molecular phylogenetic analysis of a bacterial mat ... · observations of microbial biofilms...
TRANSCRIPT

Molecular phylogenetic analysis of a bacterial mat community, Le Grotte di Frasassi, Italy
Bess Koffman Senior Integrative Exercise
10 March 2004
Submitted in partial fulfillment of the requirements for a Bachelor of Arts degree
from Carleton College, Northfield, Minnesota

Table of Contents
Abstract…………………………………………………………………………………….i Keywords…………………………………………………………………………………..i Introduction………………………………………………………………………………..1 Methods Cave description and sampling……………………………………………………4 DNA extraction, PCR amplification, and cloning of 16S rRNA genes…………...4 Phylogenetic analysis……………………………………………………………...6 Results Sampling environment and mat structure…………………………………………8 Phylogenetic analysis of 16S rRNA genes derived from mat……………………..8 Discussion………………………………………………………………………………..10 Acknowledgments………………………………………………………………………..14 References Cited…………………………………………………………………………15

i
Molecular phylogenetic analysis of a bacterial mat community, Le Grotte di Frasassi, Italy
Bess Koffman
Carleton College Senior Integrative Exercise Advisor: Jenn Macalady
10 March 2004 Abstract The Frasassi Caves are a currently forming limestone karst system in which biogenic sulfuric acid may play a significant role. High concentrations of sulfide have been found in the Frasassi aquifer, and gypsum deposits point to the presence of sulfur in the cave. White filamentous microbial mats have been observed growing in shallow streams in Grotta Sulfurea, a cave at the level of the water table. A mat was sampled and used in a bacterial phylogenetic study, from which eleven 16S ribosomal RNA (rRNA) gene clones were sequenced. The majority of 16S clones were affiliated with the δ-proteobacteria subdivision of the Proteobacteria phylum, and many grouped with 16S sequences from organisms living in similar environments. This study aims to extend our knowledge of bacterial diversity within relatively simple geochemical environments, and improve our understanding of the biological role in limestone corrosion. Georef Keywords: Sulfide oxidation, sulfate reduction, caves, bacteria, phylogeny, karst

1
Introduction Karst is the name given to large-scale limestone porosity—caves, sinkholes, and
other effects of significant limestone dissolution. Karst usually occurs as water
percolates through limestone. The water dissolves calcium carbonate, CaCO3, and reacts
with it to form bicarbonate, HCO3-; eventually pockets and large cavities form. However,
karst processes are not as simple as this popular explanation. In many cases, geologists
have found evidence of sulfur in limestone karst cave systems—often as gypsum
crystallized on the cave walls (Northrup and Lavoie, 2001). Some of the largest
limestone caves in the world, including Cueva de la Villa Luz in Tabasco, Mexico and
Lechuguilla Cave of Carlsbad Caverns National Park in New Mexico, seem to have been
formed through processes involving both bicarbonate and biogenic sulfuric acid
(Cunningham et al., 1995; Northrup and Lavoie, 2001).
Sulfuric acid, as a strong acid, can dissolve limestone more effectively than can
bicarbonate or its protonated cousin, carbonic acid. Sulfuric acid forms through the
oxidation of hydrogen sulfide, H2S, to sulfate, SO42- (Fig. 1a). When sulfate reacts with
water, it forms sulfuric acid, H2SO4. When sulfuric acid reacts with calcium carbonate,
CaCO3, it makes gypsum, CaSO4 (Fig. 1b). Water and carbon dioxide released through
the formation of gypsum react with calcium carbonate to make bicarbonate or carbonic
acid (Fig. 1c) (Galdenzi et al., 1999).
The Frasassi cave in Italy is a well-known and unusual karst limestone system, in
that, unlike most other well-known limestone caves, it is currently undergoing formation.
Observations of microbial biofilms covering the limestone surfaces within the cave and
of macroscopic microbial mats floating in cave streams have led researchers to postulate
that H2S oxidation is the driving factor behind karstification (Galdenzi et al., 1999).

2
Understanding the role of microorganisms—in particular, sulfur and sulfide oxidizers and
sulfate reducers—in the Frasassi cave may unearth biogeochemical interactions with a
broader application to karst systems.
A valuable aspect of working in Frasassi is that the geochemistry is simpler than
in many other environments (such as a lake or soil sample), and microbial metabolism is
limited by the absence of sunlight and organic material from the surface. Stable isotope
ratio analysis conducted by Galdenzi et al. on organic samples found a distinct
fractionation signature with consistently lighter isotopes of carbon and nitrogen in the
sulfidic sections of the caves compared with those at the caves’ entrances and the surface
(1999). This suggests that all organic compounds present in the cave are autochthonous,
and is consistent with our picture of the cave as an ecosystem completely isolated from
surface nutrients and photosynthesis. For this reason Frasassi is an ideal environment
that allows us to see discrete links between in situ organisms and the cave’s
geochemistry.
Given the geochemical and metabolic requirements of organisms such as those in
Frasassi, it is difficult to follow the traditional microbiological method of culturing
organisms in the lab in order to study their morphologies. This technique places heavy
selective pressure on organisms, with the result that what grows is often out of proportion
with the original microbial mat. Angert et al. note that typically less than 1% of all
microbes from a particular environment can be grown in the lab using standard
enrichment techniques (1998). In particular, organisms that rely on symbiosis or
syntrophy, whereby a substance is catabolized through the metabolic efforts of two or
more organisms that could not catabolize the substance on their own, may not grow at all

3
in the lab (Madigan et al., 2003). These two relationships often produce microbial mats
with intricate layering according to molecule-scale gradients of substances such as
oxygen and hydrogen sulfide (Stal, 2001). Therefore, it is important to rely not only on
culturing but on molecular phylogenetic work to identify organisms and determine
evolutionary relationships.
Some genes change over time faster than others depending on the necessity of
their function to the organism. Genes that code for the most important functions in an
organism are highly conserved and thus are used for molecular work. The 16S rRNA
gene codes for ribosomal RNA, a molecule that plays an important structural and
functional role in the ribosome (Madigan et al., 2003). For this reason it is highly
conserved and is the gene most often used for phylogenetic work. Using divergences
within this one gene, researchers can create phylogenetic trees that show evolutionary
relationships. On these trees, internal nodes represent common ancestors, and the lengths
of branches correspond to evolutionary changes, or more specifically, base pair
substitutions per site. Widespread use of 16S rDNA sequences for phylogenetic analysis
has resulted in a systematic and reliable way to diagram evolutionary relationships and
survey microorganisms present in natural environments (Hall, 2001a).
This paper presents a molecular phylogenetic analysis of 16S rDNA from bacteria
growing in a cave stream microbial mat in the Frasassi Grotta Sulfurea. Results suggest
that the mat contains organisms closely related to known sulfur oxidizing and sulfur
reducing bacteria. These bacteria likely play a significant role in the production of
sulfuric acid and subsequent karstification.

4
Methods Cave description and sampling
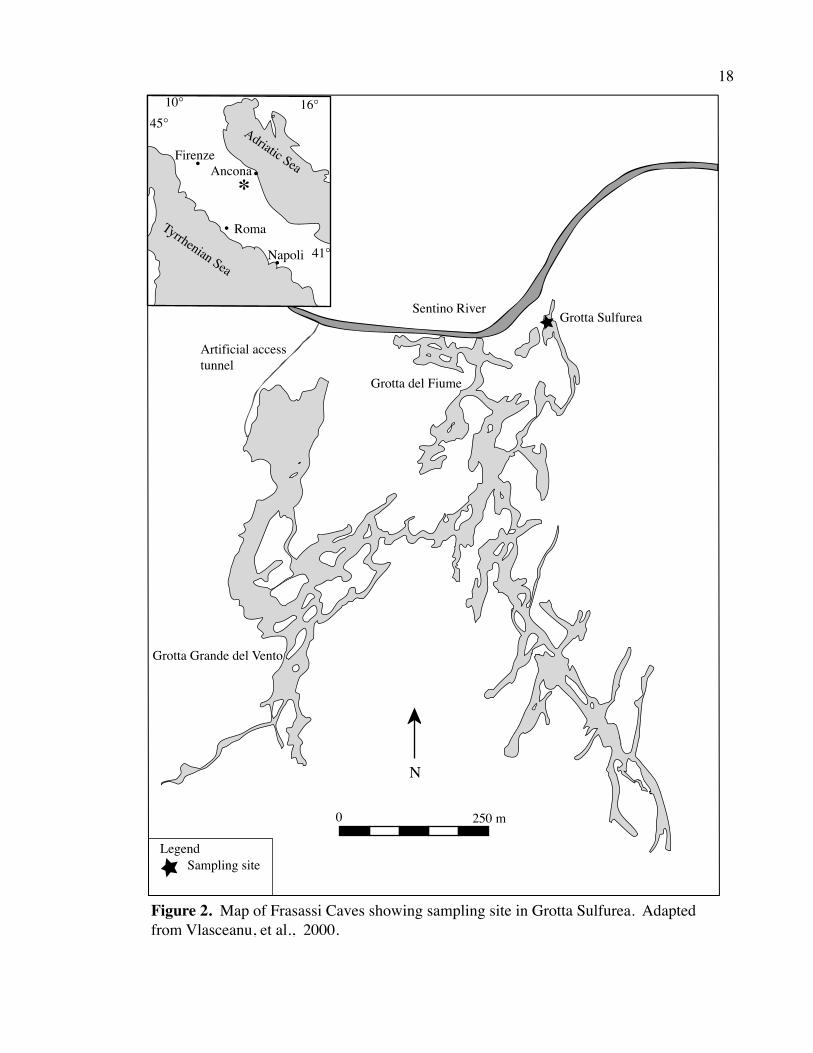
Le Grotte di Frasassi (Frasassi Caves) are located in the Marche region of Italy,
approximately 150 km NE of Rome, in the Sentino river gorge near the village of Genga.
The cave, at about 200-360 m above sea level, includes four main layers within the
massive Calcare Massiccio limestone formation (Galdenzi and Maruoka, 2003). Frasassi
includes over 100 caves in a network of about 35 cave passages (Fig. 2). The highly
permeable Massiccio (which is 600-1000 m thick) rests on the Anidriti del Burano, an
evaporite formation estimated to be 2000 m thick (Galdenzi and Maruoka, 2003). This is
thought to be the sulfur source for the Frasassi caves (Galdenzi and Maruoka, 2003).
Sodium, chloride and sulfur ions (i.e., sulfate and sulfide) are brought to the cave by
groundwater (Fig. 3) (Galdenzi et al., 1999). Sulfide concentrations of up to 0.4 mmol/l
and sulfate concentrations of up to 2.5 mmol/l have been found in the Frasassi aquifer
(Galdenzi et al., 1999). The cave streams keep a constant temperature of 13˚ C and
range in depth from 10 cm to several meters, with flow rates ranging from 0.5 to 50 l/s
(Vlasceanu et al., 2000). Galdenzi et al. found that conductivity of the streams was 1200-
1900 µS and that limestone dissolution occurred both in the streams and in the air at a
rate of 50 µm/yr over the study period of 5 years (1999).
In 2002, Jenn Macalady1 collected samples (“GS tm”) from a macroscopic
filamentous white mat in a cave stream of Grotta Sulfurea, at about 200 m elevation (the
water table) (Fig. 4). Samples were frozen at -20˚ C until transport to the U.S. on dry ice.
Upon arrival in the laboratory, they again were stored at -20˚ C.
1 Assistant Professor, Department of Geology, Carleton College.

