molecular genetic characterisation of mirafiori lettuce ...€¦ · rocky balboa, 2006 . iv . v...
TRANSCRIPT

i
Molecular genetic characterisation of Mirafiori lettuce
big-vein virus and Lettuce big-vein associated virus and
their vector Olpidium virulentus associated with
Lettuce Big-Vein Disease and the determination of
their inoculum sources in Western Australia.
This thesis is presented for the degree of Doctor of Philosophy of The
University of Western Australia
School of Plant Biology
Plant Pathology
The University of Western Australia
December, 2009
LINDA DIANNE MACCARONE
B. Sc (Cons. Biol.) (Murdoch University)
Hons (Biol. Sc.) (Murdoch University)

ii

iii
“... it ain‟t how hard you hit; it‟s about how hard you can get hit
and keep moving forward...”
Rocky Balboa, 2006

iv

v
Abstract
Lettuce Big-Vein Disease (LBVD) is a common chytrid- and soil-borne virus disease
that affects lettuce in Western Australia. It causes chlorophyll clearing of the veins of
the lettuce leaf, leaf crinkling and decreased head size, which together result in
decreased yield and quality of lettuce. The phylogenetic relationships of Lettuce big-
vein associated virus (LBVaV) and Mirafiori lettuce big-vein virus (MLBVV) and their
vector Olpidium virulentus, associated with LBVD in Western Australia were
established. Sources of inoculum in commercial lettuce seedling nurseries and
commercial lettuce growing fields in south-western Australia were identified with an
aim of improving strategies to manage the disease.
Lettuce plants showing symptoms of lettuce big-vein disease were collected from the
Perth Metropolitan region of south-west Australia and DNA extraction was done on
their roots. When PCR primers designed specifically in this study were used to amplify
the rDNA ITS region of O. brassicae and O. virulentus in extracts, only O. virulentus
was detected. Phylogenetic analysis of the complete rDNA-ITS region sequences of the
five Australian isolates and 10 others was done. The Australian isolates fitted within
two clades of O. virulentus (I and II), and within clade I, into two of its four sub-clades
(Ia and Id). LBVaV and MLBVV were both detected when symptomatic lettuce leaf
tissue samples corresponding to the root samples of these plants were tested using virus-
specific primers in Reverse Transcription-PCR. The presence of both viruses was found
to be associated with O. virulentus occurrence.

vi
MLBVV, genus Ophiovirus, was detected in samples of big-vein diseased lettuce plants
collected from seven different farms in south-west Australia. The complete coat protein
(CP) gene encoding sequences of 13 isolates were obtained from these samples.
Phylogenetic analysis of these samples and others from five continents revealed that
within each of the two major MLBVV clades (A and B) there were two sub-clades.
Sub-clade A1 contained 13 isolates from Australia, six from Europe and one from
Japan; A2 comprised one isolate from Australia and six from Europe; while B1 and B2
comprised two and five European isolates, respectively, all from Spain. The presence of
Australian isolates in two separate sub-clades suggests that there have apparently been
at least two separate introductions of MLBVV to the isolated Australian continent.
LBVaV, genus Varicosavirus, was detected in leaf samples of LBVD affected plants
collected from four different lettuce growing properties in south-west Australia. The
complete CP gene encoding sequence of seven isolates was obtained from these
samples. Phylogenetic analysis of these isolates and others from five continents, and
Tobacco stunt virus (TStV) isolates from Japan, revealed two Clades, I and II. Clade I
contained isolates only from Europe whereas Clade II contained both LBVaV and TStV
isolates from Australia and Japan and formed three distinct Sub-clades. Sub-clade IIa
contained Australian LBVaV isolates, Sub-clade IIb contained Japanese LBVaV
isolates and Sub-clade IIc contained all of the five TStV isolates which were all from
Japan. Due to the high level of genome conservation indicated with this study, further
evidence is given to support TStV as a tobacco strain of LBVaV rather than a distinct
species in the genus Varicosavirus.

vii
As previous studies have only determined the presence of LBVaV in lettuce seedling
nurseries and in commercial lettuce growing fields, this study also confirmed the
presence of LBVaV and, additionally, the presence of MLBVV in both situations by
using RT-PCR and DAS-ELISA. Lettuce seed coat contamination with viruliferous O.
virulentus resting spores was investigated as a potential source of inoculum. O.
virulentus, LBVaV and MLBVV were all detected by PCR on the lettuce seed coat.
Seeds harvested from lettuce plants that had previously tested positive for LBVaV and
MLBVV were either surfaced sterilised with sodium hypochlorite or trisodium
phosphate, with both, or left unsurfaced sterilised and sown into disinfested potting mix.
None of the experimental plants tested positive to LBVaV or MLBVV by DAS-ELISA
which suggests that either the propagule numbers were too low to cause systemic
infection or the detection method by DAS-ELISA was not sufficiently sensitive as it had
been shown by RT-PCR that all pathogens associated with LBVD could be detected on
lettuce seed coats.
In a study of alternative hosts of LBVaV and MLBVV, two known hosts Sonchus
oleraceous and S. asper as well as three unreported hosts S. hydrophilus, Reichardia
sp. and Actites sp. were used as bait plants grown in infested LBVD soil under
experimental conditions to determine if they were susceptible to both viruses. All
species were found to be susceptible to LBVaV and MLBVV and, additionally, one
Arctotheca calendula plant collected from a commercial lettuce growing field tested
positive for MLBVV. Of the species tested, there was a higher incidence of LBVaV
and/or MLBVV detected in the root tissue of the experimental plants than in the leaf
tissue of the same plants. S. asper was determined as the most susceptible of the species
tested for LBVaV and MLBVV.

viii
Additional knowledge regarding the genetic characteristics of O. virulentus, LBVaV
and MLBVV, as well as more information regarding disease epidemiology such as
sources of inoculum of the organisms associated with LBVD, either by vector-assisted
seed transmission or by alternative hosts possibly acting as disease reservoirs as
determined in this study, is expected to help in establishing more effective methods of
control for the disease as well as enhancing the current Integrated Disease Management
Strategy which has already been established for the disease.

ix
Table of Contents
Abstract ______________________________________________________________ v
Table of Contents ______________________________________________________ ix
Acknowledgements ____________________________________________________xiii
Candidate’s Declaration _______________________________________________ xiv
List of Abbreviations __________________________________________________ xv
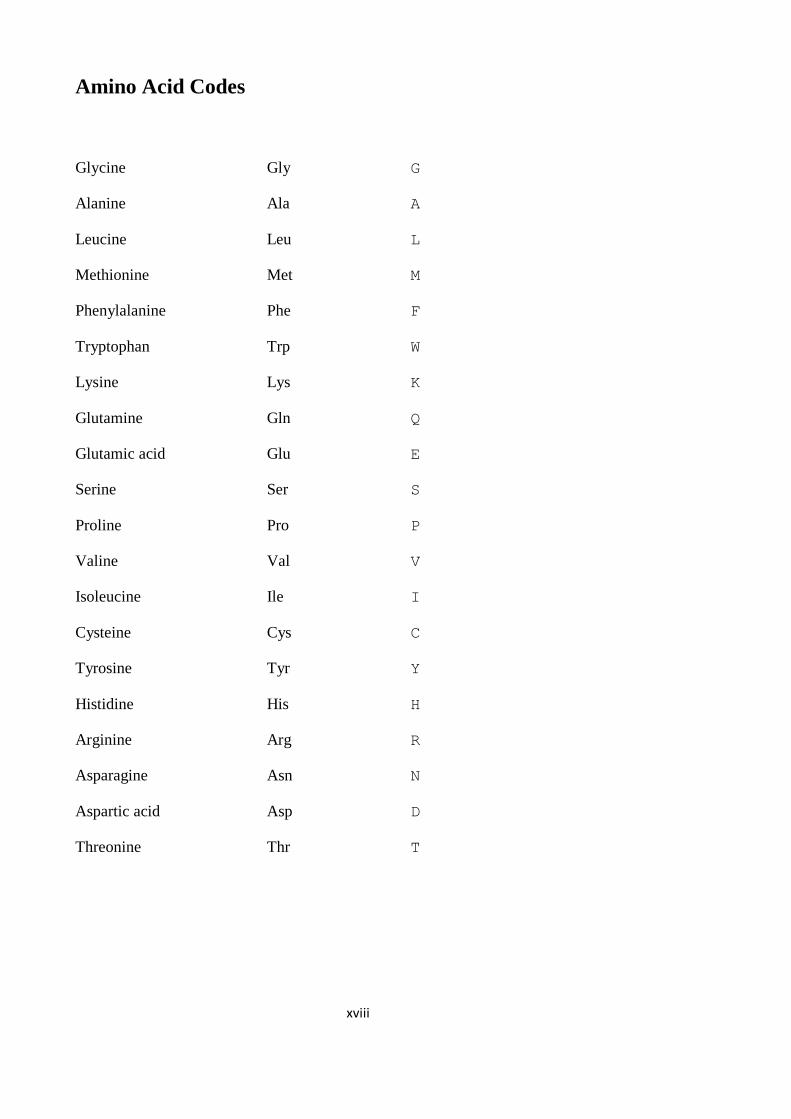
Amino Acid Codes ___________________________________________________ xviii
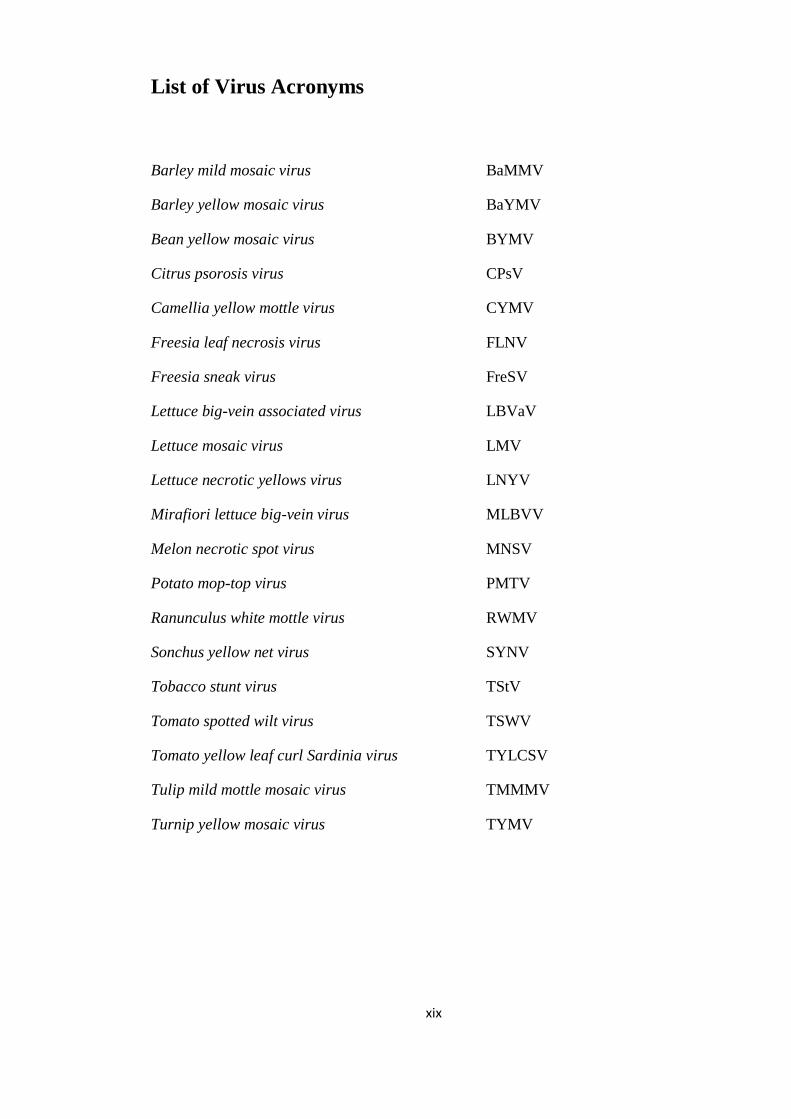
List of Virus Acronyms ________________________________________________ xix
Papers arising from this study ___________________________________________ xx
Oral presentations ____________________________________________________ xx
Poster presentations ___________________________________________________ xxi
Other Publications and Media Interest ____________________________________ xxi
Chapter 1 General Introduction and Literature Review________________________ 1
1.1 General Introduction _______________________________________________ 1
1.2 Lettuce production in Australia _____________________________________ 4
1.3 Lettuce production in Western Australia ______________________________ 7
1.4 Commonly occurring diseases of lettuce in Australia ____________________ 8
1.5 Chytrid and Plasmodiophorid vectors of plant viruses __________________ 11
1.6 Lettuce Big Vein Disease ________________________________________ 12
1.7 Control of LBVD _______________________________________________ 15
1.8 Specific research aims ___________________________________________ 18
1.9 Structure ______________________________________________________ 20
Chapter 2: General Materials and Methods ________________________________ 24
2.1 Viruses and Inoculation __________________________________________ 24
2.2 Nucleic acid extraction __________________________________________ 24
2.2.1 DNA extraction from Olpidium spp. _____________________________ 24
2.2.2 RNA extraction from viral species ______________________________ 25
2.3 Polymerase Chain Reaction (PCR) _________________________________ 25
2.3.1 PCR for Olpidium spp. amplification ____________________________ 25
2.3.2 Reverse Transcription Polymerase Chain Reaction (RT-PCR) for viral
amplification ___________________________________________________ 26
2.4 Primers _______________________________________________________ 27
2.5 Agarose gel electrophoresis _______________________________________ 27
2.6 Purification of PCR product for cloning or direct sequencing ____________ 30
2.7 DNA and RNA quantification _____________________________________ 30
2.8 Cloning_______________________________________________________ 30
2.8.1 Ligation using pGEM®
-T Easy Vector ___________________________ 30
2.8.2 Transformation using E. coli ___________________________________ 31
2.9 Screening for recombinant plasmids ________________________________ 32

x
2.10 Plasmid preparation and analysis __________________________________ 33
2.11 Restriction digest of plasmid _____________________________________ 33
2.12 Sequencing ___________________________________________________ 33
2.13 Virus detection by Double Antibody Sandwich Enzyme Linked Immuno-
Sorbent Assay (DAS-ELISA) _________________________________________ 35
2.13.1 Coating plates ______________________________________________ 36
2.13.2 Sample Preparation _________________________________________ 36
2.13.3 Conjugation _______________________________________________ 37
2.13.4 Substrate __________________________________________________ 37
Chapter 3 - Molecular genetic characterization of Olpidium virulentus isolates
associated with big-vein diseased lettuce plants ______________________________ 39
Abstract __________________________________________________________ 40
Introduction_______________________________________________________ 41
Materials and Methods ______________________________________________ 43
Collection of field samples _________________________________________ 43
DNA extraction, PCR and agarose gel electrophoresis ___________________ 43
Virus RNA extraction and Multiplex RT-PCR __________________________ 44
Cloning and sequencing of PCR amplicons ____________________________ 45
Analysis of Sequence Data _________________________________________ 46
Results___________________________________________________________ 47
PCR DNA extracts from roots ______________________________________ 47
Nucleotide sequence identities ______________________________________ 47
Phylogenetic analysis _____________________________________________ 47
RT-PCR RNA extracts from leaves __________________________________ 48
Discussion ________________________________________________________ 49
Acknowledgements_________________________________________________ 51
Literature cited ____________________________________________________ 52
Chapter 4 Comparison of the coat protein genes of Mirafiori lettuce big-vein virus
isolates from Australia with those of isolates from other continents _____________ 62
Introduction_______________________________________________________ 62
Materials and Methods ______________________________________________ 63
Sequence Properties________________________________________________ 64
Acknowledgements_________________________________________________ 66
Literature cited ____________________________________________________ 67
Chapter 5 - Comparison of the coat protein genes of Lettuce big-vein associated virus
isolates from Australia with those of isolates from other continents _____________ 71
Summary _________________________________________________________ 71
Acknowledgements_________________________________________________ 79
References ________________________________________________________ 79

xi
Chapter 6 – Commercial Lettuce Seedling Nurseries _________________________ 87
6. 1 Introduction ___________________________________________________ 87
6.1.1 Aims _____________________________________________________ 91
6.2 Materials and Methods___________________________________________ 92
6.2.1 Field collection of samples ____________________________________ 92
6.2.2 Collection of soil samples from a commercial seedling nursery _______ 93
6.2.3 Growth of lettuce seedlings in nursery soil samples _________________ 94
6.2.4 Source of seed ______________________________________________ 94
6.2.5 Surface sterilisation of seed ___________________________________ 96
6.2.6 Growing conditions __________________________________________ 97
3.2.7 ELISA and antibodies ________________________________________ 97
6.2.8 RNA extraction _____________________________________________ 98
6.2.9 Reverse transcription (RT) ____________________________________ 98
6.2.10 DNA extraction ____________________________________________ 99
6.2.11 PCR and agarose gel electrophoresis for RNA extracts _____________ 99
6.2.12 PCR and agarose gel electrophoresis for DNA extracts ____________ 100
6.2.13 Controls for DAS-ELISA and PCR ___________________________ 100
6.3 RESULTS ___________________________________________________ 102
6.3.1 Samples collected from commercial lettuce growing fields __________ 102
6.3.2 Samples collected from seedling nurseries and tested by PCR and DAS-
ELISA _______________________________________________________ 104
6.3.3 Surface sterilisation of seed __________________________________ 107
6.4 DISCUSSION ________________________________________________ 108
Chapter 7 Alternative Hosts ____________________________________________ 112
7.2 Materials and Methods__________________________________________ 116
7.2.1 Determining alternative hosts for LBVV and MLBVV _____________ 116
7.2.2 Samples collected from the field _______________________________ 116
7.2.3 Source of seedlings and inoculum for initial experiment ____________ 117
7.2.4 Growing conditions for inoculated seedlings for the initial experiment _ 117
7.2.5 Testing by DAS-ELISA of field collected samples and inoculated seedlings
used in the initial experiment ______________________________________ 118
7.2.6 Source of seed _____________________________________________ 119
7.2.7 Germination of seeds and inoculation of seedlings used to determine
alternative hosts ________________________________________________ 119
7.2.8 Sampling of plant tissue for DAS-ELISA ________________________ 120
7.2.9 RNA extraction of leaf tissue _________________________________ 120
7.2.10 Reverse Transcription Polymerase Chain Reaction (RT-PCR) ______ 120
7.3 Results ______________________________________________________ 122
7.3.1 Results of initial experiments _________________________________ 122
7.3.2 Field samples ______________________________________________ 123
7.3.3 Growth room experiment ____________________________________ 124
7.3.4 Observed virus symptoms in plant leaves ________________________ 127

xii
7.3.5 Confirmation of DAS-ELISA results by RT-PCR __________________ 130
7.4 Discussion ____________________________________________________ 133
Chapter 8 – General Discussion _________________________________________ 138
Appendix 1 – Amount of PCR product or plasmid used in sequencing reactions __ 152
Appendix 2 – The reagents used in each of the different DAS-ELISA solutions ___ 154
Appendix 3 - O. virulentus ITS sequences (5' to 3') _________________________ 155
Appendix 4 - MLBVV CP sequences (5' to 3') ______________________________ 158
Appendix 5 - LBVaV CP sequences (5' to 3') ______________________________ 167
Appendix 6 - Locations where MLBVV isolates were collected________________172
Appendix 7 - Locations where LBVaV isolates were collected_________________173
Appendix 8 - MLBVV amino acid and nucleotide distances__________________174
Appendix 9 - LBVaV amino acid and nucleotide distances___________________175
References_________________________________________________________176

xiii
Acknowledgements
Sincere thanks must go to my supervisors; Prof. Martin Barbetti for initially giving me
the opportunity to undertake this research, Adj. Prof Roger Jones for the many hours
you spent correcting this manuscript and Prof Krishnapillai Sivasithamparam for
keeping me motivated and inspired, especially towards the end.
Thank-you to all the people in the Plant Pathology group I have had the pleasure of
working with, especially Li Hua, Margaret Collins and Harsh Garg. I have truly valued
your friendship and knowledge and your understanding has often been what has helped
me through the many challenging times.
Thank-you to all the people at DAWFA who have helped me during this project; Stuart
Vincent and Eva Gadja for watering my plants, Gail Burchell for your help in editing
my reference lists, Brenda Coutts and Belinda Cox for sharing your knowledge and
providing support and especially Monica Kehoe for your „expert computer technical
knowledge‟ and a friendship which I am grateful.
Also, I would like to thank Steve Wylie and Craig Webster from the SABC for all of
your sequencing advice and help with sequence analysis.
Finally, I would like to thank my parents Ros and Joe Maccarone for keeping me and
my dogs (Zuko and Milly) housed and fed for the past two and a half years. I appreciate
it more than I let on and am forever in your debt. Now I can get a „real‟ job and move
out of home... again.
LDM

xiv
Candidate’s Declaration
The work presented in this thesis is entirely my own, unless specifically acknowledged
otherwise.
Linda Dianne Maccarone

xv
List of Abbreviations
3′ hydroxyl-terminus of DNA molecule
5′ phosphate-terminus of DNA molecule
½ one half
¼ one quarter
⅛ one eighth
°C degrees Celcius
ABS Australian Bureau of Statistics
Amp ampicillin
bp base pairs
BSA Bovine serum albumin
Ca(OCl)2 calcium hypochlorite
CBD Central Business District
CP coat protein
cDNA complimentary deoxyribonucleic acid
CALM Conservation and Land Management
cv. cultivar
DI deionised
DAFWA Department of Agriculture and Food, Western Australia
DAS-ELISA Double antibody sandwich enzyme-linked immunosorbent assay
ddH20 double distilled water
Dn non-silent mutation
DNA deoxyribonucleic acid
Ds silent mutation
EDTA ethylene diamine tetra-acetic acid
EtBr ethidium bromide
g gram
ha hectares

xvi
h hour
ITS Internally transcribed spacer region
kb kilobase
km kilometre
LBVD Lettuce Big-Vein Disease
L litre
LB medium Luria-Bertani medium
MgCl2 magnesium chloride
µg microgram
µL microlitre
µm micrometre
µM micromolar
mg milligram
min minute
mL millilitre
mm millimeter
mM millimolar
M molar
NaAC sodium acetate
nad4 mitochondrial NADH dehydrogenase subunit 4
NaOCl sodium Hypochlorite
ng nanogram
NFT Nutrient Film Technique
PNP para-nitrophenyl phosphate
pers. comm. Personal communication
PBST phosphate buffered saline with tween 20
pmol pico mole
PCR polymerase chain reaction
RT-PCR reverse transcriptase polymerase chain reaction

xvii
RNA ribonucleic acid
rDNA ribosomal deoxyribose nucleic acid
SABC State Agricultural Biotechnology Centre
sec second
SNPs single nucleotide polymorphisms
TBE buffer tris-boric acid-EDTA electrophoresis buffer
Na3PO4 trisodium phosphate
Taq Thermus aquaticus DNA polymerase
t tonnes
U units
UK United Kingdom
USA United States of America
UV ultraviolet
V volts
WA Western Australia
Code for degenerate oligonucleotides
A Adenosine M AC V ACG
C Cytosine R AG H ACT
G Guanine W AT D AGT
I Inosine S CG B CGT
T Thymine Y CT N AGCT
U Uracil K GT
xviii
Amino Acid Codes
Glycine Gly G
Alanine Ala A
Leucine Leu L
Methionine Met M
Phenylalanine Phe F
Tryptophan Trp W
Lysine Lys K
Glutamine Gln Q
Glutamic acid Glu E
Serine Ser S
Proline Pro P
Valine Val V
Isoleucine Ile I
Cysteine Cys C
Tyrosine Tyr Y
Histidine His H
Arginine Arg R
Asparagine Asn N
Aspartic acid Asp D
Threonine Thr T
xix
List of Virus Acronyms
Barley mild mosaic virus BaMMV
Barley yellow mosaic virus BaYMV
Bean yellow mosaic virus BYMV
Citrus psorosis virus CPsV
Camellia yellow mottle virus CYMV
Freesia leaf necrosis virus FLNV
Freesia sneak virus FreSV
Lettuce big-vein associated virus LBVaV
Lettuce mosaic virus LMV
Lettuce necrotic yellows virus LNYV
Mirafiori lettuce big-vein virus MLBVV
Melon necrotic spot virus MNSV
Potato mop-top virus PMTV
Ranunculus white mottle virus RWMV
Sonchus yellow net virus SYNV
Tobacco stunt virus TStV
Tomato spotted wilt virus TSWV
Tomato yellow leaf curl Sardinia virus TYLCSV
Tulip mild mottle mosaic virus TMMMV
Turnip yellow mosaic virus TYMV

xx
Papers arising from this study
Maccarone, L.D., Barbetti, M., Sivasithamparam, K. and Jones, R.A.C. 2010. Molecular
genetic characterisation of Olpidium virulentus isolates associated with big-vein
diseased lettuce plants. Plant Disease 94:563-569.
Maccarone, L.D., Barbetti, M., Sivasithamparam, K. and Jones, R.A.C. 2010.
Comparison of the coat protein genes of Mirafiori lettuce big-vein associated virus
isolates from Australia with those of isolates from other continents. Accepted in
Archives of Virology 7th
June, 2010
Maccarone, L.D., Barbetti, M., Sivasithamparam, K. and Jones, R.A.C. 2010.
Comparison of the coat protein genes of Lettuce big-vein associated virus isolates from
Australia with those of isolates from other continents Archives of Virology DOI
10.1007/s00705-010-0641-0
Oral presentations
School of Plant Biology Rottnest Island Postgraduate Summer School, Western
Australia 2007
Maccarone, L.D., Jones, R.A.C., Barbetti, M. and Sivasithamparam, K. 2007.
Prevention and spread of Lettuce Big-Vein Disease by improved production practices in
nurseries. In University of Western Australia, School of Plant Biology, 2007 Rottnest
Island Postgraduate Summer School Proceedings, page 30
Australasian Plant Pathology Society, Plant Health Research Symposium, South Perth,
Western Australia 2008
Maccarone, L.D., Jones, R.A.C., Sivasithamparam, K., Barbetti, M. 2008. Phylogenetic
analysis of Olpidium virulentus, Lettuce big-vein virus and Mirafiori lettuce big-vein
virus: the disease causing agents of Big-Vein Disease in lettuce in Western Australia

xxi
School of Plant Biology Rottnest Island Postgraduate Summer School, Western
Australia 2009
Maccarone, L.D., Sivasithamparam, K., Barbetti, M. and Jones, R.A.C. 2009
Phylogenetic analysis of Olpidium virulentus, Lettuce big-vein virus and Mirafiori
lettuce big-vein virus isolates from Western Australia
Poster presentations
Joint meeting American Pytopathological Society/Canadian Phytopathological
Society/Mycological Society of America, Quebec City, QC, Canada 2006
Maccarone, L.D., Jones, R.A.C., Latham, L.J., Sivasithamparam, K. and Barbetti, M.J.
2006. The role of nursery practices in the survival and production spread of organisms
associated with Big-Vein Disease in lettuce in Western Australia. Phytopathology 96:
71-72.
Australiasian Plant Virology Workshop, Rottnest Island, Western Australia 2006
Maccarone, L.D., Jones, R.A., Barbetti, M and Sivasithamparam, K., (2006) The role of
nursery practices in the survival and spread of Big-Vein Disease of lettuce in Western
Australia, in 7th Australasian Plant Virology Workshop Program Book, page 66
Other Publications and Media Interest
Newspaper article and front cover by Bart McGann in Countryman Horticulture, January 2007.
Tip of the iceberg – Researcher Linda Maccarone is working to stop the spread of Lettuce Big-
vein Disease, cover
Lettuce under attack – Nurseries focus of disease battle, page 3
University of Western Australia Institute of Agriculture newsletter, Number 6, December, 2008
Plant Health Research Symposium

xxii
http://www.uwa.edu.au/__data/assets/pdf_file/0009/132867/UWA_IOA_newsletter_Dec_08.pd
f

1
Chapter 1 General Introduction and Literature Review
1.1 General Introduction
Lettuce big-vein disease (LBVD) was first reported from California (Jagger and
Chandler 1934). It occurs widely in regions of the world with temperate or
Mediterranean-type climates, and at high altitudes in subtropical regions and is common
in Australia and in Western Australia. LBVD is now known to be associated with a
complex of two viruses, Lettuce big-vein associated virus (LBVaV; genus
Varicosavirus) and Mirafiori lettuce big-vein virus (MLBVV; genus Ophiovirus)
(Roggero et al. 2000) which are vectored by the root-infecting chytrid Olpidium
virulentus. Traditionally, it has been accepted that LBVaV was the virus responsible for
LBVD symptoms however, Lot et al. (2002) and Sasaya et al. (2006) described
MLBVV as being the causal agent of big-vein symptoms of lettuce. Also, until recent
molecular studies of the chytrid vector, it has been thought that the vector for both
LBVaV and MLBVV was O. brassicae when in fact it was determined to be O.
virulentus (Koganezawa et al. 2005; Sasaya and Koganezawa 2006).
Motile zoospores O. virulentus transmit LBVD to the roots of healthy plants and, in the
absence of susceptible hosts, its resting spores retain ability to harbour and transmit
LBVD for decades in infested soil (Campbell 1985; Barr 1988; Lot et al. 2002; Rochon
et al. 2004). Methyl bromide was formerly used widely as a fungicide to control the
Olpidium vector, but has since been banned in some countries including Australia.

2
In Australia, LBVD is a widespread problem in commercial lettuce producing areas. In
south-west Australia, the lettuce production system on the Swan Coastal Plain is
intensive, often uses irrigated land that is contaminated with LBVD and sometimes
involves little rotation with other crops. In Western Australia, LBVD often results in
decreased yields of produce from infected plants in winter growing seasons and this loss
has been shown to be up to 90% of the crop (Latham and Jones 2004; Latham et al.
2004). Symptoms of this disease are the obvious chlorophyll clearing which causes the
„big-vein‟ leaf symptoms from which the disease gets its name, crinkled leaves and a
reduced head size which is often oblong in shape (Fig. 1.1) rather than round in shape as
is seen in healthy lettuce plants.
Fig. 1.1: Lettuce plants grown on a commercial lettuce growing property in Western
Australia showing symptoms of LBVD. The lettuce plant on the left is not showing any
symptoms of LBVD. The plant in the centre has severe symptoms such as leaf crinkling
and which has prevented head formation. The plant on the right shows the „oblong‟
shaped head seen with LBVD.

3
Latham et al. (2004) demonstrated up to 31% incidence of LBVD in lettuce seedlings
collected from a commercial lettuce seedling nursery known to have LBVD. These
seedlings were collected over an 18 month period and tested by DAS-ELISA to
determine LBVaV infection. In the same study, contamination of LBVaV was detected
in „bait‟ plants that were grown in potting mix over variable composting time periods, in
soil that was collected from under nursery benches and also in bark which was collected
from the potting mix supplier‟s site. Given the urban sprawl experienced in the
metropolitan region of Perth, lettuce producers are being forced further from the city
and are cultivating virgin land. Given the persistent nature of O. virulentus resting
spores, it is important that in an integrated disease management strategy for this disease,
that commercial lettuce seedling nurseries produce seedlings that are LBVD-free (Jones,
2004). This could help to reduce the spread of this disease in currently infested areas
and also to new land. A thorough survey of commercial lettuce seedling nurseries,
potting mix and pine bark suppliers in Western Australia would give a greater
understanding in regards to the source of LBVD infection in Western Australia.
The overall aims of this study were:
Determine that O. virulentus is the vector of LBVaV and MLBVV in Western
Australia, not O. brassicae
Isolate O. virulentus, LBVaV and MLBVV from Western Australian lettuce
growing areas and sequence these isolates to determine phylogenetic
relationships between each other and other isolates available on GenBank
Determine the presence of LBVaV and MLBVV in commercial lettuce seedling
nurseries as only LBVaV had been detected previously and investigate other

4
possible sources of infection outside the seedling nursery and commercial
growing environments
Assess known and unknown alien and native hosts for their ability to transmit
LBVD or act as disease reservoirs.
This review presents an overall picture of LBVD, the viruses and the vectors associated
with the disease in the literature. It also covers, in general, various sources of inoculum
of the viruses and the vector to relate them to disease management. A detailed
evaluation of the relevant results from previous studies is presented in the introduction
and discussion sections of relevant chapters of this thesis.
1.2 Lettuce production in Australia
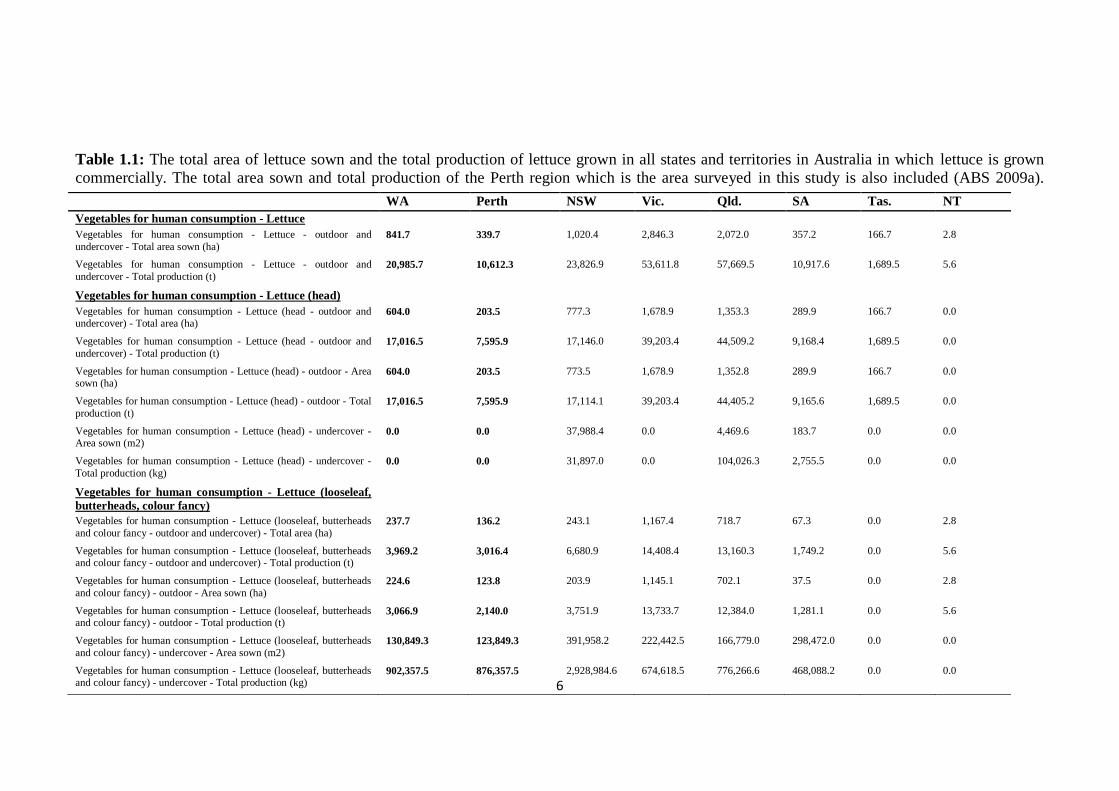
Lettuce is commercially grown in all states and territories within Australia except in the
Australian Capital Territory. In the 2007-08 financial year, a total of 7307ha of land
sown (Table 1.1). Of this total cultivation area, 4870ha were used in the production of
outdoor and undercover-grown iceberg or „head‟ type lettuce and 2437ha were used for
the production of outdoor and under-cover grown looseleaf, butterhead and coloured
fancy lettuce types (Australian Bureau of Statistics (ABS) 2009a). This area over the
same time period, produced a total of 168 707t of lettuce, 128 733t which was „head‟
lettuce and 39 977t which was looseleaf, butterhead and coloured fancy lettuce (Table
1.1). The state of Queensland is Australia‟s major lettuce producer, producing 34.2% of
all lettuce during the 2007-08 financial year (ABS 2009a) (Table 1.1).
During the 2005-06 financial year, the gross value of lettuce production in Australia
was $159m. In the 2006-07 financial year, this figure was raised to $283m but then fell

5
to $168m during the 2007-08 financial year (Table 1.2a). The gross value of lettuce
production is based on the wholesale prices realised in the marketplace whereas the
local value (Table 1.2a) is the value placed on the lettuce crop at the point of production
(at the farm gate) by deducting the marketing costs from the gross value (ABS 2009b).
Of the total lettuce value during the 2007-08 financial year, Queensland, the country‟s
major producer of lettuce was responsible for 39% of the value (Table 1.2b).
6
Table 1.1: The total area of lettuce sown and the total production of lettuce grown in all states and territories in Australia in which lettuce is grown
commercially. The total area sown and total production of the Perth region which is the area surveyed in this study is also included (ABS 2009a).
WA Perth NSW Vic. Qld. SA Tas. NT
Vegetables for human consumption - Lettuce Vegetables for human consumption - Lettuce - outdoor and
undercover - Total area sown (ha) 841.7 339.7 1,020.4 2,846.3 2,072.0 357.2 166.7 2.8
Vegetables for human consumption - Lettuce - outdoor and
undercover - Total production (t) 20,985.7 10,612.3 23,826.9 53,611.8 57,669.5 10,917.6 1,689.5 5.6
Vegetables for human consumption - Lettuce (head) Vegetables for human consumption - Lettuce (head - outdoor and undercover) - Total area (ha)
604.0 203.5 777.3 1,678.9 1,353.3 289.9 166.7 0.0
Vegetables for human consumption - Lettuce (head - outdoor and
undercover) - Total production (t) 17,016.5 7,595.9 17,146.0 39,203.4 44,509.2 9,168.4 1,689.5 0.0
Vegetables for human consumption - Lettuce (head) - outdoor - Area sown (ha)
604.0 203.5 773.5 1,678.9 1,352.8 289.9 166.7 0.0
Vegetables for human consumption - Lettuce (head) - outdoor - Total
production (t) 17,016.5 7,595.9 17,114.1 39,203.4 44,405.2 9,165.6 1,689.5 0.0
Vegetables for human consumption - Lettuce (head) - undercover - Area sown (m2)
0.0 0.0 37,988.4 0.0 4,469.6 183.7 0.0 0.0
Vegetables for human consumption - Lettuce (head) - undercover -
Total production (kg) 0.0 0.0 31,897.0 0.0 104,026.3 2,755.5 0.0 0.0
Vegetables for human consumption - Lettuce (looseleaf,
butterheads, colour fancy)
Vegetables for human consumption - Lettuce (looseleaf, butterheads
and colour fancy - outdoor and undercover) - Total area (ha) 237.7 136.2 243.1 1,167.4 718.7 67.3 0.0 2.8
Vegetables for human consumption - Lettuce (looseleaf, butterheads and colour fancy - outdoor and undercover) - Total production (t)
3,969.2 3,016.4 6,680.9 14,408.4 13,160.3 1,749.2 0.0 5.6
Vegetables for human consumption - Lettuce (looseleaf, butterheads
and colour fancy) - outdoor - Area sown (ha) 224.6 123.8 203.9 1,145.1 702.1 37.5 0.0 2.8
Vegetables for human consumption - Lettuce (looseleaf, butterheads and colour fancy) - outdoor - Total production (t)
3,066.9 2,140.0 3,751.9 13,733.7 12,384.0 1,281.1 0.0 5.6
Vegetables for human consumption - Lettuce (looseleaf, butterheads
and colour fancy) - undercover - Area sown (m2) 130,849.3 123,849.3 391,958.2 222,442.5 166,779.0 298,472.0 0.0 0.0
Vegetables for human consumption - Lettuce (looseleaf, butterheads and colour fancy) - undercover - Total production (kg)
902,357.5 876,357.5 2,928,984.6 674,618.5 776,266.6 468,088.2 0.0 0.0

7
Table 1.2a: The total value of lettuce grown in Australia during the financial years
2005-06, 2006-07 and 2007-08. Both gross values and the value of the produce at the
farm gate (local value) are shown (ABS, 2009b).
Year Gross value ($m) Local value ($m)
2006 159.1 123.7
2007 282.9 225.1
2008 168 128.8
Table 1.2b: The total value of lettuce grown in all states and territories within Australia
where lettuce is grown commercially for the financial year 2007-08. Both gross values
and the value of the produce at the farm gate (local value) are shown (ABS, 2009b).
State Gross value ($m) Local value ($m)
NSW 22.9 15.6
Vic. 49.2 41.0
Qld. 65.6 48.1
SA 13.4 11.6
WA 14.5 10.5
Tas. 2.3 2
NT 0 0
1.3 Lettuce production in Western Australia
In Western Australia, a total of 842ha was sown for both outdoor and undercover lettuce
production in the 2007-08 financial year which was 11.5% of the total area under lettuce
production in Australia (Table 1.1). During 2007-08, Western Australia produced 20
986t (12.4%) of the total lettuce produced in Australia. In this same time period, the
gross value of lettuce production in Western Australia was $14.5m which was 8.6% of
the National total (ABS 2009) (Table 1.2b).

