morphological adaptations of hot springs panic grass
TRANSCRIPT

ARTICLE IN PRESS
0306-4565/$ - se
doi:10.1016/j.jth
�CorrespondE-mail addr
Journal of Thermal Biology 33 (2008) 106–116
www.elsevier.com/locate/jtherbio
Morphological adaptations of hot springs panic grass(Dichanthelium lanigunosum var sericeum (Schmoll) to thermal stress
Gary M. Banowetza,�, Mark D. Azevedoa, Richard Stoutb
aUSDA/ARS, 3450 S.W. Campus Way, Corvallis, OR 97331, USAbDepartment of Plant Sciences and Plant Pathology, Montana State University, Bozeman, MT 59717-3150, USA
Received 10 May 2007; accepted 15 August 2007
Abstract
Although growth and productivity of most crops are significantly reduced by exposure to temperatures in excess of 35 1C, some plants
thrive in geothermally heated soils characterized by chronic temperatures in excess of 40 1C. The morphological adaptations that enable
these plants to colonize thermal environments are poorly understood. We characterized trichome development, silica accumulation,
stomatal density, and waxy cuticle development in populations of Dichanthelium lanuginosum var sericeum (Schmoll) isolated from
thermal and non-thermal environments to determine whether morphological changes existed in populations adapted to thermal
environments. Plants isolated from thermal environments of Yellowstone National Park developed an extensive series of trichomes when
exposed to chronic temperatures of 45/35 1C (day/night). In contrast, isolates from non-thermal environments of western Oregon showed
minimal trichome development during exposure to the elevated temperatures. Leaves that developed during exposure to elevated
temperatures had reduced thickness, though the reduction was less marked in plants from populations isolated from thermal
environments. Plants isolated from thermal environments also had greater numbers of stomata on adaxial leaf surfaces relative to
biotypes isolated from non-thermal environments. These results suggest phenotypic characteristics that may be useful in selection of new
varieties of crop species with improved tolerance to supraoptimal temperatures.
r 2007 Elsevier Ltd. All rights reserved.
Keywords: Leaf pubescence; Silica; Abiotic stress
1. Introduction
Plant adaptive responses to extreme soil and airtemperatures are critical to survival, plant productivity,and the capacity to compete successfully for resourceswithin the environment. Adaptive responses occur at manylevels of plant organization (Feder and Hofmann, 1999)and include heritable changes in stomatal physiology(Radin et al., 1994), increased photorespiration (Weis andBerry, 1988), altered fatty acid content of chloroplastmembranes (Murakami et al., 2000), enhanced resistance toloss of intrinsic protein structure from photo-oxidativedamage (Camejo et al., 2006), and stabilizing factorsincluding molecular chaperones (Feder and Hofmann,1999; Joshi et al., 1997; Ristic et al., 1998). Despite these
e front matter r 2007 Elsevier Ltd. All rights reserved.
erbio.2007.08.006
ing author. Tel.: +1541 750 8722; fax: +1 541 750 8750.
ess: [email protected] (G.M. Banowetz).
adaptive responses, most plants suffer reduced productivitywhen exposed to prolonged temperatures in excess of35 1C. Certain plants, however, successfully colonizeenvironments characterized by chronic exposure to soiland air temperatures in excess of 40–45 1C. The mechan-isms that permit plant survival in these extreme environ-ments are poorly understood.One environment where plant colonization is limited is
geothermally heated soil adjacent to geysers and hotsprings such as those found in Yellowstone National Parkin the US. Rhizospere temperatures in these soils arechronically in the range of 40–57 1C. The predominantspecies that colonizes these soils is Dichanthelium lanugi-
nosum var sericeum (Schmoll) (Stout et al., 1997), a C3
perennial grass that has wide geographical distribution inNorth America. D. lanuginosum also occurs in non-geothermal environments where they tend to existas isolated populations. Autogamy, the predominant

ARTICLE IN PRESSG.M. Banowetz et al. / Journal of Thermal Biology 33 (2008) 106–116 107
reproductive method in D. lanuginosum, increases thegenetic isolation of populations (Spellenberg, 1975) andpermits precise adaptation to specific environments(Stebbins, 1957). The species is considered an assemblageof intergradient forms and includes populations withcontrasting tolerance to high environmental temperatures(Spellenberg, 1975). Comparisons of heat-tolerance me-chanisms in D. lanuginosum individuals collected fromdiverse environments are limited. Stout et al. (1997)demonstrated that D. lanuginosum plants from Yellow-stone National Park expressed small molecular weightheat-shock proteins (HSPs) when exposed to 45 1C, but notwhen plants were grown or harvested at 25 and 32 1C.Expression of small molecular weight HSPs has beencorrelated with increased thermotolerance (Waters et al.,1996). Banowetz et al. (2007) found that populations of thisgrass that lacked the capacity to adapt to elevatedtemperatures produced greater levels of cellular chelationactivity associated with reduced quantities of superoxidedismutase activity.