5
DNA extraction, PCR amplification, and cloning of 16S rRNA genes Total community DNA was extracted from approximately 30 g (wet wt) frozen
sample using a MoBio Ultraclean Mega Prep Soil DNA kit, following the manufacturer’s
procedure. Presence of DNA was confirmed using agarose gel electrophoresis (1% agar)
with a secondary ethidium bromide staining solution. 16S rRNA genes were amplified
using the polymerase chain reaction (PCR) with 8-27 forward (5´-
AGAGTTTGATCCTGGCTCAG) bacterial and 1510-1492 reverse (5´-
GGTTACCTTGTTACGACTT) universal primers (Fig. 5). The PCR cycle was as
follows: initial denaturation at 94° C for 5 min., denaturation at 94° C for 1 min.,
annealing at 45° C for 45 sec., extension at 72° C for 1.5 min., final extension at 72° C
for 20 min. and storage at 4° C; the PCR ran for 30 cycles. Tests were run against
archaeal and blank controls, and against bacterial DNA extracted from a local stream
microbial mat as a positive control. Clones were transformed with a TOPO TA Cloning®
Kit for Sequencing, according to the manufacturer’s procedure. I used chemically
competent Escherichia coli, which were cultured in Ampicillin LB broth overnight and
then inoculated onto agarose plates.
Colonies from these smear plates and from later streak plates were sampled
randomly. Plasmids were extracted from cells in each colony and 16S rDNA inserts
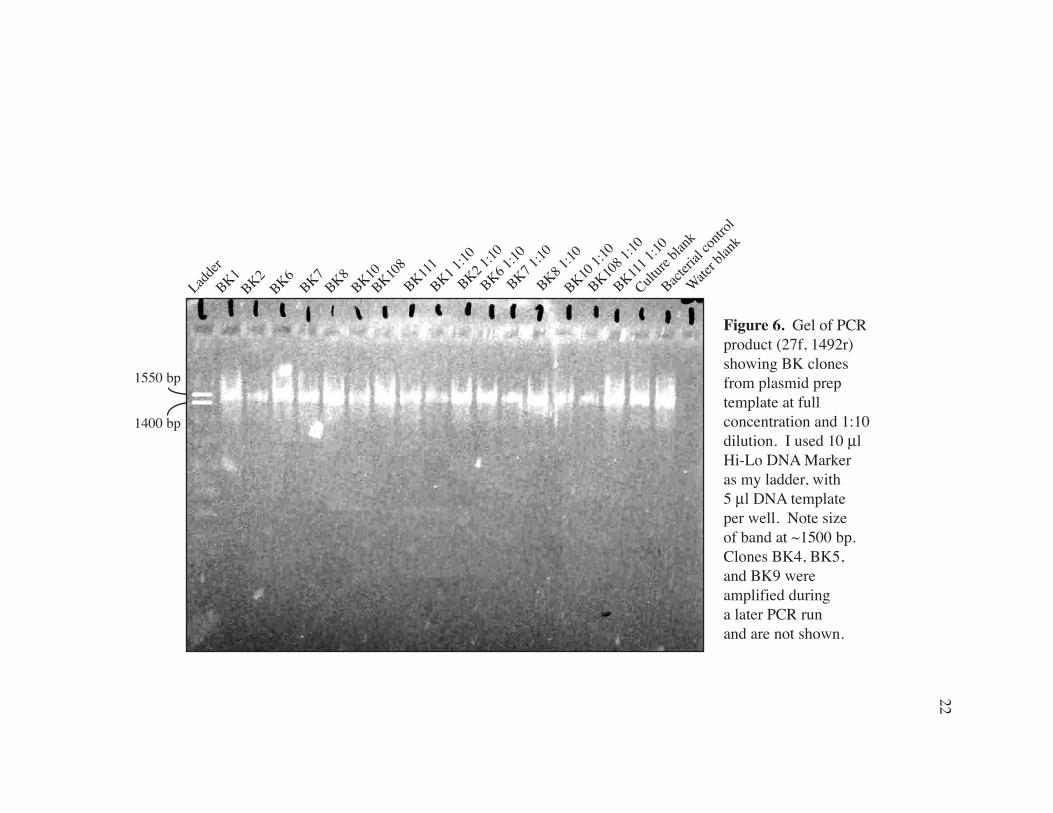
purified using a Wizard® Plus SV Minipreps DNA Purification System (Promega). I
checked for concentration using agarose gel electrophoresis with 3.6 mL culture,
following the manufacturer’s procedure (Fig. 6). PCR with 27f and 1492 primers was
used to amplify the inserts, with the following PCR conditions: initial denaturation at 4°
C for 10 min., denaturation at 94º C for 30 sec., annealing at 47º C for 1 min., extension

6
at 72º C for 2 min., final extension at 72º C for 10 min. and storage at 4º C; the PCR ran
for 30 cycles.
The 16S gene inserts were sequenced at the University of Wisconsin-Madison’s
Biotechnology Center, using cycle sequencing with thermostable polymerases and
fluorescently labeled dideoxy terminators. The primers used were T3 (5’-
ATTAACCCTCACTAAAGGGA) and T7 (5’-TAATACGACTCACTATAGGG).
Phylogenetic analysis
The T3 reverse-complement and T7 ends of each sequence were matched
manually using the multiple alignment program Se-Al (Rambaut, 1996). Sequences were
submitted to the Ribosomal Database Project’s (RDP) online CHECK_CHIMERA
software to test for possible PCR artifacts (sequences recombined from more than one
organism’s 16S gene) as mediated by Taq polymerase (Maidak et al., 1997). In order to
compare the BK clones with evolutionarily similar sequences, my advisor Jenn Macalady
and I searched the ARB 16S rDNA database, which contains over 5000 bacterial
sequences (Ludwig and Strunk, unpublished data). Sequences similar to each of the BK
clones were used for automatic and manual alignments in Seaview and BioEdit (Galtier
et al., 1996; Hall, 2001b; Ludwig and Strunk, unpublished data). Aligning sequences
well is crucial to creating reasonable phylogenies, as poor alignments can lead to
misleading or meaningless trees (Hall, 2001a). I used the Blast program on the website
for the National Center of Biotechnology Information (NCBI) to calculate percent
similarity between cultivated species and my clones (Altschul et al.).
Tree-building methods vary depending on their assumptions about genetic
evolution. Since we as scientists do not know exactly how evolution works, it is

7
important to use multiple methods to construct trees and analyze phylogenetic
relationships. I used both the neighbor-joining algorithm in ARB and Bayesian analysis
to analyze my sequences (Huelsenbeck and Hall, 2000; Ludwig and Strunk, unpublished
data). Neighbor-joining uses a distance matrix derived from a multiple alignment. The
matrix records the calculated distance, or fraction of differences, between each pair of
sequences or lineal groups. In this way it can give a value and a corresponding branch
length to the calculated evolutionary distance (Hall, 2001a). Neighbor-joining is the only
practical tree-building algorithm to use with large data sets, such as the one in ARB.
Large data sets are valuable because they provide a broad context for the placement of
new sequences.
My sequences were aligned using the ARB aligner, checked manually, and added
to the database using ARB’s “QuickAdd” parsimony method. Parsimony is a “minimum
change” method that assumes that the most likely tree is the one with the least changes
between related taxa (Hall, 2001a). It produces trees that show approximate placement of
the organisms and is useful when adding a limited number of sequences to a neighbor-
joining tree. The ARB tree was rooted using several archaeal species.
Trees specific to each bacterial lineage were created from the ARB libraries using
Bayesian inference in MrBayes and were formatted graphically using PAUP*
(Huelsenbeck and Hall, 2000; Swofford, 1999). Bayesian analysis uses posterior
probabilities to find the best set of trees for the given sequence alignment and a
postulated model of evolution that maximizes the probability of observing the given data.
Rather than arriving at a single best tree, Bayesian inference samples trees using the
Markov chain Monte Carlo method, which produces a set of converging consensus

8
trees—trees for which accepting or rejecting any particular change becomes essentially
random. The frequency of any particular tree being sampled is related to the probability
that it is the best tree for the data. Bayesian analysis is quite powerful and is becoming
increasingly popular among phylogeneticists (Hall, 2001a).
I used MrBayes to run Bayesian inference searches for 105 cycles with four
Markov chains to check for convergence. Trees were saved every 100 generations; the
first 350 trees from each cold chain (the Markov chain used for sampling) were
discarded. These were rooted using monophyletic bacterial outgroups.
Results Sampling environment and mat structure
On the sampling expedition of October 2003 to Grotta Sulfurea, Jenn Macalady
and I measured the geochemistry of the shallow flowing cave stream containing the white
microbial mat sampled in 2002. We found cave water temperature to be 13.9-14.6˚ C.
The water had pH = 7.0 ± 0.03. The white filamentous microbial mat was floating on or
below the surface of the stream, and in places was directly on top of the black sulfidic
mud that lined the streambed. The strong “rotten egg” smell of hydrogen sulfide was
quite recognizable.
Phylogenetic analysis of 16S rRNA genes derived from mat From the Grotta Sulfurea cave microbial mat samples, eleven 16S rRNA clones
were sequenced. None of the sequences were positively identified as chimeric using the
RDP software CHECK_CHIMERA. Two sequences (clones BK9 and BK111) were over
99.5% identical and so only one of these (BK9) was used for phylogenetic work. The
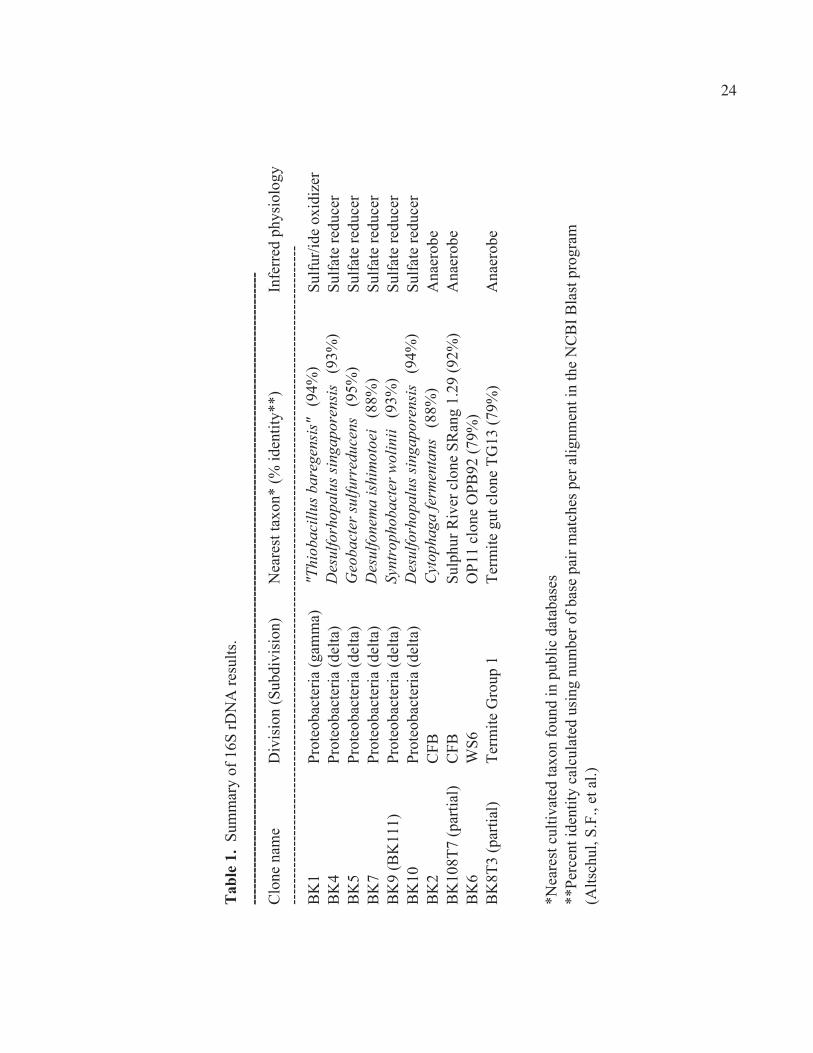
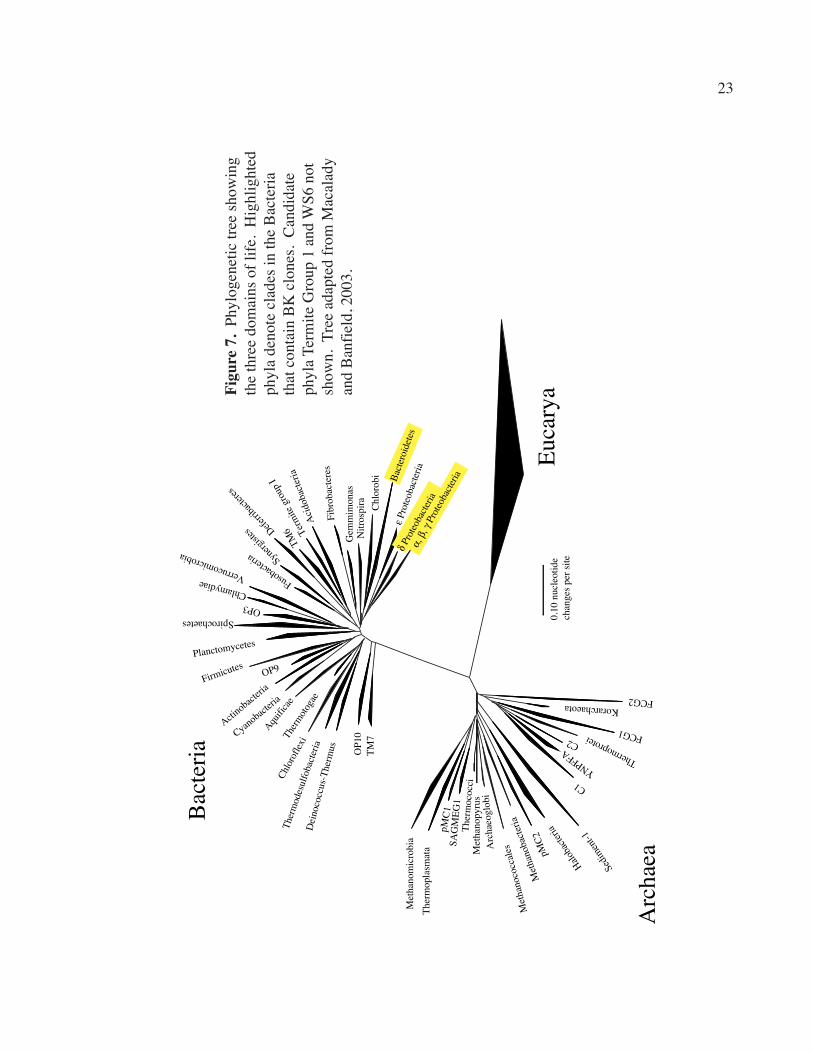
remaining ten sequences fit into four phyla, all in the Bacterial domain (Fig. 7):
Proteobacteria (6 sequences), Cytophaga/Flavobacteria/Bacteroides (CFB) (2
sequences), WS6 (1 sequence), and Termite Group 1 (1 sequence). Of the bacteria in the