8
ABS recognises Statistical Divisions for the purpose of generating data. All of the
properties surveyed in this study except for the one property surveyed in Manjimup
approximately 300km south of Perth (which is the South West Statistical Division) are
found in the Statistical Division of Perth (ABS 2009a). During the 2007-08 financial
year, the area of lettuce production in the Perth region was 40.4% of the total area under
lettuce production in the state (Table 1.1). This area was responsible for 50.6% of the
total lettuce production for Western Australia. This study surveyed commercial lettuce
growing properties that grew their crops outdoors and produced „head‟ type lettuce as
well as looseleaf, butterhead and coloured fancy lettuce. The Perth region produced
9736t of outdoor-grown lettuce which was 48% of Western Australia‟s total outdoor-
grown lettuce production (ABS 2009a) (Table 1.1).
1.4 Commonly occurring diseases of lettuce in Australia
Other than LBVD, there are many other common virus, fungus and bacterial diseases
which affect lettuce production in Australia. These diseases cause significant loss to the
quality of the lettuce often resulting in a crop which cannot be harvested or a crop with
reduced yield. Of the bacterial diseases that affect lettuce, Dry Leaf Spot, which is
caused by Xanthomonas campestris pv vitians, is a common lettuce disease in Western
Australia and causes pale brown watermarked areas on the infected lettuce leaf. In
Western Australia, one of the most common sources of disease comes from two
Sclerotinia spp., S. minor which causes Lettuce Drop and S. sclerotiorum which causes
Head Rot. Both S. minor and S. sclerotiorum ensure the lettuce plants are un-
harvestable. Downy Mildew is also a common disease of lettuce in Western Australia
and is caused by the oomycete pathogen Bremia lactucae and causes brown patches on

9
the upper leaf surface and white hyphae on the under-side of the leaf. Crown Rot which
is also known as Grey Mould is caused by the fungus Botrytis cinerea and Bottom Rot
which is caused by the fungus Rhizoctonia solani are also present in Western Australia
and cause similar symptoms of brown rot at ground level often at the time of harvest.
Septoria Leaf Spot is another fungal disease of lettuce in Western Australia which is
caused by the fungus Septoria lactucae and causes symptoms such as angular pale
brown lesions which as they mature, develop pin-head black spots (Floyd 2000; Napier
2004).
Of the commonly occurring virus diseases of lettuce in Western Australia, Lettuce
necrotic yellows virus (LNYV) is a member of the genus Cytorhabdovirus and was first
described in Australia in 1954 by Stubbs and Grogan (1963). LNYV is transmitted by
the aphid vector Hyperomyzus lactucae and causes symptoms in lettuce such as vein
yellowing which is usually followed by chlorosis and leaf curling (Stubbs and Grogan
1963; Francki and Randles, 1970). LNYV is widespread throughout Australia and has
also been reported in New Zealand (Fry et al. 1973).
Weed species such as S. oleraceous, Reichardia picroides and R. tingitana and
Australian endemic species such as S. hydrophilus and Embergeria megalocarpa have
been demonstrated to be infected with this virus and it has been suggested that the
ability of such species to support colonies of the vector H. lactucae provide a possible
inoculum source of LNYV (Randles and Carver 1971). Given the ability to infect
endemic plants and the limited distribution of LNYV, Randles and Carver (1971)
suggested that LNYV may have originated from plant species endemic to Australia
and/or New Zealand.

10
Tomato spotted wilt virus (TSWV) is another commonly occurring virus among lettuce
crops and is a member of the genus Tospovirus and is vectored by many thrip species
although most efficiently by the Western Flower Thrip Frankliniella occidentalis (Zitter
and Daughtrey 1989). Like LNYV, TSWV was also first described in Australia,
although earlier, in 1915 (Brittlebank 1919). TWSV is now common in temperate,
subtropical and tropical regions around the world however, a lettuce strain of the virus
is usually found in vegetables whereas an impatiens strain is usually found in
ornamentals (Zitter and Daughtrey 1989). TSWV causes severe symptoms in lettuce
which often result in plant death. Initial infection symptoms are necrotic spotting which
may be followed by wilting, yellowing and necrotic spotting (Cho et al. 1989). TSWV
has more than 900 known host species which belong to families Solanaceae, Asteraceae
and Legumaceae. S. oleraceous and Arctotheca calendula, common weed species found
amongst lettuce growing fields are known to be hosts of TSWV (Peters 1998) and act as
inoculum sources for the virus (Wilson 1998).
Lettuce mosaic virus (LMV) is a member of the genus Potyvirus and is a common
pathogen of lettuce in Western Australia which causes restricted growth and therefore
reduced yield (Napier 2004). LMV is commonly transmitted by the Green Peach aphid
Myzus persicae however a small percentage of the virus is known to be seed-borne
(Ryder 1973). Symptoms associated with LMV infection in lettuce are necrotic or
chlorotic local lesions and streaking, mosaic, leaf yellowing and leaf malformation. As
well as being a host for other plant viruses that infect lettuce (Peters 1998; Randles and
Carver 1971; Coutts et al. 2004, Chapter 7), S. oleraceous is also a host of LMV and is
known to act as a reservoir and inoculum source (Tomlinson 1970). Moreno et al.

11
(2007) in a laboratory environment demonstrated 37.5% infection of LMV vectored by
M. persicae from Sonchus to lettuce.
1.5 Chytrid and Plasmodiophorid vectors of plant viruses
Uniflagellate chytrids (class Chytirdiomycetes), O. brassicae and O. bornovanus and
biflagellate plasmodiophorids (class Plasmodiophoromycetes), Polymyxa graminis, P.
betae and Spongospora subterranea are known to transmit viruses in the soil to plant
roots (Matthews 1981; Bos 1999). O. bornovanus is known to vector six different virus
species, O. brassicae 10, P. graminis 15, P. betae four and S. subterranea two and each
vector species is able to transmit viruses from a range of different genera (Rochon et al.
2004). However, Sasaya and Koganezawa (2006) determined that O. virulentus was
also chytrid vector responsible for transmitting TStV and MLBVV. This study (Chapter
6) has also demonstrated that LBVaV, although once thought to be vectored by O.
brassicae (Campbell and Grogan 1962) is associated with O. virulentus but not with O.
brassicae.
Soil and vector-borne viruses can be transmitted by chytrid or plasmodiophorid vectors
either externally (in vitro) on the zoospore or resting spore or internally (in vivo) within
the zoospore or resting spore (Matthews 1981; Bos 1999). In the event of in vitro virus
transmission, the virus particles are most likely carried on the zoospore flagellum and
when the zoospore attaches to the host root, the flagellum is withdrawn into the
zoospore prior to release of the cytoplasmic contents into the host epidermal cells and it
is likely that this is when the virus is able to enter the host root (Bos 1999).

12
In the case of in vivo virus acquisition by the vector, the virus enters the thallus and
remains in the zoospore or the resting spore where it does not multiply until it is
released into the host root cells (Bos 1999). For the virus, this is an effective means of
transmission as viruliferous resting spores from dry crop debris, for example, can lay
dormant in the soil until environmental conditions are favourable for zoospore
germination (Bos 1999). In this case crop infestation is usually observed after periods of
rain or irrigation (Bos 1999).
1.6 Lettuce Big Vein Disease
To elaborate on what has already been discussed in relation to LBVD previously in this
chapter, one of the most highly contested aspects of this disease has been determining
which species of Olpidium is the vector for both LBVaV and MLBVV. O. brassicae
was first described in 1939 (Sampson 1939) and then first associated with big-vein
disease of lettuce by Fry (1958). It was in 1961 that graft transmission of big-vein
disease in lettuce was first reported and it was suggested that the disease was caused by
a virus (Campbell et al. 1961). However, in 1962, it was suggested by Campbell and
Grogan, (1962) that O. brassicae was the vector of a virus that caused big-vein disease
in lettuce (Table 1.3).
In that same year, Sahtiyanci (1962) proposed that based on differing hosts ranges that
there were infact two separate species of Olpidium, a crucifer infecting strain, O.
brassicae and a non-crucifer infecting strain, O. virulentus. Sahtiyanci (1962) also
found that O. brassicae was heterothallic whereas O. virulentus was homothallic.
Sahtiyanci (1962) also proposed that due to O. brassicae, O. virulentus and O.

13
bornovanus sporangia having more than one exit tube, they should be reassigned to the
genus Pleotrachelus. Campbell and Lin (1976) disputed this work on the basis that they
found O. brassicae sporangia to have single exit tubes and hence should remain in the
genus Olpidium and Campbell and Lin (1976) also disputed the separation of two
strains of O. brassicae, because even though they determined different host ranges, they
were unable to infect plants with single-sporangial isolates of each strain. Campbell and
Lin (1976) did however also find O. virulentus to be homothallic but could not
determine the nature of O. brassicae in this regard and concluded that there was not
enough specialisation to separate O. virulentus from O. brassicae. As a consequence, O.
brassicae has continued to be traditionally regarded as the vector of LBVaV and
MLBVV. However, in 2006, Sasaya and Koganezawa (2006) using molecular analysis
techniques determined that O. brassicae and O. virulentus are in fact different species
and not strains of the same taxon. They also determined that O. virulentus, not O.
brassicae is the vector of MLBVV and that both Olpidium spp. have different host
ranges, with O. brassicae commonly infecting crucifers (cabbage and mustard) while O.
virulentus commonly infects noncrucifers (lettuce and tomato).

14
Table 1.3: Timeline of events associated with LBVD, from the first initial description
of the disease in 1934 to molecular diagnostic techniques in 2006 that relate to the
research objectives discussed in this thesis.
Year Discovery Reference
1934 First description of big-vein of lettuce Jagger and
Chandler (1934)
1939 First description of O. brassicae Sampson (1939)
1958 First reports of the relationship of O. brassicae to the big-
vein disease of lettuce
Fry (1958) and
Grogan et al.
(1958)
1961 Graft transmission of big-vein suggests it is caused by a virus Campbell et al.
(1961)
1962 Big-Vein Virus is transmitted to lettuce by Olpidium Campbell and
Grogan (1962)
1962 Big-Vein Virus survives in the resting sporangia of Olpidium
however, does not multiply within the vector
Campbell (1962)
1962 Suggestion that O. brassicae and O. virulentus are different
species based on host range (O. brassicae infests crucifers
whilst O. virulentus infests noncrucifers) and that both
should be moved to genus Pleotrachelus based on
morphological characteristics.
Sahtiyanci
(1962)
1970 Different isolates of O. brassicae have different host ranges
and also differ in their ability to transmit Tobacco necrosis
virus (TNV).
Temmink et al.
(1970)
1976 The designation made by Sahtiyanci (1962) is challenged
and it is determined that Olpidium sp. not be reassigned to
Pleotrachelus and that there is not enough specialisation to
separate O. brassicae and O. virulentus into separate species
Campbell and
Lin (1976)
1985 Longevity of viruliferous O. brassicae resting spores in air-
dried soil was determined to be between 20.8 and 22.5 years
and the persistence of the big-vein agent within these spores
was determined to be between 18.7 and 20.8 years
Campbell (1985)
2000 An Ophiovirus is isolated from lettuce plants infected with
LBVD and named Mirafiori lettuce virus (MiLV). Some
plants infected with this virus showed big-vein symptoms
Roggero et al.
(2000)

15
2002 O. brassicae is determined as the vector of both viruses
associated with LBVD and MLBVV is determined as the
virus which causes big-vein symptoms.
Lot et al. (2002)
2004 Zoospores of O. brassicae and O. virulentus are determined
to differ morphologically and with host range. O. virulentus
transmits Tobacco necrosis virus (TNV) whilst O. brassicae
did not.
Koganezawa et
al. (2004)
2004 Commercial seedling nurseries in Western Australia are
determined as sources of LBVD inoculum
Latham et al.
(2004)
2006 Molecular analysis places O. brassicae and O. virulentus as
distinct species. O. virulentus is determined as the vector of
MLBVV and TStV
Sasaya and
Koganezawa
(2006)
1.7 Control of LBVD
Previous work carried out on the control of LBVD has been concerned with controlling
O. brassicae. As it is now accepted that O. virulentus is the chytrid vector of LBVaV
and MLBVV, it is reasonable to assume that O. virulentus and O. brassicae have similar
lifecycles and that all work done on previous control methods assuming the vector to be
O. brassicae can be directly related to O. virulentus. All references in this chapter made
to reports prior to Sasaya and Koganezawa (2006), although they mention O. brassicae,
are likely, in fact, to be dealing with O. virulentus.
Methods of control of LBVD concern both the resting spore stage of the vector and the
zoospore stage. Westerlund et al. (1978) demonstrated the importance of the soil-water
relationship required for O. brassicae germination. It was shown that zoospores were
released from sporangia when the soil was saturated and that a high level of moisture in

16
the soil also facilitated zoospore movement. This is an important consideration relating
the spread of LBVD in Western Australia as irrigation is common, often leaving the soil
saturated for a period. Westerlund et al. (1978) also made comparisons between O.
brassicae and Phytophthora in relation to zoospore movement in the soil and the water
requirements for zoospore production. This is an important comparison as similar
comparisons are suggested in the general discussion of this thesis. Jones (2004)
suggested that to reduce the spread of LBVD in the field, lettuce should be planted on
well drained soil and in areas where overhead irrigation is applied, soil should be
covered with black plastic mulch or lettuce plants grown on raised beds to reduce
excessive moisture levels in the soil and therefore reduce zoospore activity.
Given the thick cell-walled nature of O. brassicae resting spores and their persistence in
the soil for greater than 20 years, their control is most challenging. Previously, the use
of the volatile chemical methyl bromide has been used to control O. brassicae resting
spores. However, after initial control using up to two times the normal dose used on a
field, big-vein symptoms appeared in plants after the third year (Campbell et al. 1980;
White 1980). It also took three years from the first application of methyl bromide for
bromide levels in lettuce leaf tissue to be low enough to be suitable for human
consumption (White 1983). However, methyl bromide was recognised as an ozone
depleting substance and has been banned in Australia and in 159 other countries
worldwide who signed “The Montreal Protocol on Substances that Deplete the Ozone
Layer” in 1997 for a complete phase out of methyl bromide for non-quarantine purposes
by 2005.

17
Other forms of chemical control which are aimed at controlling O. brassicae zoospores
have been more effective. For example, Tomlinson (1988), found that growing a lettuce
crop using the recirculated Nutrient Film Technique (NFT) showed up to 100%
infection as conditions were ideal for zoospore movement. NFT however, provides an
ideal and effective way of delivering surfactant to the crop. It was demonstrated that a
20µL/mg concentration of Agral delivered through NFT suppressed viruliferous
zoospores for the life of the crop (Tomlinson 1988).
The use of resistant lettuce cultivars is also a means of controlling LBVD. In Western
Australia, different cultivars are grown at different times of the year and these seem to
have an effect on the incidence of LBVD. For example, in summer crisphead lettuce cv.
Raider and Aztec Sun are commonly grown; in spring and autumn cv. Jefferson,
Silverado and Marksman are grown whereas, in winter, cv. Titanic and Patagonia are
grown. (Funnekotter, pers. comm. 2006). Latham and Jones (2004) determined that by
placing black plastic mulch (to decrease soil moisture and increase temperature,
conditions that are not conducive to the multiplication and spread of LBVD) on the soil
of lettuce crops and by growing the partially resistant breeding line LE169, the
incidence of LBVD was reduced. When the susceptible cv. Oxley was compared to
LE169 in the same study, LE169 had an increased head weight of 17-48%. Complete
resistance to LBVD has only been reported in wild lettuce, L. virosa. However, the
mechanism of resistance to LBVD in lettuce is not known (Bos and Huijberts 1990;
Hayes et al. 2004) so more genetic information regarding O. virulentus, LBVaV and
MLBVV is needed in order to evaluate such mechanisms.

18
Based on previous work described here regarding the control of LBVD, and previous
integrated control methods suggested by Campbell et al. (1980), Jones (2004)
developed an Integrated Disease Management Strategy for control of LBVD in both the
seedling nursery environment and in commercial lettuce production areas. Several
methods of control were suggested in four different categories: Firstly, phytosanitary
cultural control methods such as ensuring seedling nurseries provide disease-free
seedlings to growers and avoid LBVD introductions via machinery and/or contaminated
water sources. Secondly, agronomic cultural control such as avoiding poorly drained
soil and manipulating the planting date. Thirdly, host resistance by deploying partially
resistant genotypes and finally, chemical control such as treating seedling nursery
potting mix and farming land with fungicides. Whilst this IDM strategy is
comprehensive, more information is needed with regards inoculum sources both in
commercial seedling nurseries and lettuce farms in order to better structure an IDM
strategy and make it region-specific. Also, Jones (2004) suggests that there might be
some vector-assisted seed transmission of the disease, so determining if this could
provide another source of inoculum is important for both lettuce seed producers and
commercial lettuce growers.
1.8 Specific research aims
To build on the overall aims of this study which were to determine O. virulentus as the
vector of LBVaV and MLBVV in Western Australia, determine the phylogenetic
relationships of O. virulentus, LBVaV and MLBVV isolates in Western Australia both
between each other and between other isolates of the same species found on GenBank
and to investigate possible sources of LBVD inoculum both in lettuce seedling nurseries

19
and in commercial lettuce growing fields, the specific aims relating to this study are as
follows:
Develop molecular techniques to detect both O. brassicae and O. virulentus
from lettuce tissue infected with LBVD
Determine if in Western Australia, the vector of LBVaV and MLBVV which
causes LBVD is O. brassicae or O. virulentus
Determine the phylogenetic relationship of the rDNA ITS region of Western
Australian Olpidium isolated from the roots of lettuce plants infected with
LBVD and compare them to each other and also to other Olpidium sequences
from Europe and Japan which are available on GenBank.
Develop molecular techniques to detect MLBVV from lettuce tissue infected
with LBVD
Determine the phylogenetic relationship of Western Australian isolates of the
CP gene of MLBVV and compare them to each other and also the same gene
sequence available on GenBank from five different continents
Develop molecular techniques to detect LBVaV from lettuce tissue infected with
LBVD
Determine the phylogenetic relationship of Western Australian isolates of the
CP gene of LBVaV and compare them to each other and also the same gene
sequence available on GenBank from five different continents
Determine the presence of LBVaV and MLBVV on commercial lettuce growing
farms in the Swan Coastal Plain of south-west Australia by testing samples by
DAS-ELISA.

20
Determine the presence of LBVaV and MLBVV in a commercial seedling
nursery in the Perth metropolitan region by testing samples by DAS-ELISA and
PCR.
Determine whether transmission of LBVD occurs through lettuce seed coat
contamination by resting spores of viruliferous O. virulentus by: a) detecting the
presence of LBVaV and/or MLBVV on the surface of lettuce seeds b)
determining whether sowing infected seed can result in infection with LBVaV
and/or MLBVV in lettuce plants grown from seeds harvested from LBVD-
infected plants.
to identify alternative weed hosts for LBVaV and MLBVV growing on the
Swan Coastal Plain of south-west Australia
to establish whether two native species of Asteraceae are hosts of to LBVaV and
MLBVV
to determine whether natural weed species collected in LBVD-affected lettuce
fields in Western Australia were infected with LBVaV and/or MLBVV
1.9 Structure
This thesis is in accordance with the Graduate Research School of the University of
Western Australia styles and format regulations, and is presented as one chapter of
general materials and methods used in this study, a series of three research papers and
two chapters that resulted from this study. Each of the three research papers can be read
as a separate entity or part of this thesis as a whole. Chapter 3 and Chapter 4 were
prepared for submission to Plant Disease and are presented in the format required for

21
that journal. Chapter 3 was accepted for publication on 19th
November, 2009 whilst
Chapter 4 was submitted on 18th
November, 2009. Chapter 5 was prepared and has been
submitted short communication to Archives of Virology on 30th
November, 2009 and is
presented in the format required for that journal. Each of these three chapters has a
separate abstract, introduction, materials and methods, results, discussion and reference
sections. There is some repetition especially in the introduction and discussion and
referencing sections however this is unavoidable given the related nature of each
chapter. Chapters 6 and 7 represent work which is yet to be transformed into a
publication format other than the requirements of this thesis. Chapter 8 includes a
general discussion and conclusions linking all of the chapters together as one coherent
body of research. A complete reference list which encompasses contributions of
previous work reported in all eight chapters closes this thesis. Relevant appendices are
also included.
In the current chapter, the aims, background and purpose of this research are presented.
The purpose of this research and research questions which were addressed in this study
are also presented.
In the second chapter (Chapter 2: General Materials and Methods) a general description
of all of the different protocols used within this study is provided. It includes growing
conditions of plants as well and molecular techniques that were used.
In the third chapter (Chapter 3: Molecular genetic characterization of Olpidium
virulentus isolates associated with big-vein diseased lettuce plants), five complete and
two partial rDNA ITS region sequences of O. virulentus were sequenced and compared

22
to sequences of other O. virulentus and O. brassicae sequences for the same region
were compared and this chapter presents the phylogenetic relationship amongst isolates
of O. brassicae, O. bornovanus and O. virulentus from Europe, Japan and Australia.
This chapter presents the first recorded rDNA ITS sequences of O. virulentus isolates
from Australia. The nucleotide sequences for these Western Australian isolates are
presented in the appendix.
A phylogenetic analysis of MLBVV is presented next (Chapter 4: Molecular genetic
characterization of Mirafiori lettuce big-vein virus isolates from five continents). This
chapter presents the phylogenetic relationships of the CP gene of MLBVV using
isolates from five continents, 13 of which were complete CP genes isolated in this
study. The complete nucleotide and amino acid sequences for the Western Australian
isolates are presented in the appendix.
The final research paper presented in this thesis presents the phylogenetic relationships
of the CP gene of LBVaV using isolates from five continents, seven of which were
isolated in this study (Chapter 5: Molecular genetic characterization of Lettuce big-vein
associated virus isolates from five continents). The complete nucleotide and amino acid
sequences of these isolates are included in the appendix.
The role of commercial seedling nurseries in the spread of LBVD is discussed in
Chapter 6. Chapter 6 (Nursery), for the first time reports the externally seed-borne
nature of viruliferous O. virulentus resting spores and highlights the importance of
disease-free seed in any integrated disease management strategy that would be
suggested for control of this disease. This chapter also demonstrates the presence of

23
MLBVV in commercial seedling nurseries for the first time in Western Australia as
previous studies did not include MLBVV as it had not yet been discovered as the causal
agent of LBVD.
In Chapter 7 (Alternative hosts for LBVaV and MLBVV), known and unknown hosts
were assessed both in the field and under experimental conditions for the presence of
LBVaV and MLBVV. Native Asteraceae, S. hydrophilus and Actites sp. were
determined as new hosts for LBVaV and MLBVV. Alien species Reichardia sp. and A.
calendula were determined as new hosts of MLBVV whilst Reichardia sp. was also
determined as a host of LBVaV.
The final chapter presents a detailed discussion and conclusion linking the findings of
each separate research paper and chapter reported in this thesis to the overall aims of
this study.

24
Chapter 2: General Materials and Methods
2.1 Viruses and Inoculation
Seeds used in this study were grown in seedling trays in potting mix in an insect proof,
controlled environment room at the Department of Agriculture and Food, South Perth,
Western Australia or in a similar growth room at the University of Western Australia
which was kept at a constant temperature of 18°C with a 12h photoperiod.
Approximately 4 weeks after germination, experimental seedlings were transplanted
into 140mm pots and inoculated with a 1:1 mixture of potting mix and soil infected with
LBVD. Plants were grown the same conditions as for germinating seeds and were
watered daily. Further materials and methods regarding growing conditions and soil
inoculum are given in Chapters 6 and 7.
2.2 Nucleic acid extraction
2.2.1 DNA extraction from Olpidium spp.
The Qiagen DNeasy Plant Mini Kit (Qiagen Inc., Valencia, CA ) was used on plant root
tissue according to the protocol supplied. This method gave consistently high yields of
DNA which could be used for amplification.

25
2.2.2 RNA extraction from viral species
Two different methods of RNA extraction were used which gave consistently high
yields of RNA which could be used for amplification. The Qiagen RNeasy Plant Mini
Kit (Qiagen Inc., Valencia, CA ) and the UltraClean™ Plant RNA Isolation Kit (MoBio
Laboratories, Inc. ) were used on plant root and leaf tissue as per the manufacturer‟s
protocol for each kit.
2.3 Polymerase Chain Reaction (PCR)
2.3.1 PCR for Olpidium spp. amplification
The 5 × polymerisation buffer used for this PCR mixture was purchased from Fisher
Biotec, Perth. The reagents used for a standard reaction were as follows:
Reagent Volume (µL)
5 × polymerisation buffer 20
50mM MgCl2 5 (2.5mM)
Taq polymerase (5.5 U/µL) 0.5
Primers (10pmol/µL) 0.5 each
cDNA template 2/reaction
Water To 100µL
The PCR reactions were then placed in the thermocycler under the following
temperature conditions:
94ºC for 1min
Then 35 cycles of:

26
94ºC for 30sec (denaturation)
60ºC for 30sec (annealing)
72ºC for 1min (extension)
Then a final extension at 72ºC before being held at 14ºC
2.3.2 Reverse Transcription Polymerase Chain Reaction (RT-PCR) for viral
amplification
OneStep RT-PCR kit was purchased from Qiagen, Australia. The following reagents
were used for each single reaction:
Reagent Volume
(µL)
5 × Polymerisation
buffer
2
10mM dNTPs 0.4
Primers (10pmol/µL) 0.2
Enzyme (5U/µL) 0.4
Rnasin 0.05
RNA template 0.5/tube
Water To 10µL
The PCR reactions were then placed in the thermocycler under the following
temperature conditions:
50°C for 30min
95°C for 15min
Then 30 cycles of:
94°C for 1min (denaturation)
50-57°C for 1min (annealing)

27
72°C for 1min (extension)
Then a final extension at 72°C for 10min before being held at 14°C
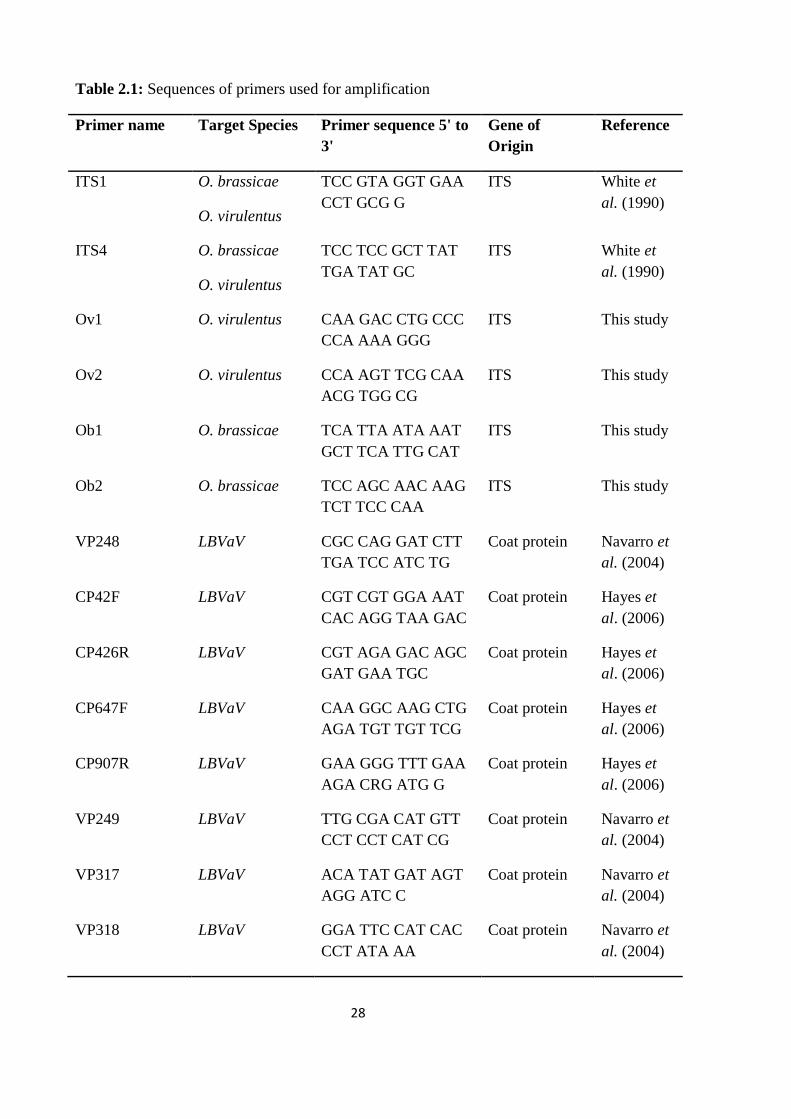
2.4 Primers
Primers used for amplification were either designed specifically by aligning known
sequences on Genbank or chosen from already known primer sequences (Table 2.1). All
primers were purchased from GeneWorks Pty Ltd, South Australia.
2.5 Agarose gel electrophoresis
PCR amplicons were visualised on 1% agarose gels by electrophoresis. Tris-boric acid-
EDTA (TBE) electrophoresis buffer was used (Sambrook and Russell, 2001). The gels
were run with 8µL of PCR product and 1µL of sucrose-based loading buffer (Sambrook
and Russell, 2001) and 6µL of 100bp ladder molecular weight marker (Axygen Corp.)
was used as a size standard. The gels were run on either Bio-Rad Mini-Sub Cells™ or
Bio-Rad Wide Mini-Sub Cells™ at 80V until the products were separated. The gels
were then stained in a 1µg/mL Ethidium Bromide (EtBr) solution for 15–20min and
then visualised on a transilluminator.
28
Table 2.1: Sequences of primers used for amplification
Primer name Target Species Primer sequence 5' to
3'
Gene of
Origin
Reference
ITS1 O. brassicae
O. virulentus
TCC GTA GGT GAA
CCT GCG G
ITS White et
al. (1990)
ITS4 O. brassicae
O. virulentus
TCC TCC GCT TAT
TGA TAT GC
ITS White et
al. (1990)
Ov1 O. virulentus CAA GAC CTG CCC
CCA AAA GGG
ITS This study
Ov2 O. virulentus CCA AGT TCG CAA
ACG TGG CG
ITS This study
Ob1 O. brassicae TCA TTA ATA AAT
GCT TCA TTG CAT
ITS This study
Ob2 O. brassicae TCC AGC AAC AAG
TCT TCC CAA
ITS This study
VP248 LBVaV CGC CAG GAT CTT
TGA TCC ATC TG
Coat protein Navarro et
al. (2004)
CP42F LBVaV CGT CGT GGA AAT
CAC AGG TAA GAC
Coat protein Hayes et
al. (2006)
CP426R LBVaV CGT AGA GAC AGC
GAT GAA TGC
Coat protein Hayes et
al. (2006)
CP647F LBVaV CAA GGC AAG CTG
AGA TGT TGT TCG
Coat protein Hayes et
al. (2006)
CP907R LBVaV GAA GGG TTT GAA
AGA CRG ATG G
Coat protein Hayes et
al. (2006)
VP249 LBVaV TTG CGA CAT GTT
CCT CCT CAT CG
Coat protein Navarro et
al. (2004)
VP317 LBVaV ACA TAT GAT AGT
AGG ATC C
Coat protein Navarro et
al. (2004)
VP318 LBVaV GGA TTC CAT CAC
CCT ATA AA
Coat protein Navarro et
al. (2004)
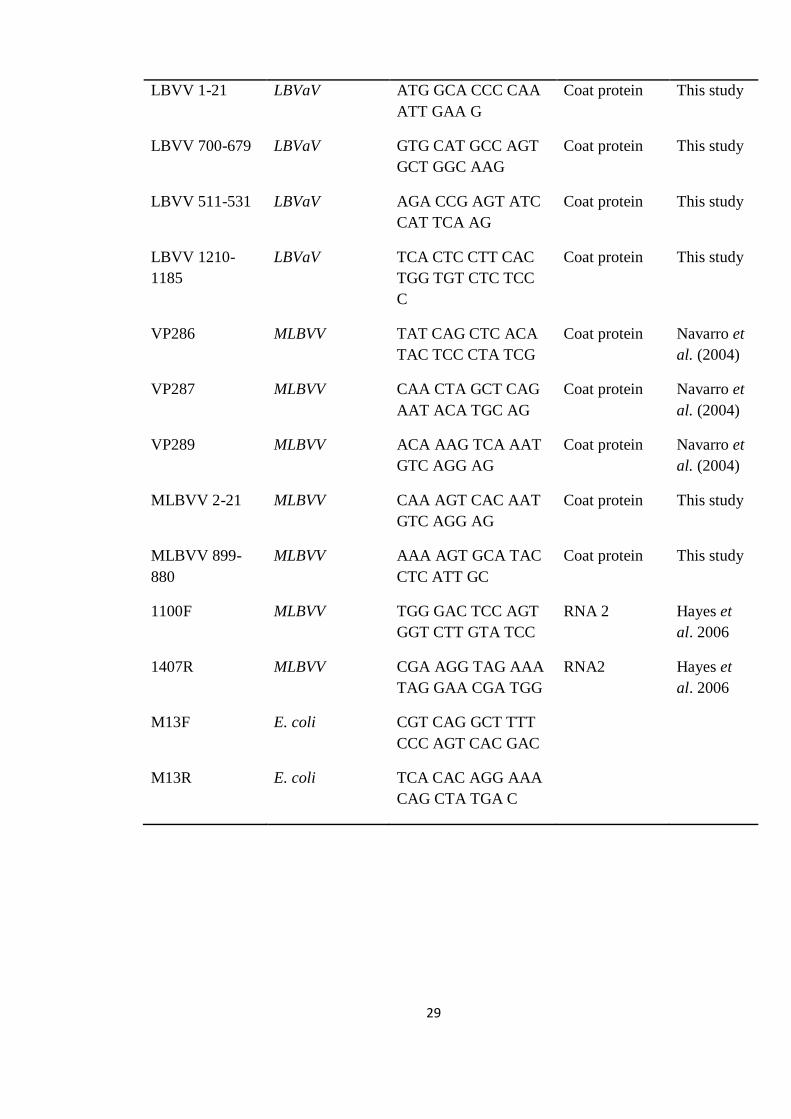
29
LBVV 1-21 LBVaV ATG GCA CCC CAA
ATT GAA G
Coat protein This study
LBVV 700-679 LBVaV GTG CAT GCC AGT
GCT GGC AAG
Coat protein This study
LBVV 511-531 LBVaV AGA CCG AGT ATC
CAT TCA AG
Coat protein This study
LBVV 1210-
1185
LBVaV TCA CTC CTT CAC
TGG TGT CTC TCC
C
Coat protein This study
VP286 MLBVV TAT CAG CTC ACA
TAC TCC CTA TCG
Coat protein Navarro et
al. (2004)
VP287 MLBVV CAA CTA GCT CAG
AAT ACA TGC AG
Coat protein Navarro et
al. (2004)
VP289 MLBVV ACA AAG TCA AAT
GTC AGG AG
Coat protein Navarro et
al. (2004)
MLBVV 2-21 MLBVV CAA AGT CAC AAT
GTC AGG AG
Coat protein This study
MLBVV 899-
880
MLBVV AAA AGT GCA TAC
CTC ATT GC
Coat protein This study
1100F MLBVV TGG GAC TCC AGT
GGT CTT GTA TCC
RNA 2 Hayes et
al. 2006
1407R MLBVV CGA AGG TAG AAA
TAG GAA CGA TGG
RNA2 Hayes et
al. 2006
M13F E. coli CGT CAG GCT TTT
CCC AGT CAC GAC
M13R E. coli TCA CAC AGG AAA
CAG CTA TGA C

30
2.6 Purification of PCR product for cloning or direct sequencing
PCR product was purified by ethanol precipitation as follows:
The following was added to each reaction:
10% vol of 3M NaAC, pH5.2
2.5 volumes of cold 100% ethanol
Then incubated at -20ºC for 15min to precipitate DNA
Precipitated DNA was recovered by centrifugation at 4ºC at 20,000 × g for 10min
Supernatant was discarded before the pellet was washed with 300µL 70% ethanol
Centrifugation again at 4ºC at 20,000 × g for 2min
Supernatant was discarded and the DNA pellet was dried on a 37°C heat block
DNA pellet was resuspended in 10µL ddH20
2.7 DNA and RNA quantification
Samples of DNA and RNA were quantified using a Nanodrop UV-Vis
spectrophotometer (Nanodrop Technologies) following manufacturer‟s protocol.
2.8 Cloning
2.8.1 Ligation using pGEM®
-T Easy Vector
Standard ligation reactions were set up using pGEM®
-T Easy Vector (Promega Corp,
Madison Wisconsin) as recommended in the vector protocol. This was carried out as
follows:

31
5µL of 2 × rapid ligation buffer, 1µL of pGEM®
-T Easy Vector, 3µL of purified
PCR product and 1µL T4 DNA ligase were added and incubated in a 14ºC water
bath overnight.
A negative control was set up using a vector that did not contain an insert.
2.8.2 Transformation using E. coli
After the ligation reaction, the transformation into JM109 competent E. coli cells was
completed as follows:
5µL of the ligation was added to 40µL of JM109 competent cells and incubate
on ice for 30min
Cells were heat shocked in a water bath at exactly 42°C for 45sec
Cells were transferred immediately to ice for 2min before adding 900µL of LB
broth to each tube
Tubes were placed on a shaker at 37°C for 1.5h
After incubation, the cultures were centrifuged @ 20 000 × g for 1min to pellet
the cells
Most of the supernatant was discarded leaving approximately 150µL in the tube. The
pellet was then resuspended before the entire volume of culture was spread on LB plates
containing 100mg/mL ampicillin. The plates were then wrapped in Parafilm and stored
upside-down at 37°C for 12-16h.

