morphology and accommodative function of the …aging.wisc.edu/pdfs/2368.pdf · morphology and...
TRANSCRIPT

1 Vitreous Zonule
Morphology and accommodative function of the vitreous zonule in human and monkey eyes Lütjen-Drecoll, Elke1; Kaufman, Paul2,4; Wasielewski, Rainer1; Ting-Li Lin3; Croft, Mary Ann2 1Institute of Anatomy II, University of Erlangen-Nuremberg, 91054 Erlangen, Germany; 2Department of Ophthalmology and Visual Sciences, University of Wisconsin, Madison, WI 53792; 3Biostatistics and Medical Informatics, University of Wisconsin, Madison, WI 53792; and 4Wisconsin National Primate Research Center, University of Wisconsin, Madison, WI 53792
Running title: Vitreous Zonule Function in Human and Monkey Eyes To whom correspondence should be addressed:
Mary Ann Croft, MS Department of Ophthalmology and Visual Sciences University of Wisconsin Clinical Sciences Center 600 Highland Avenue Madison, WI 53792-3220 Telephone: 608-263-7738 Fax: 608-263-0543 Email: [email protected]
Word Count 4434
Support:
This work was funded in part by NEI grants RO1 EY10213 & R21EY018370 to
PLK, the Ocular Physiology Research & Education Foundation; and DFG DR
124/7 to ELD. We also acknowledge the Wisconsin National Primate Research
Center, University of Wisconsin-Madison base grant # 5P51 RR 000167 and the
Core Grant for Vision Research grant # P30 EY016665.
Page 1 of 46 IOVS IOVS Papers in Press. Published on October 8, 2009 as Manuscript iovs.09-4008
Copyright 2009 by The Association for Research in Vision and Ophthalmology, Inc.

2 Vitreous Zonule
Abstract
Purpose: To explore the attachments of the posterior zonule and vitreous in
relation to accommodation and presbyopia in monkeys and humans.
Methods: We used novel SEM and UBM techniques to visualize the anterior,
intermediate and posterior vitreous zonule and their connections to the ciliary
body, vitreous membrane, lens capsule and ora serrata, and to characterize their
age-related changes and correlate them with loss of accommodative forward
movement of the ciliary body. We injected -chymotrypsin focally to lyse the
vitreous zonule and determined the effect on movement of the accommodative
apparatus in monkeys.
Results: The vitreous attaches to the peripheral lens capsule and the ora serrata
directly. The pars plana zonule and the posterior tines of the anterior zonule are
separated from the vitreous membrane except for strategically placed
attachments, collectively termed “vitreous zonule,” that might modulate and
smooth the forward and backward movements of the entire system. Age-
dependent changes in these relationships correlated significantly with loss of
accommodative amplitude. Lysis of the intermediate vitreous zonule partially
restored accommodative movement.
Conclusions: The vitreous zonule system may help to smoothly translate to the
lens the driving forces for accommodation and disaccommodation generated by
the ciliary muscle, while maintaining visual focus and protecting the lens capsule
and ora serrata from acute tractional forces. Stiffening of the vitreous zonular
system may contribute to age-related loss of accommodation and offer a
therapeutic target for presbyopia.
Page 2 of 46IOVS

3 Vitreous Zonule
Introduction
The age-related loss of accommodation (presbyopia) in human and nonhuman
primates clearly involves loss of deformability of the crystalline lens. In addition,
ciliary muscle mobility, as measured by both histologic [1, 2] and
ultrasonographic (monkey) [3] and MRI (human) [4] imaging techniques, also
diminishes with age. However, the isolated ciliary muscle in nonhuman primates
maintains its full ability to contract in response to appropriate pharmacological
stimulation throughout life, [1, 2, 5] and its contractile machinery and receptor
biology also remain normal. [6, 7] Therefore, posterior restriction of muscle
movement in the aging eye has been hypothesized also to play a role in the
pathophysiology of presbyopia, at least in monkeys. [8]
Additionally, there are zonular attachments to the vitreous membrane in both
species that have not yet been fully investigated. Earlier studies focused mainly
on the anterior hyaloid membrane. Wieger [9] was among the first to describe
attachments of the zonules to the anterior vitreous membrane. They were later
termed Wieger’s ligament. [10-12] Streeten [13] doubted the strength of Wieger’s
ligament, since following lens extraction it was not uncommon for a superficial
flap of either the capsule or anterior hyaloid to be torn from Wieger’s area. [13]
Bernal et al. [14], using a combination of environmental scanning electron
Page 3 of 46 IOVS

4 Vitreous Zonule
microscopy (ESEM) combined with a custom-made manual lens stretcher to
facilitate “inside microscopy,” were able to further analyze the complex
connection between the anterior vitreous membrane and zonule. [14]
Attachments between the posterior pars plana zonule and the vitreous
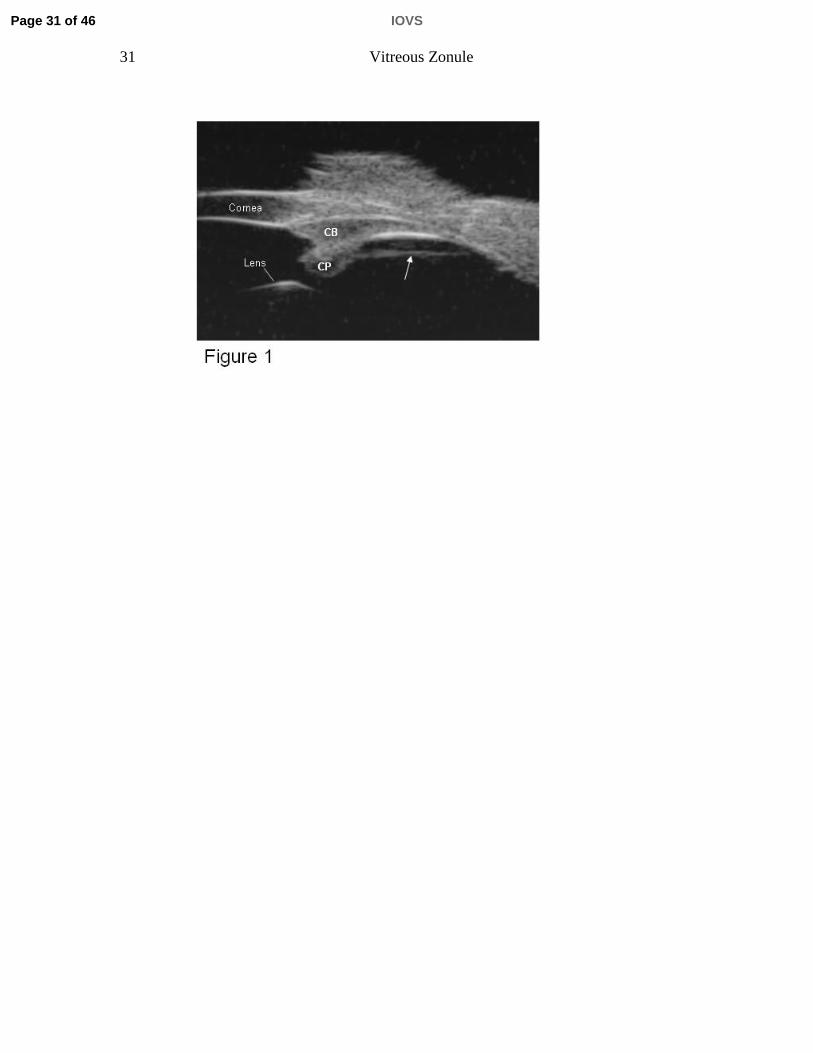
membrane have been observed in histological sections and SEM. [15, 16] [17]
Improved in vivo imaging by ultrasound biomicroscopy (UBM) allows visualization
of zonular fibers in the living eye. [18] Using UBM, we observed a straight line
parallel to but separated from the pars plana by a cleft about 2 mm wide
extending from the region of the ciliary processes to the region of the ora serrata
[Fig. 1]. There has been debate as to whether the structure observed by UBM
was: 1) the pars plana zonule that in the living eye was normally separated from
the pars plana ciliary epithelium, but after fixation for histology or air drying for
scanning electron microscopy (SEM) became apposed to the pars plana ciliary
epithelium (CE); [19] 2) the vitreous membrane [20]; 3) another component of the
vitreo-zonular system that was not previously well-described or characterized.
We have investigated the zonule, the posterior vitreous, and their attachments by
novel scanning SEM and UBM techniques, and we now can hypothesize how
this system might affect ciliary muscle movement. Additionally, we provide
preliminary physiological evidence that ciliary muscle movement might be
increased by perturbing the system.
Page 4 of 46IOVS

