motion under the microscope: modern techniques...
TRANSCRIPT

MOTION UNDER THE MICROSCOPE: MODERN TECHNIQUES FOR STUDYING CELL ADHESION AND MOTILITY
Paweł Pomorski
Laboratory of the Molecular Basis of Cell Motility
Nencki Institute of Experimental Biology, Warsaw, Poland
Ability to move is one of the fundamental functions of the living cells. It is due to the motility that organism develops, immune system can work, organs are able to regenerate and wound heal. In the same time motility studies are among methodologically most difficult ones. Biochemical processes underlying motility are notoriously unsynchronized and motile cells are usually not very numerous. Current paper reviews microscope techniques developed to solve those problems. We discuss basic measurements, parameterizing motility and substratum adhesion. Classical, structural microscopy used for motility studies are also sketched shortly. We describe also use of molecular probes for signaling studies in motile cells as well as we mention about microscopic experimental techniques, allowing experiments on single cells.

Ruch i adhezja komórek – metody mikroskopowe
Paweł Pomorski 18.06.2013

1. Obserwacja i parametryzacja ruchu komórek
2. Pomiar adhezji komórek 3. Zastosowanie w praktyce
doświadczalnej

ŚLEDZENIE KOMÓREK

Kontrast fazowy
0
20
40
60
80
100
120
140
160
180
200
0 200 400 600 800 1000
DIC Nomarski
60
80
100
120
140
160
180
200
0 50 100 150 200 250 300 350 400

M. fluorescencyjna
0
20
40
60
80
100
120
140
160
0 50 100 150 200

CrossCorrelation Tracking

CrossCorrelation Tracking

-170.5764 -85.2882 85.2882 170.5764
-170.5764
-85.2882
0
85.2882
170.5764
Trajektorie

Trajektorie
-170.5764 -85.2882 0 85.2882 170.5764
-170.5764
-85.2882
0
85.2882
170.5764

Confocal microscope Leica TCS LSI - practical applications 1Płatek R., 2Korczyński J. and 1Skup M.
1Laboratory for Reinnervation Processes and 2Laboratory for Confocal Microscopy
Nencki Institute of Experimental Biology, Warsaw, Poland.
Conventional confocal microscopy is dedicated mainly to proceed with thin tissue sections and cells seeded on a cover slip. The Leica TCS LSI macro confocal is the first super zoom microscope that combines all benefits of traditional confocal microscopy with large scale imaging of anesthetized, alive objects as well as unfixed/fixed objects post mortem. In our experiments we have tested Leica TCS LSI for scanning of (1) murine brains and spinal cords in vivo, and dissected in toto immediately post mortem, and (2) fixed rat spinal cords. We used transgenic mice expressing green fluorescent protein (GFP) under PLP promoter, to visualize oligodendrocytes, and rats with spinal cords transduced with AAV vector coding for enhanced GFP under mCMV promoter, to visualize neurons and glia. Super zoom confocal microscopy let us observe general distribution of GFP- expressing cells in whole organs as well as to focus on single cells and fibers. We were able to discriminate well between main morphological features of these cells. In murine brains we could visualize myelinated axons and oligodendrocytes, whereas in rat spinal cords the extent of eGFP expressing cells and their fibers traversing along entire spinal cord could be traced. The labeled objects could be visualized from the regions lying within a range of 100 μm from the surface of the brain/spinal cord. The details on the procedures applied, benefits and limitations of the method will be presented.

Mikroskop konfokalny Leica TCS LSI -
- przykłady zastosowania
Mikroskopia konfokalna - Praktyczny Kurs Badań In vivo
18-19.06.2013
Rafał Płatek; Pracownia Procesów Reinerwacyjnych

Mikroskopia konfokalna

?
Mikroskopia konfokalna

Scanner Method
Confocal channels
Scanner galvo (x,y)
Sequential scan
Channel multiplexing
Scan formats [pixel]
Image depth [bit]
Spectral detection
Spectral bandwidth [nm]
Detector
Detector type
true confocal
1
yes
1 – 8 sequential
128, 256, 512,1024, 2048
8 or 12, switchable
yes
430 – 750
1
ultra high dynamic PMT
Confocal Zoom
Zoom range [x]
Optical Zoom (Z16 APO)
continuously variable
1x – 16x
0.57 – 9.2x
Laser type
Number of lasers
Laser options [nm]
Excitation attenuation
solid state
max 4
405, 488, 532 or 561, 635
AOTF
Macro-Objectives
Working distance [mm]
Micro-Objectives
1x, 2x, 5x
97/39/19
10x, 20x, 40x, 63x
Mikroskopia konfokalna Parametry Leica TCS LSI

