is consis-tently apparent at 5-back but not 1-back intervalsThis finding suggests that the deficit should becharacterised as a memory encoding problemHowever the present results do not allow us todetermine whether the difficulty occurs at memoryencoding or at subsequent recognition (see Epsteinamp Kanwisher 2001) If GR and CO have a generalproblem with recognition of topographical materi-als then they should not be able to recognise scenes
EPSTEIN ET AL
500 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 10a Results of Experiment 3 for GR (top) CO (middle)and for 10 age-matched normal controls Performance measure isproportion of repeated stimuli correctly identified as such correctedfor guessing using the standard formula
Figure 10b Difference between Lego layout performance andLego object performance for the 1-back and 5-back conditionsError bars indicate one 1 SD
and landmarks that were learned prior to theirinjury We tested this possibility in the nextexperiment
Experiment 4 Famous landmarksfacesrecognition task
In this experiment we examined GR and COrsquosability to recognise the visual appearances of placeslearned prior to injury Subjects saw a series offamous landmarks intermixed with nonfamouscontrols (Imaging experiments have demonstratedthat landmarks of this type can activate the PPAalmost as strongly as complete scenes Epstein etal 1999 Epstein amp Kanwisher 1998) They wererequired to say whether they knew each specificlandmark and to identify the ones they knew Sub-jects were instructed to respond affirmatively if theywere fairly certain that they recognised the land-mark but not if it only looked generally familiarFor purposes of comparison subjects also per-formed an identical test in which the stimuli werefamous and nonfamous faces In both testsdistractor items were designed to be visually andconceptually similar to the target items (see Figure11) If parahippocampal cortex is required for placerecognition then one might expect GR and CO toperform poorly compared to normals on the land-mark recognition test
MethodEach of the two experiments (famous faces famouslandmarks) consisted of 48 trials Subjects beganeach trial by pressing the space bar after which ablack-and-white photograph of a face (in thefamous faces experiment) or a landmark (in thefamous landmarks experiment) appeared on thescreen Subjects were told to press the ldquojrdquo key if theyrecognised the specific face or landmark and the ldquof rdquokey if they did not taking as much time at theyneeded to make this discrimination but no extratime Subjects were told to report that they recog-nised a face or landmark if they felt that they knewit even if they could not retrieve its name If theydid know the name they were to say it after pressingthe button Otherwise they were to say whateverthey did remember about the item
Half of the photographs were of famous and halfof nonfamous people and landmarks The famousfaces included public figures (eg George BushRichard Nixon Prince Charles) and movie stars(eg Harrison Ford Clark Gable Marlon Brando)who were prominent prior to the time of GR andCOrsquos injuries the famous landmarks were build-ings and monuments from all over the world (egEiffel Tower US Capitol Building Great Sphinx)The nonfamous stimuli were chosen to match thefamous stimuli as closely as possible Each famousstimulus had a ldquomatchrdquo For example George Bushwas matched to a man of similar age in a coat andtie and the Taj Mahal was matched to a visually
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 501
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 4 Full results of Experiment 3 Mean proportion correct (corrected for guessing) SDs in parentheses
Lego Land- Lego Clay Nonsense Non-scenes scapes Houses objects objects objects Words Faces
1-back GR 100 094 100 100 100 094 100 100CO 091 085 091 100 100 079 074 100Normals 096(008) 095(008) 093(014) 096(008) 096(006) 099(003) 098(004) 097(011)
3-back GR ndash003 068 026 047 083 083 075 052CO 066 081 037 010 094 069 095 081Normals 066(032) 077(021) 072(015) 077(020) 086(016) 083(015) 083(017) 072(020)
5-back GR 010 034 044 047 072 083 094 053CO 031 073 075 088 081 064 100 081Normals 076(026) 077(016) 068(021) 066(016) 091(014) 083(018) 093(011) 078(021)
similar Asian palace that is not famous (see Figure11)
Results and discussionResults for GR and CO as well as for five age-matched controls are shown in Figure 12 Both
GR and CO could recognise the famous landmarksas well normals In addition both GR and CO wereusually able to name the landmarks they success-fully identified
Thus GR and CO can recognize landmarkslearned prior to injury demonstrating that at leastsome of their place recognition abilities are intactWhen combined with the fact that GR and COwere able to produce detailed maps of places theylived in prior to their injury (see Case Description)these results suggest that their deficit is primarily inencoding or consolidation of new informationrather than in recognition or recall of old informa-tion Note however that these results do not provethat their place recognition abilities are entirelynormal GR and CO might suffer from a mild rec-ognition deficit that affects identification of newly-learned stimuli like the Lego layouts in Experiment3 but not overlearned stimuli like the famous land-marks Furthermore it is possible that GR and COmight be impaired at recognising premorbidlylearned places that are defined primarily by theirspatial layout even though they are unimpaired atrecognising premorbidly learned places that aredefined primarily by a single object-like landmarkor building (see General Discussion) Neverthelessthe trend of the present evidence is to suggest thatGR and CO have particular problems with noveltopographical information
In the famous-face task GRrsquos performance wasin the normal range However he sometimes haddifficulty naming the faces even though he clearly
EPSTEIN ET AL
502 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 11 Stimuli for Experiment 4 Each famous landmark(Arc de Triomphe Taj Mahal) and famous face (Barbara BushYasser Arafat) was matched to a nonfamous building or face thatalso appeared in the experiment
FA
CE
SL
AN
DM
AR
KS
Figure 12 Results of Experiment 4 Performance measure isproportion of famous stimuli reported as ldquoknownrdquo corrected forguessing using the standard formula
recognised them For example he described Ron-ald Reagan as ldquothe gipperrdquo Nancy Reagan as ldquothewife of the gipperrdquo and Clark Gable as ldquothe movieactor who said lsquoScarlett I donrsquot give a damnrsquordquo COhad a great deal of difficulty identifying the famousfaces This result is not surprising given hisextremely low performance on the Benton FaceMatching Task In addition to his topographicproblems CO appears to suffer fromprosopagnosia
GENERAL DISCUSSION
We tested two parahippocampal patients (GR andCO) on four experiments designed to measure per-formance as a function of both task (perceptionencoding and recognition) and stimulus type(scenes objects faces and words) The only taskthat revealed a clear deficit was an n-back recogni-tion memory test (Experiment 3) In order to suc-cessfully perform this task subjects had to encode adurable representation of a visual stimulus and useit for later recognition GR and CO were signifi-cantly more impaired on this task when the stimuliwere scene-like spatial layouts (ie Lego scenesand real scenes for GR) than when they wereobjects No comparable deficits were found in otherexperiments that tested layout perception (Experi-ments 1 and 2) and placelandmark recognition(Experiment 4) Taken together these results indi-cate that GR and CO are impaired in their ability toencode novel information about the geometry ofsurrounding space into memory but are lessimpaired at their ability to encode the geometry ofnovel objects
We argue that this specific topographical defi-citmdashinability to encode the spatial layout of novelscenesmdashresults from damage to the para-hippocampal place area Although the damage inboth patients extends beyond the parahippo-campal-lingual territory normally associated withthe PPA this region is clearly lesioned bilaterally inGR and in the right hemisphere in CO Further-more functional imaging demonstrated that GRhas no functioning PPA Thus the current resultsand the earlier fMRI work (Epstein et al 1999
Epstein amp Kanwisher 1998) converge to implicateparahippocampal cortex in the encoding of infor-mation about scene geometry into memory
Our assignment of the function of memoryencoding to parahippocampal cortex is consistentwith the results of earlier case reports of patientswith parahippocampal damage who have naviga-tional difficulties in novel but not familiar environ-ments (Habib amp Sirigu 1987 Ross 1980 see alsoTeng amp Squire 1999) As with GR and CO thesepatients were either less impaired or unimpaired onnontopographical memory tasks Patients have alsobeen described who exhibit the opposite pattern ofimpairment an inability to learn faces (sometimescalled ldquoprosopamnesiardquo Cipolotti Robinson Blairamp Frith 1999 Tippett Miller amp Farah 2000) orfaces and verbal materials (Maguire amp Cipolotti1998) with an unimpaired ability to learn topo-graphical materials Taken together these resultsargue for a high degree of domain-specificity in theneural systems involved in learning of new informa-tion (Gallistel 1995) The brain may have separatemechanisms for learning faces places and wordswhich can be differentially impaired after damageto specific cortical regions (Cipolotti et al 1999Incisa della Rochetta et al 1996) The presentresults suggest that the anatomical locus of themechanism dedicated to learning new topographi-cal information is in the parahippocampal cortex(Aguirre amp DrsquoEsposito 1999)
In contrast to their impaired ability to encodenew topographical information both GR and COwere able to produce detailed maps of their previousdwellings and performed normally on the famouslandmark recognition test of Experiment 4 Theseresults are generally consistent with Teng andSquirersquos (1999) report of a patient with extensivemedial temporal damage who could recallpremorbidly learned but not postmorbidly learnedtopographical information (see also Ross 1980)Thus our results provide further evidence thatmedial temporal regions such as parahippocampalcortex may be more critical for encoding or consoli-dating new information into memory than for recallof old information
One issue that our experiments do not entirelyresolve is the precise nature of the topographical
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 503
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
information encoded by parahippocampal cortexClearly this region is critical for encoding informa-tion about the geometry of the currently visiblescene even when the ldquoscenerdquo is made out of Legoblocks and therefore has no immediate navigationalrelevance However it is unclear whether this spe-cific deficit can account for all of the real-worldnavigational difficulties encountered by GR andCO GR in particular shows such a catastrophicloss of navigational ability that we suspect he is alsoimpaired at encoding the spatial relationshipsbetweenthe current scene and other locations in theworld CO probably also suffers from such difficul-ties as he reports that he has trouble rememberinghow places relate to each other in large-scale spaceWhen he was tested on a simple 3-room videogameenvironment he not only was unable to rememberthe individual rooms (claiming that there were 10rooms when there were only 3) but he also had nomemory for the spatial relationships between themThese spatial deficits may be the result of damage tothe posterior right hippocampus (in the case of CO)or of disconnection of the posterior right hippo-campus from the parahippocampal regions thatprovide much of its cortical input (in both patientsSuzuki amp Amaral 1994) In either case the abilityto form a cognitive map of new environments likethe lab or videogame space could be impaired(Maguire et al 1998 OrsquoKeefe amp Nadel 1978)However it is also possible that parahippocampalcortex itself may be critical for encoding not onlythe geometry of individual scenes but also the spa-tial relationships between the locations of thosescenes In this case the spatial deficits of GR andCO could be attributed directly to parahippo-campal damage rather than to hippocampal dam-age or disconnection
The latter scenario would be consistent withAguirre and DrsquoEspositorsquos (1999) and Barrash etalrsquos (2000) suggestion that parahippocampal cortexis critically involved in combining visual informa-tion about the appearance of scenes and landmarkswith spatial information about their locations into aunified topographical representation Howeverboth of these authors argue that the representationsof individual scenes (Barrash et al) and landmarks(Aguirre amp DrsquoEsposito) are supported not by
parahippocampal cortex but by more posterior lin-gual-fusiform regions In contrast we believe thatthe neuroimaging data indicate that parahippo-campal cortex is involved in the encoding of thegeometric structure of individual scenes althoughit may also play a role in combining these represen-tations with other spatial representations
Interestingly GR and CO were not impaired onrecognition of famous landmarks that were mostlybuildings Thus our data do not preclude the possi-bility that there may be a separate lingual orfusiform region involved in processing informationabout buildings (Aguirre Zarahn amp DrsquoEsposito1998 Chao Haxby amp Martin 1999 IshaiUngerleider Martin Schouten amp Haxby 1999)which might have been preserved in GR and COdespite extensive damage to other regions of thelingual and fusiform gyrus Alternatively GR andCOrsquos preserved building recognition abilities mightbe mediated by interactions between cortical areasthat perform shape analysis for any object-likestimulus type (Malach et al 1995Kanwisher et al1996) and anterior temporal regions in which long-term topographical and semantic memories arestored (McCarthy et al 1996)
Is perception preserved
In contrast to their inability to encode new scenesinto memory GR and CO have substantiallyretained their ability to perceive the spatial layout ofthe currently visible scene (Experiments 1 and 2)GR can follow a route marked on a floor planwhich indicates a good on-line appreciation of hiscurrent spatial surroundings Furthermore neitherGR or CO have any trouble moving through theenvironmentmdashthey move with confidence and donot bump into doors or walls Problems of the lattertype are usually associated with damage to the pari-etal lobes which are intact in GR and CO Thus atfirst glance GR and CO do not appear to haveperceptual impairments (beyond their visual fieldrestrictions)
However GR does complain of a perceptualdeficit which he describes as an inability to ldquotakeinrdquo scenes when they are too complex Similarreports of an inability to comprehend the
EPSTEIN ET AL
504 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
ldquoglobalityrdquo of a scene were made by some of theparahippocampally-damaged patients studied byHabib and Sirigu (1987) We were not able todetermine the precise nature of this deficit whichmay be partly attributable to his visual field restric-tions It appears that his deficit is more salient in thereal-world environments than in the picture-per-ception tasks of Experiments 1 and 2 perhapsbecause he finds it more difficult to organise visualinput when immersed in it than when viewing pho-tographs that subtend only part of the visual field
We suggest that GR can perceive parts of thescene when he attends to them but may be unableto integrate information over time into a coherentrepresentation of a scene or place The fact that GRand CO are impaired for Lego layouts at 5-backintervals in Experiment 3 is consistent with thishypothesis The time between the first and secondpresentations of a stimulus is very short in this case(and even shorter with 3-back intervals where GRalso shows evidence of a deficit) In both patientsthe representation of layout information formed byperception of a Lego scene must either have anabnormally short persistence be abnormally sus-ceptible to interference or both If so they mightfind it impossible to sustain a spatial frame orschema long enough to use it to organise informa-tion obtained from many different glances at a com-plex scene (Intraub 1997) The result would be aninability to understand the ldquoglobalityrdquo of a sceneeven though all the details were perfectlyperceived
Non-topographical mnemonic abilities
Although not as severe as their topographical mem-ory problems GR and CO do have some memorydifficulties in other domains For example bothperformed poorly on the Rey AVLT whichrequired them to remember a list of unrelatedwords Both also report some subtle episodic mem-ory problems (see Case Description) COrsquos prob-lems could partly result from damage to the righthippocampusmdashindeed CO appeared to have themore acute episodic memory problems HoweverGR has no hippocampal damage so the fact that hedisplays episodic memory problems suggests thatparahippocampal cortex is involved in the forma-
tion of episodic memories (cf Wagner et al1998)
In previous work (Epstein et al 1999) weargued that spatial context information encoded byparahippocampal cortex might play a role in estab-lishing event memories by encoding informationabout where events take place The inability toencode this information might cause problems intasks such as the Rey AVLT where one has to recalla list of unrelated words If spatial (and perhapstemporal) context information is not encoded itwill be harder subsequently to recall the distinctevent of hearing each word (Tulving 1983) Incontrast lack of spatial context information wouldbe less of a problem in tasks where one is onlyrequired to recognise previously encountered verbalmaterials but not to reproduce the spatiotemporalcontext in which they were experienced IndeedGR and CO performed as well as normals on thenonsense words in Experiment 4 where recogni-tion memory but not episodic memory was requiredfor successful performance Interestingly GR andCO also did relatively well on the Logical Memorysubtest of the WMS-III perhaps because the nar-rative framework of a story provides sufficient con-textual information to overcome the absence ofspatial context and mediate successful verbalretrieval
Differences between GR and CO
Although we have discussed GR and CO togetherthere are a number of relevant differences betweenthem One open question is whether these differ-ences can be attributed to the different effects ofunilateral and bilateral parahippocampal damageIn general GRrsquos topographical difficulties weremore severe than COrsquos CO did surprisingly well onthe Warrington Topographical Memory Test inwhich he was required to remember detailed real-world city scenes and also performed substantiallybetter on real scenes than Lego scenes in Experi-ment 3 Insofar as CO has intact left para-hippocampal cortex this suggests that the left PPAmight be sufficient to encode a certain amount of(perhaps semantic) information about real-worldscenes but the right PPA is necessary to encode
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 505
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)

val but are unable to form a more durable memorytrace for the stimulus This possibility was tested inthe current experiment which was designed toexamine GR and COrsquos ability to learn new visualinformation
We devised a continuous n-back recognitionmemory test in which subjects viewed a sequence oforiginally novel stimuli some of which wererepeated within the sequence at intervals of 1 3 or5 intervening images For each stimulus subjects
reported whether they had seen it before in theexperiment or not In different runs in the experi-ment the stimuli were black-and-white photo-graphs of (1) Lego scenes (2) Lego objects (3) real-world landscapes (4) novel objects made out ofSculpey clay (5) unfamiliar houses (6) unfamiliarfaces as well as (7) line drawings of nonsenseobjects and (8) pronounceable pseudowords (Fig-ure 9) Thus we were able to test GR and COrsquos rec-ognition memory for different kinds of materials atdifferent time intervals in a paradigm in which thepresence of intervening items between the twoappearances of each stimulus made rehearsal diffi-cult Note that because none of the specific stimuliused in this experiment were familiar in advancethis task did not require an episodic memory com-ponent and could be done on the basis of pure visualfamiliarity with the particular visual stimulus
