mouse model of pulmonary cavitary tuberculosis and ... · mouse model of pulmonary cavitary...
TRANSCRIPT

RESEARCH ARTICLE
Mouse model of pulmonary cavitary tuberculosis and expressionof matrix metalloproteinase-9Alvaro A. Ordonez1,2,3,*, Rokeya Tasneen2,4,*, Supriya Pokkali1,2,3, Ziyue Xu5, Paul J. Converse2,4,Mariah H. Klunk1,2,3, Daniel J. Mollura5, Eric L. Nuermberger2,4,‡,§ and Sanjay K. Jain1,2,3,‡,§
ABSTRACTCavitation is a key pathological feature of human tuberculosis (TB),and is a well-recognized risk factor for transmission of infection,relapse after treatment and the emergence of drug resistance.Despite intense interest in the mechanisms underlying cavitationand its negative impact on treatment outcomes, there has beenlimited study of this phenomenon, owing in large part to thelimitations of existing animal models. Although cavitation does notoccur in conventional mouse strains after infection withMycobacterium tuberculosis, cavitary lung lesions haveoccasionally been observed in C3HeB/FeJ mice. However, todate, there has been no demonstration that cavitation can beproduced consistently enough to support C3HeB/FeJ mice as anew and useful model of cavitary TB. We utilized serial computedtomography (CT) imaging to detect pulmonary cavitation inC3HeB/FeJ mice after aerosol infection with M. tuberculosis.Post-mortem analyses were performed to characterize lung lesionsand to localize matrix metalloproteinases (MMPs) previouslyimplicated in cavitary TB in situ. A total of 47-61% of infectedmice developed cavities during primary disease or relapse afternon-curative treatments. Key pathological features of human TB,including simultaneous presence of multiple pathologies, werenoted in lung tissues. Optical imaging demonstrated increasedMMP activity in TB lesions and MMP-9 was significantly expressedin cavitary lesions. Tissue MMP-9 activity could be abrogated byspecific inhibitors. In situ, three-dimensional analyses of cavitarylesions demonstrated that 22.06% of CD11b+ signal colocalizedwith MMP-9. C3HeB/FeJ mice represent a reliable, economicaland tractable model of cavitary TB, with key similarities to humanTB. This model should provide an excellent tool to betterunderstand the pathogenesis of cavitation and its effects on TBtreatments.
KEY WORDS: Tuberculosis, Cavity, Computed tomography, Matrixmetalloproteinases, Mice
INTRODUCTIONCavitation of pulmonary lesions is a defining event in individualswith tuberculosis (TB), with negative implications for the individualand for society (Benator et al., 2002; Blumberg et al., 2003; Dormanet al., 2009). For the individual, cavitary TB is associated with poortreatment outcomes, including delayed sputum culture conversion,relapse after treatment and development of drug resistance. Forsociety, cavitation greatly increases the risk of person-to-persontransmission. For these reasons, more effective means to treat andprevent cavitary TB are crucial for efforts to shorten the duration ofTB treatments, limit the development of drug resistance and reducetransmission. Such efforts would be aided immensely by betterunderstanding of the processes leading to cavitation, and the factorsdriving reduced treatment responses in patients with cavitary TB.
Cavitation is a consequence of the distinctive caseation necrosisthat is the pathological hallmark of human TB. As a result, efforts todevelop animal models of cavitary TB have focused on species thatexhibit caseation necrosis upon infection with Mycobacteriumtuberculosis complex, especially rabbits (Converse et al., 1996;Manabe et al., 2003; Nedeltchev et al., 2009; Subbian et al., 2011;Via et al., 2012; Wells and Lurie, 1941; Yamamura, 1958) and non-human primates (NHPs) (Capuano et al., 2003; Via et al., 2013).However, use of these larger animals is often cost prohibitive, and,in the case of rabbits, is challenged by the limited availability ofreagents. Furthermore, these models require infection with specificstrains (Converse et al., 1996; Subbian et al., 2011; Via et al., 2013;Wells and Lurie, 1941), prior sensitization with heat-killed bacilli,and/or bronchoscopic infection to create cavitary disease(Nedeltchev et al., 2009; Yamamura, 1958). Even then, cavitationcan be an infrequent event (Lin et al., 2013). A reliable murinemodel of cavitary TB would be much more economical and afford awide array of reagents for investigation of disease mechanisms.
Mice have been overlooked as models of cavitary TB becausecommonly used mouse strains do not exhibit caseous pathologyafter infection with M. tuberculosis. However, C3HeB/FeJmice were discovered to develop caseous lung lesions afterM. tuberculosis infection (Pan et al., 2005); these lesions are nowknown to be hypoxic (Driver et al., 2012; Harper et al., 2012) and toundergo calcification similar to human TB (Ordonez et al., 2015a).C3HeB/FeJ mice are not deficient in the activation of Th1 cytokine-producing T-cells or their migration to the lungs, and they are ableto control infection with Bacille Calmette–Guerin vaccine strains(Yan et al., 2007). Their susceptibility to M. tuberculosis infectionis determined primarily at the sst1 locus, which regulates themacrophage innate immune response to infection with intracellularpathogens (Pan et al., 2005; Pichugin et al., 2009). Macrophages ofC3HeB/FeJ mice have a reduced ability to restrain multiplication ofM. tuberculosis and these cells preferentially undergo cell necrosisrather than apoptosis, which is associated with activation of the typeI interferon pathway and an exaggerated host inflammatory responseReceived 1 April 2016; Accepted 20 May 2016
1Center for Infection and Inflammation Imaging Research, Johns HopkinsUniversity School of Medicine, Baltimore, MD 21287, USA. 2Center for TuberculosisResearch, Johns Hopkins University School of Medicine, Baltimore, MD 21287,USA. 3Department of Pediatrics, Johns Hopkins University School of Medicine,Baltimore, MD 21287, USA. 4Department of Medicine, Johns Hopkins UniversitySchool of Medicine, Baltimore, MD 21287, USA. 5Center for Infectious DiseaseImaging, National Institutes of Health, Bethesda, MD 20892, USA.*These authors contributed equally to this work‡These authors contributed equally to this work
§Authors for correspondence ([email protected]; [email protected])
S.K.J., 0000-0001-9620-7070
This is an Open Access article distributed under the terms of the Creative Commons AttributionLicense (http://creativecommons.org/licenses/by/3.0), which permits unrestricted use,distribution and reproduction in any medium provided that the original work is properly attributed.
779
© 2016. Published by The Company of Biologists Ltd | Disease Models & Mechanisms (2016) 9, 779-788 doi:10.1242/dmm.025643
Disea
seModels&Mechan
isms

(He et al., 2013; Pan et al., 2005). Mounting evidence that type-I-interferon signaling pathways are upregulated in active TB (Berryet al., 2010), as well as evidence associating TB withpolymorphisms in SP110, the closest human homolog of thecandidate Ipr1 gene of the mouse sst1 locus, indicates the likelyrelevance of this mouse strain (Abhimanyu et al., 2011; He et al.,2013; Tosh et al., 2006). Reports of occasional cavitation in C3HeB/FeJ mice (Driver et al., 2012; Lanoix et al., 2015; Ordonez et al.,2015b) support further investigation of their potential as a cavitaryTB model. However, to date, there has been no demonstration thatcavitation can be produced consistently enough to support suchusage. For this study, we utilized serial computed tomography (CT)imaging to detect pulmonary lesions and cavitation in C3HeB/FeJmice after aerosol infection with M. tuberculosis while evaluatingthe impact of methods to promote cavity formation. Detailed post-mortem histopathological and immunological analyses were alsoperformed to characterize the TB lesions and to probe for matrixmetalloproteinases (MMPs) that are implicated in the process ofcavitation (Ong et al., 2014; Salgame, 2011).
