neogene sloth assemblages (mammalia, pilosa) of...
TRANSCRIPT

NEOGENE SLOTH ASSEMBLAGES (MAMMALIA,
PILOSA) OF THE COCINETAS BASIN (LA GUAJIRA,
COLOMBIA): IMPLICATIONS FOR THE GREAT
AMERICAN BIOTIC INTERCHANGE
by ELI AMSON1,2 ,3 , JUAN D. CARRILLO2 and CARLOS JARAMILLO1
1Smithsonian Tropical Research Institute, Box 0843-03092, Balboa-Ancon, Panama; [email protected] Institut und Museum der Universitat Zurich, Karl Schmid-Strasse 4, Z€urich, CH-8006, Switzerland; [email protected] address: AG Morphologie und Formengeschichte, Bild Wissen Gestaltung – ein Interdisziplin€ares Labor & Institut f€ur Biologie, Humboldt-Universit€at,
Philippstrasse, 12/13, D-10115, Berlin, Germany; [email protected]
Typescript received 19 February 2016; accepted in revised form 8 May 2016
Abstract: We describe sloth assemblages from the Cocin-
etas Basin (La Guajira peninsula, Colombia), found in the
Neogene Castilletes and Ware formations, located in north-
ernmost South America, documenting otherwise poorly
known biotas. The tentative referral of a specimen to a small
megatherioid sloth, Hyperleptus?, from the early–middle
Miocene Castilletes Formation, suggests affinities of this
fauna with the distant Santa Cruz Formation and documents
a large latitudinal distribution for this taxon. The late Plio-
cene Ware Formation is much more diverse, with five dis-
tinct taxa representing every family of ‘ground sloths’. This
diversity is also remarkable at the ecological level, with sloths
spanning over two orders of magnitude of body mass and
probably having different feeding strategies. Being only a few
hundred kilometres away from the Isthmus of Panama, and
a few hundred thousand years older than the classically
recognized first main pulse of the Great American Biotic
interchange (GABI 1), the Ware Formation furthermore doc-
uments an important fauna for the understanding of this
major event in Neogene palaeobiogeography. The sloths for
which unambiguous affinities were recovered are not closely
related to the early immigrants found in North America
before GABI 1.
Key words: Cocinetas Basin, Great American Biotic Inter-
change, Neotropics, palaeobiodiversity, Pilosa, sloth.
SLOTHS (Tardigrada) are represented today by two gen-
era that are ecologically restricted to a peculiar suspen-
sory mode of life, and geographically restricted to the
Neotropical region (Nowak & Paradiso 1983). However,
this modern distribution is depauperate when compared
to the recent past, as their fossil record includes a wide
range of body sizes (up to several metric tones; Fari~na
et al. 1998; Raj Pant et al. 2014), various feeding ecolo-
gies (Bargo & Vizca�ıno 2008), and a broad distribution
from Alaska (Stock 1942) to southernmost South Amer-
ica (and possibly Antarctica; Carlini et al. 1990; Gelfo
et al. 2015 and references therein). Their geographical
distribution was particularly broad during the Pleis-
tocene, which is partly the result of a major event in
Neogene palaeobiogeography: the Great American Biotic
Interchange (GABI; Wallace 1876). As South and North
America were isolated during most of the Cenozoic,
very distinct faunas populated the two landmasses
(Simpson 1980). This isolation was ruptured by the
northward migration of South American taxa and
southward migration of North American taxa (Webb
1985; Cione et al. 2015). In the classical understanding,
this process started with single-taxon dispersals (Wood-
burne 2010), involving the herald taxa sensu Webb
(2006) as early as 20.9 Ma (Bloch et al. 2016). This was
followed by the main phases of the GABI (or pulses,
GABI 1–4; sensu Woodburne 2010), occurring from 2.6–2.4 Ma, and involving the legion taxa sensu Webb
(2006). These migrations have mostly been assumed to
have been driven by the closure of the Central Ameri-
can Seaway and the formation of the Isthmus of
Panama, the most likely route of migration. However,
habitat and climatic changes probably played a much
more important role in mammal migration patterns
(Bacon et al. 2016). Indeed, the timing of the formation
of the Isthmus is nowadays challenged by new geologi-
cal data (Montes et al. 2015) and the onset of GABI by
palaeontological (Bloch et al. 2016) and molecular data
(Bacon et al. 2015), that suggest a complex process that
started at the Oligocene–Miocene transition.
© The Palaeontological Association doi: 10.1111/pala.12244 1
[Palaeontology, 2016, pp. 1–20]

All four families of ‘ground sloths’ took part in all key
steps of the interchange, pre- and post-GABI 1 sensu
Woodburne (2010). The earliest immigrants are Thi-
nobadistes (Mylodontidae) and Pliometanastes (Megalony-
chidae), which are present as herald taxa in North
America by c. 9 Ma (Webb 1989). Two other herald
taxa, Megalonyx (Megalonychidae) and ‘Glossotherium
chapadmalense’ (Mylodontidae; and the subsequent
Paramylodon; see McAfee 2009) are in North America by
c. 7 and 5 Ma respectively (Woodburne 2010). Finally,
subsequent sloths are involved in the main phase of
GABI, including Eremotherium (Megatheriidae) and
Nothrotheriops (Nothrotheriidae).
Intensive fieldwork recently undertaken in the Cocine-
tas Basin of the La Guajira peninsula of Colombia (the
northernmost continental region of South America)
resulted in the collection of thousands of specimens from
the Jimol, Castilletes and Ware formations (Moreno et al.
2015). At latitude 12°S, the Cocinetas Basin stands out in
delivering rich tropical South American palaeobiotas (Jar-
amillo et al. 2015) in the most biodiverse region of the
world (Wallace 1876), otherwise characterized by a poor
fossil record. Moreover, the Ware Formation in particular
is critical for the understanding of the GABI (Jaramillo
et al. 2015). Firstly, it is geographically near to the Isth-
mus of Panama (only 700 km away). Secondly, it is just
slightly older (c. 2.8 Ma) than GABI 1 (2.6–2.4 Ma). The
Ware Formation and its tardigradan assemblage have
therefore the potential to improve our understanding of
the dynamics of GABI in the Neotropics.
GEOLOGICAL SETTING
The Neogene stratigraphy of the Cocinetas Basin, includ-
ing the Jimol, Castilletes and Ware formations, was
recently revised (Moreno et al. 2015). Of those forma-
tions, only the Castilletes and Ware formations contain
mammalian assemblages. Both crop out along the West
coast of the La Guajira peninsula (Colombia) at various
localities, of which one from the Castilletes Formation
and several from the Ware Formation delivered specimens
ascribed to sloths (Fig. 1). The environment of deposition
of the Castilletes Formation ranges from shallow marine
to fluvio-deltaic, and it has been dated as 16.7–14.2 Ma
(late early Miocene to early middle Miocene, upper Bur-
digalian–Langhian, SALMA Santacrucian/Colloncuran;
Hendy et al. 2015; Moreno et al. 2015). The base of the
Ware Formation is dominated by fluvio-deltaic environ-
ment deposits, while the top is dominated by open-ocean
shoreface and nearshore deposits. Its age ranges from 3.40
to 2.78 Ma, late Pliocene (Piacenzian, SALMA Chapad-
malalan/Marplatan; Hendy et al. 2015; Moreno et al.
2015).
MATERIAL AND METHOD
We follow the recommendations on open nomenclature of
Bengston (1988). For dental terminology, we follow Gaudin
(2004), that is, the mesialmost tooth is designated as the
caniniform (C1 and c1 for the upper and lower respec-
tively) even if its shape departs from that of a canine-like
tooth. The following (more distal) teeth are designated as
molariforms (M1–4 and m1–3 for the upper four and three
lower molariforms respectively). Standard measurements
were taken with digital calipers when lower than 15 cm and
with a tape measure otherwise. In addition to standard
digital photographs, stereophotographs were taken using a
custom-built seesaw.
A phylogenetic analysis was conducted to suggest a
hypothesis regarding the affinities of one specimen (MUN
STRI 36643). The data matrix of Gaudin (2004) was used,
pruning all non-mylodontid taxa except Bradypus and
Megalonyx, which are used in the present analysis as out-
groups. Along with MUN STRI 36643, another additional
terminal taxon, Kiyumylodon, was included. This recently
described mylodontine was coded based on Rinderknecht
et al. (2007). The initial ordination of the characters of
Gaudin (2004) were kept. The edited data matrix is avail-
able in the Dryad Digital Repository (Amson et al. 2016a).
The algorithm employed was the exhaustive search (Branch
and bound) of PAUP 4.0b10 (Swofford 2002).
All specimens have STRI catalogue numbers and are
associated with a locality number and stratigraphic data.
F IG . 1 . Geographical distribution of tardigradan specimens in
the Cocinetas Basin. The magnified region is indicated on the
inset map by a rectangle. The cross marks the locality of the tar-
digradan specimen from the Castilletes Formation. The arrows
point to the Ware Formation outcrops. Scale bar represents
5 km. Colour online.
2 PALAEONTOLOGY

These are accessible through the STRI PaleoDatabase
(http://biogeodb.stri.si.edu/jaramillo/fossildb). Specimens
are deposited at the palaeontological collections of
MAPUKA (MUN), Universidad del Norte, Barranquilla,
Colombia.
In order to estimate body mass of several specimens, we
used the published regression equations of Janis (1990) (in
Fari~na et al. 1998). One of these equations involves the
occlusal area of first lower molariform (herein designated
as c1 for Pliomegatherium lelongi, see below). This area was
measured on a photograph (with scale) of the specimen
with the program Fiji (Schindelin et al. 2012).
Institutional abbreviations. F:AM, Frick Collection of the Ameri-
can Museum of Natural History, New York, USA; MCL, Museu
de Ciencias Naturais da Pontif�ıcia, Universidade Cat�olica de
Minas Gerais, Belo Horizonte, Brazil; MMP, Museo Municipal
de Ciencias Naturales ‘Lorenzo Scaglia’, Mar del Plata, Argen-
tina; MUN, Mapuka Museum, Universidad del Norte, Barran-
quilla, Colombia; MUT, Museo Universitario, Tarija, Bolivia;
STRI, Smithsonian Tropical Research Institute, ciudad de
Panam�a, Panam�a.
Other abbreviations. AP, anteroposterior; CC, craniocaudal; DV,
dorsoventral; GABI, Great American Biotic Interchange; LL, labi-
olingual; MD, mesiodistal; ML, mediolateral; PD, proximodistal.
Teeth: C1/c1, upper/lower caniniform; M1–4/m1–3, upper four/
lower three teeth distal to the caniniform; ‘A’ preceding one of
the former, the corresponding alveolus.
SYSTEMATIC PALAEONTOLOGY
MAMMALIA Linnaeus, 1758
XENARTHRA Cope, 1889
TARDIGRADA Latham & Davies, 1795
[= PHYLLOPHAGA Owen, 1842; = FOLIVORA Delsuc et al.,
2001]
Gen. et sp. indet.
Figure 2
Referred material. MUN STRI 20108, locality 390022, Ware For-
mation.
Description. MUN STRI 20108 consists of a left patella (maxi-
mum PD length = 66.7 mm; maximum ML width = 53.8 mm;
AP depth at centre of articular facet = 22.8 mm; apex PD length
(taken from distal border of articular facet) = 26.9 mm; facet
ML width = 52.7 mm). It represents a small-sized sloth,
approaching the dimensions of Nothrotherium for instance. The
articular surface is strongly convex mediolaterally (Fig. 2C). This
indicates a strongly developed medial trochlear ridge of the
femur. The medial portion of the articular surface is much smal-
ler than the lateral one, which is reminiscent of Hapalops
(Fig. 2B). The proximal side bears two grooves (Fig. 2C), as in
Mionothropus (De Iuliis et al. 2011). The apex is strongly tapered
(Fig. 2A, B), as in Thalassocnus (Amson et al. 2015), but it dif-
fers from that of the latter in being bent laterally.
Remarks. We will conservatively not consider MUN STRI
20108 to represent a taxon of its own in the diversity
count, as the possibility that it represents one of the taxa
recognized below cannot be excluded.
MYLODONTIDAE Gill, 1872
Gen. et sp. indet.
Figure 3
Referred material. MUN STRI 38047, locality 470062, Ware For-
mation.
Description. MUN STRI 38047 consists of an eroded posterior
dentary bearing three partial molariforms here referred to as m1–3 (m2 MD length = 8.1 mm; m3 MD length = 16.5 mm; DV
height at coronoid process = c. 56 mm; DV height at condyloid
process = 45.9 mm). The ontogenetic stage cannot be precisely
assessed. Based on the mesiodistal length of the last molariform, it
is larger than the holotype of the small-sized ‘Brievabradys’ laven-
tensis (Villarroel 2000; this species is probably a synonym of Glos-
sotheriopsis pascuali or another species of Glossotheriopsis, H. G.
McDonald, pers. comm. 2015), and approaches the dimensions of
a juvenile Glossotherium chapadmalense (Anaya & MacFadden
1995; mentioned therein as Glossotheridium chapadmalense, see
McAfee 2009). The m1 only preserves a rounded portion of distal
outer layer of the tooth (Fig. 3B). The m2–3 only preserve the lin-gual half of each tooth. The outline of m2 was most likely
F IG . 2 . Tardigrada gen. et sp. indet. from the Ware Formation,
left patella (MUN STRI 20108). A, anterior view (proximal towards
top). B, posterior view (proximal towards top). C, proximal view
(anterior towards top). Scale bar represents 2 cm. Colour online.
AMSON ET AL . : NEOGENE SLOTHS FROM THE COCINETAS BAS IN 3