9
Proteobacteria phylum, 1 fit in the γ-proteobacteria subdivision, and the remaining 5 in
the δ-proteobacteria subdivision (Table 1). Preliminary results of further phylogenetic
work on clones from cave wall microbial films in Grotta Sulfurea (“GSgm”) are also
shown on ARB trees (Figs. 8, 10, 12, and 14).
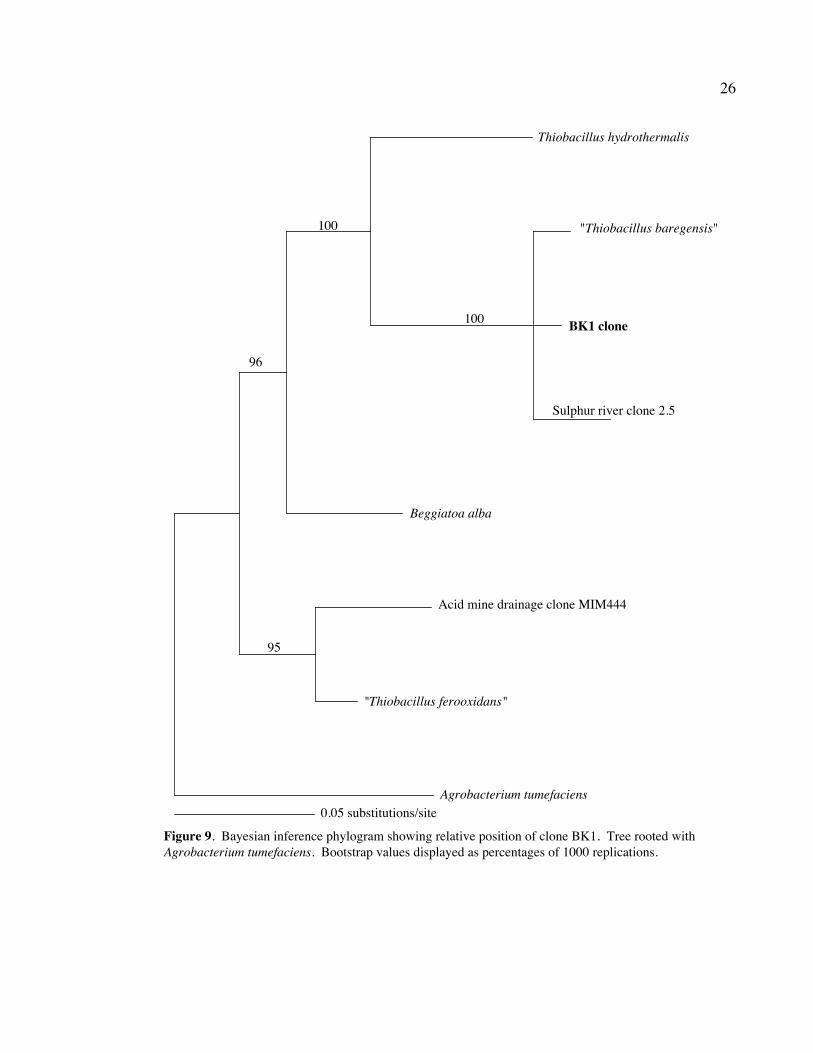
Clone BK1 fit well with the γ-proteobacterium “Thiobacillus baregensis,” a
sulfur oxidizing bacterium found in sulfurated thermal waters in Bareges, France (Hedoin
et al., unpublished 1996). It also grouped with a number of Sulphur River clones, clones
isolated from a white filamentous microbial mat in Sulphur River, Parker Cave, Kentucky
(Angert et al., 1998). Clone BK1 grouped most notably with clone SRang 2.5, which
itself grouped closely with “T.barengensis” (Figs. 8 and 9).
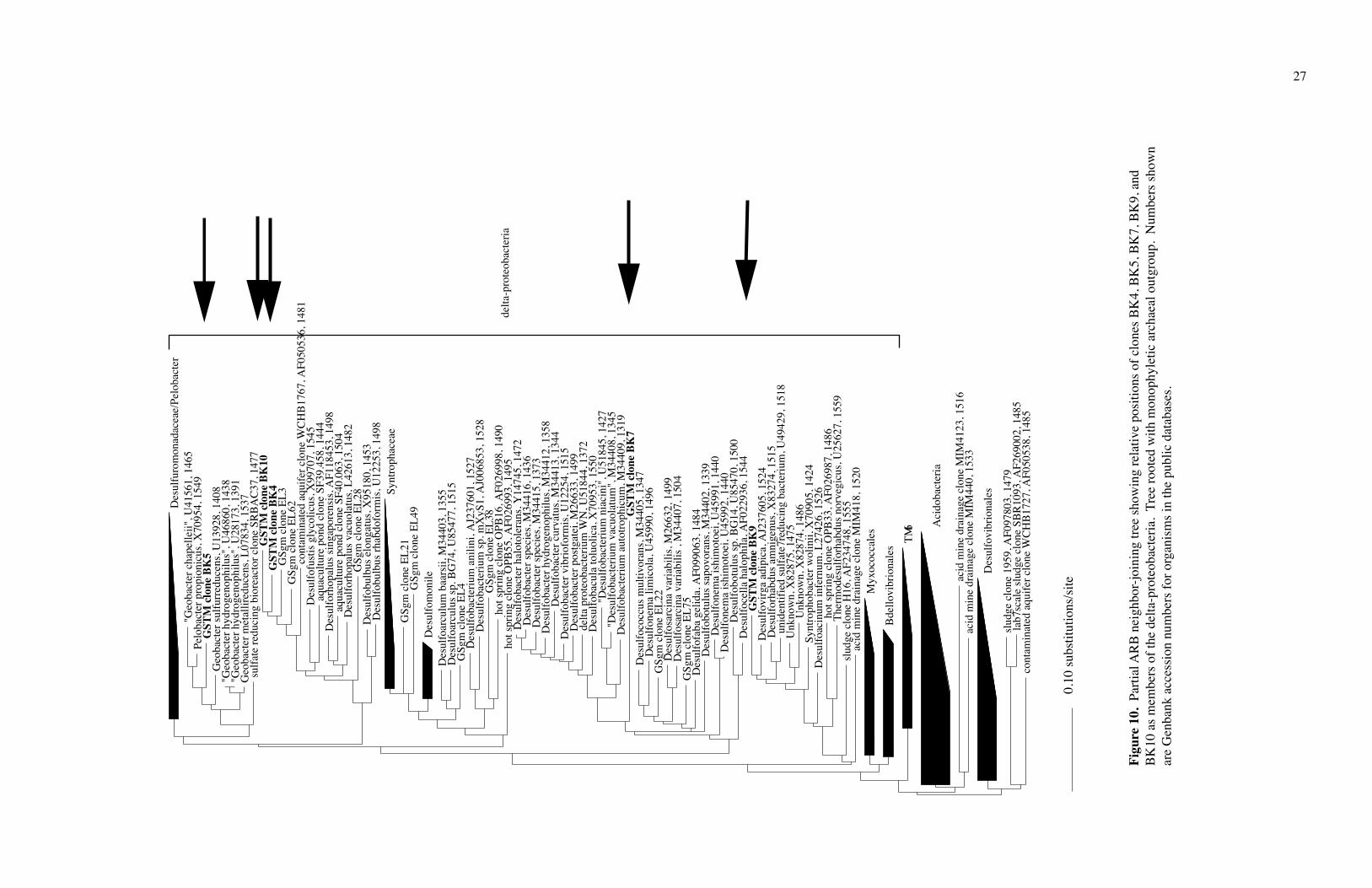
The clones BK4, BK5, BK7, BK9, and BK10 grouped in the δ-proteobacteria, a
lineage of predominantly sulfur-reducing bacteria (Figs. 10 and 11). Clones BK4 and
BK10 clustered together, with their closest cultivated neighbor being Desulfosarcina
singaporensis, a sulfate-reducing bacterium isolated from sulfide-rich black marine mud
(Lie et al., 1999). Clone BK5 was closely related to Geobacter sulfurreducens and to
Geobacter metallireducens, both metal ion reducers (Methe et al., 2003). Clone BK7 fit
near Desulfonema ishimotoei, a sulfate reducer isolated from organic-rich sulfidic marine
and freshwater sediment (Fukui et al., 1999). Clone BK9 was similar to Syntrophobacter
wolinii, a sulfate reducer isolated from a culture enriched from anaerobic granular sludge
(Harmsen et al., 1993).
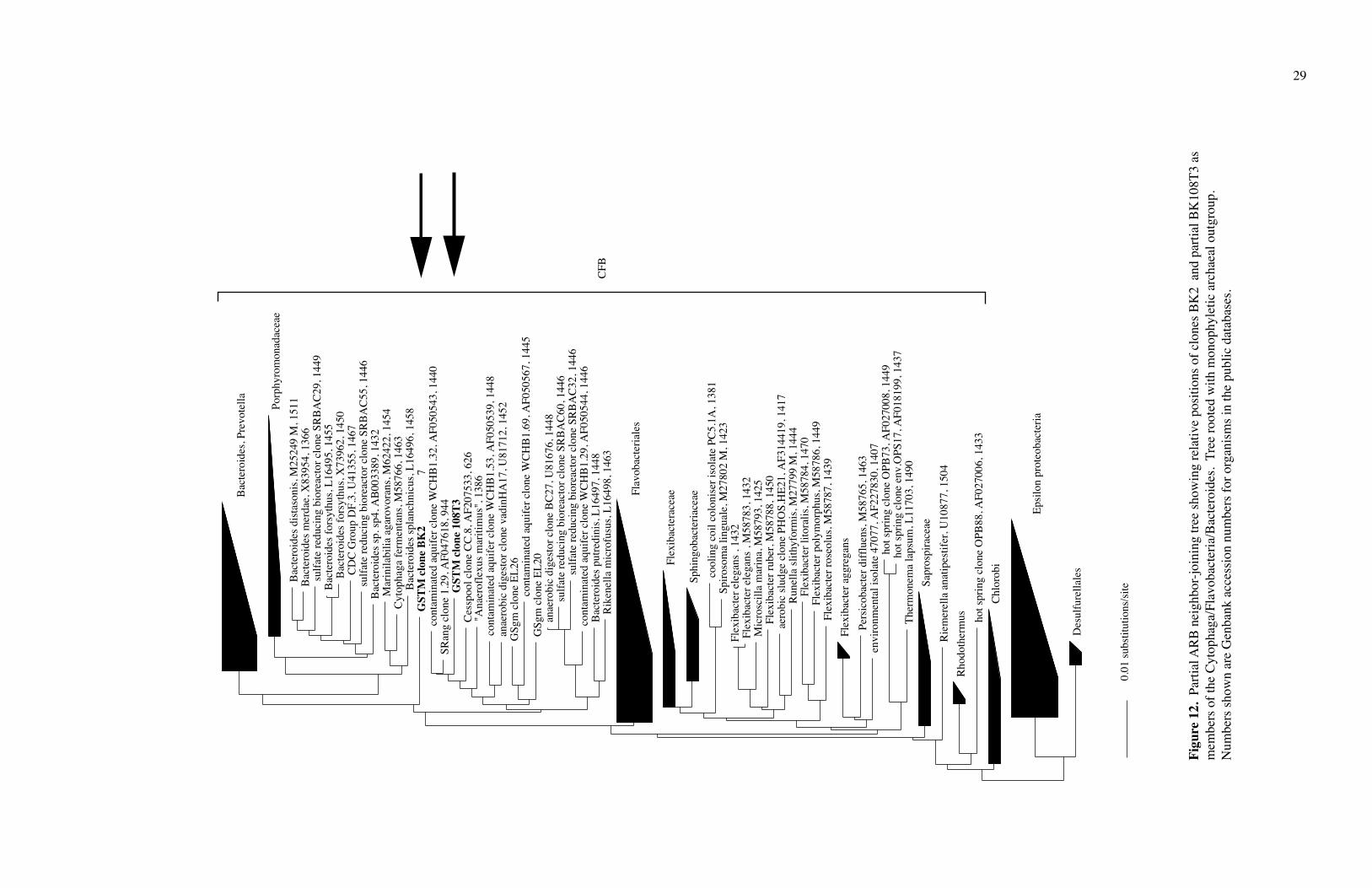
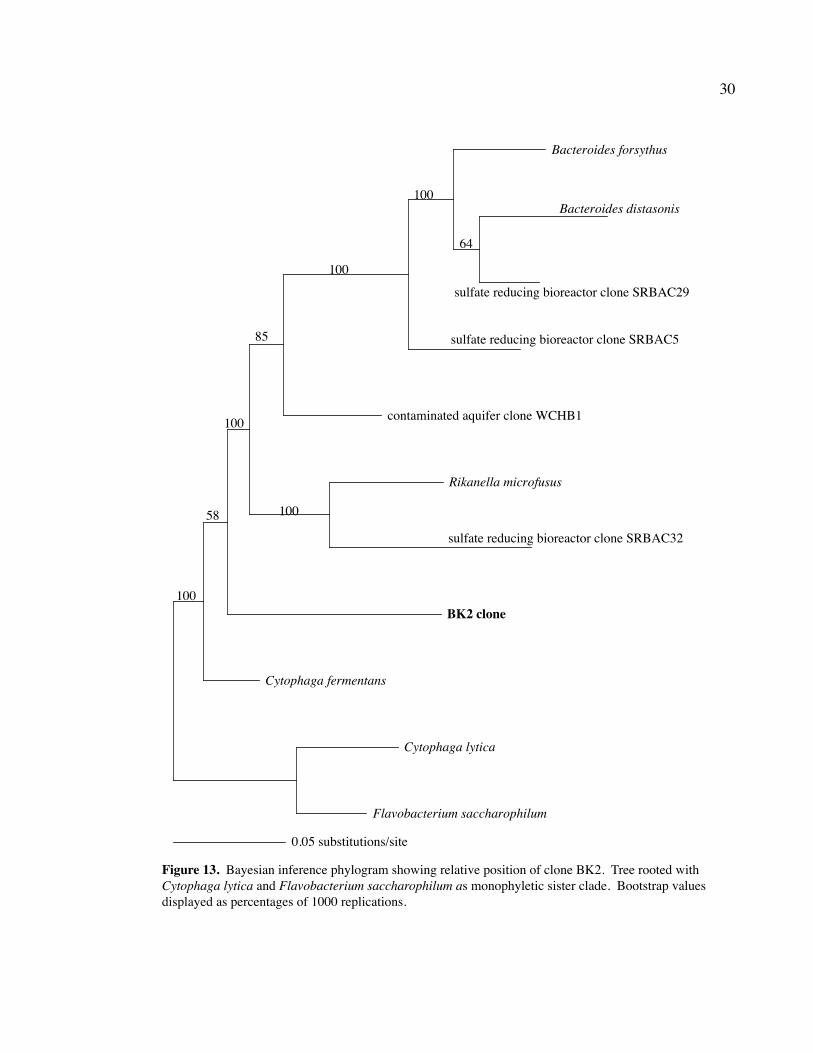
Clone BK2 and a partial sequence of BK108 (T3) fit in the CFB group (Figs. 12
and 13). Clone BK2 was most similar to Cytophaga fermentans. Clone BK108T3 was
closely associated with the Sulphur River clone SRang 1.29, the contaminated aquifer