Model doświadczalny

Lezja
Miejsce iniekcji - transdukowane komórki
Rdzeń kręgowy szczura preparat - R. Platek
zdjęcie – J.Korczynski + R. Platek
Dane niepublikowane
Instytut IBD Nenckiego
Aksony transdukowanych komórek
Miejsce iniekcji - transdukowane komórki
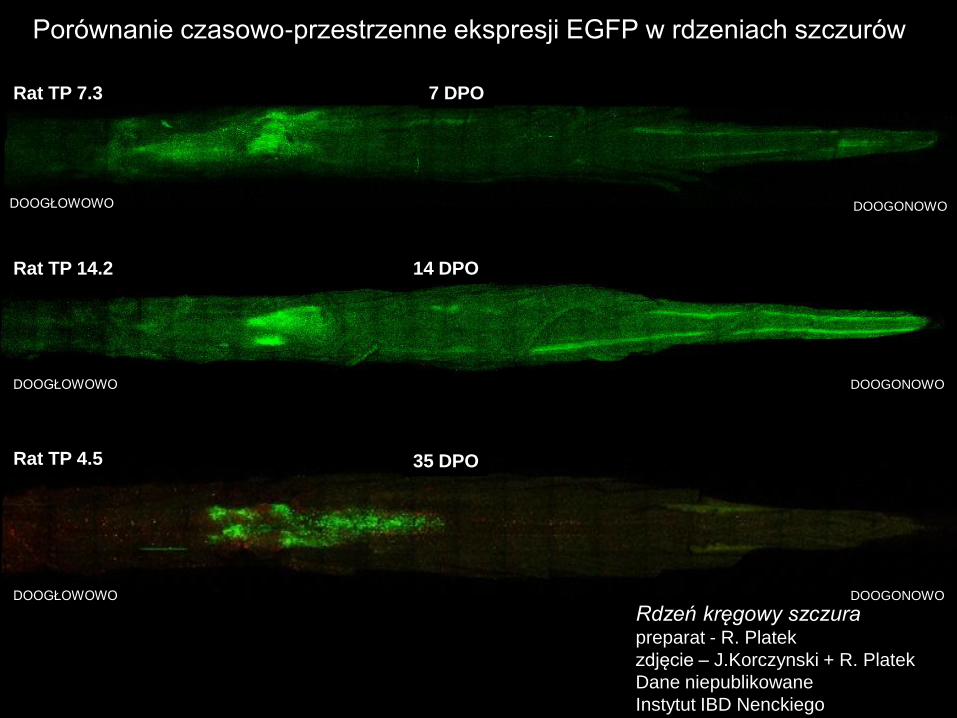
DOOGŁOWOWO DOOGONOWO
7 DPO Rat TP 7.3
DOOGŁOWOWO DOOGONOWO
14 DPO Rat TP 14.2
DOOGŁOWOWO DOOGONOWO
35 DPO Rat TP 4.5
Porównanie czasowo-przestrzenne ekspresji EGFP w rdzeniach szczurów
Rdzeń kręgowy szczura preparat - R. Platek
zdjęcie – J.Korczynski + R. Platek
Dane niepublikowane
Instytut IBD Nenckiego
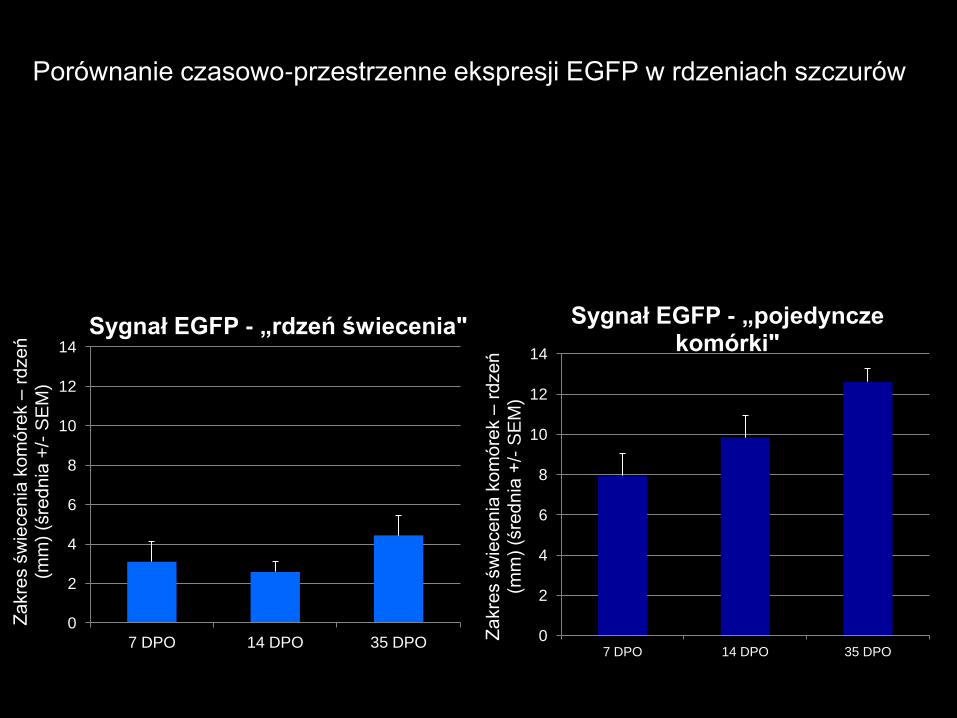
0
2
4
6
8
10
12
14
7 DPO 14 DPO 35 DPO
Za
kre
s ś
wie
ce
nia
ko
mó
rek –
rd
ze
ń
(mm
) (ś
red
nia
+/-
SE
M)
Sygnał EGFP - „rdzeń świecenia"
0
2
4
6
8
10
12
14
7 DPO 14 DPO 35 DPO
Za
kre
s ś
wie
ce
nia
ko
mó
rek –
rd
ze
ń
(mm
) (ś
red
nia
+/-
SE
M)
Sygnał EGFP - „pojedyncze komórki"
Porównanie czasowo-przestrzenne ekspresji EGFP w rdzeniach szczurów
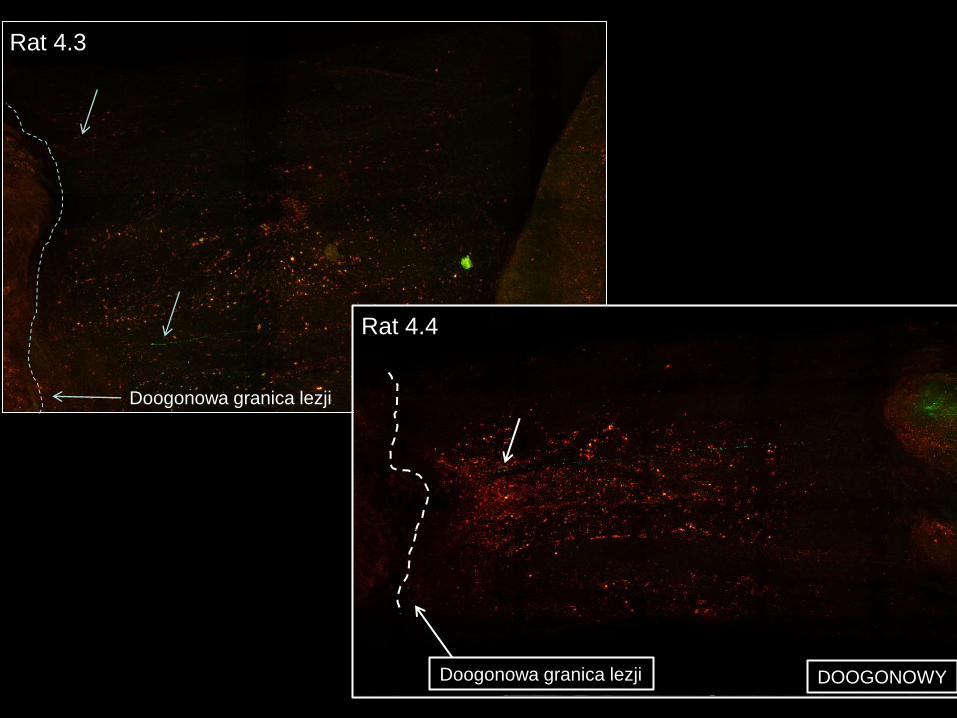
Rat 4.3
Doogonowa granica lezji DOOGONOWY
DOOGONOWY Doogonowa granica lezji
Rat 4.4

