molecular virology of hepatitis c virus - journal of general virology
TRANSCRIPT

Downloaded from www.microbiologyresearch.org by
IP: 144.76.74.74
On: Thu, 01 Feb 2018 15:12:24
Journal of General Virology (1997), 78, 2397–2410. Printed in Great Britain. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Molecular virology of hepatitis C virus
Berwyn Clarke
Hepatitis Antiviral Research, Virology Research Unit, GlaxoWellcome Medicines Research Centre, Gunnels Wood Road, Stevenage,Herts SG1 2NY, UK
IntroductionTowards the end of the 1980s a significant number of cases
of parenterally transmitted viral hepatitis could not be ascribedto any of the then known hepatic viruses (hepatitis A virus,hepatitis B virus and hepatitis delta virus). The discovery andcharacterization of a novel RNA virus with characteristicstypical of the family Flaviviridae (Choo et al., 1989) providedthe tools to study the epidemiological importance of this agentin these cases of non-A non-B hepatitis (NANBH). The virus,known as hepatitis C virus (HCV) was shown to be the causeof most cases of NANBH. Over 50% of patients becomechronic carriers (Kuo et al., 1989). The persistent infectioncommonly results in chronic active hepatitis which may lead toliver cirrhosis and hepatocellular carcinoma (Saito et al., 1990).Random screening of blood donor populations has indicatedthat worldwide there could be as many as 500 million chroniccarriers of the virus (Dhillon & Dusheiko, 1995) and thishighlights HCV as a major human pathogen.
Basic virologyCurrent approaches to the treatment of NANBH are limited
to the use of α-interferon (α-IFN) either alone (e.g. Davis et al.,1989) or in combination with other agents such as ribavirin(e.g. Brillanti et al., 1995), but the efficacy of these therapies isextremely poor and post-therapy relapse rate is very high suchthat sustained biochemical response is observed in less than20% of patients. Consequently there is an urgent need for thedevelopment of novel therapeutics. Unfortunately, approachesto the development of in vitro replication systems forpropagation of this virus have been particularly difficult withreliable systems for efficient long-term virus replication beingirreproducible. Nevertheless, reports of virus replication havebeen made using systems from hepatic tissue (Ito et al., 1996)and peripheral blood mononuclear cells (PBMC) (Willems et al.,1994 ; Zignego et al., 1992) while in vitro infection systemshave been reported from human T and B cell lines (Bertoliniet al., 1993 ; Shimizu et al., 1992), human foetal liver cells(Iacovacci et al., 1993) and chimpanzee hepatocytes (Lanfordet al., 1994). One of the most promising in vitro infection systemso far reported uses Daudi cells (a human B-cell line) (Nakajima
Author for correspondence: Berwyn Clarke.
Fax 44 1438 764279. e-mail BEC35328!ggr.co.uk
et al., 1996) where virus replication was reported to bedetectable up to a year post-infection and where there wasapparent selection of one particular viral sequence from thequasispecies present in the inoculum, indicative of selection fora cell-culture adapted virus. Very similar data were reported(Mizutani et al., 1995, 1996 ; Sugiyama et al., 1997) using ahuman T cell leukaemia virus type I-infected T cell line, MT-2.Again, in this study, there was apparent selection from the viralquasispecies of a single genomic sequence, presumably repre-senting a virus which was naturally adapted for growth in thiscell-line. Whether the viruses in these studies, which areapparently adapted for tissue culture growth, can replicate inother cell-lines remains to be established.
This system is one of the few in which the virion particlehas been visualized (Shimizu et al., 1996b). The size of thevirion had been previously estimated to be ! 80 nm byultracentrifugation (Bradley et al., 1985) while filtration studiessuggested 30–60 nm (He et al., 1987). Several studies haveshown that the buoyant density of HCV in infectious serum islower than that in non-infectious serum (Hijikata et al., 1993 c ;Bradley et al., 1991) and the density (! 1±06 g}ml) suggestedan association with the low density lipoprotein (LDL) fraction(Thomssen et al., 1992), which was subsequently verified byPrince et al. (1996). Typically, HCV-infected serum alsocontains a higher density fraction in which the virus iscomplexed with anti-viral antibodies (Hijikata et al., 1993 c). Aswould be expected, virus in the lipoprotein fraction appears tobe relatively more infectious than the antibody-coated fractionand has also been more amenable to electron-microscopicexamination. Using very low density lipoprotein (VLDL)-associated virus, highly purified particles of 60–70 nm havebeen observed which appear to have prominent (6–8 nm)spikes on their surface (Kaito et al., 1994 ; Prince et al., 1996),probably representing glycoprotein portions of the viralenvelope. The observation of an association between HCV andlipoproteins may be critical for the virus replication cyclebecause LDL and VLDL are taken up by hepatocytes throughthe LDL receptor, which may provide a unique and efficientmode of virus entry.
Molecular analysis of the HCV genomeThe genomic organization of HCV is illustrated schema-
tically in Fig. 1. Comparative analysis of the genomes ofseveral HCV strains (e.g. Kato et al., 1990 ; Choo et al., 1991 ;
0001-4901 # 1997 SGM CDJH

Downloaded from www.microbiologyresearch.org by
IP: 144.76.74.74
On: Thu, 01 Feb 2018 15:12:24
B. ClarkeB. Clarke
Fig. 1. Genomic organization of HCV RNA.
Okamoto et al., 1991 ; Takamizawa et al., 1991) indicated thevirus to be closely related to both the pestivirus and flavivirusgenera within the family Flaviviridae. The viral genome is asingle-stranded RNA molecule approximately 9±5 kb in lengthwhich is positive sense and possesses a unique open readingframe, coding for a single polyprotein, flanked by untranslatedregions at both its 5« and 3« ends. The length of thepolyprotein-encoding region varies according to the isolateand genotype of the virus from 3008 to 3037 amino acids (seeChamberlain et al., 1997). Apart from differences in lengthHCV genotypes show diversities of around 30% in thenucleotide sequences of their whole genomes and compre-hensive analysis of these sequences has revealed the existenceof at least 6 genotypes and more than 30 subtypes throughoutthe world (see Bukh et al., 1995 ; Simmonds, 1995). Theimportance of the genomic heterogeneity lies in the fact thatsome genotypes appear to be associated with more severepathology and are more refractory to treatment by currenttherapies (Bukh et al., 1995). However, other studies show nosignificant association between genotype and disease pro-gression (e.g. Benvegnu et al., 1997).
5« Untranslated region (5«UTR)
An obvious characteristic of HCV is the presence of a longuntranslated region (UTR) at the 5« end of the genome anddetailed molecular analysis indicates that polyprotein synthesisis initiated at nucleotide 342 (Han et al., 1991). This region ofthe genome was predicted to be capable of forming extensivesecondary structures (Tsukiyama-Kohara et al., 1992) whichwere biochemically confirmed by Brown et al. (1992). Fur-thermore, a survey of the sequences within the 5«UTR from 39different isolates and genotypes of HCV showed that therewas remarkable sequence conservation (Bukh et al., 1992). Infact, this region is the most conserved region of the wholegenome, a characteristic which has allowed it to be used as adiagnostic marker for HCV by PCR (Smith et al., 1995). Withinthis region the majority of genotypes of HCV possess fiveAUG codons which are not used for initiation of translation. Inthese respects, the presence of a long, highly ordereduntranslated region containing non-functional initiationcodons closely resembles the situation in the picornaviruses. Itis now well documented that picornaviral genomes aretranslated by a mechanism allowing ribosomes to bindinternally on the genome and initiate translation at specificAUG codons (Jackson et al., 1990). These internal ribosomal
entry sites (IRES) are dependent on the highly ordered structureof the 5«UTR regions and allow viral genome expression usinga cap-independent mechanism.
Because of the obvious similarities with the picornaviral5«UTR, several laboratories have looked for IRES elements inthe HCV genome. Initially, Tsukiyama-Kohara et al. (1992)demonstrated that, using in vitro expression systems, the HCV5«UTR could regulate translation initiation in a cap-inde-pendent manner, indicative of an internal initiation mechanism,and that it was possible to remove up to 101 nucleotides fromthe 5« end of the RNA without affecting the IRES function.Further studies from Wang et al. (1993) and Kettinen et al.(1994) confirmed the IRES activity but their mapping studiesindicated that only 40 nucleotides could be removed, whilesubsequent work by Fukushi et al. (1994) indicated that RNAstarting at nucleotide 62 was inactive as an IRES element.Analysis of the model of the structure of the HCV 5«UTRproposed by Brown et al. (1992) (see Fig. 2) indicates that astem–loop structure is formed at nucleotide 46 which sub-sequent deletion mutagenesis experiments have shown to becritical for IRES function (Fukushi et al., 1994 ; Rijnbrand et al.,1995 ; Honda et al., 1996b). Although an additional hairpinloop is predicted to be located nearer the 5« end of the RNA(see Fig. 2), this structure is not essential for IRES function andprobably fulfils a different replicative function.
In contrast to the general agreement on the presence of afunctional IRES in HCV, Han et al. (1991) reported the presenceof a subgenomic RNA initiating at nucleotide 145 which, giventhe highly ordered secondary structure of the full-lengthmolecule, might be necessary to allow conventional scanningtranslation. This observation has not been substantiated byother groups and, indeed, Honda et al. (1996b) clearlydemonstrated efficient IRES-directed translation from full-length genomic RNA using both in vitro systems andtransiently transfected cells.
Despite these reproducible reports of IRES activity in theHCV 5«UTR Yoo et al. (1992) were unable to show any IRESeffect using both in vitro and in vivo systems. The reasons forthis discrepancy were unclear until studies by Kettinen et al.(1994) and Reynolds et al. (1995) indicated that the IRES ofHCV was unique in that its 3« boundary did not lie in the5«UTR but was located within the actual coding sequence ofthe capsid protein. Detailed mapping studies (Reynolds et al.,1995) have subsequently shown that the optimal IRES activityresides in a segment from nucleotides 40 to 370, thus
CDJI

Downloaded from www.microbiologyresearch.org by
IP: 144.76.74.74
On: Thu, 01 Feb 2018 15:12:24
Review: Molecular virology of HCVReview: Molecular virology of HCV
Fig. 2. Model secondary and tertiary structure of the HCV 5«UTR fromHonda et al. (1996a).
incorporating at least 29 nucleotides from the capsid proteincoding region. Extension of the computer-generated model ofthe 5«UTR into the coding region indicates that this region ispredicted to form a stable hairpin structure as shown in Fig. 2.Since the studies of Yoo et al. (1992) did not maintain theintegrity of the 3« end of the UTR this may have beenresponsible for the lack of IRES activity reported. Furtherstudies by Honda et al. (1996a) have subsequently shown thatan in-frame 12 nucleotide insertion placed within the capsid-coding region 9 nucleotides downstream of the initiator AUGstrongly inhibited translation while multiple silent mutationswithin the first 42 nucleotides had the same effect. Howeverthe stem–loop structure in which the AUG is located(stem–loop IV, Fig. 2) is not essential for IRES activity but isable to regulate the efficiency of cap-independent translationboth in vitro and in vivo (Honda et al., 1996a).