10
clone WCHB1.32, and the Cesspool Cave clone CC.8. The only cultivated bacteria in the
same group as clone BK108T3 were the fermenter Bacteroides putredinis and the termite
gut bacterium Rikenella microfusus, both anaerobes (Madigan et al., 2003; Ohkuma et al.,
2002).
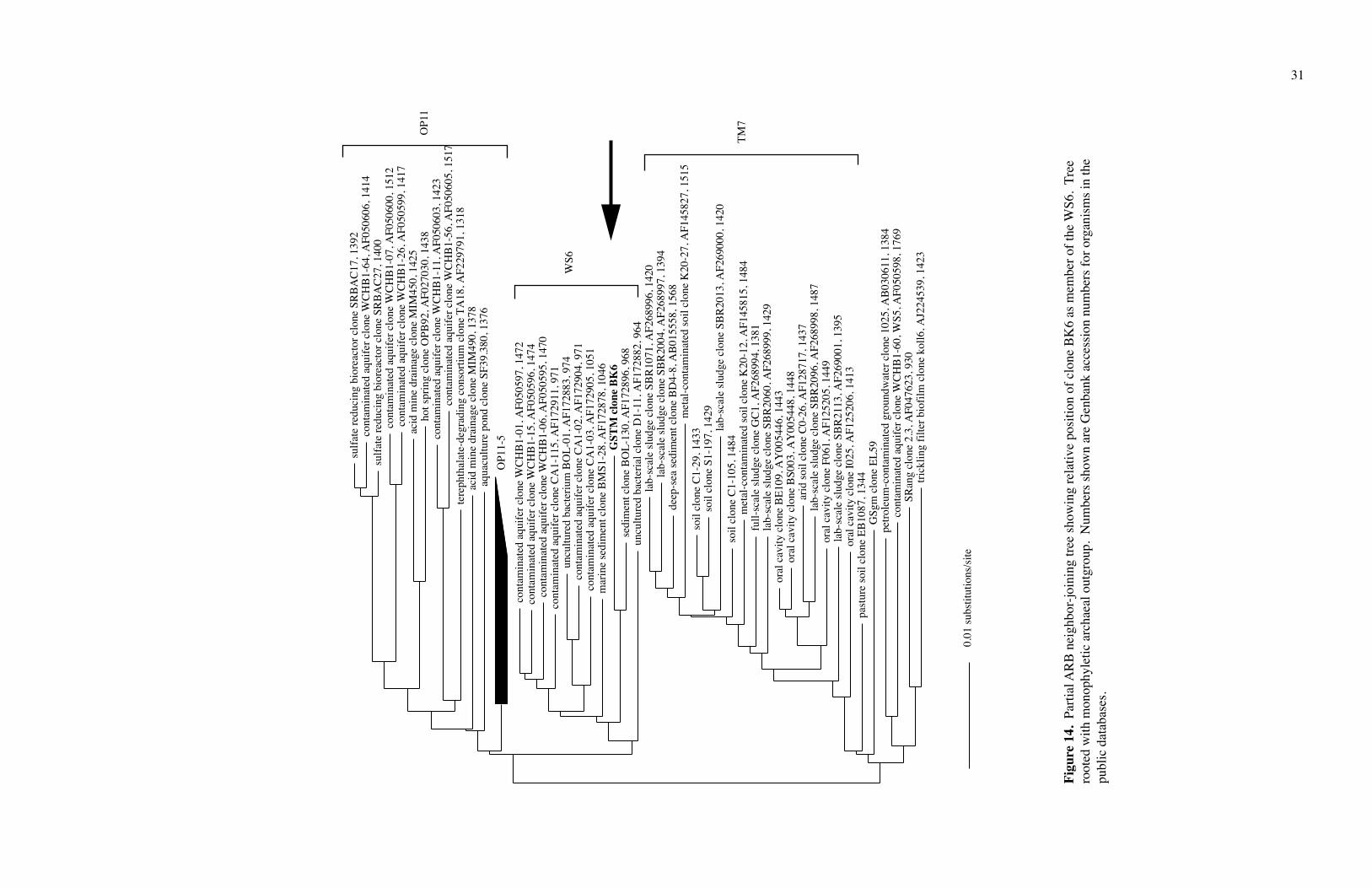
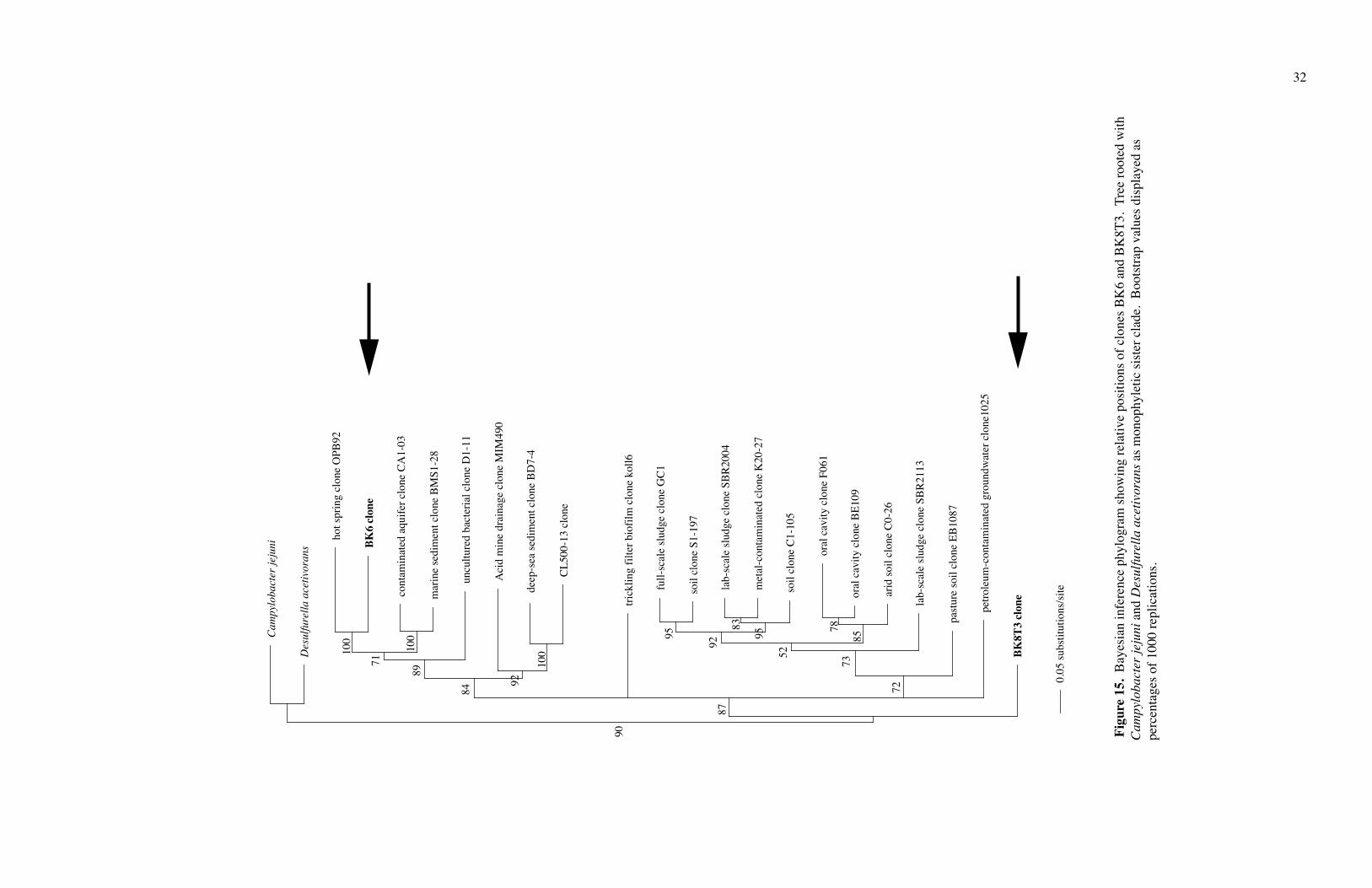
Clone BK6 fit in the WS6 phylum, near the OP11 clone OPB92 (Figs. 14 and 15).
Clone BK8 fit in Termite Group 1 (Figs. 15 and 16). Since I had only a partial sequence
of Clone BK8 (T3), I could not do rigorous phylogenetic work on it. The phyla WS6 and
Termite Group 1 are newly defined and contain no cultivated species.
Discussion The macroscopic filamentous white microbial mat found in the cave stream of
Grotta Sulfurea represents a distinct community isolated from photosynthetically derived
carbon. The observed high sulfide and sulfate levels in the water and air are sufficient to
support a sulfide-metabolizing community (Madigan et al., 2003). Given the sequence
similarities of 88-94% between the Grotta Sulfurea BK clones and known sulfur/ide
oxidizers and sulfate reducers (Table 1), it is reasonable to believe that my 16S clone
sequences came from respective sulfur/ide oxidizers and sulfate reducers, and that the
microbial community in turn includes both these types of bacteria. Since there is no input
of organic material from the surface to Grotta Sulfurea, chemolithoautotrophy likely
forms the base of the food chain. Sulfur oxidizing autotrophic bacteria thus likely
comprise a significant portion of the base-level biomass, a biomass capable of supporting
fifteen invertebrate species, of which seven are endemic to Frasassi (Galdenzi et al.,
1999).
A sulfur-rich environment such as Frasassi would be expected to support both
types of metabolism—especially where sulfur/ide oxidizers and sulfate reducers may