Although many different stimulus classes weretested in this experiment the critical comparison isbetween the Lego layouts and Lego objectsbecause these are best matched with regard tomaterials and differ primarily in their geometricstructure (see Introduction) We predicted that ifGR and CO showed a memory deficit for this taskit would be greater for the Lego layouts than for the
EPSTEIN ET AL
498 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 8 Results of Experiment 2 Performance over ldquosamerdquotrials and ldquodifferentrdquo trials is averaged together Error bars show1 SD
Figure 9 Stimuli for Experiment 3 Top row Lego layouts real-world scenes houses Bottom row Lego objects clay objects nonsenseobjects faces In an eighth condition (not shown) subjects viewed pronounceable nonwords such as ldquowilchrdquo or ldquothipperrdquo
Lego objects In addition a deficit for materialsthat convey information about the layout of sur-rounding space might also be apparent when GRand CO are tested on more realistic stimuli If soone would expect them to do poorly on the land-scapes but relatively well on the clay objects linedrawings of nonsense objects faces andpseudowords
MethodFor each subject a complete experimental sessionconsisted of being run once on each of the eightstimulus classes Each run consisted of 90 trials ineach trial a stimulus was presented and subjectsreported whether or not they had seen it before inthe run (or indeed ever) by pressing the ldquojrdquo key ifthey had or the ldquof rdquo key if they had not Each stimu-lus was on the screen for either 1 2 or 3 s (see fol-lowing) and appeared promptly after the keypressreport for the previous trial Individual stimuliappeared either once (14 stimuli) twice (30 stim-uli) or four times (4 stimuli) within a given run Ofthe stimuli that were presented twice 10 wererepeated immediately (1-back) 10 were repeatedwith a lag of two intervening items (3-back) and 10were repeated with a lag of four intervening items(5-back) In order to ensure that between-subjectdifferences in performance were not due to stimulusdifferences the same stimuli were assigned to thesame conditions each time the experimentwas run
GR performed the experiment twice once inAugust 1998 and again 9 months later Because ofa computer error he was only tested on six of theeight categories (all except Lego layouts and Legoobjects) at the later date CO was also tested twiceonce in February 1999 and again 6 months laterReported results for GR and CO are the average oftheir two testing sessions Normal subjects wereonly tested once Stimulus presentation time was 1 sin GRrsquos first testing session In pilot testing COfound this too fast for the critical categories of Lego
layouts and Lego objects so presentation time wasextended to 2 s for these categories in his first test-ing session All normal subjects were tested at thesepresentation rates (2 s for Lego layouts and objects1 s for other stimuli) In COrsquos second testingsession presentation time was lengthened to 3 s forall stimulus categories to ensure that any deficitsfound were memory deficits rather than problemswith the nonmemory aspects of the task
Results and discussionResults from GR CO and 10 elderly normal con-trols are shown in Figure 10 and Table 4 Figure 10ashows results from the main comparison betweenLego layouts and Lego objects Both normal sub-jects and patients performed near ceiling for bothLego layouts and Lego objects at 1-backinterstimulus intervals verifying that GR and COcould successfully perceive the pictures as they werebeing presented In contrast there was a strikingdifference between the two patients and the normalsubjects at 5-back intervals Whereas normals didon average somewhat better on Lego layouts thanLego objects both GR and CO showed the oppo-site pattern doing much worse on Lego layoutsthan Lego objects The 3-back condition showed amixed pattern Whereas GR did better on Legoobjects than Lego layouts in this condition (similarto his performance at 5-back intervals) CO didbetter on Lego layouts than Lego objects COrsquos lowperformance on Lego objects in the 3-back condi-tion appears to be an anomaly as it is much lowerthan his performance for comparable stimuli in the1-back and 5-back conditions
Figure 10b shows the difference in performancebetween Lego layouts and Lego objects for thecritical 1-back and 5-back intervals At 5-backintervals CO was significantly impaired comparedto the normal subjects (z = ndash250) whereasGRrsquos impairment fell just short of significance (z =ndash175)4
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 499
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
4In fact these values may underestimate GR and COrsquos relative impairment for Lego layouts In the 5-back condition 4 of the 10
normal subjects had perfect performance on the Lego layouts but all scored below ceiling on the Lego objects Thus the true advantagefor Lego layouts over Lego objects for normal subjects is probably greater than that observed here in which case GR and COrsquos lowerperformance for Lego layouts compared to Lego objects would be even more abnormal Furthermore post-hoc analyses revealed thatone of the normal subjects showed a difference in performance between Lego layouts and objects that was over 2 SD below the mean ofthe other nine subjects When this outlying subject was excluded z-scores were ndash322 for CO and ndash232 for GR
Table 4 shows results for all eight stimulusclasses Although direct comparisons between sub-jects should be treated with caution since viewingtime for different items was not the same for eachsubject we do note a few interesting patterns Firstof all there is little or no evidence that GR or CO
show a memory deficit for nonsense words clayobjects or line drawings of nonsense objects GRappears to be slightly impaired on faces and housesbut CO performs normally on these categoriesStrikingly GR shows a severe deficit for the reallandscapes performing more than 40 percentagepoints lower than normals Thus even thoughthere is some evidence that GR might have someimpairment for nonscene stimuli he clearly is muchmore impaired for scene-like stimuli such as theLego scenes and landscapes CO does not show asimilar deficit for the landscapes However itshould be kept in mind that in one of his two testingsessions CO viewed each picture for 3 s which wasmuch longer than the 1 second used for all non-Lego stimuli for GR and normals Thus COrsquosapparently ldquonormalrdquo performance in these condi-tions might be overestimated
Taken together the results of this experimentindicate that GR and CO are selectively impaired atprocessing scene-like stimuli This deficit is consis-tently apparent at 5-back but not 1-back intervalsThis finding suggests that the deficit should becharacterised as a memory encoding problemHowever the present results do not allow us todetermine whether the difficulty occurs at memoryencoding or at subsequent recognition (see Epsteinamp Kanwisher 2001) If GR and CO have a generalproblem with recognition of topographical materi-als then they should not be able to recognise scenes
EPSTEIN ET AL
500 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 10a Results of Experiment 3 for GR (top) CO (middle)and for 10 age-matched normal controls Performance measure isproportion of repeated stimuli correctly identified as such correctedfor guessing using the standard formula
Figure 10b Difference between Lego layout performance andLego object performance for the 1-back and 5-back conditionsError bars indicate one 1 SD
and landmarks that were learned prior to theirinjury We tested this possibility in the nextexperiment
Experiment 4 Famous landmarksfacesrecognition task
In this experiment we examined GR and COrsquosability to recognise the visual appearances of placeslearned prior to injury Subjects saw a series offamous landmarks intermixed with nonfamouscontrols (Imaging experiments have demonstratedthat landmarks of this type can activate the PPAalmost as strongly as complete scenes Epstein etal 1999 Epstein amp Kanwisher 1998) They wererequired to say whether they knew each specificlandmark and to identify the ones they knew Sub-jects were instructed to respond affirmatively if theywere fairly certain that they recognised the land-mark but not if it only looked generally familiarFor purposes of comparison subjects also per-formed an identical test in which the stimuli werefamous and nonfamous faces In both testsdistractor items were designed to be visually andconceptually similar to the target items (see Figure11) If parahippocampal cortex is required for placerecognition then one might expect GR and CO toperform poorly compared to normals on the land-mark recognition test
MethodEach of the two experiments (famous faces famouslandmarks) consisted of 48 trials Subjects beganeach trial by pressing the space bar after which ablack-and-white photograph of a face (in thefamous faces experiment) or a landmark (in thefamous landmarks experiment) appeared on thescreen Subjects were told to press the ldquojrdquo key if theyrecognised the specific face or landmark and the ldquof rdquokey if they did not taking as much time at theyneeded to make this discrimination but no extratime Subjects were told to report that they recog-nised a face or landmark if they felt that they knewit even if they could not retrieve its name If theydid know the name they were to say it after pressingthe button Otherwise they were to say whateverthey did remember about the item
Half of the photographs were of famous and halfof nonfamous people and landmarks The famousfaces included public figures (eg George BushRichard Nixon Prince Charles) and movie stars(eg Harrison Ford Clark Gable Marlon Brando)who were prominent prior to the time of GR andCOrsquos injuries the famous landmarks were build-ings and monuments from all over the world (egEiffel Tower US Capitol Building Great Sphinx)The nonfamous stimuli were chosen to match thefamous stimuli as closely as possible Each famousstimulus had a ldquomatchrdquo For example George Bushwas matched to a man of similar age in a coat andtie and the Taj Mahal was matched to a visually
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 501
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 4 Full results of Experiment 3 Mean proportion correct (corrected for guessing) SDs in parentheses
Lego Land- Lego Clay Nonsense Non-scenes scapes Houses objects objects objects Words Faces
1-back GR 100 094 100 100 100 094 100 100CO 091 085 091 100 100 079 074 100Normals 096(008) 095(008) 093(014) 096(008) 096(006) 099(003) 098(004) 097(011)
3-back GR ndash003 068 026 047 083 083 075 052CO 066 081 037 010 094 069 095 081Normals 066(032) 077(021) 072(015) 077(020) 086(016) 083(015) 083(017) 072(020)
5-back GR 010 034 044 047 072 083 094 053CO 031 073 075 088 081 064 100 081Normals 076(026) 077(016) 068(021) 066(016) 091(014) 083(018) 093(011) 078(021)
similar Asian palace that is not famous (see Figure11)
Results and discussionResults for GR and CO as well as for five age-matched controls are shown in Figure 12 Both
GR and CO could recognise the famous landmarksas well normals In addition both GR and CO wereusually able to name the landmarks they success-fully identified
Thus GR and CO can recognize landmarkslearned prior to injury demonstrating that at leastsome of their place recognition abilities are intactWhen combined with the fact that GR and COwere able to produce detailed maps of places theylived in prior to their injury (see Case Description)these results suggest that their deficit is primarily inencoding or consolidation of new informationrather than in recognition or recall of old informa-tion Note however that these results do not provethat their place recognition abilities are entirelynormal GR and CO might suffer from a mild rec-ognition deficit that affects identification of newly-learned stimuli like the Lego layouts in Experiment3 but not overlearned stimuli like the famous land-marks Furthermore it is possible that GR and COmight be impaired at recognising premorbidlylearned places that are defined primarily by theirspatial layout even though they are unimpaired atrecognising premorbidly learned places that aredefined primarily by a single object-like landmarkor building (see General Discussion) Neverthelessthe trend of the present evidence is to suggest thatGR and CO have particular problems with noveltopographical information
In the famous-face task GRrsquos performance wasin the normal range However he sometimes haddifficulty naming the faces even though he clearly
EPSTEIN ET AL
502 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 11 Stimuli for Experiment 4 Each famous landmark(Arc de Triomphe Taj Mahal) and famous face (Barbara BushYasser Arafat) was matched to a nonfamous building or face thatalso appeared in the experiment
FA
CE
SL
AN
DM
AR
KS
Figure 12 Results of Experiment 4 Performance measure isproportion of famous stimuli reported as ldquoknownrdquo corrected forguessing using the standard formula
recognised them For example he described Ron-ald Reagan as ldquothe gipperrdquo Nancy Reagan as ldquothewife of the gipperrdquo and Clark Gable as ldquothe movieactor who said lsquoScarlett I donrsquot give a damnrsquordquo COhad a great deal of difficulty identifying the famousfaces This result is not surprising given hisextremely low performance on the Benton FaceMatching Task In addition to his topographicproblems CO appears to suffer fromprosopagnosia
GENERAL DISCUSSION
We tested two parahippocampal patients (GR andCO) on four experiments designed to measure per-formance as a function of both task (perceptionencoding and recognition) and stimulus type(scenes objects faces and words) The only taskthat revealed a clear deficit was an n-back recogni-tion memory test (Experiment 3) In order to suc-cessfully perform this task subjects had to encode adurable representation of a visual stimulus and useit for later recognition GR and CO were signifi-cantly more impaired on this task when the stimuliwere scene-like spatial layouts (ie Lego scenesand real scenes for GR) than when they wereobjects No comparable deficits were found in otherexperiments that tested layout perception (Experi-ments 1 and 2) and placelandmark recognition(Experiment 4) Taken together these results indi-cate that GR and CO are impaired in their ability toencode novel information about the geometry ofsurrounding space into memory but are lessimpaired at their ability to encode the geometry ofnovel objects
We argue that this specific topographical defi-citmdashinability to encode the spatial layout of novelscenesmdashresults from damage to the para-hippocampal place area Although the damage inboth patients extends beyond the parahippo-campal-lingual territory normally associated withthe PPA this region is clearly lesioned bilaterally inGR and in the right hemisphere in CO Further-more functional imaging demonstrated that GRhas no functioning PPA Thus the current resultsand the earlier fMRI work (Epstein et al 1999
Epstein amp Kanwisher 1998) converge to implicateparahippocampal cortex in the encoding of infor-mation about scene geometry into memory
Our assignment of the function of memoryencoding to parahippocampal cortex is consistentwith the results of earlier case reports of patientswith parahippocampal damage who have naviga-tional difficulties in novel but not familiar environ-ments (Habib amp Sirigu 1987 Ross 1980 see alsoTeng amp Squire 1999) As with GR and CO thesepatients were either less impaired or unimpaired onnontopographical memory tasks Patients have alsobeen described who exhibit the opposite pattern ofimpairment an inability to learn faces (sometimescalled ldquoprosopamnesiardquo Cipolotti Robinson Blairamp Frith 1999 Tippett Miller amp Farah 2000) orfaces and verbal materials (Maguire amp Cipolotti1998) with an unimpaired ability to learn topo-graphical materials Taken together these resultsargue for a high degree of domain-specificity in theneural systems involved in learning of new informa-tion (Gallistel 1995) The brain may have separatemechanisms for learning faces places and wordswhich can be differentially impaired after damageto specific cortical regions (Cipolotti et al 1999Incisa della Rochetta et al 1996) The presentresults suggest that the anatomical locus of themechanism dedicated to learning new topographi-cal information is in the parahippocampal cortex(Aguirre amp DrsquoEsposito 1999)
In contrast to their impaired ability to encodenew topographical information both GR and COwere able to produce detailed maps of their previousdwellings and performed normally on the famouslandmark recognition test of Experiment 4 Theseresults are generally consistent with Teng andSquirersquos (1999) report of a patient with extensivemedial temporal damage who could recallpremorbidly learned but not postmorbidly learnedtopographical information (see also Ross 1980)Thus our results provide further evidence thatmedial temporal regions such as parahippocampalcortex may be more critical for encoding or consoli-dating new information into memory than for recallof old information
One issue that our experiments do not entirelyresolve is the precise nature of the topographical
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 503
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
information encoded by parahippocampal cortexClearly this region is critical for encoding informa-tion about the geometry of the currently visiblescene even when the ldquoscenerdquo is made out of Legoblocks and therefore has no immediate navigationalrelevance However it is unclear whether this spe-cific deficit can account for all of the real-worldnavigational difficulties encountered by GR andCO GR in particular shows such a catastrophicloss of navigational ability that we suspect he is alsoimpaired at encoding the spatial relationshipsbetweenthe current scene and other locations in theworld CO probably also suffers from such difficul-ties as he reports that he has trouble rememberinghow places relate to each other in large-scale spaceWhen he was tested on a simple 3-room videogameenvironment he not only was unable to rememberthe individual rooms (claiming that there were 10rooms when there were only 3) but he also had nomemory for the spatial relationships between themThese spatial deficits may be the result of damage tothe posterior right hippocampus (in the case of CO)or of disconnection of the posterior right hippo-campus from the parahippocampal regions thatprovide much of its cortical input (in both patientsSuzuki amp Amaral 1994) In either case the abilityto form a cognitive map of new environments likethe lab or videogame space could be impaired(Maguire et al 1998 OrsquoKeefe amp Nadel 1978)However it is also possible that parahippocampalcortex itself may be critical for encoding not onlythe geometry of individual scenes but also the spa-tial relationships between the locations of thosescenes In this case the spatial deficits of GR andCO could be attributed directly to parahippo-campal damage rather than to hippocampal dam-age or disconnection
The latter scenario would be consistent withAguirre and DrsquoEspositorsquos (1999) and Barrash etalrsquos (2000) suggestion that parahippocampal cortexis critically involved in combining visual informa-tion about the appearance of scenes and landmarkswith spatial information about their locations into aunified topographical representation Howeverboth of these authors argue that the representationsof individual scenes (Barrash et al) and landmarks(Aguirre amp DrsquoEsposito) are supported not by
parahippocampal cortex but by more posterior lin-gual-fusiform regions In contrast we believe thatthe neuroimaging data indicate that parahippo-campal cortex is involved in the encoding of thegeometric structure of individual scenes althoughit may also play a role in combining these represen-tations with other spatial representations