5 Vitreous Zonule
Materials and Methods
Morphology
Monkey Eyes
Twenty-seven normal eyes of 22 rhesus monkeys (Macaca mulatta) of either sex
and of 1 cynomolgus monkey (Macaca fascicularis), ranging in age from 3.5 to
28 years, were obtained from caged colonies of the Wisconsin National Primate
Research Center, the Harlow Primate Laboratory, and Covance Incorporated (all
in Madison/WI/USA) and analyzed by SEM. The animals were euthanized in
conjunction with various nonocular protocols. All procedures conformed to the
ARVO Statement for the Use of Animals in Research and were in accordance
with institutionally approved animal protocols.
Before sacrifice, the animals were deeply anesthetized after ketamine induction
(3-30 mg/kg i.m.) with pentobarbital Na (10-15 mg/kg i.v. supplemented with 0.5-
10 mg/kg i.v., as needed) and then perfused transcardially with 1 liter of 0.1
molar PBS (phosphate-buffered saline) followed by paraformaldehyde (PFA) 4%
for 10-15 minutes. Following systemic perfusion fixation, the eyes were
enucleated, slits were cut in the posterior sclera, and a window was cut in the
anterior cornea to enhance fixative penetration while preserving the architecture
of the ciliary muscle and its posterior attachment to the elastic lamina of Bruch’s
membrane [1] [21] The entire eyes were then placed in lto’s fixative [22] and sent
to Erlangen.
Page 5 of 46 IOVS

6 Vitreous Zonule
Before preparation for SEM, the eyes were placed in cacodylate buffer overnight.
Small pie-shaped sectors were cut from all four quadrants (of the anterior globe).
These sectors contained the ciliary body; the adjacent cornea, sclera, and
neighboring part of the lens; and the vitreous membrane.
Visualization of the vitreous membrane and the zonule is difficult and requires
several approaches, as the consistency of the vitreous does not allow good
preparation in non-critical point dried specimens. However, after critical point
drying the delicate membrane is extremely rigid and friable. Without
“customized” preparation methods, the vitreous membrane completely covers the
ciliary body and zonule, as the spaces between the structures collapse during the
drying process.
Ultimately, three different approaches were used to visualize the different
portions of the zonular connections to the vitreous membrane.
I. To analyze the zonular connections to the anterior vitreous membrane
and the posterior lens capsule, the lens was kept in place. The anterior
vitreous membrane was carefully reflected posteriorly, trying not to
disrupt its connections to the adjacent structures. In some specimens
the ciliary processes were separated from each other to visualize the
connection of the zonule to the ciliary epithelium. In other specimens
the tips of the processes were excised for the same purpose. After
Page 6 of 46IOVS

7 Vitreous Zonule
critical point drying, the specimens were viewed from a sagittal, an
“anterior chamber,” and a posterior chamber approach.
II. For visualization of the zonular connections in the area between the
pars plana zonule and the vitreous membrane over its entire anterior-
posterior sagittal length, specimens were viewed from their sagittal
aspect, and the vitreous membrane was carefully lifted from the pars
plana region. Using this approach, it was obvious that overlying most
of the pars plana region there was a cleft between the pars plana
zonule and the neighboring vitreous that disappeared in the most
posterior pars plana in the region of the ora serrata.
III. To analyze this area of connection in more detail, either the anterior or
posterior part of the vitreous membrane was carefully scrolled. The
critical point dried specimens were then investigated either from a
sagittal or an intraocular “enface” perspective.
For critical point drying, following the various preparatory protocols, the
specimens were carefully dehydrated in graded alcohol and acetone and finally
dried in a critical point dryer (cf. Rohen 1979). [19] The specimens were then put
on a disk, coated with gold-palladium and examined with the Stereoscan
scanning electron microscope (Cambridge, UK, Ltd.).
Human Eyes
Page 7 of 46 IOVS

8 Vitreous Zonule
Specimens of each quadrant of 8 human eyes aged 55-100 years (mean ± SD
67 ± 15.3 years) with postmortem times between 7 and 18 hours (mean ± SD
11.2 ± 3.5 hours) were evaluated in the same manner as the monkey eyes. The
eyes were obtained by the Department of Anatomy of the University of Erlangen-
Nürnberg, after appropriate consent. Immediately after enucleation the anterior
portion of the cornea was removed, slits were cut into the sclera, and the eyes
immersion-fixed in Ito’s solution for 3-5 hours. The anterior eye segment was
then dissected and post-fixed in 1% osmic acid for 3 hours. After that, small pie-
shaped sectors were cut from the anterior eye segment and prepared as
described above.
Accommodation in Live Monkeys
A Hartinger coincidence refractometer (Jena, Germany) was used to measure
resting refractive error and accommodation in response to electrical stimulation
of the Edinger-Westphal (E-W) nucleus via an implanted electrode. [3]
Supramaximal stimulus settings were chosen (as defined below) that induced
maximum forward ciliary body movement and maximum accommodation,
allowing comparisons to be made of accommodative responses between young
and older eyes. Maximal stimulus is defined as the level of E-W stimulus current
necessary to induce maximum accommodative change, measured
refractometrically. Supramaximal stimulus is a level of E-W stimulus current
~25% (or ~0.10 to 0.20 mA) above the maximal stimulus that ensures maximum
forward ciliary body movement.
Page 8 of 46IOVS

9 Vitreous Zonule
Ultrasound Biomicroscopy in Live Monkeys
A 50 MHz UBM instrument (Model # 840, Humphrey Instruments, San Leandro,
CA) was used to image the posterior vitreous zonule and ciliary body, and the
images were then used to measure the width of the cleft between the pars plana
and the vitreous membrane/vitreous zonule in the resting eye (Fig. 1). In addition,
a 35 mHz UBM instrument (Model # MHF-1 Ultraview System Model P60, E-
technologies, Bettendorf, IA) was used to collect images encompassing the
entire sagittal extent of the ciliary body to the region of the ora serrata. Dynamic
UBM images were obtained during central stimulation of accommodation and
then recorded to videotape. [3] From these images, measurements were taken of
the distance between the scleral spur and the posterior insertion point of the
posterior vitreous zonule at rest and during supramaximal central stimulation.
The eye was stabilized with extraocular muscle sutures during UBM imaging to
dampen or eliminate convergence eye movement. The transducer was held in
place by a stabilization arm to eliminate any change in angle of the transducer to
the eye during accommodation, and the anterior and posterior ends of the ciliary
body were oriented in a sagittal direction within all images to ensure stability and
reproducibility. The temporal region was selected as providing the best
ultrasound imagery, with clear and distinct edges of the ciliary body and posterior
vitreous zonule. Comparisons were made of the angle between the inner aspect
of the cornea and the anterior aspect of the muscle apex (clinically called the
Page 9 of 46 IOVS

10 Vitreous Zonule
ciliary face and not including the ciliary processes) in the unaccommodated vs.
the accommodated state; the narrowing of this angle was used as a surrogate
indicator of forward ciliary body movement. [3] With respect to comparing the
angle measurement in the unaccommodated and accommodated states, the
location of the anterior aspect of the muscle apex was far enough inward to
reflect the translocation of the innermost aspect of the muscle.
Lysis/Disruption of the Posterior Vitreous Zonule
Surgical Procedure
In 7 rhesus monkey eyes (aged 6-9 (n=4), 15, 25 and 26 years) and 1
cynomolgus monkey (aged 3.5 years), lysis of the posterior zonule was
attempted by insertion of 8-16 µl of 83 units/ml -chymotrypsin in heavy sucrose
medium via a 1 mm incision through the sclera and the choroid in the mid-
region of the pars plana. The eye was oriented so that the solution fell by gravity
onto the strands of the zonule. The contralateral eyes served as controls. One of
these 7 rhesus monkeys, aged 25 years, underwent bilateral extracapsular lens
extraction (ECLE) [23] 19 months before unilateral -chymotrypsin injection and
transcorneal lens capsulectomy/vitrectomy. Three of the 7 rhesus monkeys, aged
6-9 years, were pseudophakic following bilateral ECLE 8 months before
unilateral -chymotrypsin injection.
Ultrasound Biomicroscopy
Page 10 of 46IOVS