subcircular. The m3 is bilobate, and the relative size of the lobes is
hard to assess due to their preservation. Such a disposition is typi-
cally found in mylodontids. It is noteworthy that geologically
older forms of uncertain status, such as Orophodon, also feature
such a disposition (Pujos & De Iuliis 2007).
The coronoid process is small and rounded dorsally (Fig. 3A).
The condyloid process has a short neck and a ventral position,
roughly at the level of the tooth row (diagnostic of the Mylodon-
tidae; Saint-Andr�e et al. 2010). The posteromedial opening of the
mandibular canal is located ventral to the anterior edge of the
dorsal tip of the coronoid process. The smaller posterolateral
opening of the mandibular canal is located at the level of the ante-
rior root of the coronoid process. Although eroded, the angular
process appears regularly rounded, differing from the posteriorly
pointed process usually seen in mylodontids.
Remarks. We will conservatively not consider MUN STRI
38047 to represent a taxon of its own in the diversity count,
as the possibility that it represents a juvenile individual of
the Lestodontini recognized below cannot be excluded.
MYLODONTINAE Gill, 1872
LESTODONTINI Ameghino, 1889 sensu Gaudin (2004)
Gen. et sp. nov.
Figure 4
Referred material. MUN STRI 36643, locality 470060, and tenta-
tively MUN STRI 20424, locality 390023 and MUN STRI 34353,
locality 470060 (see remarks below), all Ware Formation.
Description. MUN STRI 36643 consists of an edentulous partial
horizontal ramus of the dentary (Fig. 4A, B; Ac1 MD
length = 24.4 mm; Ac1 LL width = 17.8 mm; Am1 MD length
= 19.7 mm; Am1 LL width = 18.2 mm; Am2 MD
length = 24.6 mm; Am2 LL width = 19.5 mm; ramus DV depth
between c1 and m1 = c. 54 mm). The complete alveoli of the
three most mesial teeth and the mesial half of a more distal alve-
olus are preserved. Based on the depth of the horizontal ramus,
MUN STRI 36643 is roughly 20% smaller than Thinobadistes
segnis (Webb 1989), hence much smaller than Lestodon armatus
(Lydekker 1894) and much larger than Glossotheriopsis pascuali
(McDonald 1997) or ‘Brievabradys’ laventensis (the latter species
probably being either a synonym of the former or another spe-
cies of Glossotheriopsis, H. G. McDonald, pers. comm. 2015).
The c1 alveolus is slightly deflected labially, as in Thinobadistes
and Bolivartherium urumaquensis (Carlini et al. 2006a; Fig. 4A,
B). This deflection is accompanied by a labial bulge of the den-
tary, and, in occlusal view, by the presence of concavities on the
labial side of the dentary directly distal and mesial to the alveo-
lus of the caniniform, as is Thinobadistes. However, the mesial
concavity is more marked in Thinobadistes, an almost right angle
marking the posterolateral edge of the spout. In Lestodon and
Lestobradys (Rinderknecht et al. 2010), the caniniform is more
strongly deflected labially. There is no marked diastema in MUN
STRI 36643, as in Thinobadistes, Glossotherium (Owen 1842),
Kiyumylodon (Rinderknecht et al. 2007), and Pseudoprepotherium
(Hirschfeld 1985), and differing from Lestodon (in which a dia-
stema is present). In occlusal view, the outline of the c1 alveolus
is triangular with rounded edges. It bears a low longitudinal
ridge on its lingual wall, as in Lestodon and Lestobradys. In the
latter genera, however, the alveolus projects dorsally and is
somewhat twisted medially. In MUN STRI 36643, the c1 alveo-
lus is larger than those of m1 and m2, as in Lestodon and Thi-
nobadistes. An enlarged and triangular caniniform is considered
to be diagnostic of the clade formed by Thinobadistes and Lesto-
don (Lestodontini sensu Gaudin (2004); Lestodontinae sensu
Webb (1989)). However, the caniniform is relatively not as
enlarged as in the latter taxa. The m1 of the specimen F:AM
102658, referred to Thinobadistes segnis (Webb 1989) is 69% as
large as the c1 (mesiodistal length), while in MUN STRI 36643
it is 81% as large.
In occlusal view, the alveoli of m1 and m2 are oval, with their
main axis directed mesiolabially. A very slight longitudinal ridge
is found on the labial wall of m1 alveolus. The alveolus of m3,
although fragmentary, preserves part of a strong labial longitudi-
nal ridge, indicating that the tooth was lobate. A lower dentition
comprising a caniniform, mesial molariforms with subcircular
cross-section, and a lobate distal molariform, is a disposition
typically found in some mylodontines. The cross-section of m1
and m2 is oval in Kiyumylodon and Lestodon, but subtriangular
in Thinobadistes (Webb 1989).
A large mental foramen (dorsoventral height = 11.3 mm) is
found just anterior to the alveolus of the caniniform.
MUN STRI 20424 consists of a proximal fragment of the left
tibia (Fig. 4C–E; ML width = 115.2 mm; AP depth between
condyles = 63.8 mm; length from posterior edge of lateral con-
dyle to anterior edge of medial condyle = 93.8 mm). The speci-
men is from an adult, as shown by the epiphyseal closure.
F IG . 3 . Mylodontidae gen. et sp. indet. from the Ware Forma-
tion, partial left dentary with fragmentary m1–3 (MUN STRI
36643). A, lingual view (occlusal towards top). B, occlusal view
(labial towards top). Scale bar represents 5 cm. Colour online.
4 PALAEONTOLOGY
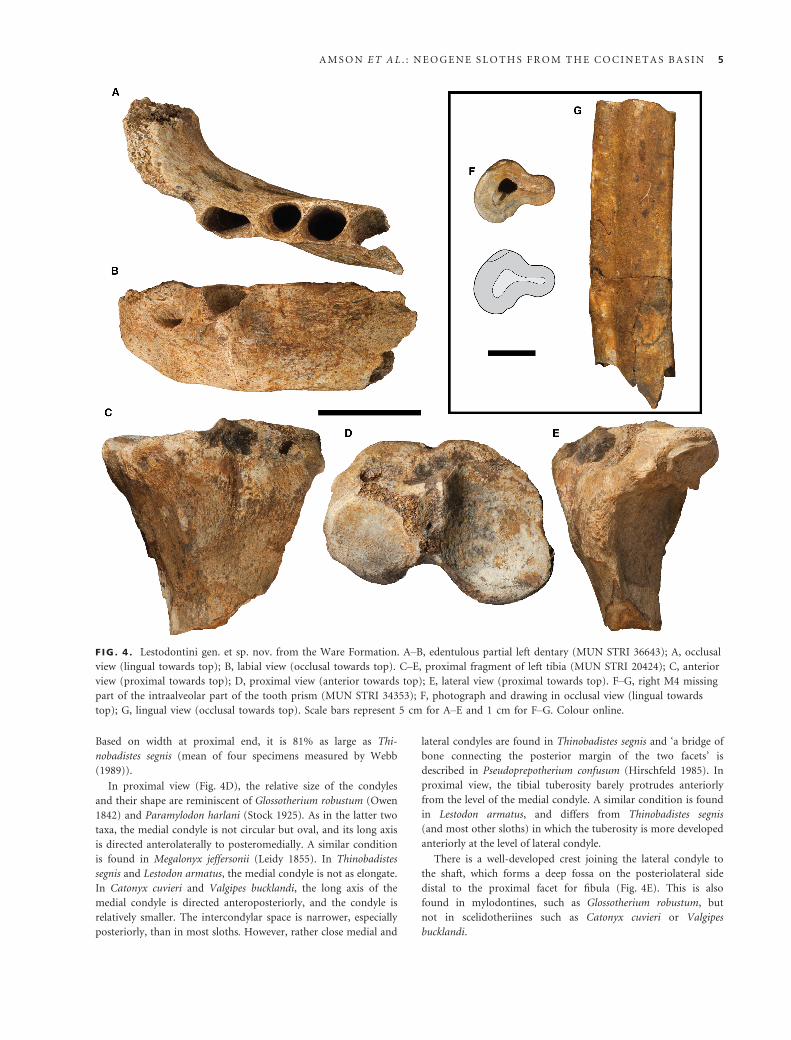
Based on width at proximal end, it is 81% as large as Thi-
nobadistes segnis (mean of four specimens measured by Webb
(1989)).
In proximal view (Fig. 4D), the relative size of the condyles
and their shape are reminiscent of Glossotherium robustum (Owen
1842) and Paramylodon harlani (Stock 1925). As in the latter two
taxa, the medial condyle is not circular but oval, and its long axis
is directed anterolaterally to posteromedially. A similar condition
is found in Megalonyx jeffersonii (Leidy 1855). In Thinobadistes
segnis and Lestodon armatus, the medial condyle is not as elongate.
In Catonyx cuvieri and Valgipes bucklandi, the long axis of the
medial condyle is directed anteroposteriorly, and the condyle is
relatively smaller. The intercondylar space is narrower, especially
posteriorly, than in most sloths. However, rather close medial and
lateral condyles are found in Thinobadistes segnis and ‘a bridge of
bone connecting the posterior margin of the two facets’ is
described in Pseudoprepotherium confusum (Hirschfeld 1985). In
proximal view, the tibial tuberosity barely protrudes anteriorly
from the level of the medial condyle. A similar condition is found
in Lestodon armatus, and differs from Thinobadistes segnis
(and most other sloths) in which the tuberosity is more developed
anteriorly at the level of lateral condyle.
There is a well-developed crest joining the lateral condyle to
the shaft, which forms a deep fossa on the posteriolateral side
distal to the proximal facet for fibula (Fig. 4E). This is also
found in mylodontines, such as Glossotherium robustum, but
not in scelidotheriines such as Catonyx cuvieri or Valgipes
bucklandi.
F IG . 4 . Lestodontini gen. et sp. nov. from the Ware Formation. A–B, edentulous partial left dentary (MUN STRI 36643); A, occlusal
view (lingual towards top); B, labial view (occlusal towards top). C–E, proximal fragment of left tibia (MUN STRI 20424); C, anterior
view (proximal towards top); D, proximal view (anterior towards top); E, lateral view (proximal towards top). F–G, right M4 missing
part of the intraalveolar part of the tooth prism (MUN STRI 34353); F, photograph and drawing in occlusal view (lingual towards
top); G, lingual view (occlusal towards top). Scale bars represent 5 cm for A–E and 1 cm for F–G. Colour online.
AMSON ET AL . : NEOGENE SLOTHS FROM THE COCINETAS BAS IN 5