RESULTSIncidence of cavitary diseaseIn a series of experiments, C3HeB/FeJ mice infected withM. tuberculosis H37Rv were imaged using CT at predeterminedtime points (Table 1). In one experiment, 4 of 9 untreated miceimaged at 8 weeks post-infection (wpi) had cavitary lesions. In otherexperiments, 14 of 31 mice and 10 of another 20 mice had cavities at10 and 14 wpi, respectively. Thus, the combined proportion ofuntreatedmice developing cavities by 8-14 weeks post-infectionwas47% (28 of 60). Because cavitary disease is classically associatedwith reactivation TB, we also evaluated the incidence of cavitationduring relapse after non-curative drug treatment. At 6 weeks afterinfection with M. tuberculosis H37Rv, mice received standard TBtreatment [8 weeks of rifampin, isoniazid and pyrazinamide (RHZ),then another 4 weeks of RH]. CT was performed 4 weeks aftercompletion of treatment (22 wpi), at which time 11 of 18 (61%)micehad cavities (Table 1) (Ordonez et al., 2015b). These mice (withcavitation) appeared substantially healthier than those that developedcavities in the context of untreated infection.Prior studies in rabbits and NHPs induced cavitation by
sensitizing with heat-killed bacilli prior to challenge with viableM. tuberculosis (Nedeltchev et al., 2009; Yamamura, 1958) orinfections with Beijing lineage M. tuberculosis (Subbian et al.,2011; Via et al., 2012, 2013). We therefore compared cavitationin sensitized or un-sensitized mice following infection withM. tuberculosis HN878. Unexpectedly, sensitization protectedagainst early death and cavitation (Fig. S1). Survival was higheramong sensitized mice (median 136 days) compared to un-sensitized mice [median 107 days; log-rank (Mantel-Cox) testP<0.01]. One-quarter of un-sensitized mice died within 44 days
post-infection (dpi), whereas no sensitized mouse died before93 dpi. The improved survival among sensitized mice wasassociated with better immune containment of infection. At 6 wpi,the pulmonary bacterial burden was lower in sensitized [5.97±0.14log10 colony forming units (CFU)] versus un-sensitized (7.77±1.83log10 CFU)mice [P=0.14;Mann–Whitney test; two of the four micein the un-sensitized group had a very high bacterial burden (>9.3log10 CFU)]. In the absence of drug treatment, none of 15mice in thesensitized group developed cavities versus 2 (40%) of 5 survivingmice in the un-sensitized group by 14 wpi (Fisher’s exact test,P=0.05). Among mice treated with RHZ for 8 weeks beginning at6 wpi, cavitation rates during the 24-week post-treatment follow-upperiod were 13% (2 of 15) in the sensitized group and 53% (8 of 15)in the un-sensitized group (Fisher’s exact test, P=0.05).
Cavitary lesionsPulmonary cavities were clearly visualized on CT imaging of liveM. tuberculosis-infected animals (Fig. 1A). On post-mortemanalysis, cavities were surrounded by a fibrotic capsule and oftencontained residual, partially liquefied caseum (Fig. 1B). Serial CTimaging demonstrated the development of cavities within enlargingnecrotic masses and the close association of cavities with majorairways (Fig. 1C). High-power microscopic views demonstratedmixed inflammatory cellular infiltrates lining the cavity wall,numerous extracellular acid-fast bacilli, as well as intracellularbacilli inside neutrophils and foamy macrophages, and a fibroticrim surrounding the cavity. Immunohistochemistry demonstrated ahigh density of CD11b+ and Gr-1+ cells, with morphologicalcharacteristics of macrophages and neutrophils, respectively.CD11b+ multinucleated giant cells were also observed (Fig. 1D).A similar architecture surrounding the caseous core – consisting ofcellular infiltrate and CD11b+ multinucleated giant cells, numerousacid-fast bacilli, and fibrosis – was also noted in necrotic but non-cavitary lesions (Fig. S2).
We investigated the relationship between major airways andcavitary lesions (Fig. 2, Movie 1). CT imaging (Fig. 2A) andconsecutive post-mortem histopathological sections from the sameanimal (Fig. 2B,C) demonstrated direct communication between theairway and the partially evacuated cavity. Debris was noted in theairways (Fig. 2C, inset). Another lesion evacuating into the airwaysis shown in Fig. S3. Cellular and necrotic debris or casts were seeninside the airways, with numerous acid-fast bacilli.
Lesion heterogeneity and simultaneous presence ofmultiple lung pathologiesLesions in various stages of development were noted simultaneouslyduring the course of infection (Fig. S4). By 2-4 wpi, the intra-alveolar spaces were occupied by a cellular infiltrate (Fig. S4A) thatevolved into organized granulomaswith fibrosis and central necrosis(4-6 wpi; Fig. S4B,C). At later stages (8-14 wpi), granulomasbecame organized, with central caseous necrosis and increasingnumbers of extracellular bacilli (Fig. S4D,F). The associated fibrosisprogressed from small collagen fibers in the alveolar walls to anorganized fibrotic ring surrounding the granuloma. Multiplepathologies – caseous pneumonia, necrotizing granulomas andcavitary lesions – were often seen alongside cellular lesions withinthe same lung (Fig. 3), as previously described for C3HeB/FeJ mice(Irwin et al., 2015), but with the added feature of cavitation.
MMP-9 expressionDestruction of lung extracellular matrix is necessary for theformation of cavities, and MMPs, a family of zinc-dependent
Table 1. Incidence of cavitary disease assessed by CT imaging afteraerosol infection with M. tuberculosis
Experimentalgroup
Pulmonary bacterial burden(log10 CFU)
Proportion of micewith cavities
Implantation (day1 post infection) 6 wpi
Untreated* 2.05±0.18 7.96±0.62 28 out of 60 (47%)Relapse‡ 2.00±0.13 8.05±0.02 11 out of 18 (61%)
*Cavitation noted 8-14 wpi.‡Cavitation assessed 4 weeks after completion of non-curative TB treatment.
780
RESEARCH ARTICLE Disease Models & Mechanisms (2016) 9, 779-788 doi:10.1242/dmm.025643