Apart from the mapping studies delineating the 5« and 3«boundaries of the IRES, several groups have reported therequirement for specific structural regions within the elementitself. Fukushi et al. (1994) reported that the complete 5«UTRcontaining all stem–loop structures was necessary for full IRESfunction while Rijnbrand et al. (1995) demonstrated that onlythe most 5« located hairpin structure could be removed.Subsequent reports by Wang et al. (1994, 1995) and Le et al.(1995) indicated that, as well as simple hairpin–loop structures,
more complex tertiary structures such as pseudoknots werealso likely to be important.
The requirement for elements of the coding region to beinvolved in optimal IRES function has not been previouslyreported in any other RNA. Interestingly, Simmonds et al.(1993) observed that there appeared to be selective pressure onthe HCV genome to maintain particular sequences up to the50th codon of the core protein, including wobble positions.The fact that this area of the genome appears to be involved inhighly ordered secondary structure would seem to provide alogical explanation for this extreme sequence conservation.Further studies by Reynolds et al. (1995) also revealed otherunusual characteristics of the HCV IRES. In most studies onIRES function, mutation of the authentic initiator AUG residuehas a dramatic inhibitory effect on initiation of translation.Systematic mutational analysis of the AUG codons within theputative IRES element of HCV revealed that the authenticinitiator AUG at 342 could be mutated to AUU or CUG withvery little effect on translation efficiency. Furthermore, Rijn-brand et al. (1996) showed that insertion of additional AUGresidues within 8 nucleotides either side of the authentic AUGhad no effect on its correct recognition by ribosomes.
The function of the HCV IRES, therefore, seems to be toprovide a structurewhich is able to specifically direct ribosomesto the AUG codon at position 342 for translation initiation. Asin other IRES examples, this interaction probably involvescomplex interactions with cellular protein cofactors andevidence has recently been reported for such specific factorswhich may critical in regulating translation (Yen et al., 1995 ;Fukushi et al., 1997).
A consequence of the cap-independent translation providedby the IRES function is that some picornaviruses are able toshut-off conventional cap-dependent scanning translation byproteolysis of eukaryotic translation initiation factor eIF4-F(Lloyd et al., 1985) by viral protein 2A. Although HCV is notknown to have any obvious homologue to 2A, it is clear thatcap-independent translation from the HCV IRES can occur inthe presence of 2A from either Coxsackievirus (Tsukiyama-Kohara et al., 1992) or poliovirus (Rijnbrand et al., 1995). Theseobservations are further indications of functional IRES activityin HCV but the significance of the resistance to inhibition ofconventional translation mechanisms is, as yet, unclear.
Recent work has also confirmed the presence of a functionalIRES element in the 5«UTR of the pestiviruses (Poole et al.,1995) which has close structural similarity to the proposedstructure of the HCV IRES (Brown et al., 1992 ; Le et al., 1995),further demonstrating the relationship between these twogenera of the family Flaviviridae.
Virus-encoded proteinsThe nascent viral polyprotein is processed by a combination
of host and viral proteinases into the mature viral proteins (e.g.Hijikata et al., 1991, 1993b ; Grakoui et al., 1993 c) (Fig. 3) ; at
CDJJ

Downloaded from www.microbiologyresearch.org by
IP: 144.76.74.74
On: Thu, 01 Feb 2018 15:12:24
B. ClarkeB. Clarke
Fig. 3. Proteolytic processing of the HCV polyprotein.
least 10 distinct viral proteins have so far been identified whichare arranged as shown in Fig. 1, with the structural proteinslocated in the amino-terminal one-third and the replicativeenzymes located within the carboxy-terminal two-thirds of thepolyprotein.
Core
The protein located at the amino terminus of the poly-protein is highly basic in nature and is considered likely to bethe viral capsid protein. It is released from the viral polyproteinby nascent proteolytic cleavage at amino acid 191 by hostproteases (Hijikata et al., 1991 ; Ralston et al., 1993). The full-length protein, known as P21, has been identified by both invitro and in vivo expression (Hijikata et al., 1991 ; Lo et al., 1994 ;Harada et al., 1991) but a second species (P19) generated by asecondary cleavage at amino acid 173 is the major productobserved following expression in mammalian cells (Santoliniet al., 1994 ; Moradpour et al., 1996). Both P21 and P19 arelocated in the endoplasmic reticulum (ER) membrane and theconversion of P21 to P19 is presumably mediated bymembrane-associated cellular enzymes. A third collinearspecies of core which can also be detected in expression studies(Lo et al., 1995) is approximately 151 amino acids long (P16)and appears to be localized in the nucleus and more specificallyin the nucleolus. The relative expression of P16 varies betweendifferent strains of HCV and, perhaps surprisingly, is closelyassociated with the nature of the amino acid at position 9 of theprotein (Lo et al., 1994). The distinct difference in subcellularlocalization between P16 and P19}P21 suggests that thebiological role of P16 may be different. In particular, thenucleolar localization of P16 may be due to its ability to bindto ribosomes, which are assembled in the nucleus (Santoliniet al., 1994). Apart from its highly basic nature the core proteinalso possesses a number of distinct hydrophobic regions(residues 121–151, 170–191) which are involved in theassociation of P21 and P19 with the ER. It is likely that thecarboxy-terminal hydrophobic region (170–191) plays acritical role in the translocation of the viral structuralglycoproteins into the ER where the enzymes responsible forpeptide processing and glycosylation are located. Recentreports have indicated additional cleavage species at 179 and
182 which are not dependent on prior cleavage at 191 (Hussyet al., 1996). It is currently unclear what roles these diverse coreproteins play in virusmorphogenesis. Since virion nucleocapsidformation involves multimerization of core protein and itsinteraction with viral RNA, these characteristics of the HCVcore protein have been extensively studied and specificfunctions mapped to discrete portions of the molecule(Santolini et al., 1994 ; Matsumoto et al., 1996). The biologicalfunctions of the core found in the nucleus, if this also occurs innatural virus replication, are still unclear. Several studies havereported the suppression, by core, of transcription of severalhost genes as well as interference in expression of co-infectinggenomes of hepatitis B and human immunodeficiency viruses(Shih et al., 1993 ; Srinivas et al., 1996). Perhaps the mostinteresting recent observations have been that core canspecifically suppress apoptotic cell death in artificial systems(Ray et al., 1996) and also specifically interact with thecytoplasmic tail of the lymphotoxin-β (receptor (LTR), amember of the tumour necrosis factor family (Matsumoto et al.,1997). Since LTR is known to be involved in apoptoticsignalling this strongly suggests that core may have animmunomodulatory function and play a critical role in theestablishment of persistence and in disease pathogenesis.
Finally, a recent report (Barba et al., 1997) shows anassociation between the core protein and the surface of lipiddroplets within the cytoplasm. Analysis of the triglyceridepopulations within the cell indicates that core protein ex-pression stimulates a change in cellular metabolism of tri-glycerides. Since a characteristic of HCV infection is liversteatosis it is plausible that this occurs as a result of the directeffect of the core protein on lipid metabolism.
E1/E2
The major viral structural proteins are the glycoproteins E1and E2, which are released from the viral polyprotein by theaction of host-cell signal peptidases. Analysis of the aminotermini of both E1 (gp35) and E2 (gp70) indicates that they arecleaved at amino acids 383 and 746 respectively (Hijikata et al.,1991 ; Grakoui et al., 1993 c ; Mizushima et al., 1994). Bothproteins are heavily glycosylated with 5}6 and 11 N-linkedglycosylation sites respectively (Miyamura & Matsuura, 1993)and E2 is sometimes found extended at its carboxy terminus toinclude a smaller protein known as p7 such that multiple E2species can be observed after expression in eukaryotic systems(Lin et al., 1994a ; Mizushima et al., 1994 ; Selby et al., 1994 ;Lanford et al., 1993 ; Grakoui et al., 1993 c). Unlike the othercleavages within the structural region, proteolytic cleavagebetween E2}p7 and p7}NS2 appears to occur post-trans-lationally but the biological function of these diverse E2species is currently unknown.
Numerous studies have reported interactions between theviral structural and non-structural proteins and it is clear thatcomplex protein–protein association is a critical part of virus
CEAA

Downloaded from www.microbiologyresearch.org by
IP: 144.76.74.74
On: Thu, 01 Feb 2018 15:12:24
Review: Molecular virology of HCVReview: Molecular virology of HCV
replication. Although E1 has been shown to associate throughits carboxy terminus with the core protein (Lo et al., 1996) andE2 with NS2 (Matsuura et al., 1994 ; Selby et al., 1994), thefocus of most studies have been on the direct interactionbetween E1 and E2 themselves since this interaction is likely tobe a critical step in virus morphogenesis. Several studies haveshown convincing evidence that E1and E2 form complexes(Dubuisson et al., 1994 ; Grakoui et al., 1993 c ; Lanford et al.,1993) which are mainly non-covalently associated (Ralston etal., 1993) but in some lysates disulphide-linked complexes havealso been reported (Grakoui et al., 1993 c). Dubuisson et al.(1994) analysed these complexes and showed that the non-covalent forms were slow-forming stable species while thedisulphide-linked forms were rapidly forming misfolded aggre-gates. Nevertheless, disulphide-dependent folding of bothE1 and E2 appears to be a necessary prerequisite to complexformation and kinetic analysis indicates that the slow rate offolding of E1 may be the rate-limiting step in the process(Dubuisson et al., 1996). The efficient folding and assembly ofthe heterodimeric complexes has been shown to occur withinthe ER and is dependent on an initial prolonged associationwith the chaperone calnexin (Dubuisson et al., 1996). Thesecomplexes then slowly mature into non-calnexin boundcomplexes which may represent pre-budding native hetero-dimers (Deleersnyder et al., 1997).
Apart from the interest in understanding the biologicalprocesses of assembly and morphogenesis both E1 and E2 havebeen extensively studied in terms of antigenic variation andmechanisms of persistence, and are obviously major com-ponents of prototype vaccine studies for HCV. E2 representsthe most variable region of the HCV genome (Kato et al., 1992 ;Weiner et al., 1991) and the variation is assumed to be causedby random mutation and selection of mutants capable ofescaping from neutralizing antibodies produced in the host.Furthermore, antibodies against E2 correlate with protectionfrom HCV challenge in chimpanzees and assays are nowavailable to directly assay for neutralizing antibodies usingcell-binding of structural proteins (Rosa et al., 1996) or in vitroculture (Shimizu et al., 1996 c). Within the E2 sequence areregions of extreme hypervariability (HVR) which have beenthe focus of more detailed study and one of these regions knowas HVR-1 represents the amino-terminal 34 amino acids withinE2, spanning residues 383–414 (e.g. Ogata et al., 1991 ; Weineret al., 1991 ; Kumar et al., 1994). This region has been suggestedto be particularly important in HCV neutralization because ofits extreme variability and the fact that this variability was notobserved in a patient with agammaglobulinaemia even over aperiod of 2±5 years (Kumar et al., 1993). Antibodies to HVR-1in patient sera have been shown to neutralize binding of E2(Zibert et al., 1995) while peptide antibodies raised to thisregion are able to block infection in tissue culture (Shimizu etal., 1996 c) and have shown efficacy in chimpanzee challengeexperiments (Farci et al., 1996). Despite this, the extremeheterogeneity in the sequence of this epitope in diverse virus
strains means that its utility in a prophylactic vaccine strategyremains doubtful.