The proximal facet for the fibula faces posterodistally. It faces
slightly more laterally in Glossotherium robustum and, conversely,
it faces slightly more distally in Catonyx cuvieri. There is a well-
developed facet for the cyamo-fabella as in other sloths, such as
mylodontids and megatheriids. In MUN STRI 20424, this facet
faces posteriorly and has a triangular outline.
On the anterior side of the bone, there is a crest issuing from
the tibial tuberosity that is directed mediodistally (Fig. 4C). A
similar crest is found in Glossotherium robustum (Owen 1842).
In Lestodon armatus, it is more distally directed (Gervais 1873).
MUN STRI 34353 consists of a bilobate tooth formed by con-
centric layers of orthodentine (Fig. 4F, G; MD length = 17.3 mm;
Max LL width of mesial lobe = 13.7 mm; Max LL width of distal
lobe = 6.5 mm). The inner nucleus of vasodentine is almost
entirely missing (Fig. 4F). Only patches of cementum are pre-
served at various heights along the tooth. The larger mesial lobe is
compressed mostly mesiodistally, but with a diagonal direction as
well, giving the tooth an asymmetrical shape in both the labiolin-
gual and mesiodistal directions. The smaller distal lobe is
strongly compressed labiolingually. Well-marked labial and lin-
gual sulci at the level of junction between the lobes are found
along the whole length of the tooth (Fig. 4G). The resulting
morphology is reminiscent of the right M4 of Pleurolestodon
dalenzae (Saint-Andr�e et al. 2010). The long and constricted
distal lobe of M4 is a diagnostic feature of this species, as the
other valid species of the genus, P. acutidens (Saint-Andr�e
et al. 2010), lacks the strong labiolingually compression of the
distal lobe (Rovereto 1914). However, the distal lobe is longer
and less constricted in P. dalenzae than in MUN STRI 34353,
forming a ‘secondary’ rounded nucleus. The labial side of the
mesial lobe forms a somewhat more acute curve in P. dalen-
zae than in MUN STRI 34353. In addition, the holotype of
P. dalenzae (MNHN-BOL V 3348) is larger (mesiodistal length
1.27 times greater).
It is noteworthy that an approaching morphology is featured
by the most distal lower molariform (m3) of Pseudopre-
potherium confusum (Hirschfeld 1985), but the latter bears a
mesiolabially directed facet on its mesial lobe. Moreover, the
whole vasodentine nucleus is relatively smaller in the m4 of
P. confusum.
The occlusal surface is not planar, the lingual part of the
mesial lobe being a bit less worn down than the rest of the
tooth. The lingual part of the mesial lobe bears a minute wear
facet that mostly faces occluso-lingually.
Remarks. We performed a phylogenetic analysis including
as the ingroup MUN STRI 36643 coded as a terminal
taxon and the mylodontids included in Gaudin (2004).
Three equally parsimonious trees were recovered. Their
topology is the same as the one recovered by Gaudin
(2004), as their difference lies in the position of MUN
STRI 36643. The latter is indeed either sister-group of
Lestodon, or Thinobadistes, or of the clade formed by the
two latter genera (= Lestodontini), as shown by the poly-
tomy of the strict consensus of these three trees (Fig. 5).
In a second analysis we included the mylodontine
Kiyumylodon, also known from an isolated mandible. A
total of 18 equally parsimonious trees were obtained.
Their consensus is much less resolved than the first analy-
sis, with the Mylodontinae, except for Pseudopre-
potherium, forming a polytomy. The only relationship
that is resolved within the latter polytomy is the
Lestodontini, recovered as in the previous analysis com-
prising MUN STRI 36643. As a result, MUN STRI 36643
should be considered as a Lestodontini sensu Gaudin
(2004). Differing from Lestodon and Thinobadistes, MUN
STRI 36643 most likely represents an undescribed taxon,
nov. gen et sp. but we refrain from formally erecting a new
taxon, pending the discovery of more complete material.
MUN STRI 20424 and MUN STRI 34353 can only be
safely ascribed to a medium-sized Mylodontinae gen. et sp.
indet. We tentatively ascribe them to the same taxon as
MUN STRI 36643, as no features appear be incompatible.
SCELIDOTHERIINAE Ameghino, 1904
Gen. et sp. indet.
Figure 6
Referred material. MUN STRI 16535, locality 390025, Ware For-
mation.
F IG . 5 . Hypothesis of phylogenetic affinities of MUN STRI
36643 among Mylodontidae. Strict consensus tree obtained with
PAUP 4.0b10 (Swofford 2002) that has a length of 550, a consis-
tency index of 0.59, and retention index of 0.55.
6 PALAEONTOLOGY

Description. MUN STRI 16535 consists of a complete left ulna
(see 3D model in Amson et al. 2016b). The specimen is from an
adult, as shown by the epiphyseal closure (PD length = 280 mm;
AP depth at coronoid process = 93.3 mm). MUN STRI 16535 is
shorter than all scelidotheriines measured by McDonald (1987),
which span from 313 to 476 mm. The four specimens of Sceli-
dotherium leptocephalum measured by Mi~no-Boilini et al. (2014)
are also longer, the smaller measuring 370 mm. It is however
longer than that of Pseudoprepotherium confusum (c. 20 cm;
Hirschfeld 1985).
In lateral view (Fig. 6C), the posterior end of the olecranon
process does not reach the level of the anterior end proximally,
which is reminiscent of Glossotherium robustum (Owen 1842)
and Scelidotherium leptocephalum (Mi~no-Boilini et al. 2014) and
differs from those of Catonyx tarijensis (McDonald 1987), Cato-
nyx cuvieri, Valgipes bucklandi (Cartelle et al. 2009) and Paramy-
lodon harlani (Stock 1925). The most reminiscent olecranon
process is found in Scelidotherium leptocephalum, which also fea-
tures a characteristic pointed proximal end.
The anconeal process is very slightly broken, but is obviously
weakly developed (Fig. 6C), which differs from Glossotherium
robustum, Catonyx cuvieri and Valgipes bucklandi (Cartelle et al.
2009), and is reminiscent of Scelidotherium leptocephalum,
Paramylodon harlani and Catonyx tarijensis.
The radial facet is restricted to a small area continuous with
the most lateral part of the humeral facet (Fig. 6A).
The ulna is more elongate (depth at coronoid process/proxi-
modistal length = 0.33) when compared to Glossotherium robus-
tum (0.45; Owen 1842) and Paramylodon harlani (0.41; Stock
1925), and slightly stouter than Valgipes bucklandi (0.26; MCL
22464) and Pseudoprepotherium confusum (0.26; Hirschfeld
1985), its proportions being more reminiscent of those of Sceli-
dotherium leptocephalum and Scelidodon chiliense (both 0.32;
MMP 549S and MUT 1510 respectively; McDonald 1987).
A generic attribution is precluded by the paucity of postcra-
nial data for Neogene taxa (Mi~no-Boilini et al. 2011).
MEGATHERIOIDEA Gray, 1821
Hyperleptus? Ameghino, 1891
Figure 7
Referred material. MUN STRI 37413, locality 130024, Castilletes
Formation.
Description. Bone fragment with three poorly preserved molari-
forms (M2 MD length = c. 6 mm; M2 LL width = c. 10 mm;
M3 MD length = 6.8 mm; M3 LL width = c. 8 mm; M4 MD
length = 6.8 mm; M4 LL width = 6.2 mm). The different types
of dental tissues are not discernable. There is no diastema
(Fig. 7A). On the lingual side of the middle and mesial teeth,
F IG . 6 . Scelidotheriinae gen. et sp. indet. from the Ware For-
mation, left ulna (MUN STRI 16535). A, anterior view (proxi-
mal towards top). B, posterior view (proximal towards top). C,
lateral view (proximal towards top). Scale bar represents 10 cm.
Colour online.
AMSON ET AL . : NEOGENE SLOTHS FROM THE COCINETAS BAS IN 7

the bone projects medially. Attributable to a small portion of
the posterior end of the palatine, this indicates that the specimen
represents upper teeth and maxilla. The occlusal surface of the
teeth does not reach the most ventral level of the bone, possibly
suggesting a very immature individual or a very abrasive diet
(Fig. 7B). Furthermore, each tooth is surrounded by a large
space, which would tend to corroborate the first hypothesis. The
M2 is poorly preserved. Its outline and that of M3, although not
entirely preserved, are clearly compressed mesiodistally. The M3
preserves mesial and distal ridges, a synapomorphy of megatheri-
oids (Gaudin 2004). Conversely, the outline of M4 (the only one
entirely preserved) is subcircular, a singular trait in sloths. The
mesial half of its occlusal surface is covered by a labionlingual
ridge. Although mostly broken, there was most likely a lower dis-
tal ridge as well. Hyperleptus garzonianus is characterized by a
rounded M4 and mesiodistally compressed M1–3 (Ameghino
1891). However in Hyperleptus garzonianus the M4 is not as
circular as in MUN STRI 37413, being slightly wider labiolin-
gually than long mesiodistally (see measurements of Scott
(1903–1904)).
Remarks. A sub-circular outline of M4 seems to be a
derived trait found of Hyperleptus garzonianus. As the M4
of MUN STRI 37413 also departs from the plesiomorphic
strongly elliptical condition of megatherioids, a close
affinity with the latter species is likely. But the fragmen-
tary nature of MUN STRI 37413 and the fact that its M4
seems even more circular than that of Hyperleptus garzo-
nianus preclude a definitive attribution to this species.
Hyperleptus is known from the Santa Cruz Formation
(Scott 1903–1904), and is one of a number of small
megatherioids of uncertain affinity, which also includes
Pelecyodon, Schismotherium, Analcimorphus and Hapalops
(Gaudin 2004).
MEGALONYCHIDAE Gervais, 1855
Gen. et sp. nov.
Figure 8
Referred material. MUN STRI 16601, locality 390077 and MUN
STRI 34226, locality 470060, both Ware Formation.
Description. MUN STRI 16601 consists of a left patella repre-
senting a relatively large-sized taxon (Fig. 8D–E; maximum PD
length = 112.3 mm; maximum ML width = 92.7 mm; AP depth
at centre of articular facet = 32.2 mm; apex PD length (taken
from distal border of articular facet) = 47.8 mm; facet ML
width = 86.4 mm). It is slightly smaller than the specimen of
Megalonyx jeffersonii measured by Leidy (1855), but much larger
than Pliomorphus mutilatus, as shown by the width of the patel-
lar trochlea of the femur (Brandoni 2009). The tapered apex
gives the patella a teardrop shape in anterior view (Fig. 8D), as
commonly seen in sloths. The articular facet is only very weakly
convex mediolaterally (Fig. 8E), precluding the attribution to a
pedolateral sloth (Mylodontidae or Megatheria) or a glyptodont.
Moreover, the lateral half of the facet is shorter proximodistally
than the medial half, as in Megalonyx jeffersonii (Leidy 1855).
However, in anterior view, the general shape differs from that of
Megalonyx jeffersonii, in that both its proximal border and apex
are more tapered.
MUN STRI 34226 consists of an ulna preserving the proximal
epiphysis and part of the diaphysis (Fig. 8A–C; AP depth at
coronoid process = 83.0 mm; PD length from proximal end to
level of coronoid process = 62.2 mm). The specimen is from an
F IG . 7 . Megatherioidea Hyperleptus? (MUN STRI 37413) from
the Castilletes Formation. A, photograph and drawing in occlu-
sal view (labial towards top). B, lingual view (occlusal towards
top). Scale bar represents 2 cm. Colour online.
8 PALAEONTOLOGY

