molecular mechanisms of pediatric kidney disease nader gordjani md phd professor of pediatrics...
TRANSCRIPT

Molecular Mechanisms of Pediatric Kidney Disease
Nader Gordjani MD PhDProfessor of Pediatrics
Universities of Freiburg and Frankfurt

Molecular origins of renal diseases
H2O
Hypophasphatemic rickets
Bartter-Syndrome
Hypokalemic alkalosis:
Gitelman-S.
Diabetes insipidus renalis
NEPHROTIC SYNDROME ?
CYSTÎC KIDNEYS ?

kidney Glomerular structure
Protein loss

Lesions sites of the glomerulus
Filtration barrier
TH-Lypmphocytes
Capillaries
Mesangial cells
s. Biopsie
Deposits of antibodiesand complement
Podocyte
Autoantibodies
Nephrotic Syndrome
Nephritic Syndrome
kidney

Proteinuria > 40 mg/m2 * h
> 1000 mg/m2 * 24 h
Plasma Albumin < 25 g/l
Nephrotic Syndrome in Children
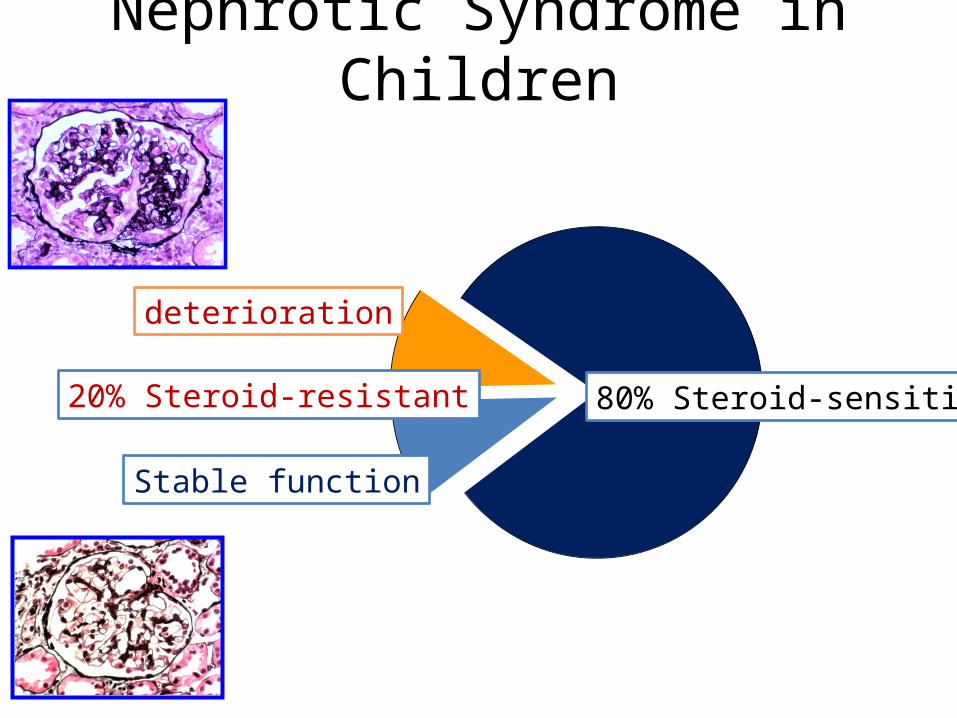
80% Steroid-sensitive
Stable function
20% Steroid-resistant
deterioration
Nephrotic Syndrome in Children

Somlo S Nat Genet 24: 333 (2000)
The glomerulus in health & NS

Somlo S Nat Genet 24: 333 (2000)
Morphology of the Filtration Barriere
normal
Nephroc syndrome