Leica TCS LSI – obrazowanie in vivo

Mózg mysi postmortem. preparat - R.Platek,
zdjęcie - J.Korczynski + R.Platek
Dane nieopublikowane
Instytut IBD Nenckiego

komórka mitralna
kłębuszek (glomerulus)
opuszka węchowa
kość
nabłonek węchowy
węchowe komórki receptorowe
węchowe komórki receptorowe są aktywowane i przesyłają pobudzenie
sygnał przekazywany jest w kłębuszkach
sygnał przesyłany jest do wyższych struktur mózgowych
Budowa opuszki węchowej

Źródło - Wikimedia Commons
Czerwony – warstwa
komórek mitralnych z
ciałami komórek
mitralnych i ziarnistych
w tej warstwie
Zielony – warstwa
komórek ziarnistych z
ciałami niedojrzałych,
migrujących
neuroblastów oraz
komórek ziarnistych.
Niebieski – warstwa
kłębuszkowa z ciałami komórek
około kłębuszkowych – miejsce
połączenia aksonów receptorów
węchowych z apikalnymi
dendrytami komórek mitralnych
Pseudokolory naniesione Photoshop-
em w celu pokazania 3 głównych
warstw anatomicznych
Budowa opuszki węchowej – dorosła mysz
Przekrój poprzeczny
przez opuszkę węchową
dorosłego samca myszy
(szczep: C57BL/6j).
Zdjęcie z mikroskopu
konfokalnego -
znakowanie jądrowe
TOTO3

Analiza całych struktur: kłębuszki opuszek węchowych u
myszy PLP
Mózg myszy PLP3 postmortem. preparat - R.Platek,
zdjęcia - J.Korczynski + R.Platek
Nieopublikowane dane
Instytut IBD Nenckiego

Podsumowanie i wnioski
Zalety mikroskopu konfokalnego TCS LSI:
• szybka ocena skuteczności zastosowanej metody doświadczalnej,
• szybkie pozyskanie wyniku w analizach nie wymagających bardzo
dużych powiększeń,
• analiza wyników na dużą skalę – skanowanie dużych powierzchni
(in vivo oraz in vitro) – trudne do przeprowadzenia na pojedynczych
skrawkach,
• duża elastyczność obrazowania – użycie obiektywów makro (1x, 2x i 5x)
w połączeniu z elektronicznym zoom-em.