adult, as shown by the epiphyseal closure. The most distal part
of the preserved diaphysis is anteroposteriorly deep, suggesting
that roughly at least a fifth of the diaphysis is missing.
The olecranon process is directed posteroproximally, forming
an angle of roughly 45° with the proximodistal axis of the bone
(Fig. 8A–C), as in Megalonyx jeffersonii (see for instance Leidy
1855, pls IX, X). However, it differs from the latter genus by lack-
ing the strong medial inflection that affects the whole tuberosity
(Fig. 8A). Indeed, the olecranon process only slightly protrudes
medially in MUN STRI 34226. The presence of a small tuberosity
on the medial side of the tip of the olecranon process is reminis-
cent of Megalonyx jeffersonii, and Nothrotheriops shastensis as well.
The articular surface for the humeral trochlea is not entirely
preserved, but one can observe that it is weakly expanded laterally
and posteriorly (Fig. 8A, C), a characteristic feature of Megalonyx
jeffersonii, and differing from Nothrotheriops shastensis for
instance. As in both latter genera, the anconeal process is weakly
developed anteriorly. It appears even narrower mediolaterally than
in these two genera. Although fragmentary, it can be discerned
that the radial facet is continuous with the humeral facet.
Intraspecific variation regarding this trait is described in Mega-
lonyx jeffersonii (McDonald 1977). A large rounded pit is present
distal to the radial facet. A strong crest, issued from the process
for the radial notch, joins rapidly the anterior edge of the shaft.
The shaft is slightly bent medially (Fig. 8A, B). Its lateral side
bears a strong muscular crest issued from the posterior end and
directed anterodistally.
Remarks. MUN STRI 16601 and MUN STRI 34226 were
found isolated and each can be ascribed to an undescribed
Megalonychidae. We conservatively ascribe both specimens
to the same Megalonychidae gen. et sp. nov. in order to
avoid overestimating the taxonomic diversity of the Ware
Formation.
MEGATHERIA sensu Gaudin (2004)
MEGATHERIIDAE Gray, 1821
MEGATHERIINAE Gray, 1821
Pliomegatherium lelongi Kraglievich, 1930
Figure 9
Referred material. MUN STRI 36685, locality 390024, and tenta-
tively MUN STRI 19747, locality 390017 and MUN STRI 20446,
locality 390026, all Ware Formation.
Description. MUN STRI 36685 consists of a partial mandible
with c1 (mesialmost tooth, molarized in megatheriines),
eroded below the level of the alveolar plane (Fig. 9A–B; maxi-
mum MD length of c1 = 32.3 mm; maximum LL width of
c1 = c. 36 mm; LL width at distal end = 33.8 mm; maximum
MD length of alveolus of c1 = 37.4 mm). It is an adult
F IG . 8 . Megalonychidae gen. et sp. nov. from the Ware Forma-
tion. A–C, left ulna (MUN STRI 34226); A, anterior view (proximal
towards top); B, posterior view (proximal towards top); C, lateral
view (proximal towards top). D–E, left patella (MUN STRI 16601);
D, anterior view (proximal towards top); E, posterior view (proxi-
mal towards top). Scale bar represents 10 cm. Colour online.
AMSON ET AL . : NEOGENE SLOTHS FROM THE COCINETAS BAS IN 9

F IG . 9 . Pliomegatherium lelongi from the Ware Formation. A–B, partial mandible with c1 (MUN STRI 36685); A, photograph and
drawing in occlusal view (anterior towards top); B, right lateral view (occlusal towards top). C, partial caudal vertebra (MUN STRI
19747) in dorsal view (cranial towards top). Scale bar represents 5 cm. Colour online.
10 PALAEONTOLOGY

individual, as indicated by the thorough closure of the symph-
ysis, and the very short distance between the labial side of c1
alveolus and lateral side of horizontal ramus (De Iuliis 1996).
Based on the tooth dimensions, it is a medium-sized megath-
eriine, similar to the holotype of Pliomegatherium lelongi
(Brandoni 2006), and smaller than Pyramiodontherium brevi-
rostrum (Carlini et al. 2002; and, by extension, also smaller
than Megatheriops rectidens, see Pujos et al. 2013). The cross-
section of c1 is trapezoidal, with the mesial side slightly nar-
rower labiolingually than the distal side (Fig. 9A). This is
characteristic of Pliomegatherium lelongi (Brandoni 2006), and
differs from the mesiolabially constricted teeth of early
megatheriines such as Megathericulus, and square cross-section
of the teeth of Megatherium americanum, Eremotherium lauril-
lardi and Eremotherium eomigrans. The dorsal edge of the pre-
served portion of the spout is not preserved (Fig. 9B), which
prevents us from precisely assessing the relative development of
the ventral bulge of the dentary (diagnostic within the Megatheri-
inae subfamily; De Iuliis 1996). The posterior end of the symph-
ysis is preserved (Fig. 9A). It forms a partially closed notch
(c. 80°). This is reminiscent of Pliomegatherium lelongi (Brandoni
2006) and Eremotherium laurillardi, and differs from the more
open symphysis of Megathericulus patagonicus, and more closed
one of Megatherium americanum. It is located approximately at
the level of the mid-mesiodistal length of c1, roughly as in Ere-
motherium laurillardi, Pliomegatherium lelongi and Pyramiodon-
therium bergi (Brandoni 2006), but differs from Megathericulus
patagonicus, Eomegatherium andinum, and Anisodontherium
halmyronomumin, in which it is more anterior (Pujos et al. 2013;
Tejada-Lara et al. 2015). In adult Megatherium americanum, it is
more posterior (De Iuliis 1996).
MUN STRI 19747 consists of a centrum with a transverse
process from a caudal vertebra (Fig. 9C; CC length of cen-
trum = 59.8 mm; DV height of centrum at cranial side = 57.3
mm; ML of centrum at caudal side = 77.6 mm). The dimen-
sions would fit a medium-sized Megatheriinae, such as the sev-
enth caudal vertebra of Megatherium urbinai. The cross-sectional
shape of the centrum is slightly flattened dorsoventrally, indicat-
ing proximity to the cranial region of the series. The cranial and
caudal facets of the centrum are only slightly oblique. The trans-
verse process is oriented laterocaudally, forming an angle of
roughly 40° with the sagittal plane. It ends with a globular
tuberosity that is located caudal to the level of the centrum.
Moreover, the body of the transverse process is triangular in
cross-section. Such a condition is found in Megatherium (Owen
1861; Pujos & Salas 2004). In Megalonyx, the transverse pro-
cesses are also strongly oriented laterocaudally, but they are
more dorsoventrally flattened. In glyptodonts, the transverse
processes are either oriented only laterally or laterocranially (e.g.
Glyptotherium texanum, Gillette & Ray 1981).
MUN STRI 20446 consists of a centrum of caudal vertebra
(CC length of centrum = 57.9 mm; DV height of centrum at
cranial side = 56.2 mm; ML of centrum at caudal side =c. 71 mm; maximum ML width (one side doubled) = 170 mm).
The cross-section of the centrum is slightly flattened dorsoven-
trally, indicating proximity to the cranial region of the series.
The cranial and caudal facets of the centrum are only slightly
oblique to each other (i.e. the cranial is set slightly dorsal to the
caudal). The four hypophyseal processes and their facets for the
haemal arches are well developed.
Remarks. Pliomegatherium lelongi is otherwise recorded in
the ‘conglomerado os�ıfero’, Ituzaing�o Formation, Entre
R�ıos Province, Argentina (upper Miocene – Pliocene; Bran-doni 2006). Pliomegatherium sp. is also reported in the
‘Barrancas de San Gregorio’, San Jos�e member of Raig�on
Formation (Perea et al. 2013), San Jos�e Department, Uru-
guay (Mones 1988). These localities are at a latitude of
c. 32–34°S, and are c. 5000–5500 km from the localities of
the Ware Formation. The geographical range of P. lelongi is
hence drastically expanded, with specimens being found
across the northern two-thirds of South America. The
broad geographical distribution seems to be common in
the subfamily, as exemplified by the Panamerican Eremoth-
erium laurillardi (Cartelle & De Iuliis 1995). Based on the
dimensions of the c1 of MUN STRI 36685, the body mass
of this individual is estimated to be between c. 1000 and
4400 kg, with a mean value of 2400 kg (Table 1). This is
consistent with the 2500 kg estimate given for another
medium-sized megatheriine, Pyramiodontherium scillatoy-
anei (De Iuliis et al. 2004).
MUN STRI 19747 and MUN STRI 20446 can only be
safely ascribed to Megatheriinae indet. and Xenarthra
indet., respectively. But, given the similar morphology
and dimensions of the centra, and the fact that no fea-
tures appear to be incompatible, we tentatively ascribe
them to the same taxon as MUN STRI 36685.
TABLE 1 . Body size estimations for the two size extremes of the tardigradan assemblage of the Ware Formation.
Measurements (cm/cm2) Prediction (kg)
MUN STRI 36685
c1 length (FLML) log mass = log FLML * 3.263 + 1.337 3.23 996
c1 width (FLMW) log mass = log FLMW * 2.909 + 2.03 3.60 4449
c1 area (FLMA) log mass = log FLMA * 1.553 + 1.701 10.042 1806
Mean of predictions 2417
MUN STRI 12924
Occipital height (OCH) log mass = log OCH * 2.783 � 0.42 5.37 41
The equations are based on Janis (1990) (in Fari~na et al. 1998). First lower molar (FLM) corresponds to c1 in Pliomegatherium lelongi
(MUN STRI 36685).
AMSON ET AL . : NEOGENE SLOTHS FROM THE COCINETAS BAS IN 11

NOTHROTHERIIDAE Ameghino, 1920
NOTHROTHERIINAE Ameghino, 1920
cf. Nothrotherium Lydekker, 1889
Figures 10, 11
Referred material. MUN STRI 12924, locality 390024, Ware For-
mation.
Description. MUN STRI 12924 consists of a fragmentary basicra-
nium, with the posterior portion of the skull roof (Fig. 10; see
3D model in Amson et al. (2016b); ML width at postorbital con-
striction = 54.5 mm; ML width at mastoid processes = 82.0 mm;
ML width at root of zygomatic process of squamosal = 95.1 mm;
maximum ML width of occipital condyles = 56.5 mm; occipital
height = 53.7 mm; maximum ML width of foramen mag-
num = 30.0 mm; maximum DV height of foramen magnum =c. 28 mm). Two foramina are present at the posterior edge of the
basioccipital, which indicate the incomplete ossification and hence
immaturity of the specimen. Moreover, some of the sutures appear
thoroughly closed (e.g. interfrontals, interparietals), while others are
not (e.g. supraoccipital-exoccipital). A subadult stage is hence
F IG . 10 . Cf. Nothrotherium from the Ware Formation, partial basicranium (MUN STRI 12924), general views (photograph and
drawing). A, dorsal view (right lateral towards top). B, left lateral view (dorsal towards top). C, occipital view (dorsal towards top).
Abbreviations: ape., anterior process of entotympanic; coa., canal for the occipital artery; eoc., external occipital crest; exc., exoccipital
crest; f., foramen; frag., fragment; nc., nuchal crest; oc., occipital condyle; Occ., occipital bone; p., process; sf., stylomastoid foramen.
Scale bars represent 5 cm. Colour online.
12 PALAEONTOLOGY