32
2.9 Screening for recombinant plasmids
After incubation, the colonies on the plates were screened for recombinant plasmids by
PCR. Ten colonies on each plate were chosen at random and suspended in 50µL of
water. PCR were set up as described in 2.3.1 above and 1µL of cell suspension was
added directly to the reaction mix along with M13 forward and reverse primers (Table
2.1) to amplify the cloned fragments.
The cycling conditions were as follows:
94°C for 3min
Then 25 cycles of:
94°C for 10sec (denaturation)
55°C for 30sec (annealing)
72°C for 1min (extension)
Then held at 14°C
The resulting PCR products were separated on a 1% DNA grade agarose (Fisher
Biotech) gel with TBE electrophoresis buffer and visualised on a transilluminator.
Colonies that contained an insert of the expected size were selected for plasmid
extraction.

33
2.10 Plasmid preparation and analysis
Recombinant plasmids were inoculated into 5mL of LB broth with ampicillin (100
mg/L) and incubated on a shaker at 37ºC overnight. The plasmid was then purified from
the culture using an Aurum™ Bio-Rad Plasmid Extraction Kit following the
manufacturer‟s protocol.
2.11 Restriction digest of plasmid
The presence and size of inserts was confirmed with a restriction digest of the purified
plasmid DNA by EcoR1 as follows:
10µL of purified plasmid DNA was added to 2µL of 10 × EcoR1 buffer, 1µL of
EcoR1 and 7µL of water
Tubes were then incubated on a 37°C heat block for 1h
DNA fragments were separated on a 1% agarose gel beside a 100bp molecular
weight marker
Gel was stained with EtBr and visualised on a transilluminator
2.12 Sequencing
Sequencing was done either directly from PCR product or from plasmids containing an
insert of target DNA. Dideoxy-termination sequencing was performed using the
sequencing facilities at the SABC and Royal Perth Hospital. Applied Biosystems

34
Industries (ABI) Big Dye Version 3.1 chemistry was used using ½, ¼ and ⅛ sequencing
reactions. When a plasmid containing an insert was sequenced, M13F and M13R
primers were used and when sequencing was done directly from a PCR product, the
primers used to amplify the product were used. The ½, ¼ and ⅛ reactions were made in
200µL PCR tubes as follows:
Reagent ½ reaction ¼ reaction ⅛ reaction
Dye terminator mix 4µL 2µL 1µL
5 × buffer - 1µL 1.5µL
3.2pmol/ µL F primer 1µL 1µL 1µL
3.2pmol/ µL R primer 1µL 1µL 1µL
200-500ng plasmid
10-40ng PCR product
Water To 10µL To 10µL To 10µL
The exact amounts of either PCR product or plasmid that was used in each of the
sequencing reactions are listed in Appendix 1. The reactions were then put in the
thermocycler and incubated under the following conditions:
96°C for 2min
Then 25 cycles of:
96°C for 10sec (denaturation)
55°C for 5sec (annealing)
60°C for 4min (extension)
Then held at 14°C
The post-reaction purification was adapted from the Big Dye Terminator Version 3.1
Cycle Sequencing protocol from Applied Biosystems. Each separate sequencing
reaction was purified as follows:

35
In a 0.65mL microcentrifuge tube, 25µL of 100% ethanol, 1µL of 3M sodium
acetate pH5.2 and 1µL of 125mM EDTA were added
The entire 10µL sequencing reaction was added to this and incubated at room
temperature for 20min
Centrifugation @ 20 000 × g for 30min
Supernatant was removed by pipette, making sure no excess liquid was left on
the pellet.
Pre-moulded tissues (Kimwipes) were used to blot excess liquid in the tube
being careful not to touch the pellet at the bottom of the tube.
The pellet was washed by adding 125µL of 70% ethanol
Centrifugation at 20 000 × g for 5min.
Supernatant was removed as before and the pellet dried on a 37°C heat block.
The sequences were loaded in an ABI 377XL capillary sequencing machine.
2.13 Virus detection by Double Antibody Sandwich Enzyme Linked
Immuno-Sorbent Assay (DAS-ELISA)
Leaf and root samples were tested by DAS-ELISA using a protocol developed from
Clarke and Adams, (1977). Reagents and amounts of each reagent used to make buffer
solutions and wash solutions used in this protocol are given in Appendix 2.

36
2.13.1 Coating plates
Nunc 96 well maxisorb plates were used. The inside 60 wells of the plate are used so
that the outside wells could act as a buffer. The amount of coating buffer required was
calculated by the following formula:
N = number of plates to be coated
0.1 is the volume (mL) of solution to be put in each well of the microtitre plate and
66 is the number of wells to be coated.
N × 0.1 × 66 + N = mL coating buffer
The amount of antibody required for the coating step was calculated by:
mL coating buffer × dilution factor of antibody × 1000 = µL antibody
100µL coating buffer/antibody solution was pipetted into each well except outside wells
before 200µL water was pipette into the outside wells. The plates were then incubated
for 4h at 37°C.
2.13.2 Sample Preparation
Plant samples were placed in grinding block with 1000µL Phosphate Buffered Saline
with Tween (PBST). Enough plant material was used to have a sample dilution of 1:20.
The samples in PBST were then ground using ball bearing. The plates were then washed

37
in wash buffer 3 times and left to incubate at room temperature for 3min each time. The
plates were then dried and 200µL of each sample was then pipetted into paired, coated
wells before the plates were left to incubate over night at 4°C.
2.13.3 Conjugation
The amount of PBST (mL) and the amount of conjugate (µL) used in this step was
calculated as described in 2.13.1. Bovine serum albumin (BSA) was then added to this
solution at a ratio of 2% using the following formula:
Amount of BSA (g) = mL PBST × 0.002
100µL PBST/conjugate/BSA solution was then pipetted into each coated well and then
100µL of water into the outside outside wells. The plates were then incubated for 4
hours at 37°C before being washed with wash buffer 3 times and leaving the wash
buffer to incubate at room temperature for 3min each time. Saline was then used for a
final wash and also left to incubate at room temperature for 3min. The plates were then
dried.
2.13.4 Substrate
The volume of substrate buffer (mL) was calculated by the following formula:
N = number of plates to be coated
0.1 is the volume (mL) of solution to be put in each well of the microtitre plate

38
72 is the number of wells to be coated.
N × 72 × 0.2 + N = mL substrate buffer
Para-nitrophenyl phosphate (PNP) was then added to this solution at a ratio of 0.06% by
using the following formula:
Amount of PNP (g) = mL substrate buffer × 0.0006
200µL substrate buffer/PNP solution was then pipette into the first column of wells and
all coated wells. Plates were then incubated at room temperature for 1h before reading
the plate on BioRad Plate Reader.

39
Chapter 3 - Molecular genetic characterization of Olpidium
virulentus isolates associated with big-vein diseased lettuce
plants
L. D. Maccarone, School of Plant Biology, Faculty of Natural and Agricultural
Sciences, University of Western Australia, Stirling Highway, Crawley, WA 6009; M. J.
Barbetti, School of Plant Biology, Faculty of Natural and Agricultural Sciences,
University of Western Australia, Stirling Highway, Crawley, WA 6009, and
Agricultural Research Western Australia, Department of Agriculture and Food, Locked
Bag No.4, Bentley Delivery Centre, WA 6983; K. Sivasithamparam, School of Plant
Biology, Faculty of Natural and Agricultural Sciences, University of Western Australia,
Stirling Highway, Crawley, WA 6009; R. A. C. Jones, School of Plant Biology,
Faculty of Natural and Agricultural Sciences, University of Western Australia, Stirling
Highway, Crawley, WA 6009 and Agricultural Research Western Australia,
Department of Agriculture and Food, Locked Bag No.4, Bentley Delivery Centre, WA
6983
Corresponding author email: [email protected]
Key words: Olpidium brassicae, Chytridiomycete, fungal vector, internal transcribed
spacer sequence, Lettuce big-vein associated virus, Mirafiori lettuce big-vein virus

40
Abstract
Lettuce plants showing symptoms of lettuce big-vein disease were collected from fields
in the Perth Metropolitan region of south-west Australia. When root extracts from each
plant were tested by PCR using primers specific to the rDNA internal transcribed spacer
(ITS) region of Olpidium brassicae and O. virulentus, only O. virulentus was detected
in each of them. The nucleotide sequences of the complete rDNA ITS regions of
isolates from five of the root samples and 10 isolates of O. virulentus from Europe and
Japan showed 97.9 - 100% identities. However, with the six O. brassicae isolates their
identities were 76.9 - 79.4%. On phylogenetic analysis of the complete rDNA-ITS
region sequences of the five Australian isolates and 10 others, the Australian isolates
fitted within two clades of O. virulentus (I and II), and within clade I into two of its four
sub-clades (Ia and Id). Japanese isolates had greatest sequence diversity fitting into
both clades and into all of clade I sub-clades except Ib, while European isolates were
restricted to sub-clades Ib and Id. When the partial rDNA-ITS region sequences of two
additional south-west Australian isolates, four from Europe and four from the Americas
were included in the analyses, the Australian isolates were within O. virulentus sub-
clades Ia and Id, the European isolates within sub-clade Ic, and the American isolates
within sub-clades Ia and Ib. These findings suggest that there may have been at least
three separate introductions of O. virulentus into the isolated Australian continent since
plant cultivation was introduced following its colonization by Europeans. They also
have implications regarding numbers of different introductions to other isolated regions.
Lettuce big-vein associated virus and Mirafiori lettuce big-vein virus were both detected
when symptomatic lettuce leaf tissue samples corresponding to the root samples from
south-west Australia were tested using virus-specific primers in Reverse Transcription-
PCR, so presence of both viruses was associated with O. virulentus occurrence.

41
Introduction
Lettuce big-vein disease (LBVD) was first found in California (10). It occurs widely in
regions of the world with temperate or Mediterranean-type climates and at high altitudes
in subtropical regions. LBVD is now known to be associated with a complex of two
viruses, Lettuce big-vein associated virus (LBVaV; genus Varicosavirus) and Mirafiori
lettuce big-vein virus (MLBVV; genus Ophiovirus) (17,18,19). Its reported natural host
range is limited to lettuce (Lactuca sativa), endive (Cichorium endive), spiny sowthistle
(Sonchus asper) and common sowthistle (Sonchus oleraceus) (2,12). The vector for
both viruses is a root-infecting chytrid belonging to the genus Olpidium
(Chytridiomycota). Motile zoospores of this chytrid transmit MLBVV and LBVaV to
the roots of healthy plants and, in the absence of susceptible hosts, its resting spores
retain ability to transmit for decades in infested soil (1,17,20,23). In addition to
damaging their quality through the presence of obvious „big-vein‟ leaf symptoms
(chlorophyll clearing around leaf veins that appear enlarged) from which the disease
gets its name, LBVD decreases yield of produce from infected plants substantially
(14,15). Symptom expression is most obvious at low temperatures and low light
intensities (2,3,4,17,27,28).
Olpidium spp. are obligate plant pathogens which vector several different soil-
borne plant viruses. They are found commonly throughout the world infecting the roots
of wild and domesticated plants and until recently only two species were recognised as
vectors, O. brassicae and O. bornovanus (= O. radicale) (2,3,9,20). Sahtiyanci (21)
separated O. brassicae (formerly Pleotrachelus brassicae) into two species, O.
brassicae being crucifer-infecting and O. virulentus non-crucifer infecting.
Morphologically, O. brassicae and O. virulentus varied slightly. Pleomorphism was
present in both species with few variations in O. virulentus but more variation in O.

42
brassicae. Also, the zoosporangium of O. brassicae was larger than that of O. virulentus
(21). However, this study was overlooked by researchers investigating LBVD, who
continued to refer to a non-crucifer infecting strain of O. brassicae as the vector of
LBVaV (2,3,4,5,17,20). Recently, molecular and host range studies confirmed that O.
virulentus was sufficiently different to be considered a distinct species that differs from
O. brassicae in infecting lettuce but not Brassica species whereas O. brassicae infects
Brassica species but not lettuce (7,13,23). Nucleotide sequence data for the rDNA
internal transcribed spacer (ITS) region of its Olpidium sp. vector confirmed that it was
O. virulentus that transmitted MLBVV and the Tobacco stunt virus (TStV) strain of
LBVaV to lettuce roots (23).
In Australia, LBVD is a widespread problem in commercial lettuce producing
areas. In south-west Australia, the lettuce production system on the Swan Coastal Plain
is intensive and often uses irrigated land that is contaminated with LBVD, sometimes
without crop rotation. Also, winter plantings often have >90% of plants with symptoms
of the disease (12,14,15). However, no nucleotide sequences of Olpidium spp. are
available from the Australian continent. This paper reports studies that determined the
nucleotide sequences of the rDNA ITS region of south-west Australian Olpidium sp.
isolates obtained from the roots of lettuce plants with LBVD, and compared their
sequence identities and phylogenies with those of the rDNA ITS region of Olpidium
spp. isolates from elsewhere.

43
Materials and Methods
Collection of field samples
Whole lettuce plants showing LBVD symptoms were sampled from two crops growing
in the northern Perth Metropolitan region of south-west Australia. They came from two
different commercial lettuce growing farms, four samples from one farm (labelled Nan
6, Nan 7, Nan 8 and T1 23) and three from the other (labelled Col 1, Col 2 and Col 3).
On the same day, roots from each sample were washed and ground in liquid nitrogen
before DNA was extracted and stored at -20°C.
DNA extraction, PCR and agarose gel electrophoresis
Total DNA was extracted from the lettuce root tissue from samples Nan 6 - 8, T1 23
and Col 1 - 3 using a Qiagen DNeasy Plant Mini Kit (Qiagen Inc., Valencia, CA)
according to manufacturer‟s instructions. No cultures of O. brassicae or O. virulentus
were available to provide positive control extracts. Primers used to amplify target
Olpidium spp. came from White et al. (30) (ITS1 and ITS4) or were designed from
within the rDNA-ITS region by aligning already known sequences using Bioedit (6)
(Ob1, Ob2, Ov1 and Ov2). For amplification of O. brassicae, the primers pairs used
were Ob1 Fwd (5' TCA TTA ATA AAT GCT TCA TTG CAT 3') with ITS4 Rev (5'
TCC TCC GCT TAT TGA TAT GC 3'), and ITS1 Fwd (5' TCC GTA GGT GAA CCT
GCG G 3') (30) with Ob2 Rev (5' TCC AGC AAC AAG TCT TCC CAA 3'). For O.
virulentus amplification, the primer pairs used were Ov1 Fwd (5' CAA GAC CTG CCC
CCA AAA GGG 3') with ITS4, and ITS1 with Ov2 Rev (5' CCA AGT TCG CAA ACG
TGG CG 3'). All primer pair combinations were used on tissue extracts from each
sample. PCR reactions were done using Taq DNA polymerase and 5 × polymerisation
buffer (Fisher Biotec, Perth, Western Australia).

44
PCR amplification conditions consisted of 1 min denaturation at 94°C followed
by 35 cycles of 94°C for 30 sec, 60°C for 30 sec and 72°C for 1 min followed by a final
extension at 72°C for 10 min before being held at 14°C. PCR amplicons were visualised
by electrophoresis on 1% agarose gels. Tris-boric acid-EDTA (TBE) electrophoresis
buffer was used (22).
Virus RNA extraction and Multiplex RT-PCR
Symptomatic leaf tissue samples corresponding to six of the seven root samples were
used for RNA extraction (isolate T1 23 was not available). Total RNA was extracted
from each sample using an UltraClean™ Plant RNA Isolation Kit (MoBio Laboratories,
Inc.) according to manufacturer‟s instructions. RNA extracts were stored at -80°C.
Three primer pairs developed by Navarro et al. (18) were used for multiplex RT-PCR:
VP248 Fwd (5' CGC CAG GAT CTT TGA TCC ATC TG 3') with VP249 Rev (5' TTG
CGA CAT GTT CCT CCT CAT CG 3') to amplify LBVaV, VP286 Fwd (5' TAT CAG
CTC ACA TAC TCC CTA TCG 3') with VP287 Rev (5' CAA CTA GCT CAG AAT
ACA TGC AG 3') to amplify MLBVV, and control primers VP383 Fwd (5' AGC GTG
CTA ATC CCT ATG TTC AT 3') with VP389 Rev (5' AAT GAA AAT CTT AAA
AGC CGT AG 3') to amplify the mitochondrial NADH dehydrogenase subunit 4 (nad4)
of lettuce. Reverse Transcription PCR (RT-PCR) was done using a OneStep RT-PCR
kit (Qiagen, Australia) according to manufacturer‟s instructions.
PCR amplification conditions consisted of 50°C for 30 min, 95°C for 15 min
followed by 30 cycles of 94°C for 1 min, 55°C for 1 min and 72°C for 1 min followed
by a final extension at 72°C for 10 min before being held at 14°C.

45
Cloning and sequencing of PCR amplicons
PCR amplicons of Olpidium were purified by ethanol precipitation and then quantified
using a Nanodrop UV-Vis spectrophotometer following the manufacturer‟s protocol.
Next, standard ligation and transformation reactions were done into JM109 competent
cells using a pGEM®-T Easy Vector (Promega Corp, Madison Wisconsin) as
recommended in the manufacturer‟s protocol.
Screening for recombinant plasmids was done using M13 primers Fwd (5' CGT
CAG GCT TTT CCC AGT CAC GAC 3') and Rev (5' TCA CAC AGG AAA CAG
CTA TGA C 3'). Amplification conditions consisted of 1 min denaturation at 94°C
followed by 25 cycles of 94°C for 30 sec, 55°C for 30 sec and 72°C for 30 sec before
being held at 14°C. Amplicons were then visualised on 1% agarose gels as described
above. Recombinant plasmids were inoculated into 5mL of Luria Bertani (LB) broth
with ampicillin (100 mg/L) and incubated on a shaker at 37°C overnight. Plasmids were
then purified from the culture using an Aurum™ Bio-Rad Plasmid Extraction Kit
following the manufacturer‟s instructions. Presence and size of inserts was confirmed
with a restriction digest of the purified plasmid DNA by EcoR1 (10µL of purified
plasmid DNA, 2µL of 10 x EcoR1 buffer, 1µL of EcoR1 and 7µL of water), incubating
on a 37°C heat block for 1 h and visualising on 1% agarose gel.
Dideoxy-termination sequencing was done at the Western Australian State
Agricultural Biotechnology Centre (SABC), Murdoch University, Perth, or Royal Perth
Hospital, Perth. In both instances, Big Dye Version 3.1 chemistry was used. The
complete rDNA-ITS sequences of isolates Nan 6-8, T1 23 and Col 1 (each 632bp) and
partial sequences of isolates Col 2 (577bp) and Col 3 (503bp) were submitted to
Genbank (Table 1).

46
Analysis of Sequence Data
Nucleotide sequence data for the 21 complete rDNA-ITS regions (632bp) of
Olpidium spp. and the ten partial rDNA-ITS regions (503-577bp) was assembled and
analysed using Molecular Evolutionary Genetics Analysis (MEGA) program Version
4.1 (24). Sequence data for the rDNA ITS region of six full length sequences of O.
brassicae, ten full length sequences of O. virulentus from Europe and Japan, and eight
partial sequences from the Americas and Europe were retrieved from Genbank (Table
1). Isolates SP-010 from Spain (EU981907) and PT-01 from Portugal (EU981906) were
excluded from this analysis as they were only 176bp in length (8).
The complete Olpidium rDNA-ITS region sequences from five Australian
isolates and 16 isolates from other continents were analysed in direct pairwise
comparisons to establish percentage indentities between them. Nucleotide sequence
data for these 21 complete Olpidium rDNA-ITS region sequences were then aligned in
MEGA 4.1 using Clustal W (26) and bootstrapped with 1000 resamplings. This process
was then repeated to also include the two partial sequences from south-west Australia
and the other eight partial sequences. Evolutionary history was inferred using the
Neighbour-Joining method initially for the 21 full length sequences only and then for all
31 sequences. Evolutionary distances were computed using the Maximum Composite
Likelihood method. All positions containing gaps and missing data were eliminated
from the dataset (complete deletion option). Two sequences of O. bornovanus were
then added to the alignment and a radiation diagram constructed to show the relative
genetic distance between the different species of Olpidium reported to be virus vectors.

47
Results
PCR DNA extracts from roots
PCR amplification of the rDNA-ITS nucleotide sequence from all seven root samples
using primers designed specifically for O. virulentus always revealed presence of this
Olpidium species. However, primers designed specifically for O. brassicae produced
only one very faint amplicon from one sample. Subsequent sequencing of this band
failed suggesting that O. brassicae was absent which is consistent with recent findings
that this species does not infect lettuce (7,13,23).
Nucleotide sequence identities
Of the 21 Olpidium complete rDNA-ITS region nucleotide sequences analysed, those of
the five south-west Australian isolates showed greatest nucleotide identity with those of
O. virulentus (four isolates from Europe and six from Japan) (Table 2). Overall, there
was 97.9 - 100% identity between the 15 isolates of O. virulentus, and 99.3 - 100%
identity between the seven isolates of O. brassicae. However, the identity between O.
virulentus and O. brassicae sequences was only 76.9 - 79.4%.
Phylogenetic analysis
The five Olpidium sp. isolates with complete rDNA-ITS nucleotide sequences from
south-west Australia all grouped with O. virulentus (Fig. 1). They fitted into two O.
virulentus clades, I and II. Although the rDNA-ITS region of Olpidium is highly
conserved, it was also possible to distinguish sub clades within clade I despite the small
overall sequence divergence. Four isolates fitted into clade I within two of its four sub-
clades (Ia-d). Nan 6 and Nan 7 grouped together with Japanese isolates KZ-1 and HY-1
within sub-clade Ia while T1 23 and Col 1 grouped with Japanese isolate WOms-3 and
UK isolate GBR1 within sub-clade Id. Sub-clade Ib contained one isolate each from the

48
Netherlands (NLD4), Great Britain (GBR10) and Italy (ITA1). The third sub-clade, Ic,
contained two isolates from Japan (CH-1 and TAK-1). Australian isolate Nan 8 grouped
with Japanese isolate WT-1, and were more distant genetically from the others forming
a distinct clade on their own (clade II).
When the ten partial sequences are added to the phylogeny (Fig. 2), sub-clade Ia
then also included an Australian isolate (Col 3) and one from Guatemala (GT-08), sub-
clade Ib contained two isolates from Mexico (MX-05 and MX-06) and one from the
USA (633), four isolates from Spain (SP-06 to SP-09) were in sub-clade Ic, and the
remaining Australian isolate (Col 2) fitted into sub-clade Id. None of the partial
sequences fell within clade II.
When the relative genetic distances between the three viral vector species of
Olpidium were compared, O. virulentus and O. brassicae were much closer to one
another than either was to O. bornovanus (Fig. 3).
RT-PCR RNA extracts from leaves
When leaf tissue from samples Nan 6-8 and Col 1 were tested by RT-PCR using
primers specific for LBVaV or MLBVV, a 496bp fragment was amplified indicating
presence of MLBVV, a 292bp fragment indicating presence of LBVaV and a 360bp
control fragment indicating presence of the nad4 gene of lettuce (Fig. 4). This shows
that both viruses were present in each of the samples. Leaf tissue from samples Col 2
and Col 3 were also tested using the same method. Col 2 contained both viruses whilst
MLBVV was the only virus present in Col 3. As O. virulentus, but not O. brassicae,
was associated with the two viruses in the same lettuce plants, this indicates that it may
have been responsible for the transmission of LBVaV and MLBVV on the two farms
originally sampled.

49
Discussion
This study revealed that O. virulentus, but not O. brassicae, was associated with
samples of lettuce plants with LBVD collected from two farms in south-west Australia,
and that the same samples were infected either with LBVaV and MLBVV, or MLBVV
alone. This association of O. virulentus with the two viruses is consistent with previous
research which suggested that O. virulentus, rather than O. brassicae, was the chytrid
vector of MLBVV and the TStV strain of LBVaV (23), and that O. brassicae does not
infect lettuce (7,13,23). However, it contrasts with previous publications (e.g.
4,17,25,27) which named O. brassicae as vector of LBVaV and MLBVV.
Sasaya and Koganezawa (23) reported that the extent of nucleotide sequence
identity in the complete rDNA-ITS region of O. virulentus and O. brassicae isolates
was 98.5 – 100% and 99.3 - 100%, respectively, while it was 79.7-81.8% between the
two species. This is consistent with the percentage identities found in our study within
isolates of O. virulentus from Australia, Europe and Japan (97.9 - 100%), and of O.
brassicae from Europe and Japan (99.3 – 100%). However, the percentage rDNA-ITS
region nucleotide sequence identity between O. brassicae and O. virulentus isolates that
we found was somewhat greater (76.9 - 79.4%). This degree of sequence divergence
supports the conclusion made by Sahtiyanci (21), based on morphological evidence, and
Sasaya and Koganezawa (23), based on molecular evidence, that they are distinct
Olpidium species, not simply different strains of the same species. When the nucleotide
sequences from the complete rDNA-ITS region of five O. virulentus isolates from
samples of lettuce roots from Australia and those of the complete rDNA-ITS region of
10 O. virulentus isolates from Europe and Japan were analysed phylogenetically, two
clades were found (I and II) and, within clade I, four sub-clades were distinguishable
(Ia-Id). Australian isolates grouped within both clades and within clade I, sub-clades Ia

50
and Id. Japanese isolates fitted within both clades and in all sub-clades except 1b, while
European isolates only fitted in sub-clades Ib and 1d. When the partial rDNA-ITS
region sequences of two additional south-west Australian isolates, four from Europe and
four from the Americas were included in the analyses, the Australian isolates were
within sub-clades Ia and Id, the European isolates within sub-clade Ic, and the American
isolates within sub-clades Ia and Ib One of the Australian farms sampled contained
isolate sequences that fitted not only into both of the clades but also into two sub-clades
(Ia and Id) while the second farm contained isolates within the same sub-clades. This
suggests that since European settlement of south-west Australia more than 160 years
ago and the subsequent introduction of plant cultivation to the region, three separate
introductions of O. virulentus have occurred. The data indicate that there have been four
separate introductions to Japan. Although Japanese isolates showed the greatest
nucleotide sequence diversity for the rDNA-ITS region of O. virulentus, this does not
mean that this species originated in Japan as it is not a recognised centre of crop
domestication. Lettuce was probably domesticated first in Egypt (16). From there,
lettuce cultivation spread through the Mediterranean region, first into Italy and then to
Greece (29). Assuming that O. virulentus evolved with lettuce in its original centre of
domestication within the Mediterranean Basin, greater nucleotide sequence diversity of
O. virulentus would be expected in this region than elsewhere. However, unlike
Japanese and Australian isolates, European isolates did not fit into both clades, only into
three sub clades of clade I. Sequencing of more isolates from the Mediterranean region
and Europe in general is required to reveal the extent of diversity in the O. virulentus
population in that part of the world. Further sequencing of the rDNA-ITS region of O.
virulentus isolates from the Americas is needed to comment on the number of possible

51
introductions there, but more sequence diversity would be expected than that in
Australia given the longer history of plant cultivation since European colonization.
Methyl bromide was formerly used widely as a fungicide to control Olpidium
spp., but is no longer available in some countries, including Australia. Environmentally
friendly cultural (agronomic) and phytosanitary control measures against Olpidium
vectors were included within the LBVD integrated disease management strategies for
nurseries, nutrient film, infested land and uninfested land of Jones (12). For example,
when infested land is used, decreasing irrigation and /or placing plastic mulch on the
soil surface when overhead irrigation is used diminishes the mobility of viruliferous O.
virulentus zoospores (14).
Acknowledgements
We thank Stephen Wylie and Craig Webster for their advice on nucleotide sequencing,
local farmers for allowing sampling on their farms, and Frances Brigg and Julie
Uhlmann for operating sequencing machines and the SABC for providing laboratory
facilities. Linda Maccarone received an Australian Postgraduate Award (Industry)
jointly funded by the Australian Research Council and the Department of Agriculture
and Food Western Australia.

52
Literature Cited
1. Barr, D. J. S. 1988. Zoosporic plant parasites as fungal vectors of viruses: taxonomy
and life cycle of species involved. Pages 123-137 in: Viruses with fungal vectors. J.
I. Cooper and M.J.C. Asher, eds. Association of Applied Biologists, Wellesbourne,
UK.
2. Campbell, R.N. 1985. Longevity of Olpidium brassicae in air-dried soil and the
persistence of the lettuce big-vein agent. Can. J. Bot. 63: 2288-2289.
3. Campbell, R. N. 1996. Fungal transmission of plant viruses. Ann. Rev. Phytopathol.
34: 87-108.
4. Campbell, R. N. and Grogan R. G. 1964. Acquisition and transmission of lettuce
big-vein virus by Olpidium brassicae. Phytopathology 54: 681-690.
5. Campbell, R. N., and Lin M. T., 1976. Morphology and thermal death point of
Olpidium brassicae. Am. J. Bot. 63: 826-832.
6. Hall, T. A. 1999. BioEdit: a user-friendly biological sequence alignment editor and
analysis program for Windows 95/98/NT. Nucl. Acid. Sym. Ser. 41: 95-98.
7. Hartwright, L. M., Hunter, P. J., and Walsh J. A. 2009. A comparison of Olpidium
isolates from a range of host plants using internal transcribed spacer sequence
analysis and host range studies. Mycol. Res. (in press) doi:
10.1016/j.mycres.2009.09.008.
8. Herrera-Vasquez, J. A., Cebrian, M. C., Alfaro-Fernandez, A., Cordoba-Selles, M.
D., and Jorda, C. (2009). Multiplex PCR assay for the simultaneous detection and

53
differentiation of Olpidium bornovanus, Olpidium brassicae and Olpidium
virulentus. Mycol. Res. 113:602-610
9. Herrera-Vasquez, J. A., Cordoba-Selles, M. C., Cebrian, M. C., Rosello, J. A., and
Jorda, C. 2010. Genetic diversity of Melon necrotic spot virus and Olpidium isolates
from different origins. Plant Pathol. (in press)
10. Jagger, I. C., and Chandler, N. 1934. Big-vein, a disease of lettuce. Phytopathology
24: 1253-1256.
11. James, T. Y., Letcher, P. M., Longcore, J. E., Mozley-Standridge, S. E., Porter, D.,
Powell, M. J., Griffith, G. W., and Vilgalys, R. (2006) A molecular phylogeny of
flagellated fungi (Chytridiomycota) and description of a new phylum
(Blastocladiomycota). Mycologia 98:860-871.
12. Jones, R. A. C., 2004. Using epidemiological information to develp effective
integrated virus disease management strategies. Virus Res. 100: 5-30
13. Koganezawa, H., Inoue, H., and Sasaya, J. 2005. Host specificity and multiplication
of eight isolates of Olpidium brassicae sensu lato and its related Olpidium spp. Bull.
Natl. Agric. Res. Cent. West Reg. 4:39-40.
14. Latham, L. J., and Jones, R. A. C. 2004. Deploying partially resistant genotypes and
plastic mulch on the soil surface to suppress spread of lettuce big-vein disease in
lettuce. Aust. J. Agr. Res. 55: 131-138
15. Latham, L. J., Jones, R. A. C., and McKirdy, S. J. 2004. Lettuce big-vein disease:
sources, patterns of spread, and losses. Aust. J. Agr. Res. 55: 125-130
16. Lindqvist K, 1960. On the origin of cultivated lettuce. Hereditas 46: 319-350

54
17. Lot, H., Campbell, R. N., Souche, S., Milne, R. G., and Roggero, P. 2002.
Transmission by Olpidium brassicae of Mirafiori lettuce virus and Lettuce big-vein
virus, and their roles in lettuce big-vein etiology. Phytopathology 92: 288-293.
18. Navarro, J. A., Botella, F., Maruhenda, A., Sastre, P., Sànchez-Pina, M. A., and
Pallas. V. 2004. Comparative infection progress analysis of Lettuce big-vein virus
and Mirafiori lettuce virus in lettuce crops by developed molecular diagnosis
techniques. Phytopathology 94: 470-477
19. Roggero, P., Ciuffo, M., Vaira, A. M., Accotto, G. P., Masenga, V., and Milne, R.
G., 2000. An ophiovirus isolated from lettuce with big-vein symptoms. Arch. Virol.
145: 2629-2642
20. Rochon, D‟A., Kakani, K., Robbins, M., and Reade, R. 2004. Molecular aspects of
plant virus transmission by Olpidium and Plasmodiophorid vectors. Ann. Rev.
Phytopathol. 42: 211-241.
21. Sahtiyanci, S. 1962. Studien über einige wurzelparasitäre Olpidiaceen. Arch.
Microbiol. 41: 187–228.
22. Sambrook, J., and Russell, D. W. 2001. Molecular cloning: A laboratory manual.
New York: Cold Spring Harbour Laboratory Press.
23. Sasaya, T., and Koganezawa, H. 2006. Molecular analysis and virus transmission
tests place Olpidium virulentus, a vector of Mirafiori lettuce big-vein virus and
Tobacco stunt virus, as a distinct species rather than a strain of Olpidium brassicae.
J. Gen. Plant Pathol. 72: 20-25.

55
24. Tamura, K., Dudley, J., Nei, M., and Kumar, S. 2007. MEGA4: Molecular
Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 24:
1596-1599.
25. Teakle, D. S. 1960. Association of Olpidium brassicae and tobacco necrosis virus.
Nature 188: 431-432.
26. Thompson, J. D., Higgins, D. G., and Gibson, T. J. 1994. CLASTAL W: Improving
the sensitivity of progressive multiple sequence alignment through sequence
weighting, position-specific gap penalties and weight matrix choice. Nucl. Acid.
Res. 22: 4673-4680.
27. Walsh, J. A. 1994. Effects of some biotic and abiotic factors on symptom expression
of lettuce big-vein virus in lettuce (Lactuca sativa). J. Hortic. Sci. 69: 21-28.
28. Westerlund, F. V., Campbell, R. N., and Grogan, R. G. 1978. Effect of temperature
on transmission, translocation, and persistence of the lettuce big-vein agent and big-
vein symptom expression. Phytopathology 68: 921-926.
29. Whitaker, T. W. 1969. Salads for everyone – A look at the lettuce plant. Econ. Bot.
23: 261-264
30. White, T. J., Bruns, T., Lee, S., and Taylor, J. W. 1990. Amplification and direct
sequencing of fungal ribosomal RNA genes for polygenetics. Pages 315-322 in:
PCR Protocols: A Guide to Methods and Applications. M. A. Inns, D. H. Gelfand, J.
J. Sninsky and T. J. White, eds., Academic Press, Inc., New York.