Geheimnisse des „Podozyten“ Molecular structures of the Podocyte
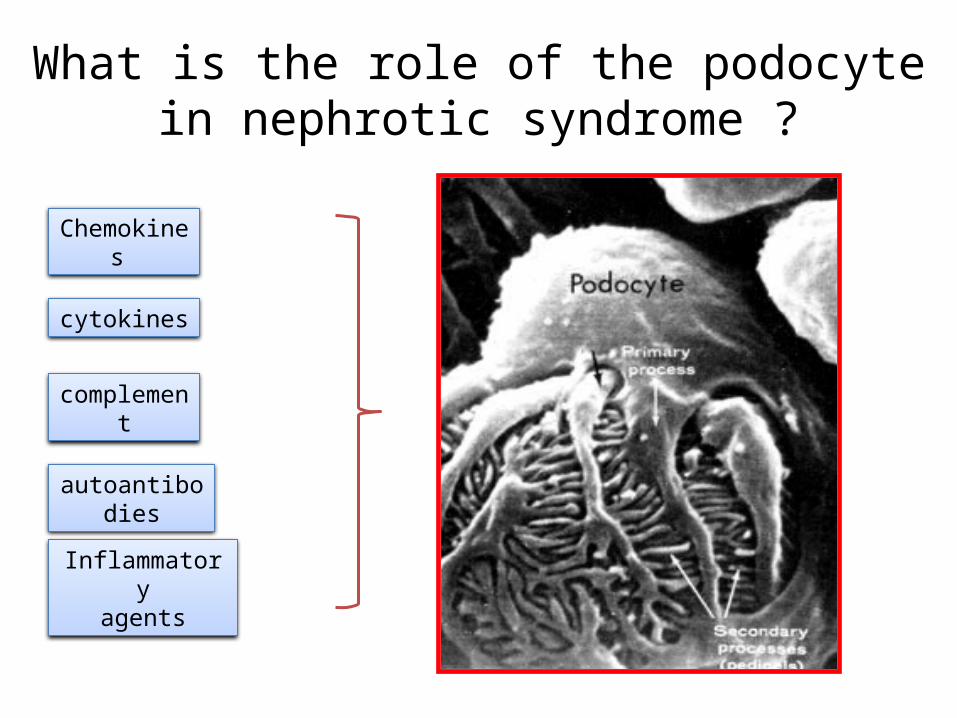
What is the role of the podocyte in nephrotic syndrome ?
Chemokines
cytokines
complement
autoantibodies
Inflammatoryagents

Das kongenitale NS vom finnischen Typ Nephrin
Congenital nephrotic syndrome Finnish Type

Familiäres Steroid-resistentes NS Podocin
Familial Steroid resistant Nephrotic Syndrome

Die familiäre Glomerulosklerose
a-act 4
Familial Glomerulosclerosis

What is the role of the podocyte in nephrotic syndrome and as target of immunmodulation?
Chemokines
cytokines
complement
autoantibodies
Inflammatoryagents
Imm
unos
uppr
essi
on

Ca2+- Signalling in the cell
Stimulation of:
Signalling pathwaysGene expressionChannel opening/closing …
0.0
1.0
2.0
3.0
fluo
resc
en
ce r
atio
34
0/3
80
ATP 10-4 M
C a2+ 10-6 M
3 m in

Calcium signalling in the Podocyte
• Fluorescence microscopy with Fura-2 AM Single-Photon TubeVideo-ImagingLaser-Scanning
RT-PCR• Isolation of rat glomeruli by sieving method• Statistical analysis: student´s t-test

Mouse Podocytes in Culture
33o C 37o C

CCR
3
CCR
4
CCR
1
CCR
2
CCR
5
GAP
DH
334
605
409
563
410
334
500bp
CC-receptors of cultured mouse podocytes
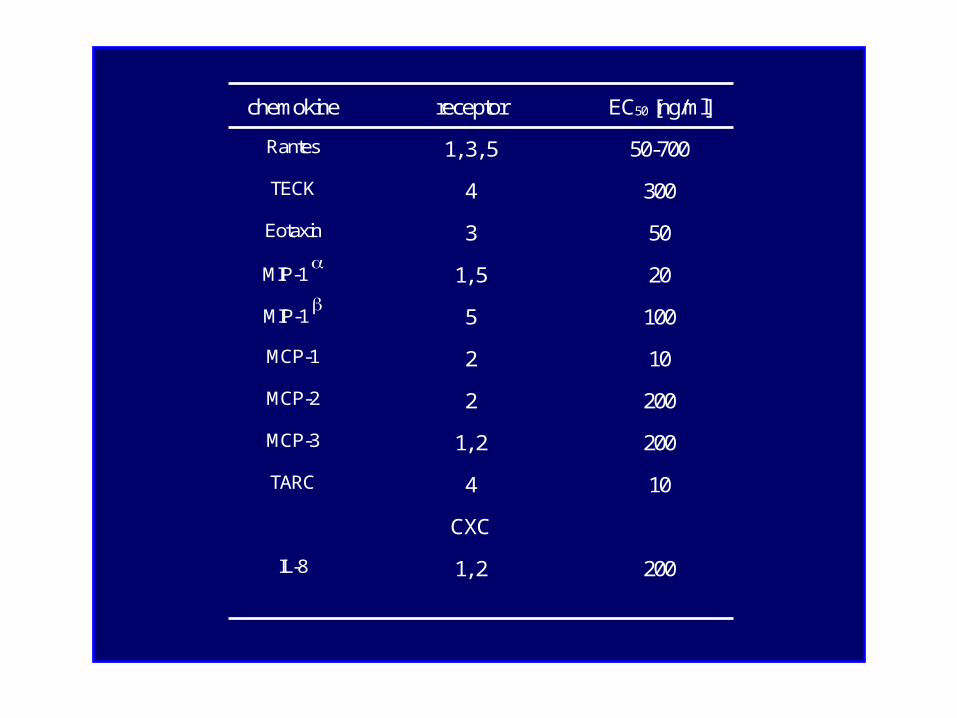
chemokine receptor EC50 [ng/ml]
Rantes 1, 3, 5 50-700
TECK 4 300
Eotaxin 3 50
MIP-1 1, 5 20
MIP-1 5 100
MCP-1 2 10
MCP-2 2 200
MCP-3 1, 2 200
TARC 4 10
CXC
IL-8 1, 2 200