inferred for this specimen. The size of the specimen falls within
the range of the adult specimens of Nothrotherium maquinense
measured by Cartelle & Fonseca (1983), and is slightly larger than
Mionothropus cartellei (De Iuliis et al. 2011) and Pronothrotherium
typicum (Patterson et al. 1992).
Only a dorsal portion of the posterior half of the frontals is pre-
served (Fig. 10A, B). There is no sagittal crest (diagnostic of the
Nothrotheriidae; De Iuliis et al. 2011), but this could be due to
the immaturity of the specimen. The frontoparietal suture is at the
level of the root of the zygomatic process of the squamosal. Part of
the alisphenoid is present on both sides. It contacts the parietal
(diagnostic of the Nothrotheriinae except Mionothropus cartellei,
De Iuliis et al. 2011). The temporal line joins the dorsal edge of
the zygomatic process of the squamosal posteriorly, as in
Mionothropus cartellei, Nothrotherium maquinense and Nothrothe-
riops shastensis, but not in Pronothrotherium typicum (De Iuliis
et al. 2011). The parietals are globose and posteriorly depressed
(diagnostic of Nothrotherium maquinense, Paula Couto 1971).
The root of the zygoma is directed anteriorly (diagnostic of
the Nothrotheriinae; De Iuliis et al. 2011). Lateral to it, there is
a bulge (diagnostic of the Nothrotheriinae except Mionothropus
cartellei, De Iuliis et al. 2011), although it does not appear as
developed as in Pronothrotherium typicum, Nothrotherium
maquinense and Nothrotheriops shastensis.
Anterior to the carotid foramen, and medial to the Glaserian
fissure, the bone has an irregular surface and displays a broken
ridge (Fig. 11). Medial to it, and just anterior to the basioccipi-
tal tuber, the basisphenoid shows a rugose surface. This region
most likely represents the location of the posterior part of the
ossified pterygoid bulla, which presumably would not have been
thoroughly sutured yet to the basiphenoid in this immature
specimen. An ossified pterygoid bulla is present in Nothrother-
ium maquinense and Nothrotheriops shastensis.
The ectotympanic is missing on both sides, except maybe for
a small portion of the anterior crus on the left side. The ento-
tympanic is well preserved on both sides. Anterolateral and pos-
teromedial portions can be distinguished, as described in
Pronothrotherium typicum (Patterson et al. 1992) and Mionothro-
pus cartellei (De Iuliis et al. 2011). The posteromedial portion is
much smaller than the anterolateral one, differing in that regard
from Pronothrotherium typicum (Patterson et al. 1992). A similar
disposition is found in Nothrotherium maquinense (MCL 21703),
but in this species the posteromedial portion seems more dorsally
elevated. In MUN STRI 12924, the anterolateral portion forms a
A B C
F IG . 11 . Cf. Nothrotherium from the Ware Formation, partial basicranium (MUN STRI 12924), close-up of the left auditory region
in ventral view (anterior towards top). A–B, left and right stereophotographs. C, same view as B but labelled. Abbreviations: ant. cr. fr.,
anterior crus fragment; ant. p. ent., anterior process of entotympanic; Bo., basioccipital; Bs., basisphenoid; car. f. + mlf., carotid fora-
men and medial lacerate foramen; ent. p. ali., entoglenoid process of alisphenoid; exo. cr., exoccipital crest; f., foramen; gl., Glaserian
fissure; mast. p., mastoid process; pet. fr., petrosal fragment; p., process; post. p. ent., posterior process of entotympanic; th., tympa-
nohyoid; sm., superficies meatus; ssp., scar of suture for pterygoid bulla; sth. fos., stylohyal fossa; stylo. f., stylomastoid foramen; su.
int. car. art., sulcus for internal carotid artery. Scale bar represents 2 cm. Colour online.
AMSON ET AL . : NEOGENE SLOTHS FROM THE COCINETAS BAS IN 13

ventrally expanded crest-like process. It is sigmoidal in ventral
view. This process appears to be particularly vertical, not being
inclined medially. The posterior end of the entotympanic forms
the medial part of the stylohyoid fossa. The junction between the
posteromedial and anterolateral portions of the entotympanic is
marked by a shallow sulcus, presumably accommodating, as in
Mionothropus cartellei, the internal carotid artery. The jugular
foramen (or posterior lacerate foramen) is large, extending poste-
riorly up to the level of the hypoglossal foramen (diagnostic of
the Nothrotheriidae; De Iuliis et al. 2011). Both of these foram-
ina somewhat share the same excavation and are located espe-
cially close to one another, which is very reminiscent of
Nothrotherium maquinense and Nothrotheriops shastensis, and dif-
fers from the more separate foramina of Mionothropus cartellei
and Pronothrotherium typicum. Posteromedial to these foramina,
there is a globular paracondylar process, very reminiscent of
Nothrotherium maquinense, and differing from the more elongate
process of Nothrotheriops shastensis. The stylomastoid foramen is
found at the dorsal end of a sulcus formed laterally by the mas-
toid process and medially by a well-developed ridge extending
from the nuchal crest (possibly the exoccipital crest sensu Gaudin
1995). This sulcus, terminating dorsally by a foramen, may repre-
sent a widely exposed canal for the occipital artery, a synapomor-
phy of the Nothrotheriidae (Gaudin 1995). A groove also leading
dorsally to the mastoid foramen is described in Acratocnus odon-
trigonus and Schismotherium fractum (Patterson et al. 1992).
Most of the preserved articular surface of the stylohyoid fossa is
formed by the posterodorsal side of the entotympanic. The rest
of the preserved articular surface is a narrow strip, presumably
formed by the tympanohyoid, as in Mionothropus cartellei for
instance (De Iuliis et al. 2011).
The occipital condyles are sessile (Fig. 10C; diagnostic of the
Nothrotheriidae; De Iuliis et al. 2011). They have a peculiar (ab-
normal?) morphology, being marked at mid-height by a medio-
laterally oriented depression. There is a distinct external occipital
crest, developed to an extent very similar to that of a juvenile
specimen of Nothrotherium maquinense (MCL 1020). The nuchal
crest, however, is inconspicuous.
Remarks. Nothrotherium is represented by the Pleistocene
N. maquinense and N. escrivanense, which are restricted to
Brazilian localities. While we provisionally ascribe MUN
STRI 12924 to cf. Nothrotherium due to the fragmentary
nature and immaturity of the specimen, it is likely to rep-
resent a closely related new taxon. Based on the occipital
height of this specimen, the body mass of the individual is
estimated to be c. 41 kg (Table 1). We estimate this speci-
men to be subadult, and consider it unlikely that this indi-
vidual would have reached 100 kg at maturity.
DISCUSSION
Mammalian assemblage of the Castilletes Formation
Although not the main focus of the present contribution,
the Castilletes Formation is discussed here because of an
important modification to its faunal composition. The
specimen previously referred to Megatheriidae (MUN
STRI 16777; Moreno et al. 2015), the only sloth formerly
recognized in the formation, is actually an astrapothere.
However, the Castilletes Formation does contain a
tardigrade, as MUN STRI 37413 represents a small
megatherioid. Tentatively referred to Hyperleptus?, this
record could be consistent with the geological age of the
Formation as it is otherwise known from the Santacrucian
fauna. A sparassodont from the Castilletes formation,
Lycopsis padillai (Su�arez et al. 2015), also suggests affini-
ties with the Santacrucian fauna. On the other hand,
crocodilian taxa of the Castilletes Formation are otherwise
known from Laventan and Huayquerian faunas (Moreno-
Bernal et al. 2016). This emphasizes the importance of
the detailed description of the rest of the mammalian
fauna from the Castilletes Formation.
Diversity of the tardigradan assemblage of the Ware
Formation
A greater taxonomic diversity in the lower latitudes is a
global pattern broadly found today among eukaryotes
(Hillebrand 2004), and it is the result of a long and com-
plex history, as shown in particular for the Neotropics
(Jaramillo et al. 2006). Recent studies have shown how
Neotropical Miocene formations can preserve hyper-
diverse vertebrate assemblages (for crocodilians, see
Scheyer et al. 2013; Salas-Gismondi et al. 2015). However,
taxonomic richness of fossil tropical assemblages is still
severely underestimated (Carrillo et al. 2015). The Ware
Formation gives an important insight into the otherwise
poorly documented diversity at low latitudes during the
Pliocene. The conservative diversity count for the tardi-
gradan assemblage of the Ware Formation is of five
distinct taxa. Each of the four families of ‘ground sloths’
(Mylodontidae, Megatheriidae, Nothrotheriidae and
Megalonychidae) is represented. When compared with
assemblages from other Pliocene Neotropical formations
(Table 2), the Ware assemblage has a rather diverse tardi-
gradan composition. The San Gregorio Formation is a
geographically nearby formation that records a roughly
synchronous continental fauna, and yet only one sloth
has been reported (Vucetich et al. 2010), most likely due
to taphonomic and/or sampling biases. When compared
to older formations, the Ware Formation’s tardigradan
diversity is equalled by the Urumaco Formation (late
Miocene; where the taxa might not have been coeval,
Carlini et al. 2006a) and surpassed by the better-known
Laventan fauna. An example of an extensively sampled
Pliocene formation that is found at higher latitudes is the
Monte Hermoso Formation. Although it records five
species in three families, only two to three species are
14 PALAEONTOLOGY

found in the same facies (Tomassini et al. 2013), making
it roughly half as diverse as the Ware Formation. When
compared to other temperate Pliocene assemblages
(Table 2), a similar conclusion can be drawn, except for
the Uqu�ıa Formation. The astonishing 22 species of sloth
of the Late Miocene Ituzaing�o Formation (‘Conglomerado
os�ıfero, mesopotamian’) has to be taken with caution, as
a recent and unfinished taxonomic revision (every family
except the Mylodontidae) has revealed a lower diversity
than previously reported (Brandoni 2013).
Another aspect of the diversity of the sloths of the
Ware Formation dwells in their ecologies. With a body
mass estimated to be c. 2400 kg, Pliomegatherium lelongi
is a megaherbivore sensu Owen-Smith (1987). While the
closely related Megatherium americanum is reconstructed
as a selective feeder (Bargo & Vizca�ıno 2008), broad geo-
graphical distribution is common in the subfamily, which
is usually considered to be indicative of generalist habits.
In contrast, the specimen ascribed to cf. Nothrotherium is
a small-sized sloth weighing roughly 41 kg. Although not
much can be argued about the ecology of the latter, the
range of two orders of magnitude in body size featured
by this tardigradan assemblage suggests that disparate eco-
logical niches were occupied. In addition to intermediate
body size, the other sloths recovered feature other feeding
ecologies. Based on the morphology of its close relatives,
Lestodon and Thinobadistes, Lestodontini gen. et sp nov.
was most likely wide-muzzled. This is argued to correlate
with another feeding ecology, namely bulk-feeding (Bargo
& Vizca�ıno 2008; Czerwonogora et al. 2011). Furthermore,
Scelidotheriinae gen. et sp. indet. was most likely as nar-
row-muzzled as Scelidotherium leptocephalum, which has
been interpreted as a mixed or selective-feeder with possi-
ble uprooting habits (Bargo & Vizca�ıno 2008). Similarly,
niche partitioning was previously argued to explain the
high diversity of Neotropical crocodilian communities
(Scheyer et al. 2013; Salas-Gismondi et al. 2015).
Ware Formation and GABI
Sloths are main elements in the American Biotic Inter-
change, as they are present in North America as early as
c. 9 Ma (Hemphillian; Woodburne 2010), and as they
take part in the main pulses of GABI migration. Addi-
tionally, it was recently emphasized how the Ware Forma-
tion can enlighten our understanding of the GABI, with
the example of the carnivoran procyonid Chapalmalania
sp., found in Ware, which is suggestive that procyonids,
North American immigrants, dispersed into South Amer-
ica in two separate events (Forasiepi et al. 2014).
All four families of ‘ground sloths’ that participated in
GABI (Mylodontidae, Megatheriidae, Nothrotheriidae,
Megalonychidae) were present in the Ware formation. A
taxon closely related to Ware’s Pliomegatherium lelongi
(MUN STRI 36685), the megatheriine Eremotherium, is
found in North America c. 2.6 Ma (Woodburne, 2010).
This taxon is widespread in both South and North Amer-
ica while its sister taxa, Megatherium, is restricted to South
America. The relationships of P. lelongi to both Mega-
therium and Eremotherium are unresolved, and therefore,
it is not possible to assess whether the P. lelongi clade
participated in GABI. Megatheriinae are also represented
in northern South America by Proeremotherium and
Urumaquia (Carlini et al. 2006b, 2008), but the recovered
elements of those taxa do not include a mandible or lower
teeth that would have allowed a comparison with MUN
STRI 36685.
TABLE 2 . Comparison of the sloth diversity in Neotropical and Temperate Pliocene formations.
Epoch Formation Latitude (°S) Environment Taxonomic
diversity Species
(families)
References
Neotropical formations
Late Pliocene Ware 11.8 Fluvio-deltaic 5 (1) Moreno et al. 2015;
present study
San Gregorio 11.3 Alluvial fans 1 (1) Vucetich et al. 2010
Early Pliocene Codore 11.3 Fluvio-deltaic with
marine inundation
3 (3) Carlini et al. 2006a, b
Temperate formations
Pliocene Uqu�ıa �23 to �24 Fluvial 5 (3)* Reguero et al. 2007
Corral Quemado �27 – 1 (1) Reguero et al. 2007
Inchasi �19.7 Fluvial 3 (2) Anaya & MacFadden 1995
Monte Hermoso �39 Fluvial 5 (3)* Tomassini et al. 2013
Montehermosan/
Chapadmalalan
Andalhual�a �27 – 3 (2) Reguero & Candela 2011
*Taxa most likely not coeval.
AMSON ET AL . : NEOGENE SLOTHS FROM THE COCINETAS BAS IN 15