11
form close symbiotic relationships. This has been seen in hydrothermal vent
communities, where bacteria (especially sulfur/ide oxidizers and sulfate reducers) play an
important role in mineralization of vent chimneys (Lovley et al., 2000; McCollom and
Shock, 1997). In fact, sulfur isotope ratios analyzed by Galdenzi and Maruoka (2003)
suggest that sulfate reducing bacteria are present in Frasassi. They found sulfur isotope
values of δ34S ~ -19.60 ‰ in gypsum deposits in the cave, δ34S ~ -14 in H2S and δ34S ~
+21.00 ‰ in sulfate within sulfidic groundwater (Galdenzi and Maruoka, 2003).
Galdenzi and Maruoka (2003) propose that the size of these isotope fractionations can
best be explained by the sulfur cycling through sulfate reducing bacteria, which are
known to produce high fractionations. The presence of sulfate reducers, shown through
phylogenetic work, fits with the sulfur isotope analysis.
Not all clones from similar microbial mats in sulfidic streams have this split
between sulfur/ide oxidation and sulfate reduction. The Sulphur River clones from a
white filamentous mat in Parker Cave, Kentucky are predominantly ε- and γ-
proteobacteria, both lineages known for their sulfur-oxidizing metabolism (Angert et al.,
1998). Similarly, clones from a white filamentous microbial mat in the sulfidic waters of
Lower Kane Cave, Wyoming are almost entirely from the ε-proteobacteria (Engel et al.,
2003). It would seem that these results would be more expected for the Frasassi Grotta
Sulfurea BK clones given the similarities in limestone geology and microbial mat
morphology. There was one γ-proteobacteria, but the majority of clones (5) were δ-
proteobacteria, putative sulfate reducers.
The differences between the Frasassi Grotta Sulfurea clones and those of Sulphur
River and Lower Kane Cave may be due to a difference in sampling technique or an

12
introduced experimental bias. In the DNA extraction there may be lysing bias, whereby
certain cells are more easily opened and their DNA extracted. Similarly, the PCR
primers may anneal with certain sequences better than others. This latter potential source
of difference can be eliminated entirely from the Lower Kane Cave clones, as they were
PCR amplified using the same primers as for the BK clones (27f, 1492r) (Engel et al.,
2003). The Sulphur River clones were amplified using a 515 forward primer and the
same1492 reverse primer (Angert et al., 1998). It is also possible that differences in the
PCR programs used could have affected the results.
Even considering these possible sources of disparity between the Grotta Sulfurea
BK clones and the Sulphur River and Lower Kane Cave clones, a difference between the
phylogenetic results found in each location is evident. Why were there no sulfate
reducers found in either the Sulphur River or the Lower Kane Cave clones? Since sulfur
oxidizers produce sulfate, it would seem that their presence would provide the sulfate
necessary for sulfate reduction metabolism. This apparent difference between the sulfur-
cycling species present in each environment needs to be investigated further.
The presence of only Bacteria in my clone libraries can be attributed to the use of
bacterial PCR primers, which amplify only the bacterial 16S rRNA gene. We would
need to use archaeal primers and sequence a greater number of bacterial clones in order
to explore the full diversity of the Grotta Sulfurea stream microbial mat. Since 16S genes
evolve slowly relative to most other genes, they often do not keep up with changes in the
genes that code for proteins such as those involved in metabolism (Woese, 1987). It
would therefore be germane to sequence protein-coding genes—and even better, entire
genomes. Another important and ongoing aspect of this research is Fluorescence In-Situ

13
Hybridization (FISH), which will help us visualize the population numbers and
distribution of organisms between the two prokaryotic domains.
This study expands the known diversity of the Frasassi Grotta Sulfurea cave
stream microbial mat community based on phylogenetic analysis, and is an excellent
launching point for further physiological and phylogenetic study. The high percent
similarities between my 16S clones and the 16S genes of known sulfur/ide oxidizers and
sulfate reducers point to a biological source of sulfuric acid through biogenic sulfur
oxidation, which implies a crucial role played by bacteria in carving the Frasassi caves.
The presence of both sulfur/ide oxidizers and sulfate reducers fits with sulfur isotope
fractionation data and testifies to the complex sulfur cycle of the Frasassi cave system.

14
Acknowledgments Alessandro Galdenzi accompanied Jenn Macalady and me to Grotta Sulfurea in
October 2003, and for this I am grateful. Alessandro Montanari kindly shipped our
samples from Italy. The computer analysis work would have been impossible without the
help of Doug Foxgrover, who worked hard to install ARB on Jenn’s computer. Leah
Morgan answered my many questions about computer programs and let me use her
computer for BioEdit. My academic advisor Bereket Haileab provided moral support and
perspective. Cam Davidson tutored me in Adobe Illustrator. Tim Vick offered constant
technical support. Funding for this project came from the Bernstein Student Research
Endowment and the NSF Biogeosciences Program. This work builds on research done by
Jenn Macalady, Alessandro Galdenzi2, and Teruyuki Maruoka,3 among others. Finally, I
would like to thank Jenn Macalady, my advisor on this project. Without her vision,
expertise, and support, this research would not have happened.
2 Instituto Italiano di Speleologia, Frasassi Section. 3 Department of Geological Sciences, University of Vienna.

15
References Cited Altschul, S. F., Madden, T. L., Schaffer, A. A., Zhang, J., Zhang, Z., Miller, W., and
Lipman, D. J., Blast 2.04 of the NCBI Toolkit. Angert, E. R., Northup, D. E., Reysenbach, A.-L., Peek, A. S., Goebel, B. M., and Pace,
N. R., 1998, Molecular phylogenetic analysis of a bacterial community in Sulphur River, Parker Cave, Kentucky: American Mineralogist, v. 83, p. 1583-1592.
Cunningham, K. I., Northup, D. E., and Pollastro, R. M., 1995, Bacteria, Fungi, and Biokarst in Lechuguilla Cave, Carlsbad-Caverns-National-Park, New-Mexico: Environmental Geology, v. 25, no. 1, p. 208.
Engel, A. S., Lee, N., Porter, M. L., Stern, L. A., Bennett, P. C., and Wagner, M., 2003, Filamentous "Epsilonproteobacteria" dominate microbial mats from sulfidic cave springs: Applied and Environmental Microbiology, v. 69, no. 9, p. 5503-5511.
Fukui, M., Teske, A., Assmus, B., Muyzer, G., and Widdel, F., 1999, Physiology, phylogenetic relationships, and ecology of filamentous sulfate-reducing bacteria (genus desulfonema): Arch. Microbiol., v. 172, p. 193-203.
Galdenzi, S., and Maruoka, T., 2003, Gypsum deposits in the Frasassi Caves, central Italy: Journal of Cave and Karst Studies, v. 65, no. 2, p. 111-125.
Galdenzi, S., Menichetti, M., Sarbu, S., and Rossi, A., 1999, Frasassi caves: a biogenic hypogean karst system?: Colloque europeen-Karst 99-European Conference, p. 101-106.
Galtier, N., Gouy, M., and Gautier, C., 1996, SEAVIEW and PHYLO_WIN: two graphic tools for sequence alignment and molecular phylogeny.: Comput Appl Biosci, v. 12, no. 6, p. 543-548.
Hall, B. G., 2001, Phylogenetic Trees Made Easy: A How-To Manual for Molecular Biologists: Sunderland, M.A., Sinauer Associates, Inc.
Hall, T., 2001, BioEdit: Raleigh, N.C., North Carolina State University. Harmsen, H. J., Wullings, B., Akkermans, A. D., Ludwig, W., and Stams, A. J., 1993,
Phylogenetic analysis of Syntrophobacter wolinii reveals a relationship with sulfate-reducing bacteria: Arch. Microbiol., v. 160, no. 3, p. 238-240.
Hedoin, H., Dauga, C., Abadie, M., Kaiser, P., and Cerning, J., unpublished 1996, Candidatus Thiobacillus baregensis a new sulfoxidizing bacteriumfrom sulfurated thermal waters of Bareges (France). Applied and Environmental Microbiology, v. In press (unpublished as of 10 March 2004).
Huelsenbeck, J. P., and Hall, B., 2000, MrBayes: A program for the Bayesian inference of phylogeny: Rochester, N.Y., University of Rochester.
Invitrogen life technologies, 2003, TOPO TA Cloning Kit for Sequencing, Version K: Invitrogen Corporation.
Komatsoulis, G. A., and Waterman, M. S., 1997, A new computational method for detection of chimeric 16S rRNA artifacts generated by PCR amplification from mixed bacterial populations: Applied and Environmental Microbiology, v. 63, no. 6, p. 2238-2346.
Lie, T. J., Clawson, M. L., Godchaux, W., and Leadbetter, E. R., 1999, Sulfidogenesis from 2-aminoethanesulfonate (taurine) fermentation by a morphologically unusual sulfate-reducing bacterium, Desulforhopalus singaporensis sp. nov.: Applied and Environmental Microbiology, v. 65, p. 3328-3334.
Ludwig, and Strunk, unpublished data, ARB: Muenchen.