0.0
1.0
2.0
3.0flu
ores
cenc
e ra
tio 3
40/3
80
ATP 10-4 M
C a2+ 10-6 M
3 m in

0.0
0.5
1.0
1.5
2.0flu
ore
scen
ce r
atio
34
0/3
80
ATP 100 µm ol/l
R AN TES 0.4 µg/m l
M C P-1 0.2 µg/m l
M IP-1 60 ng/m l
5 m in

0.0
0.2
0.4
0.6
0.8
1.0flu
ores
cenc
e ra
tio 3
40 /
380
nm
400 700 1000
RANTES
ng / m l
(n=39) (n=11) (n=4)

0.0
0.1
0.2
0.3
0.4flu
ore
sce
nce
ratio
34
0 / 3
80
nm
M C P 1 M IP 1 Eotaxin IL - 8-

R AN TES 0.7µg/m l
ATP 10-4M0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5flu
ores
cenc
e ra
tio 3
40/3
80
C a2+ 10-6 M
3 m in

0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
f
luor
esce
nce
rat
io 3
40/3
80 n
m
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
f
luor
esc
ence
rat
io 3
40/
380
nm
(n=5) (n=8)
CyA
ATP A II
CyA

(n=6)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
flu
ore
sce
nce
ra
tio 3
40
/38
0 n
m
RANTES 700 ng/m l
CyA 10 -5 M

Freshly isolated rat glomerulum
25 µm 25 µm
decapsulated
25 µm 25 µm
decapsulated

Immunofluorescence staining for podosynapsinin freshly isolated rat glomerulum
25 µm

Immunofluorescence staining for WT-1in freshly isolated rat glomerulum

Freshly isolated rat glomerulum
0
0.5
1
1.5
2
2.5
3
3.5
4flu
ores
cenc
e ra
tio 3
45/3
80 n
m
AC H 1 µm ol/l
M C P-1 20 ng/m l
10 m in
C YA 10 nm ol/l
R AN TES 70 ng/m l

ConclusionsChemokines induce an increase of [Ca2+]i in mouse podocytes by release from cytosolic Ca2+-stores and thus stimulate Ca2+-mediated signal transduction.
Therefore podocytes are target cells of these proinflammatory factors.
ATP also causes a characteristic rise of [Ca2+]i, which is mediated by Ca2+-influx from the extracellular space in addition to store release.
Cyclosporine A did not influence the chemokine- or ATP-associated Ca2+-effects.
These findings could partly be confirmed in podocytes from freshly isolated intact rat glomeruli.

Physiology
Andreas BenesicRuth FreudingerMichael GekleGerald Schwerdt
Pediatrics
Nader GordjaniAntje KirchhoffBrigitte Wollny
University of Würzburg
University of Freiburg
Physiologie
Rainer GregerHermann PavenstädtJens LeipzigerRoland NitschkeViktoria Munzinger

Rainer Greger1946 - 2004
Institute of Physiology/Freiburg

What causes Cyst formation in kidneys?

… and damage in other organs?
M. Waters et al. Pediatr Nephrol (2011) 6:1039–1056

… and other organs?
M. Waters et al. Pediatr Nephrol (2011) 6:1039–1056

Hildebrandt F et al. JASN 2009;20:23-35
Juvenile/infantile Nephronophthisis

Hildebrandt F et al. JASN 2009;20:23-35
Ciliar defects: a unifying theory of cystic kidney disease?