Nothrotheriops is recorded in North America during the
Pleistocene (c. 1.4 Ma, Woodburne 2010). However, coe-
val Nothrotherium species, which share diagnostic features
with the specimen from Ware, are restricted to Brazil.
Therefore, it seems that Ware’s Nothrotherium is a mem-
ber of the South American Nothrotherium that did not
participate in GABI.
Two other sloths of the Formation, Megalonychidae gen.
et sp. nov. and Lestodontini gen. et sp. nov. (Mylodonti-
dae), have ambiguous affinities. Megalonychidae represents
one of the earliest ‘ground sloth’ families to migrate to
North America (c. 9 Ma) and includes the early immi-
grants Pliometanastes, and subsequently Megalonyx and
Meizonyx (Woodburne 2010). The Megalonychidae gen. et
sp. nov. of Ware is reminiscent of Megalonyx (and by
extension Pliometanastes, of which the postcranium is
hardly distinguishable; McDonald & Naples 2007), but also
features clear differences. A comparison with Meizonyx is
impossible due to the incompatibility of the elements
recovered. At this point, it is not possible to assess if the
Megalonychidae gen. et sp. nov. of Ware is more closely
related to North American or South American megalony-
chids, as South American large-sized megalonychids are
still poorly documented (e.g. Megalonychops primigenius,
known by a fragmentary humerus from the Ituzaing�o For-
mation, late Miocene of Argentina; Brandoni 2013).
Lestodontini gen. et sp. nov. belongs to Mylodontinae,
a subfamily of Mylodontidae that includes Thinobadistes,
one of the earliest migrants to North America c. 9 Ma
(Woodburne 2010). The subfamily continued taking
part in GABI with the late Pliocene – late Pleistocene
Paramylodon harlani (and a specimen of ‘Glossotherium
chapadmalense’; McAfee 2009). Unfortunately, Ware’s
Lestodontini gen. et sp. nov. is not complete enough to
provide synapomorphies that could have discriminated
between South and North American affinities. As empha-
sized by the phylogenetic analysis (see above), Lestodon-
tini gen. et sp. nov. is found within a clade with
unresolved relationships with both the early immigrant
Thinobadistes and Lestodon, known from the Pleistocene
of temperate South American. Lestodontinae is also rep-
resented in northern South America by Bolivartherium
(Urumaco and Codore formations; Carlini et al. 2006a).
A subfamily of Mylodontidae, the Scelidotheriinae, did
not take part in GABI (Mi~no-Boilini et al. 2014) but has
been found in Ware. This subfamily had a wide distribu-
tion during the Pliocene and Pleistocene being recorded in
multiple localities (from �20 to �39°, Paleobiology Data-
base and the related references included therein: Rusconi
1954; MacFadden et al. 1993; Rodr�ıguez-Brizuela & Tauber
2006; Reguero & Candela 2011). The non-migrating sceli-
dotheriine contrasts with other taxa in Ware, whose rela-
tives took part in GABI (Woodburne 2010). The record of
Ware, in the vicinity of the Isthmus of Panama, further
raises the question of why Scelidotheriinae was not
involved in GABI. The specialized niche possibly associated
with derived muzzle shape is most likely a contributing fac-
tor (McDonald 2005). Scelidotheriines are indeed charac-
terized by long and narrow premaxillae, also acquired,
although under other modalities, in non-migrant members
of other subfamilies such as Megatherium and Mylodon.
CONCLUSIONS
The Cocinetas Basin comprises two units that record con-
tinental Neogene biotas, the Castilletes and Ware forma-
tions. Both delivered rich mammalian assemblages
including tardigradan taxa. Sloths are represented in the
early–middle Miocene Castilletes Formation by one frag-
mentary specimen. Referred to a small-sized megatheri-
oid, this record could suggest affinities with the distant
Santacrucian fauna.
The sloths from the late Pliocene Ware Formation are
represented by 12 specimens that belong to 5 taxa from
the 4 families of ‘ground sloths’. Their body sizes span
over two orders of magnitude, and they most likely
featured various feeding strategies, documenting a highly
diverse assemblage of sloths from the Neotropics.
Although geographically close to the Isthmus of Panama,
and temporally preceding GABI 1 by only 0.2–0.4 my, the
sloths for which unambiguous affinities were recovered
are not closely related to the early immigrants found in
North America before the first main pulse of the Great
American Biotic interchange.
Acknowledgements. Our gratitude goes to all the other members
of the field campaigns, and in particular to J. W. Moreno-Bernal
(STRI), who found several of the specimens presented here. The
constant help of L. Londo~no (STRI) is also acknowledged. We
thank C. Montes (Universidad de los Andes) and J. Escobar
(Universidad del Norte) for welcoming EA to the collections
under their care. M. R. S�anchez-Villagra is greatly acknowledged
for helping to conceive the study and improve the manuscript.
H. G. McDonald (Museum Management Program, National
Park Service) and A. A. Carlini (Museo de la Plata) are thanked
for their useful insights. B. Schubert (East Tennessee State
University) is acknowledged for providing pictures of Megalonyx’
caudal series. R. K. McAfee and an anonymous reviewer are
warmly thanked for the improvement they brought to the
manuscript. EA was granted a short-term postdoctoral fellowship
of the Smithsonian Tropical Research Institute, and was subse-
quently funded by the Alexander von Humboldt Foundation. JC
was supported by Swiss National Fund SNF 31003A-149605 to
M. R. S�anchez-Villagra. The Smithsonian Institution, the
National Geographic Society, the Anders Foundation, Gregory
D. and Jennifer Walston Johnson, Universidad del Norte, the
Lab of M. R. S�anchez-Villagra at the Pal€aontologisches Institut
und Museum der Universit€at Z€urich, and the National Science
Foundation (Grant EAR 0957679) also helped to support this
16 PALAEONTOLOGY

work. Thanks to Carlos Rosero for managing all the logistics in
the field. Thanks to the communities of Warpana, Patajau, Aule-
chit, Nazareth, Wososopo, Sillamana, Paraguachon, La Flor de la
Guajira, and Ipapura. Thanks to the Colombian National Police
(Castilletes base) and the Colombian Army (La Flor de la
Guajira and Cerro de la Teta). We express special thanks to our
drivers, Grillo, Lalo and Medardo.
DATA ARCHIVING STATEMENT
Data for this study are available in the Dryad Digital Repository:
http://dx.doi.org/10.5061/dryad.57025
Locality and stratigraphical data are available in the Smithsonian
Tropical Research Institute PaleoDatabase: http://biogeodb.stri.si.edu/
jaramillo/fossildb
3D models are available in MorphoMuseuM: http://dx.doi.org/
10.18563/m3.2.1.e3
Editor. Hannah O’Regan
REFERENCES
AMEGHINO, F. 1889. Contribuci�on al conocimiento de los
mam�ıferos f�osiles de la Rep�ublica Argentina. Actas de la Aca-
demia Nacional de Ciencias de C�ordoba, 6, 1–1027.-1891. Caracteres diagn�osticos de cincuenta especies nuevas
de mam�ıferos f�osiles argentinos. Revista Argentina de Historia
Natural, 1, 129–167.-1904. Nuevas especies de mam�ıferos cret�aceos y terciarios
de la Rep�ublica Argentina. Anales de la Sociedad Cient�ıfica
Argentina, 58, 225–291.-1920. Sur les �edent�es fossiles de l’Argentine. Examen cri-
tique, r�evision et correction de l’ouvrage de M. R. Lydekker
‘The extinct Edentates of Argentine’ etc. (Ouvrage �a demi-
in�edit avec des notes additionnelles sur quelques ongul�es et
carnassiers. Obras Completas y Correspondencia Cient�ıfica de
Florentino Ameghino, 11, 447–909.AMSON, E., ARGOT, C., McDONALD, H. G. and DE
MUIZON, C. 2015. Osteology and functional morphology
of the hind limb of the marine sloth Thalassocnus (Mammalia,
Tardigrada). Journal of Mammalian Evolution, 22, 355–419.-CARRILLO, J. D. and JARAMILLO, C. 2016a. Data
from: Neogene sloth assemblages (Mammalia, Pilosa) of the
Cocinetas Basin (La Guajira, Colombia): implications for the
Great American Biotic Interchange. Dryad Digital Repository.
doi: 10.5061/dryad.57025
---2016b. 3D models related to the publication:
Neogene sloth assemblages (Mammalia Pilosa) of the Cocine-
tas Basin (La Guajira, Colombia): implications for the Great
American Biotic Interchange. MorphoMuseuM, 2, e3. doi:
10.18563/m3.2.1.e3
ANAYA, F. and MACFADDEN, B. J. 1995. Pliocene mam-
mals from Inchasi, Bolivia: the endemic fauna just before the
great American interchange. Bulletin of the Florida Museum of
Natural History, 39, 87–140.BACON, C. D., S ILVESTRO, D., JARAMILLO, C.,
SMITH, B. T., CHAKRABARTY, P. and ANTONELLI ,
A. 2015. Biological evidence supports an early and complex
emergence of the Isthmus of Panama. Proceedings of the
National Academy of Sciences, 112, 6110–6115.-MOLNAR, P., ANTONELLI , A., CRAWFORD, A.
J., MONTES, C. and VALLEJO-PAREJA, M. C. 2016.
Quaternary glaciation and the Great American Biotic Inter-
change. Geology, 44, 375–378.BARGO, M. S. and VIZCA�INO, S. F. 2008. Paleobiology of
Pleistocene ground sloths (Xenarthra, Tardigrada): biome-
chanics, morphogeometry and ecomorphology applied to the
masticatory apparatus. Ameghiniana, 45, 175–196.BENGSTON, P. 1988. Open nomenclature. Palaeontology, 31,
223–227.BLOCH, J. I., WOODRUFF, E. D., WOOD, A. R., RIN-
CON, A. F., HARRINGTON, A. R., MORGAN, G. S.,
FOSTER, D. A., MONTES, C., JARAMILLO, C. A.,
JUD, N. A., JONES, D. S. and MACFADDEN, B. J.
2016. First North American fossil monkey and early Miocene
tropical biotic interchange. Nature, 533, 243–246.BRANDONI, D. 2006. A review of Pliomegatherium KRAGLIE-
VICH, 1930 (Xenarthra: Phyllophaga: Megatheriidae). Neues
Jahrbuch f€ur Geologie und Pal€aontologie Monatshefte, 4, 212–224.
-2009. Descripci�on del f�emur de Protomegalonyx chasicoensis
Scillato-Yan�e (Mammalia, Xenarthra, Megalonychidae) de la
Formaci�on Arroyo Chasico (Mioceno tard�ıo), provincia de
Buenos Aires, Argentina. Consideraciones sistem�aticas sobre
los perezosos. Ameghiniana, 46, 513–521.-2013. Los Tardigrada (Mammalia, Xenarthra) del Mioceno
tard�ıo de Entre R�ıos, Argentina. El Ne�ogeno de la Mesopotamia
argentina. Asociaci�on Paleontol�ogica Argentina, Publicaci�on
Especial, 14, 135–144.CARLINI , A. A., PASCUAL, R., REGUERO, M. A.,
SCILLATO-YAN�E, G. J., TONNI , E. P. and VIZ-
CA�INO, S. F. 1990. The first Paleogene land placental
mammal from Antarctica: its paleoclimatic and paleobrogeo-
graphical bearings. In COX, B. and REVEAL, J. (eds).
Abstracts IV International Congress of Systematic and evolution-
ary biology. University of Maryland, Baltimore, MD, 325 pp.
- BRANDONI, D., SCILLATO-YAN�E, G. J. and
PUJOS, F. 2002. Una nueva especie de megaterino (Xenar-
thra, Megatheriidae) del Mioceno Tard�ıo-Plioceno de Cata-
marca, Argentina. Ameghiniana, 39, 367–377.-SCILLATO-YAN�E, G. J. and S �ANCHEZ, R. 2006a.
New Mylodontoidea (Xenarthra, Phyllophaga) from the Mid-
dle Miocene–Pliocene of Venezuela. Journal of Systematic
Palaeontology, 4, 255–267.-BRANDONI, D. and S �ANCHEZ, R. 2006b. First
Megatheriines (Xenarthra, Phyllophaga, Megatheriidae) from
the Urumaco (Late Miocene) and Codore (Pliocene) Forma-
tions, Estado Falc�on, Venezuela. Journal of Systematic Palaeon-
tology, 4, 269–278.---2008. Additions to the knowledge of Urumaquia
robusta (Xenarthra Phyllophaga, Megatheriidae) from the Uru-
maco Formation (Late Miocene), Estado Falc�on, Venezuela.
Pal€aontologische Zeitschrift, 82, 153–162.CARRILLO, J. D., FORASIEPI , A., JARAMILLO, C. and
S �ANCHEZ-VILLAGRA, M. R. 2015. Neotropical mammal
diversity and the great American biotic interchange: spatial
AMSON ET AL . : NEOGENE SLOTHS FROM THE COCINETAS BAS IN 17

