neospora caninum: infection induces high lysosomal activity
TRANSCRIPT
Experimental Parasitology 134 (2013) 409–412
Contents lists available at SciVerse ScienceDirect
Experimental Parasitology
journal homepage: www.elsevier .com/locate /yexpr
Neospora caninum: Infection induces high lysosomal activity
0014-4894/$ - see front matter � 2013 Elsevier Inc. All rights reserved.http://dx.doi.org/10.1016/j.exppara.2013.04.008
⇑ Corresponding author. Address: Centro de Ciências Agrárias, Ambientais eBiológicas, UFRB, Rua Rui Barbosa, 710, Centro, 44.380-000 Cruz das Almas, Bahia,Brasil.
E-mail address: [email protected] (A.M. Pinheiro).
Alexandre M. Pinheiro a,⇑, Cláudia Valle Cabral D. Santos b, Luiz Erlon A. Rodrigues b
a Laboratory of Veterinary Biochemistry and Immunology, CCAAB – Centro de Ciências Agrárias, Ambientais e Biológicas, Universidade Federal do Recôncavo da Bahia, Brazilb Laboratory of Basic Research, EBMSP – Fundação Bahiana para Desenvolvimento das Ciências, Salvador, Bahia, Brazil
h i g h l i g h t s
� The activity of acid phosphatase wasassayed in the medium of cellcultures.� Neospora caninum induces high
production of acid phosphatase.� Lysosomal compartment could play
an important role in cell infection byN. caninum.
g r a p h i c a l a b s t r a c t
a r t i c l e i n f o
Article history:Received 31 July 2012Received in revised form 28 January 2013Accepted 24 April 2013Available online 3 May 2013
Keywords:Neospora caninumLysosomal activityAstrocytes
a b s t r a c t
Neospora caninum is a protozoan that causes abortion in cattle and neuromuscular lesions in dogs, mak-ing it an important target of veterinary medicine. Lysosomes are cellular organelles responsible forimportant biological functions as cellular defense mechanisms. The aim of this work was to evaluatethe lysosomal stability of rat gliocytes infected in vitro with N. caninum. Rat glial cultures were infectedat a ratio of 1:1 (cell/parasite). The enzymatic activity of acid phosphatase (orthophosphoric-monoesterphosphohydrolase, EC 3.1.3.2) was assayed in the medium of control and infected cell cultures. The activ-ity observed at 24 h of incubation was 0.4 ± 0.08 mU/mg/min for control cells and 1.3 ± 0.5 mU/mg/minfor infected cells. After 72 h, control and infected cells exhibited activities of 1.3 ± 0.5 and 4.1 ± 0.9 mU/mg/min, respectively. These results suggested that lysosomal compartment plays an important role in themechanisms of cellular infection by N. caninum.
� 2013 Elsevier Inc. All rights reserved.
1. Introduction
Neospora caninum has been identified as an agent responsiblefor abortion in cattle and neuromuscular alterations in dogs (Du-bey, 2003; Innes et al., 2005; Dubey et al., 2006). Nowadays thisparasite continues to be a major problem in cattle (Dubey and Sch-ares, 2011). Various animal species are intermediate hosts for thisApicomplexa protozoan, but only dogs, coyotes (Canis latrans) anddingoes (Canis lupus dingo) are identified as definitive hosts (McAll-
ister et al., 1998; Dubey et al., 2002; Gondim et al., 2004; Kinget al., 2010). This parasite has a tropism for the central nervous sys-tem (CNS), infecting and eliciting a response from glial cells (Pinhe-iro et al., 2006a).