NS2
The NS2 protein has been shown to be a transmembraneprotein with its carboxy terminus translocated into the lumenof the ER while its amino terminus lies in the cytosol (Santoliniet al., 1995). Although immunoprecipitation studies (Grakouiet al., 1993b ; Matsuura et al., 1994) have shown that NS2 isclosely associated with the structural proteins the biologicalfunction of the majority of the NS2 protein is still unclear.However, detailed study of the proteolytic processing of thisregion of the polyprotein has revealed an unexpected pro-teolytic function contributed partially by the carboxy terminusof NS2.
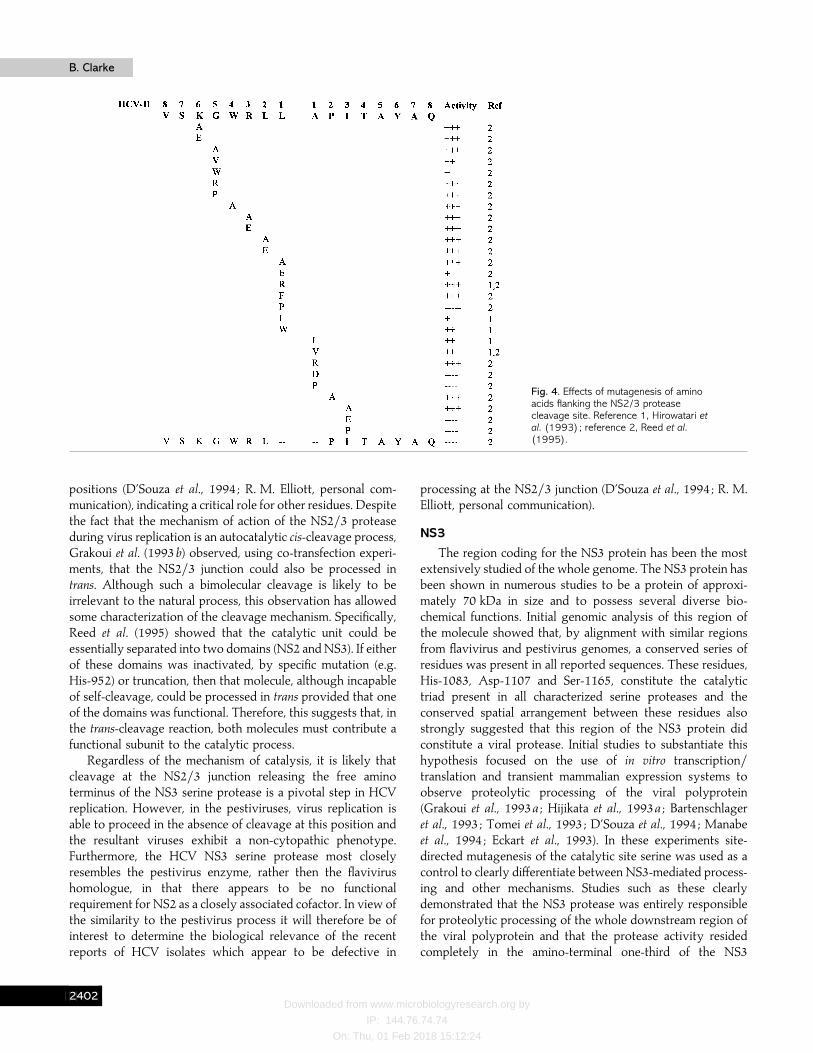
By analogy with the genus Flavivirus it would be assumedthat the NS3 serine protease was responsible for cleavage atthe junction between NS2 and NS3 in HCV. Surprisingly,mutation of the active site Ser-1165 in NS3, althoughabolishing processing at downstream sites, did not affectproteolytic cleavage at the NS2}NS3 junction. The first reportof the existence of a second virus-encoded protease was madeby Hijikata et al. (1993a) who showed that the proteaseresponsible for cleavage at this position comprised elements ofboth the NS2 and NS3 proteins. Further mapping studies(Hijikata et al., 1993a ; Grakoui et al., 1993b) delineated theproteolytic domain responsible for this activity to amino acids827–1207. This region encompasses the carboxy terminus ofNS2 and the amino terminus of NS3 and, therefore, overlapsthe active domain of the serine protease. Apart from internalautocatalytic cleavage of itself, no other proteolytic processingfunctions have been ascribed to this enzyme. It has no knownhomology to other classes of proteases but has been proposedto be a metalloprotease since its activity can be inhibited byEDTA and stimulated by zinc (Zn#+) ions. Mutational analysisof the region surrounding the cleavage site (Hirowatari et al.,1993 ; Reed et al., 1995) (see Fig. 4) has shown that the NS2}3protease is remarkably tolerant to amino acid mutations in theregion from P5 to P3«, apart from substitutions such as prolinewhich markedly affect the conformation of the substrate.However, Reed et al. (1995) noted a preference for hydrophobicamino acids at the P1 and P1« positions similar to that shownby neutral metalloproteases. In particular, the preferencesmarkedly resembled those of endopeptidase 24.15, a mam-malian enzyme involved in processing of peptide hormones(Erdos & Skidgel, 1989). Despite this similarity there is,currently, no solid evidence to suggest that theNS2}3 proteaseis a metalloprotease.
Several groups have attempted to identify those regions ofthe enzyme involved in catalytic activity and mutation of His-952 and Cys-993 in NS2 has shown them to be involved.Nevertheless, other reports have also shown lack of NS2}3protease activity even with wild-type residues at these
CEAB
Downloaded from www.microbiologyresearch.org by
IP: 144.76.74.74
On: Thu, 01 Feb 2018 15:12:24
B. ClarkeB. Clarke
Fig. 4. Effects of mutagenesis of aminoacids flanking the NS2/3 proteasecleavage site. Reference 1, Hirowatari etal. (1993); reference 2, Reed et al.(1995).
positions (D’Souza et al., 1994 ; R. M. Elliott, personal com-munication), indicating a critical role for other residues. Despitethe fact that the mechanism of action of the NS2}3 proteaseduring virus replication is an autocatalytic cis-cleavage process,Grakoui et al. (1993b) observed, using co-transfection experi-ments, that the NS2}3 junction could also be processed intrans. Although such a bimolecular cleavage is likely to beirrelevant to the natural process, this observation has allowedsome characterization of the cleavage mechanism. Specifically,Reed et al. (1995) showed that the catalytic unit could beessentially separated into two domains (NS2 and NS3). If eitherof these domains was inactivated, by specific mutation (e.g.His-952) or truncation, then that molecule, although incapableof self-cleavage, could be processed in trans provided that oneof the domains was functional. Therefore, this suggests that, inthe trans-cleavage reaction, both molecules must contribute afunctional subunit to the catalytic process.
Regardless of the mechanism of catalysis, it is likely thatcleavage at the NS2}3 junction releasing the free aminoterminus of the NS3 serine protease is a pivotal step in HCVreplication. However, in the pestiviruses, virus replication isable to proceed in the absence of cleavage at this position andthe resultant viruses exhibit a non-cytopathic phenotype.Furthermore, the HCV NS3 serine protease most closelyresembles the pestivirus enzyme, rather then the flavivirushomologue, in that there appears to be no functionalrequirement for NS2 as a closely associated cofactor. In view ofthe similarity to the pestivirus process it will therefore be ofinterest to determine the biological relevance of the recentreports of HCV isolates which appear to be defective in
processing at the NS2}3 junction (D’Souza et al., 1994 ; R. M.Elliott, personal communication).
NS3
The region coding for the NS3 protein has been the mostextensively studied of the whole genome. The NS3 protein hasbeen shown in numerous studies to be a protein of approxi-mately 70 kDa in size and to possess several diverse bio-chemical functions. Initial genomic analysis of this region ofthe molecule showed that, by alignment with similar regionsfrom flavivirus and pestivirus genomes, a conserved series ofresidues was present in all reported sequences. These residues,His-1083, Asp-1107 and Ser-1165, constitute the catalytictriad present in all characterized serine proteases and theconserved spatial arrangement between these residues alsostrongly suggested that this region of the NS3 protein didconstitute a viral protease. Initial studies to substantiate thishypothesis focused on the use of in vitro transcription}translation and transient mammalian expression systems toobserve proteolytic processing of the viral polyprotein(Grakoui et al., 1993a ; Hijikata et al., 1993a ; Bartenschlageret al., 1993 ; Tomei et al., 1993 ; D’Souza et al., 1994 ; Manabeet al., 1994 ; Eckart et al., 1993). In these experiments site-directed mutagenesis of the catalytic site serine was used as acontrol to clearly differentiate between NS3-mediated process-ing and other mechanisms. Studies such as these clearlydemonstrated that the NS3 protease was entirely responsiblefor proteolytic processing of the whole downstream region ofthe viral polyprotein and that the protease activity residedcompletely in the amino-terminal one-third of the NS3
CEAC

Downloaded from www.microbiologyresearch.org by
IP: 144.76.74.74
On: Thu, 01 Feb 2018 15:12:24
Review: Molecular virology of HCVReview: Molecular virology of HCV
Fig. 5. Cleavage site specificities of HCV NS3 protease.