and temporal variation in South America’s fossil record. Fron-
tiers in Genetics, 5, 451.
CARTELLE, C. and FONSECA, J. S. 1983. Contribuic�~ao ao
melhor conhecimento da pequena preguic�a terr�ıcola
Nothrotherium maquinense (Lund) Lydekker, 1889. Lundiana,
2, 127–181.-and DE IULIIS , G. 1995. Eremotherium laurillardi: the
Panamerican Late Pleistocene megatheriid sloth. Journal of
Vertebrate Paleontology, 15, 830–841.--and FERREIRA R. L. 2009. Systematic revision of
tropical Brazilian scelidotheriine sloths (Xenarthra, Mylodon-
toidea). Journal of Vertebrate Paleontology, 29, 555–566.CIONE, A. L., GASPARINI , G. M., SOIBELZON, E.,
SOIBELZON, L. H. and TONNI , E. P. 2015. The Great
American biotic interchange. A South American perspective.
Springer, Dordrecht, 97 pp.
COPE, E. D. 1889. The Edentata of North America. The Ameri-
can Naturalist, 23, 657–664.CZERWONOGORA, A., FARI ~NA, R. A. and TONNI , E.
P. 2011. Diet and isotopes of Late Pleistocene ground sloths:
first results for Lestodon and Glossotherium (Xenarthra, Tardi-
grada). Neues Jahrbuch f€ur Geologie und Pal€aontologie –Abhandlungen, 262, 257–266.
DE IULIIS , G. 1996. A systematic review of the Megatheriinae
(Mammalia: Xenarthra: Megatheriidae). Unpublished PhD
thesis, University of Toronto, Toronto, 781 pp.
-R�E, G. H. and VIZCA�INO, S. F. 2004. The Toro Negro
megatheriine (Mammalia, Xenathra): a new species of Pyra-
miodontherium and a review of Plesiomegatherium. Journal of
Vertebrate Paleontology, 24, 214–227.-GAUDIN, T. J. and VICARS, M. J. 2011. A new genus
and species of nothrotheriid sloth (Xenarthra, Tardigrada,
Nothrotheriidae) from the Late Miocene (Huayquerian) of
Peru. Palaeontology, 54, 171–205.DELSUC, F., CATZEFLIS , F. M., STANHOPE, M. J. and
DOUZERY, E. J. 2001. The evolution of armadillos, anteaters
and sloths depicted by nuclear and mitochondrial phylogenies:
implications for the status of the enigmatic fossil Eurotamand-
ua. Proceedings of the Royal Society B, 268, 1605–1615.FARI ~NA, R. A., VIZCA�INO, S. F. and BARGO, M. S.
1998. Body mass estimations in Lujanian (late Pleistocene–early Holocene of South America) mammal megafauna. Mas-
tozoolog�ıa Neotropical, 5, 87–108.FORASIEPI , A. M., SOIBELZON, L. H., GOMEZ, C. S.,
S �ANCHEZ, R., QUIROZ, L. I., JARAMILLO, C. and
S �ANCHEZ-VILLAGRA, M. R. 2014. Carnivorans at the
Great American Biotic Interchange: new discoveries from the
northern neotropics. Naturwissenschaften, 101, 965–974.GAUDIN, T. J. 1995. The ear region of edentates and the phy-
logeny of the Tardigrada (Mammalia, Xenarthra). Journal of
Vertebrate Paleontology, 15, 672–705.-2004. Phylogenetic relationships among sloths (Mammalia,
Xenarthra, Tardigrada): the craniodental evidence. Zoological
Journal of the Linnean Society, 140, 255–305.GELFO, J. N., M €ORS, T., LORENTE, M., L �OPEZ, G. M.
and REGUERO, M. 2015. The oldest mammals from
Antarctica, early Eocene of the La Meseta Formation, Seymour
Island. Palaeontology, 58, 101–110.
GERVAIS , P. 1855. Recherches sur les mammif�eres fossiles de
l’Am�erique M�eridionale. Annales des Sciences Naturelle. Zoolo-
gie et Biologie Animale, 3, 330–338.-1873. M�emoires sur plusieurs esp�eces de mammif�eres fos-
siles propres d’Am�erique M�eridionale. M�emoires de la Soci�et�e
G�eologique de France, S�erie 2, 9, 1–44.GILL , T. 1872. Arrangement of the families of mammals with
analytical tables. Smithsonian Miscellaneous Collections, 11, 1–98.GILLETTE, D. D. and RAY, C. E. 1981. Glyptodonts of
North America. Smithsonian Contributions to Paleobiology, 40,
1–255.GRAY, J. E. 1821. On the natural arrangement of vertebrose
animals. London Medical Repository, 15, 296–310.HENDY, A. J. W., JONES , D. S., MORENO, F., ZAP-
ATA, V. and JARAMILLO, C. 2015. Neogene molluscs,
shallow marine paleoenvironments, and chronostratigraphy of
the Guajira Peninsula, Colombia. Swiss Journal of Palaeontol-
ogy, 134, 45–75.HILLEBRAND, H. 2004. On the generality of the latitudinal
diversity gradient. The American Naturalist, 163, 192–211.HIRSCHFELD, S. E. 1985. Ground sloths from the Friasian
La Venta fauna, with additions to the Pre-Friasian Coyaima
fauna of Colombia, South America. University of California
Publications in Geological Science, 128, 1–91.JARAMILLO, C., RUEDA, M. J. and MORA, G. 2006. Ceno-
zoic plant diversity in the Neotropics. Science, 311, 1893–1896.-MORENO, F., HENDY, A. J. W., S �ANCHEZ– VIL-
LAGRA, M. R. and MARTY, D. 2015. Preface: La Guajira,
Colombia: a new window into the Cenozoic neotropical biodi-
versity and the Great American Biotic Interchange. Swiss Jour-
nal of Palaeontology, 134, 1–4.KRAGLIEVICH, L. 1930. La Formaci�on Friaseana del R�ıo
Fr�ıas, R�ıo F�enix, Laguna Blanca, etc., y su fauna de mam�ıferos.
Physis. Revista de la Sociedad Argentina de Ciencias Naturales,
10, 127–161.LATHAM, J. and DAVIES, H. 1795. Faunula indica, id est
Catalogus Animalium Indiae orientalis. In FORSTER, J. R.
(ed.). Faunula Indica. Joanne Reinholdo, Gebauer, Halle, 38
pp.
LEIDY, J. 1855. A memoir on the extinct sloth tribe of North
America. Smithsonian Contributions to Knowledge, 7, 1–68.LINNAEUS, C. 1758. Systema Naturæ per Regna Tria Naturæ,
Secundum Classes, Ordines, Genera, Species, cum Characteribus,
differentiis, synonymis, locis. Vol 1: Regnum animale. 10th edn.
Laurentii Salvii, Stockholm, 823 pp.
LYDEKKER, R. 1889. Palaezoology Vertebrate. In NICHOL-
SON, H. A. and LYDEKKER, R. (eds). A manual of zoology
for the use of students, with a general introduction on the princi-
ples of zoology, Vol. 2, William Blackwood & Sons, Edinburgh,
1624 pp.
-1894. Contributions to a knowledge of the fossil vertebrates
of Argentina. Anales del Museo de la Plata, Paleontologia
Argentina, 3, 1–103.MACFADDEN, B. J., ANAYA, F. and ARGOLLO, J. 1993.
Magnetic polarity stratigraphy of Inchasi: a Pliocene mammal-
bearing locality from the Bolivian Andes deposited just before
the Great American Interchange. Earth & Planetary Science
Letters, 114, 229–241.
18 PALAEONTOLOGY