Disea
seModels&Mechan
isms

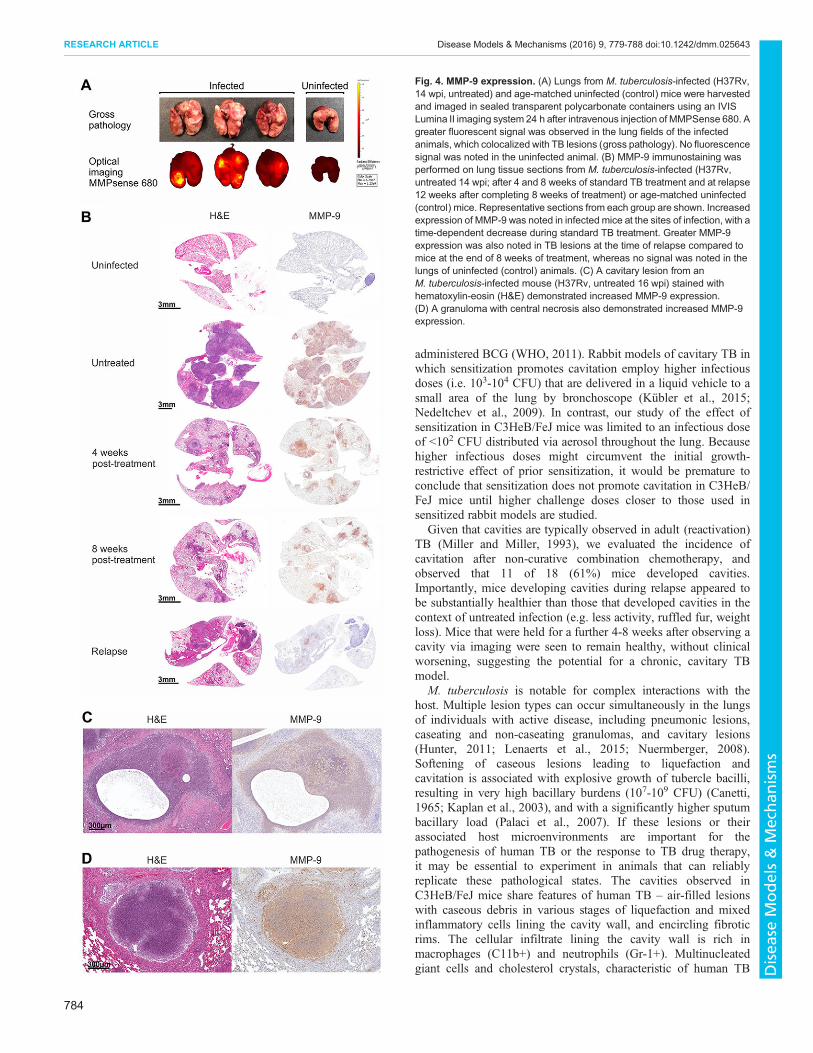
proteases, are known to degrade components of the extracellularmatrix (Greenlee et al., 2007). MMP activity was qualitativelyevaluated in the lungs of M. tuberculosis-infected mice withMMPSense 680, an in vivo imaging agent activated by key MMPs,including by MMP-9. Fluorescent optical imaging clearlydemonstrated increased MMP activity, which colocalized with TBlesions (Fig. 4A). Immunohistochemical analysis revealedincreased expression of MMP-9 in the lungs of infected mice,with the signal colocalizing to TB lesions. Time-dependentdecreases in MMP expression were observed during standard TBtreatment, followed by increased expression in lesions of relapsingmice (Fig. 4B). No signal was noted in the lungs of age-matcheduninfected mice. High-power views demonstrated robust MMP-9
expression surrounding cavities and in and around necrotic lesions(Fig. 4C,D). To complement immunohistochemistry, we measuredMMP-9 activity in lung homogenates by using gelatin zymography(Fig. S5). MMP-9 activity was noted in lungs from infected mice,but not from uninfected controls. Furthermore, treatment of lunghomogenates from M. tuberculosis-infected animals with knownMMP inhibitors [EDTA (10 mM) and doxycycline (0.25 mM)](Gendron et al., 1999; Liu et al., 2003), as well as with a highlyspecific MMP-9 inhibitor, SB-3CT (500 µM) (Gu et al., 2005),abrogated MMP activity. MMP-9 signal also colocalized withCD11b+ cells, which were abundant in the areas surroundingcavities and necrotic lesions (Fig. 5A,B). MMP-9 signal alsocolocalized with CD11+ cells in pneumonic areas (Fig. 5C). Finally,
Fig. 1. Cavitary lesions in C3HeB/FeJmice. (A) Transverse, coronal andsagittal CT images from arepresentative Mycobacteriumtuberculosis-infected (HN878, 29 wpi,after standard TB treatment from 6-14wpi) mouse demonstrating a thick-walled lung cavity (yellow arrow) areshown. (B) Post-mortem examination ofthe same animal demonstrated a cavitywith residual, partially liquefiedcaseous material (arrow), a thin cellularrim seen with hematoxylin-eosin (H&E)stain and a fibrotic capsule (Masson’strichrome stain; blue). (C) Serial CTimaging of another representativemouse infected with M. tuberculosisHN878 that received standard TBtreatment from 6-14 wpi, showing adecrease in the size of an inflammatorymass in the left lung between 9-14 wpi,followed by an increase in the size ofthe mass and evolution of a cavity(yellow arrows) 12 weeks afterdiscontinuation of treatment (26 wpi).Three-dimensional reconstruction ofthe skeleton (gray), airways (blue) andcavity (yellow) derived from the CTimages is also shown. (D) Low-powerview of a representative cavity withnecrotic content from anotherM. tuberculosis-infected mouse(H37Rv, 14 wpi) is shown. High-powerview of the cavity wall (inset)demonstrates inflammatory cells (H&E)with numerous extracellular andintracellular acid-fast bacilli, revealedwith Ziehl-Neelsen stain (AFB), andcollagen deposition (Masson’strichrome stain, blue).Immunohistochemistry analysis revealsa high density of CD11b+ (red) andsome Gr-1+ (red) cells. Multinucleatedgiant (MNG) cells, which are CD11b+,can also be observed in the infectedtissues. Black arrows, MNG. He, heart.
781
RESEARCH ARTICLE Disease Models & Mechanisms (2016) 9, 779-788 doi:10.1242/dmm.025643
Disea
seModels&Mechan
isms

MMP-9 signal colocalized with the majority of M. tuberculosis-infected cells in areas surrounding cavities and pneumonic areas.
Spatial localization of MMP signal with inflammatory cellsWe stained and digitally scanned 36 consecutive sections through apulmonary cavitary lesion from a representative M. tuberculosis-infected mouse (Fig. S6, Movie 2). MMP-9 demonstratedsignificant colocalization with CD11b+. Quantitative analysesdemonstrated that 22.06% of CD11b+ versus 3.75% of Gr-1+cells colocalized with the MMP-9 signal (Chi-square test withYates’ correction, P<0.01).
MMP-7 expressionBecause necrotic TB lesions in this model are hypoxic (Driver et al.,2012; Harper et al., 2012) and MMP-7 is induced by hypoxia
(Burke et al., 2003), we also investigated MMP-7 expression.Similar to MMP-9, immunohistochemical analysis revealedincreased expression of MMP-7 in TB lesions and time-dependent decreases in MMP-7 expression during standard TBtreatment, followed by increased expression in lesions duringrelapse (Fig. S7). No signal was noted in the lungs of age-matchedcontrols. High-power views also demonstrated MMP-7 expressionsurrounding cavities and in and around necrotic lesions. Spatiallocalization demonstrated that 16.74% of CD11b+ versus 2.37% ofGr-1+ cells colocalized with the MMP-7 signal (Chi-square testwith Yates’ correction, P<0.01).
DISCUSSIONAnimal models have key roles in the study of TB pathogenesis andin the development of new therapeutic and preventative measures.
Fig. 2. Communication of the cavitary lesion with airways. (A) Transverse, coronal and sagittal views, and three-dimensional reconstruction derived from CTimages from a representative M. tuberculosis-infected (H37Rv, 8 wpi; without TB treatment) mouse are shown. The airways (blue) directly communicate withthe cavity (yellow, arrows). (B) Post-mortem three-dimensional alignment of 98 hematoxylin-eosin (H&E)-stained consecutive histopathological sections ofthe lung from the same animal shows the cavity (arrow). (C) Consecutive H&E-stained histopathological sections from the same animal demonstrate thecommunication between a caseating granuloma with cavitation and an adjacent airway. Arrows, airway communicating with the cavity. The (enlarging) lumen ofthe cavity is seen in the consecutive serial sections, with partially liquefied retained caseum at the periphery. High-power views (insets) of each sectiondemonstrate intraluminal cellular and necrotic debris within the airway. He, heart.