Podziękowania:
Marian Kawczynski,
Kaw.aska, Zalesie Górne, Polska
• prof. Wanda Kłopocka Pracownia Mikroskopii Konfokalnej, Nencki
• Jarek Korczyński Pracownia Mikroskopii Konfokalnej, Nencki
• prof. Małgorzata Skup Pracownia Procesów Reinerwacyjnych, Nencki

Dziękuję za uwagę

Cell cycle analysis using time-lapse microscopy
Grażyna Mosieniak
Laboratory of Molecular Bases of Aging
Nencki Institute of Experimental Biology, Warsaw, Poland
Among many different techniques used in microscopy in order to visualize biological processes, time-laps microscopy gives unique opportunity to observe living cells on single cell level in time. One of the important processes that gain special interest is mitosis. Equal and undisturbed distribution of genetic material into two daughter cell that take place during mitosis is prerequisite of genome stability. In contrary, mitosis disturbances give rise to chromosomal instability, aneuploidy and cancer. Morphological changes that characterize mitotic cells enables to monitor cell division as well as tracking the fate of daughter cells. Moreover, transfection of the cells with vectors coding fluorescent proteins which expression changes during cell cycle make possible to follow the cell cycle progression of individual cell. One of the very interesting process that gain attention in cancer biology research is senescence of cancer cells. This process is induced during chemotherapy and thus it determine the outcome of it. Using video-microscopy we were able to show that aberrant mitosis could be the primary cause of senescence of cancer cells. Moreover video-microscopy enables to observe the fate of senescent cells and help to answer the question whether senescence of cancer cells lead to permanent growth arrest or is transient and could results in regrowth of cancer cells after therapy.

Cell cycle analysis using time-lapse microscopy
Grażyna Mosieniak
Laboratory of Molecular Bases of Aging

2N
2N-4N
4N
4N
CYKL KOMÓRKOWY
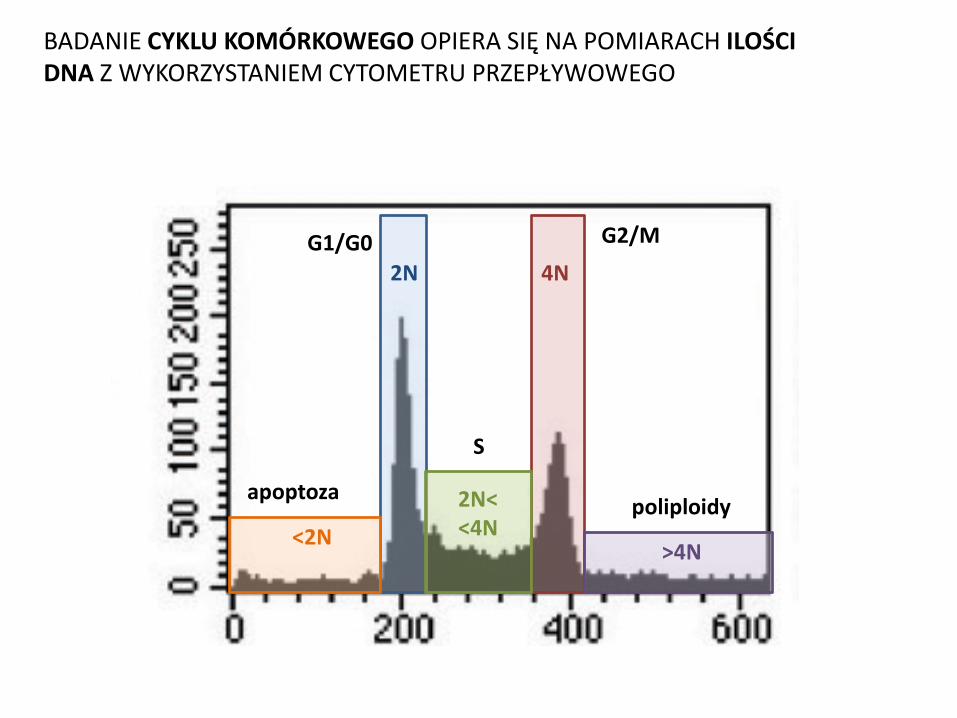
G1/G0 G2/M 2N 4N
2N< <4N <2N
S
apoptoza
>4N
poliploidy
BADANIE CYKLU KOMÓRKOWEGO OPIERA SIĘ NA POMIARACH ILOŚCI DNA Z WYKORZYSTANIEM CYTOMETRU PRZEPŁYWOWEGO