Interestingly GR and CO were not impaired onrecognition of famous landmarks that were mostlybuildings Thus our data do not preclude the possi-bility that there may be a separate lingual orfusiform region involved in processing informationabout buildings (Aguirre Zarahn amp DrsquoEsposito1998 Chao Haxby amp Martin 1999 IshaiUngerleider Martin Schouten amp Haxby 1999)which might have been preserved in GR and COdespite extensive damage to other regions of thelingual and fusiform gyrus Alternatively GR andCOrsquos preserved building recognition abilities mightbe mediated by interactions between cortical areasthat perform shape analysis for any object-likestimulus type (Malach et al 1995Kanwisher et al1996) and anterior temporal regions in which long-term topographical and semantic memories arestored (McCarthy et al 1996)
Is perception preserved
In contrast to their inability to encode new scenesinto memory GR and CO have substantiallyretained their ability to perceive the spatial layout ofthe currently visible scene (Experiments 1 and 2)GR can follow a route marked on a floor planwhich indicates a good on-line appreciation of hiscurrent spatial surroundings Furthermore neitherGR or CO have any trouble moving through theenvironmentmdashthey move with confidence and donot bump into doors or walls Problems of the lattertype are usually associated with damage to the pari-etal lobes which are intact in GR and CO Thus atfirst glance GR and CO do not appear to haveperceptual impairments (beyond their visual fieldrestrictions)
However GR does complain of a perceptualdeficit which he describes as an inability to ldquotakeinrdquo scenes when they are too complex Similarreports of an inability to comprehend the
EPSTEIN ET AL
504 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
ldquoglobalityrdquo of a scene were made by some of theparahippocampally-damaged patients studied byHabib and Sirigu (1987) We were not able todetermine the precise nature of this deficit whichmay be partly attributable to his visual field restric-tions It appears that his deficit is more salient in thereal-world environments than in the picture-per-ception tasks of Experiments 1 and 2 perhapsbecause he finds it more difficult to organise visualinput when immersed in it than when viewing pho-tographs that subtend only part of the visual field
We suggest that GR can perceive parts of thescene when he attends to them but may be unableto integrate information over time into a coherentrepresentation of a scene or place The fact that GRand CO are impaired for Lego layouts at 5-backintervals in Experiment 3 is consistent with thishypothesis The time between the first and secondpresentations of a stimulus is very short in this case(and even shorter with 3-back intervals where GRalso shows evidence of a deficit) In both patientsthe representation of layout information formed byperception of a Lego scene must either have anabnormally short persistence be abnormally sus-ceptible to interference or both If so they mightfind it impossible to sustain a spatial frame orschema long enough to use it to organise informa-tion obtained from many different glances at a com-plex scene (Intraub 1997) The result would be aninability to understand the ldquoglobalityrdquo of a sceneeven though all the details were perfectlyperceived
Non-topographical mnemonic abilities
Although not as severe as their topographical mem-ory problems GR and CO do have some memorydifficulties in other domains For example bothperformed poorly on the Rey AVLT whichrequired them to remember a list of unrelatedwords Both also report some subtle episodic mem-ory problems (see Case Description) COrsquos prob-lems could partly result from damage to the righthippocampusmdashindeed CO appeared to have themore acute episodic memory problems HoweverGR has no hippocampal damage so the fact that hedisplays episodic memory problems suggests thatparahippocampal cortex is involved in the forma-
tion of episodic memories (cf Wagner et al1998)
In previous work (Epstein et al 1999) weargued that spatial context information encoded byparahippocampal cortex might play a role in estab-lishing event memories by encoding informationabout where events take place The inability toencode this information might cause problems intasks such as the Rey AVLT where one has to recalla list of unrelated words If spatial (and perhapstemporal) context information is not encoded itwill be harder subsequently to recall the distinctevent of hearing each word (Tulving 1983) Incontrast lack of spatial context information wouldbe less of a problem in tasks where one is onlyrequired to recognise previously encountered verbalmaterials but not to reproduce the spatiotemporalcontext in which they were experienced IndeedGR and CO performed as well as normals on thenonsense words in Experiment 4 where recogni-tion memory but not episodic memory was requiredfor successful performance Interestingly GR andCO also did relatively well on the Logical Memorysubtest of the WMS-III perhaps because the nar-rative framework of a story provides sufficient con-textual information to overcome the absence ofspatial context and mediate successful verbalretrieval
Differences between GR and CO
Although we have discussed GR and CO togetherthere are a number of relevant differences betweenthem One open question is whether these differ-ences can be attributed to the different effects ofunilateral and bilateral parahippocampal damageIn general GRrsquos topographical difficulties weremore severe than COrsquos CO did surprisingly well onthe Warrington Topographical Memory Test inwhich he was required to remember detailed real-world city scenes and also performed substantiallybetter on real scenes than Lego scenes in Experi-ment 3 Insofar as CO has intact left para-hippocampal cortex this suggests that the left PPAmight be sufficient to encode a certain amount of(perhaps semantic) information about real-worldscenes but the right PPA is necessary to encode
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 505
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)

Lego objects In addition a deficit for materialsthat convey information about the layout of sur-rounding space might also be apparent when GRand CO are tested on more realistic stimuli If soone would expect them to do poorly on the land-scapes but relatively well on the clay objects linedrawings of nonsense objects faces andpseudowords
MethodFor each subject a complete experimental sessionconsisted of being run once on each of the eightstimulus classes Each run consisted of 90 trials ineach trial a stimulus was presented and subjectsreported whether or not they had seen it before inthe run (or indeed ever) by pressing the ldquojrdquo key ifthey had or the ldquof rdquo key if they had not Each stimu-lus was on the screen for either 1 2 or 3 s (see fol-lowing) and appeared promptly after the keypressreport for the previous trial Individual stimuliappeared either once (14 stimuli) twice (30 stim-uli) or four times (4 stimuli) within a given run Ofthe stimuli that were presented twice 10 wererepeated immediately (1-back) 10 were repeatedwith a lag of two intervening items (3-back) and 10were repeated with a lag of four intervening items(5-back) In order to ensure that between-subjectdifferences in performance were not due to stimulusdifferences the same stimuli were assigned to thesame conditions each time the experimentwas run
GR performed the experiment twice once inAugust 1998 and again 9 months later Because ofa computer error he was only tested on six of theeight categories (all except Lego layouts and Legoobjects) at the later date CO was also tested twiceonce in February 1999 and again 6 months laterReported results for GR and CO are the average oftheir two testing sessions Normal subjects wereonly tested once Stimulus presentation time was 1 sin GRrsquos first testing session In pilot testing COfound this too fast for the critical categories of Lego
layouts and Lego objects so presentation time wasextended to 2 s for these categories in his first test-ing session All normal subjects were tested at thesepresentation rates (2 s for Lego layouts and objects1 s for other stimuli) In COrsquos second testingsession presentation time was lengthened to 3 s forall stimulus categories to ensure that any deficitsfound were memory deficits rather than problemswith the nonmemory aspects of the task
Results and discussionResults from GR CO and 10 elderly normal con-trols are shown in Figure 10 and Table 4 Figure 10ashows results from the main comparison betweenLego layouts and Lego objects Both normal sub-jects and patients performed near ceiling for bothLego layouts and Lego objects at 1-backinterstimulus intervals verifying that GR and COcould successfully perceive the pictures as they werebeing presented In contrast there was a strikingdifference between the two patients and the normalsubjects at 5-back intervals Whereas normals didon average somewhat better on Lego layouts thanLego objects both GR and CO showed the oppo-site pattern doing much worse on Lego layoutsthan Lego objects The 3-back condition showed amixed pattern Whereas GR did better on Legoobjects than Lego layouts in this condition (similarto his performance at 5-back intervals) CO didbetter on Lego layouts than Lego objects COrsquos lowperformance on Lego objects in the 3-back condi-tion appears to be an anomaly as it is much lowerthan his performance for comparable stimuli in the1-back and 5-back conditions
Figure 10b shows the difference in performancebetween Lego layouts and Lego objects for thecritical 1-back and 5-back intervals At 5-backintervals CO was significantly impaired comparedto the normal subjects (z = ndash250) whereasGRrsquos impairment fell just short of significance (z =ndash175)4
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 499
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
4In fact these values may underestimate GR and COrsquos relative impairment for Lego layouts In the 5-back condition 4 of the 10
normal subjects had perfect performance on the Lego layouts but all scored below ceiling on the Lego objects Thus the true advantagefor Lego layouts over Lego objects for normal subjects is probably greater than that observed here in which case GR and COrsquos lowerperformance for Lego layouts compared to Lego objects would be even more abnormal Furthermore post-hoc analyses revealed thatone of the normal subjects showed a difference in performance between Lego layouts and objects that was over 2 SD below the mean ofthe other nine subjects When this outlying subject was excluded z-scores were ndash322 for CO and ndash232 for GR
Table 4 shows results for all eight stimulusclasses Although direct comparisons between sub-jects should be treated with caution since viewingtime for different items was not the same for eachsubject we do note a few interesting patterns Firstof all there is little or no evidence that GR or CO
show a memory deficit for nonsense words clayobjects or line drawings of nonsense objects GRappears to be slightly impaired on faces and housesbut CO performs normally on these categoriesStrikingly GR shows a severe deficit for the reallandscapes performing more than 40 percentagepoints lower than normals Thus even thoughthere is some evidence that GR might have someimpairment for nonscene stimuli he clearly is muchmore impaired for scene-like stimuli such as theLego scenes and landscapes CO does not show asimilar deficit for the landscapes However itshould be kept in mind that in one of his two testingsessions CO viewed each picture for 3 s which wasmuch longer than the 1 second used for all non-Lego stimuli for GR and normals Thus COrsquosapparently ldquonormalrdquo performance in these condi-tions might be overestimated
Taken together the results of this experimentindicate that GR and CO are selectively impaired atprocessing scene-like stimuli This deficit is consis-tently apparent at 5-back but not 1-back intervalsThis finding suggests that the deficit should becharacterised as a memory encoding problemHowever the present results do not allow us todetermine whether the difficulty occurs at memoryencoding or at subsequent recognition (see Epsteinamp Kanwisher 2001) If GR and CO have a generalproblem with recognition of topographical materi-als then they should not be able to recognise scenes
EPSTEIN ET AL
500 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 10a Results of Experiment 3 for GR (top) CO (middle)and for 10 age-matched normal controls Performance measure isproportion of repeated stimuli correctly identified as such correctedfor guessing using the standard formula
Figure 10b Difference between Lego layout performance andLego object performance for the 1-back and 5-back conditionsError bars indicate one 1 SD
and landmarks that were learned prior to theirinjury We tested this possibility in the nextexperiment
Experiment 4 Famous landmarksfacesrecognition task
In this experiment we examined GR and COrsquosability to recognise the visual appearances of placeslearned prior to injury Subjects saw a series offamous landmarks intermixed with nonfamouscontrols (Imaging experiments have demonstratedthat landmarks of this type can activate the PPAalmost as strongly as complete scenes Epstein etal 1999 Epstein amp Kanwisher 1998) They wererequired to say whether they knew each specificlandmark and to identify the ones they knew Sub-jects were instructed to respond affirmatively if theywere fairly certain that they recognised the land-mark but not if it only looked generally familiarFor purposes of comparison subjects also per-formed an identical test in which the stimuli werefamous and nonfamous faces In both testsdistractor items were designed to be visually andconceptually similar to the target items (see Figure11) If parahippocampal cortex is required for placerecognition then one might expect GR and CO toperform poorly compared to normals on the land-mark recognition test
MethodEach of the two experiments (famous faces famouslandmarks) consisted of 48 trials Subjects beganeach trial by pressing the space bar after which ablack-and-white photograph of a face (in thefamous faces experiment) or a landmark (in thefamous landmarks experiment) appeared on thescreen Subjects were told to press the ldquojrdquo key if theyrecognised the specific face or landmark and the ldquof rdquokey if they did not taking as much time at theyneeded to make this discrimination but no extratime Subjects were told to report that they recog-nised a face or landmark if they felt that they knewit even if they could not retrieve its name If theydid know the name they were to say it after pressingthe button Otherwise they were to say whateverthey did remember about the item
Half of the photographs were of famous and halfof nonfamous people and landmarks The famousfaces included public figures (eg George BushRichard Nixon Prince Charles) and movie stars(eg Harrison Ford Clark Gable Marlon Brando)who were prominent prior to the time of GR andCOrsquos injuries the famous landmarks were build-ings and monuments from all over the world (egEiffel Tower US Capitol Building Great Sphinx)The nonfamous stimuli were chosen to match thefamous stimuli as closely as possible Each famousstimulus had a ldquomatchrdquo For example George Bushwas matched to a man of similar age in a coat andtie and the Taj Mahal was matched to a visually
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 501
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 4 Full results of Experiment 3 Mean proportion correct (corrected for guessing) SDs in parentheses
Lego Land- Lego Clay Nonsense Non-scenes scapes Houses objects objects objects Words Faces
1-back GR 100 094 100 100 100 094 100 100CO 091 085 091 100 100 079 074 100Normals 096(008) 095(008) 093(014) 096(008) 096(006) 099(003) 098(004) 097(011)
3-back GR ndash003 068 026 047 083 083 075 052CO 066 081 037 010 094 069 095 081Normals 066(032) 077(021) 072(015) 077(020) 086(016) 083(015) 083(017) 072(020)
5-back GR 010 034 044 047 072 083 094 053CO 031 073 075 088 081 064 100 081Normals 076(026) 077(016) 068(021) 066(016) 091(014) 083(018) 093(011) 078(021)
similar Asian palace that is not famous (see Figure11)
Results and discussionResults for GR and CO as well as for five age-matched controls are shown in Figure 12 Both
GR and CO could recognise the famous landmarksas well normals In addition both GR and CO wereusually able to name the landmarks they success-fully identified
Thus GR and CO can recognize landmarkslearned prior to injury demonstrating that at leastsome of their place recognition abilities are intactWhen combined with the fact that GR and COwere able to produce detailed maps of places theylived in prior to their injury (see Case Description)these results suggest that their deficit is primarily inencoding or consolidation of new informationrather than in recognition or recall of old informa-tion Note however that these results do not provethat their place recognition abilities are entirelynormal GR and CO might suffer from a mild rec-ognition deficit that affects identification of newly-learned stimuli like the Lego layouts in Experiment3 but not overlearned stimuli like the famous land-marks Furthermore it is possible that GR and COmight be impaired at recognising premorbidlylearned places that are defined primarily by theirspatial layout even though they are unimpaired atrecognising premorbidly learned places that aredefined primarily by a single object-like landmarkor building (see General Discussion) Neverthelessthe trend of the present evidence is to suggest thatGR and CO have particular problems with noveltopographical information
In the famous-face task GRrsquos performance wasin the normal range However he sometimes haddifficulty naming the faces even though he clearly
EPSTEIN ET AL
502 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 11 Stimuli for Experiment 4 Each famous landmark(Arc de Triomphe Taj Mahal) and famous face (Barbara BushYasser Arafat) was matched to a nonfamous building or face thatalso appeared in the experiment
FA
CE
SL
AN
DM
AR
KS
Figure 12 Results of Experiment 4 Performance measure isproportion of famous stimuli reported as ldquoknownrdquo corrected forguessing using the standard formula
recognised them For example he described Ron-ald Reagan as ldquothe gipperrdquo Nancy Reagan as ldquothewife of the gipperrdquo and Clark Gable as ldquothe movieactor who said lsquoScarlett I donrsquot give a damnrsquordquo COhad a great deal of difficulty identifying the famousfaces This result is not surprising given hisextremely low performance on the Benton FaceMatching Task In addition to his topographicproblems CO appears to suffer fromprosopagnosia
GENERAL DISCUSSION
We tested two parahippocampal patients (GR andCO) on four experiments designed to measure per-formance as a function of both task (perceptionencoding and recognition) and stimulus type(scenes objects faces and words) The only taskthat revealed a clear deficit was an n-back recogni-tion memory test (Experiment 3) In order to suc-cessfully perform this task subjects had to encode adurable representation of a visual stimulus and useit for later recognition GR and CO were signifi-cantly more impaired on this task when the stimuliwere scene-like spatial layouts (ie Lego scenesand real scenes for GR) than when they wereobjects No comparable deficits were found in otherexperiments that tested layout perception (Experi-ments 1 and 2) and placelandmark recognition(Experiment 4) Taken together these results indi-cate that GR and CO are impaired in their ability toencode novel information about the geometry ofsurrounding space into memory but are lessimpaired at their ability to encode the geometry ofnovel objects
We argue that this specific topographical defi-citmdashinability to encode the spatial layout of novelscenesmdashresults from damage to the para-hippocampal place area Although the damage inboth patients extends beyond the parahippo-campal-lingual territory normally associated withthe PPA this region is clearly lesioned bilaterally inGR and in the right hemisphere in CO Further-more functional imaging demonstrated that GRhas no functioning PPA Thus the current resultsand the earlier fMRI work (Epstein et al 1999