The objective of this study was to compare morpholo-gical characteristics commonly associated with adaptationto growth at high temperature in individuals fromD. lanuginosum populations collected from contrastingenvironments ranging from geothermal areas of Yellow-stone National Park to non-thermal environments in theCascade Mountains of Oregon. We hypothesized thatindividuals collected from non-thermal areas would haveless genetic adaptation to growth and survival at elevatedtemperatures and that quantifiable differences between thegenotypes would be apparent at multiple levels of plantorganization. We examined leaf pubescence and morphol-ogy, stomatal density, temperature-associated leaf andtiller development, and accumulation of silica and silica-containing bodies in these isolates.
2. Materials and methods
2.1. Plant material and growth conditions
D. lanuginosum plants were collected from two sites withthermally heated soils in Yellowstone Park includingRabbit Creek, characterized by warm moist soils, and asite in the Norris Basin characterized by warm, dry soils.The two sites are approximately 32 km distant from eachother. Specimens also were collected from a site that wasnot thermally heated adjacent to the Umpqua River,Douglas, County, OR, and from Belknap Springs, a sitewith hot springs in Lane County, OR. The plants weretransferred to flats that contained coarse vermiculite,fertilized weekly with 18:18:18 (N:P:K) liquid fertilizer(243 ppmN) and grown in a greenhouse to permitacclimation. For the growth comparison study, singleramets of greenhouse-grown plants from each collectionsite were transferred to 15 cm pots containing vermiculite(12 pots/site). Plants utilized for leaf morphology examina-tion and silica content evaluation were prepared by
transplanting 12 ramets from each population to flatscontaining vermiculite (2 flats/population). The pots andflats were fertilized as above and placed into a growthchamber that provided a 12 h photoperiod, 25/20 1C (day/night) with 500 mmol/m2/s photosynthetically active radia-tion (PAR) and allowed to acclimate for 1 week. Heattreatment consisted of transferring half of the total numberof pots and flats to a second growth chamber (35/25 1Cday/night, 12 h photoperiod, 500 mmol/m2/s PAR). Thetemperature of the second chamber was then increased in2 1C increments/day until a final day/night temperatureregime of 45/35 1C was achieved. Plants maintained atelevated temperatures were watered twice daily, while theambient condition-grown plants were watered daily. Bothheat-acclimated and ambient treated plants were fertilizedweekly with equivalent amounts of fertilizer. Plant growthwas quantified after 8 weeks at the elevated temperature.Morphology and silica content data were collected fromleaves that emerged during the elevated temperature regime(about 4 weeks at the final elevated temperatures). Anotherset of plants was maintained in a glasshouse under naturalspring/summer lighting conditions (Corvallis, Oregon44.31N, 123.151W). Unless otherwise noted, penultimatefully expanded leaves detached from replicate plants wereused for physiological measurements.
2.2. Microscopy
2.2.1. Leaf cross sections
Three leaf sections collected from the base of penulti-mate leaves on vegetative tillers from plants maintained at45/35 or 25/20 1C were fixed in formalin-acid-alcohol(FAA), embedded in Leica Historesins glycol methacry-late plastic, sectioned at 5 mm on a rotary microtome,stained with 0.5% TBO (pH ¼ 4.2) in citrate buffer, andexamined using differential interference contrast lightmicroscopy. Photomicrographs were prepared at 100�magnification.
2.2.2. Leaf clearing and polarized light microscopy
Penultimate leaves from vegetative tillers were cleared byboiling in a 1:1:4 mixture of lactic acid:glycerol:ethanol for30min, which included replacement of the clearing solutionafter the first 15min, washed in deionized water, andincubated in methanol at 22 1C for 16 h to remove residualleaf pigmentation (O’Brien and McCully, 1981). Leafsections (1 cm) of cleared leaf tissues were rinsed indeionized water, mounted on glass slides with glycerol,and observed with phase contrast and simple polarizedlight microscopy at 160–400� magnifications. Spodo-grams of leaf sections were prepared to confirm clearedtissue observations (Parry and Smithson, 1958). Sampleswere mounted under a coverslip on glass slides and heatedat 500 1C for 4 h. After 16 h of cooling, glycerin was addedas a mounting medium and the remaining silica structureswere observed using phase contrast and polarized lightmicroscopy. Photomicrographs of the same field of view

ARTICLE IN PRESSG.M. Banowetz et al. / Journal of Thermal Biology 33 (2008) 106–116108
were prepared using both phase contrast and polarizedlight for visualization of silica-containing cells.
2.2.3. Scanning electron microscopy
Three penultimate leaves from vegetative tillers of plantsgrown at 45/35 or 25/20 1C were collected from eachpopulation, pressed flat, and air dried at 40 1C overnight.Leaf sections (1 cm) were mounted on edge (vertically) onaluminum mounts (Pelco # 16262) to permit both edge andsurface views using specimen tilt and rotation options. Leafspecimens were glued onto the mounts using DUCOcement (Pelco # 14445), coated with �10nm of 60/40wt%
B.T.
S
V.B.
V.B.
V.B.
B.T.
S.
S
S.C.
S.C.
S.C.