11 Vitreous Zonule
Maximum accommodative responses were induced by corneal iontophoresis of
40% carbachol in agar (a supramaximal dose) [24] in 3 monkeys before and after
-chymotrypsin injection. In 3 other rhesus monkeys, maximal accommodative
responses were induced by supramaximal E-W stimulation before and after -
chymotrypsin injection. UBM imaging was undertaken in the resting and
accommodated state before and after -chymotrypsin injection in these 6 monkeys.
Three of these monkeys were randomly selected to undergo morphological
examination (below).
Morphology
Three of the pseudophakic rhesus (aged 6-9 years), the aphakic rhesus, and the
1 normal phakic 3.5-year-old cynomolgus monkey were studied. Specimens from
the region of injection of -chymotrypsin were prepared for SEM as described
above. In addition, 1-2 mm wide specimens of the anterior eye segment that
included the entire anterior-posterior extent of the ciliary body and the adjacent
cornea and sclera were embedded in Epon, and 1 µm semithin sections were cut
and stained with toluidine blue. In these semithin sections, the apical position and
width of the ciliary muscle were determined as described previously. [21] The
opposite eye served as control.
Statistical Analysis
A two-tailed paired t-test was used to detect significant differences. Multiple
regression analysis was performed to evaluate the relationships between
Page 11 of 46 IOVS

12 Vitreous Zonule
accommodative amplitude and age or the forward movement of the posterior
insertion zone. Stepwise regression analysis based on Akaike information
criterion (AIC) was used for model selection. [25] A p-value 0.05 was
considered significant; 0.05 p 0.10 was considered to indicate a trend, given
the small number of monkeys in some groups. Throughout this paper, mean ±
s.e.m. was reported. However, with respect to the age range and post mortem
time range of the 8 human eyes that were studied the mean ± standard deviation
are reported (see above).
All procedures conformed to the ARVO statement for the Use of Animals in
Research and were in accordance with institutionally approved animal protocols.
Results
Morphology in Monkey Eyes
Anterior Vitreous Zonule
An overview of the insertion of the vitreous membrane and posterior tines of the
anterior zonule indicated that both systems inserted into the posterior lens
capsule at the lens periphery (Fig. 2 A, B). Following careful lifting of the vitreous
membrane from the zonule, it could be shown that both the zonule and the
vitreous inserted nearly at the same place but the vitreous zonule covered the
insertion of the zonular plate and inserted somewhat more centrally. The majority
of the posterior zonular tines were not attached to the vitreous membrane but
formed a zonular plate running from the zonular plexus to the posterior lens
Page 12 of 46IOVS

13 Vitreous Zonule
capsule (Fig. 2 A). As the vitreous could easily be lifted from this zonular plate,
we assume that in the live monkey the zonular plate and vitreous are separated
by an aqueous-filled small cleft in this region. There were, however, strands of
zonular fibers, deriving from the zonular plexus that inserted into the vitreous
membrane at fairly regular intervals of about 50 to 75 µm (Fig. 2 C). These
connections were termed “anterior vitreous zonule.”
Intermediate and Posterior Vitreous Zonule
After careful elevation of the vitreous membrane overlying the middle part of the
pars plana, taking care not to remove the vitreous membrane itself, we could
clearly identify the presence of a cleft between the vitreous membrane and the
pars plana zonules. This cleft was bridged by zonular fiber bundles, here called
intermediate vitreous zonules. These were spaced uniformly around the entire
circumference of the pars plana region, separated by approximately the width of
a ciliary process (Fig. 3 A, B). Indeed, each intermediate vitreous zonular fiber
bundle split anteriorly and formed a fork whose tines inserted into the zonular
plexus on either side of an individual ciliary process (Fig. 3 B). The intermediate
vitreous zonular bundles had no direct contact with the pars plana zonular
apparatus.
In contrast, posteriorly in the ora serrata region, each intermediate vitreous
zonular fiber bundle split into several fine fibrils that became part of the vitreous
membrane (Fig. 4 A). The vitreous membrane, with its brush-like posterior
Page 13 of 46 IOVS

14 Vitreous Zonule
extensions of the intermediate vitreous zonule, was attached to the pars plana
zonule by numerous oblique-running fine fibrils (Fig. 4 B). Thus, at the site of
attachment of the vitreous to the posterior pars plana and ora region, a multi-
layered, interconnected sponge-like ring, termed the posterior vitreous zonule,
was formed.
Morphology in Human Eyes
In general, the SEM appearance and anatomic relationships of the vitreous
zonule and vitreous membrane were similar in humans and monkeys. The most
striking difference involved the posterior vitreous zonule and its attachment to the
pars plana zonule in the region of the ora serrata. In the monkey, the sponge-like
insertion encompassed a sagittal length approximately 1/6 ( 0.5 mm) the
distance between the scleral spur and ora. In the human, the insertion was
disproportionately thicker and encompassed between 1/3 and 1/2 ( 3.4 mm) the
sagittal distance between the scleral spur and the ora (Fig. 5 A). SEM of the
posterior vitreous zonule showed that this system is more complex in the human
eye than in the monkey eye. In the entire area of attachment, vitreous zonular
fibers ran from the posterior zonular insertion anteriorly to the vitreous membrane
and also from the more anterior pars plana zonule into the posterior vitreous
zonule, thereby forming a lattice-like structure (Fig. 5 B).
Ultrasound Biomicroscopy in Live Monkeys
Page 14 of 46IOVS

15 Vitreous Zonule
1) UBM imaging, involving slightly oblique rather than absolutely perpendicular
cuts, revealed that the pars plana zonule lay immediately adjacent to the pars
plana ciliary epithelium (Fig. 6 A, B).
2) Nearly serial cuts revealed two distinct linear structures separated from the
pars plana by a wide cleft: the vitreous zonule extending from the ora serrata
region to the valleys of the pars plicata, and the vitreous membrane branching
from and lying internal to the vitreous zonule as both extended anteriorly from the
ora region (Fig. 7 A, B). The relative prominence of each linear structure varied
from cut to cut.
3). The apparent discontinuities in the vitreous zonule in some of the
perpendicular sections (Fig. 7 A) most likely represented the forking of the
vitreous zonule fibers as they approached the posterior aspects of the ciliary
processes as seen by SEM (Fig. 3 B). The similarity of the SEM and UBM
images is easily seen in Fig. 8.
Age-related Changes in Live Monkeys
In the resting eye, the width of the cleft between the pars plana region of the
ciliary body and the intermediate vitreous zonular bundles, at the cleft’s widest
point, increased with age by 0.012 ± 0.0022 mm/year (Fig. 9 A-D); p=0.001,
r=0.79, n=19). Additionally, the thickness of the ciliary muscle at the point of the
maximum cleft width decreased with age, and the curvature of the pars plana
and the posterior pars plicata increased with age (Fig. 9 A-C). Further, with
Page 15 of 46 IOVS