molecule. The overall proteolytic pathway for the whole viralgenome is illustrated in Fig. 3 and shows that the NS3 proteasemediates proteolysis at the NS3}NS4A, NS4A}NS4B,NS4B}NS5A and NS5A}NS5B junctions to release the matureNS3, NS4A, NS4B, NS5A and NS5B proteins. However, thereare subtle biochemical differences in the exact mechanisms bywhich NS3 mediates these cleavages. Firstly, it was observedthat cleavage of NS3 from NS4A was a spontaneous rapidautocatalytic event which, under normal conditions, could onlybe mediated in cis whereas all other cleavages could be carriedout in trans if exogenous NS3 was added to substratescontaining target cleavage sites (D’Souza et al., 1995 ; Tomei etal., 1993 ; Bartenschlager et al., 1994 ; Lin et al, 1994b).Furthermore, efficient cleavage at the trans sites was dependenton the presence of the NS4A protein itself as a cofactor for NS3(Bartenschlager et al., 1994 ; Failla et al., 1994 ; Lin et al., 1994b).However, the relative dependence on the presence of NS4Aseemed to vary according to the particular cleavage site suchthat cleavage at NS5A}5B was relatively unaffected by thepresence of NS4A whereas cleavage at NS4B}5A wasabsolutely dependent on its presence. These observationssuggested a close association between NS3 and NS4A at themolecular level and several studies confirmed the formation ofa stable complex between the two species (Bartenschlager et al.,1995 ; Failla et al., 1995 ; Lin et al., 1995 ; Tanji et al., 1995a).Detailed biochemical and mutagenesis experiments indicatedthat the point of association between NS3 and NS4A lay in theamino-terminal region of the NS3 molecule between residues15 and 22 (Satoh et al., 1995 ; Koch et al., 1996) and involvedthe central region of NS4A in which residues 22 to 31 werecritical (Bartenschlager et al., 1995 ; Lin et al., 1995 ; Failla et al.,1995 ; Shimizu et al., 1996a ; Butkiewicz et al., 1996). The recentelucidation of the crystal structure of the NS3}4A complex hasconfirmed these observations indicating that NS4A forms animportant structural feature of the active complex (Kim et al.,1996). A further unexpected feature of the crystal structure was
the fact that a zinc atom appears to have a critical structural role(Kim et al., 1996 ; Love et al., 1996). This atom is closely bondedto four protease residues at Cys-97, Cys-99, Cys-145 and His-149 (via a water molecule). Although not located close to theactive site of the enzyme, mutation of these residues has adramatic effect on the proteolytic activity (Stempniak et al.,1997). Most of the original studies assessing the proteolyticactivity of the NS3 protease used large precursors mimickingthe natural substrates and amino-terminal analysis of theprocessed products revealed the specificity of the enzyme. Ascan be seen from Fig. 5, comparative analysis of the sequencesurrounding the cleavage sites shows functionally importantconservation at residues P1}P1« and an acidic preference at P6with a significant degree of variation at the other residuesparticularly on the carboxy side of the scissile bond. Inter-estingly, there is a significant difference between the P1 residueat the cis site (NS3}4A) which is always threonine and the transsites which is always cysteine. Subsequent mutagenesisexperiments using either natural substrates or syntheticpeptides have allowed detailed analysis of the ability of theenzyme to cleave substrates with amino acid changes atspecific positions around the cleavage sites and these largelysupport the observation that residues P1}P1« and P6 arecritical for efficient proteolysis (Kolykhalov et al., 1994 ;Komoda et al., 1994 ; Steinkuhler et al., 1996).
Analysis of the remainder of the sequence of the NS3protein also revealed the presence of other motifs characteristicof NTPase and RNA helicase enzymatic functions. Expressionof this portion of the NS3 protein in bacteria has alloweddetailed evaluation of the presence of these activities. Initialstudies (Suzich et al., 1993 ; D’Souza et al., 1995) confirmed thepresence of the NTPase activity on the carboxy-terminaldomain of the full-length NS3 protein and compared theactivity with the related pestivirus and flavivirus enzymes.Subsequent studies (Kim et al., 1995 ; Gwack et al., 1996)confirmed the presence of the RNA helicase activity and alsoreported detailed evaluation of the biochemical specificity andkinetic characteristics of the enzyme (Preugschat et al., 1996).The purified enzyme was shown to actively bind RNAsubstrates with a minimal RNA binding size of between 7 and20 nucleotides and to unwind both RNA}RNA, RNA}DNAand DNA}DNA heteroduplexes. Further analysis (Preugschatet al., 1996 ; Tai et al., 1996) indicated that the enzyme had apreference for duplex molecules containing single-stranded 3«regions implying a 3« to 5« directionality and also that theenzyme bound preferentially to the poly(rU) sequence near the3« end of the viral genome (Kanai et al., 1995 ; Tanaka et al.,1995). Taken together, all these observations suggest that theHCV NS3 RNA helicase is unique among RNA helicasescharacterized so far including the related pestivirus enzyme.Further insight into the role that the RNA helicase plays invirus replication has recently been derived through thecompletion of the first crystal structure (Yao et al., 1997). Thestructure, refined to 2±1 AI , reveals that the molecule has
CEAD

Downloaded from www.microbiologyresearch.org by
IP: 144.76.74.74
On: Thu, 01 Feb 2018 15:12:24
B. ClarkeB. Clarke
distinct NTPase and RNA binding domains and supports amechanism of helicase activity involving initial recognition ofa 3« single-stranded region on the nucleic acid substrate by aconserved arginine-rich sequence on the RNA-binding domain.The RNA binding domain then appears to undergo a rotationalevent involved in physically unwinding the helicase and it islikely that this rotation is coupled to NTP hydrolysis throughthe other domain.
Despite the fact that there are three known catalyticactivities in the NS3 protein there is no evidence to suggestthat the two domains are separated by proteolysis in vivo. Thiscould mean that there is a functional interdependence betweenthe helicase and protease functions. Recent observations fromMorgenstern et al. (1997) have shown evidence to support thisin that all three activities of NS3 can be modulated by theaddition of polynucleotides to the assay systems. Since theseenzymes are likely to be components of a complex replicationstructure it is reasonable to assume that the interplay betweenthe various domains of NS3, as well as other viral proteins,may be critical in regulating virus replication.
Apart from the reported biochemical functions of the NS3protein there are also a number of intriguing reports on thepossible role of NS3 in disease. In one study (Sakamuro et al.,1995) expression of the NS3 protein in NIH 3T3 cells resultedin cellular transformation and, on subsequent inoculation intonude mice, tumorigenesis. This may be related to a number ofother reports (Borowski et al., 1996, 1997) that the NS3 proteinis able to specifically interact with the catalytic subunit ofprotein kinase A. This molecule is involved in intracellularsignal transduction processes and so interference by NS3 inthese pathways would be expected to have a dramatic effect onnormal cellular functions and would be closely associated withpathogenic mechanisms.
NS4
The NS4 region of the polyprotein comprises two proteins,namely NS4A and NS4B. Both of these are released from theviral polyprotein by the NS3 serine protease by cis cleavage atthe NS3}NS4A and trans cleavage at the NS4A}NS4B andNS4B}NS5A junctions. NS4A is a small protein, approxi-mately 8 kDa in size, and appears to have diverse functionssuch as anchorage of replication complexes and as a cofactorfor the NS3 protease (see previous). Currently, there is noascribed function for the NS4B protein but it is likely that itplays an integral role within HCV replication complexes.
NS5A and NS5B
The NS5 region of the polyprotein is composed of twomajor proteins, NS5A (p56) and NS5B (p65), which arereleased as mature products by the action of the NS3 proteasein conjunction with NS4A. Early studies suggested that aprecursor of NS5A existed (p58) from which a small third
protein was released at the amino terminus resulting in matureNS5A (p56) and a small protein fragment NS5C (p2) (Hijikataet al., 1993b). Subsequent work indicated that this was incorrectand that p56 and p58 were both full-length NS5A and thatboth were phosphoproteins with a degree of hyperphos-phorylation reflecting the difference in size between p58 andp56 (Kaneko et al., 1994). Both forms of NS5A are phos-phorylated at serine residues and phosphorylation occurs afterthe mature NS5A protein is released from the polyprotein.Basal phosphorylation has been shown to occur in two regions(2200–2250 and the carboxy terminus of NS5A) and isindependent of the presence of any other viral proteins (Tanjiet al., 1995b). In contrast, hyperphosphorylation is extremelydependent on the presence of serine residues 2197, 2201 and2204 and is enhanced by the presence of NS4A as a cofactor(Kaneko et al., 1994), again indicating the multifunctionalnature of the NS4A protein (Tanji et al., 1995a). Furtheranalysis has also shown that the NS4A-dependent phos-phorylation is dependent on association between NS4A andamino acids 2135–2139 from NS5A (Asabe et al., 1997).Sequence comparison of the regions surrounding the sites ofphosphorylation indicates an extremely high level of con-servation between different strains of the virus but thebiological significance of the phosphorylation is still unclear.However, the sites do not conform to recognized consensusmotifs for known serine kinases. This indicates that either anunknown serine kinase is involved or that the protein has auto-catalytic kinase activity. The functional role of either form ofNS5A is currently unclear although both forms possess nuclearlocalization signals and are detected in the nuclear periplasmicmembrane by immunofluoresence. Similarly, recent studieshave reported a similar localization of the NS5B protein,suggesting that NS5A and NS5B may be closely associatedcomponents of a membrane-bound replication complex(Hwang et al., 1997). Apart from the probable role of NS5A inthe replication cycle, recent evidence has suggested that it maybe a critical factor in determining the susceptibility of the virusto treatment with IFN. It was initially reported that IFNsensitivity correlated with mutations within a discrete regionof NS5A which was subsequently named the IFN sensitivitydetermining region (ISDR) (Enomoto et al., 1995, 1996).Subsequent analysis indicated that the likely mechanism bywhich this occurred was through a direct interaction of NS5Awith the IFN-induced protein kinase, PKR, a mediator of IFN-induced antiviral resistance (Gale et al., 1997). Since PKR is acritical factor in the response to IFN (Katze, 1995) itsinactivation by NS5A may be a major mechanism by whichHCV evades the host immune response.
The sequence of the NS5B protein is highly conserved, notonly between different strains of HCV but also in pestiviruses,flaviviruses and even in other RNA viruses. In particular, theamino acid motif G-D-D is totally conserved in HCV,flaviviruses, poliovirus and tobacco mosaic virus (Kamer &Argos, 1984). This motif is a characteristic of all known RNA-
CEAE

Downloaded from www.microbiologyresearch.org by
IP: 144.76.74.74
On: Thu, 01 Feb 2018 15:12:24
Review: Molecular virology of HCVReview: Molecular virology of HCV
dependent RNA polymerases and so the function of NS5B inHCV has been speculated to be the viral polymerase. Insupport of this Behrens et al. (1995) recently presented evidenceof RNA-dependent RNA synthesis in extracts of baculovirus-infected insect cells expressing HCV proteins. As yet it isunclear whether NS5B alone can perform this role or whetherit is merely a critical component of a multimolecular replicationcomplex.
3« Untranslated region (3«UTR)
A significant recent finding in the HCV area has been therevelation that the considered 3« terminal region of thegenome was incorrect. A number of studies had suggested thatthe 3« terminus of the genome terminated in a poly(U) tract(e.g. Kato et al., 1990 ; Takamizawa et al., 1991 ; Okamoto et al.,1992) or, in a single report (Han et al., 1991), as a poly(A) tract.However, more detailed analysis of the 5« end of the negativeanti-genomic RNA strand in infected liver cells revealed thepresence of a novel 98 nucleotide sequence downstream of thepresumed genomic terminus (Tanaka et al., 1995 ; Kolykhalovet al., 1996). Detailed sequence analysis of the complete new3«UTR shows that it can be considered as a tripartite structurecomprising the conventional 3« end, a poly(U) tract and thenew highly conserved sequence known as the 3«X tail.Interestingly, the poly(U) region appears to be extremelyheterogeneous between different virus isolates and even withinthe same infected liver. In contrast, the new sequence has beenshown to be extremely highly conserved even between thetwo most genetically divergent HCV genotypes, 1B and 2A(Tanaka et al., 1996), although Yamada et al. (1996) haverecently reported that some genotype 1B isolates possess anadditional two uridine residues at the extreme 3« terminus.Furthermore, computer modelling has predicted that thissequence can fold into an elaborate stem–loop structure(Kolykhalov et al., 1996) suggestive of a critical functionalcomponent in virus replication as shown for many positive-strand viruses. Surprisingly, the only reported infectious clonefor HCV (Yoo et al., 1995) represents a virus which lacks thisnew region. The levels of genomic replication in this reportwere not high, however, and ligation of this new 3« end to theclone may significantly improve replicative efficiency.