Epstein amp Kanwisher 1998) converge to implicateparahippocampal cortex in the encoding of infor-mation about scene geometry into memory
Our assignment of the function of memoryencoding to parahippocampal cortex is consistentwith the results of earlier case reports of patientswith parahippocampal damage who have naviga-tional difficulties in novel but not familiar environ-ments (Habib amp Sirigu 1987 Ross 1980 see alsoTeng amp Squire 1999) As with GR and CO thesepatients were either less impaired or unimpaired onnontopographical memory tasks Patients have alsobeen described who exhibit the opposite pattern ofimpairment an inability to learn faces (sometimescalled ldquoprosopamnesiardquo Cipolotti Robinson Blairamp Frith 1999 Tippett Miller amp Farah 2000) orfaces and verbal materials (Maguire amp Cipolotti1998) with an unimpaired ability to learn topo-graphical materials Taken together these resultsargue for a high degree of domain-specificity in theneural systems involved in learning of new informa-tion (Gallistel 1995) The brain may have separatemechanisms for learning faces places and wordswhich can be differentially impaired after damageto specific cortical regions (Cipolotti et al 1999Incisa della Rochetta et al 1996) The presentresults suggest that the anatomical locus of themechanism dedicated to learning new topographi-cal information is in the parahippocampal cortex(Aguirre amp DrsquoEsposito 1999)
In contrast to their impaired ability to encodenew topographical information both GR and COwere able to produce detailed maps of their previousdwellings and performed normally on the famouslandmark recognition test of Experiment 4 Theseresults are generally consistent with Teng andSquirersquos (1999) report of a patient with extensivemedial temporal damage who could recallpremorbidly learned but not postmorbidly learnedtopographical information (see also Ross 1980)Thus our results provide further evidence thatmedial temporal regions such as parahippocampalcortex may be more critical for encoding or consoli-dating new information into memory than for recallof old information
One issue that our experiments do not entirelyresolve is the precise nature of the topographical
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 503
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
information encoded by parahippocampal cortexClearly this region is critical for encoding informa-tion about the geometry of the currently visiblescene even when the ldquoscenerdquo is made out of Legoblocks and therefore has no immediate navigationalrelevance However it is unclear whether this spe-cific deficit can account for all of the real-worldnavigational difficulties encountered by GR andCO GR in particular shows such a catastrophicloss of navigational ability that we suspect he is alsoimpaired at encoding the spatial relationshipsbetweenthe current scene and other locations in theworld CO probably also suffers from such difficul-ties as he reports that he has trouble rememberinghow places relate to each other in large-scale spaceWhen he was tested on a simple 3-room videogameenvironment he not only was unable to rememberthe individual rooms (claiming that there were 10rooms when there were only 3) but he also had nomemory for the spatial relationships between themThese spatial deficits may be the result of damage tothe posterior right hippocampus (in the case of CO)or of disconnection of the posterior right hippo-campus from the parahippocampal regions thatprovide much of its cortical input (in both patientsSuzuki amp Amaral 1994) In either case the abilityto form a cognitive map of new environments likethe lab or videogame space could be impaired(Maguire et al 1998 OrsquoKeefe amp Nadel 1978)However it is also possible that parahippocampalcortex itself may be critical for encoding not onlythe geometry of individual scenes but also the spa-tial relationships between the locations of thosescenes In this case the spatial deficits of GR andCO could be attributed directly to parahippo-campal damage rather than to hippocampal dam-age or disconnection
The latter scenario would be consistent withAguirre and DrsquoEspositorsquos (1999) and Barrash etalrsquos (2000) suggestion that parahippocampal cortexis critically involved in combining visual informa-tion about the appearance of scenes and landmarkswith spatial information about their locations into aunified topographical representation Howeverboth of these authors argue that the representationsof individual scenes (Barrash et al) and landmarks(Aguirre amp DrsquoEsposito) are supported not by
parahippocampal cortex but by more posterior lin-gual-fusiform regions In contrast we believe thatthe neuroimaging data indicate that parahippo-campal cortex is involved in the encoding of thegeometric structure of individual scenes althoughit may also play a role in combining these represen-tations with other spatial representations
Interestingly GR and CO were not impaired onrecognition of famous landmarks that were mostlybuildings Thus our data do not preclude the possi-bility that there may be a separate lingual orfusiform region involved in processing informationabout buildings (Aguirre Zarahn amp DrsquoEsposito1998 Chao Haxby amp Martin 1999 IshaiUngerleider Martin Schouten amp Haxby 1999)which might have been preserved in GR and COdespite extensive damage to other regions of thelingual and fusiform gyrus Alternatively GR andCOrsquos preserved building recognition abilities mightbe mediated by interactions between cortical areasthat perform shape analysis for any object-likestimulus type (Malach et al 1995Kanwisher et al1996) and anterior temporal regions in which long-term topographical and semantic memories arestored (McCarthy et al 1996)
Is perception preserved
In contrast to their inability to encode new scenesinto memory GR and CO have substantiallyretained their ability to perceive the spatial layout ofthe currently visible scene (Experiments 1 and 2)GR can follow a route marked on a floor planwhich indicates a good on-line appreciation of hiscurrent spatial surroundings Furthermore neitherGR or CO have any trouble moving through theenvironmentmdashthey move with confidence and donot bump into doors or walls Problems of the lattertype are usually associated with damage to the pari-etal lobes which are intact in GR and CO Thus atfirst glance GR and CO do not appear to haveperceptual impairments (beyond their visual fieldrestrictions)
However GR does complain of a perceptualdeficit which he describes as an inability to ldquotakeinrdquo scenes when they are too complex Similarreports of an inability to comprehend the
EPSTEIN ET AL
504 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
ldquoglobalityrdquo of a scene were made by some of theparahippocampally-damaged patients studied byHabib and Sirigu (1987) We were not able todetermine the precise nature of this deficit whichmay be partly attributable to his visual field restric-tions It appears that his deficit is more salient in thereal-world environments than in the picture-per-ception tasks of Experiments 1 and 2 perhapsbecause he finds it more difficult to organise visualinput when immersed in it than when viewing pho-tographs that subtend only part of the visual field
We suggest that GR can perceive parts of thescene when he attends to them but may be unableto integrate information over time into a coherentrepresentation of a scene or place The fact that GRand CO are impaired for Lego layouts at 5-backintervals in Experiment 3 is consistent with thishypothesis The time between the first and secondpresentations of a stimulus is very short in this case(and even shorter with 3-back intervals where GRalso shows evidence of a deficit) In both patientsthe representation of layout information formed byperception of a Lego scene must either have anabnormally short persistence be abnormally sus-ceptible to interference or both If so they mightfind it impossible to sustain a spatial frame orschema long enough to use it to organise informa-tion obtained from many different glances at a com-plex scene (Intraub 1997) The result would be aninability to understand the ldquoglobalityrdquo of a sceneeven though all the details were perfectlyperceived
Non-topographical mnemonic abilities
Although not as severe as their topographical mem-ory problems GR and CO do have some memorydifficulties in other domains For example bothperformed poorly on the Rey AVLT whichrequired them to remember a list of unrelatedwords Both also report some subtle episodic mem-ory problems (see Case Description) COrsquos prob-lems could partly result from damage to the righthippocampusmdashindeed CO appeared to have themore acute episodic memory problems HoweverGR has no hippocampal damage so the fact that hedisplays episodic memory problems suggests thatparahippocampal cortex is involved in the forma-
tion of episodic memories (cf Wagner et al1998)
In previous work (Epstein et al 1999) weargued that spatial context information encoded byparahippocampal cortex might play a role in estab-lishing event memories by encoding informationabout where events take place The inability toencode this information might cause problems intasks such as the Rey AVLT where one has to recalla list of unrelated words If spatial (and perhapstemporal) context information is not encoded itwill be harder subsequently to recall the distinctevent of hearing each word (Tulving 1983) Incontrast lack of spatial context information wouldbe less of a problem in tasks where one is onlyrequired to recognise previously encountered verbalmaterials but not to reproduce the spatiotemporalcontext in which they were experienced IndeedGR and CO performed as well as normals on thenonsense words in Experiment 4 where recogni-tion memory but not episodic memory was requiredfor successful performance Interestingly GR andCO also did relatively well on the Logical Memorysubtest of the WMS-III perhaps because the nar-rative framework of a story provides sufficient con-textual information to overcome the absence ofspatial context and mediate successful verbalretrieval
Differences between GR and CO
Although we have discussed GR and CO togetherthere are a number of relevant differences betweenthem One open question is whether these differ-ences can be attributed to the different effects ofunilateral and bilateral parahippocampal damageIn general GRrsquos topographical difficulties weremore severe than COrsquos CO did surprisingly well onthe Warrington Topographical Memory Test inwhich he was required to remember detailed real-world city scenes and also performed substantiallybetter on real scenes than Lego scenes in Experi-ment 3 Insofar as CO has intact left para-hippocampal cortex this suggests that the left PPAmight be sufficient to encode a certain amount of(perhaps semantic) information about real-worldscenes but the right PPA is necessary to encode
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 505
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
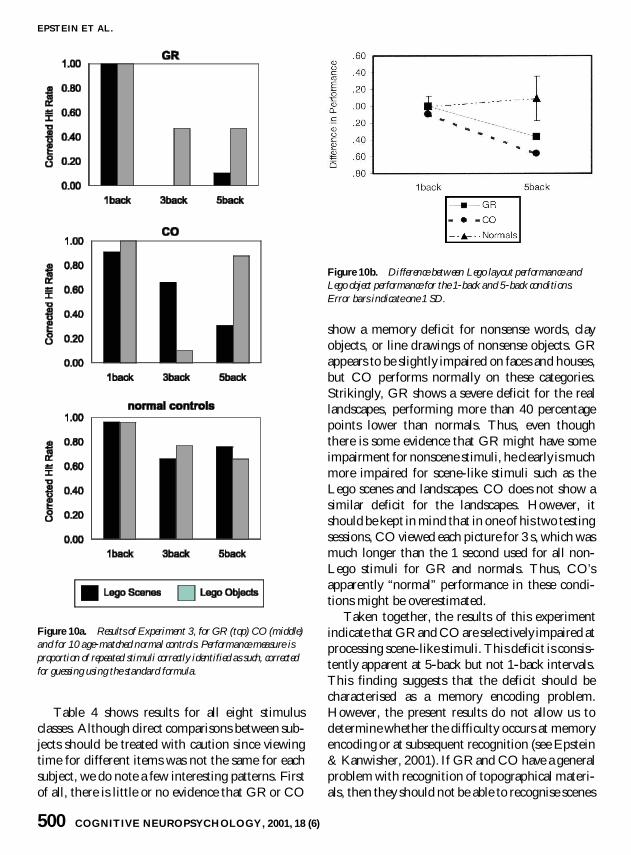
Table 4 shows results for all eight stimulusclasses Although direct comparisons between sub-jects should be treated with caution since viewingtime for different items was not the same for eachsubject we do note a few interesting patterns Firstof all there is little or no evidence that GR or CO
show a memory deficit for nonsense words clayobjects or line drawings of nonsense objects GRappears to be slightly impaired on faces and housesbut CO performs normally on these categoriesStrikingly GR shows a severe deficit for the reallandscapes performing more than 40 percentagepoints lower than normals Thus even thoughthere is some evidence that GR might have someimpairment for nonscene stimuli he clearly is muchmore impaired for scene-like stimuli such as theLego scenes and landscapes CO does not show asimilar deficit for the landscapes However itshould be kept in mind that in one of his two testingsessions CO viewed each picture for 3 s which wasmuch longer than the 1 second used for all non-Lego stimuli for GR and normals Thus COrsquosapparently ldquonormalrdquo performance in these condi-tions might be overestimated
Taken together the results of this experimentindicate that GR and CO are selectively impaired atprocessing scene-like stimuli This deficit is consis-tently apparent at 5-back but not 1-back intervalsThis finding suggests that the deficit should becharacterised as a memory encoding problemHowever the present results do not allow us todetermine whether the difficulty occurs at memoryencoding or at subsequent recognition (see Epsteinamp Kanwisher 2001) If GR and CO have a generalproblem with recognition of topographical materi-als then they should not be able to recognise scenes
EPSTEIN ET AL
500 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 10a Results of Experiment 3 for GR (top) CO (middle)and for 10 age-matched normal controls Performance measure isproportion of repeated stimuli correctly identified as such correctedfor guessing using the standard formula
Figure 10b Difference between Lego layout performance andLego object performance for the 1-back and 5-back conditionsError bars indicate one 1 SD
and landmarks that were learned prior to theirinjury We tested this possibility in the nextexperiment
Experiment 4 Famous landmarksfacesrecognition task
In this experiment we examined GR and COrsquosability to recognise the visual appearances of placeslearned prior to injury Subjects saw a series offamous landmarks intermixed with nonfamouscontrols (Imaging experiments have demonstratedthat landmarks of this type can activate the PPAalmost as strongly as complete scenes Epstein etal 1999 Epstein amp Kanwisher 1998) They wererequired to say whether they knew each specificlandmark and to identify the ones they knew Sub-jects were instructed to respond affirmatively if theywere fairly certain that they recognised the land-mark but not if it only looked generally familiarFor purposes of comparison subjects also per-formed an identical test in which the stimuli werefamous and nonfamous faces In both testsdistractor items were designed to be visually andconceptually similar to the target items (see Figure11) If parahippocampal cortex is required for placerecognition then one might expect GR and CO toperform poorly compared to normals on the land-mark recognition test
MethodEach of the two experiments (famous faces famouslandmarks) consisted of 48 trials Subjects beganeach trial by pressing the space bar after which ablack-and-white photograph of a face (in thefamous faces experiment) or a landmark (in thefamous landmarks experiment) appeared on thescreen Subjects were told to press the ldquojrdquo key if theyrecognised the specific face or landmark and the ldquof rdquokey if they did not taking as much time at theyneeded to make this discrimination but no extratime Subjects were told to report that they recog-nised a face or landmark if they felt that they knewit even if they could not retrieve its name If theydid know the name they were to say it after pressingthe button Otherwise they were to say whateverthey did remember about the item
Half of the photographs were of famous and halfof nonfamous people and landmarks The famousfaces included public figures (eg George BushRichard Nixon Prince Charles) and movie stars(eg Harrison Ford Clark Gable Marlon Brando)who were prominent prior to the time of GR andCOrsquos injuries the famous landmarks were build-ings and monuments from all over the world (egEiffel Tower US Capitol Building Great Sphinx)The nonfamous stimuli were chosen to match thefamous stimuli as closely as possible Each famousstimulus had a ldquomatchrdquo For example George Bushwas matched to a man of similar age in a coat andtie and the Taj Mahal was matched to a visually
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 501
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 4 Full results of Experiment 3 Mean proportion correct (corrected for guessing) SDs in parentheses
Lego Land- Lego Clay Nonsense Non-scenes scapes Houses objects objects objects Words Faces
1-back GR 100 094 100 100 100 094 100 100CO 091 085 091 100 100 079 074 100Normals 096(008) 095(008) 093(014) 096(008) 096(006) 099(003) 098(004) 097(011)
3-back GR ndash003 068 026 047 083 083 075 052CO 066 081 037 010 094 069 095 081Normals 066(032) 077(021) 072(015) 077(020) 086(016) 083(015) 083(017) 072(020)
5-back GR 010 034 044 047 072 083 094 053CO 031 073 075 088 081 064 100 081Normals 076(026) 077(016) 068(021) 066(016) 091(014) 083(018) 093(011) 078(021)
similar Asian palace that is not famous (see Figure11)
Results and discussionResults for GR and CO as well as for five age-matched controls are shown in Figure 12 Both
GR and CO could recognise the famous landmarksas well normals In addition both GR and CO wereusually able to name the landmarks they success-fully identified
Thus GR and CO can recognize landmarkslearned prior to injury demonstrating that at leastsome of their place recognition abilities are intactWhen combined with the fact that GR and COwere able to produce detailed maps of places theylived in prior to their injury (see Case Description)these results suggest that their deficit is primarily inencoding or consolidation of new informationrather than in recognition or recall of old informa-tion Note however that these results do not provethat their place recognition abilities are entirelynormal GR and CO might suffer from a mild rec-ognition deficit that affects identification of newly-learned stimuli like the Lego layouts in Experiment3 but not overlearned stimuli like the famous land-marks Furthermore it is possible that GR and COmight be impaired at recognising premorbidlylearned places that are defined primarily by theirspatial layout even though they are unimpaired atrecognising premorbidly learned places that aredefined primarily by a single object-like landmarkor building (see General Discussion) Neverthelessthe trend of the present evidence is to suggest thatGR and CO have particular problems with noveltopographical information
In the famous-face task GRrsquos performance wasin the normal range However he sometimes haddifficulty naming the faces even though he clearly
EPSTEIN ET AL
502 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 11 Stimuli for Experiment 4 Each famous landmark(Arc de Triomphe Taj Mahal) and famous face (Barbara BushYasser Arafat) was matched to a nonfamous building or face thatalso appeared in the experiment
FA
CE
SL
AN
DM
AR
KS
Figure 12 Results of Experiment 4 Performance measure isproportion of famous stimuli reported as ldquoknownrdquo corrected forguessing using the standard formula
recognised them For example he described Ron-ald Reagan as ldquothe gipperrdquo Nancy Reagan as ldquothewife of the gipperrdquo and Clark Gable as ldquothe movieactor who said lsquoScarlett I donrsquot give a damnrsquordquo COhad a great deal of difficulty identifying the famousfaces This result is not surprising given hisextremely low performance on the Benton FaceMatching Task In addition to his topographicproblems CO appears to suffer fromprosopagnosia
GENERAL DISCUSSION
We tested two parahippocampal patients (GR andCO) on four experiments designed to measure per-formance as a function of both task (perceptionencoding and recognition) and stimulus type(scenes objects faces and words) The only taskthat revealed a clear deficit was an n-back recogni-tion memory test (Experiment 3) In order to suc-cessfully perform this task subjects had to encode adurable representation of a visual stimulus and useit for later recognition GR and CO were signifi-cantly more impaired on this task when the stimuliwere scene-like spatial layouts (ie Lego scenesand real scenes for GR) than when they wereobjects No comparable deficits were found in otherexperiments that tested layout perception (Experi-ments 1 and 2) and placelandmark recognition(Experiment 4) Taken together these results indi-cate that GR and CO are impaired in their ability toencode novel information about the geometry ofsurrounding space into memory but are lessimpaired at their ability to encode the geometry ofnovel objects