25° C/ 20° C
Fig. 1. Changes in the adaxial leaf surface of Dichanthelium lanuginosum selecti
Scanning electron micrographs of the adaxial leaf surface of D. lanuginosum co
and two Yellowstone National Park (KD8, panels E and F; KD9, panels G an
that developed at 25/20 1C (day/night) with 12 h of 500mmol/m2/s photosynth
leaves that emerged from plants adapted to 45/35 1C in identical lighting condit
B.T. ¼ bicellular trichome, S.C. ¼ bilobate silica cell, V.B. ¼ vascular bundle.
gold/palladium alloy applied in an Edwards S150B SputterCoating system initiated from a vacuum of 1.5 Torrvacuum, and carried out at 5mb argon pressure using a20mA plasma current at 1.5 kV for 60 s. The specimenswere observed and images were collected using an Amray3300FE scanning electron microscope. The experiment wasrepeated two times.
2.2.4. Mineral analysis
Soil samples collected from each geographic collectionsite immediately adjacent to D. lanigunosum plants wereplaced into 15ml plastic centrifuge tubes and stored at
S.T.
V.B.
S.T.
S.T.
S.
S.
V.B.
S.T.
S.
V.B.
S.C.
V.B.
45° C/ 35° C
S.
ons with contrasting thermotolerance when grown at elevated temperature.
llected from two Oregon (KD 10, panels A and B; KD7, panels C and D)
d H) populations. Panels A, C, E, and G are photomicrographs of leaves
etically active radiation. Panels B, D, F, and H are photomicrographs of
ions. Bar ¼ 100mm. Abbreviations: S. ¼ stomate, S.T. ¼ simple trichome,

ARTICLE IN PRESSG.M. Banowetz et al. / Journal of Thermal Biology 33 (2008) 106–116 109
�20 1C until they were analyzed for mineral content.Approximately 250mg of ground oven dry soil was usedfor each analysis. Three samples of each leaf blade (minusthe leaf sheaths) from four vegetative tillers were collectedfrom each D. lanigunosum population grown at either 45/35or 25/20 1C. Leaf samples were dried at 90 1C for 16 h andground in a tissue grinder.
Minerals from both soil and leaf samples were extractedutilizing microwave-assisted acid digestion (EPA method3052) with an Ethos D microwave station (Milestone,Monroe, CT). Soil extracts were analyzed for Ni, Zn, As,Se, Ag, Cd, Ba, Hg, and Pb, while leaf extracts wereanalyzed for Si by using inductive coupled plasma optical
25° C/ 20° C
V.B.
S.T.
B.T.
B.T.
V.B.
B.T.
S.T.
V.B.
V.B.
S.
V.B.S.
V.B.
S.
B.T.
S.
S.C.
Fig. 2. Comparison of the abaxial leaf surfaces of Dichanthelium lanuginosu
temperatures. Scanning electron micrographs of abaxial leaf surface of D. lanug
and D) and two Yellowstone National Park (KD8, panels E and F; KD9, pa
developed at 25/20 1C (day/night) with 12 h of 500mmol/m2/s photosynthetica
plants adapted to 45/35 1C in identical lighting conditions. Note increased simp
Bar ¼ 100mm. Abbreviations: S. ¼ stomate, S.T. ¼ simple trichome, B.T. ¼ b
emission spectroscopy (Perkin-Elmer Life and AnalyticalSciences, Shelton, CT).
3. Results
3.1. Temperature and leaf pubescence
Scanning electron micrographs showed that leaf pub-escence (trichome development) increased on abaxial andadaxial leaf surfaces in response to growth at elevatedtemperatures (Figs. 1 and 2). Leaves from plants grown at45/35 1C developed thicker trichomes at an increaseddensity along with a surface of roughened cuticular waxes.
V.B. S.T.
S.T.
V.B.
S.T.
S.T.
B.T.
V.B.
B.T.
S.C.
S.
B.T.
S.C.
B.T.V.B.
45° C/ 35° C
m selections with contrasting thermotolerance during growth at elevated
inosum collected from two Oregon (KD 10, panels A and B; KD7, panels C
nels G and H) populations. Panels A, C, E, and G represent leaves that
lly active radiation. Panels B, D, F, and H are leaves that emerged from
le trichome development and density in leaves that developed at 45/35 1C.
icellular trichome, S.C. ¼ bilobate silica cell, V.B. ¼ vascular bundle.

ARTICLE IN PRESSG.M. Banowetz et al. / Journal of Thermal Biology 33 (2008) 106–116110
The increased pubescence was greatest in D. lanuginosum
isolates from the thermal environments of YellowstonePark and least in isolates from non-thermal environmentsof western Oregon. The entire epidermis of thermallyadapted plants was coated with a roughened waxy cuticle(Fig. 3). The leaf surface of plants grown at 25/20 1C had asmoother coating of wax that did not cover the entireepidermis. Increased trichome density was especiallyapparent in side views of leaf surfaces from heat-adaptedplants where isolates from the thermal environments ofYellowstone National Park had increased pubescencerelative to the other isolates (Fig. 4).
25° C/ 20° C
S.T.
S.
S.
S.
S.
B.T.
B.T.
V.B.
B.T.