56
Table 1: Details of isolates of three vector Olpidium spp. used to determine relationships
using the nucleotide sequences of their rDNA ITS regions
Species Accession
number
Isolate Geographic origin Isolated from root
of:
Reference
O. bornovanus AB205214 NA Nagasaki, Japan Melon 23
O. bornovanus AB205215 CH Chiba, Japan Melon 23
O. brassicae AB205209 CBG-3 Nagano, Japan Cabbage 23
O. brassicae AB205210 NKa-3 Nagano, Japan Cabbage 23 O. brassicae AB205211 YR-2 Shimane, Japan Cabbage 23
O. brassicae AB205212 ZE-1 Zentsuji, Japan Cabbage 23
O. brassicae AB205213 EH-2 Iyo, Japan Cabbage 23
O. brassicae AY373015 GBR7 Warwickshire, UK Cauliflower 7
O. virulentus AB205203 WT-1 Wakayama, Japan Lettuce 23
O. virulentus AB205204 HY-1 Hyougo, Japan Lettuce 23 O. virulentus AB205205 KZ-1 Kagawa, Japan Lettuce 23
O. virulentus AB205206 TAK-1 Takamatsu, Japan Tobacco 23
O. virulentus AB205207 CH-1 Chunan, Japan Tobacco 23
O. virulentus AB205208 WOms-3 Toyama, Japan Welsh onion 23 O. virulentus AY373011 GBR1 Warwickshire, UK Lettuce 7
O. virulentus AY373012 ITA1 Mirafiori, Italy Lettuce 7
O. virulentus AY373013 NLD4 De Lier, the
Netherlands
Lettuce
7
O. virulentus AY373014 GBR10 Humberside, UK Cucumber 7
O. virulentus AY997067 633 Duke University
Herbarium, USA
unknown 11
O. virulentus EU981898 GT-8 Zacapa, Guatemala Cantaloupe 9 O. virulentus EU981899 MX-06 Sinaloa, Mexico Cantaloupe 9
O. virulentus EU981900 MX-05 Colima, Mexico Cantaloupe 9
O. virulentus EU981901 SP-08 Castellon, Spain Lettuce 9
O. virulentus EU981902 SP-09 Murcia, Spain Tomato 9 O. virulentus EU981903 SP-07 Alicante, Spain Watermelon 9
O. virulentus EU981905 SP-06 Seville, Spain Cantaloupe 9
O. virulentus GQ304519 Nan6 Perth, Western
Australia
Lettuce This study
O. virulentus GQ304520 Nan7 Perth, Western
Australia
Lettuce This study
O. virulentus GQ304521 Nan8 Perth, Western
Australia
Lettuce This study
O. virulentus GQ304522 T123 Perth, Western
Australia
Lettuce This study
O. virulentus GQ304523 Col1 Perth, Western
Australia
Lettuce This study
O. virulentus GQ328050 Col2 Perth, Western
Australia
Lettuce This study
O. virulentus GQ328051 Col3 Perth, Western
Australia
Lettuce This study
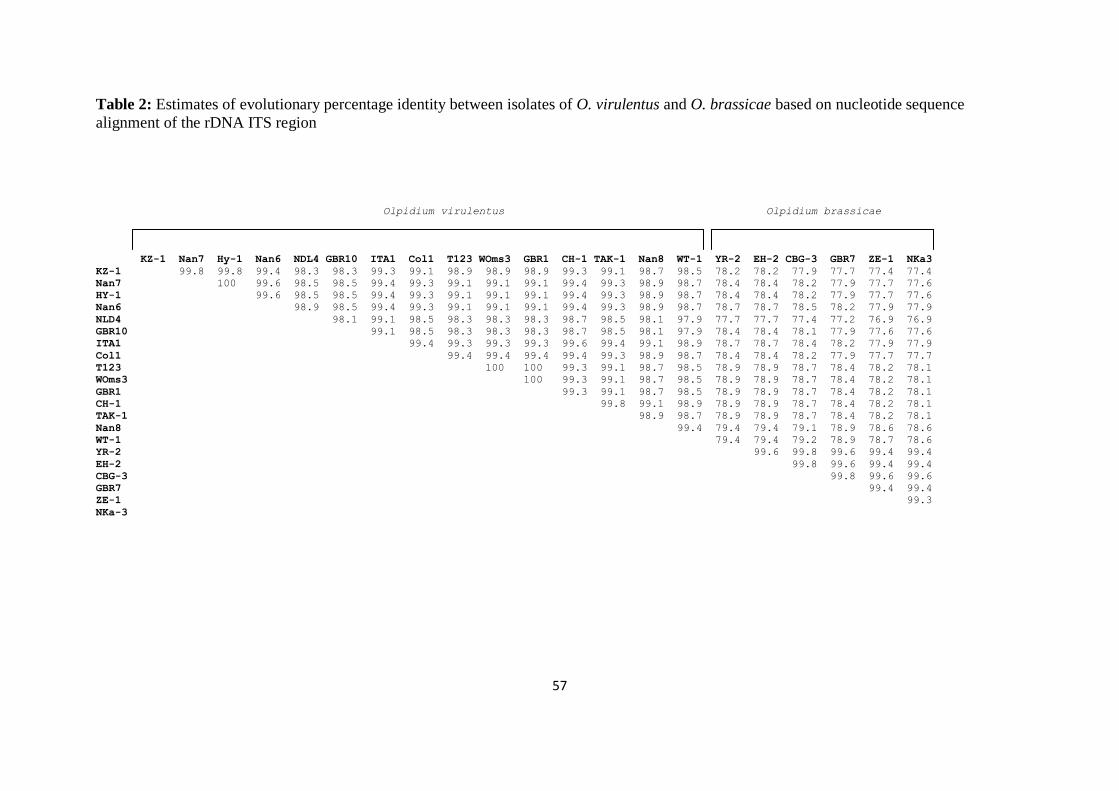
57
Table 2: Estimates of evolutionary percentage identity between isolates of O. virulentus and O. brassicae based on nucleotide sequence
alignment of the rDNA ITS region
Olpidium virulentus Olpidium brassicae
KZ-1 Nan7 Hy-1 Nan6 NDL4 GBR10 ITA1 Col1 T123 WOms3 GBR1 CH-1 TAK-1 Nan8 WT-1 YR-2 EH-2 CBG-3 GBR7 ZE-1 NKa3
KZ-1 99.8 99.8 99.4 98.3 98.3 99.3 99.1 98.9 98.9 98.9 99.3 99.1 98.7 98.5 78.2 78.2 77.9 77.7 77.4 77.4
Nan7 100 99.6 98.5 98.5 99.4 99.3 99.1 99.1 99.1 99.4 99.3 98.9 98.7 78.4 78.4 78.2 77.9 77.7 77.6
HY-1 99.6 98.5 98.5 99.4 99.3 99.1 99.1 99.1 99.4 99.3 98.9 98.7 78.4 78.4 78.2 77.9 77.7 77.6
Nan6 98.9 98.5 99.4 99.3 99.1 99.1 99.1 99.4 99.3 98.9 98.7 78.7 78.7 78.5 78.2 77.9 77.9
NLD4 98.1 99.1 98.5 98.3 98.3 98.3 98.7 98.5 98.1 97.9 77.7 77.7 77.4 77.2 76.9 76.9
GBR10 99.1 98.5 98.3 98.3 98.3 98.7 98.5 98.1 97.9 78.4 78.4 78.1 77.9 77.6 77.6
ITA1 99.4 99.3 99.3 99.3 99.6 99.4 99.1 98.9 78.7 78.7 78.4 78.2 77.9 77.9
Col1 99.4 99.4 99.4 99.4 99.3 98.9 98.7 78.4 78.4 78.2 77.9 77.7 77.7
T123 100 100 99.3 99.1 98.7 98.5 78.9 78.9 78.7 78.4 78.2 78.1
WOms3 100 99.3 99.1 98.7 98.5 78.9 78.9 78.7 78.4 78.2 78.1
GBR1 99.3 99.1 98.7 98.5 78.9 78.9 78.7 78.4 78.2 78.1
CH-1 99.8 99.1 98.9 78.9 78.9 78.7 78.4 78.2 78.1
TAK-1 98.9 98.7 78.9 78.9 78.7 78.4 78.2 78.1
Nan8 99.4 79.4 79.4 79.1 78.9 78.6 78.6
WT-1 79.4 79.4 79.2 78.9 78.7 78.6
YR-2 99.6 99.8 99.6 99.4 99.4
EH-2 99.8 99.6 99.4 99.4
CBG-3 99.8 99.6 99.6
GBR7 99.4 99.4
ZE-1 99.3
NKa-3

58
KZ-1
Nan 7
HY-1
Nan 6
NLD4
GBR10
ITA1
CH-1
TAK-1
Col 1
T1 23
WOms-3
GBR1
Nan 8
WT-1
YR-2
GBR7
EH-2
CBG-3
ZE-1
NKa-3 85 55
100
86
84
66
51
56
68
0.02
I
II
Ia
Ib
Ic
Id
Olpidium
virulentus
Olpidium
brassicae
Fig 1: Unrooted neighbour-joining relationship dendrogram for the complete 632bp
rDNA ITS nucleotide sequences of five Olpidium isolates sequenced in this study and
16 others from Europe and Japan available on Genbank. The phylogenetic tree was
generated using MEGA 4.1 using the default parameters. The tree branches were
bootstrapped with 1000 replications. Numbers at nodes indicate bootstrap scores
>50%. The scale bar represents a genetic distance of 0.02 for horizontal branch
lengths. Table 1 shows isolate designations.

59
KZ-1
Nan 7
HY-1
GT-08
Col 3
Nan 6
NLD4
ITA1
MX-O5
MX-O6
GBR10
633
SP-O6
SP-O7
CH-1
TAK-1
SP-O9
SP-O8
Col 2
Col 1
WOms-3
GBR1
T1 23
Nan 8
WT-1
YR-2
EH-2
CBG-3
GBR7
ZE-1
NKa-3 84
58
56
99
97
86
81
68
64
50
68
0.02
I
II
Ia
Ib
Ic
Id
Olpidium
virulentus
Olpidium
brassicae
Fig 2: Unrooted neighbour-joining relationship dendrogram for rDNA ITS nucleotide
sequences of the five complete and two partial Olpidium isolates sequenced in this
study and 16 complete and eight partial ones from Europe, Japan and the Americas
available on Genbank. The phylogenetic tree was generated using MEGA 4.1 using
the default parameters. The tree branches were bootstrapped with 1000 replications.
Numbers at nodes indicate bootstrap scores >50%. The scale bar represents a genetic
distance of 0.02 for horizontal branch lengths. Table 1 shows isolate designations.
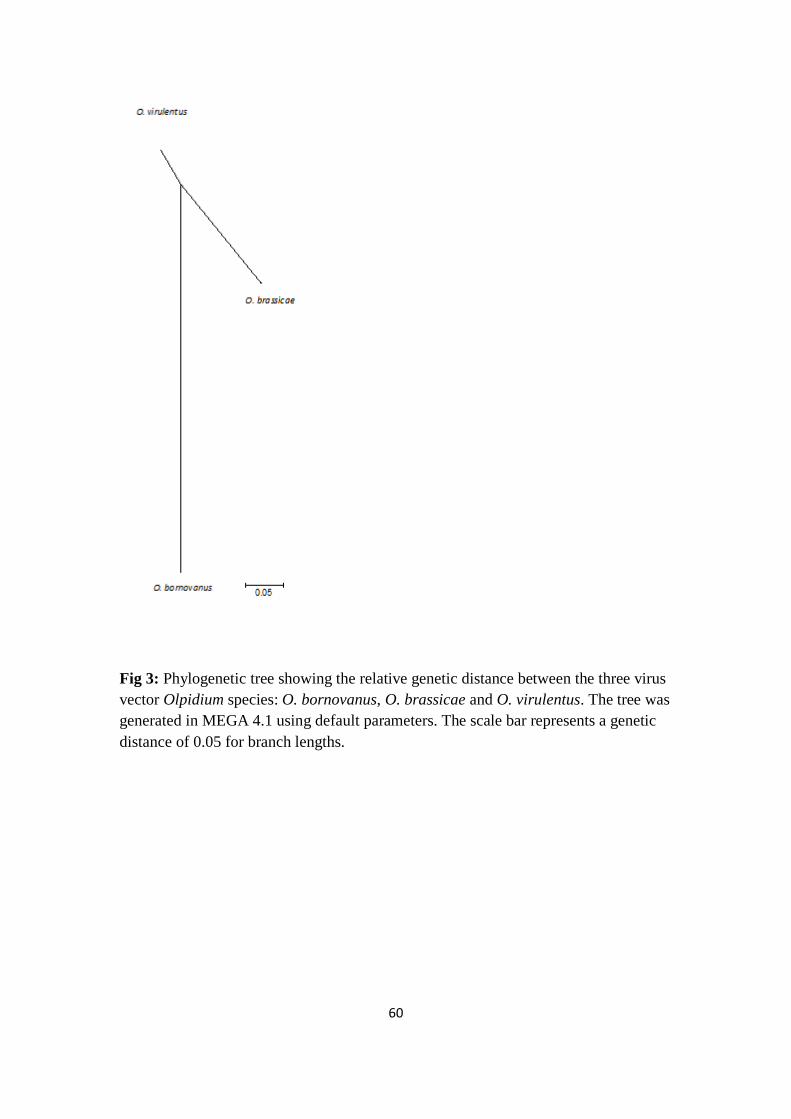
60
Fig 3: Phylogenetic tree showing the relative genetic distance between the three virus
vector Olpidium species: O. bornovanus, O. brassicae and O. virulentus. The tree was
generated in MEGA 4.1 using default parameters. The scale bar represents a genetic
distance of 0.05 for branch lengths.

61
Fig 4: Agarose gel resulting from a multiplex RT-PCR to detect the presence of
MLBVV, LBVaV and control gene nad4 in four samples of lettuce with LBVD from
south-west Australia. All samples had mixed infection with LBVaV and MLBVV,
and were positive for nad4. Lane 1: isolate Nan 6, Lane 2: Nan 7, Lane 8: Nan 8,
Lane 4: Col 1. Lane M represents a 100bp molecular weight marker.
496bp MLBVV
360bp nad4
292bp LBVaV
M 1 2 3 4

62
Chapter 4 - Comparison of the coat protein genes of
Mirafiori lettuce big-vein virus isolates from Australia with
those of isolates from other continents
Linda D. Maccarone1, Martin J. Barbetti
1, Krishnapillai Sivasithamparam
1,
Roger A. C. Jones1,2,3
1School of Plant Biology, Faculty of Natural and Agricultural Sciences, University of
Western Australia, Crawley, WA 6009, Australia
2 Department of Agriculture and Food, Locked bag 4, Bentley Delivery Centre, WA
6983, Australia
3Corresponding author email: [email protected]
Introduction
Mirafiori lettuce big-vein virus (MLBVV; genus Ophiovirus) commonly occurs in
lettuce plants in mixed infection with Lettuce big-vein associated virus (LBVaV;
genus Varicasavirus) [e.g., 4, 5, 6, 9]. Both are transmitted by Olpidium virulentus [4,
10]. MLBVV causes lettuce big-vein disease (LBVD), decreasing lettuce yield and
quality [e.g., 3, 9]. It has filamentous, circularised particles [2, 9, 11]. Its known
natural host range includes lettuce, endive (Cichorium endive), spiny sowthistle
(Sonchus asper) and common sowthistle (Sonchus oleraceus) [3, 7]. When Navarro
et al. [8] analysed the complete CP nucleotide sequences of 17 MLBVV isolates
mostly from Europe, they found identities of 88.0 to 99.8% (nucleic acids) and 92.9-
99.8% (amino acids), and when these sequences were analysed phylogenetically, two
isolate groupings were found (A and B). When three further European sequences

63
from sowthistle were included, they fitted into group B [7]. This paper compares the
CPs of 13 previously unreported isolates from Australia with those of 26 previously
sequenced isolates from different continents.
Materials and Methods
Leaves with LBVD symptoms were sampled from seven farms (A-G) in four districts
in the Perth Metropolitan region. Thirteen samples were collected, one each from
farms B, C, D and F, two each from A and G, and five from E (Supplementary Fig. 1).
Procedures and conditions used for sample storage, RNA extraction, RT-PCR, PCR
amplification, electrophoresis on agarose gels, amplicon quantification, cloning and
sequencing were described previously [5]. The CP primer pairs specific for MLBVV
used were those of Navarro et al. [8], fwd VP286 (5' TAT CAG CTC ACA TAC TCC
CTA TCG 3') and rev VP289 (5' ACA AAG TCA AAT GTC AGG AG 3'), and
primers developed with sequences from GenBank using Bioedit, fwd M2-21 (5' CAA
AGT CAC AAT GTC AGG AG 3') and rev M899-880 (5' AAA AGT GCA TAC
CTC ATT GC 3'). The PCR reaction was done with an annealing temperature of
55°C for both primer pairs. Complete CP sequences of were submitted to GenBank
(Table 1). They contained one ORF 1,311bp in length encoding 437 amino acids. In
addition, 23 complete and three partial CP sequences of other isolates were retrieved
from GenBank. The partial sequences were DQ530356 and DQ530358 (both
1261bp), and DQ530357 (1260bp) from Brazil. Other partial MLBVV sequences
available were too short to use. The 36 complete CP sequences were assembled and
analysed for nucleotide and amino acid identities, and phylogeny as described
previously [5]. This process was then repeated to include the partial sequences. Next,

64
with the complete CP sequences, the mean number of non-silent substitutions (Dn)
and silent substitutions (Ds), and Dn/Ds ratios were determined [5].
Sequence properties
When the 36 complete CP nucleotide sequences were analysed phylogenetically, all
were within clades A and B. Also, each clade contained two sub-clades (Fig. 1;
Supplementary Fig. 2). This clade sub-division was also evident in earlier trees, in
clade A [8] and both clades [7]. Phylogenetic analysis of amino acid sequences from
the 36 complete CPs revealed a similar pattern. Sub-clade A1 included 12 of the 13
new Australian isolates, the Australian isolate sequenced previously [AUS1], one
Japanese isolate and six European isolates; A2 included new Australian isolate
AUSE2, two South American isolates (from Argentina) and six European isolates;
while B1 and B2 included two and five European isolates (all from Spain),
respectively. Despite being in a different sub-clade, isolate AUSE2 was originally
collected from the same farm as isolates AUSE1, AUSE3, AUSE4 and AUSE6.
Spanish isolates SON1-4 in sub-clade B2 were from sowthistle while all other isolates
in both clades were from lettuce. The three partial nucleotide sequences from Brazil
all grouped with the two complete South American sequences in sub-clade A2.
The nucleotide sequence identities of the 14 Australian isolates were 96.3–
99.9%. New isolate AUSE2 diverged most from the other 13 which had 99.1-99.9%
identities. AUSE2 was closest to the three Brazilian isolates in sub-clade A2, while
the other Australian isolates were closer to the European and Japanese isolates in sub-
clade A1. The nucleotide sequence identities of all 36 complete CPs were 85.4–
100%. Those of clade A and B isolates were 95.8–99.9% and 98.4–100%,
respectively. Isolate pairs SON1 and SON4, and SON3 and ALM5 in sub-clade B2

65
had identical sequences. Thus, the degree of nucleotide sequence divergence
resembled that reported previously, 88.0-99.8% (all isolates), 95.8-99.5% (clade A)
and 98.7-99.8% (clade B) [8]. The sequence identities of sub-clades were 98.1-99.9%
(A1), 96.1-99.5% (A2), 99.8% (B1) and 98.4-100% (B2). Overall, the identities of
Spanish isolates SON1 and SON4 (in sub-clade B2) and ALM3 (in sub-clade A2)
were most divergent, differing by 85.4%. Amino acid identities were 92.8 to 100%
(all isolates), 95.3 to 100% (clade A) and 99.1 to 100% (clade B), again resembling
values reported previously, 92.9-99.8% (all isolates), 95.8-99.8% (clade A) and 99.3-
99.5% (clade B) [8].
Assuming that MLBVV evolved with lettuce within its domestication centre in
the Mediterranean region, greatest genetic diversity would be expected there. This
was reflected in the results with the 11 Spanish isolates which were in all sub-clades
apart from A1. Occurrence of Australian isolates within two sub-clades (A1 and A2)
suggests two separate introductions. Presence of European isolates in both of these
sub-clades might reflect introduction from there. Since few isolates from Asia and
South America have been sequenced so far, their occurrence only in sub-clades A1
and A2, respectively, presumably reflects insufficient sequencing rather than single
introductions. No African isolates have been sequenced and the two partial North
American CP sequences on GenBank (DQ182564 and DQ191062) [1] are too short
(409bp) to draw conclusions.
Navarro et al. [8] reported mean Dn/Ds ratios for MLBVV CP sequences of
0.095 (all 17 sequences), 0.153 (clade A) and 0.211 (clade B). Here, the mean Dn/Ds
ratios were 0.112 (all 36 complete CPs), 0.076 (Australian isolates), 00.187 (clade A)
and 0.063 (clade B). Thus, most nucleotide substitutions are silent, mean Dn/Ds ratios
<1 suggesting selection for amino acid conservation. As overall mean Dn/Ds ratio for

66
clade B was lower than for clade A, this suggests more positive selection pressure on
the nucleotide sequence of clade A. Navarro et al. [8] based their Dn/Ds ratio for
clade B on only three CP amino acid sequences which might explain the difference
between our ratio (0.063) and theirs (0.211).
The 39 amino acid CP sequences varied considerably between clades A and B,
especially in their N-terminus and central regions (Supplementary Table 1). There
were 93 amino acid substitutions across the entire sequence, 18 of which were only in
clade B. Of these 18, 14 were in all clade B isolates, two only in sub-clade B1 isolates
ALM4 and ALM1 (codons 114 and 215), and one each only in ALM1 (codon 181)
and sub-clade B2 isolate SON2 (codon 97). Codon 305 showed a change from Ile to
Val present in all isolates except MUR1, LP1 and LP2 in sub-clade A2 and all isolates
in clade B. The 13 Australian isolates had only eight substitutions overall, which,
unlike those in the other isolates, varied most in their C-terminal regions (codon
positions 287-414). These changes were in AUSE6 at codon 7; AUSF1 at codons 118,
287 and 414; AUSG2 at codon 291; AUSA2 at codon 329; AUSA1 at codon 338; and
AUSE1 at codon 390. Isolate AUSE2 had two substitutions which were common to
most sub-clade A2 isolates at codons 305 (Ile to Val) and 314 (Thr to Ile), but absent
from sub-clade A1.
Acknowledgements
We thank Stephen Wylie, Craig Webster, Rohan Prince and Denis Phillips for advice,
the State Agricultural Biotechnology Centre for laboratory facilities and local farmers
for allowing sampling. L.M. received an Australian Postgraduate Award Industry
from the Australian Research Council.

67
References
1. Hayes RJ, Wintermantel WM, Nicely PA, Ryder EJ (2006) Host resistance to
Mirafiori lettuce big-vein virus and Lettuce big-vein associated virus and virus
sequence diversity and frequency in California. Plant Dis 90:233-239
2. Kawazu Y, Sasaya T, Morikawa T, Sugiyama K, Natsuaki T (2003) Nucleotide
sequence of the coat protein gene of Mirafiori lettuce virus. J Gen Plant Pathol
69:55–60
3. Latham LJ, Jones RAC, McKirdy SJ (2004) Lettuce big-vein disease: sources,
patterns of spread, and losses. Aust J Agric Res 55:125-130
4. Maccarone LD, Barbetti MJ, Sivasithamparam K, Jones RAC (2010) Molecular
genetic characterization of Olpidium virulentus isolates associated with big-vein
diseased lettuce plants. Plant Disease 94:563-569.
5. Maccarone LD, Barbetti MJ, Sivasithamparam K, Jones RAC (2010) Comparison
of the coat protein genes of Lettuce big-vein associated virus isolates from
Australia with those of isolates from other continents. Arch. Virol. (in press) DOI.
10.1007/s00705-010-0641-0
6. Navarro JA, Botella F, Maruhenda A, Sastre P, Sanchez-Pina MA, Pallas V (2004)
Comparative infection progress analysis of Lettuce big-vein virus and Mirafiori
lettuce virus in lettuce crops by developed molecular diagnosis techniques.
Phytopathology 94:470-477
7. Navarro JA, Botella F, Maruhenda A, Sastre P, Sanchez-Pina MA, Pallas V
(2005a) Identification and partial characterisation of Lettuce big-vein associated
virus and Mirafiori lettuce big-vein virus in common weeds found amongst
Spanish lettuce crops and their role in lettuce big-vein disease transmission.
European J Plant Path 113:25-34

68
8. Navarro JA, Torok VA, Vetten HJ, Pallas V (2005b) Genetic variability in the coat
protein genes of Lettuce big-vein associated virus and Mirafiori lettuce big-vein
virus. Arch Virol 150:681-694
9. Roggero P, Ciuffo M, Vaira AM, Accotto GP, Masenga V, Milne RG (2000) An
ophiovirus isolated from lettuce with big-vein symptoms. Arch Virol 145:2629-
2642
10. Sasaya T, Koganezawa H (2006) Molecular analysis and virus transmission tests
place Olpidium virulentus, a vector of Mirafiori lettuce big-vein virus and Tobacco
stunt virus, as a distinct species rather than a strain of Olpidium brassicae. J Gen
Plant Pathol 72:20-25
11. van der Wilk F, Dullemans AM, Verbeek M, van den Heuvel JFJM (2002)
Nucleotide sequence and genomic organization of an ophiovirus associated with
lettuce big-vein disease. J Gen Virol 83:2869–2877

69
Table 1. Isolates of Mirafiori lettuce big-vein virus used in sequence comparisons
Virus isolate Accession
number
Isolate location Species
isolated from
Reference
LS301-0 AF525935 Netherlands Lettuce [11]
JPN1 AF532872 Hyogo, Japan Lettuce [2]
MUR1 AY366415 Murcia, Spain Lettuce [8]
GAL1 AY366416 Galicia, Spain Lettuce [8]
ALM1 AY366417 Almeria, Spain Lettuce [8]
ALM2 AY366418 Almeria, Spain Lettuce [8]
GER3 AY581598 Frankfurt, Germany Lettuce [8]]
DEN1 AY581692 Denmark Lettuce [8]
HOL2 AY581693 Netherlands Lettuce [8]
UK1 AY581694 United Kingdom Lettuce [8]
GER1 AY581695 Fischenich, Germany Lettuce [8]
AUS1 AY581696 Western Australia Lettuce [8]
GER2 AY581697 Reskia, Germany Lettuce [8]
ITA1 AY581699 Italy Lettuce [8]
ALM3 AY581700 Almeria, Spain Lettuce [8]
ALM4 AY581701 Almeria, Spain Lettuce [8]
ALM5 AY581702 Almeria, Spain Lettuce [8]
SON1 AY839624 Almeria, Spain Sowthistle [7]
SON2 AY839625 Almeria, Spain Sowthistle [7]
SON3 AY839626 Almeria, Spain Sowthistle [7]
SON4 AY839627 Almeria, Spain Sowthistle Unpublished
LP1 FJ864681 Argentina Lettuce Unpublished
LP2 FJ864680 Argentina Lettuce Unpublished
58 a DQ530358 Brazil Lettuce Unpublished
60 a DQ530357 Brazil Lettuce Unpublished
61 a
DQ530356 Brazil Lettuce Unpublished
AUSG2 GU193114 Perth, Western
Australia
Lettuce This study
AUSE3 GU193115 Perth, Western
Australia
Lettuce This study
AUSE6 GU193116 Perth, Western
Australia
Lettuce This study
AUSF1 GU193117 Perth, Western
Australia
Lettuce This study
AUSB2 GU193118 Perth, Western
Australia
Lettuce This study
AUSA1 GU193119 Perth, Western
Australia
Lettuce This study
AUSE4 GU193120 Perth, Western
Australia
Lettuce This study
AUSG1 GU193121 Perth, Western
Australia
Lettuce This study
AUSE1 GU193122 Perth, Western
Australia
Lettuce This study
AUSD1 GU193123 Perth, Western
Australia
Lettuce This study
AUSA2 GU193124 Perth, Western
Australia
Lettuce This study
AUSC1 GU193125 Perth, Western
Australia
Lettuce This study
AUSE2 GU193126 Perth, Western
Australia
Lettuce This study
a Partial MLBVV CP sequence

70
AUSA1
AUSE4
AUSB2
AUSE3
AUSE6
AUSF1
AUSG2
AUSE1
AUSD1
AUSA2
AUSG1
AUSC1
AUS1
GER1
LS301-O
JPN1
HOL2
UK1
DEN1
GER2
A1
LP1
LP2
MUR1
ALM2
ITA1
GER3
ALM3
GAL1
AUSE2
58
60
61
A2
A1
ALM4
ALM1B1
SON1
SON4
ALM5
SON3
SON2
B2
B
99
99
97
75
99
55
98
99
77
99
99
99
66
66
60
99
99
58
74
84
61
71
0.01
CPsV
0.2
Fig 1: Phylogenetic relationships among complete coat protein (CP) nucleotide
sequences of 36 Mirafiori lettuce big-vein virus (MLBVV) isolates from Australia,
Europe, Japan and South America, and three partial MLBVV CP sequences from
Brazil. Trees generated using MEGA 4.1 using the default parameters. Tree branches
were bootstrapped with 1000 replications. Numbers at nodes indicate bootstrap scores
>50%. The scale bar represents a genetic distance of 0.01 for horizontal branch
lengths. For isolate designations, see Table 1. Inset shows isolates rooted with Citrus
psorosis virus (CPsV), accession number AF036338

71
Chapter 5 - Comparison of the coat protein genes of Lettuce
big-vein associated virus isolates from Australia with those of
isolates from other continents
Linda D. Maccarone1, Martin J. Barbetti
1, Krishnapillai Sivasithamparam
1,
Roger A. C. Jones1,2,3
1School of Plant Biology, Faculty of Natural and Agricultural Sciences, University of
Western Australia, Stirling Highway, Crawley, WA 6009, Australia
2Agricultural Research Western Australia, Department of Agriculture and Food
Western Australia, Locked bag No.4, Bentley Delivery Centre, WA 6983, Australia
3Corresponding author email: [email protected]
Summary
The complete coat protein (CP) nucleotide sequences of seven Lettuce big-vein
associated virus (LBVaV) isolates from Australia were compared to those of 22 other
LBVaV and five tobacco stunt virus (TStV) isolates. On phylogenetic analysis, clade I
contained only LBVaV isolates from Europe, sub-clade IIa only Australian LBVaV
isolates, IIb only Japanese LBVaV isolates, and IIc only TStV isolates from Japan. In
the amino acid sequences deduced, the central region of the gene was most divergent.
Mean Dn/Ds ratios were 0.283 and 0.124 for clades I and II, respectively. The
suggestion that TStV is a strain of LBVaV was supported.

72
Lettuce big-vein associated virus (LBVaV) and tobacco stunt virus (TStV) belong to
the genus Varicosavirus of which LBVaV is type species [1]. LBVaV was formerly
considered the cause of lettuce big-vein disease (LBVD) [16] until another virus
occurring in mixed infection, Mirafiori lettuce big-vein virus (MLBVV; genus
Ophiovirus), was found responsible for the symptoms [5, 7]. TStV infects tobacco
causing severe symptoms [4]. LBVaV and TStV have rod shaped particles 320-360
(LBVaV) or 300-340 (TStV) nm in length [4, 16]. Both have a CP 1194 nucleotides
long that codes for 397 amino acids [12] and are serologically related [4, 16]. Both
were considered different species despite their similar morphology, serology and
vector relationships because TStV does not to infect lettuce but infects tobacco whilst
LBVaV does the opposite [4]. Subsequently, Sasaya et al. [12] suggested that TStV
is a tobacco infecting strain of LBVaV. TStV and MLBVV are transmitted by the
chytrid Olpidium virulentus [13]. Sasaya et al. [12] reported that the complete
nucleotide and amino acid CP sequence identities between eight LBVaV and five
TStV isolates were 95.6-96.5% and 97.2-98.7%, respectively. When Navarro et al. [9]
analysed the CP sequences of 17 LBVaV isolates, they found identities of 95.2-99.8%
(nucleotides) and 98-100% (amino acids), and when they subjected 13 complete and
four partial LBVaV CP sequences to phylogenetic analysis, three isolate groupings
were found. This paper compares the CPs of seven previously unreported isolates
from Australia with those of 30 LBVaV and five TStV isolates from different
continents.
Lettuce leaves with LBVD symptoms were sampled from four farms (A, B, E
and G) in the northern Perth Metropolitan region, south-west Australia. Seven
samples were collected, one each from A and G, two from B and three from E. On the
collection day, each sample was ground in liquid nitrogen before storage at -20°C

73
before RNA extraction. Total RNA was extracted from each sample using an
UltraClean™ Plant RNA Isolation Kit (MoBio Laboratories, Inc.) according to
manufacturer‟s instructions. RNA extracts were stored at -80°C. Primer pairs specific
for the CP of LBVaV (fwd L1-21 5' ATG GCA CCC CAA ATT GAA G 3', rev
L700-679 5' GTG CAT GCC AGT GCT GGC AAG 3' and fwd L511-5 31 5' AGA
CCG AGT ATC CAT TCA AG 3', rev L1210-1185 5' TCA CTC CTT CAC TGG
TGT CTC TCC C 3') were developed by aligning known CP sequences from
GenBank in Bioedit. To amplify the entire CP sequence, both primer sets were used
on each sample. Reverse Transcription PCR (RT-PCR) was done using a OneStep
RT-PCR kit (Qiagen, Australia) according to manufacturer‟s instructions. PCR
amplification conditions consisted of 50°C for 30 min, 95°C for 15 min, followed by
30 cycles of 94°C for 1 min, 57°C for 1 min, 72°C for 1 min, then a final extension at
72°C for 10 min before being held at 14°C. PCR amplicons were visualised by
electrophoresis on 1% agarose gels. Tris-boric acid-EDTA (TBE) electrophoresis
buffer was used [10].
PCR amplicons were purified by ethanol precipitation and quantified using a
Nanodrop UV-Vis spectrophotometer following the manufacturer‟s protocol.
Standard ligation and transformation reactions were done into JM109 competent cells
using a pGEM®-T Easy Vector (Promega Corp, Madison, Wisconsin) as
recommended by the manufacturer. Screening for recombinant plasmids used M13
primers: M13 fwd (5' CGT CAG GCT TTT CCC AGT CAC GAC 3') and M13 rev (5'
TCA CAC AGG AAA CAG CTA TGA C 3'). Amplification conditions were 1 min
denaturation at 94°C followed by 25 cycles of 94°C for 30 sec, 55°C for 30 sec, 72°C
for 30 sec before being held at 14°C. Recombinant plasmids were inoculated into 5
mL of Luria-Bertani (LB) broth with ampicillin (100 mg/L) and incubated on a shaker

74
at 37°C overnight. The plasmid was then purified from the culture using an Aurum™
Bio-Rad Plasmid Extraction Kit following the manufacturer‟s protocol. Presence and
size of inserts was confirmed with a restriction digest of purified plasmid DNA by
EcoR1 (10 µL of purified plasmid DNA, 2 µL of 10 x EcoR1 buffer, 1 µL of EcoR1
and 7µ L of water). Dideoxy-termination sequencing was done at the State
Agricultural Biotechnology Centre (SABC), Murdoch University using BigDye
terminator V3.1 chemistry and an Applied Biosystems/Hitachi 37030 DNA Analyser.
The complete CP sequences of each isolate were submitted to GenBank (Table 1).
One clone was sequenced per isolate.
The complete CP genes of the seven new Australian LBVaV isolates
contained one ORF 1194 bp in length that coded for 397 amino acids. In addition, 22
complete and eight partial CP sequences of other LBVaV isolates and complete CP
sequences of five TStV isolates were retrieved from GenBank (Table 1). The partial
sequences were four from Brazil [DQ530352 (889 bp), DQ530353 (882 bp),
DQ530354 (866 bp) and DQ530355 (876 bp)] and one each from USA (AY496053),
UK (AY496054), Netherlands (AY496056) and Australia (AY496055) (all 1010bp in
length). Complete CP nucleotide sequences AB050272 (isolate LBVaV-CP) and
AB114138 (isolate Ka), both from Kagawa Japan, were identical so only the former
was included. A partial LBVaV sequence from Salinas, California (DQ182566) was
excluded as it was only 302 bp [2]. The 34 complete LBVaV and TStV CP sequences
were assembled and analysed using Molecular Evolutionary Genetics Analysis
(MEGA) program Version 4.1 with Clustal W [15] and bootstrapped with 1000 re-
samplings [14]. These sequences were analysed in direct pairwise comparisons to
establish percentage identities. Evolutionary history was inferred by the Neighbour-
Joining method. Evolutionary distances between nucleotide sequences were computed

75
using the Maximum Composite Likelihood method. This was done first with the 29
complete CP sequences of LBVaV and five of TStV, and then repeated to include the
eight partial sequences after trimming complete sequences to reflect the length of the
partial sequences. Evolutionary distances of deduced amino acid sequences were
computed for the complete CP sequences using the Poisson correction method in
MEGA4.1. Any positions containing gaps and missing data were eliminated.
Nucleotide substitutions for the CP region were inferred using the SLAC model [3].
Next, the mean number of non-silent substitutions (Dn) and silent substitutions (Ds),
and Dn/Ds ratios were determined to assess the selective constraints on the CP gene
[3]. Dn/Ds ratios for the sequences of complete CP genes were estimated first for the
seven Australian isolates alone and then for all complete nucleotide sequences.
The sequence identities of the seven new Australian isolates were 95.6-99.7%
(nucleotides) and 98.2-99.7% (amino acids). They formed a geographically distinct
group most closely related to each other and to LBVaV isolates from Japan. The
complete sequence identities of all 29 isolates of LBVaV and five of TStV ranged
from 93.6-99.7% (nucleotides) and 94.3-100% (amino acids). These values for both
nucleotide and amino acid sequence identity were the same when comparing the 29
LBVaV isolates alone. The sequence identities of the five TStV isolates alone were
95.7-99.7% (nucleotides) and 97.2-100% (amino acids). LBVaV isolate UK2 was
most divergent with 93.6% identities to Japanese LBVaV Hy and TStV Na isolates.
These sequence identities resemble those reported previously [9, 12]. Thus, our data
showing that the CP of LBVaV and TStV is highly conserved confirmed data from
previous work with fewer isolates and supported TStV being a strain of LBVaV [12].
When the complete CP nucleotide sequences (29 of LBVaV and five of TStV)
were subjected to phylogenetic analysis, all were within two clades (I and II), II being

76
resolved into three distinct sub-clades (IIa-c) (Fig. 1A). All clades and sub-clades
were geographically distinct as regards the origins of the isolates within them. Clade I
contained 12 lettuce and six sowthistle isolates of LBVaV, all from Europe. Within
clade II, sub-clade IIa only contained the seven new Australian LBVaV isolates, IIb
the four LBVaV isolates, and IIc the five Japanese TStV isolates. Sub-clades IIb and
IIc were closer to each other than to sub-clade IIa containing Australian isolates. The
groupings obtained not only agreed with the three groupings of Sasaya et al. [12], (i)
five TStV isolates, (ii) four Japanese LBVaV isolates and (iii) four Spanish LBVaV
isolates, but also added 16 extra European isolates to (iii) and revealed a new
grouping containing only Australian isolates. Our findings also agree with those of
Navarro et al. [9] in which, apart from one Japanese LBVaV isolate, the only
complete CP sequences examined were 12 from Europe. Moreover, in our study,
inclusion together in one analysis of the complete CP sequences of seven additional
isolates with those of all other 27 isolates now available on GenBank allowed greater
resolution revealing presence of two clades, and three sub-clades containing isolates
from geographically distant regions (Europe, Japan, Australia). In addition, although
lettuce isolates were present in both clades and two of the three sub-clades, sowthistle
isolates (SON1-5) were all together in one clade (I), while tobacco isolates were
isolated within a sub-clade of their own (IIc). When the eight partial CP sequences
were added to the 34 complete sequences and aligned, the single sequences of
Brazilian isolate 106 and those from Australia and the Netherlands all grouped with
the Australian isolates within sub-clade IIa (Fig. 1B). The other three Brazilian
sequences and the UK and USA sequences were in clade I, the latter being most
closely related to UK2, but the former to Spanish isolates SON1 and GAL1. None
grouped with sub-clades IIb or IIc.