782
RESEARCH ARTICLE Disease Models & Mechanisms (2016) 9, 779-788 doi:10.1242/dmm.025643
Disea
seModels&Mechan
isms

However, considering the importance of cavitary pulmonary TB,the development of models representing this pathological featurehas been remarkably limited. Our results demonstrate, for the firsttime, that C3HeB/FeJ mice can serve as a reliable model ofcavitary TB to complement models using larger species (Capuanoet al., 2003; Converse et al., 1996; Manabe et al., 2003; Nedeltchevet al., 2009; Subbian et al., 2011; Via et al., 2012, 2013; Wells andLurie, 1941; Yamamura, 1958). In addition to being lesseconomical and less tractable than mice, larger species haveother disadvantages. Rabbits are relatively resistant toM. tuberculosis infection and have traditionally requiredinfections with Mycobacterium bovis (Converse et al., 1996;Wells and Lurie, 1941) or specific strains of M. tuberculosis(Subbian et al., 2011), or sensitization with heat-killed bacilli priorto bronchoscopic infection to create cavities (Nedeltchev et al.,2009; Yamamura, 1958). Cavitation occurs in NHPs afterM. tuberculosis infection (Capuano et al., 2003; Lin et al., 2013;Via et al., 2013), but the frequency of cavitation may not be veryhigh [e.g. 4 of 26 (15%) among bronchoscopically infectedcynomolgus macaques] (Lin et al., 2013). Up to 50% (i.e. 3 of 6)marmosets developed cavities after aerosol infection, but only withone of three strains of M. tuberculosis complex (Via et al., 2013).Reliance upon M. bovis or specific M. tuberculosis lineages couldlimit generalizability to human TB. One example is the intrinsicresistance of M. bovis to pyrazinamide – a first-line TB drug that isalso a critical component of novel drug regimens underdevelopment. The impact of caseous pathology onpyrazinamide’s activity and its contribution to combinationchemotherapy has recently been elucidated in M. tuberculosis-infected C3HeB/FeJ mice (Lanoix et al., 2016a,b). Similarly,sensitization with heat-killed bacilli and bronchoscopic M.tuberculosis infection promotes cavitation in rabbits, but alsoincreases the cost and complexity of the model.We utilized serial CT imaging to rigorously quantify and
characterize pulmonary cavitation in C3HeB/FeJ mice afteraerosol infection with two M. tuberculosis strains. Radiologicalimaging has been used extensively to assess pulmonary TB (Daviset al., 2009; Kübler et al., 2015; Lin et al., 2013; Ordonez et al.,
2015b; Via et al., 2012) and, more recently, as a biomarker oftreatment responses in humans (Chen et al., 2014; Salazar-Austinet al., 2015). It is noninvasive and provides rapid, three-dimensionalviews of the whole organ. It also has the fundamental advantageof longitudinal assessments to monitor disease progression in thesame individual, which could also provide new insights into thepathophysiology of disease that are not feasible with conventionalmethods. We observed pulmonary cavities in 47-61% of C3HeB/FeJ mice, a rate that may be ideal to study the factors associated withcavitation and the impact of cavitation on treatment outcomes. Thecavitation rate after M. tuberculosis HN878 (East Asian lineage)infection (40%) was similar to that observed with M. tuberculosisH37Rv (Euro-American lineage) (47%), although mice infectedwith HN878 were more likely to require euthanasia prior to the firstimaging time point. Further work is needed to determine whetherother M. tuberculosis strains cause cavitation at similar rates. Wealso evaluated the effect of sensitization with heat-killed bacilli priorto infection with M. tuberculosis HN878 (Beijing subfamily).Unlike prior observations in rabbits (Nedeltchev et al., 2009;Yamamura, 1958), sensitization was protective in C3HeB/FeJ mice,leading to increased survival, lower pulmonary bacterial burdens,and lower rates of cavitation during progressive disease and relapsecompared to un-sensitized animals. It is likely that the lower rates ofcavitation in sensitized mice was the result of an immunizing effectof the sensitization procedure that restricted initial bacterial growth,as evidenced by the lower lung CFU counts of sensitized mice, andreduced the number and size of caseating lesions in favor of cellulargranulomas that do not caseate and thus do not cavitate (Irwin et al.,2015). BCG vaccination has been associated with reduced bacillaryburden at the early stages of infection in M. tuberculosis-infectedC3HeB/FeJ mice, with delayed formation of necrotic granulomas(Yan et al., 2007). Therefore, it is likely that immunization in thismodel may delay, but not prevent, the formation of cavitarygranulomas, as well. Additional longitudinal studies using liveimaging are required to clarify the impact of immunization on theprogression trajectories of individual granulomas. Similar responsesthat limit M. tuberculosis burden at the site of primary infection(mostly lungs) are noted in primary disease in children who were
Fig. 3. Lesion heterogeneity.Histopathological section from the lung of a representativeM. tuberculosis-infected (H37Rv, 14 wpi; without TB treatment) mouseis shown. Higher-power views (insets) demonstratemultiple pathologies – granulomaswith central caseation (A,D), pneumonia (B) and cavitation (C) – in differentareas of the same lung tissue simultaneously.
783
RESEARCH ARTICLE Disease Models & Mechanisms (2016) 9, 779-788 doi:10.1242/dmm.025643
Disea
seModels&Mechan
isms
administered BCG (WHO, 2011). Rabbit models of cavitary TB inwhich sensitization promotes cavitation employ higher infectiousdoses (i.e. 103-104 CFU) that are delivered in a liquid vehicle to asmall area of the lung by bronchoscope (Kübler et al., 2015;Nedeltchev et al., 2009). In contrast, our study of the effect ofsensitization in C3HeB/FeJ mice was limited to an infectious doseof <102 CFU distributed via aerosol throughout the lung. Becausehigher infectious doses might circumvent the initial growth-restrictive effect of prior sensitization, it would be premature toconclude that sensitization does not promote cavitation in C3HeB/FeJ mice until higher challenge doses closer to those used insensitized rabbit models are studied.
Given that cavities are typically observed in adult (reactivation)TB (Miller and Miller, 1993), we evaluated the incidence ofcavitation after non-curative combination chemotherapy, andobserved that 11 of 18 (61%) mice developed cavities.Importantly, mice developing cavities during relapse appeared tobe substantially healthier than those that developed cavities in thecontext of untreated infection (e.g. less activity, ruffled fur, weightloss). Mice that were held for a further 4-8 weeks after observing acavity via imaging were seen to remain healthy, without clinicalworsening, suggesting the potential for a chronic, cavitary TBmodel.
M. tuberculosis is notable for complex interactions with thehost. Multiple lesion types can occur simultaneously in the lungsof individuals with active disease, including pneumonic lesions,caseating and non-caseating granulomas, and cavitary lesions(Hunter, 2011; Lenaerts et al., 2015; Nuermberger, 2008).Softening of caseous lesions leading to liquefaction andcavitation is associated with explosive growth of tubercle bacilli,resulting in very high bacillary burdens (107-109 CFU) (Canetti,1965; Kaplan et al., 2003), and with a significantly higher sputumbacillary load (Palaci et al., 2007). If these lesions or theirassociated host microenvironments are important for thepathogenesis of human TB or the response to TB drug therapy,it may be essential to experiment in animals that can reliablyreplicate these pathological states. The cavities observed inC3HeB/FeJ mice share features of human TB – air-filled lesionswith caseous debris in various stages of liquefaction and mixedinflammatory cells lining the cavity wall, and encircling fibroticrims. The cellular infiltrate lining the cavity wall is rich inmacrophages (C11b+) and neutrophils (Gr-1+). Multinucleatedgiant cells and cholesterol crystals, characteristic of human TB
Fig. 4. MMP-9 expression. (A) Lungs from M. tuberculosis-infected (H37Rv,14 wpi, untreated) and age-matched uninfected (control) mice were harvestedand imaged in sealed transparent polycarbonate containers using an IVISLumina II imaging system 24 h after intravenous injection of MMPSense 680. Agreater fluorescent signal was observed in the lung fields of the infectedanimals, which colocalized with TB lesions (gross pathology). No fluorescencesignal was noted in the uninfected animal. (B) MMP-9 immunostaining wasperformed on lung tissue sections from M. tuberculosis-infected (H37Rv,untreated 14 wpi; after 4 and 8 weeks of standard TB treatment and at relapse12 weeks after completing 8 weeks of treatment) or age-matched uninfected(control) mice. Representative sections from each group are shown. Increasedexpression of MMP-9 was noted in infected mice at the sites of infection, with atime-dependent decrease during standard TB treatment. Greater MMP-9expression was also noted in TB lesions at the time of relapse compared tomice at the end of 8 weeks of treatment, whereas no signal was noted in thelungs of uninfected (control) animals. (C) A cavitary lesion from anM. tuberculosis-infected mouse (H37Rv, untreated 16 wpi) stained withhematoxylin-eosin (H&E) demonstrated increased MMP-9 expression.(D) A granuloma with central necrosis also demonstrated increased MMP-9expression.
784
RESEARCH ARTICLE Disease Models & Mechanisms (2016) 9, 779-788 doi:10.1242/dmm.025643
Disea
seModels&Mechan
isms

lesions (Hunter, 2011), are also observed. CT imaging revealedcavities communicating with major airways containingintraluminal cellular and necrotic debris with numerous intra-and extra-cellular acid-fast bacilli, indicating that the cavities inthis model developed when liquefied caseum was evacuated fromthese lesions through the airways. Serial imaging and necropsiesrevealed cavities within the necrotic center of organized caseouslesions situated within larger zones of caseous pneumonia,findings described in other models of cavitary TB (Kübler et al.,2015; Via et al., 2013), and in affected humans (Canetti, 1955).Moreover, lesions in various stages of development with multiplepathologies – pneumonia, necrosis and cavitation – were oftenseen within the lung tissue of the same mouse, suggesting lesion-specific progression of pathology as described in humans andNHPs (Canetti, 1955; Kaushal et al., 2012). However, thedevelopment of necrotic granulomas from progressive andcoalescing pneumonic lesions cannot be entirely ruled out. Thepresence of multiple different pulmonary lesions in C3HeB/FeJmice was described recently (Irwin et al., 2015), but cavitation wasa rare event under the conditions evaluated. Because several TBdrugs with differing physicochemical properties have been shownto partition differently into these lesions in a manner similar toobservations in humans and/or rabbits (DeMarco et al., 2015;Irwin et al., 2016; Kjellsson et al., 2012; Lanoix et al., 2016b;Prideaux et al., 2015; Weinstein et al., 2012), and because theactivity of some drugs might be modulated by conditions withinlarge caseous lesions (Lanoix et al., 2015, 2016a), the moretractable and economical C3HeB/FeJ mouse model that exhibits
these multiple lesion types provides an important new tool for TBdrug development research.