ConclusionThe identification of HCV as the aetiological agent of
NANB hepatitis is certainly the most significant recentdevelopment in any area of viral disease. The clinicalimportance of the disease and the need to rapidly identify newtherapeutic approaches have resulted in intensive study of themolecular properties of the virus. Although there are still largegaps in our understanding of some aspects of the virus and its
replication this review has attempted to provide a snapshot ofthe current state of the art.
I would like to thank my colleagues at GlaxoWellcome, both past andpresent, for valuable contributions to this review, particularly TonyCarroll and Eddy Littler for critically reading the manuscript. Similarly, toRichard Elliott as well as providing access to unpublished information.
ReferencesAsabe, S. I., Tanji, Y., Satoh, S., Kaneko, T., Kimura, K. & Shimotohno,K. (1997). The N-terminal region of hepatitis C virus-encoded NS5A isimportant for NS4A-dependent phosphorylation. Journal of Virology 71,790–796.
Barba, G., Harper, F., Harada, T., Kohara, M., Goulinet, S., Matsuura,Y., Eder, G., Schaff, Z., Chapman, M. J., Miyamura, T. & Brechot, C.(1997). Hepatitis C virus core protein shows a cytoplasmic localizationand associates to cellular lipid storage droplets. Proceedings of the NationalAcademy of Sciences, USA 94, 1200–1205.
Bartenschlager, R., Ahlborn-Laake, L., Mous, J. & Jacobsen, H.(1993). Nonstructural protein 3 of the hepatitis C virus encodes a serine-type proteinase required for cleavage at the NS3}4 and NS4}5 junctions.Journal of Virology 67, 3835–3844.
Bartenschlager, R., Ahlborn-Laake, L., Mous, J. & Jacobsen, H.(1994). Kinetic and structural analyses of hepatitis C virus polyproteinprocessing. Journal of Virology 68, 5045–5055.
Bartenschlager, R., Lohmann, V., Wilkinson, T. & Koch, J. O. (1995).Complex formation between the NS3 serine-type proteinase of thehepatitis C virus and NS4A and its importance for polyproteinmaturation. Journal of Virology 69, 7519–7528.
Behrens, S. E., Tomei, L. & De Francesco, R. (1996). Identification andproperties of the RNA-dependent RNA polymerase of hepatitis C virus.EMBO Journal 15, 12–22.
Benvegnu, L., Pontisso, P., Cavalletto, D., Noventa, F., Chemello, L. &Alberti, A. (1997). Lack of correlation between hepatitis C virusgenotypes and clinical course of hepatitis C virus-related cirrhosis.Hepatology 25, 211–215.
Bertolini, L., Iacovacci, S., Ponzetto, A., Gorini, G., Battaglia, M. &Carloni, G. (1993). The human bone-marrow-derived B cell line CE,susceptible to hepatitis C virus infection. Research in Virology 144,281–285.
Borowski, P., Heiland, M., Oehlmann, K., Becker, B., Kornetsky, H.,Feucht, H. & Laufs, R. (1996). Non-structural protein 3 of hepatitis Cvirus inhibits phosphorylation mediated by cAMP-dependent proteinkinase. European Journal of Biochemistry 237, 611–618.
Borowski, P., Oehlmann, K., Heiland, M. & Laufs, R. (1997).Nonstructural protein 3 of hepatitis C virus blocks the distribution of thefree catalytic subunit of cyclic AMP-dependent protein kinase. Journal ofVirology 71, 2838–2843.
Bradley, D. W., McCaustland, K. A., Cook, E. H., Schable, C. A., Ebert,J. W. & Maynard, J. E. (1985). Post-transfusion non-A, non-B hepatitisin chimpanzees. Physicochemical evidence that a tubule-forming agent isa small, enveloped virus. Gastroenterology 85, 773–779.
Bradley, D. W., McCaustland, K. A., Krawczynski, K., Spelbring, J.,Humphrey, C. & Cook, E. H. (1991). Hepatitis C virus : buoyant densityof the Factor VIII-derived isolate in sucrose. Journal of Medical Virology34, 206–208.
Brillanti, S., Miglioli, M. & Barbara, L. (1995). Combination antiviraltherapy with ribavirin and interferon alpha in interferon alpha relapsersand non-responders : Italian experience. Journal of Hepatology 23(Suppl. 2),17–21.
CEAF

Downloaded from www.microbiologyresearch.org by
IP: 144.76.74.74
On: Thu, 01 Feb 2018 15:12:24
B. ClarkeB. Clarke
Brown, E. A., Zhang, H., Ping, L.-H. & Lemon, S. M. (1992). Secondarystructure of the 5« nontranslated regions of the hepatitis C virus andpestivirus genomic RNAs. Nucleic Acids Research 20, 5041–5045.
Bukh, J., Purcell, R. H. & Miller, R. H. (1992). Sequence analysis of the5« non-coding region of hepatitis C virus. Proceedings of the NationalAcademy of Sciences, USA 89, 4942–4946.
Bukh, J., Miller, R. H. & Purcell, R. H. (1995). Genetic heterogeneity ofhepatitis C virus quasispecies and genotypes (Review). Seminars in LiverDisease 15, 41–63.
Butkiewicz, N. J., Wendel, M., Zhang, R. M., Jubin, R., Pichardo, J.,Smith, E. B., Hart, A. M., Ingram, R., Durkin, J., Mui, P. W., Murray,R. G., Ramanathan, L. & Dasmahapatra, B. (1996). Enhancement ofhepatitis C virus NS3 proteinase activity by association with NS4A-specific synthetic peptides – identification of sequence and criticalresidues of NS4A for the cofactor activity. Virology 225, 328–338.
Chamberlain, R. W., Adams, N., Saeed, A. A., Simmonds, P. & Elliott, R.M. (1997). Complete nucleotide sequence of a type 4 hepatitis C virusvariant, the predominant genotype in the Middle East. Journal of GeneralVirology 78, 1341–1347.
Choo, Q.-L., Kuo, G., Weiner, A. J., Overby, L. R., Bradley, D. W. &Houghton, M. (1989). Isolation of a cDNA clone derived from a blood-borne non-A, non-B viral hepatitis genome. Science 244, 359–362.
Choo, Q.-L., Richman, K. H., Han, J. H., Berger, K., Lee, C., Dong, C.,Gallegos, C., Coit, D., Medina-Selby, A., Barr, P. J., Weiner, A. J.,Bradley, D. W., Kuo, G. & Houghton, M. (1991). Genetic organizationand diversity of the hepatitis C virus. Proceedings of the National Academyof Sciences, USA 88, 2451–2455.
Davis, G. L., Balart, L. A., Schiff, E. R., Lindsay, K., Bodenheimer, H. C.,Perrillo, R. P., Carey, W., Jacobson, I. M., Payne, J., Dienstag, J. L., VanThiel, D. H., Tamburra, C., Lefkowitch, J., Albrecht, J., Meschievitz, C.,Ortego, T. J. & Gibas, A. (1990). Treatment of chronic hepatitis C withrecombinant interferon alpha : a randomized double-blind, placebo-controlled trial. New England Journal of Medicine 321, 1501–1506.
Deleersnyder, V., Pillez, A., Wychowski, C., Blight, K., Xu, J., Hahn,Y. S., Rice, C. M. & Dubuisson, J. (1997). Formation of native hepatitis Cvirus glycoprotein complexes. Journal of Virology 71, 697–704.
Dhillon, A. P. & Dusheiko, G. M. (1995). Pathology of hepatitis C virusinfection. Histopathology 26, 297–309.
D’Souza, E. D. A., O’Sullivan, E., Amphlett, E. M., Rowlands, D. J.,Sangar, D. V. & Clarke, B. E. (1994). Analysis of NS3-mediatedprocessing of the hepatitis C virus non-structural region in vitro. Journalof General Virology 75, 3469–3476.
D’Souza, E. D. A., Grace, K., Sangar, D. V., Rowlands, D. J. & Clarke,B. E. (1995). In vitro cleavage of hepatitis C virus polyprotein substratesby purified recombinant NS3 protease. Journal of General Virology 76,1729–1736.
Dubuisson, J. & Rice, C. M. (1996). Hepatitis C virus glycoproteinfolding : disulphide bond formation and association with calnexin. Journalof Virology 70, 778–786.
Dubuisson, J., Hsu, H. H., Cheung, R. C., Greenberg, H. B., Russell,D. G. & Rice, C. M. (1994). Formation and intracellular localization ofhepatitis C virus envelope glycoprotein complexes expressed byrecombinant vaccinia and Sindbis viruses. Journal of Virology 68,6147–6160.
Eckart, M. R., Selby, M., Masiarz, F., Lee, C., Berger, K., Crawford, K.,Kuo, G., Houghton, M. & Choo, Q.-L. (1993). The hepatitis C virusencodes a serine protease involved in processing of the putative non-structural proteins from the viral polyprotein precursor. Biochemical andBiophysical Research Communications 192, 399–406.
Enomoto, N., Sakuma, I., Asahina, Y., Kurosaki, M., Murakami, T.,
Yamamoto, C., Izumi, N., Marumo, F. & Sato, C. (1995). Comparison offull length sequences of interferon-sensitive and resistant hepatitis C virus1b : sensitivity to interferon is conferred by amino acid substitutions inthe NS5A region. Journal of Clinical Investigation 96, 224–230.
Enomoto, N., Sakuma, I., Asahina, Y., Kurosaki, M., Murakami, T.,Yamamoto, C., Ogura, Y., Izumi, N., Marumo, F. & Sato, C. (1996).Mutations in the nonstructural protein NS5A gene and response tointerferon in patients with chronic hepatitis C virus 1b infection. NewEngland Journal of Medicine 334, 77–81.
Erdos, E. G. & Skidgel, R. A. (1989). Neutral endopeptidase 24.11(enkephalinase) and related regulators of peptide hormones. FASEBJournal 3, 145–151.
Failla, C., Tomei, L. & De Francesco, R. (1994). Both NS3 and NS4Aare required for proteolytic processing of hepatitis C virus nonstructuralproteins. Journal of Virology 68, 3753–3760.