77
Assuming that LBVaV evolved with lettuce in its centre of domestication in
the Mediterranean region, greatest genetic diversity would be expected there. This
was reflected in the results with the 17 Spanish isolates. Occurrence of all Australian
sequences within the same sub-clade (IIa) suggests only one introduction of LBVaV
into Australia since European settlement approximately 200 years ago. Presence of a
partial CP sequence from the Netherlands in this sub-clade might reflect its
introduction from Europe. Occurrence of only Japanese LBVaV sequences in sub-
clade IIb and Japanese TStV sequences in sub-clade IIc suggests two introductions to
Japan, but does not indicate their origin. Cultivated tobacco is from the Americas but
TStV has not been found there as yet. Further complete CP sequences of LBVaV
from North America, South America, Asia (outside Japan) and Europe (outside Spain)
would help identify the geographical origins of isolates found in different parts of the
world. Identification and sequencing of TStV isolates from countries other than Japan
would do the same for the tobacco form of LBVaV. Further sequencing might also
permit resolution of clade I into 2-3 sub-clades (Fig. 1B).
When the 34 complete amino acid sequences were compared, the central
region of the CP gene was most divergent with few amino acid substitutions towards
the N-terminus and C-terminus regions (Table 2). When the partial sequences were
included, there were 79 amino acid substitutions across the entire CP of 397
nucleotides, the partial sequences contributing an additional four. Forty six of these
substitutions occurred between amino acid positions 200 and 300. Among the isolates
in clade I, there were 55 amino acid substitutions over the entire CP gene sequence
and 48 of them were unique. There were 39 substitutions between positions 200 and
300. There were 31 amino acid substitutions in clade II which were relatively evenly
spread throughout the CP gene except for a cluster at the C-terminus. The Australian

78
LBVaV isolates in sub-clade IIa showed 13 amino acid substitutions, 10 of which
were found only in sub-clade IIa; six substitutions were in a cluster between positions
214 and 243. The Japanese LBVaV isolates in sub-clade IIb showed seven amino acid
substitutions, those at positions 110, 183 and 394 being unique. In sub-clade IIc, there
were 13 amino acid substitutions nine of which were unique, those at positions 53, 97,
183 and 393 being present in all isolates.
Navarro et al. [9] reported a mean Dn/Ds ratio of 0.132 for LBVaV CP gene
sequences. In our study, a mean Dn/Ds ratio of 0.187 was obtained for all 34 complete
CP sequences indicating that the majority of the nucleotide substitutions were silent.
The mean Dn/Ds ratios for clades I and II were 0.283 and 0.124 respectively, while
those for sub-clades IIa, IIb and IIc were 0.274, 0.060 and 0.156, respectively. Mean
Dn/Ds ratios <1 suggest selection for amino acid conservation. Genome conservation
is expected as genetic stability is commonly seen in natural plant virus populations so
the low mean Dn/Ds ratios in clades I and II reflect selection against nucleotide
changes causing amino acid substitutions, especially in sub-clades IIb and IIc. There
were 74 amino acid substitutions in the complete CP sequences, 52 were in clade I.
with 47 of them being unique. This greater level of amino acid substitution could also
explain the higher Dn/Ds ratio for clade I. Similarly, the greater Dn/Ds ratio for sub-
clade IIa might be explained by 10 of the 13 amino acid substitutions in the complete
CP sequences being unique to this sub-clade. Only three unique amino acid
substitutions were found in sub-clade IIb which had the lowest Dn/Ds ratio. The CPs
of vector-borne plant viruses are subject to greater stabilising selection than those of
viruses transmitted otherwise because interaction between the CP and cellular
receptors on vectors places greater selective constraint on the amino acid sequence
than host-virus interactions. For example, in Melon necrotic spot virus vectored by

79
Olpidium bornovanus just one amino acid substitution from Ile to Phe at position 300
resulted in loss of specific binding and fungal transmission as the CP acts as a ligand
to the vector zoospore [6]. A similar scenario might also occur within LBVaV,
explaining its high degree of CP sequence conservation.
Acknowledgements
We thank Stephen Wylie, Craig Webster and Denis Phillips for advice, the SABC for
laboratory facilities and local farmers for allowing sampling. Linda Maccarone
received an Australian Postgraduate Award from the Australian Research Council.
References
1. Brunt AA, Milne RG, Sasaya T, Verbeek M, Vetten HJ, Walsh JA (2005)
Varicosavirus. In: Fauquet CM, Mayo MA, Maniloff J, Desselberger U, Ball LA (eds)
Virus Taxonomy. Eighth Report of the International Committee on Taxonomy of
Viruses. Elsevier/Academic Press, London, pp. 669-671
2 Hayes RJ, Wintermantel WM, Nicely PA, Ryder EJ (2006) Host resistance to
Mirafiori lettuce big-vein virus and Lettuce big-vein associated virus and virus
sequence diversity and frequency in California. Plant Dis. 90:233-239
3. Kosakovsky-Pond SL, Frost SDW (2005) Datamonkey: rapid detection of selective
pressure on individual sites of codon alignments. Bioinformatics 21:2531-2533
4. Kuwata S, Kubo S (1986) Tobacco stunt virus. no 313Association of Applied
Biologist, Wellesbourne, UK, p4

80
5. Lot H, Campbell RN, Souche S, Milne RG, Roggero P (2002) Transmission by
Olpidium brassicae of Mirafiori lettuce virus and Lettuce big-vein virus, and their
roles in lettuce big-vein etiology. Phytopathology 92:288–293
6. Mochizuki T, Ohnishi J, Ohki T, Kanda A, Tsuda S (2008) Amino acid substitution
in the coat protein of Melon necrotic spot virus causes loss of binding to the surface of
Olpidium bornovanus zoospores. J. Gen. Plant Pathol. 74:176-181
7. Navarro JA, Botella F, Maruhenda A, Sastre P, Sánchez-Pina MA Pallas V (2004)
Comparative infection progress analysis of Lettuce big-vein virus and Mirafiori
lettuce virus in lettuce crops by developed molecular diagnosis techniques.
Phytopathology 94:470-477
8. Navarro JA, Botella F, Marhuenda A, Sastre P, Sánchez-Pina MA, Pallas V (2005a)
Identification and partial characterisation of Lettuce big-vein associated virus and
Mirafiori lettuce big-vein virus in common weeds found amongst Spanish lettuce
crops and their role in lettuce big-vein disease transmission. Eur. J. Plant Path.
113:25-34
9. Navarro JA, Torok VA, Vetten HJ, Pallas V (2005b) Genetic variability in the coat
protein genes of Lettuce big-vein associated virus and Mirafiori lettuce big-vein virus.
Arch. Virol. 150:681-694
10. Sambrook J, Russell DW (2001) Molecular cloning: A laboratory manual. Cold
Spring Harbour Laboratory Press, New York
11. Sasaya T, Ishikawa K, Koganezawa H (2001) Nucleotide sequence of the coat
protein gene of Lettuce big-vein virus. J. Gen. Virol. 82:1509-1515
12. Sasaya T, Ishikawa K, Kuwata S, Koganezawa H (2005) Molecular analysis of
coat protein coding region of tobacco stunt virus shows that it is a strain of Lettuce
big-vein virus in the genus Varicosavirus. Arch. Virol. 150:1013-1021

81
13. Sasaya T, Koganezawa H (2006) Molecular analysis and virus transmission tests
place Olpidium virulentus, a vector of Mirafiori lettuce big-vein virus and Tobacco
stunt virus, as a distinct species rather than a strain of Olpidium brassicae. J. Gen.
Plant Pathol. 72:20-25
14. Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4: Molecular Evolutionary
Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 24:1596-1599
15. Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W: improving the
sensitivity of progressive multiple sequence alignment through sequence weighting,
position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 22:4673-
4680
16. Vetten HJ, Lesemann DE, Dalchow J (1987) Electron microscopical and
serological detection of virus-like particles associated with lettuce big vein disease. J.
Phytopathol. 120:53-59
Figure Legend
Fig. 1. Phylogenetic relationships among complete coat protein (CP) nucleotide
sequences of 29 Lettuce big-vein associated virus (LBVaV) isolates from Europe,
Australia and Japan, and five tobacco stunt virus (TStV) isolates from Japan (A), and
these same sequences with eight partial LBVaV CP sequences included (B). Trees
generated with MEGA 4.1 using default parameters. Tree branches were bootstrapped
with 1000 replications. Numbers at nodes indicate bootsrap scores >50% (a) or all
scores (b). The scale bar represents a genetic distance of 0.005 for horizontal branch
lengths. For isolate designations, see Table 1. Inset shows isolates rooted with
sonchus yellow net virus (SYNV), accession number M17210.

82
Table 1 Isolates of Lettuce big-vein associated virus (LBVaV) and tobacco stunt virus (TStV)
used in sequence comparisons.
Virus isolate Accession
number
Geographic Origin Species
isolated
from
Reference
LBVaV-CP* AB050272* Kagawa, Japan lettuce [9, 11, 12]
No2† AB190521 Takamatsu, Japan tobacco [12]
No89† AB190522 Chunan, Japan tobacco [12]
No88† AB190523 Chunan, Japan tobacco [12]
Kan† AB190524 Kannonji, Japan tobacco [12]
Na† AB190525 Fukue, Japan tobacco [12]
A AB190526 Kannonji, Japan lettuce [12]
Hy AB190527 Nandan, Japan lettuce [12]
Wa AB190528 Hiochigawa, Japan lettuce [12]
MUR1 AY366411 Murcia, Spain lettuce [9]
GAL1 AY366412 Galicia, Spain lettuce [9]
ALM1 AY366413 Pulpi, Almeria, Spain lettuce [9]
ALM2 AY366414 Pulpi, Almeria, Spain lettuce [9]
USA** AY496053 Salinas, California, USA lettuce [9]
UK** AY496054 Wellesbourne, UK lettuce [9]
AUS** AY496055 Perth, south-west
Australia
lettuce [9]
NL** AY496056 De Lier, The Netherlands lettuce [9]
ALM3 AY581683 Pulpi, Almeria, Spain lettuce [9]
ALM4 AY581685 Pulpi, Almeria, Spain lettuce [9]
ALM5 AY581686 Pulpi, Almeria, Spain lettuce [9]
ALM6 AY581687 Pulpi, Almeria, Spain lettuce [9]
ALM7 AY581688 Pulpi, Almeria, Spain lettuce [9]
GRA1 AY581689 Granada, Spain lettuce [9]
UK2 AY581690 UK lettuce [9]
MUR2 AY581691 Aguilas, Murcia, Spain lettuce [9]
SON1 AY581681 Almeria, Spain sowthistle [8]
SON2 AY839619 Almeria, Spain sowthistle [8]
SON3 AY839620 Almeria, Spain sowthistle [8]
SON4 AY839621 Almeria, Spain sowthistle [8]
SON5 AY839622 Granada, Spain sowthistle [8]
SON6 AY839623 Granada, Spain sowthistle [8]
107** DQ530352 Bauru, Brazil lettuce Unpublished
104** DQ530353 Campinas, Brazil lettuce Unpublished
105** DQ530354 Bauru, Brazil lettuce Unpublished
106** DQ530355 Mogi das Cruzes, Brazil lettuce Unpublished
AUSE4 GU220721 Perth, south-west
Australia
lettuce This Study
AUSE5 GU220722 Perth, south-west
Australia
lettuce This Study
AUSA5 GU220723 Perth, south-west
Australia
lettuce This Study
AUSB1 GU220724 Perth, south-west
Australia
lettuce This Study

83
AUSB2 GU220725 Perth, south-west
Australia
lettuce This Study
AUSG1 GU220726 Perth, south-west
Australia
lettuce This Study
AUSE6 GU220727 Perth, south-west
Australia
lettuce This Study
*Complete LBVaV CP nucleotide sequences AB050272 (isolate LBVaV-CP) and
AB114138 (isolate Ka), both from Kagawa in Japan, were identical.
**Partial LBVaV CP sequences.
† TStV isolates.
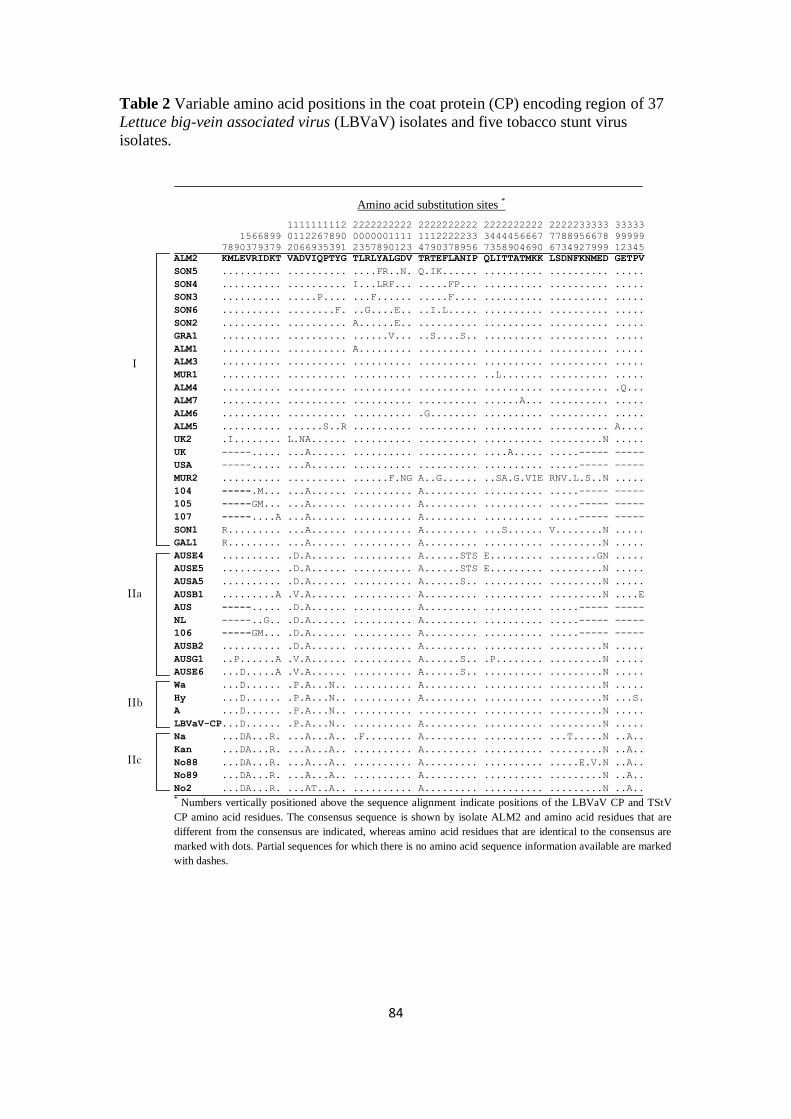
84
Table 2 Variable amino acid positions in the coat protein (CP) encoding region of 37
Lettuce big-vein associated virus (LBVaV) isolates and five tobacco stunt virus
isolates.
1111111112 2222222222 2222222222 2222222222 2222233333 33333
1566899 0112267890 0000001111 1112222233 3444456667 7788956678 99999
7890379379 2066935391 2357890123 4790378956 7358904690 6734927999 12345
ALM2 KMLEVRIDKT VADVIQPTYG TLRLYALGDV TRTEFLANIP QLITTATMKK LSDNFKNMED GETPV
SON5 .......... .......... ....FR..N. Q.IK...... .......... .......... .....
SON4 .......... .......... I...LRF... .....FP... .......... .......... .....
SON3 .......... .....P.... ...F...... .....F.... .......... .......... .....
SON6 .......... ........F. ..G....E.. ..I.L..... .......... .......... .....
SON2 .......... .......... A......E.. .......... .......... .......... .....
GRA1 .......... .......... ......V... ..S....S.. .......... .......... .....
ALM1 .......... .......... A......... .......... .......... .......... .....
ALM3 .......... .......... .......... .......... .......... .......... .....
MUR1 .......... .......... .......... .......... ..L....... .......... .....
ALM4 .......... .......... .......... .......... .......... .......... .Q...
ALM7 .......... .......... .......... .......... ......A... .......... .....
ALM6 .......... .......... .......... .G........ .......... .......... .....
ALM5 .......... ......S..R .......... .......... .......... .......... A....
UK2 .I........ L.NA...... .......... .......... .......... .........N .....
UK -----..... ...A...... .......... .......... ....A..... .....----- -----
USA -----..... ...A...... .......... .......... .......... .....----- -----
MUR2 .......... .......... ......F.NG A..G...... ..SA.G.VIE RNV.L.S..N .....
104 -----.M... ...A...... .......... A......... .......... .....----- -----
105 -----GM... ...A...... .......... A......... .......... .....----- -----
107 -----....A ...A...... .......... A......... .......... .....----- -----
SON1 R......... ...A...... .......... A......... ...S...... V........N .....
GAL1 R......... ...A...... .......... A......... .......... .........N .....
AUSE4 .......... .D.A...... .......... A......STS E......... ........GN .....
AUSE5 .......... .D.A...... .......... A......STS E......... .........N .....
AUSA5 .......... .D.A...... .......... A......S.. .......... .........N .....
AUSB1 .........A .V.A...... .......... A......... .......... .........N ....E
AUS -----..... .D.A...... .......... A......... .......... .....----- -----
NL -----..G.. .D.A...... .......... A......... .......... .....----- -----
106 -----GM... .D.A...... .......... A......... .......... .....----- -----
AUSB2 .......... .D.A...... .......... A......... .......... .........N .....
AUSG1 ..P......A .V.A...... .......... A......S.. .P........ .........N .....
AUSE6 ...D.....A .V.A...... .......... A......S.. .......... .........N .....
Wa ...D...... .P.A...N.. .......... A......... .......... .........N .....
Hy ...D...... .P.A...N.. .......... A......... .......... .........N ...S.
A ...D...... .P.A...N.. .......... .......... .......... .........N .....
LBVaV-CP...D...... .P.A...N.. .......... A......... .......... .........N .....
Na ...DA...R. ...A...A.. .F........ A......... .......... ...T.....N ..A..
Kan ...DA...R. ...A...A.. .......... A......... .......... .........N ..A..
No88 ...DA...R. ...A...A.. .......... A......... .......... .....E.V.N ..A..
No89 ...DA...R. ...A...A.. .......... A......... .......... .........N ..A..
No2 ...DA...R. ...AT..A.. .......... A......... .......... .........N ..A.. * Numbers vertically positioned above the sequence alignment indicate positions of the LBVaV CP and TStV
CP amino acid residues. The consensus sequence is shown by isolate ALM2 and amino acid residues that are
different from the consensus are indicated, whereas amino acid residues that are identical to the consensus are
marked with dots. Partial sequences for which there is no amino acid sequence information available are marked
with dashes.
Amino acid substitution sites *
I
IIa
IIb
IIc

85
Fig. 1A
SON5
SON4
SON3
SON6
SON2
GRA1
ALM1
ALM3
MUR1
ALM4
ALM7
ALM6
ALM2
ALM5
UK2
MUR2
SON1
GAL1
I
AUSE4
AUSE5
AUSA5
AUSB1
AUSB2
AUSG1
AUSE6
IIa
Wa
Hy
LBVaV CP
A
IIb
Na
Kan
No88
No89
No2
IIc
II 87 50
100
93
81
55
100
100
99
99
94
98
85
62
96
86
96
74
62
64
66
98
63
65
50
52
0.005
SYNV
0.1

86
Fig. 1B
SYNV
0. 2
AUSE4
AUSE5
AUSA5
AUSB1
AUS
NL
106
AUSB2
AUSG1
AUSE6
IIa
Wa
Hy
A
LBVaV CP
IIb
Na
Kan
No88
No89
No2
IIc
II
UK2
UK
USA
ALM2
ALM5
ALM4
ALM6
MUR1
ALM7
ALM3
ALM1
GRA1
SON6
SON2
SON3
SON5
SON4
105
107
SON1
104
GAL1
MUR2
85 49
100
65
46
54
100
100
73 94
98
97
86
51
54
56
83
92
73
69 99
78
84
83
77
64
69
68
44
70
69
93
47
29
62
45
0.005
I

87
Chapter 6 – Commercial Lettuce Seedling Nurseries
6. 1 Introduction
Western Australia is unique in that its isolation from the other areas of Australia by
desert and distance means that many plant pathogens either do not occur or can be
eradicated following their introduction (McKirdy et al. 2001; Woods et al. 2001 and
Mackie et al. 2002). However, many important soil-borne pathogens of commercial
horticultural crops such as Phytophthora spp. and Pythium spp. and the vector and
viruses that cause LBVD are already well established (Davison et al. 2006; Latham et
al. 2004). For example, Phytophthora cinnamomi, the causal agent of jarrah (Eucalyptus
marginata) dieback is a soil-borne plant pathogen that is spread through contamination
from within nursery environments by motile zoospores in water, infested potting mix
and soil, dust on seed for propagation material, mud on vehicle tyres and on cultivation
implements (Sivasithamparam and Goss 1981). In order to prevent spread of pathogens
both within the nursery and onto producer‟s farms, there is a need to use seed that is
free from spore contamination, sterilised potting mixes, new or sterilised pots and
containers, free draining benches to grow plants and water decontaminated by
chlorination or exposure to ultra-violet light before coming into contact with plants
(Sivasithamparam and Goss 1981).
O. virulentus (the Chytrid vector of LBVaV and MLBVV) is holocarpic and produces
many zoosporangia and resting spores in the root tissue of infested host plants. The
zoosporangia of O. virulentus release posteriorly-uniflagellate zoospores that are motile

88
within water that is trapped between roots and soil particles. These zoospores are able to
infect roots of other host plants (Campbell 1996). The resting spores of O. virulentus
contaminate the soil when roots of infested plants die and decay and these resting spores
can remain viruliferous in the soil for up to 20 years (Campbell 1985). In lettuce
(Lactuca sativa) seedling nurseries, LBVD is spread particularly when seedling trays
are not placed on free-draining benches as the water laying on benches gives the
viruliferous O. virulentus zoospores the ability to move and infect seedlings in
neighbouring trays (Jones 2004).
On the Swan Coastal Plain of south-west Australia, most of the commercial lettuce
growing farms source their seedlings from three major lettuce seedling producers rather
than direct seeding or growing and transplanting their own seedlings. Latham et al.
(2004) reported that a lettuce seedling nursery was inadvertently on-selling LBVD
infected lettuce seedlings to commercial lettuce growers. They recorded up to 31%
incidence of LBVD on lettuce seedlings purchased from this seedling nursery. LBVD
was also evident in „bait‟ plants grown in potting mix and dirt collected from under the
nursery benches. While LBVD was detected in nursery potting mix nine weeks after
composting, no evidence was found of LBVD contamination on seedling trays, peat or
pelleted seed. It was also found that the potting mix imported to prepare the composted
final commercial mix in which seeds are sown was also contaminated.
Mature lettuce seed has three distinguishable layers, the pericarp which is the outermost
layer containing longitudinal „ribs‟ which is made out of sclerenchymatous tissue, the
integument which is also made up of dead cells and is only a few cell layers thick, and
the innermost layer, the endosperm, that tightly surrounds the embryo and is only two

89
cell layers thick (Borthwick and Robbins 1928; Paulson and Srivastava 1968; Jones,
1974; Nijsse et al. 1998) (Fig. 6.1 and Fig. 6.2). O. virulentus resting spores are able to
be carried by dust and have to potential to contaminate the seed coat of lettuce (Jones
2004) by lodging amongst the many „ribs‟ on the seed coat (Fig. 6.2).
Lettuce seedling nurseries compost a mixture of imported potting mix and peat for
various amounts of time allowing the temperature of the compost to reach a maximum
of 65°C. The nurseries surveyed make their final compost mix and then mix with peat to
make their commercial mix into which the lettuce seeds are sown. Seedling trays are
treated with chlorine and pelleted lettuce seeds are sown. With pelleting, lettuce seeds
are coated with an inert substance which gives them a round uniform shape and makes
them easier to handle. The seedling trays are placed in a 16-17°C growth chamber
maintained with 80% humidity for 48h. Lettuce seedlings are then grown outdoors from
the cotyledon stage onwards and watered daily.
Given the rapidly growing urban sprawl occurring in the outer metropolitan region of
Perth, vegetable producers are moving further from the city and cultivating virgin land.
In order to prevent the spread of LBVD onto these virgin lands, which have never
grown lettuce, an efficient integrated disease management strategy, such as the one
developed by Jones (2004), is needed and requires lettuce seedling nurseries to sell an
LBVD-free product to commercial lettuce growers. This chapter examines a lettuce
seedling nursery for the presence of LBVD and also investigates the possible seed coat
contamination of lettuce seeds by viruliferous O. virulentus resting spores.

90
Fig. 6.1: Schematic drawing of a lettuce seed showing the three layers surrounding the
embryo, endosperm, integument and „ribs‟ on the pericarp. Adapted from Nijsse et al.
(1998).
Fig. 6.2: Lettuce seed viewed under a light microscope at 4×magnification showing the
„ribs‟ on the seed coat. Scale bar represents 100.0µm
Pappus
P ericarp
Embryo
Integument
Endosperm
3.8mm

91
6.1.1 Aims
The aims of this study were to:
Determine the presence of LBVaV and MLBVV on commercial lettuce growing
farms in the Swan Coastal Plain of south-west Australia by testing samples by
DAS-ELISA.
Determine the presence of LBVaV and MLBVV in a commercial seedling
nursery in the Perth metropolitan region by testing samples by DAS-ELISA and
PCR.
Determine whether transmission of LBVD occurs through lettuce seed coat
contamination by resting spores of viruliferous O. virulentus by: a) detecting the
presence of LBVaV and/or MLBVV on the surface of lettuce seeds b)
determining whether sowing infected seed can result in infection with LBVaV
and/or MLBVV in lettuce plants grown from seeds harvested from LBVD-
infected plants.

92
6.2 Materials and Methods
6.2.1 Field collection of samples
Samples used were collected in the winter growing season of 2006 in south-western
Australia. Fourteen different commercial lettuce growing properties were visited and
sampled and 26 leaf samples taken in total. In each instance one whole lettuce leaf
represented one sample. The lettuce growing farms were assigned letter codes to protect
their anonymity (Table 6.3). Eleven of the commercial lettuce growing properties were
located in the Wanneroo to Gingin region up to 98km north of the Perth Central
Business District (CBD), two in the Peel region up to 43km south of the Perth CBD and
at Manjimup located approximately 300km south of the Perth CBD. In addition, four
lettuce plants of unknown origin, but showing LBVD symptoms (Fig. 6.3), were
purchased from a local supermarket and also tested. Of all the lettuce leaf samples
collected, three were coloured fancy lettuce cv. Lollo Rossa, 18 were crisphead/iceberg
type lettuce of unknown cultivar and three were cos type lettuce of unknown cultivar. In
addition, two leaf samples from separate S. oleraceous plants growing amongst a lettuce
crop from farm H, in which LBVD was evident, were also sampled. Leaf samples were
kept in separate zip lock bags and stored at 4°C until tested.

93
Fig 6.3: An example of a field grown crisphead type lettuce from which a leaf sample
was taken. The arrow points to obvious vein chlorosis. This particular plant was grown
in a field on Farm A.
6.2.2 Collection of soil samples from a commercial seedling nursery
One commercial seedling nursery in the Perth Metropolitan region was sampled once in
2006. The lettuce seedling nursery was also assigned a code to protect its anonymity
and will be described only as Nursery 1. No lettuce tissue samples were taken from the
nursery, rather soil and compost which could provide an inoculum source. In addition,
samples of the final compost, commercial mix and soil samples from two lettuce
seedling growing areas were all randomly taken. The samples were collected into large
bags and stored at room temperature until used. Of the two lettuce seedling growing
areas sampled, one was described as the „lettuce area‟ and had been used for growing
lettuce for between 8 and 10 years whilst the other was described as the „newer area‟
and had been used for growing lettuce seedlings for approximately two years.

94
6.2.3 Growth of lettuce seedlings in nursery soil samples
Lettuce cv. Great Lakes seeds were sown into the soil samples collected from the
nursery in 140mm pots. At least three seeds were sown per pot and once germination
has occurred, the seedlings were culled to one seedling per pot. The pots were kept in an
environment controlled growth facility at the University of Western Australia and
maintained on free draining benches at 18°C day/13°C night with a 12 hour
photoperiod. The youngest leaf of each of the seedlings was sampled five weeks after
sowing and then tested by either DAS-ELISA or PCR for both LBVaV and MLBVV.
Four lettuce seedlings grown in the final compost and six lettuce seedlings grown in the
commercial mix were tested by PCR, whilst five lettuce seedlings grown in soil
collected from the „newer lettuce growing area‟ were tested by DAS-ELISA. Four
lettuce seedlings grown in soil collected from the „lettuce growing area‟ were tested by
both PCR and DAS-ELISA. Appropriate controls for this experiment are described in
6.2.13.
6.2.4 Source of seed
The seeds used in these experiments came from two separate sources. Source 1: the
seeds used for PCR detection of LBVaV and MLBVV on the seed surface came from
lettuce cv. Lollo Rossa plants which were grown a glasshouse (Fig 6.4) which had all
previously tested positive for both LBVaV and MLBVV by PCR. Source 2: seeds
collected from lettuce cv. Great Lakes plants, that had been grown in soil infested with
LBVD collected from commercial lettuce Farm A, were used in the soil bioassay. The
lettuce plants from both sources used for seed production were grown in the same

95
conditions as those described in 6.2.3 above. For use as a negative control, seeds were
also collected from lettuce cv. Great Lakes plants that had been grown in sterilised
potting mix and had shown no visible symptoms of LBVD. Both LBVaV and MLBVV
were previously detected by PCR in the plants which were grown in LBVD-infested soil
but not in the plants grown in sterile potting mix.
Fig. 6.4: Lettuce cv. Lollo Rossa plant which had tested positive for LBVD showing
flower buds and one yellow flower. Seeds were harvested from this plant and used in
PCR detection of O. virulentus, LBVaV and MLBVV. The purple flowers in the
background are of Chicorum endiva and were not used in this study.

96
6.2.5 Surface sterilisation of seed
Two methods were used to surface sterilise the lettuce seeds. To eliminate resting
spores of Olpidium sp., seeds were surface sterilised with sodium hypochlorite
(NaOCl). Seeds from plants that tested positive for LBVaV and MLBVV, and seeds
from plants that tested negative for both viruses were placed in separate seed packets
(approximately 10 seeds per packet) folded from Whatman filter paper and then
immersed in a 2% aqueous NaOCl solution on a shaker for 20 mins at room
temperature. The seeds were then rinsed twice in sterile DI water for 1 min before being
air-dried on the open seed packets in a laminar flow cabinet. This method of seed
surface sterilisation was also used in the initial experiment.
To eliminate potential virus particle contamination of seed coats, seeds were surface
sterilised with trisodium phosphate (Na3PO4). For this, seeds were again placed in
separate filter paper seed packets (approximately 10 seeds per packet) and placed in
10% aqueous Na3PO4 solution for 2h at room temperature before being rinsed six times
for 1 min in distilled water. The seeds were then air-dried on the filter paper packets.
Seeds were also surface sterilised with both NaOCl and Na3PO4. For each treatment, 10
seeds from lettuce plants which had previously tested positive for LBVaV and MLBVV,
and 10 seeds from lettuce plants which had previously tested negative for LBVaV and
MLBVV, were used. Five seeds from lettuce plants which had previously tested
positive for both viruses were left un-surface sterilised as were five seeds from lettuce
plants which previously tested negative for both viruses. Not all of the 10 seeds for each

97
treatment germinated however the number of plants germinated for each treatment is
given in Table 6.3.
6.2.6 Growing conditions
The potting mix was autoclaved three times at 121°C for 40 mins. Lettuce cv. Great
Lakes seeds were sown into autoclaved potting mix in new 140mm diameter plastic
pots. The pots were placed in a glasshouse in which temperature was kept at
approximately 20°C on free draining benches at DAFWA, South Perth and seed
germination was observed 16 days after sowing. Leaf tissue samples were taken from
the youngest leaf 70 days after the seeds were sown.
3.2.7 ELISA and antibodies
Leaf samples (6.2.6) were tested on the day of collection by DAS-ELISA (Clark and
Adams 1977), Chapter 2 (2.14). Virus specific polyclonal antisera for LBVaV (Prime
Diagnostics, Wageningen, The Netherlands) and MLBVV (DSMZ, Braunschweig,
Germany) were used. Samples were considered positive if the absorbance reading was ≥
three times the absorbance of the healthy control sample. The controls used for this
experiment are described in 6.2.13.

98
6.2.8 RNA extraction
RNA extraction was done in order to amplify LBVaV and MLBVV. The RNA of 22
non-surface sterilised lettuce cv. Lollo Rossa seeds was extracted using a Qiagen
RNeasy Plant Mini Kit as described in Chapter 2 (2.2.2) before RT (reverse
transcription) and subsequent PCR. RNA from leaf samples of lettuce bait plants grown
in final compost and commercial mix collected from Nursery 1 were also extracted
using the same protocol.
6.2.9 Reverse transcription (RT)
The RT method used to synthesise first strand cDNA from the RNA extract used M-
MLV reverse transcriptase (Promega, Inc., Madison, WI) where 12µL of RNA was
added to 3µL of random primers and incubated at 70°C for 5min before the tubes were
put on ice for 10min and then centrifuged for 30sec at approximately 14 000 × g. Then
added to each primer/template reaction was 5µL of M-MLV 5 × reaction buffer, 1.25µL
of each 10mM dATP, 10mM dCTP, 10mM dGTP and 10mM dTTP, 0.5µL of
ribonuclease inhibitor, 1µL of M-MLV RT and 13.5µL of nuclease-free water. The
reactions were then incubated at 37°C for one hour and then used immediately in PCR
as described in Chapter 2 (2.3.1) and then stored at 80°C.

99
6.2.10 DNA extraction
In the initial experiment, DNA extraction was done in order to amplify and therefore
determine the presence of O. virulentus resting spores. The seeds harvested were from a
plant grown in a glasshouse where the majority of the plants had tested positive for
LBVaV and MLBVV and therefore it was assumed that O. virulentus resting spores
were present. Approximately 200 of these seeds were separated into four separate
1.5mL Eppendorf tubes so that there were approximately 50 seeds in each tube and
homogenised with a plastic pestle in liquid nitrogen before DNA extraction. The tubes
containing the homogenised lettuce seed samples were labelled Seed 1, Seed 2, Seed 3
and Seed 4. DNA extraction was done using Qiagen DNeasy Plant Mini Kit as
described in Chapter 2 (2.2.1). As this was an initial experiment there were no positive
or negative seed controls available.
6.2.11 PCR and agarose gel electrophoresis for RNA extracts
Promega GoTaq® Flexi chemistry was used for PCR amplification. A 100µL master
mix was made following the manufacturer‟s instructions by adding 20µL of 5 ×
GoTaq® Flexi buffer, 4µL 25mM MgCl2, 2µL 10mM dNTPs, 5µL 10pmol/µL forward
primer, 5µL 10pmol/µL reverse primer, 0.5µL GoTaq® DNA polymerase, 10µL
template cDNA and then the total volume was made up to 100µL by adding 53.5µL of
nuclease free water.
The primers used to amplify LBVaV were CP42F forward (5' CGT CGT GGA AAT
CAC AGG TAA GAC 3') and CP426R reverse (5' CGT AGA GAC AGC GAT GAA

100
TGC 3') and CP647F forward (5' CAA GGC AAG CTG AGA TGT TGT TCG 3') and
CP907R reverse (5' GAA GGG TTT GAA AGA CRG ATG G 3') (Hayes et al. 2006).
The primers used to amplify MLBVV were 1100F forward (5' TGG GAC TCC AGT
GGT CTT GTA TCC 3') and 1407R reverse (5' CGA AGG TAG AAA TAG GAA
CGA TGG 3') (Hayes et al. 2006). The PCR conditions consisted of an initial
denaturation at 95°C for 5min followed by 35 cycles of denaturation at 95°C for 30sec,
annealing at 52°C for 30sec and extension at 72°C for 1min. This was followed by a
final extension at 72°C for 10min before being held at 4°C. Agarose gel electrophoresis
was done as described in Chapter 2 (2.5).
6.2.12 PCR and agarose gel electrophoresis for DNA extracts
The PCR for DNA extracts was done as described in Chapter 2 (2.3.1). The primers
used were Ov1 forward (5' CAA GAC CTG CCC CCA AAA GGG 3') and Ov2 reverse
(5' CCA AGT TCG CAA ACG TGG CG 3'). Agarose gel electrophoresis was done as
described in Chapter 2 (2.5). As this was an initial experiment, there were no positive or
negative seed controls available so a lettuce root extract which had previously tested
positive for O. virulentus was used as a positive control and a negative water control
was used to show that there was no contamination in PCR reagents.
6.2.13 Controls for DAS-ELISA and PCR
The positive control used for testing for LBVaV and MLBVV by PCR and DAS-
ELISA, came from a lettuce plant which had previously tested positive to both LBVaV
and MLBVV by both PCR and DAS-ELISA. The positive control was grown in soil

101
that had been collected from farm A and was maintained in the same growth room
under the same conditions as the other experimental plants. A negative control lettuce
cv. Silverado plant was grown in sterilised potting mix and tested periodically by
ELISA and PCR to ensure it was not infected with either virus. It was maintained in the
same manner as was the positive control and the other experimental plants.
For the experiment regarding seed coat contamination, a sample of the youngest leaf of
a lettuce plant visibly infected with LBVD was collected from a lettuce crop grown at
the DAFWA Medina Research Station, Medina, Western Australia (approximately
40km south of the Perth CBD) and this was used as the positive control. The leaves
used as a negative control were from a plant grown in sterilised soil and maintained in a
separate glasshouse to the experimental plants to prevent any cross-contamination.