16
Madigan, M. T., Martinko, J. M., and Parker, J., 2003, Brock Biology of Microorganisms: Upper Saddle River, NJ, Pearson Education, Inc.
Maidak, B. L., Olsen, G. J., Larsen, N., Overbeek, R., McCaughey, M. J., and Woese, C. R., 1997, The RDP (Ribosomal Database Project): Nucleic Acids Research, v. 25, no. 1, p. 109-111.
Methe, B. A., Nelson, K. E., Eisen, J. A., Paulsen, I. T., Nelson, W., Heidelberg, J. F., Wu, D., Wu, M., Ward, N., Beanan, M. J., Dodson, R. J., Madupu, R., Brinkac, L. M., Daugherty, S. C., DeBoy, R. T., Durkin, A. S., Gwinn, M., Kolonay, J. F., Sullivan, S. A., Haft, D. H., Selengut, J., Davidsen, T. M., Zafar, N., White, O., Tran, B., Romero, C., Forberger, H. A., Weidman, J., Khouri, H., Feldblyum, T. V., Utterback, T. R., Van Aken, S. E., Lovley, D. R., and Fraser, C. M., 2003, Genome of Geobacter sulfurreducens: Metal reduction in subsurface environments: Science, v. 302, no. 5652, p. 1967-1969.
Northrup, D., and Lavoie, K., 2001, Geomicrobiology of Caves: A Review: Geomicrobiology Journal, v. 18, p. 199-222.
Ohkuma, M., Noda, S., Hongoh, Y., and Kudo, T., 2002, Diverse bacteria related to the bacteroides subgroup of the CFB phylum within the gut symbiotic communities of various termites: Biosci Biotechnol Biochem., v. 66, no. 1, p. 78-84.
Rambaut, A., 1996, Se-Al: Sequence Alignment Editor: Oxford, UK. Stal, L. J., 2001, Coastal microbial mats: the physiology of a small-scale ecosystem:
South African Journal of Botany, v. 67, no. 3, p. 399-410. Swofford, D. L., 1999, PAUP*. Phylogenetic Analysis Using Parsimony (*and Other
Methods). Version 4.: Sunderland, M.A., Sinauer Associates. Vlasceanu, L., Sarbu, S., Engel, A., and Kinkle, B., 2000, Acidic Cave-Wall Biofilms
Located in the Frasassi Gorge, Italy: Geomicrobiology Journal, v. 17, p. 125-139.

(a) H2S + 2 O2 2 H+ + SO42-
(b) 2 H+ + SO42- + CaCO3 + H2O CaSO4 * 2 H2O + CO2
(c) CO2 + CaCO3 + H2O Ca2+ + 2 HCO3-
Figure 1. Various pathways for limestone corrosion: (a) oxidation of sulfide tosulfate, which can bond with water to form sulfuric acid, H2SO4; (b) formation of gypsum, which frees CO2 to form bicarbonate; (c) typical karst-style formation
of bicarbonate. This can become protonated to form carbonic acid, H2CO3.
Adapted from Galdenzi et al., 1999.
17
Grotta SulfureaSentino River
Grotta Grande del Vento
Grotta del Fiume
Ancona*
Firenze
Roma
Napoli
Artificial accesstunnel
N
250 m0
Adriatic Sea
Tyrrhenian Sea
LegendSampling site
45°10° 16°
41°
Figure 2. Map of Frasassi Caves showing sampling site in Grotta Sulfurea. Adapted from Vlasceanu, et al., 2000.
18

Figure 3. Schematic cross section of Frasassi Gorge showing water table and movement of ground water. Adapted from Galdenzi et al., 1999.
~100 m elevation
19

Figure 4. Photographs from Frasassi Gorge and Caves: a) entrance to Grotta Sulfurea from SS 76 in Genga; b) Sandro Galdenzi lowering the rope into the first chute of Grotta Sulfurea; c) sampling the white mat; d) white mat (scale bar is about 0.3 m); e) measuring pH of drips from wall gypsum crystals. Photos by Jenn L. Macalady.
a)
b)
c)
e) d)
20
Clo
ne n
ame
Div
isio
n (S
ubdi
visi
on)
Nea
rest
taxo
n* (
% id
entit
y**)
Infe
rred
phy
siol
ogy
BK
1Pr
oteo
bact
eria
(ga
mm
a)"T
hiob
acill
us b
areg
ensi
s"
(94%
)Su
lfur
/ide
oxid
izer
BK
4Pr
oteo
bact
eria
(de
lta)
Des
ulfo
rhop
alus
sin
gapo
rens
is
(93%
)Su
lfat
e re
duce
rB
K5
Prot
eoba
cter
ia (
delta
)G
eoba
cter
sul
furr
educ
ens
(95
%)
Sulf
ate
redu
cer
BK
7Pr
oteo
bact
eria
(de
lta)
Des
ulfo
nem
a is
him
otoe
i (8
8%)
Sulf
ate
redu
cer
BK
9 (B
K11
1)Pr
oteo
bact
eria
(de
lta)
Synt
roph
obac
ter
wol
inii
(93%
)Su
lfat
e re
duce
rB
K10
Prot
eoba
cter
ia (
delta
)D
esul
forh
opal
us s
inga
pore
nsis
(9
4%)
Sulf
ate
redu
cer
BK
2C
FBC
ytop
haga
ferm
enta
ns
(88%
)A
naer
obe
BK
108T
7 (p
artia
l)C
FBSu
lphu
r R
iver
clo
ne S
Ran
g 1.
29 (
92%
)A
naer
obe
BK
6W
S6O
P11
clon
e O
PB92
(79
%)
BK
8T3
(par
tial)
Ter
mite
Gro
up 1
Ter
mite
gut
clo
ne T
G13
(79
%)
Ana
erob
e
*Nea
rest
cul
tivat
ed ta
xon
foun
d in
pub
lic d
atab
ases
**Pe
rcen
t ide
ntity
cal
cula
ted
usin
g nu
mbe
r of
bas
e pa
ir m
atch
es p
er a
lignm
ent i
n th
e N
CB
I B
last
pro
gram
(A
ltsch
ul, S
.F.,
et a
l.)
Tab
le 1
. Su
mm
ary
of 1
6S r
DN
A r
esul
ts.
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
----
---
24
LacZ
α-cc
dB
Kan
amyc
in
Am
pici
llin
pUC
ori
P lac
CA
CA
CA
GG
AA
A
CA
GC
TATG
A
CC
ATG
ATT
AC
G
CC
AA
GC
TCA
G
AA
TTA
AC
CC
TC
AC
TAA
AG
GG
TGTG
TCC
TT
TGTC
GA
TAC
TG
GTA
CTA
ATG
C
GG
TTC
GA
GT
CTT
AA
TTG
GG
A
GTG
ATT
TCC
GA
CTA
GTC
CT
GC
AG
GTT
TAA
A
CG
AA
TTC
GC
C
CTT
G
GG
C
GA
ATT
CG
CG
GC
TGA
TCA
GG
A
CG
TCC
AA
ATT
TG
CTT
AA
GC
G
GG
A
TT
CC
CG
C
TTA
AG
CG
CC
CC
GC
TAA
ATT
C
AA
TTC
GC
CC
TA
TAG
TGA
GT
CG
TATT
AC
AA
TT
CA
CTG
GC
C
GTC
GTT
TTA
CG
GC
GA
TTTA
A
GTT
AA
GC
GG
G
ATA
TCA
CTC
A
GC
ATA
ATG
TT
AA
GTG
AC
CG
G
CA
GC
AA
AA
TG
A A
PC
R
Pro
duct
T3
prim
ing
site
T7
prim
ing
site
Figu
re 5
. M
ap o
f pC
R®
4-
TOPO
® pl
asm
id, s
how
ing
nucl
eotid
e se
quen
ce su
rrou
ndin
g th
e cl
onin
g si
te w
ith b
ase
pair
num
bers
giv
en o
n th
e le
ft.
Plas
mid
con
tain
s 395
4 ba
se p
airs
w
ithou
t the
PC
R p
rodu
ct.
Ada
pted
fr
om In
vitro
gen
(200
3).
201
261
311
Eco
R I
Eco
R I
Pm
e I
Pst
I
LacZ
α in
itiat
ion
codo
n
21
1550 bp
1400 bp
Ladde
r
BK1BK2
BK6BK7
BK8BK10
BK1 1:10
BK2 1:10
BK6 1:10
BK7 1:10
BK8 1:10
BK10 1:
10
Water b
lank
Bacteri
al co
ntrol
Culture
blan
k
BK108
BK111BK111
1:10
BK108 1
:10
Figure 6. Gel of PCR product (27f, 1492r) showing BK clones from plasmid prep template at full concentration and 1:10 dilution. I used 10 µl Hi-Lo DNA Marker as my ladder, with 5 µl DNA template per well. Note size of band at ~1500 bp. Clones BK4, BK5, and BK9 were amplified during a later PCR run and are not shown.
22
TM
7 O
P10
Dei
noco
ccus
-The
rmus
Chl
orof
lexi T
herm
otoga
e
The
rmod
esul
foba
cter
ia
Aqu
ifica
e
Cyanobacteri
a
Actinobact
eria
OP9 Firmicutes
Spirochaetes
Planctomycetes
OP3 Chlamydiae Verrucomicrobia
Defe
rriba
cteres
Synerg
istes
Fusobacteria
TM
6 Ter
mite
grou
p I
Aci
doba
cter
ia
Fib
roba
cter
es
Gem
mim
onas
Nitr
ospi
ra C
hlor
obi
ε Pro
teob
acte
ria
Euca
rya
FCG2 Korarchaeota
FCG1
Thermoprotei C2
YNPFFA
C1
Sedim
ent-1
Halo
bacte
ria p
MC2
Met
hano
bact
eria
Met
hano
cocc
ales A
rcha
eogl
obi
Met
hano
pyru
s T
herm
ococ
ci
SA
GM
EG1
pM
C1
The
rmop
lasm
ata
Met
hano
mic
robi
a
0.10
nuc
leot
ide
chan
ges p
er si
te
Bac
teria
Arc
haea
Bac
tero
idet
es δ
Prot
eoba
cteria
α, β
, γ P
roteo
bacte
ria
Figu
re 7
. Ph
ylog
enet
ic tr
ee sh
owin
g th
e th
ree
dom
ains
of l
ife.
Hig
hlig
hted
ph
yla
deno
te c
lade
s in
the
Bac
teria
th
at c
onta
in B
K c
lone
s. C
andi
date
ph
yla
Term
ite G
roup
1 a
nd W
S6 n
ot
show
n. T
ree
adap
ted
from
Mac
alad
y an
d B
anfie
ld, 2
003.
23