Failla, C., Tomei, L. & De Francesco, R. (1995). An amino-terminaldomain of the hepatitis C virus NS3 protease is essential for interactionwith NS4A. Journal of Virology 69, 1769–1777.
Farci, P., Shimoda, A., Wong, D., Cabezon, T., De Gioannis, D.,Strazzera, A., Shimizu, Y., Shapiro, M., Alter, H. J. & Purcell, R. H.(1996). Prevention of hepatitis C virus infection in chimpanzees byhyperimmune serum against the hypervariable region 1 of the envelope2 protein. Proceedings of the National Academy of Sciences, USA 93,15394–15399.
Fukushi, S., Katayama, K., Kurihara, C., Ishiyama, N., Hoshino, F. B.,Ando, T. & Oya, A. (1994). Complete 5« noncoding region is necessaryfor the efficient internal initiation of hepatitis C virus RNA. Biochemicaland Biophysical Research Communications 199, 425–432.
Fukushi, S., Kurihara, C., Ishiyama, N., Hoshino, F. B., Oya, A. &Katayama, K. (1997). The sequence element of the internal ribosomeentry site and a 25-kilodalton cellular protein contribute to efficientinternal initiation of translation of hepatitis C virus RNA. Journal ofVirology 71, 1662–1666.
Gale, M. J., Korth, M. J., Tang, N. M., Tan, S.-L., Hopkins, D. A., Dever,T. E., Polyak, S. J., Gretch, D. R. & Katze, M. G. (1997). Evidence thathepatitis C virus resistance to interferon is mediated through repressionof the PKR protein kinase by the nonstructural 5A protein. Virology 230,217–227.
Grakoui, A., McCourt, D. W., Wychowski, C., Feinstone, S. M. & Rice,C. M. (1993a). Characterization of the hepatitis C virus-encoded serineproteinase : determination of proteinase-dependent polyprotein cleavagesites. Journal of Virology 67, 2832–2343.
Grakoui, A., McCourt, D. W., Wychowski, C., Feinstone, S. M. & Rice,C. M. (1993b). A second hepatitis C virus-encoded protease. Proceedingsof the National Academy of Sciences, USA 90, 10583–10587.
Grakoui, A., Wychowski, C., Lin, C., Feinstone, S. M. & Rice, C. M.(1993c). Expression and identification of hepatitis C virus polyproteincleavage products. Journal of Virology 67, 1385–1395.
Gwack, Y., Kim, D. W., Han, J. H. & Choe, J. (1996). The hepatitis Cvirus NS3 protein. Biochemical and Biophysical Research Communications225, 654–659.
Han, J. H., Shyamala, V., Richman, K. H., Brauer, M. J., Irvine, B.,Urdea, M. S., Tekamp Olson, P., Kuo, G., Choo, Q. L. & Houghton, M.(1991). Characterization of the terminal regions of hepatitis C viralRNA: identification of conserved sequences in the 5« untranslated regionand poly(A) tails at the 3« end. Proceedings of the National Academy ofSciences, USA 88, 1711–1715.
Harada, S., Watanabe, Y., Takeuchi, K., Suzuki, T., Katayama, T.,Takebe, Y., Saito, I. & Miyamura, T. (1991). Expression of processed
CEAG

Downloaded from www.microbiologyresearch.org by
IP: 144.76.74.74
On: Thu, 01 Feb 2018 15:12:24
Review: Molecular virology of HCVReview: Molecular virology of HCV
core protein of hepatitis C virus in mammalian cells. Journal of Virology65, 3015–3021.
He, L. F., Alling, D., Popkin, T., Shapiro, M., Alter, H. J. & Purcell,R. H. (1987). Determining the size of non-A, non-B hepatitis virus byfiltration. Journal of Infectious Diseases 156, 636–640.
Hijikata, M., Kato, N., Ootsuyama, Y., Nakagawa, M. & Shimotohno, K.(1991). Gene mapping of the putative structural region of the hepatitisC virus genome by in vitro processing analysis. Proceedings of the NationalAcademy of Sciences, USA 88, 5547–5551.
Hijikata, M., Mizushima, H., Akagi, T., Mori, S., Kakiuchi, N., Kato, N.,Tanaka, T., Kimura, K. & Shimotohmo, K. (1993a). Two distinctproteinase activities required for the processing of a putative non-structural precursor protein of hepatitis C virus. Journal of Virology 67,4665–4675.
Hijikata, M., Mizushima, H., Tanji, Y., Komoda, Y., Hirowatari, Y.,Akagi, T., Kato, N., Kimura, K. & Shimotohno, K. (1993b). Proteolyticprocessing and membrane association of putative nonstructural proteinsof hepatitis C virus. Proceedings of the National Academy of Sciences, USA90, 10773–10777.
Hijikata, M., Shimizu, Y. K., Kato, H., Iwamoto, A., Shih, J. W., Alter,H. J., Purcell, R. H. & Yoshikura, H. (1993c). Equilibrium centrifugationstudies of hepatitis C virus : evidence for circulating immune complexes.Journal of Virology 67, 1953–1958.
Hirowatari, Y., Hijikata, M., Tanji, Y., Nyonoya, H., Mizushima, H.,Kimura, K., Tanaka, T., Kato, N. & Shimotohno, K. (1993). Twoproteinase activities in HCV polypeptide expressed in insect cells usingbaculovirus vector. Archives of Virology 133, 349–356.
Honda, M., Brown, E. A. & Lemon, S. M. (1996a). Stability of astem–loop involving the initiator AUG controls the efficiency of internalinitiation of translation on hepatitis C virus RNA. RNA 2, 955–968.
Honda, M., Ping, L.-H., Rijnbrand, R. C. A., Amphlett, E., Clarke, B. E.,Rowlands, D. J. & Lemon, S. M. (1996b). Structural requirements forinitiation of translation by internal ribosome entry within genome-lengthhepatitis C virus RNA. Virology 222, 31–42.
Hussy, P., Langen, H., Mous, J. & Jacobsen, H. (1996). Hepatitis Cvirus core protein : carboxy-terminal boundaries of two processed speciessuggest cleavage by a signal peptide peptidase. Virology 224, 93–104.
Hwang, S. B., Park, K.-J., Kim, Y.-S., Sung, Y. C. & Lai, M. M. C. (1997).Hepatitis C virus NS5B protein is a membrane-associated phosphoproteinwith a predominantly perinuclear localization. Virology 227, 439–446.
Iacovacci, S., Sargiacomo, M., Parolini, I., Ponzetto, A., Peschle, C. &Carloni, G. (1993). Replication and multiplication of hepatitis C virusgenome in human foetal liver cells. Research in Virology 144, 275–279.
Ito, T., Mukaigawa, J., Zuo, J., Hirabayashi, Y., Mitamura, K. & Yasui,K. (1996). Cultivation of hepatitis C virus in primary hepatocyte culturefrom patients with chronic hepatitis C results in release of high titreinfectious virus. Journal of General Virology 77, 1043–1054.
Jackson, R. J., Howell, M. T. & Kaminski, A. (1990). The novelmechanism of initiation of picornavirus RNA translation. Trends inBiochemical Sciences 15, 477–483.
Kaito, M., Watanabe, S., Tsukiyama-Kohara, K., Yamaguchi, K.,Kobayashi, Y., Konishi, M., Yokoi, M., Ishida, S., Suzuki, S. & Kohara,M. (1994). Hepatitis C virus particle detected by immunoelectronmicroscopic study. Journal of General Virology 75, 1755–1760.
Kamer, G. & Argos, P. (1984). Primary structural comparison of RNA-dependent polymerases from plant, animal and bacterial viruses. NucleicAcids Research 12, 7269–7282.
Kanai, A., Tanabe, K. & Kohara, M. (1995). Poly(U) binding activity ofhepatitis C virus NS3 protein, a putative RNA helicase. FEBS Letters 376,221–224.
Kaneko, T., Tanji, Y., Satoh, S., Hijikata, M., Asabe, S., Kimura, K. &Shimotohno, K. (1994). Production of two phosphoproteins from theNS5A region of the hepatitis C viral genome. Biochemical and BiophysicalResearch Communications 205, 320–326.
Kato, N., Hijikata, M., Ootsuyama, Y., Nakagawa, M., Ohkoshi, S.,Sugimura, T. & Shimotohno, K. (1990). Molecular cloning of the humanhepatitis C virus genome from Japanese patients with non-A, non-Bhepatitis. Proceedings of the National Academy of Sciences, USA 87,9524–9528.
Kato, N., Ootsuyama, Y., Ohkoshi, S., Nakazawa, T., Sekiya, H.,Hijikata, M. & Shimotohno, K. (1992). Distribution of plural HCV typesin Japan. Biochemical and Biophysical Research Communications 189,119–127.
Katze, M. G. (1995). Regulation of the interferon-induced PKR: canviruses cope? Trends in Microbiology 3, 75–78.
Kettinen, H. K., Grace, K., Grunert, S., Clarke, B. E., Rowlands, D. &Jackson, R. (1994). Mapping of the internal ribosome entry site at the5« end of the hepatitis C virus. In Proceedings of the International Symposiumon Viral Hepatitis and Liver Disease, Tokyo, pp. 125–131. Edited by K.Nishioka, H. Suziki, S. Mishiro & T. Oda. Tokyo : Springer-Verlag.
Kim, D. W., Gwack, Y., Han, J. H. & Choe, J. (1995). C-terminal domainof the hepatitis C virus NS3 protein contains an RNA helicase activity.Biochemical and Biophysical Research Communications 215, 160–166.
Kim, J. L., Morgenstern, K. A., Lin, C., Fox, T., Dwyer, M. D., Landro,J. A., Chambers, S. P., Markland, W., Lepre, C. A., O’Malley, E. T.,Harbeson, S. L., Rice, C. M., Murcko, M. A., Caron, P. R. & Thomson,J. A. (1996). Crystal structure of the hepatitis C virus NS3 proteasedomain complexed with a synthetic NS4A cofactor peptide. Cell 87,343–355.
Koch, J. O., Lohmann, V., Herian, U. & Bartenschlager, R. (1996). Invitro studies on the activation of the hepatitis C virus NS3 proteinase bythe NS4A cofactor. Virology 221, 54–66.
Kolykhalov, A. A., Agapov, E. V. & Rice, C. M. (1994). Specificity of thehepatitis C virus NS3 serine protease : effects of substitutions at the3}4A, 4A}4B, 4B}5A and 5A}5B cleavage sites on polyproteinprocessing. Journal of Virology 68, 7525–7533.
Kolykhalov, A. A., Feinstone, S. M. & Rice, C. M. (1996). Identificationof a highly conserved sequence element at the 3« terminus of hepatitis Cvirus genome RNA. Journal of Virology 70, 3363–3371.
Komoda, Y., Hijikata, M., Sato, S., Asabe, S.-I., Kimura, K. &Shimotohno, K. (1994). Substrate requirements of hepatitis C virusserine proteinase for intermolecular polypeptide cleavage in E. coli.Journal of Virology 68, 7351–7357.