102
6.3 RESULTS
6.3.1 Samples collected from commercial lettuce growing fields
Of the 22 field collected samples, (20 lettuce and 2 sowthistle) and the 4 shop bought
lettuce samples, 23 tested positive by DAS-ELISA for MLBVV and 19 tested positive
for LBVaV. All of the four shop bought samples had a mixed infection with both
LBVaV and MLBVV, as did four samples from farm H, two samples from farm A and
one sample each from farms D, E, G, I, J, K, O and P (Table 6.1). One of the sowthistle
samples tested positive only for LBVaV whilst two samples from farm L, one sample
from farm M and two samples from farm N had an infection with only MLBVV. Two
of the samples, one sowthistle from farm H and one lettuce cv. Lollo Rossa from Farm
N, tested negative for both viruses (Table 6.1).
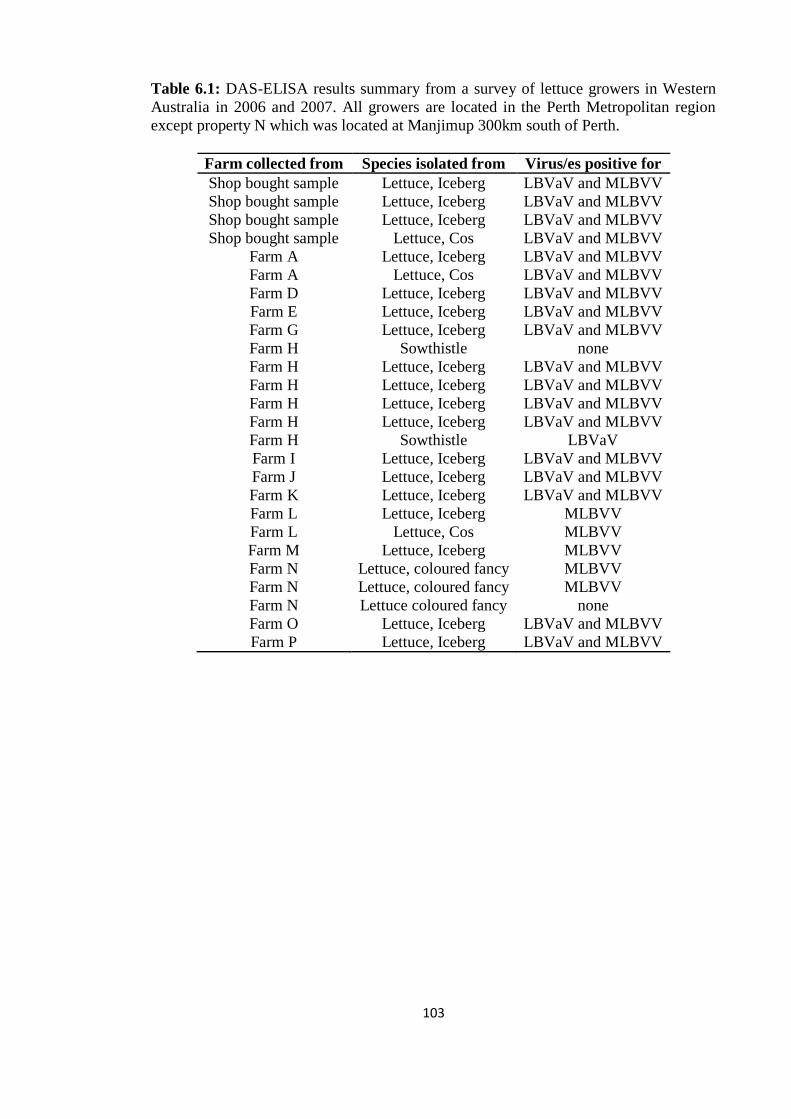
103
Table 6.1: DAS-ELISA results summary from a survey of lettuce growers in Western
Australia in 2006 and 2007. All growers are located in the Perth Metropolitan region
except property N which was located at Manjimup 300km south of Perth.
Farm collected from Species isolated from Virus/es positive for
Shop bought sample Lettuce, Iceberg LBVaV and MLBVV
Shop bought sample Lettuce, Iceberg LBVaV and MLBVV
Shop bought sample Lettuce, Iceberg LBVaV and MLBVV
Shop bought sample Lettuce, Cos LBVaV and MLBVV
Farm A Lettuce, Iceberg LBVaV and MLBVV
Farm A Lettuce, Cos LBVaV and MLBVV
Farm D Lettuce, Iceberg LBVaV and MLBVV
Farm E Lettuce, Iceberg LBVaV and MLBVV
Farm G Lettuce, Iceberg LBVaV and MLBVV
Farm H Sowthistle none
Farm H Lettuce, Iceberg LBVaV and MLBVV
Farm H Lettuce, Iceberg LBVaV and MLBVV
Farm H Lettuce, Iceberg LBVaV and MLBVV
Farm H Lettuce, Iceberg LBVaV and MLBVV
Farm H Sowthistle LBVaV
Farm I Lettuce, Iceberg LBVaV and MLBVV
Farm J Lettuce, Iceberg LBVaV and MLBVV
Farm K Lettuce, Iceberg LBVaV and MLBVV
Farm L Lettuce, Iceberg MLBVV
Farm L Lettuce, Cos MLBVV
Farm M Lettuce, Iceberg MLBVV
Farm N Lettuce, coloured fancy MLBVV
Farm N Lettuce, coloured fancy MLBVV
Farm N Lettuce coloured fancy none
Farm O Lettuce, Iceberg LBVaV and MLBVV
Farm P Lettuce, Iceberg LBVaV and MLBVV

104
6.3.2 Samples collected from seedling nurseries and tested by PCR and DAS-
ELISA
Lettuce leaf tissue samples from bait plants grown in soil collected from Nursery 1
(final compost, final mix and soil from the lettuce seedling growing area) were taken
five weeks after germination. All 19 leaf samples tested positive by PCR for MLBVV.
LBVaV was amplified in four out of the six plants grown in the commercial mix and in
four out of the four plants grown in the soil collected from the lettuce seedling growing
area. LBVaV was not detected in plants grown in the final compost (Table 6.2). The
bands obtained using LBVaV primer pair CP42F and CP426R were of expected size
(385bp) as were the bands obtained using LBVaV primer pair CP647F and CP907R
(261bp) and MLBVV primer pair 1100F and 1407R (308bp) (Fig. 6.3). None of these
19 samples tested positive for LBVaV or MLBVV by DAS-ELISA.
Table 6.2: The areas from where samples were taken at Nursery 1 and the number of
lettuce seedlings grown in each sample and the number of plants showing the presence
of LBVaV and/or MLBVV as tested by PCR.
Sample taken Number of seedlings
grown
DAS-
ELISA/PCR
LBVaV MLBVV
Final compost 4 PCR 0/4 4/4
Commercial
mix
6 PCR 4/6 6/6
Newer area 5 DAS-ELISA 0/5 5/5
Lettuce area 4 DAS-ELISA/PCR 4/4 4/4

105
Fig. 6.3: Amplification of LBVaV and MLBVV isolated from leaves of lettuce which
had been grown in soil collected from the lettuce seedling growing area in Nursery 1.
Lane M: 6µL of 100bp molecular weight marker (Fisher Biotech), Lane 1: LBVaV
amplicon using primers CP42F and CP426R, Lane 2: LBVaV amplicons using primers
CP647F and CP907R, Lane 3: MLBVV amplicon using primers 1100F and 1407R,
Lane 4: negative control using LBVaV primers CP647F and CP907R, Lane 5: negative
control using MLBVV primers 1100F and 1407R, Lane 6: positive control using
LBVaV primers CP647F and CP907R, Lane 7: positive control using MLBVV primers
1100F and 1407R.
6.3.4 Seeds tested by PCR for LBVaV, MLBVV and O. virulentus
PCR and agarose gel electrophoresis confirmed the presence of LBVaV and MLBVV
from the RNA extracts obtained from 22 bulked lettuce seeds. The expected fragment
sizes were obtained for each primer pair; LBVaV primer pair CP647F and CP907R
(261bp) and MLBVV primer pair 1100F and 1407R (308bp) (Fig. 6.4). DNA extraction
and then PCR amplification of the four bulked lettuce seed samples, Seed 1, Seed 2,
Seed 3 and Seed 4, revealed O. virulentus amplicons in two out of the four homogenised
seed samples (Seed 3 and Seed 4). The amplicons were of expected size (361bp).
M 1 2 3 4 5 6 7
500bp
385bp
308bp
261bp

106
Fig. 6.4: Amplification of LBVaV and MLBVV from 22 bulked lettuce seeds. Lane M:
6µL 100bp molecular weight marker (Fisher Biotech), Lane 1: LBVaV amplicons using
primers CP647F and CP907R, Lane 2: MLBVV amplicon using primers 1100F and
1407R. White oval shows the location of the faint band in Lane 2.
Fig. 6.5: Amplification of O.virulentus from lettuce seeds using specific primers
Ov1(forward) and Ov2 (reverse) giving the expected fragment size of 361bp. Lane M:
6µL 100bp molecular weight marker (Fisher Biotech), Lane 1: cDNA from Seed 1,
Lane 2: cDNA from Seed 2, Lane 3: cDNA from Seed 3, Lane 4: cDNA from Seed 4,
Lane 5: negative water control, Lane 6: positive O. virulentus control.
M 1 2
500bp
308bp
261bp
M 1 2 3 4 5 6
500bp
361bp

107
6.3.3 Surface sterilisation of seed
All of the lettuce leaf tissue samples from plants grown from surface sterilised seed that
were tested by DAS-ELISA were negative for both LBVaV and MLBVV (Table 6.3).
In contrast, the positive control collected from the DAFWA Medina Research Station
gave a high absorbance reading and therefore tested positive for both viruses. This also
demonstrates that autoclaving potting mix three times at 121°C for 40 mins was an
effective sterilisation method as neither virus was detected in any of the plants grown in
this autoclaved potting mix.
Table 6.3: Method of surface sterilisation used on lettuce cv. Great Lakes seeds
harvested from plants that were either positive or negative for LBVaV and MLBVV
when tested by DAS_ELISA, and the number of plants grown for each treatment and
the number of plants in each treatment that tested positive for LBVaV or MLBVV by
DAS-ELISA
Method of
Surface
sterilisation
Positive/negative for
LBVaV and MLBVV
Number of plants
grown/treatment
LBVaV MLBVV
No surface
sterilisation
positive 4 0/4 0/4
No surface
sterilisation
negative 2 0/2 0/2
Na3PO4 positive 6 0/6 0/6 Na3PO4 negative 9 0/9 0/9 NaOCl positive 10 0/10 0/10 NaOCl negative 5 0/5 0/5
Na3PO4/ NaOCl positive 5 0/5 0/5 Na3PO4/ NaOCl negative 5 0/5 0/5

108
6.4 DISCUSSION
LBVaV and MLBVV were readily detected by DAS-ELISA in 18 lettuce samples
collected from plants almost at harvest stage. MLBVV was the only virus detected in
six of the lettuce samples but for one lettuce sample both viruses were detected. LBVaV
was the only virus detected in the first of the Sowthistle samples whilst the second
Sowthistle sample tested negative for both viruses. All of these plants with the
exception of the negative control plant and the two sowthistle samples, showed
symptoms for LBVD and were collected across 13 different commercial lettuce growing
properties and with four samples purchased from a local fruit and vegetable retailer.
DAS-ELISA however, failed to detect LBVaV or MLBVV in lettuce samples taken five
weeks after sowing from bait plants grown in soil collected from a contaminated
commercial lettuce seedling nursery. As O. virulentus, LBVaV and MLBVV were
detected in these samples by RT-PCR, this suggests that the method of testing for both
LBVaV and MLBVV by RT-PCR is more sensitive than testing by DAS-ELISA, as
both viruses were detected much earlier despite the fact that tests using DAS-ELISA
seemed to be more reliable when using leaf tissue from older plants. Roggero et al.
(2003) also reported a lack of sensitivity in MLBVV antiserum when detecting
MLBVV in low concentrations in leaf tissue. This comparison of sensitivity was also
observed by Schenk et al. (1995) who tested the movement of Barley mild mosaic virus
and Barley yellow mosaic virus which is vectored by the fungus Polymyxa graminis,
and found that both these viruses could be detected in roots by RT-PCR one day after
inoculation with viruliferous zoospores whereas it took 3-4 weeks before the viruses
could be detected in the same root tissue by DAS-ELISA.

109
Previous studies on the spread of LBVD through infected seedlings being on-sold from
nurseries to lettuce producers detected LBVaV by DAS-ELISA in lettuce leaf samples
but did not involve tests for MLBVV as this work was carried out in 1999-2000 which
was before MLBVV had been discovered in association with LBVD (Latham et al.
2004). This study (Chapter 6) has shown that both viruses involved in the LBVD
complex, LBVaV and MLBVV can be detected by DAS-ELISA. This has provided the
first evidence that both viruses are associated with LBVD in Western Australia, which
reflects the situation found previously in European temperate and Mediterranean
climates (Roggero et al. 2000), Brazil (Colariccio et al. 2003), Japan (Navarro et al.
2004) and in North America (Hayes et al. 2005). MLBVV is considered as the
symptom-causing virus whereas LBVaV is considered to be latent (Roggero et al.
2000). Given the high sensitivity of RT-PCR, some random testing of lettuce seedlings
by this method at the transplant stage could now be included within the integrated
disease management strategy suggested by Jones (2004) to more reliably ensure lettuce
seedling nurseries are not selling diseased seedlings to lettuce producers. However, the
cost of RT-PCR exceeds that of DAS-ELISA so this may be the method of choice
instead, possibly by growing batches of seedlings from nurseries in autoclaved potting
mix for several weeks until they can be reliably tested for the presence of LBVaV and
MLBVV using DAS-ELISA.
Seeds can be inadvertently contaminated with viruliferous O. virulentus resting spores.
This was demonstrated by RT-PCR amplification of O. virulentus, LBVaV and
MLBVV from extracts of lettuce seeds harvested from LBVD infected plants. Even
though the vector, LBVaV and MLBVV were all amplified on lettuce seeds, it is
suggested by the seed coat contamination experiment that if lettuce seeds come into

110
contact with viruliferous O. virulentus resting spores, the number of propagules present
on the seed is likely to be inadequate to cause systemic infection of the lettuce plant. It
is also possible that the temperature, and possibly the potting mix biology/chemistry
which may have been altered during sterilisation, were not conducive to systemic
infection. However, further detection by PCR (on both the roots and leaves) may reveal
the presence of LBVaV and MLBVV that DAS-ELISA failed to detect in lettuce leaf
tissue.
This study is the first to demonstrate the external contamination of lettuce seeds by O.
virulentus, LBVaV and MLBVV. A similar phenomenon has been previously reported
with Melon necrotic spot virus (MNSV) in melon which is also vectored by another
chytrid, Olpidium bornovanus (Faruki 1981; Campbell et al. 1996). Faruki (1981) found
that O. bornovanus zoospores acquired and transmitted MNSV that contaminated the
surface of melon seed and found that while MNSV occurred in high incidence on the
surface of the seed it would not infect the seedling unless the vector was present,
suggesting vector-dependent seed transmission. Incidence of MNSV transmission when
externally contaminated melon seeds were sown in the presence of O. bornovanus was
demonstrated to be 10-40% (Faruki 1981). However, Campbell et al. (1996) reported
transmission of MNSV in melon seeds in the absence of O. bornovanus and their
findings suggest that seed transmission can be independent of the vector.
In conclusion, this study has determined the presence of both LBVaV and MLBVV on
commercial lettuce growing farms the Swan Coastal Plain of south-west Australia by
testing samples by DAS-ELISA as well as the presence of LBVaV and MLBVV at
different stages of production in commercial seedling nurseries in the Perth

111
metropolitan region by testing „bait‟ lettuce samples by DAS-ELISA and/or PCR. This
study has also determined a), the presence of O. virulentus resting spores, LBVaV and
MLBVV on the surface of lettuce seeds and, b), that even though O. virulentus resting
spores, LBVaV and MLBVV are present on lettuce seed, the number of propagules was
shown to be too low to cause systemic infection. However, by increasing the sample
size and growing a greater number of non-surface sterilised seeds harvested from plants
which have tested positive for LBVD into autoclaved soil, and testing these plants by
the more sensitive PCR method described in this chapter, the proposed vector-assisted
seed transmission is likely to be established as a source of effective LBVD inoculum.

112
Chapter 7 Alternative Hosts
7.1 Introduction
LBVaV and MLBVV are not limited to lettuce, although both viruses have a narrow
host range. Their chytrid vector, earlier identified as O. brassicae and now determined
to be O. virulentus (Sasaya and Koganezawa 2006) has a wide host range (Campbell
1965). Other known natural hosts of LBVaV and MLBVV come from the family
Asteraceae and include: S. oleraceous, S. asper, C. endiva (endive) and chickory
(Campbell 1965). Species known as experimental hosts are Chenopodium
amaranticolor, C. quinoa, Nicotiana benthamiana, N. clevlandii, N. occidentalis and
Tetragonia tetragonioides (Huijberts et al. 1990).
Weed species found in commercial production areas play an important role in the
survival and spread of many important plant viral pathogens because the virus/vector
inoculum potential of weeds may be increased by close association with nearby
susceptible crops (Bos, 1999). In Western Australia, Coutts et al. (2004) demonstrated
in a lettuce field, that clustering of LNYV in lettuce plants occurred at the crop edge
where there was an S. oleraceous source and that there was few incidences of LNYV
further away from the S. oleraceous source. S. asper has been shown to be a virus
reservoir with natural infections of Tomato yellow leaf curl Sardinia virus, which is
vectored by the whitefly Bemisia tabaci occurring in tomato growing fields (Fanigliulo
et al. 2007).
.

113
S. oleraceous and S. asper are both introduced weed species. Both occur in lettuce
growing fields in Western Australia with S. oleraceous being the most common species
observed during this study. S. oleraceous is also commonly found growing in lettuce
growing fields in Spain where Navarro et al. (2005) confirmed the presence of LBVaV
and MLBVV in S. oleraceous leaf tissue collected from lettuce fields.
Determining what alternative hosts occur in the Western Australian lettuce production
system is important in helping to understand the epidemiology of LBVD so that further
control measures can be established. Even though infected soil is considered the main
infection source of LBVD, the elimination of such weeds could remove this additional
virus infection source by reducing both the survival and the activity of the chytrid
vector (Jones, 2004).
In this chapter, both native and alien species from the family Asteraceae (Daisy Family)
were tested for susceptibility to LBVD. The alien species tested were S. oleraceous, S.
asper (rough sowthistle), Arctotheca calendula (capeweed) and Reichardia tingitana
(false sowthistle). The native species tested were S. hydrophilus (native sowthistle) and
Actites sp. (dune thistle). S. oleraceous and A. calendula are found in almost all
horticultural and agricultural areas of south-west Australia including the Swan Coastal
Plain. Both of these species are commonly found on disturbed sites such as cultivated
areas and road verges (CALM 2007) and during this study, both species were
commonly found growing amongst lettuce crops. S. asper is not as common as S.
oleraceous but is also found on the Swan Coastal Plain on disturbed sites (CALM
2007). Reichardia spp. are also found on the Swan Coastal Plain but are more common
in the Southwest Botanical Province (CALM 2007). They grow in sandy soils, alluvial

114
river flats and coastal dunes (CALM 2007). S. hydrophilus is also common along the
Swan Coastal Plain. However, it occurs near temporarily wet ground such as on the
edges of lakes and streams (CALM 2007). Actites spp. are also common to the Swan
Coastal Plain and occur in calcareous sand, coastal dunes, cliffs and winter-wet plains
(CALM 2007). During this study S. hydrophilus and Actites sp. and Reichardia sp. were
not observed in lettuce growing areas.

115
7.1.1 Aims
The aims of this study were
to identify alternative weed hosts for LBVaV and MLBVV growing on the
Swan Coastal Plain of south-west Australia
to establish whether two native species of Asteraceae are hosts of LBVaV and
MLBVV
to determine whether natural weed species collected in LBVD-affected lettuce
fields in Western Australia were infected with LBVaV and/or MLBVV

116
7.2 Materials and Methods
7.2.1 Determining alternative hosts for LBVV and MLBVV
Species chosen for this experiment on alternative hosts included two known hosts S.
asper and S. oleraceous as well as three untested but potential hosts, S. hydrophilus,
Reichardia sp. and Actites sp.. S. hydrophilus was chosen as a species to test for
susceptibility to LBVaV and MLBVV as it is a member of the same genus as the known
LBVaV and MLBVV hosts S. asper and S. oleraceous. An initial experiment revealed
infection of both viruses in S. hydrophilus. Therefore, Reichardia sp., Actites sp. and A.
calendula were additionally chosen as they are also members of the family Asteraceae,
a family within which all other known natural hosts are found, and also because seeds
for these species were readily available (7.2.7).
7.2.2 Samples collected from the field
A. calendula and S. oleraceous plants were collected at random from four different
commercial lettuce growing farms. Those sampled were from farms B, C, E and H. In
each instance, only young inside leaves of five week old plants were sampled from each
plant. There were no root samples collected for testing. Three A. calendula samples
were taken from Farm B, one from Farm C and three samples from Farm E. One S.
oleraceous sample was taken from Farm B, two samples were taken from Farm E and
two samples were taken from Farm H. None of the samples of either species showed
disease symptoms.

117
7.2.3 Source of seedlings and inoculum for initial experiment
A small scale experiment was initially conducted to determine if S. hydrophilus is a host
of MLBVV and LBVaV. Seedlings of S. hydrophilus were obtained from Geographe
Community Landcare Nursery in Dunsborough, Western Australia. Infested soil was
collected from Farm A, from a field in which lettuce plants had shown symptoms of
LBVD.
7.2.4 Growing conditions for inoculated seedlings for the initial experiment
Thirty three S. hydrophilus seedlings were grown as bait plants in soil infested with
LBVD. The seedlings were transplanted into separate 140mm diameter pots so that
there was one seedling per pot which contained one part potting mix and one part
LBVD infested soil (7.2.3). They were grown in a controlled environment growth room
facility at DAFWA, South Perth (Fig. 7.1). The seedlings were exposed to a 12h
photoperiod and maintained at a constant temperature of 18°C. The pots were placed on
free draining benches. Negative controls were set up by transplanting 25 seedlings so
that there was one seedling per pot into pots containing only potting mix. They were
transplanted at the same time and maintained under the same temperature and light
conditions as the experimental seedlings. Also, approximately five lettuce seedlings
were transplanted into infested soil at the same time to act as positive controls.

118
Fig. 7.1: S. hydrophilus seedlings used in the initial experiment that were inoculated by
growing them in soil contaminated with LBVD pathogens and grown in 140mm
diameter pots on free-draining benches in a controlled environment room. Seeds were
harvested from these plants (7.2.6).
7.2.5 Testing by DAS-ELISA of field collected samples and inoculated seedlings
used in the initial experiment
The field collected leaf samples were stored at 4°C in sealed plastic bags on the day of
collection. The following day, the samples were analysed by DAS-ELISA, (Clark and
Adams, 1977) as described in Chapter 2 (2.14). After a 46 day growth period, the
youngest leaf on each S. hydrophilus seedling was sampled and tested for LBVaV and
MLBVV by DAS-ELISA also as described in Chapter 2 (2.14). The antisera used and
the determination of positive results, was the same as described in Chapter 6 (6.2.7).

119
7.2.6 Source of seed
S. hydrophilus seeds were collected from mature plants from the initial experiment
(7.2.4). All other seeds, of Actites sp., Reichardia sp., S. oleraceous and S. asper, were
obtained from Kathryn McCarren at CSIRO Laboratories, Floreat, Western Australia
and were of Western Australian origin.
7.2.7 Germination of seeds and inoculation of seedlings used to determine
alternative hosts
Thirty seeds of each species listed in 7.2.1 were sown and germinated in a glasshouse.
Approximately three weeks after germination, the seedlings were transplanted into
separate 140mm pots where there was one seedling per pot grown in soil that contained
one part potting mix and one part LBVD infested soil. The LBVD infested soil was
from Farm A (7.2.3). The plants in pots were grown as described in 7.2.4. However, due
to technical failures associated with the controlled environment room, a constant
temperature and light intensity could not be maintained throughout the duration of the
experiment, rather the plants were sometimes exposed to periods of darkness and high
temperatures (occasionally exceeding 30°C) which reflected the outside temperature of
the day. One negative control plant for each species was transplanted at the same time
by sowing in sterile soil which had been autoclaved three times at 120°C for 40mins.
These plants were kept in a different growth room to prevent cross-contamination and
tested periodically for the presence of LBVaV and MLBVV by DAS-ELISA.

120
7.2.8 Sampling of plant tissue for DAS-ELISA
After approximately one month of growing in infested soil, the youngest leaf of each
plant and the roots of each plant were sampled separately and each sample tested
individually for the presence of LBVV and MLBVV by DAS-ELISA as described in
Chapter 2 (2.14) and 7.2.5. The root samples were washed to remove any soil and the
possible contamination of viruliferous zoospores on the root surface or in the soil.
7.2.9 RNA extraction of leaf tissue
To re-affirm ELISA results, one leaf sample from each of the negative control plants,
Actites sp., Reichardia sp., S. asper, S. hydrophilus and S. oleraceous, was taken for
RNA extraction and testing by RT-PCR for the presence of LBVaV and MLBVV. Also
tested were plants from each species that were grown in infested soil and showing viral
infection symptoms; Actites sp. plant 19, Reichardia sp. plant 5, S. asper plants 9 and
17, S. hydrophilus plant 6 and S. oleraceous plant 4. Also sampled as a positive control,
was one lettuce plant showing LBVD symptoms that had been grown in LBVD infested
soil and one lettuce plant that had been grown with the other negative control plants.
7.2.10 Reverse Transcription Polymerase Chain Reaction (RT-PCR)
The RNA extracts from each leaf sample described in 7.2.9 were tested for the presence
of both LBVaV and MLBVV by RT-PCR. A OneStep RT-PCR kit from Qiagen,
Australia was used. The primers chosen for this assay were VP248/VP249 (LBVaV)
and VP286/VP287 (MLBVV) (Navarro et al. 2004). The sequences for these primers

121
are as listed in Table 2.1. The PCR reactions were placed in a thermocycler with
conditions as described in Chapter 2 (2.3.2) and then run on a 1% agarose gel and
viewed using a transilluminator as described in Chapter 2 (2.5).

122
7.3 Results
7.3.1 Results of initial experiments
Of the 33 S. hydrophilus seedlings grown in infested soil, 28 tested positive by DAS-
ELISA for LBVaV and 4 tested positive by DAS-ELISA for MLBVV. Of the 25 S.
hydrophilus seedlings grown as negative controls in potting mix, 10 tested positive by
DAS-ELISA for LBVaV and 5 tested positive by DAS-ELISA for MLBVV (Table 7.1).
As the negative controls tested positive ie. were three times the average absorbance
reading of the test negative controls, there was no distinct negative control. However,
the positive controls were determined by comparing their absorbance to the level
observed with the positive lettuce control sample and also to the samples on the ELISA
plate where there was very little absorbance observed.
Table 7.1: Detection of LBVaV and MLBVV in initial experiments where S.
hydrophilus seedlings were transplanted into pots containing a mixture of potting mix
and LBVD pathogen infested soil and tested by DAS-ELISA for both LBVaV and
MLBVV.
Treatment LBVaV MLBVV
Potting mix 10/25 5/25
Potting mix and soil inoculum 28/30 4/33

123
7.3.2 Field samples
A sample from one A. calendula plant (E7) of the seven tested, was positive for
MLBVV by DAS-ELISA but not for LBVaV. This single detection is a tentative first
report of A. calendula as a host of MLBVV. Samples from four of the six S. oleraceous
plants collected tested positive for LBVaV (E10, E11, H1 and H2). Samples from plants
E11 and H2 also tested positive for MLBVV indicating a mixed infection. Plant
samples B3, B4, B5, B6, E8 and E9 did not test positive by DAS-ELISA for either
LBVaV or MLBVV (Table 7.2). Neither the plants that tested positive nor the plants
that tested negative showed any visible LBVD symptoms. The same positive control as
described in 7.2.4 was used in this experiment and positive absorbance readings were
based on this control.
Table 7.2: Presence of LBVaV and/or MLBVV using DAS-ELISA test on field
collected samples of A. calendula and S. oleraceous.
Sample namea
Species sampled LBVaV MLBVV
B3 A. calendula - -
B4 A. calendula - -
B5 A. calendula - -
B6 S. oleraceous - -
C2 A. calendula - -
E7 A. calendula - +
E8 A. calendula - -
E9 A. calendula - -
E10 S. oleraceous + -
E11 S. oleraceous + +
H1 S. oleraceous + -
H2 S. oleraceous + +
Total 4/12 3/12 aLetters indicate separate commercial lettuce growing properties that leaf samples were taken from and
the numbers represent the sample number.

124
7.3.3 Growth room experiment
Of the weed species tested, the Reichardia sp. tested positive for LBVaV in 27 of the 28
root samples tested, but only one out of the 28 corresponding leaf samples tested
positive. When the Reichardia sp. was tested for MLBVV, 26 of the 28 root samples
tested positive and 16 of the 28 leaf samples also tested positive. Of the leaf samples,
there were no plants with mixed infections, one plant with LBVaV only, 16 plants with
MLBVV only and eleven plants which did not test positive for either virus. Of the root
samples, 25 were recorded as having mixed infections whilst 2 had LBVaV alone and
one plant tested positive for only MLBVV. No Reichardia sp. root samples were
negative for both viruses (Table 7.3).
S. asper showed 100% infection for LBVaV in the root samples whilst seven out of the
22 corresponding leaf samples also tested positive for LBVaV. When tested for
MLBVV, S. asper also had 100% infection in root samples and 15 of the leaf samples
also tested positive for MLBVV. Six of the S. asper leaf samples had a mixed infection
of both viruses, two had LBVaV alone and eight tested positive for only MLBVV. Six
of the leaf samples tested negative for both LBVaV and MLBVV. Of the S. asper root
samples, 22 tested positive for both LBVaV and MLBVV. There were no samples in
which either virus was detected alone or not at all (Table 7.3).
S. oleraceous tested positive for LBVaV in 27 of the 30 root samples tested with five
out of the 30 corresponding leaf samples also testing positive. When tested for
MLBVV, S. oleraceous root samples all tested positive as did 26 of the 30 leaf samples.
Of the leaf samples, five had mixed infections, no samples were positive for LBVaV

125
alone, 13 samples were infected only with MLBVV and two leaf samples tested
negative for both viruses. Of the root samples, 28 had mixed infections, no samples
were positive for LBVaV alone, three samples were infected only with MLBVV and no
samples tested negative for both viruses (Table 7.3).
Of the native species tested, Actites sp. and S. hydrophilus, Actites sp. tested positive for
LBVaV in 25 of the 30 root samples and 5 of the 30 corresponding leaf samples. When
tested for MLBVV, Actites sp. showed 100% infection in the root samples and 17 of the
30 corresponding leaf samples tested positive. Of the leaf samples, one had a mixed
infection, four samples were positive for LBVaV alone, 15 samples were infected only
with MLBVV and seven leaf samples tested negative for both viruses. Of the root
samples, 25 had mixed infections, no samples were positive for LBVaV alone, five
samples were infected only with MLBVV and no samples tested negative for both
viruses (Table 7.3).
S. hydrophilus tested positive for LBVaV in 26 of the 30 root samples but failed to test
positive in any leaf tissue samples. When tested for MLBVV, S. hydrophilus tested
positive for 28 of the 30 root samples and tested positive and 16 of the 30 leaf samples
also tested positive. Of the leaf samples, none had mixed infections or were positive for
LBVaV alone, 16 samples were infected only with MLBVV and 13 leaf samples tested
negative for both viruses. Of the root samples, 16 had mixed infections, no samples
were positive for LBVaV alone, 13 samples were infected only with MLBVV and two
tested negative for both viruses (Table 7.3).

126
Table 7.3: The incidence of LBVaV and MLBVV tested by DAS-ELISA on leaf and root tissue of three alien and two native species of Asteraceae.
Plant species Conservation
status
Tissue
tested
LBVaV and
MLBVV
LBVaV
only
MLBVV
only
Neither
virus
Total
LBVaV
Total
MLBVV
Actites sp. Not threatened
Native host
Leaf
Root
1/30
25/30
4/30
0/30
15/30
5/30
7/30
0/30
5/30
25/30
16/30
30/30
Reichardia sp. Alien Leaf
Root
0/28
25/28
1/28
2/28
16/28
1/28
11/28
0/28
1/28
27/28
16/28
26/28
S. asper Alien Leaf
Root
6/22
22/22
2/22
0/22
8/22
0/22
6/22
0/22
7/22
22/22
15/22
22/22
S. hydrophilus Not threatened
Native host
Leaf
Root
0/30
16/30
0/30
0/30
16/30
13/30
13/30
2/30
0/30
15/30
16/30
28/30
S. oleraceous Alien Leaf
Root
5/30
28/30
0/30
0/30
22/30
3/30
3/30
0/30
5/30
26/30
27/30
30/30

127
7.3.4 Observed virus symptoms in plant leaves
The plants which were infected with LBVaV and MLBVV showed a wide range of
symptoms which were different from those previously associated with LBVD in lettuce
(Table 7.4). Actites sp. (plant number 19) tested positive for both LBVaV and MLBVV
and showed necrotic petiole streaking which was not present in the negative control
(Fig. 7.2a and b). This symptom was observed in many of the Actites sp. plants infected
with LBVD. The Reichardia sp. (Fig. 7.2c) showed the widest range of symptoms when
infected with LBVD including vein clearing, interveinal chlorosis and necrotic midrib
streaking (Fig. 7.2d) and necrotic leaf spotting and yellow blotching, (Fig. 7.2e). S.
hydrophilus (Fig. 7.2f) showed mild vein clearing when infected with LBVaV and
MLBVV (Fig. 7.2g). Necrotic petiole streaking was also observed in S. hydrophilus.
Two S. asper plants (Fig. 7.2h) showed severe vein clearing and vein banding (Fig.
7.2i). Both of these plants with symptoms were positive for both viruses in the root
tissue but only positive for MLBVV in the leaf tissue. Mild vein clearing and necrotic
stem streaking was also observed in S. asper (Fig. 7.2j). In S. oleraceous (Fig. 7.2k),
mild vein clearing was observed and interveinal chlorosis was present when only
LBVaV infection was detected.

128
Table 7.4: Plant foliage symptoms on the three alien and two native Asteraceae species
grown in soil infested with LBVD pathogens.
Plant species Symptoms on foliage
Actites sp. Necrotic petiole streaking
Reichardia sp. Necrotic vein spotting
Interveinal chlorosis
Yellow blotching
Vein clearing
Necrotic midrib streaking
S. asper Necrotic petiole streaking
Mild vein clearing
Severe vein clearing
Vein banding
S. hydrophilus Mild vein clearing
Necrotic petiole streaking
S. oleraceous Interveinal chlorosis
Mild vein clearing

129
Fig. 7.2: Response of Actites sp., Reichardia sp., S. hydrophilus, S. asper and S. oleraceous to inoculation with LBVaV and MLBVV a) Actites sp. negative control, b)
Actites sp. necrotic petiole streaking c) Reichardia sp. negative control d) Reichardia sp. interveinal chlorosis, vein clearing and necrotic midrib streaking e) Reichardia sp.
yellow blotching and necrotic vein spotting on midrib f) S. hydrophilus negative control g) S. hydrophilus mild vein clearing h) S. asper negative control i) S. asper vein
clearing and vein banding j) S. asper necrotic petiole streaking k) S. oleraceous negative control l) S. oleraceous interveinal chlorosis

130
7.3.5 Confirmation of DAS-ELISA results by RT-PCR
Of the six leaf samples that were grown in potting mix only and in a separate glasshouse
to the experimental plants, five tested negative to both LBVaV and MLBVV by RT-
PCR whilst S. asper tested positive for both MLBVV and LBVaV. Of the eight
experimental plants, Actites sp. plant 19, Reichardia sp. plants 5 and 27, S. asper plant
9, S. hydrophilus plant 6 and S. oleraceous plant 4 all tested positive for LBVaV whilst
S. asper plant 17 was negative for LBVaV (Fig. 7.3). All amplicons using LBVaV
primer pair VP248 (forward) and VP249 (reverse) gave the expected fragment size of
296bp (Fig. 7.3). Of the eight experimental plants, only four tested positive for
MLBVV; L. sativa, S. hydrophilus plant 6, Reichardia sp. plant 27 and Actites sp. plant
19 (Fig 7.4). All amplicons using MLBVV specific primer pair VP286 (forward) and
VP287 (reverse) gave the expected fragment size of 469bp (Fig. 7.4). The RT-PCR
results agree with the results for each of the plants when tested for both LBVaV and
MLBVV by DAS-ELISA (Table 7.5).
Table 7.5: Positive (+) and negative (-) results obtained for each of the experimental
plants that were tested for LBVaV and MLBVV by both RT-PCR and DAS-ELISA
RT-PCR DAS-ELISA
Species LBVaV MLBVV LBVaV MLBVV
L. sativa –ve - - - -
L. sativa +ve + + + +
S. hydrophilus –ve - - - -
S.hydrophilus 6 + + + +
S. asper –ve + + + +
S. asper 9 + - + -
S. asper 17 - + - +
Reichardia sp. –ve - - - -
Reichardia sp. 5 + - + -
Reichardia sp. 27 + + + +
Actites sp. –ve - - - -
Actites sp. 19 + + + +
S. oleraceous –ve - - - -
S. oleraceous 4 + - + -
M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16

131
Fig 7.3: Amplification of LBVaV isolated from leaves of various native and alien
Asteraceae species which had been grown in soil infested with LBVD and leaf samples
of the same species grown in potting mix only (negative control plants). Primers used
were VP248 and VP249 giving a 296bp amplicons. Lane M: 6µL of 100bp molecular
weight marker (Fisher Biotech), Lane 1: L. sativa negative control, Lane 2: L. sativa,
Lane 3: S. hydrophilus negative control, Lane 4: S. hydrophilus plant 6, Lane 5: S. asper
negative control, Lane 6: S. asper plant 9, Lane 7: S. asper plant 17, Lane 8: Reichardia
sp. negative control, Lane 9: Reichardia sp. plant 5, Lane 10: Reichardia sp. plant 27,
Lane 11: Actites sp. negative control, Lane 12: Actites sp. plant 19, Lane 13: S.
oleraceous negative control, Lane 14: S. oleraceous plant 4, Lane 15: water negative
control, Lane 16: cDNA positive LBVaV control.
500bp
296bp
M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
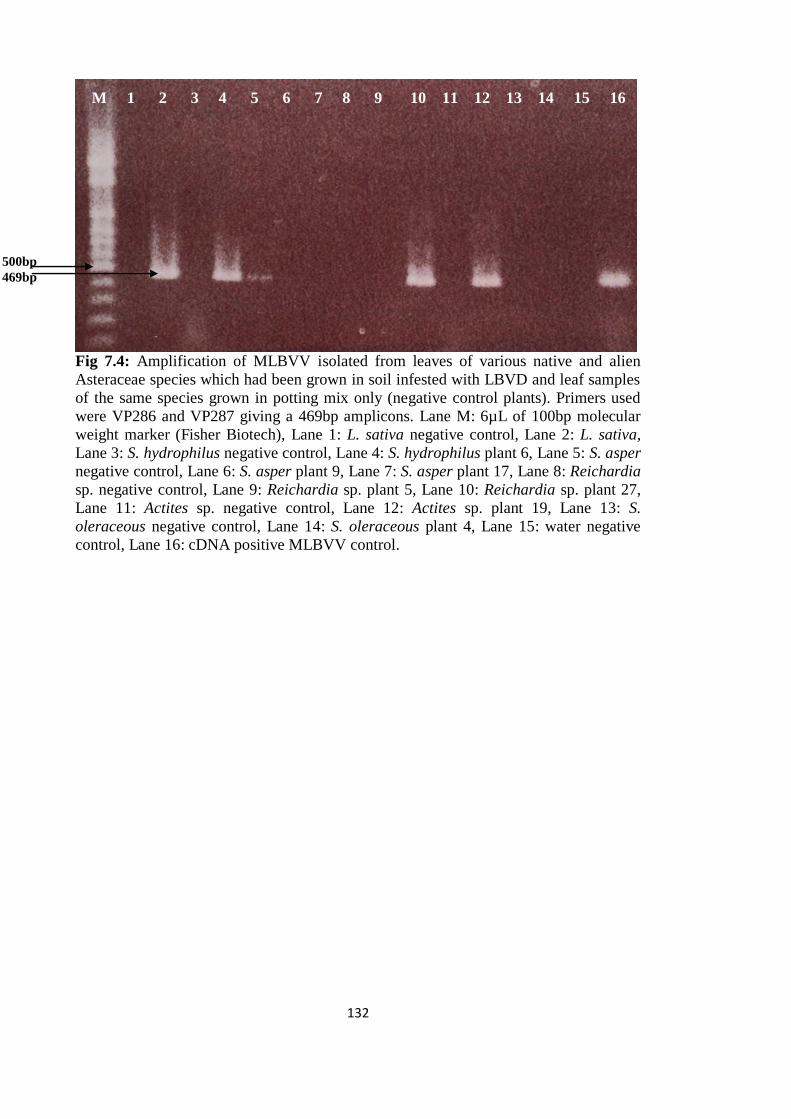
132
Fig 7.4: Amplification of MLBVV isolated from leaves of various native and alien
Asteraceae species which had been grown in soil infested with LBVD and leaf samples
of the same species grown in potting mix only (negative control plants). Primers used
were VP286 and VP287 giving a 469bp amplicons. Lane M: 6µL of 100bp molecular
weight marker (Fisher Biotech), Lane 1: L. sativa negative control, Lane 2: L. sativa,
Lane 3: S. hydrophilus negative control, Lane 4: S. hydrophilus plant 6, Lane 5: S. asper
negative control, Lane 6: S. asper plant 9, Lane 7: S. asper plant 17, Lane 8: Reichardia
sp. negative control, Lane 9: Reichardia sp. plant 5, Lane 10: Reichardia sp. plant 27,
Lane 11: Actites sp. negative control, Lane 12: Actites sp. plant 19, Lane 13: S.
oleraceous negative control, Lane 14: S. oleraceous plant 4, Lane 15: water negative
control, Lane 16: cDNA positive MLBVV control.
M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
500bp
469bp

133
7.4 Discussion
Four previously unreported hosts of MLBVV were determined in this study; A.
calendula, S. hydrophilus, Actites sp. and Reichardia sp. Three previously unreported
hosts of LBVaV were also determined in this study; S. hydrophilus, Actites sp. and
Reichardia sp. The report of infection in A. calendula is tentative as MLBVV was only
detected in one out of six samples. Of the six different Asteraceae species tested in this
experiment, S. asper was the most susceptible to LBVaV and MLBVV as it showed
100% detection from root tissue, with the Reichardia sp. being the second most
susceptible to LBVaV with 96% recovery from root tissue. The Actites sp. and S.
oleraceous are also highly susceptible to MLBVV also showing 100% infection in root
tissue.
There is very little information regarding the function of virus resistance and movement
in roots as it is commonly the leaf tissue that is studied. However, Germundsson et al.
(2002) suggested that virus infection in plant roots could induce systemic resistance and
prevent virus movement into the leaf tissue. Germundsson et al. (2002) inoculated the
roots of CP-transgenic N. benthamiana with Spongospora subterranea zoospores that
were carrying Potato mop-top virus (PMTV) and found that whilst there was PMTV
infection in the roots (as tested by ELISA and PCR), there was no systemic movement
to the above ground parts of the plant. Due to this finding, they concluded that roots and
leaves provide different expression of virus resistance. This is similar to the situation
seen in this study where in every species tested for LBVaV and MLBVV, infection was
more common in the root tissue of the plant than in the leaf tissue. This was most
obvious with the Reichardia sp. that was infected with LBVaV with 27/28 of the root

134
samples testing positive but with only 1/28 of the leaf samples testing positive. If such a
mechanism exists in regard to the systemic spread of LBVaV and MLBVV, then this
would be an important mechanism to exploit in the development of CP gene-mediated
resistant cultivars.
The diverse symptoms observed in the five Asteraceae species grown in LBVD infested
soil in the controlled environment growth room are largely due to the way in which the
virus/es interact with each of the different host species used. Both virus and host are
genetically variable. This genetic variability and the surrounding environmental
conditions affect their interactions. The extent of the severity of virus diseases depends
both upon the virus pathogenicity as well as the susceptibility of the host, which differs
depending on the species or cultivar (Bos 1999). The infected S. oleraceous plants
collected in the lettuce growing field were symptomless whereas the infected S.
oleraceous grown in the controlled environment room showed symptoms of interveinal
chlorosis and mild vein clearing. This can perhaps be explained by the low temperatures
and low light intensity which the controlled environment room plants were subjected to,
condition conducive for optimum symptom expression (Walsh 1994), rather than the
warmer and higher light intensity summer growing conditions in which the field
collected S. oleraceous and A. calendula were grown under. Navarro et al. (2005) found
that S. oleraceous that was infected with LBVD was symptomless whilst Campbell
(1965) described yellow vein banding and chlorosis. It is unclear which virus caused the
symptoms observed by Campbell (1965) as these were observed before the discovery of
MLBVV as the symptom causing virus associated with LBVD (Lot et al. 2002).