CYKL KOMÓRKOWY

RIEDER C.L., KHODJAKOV A., 2003 Science 300, 91-96
Mitoza dziś …
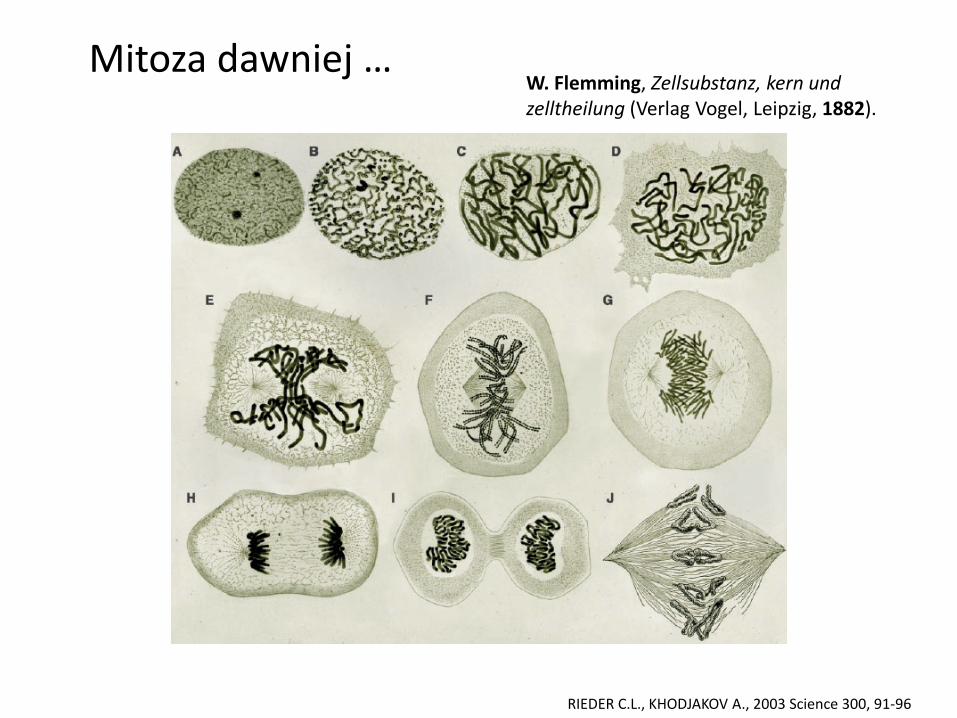
RIEDER C.L., KHODJAKOV A., 2003 Science 300, 91-96
Mitoza dawniej … W. Flemming, Zellsubstanz, kern und zelltheilung (Verlag Vogel, Leipzig, 1882).

Lata 20-te XX w

CZAS
VIDEO-MIKROSKOPIA UMOŻLIWIA ŚLEDZENIE LOSÓW KOMÓRKI
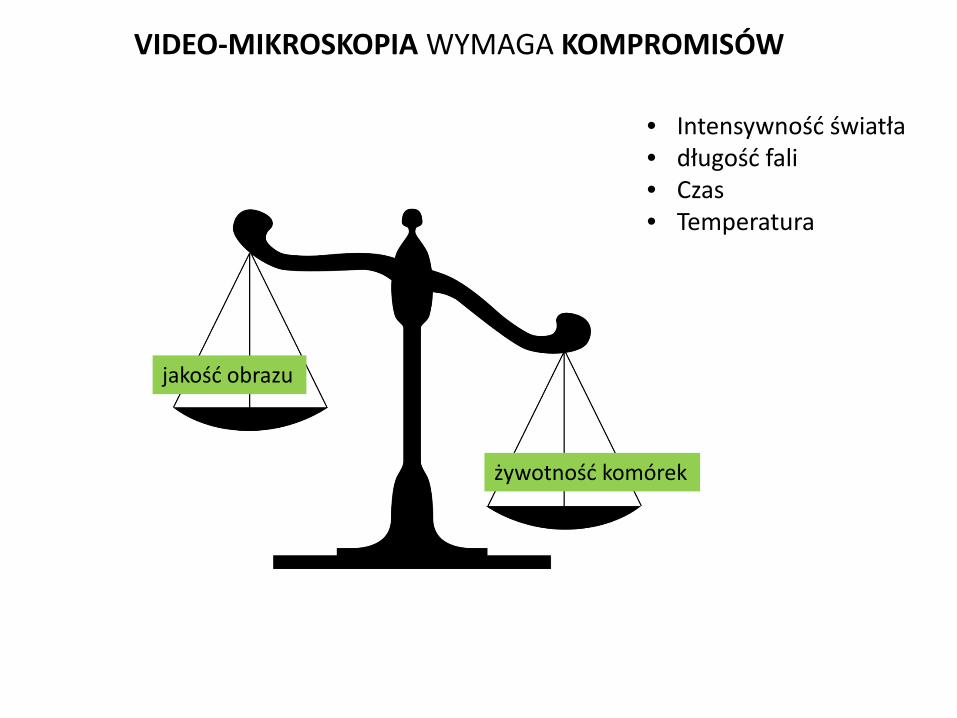
jakość obrazu
żywotność komórek
VIDEO-MIKROSKOPIA WYMAGA KOMPROMISÓW
• Intensywność światła • długość fali • Czas • Temperatura