One of the most important cells of the CNS is astrocyte. Thesecells supply energy to neurons, regulate extracellular homeostasis,and also participate in the formation and maintenance of thehemato-encephalic barrier, regulation of synaptic transmission,CNS neurogenesis and defense, and expression of receptors andseveral chemical messengers (van Wagoner and Benveniste,1999; Baumann and Pham-Dinh, 2001; Seifert et al., 2006). Thesecells are emerging as mediator of brain development, functionand plasticity (Stipursky et al., 2012). Microglia cells are activephagocytes and multiply when activated by various stimuli, such

410 A.M. Pinheiro et al. / Experimental Parasitology 134 (2013) 409–412
as lesions (Kreutzberg, 1996). Activated microglia releases variousneurotrophic factors and cytokines that can affect neurogenesis(Wake et al., 2012).
It is well known that lysosomal compartment consists of pri-mary lysosomes, auto and heterophagic vesicles, secondary lyso-somes (phagosomes), and residual corpuscles. It is amultifunctional cellular organelle, rich in 50 hydrolytic enzymes,and responsible for degrading all cellular constituents and macro-molecules that come from extracellular space, by endocytosis. Inaddition to its primary function, the lysosomal compartment is in-volved in cellular defense mechanisms against toxic and infectionagents (Cuervo and Dice, 2000; Hafner Cesen et al., 2012). One ofthese enzymes is acid phosphatase, a marker lysosomal that partic-ipates with other acid hydrolases in the digestion of biologic mate-rial delivered to lysosomes.
Once N. caninum multiply inside cellular vesicles using plasmamembranes of host cells, their inclusions in membrane-derivedvesicles may help them to escape, neutralizing immune responsesand lysosomal attack (Buxton et al., 2002; Dubey et al., 2002). Forthis reasons the aim of this study was to verify the stability of lyso-somal membranes in rat gliocytes infected in vitro with N. caninum.
2. Material and methods
2.1. Neuroglia cultures
Astrocytes cultures were obtained as describe to Pinheiro et al.(2006a). Briefly, cortex of newborn rats (<48 h) were mechanicallydissociated through an 80-lm filter and 5 � 105 cells were sus-pended in 1.0 mL of Dulbecco’s Modified Eagle’s Medium (DMEM),supplemented with 10% (v/v) of fetal bovine serum (Cultilab, Bra-zil), 1 mM pyruvic acid and 2 mM glutamine (Sigma, St. Louis,MO, USA), and seeded into 40-mm culture dishes (TPP, Switzer-land). Cultures were incubated at 37 �C in a humidified atmospherewith 5% CO2 and the medium, was changed every 48 h. Experi-ments were performed on three different cultures at 2 weeks,when the astroglial monolayer was confluent. This procedure al-lows the detection of at least 86% of GFAP (Glial Fibrillary AcidicProtein) positive cells, which is a specific marker of astrocytes.
2.2. Infection of glial cultures with N. caninum
Cells were infected with tachyzoites of N. caninum (NC Bahiastrain – Gondim et al., 2001) that were maintained in Vero cellswith regular changes of DMEM medium, supplemented with 10%fetal bovine serum, and purified using a 5-lm filter (Millipore –Carrigtwohill Co., Ireland). The parasites were counted and the glialcells were infected at a ratio of 1:1 (glia/parasite).
Fig. 1. Immunocytochemistry of uninfected glial cell cultures at 24 h of incubation.Astrocytes cell bodies are show in red stained with GFAP. Cell nuclei were stainedwith Hoechst 33258 to appear blue. Magnification – 200�. (For interpretation of thereferences to colour in this figure legend, the reader is referred to the web version ofthis article.)
2.3. Immunocytochemistry
Control and infected cells were fixed in ice-cold methanol for20 min at �20 �C. In order to detect the astroglial marker GFAP,cells were incubated with rabbit anti-GFAP IgG antibody (DAKO,Denmark), which was diluted 1:500 in PBS, in a humid chamberovernight at 4 �C. The labeled cells were then incubated with TRITC(tetramethyl-rhodamine isothiocyanate) conjugated to anti-IgG(Cappel, Durham, Canada), which was also diluted 1:500, for 2 hat 37 �C. Nuclei were stained after incubation with the nucleardye Cell stain (C25H27Cl3N6O – Hoechst 33258) for 10 min at roomtemperature. Normal rabbit serum was used as negative control(Pinheiro et al., 2006b).