135
In the initial experiment, 28 out of 30 S. hydrophilus leaf tissue samples tested positive
for LBVaV and 16 out of 30 tested positive for MLBVV. However, in the following
experiment, S. hydrophilus leaf tissue samples failed to test positive for LBVaV and
only MLBVV was detected and even then in only four out of the 30 samples. The
inconsistency of these results may be associated with fluctuating temperatures within
the environment controlled growth facility. During the initial experiment, growth room
conditions remained constant. However, during the latter experiment, there were
fluctuations in the temperature due to mechanical failure. The experiment was done in
summer (January and February, 2008). During this period of mechanical failure,
daytime outside temperatures occasionally exceeded 40°C. In addition, Olpidium sp.
isolates infected with LBVaV have been reported to reproduce better between 10-16°C
than at 22°C and are known to reproduce poorly at 27°C (Fry and Campbell, 1966).
Teakle and Thomas (1985) showed that Olpidium sp. associated with LBVD multiplied
at 20°C but not at 29°C. Thus, temperature may have not only affected the interaction of
the host and the viruses but also the activities of the vector.
In the initial experiment, the negative control plants grown in un-infested potting mix
showed a higher number of S. hydrophilus plants infected with MLBVV in their leaves
than those grown in soil infested with LBVD. Four of the 33 plants grown in the
infested soil inoculum tested positive for MLBVV whereas five of the 25 plants grown
in a separate growth room tested positive for MLBVV. LBVaV was also detected in ten
of the 25 S. hydrophilus plants grown in un-infested potting mix. In each DAS-ELISA
test there were clear negative and positive control absorbance readings from the plate so
it is unlikely that these results were due to false positives. The growth room had
previously been used for growing LBVD infected lettuce and viruliferous O. virulentus

136
resting spores could have still been present on the benchtop and on the floor of the
growth room. Between experiments, the growth room had not been disinfected but just
swept to remove plant debris and soil.
That such contamination had occurred is supported by the observation in a previous
experiment using LBVD pathogen infested soil in that particular growth room, where a
lettuce seed which had germinated in the crack between the floor and the wall of the
growth room showed visible vein chlorosis, consistent with LBVD. It is also possible
that the potting mix used was already infested with viruliferous O. virulentus resting
spores. It is noteworthy in that in another experiment in this study, when potting mix
was sterilised by autoclaving it three times at 120°C for 40mins (Chapter 6, 6.3.5),
lettuce plants grown in new pots tested negative by DAS-ELISA for both LBVaV and
MLBVV. The negative control S. asper plant which was grown in a growth room
separate from the others also tested positive for both LBVaV and MLBVV, suggesting
not only is it the most susceptible species to both viruses, but also that the potting mix
used was in fact inadvertantly contaminated with viruliferous O. virulentus resting
spores.
This study also highlights the pathogenicity of O. virulentus, LBVaV and MLBVV and
the need for strict sanitation practices in lettuce production. The results from these
experiments have direct relevance for lettuce farmers and those involved in the
production of lettuce seedlings in a nursery environment. Jones (2004) proposed
integrated disease management strategies to minimise LBVD infection in lettuce crops,
lettuce seedling nurseries and nutrient film growing conditions. Jones (2004) suggested
that in areas of infested land, seedling nurseries and where nutrient film techniques are

137
employed, that weeds and „volunteer‟ crop plants be removed by selective herbicide or
by hand in order to remove both the internal and the external sources of the viruses and
the vector. It was also suggested that in seedling nurseries, all potting media, benches,
seedling trays and floors should be sterilised to avoid introducing new infections via the
vector‟s resting spores (Jones 2004).
It has been shown that one of the most common weeds found in lettuce growing fields
in south-west Australia, S. oleraceous, is highly susceptible to both LBVaV and
MLBVV with 26/30 root samples testing positive to and 30/30 root samples testing
positive to MLBVV. A. calendula is also a common weed in Western Australia and a
field sample of it tested positive for MLBVV on one occasion. Even though in some
cases the viruses are not moving systemically throughout the plant, these weeds can still
effectively act as reservoirs by providing a host for the Olpidium vector and the virus
species. In a system where crop rotation is being administered or where land is left
vacant, agricultural weeds may increase persistence of LBVD that is occurring anyway
through dormant resting spores (Navarro et al. 2005). By employing weed removal into
an Integrated Disease Management Strategy, the number of viruliferous resting spores
would be greatly reduced. These results have implications in the strategies for the
management of the disease in the Western Australian environment and therefore will be
discussed in greater detail in the General Discussion (Chapter 8).

138
Chapter 8 – General Discussion
This study has determined not only the identity and the phylogeny of MLBVV and
LBVaV and their vector O. virulentus, all associated with LBVD in Western Australia,
but also some aspects of the sources of inoculum of the viruses and the vector. A better
understanding of the pathogen/vector and the nature of the disease is expected to help in
improving strategies to minimise spread and/or the severity of the disease in Western
Australia and widely in the world.
The diagnosis of MLBVV associated with LBVD in Western Australia was not
unexpected as it has been established to be the causal agent of LBVD by previous work
done in Europe (van der Wilk et al. 2002; Navarro et al. 2004; Navarro et al. 2005a),
Japan, (Kawazu et al. 2003) and America (Hayes et al. 2006; Sanches 2006). It is
interesting that in this study lettuce, S. oleraceous and one plant of A. calendula were
the only species among those tested found naturally infected in the field. This indicates
that the natural host range of LBVaV and MLBVV may be limited although the disease
is widespread as suggested previously by others (Sahtiyacnci 1962; Campbell and Lin
1976; Hartwright et al. 2009). It not only confirms that LBVD is a serious disease of the
lettuce crop but also that it is able to spread and survive in the Western Australian
environment effectively as found previously (Latham et al. (2004); Latham and Jones
2004) .
The complete coat protein (CP) coding region sequences of 13 Western Australian
MLBVV isolates all fitted within clade A following phylogenetic examination of
MLBVV in this study involving isolates from across five continents which revealed two

139
clades, A and B. Twelve of the 13 Western Australian isolates belonged in
geographically distinct sub-clade A1 which contained no other isolates whilst the 14th
isolate from Western Australia fitted into sub-clade A3 which also included isolates
from Europe and Japan. This indicates that there were probably at least two
introductions of MLBVV into Western Australia. Clade B was found to contain only
isolates of MLBVV from Spain reflecting the Mediterranean region as the centre of
domestication of the crop (Whitaker 1969). Navarro et al. (2005b) previously reported
clade A and clade B but did not discriminate sub-clades as done in this study. It is
important to note that all indications in this thesis only relate to Western Australia in
relation to information on GenBank from elsewhere because concurrent work was not
carried out in relation to the distribution of this virus elsewhere in Australia.
It is also noteworthy that the CP coding region of MLBVV was highly conserved across
five continents. Considering that lettuce originated in Egypt and then moved next into
the southern region of the Mediterranean basin (Lindqvist 1960), it is likely that the
virus and the host were spread elsewhere from this region together. This may explain
why the CP coding region is so highly conserved.
The 12 Australian isolates that were more highly conserved than the rest may indicate
that the vector and isolates within sub-clade A1 arrived together and spread from a
common source within the Australia. Therefore, the distribution patterns would be
expected to be rather limited given the limited areas within Australia where there are
horticultural industries and the advent of agriculture only 200 years ago following
European colonisation. However, presence of the isolate in sub-clade A3 indicates a
second introduction of the virus. Determining the incidence of these two sub-clade

140
introductions would involve comparing isolates of MLBVV from horticultural areas
within Western Australia, South Australia, Victoria, Queensland and Tasmania where
lettuce is commercially grown to see if both members of sub-clades occur across the
continent.
This study also reports for the first time, the ability of two Australian native plant
species Actites sp. and S. hydrophilus to be hosts for MLBVV. As the screening of
native plants in this study was limited to available seed, it is not known how many other
native Australian plants could be symptomatic or asymptomatic carriers of MLBVV.
Such knowledge is important because south-west Australia experiences a Mediterranean
climate in which in summer all annual plants die unless they are irrigated [Reichardia
sp. are annual or perennial, S. asper are annual or biennial, S. hydrophilus is either
annual, biennial or perennial, S. oleraceous is annual and Actites sp. are perennial,
(CALM 2007)]. The perpetuation of the virus in these annual plants could be limited to
situations where they are externally seed-borne in the resting spores of O. virulentus
which may not account for the widespread occurrence of this virus. Even if there was
only limited carry-over of the virus on seed, the fact that Australian native plants can be
a reservoir for MLBVV could be significant for both the Australian environment and for
commercial lettuce production. Crop viruses such as Bean yellow mosaic virus (BYMV)
in Western Australia have been shown to cause symptoms such as generalised leaf
distortion, vein clearing, chlorotic mosaic and severe plant dwarfing in the native plant
Kennedia prostrata (Webster et al. 2007). Webster et al. (2007) also suggested that
humans facilitate the frequency by which native plant communities come into contact
with crop viruses and that this could affect biodiversity.

141
The exact role of LBVaV in LBVD still remains unclear as it is generally considered
that infection with this virus does not induce symptoms in lettuce (Colariccio et al.
2005; Roggero et al. 2003). Even when lettuce is infected, it is an asymptomatic carrier
of LBVaV (Lot et al. 2002). It is also not known whether this virus has an additive or
synergistic effect on the expression of the disease in the presence of MLBVV. However,
while Colariccio et al. (2005) and Roggero et al. (2003) both report no evidence of
synergism, Roggero et al. (2003) suggest this lack of synergism may reflect the poor
detection of the two viruses rather than proof of LBVaV on its own causing the
symptoms or adding to the level of symptom expression. In tests in which LBVaV and
MLBVV were baited out of infested soil using seedlings, 88% of the soils assayed in
this study from nurseries were positive for MLBVV and 69% of the samples were
positive for both LBVaV and MLBVV, suggesting a receptiveness from lettuce to both
of these viruses even though there is little indication that the two viruses are interacting
with each other.
This study was successful in obtaining seven complete CP coding region sequences of
different LBVaV isolates which resulted in the establishment of two clades using these
sequences and those available on GenBank. All isolates from Western Australia were
distinct from the others but were most closely related to the Japanese isolates of LBVaV
and TStV (clade II) than to LBVaV isolates from Europe (clade I). This study also
showed that the CP of LBVaV is even more highly conserved than the highly conserved
CP region of MLBVV. This may indeed reflect the possibility of LBVaV having existed
in Australia even before lettuce was introduced as a crop. This is similar to the origin
reported for LNYV. LNYV was first described in Australia and has been considered as
originating from plant species endemic to Australia (Randles and Carver 1971). Also

142
LBVaV may have stabilised as a latent virus in a variety of hosts including lettuce and
with little selection pressure by the host to modify itself to the extent that MLBVV may
have had. Another possibility for the highly conserved nature of the LBVaV CP coding
region is that it is a recent introduction and evolutionary processes are yet to have
occurred.
O. virulentus was associated with both LBVaV and MLBVV in this study but, what was
interesting, was that O. brassicae was only detected in one root extract out of more than
80 that were tested throughout the course of this study. This is also the first report of
LBVaV being associated with O. virulentus as previous work done by Sasaya and
Koganezawa (2006) investigated MLBVV and TStV, but not LBVaV.
It is not clear whether O. brassicae was associated with other hosts in this study. In
early studies prior to the molecular detection of O. virulentus, Olpidia commonly
carried on grasses and field crops, especially cucurbits, were not only misidentified as
O. brassicae but also as the vector of LBVaV and MLBVV. Considering recent work,
especially from Spain (Herrera-Vasquez et al. 2009), it is clear that O. brassicae is not
common in horticultural fields in several countries and even where it did occur (which
was only 4.5% of the plants studied), it occurred only on broccoli and cabbage both
belonging to the family Cruciferae. Hartwright et al. (2009) have described three
different Olpidium isolates based on phylogenetic analysis of the ITS region sequence
and host range. O. brassicae was found to infect broccoli but not lettuce or carrot whilst
one isolate failed to infect cucumber. Olpidium isolated from lettuce failed to infect
broccoli or carrot and one isolate did not infect cucumber. Olpidium isolated from carrot
infected lettuce but failed to infect broccoli or cucumber. From this study, Hartwright et

143
al. (2009) propose that the broccoli-infecting isolates still be referred to as O. brassicae
whilst the lettuce-infecting isolates be referred to O. compositae as they infect other
members of the family Compositae and that the third group of isolates found to infect
cucumber be referred to as O. cucurbitaceae. Based on morphological characteristics,
Hartwright et al. (2009) also challenge the suggestion of Sahtiyanci (1962) to include
these species of Olpidium in the genus Pleotrachelus.
O. bornovanus was observed on 53% of all the plants studied by Herrera-Vasquez
(2009) which were collected over a ten year period from Brazil, Guatemala, Honduras,
Mexico, Panama, Portugal, Spain, the Netherlands, Tunisia, Uruguay, and the USA. It
was clearly the most common of the three Olpidium species they studied. Although
more Olpidium species are described in the literature, the scientific verification of these
species is still to be carried out because most of these species have not yet been
identified through molecular methods. O. virulentus is apparently the vector for LBVD
in Western Australia. Although under natural conditions, O. virulentus has been
reported elsewhere on melon, watermelon and tomato in addition to lettuce, in this
study, under natural conditions, MLBVV was found only in lettuce, S. oleraceous and
once on A. calendula.
Complete rDNA ITS sequences of five Western Australian isolates of O. virulentus
were obtained in this study and the phylogenetic analysis showed that the Western
Australian isolates are highly conserved and this again may indicate the uniformity of
the population in Western Australia even though the sequenced isolates were found in
two sub-clades which suggests two possible introductions of O. virulentus. The first
sub-clade included four of the five isolates as well as isolates from Europe and Japan

144
whilst the second sub-clade contained one Western Australian isolate and one other
from Japan. It is interesting that Japanese isolates are represented in both sub-clades in
which the Western Australian isolates are associated and this suggests that it is possible
both came from the same source, originally in The Mediterranean. It seems unlikely that
these isolates were moved from Japan to Australia after European colonisation. Possibly
they occur concurrently in the Western Pacific Rim. This could be established only if a
wider study is carried out of the populations that occur in China and other countries in
this region where lettuce is grown.
Management strategies realistically are not about control of the disease but about
prevention of the spread of the LBVD to virgin farming land which is currently free of
the disease (if such areas exist) and the reduction of inoculum potential. So the
management of LBVD relates to two major points. Firstly, the study of the source(s) of
the inoculum to prevent the spread of the vector and/or the viruses from production
nurseries to lettuce growers who have started cultivation on virgin lands which are
assumed to be free of LBVD contamination. Secondly, a further reason why
management of LBVD is functional is evident in the work done by Latham et al. (2004)
which shows that early infection of lettuce seedlings with LBVD causes more severe
symptoms and greater yield loss than infections which occur later once the seedlings
have been planted in the field (Fig 8.1). Prevention of LBVD at the early stages lettuce
of production, for example, in commercial seedling nurseries, can increase yields
because even where the plants are subsequently exposed to field inoculum, this is likely
to cause later infections with smaller yield losses than when infections occur early as a
consequence of early seedling infection in nurseries (Latham et al. 2004).

145
Nursery Field
Commercial lettuce seedling nurseries Commercial lettuce farms
1.
Seeds 2.
Weeds
3.
Potting
mix 4.
Water
Infected seedlings
Reduced yield
New field
8.
Weed seeds
10.
Machinery
12.
Flood and
irrigation
13. Dust storms
5. Dust
9. Weeds
6. Dry crop debris
7. Infested field soil
11. Dry crop debris
Fig. 8.1: Flow chart representing the possible sources of inoculum of LBVD carried in/on O. virulentus both in the seedling nursery environment and
also in the lettuce production fields in Western Australia. These potential sources are indicated by the broken arrows whilst inoculum sources which
have been determined in this study or by Latham et al. (2004) have been indicated by solid arrows.

146
One of the main outcomes of this study is the finding that the seeds can carry
viruliferous O. virulentus resting spores externally on the seed coat. This is significant
because even with nursery hygiene, potting mix sterilisation and all other sanitary
precautions that should ideally be associated with lettuce seedling production in a
seedling nursery, lettuce seeds still can bring in the disease into the nursery (No. 1, Fig.
8.1). Surface sterilisation to eliminate viruliferous O. virulentus resting spores would be
required to overcome this. This is a significant finding in relation to the epidemiology of
LBVD. O. bornovanus which is the vector of MNSV has similarly been shown to be
externally seed-borne (Faruki 1981; Campbell et al. 1996). In addition to the
constituents of potting mix being a potential source of inoculum as reported by Latham
et al. (2004), seeds too should be considered as an alternative source of infection, which
could be negated by the use of surface sterilisation of the seeds prior to being coated by
applying methods described in Chapter 6 relating to use of a 2% aqueous NaOCl
solution and/or a10% aqueous Na3PO4 solution.
Hardy and Sivasithamparam (1988) reported that Western Australian nurseries carry
eight different species of Phytophthora and proposed that nurseries were the source of
new strains and new species of that pathogen in Western Australia. Similarly, it is
possible for production nurseries to supply lettuce seedlings grown from seeds that
could introduce new strains and/or species of viruses into the farming environment.
Although LBVaV and MLBVV have not been detected internally within lettuce seed (in
the embryo), external transmission in the vector resting spores could be serious even at
low concentrations because that can result in the introduction of LBVD to uninfected
land and production areas. In addition to the example already described in Chapter 6 of
vector dependent virus transmission in which O. bornovanus zoospores acquired and

147
transmitted MNSV that contaminated the surface of melon seed, a similar
seed/pathogen relationship is described with white rust on brassicas. Barbetti (1981)
reported that the seeds of Brassica rapa carrying oospores of Albugo candida (white
rust) borne externally could well function as the means of introducing this disease into
commercial oilseed brassica fields in areas where the disease never existed before.
Weeds may play an important role in the spread and the carry-over of the vector and the
virus despite the long term survival of viruliferous resting spores in soil. The fact that S.
oleraceous and A. calendula, two of the most common weed species in agricultural
regions of Western Australia, might be implicated in the spread and survival of LBVD
indicates that management of these weeds, even though it would not eradicate LBVD,
would help to reduce both the level of inoculum potential in the soil over a period of
years and subsequent incidence/severity of the disease (No. 9, Fig 8.1). Weed removal
was also suggested as part of disease management strategy for the control of TSWV in
Western Australia which weeds have been shown to act as virus reservoirs (Latham and
Jones 1997). Although weeds from nurseries were not tested in this study, given the
highly susceptible nature of S. oleraceous in particular to LBVD, weeds in nurseries are
a suspected source of inoculum (No. 2, Fig. 8.1) which justifies the management of
weeds already included in the disease management strategy for LBVD (Jones 2004) and
also suggested by Navarro et al. (2005b). While it is also possible that weeds or weed
seeds infected with LBVD might sometimes enter the seedling nursery environment in
potting mix purchased by the seedling nursery thereby introducing the virus (No. 3 Fig.
8.1), this remains to be determined (as indicated by the dashed arrow in Fig. 8.1).
Potting mix and peat purchased by seedling nurseries for composting and use in their

148
commercial mix have been shown to be a source of LBVD inoculum (Latham et al.
2004) and this is represented as a solid arrow in Fig. 8.1.
Previous studies done on the occurrence of LBVaV and MLBVV separately on
individual weed plants have only included S. oleraceous (Navarro et al. 2005b) and
found that LBVaV occurred as a single infection in six of the plants tested whilst
MLBVV was never found as a single infection but only as a double infection with
LBVaV in four of the plants tested. It would be valuable to know of any interaction
between the two viruses within other weed species as this would provide further
knowledge regarding LBVD epidemiology and may have additional implications on
weed removal management strategies. It is also noteworthy that while LBVaV seems
latent not only in lettuce but also in the experimental test hosts Nicotiana benthamiana
and N. clevelandii (Huijberts et al. 1990). It is also not known, as with lettuce, whether
the presence of this virus may predispose the weeds to MLBVV infection which may be
important in the epidemiology of LBVD.
Disinfection of soil and potting mix, surface disinfection of seed and the use of clean
water and clean production areas are all important in the management of LBVD in
seedling production nurseries in Western Australia (Latham et al. 2004). Latham et al.
(2004) have also shown that contaminated potting mix is a source of inoculum for
lettuce seedling nurseries (No.3, Fig. 8.1) and even though not practical on a large scale,
this study (Chapter 6) has demonstrated that autoclaving potting mix three times at
120°C for 40min is effective in eradicating O. virulentus resting spores. Sterilisation of
potting mix and soil within commercial seedling nurseries was suggested by Latham et
al. (2004) and Jones (2004) as an important disease management strategy.

149
Water is also a possible source of LBVD in seedling nurseries and the reduction of
LBVD, which is transmitted through water (No. 4 Fig. 8.1), could be managed by the
use of filtration, which has been used successfully by nurseries to manage the spread of
Phytophthora (Ufer et al. 2005), or water may be decontaminated by chlorination or by
exposure to UV light before coming into contact with susceptible plants
(Sivasithamparam and Goss 1981; Tomlinson 1988). In addition, another source of
LBVD inoculum both in the nursery environment and the field could be by wind
dispersal if O. virulentus is carried in windborne dust (Nos. 5 and 13 Fig. 8.1) and also
with viruliferous resting spores being associated with dry crop debris within the
seedling nursery (Jones, 2004), (No. 6, Fig. 8.1).
LBVD can enter commercial lettuce growing fields via a number of routes (Fig. 8.1).
The primary source of inoculum is with the infested field soil which seedlings are being
grown (No. 7, Fig. 8.1) and the level of inoculum in infested field soil may also be
increased if there is dry crop debris left in the field (No. 11, Fig. 8.1). As previously
discussed, weeds are a source of inoculum in lettuce growing fields but given that the
seeds of S. oleraceous have a similar morphology to lettuce seeds, for example
longitudinal ribs along the surface, it is reasonable to speculate that weed seeds could
also be a source of LBVD inoculum in lettuce growing fields (No. 8, Fig. 8.1).
Contaminated weed seeds could transmit LBVD not only to lettuce but also to other
susceptible weed species including A. calendula and facilitate both the spread and the
persistence of LBVD in the field. It is also possible that, as seen with studies done on
Phytophthora (Sivasithamparam and Goss 1981), O. virulentus resting spores carrying
LBVaV and/or MLBVV could be transmitted on farm machinery [also suggested by
Jones (2004) and also on the footwear of farm workers (No. 10 Fig. 8.1)]. Some

150
commercial lettuce growing farms investigated in this study cultivate more than one
farm in which lettuce is grown and machinery such as tractors, trucks and other vehicles
are driven onto and between farms without any implementation of disinfection
strategies.
If more annual and perennial native plants are discovered to be hosts of LBVaV and
MLBVV, even if they are found to be asymptomatic, then that may add another
complexity to the LBVD story in the long term management of the disease. It is
possible that native plants which have natural infections of LBVD act as reservoirs and
if they are found to be growing in native vegetation close to lettuce growing fields,
motile O. virulentus zoospores carrying both LBVaV and MLBVV could be transported
through water by a flooding event (No. 12, Fig. 8.1). As with the consideration for
contaminated water sources in the nursery environment, the same sterilisation or
filtration management options discussed earlier also apply in relation to irrigation water
used in lettuce fields (No. 12, Fig. 8.1). However, this is not likely to be practical where
irrigation water is used in large volumes.
In conclusion, this investigation has established the phylogenetic relationships of
LBVaV, MLBVV and O. virulentus isolates from Western Australia to those in other
parts of the world. This study has also associated viruliferous O. virulentus resting
spores as external contaminants of lettuce seed and identified four new hosts of
MLBVV [A. calendula (tentative host), S. hydrophilus, Reichardia sp. and Actites sp.]
along with three new hosts of LBVaV (S. hydrophilus, Reichardia sp. and Actites sp.).
This new knowledge obtained regarding the epidemiology of LBVD can help to
enhance the effectiveness of the existing integrated management strategy for this

151
disease in order to reduce its spread and impact in Western Australia both in seedling
nurseries and also in lettuce growing fields. However, it is essential that the information
presented within this thesis and previously done by others (Latham et al. 2004 and
Jones 2004) be made available to both lettuce seedling nurseries and lettuce farmers
through extension bulletins, seminars or individual farm visits so that producers have
access to the information they need for better control the disease.
152
Appendix 1 – Amount of PCR product or plasmid used in
sequencing reactions
Origin of DNA/RNA used to generate sequence data, the primers used and the amount
of each plasmid/PCR product used for each sequencing reaction.
Plant
origin
Primer set used
to generate
amplicon
Species
amplified
Plasmid or
PCR
product
Amount of
plasmid/PCR product
sequenced per reaction
(ng)
AUSA5 L1-21/L700-679 LBVaV PCR product 95
AUSA5 L511-531/L1210-
1185
LBVaV PCR product 75
AUSB1 L1-21/L700-679 LBVaV Plasmid 702
AUSB1 L511-531/L1210-
1185
LBVaV Plasmid 684
AUSB2 L1-21/L700-679 LBVaV Plasmid 664
AUSB2 L511-531/L1210-
1185
LBVaV Plasmid 740
AUSE4 L1-21/L700-679 LBVaV PCR product 72
AUSE4 L511-531/L1210-
1185
LBVaV Plasmid 698
AUSE5 L1-21/L700-679 LBVaV PCR product 159
AUSE5 L511-531/L1210-
1185
LBVaV Plasmid 684
AUSE6 L1-21/L700-679 LBVaV Plasmid 618
AUSE6 L511-531/L1210-
1185
LBVaV Plasmid 917
AUSG1 L1-21/L700-679 LBVaV Plasmid 662
AUSG1 L511-531/L1210-
1185
LBVaV Plasmid 492
AUSA1 M2-21/M899-880 MLBVV PCR product 56
AUSA1 VP286/VP289 MLBVV PCR product 66
AUSA2 M2-21/M899-880 MLBVV Plasmid 572
AUSA2 VP286/VP289 MLBVV Plasmid 636
AUSB2 M2-21/M899-880 MLBVV PCR product 120
AUSB2 VP286/VP289 MLBVV PCR product 48
AUSC1 M2-21/M899-880 MLBVV PCR product 35
AUSC1 VP286/VP289 MLBVV PCR product 47
AUSD1 M2-21/M899-880 MLBVV PCR product 49
AUSD1 VP286/VP289 MLBVV PCR product 89
AUSE1 M2-21/M899-880 MLBVV PCR product 44
AUSE1 VP286/VP289 MLBVV PCR product 40
AUSE2 M2-21/M899-880 MLBVV PCR product 37
AUSE2 VP286/VP289 MLBVV Plasmid 351

153
AUSE3 M2-21/M899-880 MLBVV PCR product 12
AUSE3 VP286/VP289 MLBVV Plasmid 432
AUSE4 M2-21/M899-880 MLBVV PCR product 41
AUSE4 VP286/VP289 MLBVV Plasmid 396
AUSE6 M2-21/M899-880 MLBVV PCR product 131
AUSE6 VP286/VP289 MLBVV Plasmid 610
AUSF1 M2-21/M899-880 MLBVV PCR product 24
AUSF1 VP286/VP298 MLBVV Plasmid 569
AUSG1 M2-21/M899-880 MLBVV PCR product 50
AUSG1 VP286/VP289 MLBVV PCR product 72
AUSG2 M2-21/M899-880 MLBVV PCR product 37
AUSG2 VP286/VP298 MLBVV Plasmid 421
Col1 Ob1/ITS4 O. virulentus Plasmid 530
Col1 ITS1/Ob2 O. virulentus PCR product 33
Col2 Ob1/ITS4 O. virulentus Plasmid 542
Col3 Ob1/ITS4 O. virulentus PCR product 33
Nan6 Ob1/ITS4 O. virulentus Plasmid 535
Nan6 ITS1/Ob2 O. virulentus PCR product 53
Nan7 Ob1/ITS4 O. virulentus Plasmid 614
Nan7 ITS1/Ob2 O. virulentus PCR product 50
Nan8 Ob1/ITS4 O. virulentus Plasmid 543
Nan8 ITS1/Ob2 O. virulentus PCR product 29
T123 Ob1/ITS4 O. virulentus PCR product 58
T123 ITS1/Ob2 O. virulentus PCR product 63

154
Appendix 2 – The reagents used in each of the different DAS-
ELISA solutions
Coating buffer in 1L dH2O
Na2CO3 1.59g
NaHCO3 2.93g
pH 9.6
PBST in 1L dH2O
NaCl 8.0g
KH2PO4 0.2g
Na2HPO4 1.15g
KCl 0.2g
PVP 20g
Tween 20 5.0g
pH 7.4
Substrate buffer in 1L dH2O
Diethanolamine 106.7mL
pH 9.8
Washing buffer in 5L dH2O
NaCl 40g
KH2PO4 1g
Na2HPO4 5.75g
KCl 1g
Tween 20 10g
Saline Wash Solution in 1L dH2O
NaCl 8.8g

155
Appendix 3 - O. virulentus ITS sequences (5' to 3')
Nan 6 (634bp) GQ304519
TCCGTAGGTGAACCTGCGGAAGGATCATTATAAAAAATTCGGGCTTGGGTTTAACCCAA
GACCTGCCCCCAAAAGGGACCTTTTGTGCACTGTAAAATTTTTTCACTTAAGCTTTGTC
TGATTGATATTTGTCCCGAAAGGGACAAGAATTTTAATCTAAAACAACTTTTAACAATG
GATCTCTTGGCTCTCGCATCGATGAAGAACGCAGCGAAATGCGATAAGTAATGTGAATT
GCAGAATTCAGTGAATCATCGAATCTTTGAACGCATATTGCGCTCTCCGGCATTCCGGA
GAGCATGCCTGTTTGAGTATTGGCCTAAACGAGAGTCTTCCTTTAAAGATATAAAAAAG
AACGTTAAGAACCCTCAGTTGAGACTTCTCGACAATTCATTGCCGAGTGTTTTAAAATG
CAATAGTACCGCCACGGTTTGCGAACTTGGAGTAATATGTGAGCCTGAAAAGGTGACCT
CTATTCTCCTCGGACCACGTAGACCTGTTAAGCGGGAAGTTTTTGAATGCCGAATTATG
GGAGACGGTGTTCAATTGTTCGAGTAAACGACGAACGAAATAAACTCAATCTCAAATCA
GGTAGGATTACCCGCTGAACTTAAGCATATCAATAAGCGGAGGA
Nan 7 (632bp) GQ304520
TCCGTAGGTGAACCTGCGGAAGGATCATTATAAAAAAGTCGGGCTTGGGTTTAACCCAA
GACCTGCCCCCAAAAGGGACCTTTTGTGCACTGTAAAATTTTTTCACTTAAGCTTTGTC
TGATGGATATTTGTACCGAAAGGGACAAGAATTTTAATCTAAAACAACTTTTAACAATG
GATCTCTTGGCTCTCGCATCGATGAAGAACGCAGCGAAATGCGATAAGTAATGTGAATT
GCAGAATTCAGTGAATCATCGAATCTTTGAACGCATATTGCGCTCTCCGGCATTCCGGA
GAGCATGCCTGTTTGAGTATTGGCCTAAACGAGAGTCTTCCTTTAAAGATATAAAAAAG
AACGTTAAGAACCCTCAGTTGAGACTTCTCGACAATTCATTGCCGAGTGTTTTAAAATG
CAATAGTACCGCCACGTTTGCGAACTTGGAGTAATATGTGAGCCTGAAAAGGTGACCTC
TATTCTCCTCGGACCACGTAGACCTGTTAAGCGGGAAGTTTTTGAATGCCGAATTATGG
GAGACGGTGTTCAATTGTTCGAGTAACGACGAACGAAATAAACTCAATCTCAAATCAGG
TAGGATTACCCGCTGAACTTAAGCATATCAATAAGCGGAGGA

156
Nan 8 (632bp) GQ304521
TCCGTAGGTGAACCTGCGGAAGGATCATTATAAAAAAGTCGGGCTTGGGTTTAACCCAA
GACCTGCCCCCAAAAGGGACCTTTTGTGCACTGTAAAATTTTTTCACTTAAACTTTGTC
TGATTGATATTTGTACCGAAAGGGACAAGAATTTTAATCTAAAACAACTTTTAACAATG
GATCTCTTGGCTCTCGCATCGATGAAGAACGCAGCGAAATGCGATAAGTAATGTGAATT
GCAGAATTCAGTGAATCATCGAATCTTTGAACGCATATTGCGCTCTCCGGCATTCCGGA
GAGCATGCCTGTTTGAGTATTGGCCTAAATGAGAATCATCCTTTAAAGATATAAAAAAG
AACGTTAAGAACCCTCAGTTGAGACTTCTCGACAATTCATTGCCGAGTGTTTTAAAATG
CAATAGTACCGCCACGTTTGCGAACTTGGAGTAATATGTGAGCCTGAAAAGGTGACCTC
TATTCTCCTCGGAGCACGTAGACCTGTTAAGCGGGAAGTTTTTGAATGCCGAATTATGG
GAGACGGTGTTCAATTGTTCGAGTAACGACGAACGAAATAAACTCAATCTCAAATCAGG
TAGGATTACCCGCTGAACTTAAGCATATCAATAAGCGGAGGA
Col 1 (633bp) GQ304523
TCCGTAGGTGAACCTGCGGAAGGATCATTATAAAAAATTCGGGCTTGGGTTTAACCCAA
GACCTGCCCCCAAAAGGGACCTTTTGTGCACTGTAAAATTTTTTCACTTAAGCTTTGTC
TGATTGATATTTGTACCGAAAGGGACAAGAATTTTAATCTAAAACAACTTTTAACAATG
GATCTCTTGGCTCTCGCATCGATGAAGAACGCAGCGAAATGCGATAAGTAATGTGAATT
GCAGAATTCAGTGAATCATCGAATCTTTGAACGCATATTGCGCTCTCCGGCATTCCGGA
GAGCATGCCTGTTTGAGTATTGGCCTAAATGAGAGTCTTCCTTTAAAGATATAAAAAAG
AACGTCAAGAACCCTCAGTTGAGACTTCTCGACAATTCATTGCCGAGTGTTTTAAAATG
CAATAGTACCGCCACGGTTTGCGAACTTGGAGTAATATGTGAGCCTGAAAAGGTGACCT
CTATTCTCCTCGTACCACGTAGACCTGTTAAGCGGGAAGTTTTTGAATGCCGAATTATG
GGAGACGGTGTTCAATTGTTCGAGTAACGACGAACGAAATAAACTCAATCTCAAATCAG
GTAGGATTACCCGCTGAACTTAAGCATATCAATAAGCGGAGGA
T123 (633bp) GQ304522
TCCGTAGGTGAACCTGCGGAAGGATCATTATAAAAAATTCGGGCTTGGGTTTAACCCAA
GACCTGCCCCCAAAAGGGACCTTTTGTGCACTGTAAAATTTTTTCACTTAAGCTTTGTC
TGATTGATATTTGTACCGAAAGGGACAAGAATTTTAATCTAAAACAACTTTTAACAATG
GATCTCTTGGCTCTCGCATCGATGAAGAACGCAGCGAAATGCGATAAGTAATGTGAATT
GCAGAATTCAGTGAATCATCGAATCTTTGAACGCATATTGCGCTCTCCGGCATTCCGGA
GAGCATGCCTGTTTGAGTATTGGCCTAAATGAGAGTCTTCCTTTAAAGATATAAAAAAG
AACGTCAAGAACCCTCAGTTGAGACTTCTCGACAATTCATTGCCGAGTGTTTTAAAATG
CAATAGTACCGCCACGGTTTGCGAACTTGGAGTAATATGTGAGCCTGAAAAGGTGACCT
CTATTCTCCTCGGACCATGTAGACTTGTTAAGCGGGAAGTTTTTGAATGCCGAATTATG
GGAGACGGTGTTCAATTGTTCGAGTAACGACGAACGAAATAAACTCAATCTCAAATCAG
GTAGGATTACCCGCTGAACTTAAGCATATCAATAAGCGGAGGA

157
Col 2 (577bp) GQ328050
CAAGACCTGCCCCCAAAAGGGACCTTTTGTGCACTGTAAAATTTTTTCACTTAAGCTTT
GTCTGATTGATATTTGTACCGAAAGGGACAAGAATTTTAATCTAAAACAACTTTTAACA
ATGGATCTCTTGGCTCTCGCATCGATGAAGAACGCAGCGAAATGCGATAAGTAATGTGA
ATTGCAGAATTCAGTGAATCATCGAATCTTTGAACGCATATTGCGCTCTCCGGCATTCC
GGAGAGCATGCCTGTTTGAGTATTGGCCTAAATGAGAGTCTTCCTTTAAAGATATAAAA
AAGAGCGTCAAGAACCCTCAGTTGAGACTTCTCGACAATTCATTGCCGAGTGTTTTAAA
ATGCAATAGTACCGCCACGTTTGCGAACTTGGAGTAATATGTGAGCCTGAAAAGGTGAC
CTCTATTCTCCTCGGAGCACGTAGACCTGTTAAGCGGGAAGTTTTTGAATGCCGAATTA
TGGGAGACGGTGTTCAATTGTTCGAGTAACGACGAACGAAATAAACTCAATCTCAAATC
AGGTAGGATTACCCGCTGAACTTAAGCATATCAATAAGCGGAGGAA
Col 3 (504bp) GQ328051
AAAGGGACCTTTTGTGCACTGTAAAATTTTTTCACTTAAGCTTTGTCTGATTGATATTT
GTACCGAAAGGGACAAGAATTTTAATCTAAAACAACTTTTAACAATGGATCTCTTGGCT
CTCGCATCGATGAAGAACGCAGCGAAATGCGATAAGTAATGTGAATTGCAGAATTCAGT
GAATCATCGAATCTTTGAACGCATATTGCGCTCTCCGGCATTCCGGAGAGCATGCCTGT
TTGAGTATTGGCCTAAACGAGAGTCTTCCTTTAAAGATATAAAAAAGAACGTTAAGAAC
CCTCAGTTGAGACTTCTCGACAATTCATTGCCGAGTGTTTTAAAATGCAATAGTACCGC
CACGTTTGCGAACTTGGAGTAATATGTGAGCCTGAAAAGGTGACCTCTATTCTCCTCGG
ACCACGTAGACCTGTTAAGCGGGAAGTTTTTGAATGCCGAATTATGGGAGACGGTGTTC
AATTGTTCGAGTAACGACGAACGAAATAAACT