Kumar, U., Brown, J., Monjardino, J. & Thomas, H. C. (1993). Sequencevariation in the large envelope glycoprotein (E2}NS1) of hepatitis Cvirus during chronic infection. Journal of Infectious Diseases 167, 726–730.
Kumar, U., Monjardino, J. & Thomas, H. C. (1994). Hypervariableregion of hepatitis C virus envelope glycoprotein (E2}NS1) in agammaglobulinemic patient. Gastroenterology 106, 1072–1075.
Kuo, G., Choo, Q.-L., Alter, H. J., Gitnick, G. L., Redeker, A. G., Purcell,R. H., Miyamura, T., Dienstag, J. L., Alter, M. J., Stevens, C. E.,Tegtmeyer, G. E., Bonino, F., Colombo, M., Lee, W.-E., Kuo, C., Berger,K., Shuster, J. R., Overby, L. R., Bradley, D. W. & Houghton, M.(1989). An assay for circulating antibodies to a major etiologic virus ofhuman non-A, non-B hepatitis. Science 244, 362–364.
Lanford, R. E., Notvall, L., Chavez, D., White, R., Frenzel, G., Simonsen,C. & Kim, J. (1993). Analysis of hepatitis C virus capsid, E1 and E2}NS1proteins expressed in insect cells. Virology 197, 225–235.
Lanford, R. E., Sureau, C., Jacob, J. R., White, R. & Fuerst, T. R. (1994).
CEAH

Downloaded from www.microbiologyresearch.org by
IP: 144.76.74.74
On: Thu, 01 Feb 2018 15:12:24
B. ClarkeB. Clarke
Demonstration of in vitro infection of chimpanzee hepatocytes withhepatitis C virus using strand-specific RT}PCR. Virology 202, 606–614.
Le, S.-Y., Sonenberg, N. & Maizel, J. V. (1995). Unusual folding regionsand ribosome landing pad within hepatitis C virus and pestivirus RNAs.Gene 154, 137–143.
Lin, C., Lindenbach, B. D., Pragal, B. M., McCourt, D. W. & Rice, C. M.(1994a). Processing of the hepatitis C virus E2–NS2 region : identifi-cation of p7 and two distinct E2-specific products with different Ctermini. Journal of Virology 68, 5063–5073.
Lin, C., Pragai, B. M., Grakoui, A., Xu, J. & Rice, C. M. (1994b).Hepatitis C virus NS3 proteinase : trans-cleavage requirements andprocessing kinetics. Journal of Virology 68, 8147–8157.
Lin, C., Thomson, J. A. & Rice, C. M. (1995). A central region in thehepatitis C virus NS4A protein allows formation of an active NS3–NS4Aserine protease complex in vivo and in vitro. Journal of Virology 69,4373–4380.
Lloyd, R. E., Etchison, D. & Ehrenfeld, E. (1985). Poliovirus proteasedoes not mediate cleavage of the 220,000 Da component of the cap-binding complex. Proceedings of the National Academy of Sciences, USA 82,2723–2727.
Lo, S.-Y., Selby, M., Tong, M. & Ou, J. H. (1994). Comparative studiesof the core gene products of two different hepatitis C virus isolates : twoalternative forms determined by a single amino acid substitution. Virology199, 124–131.
Lo, S.-Y., Masiarz, F., Hwang, S. B., Lai, M. M. C. & Ou, J.-H. (1995).Differential subcellular localisation of hepatitis C virus core geneproducts. Virology 213, 455–461.
Lo, S.-Y., Selby, M. J. & Ou, J.-H. (1996). Interaction between hepatitisC virus core protein and E1 envelope protein. Journal of Virology 70,5177–5182.
Love, R. A., Parge, H. E., Wickersham, J. A., Hostomsky, Z., Habuka,N., Moomaw, E. W., Adachi, T. & Hostomska, Z. (1996). The crystalstructure of hepatitis C virus NS3 proteinase reveals a trypsin-like foldand a structural zinc binding site. Cell 87, 331–342.
Manabe, S., Fuke, I., Tanishita, O., Kaji, C., Gomi, Y., Yoshida, S., Mori,C., Takamizawa, A., Yosida, I. & Okayama, H. (1994). Production ofnonstructural proteins of hepatitis C virus requires a putative viralprotease encoded by NS3. Virology 198, 636–644.
Matsumoto, M., Hwang, S. B., Jeng, K.-S., Zhu, N. & Lai, M. C. (1996).Homotypic interaction and multimerization of hepatitis C virus coreprotein. Virology 218, 43–51.
Matsumoto, M., Hsieh, T.-Y., Zhu, N., Vanarsdale, T., Hwang, S. B.,Jeng, K.-S., Gorbalenya, A. E., Lo, S.-Y., Ou, J.-H., Ware, C. F. & Lai, M.M. C. (1997). Hepatitis C virus core protein interacts with thecytoplasmic tail of lymphotoxin-β receptor. Journal of Virology 71,1301–1309.
Matsuura, Y., Suzuki, T., Suzuki, R., Sato, M., Aizaki, H., Saito, I. &Miyamura, T. (1994). Processing of E1 and E2 glycoproteins of hepatitisC virus expressed in mammalian and insect cells. Virology 205, 141–150.
Miyamura, T. & Matsuura, Y. (1993). Structural proteins of hepatitis Cvirus. Trends in Microbiology 1, 229–231.
Mizushima, H., Hijikata, M., Asabe, S., Hirota, M., Kimura, K. &Shimotohno, K. (1994). Two hepatitis C virus glycoprotein E2 productswith different C termini. Journal of Virology 68, 6215–6222.
Mizutani, T., Kato, N., Hirota, M., Sugiyama, K., Murakami, A. &Shimotohno, K. (1995). Inhibition of hepatitis C virus replication byantisense oligonucleotides in cultured cells. Biochemical and BiophysicalResearch Communications 212, 906–911.
Mizutani, T., Kato, N., Saito, S., Ikeda, M., Sugiyama, K. & Shimotohno,K. (1996). Characterization of hepatitis C virus replication in cloned cellsobtained from a human T-cell leukaemia virus type I infected cell line,MT-2. Journal of Virology 70, 7219–7223.
Moradpour, D., Englert, C., Wakita, T. & Wands, J. R. (1996).Characterization of cell lines allowing tightly regulated expression ofhepatitis C virus core protein. Virology 222, 51–63.
Morgenstern, K. A., Landro, J. A., Hsiao, K., Lin, C., Gu, Y., Su, S.-S. &Thomson, J. A. (1997). Polynucleotide modulation of the protease,nucleoside triphosphatase, and helicase activities of a hepatitis C virusNS3–NS4A complex isolated from transfected COS cells. Journal ofVirology 71, 3767–3775.
Nakajima, N., Hijikata, M., Yoshikura, H. & Shimizu, Y. K. (1996).Characterization of long-term cultures of hepatitis C virus. Journal ofVirology 70, 3325–3329.
Ogata, N., Alter, H. J., Miller, R. H. & Purcell, R. H. (1991). Nucleotidesequence and mutation rate of the H strain of hepatitis C virus. Proceedingsof the National Academy of Sciences, USA 88, 3392–3396.
Okamoto, H., Okada, S., Sugiyama, Y., Kurai, K., Iizuka, H., Machida,A., Miyakawa, Y. & Mayumi, M. (1991). Nucleotide sequence of thegenomic RNA of hepatitis C virus isolated from a human carrier :comparison with reported isolates for conserved and divergent regions.Journal of General Virology 72, 2697–2704.
Okamoto, H., Kurai, K., Okada, S-I., Yamamoto, K., Lizuka, H., Tanaka,T., Fukuda, S., Tsuda, F. & Mishiro, S. (1992). Full-length sequence ofa hepatitis C virus genome having poor homology to reported isolates :comparative study of four distinct genotypes. Virology 188, 331–341.
Poole, T. L., Wang, C., Popp, R. A., Potgieter, L. N. D., Siddiqui, A. &Collett, M. S. (1995). Pestivirus translation initiation occurs by internalribosomal entry. Virology 206, 750–754.
Preugschat, F., Averett, D. R., Clarke, B. E. & Porter, D. J. T. (1996). Asteady-state and pre-steady-state kinetic analysis of the NTPase activityassociated with the hepatitis C virus NS3 helicase domain. Journal ofBiological Chemistry 271, 24449–24457.
Prince, A. M., Huima-Byron, T., Parker, T. S. & Levine, D. M. (1996).Visualization of hepatitis C virions and putative defective interferingparticles isolated from low-density lipoproteins. Journal of Viral Hepatitis3, 11–17.
Ralston, R., Thudium, K., Berger, K., Kuo, C., Gervase, B., Hall, J.,Selby, M., Kuo, G., Houghton, M. & Choo, Q.-L. (1993). Charac-terization of hepatitis C virus envelope glycoprotein complexes ex-pressed by recombinant vaccinia viruses. Journal of Virology 67,6753–6761.
Ray, R. B., Meyer, K. & Ray, R. (1996). Suppression of apoptotic celldeath by hepatitis C virus core protein. Virology 176, 182–185.
Reed, K. E., Grakoui, A. & Rice, C. M. (1995). Hepatitis C virus-encodedNS2-3 protease : cleavage-site mutagenesis and requirements for bi-molecular cleavage. Journal of Virology 69, 4127–4136.
Reynolds, J. E., Kaminski, A., Kettinen, H. J., Grace, K., Clarke, B. E.,Rowlands, D. J. & Jackson, R. J. (1995). Unique features of internalinitiation of hepatitis C virus RNA translation. EMBO Journal 14,6010–6020.
Rijnbrand, R., Bredenbeek, P., Van Der Straaten, T., Whetter, L.,Inchauspe, G., Lemon, S. & Spaan, W. (1995). Almost the entire 5« non-translated region of hepatitis C virus is required for cap-independenttranslation. FEBS Letters 365, 115–119.
Rijnbrand, R. C. A., Abbink, T. E. M., Haasnoot, P. C. J., Spaan, W. J. M.& Bredenbeek, P. J. (1996). The influence of AUG codons in thehepatitis C virus 5« nontranslated region on translation and mapping ofthe translation initiation window. Virology 226, 47–56.
CEAI

Downloaded from www.microbiologyresearch.org by
IP: 144.76.74.74
On: Thu, 01 Feb 2018 15:12:24
Review: Molecular virology of HCVReview: Molecular virology of HCV
Rosa, D., Campagnoli, S., Moretto, C., Guenzi, E., Cousens, L., Chin,M., Dong, C., Weiner, A. J., Lau, J. Y. N., Choo, Q.-L., Chien, D., Pileri,P., Houghton, M. & Abrignani, S. (1996). A quantitative test to estimateneutralizing antibodies to the hepatitis C virus : cytofluorometricassessment of envelope glycoprotein 2 binding to target cells. Proceedingsof the National Academy of Sciences, USA 93, 1759–1763.