16 Vitreous Zonule
increased cleft width, accommodative amplitude declined significantly (-36.1 ±
11.1 D/mm; Fig. 9 E); p=0.007, r=0.63, n=17).
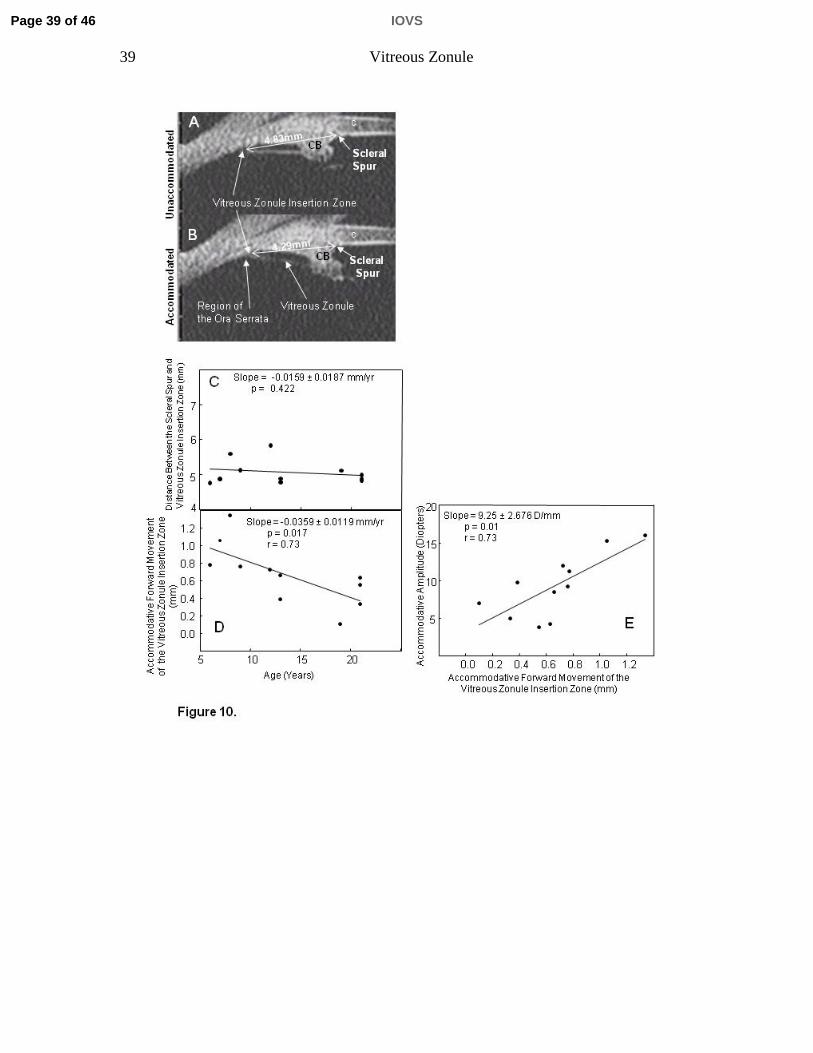
In the resting eye, the distance between the scleral spur and the posterior
insertion zone of the posterior vitreous zonule (Fig. 10 A, B) did not significantly
change with age (Fig. 10 C). During accommodation, it could clearly be seen that
the posterior insertion zone of the posterior vitreous zonule moved forward (see
Video Clip 1, Fig. 10 A, B). Thus the distance between the scleral spur and the
posterior insertion zone of the posterior zonule diminished with accommodation
(p<0.001) and did so more markedly in the young eyes (1.052 ± 0.067 mm, n=4)
than in the older eyes (0.403 ± 0.118 mm, n=4) (Table 1, Fig. 10 D).
Results of the regression analysis showed that accommodative amplitude
(p=0.0003; not shown) and accommodative forward movement of the posterior
insertion zone (p=0.017) declined significantly with age (Fig. 10 D). The
accommodative forward movement of the posterior insertion zone declined by
~60% in the older vs. the young eyes (p=0.001; Table 1). Accommodative
amplitude increased significantly with increased accommodative forward
movement of the vitreous zonule’s posterior insertion zone (Fig. 10 E).
Page 16 of 46IOVS

17 Vitreous Zonule
The results of the stepwise regression analysis showed that age better predicted
accommodative amplitude than did either the forward movement of the posterior
insertion zone or the cleft width.
Disruption of the Intermediate Vitreous Zonule in Live Monkeys
Morphology
SEM showed that the intermediate vitreous zonular bundles in the region of the
-chymotrypsin injection were absent (Fig. 11). In all 4 rhesus (aged 6-9 and 25
years) independent of phakic status, at rest the muscle in the enzyme-injected
temporal quadrant was positioned more anteriorly than the temporal quadrant in
the contralateral controls (Fig. 12). This was true to a lesser degree for the single
young cynomolgus. For all 5 monkeys considered together, the anterior
translation was 0.256 ± 0.079 mm (n=5; p=0.032; Table 2). The difference in
apical position between treated vs. control eyes became far less pronounced as
the distance from the injection site increased (Table 2). The muscle tended to be
thicker in the -chymotrypsin injected eyes than the contralateral control eyes,
but the difference was not significant.
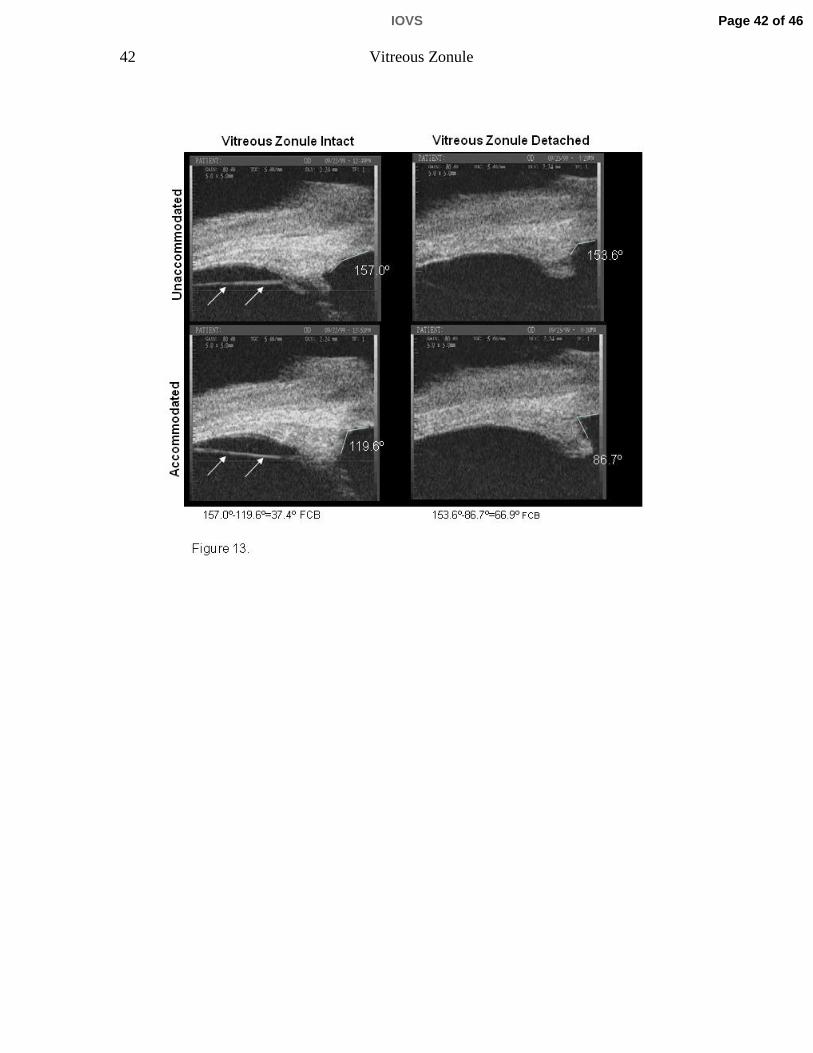
Ultrasound biomicroscopy in live monkeys
UBM imaging of the ciliary body in the temporal quadrant of 6 rhesus monkey
eyes (aged 6 to 26 years) before and after α-chymotrypsin lysis of the vitreous
zonule showed increased accommodative forward ciliary body movement post-
lysis vs. pre-lysis (p=0.044; Table 3, Fig. 13; see Video Clip 2). This was true for
Page 17 of 46 IOVS

18 Vitreous Zonule
phakic eyes, pseudophakic eyes with intact posterior capsule, and the one truly
aphakic eye without capsule. The latter eyes typically exhibited markedly
decreased accommodative ciliary body movement when the intermediate
vitreous zonule was intact. [21]
Discussion
This study describes the complexity of a vitreous zonular system that helps to
smoothly translate to the lens the driving forces for accommodation and
disaccommodation generated by ciliary muscle contraction and relaxation. Our
study also suggests how that structure allows and assists the very fine and rapid
refocusing and maintenance of focus at multiple distances required by primate
visual needs. [26]
In relation to the Helmholtz theory, [27] accommodation requires both centripetal
as well as forward ciliary body movement, and the age-related loss of
accommodation may be related to the loss of forward ciliary body movement. In
relation to the Coleman (catenary) theory of accommodation, which suggests that
accommodative shape changes in the lens are induced by vitreous pressure, [28-
30], age-related loss of ciliary body movement combined with increased lens size
[4, 31, 32] might alter the central catenary shape and thereby reduce
accommodative amplitude. We prefer to not go beyond what we have just stated
above, in order to avoid overspeculation with respect to the various theories of
Page 18 of 46IOVS