2.4. Lysosomal activity
Lysosomal activities were measured in the media of control andinfected cell cultures after 24 and 72 h of incubation. Acid phos-phatase (orthophosphoric-monoester phosphohydrolase, EC3.1.3.2) activity was measured as a surrogate for lysosomal activityaccording to the technique described by Yang et al. (1996), with acommercial diagnostic kit according to manufacture instructions(Doles Reag., Brazil). Total protein was measured using the Folin-biuret technique modified by Komsa-Penkova et al. (1996), andthe results were expressed as milliunits of specific acid phospha-tase activity (one milliunit being equivalent to the enzymatic activ-ity necessary to hydrolyze one micromole of substrate (sodiumthymolphtalein monophosphate) at 37 �C and pH 5.4 during oneminute of incubation time per milligram of total protein).
The lysosomal activity was expressed as mean ± standard devi-ation and a Student’s t-test was used to determine the statisticalsignificance between control and infected groups. P values lessthan 0.05 were deemed significant.
3. Results
Immunocytochemistry assay showed that more than 86% cellwere GFAP (a specific marker of astrocytes) positive (Fig. 1). Lyso-somal activities of control and N. caninum infected cultures, as as-sessed by measuring acid phosphatase activity, are presented inFig. 2. The mean activities for the control and infected cells at24 h of incubation with N. caninum were 0.4 ± 0.08 and1.3 ± 0.5 mU/mg/min, respectively. After 72 h of incubation, con-trol and infected groups had activities of 1.3 ± 0.5 and4.1 ± 0.9 mU/mg/min, respectively. Infected cultures exhibited spe-cific activities three times greater than those observed for the con-trol group (p < 0.01). Meanwhile the ratio of 24-hour (1.3/0.4 = 3.25) to 72-hour results (4.1/1.3 = 3.15) was identical.
4. Discussion
The aim of this work was to evaluate acid phosphatase (ortho-phosphoric-monoester phosphohydrolase, EC 3.1.3.2) activity ofrat gliocytes infected with N. caninum. This procedure allows aninvestigation of the physicochemical properties associated withstability of lysosomal membranes (Rodrigues et al., 1998). Usingrat gliocytes cultures infected with N. caninum we have shown thatthe activity of acid phosphatase was three times higher in infected
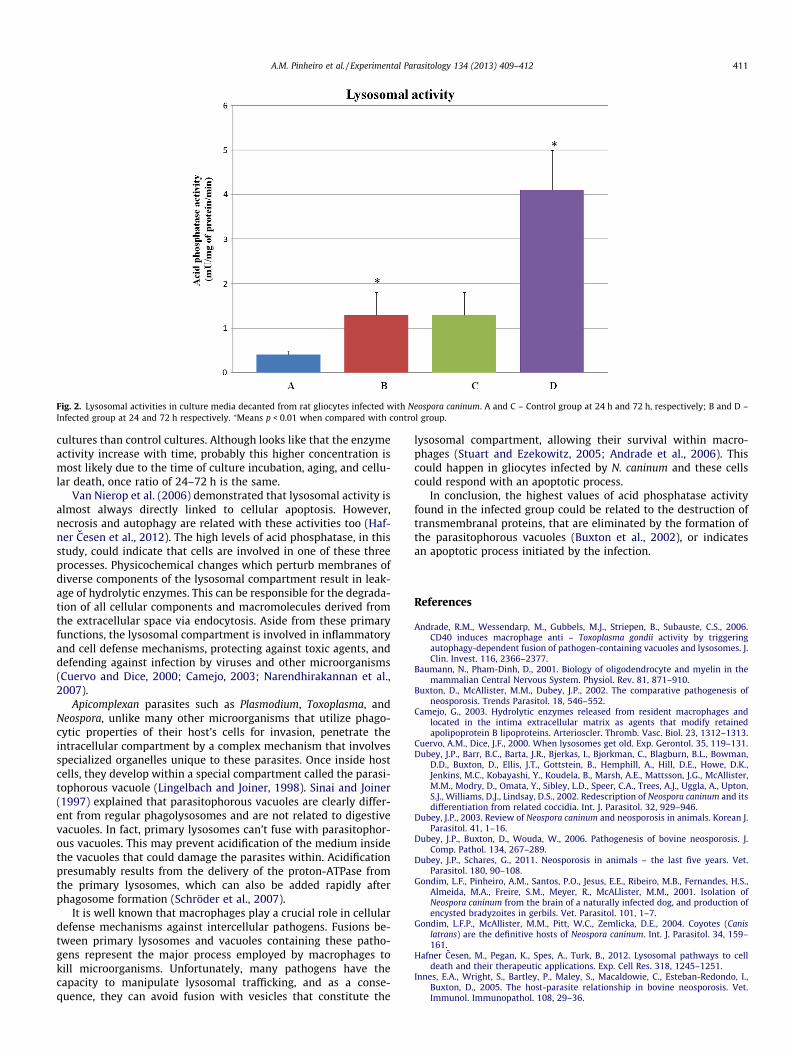
Fig. 2. Lysosomal activities in culture media decanted from rat gliocytes infected with Neospora caninum. A and C – Control group at 24 h and 72 h, respectively; B and D –Infected group at 24 and 72 h respectively. ⁄Means p < 0.01 when compared with control group.
A.M. Pinheiro et al. / Experimental Parasitology 134 (2013) 409–412 411