Fig. 3. Comparisons of epicuticular wax development on the adaxial leaf surfa
Scanning electron micrographs of adaxial leaf surfaces of D. laniginosum collec
two Yellowstone National Park (KD8, panels E and F; KD9, panels G and H)
at 25/20 1C (day/night) with 12 h of 500mmol/m2/s photosynthetically active rad
from plants adapted to 45/35 1C in identical lighting conditions. Bar ¼ 20mmtrichome, V.B. ¼ vascular bundle, R.W. ¼ roughened wax on the cuticle surfa
3.2. Cellular adaptations to temperature
Tissue sections from the base of penultimate leaves ofvegetative tillers showed that exposure of all D. lanugino-
sum plants to 45/35 1C reduced apparent leaf thickness. Thereduction was greatest in isolates from non-thermalenvironments (Fig. 5). Leaf thickness reduction of approxi-mately 32% occurred on leaves of Oregon isolatescompared with a reduction of 17% in isolates from thethermal environments of Yellowstone National Park.Differences in leaf thickness and distance between vascularbundles across each leaf appeared to be due to a reduction
S.T.
S.T.
S.T.
S.
S.
S.
S.
R. W.
R. W.
B.T.
R. W.
R. W.
45° C/ 35° C
ce of Dichanthelium lanuginosum isolates with contrasting thermotolerance.
ted from two Oregon (KD 10, panels A and B; KD7, panels C and D) and
populations. Panels A, C, E, and G are photomicrographs of leaves grown
iation. Panels B, D, F, and H are photomicrographs of leaves that emerged
. Abbreviations: S. ¼ stomate, S.T. ¼ simple trichome, B.T. ¼ bicellular
ce.
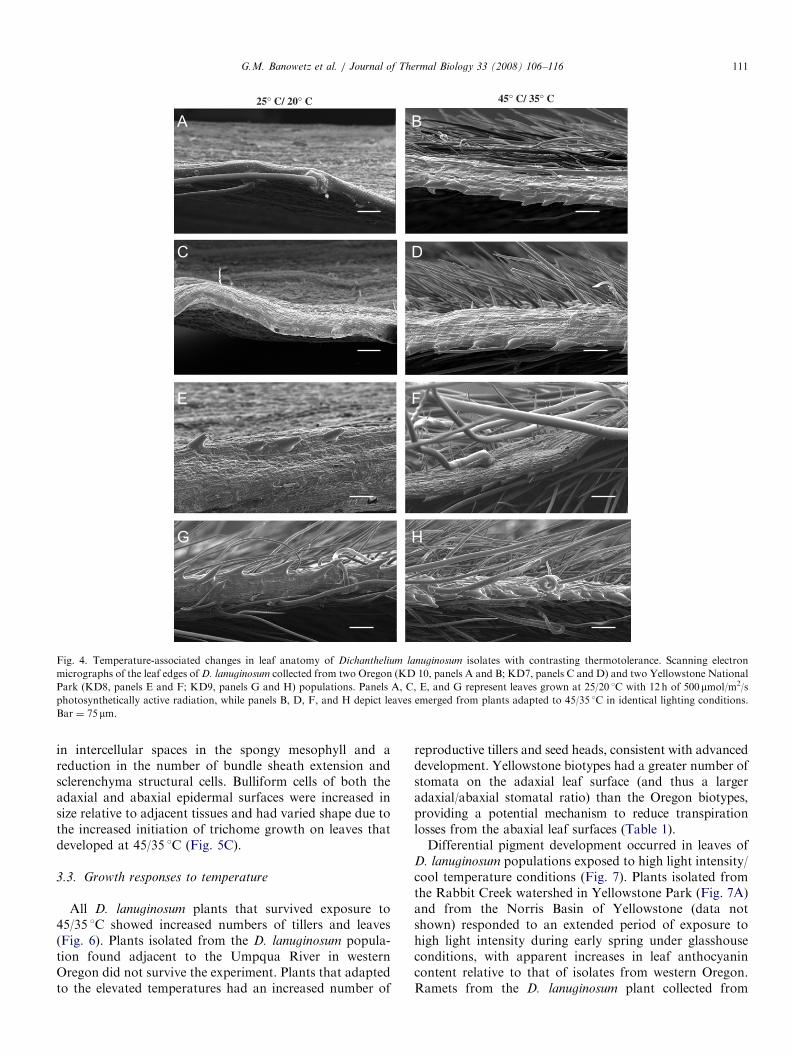
ARTICLE IN PRESS
25° C/ 20° C 45° C/ 35° C
Fig. 4. Temperature-associated changes in leaf anatomy of Dichanthelium lanuginosum isolates with contrasting thermotolerance. Scanning electron
micrographs of the leaf edges of D. lanuginosum collected from two Oregon (KD 10, panels A and B; KD7, panels C and D) and two Yellowstone National
Park (KD8, panels E and F; KD9, panels G and H) populations. Panels A, C, E, and G represent leaves grown at 25/20 1C with 12 h of 500mmol/m2/s
photosynthetically active radiation, while panels B, D, F, and H depict leaves emerged from plants adapted to 45/35 1C in identical lighting conditions.
Bar ¼ 75 mm.