We argue that this specific topographical defi-citmdashinability to encode the spatial layout of novelscenesmdashresults from damage to the para-hippocampal place area Although the damage inboth patients extends beyond the parahippo-campal-lingual territory normally associated withthe PPA this region is clearly lesioned bilaterally inGR and in the right hemisphere in CO Further-more functional imaging demonstrated that GRhas no functioning PPA Thus the current resultsand the earlier fMRI work (Epstein et al 1999
Epstein amp Kanwisher 1998) converge to implicateparahippocampal cortex in the encoding of infor-mation about scene geometry into memory
Our assignment of the function of memoryencoding to parahippocampal cortex is consistentwith the results of earlier case reports of patientswith parahippocampal damage who have naviga-tional difficulties in novel but not familiar environ-ments (Habib amp Sirigu 1987 Ross 1980 see alsoTeng amp Squire 1999) As with GR and CO thesepatients were either less impaired or unimpaired onnontopographical memory tasks Patients have alsobeen described who exhibit the opposite pattern ofimpairment an inability to learn faces (sometimescalled ldquoprosopamnesiardquo Cipolotti Robinson Blairamp Frith 1999 Tippett Miller amp Farah 2000) orfaces and verbal materials (Maguire amp Cipolotti1998) with an unimpaired ability to learn topo-graphical materials Taken together these resultsargue for a high degree of domain-specificity in theneural systems involved in learning of new informa-tion (Gallistel 1995) The brain may have separatemechanisms for learning faces places and wordswhich can be differentially impaired after damageto specific cortical regions (Cipolotti et al 1999Incisa della Rochetta et al 1996) The presentresults suggest that the anatomical locus of themechanism dedicated to learning new topographi-cal information is in the parahippocampal cortex(Aguirre amp DrsquoEsposito 1999)
In contrast to their impaired ability to encodenew topographical information both GR and COwere able to produce detailed maps of their previousdwellings and performed normally on the famouslandmark recognition test of Experiment 4 Theseresults are generally consistent with Teng andSquirersquos (1999) report of a patient with extensivemedial temporal damage who could recallpremorbidly learned but not postmorbidly learnedtopographical information (see also Ross 1980)Thus our results provide further evidence thatmedial temporal regions such as parahippocampalcortex may be more critical for encoding or consoli-dating new information into memory than for recallof old information
One issue that our experiments do not entirelyresolve is the precise nature of the topographical
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 503
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
information encoded by parahippocampal cortexClearly this region is critical for encoding informa-tion about the geometry of the currently visiblescene even when the ldquoscenerdquo is made out of Legoblocks and therefore has no immediate navigationalrelevance However it is unclear whether this spe-cific deficit can account for all of the real-worldnavigational difficulties encountered by GR andCO GR in particular shows such a catastrophicloss of navigational ability that we suspect he is alsoimpaired at encoding the spatial relationshipsbetweenthe current scene and other locations in theworld CO probably also suffers from such difficul-ties as he reports that he has trouble rememberinghow places relate to each other in large-scale spaceWhen he was tested on a simple 3-room videogameenvironment he not only was unable to rememberthe individual rooms (claiming that there were 10rooms when there were only 3) but he also had nomemory for the spatial relationships between themThese spatial deficits may be the result of damage tothe posterior right hippocampus (in the case of CO)or of disconnection of the posterior right hippo-campus from the parahippocampal regions thatprovide much of its cortical input (in both patientsSuzuki amp Amaral 1994) In either case the abilityto form a cognitive map of new environments likethe lab or videogame space could be impaired(Maguire et al 1998 OrsquoKeefe amp Nadel 1978)However it is also possible that parahippocampalcortex itself may be critical for encoding not onlythe geometry of individual scenes but also the spa-tial relationships between the locations of thosescenes In this case the spatial deficits of GR andCO could be attributed directly to parahippo-campal damage rather than to hippocampal dam-age or disconnection
The latter scenario would be consistent withAguirre and DrsquoEspositorsquos (1999) and Barrash etalrsquos (2000) suggestion that parahippocampal cortexis critically involved in combining visual informa-tion about the appearance of scenes and landmarkswith spatial information about their locations into aunified topographical representation Howeverboth of these authors argue that the representationsof individual scenes (Barrash et al) and landmarks(Aguirre amp DrsquoEsposito) are supported not by
parahippocampal cortex but by more posterior lin-gual-fusiform regions In contrast we believe thatthe neuroimaging data indicate that parahippo-campal cortex is involved in the encoding of thegeometric structure of individual scenes althoughit may also play a role in combining these represen-tations with other spatial representations
Interestingly GR and CO were not impaired onrecognition of famous landmarks that were mostlybuildings Thus our data do not preclude the possi-bility that there may be a separate lingual orfusiform region involved in processing informationabout buildings (Aguirre Zarahn amp DrsquoEsposito1998 Chao Haxby amp Martin 1999 IshaiUngerleider Martin Schouten amp Haxby 1999)which might have been preserved in GR and COdespite extensive damage to other regions of thelingual and fusiform gyrus Alternatively GR andCOrsquos preserved building recognition abilities mightbe mediated by interactions between cortical areasthat perform shape analysis for any object-likestimulus type (Malach et al 1995Kanwisher et al1996) and anterior temporal regions in which long-term topographical and semantic memories arestored (McCarthy et al 1996)
Is perception preserved
In contrast to their inability to encode new scenesinto memory GR and CO have substantiallyretained their ability to perceive the spatial layout ofthe currently visible scene (Experiments 1 and 2)GR can follow a route marked on a floor planwhich indicates a good on-line appreciation of hiscurrent spatial surroundings Furthermore neitherGR or CO have any trouble moving through theenvironmentmdashthey move with confidence and donot bump into doors or walls Problems of the lattertype are usually associated with damage to the pari-etal lobes which are intact in GR and CO Thus atfirst glance GR and CO do not appear to haveperceptual impairments (beyond their visual fieldrestrictions)
However GR does complain of a perceptualdeficit which he describes as an inability to ldquotakeinrdquo scenes when they are too complex Similarreports of an inability to comprehend the
EPSTEIN ET AL
504 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
ldquoglobalityrdquo of a scene were made by some of theparahippocampally-damaged patients studied byHabib and Sirigu (1987) We were not able todetermine the precise nature of this deficit whichmay be partly attributable to his visual field restric-tions It appears that his deficit is more salient in thereal-world environments than in the picture-per-ception tasks of Experiments 1 and 2 perhapsbecause he finds it more difficult to organise visualinput when immersed in it than when viewing pho-tographs that subtend only part of the visual field
We suggest that GR can perceive parts of thescene when he attends to them but may be unableto integrate information over time into a coherentrepresentation of a scene or place The fact that GRand CO are impaired for Lego layouts at 5-backintervals in Experiment 3 is consistent with thishypothesis The time between the first and secondpresentations of a stimulus is very short in this case(and even shorter with 3-back intervals where GRalso shows evidence of a deficit) In both patientsthe representation of layout information formed byperception of a Lego scene must either have anabnormally short persistence be abnormally sus-ceptible to interference or both If so they mightfind it impossible to sustain a spatial frame orschema long enough to use it to organise informa-tion obtained from many different glances at a com-plex scene (Intraub 1997) The result would be aninability to understand the ldquoglobalityrdquo of a sceneeven though all the details were perfectlyperceived
Non-topographical mnemonic abilities
Although not as severe as their topographical mem-ory problems GR and CO do have some memorydifficulties in other domains For example bothperformed poorly on the Rey AVLT whichrequired them to remember a list of unrelatedwords Both also report some subtle episodic mem-ory problems (see Case Description) COrsquos prob-lems could partly result from damage to the righthippocampusmdashindeed CO appeared to have themore acute episodic memory problems HoweverGR has no hippocampal damage so the fact that hedisplays episodic memory problems suggests thatparahippocampal cortex is involved in the forma-
tion of episodic memories (cf Wagner et al1998)
In previous work (Epstein et al 1999) weargued that spatial context information encoded byparahippocampal cortex might play a role in estab-lishing event memories by encoding informationabout where events take place The inability toencode this information might cause problems intasks such as the Rey AVLT where one has to recalla list of unrelated words If spatial (and perhapstemporal) context information is not encoded itwill be harder subsequently to recall the distinctevent of hearing each word (Tulving 1983) Incontrast lack of spatial context information wouldbe less of a problem in tasks where one is onlyrequired to recognise previously encountered verbalmaterials but not to reproduce the spatiotemporalcontext in which they were experienced IndeedGR and CO performed as well as normals on thenonsense words in Experiment 4 where recogni-tion memory but not episodic memory was requiredfor successful performance Interestingly GR andCO also did relatively well on the Logical Memorysubtest of the WMS-III perhaps because the nar-rative framework of a story provides sufficient con-textual information to overcome the absence ofspatial context and mediate successful verbalretrieval
Differences between GR and CO
Although we have discussed GR and CO togetherthere are a number of relevant differences betweenthem One open question is whether these differ-ences can be attributed to the different effects ofunilateral and bilateral parahippocampal damageIn general GRrsquos topographical difficulties weremore severe than COrsquos CO did surprisingly well onthe Warrington Topographical Memory Test inwhich he was required to remember detailed real-world city scenes and also performed substantiallybetter on real scenes than Lego scenes in Experi-ment 3 Insofar as CO has intact left para-hippocampal cortex this suggests that the left PPAmight be sufficient to encode a certain amount of(perhaps semantic) information about real-worldscenes but the right PPA is necessary to encode
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 505
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)

and landmarks that were learned prior to theirinjury We tested this possibility in the nextexperiment
Experiment 4 Famous landmarksfacesrecognition task
In this experiment we examined GR and COrsquosability to recognise the visual appearances of placeslearned prior to injury Subjects saw a series offamous landmarks intermixed with nonfamouscontrols (Imaging experiments have demonstratedthat landmarks of this type can activate the PPAalmost as strongly as complete scenes Epstein etal 1999 Epstein amp Kanwisher 1998) They wererequired to say whether they knew each specificlandmark and to identify the ones they knew Sub-jects were instructed to respond affirmatively if theywere fairly certain that they recognised the land-mark but not if it only looked generally familiarFor purposes of comparison subjects also per-formed an identical test in which the stimuli werefamous and nonfamous faces In both testsdistractor items were designed to be visually andconceptually similar to the target items (see Figure11) If parahippocampal cortex is required for placerecognition then one might expect GR and CO toperform poorly compared to normals on the land-mark recognition test
MethodEach of the two experiments (famous faces famouslandmarks) consisted of 48 trials Subjects beganeach trial by pressing the space bar after which ablack-and-white photograph of a face (in thefamous faces experiment) or a landmark (in thefamous landmarks experiment) appeared on thescreen Subjects were told to press the ldquojrdquo key if theyrecognised the specific face or landmark and the ldquof rdquokey if they did not taking as much time at theyneeded to make this discrimination but no extratime Subjects were told to report that they recog-nised a face or landmark if they felt that they knewit even if they could not retrieve its name If theydid know the name they were to say it after pressingthe button Otherwise they were to say whateverthey did remember about the item
Half of the photographs were of famous and halfof nonfamous people and landmarks The famousfaces included public figures (eg George BushRichard Nixon Prince Charles) and movie stars(eg Harrison Ford Clark Gable Marlon Brando)who were prominent prior to the time of GR andCOrsquos injuries the famous landmarks were build-ings and monuments from all over the world (egEiffel Tower US Capitol Building Great Sphinx)The nonfamous stimuli were chosen to match thefamous stimuli as closely as possible Each famousstimulus had a ldquomatchrdquo For example George Bushwas matched to a man of similar age in a coat andtie and the Taj Mahal was matched to a visually
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 501
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Table 4 Full results of Experiment 3 Mean proportion correct (corrected for guessing) SDs in parentheses
Lego Land- Lego Clay Nonsense Non-scenes scapes Houses objects objects objects Words Faces
1-back GR 100 094 100 100 100 094 100 100CO 091 085 091 100 100 079 074 100Normals 096(008) 095(008) 093(014) 096(008) 096(006) 099(003) 098(004) 097(011)
3-back GR ndash003 068 026 047 083 083 075 052CO 066 081 037 010 094 069 095 081Normals 066(032) 077(021) 072(015) 077(020) 086(016) 083(015) 083(017) 072(020)
5-back GR 010 034 044 047 072 083 094 053CO 031 073 075 088 081 064 100 081Normals 076(026) 077(016) 068(021) 066(016) 091(014) 083(018) 093(011) 078(021)
similar Asian palace that is not famous (see Figure11)
Results and discussionResults for GR and CO as well as for five age-matched controls are shown in Figure 12 Both
GR and CO could recognise the famous landmarksas well normals In addition both GR and CO wereusually able to name the landmarks they success-fully identified
Thus GR and CO can recognize landmarkslearned prior to injury demonstrating that at leastsome of their place recognition abilities are intactWhen combined with the fact that GR and COwere able to produce detailed maps of places theylived in prior to their injury (see Case Description)these results suggest that their deficit is primarily inencoding or consolidation of new informationrather than in recognition or recall of old informa-tion Note however that these results do not provethat their place recognition abilities are entirelynormal GR and CO might suffer from a mild rec-ognition deficit that affects identification of newly-learned stimuli like the Lego layouts in Experiment3 but not overlearned stimuli like the famous land-marks Furthermore it is possible that GR and COmight be impaired at recognising premorbidlylearned places that are defined primarily by theirspatial layout even though they are unimpaired atrecognising premorbidly learned places that aredefined primarily by a single object-like landmarkor building (see General Discussion) Neverthelessthe trend of the present evidence is to suggest thatGR and CO have particular problems with noveltopographical information
In the famous-face task GRrsquos performance wasin the normal range However he sometimes haddifficulty naming the faces even though he clearly
EPSTEIN ET AL
502 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 11 Stimuli for Experiment 4 Each famous landmark(Arc de Triomphe Taj Mahal) and famous face (Barbara BushYasser Arafat) was matched to a nonfamous building or face thatalso appeared in the experiment
FA
CE
SL
AN
DM
AR
KS
Figure 12 Results of Experiment 4 Performance measure isproportion of famous stimuli reported as ldquoknownrdquo corrected forguessing using the standard formula
recognised them For example he described Ron-ald Reagan as ldquothe gipperrdquo Nancy Reagan as ldquothewife of the gipperrdquo and Clark Gable as ldquothe movieactor who said lsquoScarlett I donrsquot give a damnrsquordquo COhad a great deal of difficulty identifying the famousfaces This result is not surprising given hisextremely low performance on the Benton FaceMatching Task In addition to his topographicproblems CO appears to suffer fromprosopagnosia
GENERAL DISCUSSION
We tested two parahippocampal patients (GR andCO) on four experiments designed to measure per-formance as a function of both task (perceptionencoding and recognition) and stimulus type(scenes objects faces and words) The only taskthat revealed a clear deficit was an n-back recogni-tion memory test (Experiment 3) In order to suc-cessfully perform this task subjects had to encode adurable representation of a visual stimulus and useit for later recognition GR and CO were signifi-cantly more impaired on this task when the stimuliwere scene-like spatial layouts (ie Lego scenesand real scenes for GR) than when they wereobjects No comparable deficits were found in otherexperiments that tested layout perception (Experi-ments 1 and 2) and placelandmark recognition(Experiment 4) Taken together these results indi-cate that GR and CO are impaired in their ability toencode novel information about the geometry ofsurrounding space into memory but are lessimpaired at their ability to encode the geometry ofnovel objects
We argue that this specific topographical defi-citmdashinability to encode the spatial layout of novelscenesmdashresults from damage to the para-hippocampal place area Although the damage inboth patients extends beyond the parahippo-campal-lingual territory normally associated withthe PPA this region is clearly lesioned bilaterally inGR and in the right hemisphere in CO Further-more functional imaging demonstrated that GRhas no functioning PPA Thus the current resultsand the earlier fMRI work (Epstein et al 1999
Epstein amp Kanwisher 1998) converge to implicateparahippocampal cortex in the encoding of infor-mation about scene geometry into memory
Our assignment of the function of memoryencoding to parahippocampal cortex is consistentwith the results of earlier case reports of patientswith parahippocampal damage who have naviga-tional difficulties in novel but not familiar environ-ments (Habib amp Sirigu 1987 Ross 1980 see alsoTeng amp Squire 1999) As with GR and CO thesepatients were either less impaired or unimpaired onnontopographical memory tasks Patients have alsobeen described who exhibit the opposite pattern ofimpairment an inability to learn faces (sometimescalled ldquoprosopamnesiardquo Cipolotti Robinson Blairamp Frith 1999 Tippett Miller amp Farah 2000) orfaces and verbal materials (Maguire amp Cipolotti1998) with an unimpaired ability to learn topo-graphical materials Taken together these resultsargue for a high degree of domain-specificity in theneural systems involved in learning of new informa-tion (Gallistel 1995) The brain may have separatemechanisms for learning faces places and wordswhich can be differentially impaired after damageto specific cortical regions (Cipolotti et al 1999Incisa della Rochetta et al 1996) The presentresults suggest that the anatomical locus of themechanism dedicated to learning new topographi-cal information is in the parahippocampal cortex(Aguirre amp DrsquoEsposito 1999)