cultures than control cultures. Although looks like that the enzymeactivity increase with time, probably this higher concentration ismost likely due to the time of culture incubation, aging, and cellu-lar death, once ratio of 24–72 h is the same.
Van Nierop et al. (2006) demonstrated that lysosomal activity isalmost always directly linked to cellular apoptosis. However,necrosis and autophagy are related with these activities too (Haf-ner Cesen et al., 2012). The high levels of acid phosphatase, in thisstudy, could indicate that cells are involved in one of these threeprocesses. Physicochemical changes which perturb membranes ofdiverse components of the lysosomal compartment result in leak-age of hydrolytic enzymes. This can be responsible for the degrada-tion of all cellular components and macromolecules derived fromthe extracellular space via endocytosis. Aside from these primaryfunctions, the lysosomal compartment is involved in inflammatoryand cell defense mechanisms, protecting against toxic agents, anddefending against infection by viruses and other microorganisms(Cuervo and Dice, 2000; Camejo, 2003; Narendhirakannan et al.,2007).
Apicomplexan parasites such as Plasmodium, Toxoplasma, andNeospora, unlike many other microorganisms that utilize phago-cytic properties of their host’s cells for invasion, penetrate theintracellular compartment by a complex mechanism that involvesspecialized organelles unique to these parasites. Once inside hostcells, they develop within a special compartment called the parasi-tophorous vacuole (Lingelbach and Joiner, 1998). Sinai and Joiner(1997) explained that parasitophorous vacuoles are clearly differ-ent from regular phagolysosomes and are not related to digestivevacuoles. In fact, primary lysosomes can’t fuse with parasitophor-ous vacuoles. This may prevent acidification of the medium insidethe vacuoles that could damage the parasites within. Acidificationpresumably results from the delivery of the proton-ATPase fromthe primary lysosomes, which can also be added rapidly afterphagosome formation (Schröder et al., 2007).
It is well known that macrophages play a crucial role in cellulardefense mechanisms against intercellular pathogens. Fusions be-tween primary lysosomes and vacuoles containing these patho-gens represent the major process employed by macrophages tokill microorganisms. Unfortunately, many pathogens have thecapacity to manipulate lysosomal trafficking, and as a conse-quence, they can avoid fusion with vesicles that constitute the
lysosomal compartment, allowing their survival within macro-phages (Stuart and Ezekowitz, 2005; Andrade et al., 2006). Thiscould happen in gliocytes infected by N. caninum and these cellscould respond with an apoptotic process.