G.M. Banowetz et al. / Journal of Thermal Biology 33 (2008) 106–116 111
in intercellular spaces in the spongy mesophyll and areduction in the number of bundle sheath extension andsclerenchyma structural cells. Bulliform cells of both theadaxial and abaxial epidermal surfaces were increased insize relative to adjacent tissues and had varied shape due tothe increased initiation of trichome growth on leaves thatdeveloped at 45/35 1C (Fig. 5C).
3.3. Growth responses to temperature
All D. lanuginosum plants that survived exposure to45/35 1C showed increased numbers of tillers and leaves(Fig. 6). Plants isolated from the D. lanuginosum popula-tion found adjacent to the Umpqua River in westernOregon did not survive the experiment. Plants that adaptedto the elevated temperatures had an increased number of
reproductive tillers and seed heads, consistent with advanceddevelopment. Yellowstone biotypes had a greater number ofstomata on the adaxial leaf surface (and thus a largeradaxial/abaxial stomatal ratio) than the Oregon biotypes,providing a potential mechanism to reduce transpirationlosses from the abaxial leaf surfaces (Table 1).Differential pigment development occurred in leaves of
D. lanuginosum populations exposed to high light intensity/cool temperature conditions (Fig. 7). Plants isolated fromthe Rabbit Creek watershed in Yellowstone Park (Fig. 7A)and from the Norris Basin of Yellowstone (data notshown) responded to an extended period of exposure tohigh light intensity during early spring under glasshouseconditions, with apparent increases in leaf anthocyanincontent relative to that of isolates from western Oregon.Ramets from the D. lanuginosum plant collected from

ARTICLE IN PRESS
B B
B
B
B B
I.S. I.S.
I.S.
I.S.I.S.
V.B.
V.B.
V.B.
V.B.
V.B. V.B.
S S
S
S
S.M.
S.M.
S.M.
S.M. S.M.
B.S.
B.S. B.S.
B.S.
B.S.
B.S.
S S
B
B
S
T
B B
S
B
T
B
Sc
Sc
Sc
Sc Sc
Sc
Sc Sc Sc
Sc
I.S.
S.M.
S
Fig. 5. Photomicrographs of leaf cross sections showing temperature-associated changes in internal leaf anatomy of Dichanthelium lanuginosum isolates
with contrasting thermotolerance. The top image in each panel is a cross section of the penultimate leaf from a vegetative tiller grown at 25/20 1C with 12 h
of 500mmol/m2/s photosynthetically active radiation (PAR). The lower image is a comparable region of a penultimate leaf from a vegetative tiller from the
same plant clone grown at 45/35 1C under identical lighting. Panel A, Dichanthelium lanuginosum collected from Belknap Hot Springs, Oregon (KD6).
Panel B, Dichanthelium lanuginosum collected from a site near the Umpqua River in western Oregon (KD 10). Panel C, Dichanthelium lanuginosum
collected from Rabbit Creek, Yellowstone National Park, Wyoming (KD 9). Photographs were prepared from differential interference contrast light
microscopy images at 100� total magnification (bar ¼ 250mm). Abbreviations: B ¼ bulliform cell, S ¼ stomate, T ¼ trichome, Sc ¼ sclerenchyma
structural cells, I.S. ¼ intercellular spaces of the spongy mesophyll, S.M. ¼ spongy mesophyll, B.S. ¼ bundle sheath including extension cells,
V.B. ¼ vascular bundle.
G.M. Banowetz et al. / Journal of Thermal Biology 33 (2008) 106–116112
Belknap Hot Springs in Oregon grown under the sametemperature and light regime did not shift from chlorophyllto anthocyanin pigmentation (Fig. 7B).
3.4. Silica formation with temperature stress
Development of silica-containing bodies and structureswas examined because silica accumulation is associatedwith tolerance to a variety of plant stresses. Spodogramsprepared from leaves of Yellowstone populations showedthat D. lanuginosum contained abundant silica bodies andsilica-containing structures (Fig. 8) where silica-containingstructures in the figure appear highly refractive underpolarized light. The spodogram in Fig. 8A provides anoverview of the carbonized structural organization of leafepidermal non-silica-containing cells outlined in black. Theimage also shows structural details of the bilobate cells of
the vascular bundle and remnants of the fragmented simpletrichomes. The birefringence observed in the same field ofview under polarized light confirmed the presence of silicain the bilobate cells, the lining of the stomatal cells, and theremnants of the simple trichome fragments (Fig. 8B).Similarly, the phase contrast image of the abaxial leafsurface of a cleared leaf shows an overview of the structuralorganization of the leaf epidermis including the simpletrichomes, stomates, and the vascular bundle (Fig. 8C).The polarized light image of the same leaf surface and leafmargin confirmed the presence of silica (Fig. 8D).The silica structures included a variety of trichomes,
spur-like structures along the leaf margin, and bilobatesilica cells in the epidermis adjacent to vascular bundles.Relative to leaf development at 25/20 1C, silica content inleaves that developed at 45/35 1C increased significantly inall populations with the exception of the one that

ARTICLE IN PRESS
25° C/ 20° C 45° C/ 35° C
Fig. 6. Temperature-associated changes in the aerial canopy of Dichanthelium lanuginosum isolates with contrasting thermotolerance. Photographs in left
panels show plants from populations grown at 25/20 1C (day/night) with 12 h of 500mmol/m2/s photosynthetically active radiation (PAR) for 8 weeks.