Destruction of lung extracellular matrix is a prerequisite for cavityformation. MMPs, a family of zinc-dependent proteases, are knownto degrade several components of the extracellular matrix (Greenleeet al., 2007). The role of MMPs in TB is well summarized in recentreviews (Ong et al., 2014; Salgame, 2011) and multiple MMPshave been associated with TB pathogenesis. In particular, previousstudies have implicated MMP-1 (collagenase), MMP-9 (gelatinase)and MMP-7 (matrilysin) in active TB and cavitation (Singh et al.,2014). We qualitatively assessed MMP activity using opticalimaging with MMPSense 680, an in vivo imaging agent activatedby key MMPs, including MMP-9. Optical imaging clearlydemonstrated MMP activity, which colocalized with TB lesionsseen on gross pathology. Although wild-type mice do not express anortholog of MMP-1 (Elkington et al., 2011), they do expressMMP-7 and MMP-9. The absence of an MMP-1 homolog in miceindicates that this MMP is not essential for cavity formation in thismodel. Therefore, other collagenases (MMP-8, MMP-13) may playa role in degrading the fibrillary collagen of the lung extracellularmatrix in this mouse model. MMP-9 is known to degrade primarilycollagen type IV present in the basement membrane of alveoli(Gioia et al., 2009). Immunohistochemistry demonstrated increasedexpression of MMP-9 surrounding cavitary lesions as well as in andaround necrotic lesions, which decreased with TB treatment. Thiswas corroborated by robust MMP-9 activity in infected lung tissues,which was abrogated by known MMP inhibitors (EDTA anddoxycycline) (Gendron et al., 1999; Liu et al., 2003), as well as by a
Fig. 5. Association of MMP expression with macrophages. Hematoxylin-eosin (H&E), Ziehl-Neelsen (AFB), macrophage (CD11b+, red) and MMP-9 (brown)staining was performed on lung tissue sections from a representative M. tuberculosis-infected (H37Rv, 14 wpi, without TB treatment) mouse with a cavitarylesion (arrow). (A) Expression of MMP-9 was noted at the sites of infection and colocalized with infected areas and the CD11b+ signal. Higher-power viewsdemonstrate CD11b+ and MMP-9 expression in the cellular rim surrounding cavities (B) and in the adjacent pneumonia (C). Numerous bacteria were noted inthese lesions.
785
RESEARCH ARTICLE Disease Models & Mechanisms (2016) 9, 779-788 doi:10.1242/dmm.025643
Disea
seModels&Mechan
isms

highly specific MMP-9 inhibitor, SB-3CT (Gu et al., 2005). Theseand other MMP-9 inhibitors warrant evaluation as inhibitors ofcavitation and lung destruction during TB infection. We and othershave previously demonstrated that necrotic TB lesions in C3HeB/FeJ mice are hypoxic (Driver et al., 2012; Harper et al., 2012), andMMP-7 is induced by hypoxia (Burke et al., 2003). Therefore, wealso investigated MMP-7 in this model, which was similarlyexpressed surrounding cavitary and in and around necrotic lesions.Other MMPs (e.g. MMP-8) (Sathyamoorthy et al., 2015) andproteases such as cathepsins (Converse et al., 1996; Kübler et al.,2016, 2015) may also have significant roles, and their evaluationwill be the focus of future studies. Our results indicate that C3HeB/FeJ mice could also be useful to develop novel host-directed TBtreatments, especially those targeting MMPs.Given the well-established role of macrophages and growing
evidence that neutrophils are also key mediators of inflammation innecrotic and cavitary TB lesions (Berry et al., 2010; Dorhoi andKaufmann, 2016; Eum et al., 2010; Marzo et al., 2014), weevaluated the spatial location of MMPs and their relationship tomacrophages and neutrophils in situ in three dimensions. BecauseMMPs are secreted extracellularly, they are not easily amenable toflow-cytometric analyses. Therefore, we immunostained anddigitally scanned 36 consecutive sections through a pulmonarycavitary lesion from a representative M. tuberculosis-infectedmouse and re-constructed a three-dimensional stack with multiplechannels. We analyzed high-resolution images from severalconsecutive sections across macroscopic dimensions to control forsampling bias. MMP-9 and MMP-7 demonstrated significantcolocalization with CD11b+ cells (Chi-square P<0.01),suggesting that macrophages are an important source of MMP-7and MMP-9 in this model, and consistent with published literature(Volkman et al., 2010; Volpe et al., 2006).In summary, we have demonstrated that C3HeB/FeJ mice
reliably develop cavitary pulmonary TB after aerosol infectionwith M. tuberculosis. Serial CT imaging could non-invasivelydetect cavitary lesions in vivo and their communication with majorairways. Post-mortem analyses demonstrated that pulmonarylesions in C3HeB/FeJ mice share several key pathological featuresof human TB, including the presence of multinucleated giantcells, and the simultaneous presence of multiple pathologies –granulomatous pneumonia, necrosis and cavitation – in the sameanimal. MMP-9 is significantly expressed in cavitary and necroticlesions, and predominantly associated with CD11b+ cells. C3HeB/FeJ mice present an economical and tractable model of cavitary TB,and warrant further development as a new tool for studying thepathogenesis of cavitation and its effects on TB treatment.
MATERIALS AND METHODSAll protocols were approved by the Johns Hopkins Biosafety, RadiationSafety, and Animal Care and Use Committees.
Mycobacterial strainsM. tuberculosis H37Rv and HN878 were used as frozen stocks preparedfrom a log-phase culture in Middlebrook 7H9 broth after mouse passage andwere diluted in phosphate buffered saline (PBS) before infection.
Animal infection and treatmentsFour- to six-week-old female C3HeB/FeJ mice (Jackson Laboratories, BarHarbor, ME) were aerosol-infected with frozen stocks of M. tuberculosisusing the Middlebrook Inhalation Exposure System (Glas-Col, Terre Haute,IN) (Harper et al., 2012). At least 3micewere sacrificed the day after infectionand at subsequent pre-determined time points to determine CFU counts in thelungs. Some cohorts received rifampin (10 mg/kg body weight), isoniazid
(10 mg/kg) and pyrazinamide (150 mg/kg) (RHZ) once daily, by gavage, 5days per week for 8-12 weeks, beginning 6 wpi, and followed aftercompleting treatment to detect cavitation during relapse. To assess theeffect of prior sensitization with mycobacterial antigens on cavity formation,some cohorts received 5 subcutaneous injections of approximately 7 log10CFU of heat-killedM. tuberculosisHN878 in incomplete Freund’s adjuvanttwice weekly beginning 45 days before aerosol infection with the same strain(Nedeltchev et al., 2009).
ImagingLive M. tuberculosis-infected mice were serially imaged within a sealedbiocontainment device (Minerve, Esternay, France) modified in-house tocomply with biosafety level-3 (BSL-3) containment (Davis et al., 2009;Ordonez et al., 2015b; Weinstein et al., 2012). Each mouse was imagedusing the NanoSPECT/CT small animal imager (Bioscan, Washington,DC). Images were reconstructed and visualized using VivoQuant 2.50(inviCRO, Boston, MA) or Amira 5.2.1 (FEI, Hillsboro, OR). A cavity wasdefined as a macroscopic region of air (density<−900 Hounsfield units)within diseased lung parenchyma.
Ex vivo optical imagingLungs from M. tuberculosis-infected (14 wpi) and age-matched uninfected(control) micewere harvested and imaged in sealed transparent polycarbonatecontainers using an IVIS Lumina II imaging system (PerkinElmer, Waltham,MA) 24 h after intravenous injection of MMPSense® 680 FluorescenceImaging Agent (PerkinElmer), an in vivo imaging agent activated primarilybyMMP-2,MMP-3,MMP-9 andMMP-13 (Jager et al., 2016). The datawereanalyzed using Living Image software (PerkinElmer).
HistopathologyLungs were harvested after systemic perfusion with PBS under deepanesthesia, fixed in 4% paraformaldehyde and sectioned to 5-μm thickness.Hematoxylin-eosin, acid-fast, Masson’s Trichrome, reticulin and PicrosiriusRed staining was performed following standard procedures (Ordonez et al.,2015b). A cavity was defined by visualization of an air-filled lesion on grossand histopathological assessment.