In conclusion, the highest values of acid phosphatase activityfound in the infected group could be related to the destruction oftransmembranal proteins, that are eliminated by the formation ofthe parasitophorous vacuoles (Buxton et al., 2002), or indicatesan apoptotic process initiated by the infection.
References
Andrade, R.M., Wessendarp, M., Gubbels, M.J., Striepen, B., Subauste, C.S., 2006.CD40 induces macrophage anti – Toxoplasma gondii activity by triggeringautophagy-dependent fusion of pathogen-containing vacuoles and lysosomes. J.Clin. Invest. 116, 2366–2377.
Baumann, N., Pham-Dinh, D., 2001. Biology of oligodendrocyte and myelin in themammalian Central Nervous System. Physiol. Rev. 81, 871–910.
Buxton, D., McAllister, M.M., Dubey, J.P., 2002. The comparative pathogenesis ofneosporosis. Trends Parasitol. 18, 546–552.
Camejo, G., 2003. Hydrolytic enzymes released from resident macrophages andlocated in the intima extracellular matrix as agents that modify retainedapolipoprotein B lipoproteins. Arterioscler. Thromb. Vasc. Biol. 23, 1312–1313.
Cuervo, A.M., Dice, J.F., 2000. When lysosomes get old. Exp. Gerontol. 35, 119–131.Dubey, J.P., Barr, B.C., Barta, J.R., Bjerkas, I., Bjorkman, C., Blagburn, B.L., Bowman,
D.D., Buxton, D., Ellis, J.T., Gottstein, B., Hemphill, A., Hill, D.E., Howe, D.K.,Jenkins, M.C., Kobayashi, Y., Koudela, B., Marsh, A.E., Mattsson, J.G., McAllister,M.M., Modry, D., Omata, Y., Sibley, L.D., Speer, C.A., Trees, A.J., Uggla, A., Upton,S.J., Williams, D.J., Lindsay, D.S., 2002. Redescription of Neospora caninum and itsdifferentiation from related coccidia. Int. J. Parasitol. 32, 929–946.
Dubey, J.P., 2003. Review of Neospora caninum and neosporosis in animals. Korean J.Parasitol. 41, 1–16.
Dubey, J.P., Buxton, D., Wouda, W., 2006. Pathogenesis of bovine neosporosis. J.Comp. Pathol. 134, 267–289.
Dubey, J.P., Schares, G., 2011. Neosporosis in animals – the last five years. Vet.Parasitol. 180, 90–108.
Gondim, L.F., Pinheiro, A.M., Santos, P.O., Jesus, E.E., Ribeiro, M.B., Fernandes, H.S.,Almeida, M.A., Freire, S.M., Meyer, R., McALlister, M.M., 2001. Isolation ofNeospora caninum from the brain of a naturally infected dog, and production ofencysted bradyzoites in gerbils. Vet. Parasitol. 101, 1–7.
Gondim, L.F.P., McAllister, M.M., Pitt, W.C., Zemlicka, D.E., 2004. Coyotes (Canislatrans) are the definitive hosts of Neospora caninum. Int. J. Parasitol. 34, 159–161.
Hafner Cesen, M., Pegan, K., Spes, A., Turk, B., 2012. Lysosomal pathways to celldeath and their therapeutic applications. Exp. Cell Res. 318, 1245–1251.
Innes, E.A., Wright, S., Bartley, P., Maley, S., Macaldowie, C., Esteban-Redondo, I.,Buxton, D., 2005. The host-parasite relationship in bovine neosporosis. Vet.Immunol. Immunopathol. 108, 29–36.

412 A.M. Pinheiro et al. / Experimental Parasitology 134 (2013) 409–412
King, J.S., Slapeta, J., Jenkins, D.J., Al-Qassab, S.E., Ellis, J.T., Windsor, P.A., 2010.Australian dingoes are definitive hosts of Neospora caninum. Int. J. Parasitol. 40,945–950.