Live imaging of influence of MMP-9 enzymatic activity on spine morphology through integrins. Izabela Rutkowska-Włodarczyk Laboratory of Neurobiology Nencki Institute of Experimental Biology, Warsaw, Poland
Synaptic plasticity can be defined as a re-organization comprising both, alterations at
the morphological level and changes at the functional level of dendritic spines, carrying
postsynaptic domains of excitatory synapses. It has been reported that motor learning leads to
rapid formation of dendritic spines (spinogenesis) in the motor cortex mice (Xu et al., 2009; Yang
et al., 2009). Changes in spine dynamics are region- and learning-specific, indicating that motor
learning causes synaptic reorganization in the corresponding motor cortex (Xu et al., 2009) and
synaptic connections are not only capable of undergoing rapid changes in response to new
experience but also can serve as substrates for long-term information storage (Yang et al.,
2009). This is why it became important to effectively quantify changes in dendritic spine
morphology. However, the results of this quantitative analysis can be largely influenced by the
the diversity in dendritic spine population. The detection of differences in spine morphology
between control and test group is often compromised by the number of dendritic spines taken
for analysis. For example, to have statistically significant detection of hidden morphological
differences between control and test groups in terms of spine head-width, length and area in
which the values of each morphometric variable under investigation grew in a 20% rate there
are required 12, 21 and 24 cells respectively with around 30 spines/cell. Simulation of changes
occurring in a subpopulation of spines reveal the strong dependence of detectability on the
statistical approach applied (Ruszczycki et al. 2012). This is way the best method to track spine
morphology changes is live imaging.

Live imaging of influence of MMP-9 enzymatic activity on spine morphology
through integrins.
Izabela Rutkowska-Wlodarczyk
Laboratory of Neurobiology
The Nencki Institute of Experimental Biology

OVERVIEW OF THE PROJECT

Changes in dendritic spine morphology underlie learning processes
(Xu et al., Nature,2009; Yang et al., Nature, 2009)

extracellurally operating protease that is expressed by the neurons and released in response to enhanced neuronal activity Michaluk et al., J. Biol. Chem., 2007
plays a key role in synaptic plasticity associated with memory and learning processes; Nagy et al, J. Neurosci., 2006;
Okulski et al., Biol. Psychiatry, 2007
was found to be present in a subset of dendritic spines bearing asymmetric synapses; Wilczynski et al., J. Cell Biol., 2008.
Matrix metallopeptidase 9
Dziembowska&Wlodarczyk, Int. J. Biochem. Cell Biol, (2012)