ImmunohistochemistryParaffin-embedded sections were rehydrated in graded alcohols, steamed incitrate buffer at pH 6 and probed at room temperature for 2 h for the following:macrophages [rat monoclonal (M1/70) to CD11b+; 1:500; Abcam],granulocytes [rat monoclonal (NIMP-R14) to Gr-1+; 1:500; Novusbio], andMMP-7 and MMP-9 (rabbit polyclonal; 1:250; Abcam). Both macrophagesand granulocytes were then processed usingVECTASTAINAP-Red, (VectorLaboratories, Burlingame, CA) whereas MMPs were processed with apolymer-HRP kit (BioGenex, San Ramon, CA) with diaminobenzidinedevelopment and Mayer hematoxylin counterstaining. Lungs from untreatedanimals, both uninfected and infected, without primary antibody served asnegative controls. Slides were scanned using anApeiro digital scanner (Leica,Buffalo Grove, IL).
Gelatin zymographyMMP-9 activity was analyzed in frozen lung tissue homogenates. Thesamples were thawed and incubated in Triton X-100 to release MMPs andrun on a 7.5% SDS-polyacrylamide gel electrophoresis with a gelatinsubstrate (Snoek-van Beurden and Von den Hoff, 2005). Recombinanthuman MMPs (rhMMP-9 and rhMMP-2; 10 ng/ml) were loaded ascontrols. Incubation of lung homogenates from M. tuberculosis-infectedanimals with inhibitors of MMP activity was also performed.
In situ spatial localization of MMP signal with inflammatory cellsWe analyzed high-resolution data from several consecutive sections acrossmacroscopic data to control for sampling bias. Thirty-six consecutivesections (each 5 µm) through a pulmonary cavity from a representativeM. tuberculosis-infected mouse were immunostained, digitally scanned athigh resolution (Aperio Digital Pathology Slide Scanner, Leica Biosystems,Buffalo Grove, IL) and reconstructed in three dimensions. Each section was
786
RESEARCH ARTICLE Disease Models & Mechanisms (2016) 9, 779-788 doi:10.1242/dmm.025643
Disea
seModels&Mechan
isms

manually reviewed for staining characteristics. To analyze the colocalizationof CD11b+ and Gr-1+ with MMPs, sequential sections were spatiallyregistered. Immunostains were first converted to luminance, and an affineregistration algorithm was utilized based on mutual information (Matlab,MathWorks, Natick, MA). Images were visualized using Amira 5.2.1 (FEI,Hillsboro, OR). A minimum of 300,000 voxels (three-dimensional pixels)were counted for each channel.
Statistical analysisSurvival was compared using the log-rank (Mantel–Cox) test. Lung CFUcounts were log-transformed and expressed as mean and standard deviation(s.d.). Group means were compared using the Mann–Whitney test.Proportions were compared using two-tailed Chi-square with Yates’correction or Fisher’s exact test. All analyses were performed using Prism6.0 (GraphPad, San Diego, CA).
Competing interestsThe authors declare no competing or financial interests.
Author contributionsA.A.O., S.P., E.L.N. and S.K.J. designed the research. A.A.O., R.T. and S.P.performed the animal experiments. M.H.K. performed imaging, and A.A.O. andS.K.J. analyzed the imaging data. S.P. performed the immunohistochemistry andother immunological assays. Z.X. and D.J.M. performed image analyses for spatiallocation of signals. A.A.O., R.T., S.P., P.J.C., E.L.N. and S.K.J. analyzed the data.A.A.O., E.L.N. and S.K.J. wrote the initial draft, and all coauthors participated in theediting of the report. E.L.N. and S.K.J. provided funding and supervised the project.
FundingThis work was supported by the National Institutes of Health [Director’sTransformative Research Award R01-EB-020539 to S.K.J., Director’s NewInnovator Award DP2-OD-006492 to S.K.J., R01-HL-116316 to S.K.J. and R01-AI-111992 to E.L.N.], the Bill and Melinda Gates Foundation [OPP1037174 to E.L.N.]and the U.S. Food and Drug Administration [U18-FD-004004 to E.L.N.].
Supplementary informationSupplementary information available online athttp://dmm.biologists.org/lookup/doi/10.1242/dmm.025643.supplemental
ReferencesAbhimanyu, Jha, P., Jain, A., Arora, K. and Bose, M. (2011). Genetic associationstudy suggests a role for SP110 variants in lymph node tuberculosis but notpulmonary tuberculosis in north Indians. Hum. Immunol. 72, 576-580.
Benator, D., Bhattacharya, M., Bozeman, L., Burman, W., Cantazaro, A.,Chaisson, R., Gordin, F., Horsburgh, C. R., Horton, J., Khan, A. et al.,Tuberculosis Trials Consortium (2002). Rifapentine and isoniazid once a weekversus rifampicin and isoniazid twice a week for treatment of drug-susceptiblepulmonary tuberculosis in HIV-negative patients: a randomised clinical trial.Lancet 360, 528-534.
Berry, M. P. R., Graham, C. M., McNab, F. W., Xu, Z., Bloch, S. A. A., Oni, T.,Wilkinson, K. A., Banchereau, R., Skinner, J., Wilkinson, R. J. et al. (2010). Aninterferon-inducible neutrophil-driven blood transcriptional signature in humantuberculosis. Nature 466, 973-977.
Blumberg, H. M., Burman, W. J., Chaisson, R. E., Daley, C. L., Etkind, S. C.,Friedman, L. N., Fujiwara, P., Grzemska, M., Hopewell, P. C., Iseman, M. D.et al., American Thoracic Society, Centers for Disease Control andPrevention and the Infectious Diseases Society (2003). American ThoracicSociety/Centers for Disease Control and Prevention/Infectious Diseases Societyof America: treatment of tuberculosis. Am. J. Respir. Crit. Care Med. 167,603-662.
Burke, B., Giannoudis, A., Corke, K. P., Gill, D., Wells, M., Ziegler-Heitbrock, L.and Lewis, C. E. (2003). Hypoxia-induced gene expression in humanmacrophages: implications for ischemic tissues and hypoxia-regulated genetherapy. Am. J. Pathol. 163, 1233-1243.
Canetti, G. (1955). The Tubercle Bacillus in the Pulmonary Lesion of Man.New York, NY: Springer Publishing Co., Inc.
Canetti, G. (1965). Present aspects of bacterial resistance in tuberculosis. Am. Rev.Respir. Dis. 92, 687-703.
Capuano, S. V., III, Croix, D. A., Pawar, S., Zinovik, A., Myers, A., Lin, P. L.,Bissel, S., Fuhrman, C., Klein, E. and Flynn, J. L. (2003). ExperimentalMycobacterium tuberculosis infection of cynomolgus macaques closelyresembles the various manifestations of human M. tuberculosis infection. Infect.Immun. 71, 5831-5844.
Chen, R. Y., Dodd, L. E., Lee, M., Paripati, P., Hammoud, D. A., Mountz, J. M.,Jeon, D., Zia, N., Zahiri, H., Coleman, M. T. et al. (2014). PET/CT imagingcorrelates with treatment outcome in patients with multidrug-resistanttuberculosis. Sci. Transl. Med. 6, 265ra166.
Converse, P. J., Dannenberg, A. M., Jr., Estep, J. E., Sugisaki, K., Abe, Y.,Schofield, B. H. and Pitt, M. L. (1996). Cavitary tuberculosis produced in rabbitsby aerosolized virulent tubercle bacilli. Infect. Immun. 64, 4776-4787.
Davis, S. L., Nuermberger, E. L., Um, P. K., Vidal, C., Jedynak, B., Pomper,M. G., Bishai, W. R. and Jain, S. K. (2009). Noninvasive pulmonary [18F]-2-fluoro-deoxy-D-glucose positron emission tomography correlates withbactericidal activity of tuberculosis drug treatment. Antimicrob. AgentsChemother. 53, 4879-4884.
DeMarco, V. P., Ordonez, A. A., Klunk, M., Prideaux, B., Wang, H., Zhuo, Z.,Tonge, P. J., Dannals, R. F., Holt, D. P., Lee, C. K. et al. (2015). Determination of[11C]rifampin pharmacokinetics within Mycobacterium tuberculosis-infectedmiceby using dynamic positron emission tomography bioimaging. Antimicrob. AgentsChemother. 59, 5768-5774.
Dorhoi, A. and Kaufmann, S. H. E. (2016). Pathology and immune reactivity:understanding multidimensionality in pulmonary tuberculosis. Semin.Immunopathol. 38, 153-166.