19 Vitreous Zonule
accommodation. We have opted to simply publish the findings and have provided
some discussion as to their meaning.
This study unveils the potential for novel mechanisms and suggests the need to
establish methods to include the vitreous membrane and vitreous zonule, along
with the anterior and posterior zonular complexes, in future finite element models
of the primate accommodation system. This manuscript also raises several
interesting questions about the future of anti-presbyopia surgery and treatment.
The presence of vitreous strands in living humans has been observed earlier by
Coleman [33] and in monkeys by Glasser et. al. [18] using ultrasound
biomicroscopy. However, neither the strands themselves nor their anatomic
relationships to other structures were definitively identified and characterized.
Indeed, some researchers have labeled the vitreous strands simply as anterior
hyaloid membrane. [20]
With respect to forward ciliary body movement, the angle measured includes the
inner aspect of the cornea and the anterior aspect of the muscle apex (not
including the ciliary processes) which is quite distinct and easily measured in the
iridectomized eye. Forward ciliary body movement as measured by ultrasound
biomicroscopy does not discern the individual contributions of the different areas
of the muscle (i.e., longitudinal, circular, reticular muscle portions) and what
occurs in the different internal anatomic regions of the muscle cannot be
Page 19 of 46 IOVS

20 Vitreous Zonule
precisely quantified. Nonetheless, the technique is a reliable indicator of overall
forward ciliary body/muscle movement (i.e., muscle apex) [3] [21] based on the
following: 1) the angle change reflects the translocation of the innermost aspect
of the muscle (apex); 2) the change in forward ciliary body movement as
measured by UBM following removal of the posterior restriction is corroborated
by the morphological findings in this paper and historically[1, 2, 5]; 3) the
qualitative examination of the dynamic video images (Video Clip 1 and 2) show
the overall muscle movement is forward and inward without isolated pockets
remaining static (with the exception of the outermost anterior longitudinal portion
of the muscle which is anchored/fixed anteriorly at the scleral spur). Therefore,
the overall forward movement of the muscle (including the muscle apex) could be
inferred from the forward ciliary body movement measured by UBM, and the
inward movement of the circular portion of the muscle could be inferred from
gonioscopically measured inward ciliary process movement. Comparisons of
forward vs. centripetal/inward ciliary body movement (measured by UBM and
goniovideography respectively) in living monkey eyes are made elsewhere and
are beyond the scope of this paper. [34]
There are only two regions in which the vitreous membrane is firmly attached to
structures of the accommodative system over the entire circumference: anteriorly
at the peripheral posterior lens capsule and posteriorly in the region of the ora
serrata. The intervening separation allows for the movement of the zonular
system needed for accommodation and disaccommodation. However, there are
Page 20 of 46IOVS

21 Vitreous Zonule
three other less-fixed fibrillar attachments, comprising the vitreous zonule, that
may stabilize and smooth the movement of the entire system and allow for finer
accommodative control and reduced stress on the ora serrata and the lens
capsule. This vitreous zonule, visualized by our improved SEM and UBM
techniques, is surprisingly robust and is intimately related to the anterior,
intermediate, and posterior vitreous membrane.
Our findings concerning the anterior vitreous zonule add new information to that
described previously for humans [9-14, 15 , 16, 17] and to goniovideography
observations previously reported for monkeys. [23] The earlier goniovideography
study included a video that showed the zonular attachments between the valleys
of the ciliary processes and Wieger’s ligament in vivo in the resting eye and
during accommodation. [23] The current SEM investigations indicate that, at most
places, the anterior vitreous membrane and posterior zonular tines of the anterior
zonular fork attaching to the posterior lens capsule are separated from each
other, presumably allowing unrestricted movement of the system. There are,
however, strands of anterior vitreous zonule that connect the zonular plexus with
the anterior vitreous membrane. This anterior vitreous zonule may help to
stabilize the anterior vitreous and thereby the position of the lens during
accommodation.
The cleft between the pars plana zonule of the ciliary body and the vitreous
membrane is bridged by bands of intermediate vitreous zonule fibers. The cleft
Page 21 of 46 IOVS

22 Vitreous Zonule
could provide a low-friction interface between the ciliary epithelium/pars plana
zonule and the vitreous membrane when the ciliary muscle moves anteriorly and
posteriorly during accommodation and disaccommodation, increasing the
efficiency of the system. The intermediate vitreous zonule may also stabilize the
vitreous membrane in its optimal contour, while dampening and smoothing the
muscle’s forward and inward movement.
The posterior vitreous zonule with its adjacent vitreous membrane, the pars
plana zonule, and the inner limiting membrane of the ciliary epithelium are joined
in an attachment zone of about 0.5 mm in monkeys and an astonishing 3-4 mm
in humans in anterior-posterior length, extending circumferentially around the eye
in the region of the ora serrata. This loose fibrillar attachment of the several
structures may also permit the forward-inward movement of the ciliary
muscle/zonular vitreous system in a dampened manner so as not to acutely
overstretch the peripheral retina and its attachment at the ora. This would
minimize the risk of tearing the peripheral retina upon accommodative effort. The
greater length and thickness of this zone in humans compared to monkeys may
reflect the need for protecting the retina over the much longer human lifespan.
With age, there is widening of the space between the ciliary body and vitreous
membrane and reduction of forward movement of the posterior insertion zone.
This could restrict forward movement of the zonular plexus upon accommodative
effort. However, our regression analysis showed that age itself was more
Page 22 of 46IOVS

23 Vitreous Zonule
important in predicting accommodative amplitude than was either the cleft width
or the forward movement of the posterior insertion zone. This is not surprising
given that there are likely other age-related changes occurring in the eye
concurrently that also affect accommodation.
Nonetheless, consistent with the hypothesis of age-related posterior restriction of
ciliary muscle/zonular movement, lysis of the posterior vitreous zonular system in
a few eyes increased the forward movement of the ciliary muscle upon central
electrical or peripheral pharmacological stimulation. While not a clinically
applicable technique, the findings and interpretations further support age-related
posterior restriction of ciliary muscle and zonular movement as both a
pathophysiologic factor and a potential therapeutic target in presbyopia.
Acknowledgements: The authors would like to thank Dr. Johannes Rohen for his immense help in preparing the specimens for SEM investigations, Elke Kretzschmar and Marco Gößwein for their technical assistance.
References 1. Lütjen-Drecoll, E., E. Tamm, and P.L. Kaufman, Age-related loss of
morphologic responses to pilocarpine in rhesus monkey ciliary muscle. Archives of Ophthalmology, 1988. 106: p. 1591-1598.
Page 23 of 46 IOVS