Integrins:
• play a role in the synaptic structural changes associated with LTP (Lee et al., 1980).
• stabilizing LTP (Staubli et al., 1990)
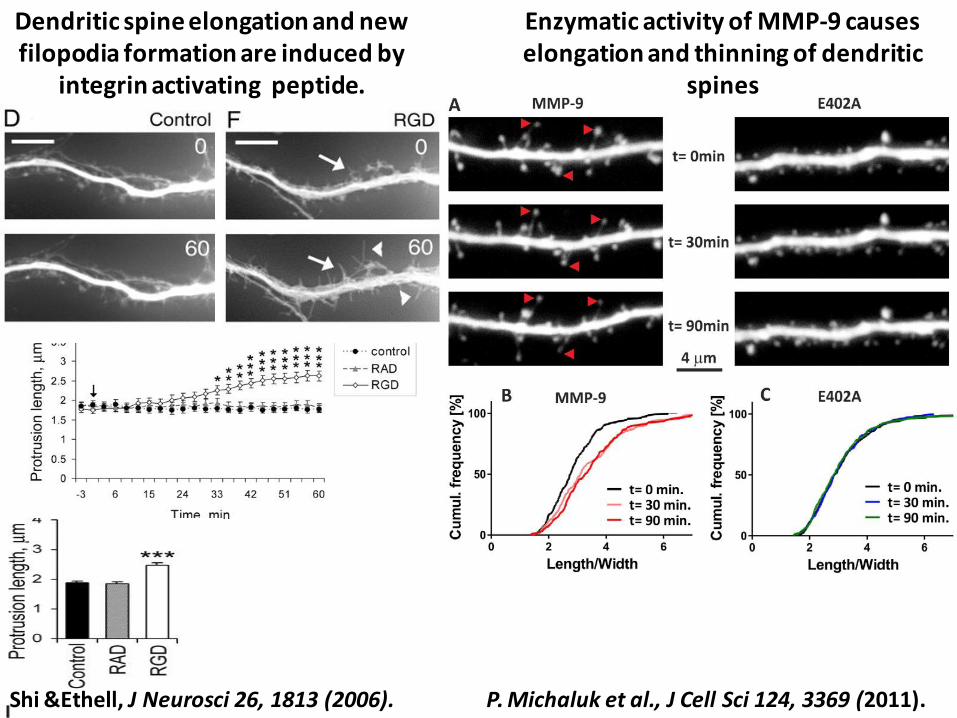
Dendritic spine elongation and new filopodia formation are induced by
integrin activating peptide.
Enzymatic activity of MMP-9 causes elongation and thinning of dendritic
spines
P. Michaluk et al., J Cell Sci 124, 3369 (2011). Shi &Ethell, J Neurosci 26, 1813 (2006).
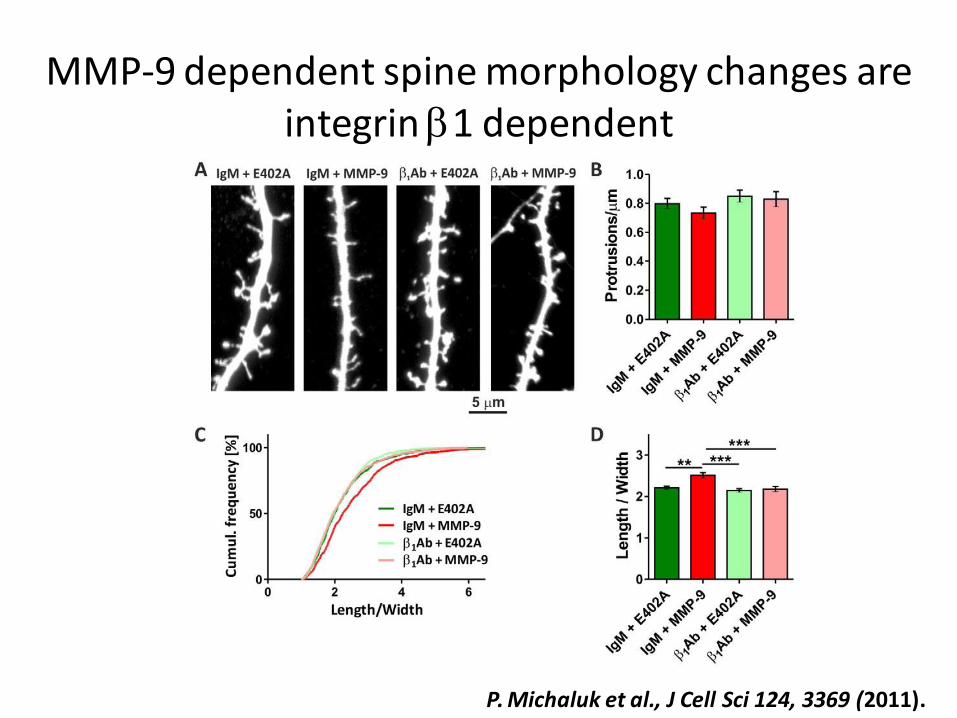
P. Michaluk et al., J Cell Sci 124, 3369 (2011).
MMP-9 dependent spine morphology changes are integrin b1 dependent

HYPOTHESIS

MMP-9 spine elongations
integrins
MMP-9-mediated spines morpholgy changes are integrins dependent
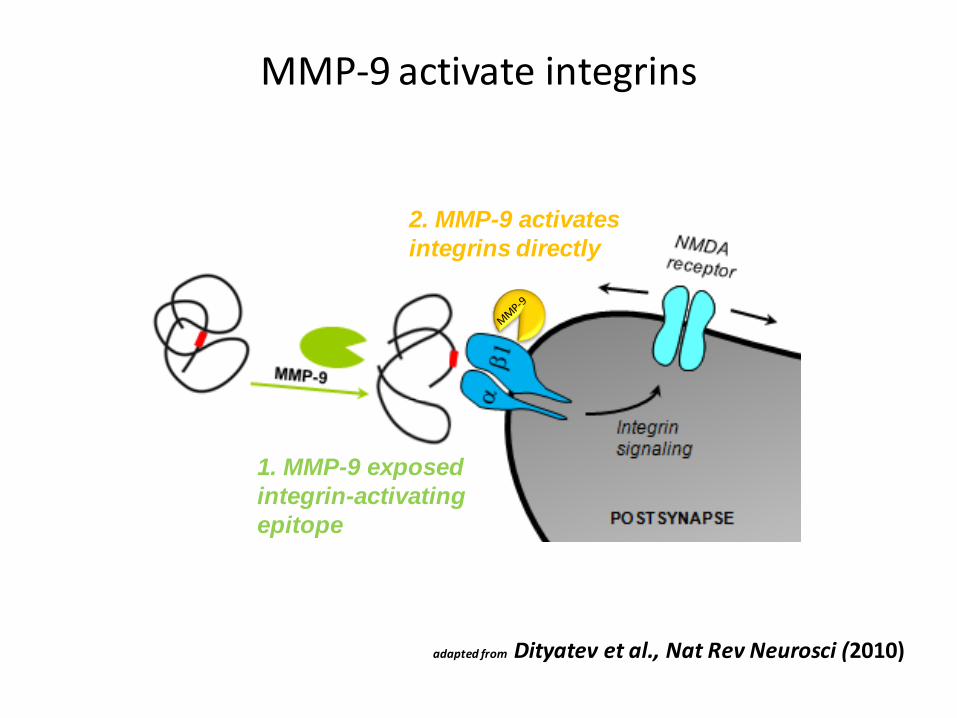
MMP-9 activate integrins
1. MMP-9 exposed
integrin-activating
epitope
2. MMP-9 activates
integrins directly
adapted from Dityatev et al., Nat Rev Neurosci (2010)