Right panels show plants from the same populations grown at 45/35 1C. Panels A and B, isolate from Belknap Hot Springs, Oregon (KD6). Panels C and
D, isolate from Deer Creek near McKenzie River, Oregon (KD 7). Panel E and F, isolate from Susan Creek near Umpqua River, Oregon (KD 10). Panel
G and H, isolate from Rabbit Creek, Yellowstone National Park, Wyoming (KD 9). Bar ¼ 2.54 cm.
G.M. Banowetz et al. / Journal of Thermal Biology 33 (2008) 106–116 113
originated from the non-thermal region near the UmpquaRiver in western Oregon (Table 2). Relative to thepopulation isolated from a site adjacent to the UmpquaRiver, the three populations that showed increased silicacontent after growth at 45/35 1C contained significantlylower silica content when grown at 25/20 1C.
4. Discussion
The capacity of D. lanuginosum to adapt to growth atelevated temperatures appears to be associated with theorigin of the population, suggestive of genotypic differ-ences in thermotolerance. Morphological adaptations in

ARTICLE IN PRESS
Table 1
Stomate density on leaves of Dichanthelium lanuginosum collected from western Oregon and Yellowstone National Park, Wyoming
Geographic origin of clone Stomata/vascular bundle/mm of leafa
Adaxial surfaceb Abaxial surface Adaxial/abaxial
Deer Creek, Oregon (Xeric site adjacent to the McKenzie, River) 62.6 39.3 1.59
Belknap Hot Springs, Oregon (adjacent to hot spring discharge) 69.5 36 1.93
Norris Basin, YNP, Wyoming (adjacent to fumaroles in 40 1C+ soil) 94.8 44.5 2.13
Rabbit Creek, YNP, Wyoming (Xeric site with active geothermal activity) 91.7 32.1 2.86
aRatios were determined by quantifying the stomata on both sides of a single vascular bundle across a 420mm microscope field of view (200� ).bMean values (n ¼ 10).
A B
Fig. 7. High light intensity/cool temperature-associated changes in leaf pigment development in isolates of Dichanthelium lanuginosum. Panel A represents
a plant collected from Rabbit Creek, Yellowstone National Park, Wyoming (KD 9) exposed to springtime greenhouse conditions of high light intensity.
Panel B depicts D. lanuginosum collected from Belknap Hot Springs in Oregon grown under identical temperature and light conditions. Bar ¼ 2.54 cm.
T.M.C.
S.T.
S.C.
V.B.
V.B.
L.M.
S.T.
V.B.
S
V.B.
S.C.S.
S.
V.B.
S.T.F. S.T.F.
V.B.
Fig. 8. Silica-containing bodies from leaves of Dichanthelium lanuginosum. Panel A is a phase contrast photomicrograph of a spodogram of the basal
section of a penultimate leaf from a vegetative tiller of D. lanuginosum. Panel B is the same field of view observed using polarized light microscopy. Panel C
is a polarized light microscopy image of the abaxial leaf surface which has been cleared to view silica-containing structures. Panel D is a phase contrast
image of a cleared leaf section showing the abaxial face of the leaf margin. Bar ¼ 25mm for panels A, B, and D, and 150mm for panel C. Abbreviations:
S. ¼ stomate, S.T. ¼ simple trichome, T.M.C. ¼ trichome mother cell, L.M. ¼ leaf margin, V.B. ¼ vascular bundle, S.C. ¼ bilobate silica cell from the
epidermis about the vascular bundle, S.T.F. ¼ simple trichome fragments.
G.M. Banowetz et al. / Journal of Thermal Biology 33 (2008) 106–116114

ARTICLE IN PRESS
Table 2
Silica content of leaf blades from isolates of Dichanthelium lanuginosum with contrasting thermotolerance
Geographic origin of clone Clone Growth temperature
(1C)
Leaf silica content (mg/g dry wt.)
95% confidence interval 73012
Susan Creek Campground, Oregon (non-thermal site) KD10 25/20 8273
45/35 7446
Deer Creek, Oregon (Xeric site adjacent to the McKenzie
River)
KD7 25/20 4238
45/35 11,495
Norris Basin, YNP, Wyoming (Xeric site with active
geothermal activity)
KD8 25/20 4736
45/35 10,540
Rabbit Creek, YNP, Wyoming (Xeric site with active
geothermal activity)
KD9 25/20 3766
45/35 13,700
Ramets of greenhouse-grown plants collected from each geographic location were propagated in a growth chamber at 25/20 1C (day/night) with 12 h of
500mmol/m2/s photosynthetically active radiation (PAR) or adapted to growth at 45/35 1C under identical conditions. Data represent means and 95%
confidence interval (n ¼ 3) after 4 weeks of growth at adapted temperatures.