McAFEE, R. K. 2009. Reassessment of the cranial characters of
Glossotherium and Paramylodon (Mammalia: Xenarthra:
Mylodontidae). Zoological Journal of the Linnean Society, 155,
885–903.McDONALD, H. G. 1977. Description of the osteology of
the extinct gravigrade edentate Megalonyx with observations
on its ontogeny, phylogeny, and functional anatomy.
Unpublished MS thesis, University of Florida, Gainesville,
326 pp.
-1987. A systematic review of the Plio-Pleistocene sceli-
dotherine ground sloth (Mammalia, Xenarthra, Mylodonti-
dae). Unpublished PhD thesis, University of Toronto,
Toronto, 478 pp.
-1997. Xenarthrans: Pilosans. In KAY, R. F., MADDEN,
R. H., CIFELLI , R. L. and FLYNN, J. J. (eds). Vertebrate
paleontology in the Neotropics. The Miocene fauna of La Venta,
Colombia. Smithsonian Institution Press, Washington, 608 pp.
-2005. Palecology of extinct xenarthrans and the great
American biotic interchange. Bulletin of the Florida Museum of
Natural History, 45, 319–340.-and NAPLES , V. L. 2007. Chapter 10. Xenarthra. In
JANIS , C. M., GUNNELL, G. F. and UHEN, M. D.
(eds). Evolution of tertiary mammals of North America, Vol. 2:
Small mammals, xenarthrans, and marine mammals. Cam-
bridge University Press, 795 pp.
MI ~NO-BOILINI , �A. R., TOMASSINI , R. L., OLIVA, C.
and DE BIANCO, T. M. 2011. Adiciones al Conocimiento
de Proscelidodon Bordas, 1935 (Mammalia, Xenarthra, Sceli-
dotheriinae). Revista Brasileira de Paleontologia, 14, 269–278.-CARLINI , A. A. and SCILLATO-YAN�E, G. J. 2014.
Revisi�on sistem�atica y taxon�omica del g�enero Scelidotherium
Owen, 1839 (Xenarthra, Phyllophaga, Mylodontidae). Revista
Brasileira de Paleontologia, 17, 43–58.MONES, A. 1988. Nuevos registros de mamiferos fosiles de la
Formacion San Jos�e (Plioceno – ¿Pleistoceno inferior?) (Mam-
malia: Xenarthra; Artiodactyla; Rodentia). Comunicaciones
Paleontologicas des Museo de Hostoria Natural de Montevideo,
20, 255–277.MONTES, C., CARDONA, A., JARAMILLO, C.,
PARDO, A., S ILVA, J. C., VALENCIA, V., AYALA, C.,
P�EREZ-ANGEL, L. C., RODR�IGUEZ-PARRA, L. A.,
RAM�IREZ, V. and NI ~NO, H. 2015. Middle Miocene clo-
sure of the Central American Seaway. Science, 348, 226–229.MORENO, F., HENDY, A. J. W., QUIROZ, L., HOYOS,
N., JONES , D. S., ZAPATA, V., ZAPATA, S., BALLEN,
G. A., CADENA, E., C �ARDENAS, A. L., CARRILLO-
BRICE ~NO, J. D., CARRILLO, J. D., DELGADO-
SIERRA, D., ESCOBAR, J., MART�INEZ, J. I., MAR-
T�INEZ, C., MONTES, C., MORENO, J., P�EREZ, N.,
S �ANCHEZ, R., SU �AREZ, C., VALLEJO-PAREJA, M.
C. and JARAMILLO, C. 2015. Revised stratigraphy of Neo-
gene strata in the Cocinetas Basin, La Guajira, Colombia.
Swiss Journal of Palaeontology, 134, 5–43.MORENO-BERNAL, J. W., HEAD, J. and JARAMILLO,
C. A. 2016. Fossil crocodilians from the High Guajira Penin-
sula of Colombia: neogene faunal change in northernmost
South America. Journal of Vertebrate Paleontology, 36 (3),
e1110586.
NOWAK, R. M. and PARADISO, J. L. 1983. Walker’s Mam-
mals of the World vols I–II. John Hopkins University Press,
Baltimore, 1306 pp.
OWEN, R. 1842. Description of the skeleton of an extinct giant
sloth, Mylodon robustus, Owen. With observations on the oste-
ology, natural affinities, and probable habits of the megatherioid
quadrupeds in general. R. & J. E. Taylor, London, 176 pp.
-1861. Memoir on the Megatherium, or giant ground-sloth of
America (Megatherium americanum, Cuvier). Williams &
Norgate, London, 84 pp.
OWEN-SMITH, N. 1987. Pleistocene extinctions: the pivotal
role of megaherbivores. Paleobiology, 13, 351–362.PATTERSON, B., SEGALL, W., TURNBULL, W. D. and
GAUDIN, T. J. 1992. The ear region in xenarthrans (=Edentata: Mammalia) Part II. Pilosa (Sloths, Anteaters),
palaeanodonts, and a miscellany. Fieldiana. Geology, New Ser-
ies, 24, 1–79.PAULA COUTO, C. DE 1971. On two small Pleistocene
ground-sloths. Anais da Academia Brasileira de Ciencias, 43,
499–513.PEREA, D., RINDERKNECHT, A., UBILLA, M.,
BOSTELMANN, T. E. and MART�INEZ, S. 2013.
Mam�ıferos y estratigraf�ıa del Ne�ogeno de Uruguay. Asociaci�on
Paleontol�ogica Argentina, Publicaci�on Especial, 14, 192–206.PUJOS, F. and SALAS , R. 2004. A new species of Megath-
erium (Mammalia: Xenarthra: Megatheriidae) from the Pleis-
tocene of Sacaco and Tres Ventanas, Peru. Palaeontology, 47,
579–604.-and DE IULIIS , G. 2007. Late Oligocene Megatherioidea
fauna (Mammalia: Xenarthra) from Salla-Luribay (Bolivia):
new data on basal sloth radiation and Cingulata-Tardigrada
split. Journal of Vertebrate Paleontology, 27, 132–144.-SALAS, R., BABY, G., BABY, P., GOILLOT, C.,
TEJADA, J. and ANTOINE, P.-O. 2013. Implication of
the presence of Megathericulus (Xenarthra: Tardigrada:
Megatheriidae) in the Laventan of Peruvian Amazonia. Journal
of Systematic Palaeontology, 11, 973–991.RAJ PANT, S., GOSWAMI, A. and FINARELLI , J. A.
2014. Complex body size trends in the evolution of sloths
(Xenarthra: Pilosa). BMC Evolutionary Biology, 14, 184.
REGUERO, M. A. and CANDELA, A. M. 2011. Late Cenozoic
mammals from the northwest of Argentina. In SALFITY, J. A.
and MARQUILLAS , R. A. (eds). Cenozoic geology of the Cen-
tral Andes of Argentina. SCS Publisher, Salta, 458 pp.
--and ALONSO R. N. 2007. Biochronology and bios-
tratigraphy of the Uqu�ıa Formation (Pliocene–early Pleis-
tocene, NW Argentina) and its significance in the Great
American Biotic Interchange. Journal of South American Earth
Sciences, 23, 1–16.RINDERKNECHT, A., PEREA, D. and McDONALD, H.
G. 2007. A new Mylodontinae (Mammalia, Xenarthra) from
the Camacho Formation (Late Miocene), Uruguay. Journal of
Vertebrate Paleontology, 27, 744–747.-BOSTELMANN, T. E., PEREA, D. and LECUONA,
G. 2010. A new genus and species of Mylodontidae (Mam-
malia: Xenarthra) from the late Miocene of southern Uruguay,
with comments on the systematics of the Mylodontinae. Jour-
nal of Vertebrate Paleontology, 30, 899–910.
AMSON ET AL . : NEOGENE SLOTHS FROM THE COCINETAS BAS IN 19

RODR�IGUEZ-BRIZUELA, R. and TAUBER, A. 2006.
Estratigraf�ıa y mam�ıferos f�osiles de la Formaci�on Toro Negro
(Ne�ogeno), Departamento Vinchina, noroeste de la provincia
de La Rioja, Argentina. Ameghiniana, 43, 257–272.ROVERETO, C. 1914. Los estratos araucanos y sus f�osiles.
Anales del Museo Nacional de Buenos Aires, 25, 1–249.RUSCONI , C. 1954. Tercera noticia sobre los vertebrados fos-
iles de las arenas de Villa Ballester. Anales de la Sociedad
Cient�ıfica Argentina, 117, 19–37.SAINT-ANDR�E, P. A., PUJOS, F., CARTELLE, C. and
DE IULIIS , G. 2010. Nouveaux paresseux terrestres (Mam-
malia, Xenarthra, Mylodontidae) du N�eog�ene de l’Altiplano
bolivien. Geodiversitas, 32, 255–306.SALAS-GISMONDI , R., FLYNN, J. J., BABY, P.,
TEJADA-LARA, J. V., WESSELINGH, F. P. and
ANTOINE, P.-O. 2015. A Miocene hyperdiverse crocodylian
community reveals peculiar trophic dynamics in proto-Ama-
zonian mega-wetlands. Proceedings of the Royal Society B, 282,
20142490.
SCHEYER, T. M., AGUILERA, O. A., DELFINO, M.,
FORTIER, D. C., CARLINI , A. A., S �ANCHEZ, R.,
CARRILLO-BRICE ~NO, J. D., QUIROZ, L. and
S �ANCHEZ-VILLAGRA, M. R. 2013. Crocodylian diversity
peak and extinction in the late Cenozoic of the northern
Neotropics. Nature Communications, 4, 1907.
SCHINDELIN, J., ARGANDA-CARRERAS, I., FRISE,
E., KAYNIG, V., LONGAIR, M., PIETZSCH, T., PRE-
IBISCH, S., RUEDEN, C., SAALFELD, S., SCHMID,
B., TINEVEZ, J.-Y., WHITE, D. J., HARTENSTEIN,
V., ELICEIRI , K., TOMANCAK, P. and CARDONA, A.
2012. Fiji: an open-source platform for biological-image analy-
sis. Nature Methods, 9, 676–682.SCOTT, W. B. 1903–1904. Mammalia of the Santa Cruz beds.
Reports of the Princeton University Expeditions to Patagonia, 5,
1–490.S IMPSON, G. G. 1980. Splendid isolation: the curious history of
South American mammals. Yale University Press, New Haven,
266 pp.
STOCK, C. 1925. Cenozoic gravigrade edentates of western
North America, with special reference to the Pleistocene
Megalonychinae and Mylodontidae of Rancho La Brea. Carne-
gie Institution of Washington Publications, 331, 1–206.-1942. A ground sloth in Alaska. Science, 95, 552–553.
SU �AREZ, C., FORASIEPI , A. M., GOIN, F. J. and JARA-
MILLO, C. 2015. Insights into the Neotropics Prior to the
Great American Biotic Interchange: new evidence of mam-
malian predators from the Miocene of Northern Colombia.
Journal of Vertebrate Paleontology, 36 (1), e1029581.
SWOFFORD, D. L. 2002. PAUP: phylogenetic analysis using
parsimony, v. 4.0b10. Sinauer Associates, Sunderland, MA.
TEJADA-LARA, J. V., SALAS-GISMONDI , R., DE
FRANCESCHI , D., ESPURT, N., URBINA, M. and
ANTOINE, P.-O. 2015. Life in Proto-Amazonia: middle
Miocene mammals from the Fitzcarrald arch (Peruvian Ama-
zonia). Palaeontology, 58, 341–378.TOMASSINI , R. L., MONTALVO, C. I., DESCHAMPS,
C. M. and MANERA, T. 2013. Biostratigraphy and
biochronology of the Monte Hermoso Formation (early Plio-
cene) at its type locality, Buenos Aires Province, Argentina.
Journal of South American Earth Sciences, 48, 31–42.VILLARROEL, C. 2000. Un nuevo Mylodontinae (Xenarthra,
Tardigrada) en la fauna de La Venta, Mioceno de Colombia:
el estado actual de la familia Orophodontidae. Revista Acade-
mia Colombiana de Ciencias, 24, 117–127.VUCETICH, M. G., CARLINI , A. A., AGUILERA, O. and
S �ANCHEZ-VILLAGRA, M. R. 2010. The Tropics as reser-
voir of otherwise extinct mammals: the case of rodents from a
new Pliocene faunal assemblage from northern Venezuela.
Journal of Mammalian Evolution, 17, 265–273.WALLACE, A. R. 1876. The geographical distribution of ani-
mals; with a study of the relations of living and extinct faunas
as elucidating the past changes of the Earth’s surface, vols 1 & 2.
Harper & Brothers, New York, 503 & 607 pp.
WEBB, S. D. 1985. Late Cenozoic mammal dispersal between
the Americas. In STEHLIN, F. G. and WEBB, S. D. (eds).
The Great American biotic interchange. Plenum Press, New
York, 532 pp.
-1989. Osteology and Relationships of Thinobadistes segnis,
the first mylodont sloth in North America. In REDFORD,
K. and EISENBERG, J. F. (eds). Adavances in neotropical
mammalogy. The Sandhill Crane Press Inc., Gainesville, 614 pp.
-2006. The great American biotic interchange: patterns and
processes. Annals of the Missouri Botanical Garden, 93, 245–257.WOODBURNE, M. O. 2010. The Great American Biotic
Interchange: dispersals, tectonics, climate, sea level and hold-
ing pens. Journal of Mammalian Evolution, 17, 245–264.
20 PALAEONTOLOGY