Komsa-Penkova, R., Spirova, R., Bechev, B., 1996. Modifications of Lowry’s methodfor collagen concentration measurement. J. Biochem. Biophys. Methods 32, 33–43.
Kreutzberg, G.W., 1996. Microglia: a sensor for pathological events in the CNS.Trends Neurosci. 19, 312–318.
Lingelbach, K., Joiner, K.A., 1998. The parasitophorous vacuole membranesurrounding plasmodium and toxoplasma: unusual compartment in infectedcells. J. Cell Sci. 111, 1467–1475.
McAllister, M.M., Dubey, J.P., Lindsay, D.S., Jolley, W.R., Wills, R.A., McGuire, A.M.,1998. Dogs are the definitive hosts of Neospora caninum. Int. J. Parasitol. 28,1473–1478.
Narendhirakannan, R.T., Subramanian, S., Kandaswamy, M., 2007. Anti-inflammatory and lysosomal stability action of Cleome gynandra L. studied inadjuvant induced arthritic rats. Food Chem. Toxicol. 45, 1001–1012.
Pinheiro, A.M., Costa, S.L., Freire, S.M., Meyer, R., Almeida, M.A.O., Tardy, M., ELBachá, R., Costa, M.F.D., 2006a. Neospora caninum: infection induced IL-10 overexpression in rat astrocytes in vitro. Exp. Parasitol. 112, 193–197.
Pinheiro, A.M., Costa, S.L., Freire, S.M., Almeida, M.A.O., Tardy, M., EL Bachá, R., Costa,M.F.D., 2006b. A stroglial cells in primary culture: a valid model to studyNeospora caninum infection in the CNS. Vet. Immunol. Immunopathol. 113,243–247.
Rodrigues, L.E.A., Paes, B.I., Jacobina, H., 1998. Role of lysosomes on humanulcerogenic gastropathies. Effect of zinc ion on the lysosomal stability. Arch.Gastroenterol. 35, 247–251.
Schröder, B., Wrocklage, C., Pan, C., Jäger, R., Kösters, B., Schäfer, H., Elsässer, H.P.,Mann, M., Hasilik, A., 2007. Integral and associated lysosomal membraneproteins. Traffic 8, 1676–1686.
Seifert, G., Schilling, K., Steinhäuser, C., 2006. Astrocyte dysfunction in neurologicaldisorders: a molecular perspective. Nat. Rev. Neurosci. 7, 194–206.
Sinai, A.P., Joiner, K.A., 1997. Safe haven: the cell biology of nonfusogenic pathogenvacuole. Annu. Rev. Microbiol. 51, 415–462.
Stipursky, J., Spohr, T.C., Sousa, V.O., Gomes, F.C., 2012. Neuron-astroglialinteractions in cell-fate commitment and maturation in the central nervoussystem. Neurochem. Res., 22 [Epub ahead of print].
Stuart, L.M., Ezekowitz, R.A.B., 2005. Phagocytosis: elegant complexity. Immunity22, 539–550.
van Nierop, K., Muller, F.J.M., Stap, J., van Noorden, C.J.F., van Eikjk, M., de Groot, C.,2006. Lysosomal destabilization contributes to apoptosis of germinal center B-lymphocytes. J. Histochem. Cytochem. 54, 1425–1453.
van Wagoner, N.J., Benveniste, E.N., 1999. Interleukin-6 expression and regulationin astrocytes. J. Neuroimmunol. 100, 124–139.
Wake, H., Moorhouse, A.J., Nabekura, J., 2012. Functions of microglia in the centralnervous system – beyond the immune response. Neuron Glia Biol. 22, 1–7.
Yang, T.T., Sinai, P., Kain, S.R., 1996. An acid phosphatase assay for quantifying thegrowth of adherent and nonadherent cells. Anal. Biochem. 241, 103–108.