24 Vitreous Zonule
2. Tamm, E., M.A. Croft, W. Jungkunz, E. Lütjen-Drecoll, and P.L. Kaufman, Age-related loss of ciliary muscle mobility in the rhesus monkey: role of the choroid. Archives of Ophthalmology, 1992. 110: p. 871-876.
3. Croft, M., Glasser A, Heatley G, McDonald J, Ebbert T, Kaufman PL, The relationship between accommodative ciliary body and lens function in rhesus monkeys. I. Normal lens, zonule and ciliary process configuration in the iridectomized eye. Investigative Ophthalmology and Visual Science, 2006. 47(3): p. 1076-1086.
4. Strenk, S.A., J.L. Semmlow, I.M. Strenk, P. Munoz, J. Gronlund-Jacob, and J.K. DeMarco, Age-related changes in human ciliary muscle and lens: a magnetic resonance imaging study. Investigative Ophthalmology and Visual Science, 1999. 40: p. 1162-1169.
5. Poyer, J.F., P.L. Kaufman, and C. Flügel, Age does not affect contractile response of the isolated monkey ciliary muscle to muscarinic agonists. Current Eye Research, 1993. 12: p. 413-422.
6. Lütjen-Drecoll, E., E. Tamm, and P.L. Kaufman, Age changes in rhesus monkey ciliary muscle: Light and electron microscopy. Experimental Eye Research, 1988. 47: p. 885-899.
7. Gabelt, B.T., P.L. Kaufman, and J.R. Polansky, Ciliary muscle muscarinic binding sites, choline acetyltransferase and acetylcholinesterase in aging rhesus monkeys. Investigative Ophthalmology and Visual Science, 1990. 31: p. 2431-2436.
8. Tamm, E., E. Lütjen-Drecoll, W. Jungkunz, and J.W. Rohen, Posterior attachment of ciliary muscle in young, accommodating old, presbyopic monkeys. Investigative Ophthalmology and Visual Science, 1991. 32(5): p. 1678-1692.
9. Wieger, G., Ueber Canalis Petiti und ein Ligamentum hyaloideocapsulare. Diss. Inaug. Strassbourg, 1883.
10. Eisner, G., Biomicroscopy of the peripheral fundus. Heidelberg, Springerverlag, 1973.
11. Vail, D., The zonula of Zinn and ligament of Wieger. Transactions of the Ophthalmological Societies of the United Kingdom, 1957. 77: p. 441-499.
12. Eisner, G., Clinical examination of the vitreous. Transactions of the Ophthalmological Societies of the United Kingdom, 1975. 95: p. 360-363.
13. Streeten, B.W. and J.O. Pulaski, Posterior zonules and the lens extraction. Archives of Ophthalmology, 1977. 96: p. 132-138.
14. Bernal, A., J.M. Parel, and F. Manns, Evidence for posterior zonular attachment on the anterior hyaloid membrane. Investigative Ophthalmology and Visual Science, 2006. 47(11): p. 4708-4713.
15. Salzmann, M., Anatomie und Histologie des menschlichen. Augapfels. Wein, Franz Deuticke, 1912.
16. Eisner, G., Clinical anatomy of the vitreous. In: Tasman W, Jaeger EA,ed. /Duane's foundations of clinical ophthalmology. 2005, Philadelphia: Lippincott Williams & Wilkins. 34.
Page 24 of 46IOVS

25 Vitreous Zonule
17. Sebag, J., ed. Vitreous-from biochemistry to clinical relevance. In: Tasman W, Jaeger EA,ed. /Duane's foundations of clinical ophthalmology. 1998 ed. Vol. 1. 1998, Lippincott Williams & Wilkins: Philadelphia. 21.
18. Glasser, A., M.A. Croft, L. Brumback, and P.L. Kaufman, Ultrasound biomicroscopy of the aging rhesus monkey ciliary region. Optometry and Vision Science, 2001. 78: p. 417-424.
19. Rohen, J.W., Scanning electron microscopic studies of the zonular apparatus in human and monkey eyes. Investigative Ophthalmology and Visual Science, 1979. 18: p. 133-144.
20. Byrne, S.F. and R.L. Green, Ultrasound of the eye and orbit. Second Edition. ed. 2002, St. Louis, MO: Mosby Inc.
21. Wasilewski, R., J.P. McDonald, G. Heatley, E. Lütjen-Drecoll, P.L. Kaufman, and M.A. Croft, Surgical intervention and accommodative responses, II. Forward ciliary body accommodative movement is facilitated by zonular attachments to the lens capsule. Investigative Ophthalmology and Visual Science, 2008. 49(12): p. 5495-5502.
22. Ito, S. and M.J. Karnovsky, Formaldehyde-glutaraldehyde fixatives containing trinitro compounds. Journal of Cell Biology, 1968. 39: p. 168a.
23. Croft, M.A., J.P. McDonald, R.J. James, G.A. Heatley, T.L. Lin, E. Lütjen-Drecoll, and P.L. Kaufman, Surgical intervention and accommodative responses, I: Centripetal ciliary body, capsule, and lens movements in rhesus monkeys of various ages. Investigative Ophthalmology and Visual Science, 2008. 49(12): p. 5484-5494.
24. Crawford, K., E. Terasawa, and P.L. Kaufman, Reproducible stimulation of ciliary muscle contraction in the cynomolgus monkey via a permanent indwelling midbrain electrode. Brain Research, 1989. 503: p. 265-272.
25. Akaike, H., A new look at the statistical model identification. IEEE Transactions on Automatic Control, 1974. 19(6): p. 716-723.
26. May, C.A., N. Neuhuber, and E. Lütjen-Drecoll, Immunohistochemical Classification and Functional Morphology of Human Choroidal Ganglion Cells. Invest. Ophthalmol. Vis. Sci., 2004: p. 361-367.
27. von Helmholtz, H.H., Helmholtz's treatise on physiological optics, in Mechanism of accommodation., J.P.C. Southall, Editor. 1909, Dover Publications: New York. p. 143-172.
28. Coleman, D.J., Unified model for accommodative mechanism. American Journal of Ophthalmology, 1970. 69: p. 1063-1079.
29. Coleman, D.J., On the hydraulic suspension theory of accommodation. Transactions of the American Ophthalmological Society, 1986. 84: p. 846-868.
30. Coleman, D.J. and F. S.K., Presbyopia, accommodation, and the mature catenary. Ophthalmology, 2001. 108(9): p. 1544-51.
31. Wendt, M., Croft MA, McDonald J, Kaufman PL, Glasser A, Lens diameter and thickness as a function of age and pharmacologically stimulated accommodation in rhesus monkeys. Experimental Eye Research, 2008. 86: p. 746-752.
Page 25 of 46 IOVS

26 Vitreous Zonule
32. Rafferty, N.S., Structure, function, and pathology, in The ocular lens, H. Maisel, Editor. 1985, Marcel Dekker: New York. p. 1-60.
33. Coleman, D.J., R.H. Silverman, F.L. Lizzi, D.Z. Reinstein, M.J. Rondeau, H.O. Lloyd, and S.W. Daly, Ultrasonography of the Eye and Orbit. 2nd ed. 2006, Philadelphia: Lippincott Williams & Wilkins.
34. Croft, M.A., J.P. McDonald, N.V. Nadkarni, T.L. Lin, and P.L. Kaufman, Age-related changes in centripetal ciliary body movement relative to centripetal lens movement in monkeys. Experimental Eye Research, 2009. In Press.
Page 26 of 46IOVS

27 Vitreous Zonule
Figure legends Figure 1. Ultrasound Biomicroscopy (UBM) overview image (E-Tech) in a live
rhesus monkey shows a prominent straight line (arrow) extending from the pars
plicata region of the ciliary body to the ora serrata region, and separated from the
pars plana epithelium by a cleft, CP=ciliary processes, CB=ciliary body.
Figure 2. A 75-year-old human eye. A) Scanning electron micrograph of a sagittal
aspect of the anterior vitreous membrane (VM), the posterior tine of the zonular
fork (asterisk), and the posterior lens capsule (LC). B) Scanning electron
micrograph of the anterior VM and its attachment to the posterior LC. C) Careful
lifting of the VM reveals that the majority of the zonule (Z) inserts directly into the
posterior LC without attachment to the vitreous membrane that itself forms a
separate layer also directly inserting into the posterior LC. However, some
bundles of zonular fibers (anterior vitreous zonule, aVZ) do insert into the anterior
VM at spatial intervals of about 75 µm.
Figure 3. A) Scanning electron micrographs of a sagittal and oblique internal view
of the ciliary body (CB), the vitreous membrane (VM), the zonular plexus (ZP),
and the zonular connections bridging the cleft between the pars plana zonules
(ppZ) and VM (rhesus monkey, aged 8 years). The bridging bundles of zonular
fibers run from the region of the ZP in the valleys of the posterior pars plicata
towards the VM in the region of the ora serrata (asterisk). These zonular bundles
were termed intermediate vitreous zonule (iVZ). B) Scanning electron micrograph
showing the inner aspect of the posterior pars plicata (ppl), pars plana (pp), VM
and iVZ of a 10-year-old rhesus monkey. Anteriorly, the intermediate vitreous
zonular bundles split into a fork whose tines insert on both sides of the processes
in the valleys of the ppl (asterisk). Posteriorly, each main bundle splits into
several smaller bundles that merge with the VM (arrows).
Page 27 of 46 IOVS