In contrast to their impaired ability to encodenew topographical information both GR and COwere able to produce detailed maps of their previousdwellings and performed normally on the famouslandmark recognition test of Experiment 4 Theseresults are generally consistent with Teng andSquirersquos (1999) report of a patient with extensivemedial temporal damage who could recallpremorbidly learned but not postmorbidly learnedtopographical information (see also Ross 1980)Thus our results provide further evidence thatmedial temporal regions such as parahippocampalcortex may be more critical for encoding or consoli-dating new information into memory than for recallof old information
One issue that our experiments do not entirelyresolve is the precise nature of the topographical
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 503
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
information encoded by parahippocampal cortexClearly this region is critical for encoding informa-tion about the geometry of the currently visiblescene even when the ldquoscenerdquo is made out of Legoblocks and therefore has no immediate navigationalrelevance However it is unclear whether this spe-cific deficit can account for all of the real-worldnavigational difficulties encountered by GR andCO GR in particular shows such a catastrophicloss of navigational ability that we suspect he is alsoimpaired at encoding the spatial relationshipsbetweenthe current scene and other locations in theworld CO probably also suffers from such difficul-ties as he reports that he has trouble rememberinghow places relate to each other in large-scale spaceWhen he was tested on a simple 3-room videogameenvironment he not only was unable to rememberthe individual rooms (claiming that there were 10rooms when there were only 3) but he also had nomemory for the spatial relationships between themThese spatial deficits may be the result of damage tothe posterior right hippocampus (in the case of CO)or of disconnection of the posterior right hippo-campus from the parahippocampal regions thatprovide much of its cortical input (in both patientsSuzuki amp Amaral 1994) In either case the abilityto form a cognitive map of new environments likethe lab or videogame space could be impaired(Maguire et al 1998 OrsquoKeefe amp Nadel 1978)However it is also possible that parahippocampalcortex itself may be critical for encoding not onlythe geometry of individual scenes but also the spa-tial relationships between the locations of thosescenes In this case the spatial deficits of GR andCO could be attributed directly to parahippo-campal damage rather than to hippocampal dam-age or disconnection
The latter scenario would be consistent withAguirre and DrsquoEspositorsquos (1999) and Barrash etalrsquos (2000) suggestion that parahippocampal cortexis critically involved in combining visual informa-tion about the appearance of scenes and landmarkswith spatial information about their locations into aunified topographical representation Howeverboth of these authors argue that the representationsof individual scenes (Barrash et al) and landmarks(Aguirre amp DrsquoEsposito) are supported not by
parahippocampal cortex but by more posterior lin-gual-fusiform regions In contrast we believe thatthe neuroimaging data indicate that parahippo-campal cortex is involved in the encoding of thegeometric structure of individual scenes althoughit may also play a role in combining these represen-tations with other spatial representations
Interestingly GR and CO were not impaired onrecognition of famous landmarks that were mostlybuildings Thus our data do not preclude the possi-bility that there may be a separate lingual orfusiform region involved in processing informationabout buildings (Aguirre Zarahn amp DrsquoEsposito1998 Chao Haxby amp Martin 1999 IshaiUngerleider Martin Schouten amp Haxby 1999)which might have been preserved in GR and COdespite extensive damage to other regions of thelingual and fusiform gyrus Alternatively GR andCOrsquos preserved building recognition abilities mightbe mediated by interactions between cortical areasthat perform shape analysis for any object-likestimulus type (Malach et al 1995Kanwisher et al1996) and anterior temporal regions in which long-term topographical and semantic memories arestored (McCarthy et al 1996)
Is perception preserved
In contrast to their inability to encode new scenesinto memory GR and CO have substantiallyretained their ability to perceive the spatial layout ofthe currently visible scene (Experiments 1 and 2)GR can follow a route marked on a floor planwhich indicates a good on-line appreciation of hiscurrent spatial surroundings Furthermore neitherGR or CO have any trouble moving through theenvironmentmdashthey move with confidence and donot bump into doors or walls Problems of the lattertype are usually associated with damage to the pari-etal lobes which are intact in GR and CO Thus atfirst glance GR and CO do not appear to haveperceptual impairments (beyond their visual fieldrestrictions)
However GR does complain of a perceptualdeficit which he describes as an inability to ldquotakeinrdquo scenes when they are too complex Similarreports of an inability to comprehend the
EPSTEIN ET AL
504 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
ldquoglobalityrdquo of a scene were made by some of theparahippocampally-damaged patients studied byHabib and Sirigu (1987) We were not able todetermine the precise nature of this deficit whichmay be partly attributable to his visual field restric-tions It appears that his deficit is more salient in thereal-world environments than in the picture-per-ception tasks of Experiments 1 and 2 perhapsbecause he finds it more difficult to organise visualinput when immersed in it than when viewing pho-tographs that subtend only part of the visual field
We suggest that GR can perceive parts of thescene when he attends to them but may be unableto integrate information over time into a coherentrepresentation of a scene or place The fact that GRand CO are impaired for Lego layouts at 5-backintervals in Experiment 3 is consistent with thishypothesis The time between the first and secondpresentations of a stimulus is very short in this case(and even shorter with 3-back intervals where GRalso shows evidence of a deficit) In both patientsthe representation of layout information formed byperception of a Lego scene must either have anabnormally short persistence be abnormally sus-ceptible to interference or both If so they mightfind it impossible to sustain a spatial frame orschema long enough to use it to organise informa-tion obtained from many different glances at a com-plex scene (Intraub 1997) The result would be aninability to understand the ldquoglobalityrdquo of a sceneeven though all the details were perfectlyperceived
Non-topographical mnemonic abilities
Although not as severe as their topographical mem-ory problems GR and CO do have some memorydifficulties in other domains For example bothperformed poorly on the Rey AVLT whichrequired them to remember a list of unrelatedwords Both also report some subtle episodic mem-ory problems (see Case Description) COrsquos prob-lems could partly result from damage to the righthippocampusmdashindeed CO appeared to have themore acute episodic memory problems HoweverGR has no hippocampal damage so the fact that hedisplays episodic memory problems suggests thatparahippocampal cortex is involved in the forma-
tion of episodic memories (cf Wagner et al1998)
In previous work (Epstein et al 1999) weargued that spatial context information encoded byparahippocampal cortex might play a role in estab-lishing event memories by encoding informationabout where events take place The inability toencode this information might cause problems intasks such as the Rey AVLT where one has to recalla list of unrelated words If spatial (and perhapstemporal) context information is not encoded itwill be harder subsequently to recall the distinctevent of hearing each word (Tulving 1983) Incontrast lack of spatial context information wouldbe less of a problem in tasks where one is onlyrequired to recognise previously encountered verbalmaterials but not to reproduce the spatiotemporalcontext in which they were experienced IndeedGR and CO performed as well as normals on thenonsense words in Experiment 4 where recogni-tion memory but not episodic memory was requiredfor successful performance Interestingly GR andCO also did relatively well on the Logical Memorysubtest of the WMS-III perhaps because the nar-rative framework of a story provides sufficient con-textual information to overcome the absence ofspatial context and mediate successful verbalretrieval
Differences between GR and CO
Although we have discussed GR and CO togetherthere are a number of relevant differences betweenthem One open question is whether these differ-ences can be attributed to the different effects ofunilateral and bilateral parahippocampal damageIn general GRrsquos topographical difficulties weremore severe than COrsquos CO did surprisingly well onthe Warrington Topographical Memory Test inwhich he was required to remember detailed real-world city scenes and also performed substantiallybetter on real scenes than Lego scenes in Experi-ment 3 Insofar as CO has intact left para-hippocampal cortex this suggests that the left PPAmight be sufficient to encode a certain amount of(perhaps semantic) information about real-worldscenes but the right PPA is necessary to encode
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 505
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)

similar Asian palace that is not famous (see Figure11)
Results and discussionResults for GR and CO as well as for five age-matched controls are shown in Figure 12 Both
GR and CO could recognise the famous landmarksas well normals In addition both GR and CO wereusually able to name the landmarks they success-fully identified
Thus GR and CO can recognize landmarkslearned prior to injury demonstrating that at leastsome of their place recognition abilities are intactWhen combined with the fact that GR and COwere able to produce detailed maps of places theylived in prior to their injury (see Case Description)these results suggest that their deficit is primarily inencoding or consolidation of new informationrather than in recognition or recall of old informa-tion Note however that these results do not provethat their place recognition abilities are entirelynormal GR and CO might suffer from a mild rec-ognition deficit that affects identification of newly-learned stimuli like the Lego layouts in Experiment3 but not overlearned stimuli like the famous land-marks Furthermore it is possible that GR and COmight be impaired at recognising premorbidlylearned places that are defined primarily by theirspatial layout even though they are unimpaired atrecognising premorbidly learned places that aredefined primarily by a single object-like landmarkor building (see General Discussion) Neverthelessthe trend of the present evidence is to suggest thatGR and CO have particular problems with noveltopographical information
In the famous-face task GRrsquos performance wasin the normal range However he sometimes haddifficulty naming the faces even though he clearly
EPSTEIN ET AL
502 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Figure 11 Stimuli for Experiment 4 Each famous landmark(Arc de Triomphe Taj Mahal) and famous face (Barbara BushYasser Arafat) was matched to a nonfamous building or face thatalso appeared in the experiment
FA
CE
SL
AN
DM
AR
KS
Figure 12 Results of Experiment 4 Performance measure isproportion of famous stimuli reported as ldquoknownrdquo corrected forguessing using the standard formula
recognised them For example he described Ron-ald Reagan as ldquothe gipperrdquo Nancy Reagan as ldquothewife of the gipperrdquo and Clark Gable as ldquothe movieactor who said lsquoScarlett I donrsquot give a damnrsquordquo COhad a great deal of difficulty identifying the famousfaces This result is not surprising given hisextremely low performance on the Benton FaceMatching Task In addition to his topographicproblems CO appears to suffer fromprosopagnosia
GENERAL DISCUSSION
We tested two parahippocampal patients (GR andCO) on four experiments designed to measure per-formance as a function of both task (perceptionencoding and recognition) and stimulus type(scenes objects faces and words) The only taskthat revealed a clear deficit was an n-back recogni-tion memory test (Experiment 3) In order to suc-cessfully perform this task subjects had to encode adurable representation of a visual stimulus and useit for later recognition GR and CO were signifi-cantly more impaired on this task when the stimuliwere scene-like spatial layouts (ie Lego scenesand real scenes for GR) than when they wereobjects No comparable deficits were found in otherexperiments that tested layout perception (Experi-ments 1 and 2) and placelandmark recognition(Experiment 4) Taken together these results indi-cate that GR and CO are impaired in their ability toencode novel information about the geometry ofsurrounding space into memory but are lessimpaired at their ability to encode the geometry ofnovel objects
We argue that this specific topographical defi-citmdashinability to encode the spatial layout of novelscenesmdashresults from damage to the para-hippocampal place area Although the damage inboth patients extends beyond the parahippo-campal-lingual territory normally associated withthe PPA this region is clearly lesioned bilaterally inGR and in the right hemisphere in CO Further-more functional imaging demonstrated that GRhas no functioning PPA Thus the current resultsand the earlier fMRI work (Epstein et al 1999
Epstein amp Kanwisher 1998) converge to implicateparahippocampal cortex in the encoding of infor-mation about scene geometry into memory
Our assignment of the function of memoryencoding to parahippocampal cortex is consistentwith the results of earlier case reports of patientswith parahippocampal damage who have naviga-tional difficulties in novel but not familiar environ-ments (Habib amp Sirigu 1987 Ross 1980 see alsoTeng amp Squire 1999) As with GR and CO thesepatients were either less impaired or unimpaired onnontopographical memory tasks Patients have alsobeen described who exhibit the opposite pattern ofimpairment an inability to learn faces (sometimescalled ldquoprosopamnesiardquo Cipolotti Robinson Blairamp Frith 1999 Tippett Miller amp Farah 2000) orfaces and verbal materials (Maguire amp Cipolotti1998) with an unimpaired ability to learn topo-graphical materials Taken together these resultsargue for a high degree of domain-specificity in theneural systems involved in learning of new informa-tion (Gallistel 1995) The brain may have separatemechanisms for learning faces places and wordswhich can be differentially impaired after damageto specific cortical regions (Cipolotti et al 1999Incisa della Rochetta et al 1996) The presentresults suggest that the anatomical locus of themechanism dedicated to learning new topographi-cal information is in the parahippocampal cortex(Aguirre amp DrsquoEsposito 1999)
In contrast to their impaired ability to encodenew topographical information both GR and COwere able to produce detailed maps of their previousdwellings and performed normally on the famouslandmark recognition test of Experiment 4 Theseresults are generally consistent with Teng andSquirersquos (1999) report of a patient with extensivemedial temporal damage who could recallpremorbidly learned but not postmorbidly learnedtopographical information (see also Ross 1980)Thus our results provide further evidence thatmedial temporal regions such as parahippocampalcortex may be more critical for encoding or consoli-dating new information into memory than for recallof old information
One issue that our experiments do not entirelyresolve is the precise nature of the topographical
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 503
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
information encoded by parahippocampal cortexClearly this region is critical for encoding informa-tion about the geometry of the currently visiblescene even when the ldquoscenerdquo is made out of Legoblocks and therefore has no immediate navigationalrelevance However it is unclear whether this spe-cific deficit can account for all of the real-worldnavigational difficulties encountered by GR andCO GR in particular shows such a catastrophicloss of navigational ability that we suspect he is alsoimpaired at encoding the spatial relationshipsbetweenthe current scene and other locations in theworld CO probably also suffers from such difficul-ties as he reports that he has trouble rememberinghow places relate to each other in large-scale spaceWhen he was tested on a simple 3-room videogameenvironment he not only was unable to rememberthe individual rooms (claiming that there were 10rooms when there were only 3) but he also had nomemory for the spatial relationships between themThese spatial deficits may be the result of damage tothe posterior right hippocampus (in the case of CO)or of disconnection of the posterior right hippo-campus from the parahippocampal regions thatprovide much of its cortical input (in both patientsSuzuki amp Amaral 1994) In either case the abilityto form a cognitive map of new environments likethe lab or videogame space could be impaired(Maguire et al 1998 OrsquoKeefe amp Nadel 1978)However it is also possible that parahippocampalcortex itself may be critical for encoding not onlythe geometry of individual scenes but also the spa-tial relationships between the locations of thosescenes In this case the spatial deficits of GR andCO could be attributed directly to parahippo-campal damage rather than to hippocampal dam-age or disconnection
The latter scenario would be consistent withAguirre and DrsquoEspositorsquos (1999) and Barrash etalrsquos (2000) suggestion that parahippocampal cortexis critically involved in combining visual informa-tion about the appearance of scenes and landmarkswith spatial information about their locations into aunified topographical representation Howeverboth of these authors argue that the representationsof individual scenes (Barrash et al) and landmarks(Aguirre amp DrsquoEsposito) are supported not by
parahippocampal cortex but by more posterior lin-gual-fusiform regions In contrast we believe thatthe neuroimaging data indicate that parahippo-campal cortex is involved in the encoding of thegeometric structure of individual scenes althoughit may also play a role in combining these represen-tations with other spatial representations
Interestingly GR and CO were not impaired onrecognition of famous landmarks that were mostlybuildings Thus our data do not preclude the possi-bility that there may be a separate lingual orfusiform region involved in processing informationabout buildings (Aguirre Zarahn amp DrsquoEsposito1998 Chao Haxby amp Martin 1999 IshaiUngerleider Martin Schouten amp Haxby 1999)which might have been preserved in GR and COdespite extensive damage to other regions of thelingual and fusiform gyrus Alternatively GR andCOrsquos preserved building recognition abilities mightbe mediated by interactions between cortical areasthat perform shape analysis for any object-likestimulus type (Malach et al 1995Kanwisher et al1996) and anterior temporal regions in which long-term topographical and semantic memories arestored (McCarthy et al 1996)
Is perception preserved
In contrast to their inability to encode new scenesinto memory GR and CO have substantiallyretained their ability to perceive the spatial layout ofthe currently visible scene (Experiments 1 and 2)GR can follow a route marked on a floor planwhich indicates a good on-line appreciation of hiscurrent spatial surroundings Furthermore neitherGR or CO have any trouble moving through theenvironmentmdashthey move with confidence and donot bump into doors or walls Problems of the lattertype are usually associated with damage to the pari-etal lobes which are intact in GR and CO Thus atfirst glance GR and CO do not appear to haveperceptual impairments (beyond their visual fieldrestrictions)