Dorman, S. E., Johnson, J. L., Goldberg, S., Muzanye, G., Padayatchi, N.,Bozeman, L., Heilig, C. M., Bernardo, J., Choudhri, S., Grosset, J. H. et al.(2009). Substitution of moxifloxacin for isoniazid during intensive phase treatmentof pulmonary tuberculosis. Am. J. Respir. Crit. Care Med. 180, 273-280.
Driver, E. R., Ryan, G. J., Hoff, D. R., Irwin, S.M., Basaraba, R. J., Kramnik, I. andLenaerts, A. J. (2012). Evaluation of a mouse model of necrotic granulomaformation using C3HeB/FeJ mice for testing of drugs against Mycobacteriumtuberculosis. Antimicrob. Agents Chemother. 56, 3181-3195.
Elkington, P., Shiomi, T., Breen, R., Nuttall, R. K., Ugarte-Gil, C. A., Walker, N. F.,Saraiva, L., Pedersen, B., Mauri, F., Lipman, M. et al. (2011). MMP-1 drivesimmunopathology in human tuberculosis and transgenic mice. J. Clin. Invest. 121,1827-1833.
Eum, S.-Y., Kong, J.-H., Hong, M.-S., Lee, Y.-J., Kim, J.-H., Hwang, S.-H., Cho,S.-N., Via, L. E. and Barry, C. E.III (2010). Neutrophils are the predominantinfected phagocytic cells in the airways of patients with active pulmonary TB.Chest 137, 122-128.
Gendron, R., Grenier, D., Sorsa, T. and Mayrand, D. (1999). Inhibition of theactivities of matrix metalloproteinases 2, 8, and 9 by chlorhexidine. Clin. Diagn.Lab. Immunol. 6, 437-439.
Gioia, M., Monaco, S., VanDenSteen, P. E., Sbardella, D., Grasso, G., Marini, S.,Overall, C. M., Opdenakker, G. and Coletta, M. (2009). The collagen bindingdomain of gelatinase A modulates degradation of collagen IV by gelatinase B.J. Mol. Biol. 386, 419-434.
Greenlee, K. J.,Werb, Z. andKheradmand, F. (2007). Matrix metalloproteinases inlung: multiple, multifarious, and multifaceted. Physiol. Rev. 87, 69-98.
Gu, Z., Cui, J., Brown, S., Fridman, R., Mobashery, S., Strongin, A. Y. andLipton, S. A. (2005). A highly specific inhibitor of matrix metalloproteinase-9rescues laminin from proteolysis and neurons from apoptosis in transient focalcerebral ischemia. J. Neurosci. 25, 6401-6408.
Harper, J., Skerry, C., Davis, S. L., Tasneen, R., Weir, M., Kramnik, I., Bishai,W. R., Pomper, M. G., Nuermberger, E. L. and Jain, S. K. (2012). Mouse modelof necrotic tuberculosis granulomas develops hypoxic lesions. J. Infect. Dis. 205,595-602.
He, X., Berland, R., Mekasha, S., Christensen, T. G., Alroy, J., Kramnik, I. andIngalls, R. R. (2013). The sst1 resistance locus regulates evasion of type Iinterferon signaling by Chlamydia pneumoniae as a disease tolerancemechanism. PLoS Pathog. 9, e1003569.
Hunter, R. L. (2011). Pathology of post primary tuberculosis of the lung: anillustrated critical review. Tuberculosis 91, 497-509.
Irwin, S. M., Driver, E., Lyon, E., Schrupp, C., Ryan, G., Gonzalez-Juarrero, M.,Basaraba, R. J., Nuermberger, E. L. and Lenaerts, A. J. (2015). Presence ofmultiple lesion types with vastly different microenvironments in C3HeB/FeJ micefollowing aerosol infection with Mycobacterium tuberculosis. Dis. Model. Mech. 8,591-602.
Irwin, S. M., Prideaux, B., Lyon, E. R., Zimmerman, M. D., Brooks, E. J.,Schrupp, C. A., Chen, C., Reichlen, M. J., Asay, B. C., Voskuil, M. I. et al.(2016). Bedaquiline and pyrazinamide treatment responses are affected bypulmonary lesion heterogeneity in Mycobacterium tuberculosis infected C3HeB/FeJ mice. ACS Infect. Dis. 2, 251-267.
Jager, N. A., de Vries, B. M.W., Hillebrands, J.-L., Harlaar, N. J., Tio, R. A., Slart,R. H. J. A., van Dam, G. M., Boersma, H. H., Zeebregts, C. J. and Westra, J.(2016). Distribution of matrix metalloproteinases in human atherosclerotic carotidplaques and their production by smooth muscle cells and macrophage subsets.Mol. Imaging Biol. 18, 238-291.
Kaplan, G., Post, F. A., Moreira, A. L., Wainwright, H., Kreiswirth, B. N.,Tanverdi, M., Mathema, B., Ramaswamy, S. V., Walther, G., Steyn, L. M. et al.(2003). Mycobacterium tuberculosis growth at the cavity surface: amicroenvironment with failed immunity. Infect. Immun. 71, 7099-7108.
Kaushal, D., Mehra, S., Didier, P. and Lackner, A. (2012). The non-human primatemodel of tuberculosis. J. Med. Primatol. 41, 191-201.
787
RESEARCH ARTICLE Disease Models & Mechanisms (2016) 9, 779-788 doi:10.1242/dmm.025643
Disea
seModels&Mechan
isms

Kjellsson, M. C., Via, L. E., Goh, A., Weiner, D., Low, K. M., Kern, S., Pillai, G.,Barry, C. E., III and Dartois, V. (2012). Pharmacokinetic evaluation of thepenetration of antituberculosis agents in rabbit pulmonary lesions. Antimicrob.Agents Chemother. 56, 446-457.
Kubler, A., Luna, B., Larsson, C., Ammerman, N. C., Andrade, B. B., Orandle,M., Bock, K. W., Xu, Z., Bagci, U., Molura, D. J. et al. (2015). Mycobacteriumtuberculosis dysregulates MMP/TIMP balance to drive rapid cavitation andunrestrained bacterial proliferation. J. Pathol. 235, 431-444.
Kubler, A., Larsson, C., Luna, B., Andrade, B. B., Amaral, E. P., Urbanowski, M.,Orandle, M., Bock, K., Ammerman, N. C., Cheung, L. S. et al. (2016).Cathepsin K contributes to cavitation and collagen turnover in pulmonarytuberculosis. J. Infect. Dis. 213, 618-627.
Lanoix, J.-P., Lenaerts, A. J. and Nuermberger, E. L. (2015). Heterogeneousdisease progression and treatment response in a C3HeB/FeJ mouse model oftuberculosis. Dis. Models Mech. 8, 603-610.
Lanoix, J.-P., Betoudji, F. and Nuermberger, E. L. (2016a). Sterilizing activity ofpyrazinamide in combination with first-line drugs in a C3HeB/FeJ mouse model oftuberculosis. Antimicrob. Agents Chemother. 60, 1091-1096.
Lanoix, J.-P., Ioerger, T., Ormond, A., Kaya, F., Sacchettini, J., Dartois, V. andNuermberger, E. (2016b). Selective inactivity of pyrazinamide againsttuberculosis in C3HeB/FeJ mice is best explained by neutral pH of caseum.Antimicrob. Agents Chemother. 60, 735-743.
Lenaerts, A., Barry, C. E., III and Dartois, V. (2015). Heterogeneity in tuberculosispathology, microenvironments and therapeutic responses. Immunol. Rev. 264,288-307.
Lin, P. L., Coleman, T., Carney, J. P. J., Lopresti, B. J., Tomko, J., Fillmore, D.,Dartois, V., Scanga, C., Frye, L. J., Janssen, C. et al. (2013). Radiologicresponses in cynomolgus macaques for assessing tuberculosis chemotherapyregimens. Antimicrob. Agents Chemother. 57, 4237-4244.
Liu, J., Xiong, W., Baca-Regen, L., Nagase, H. and Baxter, B. T. (2003).Mechanism of inhibition of matrix metalloproteinase-2 expression by doxycyclinein human aortic smooth muscle cells. J. Vasc. Surg. 38, 1376-1383.