28 Vitreous Zonule
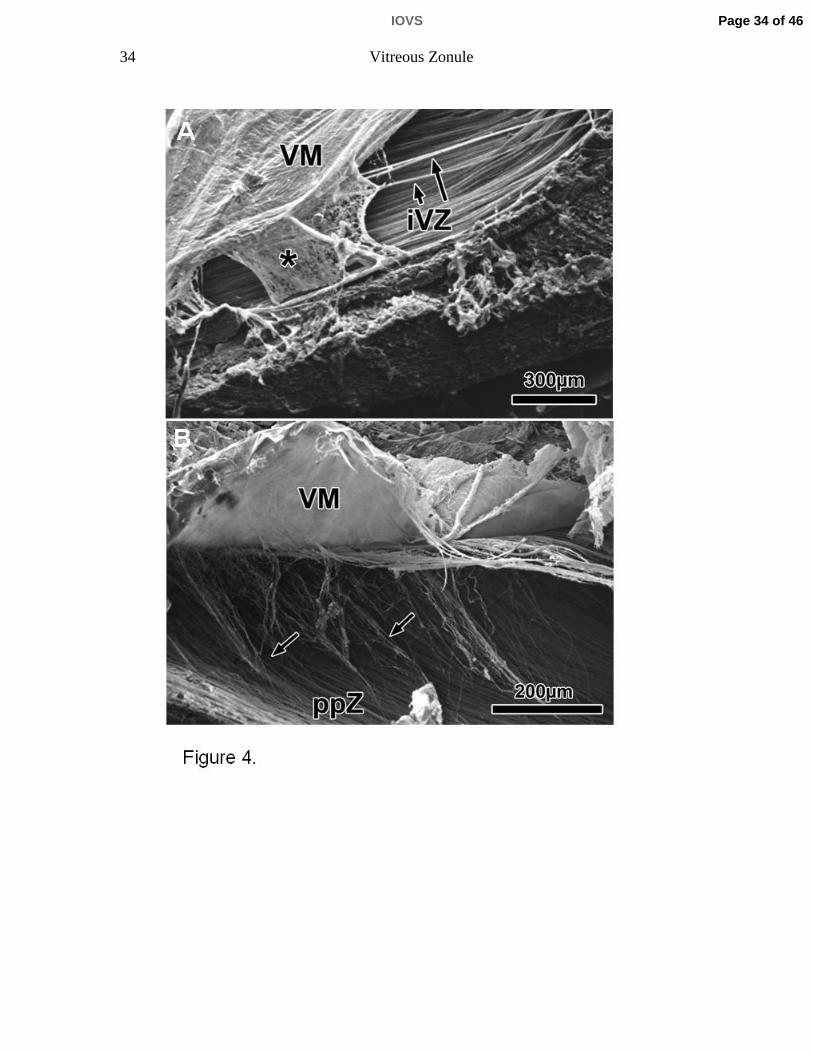
Figure 4. A) Scanning electron micrographs of sagittal sections of the ora serrata
region (rhesus monkey, aged 6 years). The vitreous membrane (VM), the
posterior extensions of the intermediate vitreous zonule (iVZ) and the pars plana
zonule form an interconnected sponge-like structure (asterisk). B) After careful
elevation of the VM, the connections of the ppZ to the posterior VM become
visible (arrows).
Figure 5. A) Histological sagittal section through a 100-year-old human eye
showing the posterior adherence of the vitreous membrane to the posterior pars
plana and ora region (asterisk). Note that this region in humans encompasses
nearly ½ the sagittal length between the scleral spur and ora serrata. B)
Scanning electron micrograph of the posterior vitreous zonule (sagittal aspect,
85-year-old human eye). Note the oblique-running fibrils between the pars plana
zonules (ppZ) and the vitreous membrane (VM); in contrast to the monkey (Fig. 4
B), these fibrils form a lattice-like structure (arrows).
Figure 6 A, B. UBM images of unaccommodated (A) and accommodated (B)
ciliary muscle in live rhesus monkey, aged 25 years. In these somewhat oblique
sections, one can see the pars plana zonules immediately adjacent to the pars
plana epithelium (arrowheads). This is more apparent in the accommodated than
in the unaccommodated state. The straight line between the ciliary processes
and the ora serrata represents the vitreous zonule. The numbers represent the
angle between the anterior face of the ciliary body and the inner surface of the
peripheral cornea, as defined by the white lines. Narrowing of the ciliary body
(CB)-Cornea angle in the accommodated vs. the unaccommodated state was
used as a surrogate indicator of forward ciliary body movement.
Figure 7. Ultrasound biomicroscopy (UBM) images were obtained in a 15-year-
old rhesus monkey. Care was taken to assure that the vitreous zonule appeared
as a continuous prominent straight line parallel to the focus line (asterisk)
Page 28 of 46IOVS

29 Vitreous Zonule
imprinted by the instrument within all images, indicating that these were true
sagittal sections. A) Anteriorly, the vitreous membrane is separated from the
vitreous zonule. The vitreous zonule fork (seen by SEM, Fig. 3 B) occurs just
posterior to its insertion to the zonular plexus and, as the fibers split at the fork,
they course out of the UBM image plane (arrowhead). Thus the white line that
represents the vitreous zonule appears to discontinue (arrowhead) immediately
posterior to reaching the ciliary process region in this image. This “drop out” of
the vitreous zonule strand is not always apparent in UBM images, due to the
orientation of the UBM probe and the close proximity of the vitreous zonule fork
to the ciliary processes (CP), as in Panel B.
Figure 8. Ultrasound biomicroscopy (UBM) image (A) analogous to SEM section
(B), and also panel A of Fig. 3 reproduced for ease of comparison, from the same
8-year-old rhesus monkey. CB=ciliary body, pp=pars plana zonule, VZ=vitreous
zonule, VM=vitreous membrane.
Figure 9. Ultrasound biomicroscopy obtained in 3 rhesus monkeys aged 6, 16,
and 25 years, showing the age-related change in cleft width (A, B, C). Cleft width
plotted vs. age (D) and accommodative amplitude (E). Note also the decreasing
width of the ciliary muscle overlying the widest part of the cleft, and the increased
curvature of the arc formed by the inner portion of the ciliary body.
Figure 10. Ultrasound biomicroscopy images in a 21-year-old rhesus monkey eye
before (A) and during (B) central electrical supramaximal stimulation of the
Edinger-Westphal nucleus. The distance between the scleral spur and the
posterior insertion zone of the vitreous zonule was measured in the
unaccommodated and accommodated states; CB=ciliary body; c=Cornea. C)
Distance between the scleral spur and vitreous zonule insertion zone in the
resting eye, and D) accommodative forward movement of the vitreous zonule
Page 29 of 46 IOVS

30 Vitreous Zonule
insertion zone, plotted vs. age. E) Accommodative amplitude plotted vs. the
forward movement of the vitreous zonule insertion zone during accommodation.
Figure 11. Scanning electron micrograph of the temporal region of a 9-year-old
rhesus monkey eye following a pars plana injection of -chymotrypsin. Note the
absence of intermediate vitreous zonular fibers in this region. VM=vitreous
membrane; ppZ=pars plana zonule; CB=ciliary body.
Figure 12. Semithin sections of the temporal regions of the right and left eye of a
9-year-old rhesus monkey. In the resting eye (A), the inner edge of the ciliary
muscle was located much more anteriorly following -chymotrypsin injection with
disruption of the intermediate vitreous zonule than in the contralateral non-
injected eye (B). (Sp=scleral spur, arrow=inner apex).
Figure 13. Ultrasound biomicroscopic images of the 25-year-old rhesus monkey
eye at rest and during supramaximal central stimulation to induce
accommodation. The left-hand panels are of the aphakic eye following ECLE.
The right-hand panels are after subsequent a-chymotrypsin lysis of the
intermediate vitreous zonule and surgical removal of the lens capsule (see
methods). The change in the ciliary body to cornea angle during maximal
accommodation (lower panels) is increased following lysis of the intermediate
vitreous zonule fibers (arrows), although the difference at rest (upper panels) is
minimal.
Page 30 of 46IOVS
31 Vitreous Zonule
Page 31 of 46 IOVS