However GR does complain of a perceptualdeficit which he describes as an inability to ldquotakeinrdquo scenes when they are too complex Similarreports of an inability to comprehend the
EPSTEIN ET AL
504 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
ldquoglobalityrdquo of a scene were made by some of theparahippocampally-damaged patients studied byHabib and Sirigu (1987) We were not able todetermine the precise nature of this deficit whichmay be partly attributable to his visual field restric-tions It appears that his deficit is more salient in thereal-world environments than in the picture-per-ception tasks of Experiments 1 and 2 perhapsbecause he finds it more difficult to organise visualinput when immersed in it than when viewing pho-tographs that subtend only part of the visual field
We suggest that GR can perceive parts of thescene when he attends to them but may be unableto integrate information over time into a coherentrepresentation of a scene or place The fact that GRand CO are impaired for Lego layouts at 5-backintervals in Experiment 3 is consistent with thishypothesis The time between the first and secondpresentations of a stimulus is very short in this case(and even shorter with 3-back intervals where GRalso shows evidence of a deficit) In both patientsthe representation of layout information formed byperception of a Lego scene must either have anabnormally short persistence be abnormally sus-ceptible to interference or both If so they mightfind it impossible to sustain a spatial frame orschema long enough to use it to organise informa-tion obtained from many different glances at a com-plex scene (Intraub 1997) The result would be aninability to understand the ldquoglobalityrdquo of a sceneeven though all the details were perfectlyperceived
Non-topographical mnemonic abilities
Although not as severe as their topographical mem-ory problems GR and CO do have some memorydifficulties in other domains For example bothperformed poorly on the Rey AVLT whichrequired them to remember a list of unrelatedwords Both also report some subtle episodic mem-ory problems (see Case Description) COrsquos prob-lems could partly result from damage to the righthippocampusmdashindeed CO appeared to have themore acute episodic memory problems HoweverGR has no hippocampal damage so the fact that hedisplays episodic memory problems suggests thatparahippocampal cortex is involved in the forma-
tion of episodic memories (cf Wagner et al1998)
In previous work (Epstein et al 1999) weargued that spatial context information encoded byparahippocampal cortex might play a role in estab-lishing event memories by encoding informationabout where events take place The inability toencode this information might cause problems intasks such as the Rey AVLT where one has to recalla list of unrelated words If spatial (and perhapstemporal) context information is not encoded itwill be harder subsequently to recall the distinctevent of hearing each word (Tulving 1983) Incontrast lack of spatial context information wouldbe less of a problem in tasks where one is onlyrequired to recognise previously encountered verbalmaterials but not to reproduce the spatiotemporalcontext in which they were experienced IndeedGR and CO performed as well as normals on thenonsense words in Experiment 4 where recogni-tion memory but not episodic memory was requiredfor successful performance Interestingly GR andCO also did relatively well on the Logical Memorysubtest of the WMS-III perhaps because the nar-rative framework of a story provides sufficient con-textual information to overcome the absence ofspatial context and mediate successful verbalretrieval
Differences between GR and CO
Although we have discussed GR and CO togetherthere are a number of relevant differences betweenthem One open question is whether these differ-ences can be attributed to the different effects ofunilateral and bilateral parahippocampal damageIn general GRrsquos topographical difficulties weremore severe than COrsquos CO did surprisingly well onthe Warrington Topographical Memory Test inwhich he was required to remember detailed real-world city scenes and also performed substantiallybetter on real scenes than Lego scenes in Experi-ment 3 Insofar as CO has intact left para-hippocampal cortex this suggests that the left PPAmight be sufficient to encode a certain amount of(perhaps semantic) information about real-worldscenes but the right PPA is necessary to encode
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 505
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)

recognised them For example he described Ron-ald Reagan as ldquothe gipperrdquo Nancy Reagan as ldquothewife of the gipperrdquo and Clark Gable as ldquothe movieactor who said lsquoScarlett I donrsquot give a damnrsquordquo COhad a great deal of difficulty identifying the famousfaces This result is not surprising given hisextremely low performance on the Benton FaceMatching Task In addition to his topographicproblems CO appears to suffer fromprosopagnosia
GENERAL DISCUSSION
We tested two parahippocampal patients (GR andCO) on four experiments designed to measure per-formance as a function of both task (perceptionencoding and recognition) and stimulus type(scenes objects faces and words) The only taskthat revealed a clear deficit was an n-back recogni-tion memory test (Experiment 3) In order to suc-cessfully perform this task subjects had to encode adurable representation of a visual stimulus and useit for later recognition GR and CO were signifi-cantly more impaired on this task when the stimuliwere scene-like spatial layouts (ie Lego scenesand real scenes for GR) than when they wereobjects No comparable deficits were found in otherexperiments that tested layout perception (Experi-ments 1 and 2) and placelandmark recognition(Experiment 4) Taken together these results indi-cate that GR and CO are impaired in their ability toencode novel information about the geometry ofsurrounding space into memory but are lessimpaired at their ability to encode the geometry ofnovel objects
We argue that this specific topographical defi-citmdashinability to encode the spatial layout of novelscenesmdashresults from damage to the para-hippocampal place area Although the damage inboth patients extends beyond the parahippo-campal-lingual territory normally associated withthe PPA this region is clearly lesioned bilaterally inGR and in the right hemisphere in CO Further-more functional imaging demonstrated that GRhas no functioning PPA Thus the current resultsand the earlier fMRI work (Epstein et al 1999
Epstein amp Kanwisher 1998) converge to implicateparahippocampal cortex in the encoding of infor-mation about scene geometry into memory
Our assignment of the function of memoryencoding to parahippocampal cortex is consistentwith the results of earlier case reports of patientswith parahippocampal damage who have naviga-tional difficulties in novel but not familiar environ-ments (Habib amp Sirigu 1987 Ross 1980 see alsoTeng amp Squire 1999) As with GR and CO thesepatients were either less impaired or unimpaired onnontopographical memory tasks Patients have alsobeen described who exhibit the opposite pattern ofimpairment an inability to learn faces (sometimescalled ldquoprosopamnesiardquo Cipolotti Robinson Blairamp Frith 1999 Tippett Miller amp Farah 2000) orfaces and verbal materials (Maguire amp Cipolotti1998) with an unimpaired ability to learn topo-graphical materials Taken together these resultsargue for a high degree of domain-specificity in theneural systems involved in learning of new informa-tion (Gallistel 1995) The brain may have separatemechanisms for learning faces places and wordswhich can be differentially impaired after damageto specific cortical regions (Cipolotti et al 1999Incisa della Rochetta et al 1996) The presentresults suggest that the anatomical locus of themechanism dedicated to learning new topographi-cal information is in the parahippocampal cortex(Aguirre amp DrsquoEsposito 1999)
In contrast to their impaired ability to encodenew topographical information both GR and COwere able to produce detailed maps of their previousdwellings and performed normally on the famouslandmark recognition test of Experiment 4 Theseresults are generally consistent with Teng andSquirersquos (1999) report of a patient with extensivemedial temporal damage who could recallpremorbidly learned but not postmorbidly learnedtopographical information (see also Ross 1980)Thus our results provide further evidence thatmedial temporal regions such as parahippocampalcortex may be more critical for encoding or consoli-dating new information into memory than for recallof old information
One issue that our experiments do not entirelyresolve is the precise nature of the topographical
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 503
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
information encoded by parahippocampal cortexClearly this region is critical for encoding informa-tion about the geometry of the currently visiblescene even when the ldquoscenerdquo is made out of Legoblocks and therefore has no immediate navigationalrelevance However it is unclear whether this spe-cific deficit can account for all of the real-worldnavigational difficulties encountered by GR andCO GR in particular shows such a catastrophicloss of navigational ability that we suspect he is alsoimpaired at encoding the spatial relationshipsbetweenthe current scene and other locations in theworld CO probably also suffers from such difficul-ties as he reports that he has trouble rememberinghow places relate to each other in large-scale spaceWhen he was tested on a simple 3-room videogameenvironment he not only was unable to rememberthe individual rooms (claiming that there were 10rooms when there were only 3) but he also had nomemory for the spatial relationships between themThese spatial deficits may be the result of damage tothe posterior right hippocampus (in the case of CO)or of disconnection of the posterior right hippo-campus from the parahippocampal regions thatprovide much of its cortical input (in both patientsSuzuki amp Amaral 1994) In either case the abilityto form a cognitive map of new environments likethe lab or videogame space could be impaired(Maguire et al 1998 OrsquoKeefe amp Nadel 1978)However it is also possible that parahippocampalcortex itself may be critical for encoding not onlythe geometry of individual scenes but also the spa-tial relationships between the locations of thosescenes In this case the spatial deficits of GR andCO could be attributed directly to parahippo-campal damage rather than to hippocampal dam-age or disconnection
The latter scenario would be consistent withAguirre and DrsquoEspositorsquos (1999) and Barrash etalrsquos (2000) suggestion that parahippocampal cortexis critically involved in combining visual informa-tion about the appearance of scenes and landmarkswith spatial information about their locations into aunified topographical representation Howeverboth of these authors argue that the representationsof individual scenes (Barrash et al) and landmarks(Aguirre amp DrsquoEsposito) are supported not by
parahippocampal cortex but by more posterior lin-gual-fusiform regions In contrast we believe thatthe neuroimaging data indicate that parahippo-campal cortex is involved in the encoding of thegeometric structure of individual scenes althoughit may also play a role in combining these represen-tations with other spatial representations
Interestingly GR and CO were not impaired onrecognition of famous landmarks that were mostlybuildings Thus our data do not preclude the possi-bility that there may be a separate lingual orfusiform region involved in processing informationabout buildings (Aguirre Zarahn amp DrsquoEsposito1998 Chao Haxby amp Martin 1999 IshaiUngerleider Martin Schouten amp Haxby 1999)which might have been preserved in GR and COdespite extensive damage to other regions of thelingual and fusiform gyrus Alternatively GR andCOrsquos preserved building recognition abilities mightbe mediated by interactions between cortical areasthat perform shape analysis for any object-likestimulus type (Malach et al 1995Kanwisher et al1996) and anterior temporal regions in which long-term topographical and semantic memories arestored (McCarthy et al 1996)
Is perception preserved
In contrast to their inability to encode new scenesinto memory GR and CO have substantiallyretained their ability to perceive the spatial layout ofthe currently visible scene (Experiments 1 and 2)GR can follow a route marked on a floor planwhich indicates a good on-line appreciation of hiscurrent spatial surroundings Furthermore neitherGR or CO have any trouble moving through theenvironmentmdashthey move with confidence and donot bump into doors or walls Problems of the lattertype are usually associated with damage to the pari-etal lobes which are intact in GR and CO Thus atfirst glance GR and CO do not appear to haveperceptual impairments (beyond their visual fieldrestrictions)
However GR does complain of a perceptualdeficit which he describes as an inability to ldquotakeinrdquo scenes when they are too complex Similarreports of an inability to comprehend the
EPSTEIN ET AL
504 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
ldquoglobalityrdquo of a scene were made by some of theparahippocampally-damaged patients studied byHabib and Sirigu (1987) We were not able todetermine the precise nature of this deficit whichmay be partly attributable to his visual field restric-tions It appears that his deficit is more salient in thereal-world environments than in the picture-per-ception tasks of Experiments 1 and 2 perhapsbecause he finds it more difficult to organise visualinput when immersed in it than when viewing pho-tographs that subtend only part of the visual field
We suggest that GR can perceive parts of thescene when he attends to them but may be unableto integrate information over time into a coherentrepresentation of a scene or place The fact that GRand CO are impaired for Lego layouts at 5-backintervals in Experiment 3 is consistent with thishypothesis The time between the first and secondpresentations of a stimulus is very short in this case(and even shorter with 3-back intervals where GRalso shows evidence of a deficit) In both patientsthe representation of layout information formed byperception of a Lego scene must either have anabnormally short persistence be abnormally sus-ceptible to interference or both If so they mightfind it impossible to sustain a spatial frame orschema long enough to use it to organise informa-tion obtained from many different glances at a com-plex scene (Intraub 1997) The result would be aninability to understand the ldquoglobalityrdquo of a sceneeven though all the details were perfectlyperceived
Non-topographical mnemonic abilities
Although not as severe as their topographical mem-ory problems GR and CO do have some memorydifficulties in other domains For example bothperformed poorly on the Rey AVLT whichrequired them to remember a list of unrelatedwords Both also report some subtle episodic mem-ory problems (see Case Description) COrsquos prob-lems could partly result from damage to the righthippocampusmdashindeed CO appeared to have themore acute episodic memory problems HoweverGR has no hippocampal damage so the fact that hedisplays episodic memory problems suggests thatparahippocampal cortex is involved in the forma-
tion of episodic memories (cf Wagner et al1998)
In previous work (Epstein et al 1999) weargued that spatial context information encoded byparahippocampal cortex might play a role in estab-lishing event memories by encoding informationabout where events take place The inability toencode this information might cause problems intasks such as the Rey AVLT where one has to recalla list of unrelated words If spatial (and perhapstemporal) context information is not encoded itwill be harder subsequently to recall the distinctevent of hearing each word (Tulving 1983) Incontrast lack of spatial context information wouldbe less of a problem in tasks where one is onlyrequired to recognise previously encountered verbalmaterials but not to reproduce the spatiotemporalcontext in which they were experienced IndeedGR and CO performed as well as normals on thenonsense words in Experiment 4 where recogni-tion memory but not episodic memory was requiredfor successful performance Interestingly GR andCO also did relatively well on the Logical Memorysubtest of the WMS-III perhaps because the nar-rative framework of a story provides sufficient con-textual information to overcome the absence ofspatial context and mediate successful verbalretrieval
Differences between GR and CO
Although we have discussed GR and CO togetherthere are a number of relevant differences betweenthem One open question is whether these differ-ences can be attributed to the different effects ofunilateral and bilateral parahippocampal damageIn general GRrsquos topographical difficulties weremore severe than COrsquos CO did surprisingly well onthe Warrington Topographical Memory Test inwhich he was required to remember detailed real-world city scenes and also performed substantiallybetter on real scenes than Lego scenes in Experi-ment 3 Insofar as CO has intact left para-hippocampal cortex this suggests that the left PPAmight be sufficient to encode a certain amount of(perhaps semantic) information about real-worldscenes but the right PPA is necessary to encode
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 505
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)

information encoded by parahippocampal cortexClearly this region is critical for encoding informa-tion about the geometry of the currently visiblescene even when the ldquoscenerdquo is made out of Legoblocks and therefore has no immediate navigationalrelevance However it is unclear whether this spe-cific deficit can account for all of the real-worldnavigational difficulties encountered by GR andCO GR in particular shows such a catastrophicloss of navigational ability that we suspect he is alsoimpaired at encoding the spatial relationshipsbetweenthe current scene and other locations in theworld CO probably also suffers from such difficul-ties as he reports that he has trouble rememberinghow places relate to each other in large-scale spaceWhen he was tested on a simple 3-room videogameenvironment he not only was unable to rememberthe individual rooms (claiming that there were 10rooms when there were only 3) but he also had nomemory for the spatial relationships between themThese spatial deficits may be the result of damage tothe posterior right hippocampus (in the case of CO)or of disconnection of the posterior right hippo-campus from the parahippocampal regions thatprovide much of its cortical input (in both patientsSuzuki amp Amaral 1994) In either case the abilityto form a cognitive map of new environments likethe lab or videogame space could be impaired(Maguire et al 1998 OrsquoKeefe amp Nadel 1978)However it is also possible that parahippocampalcortex itself may be critical for encoding not onlythe geometry of individual scenes but also the spa-tial relationships between the locations of thosescenes In this case the spatial deficits of GR andCO could be attributed directly to parahippo-campal damage rather than to hippocampal dam-age or disconnection
The latter scenario would be consistent withAguirre and DrsquoEspositorsquos (1999) and Barrash etalrsquos (2000) suggestion that parahippocampal cortexis critically involved in combining visual informa-tion about the appearance of scenes and landmarkswith spatial information about their locations into aunified topographical representation Howeverboth of these authors argue that the representationsof individual scenes (Barrash et al) and landmarks(Aguirre amp DrsquoEsposito) are supported not by
parahippocampal cortex but by more posterior lin-gual-fusiform regions In contrast we believe thatthe neuroimaging data indicate that parahippo-campal cortex is involved in the encoding of thegeometric structure of individual scenes althoughit may also play a role in combining these represen-tations with other spatial representations