Manabe, Y. C., Dannenberg, A. M., Jr., Tyagi, S. K., Hatem, C. L., Yoder, M.,Woolwine, S. C., Zook, B. C., Pitt, M. L. M. and Bishai, W. R. (2003). Differentstrains of Mycobacterium tuberculosis cause various spectrums of disease in therabbit model of tuberculosis. Infect. Immun. 71, 6004-6011.
Marzo, E., Vilaplana, C., Tapia, G., Diaz, J., Garcia, V. and Cardona, P.-J. (2014).Damaging role of neutrophilic infiltration in a mouse model of progressivetuberculosis. Tuberculosis 94, 55-64.
Miller, W. T. and Miller, W. T. Jr. (1993). Tuberculosis in the normal host:radiological findings. Semin. Roentgenol. 28, 109-118.
Nedeltchev, G. G., Raghunand, T. R., Jassal, M. S., Lun, S., Cheng, Q.-J. andBishai, W. R. (2009). Extrapulmonary dissemination of Mycobacterium bovis butnot Mycobacterium tuberculosis in a bronchoscopic rabbit model of cavitarytuberculosis. Infect. Immun. 77, 598-603.
Nuermberger, E. L. (2008). Using animal models to develop new treatments fortuberculosis. Semin. Respir. Crit. Care Med. 29, 542-551.
Ong, C. W. M., Elkington, P. T. and Friedland, J. S. (2014). Tuberculosis,pulmonary cavitation, and matrix metalloproteinases. Am. J. Respir. Crit. CareMed. 190, 9-18.
Ordonez, A. A., DeMarco, V. P., Klunk, M. H., Pokkali, S. and Jain, S. K. (2015a).Imaging chronic tuberculous lesions using sodium [18F]fluoride positron emissiontomography in mice. Mol. Imaging Biol. 17, 609-614.
Ordonez, A. A., Pokkali, S., DeMarco, V. P., Klunk, M., Mease, R. C., Foss, C. A.,Pomper, M. G. and Jain, S. K. (2015b). Radioiodinated DPA-713 imagingcorrelates with bactericidal activity of tuberculosis treatments in mice. Antimicrob.Agents Chemother. 59, 642-649.
Palaci, M., Dietze, R., Hadad, D. J., Ribeiro, F. K. C., Peres, R. L., Vinhas, S. A.,Maciel, E. L. N., do Valle Dettoni, V., Horter, L., Boom, W. H. et al. (2007).Cavitary disease and quantitative sputum bacillary load in cases of pulmonarytuberculosis. J. Clin. Microbiol. 45, 4064-4066.
Pan, H., Yan, B.-S., Rojas, M., Shebzukhov, Y. V., Zhou, H., Kobzik, L., Higgins,D. E., Daly, M. J., Bloom, B. R. and Kramnik, I. (2005). Ipr1 gene mediatesinnate immunity to tuberculosis. Nature 434, 767-772.
Pichugin, A. V., Yan, B.-S., Sloutsky, A., Kobzik, L. and Kramnik, I. (2009).Dominant role of the sst1 locus in pathogenesis of necrotizing lung granulomasduring chronic tuberculosis infection and reactivation in genetically resistanthosts. Am. J. Pathol. 174, 2190-2201.
Prideaux, B., Via, L. E., Zimmerman, M. D., Eum, S., Sarathy, J., O’Brien, P.,Chen, C., Kaya, F., Weiner, D. M., Chen, P.-Y. et al. (2015). The associationbetween sterilizing activity and drug distribution into tuberculosis lesions. Nat.Med. 21, 1223-1227.
Salazar-Austin, N., Ordonez, A. A., Hsu, A. J., Benson, J. E., Mahesh, M.,Menachery, E., Razeq, J. H., Salfinger, M., Starke, J. R., Milstone, A. M. et al.(2015). Extensively drug-resistant tuberculosis in a young child after travel toIndia. Lancet Infect. Dis. 15, 1485-1491.
Salgame, P. (2011). MMPs in tuberculosis: granuloma creators and tissuedestroyers. J. Clin. Invest. 121, 1686-1688.
Sathyamoorthy, T., Sandhu, G., Tezera, L. B., Thomas, R., Singhania, A.,Woelk,C. H., Dimitrov, B. D., Agranoff, D., Evans, C. A. W., Friedland, J. S. et al.(2015). Gender-dependent differences in plasma matrix metalloproteinase-8elevated in pulmonary tuberculosis. PLoS ONE 10, e0117605.
Singh, S., Kubler, A., Singh, U. K., Singh, A., Gardiner, H., Prasad, R.,Elkington, P. T. and Friedland, J. S. (2014). Antimycobacterial drugs modulateimmunopathogenic matrix metalloproteinases in a cellular model of pulmonarytuberculosis. Antimicrob. Agents Chemother. 58, 4657-4665.
Snoek-van Beurden, P. A. and Von den Hoff, J. W. (2005). Zymographictechniques for the analysis of matrix metalloproteinases and their inhibitors.Biotechniques 38, 73-83.
Subbian, S., Tsenova, L., Yang, G., O’Brien, P., Parsons, S., Peixoto, B., Taylor,L., Fallows,D. andKaplan, G. (2011). Chronic pulmonary cavitary tuberculosis inrabbits: a failed host immune response. Open Biol. 1, 110016.
Tosh, K., Campbell, S. J., Fielding, K., Sillah, J., Bah, B., Gustafson, P.,Manneh, K., Lisse, I., Sirugo, G., Bennett, S. et al. (2006). Variants in the SP110gene are associated with genetic susceptibility to tuberculosis in West Africa.Proc. Natl. Acad. Sci. USA 103, 10364-10368.
Via, L. E., Schimel, D., Weiner, D. M., Dartois, V., Dayao, E., Cai, Y., Yoon, Y.-S.,Dreher, M. R., Kastenmayer, R. J., Laymon, C. M. et al. (2012). Infectiondynamics and response to chemotherapy in a rabbit model of tuberculosis using[18F]2-fluoro-deoxy-D-glucose positron emission tomography and computedtomography. Antimicrob. Agents Chemother. 56, 4391-4402.
Via, L. E., Weiner, D. M., Schimel, D., Lin, P. L., Dayao, E., Tankersley, S. L., Cai,Y., Coleman,M. T., Tomko, J., Paripati, P. et al. (2013). Differential virulence anddisease progression following Mycobacterium tuberculosis complex infection ofthe common marmoset (Callithrix jacchus). Infect. Immun. 81, 2909-2919.
Volkman, H. E., Pozos, T. C., Zheng, J., Davis, J. M., Rawls, J. F. andRamakrishnan, L. (2010). Tuberculous granuloma induction via interaction of abacterial secreted protein with host epithelium. Science 327, 466-469.
Volpe, E., Cappelli, G., Grassi, M., Martino, A., Serafino, A., Colizzi, V.,Sanarico, N. and Mariani, F. (2006). Gene expression profiling of humanmacrophages at late time of infection with Mycobacterium tuberculosis.Immunology 118, 449-460.
Weinstein, E. A., Liu, L., Ordonez, A. A., Wang, H., Hooker, J. M., Tonge, P. J. andJain, S. K. (2012). Noninvasive determination of 2-[18F]-fluoroisonicotinic acidhydrazide pharmacokinetics by positron emission tomography in Mycobacteriumtuberculosis-infected mice. Antimicrob. Agents Chemother. 56, 6284-6290.
Wells, W. F. and Lurie, M. B. (1941). Experimental airborne disease: quantitativenatural respiratory contagion of tuberculosis. Am. J. Epidemiol. 34, 21-40.
WHO (2011). The Immunological Basis for Immunization Series. Module 5:Tuberculosis. Geneva, Switzerland: World Health Organization.
Yamamura, Y. (1958). The pathogenesis of tuberculous cavities. Bibl. Tuberc. 13,13-37.
Yan, B.-S., Pichugin, A. V., Jobe, O., Helming, L., Eruslanov, E. B., Gutierrez-Pabello, J. A., Rojas, M., Shebzukhov, Y. V., Kobzik, L. and Kramnik, I. (2007).Progression of pulmonary tuberculosis and efficiency of bacillus Calmette-Guerinvaccination are genetically controlled via a common sst1-mediated mechanism ofinnate immunity. J. Immunol. 179, 6919-6932.
788
RESEARCH ARTICLE Disease Models & Mechanisms (2016) 9, 779-788 doi:10.1242/dmm.025643
Disea
seModels&Mechan
isms