G.M. Banowetz et al. / Journal of Thermal Biology 33 (2008) 106–116 115
leaves that develop at elevated temperatures includeincreased pubescence and development of a waxy cuticle.Increased trichome density and pubescence are associatedwith improved thermotolerance in woody species includingthe shrub Wigandia urens (Perez-Estrada et al., 2000),Mediterranean oaks (Morales et al., 2002), grapes (Liako-poulos et al., 2006), and olives (Ennajeh et al., 2006).Increased trichome density and leaf surface developmentreflects or absorbs visible light and modifies the quality andquantity of light reaching the chloroplasts (Karabourniotisand Bornman, 1999; Liakopoulos et al., 2006). Lightreflection and shading reduce the potential for oxidativedamage to photosynthetic membranes associated withexcess light energy. The protection these adaptationsprovide to reduce oxidative damage is enhanced bybiochemical mechanisms. Banowetz et al. (2007) foundthat populations of D. lanuginosum isolated from thermalenvironments maintained greater levels of superoxidedisumtase during exposure to 45 1C, and suggested thatthe increased anti-oxidative potential was associated withthermotolerance.
Adaptation in leaf surface characteristics during pro-longed exposure to elevated temperatures potentiallyimproves thermotolerance by reducing transpiration andleaf surface temperature. Increased trichome densitywidens the air boundary layer around the stomata, anadaptation that reduces transpiration and increases photo-synthetic efficiency. It is likely that increased trichomedensity and surface area, particularly on the abaxial leafsurface, also increase dew accumulation and provide acooling effect. Leaf cooling reduces water permeability(Riederer and Schreiber, 2001). Increased trichome forma-tion occurs at the expense of bulliform cell functionbecause the trichome mother cells are derived from thebulliform cells. This is one potential reason why the abaxialleaf surface has greater trichome density than the adaxial
surface. Adaxial bulliform cells play a critical role in theleaf-rolling process, an adaptation to reduce leaf desicca-tion. The waxy cuticle coating the epidermis on the adaxialsurface provides a hydrophobic surface that reducesdesiccation (Riederer and Schreiber, 2001). In grasses, itis likely that the cuticle enables water to form droplets onthe adaxial surface of the leaf, which coalesce with otherdroplets that are channeled to the base of the plant andtaken up by the root system.Total silica content in leaves that developed at 45/35 1C
increased significantly in all populations with the exceptionof the one originating from the non-thermal region near theUmpqua River in western Oregon. This increase in totalsilica content coincided with the increased trichome densityassociated with the more thermotolerant populations.Polarized microscopy of dendograms and clear leaf tissueshowed that the trichomes were primarily composed ofsilica. It is possible that D. lanuginosum isolates fromthermal environments increase the uptake of soluble silicaand form trichomes of insoluble silica in response tothermal conditions. This increased trichome density underthermal stress appears to be genetically controlled.All YNP plant biotypes produced anthocyanin pigmen-
tation in response to the high light cool temperatureconditions, while the Oregon biotypes did not. Thisadaptation may enhance tolerance to photooxidative stressduring bright spring days and cool nights found at the highaltitude of YNP. The role of pigmentation shift duringoxidative stress responses including thermotolerance re-mains unclear (Steyn et al., 2002). Merzlyak and Chivku-nova (2000) compared light absorption by apples withcontrasting anthocyanin production and suggested thatthese pigments protected against light-induced stress. Fieldet al. (2001) proposed a similar role for anthocyanins basedon their studies of light absorption by leaves of dogwood.In contrast, Burger and Edwards (1996) did not detect any

ARTICLE IN PRESSG.M. Banowetz et al. / Journal of Thermal Biology 33 (2008) 106–116116
differences between red- and green-pigmented coleus in theamount of photoinhibition observed when plants wereexposed to visible light. Anthocyanin pigmentation did,however, reduce the apparent amount of damage whenthese plants were exposed to UV irradiation at 15 1C.
It is most likely that a combination of biochemical andmorphological adaptations contribute significantly to thethermotolerance and general stress tolerance in popula-tions of D. lanuginosum isolated from thermal environ-ments. This study did not attempt to determine whetherthese adaptations were heritable, although the possibilityexists that crosses between genotypes with contrastingthermotolerance would produce progeny with enhancedresistance to heat stress. The morphological adaptationsidentified here may serve as useful markers in subsequentgenetic attempts to improve thermotolerance in grassesgrown as crops.
Acknowledgments
We wish to express appreciation to Al Soeldner and theElectron Microscopy Center, Oregon State University(OSU), for S.E.M. images, Kathy Cook and the Micro-technique Laboratory, Dept. of Horticulture, OSU, forembedment and sectioning of leaves and Machelle NelsonUSDA-ARS, NFSPRC, Corvallis, Oregon, for silicaanalysis of soil and leaf samples. The use of trade, firm,or corporation names in this publication is for theinformation and convenience of the reader. Such use doesnot constitute an official endorsement or approval by theUnited States Department of Agriculture or the Agricul-tural Research Service of any product or service to theexclusion of others that may be suitable.
References
Banowetz, G.M., Azevedo, M.D., El-Nashaar, H.M., Martin, R.C., Stout,
R.G., 2007. Temperature-induced increase in cellular chelating
potential associated with reduced thermotolerance. J. Thermal Biol.