Ciliar defects: a unifying theory of cystic kidney disease?
Lin et al. PNAS 2002

Subcellular localization of nephrocystins
Hildebrandt F et al. JASN 2009;20:23-35
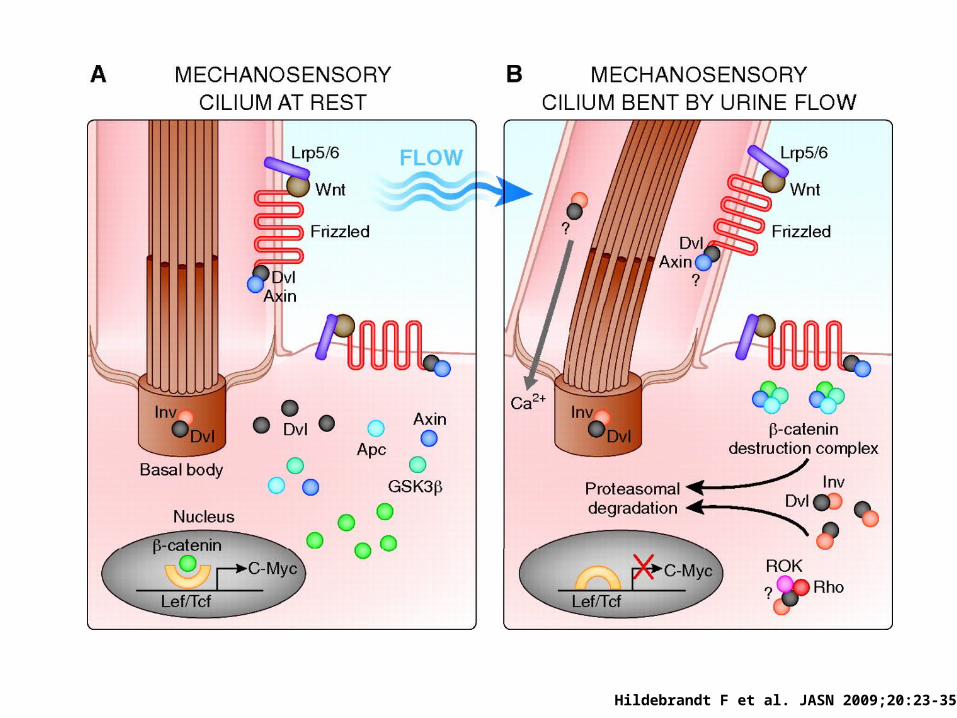
Hildebrandt F et al. JASN 2009;20:23-35

Chapin et al. JCB 2010
Ciliar defects: a unifying theory of cystic kidney disease?

Hildebrandt F et al. JASN 2009;20:23-35
Correct mitotic spindle
orientation
False mitotic spindle
orientation

Hildebrandt F et al. JASN 2009;20:23-35
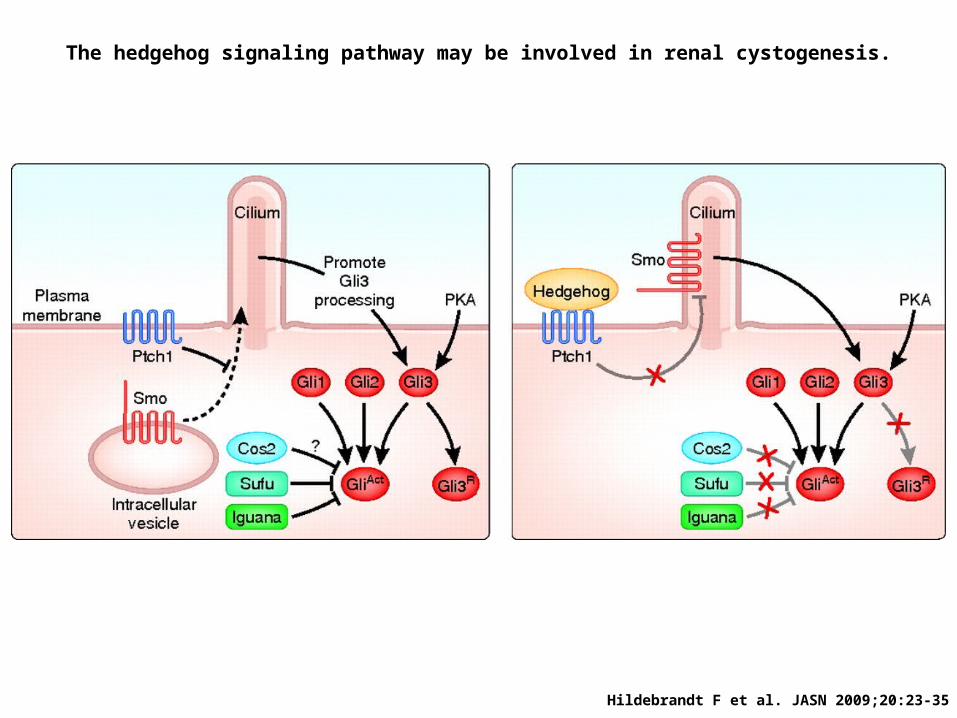
The hedgehog signaling pathway may be involved in renal cystogenesis.
Hildebrandt F et al. JASN 2009;20:23-35