Interestingly GR and CO were not impaired onrecognition of famous landmarks that were mostlybuildings Thus our data do not preclude the possi-bility that there may be a separate lingual orfusiform region involved in processing informationabout buildings (Aguirre Zarahn amp DrsquoEsposito1998 Chao Haxby amp Martin 1999 IshaiUngerleider Martin Schouten amp Haxby 1999)which might have been preserved in GR and COdespite extensive damage to other regions of thelingual and fusiform gyrus Alternatively GR andCOrsquos preserved building recognition abilities mightbe mediated by interactions between cortical areasthat perform shape analysis for any object-likestimulus type (Malach et al 1995Kanwisher et al1996) and anterior temporal regions in which long-term topographical and semantic memories arestored (McCarthy et al 1996)
Is perception preserved
In contrast to their inability to encode new scenesinto memory GR and CO have substantiallyretained their ability to perceive the spatial layout ofthe currently visible scene (Experiments 1 and 2)GR can follow a route marked on a floor planwhich indicates a good on-line appreciation of hiscurrent spatial surroundings Furthermore neitherGR or CO have any trouble moving through theenvironmentmdashthey move with confidence and donot bump into doors or walls Problems of the lattertype are usually associated with damage to the pari-etal lobes which are intact in GR and CO Thus atfirst glance GR and CO do not appear to haveperceptual impairments (beyond their visual fieldrestrictions)
However GR does complain of a perceptualdeficit which he describes as an inability to ldquotakeinrdquo scenes when they are too complex Similarreports of an inability to comprehend the
EPSTEIN ET AL
504 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
ldquoglobalityrdquo of a scene were made by some of theparahippocampally-damaged patients studied byHabib and Sirigu (1987) We were not able todetermine the precise nature of this deficit whichmay be partly attributable to his visual field restric-tions It appears that his deficit is more salient in thereal-world environments than in the picture-per-ception tasks of Experiments 1 and 2 perhapsbecause he finds it more difficult to organise visualinput when immersed in it than when viewing pho-tographs that subtend only part of the visual field
We suggest that GR can perceive parts of thescene when he attends to them but may be unableto integrate information over time into a coherentrepresentation of a scene or place The fact that GRand CO are impaired for Lego layouts at 5-backintervals in Experiment 3 is consistent with thishypothesis The time between the first and secondpresentations of a stimulus is very short in this case(and even shorter with 3-back intervals where GRalso shows evidence of a deficit) In both patientsthe representation of layout information formed byperception of a Lego scene must either have anabnormally short persistence be abnormally sus-ceptible to interference or both If so they mightfind it impossible to sustain a spatial frame orschema long enough to use it to organise informa-tion obtained from many different glances at a com-plex scene (Intraub 1997) The result would be aninability to understand the ldquoglobalityrdquo of a sceneeven though all the details were perfectlyperceived
Non-topographical mnemonic abilities
Although not as severe as their topographical mem-ory problems GR and CO do have some memorydifficulties in other domains For example bothperformed poorly on the Rey AVLT whichrequired them to remember a list of unrelatedwords Both also report some subtle episodic mem-ory problems (see Case Description) COrsquos prob-lems could partly result from damage to the righthippocampusmdashindeed CO appeared to have themore acute episodic memory problems HoweverGR has no hippocampal damage so the fact that hedisplays episodic memory problems suggests thatparahippocampal cortex is involved in the forma-
tion of episodic memories (cf Wagner et al1998)
In previous work (Epstein et al 1999) weargued that spatial context information encoded byparahippocampal cortex might play a role in estab-lishing event memories by encoding informationabout where events take place The inability toencode this information might cause problems intasks such as the Rey AVLT where one has to recalla list of unrelated words If spatial (and perhapstemporal) context information is not encoded itwill be harder subsequently to recall the distinctevent of hearing each word (Tulving 1983) Incontrast lack of spatial context information wouldbe less of a problem in tasks where one is onlyrequired to recognise previously encountered verbalmaterials but not to reproduce the spatiotemporalcontext in which they were experienced IndeedGR and CO performed as well as normals on thenonsense words in Experiment 4 where recogni-tion memory but not episodic memory was requiredfor successful performance Interestingly GR andCO also did relatively well on the Logical Memorysubtest of the WMS-III perhaps because the nar-rative framework of a story provides sufficient con-textual information to overcome the absence ofspatial context and mediate successful verbalretrieval
Differences between GR and CO
Although we have discussed GR and CO togetherthere are a number of relevant differences betweenthem One open question is whether these differ-ences can be attributed to the different effects ofunilateral and bilateral parahippocampal damageIn general GRrsquos topographical difficulties weremore severe than COrsquos CO did surprisingly well onthe Warrington Topographical Memory Test inwhich he was required to remember detailed real-world city scenes and also performed substantiallybetter on real scenes than Lego scenes in Experi-ment 3 Insofar as CO has intact left para-hippocampal cortex this suggests that the left PPAmight be sufficient to encode a certain amount of(perhaps semantic) information about real-worldscenes but the right PPA is necessary to encode
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 505
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)

ldquoglobalityrdquo of a scene were made by some of theparahippocampally-damaged patients studied byHabib and Sirigu (1987) We were not able todetermine the precise nature of this deficit whichmay be partly attributable to his visual field restric-tions It appears that his deficit is more salient in thereal-world environments than in the picture-per-ception tasks of Experiments 1 and 2 perhapsbecause he finds it more difficult to organise visualinput when immersed in it than when viewing pho-tographs that subtend only part of the visual field
We suggest that GR can perceive parts of thescene when he attends to them but may be unableto integrate information over time into a coherentrepresentation of a scene or place The fact that GRand CO are impaired for Lego layouts at 5-backintervals in Experiment 3 is consistent with thishypothesis The time between the first and secondpresentations of a stimulus is very short in this case(and even shorter with 3-back intervals where GRalso shows evidence of a deficit) In both patientsthe representation of layout information formed byperception of a Lego scene must either have anabnormally short persistence be abnormally sus-ceptible to interference or both If so they mightfind it impossible to sustain a spatial frame orschema long enough to use it to organise informa-tion obtained from many different glances at a com-plex scene (Intraub 1997) The result would be aninability to understand the ldquoglobalityrdquo of a sceneeven though all the details were perfectlyperceived
Non-topographical mnemonic abilities
Although not as severe as their topographical mem-ory problems GR and CO do have some memorydifficulties in other domains For example bothperformed poorly on the Rey AVLT whichrequired them to remember a list of unrelatedwords Both also report some subtle episodic mem-ory problems (see Case Description) COrsquos prob-lems could partly result from damage to the righthippocampusmdashindeed CO appeared to have themore acute episodic memory problems HoweverGR has no hippocampal damage so the fact that hedisplays episodic memory problems suggests thatparahippocampal cortex is involved in the forma-
tion of episodic memories (cf Wagner et al1998)
In previous work (Epstein et al 1999) weargued that spatial context information encoded byparahippocampal cortex might play a role in estab-lishing event memories by encoding informationabout where events take place The inability toencode this information might cause problems intasks such as the Rey AVLT where one has to recalla list of unrelated words If spatial (and perhapstemporal) context information is not encoded itwill be harder subsequently to recall the distinctevent of hearing each word (Tulving 1983) Incontrast lack of spatial context information wouldbe less of a problem in tasks where one is onlyrequired to recognise previously encountered verbalmaterials but not to reproduce the spatiotemporalcontext in which they were experienced IndeedGR and CO performed as well as normals on thenonsense words in Experiment 4 where recogni-tion memory but not episodic memory was requiredfor successful performance Interestingly GR andCO also did relatively well on the Logical Memorysubtest of the WMS-III perhaps because the nar-rative framework of a story provides sufficient con-textual information to overcome the absence ofspatial context and mediate successful verbalretrieval
Differences between GR and CO
Although we have discussed GR and CO togetherthere are a number of relevant differences betweenthem One open question is whether these differ-ences can be attributed to the different effects ofunilateral and bilateral parahippocampal damageIn general GRrsquos topographical difficulties weremore severe than COrsquos CO did surprisingly well onthe Warrington Topographical Memory Test inwhich he was required to remember detailed real-world city scenes and also performed substantiallybetter on real scenes than Lego scenes in Experi-ment 3 Insofar as CO has intact left para-hippocampal cortex this suggests that the left PPAmight be sufficient to encode a certain amount of(perhaps semantic) information about real-worldscenes but the right PPA is necessary to encode
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 505
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)

Lego scenes that must be recognised solely on thebasis of their spatial structure Clearly however theamount of topographical information that can beencoded using only a left PPA is strictly limitedAnother difference between the two patients is thatGR reports some kind of perceptual problem butCO does not It is presently unclear whether thesedifferences are indicative of qualitatively differentkinds of impairment after unilateral and bilaterallesions or whether the same abilities are affected inboth cases but to a more severe degree with bilaterallesions
Conclusion
We examined two patients with lesions toparahippocampal cortex Both exhibited severetopographical difficulties particularly in new envi-ronments A key component of their problem is amemory encoding deficit that is more severe forvisual stimuli that depict spaces that one can navi-gate through or act within than for other visualmaterials We found no clear evidence thatparahippocampal cortex is necessary for the initialperceptual processing of visual scenes recognitionof landmarks or recall of premorbidly learned top-ographical information Parahippocampal cortexmay be the neuroanatomical locus of a learningmechanism preferentially involved in the encodingof topographical materials into memory
Manuscript received 20 December 1999Revised manuscript received 14 June 2000
Revised manuscript accepted 20 October 2000
REFERENCES
Aguirre GK amp DrsquoEsposito M (1999) Topographicaldisorientation A synthesis and taxonomy Brain 1221613ndash1628
Aguirre GK Detre JA Alsop DC amp DrsquoEspositoM (1996) The parahippocampus subserves topo-graphical learning in man Cerebral Cortex 6 823ndash829
Aguirre GK Zarahn E amp DrsquoEsposito M (1998)Anarea within human ventral cortex sensitive to ldquobuild-
ingrdquo stimuli Evidence and implications Neuron 21373ndash383
Barrash J Damasio H Adolphs R amp Tranel D(2000) The neuroanatomical correlates of routelearning impairment Neuropsychologia 38 820ndash836
Beauchamp MS Haxby JV Rosen AC amp DeYoeEA (2000) Functional MRI correlates of recoveryof function in acquired cerebral dyschromatopsiaNeuropsychologia 38 1170ndash1179
Bohbot V Kalina M Stepaknova K Spackova NPetrides M amp Nadel L (1998) Spatial memorydeficits in patients with lesions to the right hippocam-pus and to the right parahippocampal cortexNeuropsychologia 36 1217ndash1238
Chao LL Haxby JV amp Martin A (1999) Attrib-ute-based neural substrates in temporal cortex forperceiving and knowing about objects Nature Neuro-science 2 913ndash919
Chen LL Lin LH Green EJ Barnes CA ampMcNaughton BL (1994) Head direction cells inthe rat posterior cortex I Anatomical distributionand behavioral modulation Experimental BrainResearch 101 24ndash34
Cheng K (1986)A purely geometric module in the ratrsquosspatial representation Cognition 23 149ndash178
Cipolotti L Robinson G Blair J amp Frith U (1999)Fractionation of visual memory Evidence from a casewith multiple neurodevelopmental impairmentsNeuropsychologia 37 455ndash465
Eichenbaum H Dudchenko P Wood E ShapiroM amp Tanila H (1999) The hippocampus mem-ory and place cells Is it spatial memory or a memoryspace Neuron 23 209ndash226
Epstein R Harris A Stanley D amp Kanwisher N(1999) The parahippocampal place area Recogni-tion navigation or encoding Neuron 23 115ndash125
Epstein R amp Kanwisher N (1998) A cortical repre-sentation of the local visual environment Nature 392598ndash601
Epstein R amp Kanwisher N (2001) Mnemonic func-tions of parahippocampal cortex An event-related fMRIstudy Manuscript submitted for publication
Evans AC Collins DL Mills SR Brown EDKelly RL amp Peters TM (1993) 3D statisticalneuroanatomical models from 305 MRI volumes InProceedings of the IEEE-Nuclear Science Symposiumand Medical Imaging Conference (pp 1813ndash1817)
Gallistel CR (1990) The organization of learningCambridge MA MIT Press
Gallistel CR (1995) The replacement of general-pur-pose theories with adaptive specializations In MS
EPSTEIN ET AL
506 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)
Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)

Gazzaniga (Ed) The cognitive neurosciences Cam-bridge MA MIT Press
Ghaem O Mellet E Crivello F Tzourio NMazoyer B Berthoz A amp Denis M (1997)Men-tal navigation along memorized routes activates thehippocampus precuneus and insula NeuroReport 8739ndash744
Habib M amp Sirigu A (1987) Pure topographical dis-orientation a definition and anatomical basis Cortex23 73ndash85
Hecaen H Tzortzis C amp Rondot P (1980) Loss oftopographic memory with learning deficits Cortex16 525ndash542
Hermer L amp Spelke ES (1994) A geometric processfor spatial reorientation in young children Nature370 57ndash59
Hermer L amp Spelke ES (1996) Modularity anddevelopment the case of spatial reorientation Cogni-tion 61 195ndash232
Hermer-Vasquez L Spelke ES amp Katsnelson AS(1999) Sources of flexibility in human cognitionDual-task studies of space and language CognitivePsychology 39 3ndash36
Incisa della Rocchetta A Cipolotti L amp WarringtonEK (1996) Topographical disorientation Selectiveimpairment of locomotorspace Cortex 32 727ndash735
Intraub H (1997) The representation of visual scenesTrends in Cognitive Sciences 1 217ndash222
Ishai A Ungerleider LG Martin A Schouten JLamp Haxby JV (1999) Distributed representation ofobjects in the human ventral visual pathway Proceed-ings of the National Academy of Sciences USA 969379ndash9384
Kanwisher N Chun MM McDermott J ampLedden PJ (1996) Functional imaging of humanvisual recognition Cognitive Brain Research 5 55ndash67
Kanwisher N McDermott J amp Chun MM (1997)The fusiform face area a module in humanextrastriate cortex specialized for face perceptionJournal of Neuroscience 17 4302ndash4311
Landis T Cummings JL Benson DF amp PalmerEP (1986) Loss of topographic familiarity an envi-ronmental agnosia Archives of Neurology 43 132ndash136
Levine DN Warach J amp Farah MJ (1985) Twovisual systems in mental imagery Dissociation ofldquowhatrdquo and ldquowhererdquo in imagery disorders due to bilat-eral posterior cerebral lesions Neurology 35 1010ndash1018
Maguire EA Burgess N Donnett JG FrackowiakRSJ Frith CD amp OrsquoKeefe J (1998) Knowing
where and getting there A human navigation net-work Science 280 921ndash924
Maguire EA Burke T Philips J amp Staunton H(1996) Topographical disorientation following uni-lateral temporal lobe lesions in humans Neuro-psychologia 34 994ndash1001
Maguire EA amp Cipolotti L (1998) Selective sparingof topographical memory Journal of NeurologyNeurosurgery and Psychiatry 65 903ndash909
Maguire EA Frackowiak RSJ amp Frith CD(1996) Learning to find your way A role for thehuman hippocampal region Proceedings of the RoyalSociety of London B Series [Biological Sciences] 2631745ndash1750
Maguire EA Frackowiak RSJ amp Frith CD(1997) Recalling routes around London Activationof the right hippocampus in taxi drivers Journal ofNeuroscience 17 7103ndash7110
Malach R Reppas JB Benson RB Kwong KKJiang H Kennedy WA Ledden PJ Brady TJRosen BR amp Tootell RBH (1995) Object-related activity revealed by functional magnetic reso-nance imaging in human occipital cortex Proceedingsof the National Academy of Science USA 92 8135ndash8138
Margules J amp Gallistel CR (1988) Heading in therat determination by environmental shape AnimalLearning and Behavior 16 404ndash410
McCarthy RA Evans JJ amp Hodges JR (1996)Topographic amnesia Spatial memory disorderperceptual disfunction or category-specific semanticmemory impairment Journal of Neurology Neurosur-gery and Psychiatry 60 318ndash325
Milner AD amp Goodale MA (1995)The visual brainin action Oxford Oxford University Press
OrsquoKeefe J amp Nadel L (1978)The hippocampus as a cog-nitive map Oxford Oxford University Press
Pallis CA (1955) Impaired identification of faces andplaces with agnosia for colours Journal of NeurologyNeurosurgery and Psychiatry 18 218ndash224
Pigott S amp Milner B (1993) Memory for differentaspects of complex visual scenes after unilateral tem-poral- or frontal-lobe resection Neuropsychologia 311ndash15
Rao SM Hammeke TA McQuillen MP KhatriBO amp Lloyd D (1984) Memory disturbance inchronic progressive multiple sclerosis Archives ofNeurology 41 625ndash631
Ross ED (1980) Sensory-specific and fractional disor-ders of recent memory in man I Isolated loss of visualrecent memory Archives of Neurology 37 193ndash200
COGNITIVE NEUROPSYCHOLOGY 2001 18 (6) 507
PARAHIPPOCAMPUS amp TOPOGRAPHICAL LEARNING
Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)

Spreen O amp Strauss E (1998) A compendium of neuro-psychological tests Administration norms and commen-tary New York Oxford University Press
Stark M Coslett HB amp Saffran EM (1996)Impairment of an egocentric map of locations Impli-cations for perception and action Cognitive Neuro-psychology 13 481ndash523
Suzuki WA amp Amaral DG (1994) Perirhinal andparahippocampal cortices of the macaque monkeyCortical referents Journal of Comparative Neurology350 497ndash533
Talairach J amp Tournoux P (1988) Co-planar stereo-tactic atlas of the human brain Stuttgart GermanyThieme
Teng E amp Squire LR (1999) Memory for placeslearned long ago is intact after hippocampal damageNature 400 675ndash677
Tippett LJ Miller LA amp Farah MJ (2000)Prosopamnesia A selective impairment in face learn-ing Cognitive Neuropsychology 17 241ndash256
Tulving E (1983) Elements of episodic memory OxfordClarendon Press
Wagner AD Schacter DL Rotte M KoutstaalW Maril A Dale AM Rosen BR amp BucknerRL (1998) Building memories Remembering andforgetting of verbal experiences as predicted by brainactivity Science 281 1188ndash1191
Wechsler D (1997a) Wechsler Adult Intelligence ScalemdashIII Administration and scoring manual San AntonioTX The Psychological Corporation Harcourt Braceamp Company
Wechsler D (1997b) Wechsler Memory ScalemdashIIIAdministration and scoring manual San Antonio TXThe Psychological Corporation Harcourt Brace ampCompany
Whiteley AM amp Warrington EK (1978) Selectiveimpairment of topographic memory A single casestudy Journal of Neurology Neurosurgery and Psychia-try 41 575ndash578
EPSTEIN ET AL
508 COGNITIVE NEUROPSYCHOLOGY 2001 18 (6)