HYPOTHESIS VERIFICATION
Source of MMP-9 substrates: hippocampal tissue
aaMMP-9 400ng/ml
37oC, 1h
MMP-9 Integrin conneXtion
Identification of MMP-9 substrates which serve
as integrin activators
MIX
GFP-transfected 21 DIV hipocampal culture
Zeiss LSM780

‘Live’ experiments-protocol
inhibitor MIX
0 -30 10 20 30 5


‘Live’ experiments allow to track changes in the
spines morphology
Filopodia Thin Mushroom Stubby
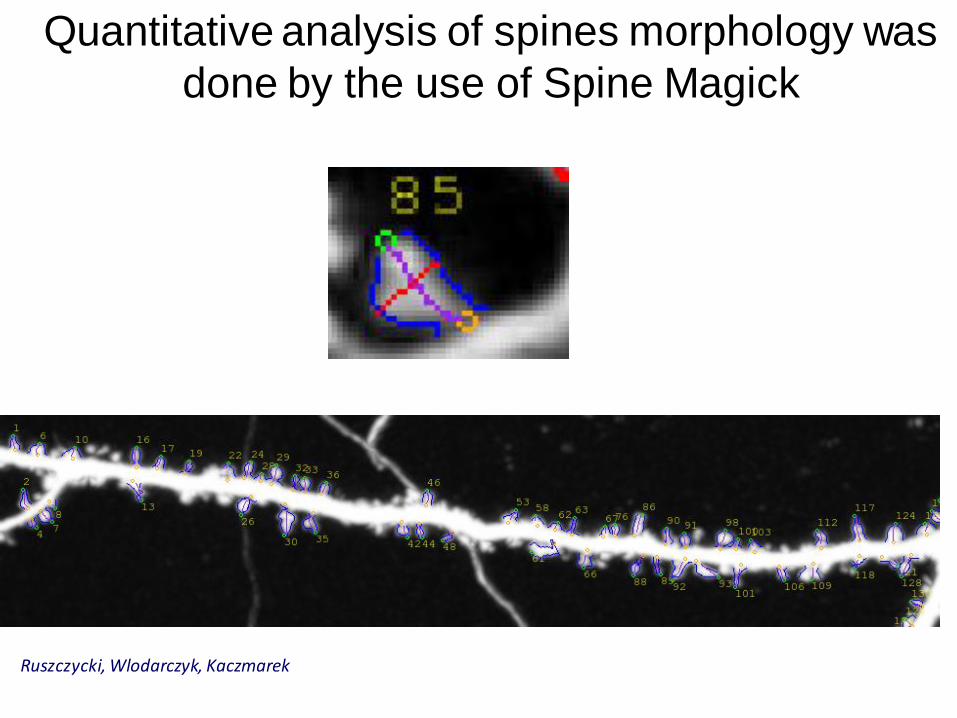
Quantitative analysis of spines morphology was
done by the use of Spine Magick
Ruszczycki, Wlodarczyk, Kaczmarek

Z-section
Halo correction:
Spine head anti-detachment:
spine heads often separated; necks barely visible
spines covered by dendrite halo
improved segmentation of spine neck
before correction after correction
Local brightness variation:
Ruszczycki, Wlodarczyk,Kaczmarek
SpineMagick

Acknowledgement
Kasia Zalewska Amanda Costa Marta Pyskaty

Thank you for your attention

The application of the FRAP technique and live cell imaging to investigate an impact of HAX-1 protein on the formation and movement of P-bodies in HeLa cell line. Ewa A. Grzybowska Cancer Center Institute, Warsaw, Poland HAX-1 is a multifunctional protein involved in several key processes like apoptosis, cell migration and regulation of calcium homeostasis, but its mechanisms of action remain unknown. HAX-1 is also known to bind to the 3'UTR of at least two mRNA targets. Our research indicate that HAX-1 co-localizes with P-body marker, decapping protein 1 (Dcp1). Using Fluorescence Recovery After Photobleaching (FRAP) technique we demonstrated that HAX-1 dynamically interchanges between P-body and the cytoplasm, but it is also bound in P-body to significant extent. Concurrently, using live-cell imaging, we showed that HAX-1 is not responsible for the immobilization of P-bodies on actin filaments.