32 Vitreous Zonule
Page 32 of 46IOVS

33 Vitreous Zonule
Page 33 of 46 IOVS
34 Vitreous Zonule
Page 34 of 46IOVS

35 Vitreous Zonule
Page 35 of 46 IOVS

36 Vitreous Zonule
Page 36 of 46IOVS

37 Vitreous Zonule
Page 37 of 46 IOVS

38 Vitreous Zonule
Page 38 of 46IOVS
39 Vitreous Zonule
Page 39 of 46 IOVS

40 Vitreous Zonule
Page 40 of 46IOVS

41 Vitreous Zonule
Page 41 of 46 IOVS
42 Vitreous Zonule
Page 42 of 46IOVS

Table 1. Accommodative Forward Movement of the Vitreous Zonule Posterior Insertion Zone in Monkey Eyes
Mean ± s.e.m. (mm) n pYoung 1 052 ± 0 067 4 0 001Young 1.052 ± 0.067 4 0.001Older 0.403 ± 0.118 4
Table 1. Data are mean ± s.e.m. (mm) accommodative forward movement of the intermediate posterior zonule’s posterior insertion zone during supramaximal central stimulation to induce accommodation in monkeys. The accommodative forward movement of the posterior insertion zone declined by ~60% in the older vs the
Table 1. Data are mean ± s.e.m. (mm) accommodative forward movement of the intermediate posterior zonule’s posterior insertion zone during supramaximal central stimulation to induce accommodation in monkeys. The accommodative forward movement of the posterior insertion zone declined by ~60% in the older vs the young monkey eyes. Young = 6 to 9, Older = 19 to 21 years old.
Page 43 of 46 IOVS

Table 2. Morphological Findings With and Without α-Chymotrypsin Treatment to Lyse the Vitreous Zonule in Monkey Eyes
A. Apical Position (mm) Muscle Width (mm)Age
Monkey (Years) Quadrant Temporal Inferior Superior Nasal Temporal Inferior Superior Nasaly ( ) p p p p1 cyno 3.50 Post Lysis 0.51 0.49 0.53 0.61 0.67 0.54 0.60 0.76
control 0.55 0.51 0.53 0.64 0.60 0.53 0.53 0.62
2 6-8 Post Lysis 0.32 0.55 0.57 0.76 0.60 0.60 0.70 0.59Control 0.59 0.72 0.55 0.47 0.68 0.51 0.59 0.60
3 9.00 Post Lysis 0.31 0.27 0.45 0.64 1.01 0.98 1.04 1.04control 0.53 0.64 0.63 0.39 0.86 0.80 0.85 0.80
4 9.50 Post Lysis 0.23 0.31 0.37 0.94 0.91 0.88 0.88control 0.76 0.85 0.85 0.55 0.67 0.72 0.73 0.79control 0.76 0.85 0.85 0.55 0.67 0.72 0.73 0.79
5* 25.00 Post Lysis 0.42 0.45 0.53 0.59 0.71 0.72 0.67 0.72control 0.64 0.56 0.58 0.47 0.63 0.73 0.71 0.64
B Diff i A i l P iti ( ) Diff i M l Width ( )B. Differences in Apical Position (mm) Differences in Muscle Width (mm)
α-Chymotrypsin Treated minus Control Eye α-Chymotrypsin Treated minus Control EyeQuadrant Temporal Inferior Superior Nasal Temporal Inferior Superior Nasal
Monkeyy1 cyno -0.04 -0.02 0 -0.03 0.07 0.01 0.07 0.14
2** -0.27 -0.17 0.02 0.29 -0.08 0.09 0.11 -0.01
3** -0.22 -0.37 -0.18 0.25 0.15 0.18 0.19 0.24
4** -0.53 -0.54 -0.48 0.27 0.19 0.15 0.09
Page 44 of 46IOVS

5* -0.22 -0.11 -0.05 0.12 0.08 -0.01 -0.04 0.08Mean -0.256* -0.242 -0.138 0.158 0.098 0.092 0.096 0.108Sem 0.079 0.094 0.092 0.072 0.057 0.042 0.039 0.041
p 0.044 0.062 0.209 NS 0.161 0.091 0.072 0.058
Table 2. Data are apical position and muscle width (A) and the differences in muscle apical position and muscle width between chymotrypsin treated and control monkey eyes (B) for the respective quadrants The eyes received Table 2. Data are apical position and muscle width (A) and the differences in muscle apical position and muscle width between ‐chymotrypsin‐treated and control monkey eyes (B) for the respective quadrants . The eyes received ‐chymotrypsin injection in the temporal quadrant to lyse the vitreous zonule (Post‐Lysis).The apical position of the muscle was farther anterior in ‐chymotrypsin injected eyes than in contralateral control eyes; the difference was most pronounced near the injection site (temporal quadrant; p=0.044, by two‐tailed paired t‐test). Overall the muscle widthtended to be thicker in the ‐chymotrypsin injected eyes than the contralateral control eyes but the change in width was not dependent upon proximity to injection site. NS=Not significant. Monkey #1 was a cynomolgus monkey and monkeys 2‐
Table 2. Data are apical position and muscle width (A) and the differences in muscle apical position and muscle width between ‐chymotrypsin‐treated and control monkey eyes (B) for the respective quadrants . The eyes received ‐chymotrypsin injection in the temporal quadrant to lyse the vitreous zonule (Post‐Lysis).The apical position of the muscle was farther anterior in ‐chymotrypsin injected eyes than in contralateral control eyes; the difference was most pronounced near the injection site (temporal quadrant; p=0.044, by two‐tailed paired t‐test). Overall the muscle widthtended to be thicker in the ‐chymotrypsin injected eyes than the contralateral control eyes but the change in width was not dependent upon proximity to injection site. NS=Not significant. Monkey #1 was a cynomolgus monkey and monkeys 2‐5 were rhesus monkeys. *Aphakic eye; **Pseudophakic eyes.
Page 45 of 46 IOVS

Table 3. Forward Ciliary Body Movement in Monkey Eyes as Measured by Ultrasound Biomicroscopy (degrees)Before and After α-Chymotrypsin
Post minus % FCB MovmentAge (Years) Monkey Pre α-Chymotrypsin Post α-Chymotrypsin Pre α-Chymotrypsin Increase
25* 1 37.4 66.9 29.5 78.9 25 1 37.4 66.9 29.5 78.915 2 30.4 44.9 14.5 47.726 3 13.2 31.3 18.1 137.18** 4 91 80.8 -10.2 -11.2
8.5** 5 61.7 100 38.3 62.18** 6 49.8 103.6 53.8 108.0
Mean 47 3 71 3 24 0 70 4Mean 47.3 71.3 24.0 70.4Sem 11.1 11.9 9.0 21.0
p= 0.044 0.02
Table 3. Data are forward ciliary body (FCB) movement in six rhesus monkey eyes before and after ‐chymotrypsin injection. FCB was determined as the narrowing of the angle that is formed between the inner aspect of the cornea and the anterior aspect of the ciliary body in the accommodated versus the unaccommodated state measured in degrees (see Fi 13) Th i d h t i i j ti i th t l d t M t t k i th i
Table 3. Data are forward ciliary body (FCB) movement in six rhesus monkey eyes before and after ‐chymotrypsin injection. FCB was determined as the narrowing of the angle that is formed between the inner aspect of the cornea and the anterior aspect of the ciliary body in the accommodated versus the unaccommodated state measured in degrees (see Fig. 13). The eyes received ‐chymotrypsin injection in the temporal quadrant. Measurements were taken in the region of the injection site. FCB movement was significantly higher Post α‐chymotrypsin vs Pre (p= 0.044) by two‐tailed paired t‐test. Monkeys 1‐3 were stimulated to accommodate by central electrical stimulation while monkeys 4‐6 were stimulated to accommodate pharmacologically by carbachol iontophoresis. *Aphakic eye (see Materials and Methods); **Pseudophakic eyes).
Table 3. Data are forward ciliary body (FCB) movement in six rhesus monkey eyes before and after ‐chymotrypsin injection. FCB was determined as the narrowing of the angle that is formed between the inner aspect of the cornea and the anterior aspect of the ciliary body in the accommodated versus the unaccommodated state measured in degrees (see Fig. 13). The eyes received ‐chymotrypsin injection in the temporal quadrant. Measurements were taken in the region of the injection site. FCB movement was significantly higher Post α‐chymotrypsin vs Pre (p= 0.044) by two‐tailed paired t‐test. Monkeys 1‐3 were stimulated to accommodate by central electrical stimulation while monkeys 4‐6 were stimulated to accommodate pharmacologically by carbachol iontophoresis. *Aphakic eye (see Materials and Methods); **Pseudophakic eyes).
Page 46 of 46IOVS