Saito, I., Miyamura, T., Ohbayashi, A., Harada, H., Katayama, T.,Kikuchi, S., Watanabe, T. Y., Koi, S., Onji, M., Ohta, Y., Choo, Q.-L.,Houghton, M. & Kuo, G. (1990). Hepatitis C virus infection is associatedwith the development of hepatocellular carcinoma. Proceedings of theNational Academy of Sciences, USA 87, 6547–6549.
Sakamuro, D., Furukawa, T. & Takegami, T. (1995). Hepatitis C virusnonstructural protein NS3 transforms NIH 3T3 cells. Journal of Virology69, 3893–3896.
Santolini, E., Migliaccio, G. & La Monica, N. (1994). Biosynthesis andbiochemical properties of the hepatitis C virus core protein. Journal ofVirology 68, 3631–3641.
Santolini, E., Pacini, L., Fipaldini, C., Migliaccio, G. & La Monica, N.(1995). The NS2 protein of hepatitis C virus is a transmembranepolypeptide. Journal of Virology 69, 7461–7471.
Satoh, S., Tanji, Y., Hijikata, M., Kimura, K. & Shimotohno, K. (1995).The N-terminal region of hepatitis C virus non-structural protein 3 (NS3)is essential for stable complex formation with NS4A. Journal of Virology69, 4255–4260.
Selby, M. J., Glazer, E., Masiarz, F. & Houghton, M. (1994). Complexprocessing and protein :protein interactions in the E2 :NS2 region ofHCV. Virology 204, 114–122.
Shih, C. M., Lo, S. J., Miyamura, T., Chen, S. Y. & Lee, Y. W. (1993).Suppression of hepatitis B virus expression and replication by hepatitis Cvirus core protein in Huh-7 cells. Journal of Virology 67, 5823–5832.
Shimizu, Y. K., Iwamoto, A., Hijikata, M., Purcell, R. H. & Yoshikura, H.(1992). Evidence for in vitro replication of hepatitis C virus genome in ahuman T-cell line. Proceedings of the National Academy of Sciences, USA 89,5477–5481.
Shimizu, Y., Yamaji, K., Masuho, Y., Yokota, T., Inoue, H., Sudo, K.,Satoh, S. & Shimotohno, K. (1996a). Identification of the sequence onNS4A required for enhanced cleavage of the NS5A}5B site by hepatitisC virus NS3 protease. Journal of Virology 70, 127–132.
Shimizu, Y. K., Feinstone, S. M., Kohara, M., Purcell, R. H. & Yoshikura,H. (1996b). Hepatitis C virus : detection of intracellular virus particles byelectron microscopy. Hepatology 23, 205–209.
Shimizu, Y. K., Igarashi, H., Kiyohara, T., Cabezon, T., Farci, P.,Purcell, R. H. & Yoshikura, H. (1996c). A hyperimmune serum againsta synthetic peptide corresponding to the hypervariable region 1 ofhepatitis C virus can prevent viral infection in cell cultures. Virology 223,409–412.
Simmonds, P. (1995). Variability of hepatitis C virus (Review).Hepatology 21, 570–583.
Simmonds, P., McOmish, F., Yap, P. L., Chan, S.-W., Lin, C. K.,Dusheiko, G., Saeed, A. A. & Holmes, E. C. (1993). Sequence variabilityin the 5« non-coding region of hepatitis C virus : identification of a newvirus type and restrictions on sequence diversity. Journal of GeneralVirology 74, 661–668.
Smith, D. B., Mellor, J., Jarvis, L. M., Davidson, F., Kolberg, J., Urdea,M., Yap, P.-L., Simmonds, P. & The International HCV CollaborativeStudy Group (1995). Variation of the hepatitis C virus 5« non-codingregion : implications for secondary structure, virus detection and typing.Journal of General Virology 76, 1749–1761.
Srinivas, R. V., Ray, R. B., Meyer, K. & Ray, R. (1996). Hepatitis C virus
core protein inhibits human immunodeficiency virus type 1 replication.Virus Research 45, 87–92.
Steinkuhler, C., Urbani, A., Tomei, L., Biasiol, G., Sardana, M., Bianchi,E., Pessi, A. & De Francesco, R. (1996). Activity of purified hepatitis Cvirus protease NS3 on peptide substrates. Journal of Virology 70,6694–6700.
Stempniak, M., Hostomska, Z., Nodes, B. R. & Hostomsky, Z. (1997).The NS3 proteinase domain of hepatitis C virus is a zinc-containingenzyme. Journal of Virology 71, 2881–2886.
Sugiyama, K., Kato, N., Mizutani, T., Ikeda, M., Tanaka, T. &Shimotohno, K. (1997). Genetic analysis of the hepatitis C virus (HCV)genome from HCV-infected human T cells. Journal of General Virology 78,329–336.
Suzich, J. A., Tamura, J. K., Palmer-Hill, F., Warrener, P., Grakoui, A.,Rice, C. M., Feinstone, S. M. & Collett, M. S. (1993). Hepatitis C virusNS3 protein polynucleotide-stimulated nucleoside triphosphatase andcomparison with the related pestivirus and flavivirus enzymes. Journal ofVirology 67, 6152–6158.
Tai, C.-L., Chi, W.-K., Chen, D.-S. & Hwang, L.-H. (1996). The helicaseactivity associated with hepatitis C virus nonstructural protein 3 (NS3).Journal of Virology 70, 8477–8484.
Takamizawa, A., Mori, C., Fuke, I., Manabe, S., Murakami, S., Fujita, J.,Onishi, E., Andoh, T., Yoshida, I. & Okayama, H. (1991). Structure andorganization of the hepatitis C virus genome isolated from humancarriers. Journal of Virology 65, 1105–1113.
Tanaka, T., Kato, N., Cho, M.-J. & Shimotohno, K. (1995). A novelsequence found at the 3« terminus of hepatitis C virus genome. Biochemicaland Biophysical Research Communications 215, 744–749.
Tanaka, T., Kato, N., Cho, M.-J., Sugiyama, K. & Shimotohno, K.(1996). Structure of the 3« terminus of the hepatitis C virus genome.Journal of Virology 70, 3307–3312.
Tanji, Y., Hijikata, M., Satoh, S., Kaneko, T. & Shimotohno, K. (1995a).Hepatitis C virus-encoded nonstructural protein NS4A has versatilefunctions in viral protein processing. Journal of Virology 69, 1575–1581.
Tanji, Y., Kaneko, T., Satoh, S. & Shimotohno, K. (1995b). Phos-phorylation of hepatitis C virus-encoded nonstructural protein NS5A.Journal of Virology 69, 3980–3986.
Thomssen, R., Bonk, S., Propfe, C., Heermann, K.-H., Kochel, H. G. &Uy, A. (1992). Association of hepatitis C virus in human sera with β-lipoprotein. Medical and Microbiological Immunology 181, 293–300.
Tomei, L., Failla, C., Santolini, E., De Francesco, R. & La Monica, N.(1993). NS3 is a serine protease required for processing of hepatitis Cvirus polyprotein. Journal of Virology 67, 4017–4026.
Tsukiyama-Kohara, K., Iizuka, N., Kohara, M. & Nomoto, A. (1992).Internal ribosome entry site within hepatitis C virus RNA. Journal ofVirology 66, 1476–1483.
Wang, C., Sarnow, P. & Siddiqui, A. (1993). Translation of humanhepatitis C virus RNA in cultured cells is mediated by an internalribosome-binding mechanism. Journal of Virology 67, 3338–3344.
Wang, C., Sarnow, P. & Siddiqui, A. (1994). A conserved helical elementis essential for internal initiation of translation of hepatitis C virus RNA.Journal of Virology 68, 7301–7304.
Wang, C., Le, S., Ali, N. & Siddiqui, A. (1995). An RNA pseudoknot isan essential structural element of the internal ribosome entry site locatedwithin the hepatitis C virus 5« non-coding region. RNA 1, 526–537.
Weiner, A. J., Brauer, M. J., Rosenblatt, J., Richman, K. H., Tung, J.,Crawford, K., Bonino, F., Saracco, G., Choo, Q. L. & Houghton, M.(1991). Variable and hypervariable domains are found in the regions of
CEAJ

Downloaded from www.microbiologyresearch.org by
IP: 144.76.74.74
On: Thu, 01 Feb 2018 15:12:24
B. ClarkeB. Clarke
HCV corresponding to the flavivirus envelope and NS1 proteins and thepestivirus envelope glycoproteins. Virology 180, 842–848.
Willems, M., Peerlinck, K., Moshage, H., Deleu, I., Van Den Eynde, C.,Vermylen, J. & Jap, S. H. (1996). Hepatitis C virus RNA in plasma andperipheral blood mononuclear cells of hemophiliacs with chronic hepatitisC : evidence for viral replication in peripheral blood mononuclear cells.Journal of Medical Virology 42, 272–278.
Yamada, N., Tanihara, K., Takada, A., Yorihuzi, T., Tsutsumi, M.,Shihmomura, H., Tsuji, T. & Date, T. (1996). Genetic organization anddiversity of the 3«non-coding region of the hepatitis C virus genome.Virology 223, 255–261.
Yao, N., Hesson, T., Cable, M., Hong, Z., Kwong, A. D., Le, H. V. &Weber, P. C. (1997). Structure of the hepatitis C virus RNA helicasedomain. Nature Structural Biology 4, 463–467.
Yen, J.-H., Chang, S. C., Hu, C.-R., Chu, S.-C., Lin, S.-S., Hsieh, Y.-S. &Chang, M.-F. (1995). Cellular proteins specifically bind to the 5«-noncoding region of hepatitis C virus RNA. Virology 208, 723–732.
Yoo, B. J., Spaete, R. R., Geballe, A. P., Selby, M., Houghton, M. &Han, J. H. (1992). 5« end-dependent translation initiation of hepatitis Cviral RNA and the presence of putative positive and negative translationalcontrol elements within the 5« untranslated region. Virology 191,889–899.
Yoo, B. J., Selby, M. J., Choe, J., Suh, B. S., Choi, S., Joh, J. S., Nuovo,G. J., Lee, H.-S., Houghton, M. & Han, J. H. (1995). Transfection of adifferentiated human hepatoma cell line (Huh7) with in vitro transcribedhepatitis C virus (HCV) RNA and establishment of a long-term culturepersistently infected with HCV. Journal of Virology 69, 32–38.
Zibert, A., Schreier, E. & Roggendorf, M. (1995). Antibodies in humansera specific to hypervariable region 1 of hepatitis C virus can block viralattachment. Virology 208, 653–661.
Zignego, A. L., Macchia, D., Monti, M., Thiers, V., Mazzetti, M., Foschi,M., Maggi, E., Romagnani, S., Gentilini, P. & Brechot, C. (1992).Infection of peripheral mononuclear blood cells by hepatitis C virus.Journal of Hepatology 15, 382–386.
CEBA