32, 12–19.
Burger, J., Edwards, G.E., 1996. Photosynthetic efficiency, and photo-
damage by UV and visible radiation, in red versus green leaf Coleus
varieties. Plant Cell Physiol. 37, 395–399.
Camejo, D., Jimenez, A., Alarcon, J.J., Torres, W., Gomez, J.M., Sevilla,
F., 2006. Changes in photosynthetic parameters and antioxidant
activities following heat-shock treatment in tomato plants. Funct.
Plant Biol. 33, 177–187.
Ennajeh, M., Vadel, A.M., Khemira, H., Ben Mimoun, M., Hellali, R.,
2006. Defense mechanisms against water deficit in two olice (Olea
europaea L.) cultivars ‘Meski’ and ‘Chemlali’. J. Hort. Sci. Biotechnol.
81, 99–104.
Feder, M.E., Hofmann, G.E., 1999. Heat-shock proteins, molecular
chaperones, and the stress response. Ann. Rev. Physiol. 61, 243–282.
Field, T.S., Lee, D.W., Holbrook, N.M., 2001. Why leaves turn red in
autumn. The role of anthocyanins in senescing leaves of red-Osier
dogwood. Plant Physiol. 127, 566–574.
Joshi, C.P., Klueva, N.Y., Morrow, K.J., Nguyen, H.T., 1997. Expression
of a unique plastid-localized heat-shock protein is genetically linked to
acquired thermotolerance in wheat. Theor. Appl. Genet. 95, 834–841.
Karabourniotis, G., Bornman, J.F., 1999. Penetration of UV-A, UN-B
and blue light through the leaf trichome layers of two xeromorphic
plants, olive and oak, measured by optical fibre microprobes. Physiol.
Plant 105, 655–661.
Liakopoulos, G., Nikolopoulos, D., Klouvatou, A., Vekkos, K.-A.,
Manetas, Y., Karabourniotis, G., 2006. The photoprotective role of
epidermal anthocyanins and surface pubescence in young leaves of
grapevine (Vitus vinifera). Ann. Bot. 98, 257–265.
Merzlyak, M.N., Chivkunova, O.B., 2000. Light–stress-induced pigment
changes and evidence for anthocyanin photoprotection in apples.
J. Photochem. Photobiol. B: Biol. 55, 155–163.
Morales, F., Abadia, A., Abadia, J., Montserrat, G., Gil-Pelegrin, E.,
2002. Trichomes and photosynthetic pigment composition changes:
responses of Quercus ilex subsp. ballota (Desf.) Samp. and Quercus
coccifera L. to Mediterranean stress conditions. Trees 16, 504–510.
Murakami, Y., Tsuyama, M., Kobayashi, Y., Kodama, H., Iba, K., 2000.
Trienoic acids and plant tolerance of high temperature. Science 287,
476–479.
O’Brien, T.P., McCully, M.E., 1981. The Study of Plant Structure:
Principles and Selected Methods. Termarcarphi Pty Ltd., Melbourne,
290p.
Parry, D.W., Smithson, F., 1958. Techniques for studying opaline silica in
grasses. Ann. Bot. 22, 543–549.
Perez-Estrada, L., Cano-Santana, Z., Oyama, K., 2000. Variation in leaf
trichomes of Wigandi urens: environmental factors and physiological
consequences. Tree Physiol. 20, 629–632.
Radin, J.W., Lu, Z., Percy, R.G., Zeiger, E., 1994. Genetic variability for
stomatal conductance in Pima cotton and its relation to improvements
of heat adaptation. Proc. Natl. Acad. Sci. 91, 7217–7221.
Ristic, Z., Yang, G., Martin, B., Fullerton, S., 1998. Evidence of
association between specific heat-shock proteins and the drought and
heat tolerance phenotype in maize. J. Plant Physiol. 153, 497–505.
Riederer, M., Schreiber, L., 2001. Protecting against water loss: analysis of
the barrier properties of plant cuticles. J. Exp. Bot. 52, 2023–2032.
Spellenberg, R., 1975. Synthetic hybridization and taxonomy of western
north American Dichanthelium group lanuginosa (Poaceae). Madrono
23, 134–153.
Stebbins, G.L., 1957. Self fertilization and population variability in the
higher plants. Am. Midl. Nat. 32, 164–180.
Stout, R.G., Summers, M.L., Kerstetter, T., McDermott, T.R., 1997.
Heat- and acid-tolerance of a grass commonly found in geothermal
areas within Yellowstone National Park. Plant Sci. 130, 1–9.
Steyn, W.J., Wand, S.J.E., Holcroft, D.M., Jacobs, G., 2002. Anthocya-
nins in vegetative tissues: a proposed unified function in photoprotec-
tion. New Phytol. 155, 349–361.
Waters, E.R., Lee, G.L., Vierling, E., 1996. Evolution, structure and
function of small heat shock proteins in plants. J. Exp. Bot. 47,
325–338.
Weis, E., Berry, J.A., 1988. In: Long, P.S., Woodward, F.I. (Eds.), Plants
and Temperature. Company of Biologists, Cambridge, pp. 329–346.