Pseu
dom
onad
acea
e 5
6
Terr
edin
ibac
ter s
peci
es, Z
3165
8, 1
494
Hal
omon
adac
eae
24
Oce
anos
piril
lum
min
utul
um, 1
500
Oce
anos
piril
lace
ae 9
Flec
toba
cillu
s sp.
, U14
584,
147
0 M
arin
obac
ter h
ydro
carb
onoc
last
icus
, X67
022,
146
1 O
cean
ospi
rillu
m ja
poni
cum
, 149
8 SA
R86
7
Sym
bion
ts 3
acid
min
e dr
aina
ge c
lone
MIM
4138
, 148
4 C
oxie
lla b
urne
tii, M
2129
1, 1
471
Rhi
pice
phal
us sa
ngui
neus
sym
bion
t, D
8455
9, 1
460
acid
min
e dr
aina
ge c
lone
MIM
451,
148
5
Mor
axel
lace
ae 3
1
Fran
cise
llace
ae 7
Psiri
cket
tsia
Sym
bion
ts 7
Leuc
othr
ix m
ucor
, X87
277,
150
5 Th
ioth
rix 6
Legi
onel
lale
s 8
4
Xan
thom
onad
ales
13
GSg
m c
lone
EL5
8G
Sgm
clo
ne E
L9G
Sgm
clo
ne E
L2G
Sgm
clo
ne E
L54
GSg
m c
lone
EL7
0G
Sgm
clo
ne E
L8 GSg
m c
lone
EL4
2en
viro
nmen
tal c
lone
gro
up 8
PCB
8con
tam
inat
ed so
il cl
one
WD
284,
AJ2
9267
5, 1
489
rape
rhiz
osph
ere
clon
e w
r000
8, A
J295
469,
134
1 Th
iotri
chac
eae
5co
al ta
r8co
ntam
inat
ed g
roun
dwat
er c
lone
482
5, A
F351
230,
144
7 ac
id m
ine
drai
nage
clo
ne M
IM44
4, 1
467
"Thi
obac
illus
ferr
ooxi
dans
" stra
in m
81 (B
G),
1329
G
STM
clo
ne B
K1
SRan
g cl
one
1.33
, AF0
4761
9, 9
61SR
ang
clon
e 1.
28, A
F047
617,
963
GSg
m c
lone
EL5
GSg
m c
lone
EL7
7"T
hiob
acill
us b
areg
ensi
s", Y
0928
0, 1
405
SRan
g cl
one
2.5,
AF0
4762
4, 9
61C
ardi
obac
teria
les
3Fr
asas
si sn
ottit
e is
olat
e 1C
("Th
ioba
cillu
s sp.
"), A
F213
054,
629
Thio
baci
llus s
p., X
9753
4, 1
446
Hal
othi
obac
illus
kel
lyi,
AF1
7041
9, 1
494
Thio
baci
llus h
ydro
ther
mal
is, M
9066
2, 1
504
Met
hylo
cocc
ales
20
Cyc
locl
astic
us p
uget
ii, U
1262
4, 1
492
Cyc
locl
astic
us p
uget
ii, L
3495
5, 1
492
Thio
mic
rosp
ira 5
Met
hylo
phag
a 5
Sole
mya
reid
i sym
bion
t 2, L
2570
9, 1
407
Sym
bion
ts 8
Ach
rom
atiu
m o
xalif
erum
, L42
543,
147
9 M
ethy
loco
ccus
4aq
uacu
lture
pon
d cl
one
SF40
.049
, 149
8 sy
mbi
ont o
f Sol
emya
vel
um, M
9041
5, 1
459
Sole
mya
occ
iden
talis
gill
sym
bion
t, U
4104
9, 1
374
acid
min
e dr
aina
ge c
lone
MIM
486,
149
5 "T
hiob
acill
us p
rosp
erus
", un
publ
is, 1
419
Ecto
thio
rhod
ospi
ra sh
apos
hnik
ovi,
M59
151,
145
1 C
hrom
atia
ceae
6
Ecto
thio
rhod
ospi
race
ae 4
Nitr
osoc
occu
s oce
anus
, M96
395,
139
2 N
itros
ococ
cus o
cean
us ,
M96
398,
134
7
Figu
re 8
. Pa
rtial
AR
B n
eigh
bor-j
oini
ng tr
ee sh
owin
g re
lativ
e po
sitio
n of
clo
ne B
K1.
Tree
con
tain
s gam
ma-
prot
eoba
cter
ial s
eque
nces
and
is ro
oted
with
a m
onop
hyle
tic a
rcha
eal o
utgr
oup.
N
umbe
rs o
n tre
e ar
e G
enba
nk a
cces
sion
num
bers
for o
rgan
ism
s in
the
publ
ic d
ata
base
s.
0.10
subs
titut
ions
/site
25
Thiobacillus hydrothermalis
BK1 clone
Sulphur river clone 2.5
Beggiatoa alba
Acid mine drainage clone MIM444
"Thiobacillus ferooxidans"
Agrobacterium tumefaciens0.05 substitutions/site
100
100
96
95
Figure 9. Bayesian inference phylogram showing relative position of clone BK1. Tree rooted with Agrobacterium tumefaciens. Bootstrap values displayed as percentages of 1000 replications.
"Thiobacillus baregensis"
26
Des
ulfu
rom
onad
acea
e/Pe
loba
cter
11
"Geo
bact
er c
hape
lleii"
, U41
561,
146
5 Pe
loba
cter
pro
pion
icus
, X70
954,
154
9 G
STM
clo
ne B
K5
Geo
bact
er su
lfurr
educ
ens,
U13
928,
140
8 "G
eoba
cter
hyd
roge
noph
ilus"
, U46
860,
143
8 "G
eoba
cter
hyd
roge
noph
ilus"
, U28
173,
139
1 G
eoba
cter
met
allir
educ
ens,
L078
34, 1
537
sulfa
te re
duci
ng b
iore
acto
r clo
ne S
RB
AC
37, 1
477
GST
M c
lone
BK
10G
STM
clo
ne B
K4
GSg
m c
lone
EL3
GSg
m c
lone
EL6
2co
ntam
inat
ed a
quife
r clo
ne W
CH
B17
67, A
F050
536,
148
1 D
esul
fofu
stis
gly
colic
us, X
9970
7, 1
545
aqua
cultu
re p
ond
clon
e SF
39.4
58, 1
444
Des
ulfo
rhop
alus
sing
apor
ensi
s, A
F118
453,
149
8 aq
uacu
lture
pon
d cl
one
SF40
.063
, 150
4 D
esul
forh
opal
us v
acuo
latu
s, L4
2613
, 148
2 G
Sgm
clo
ne E
L28
Des
ulfo
bulb
us e
long
atus
, X95
180,
145
3 D
esul
fobu
lbus
rhab
dofo
rmis
, U12
253,
149
8 Sy
ntro
phac
eae
4G
Sgm
clo
ne E
L21
GSg
m c
lone
EL4
9D
esul
fom
onile
4D
esul
foar
culu
m b
aars
ii, M
3440
3, 1
355
Des
ulfo
arcu
lus s
p. B
G74
, U85
477,
151
5 G
Sgm
clo
ne E
L4D
esul
foba
cter
ium
ani
lini,
AJ2
3760
1, 1
527
Des
ulfo
bact
eriu
m sp
. mX
yS1,
AJ0
0685
3, 1
528
GSg
m c
lone
EL3
8ho
t spr
ing
clon
e O
PB16
, AF0
2699
8, 1
490
hot s
prin
g cl
one
OPB
55, A
F026
993,
149
5 D
esul
foba
cter
hal
otol
eran
s, Y
1474
5, 1
472
Des
ulfo
bact
er sp
ecie
s, M
3441
6, 1
436
Des
ulfo
bact
er sp
ecie
s, M
3441
5, 1
373
Des
ulfo
bact
er h
ydro
geno
philu
s, M
3441
2, 1
358
Des
ulfo
bact
er c
urva
tus,
M34
413,
134
4 D
esul
foba
cter
vib
riofo
rmis
, U12
254,
151
5 D
esul
foba
cter
pos
tgat
ei, M
2663
3, 1
499
delta
pro
teob
acte
rium
WN
, U51
844,
137
2 D
esul
foba
cula
tolu
olic
a, X
7095
3, 1
550
"Des
ulfo
bact
eriu
m n
iaci
ni",
U51
845,
142
7 "D
esul
foba
cter
ium
vac
uola
tum
", M
3440
8, 1
345
Des
ulfo
bact
eriu
m a
utot
roph
icum
, M34
409,
131
9 G
STM
clo
ne B
K7
Des
ulfo
cocc
us m
ultiv
oran
s, M
3440
5, 1
347
Des
ulfo
nem
a lim
icol
a, U
4599
0, 1
496
GSg
m c
lone
EL2
2D
esul
fosa
rcin
a va
riabi
lis, M
2663
2, 1
499
Des
ulfo
sarc
ina
varia
bilis
, M
3440
7, 1
504
GSg
m c
lone
EL7
5D
esul
fofa
ba g
elid
a, A
F099
063,
148
4 D
esul
fobo
tulu
s sap
ovor
ans,
M34
402,
133
9 D
esul
fone
ma
ishi
mot
oei,
U45
991,
144
0 D
esul
fone
ma
ishi
mot
oei,
U45
992,
144
0 D
esul
fobo
tulu
s sp.
BG
14, U
8547
0, 1
500
Des
ulfo
cella
hal
ophi
la, A
F022
936,
154
4 G
STM
clo
ne B
K9
Des
ulfo
virg
a ad
ipic
a, A
J237
605,
152
4 D
esul
forh
abdu
s am
nige
nus,
X83
274,
151
5 un
iden
tifie
d su
lfate
7red
ucin
g ba
cter
ium
, U49
429,
151
8 U
nkno
wn,
X82
875,
147
5 U
nkno
wn,
X82
874,
148
6 Sy
ntro
phob
acte
r wol
inii,
X70
905,
142
4 D
esul
foac
inum
infe
rnum
, L27
426,
152
6 ho
t spr
ing
clon
e O
PB33
, AF0
2698
7, 1
486
Ther
mod
esul
forh
abdu
s nor
vegi
cus,
U25
627,
155
9 sl
udge
clo
ne H
16, A
F234
748,
155
5 ac
id m
ine
drai
nage
clo
ne M
IM41
8, 1
520
Myx
ococ
cale
s 1
2
Bde
llovi
brio
nale
s 2
TM6
9
Aci
doba
cter
ia 8
9
acid
min
e dr
aina
ge c
lone
MIM
4123
, 151
6 ac
id m
ine
drai
nage
clo
ne M
IM44
0, 1
533
Des
ulfo
vibr
iona
les
44
slud
ge c
lone
195
9, A
F097
803,
147
9 la
b7sc
ale
slud
ge c
lone
SB
R10
93, A
F269
002,
148
5 co
ntam
inat
ed a
quife
r clo
ne W
CH
B17
27, A
F050
538,
148
5
delta
-pro
teob
acte
ria
Figu
re 1
0. P
artia
l AR
B n
eigh
bor-j
oini
ng tr
ee sh
owin
g re
lativ
e po
sitio
ns o
f clo
nes B
K4,
BK
5, B
K7,
BK
9, a
nd
BK
10 a
s mem
bers
of t
he d
elta
-pro
teob
acte
ria.
Tree
root
ed w
ith m
onop
hyle
tic a
rcha
eal o
utgr
oup.
Num
bers
show
n ar
e G
enba
nk a
cces
sion
num
bers
for o
rgan
ism
s in
the
publ
ic d
atab
ases
.
0.10
subs
titut
ions
/site
27

Desulfobacter postgatei
Syntrophobacter wolinii
BK9 clone
Desulfosarcina sp.
Desulforhopalus singaporensis
BK4 clone
BK10 clone
Desulfomonile tiedjei
Nitrospina gracilis
Desulfobacterium anilini
Geobacter metaloreducens
Geobacter sulfurreducens
BK5 clone
Desulfonema ishimotoei
BK7 clone
0.05 substitutions/site
Figure 11. Bayesian inference phylogram showing relative positions of clones BK4, BK5, BK7, BK9, and BK10. Tree rooted with Desulfobacter postgatei. Bootstrap values displayed as percentages of 100 replications.
100
100
100
100
94
59
78
100
94
100
51
90
28
Bac
tero
ides
, Pre
vote
lla 6
9
Porp
hyro
mon
adac
eae
13
Bac
tero
ides
dis
taso
nis,
M25
249
M, 1
511
Bac
tero
ides
mer
dae,
X83
954,
136
6 su
lfate
redu
cing
bio
reac
tor c
lone
SR
BA
C29
, 144
9 B
acte
roid
es fo
rsyt
hus,
L164
95, 1
455
Bac
tero
ides
fors
ythu
s, X
7396
2, 1
450
CD
C G
roup
DF.
3, U
4135
5, 1
467
sulfa
te re
duci
ng b
iore
acto
r clo
ne S
RB
AC
55, 1
446
Bac
tero
ides
sp. s
p4, A
B00
3389
, 143
2 M
arin
ilabi
lia a
garo
vora
ns, M
6242
2, 1
454
Cyt
opha
ga fe
rmen
tans
, M58
766,
146
3 B
acte
roid
es sp
lanc
hnic
us, L
1649
6, 1
458
G
STM
clo
ne B
K2
7co
ntam
inat
ed a
quife
r clo
ne W
CH
B1.
32, A
F050
543,
144
0 SR
ang
clon
e 1.
29, A
F047
618,
944
GST
M c
lone
108
T3C
essp
ool c
lone
CC
.8, A
F207
533,
626
"Ana
erof
lexu
s mar
itim
us",
1386
co
ntam
inat
ed a
quife
r clo
ne W
CH
B1.
53, A
F050
539,
144
8
anae
robi
c di
gest
or c
lone
vad
inH
A17
, U81
712,
145
2 G
Sgm
clo
ne E
L26
cont
amin
ated
aqu
ifer c
lone
WC
HB
1.69
, AF0
5056
7, 1
445
GSg
m c
lone
EL2
0an
aero
bic
dige
stor
clo
ne B
C27
, U81
676,
144
8
sulfa
te re
duci
ng b
iore
acto
r clo
ne S
RB
AC
60, 1
446
sulfa
te re
duci
ng b
iore
acto
r clo
ne S
RB
AC
32, 1
446
cont
amin
ated
aqu
ifer c
lone
WC
HB
1.29
, AF0
5054
4, 1
446
Bac
tero
ides
put
redi
nis,
L164
97, 1
448
R
iken
ella
mic
rofu
sus,
L164
98, 1
463
Flav
obac
teria
les
64
Flex
ibac
tera
ceae
9
Sphi
ngob
acte
riace
ae 9
cool
ing
coil
colo
nise
r iso
late
PC
5.1A
, 138
1 Sp
iroso
ma
lingu
ale,
M27
802
M, 1
423
Flex
ibac
ter e
lega
ns ,
1432
Fl
exib
acte
r ele
gans
, M
5878
3, 1
432
Mic
rosc
illa
mar
ina,
M58
793,
142
5 Fl
exib
acte
r rub
er, M
5878
8, 1
450
aero
bic
slud
ge c
lone
PH
OS.
HE2
1, A
F314
419,
141
7 R
unel
la sl
ithyf
orm
is, M
2779
9 M
, 144
4 Fl
exib
acte
r lito
ralis
, M58
784,
147
0 Fl
exib
acte
r pol
ymor
phus
, M58
786,
144
9 Fl
exib
acte
r ros
eolu
s, M
5878
7, 1
439
Flex
ibac
ter a
ggre
gans
Pers
icob
acte
r diff
luen
s, M
5876
5, 1
463
envi
ronm
enta
l iso
late
470
77, A
F227
830,
140
7 ho
t spr
ing
clon
e O
PB73
, AF0
2700
8, 1
449
hot s
prin
g cl
one
env.
OPS
17, A
F018
199,
143
7 Th
erm
onem
a la
psum
, L11
703,
149
0 Sa
pros
pira
ceae
11
Rie
mer
ella
ana
tipes
tifer
, U10
877,
150
4 R
hodo
ther
mus ho
t spr
ing
clon
e O
PB88
, AF0
2700
6, 1
433
Chl
orob
i 1
4
Epsi
lon
prot
eoba
cter
ia 1
28
Des
ulfu
rella
les
2
CFB
Figu
re 1
2. P
artia
l AR
B n
eigh
bor-j
oini
ng tr
ee sh
owin
g re
lativ
e po
sitio
ns o
f clo
nes B
K2
and
par
tial B
K10
8T3
as
mem
bers
of t
he C
ytop
haga
/Fla
voba
cter
ia/B
acte
roid
es.
Tree
root
ed w
ith m
onop
hyle
tic a
rcha
eal o
utgr
oup.
N
umbe
rs sh
own
are
Gen
bank
acc
essi
on n
umbe
rs fo
r org
anis
ms i
n th
e pu
blic
dat
abas
es.
0.01
subs
titut
ions
/site
29
Bacteroides forsythus
Bacteroides distasonis
sulfate reducing bioreactor clone SRBAC29
sulfate reducing bioreactor clone SRBAC5
contaminated aquifer clone WCHB1
Rikanella microfusus
sulfate reducing bioreactor clone SRBAC32
BK2 clone
Cytophaga fermentans
Cytophaga lytica
Flavobacterium saccharophilum
0.05 substitutions/site
100
64
100
85
100
100
58
100
Figure 13. Bayesian inference phylogram showing relative position of clone BK2. Tree rooted with Cytophaga lytica and Flavobacterium saccharophilum as monophyletic sister clade. Bootstrap values displayed as percentages of 1000 replications.
30
sulfa
te re
duci
ng b
iore
acto
r clo
ne S
RB
AC
17, 1
392
co
ntam
inat
ed a
quife
r clo
ne W
CH
B1-
64, A
F050
606,
141
4 su
lfate
redu
cing
bio
reac
tor c
lone
SR
BA
C27
, 140
0 co
ntam
inat
ed a
quife
r clo
ne W
CH
B1-
07, A
F050
600,
151
2 co
ntam
inat
ed a
quife
r clo
ne W
CH
B1-
26, A
F050
599,
141
7 ac
id m
ine
drai
nage
clo
ne M
IM45
0, 1
425
hot s
prin
g cl
one
OPB
92, A
F027
030,
143
8 co
ntam
inat
ed a
quife
r clo
ne W
CH
B1-
11, A
F050
603,
142
3
cont
amin
ated
aqu
ifer c
lone
WC
HB
1-56
, AF0
5060
5, 1
517
tere
phth
alat
e-de
grad
ing
cons
ortiu
m c
lone
TA
18, A
F229
791,
131
8 ac
id m
ine
drai
nage
clo
ne M
IM49
0, 1
378
aqua
cultu
re p
ond
clon
e SF
39.3
80, 1
376
OP1
1-5
cont
amin
ated
aqu
ifer c
lone
WC
HB
1-01
, AF0
5059
7, 1
472
cont
amin
ated
aqu
ifer c
lone
WC
HB
1-15
, AF0
5059
6, 1
474
cont
amin
ated
aqu
ifer c
lone
WC
HB
1-06
, AF0
5059
5, 1
470
cont
amin
ated
aqu
ifer c
lone
CA
1-11
5, A
F172
911,
971
un
cultu
red
bact
eriu
m B
OL-
01, A
F172
883,
974
co
ntam
inat
ed a
quife
r clo
ne C
A1-
02, A
F172
904,
971
co
ntam
inat
ed a
quife
r clo
ne C
A1-
03, A
F172
905,
105
1 m
arin
e se
dim
ent c
lone
BM
S1-2
8, A
F172
878,
104
6 G
STM
clo
ne B
K6
sedi
men
t clo
ne B
OL-
130,
AF1
7289
6, 9
68
uncu
lture
d ba
cter
ial c
lone
D1-
11, A
F172
882,
964
la
b-sc
ale
slud
ge c
lone
SB
R10
71, A
F268
996,
142
0 la
b-sc
ale
slud
ge c
lone
SB
R20
04, A
F268
997,
139
4 de
ep-s
ea se
dim
ent c
lone
BD
4-8,
AB
0155
58, 1
568
met
al-c
onta
min
ated
soil
clon
e K
20-2
7, A
F145
827,
151
5 so
il cl
one
C1-
29, 1
433
soil
clon
e S1
-197
, 142
9 la
b-sc
ale
slud
ge c
lone
SB
R20
13, A
F269
000,
142
0 so
il cl
one
C1-
105,
148
4 m
etal
-con
tam
inat
ed so
il cl
one
K20
-12,
AF1
4581
5, 1
484
full-
scal
e sl
udge
clo
ne G
C1,
AF2
6899
4, 1
381
lab-
scal
e sl
udge
clo
ne S
BR
2060
, AF2
6899
9, 1
429
oral
cav
ity c
lone
BE1
09, A
Y00
5446
, 144
3 or
al c
avity
clo
ne B
S003
, AY
0054
48, 1
448
arid
soil
clon
e C
0-26
, AF1
2871
7, 1
437
lab-
scal
e sl
udge
clo
ne S
BR
2096
, AF2
6899
8, 1
487
or
al c
avity
clo
ne F
061,
AF1
2520
5, 1
449
lab-
scal
e sl
udge
clo
ne S
BR
2113
, AF2
6900
1, 1
395
oral
cav
ity c
lone
I025
, AF1
2520
6, 1
413
past
ure
soil
clon
e EB
1087
, 134
4 G
Sgm
clo
ne E
L59
petro
leum
-con
tam
inat
ed g
roun
dwat
er c
lone
102
5, A
B03
0611
, 138
4 co
ntam
inat
ed a
quife
r clo
ne W
CH
B1-
60, W
S5, A
F050
598,
176
9 SR
ang
clon
e 2.
3, A
F047
623,
930
trick
ling
filte
r bio
film
clo
ne k
oll6
, AJ2
2453
9, 1
423
OP1
1
WS6
TM7
Figu
re 1
4. P
artia
l AR
B n
eigh
bor-j
oini
ng tr
ee sh
owin
g re
lativ
e po
sitio
n of
clo
ne B
K6
as m
embe
r of t
he W
S6.
Tree
ro
oted
with
mon
ophy
letic
arc
haea
l out
grou
p. N
umbe
rs sh
own
are
Gen
bank
acc
essi
on n
umbe
rs fo
r org
anis
ms i
n th
e pu
blic
dat
abas
es.
0.01
subs
titut
ions
/site
31
Cam
pylo
bact
er je
juni
Des
ulfu
rella
ace
tivor
ans
hot s
prin
g cl
one
OPB
92
BK6
clon
e
cont
amin
ated
aqu
ifer c
lone
CA
1-03
mar
ine
sedi
men
t clo
ne B
MS1
-28
uncu
lture
d ba
cter
ial c
lone
D1-
11
Aci
d m
ine
drai
nage
clo
ne M
IM49
0
deep
-sea
sedi
men
t clo
ne B
D7-
4
CL5
00-1
3 cl
one
trick
ling
filte
r bio
film
clo
ne k
oll6
full-
scal
e sl
udge
clo
ne G
C1
soil
clon
e S1
-197
lab-
scal
e sl
udge
clo
ne S
BR
2004
met
al-c
onta
min
ated
clo
ne K
20-2
7
soil
clon
e C
1-10
5
oral
cav
ity c
lone
F06
1
oral
cav
ity c
lone
BE1
09
arid
soil
clon
e C
0-26
lab-
scal
e sl
udge
clo
ne S
BR
2113
past
ure
soil
clon
e EB
1087
petro
leum
-con
tam
inat
ed g
roun
dwat
er c
lone
1025
BK8T
3 cl
one
0.05
subs
titut
ions
/site
Fig
ure
15.
Bay
esia
n in
fere
nce
phyl
ogra
m sh
owin
g re
lativ
e po
sitio
ns o
f clo
nes B
K6
and
BK
8T3.
Tre
e ro
oted
with
C
ampy
loba
cter
jeju
ni a
nd D
esul
fure
lla a
cetiv
oran
s as m
onop
hyle
tic si
ster
cla
de.
Boo
tstra
p va
lues
dis
play
ed a
s pe
rcen
tage
s of 1
000
repl
icat
ions
.
90
84
92
10010
089
71
100 95
9287
83
95
52
7873
72
85
32

term
ite g
ut c
lone
TG
14, A
B00
4576
, 134
2 te
rmite
gut
clo
ne T
G33
, AB
0045
78, 1
342
term
ite g
ut c
lone
TG
25, A
B00
4577
, 134
1 te
rmite
gut
clo
ne T
G13
, AB
0045
75, 1
341
GST
M c
lone
BK
8T7
rum
en c
lone
4C
0d$3
, AB
0340
17, 1
505
anae
robi
c sl
udge
clo
ne v
adin
HB
76, U
8175
4, 1
446
lab$
scal
e sl
udge
clo
ne S
BR
2095
, AF2
6900
3, 1
453
OP5
3ho
t spr
ing
clon
e O
PB25
, OP2
, AF0
2709
7, 1
478
Figu
re 1
6. P
artia
l AR
B n
eigh
bor-j
oini
ng tr
ee sh
owin
g re
lativ
e po
sitio
n of
par
tial c
lone
BK
8T7
as m
embe
r of t
he
Term
ite G
roup
1 p
hylu
m.
Num
bers
show
n ar
e G
enba
nk a
cces
sion
num
bers
for o
rgan
ism
s in
the
publ
ic d
atab
ases
.
0.01
subs
titut
ions
/site
33