158
Appendix 4 - MLBVV CP sequences (5' to 3')
AUSG2 GU139114
ATGTCAGGAGTATACAAGGTTTCCGAAATTCAGTCTATCTTGCAAAAAGATGTAACTTC
AGAAGGAGAAACAGCTATTCTAATTTCTCTTGGTCTCATGACAAAAGAAGAGAAGCCTG
TTCCTGCAAAAATGGCCATGGTGGCATCTGCAAAAGCAAACTCAATCATCTTTGTTTCG
GAAGATGGCTCTTTGTCTTTTGAAGCTCCAAAAGAAACAGGAGAGACCAGCAAACCAGG
AGAGAAGAAAGAGGAAAAGAAGGTAGAAGTGGGAGTTAAGTTTCCTTTCAGCGCAGCCA
AAGTAAAGGAGCTAATTGAAGGGAAAAGTCTTACTTTGGATCAGGACAAAATCCAAAAA
GTGCTGGAAGAATATGTTAAGAATTTGCCAAGGACTGCTGAGACTTACAAACCAAAGGA
GATTGAGATCAAATGTTTCAAGGGTGTTGACTTCAGTATCAGCAGTTTGCTTTCTTCAG
GGACCAAAATCTTAGATGCTATTCTTTACAGTACTTACAAGGATTCAGCAGAGCACAAC
TTCATATTTGATGTGAAAGTTCTATCTCCTGATTTCATCGATAGCAAGTTACTCGTGAA
CAACATCGAAACAGGCAATCGAGCAATCAAAGCAGCTTTCTGTCTTGTTTACAATCAAG
GTGGATTGCCATCAAAGACGAGTGAAGAACGACCACTGTCTAAGTTTGTAAGAGAAACG
ATATTCCGTGAGAAAGATCTCAAAGCTAACGAGTTATGTGAATACCTATCATCAGCAGA
TCCTTCTTTGTTTCCAAGTCAAGTCTTTTTGAAAATCTCACTTGAAAACCTTCCTACTG
AGGTTTCATCACGTTGCAAGATGTCGATTGCGGGCAACAAAGCATTGAGATATGCACTC
TTAGCTCAAAAGTTTGACAAAGATGAAATTCCAGTTCCAACAGAAGTGAATCCTACAAC
TAGCTCAGAATACATGCAGAAAAAGGAGAAAATAGAAAAAGCAAAAAAGATAGTTGATG
TTCTATGTTCTCTTGCTTCTGACTTCCAGGCACAAGTGAAAATGCATCCTCTCTCCCCT
GAGAGATCATCGAGGAAGAATTTCACTCTGCAATTGACTTCTGCAATTGTTACTTCACT
TTCCTACAAAGGGAGGTTAGACATGAGAAAAGCAATCGAAGAGAAAAAGATAGAGGCTT
TCAAAAGAGATGAAAATATATTTGGAAGGTTAAATGCTCTTGGACAACCCACGTTTCCT
GTTCTGACTAACGCAGATGCTGACTTTTCTGAATTGTCAGTTGAGGCCGTGAAGACAGC
TTACGGAAAGAAA
AUSE3 GU139115
ATGTCAGGAGTATACAAGGTTTCCGAAATTCAGTCTATCTTGCAAAAAGATGTAACTTC
CGAAGGAGAAACAGCTATTCTAATTTCTCTTGGTCTCATGACAAAAGAAGAGAAGCCTG
TTCCTGCAAAAATGGCCATGGTGGCATCTGCAAAAGCAAACTCAATCATCTTTGTTTCG
GAAGATGGCTCTTTGTCTTTTGAAGCTCCAAAAGAAACAGGAGAGACCAGCAAACCAGG
AGAGAAGAAAGAGGAAAAGAAGGTAGAAGTGGGAGTCAAGTTTCCTTTCAGCGCAGCCA
AAGTAAAGGAGCTAATTGAAGGGAAAAGTCTTACTTTGGATCAGGACAAAATCCAAAAA
GTGCTGGAAGAATATGTTAAGAATTTGCCAAGGACTGCTGAGACTTACAAACCAAAGGA
GATTGAGATCAAATGTTTCAAGGGTGTTGACTTCAGTATCAGCAGTTTGCTTTCTTCAG
GGACCAAAATCTTAGATGCTATTCTTTACAGTACTTACAAGGATTCAGCAGAGCACAAC
TTCATATTTGATGTGAAAGTTCTATCTCCTGATTTCATCGATAGCAAGTTACTCGTGAA
CAACATCGAAACAGGCAATCGAGCAATCAAAGCAGCTTTCTGTCTTGTTTACAATCAAG

159
GTGGATTGCCATCAAAGACGAGTGAAGAACGACCACTGTCTAAGTTTGTAAGAGAAACG
ATATTCCGTGAGAAAGATCTCAAAGCTAACGAGTTATGTGAATACCTATCATCAGCAGA
TCCTTCTTTGTTTCCAAGTCAAGTCTTTTTGAAAATCTCACTTGAAAACCTTCCTACTG
AGGTTTCATCACGTTGCAAGATGTCGATTGCGGGCAACAAAGCAATGAGATATGCACTC
TTAGCTCAAAAGTTTGACAAAGATGAAATTCCAGTTCCAACAGAAGTGAATCCTACAAC
TAGCTCAGAATACATGCAGAAAAAGGAGAAAATAGAAAAGGCAAAAAAGATAGTTGATG
TTCTATGTTCTCTTGCTTCTGACTTCCAGGCACAAGTGAAAATGCATCCTCTCTCCCCT
GAGAGATCATCGAGGAAGAATTTCACTCTGCAATTGACTTCTGCAATTGTTACTTCACT
TTCCTACAAAGGGAGGTTAGACATGAGAAAAGCAATCGAAGAGAAAAAGATAGAGGCTT
TCAAAAGAGATGAAAATATATTTGGAAGGTTAAATGCTCTTGGACAACCCACGTTTCCT
GTTCTGACTAACGCAGATGCTGACTTTTCTGAATTGTCAGTTGAGGCCGTGAAGACAGC
TTACGGAAAGAAA
AUSE6 GU139116
ATGTCAGGAGTATACAAGATTTCCGAAATTCAGTCTATCTTGCAAAAAGATGTAACTTC
CGAAGGAGAAACAGCTATTCTAATTTCTCTTGGTCTCATGACAAAAGAAGAGAAGCCTG
TTCCTGCAAAAATGGCCATGGTGGCATCTGCAAAAGCAAACTCAATCATCTTTGTTTCG
GAAGATGGCTCTTTGTCTTTTGAAGCTCCAAAAGAAACAGGAGAGACCAGCAAACCAGG
AGAGAAGAAAGAGGAAAAGAAGGTAGAAGTGGGAGTCAAGTTTCCTTTCAGCGCAGCCA
AAGTAAAGGAGCTAATTGAAGGGAAAAGTCTTACTTTGGATCAGGACAAAATCCAAAAA
GTGCTGGAAGAATATGTTAAGAATTTGCCAAGGACTGCTGAGACTTACAAACCAAAGGA
GATTGAGATCAAATGTTTCAAGGGTGTTGACTTCAGTATCAGCAGTTTGCTTTCTTCAG
GGACCAAAATCTTAGATGCTATTCTTTACAGTACTTACAAGGATTCAGCAGAGCACAAC
TTCATATTTGATGTGAAAGTTCTATCTCCTGATTTCATCGATAGCAAGTTACTCGTGAA
CAACATCGAAACAGGCAATCGAGCAATCAAAGCAGCTTTCTGTCTTGTTTACAATCAAG
GTGGATTGCCATCAAAGACGAGTGAAGAACGACCACTGTCTAAGTTTGTAAGAGAAACG
ATATTCCGTGAGAAAGATCTCAAAGCTAACGAGTTATGTGAATACCTATCATCAGCAGA
TCCTTCTTTGTTTCCAAGTCAAGTCTTTTTGAAAATCTCACTTGAAAACCTTCCTACTG
AGGTTTCATCACGTTGCAAGATGTCGATTGCGGGCAACAAAGCAATGAGATATGCACTC
TTAGCTCAAAAGTTTGACAAAGATGAAATTCCAGTTCCAACAGAAGTGAATCCTACAAC
TAGCTCAGAATACATGCAGAAAAAGGAGAAAATAGAAAAAGCAAAAAAGATAGTTGATG
TTCTATGTTCTCTTGCTTCTGACTTCCAGGCACAAGTGAAAATGCATCCTCTCTCCCCT
GAGAGATCATCGAGGAAGAATTTCACTCTGCAATTGACTTCTGCAATTGTTACTTCACT
TTCCTACAAAGGGAGGTTAGACATGAGAAAAGCAATCGAAGAGAAAAAGATAGAGGCTT
TCAAAAGAGATGAAAATATATTTGGAAGGTTAAATGCTCTTGGACAACCCACGTTTCCT
GTTCTGACTAACGCAGATGCTGACTTTTCTGAATTGTCAGTTGAGGCCGTGAAGACAGC
TTACGGAAAGAAA

160
AUSF1 GU139117
ATGTCAGGAGTATACAAGGTTTCCGAAATTCAGTCTATCTTGCAAAAAGATGTAACTTC
CGAAGGAGAAACAGCTATTCTAATTTCTCTTGGTCTCATGACAAAAGAAGAGAAGCCTG
TTCCTGCAAAAATGGCCATGGTGGCATCTGCAAAAGCAAACTCAATCATCTTTGTTTCG
GAAGATGGCTCTTTGTCTTTTGAAGCTCCAAAAGAAACAGGAGAGACCAGCAAACCAGG
AGAGAAGAAAGAGGAAAAGAAGGTAGAAGTGGGAGTCAAGTTTCCTTTCAGCGCAGCCA
AAGTAAAGGAGCTAATTGAAGGGAAAAGTCTTACTTTGGATCAGGACAAAATCCAAGAA
GTGCTGGAAGAATATGTTAAGAATTTGCCAAGGACTGCTGAGACTTACAAACCAAAGGA
GATTGAGATCAAATGTTTCAAGGGTGTTGACTTCAGTATCAGCAGTTTGCTTTCTTCAG
GGACCAAAATCTTAGATGCTATTCTTTACAGTACTTACAAGGATTCAGCAGAGCACAAC
TTCATATTTGATGTGAAAGTTCTATCTCCTGATTTCATCGATAGCAAGTTACTCGTGAA
CAACATCGAAACAGGCAATCGAGCAATCAAAGCAGCTTTCTGTCTTGTTTACAATCAAG
GTGGATTGCCATCAAAGACGAGCGAAGAACGACCACTGTCTAAGTTTGTAAGAGAAACG
ATATTCCGTGAGAAAGATCTCAAAGCTAACGAGTTATGTGAATACCTATCATCAGCGGA
TCCTTCTTTGTTTCCAAGTCAAGTCTTTTTGAAAATCTCACTTGAAAACCTTCCTACTG
AGGTTTCATCACGTTGCAAGATGTCGATTGCGCGCAACAAAGCAATGAGATATGCACTC
TTAGCTCAAAAGTTTGACAAAGATGAAATTCCAGTTCCAACAGAAGTGAATCCTACAAC
TAGCTCAGAATACATGCAGAAAAAGGAGAAAATAGAAAAAGCAAAAAAGATAGTTGATG
TTCTATGTTCTCTTGCTTCTGACTTCCAGGCACAAGTGAAAATGCATCCTCTCTCCCCT
GAGAGATCATCGAGGAAGAATTTCACTCTGCAATTGACTTCTGCAATTGTTACTTCACT
TTCCTACAAAGGGAGGTTAGACATGAGAAAAGCAATCGAAGAGAAAAAGATAGAGGCTT
TCAAAAGAGATGAAAATATATTTGGAAGGTTAAATGCTCTTGGACAACCCACGTTTCCT
ATTCTGACTAACGCAGATGCTGACTTTTCTGAATTGTCAGTTGAGGCCGTGAAGACAGC
TTACGGAAAGAAA
AUSB2 GU139118
ATGTCAGGAGTATACAAGGTTTCCGAAATTCAGTCTATCTTGCAAAAAGATGTAACTTC
CGAAGGAGAAACAGCTATTCTAATTTCTCTTGGTCTCATGACAAAAGAAGAGAAGCCTG
TTCCTGCAAAAATGGCCATGGTGGCATCTGCAAAAGCAAACTCAATCATCTTTGTTTCG
GAAGATGGCTCTTTGTCTTTTGAAGCTCCAAAAGAAACAGGAGAGACCAGCAAACCAGG
AGAGAAGAAAGAGGAAAAGAAGGTAGAAGTGGGAGTCAAGTTTCCTTTCAGCGCAGCCA
AAGTAAAGGAGCTAATTGAAGGGAAAAGTCTTACTTTGGATCAGGACAAAATCCAAAAA
GTGCTGGAAGAATATGTTAAGAATTTGCCAAGGACTGCTGAGACTTACAAACCAAAGGA
GATTGAGATCAAATGTTTCAAGGGTGTTGACTTCAGTATCAGCAGTTTGCTTTCTTCAG
GGACCAAAATCTTAGATGCTATTCTTTACAGTACTTACAAGGATTCAGCAGAGCACAAC
TTCATATTTGATGTGAAAGTTCTATCTCCTGATTTCATCGATAGCAAGTTACTCGTGAA
CAACATCGAAACAGGCAATCGAGCAATCAAAGCAGCTTTCTGTCTTGTTTACAATCAAG
GTGGATTGCCATCAAAGACGAGTGAAGAACGACCACTGTCTAAGTTTGTAAGAGAAACG
ATATTCCGTGAGAAAGATCTCAAAGCTAACGAGTTATGTGAATACCTATCATCAGCAGA
TCCTTCTTTGTTTCCAAGTCAAGTCTTTTTGAAAATCTCACTTGAAAACCTTCCTACTG
AGGTTTCATCACGTTGCAAGATGTCGATTGCGGGCAACAAAGCAATGAGATATGCACTC

161
TTAGCTCAAAAGTTTGACAAAGATGAAATTCCAGTTCAACAGAAGTGAATCCTACAACT
AGCTCAGAATACATGCAGAAAAAGGAGAAAATAGAAAAAGCAAAAAAGATAGTTGATGT
TCTATGTTCTCTTGCTTCTGACTTCCAGGCACAAGTGAAAATGCATCCTCTCTCCCCTG
AGAGATCATCGAGGAAGAATTTCACTCTGCAATTGACTTCTGCAATTGTTACTTCACTT
TCCTACAAAGGGAGGTTAGACATGAGAAAAGCAATCGAAGAGAAAAAGATAGAGGCTTT
CAAAAGAGATGAAAATATATTTGGAAGGTTAAATGCTCTTGGACAACCCACGTTTCCTG
TTCTGACTAACGCAGATGCTGACTTTTCTGAATTGTCAGTTGAGGCCGTGAAGACAGCT
TACGGAAAGAAA
AUSA1 GU139119
ATGTCAGGAGTATACAAGGTTTCCGAAATTCAGTCTATCTTGCAAAAAGATGTAACTTC
CGAAGGAGAAACAGCTATTCTAATTTCTCTTGGTCTCATGACAAAAGAAGAGAAGCCTG
TTCCTGCAAAAATGGCCATGGTGGCATCTGCAAAAGCAAACTCAATCATCTTTGTTTCG
GAAGATGGCTCTTTGTCCTTTGAAGCTCCAAAAGAAACAGGAGAGACCAGCAAACCAGG
AGAGAAGAAAGAGGAAAAGAAGGTAGAAGTGGGAGTCAAGTTTCCTTTCAGTGCAGCCA
AAGTAAAGGAGCTAATTGAAGGGAAAAGTCTTACTTTGGATCAGGACAAAATCCAAAAA
GTGCTGGAAGAATATGTTAAGAATTTGCCAAGGACTGCTGAGACTTACAAACCAAAGGA
GATTGAGATCAAATGTTTCAAGGGTGTTGACTTCAGTATCAGCAGTTTGCTTTCTTCAG
GGACCAAAATCTTAGATGCTATTCTTTACAGTACTTACAAGGATTCAGCAGAGCACAAC
TTCATATTTGATGTGAAAGTTCTATCTCCTGACTTCATCGATAGCAAGTTACTCGTGAA
CAACATCGAAACAGGCAATCGAGCAATCAAAGCAGCTTTCTGTCTTGTTTACAATCAAG
GTGGATTGCCATCAAAGACGAGTGAAGAACGACCACTGTCTAAGTTTGTAAGAGAAACG
ATATTCCGTGAGAAAGATCTCAAAGCCAACGAGTTATGTGAATACCTATCATCAGCAGA
TCCTTCTTTGTTTCCAAGTCAAGTCTTTTTGAAAATCTCACTTGAAAACCTTCCAACTG
AGGTTTCATCACGTTGCAAGATGTCGATTGCGGGCAACAAAGCAATGAGATATGCACTC
TTAGCTCAAAAGTTTGACAAAGATGAAATTCCAGTTCCAACAGAAGTGAATCCTACAAC
TAGCTCAGAATACATGCAGAAAAAGGAGAAAATAGAAAAAGCAAAAAAGATAGTTGATG
TTCTATGTCTTCTTGCTTCTGACTTCCAGGCCCAAGTGAAAATGCATCCTCTCTCCCCT
GAGAGATCATCGAGGAAGAATTTCACTCTGCAATTGACTTCTGCAATTGTTACTTCACT
TTCCTACAAAGGGAGGTTAGACATGAGAAAAGCAATCGAAGAGAAAAAGATAGAGGCTT
TCAAAAGAGATGAAAATATATTTGGAAGGTTAAATGCTCTTGGACAACCCACGTTTCCT
GTTCTGACTAACGCAGATGCTGACTTTTCTGAATTGTCAGTTGAGGCCGTGAAGACAGC
TTACGGAAAGAAA

162
AUSE4 GU139120
ATGTCAGGAGTATACAAGGTTTCCGAAATTCAGTCTATCTTGCAAAAAGATGTAACTTC
CGAAGGAGAAACAGCTATTCTAATTTCTCTTGGTCTCATGACAAAAGAAGAGAAGCCTG
TTCCTGCAAAAATGGCCATGGTGGCATCTGCAAAAGCAAACTCAATCATCTTTGTTTCG
GAAGATGGCTCTTTGTCCTTTGAAGCTCCAAAAGAAACAGGAGAGACCAGCAAACCAGG
AGAGAAGAAAGAGGAAAAGAAGGTAGAAGTGGGAGTCAAGTTTCCTTTCAGTGCAGCCA
AAGTAAAGGAGCTAATTGAAGGGAAAAGTCTTACTTTGGATCAGGACAAAATCCAAAAA
GTGCTGGAAGAATATGTTAAGAATTTGCCAAGGACTGCTGAGACTTACAAACCAAAGGA
GATTGAGATCAAATGTTTCAAGGGTGTTGACTTCAGTATCAGCAGTTTGCTTTCTTCAG
GGACCAAAATCTTAGATGCTATTCTTTACAGTACTTACAAGGATTCAGCAGAGCACAAC
TTCATATTTGATGTGAAAGTTCTATCTCCTGACTTCATCGATAGCAAGTTACTCGTGAA
CAACATCGAAACAGGCAATCGAGCAATCAAAGCAGCTTTCTGTCTTGTTTACAATCAAG
GTGGATTGCCATCAAAGACGAGTGAAGAACGACCACTGTCTAAGTTTGTAAGAGAAACG
ATATTCCGTGAGAAAGATCTCAAAGCCAACGAGTTATGTGAATACCTATCATCAGCAGA
TCCTTCTTTGTTTCCAAGTCAAGTCTTTTTGAAAATCTCACTTGAAAACCTTCCAACTG
AGGTTTCATCACGTTGCAAGATGTCGATTGCGGGCAACAAAGCAATGAGATATGCACTC
TTAGCTCAAAAGTTTGACAAAGATGAAATTCCAGTTCCAACAGAAGTGAATCCTACAAC
TAGCTCAGAATACATGCAGAAAAAGGAGAAAATAGAAAAAGCAAAAAAGATAGTTGATG
TTCTATGTTCTCTTGCTTCTGACTTCCAGGCACAAGTGAAAATGCATCCTCTCTCCCCT
GAGAGATCATCGAGGAAGAATTTCACTCTGCAATTGACTTCTGCAATTGTTACTTCACT
TTCCTACAAAGGGAGGTTAGACATGAGAAAAGCAATCGAAGAGAAAAAGATAGAGGCTT
TCAAAAGAGATGAAAATATATTCGGAAGGTTAAATGCTCTTGGACAACCCACGTTTCCT
GTTCTGACTAACGCAGATGCTGACTTTTCTGAATTGTCAGTTGAGGCCGTGAAGACAGC
TTACGGAAAGAAA
AUSG1 GU139121
ATGTCAGGAGTATACAAGGTTTCCGAAATTCAGTCTATCTTGCAAAAAGATGTAACTTC
CGAAGGAGAAACAGCTATTCTAATTTCTCTTGGTCTCATGACAAAAGAAGAGAAGCCTG
TTCCTGCAAAAATGGCCATGGTGGCATCTGCAAAAGCAAACTCAATCATCTTTGTTTCG
GAAGATGGCTCTTTGTCTTTTGAAGCTCCAAAAGAAACAGGAGAGACCAGCAAACCAGG
AGAGAAGAAAGAGGAAAAGAAGGTAGAAGTGGGAGTCAAGTTTCCTTTCAGCGCAGCCA
AAGTAAAGGAGCTAATTGAAGGGAAAAGTCTTACTTTGGATCAGGACAAAATCCAAAAA
GTGCTAGAAGAATATGTTAAGAATTTGCCAAGGACTGCTGAGACTTACAAACCAAAGGA
GATTGAGATCAAATGTTTCAAGGGTGTTGACTTCAGTATCAGCAGTTTGCTTTCTTCAG
GGACCAAAATCTTAGATGCTATTCTTTACAGTACTTACAAGGATTCAGCAGAGCACAAC
TTCATATTTGATGTGAAAGTTCTATCTCCTGATTTCATCGATAGCAAGTTACTCGTGAA
CAACATCGAAACAGGCAATCGAGCAATCAAAGCAGCTTTCTGTCTTGTTTACAATCAAG
GTGGATTGCCATCAAAGACGAGTGAAGAACGACCACTGTCTAAGTTTGTAAGAGAAACG
ATATTCCGTGAGAAAGATCTCAAAGCTAACGAGTTATGTGAATACCTATCATCAGCAGA
TCCTTCTTTGTTTCCAAGTCAAGTCTTTTTGAAAATCTCACTTGAAAACCTTCCTACTG
AGGTTTCATCACGTTGCAAGATGTCGATTGCGGGCAACAAAGCAATGAGATATGCACTC

163
TTAGCTCAAAAGTTTGACAAAGATGAAATTCCAGTTCCAACAGAAGTGAATCCTACAAC
TAGCTCAGAATACATGCAGAAAAAGGAGAAAATAGAAAAAGCAAAAAAGATAGTTGATG
TTCTATGTTCTCTTGCTTCTGACTTCCAGGCACAAGTGAAAATGCATCCTCTCTCCCCT
GAGAGATCATCAAGGAAGAATTTCACTCTGCAATTGACTTCTGCAATTGTTACTTCACT
TTCCTACAAAGGGAGGTTAGACATGAGAAAAGCAATCGAAGAGAAAAAGATAGAGGCTT
TCAAAAGAGATGAAAATATATTTGGAAGATTAAATGCTCTTGGACAACCCACGTTTCCT
GTTCTGACTAACGCAGATGCTGACTTTTCTGAATTGTCAGTTGAGGCCGTGAAGACAGC
TTACGGAAAGAAA
AUSE1 GU139122
ATGTCAGGAGTATACAAGGTTTCCGAAATTCAGTCTATCTTGCAAAAAGATGTAACTTC
CGAAGGAGAAACAGCTATTCTAATTTCTCTTGGTCTCATGACAAAAGAAGAGAAGCCTG
TTCCTGCAAAAATGGCCATGGTGGCATCTGCAAAAGCAAACTCAATCATCTTTGTTTCG
GAAGATGGCTCTTTGTCTTTTGAAGCTCCAAAAGAAACAGGAGAGACCAGCAAACCAGG
AGAGAAGAAAGAGGAAAAGAAGGTAGAAGTGGGAGTCAAGTTTCCTTTCAGCGCAGCCA
AAGTAAAGGAGCTAATTGAAGGGAAAAGTCTTACTTTGGATCAGGACAAAATCCAAAAA
GTGCTGGAAGAATATGTTAAGAATTTGCCAAGGACTGCTGAGACTTACAAACCAAAGGA
GATTGAGATCAAATGTTTCAAGGGTGTTGACTTCAGTATCAGCAGTTTGCTTTCTTCAG
GGACCAAAATCTTAGATGCTATTCTTTACAGTACTTACAAGGATTCAGCAGAGCACAAC
TTCATATTTGATGTGAAAGTTCTATCTCCTGATTTCATCGATAGCAAGTTACTCGTGAA
CAACATCGAAACAGGCAATCGAGCAATCAAAGCAGCTTTCTGTCTTGTTTACAATCAAG
GTGGATTGCCATCAAAGACGAGTGAAGAACGACCACTGTCTAAGTTTGTAAGAGAAACG
ATATTCCGTGAGAAAGATCTCAAAGCTAACGAGTTATGTGAATACCTATCATCAGCAGA
TCCTTCTTTGTTTCCAAGTCAAGTCTTTTTGAAAATCTCACTTGAAAACCTTCCTACTG
AGGTTTCATCACGTTGCAAGATGTCAATTGCGGGCAACAAAGCAATGAGATATGCACTC
TTAGCTCAAAAGTTTGACAAAGATGAAATTCCAGTTCCAACAGAAGTGAATCCTACAAC
TAGCTCAGAATACATGCAGAAAAAGGAGAAAATAGAAAAAGCAAAAAAGATAGTTGATG
TTCTATGTTCTCTTGCTTCTGACTTCCAGGCACAAGTGAAAATGCATCCTCTCTCCCCT
GAGAGATCATCGAGGAAGAATTTCACTCTGCAATTGACTTCTGCAATTGTTACTTCACT
TTCCTACAAAGGGAGGTTAGACATGAGAAAAGCAATCGAAGAAAAAAGGATAGAGGCTT
TCAAAAGAGATGAAAATATATTTGGAAGGTTAAATGCTCTTGGACAACCCACGTTTCCT
GTTCTGACTAACGCAGATGCTGACTTTTCTGAATTGTCAGTTGAGGCCGTGAAGACAGC
TTACGGAAAGAAA

164
AUSD1 GU139123
ATGTCAGGAGTATACAAGGTTTCCGAAATTCAGTCTATCTTGCAAAAAGATGTAACTTC
CGAAGGAGAAACAGCTATTCTAATTTCTCTTGGTCTCATGACAAAAGAAGAGAAGCCTG
TTCCTGCAAAAATGGNCATGGTGGCATCTGCAAAAGCAAACTCAATCATCTTTGTTTCN
GAAGATGGCTCTTTGTCTTTTGAAGCTCCAAAAGAAACAGGAGAGACCAGCAAACCAGG
AGAGAAGAAAGAGGAAAAGAAGGTAGAAGTGGGAGTCAAGTTTCCTTTCAGCGCAGCCA
AAGTAAAGGAGCTAATTGAAGGGAAAAGTCTTACTTTGGATCAGGACAAAATCCAAAAA
GTGCTGGAAGAATATGTTAAGAATTTGCCAAGGACTGCTGAGACTTACAAACCAAAGGA
GATTGAGATCAAATGTTTCAAGGGTGTTGACTTCAGTATCAGCAGTTTGCTTTCTTCAG
GGACCAAAATCTTAGATGCTATTCTTTACAGTACTTACAAGGATTCAGCAGAGCACAAC
TTCATATTTGATGTGAAAGTTCTATCTCCTGATTTCATCGATAGCAAGTTACTCGTGAA
CAACATCGAAACAGGCAATCGAGCAATCAAAGCAGCTTTCTGTCTTGTTTACAATCAAG
GTGGATTGCCATCAAAGACGAGTGAAGAACGACCACTGTCTAAGTTTGTAAGAGAAACG
ATATTCCGTGAGAAAGATCTCAAAGCTAACGAGTTATGTGAATACCTATCATCAGCAGA
TCCTTCTTTGTTTCCAAGTCAAGTCTTTTTGAAAATCTCACTTGAAAACCTTCCTACTG
AGGTTTCATCACGTTGCAAGATGTCAATTGCGGGCAACAAAGCAATGAGATATGCACTC
TTAGCTCAAAAGTTTGACAAAGATGAAATTCCAGTTCCAACAGAAGTGAATCCTACAAC
TAGCTCAGAATACATGCAGAAAAAGGAGAAAATAGAAAAAGCAAAAAAGATAGTTGATG
TTCTATGTTCTCTTGCTTCTGACTTCCAGGCACAAGTGAAAATGCATCCTCTCTCCCCT
GAGAGATCATCGAGGAAGAATTTCACTCTGCAATTGACTTCTGCAATTGTTACTTCACT
TTCCTACAAAGGGAGGTTAGACATGAGAAAAGCAATCGAAGAGAAAAAGATAGAGGCTT
TCAAAAGAGATGAAAATATATTTGGAAGGTTAAATGCTCTTGGACAACCCACGTTTCCT
GTTCTGACTAACGCAGATGCTGACTTTTCTGAATTGTCAGTTGAGGCCGTGAAGACAGC
TTACGGAAAGAAA
AUSA2 GU139124
ATGTCAGGAGTATACAAGGTTTCCGAAATTCAGTCTATCTTGCAAAAAGATGTAACTTC
CGAAGGAGAAACAGCTATTCTAATTTCTCTTGGTCTCATGACAAAAGAAGAGAAGCCTG
TTCCTGCAAAAATGGCCATGGTGGCATCTGCAAAAGCAAACTCAATCATCTTTGTTTCG
GAAGATGGCTCTTTGTCTTTTGAAGCTCCAAAAGAAACAGGAGAGACCAGCAAACCAGG
AGAGAAGAAAGAGGAAAAGAAGGTAGAAGTGGGAGTCAAGTTTCCTTTCAGCGCAGCCA
AAGTAAAGGAGCTAATTGAAGGGAAAAGTCTTACTTTGGATCAGGACAAAATCCAAAAA
GTGCTGGAAGAATATGTTAAGAATTTGCCAAGGACTGCTGAGACTTACAAACCAAAGGA
GATTGAGATCAAATGTTTCAAGGGTGTTGACTTCAGTATCAGCAGTTTGCTTTCTTCAG
GGACCAAAATCTTAGATGCTATTCTTTACAGTACTTACAAGGATTCAGCAGAGCACAAC
TTCATATTTGATGTGAAAGTTCTATCTCCTGATTTCATCGATAGCAAGTTACTCGTGAA
CAACATCGAAACAGGCAATCGAGCAATCAAAGCAGCTTTCTGTCTTGTTTACAATCAAG
GTGGATTGCCATCAAAGACGAGTGAAGAACGACCACTGTCTAAGTTTGTAAGAGAAACG
ATATTCCGTGAGAAAGATCTCAAAGCTAACGAGTTATGTGAATACTTATCATCAGCAGA
TCCTTCTTTGTTTCCAAGTCAAGTCTTTTTGAAAATCTCACTTGAAAACCTTCCTACTG
AGGTTTCATCACGTTGCAAGATGTCAATTGCGGGCAACAAAGCAATGAGATATGCACTC

165
TTAGCTCAAAAGTTTGACAAAGATGAAATTCCAGTTCCAACAGAAGTGAATCCTACAAC
TAGCTCAGAATACATGCAGAAAAAGGAGAAAATAGAAAAAGGAAAAAAGATAGTTGATG
TTCTATGTTCTCTTGCTTCTGACTTCCAGGCACAAGTGAAAATGCATCCTCTCTCCCCT
GAGAGATCATCGAGGAAGAATTTCACTCTGCAATTGACTTCTGCAATTGTTACTTCACT
TTCCTACAAAGGGAGGTTAGACATGAGAAAAGCAATCGAAGAGAAAAAGATAGAGGCTT
TCAAAAGAGATGAAAATATATTTGGAAGGTTAAATGCTCTTGGACAACCCACGTTTCCT
GTTTTGACTAACGCAGATGCTGACTTTTCTGAATTGTCAGTTGAGGCCGTGAAGACAGC
TTACGGAAAGAAA
AUSC1 GU139125
ATGTCAGGAGTATACAAGGTTTCCGAAATTCAGTCTATCTTGCAAAAAGATGTAACTTC
CGAAGGAGAAACAGCTATTCTAATTTCTCTTGGTCTCATGACAAAAGAGGAGAAGCCTG
TTCCTGCAAAAATGGCCATGGTGGCATCTGCAAAAGCAAACTCAATCATCTTTGTTTCG
GAAGATGGCTCTTTGTCTTTTGAAGCTCCAAAAGAAACAGGAGAGACCAGCAAACCAGG
AGAGAAGAAAGAGGAAAAGAAGGTAGAAGTGGGAGTCAAGTTTCCTTTCAGCGCAGCCA
AAGTAAAGGAGCTAATTGAAGGGAAAAGTCTTACTTTGGATCAGGACAAAATCCAAAAA
GTGCTGGAAGAATATGTTAAGAATTTGCCAAGGACTGCTGAGACTTACAAACCAAAGGA
GATTGAGATCAAATGTTTCAAGGGTGTTGACTTCAGTATCAGCAGTTTGCTTTCTTCAG
GGACCAAAATCTTAGATGCTATTCTTTACAGTACTTACAAGGATTCAGCAGAGCACAAC
TTCATATTTGATGTGAAAGTTCTATCTCCTGATTTCATCGATAGCAAGTTACTCGTGAA
CAACATCGAAACAGGCAATCGAGCAATCAAAGCAGCTTTCTGTCTTGTTTACAATCAAG
GTGGATTGCCATCAAAGACGAGTGAAGAGCGACCACTGTCTAAGTTTGTAAGAGAAACG
ATATTCCGTGAGAAAGATCTCAAAGCTAACGAGTTATGTGAATACCTATCATCAGCAGA
TCCTTCTTTGTTTCCAAGTCAAGTCTTTTTGAAAATCTCACTTGAAAACCTTCCTACTG
AGGTTTCATCACGTTGCAAGATGTCGATTGCGGGCAACAAAGCAATGAGATATGCACTC
TTAGCTCAAAAGTTTGACAAAGATGAAATTCCAGTTCCAACAGAAGTGAATCCTACAAC
TAGCTCAGAATACATGCAGAAAAAGGAGAAAATAGAAAAAGCAAAAAAGATAGTTGATG
TTCTATGTTCTCTTGCTTCTGACTTCCAGGCACAAGTGAAAATGCATCCTCTCTCTCCT
GAGAGATCATCGAGGAAGAATTTCACCCTGCAATTGACTTCTGCAATTGTTACTTCACT
TTCCTACAAAGGGAGGTTAGACATGAGAAAAGCAATCGAAGAGAAAAAGATAGAGGCTT
TCAAAAGAGATGAAAATATATTTGGAAGGTTAAATGCTCTTGGACAACCCACGTTTCCT
GTTCTGACTAACGCAGATGCTGACTTTTCTGAATTGTCAGTTGAGGCCGTGAAGACAGC
TTACGGAAAGAAA

166
AUSE2 GU139126
ATGTCAGGAGTATACAAAGTTTCCGAAATTCAGTCTATCTTGCAAAAGGATGTAACTTC
CGAAGGAGAAACAGCTATTCTAATTTCTCTTGGTCTCATGACAAAAGAAGAGAAGCCTG
TTCCTGCAAAAATGGCCATGGTGGCATCTGCAAAAGCTAACTCAATCATCTTTGTTTCA
GAAGATGGCTCTCTGTCTTTTGAAGCTCCAAAGGAAACAGGAGAGACCAGCAAACCAGG
AGAAAAGAAAGAGGAAAAGAAGGTAGAAGTGGGGGTCAAGTTTCCTTTCAGTGCAGCCA
AAGTAAAGGAGCTAATCGAAGGGAAAAGTCTTACTTTGGATCAGGACAAGATCCAAAAA
GTGCTGGAAGAATATGTTAAAAATCTGCCAAGGACTGCTGAGACTTACAAACCAAAAGA
GATTGAAATCAAATGTTTCAAGGGTGTTGACTTCAGTATCAGCAGTTTGCTTTCTTCAG
GGACCAAAATCCTAGATGCTATTCTCTACAGTACTTACAAAGATTCAGCGGAGCACAAC
TTCATATTTGATGTAAAAGTTCTATCCCCTGATTTCATCGACAGCAAGTTACTCGTGAA
CAACATAGAAACAGGCAATCGAGCAATCAAAGCAGCTTTCTGTCTCGTTTACAATCAAG
GTGGATTGCCATCAAAGACGAGTGAAGAACGACCACTATCCAAGTTTGTAAGAGAAACG
ATATTCCGTGAGAAAGATCTCAAAGCTAACGAGTTATGTGAATACTTGTCATCAGCGGA
TCCTTCTTTGTTTCCAAGTCAAGTCTTTTTGAAAATCTCACTTGAGAACCTTCCTACTG
AGGTTTCATCACGCTGCAAGATGTCGATTGCAGGCAACAAAGCAATGAGATATGCACTT
TTAGCTCAAAAGTTTGACAAAGATGAAGTTCCAGTTCCAACAGAAGTGAATCCTATAAC
TAGCTCAGAATACATGCAGAAAAAGGAGAAAATAGAGAAAGCAAAAAAGATAGTTGATG
TTCTATGTTCTCTTGCTTCTGACTTCCAGGCACAAGTGAAAATGCATCCTCTCTCCCCT
GAGAGATCATCGAGGAAGAATTTCACTCTGCAATTGACTTCTGCAATTGTTACTTCACT
TTCTTACAAAGGGAGGTTAGACATGAGAAAAGCAATCGAAGAGAAAAAGATAGAGGCCT
TCAAAAGAGATGAAAATATATTCGGAAGGTTAAATGCTCTTGGACAGCCTACGTTTCCT
GTTCTGACTAACGCAGATGCTGACTTTTCCGAATTGTCAGTTGAGGCCGTGAAGACAGC
TTACGGAAAGAAA

167
Appendix 5 - LBVaV CP sequences (5' to 3')
AUSE4 GU220721
ATGGCACACCCCAAATTGAAGATGCTAGAGGCATTCAGTGACGTCGTGGAAATCACAGG
TAAGACTGCCGGGAAAGAATCCTGGGATGATGAAAGCACAATAGCTATGCCCTCCTACA
AACTTTCCGTACTGTCCGACGCCGATGCTGTTCGTGAGGTAAAGATCTTCCTGACAGGG
CTCTTTGTGAGGTCCTCTCCGAGGGCGATTGCTGCAGCTCTCATCATGACATGGAACAT
GAGGTCTGTTGATCCCGTGGCTGTCAGAATATTCCCCGCAAAGGACAAGGGGAAAGACA
CAGCGGATGTGGATGTCAAGAATCTGGAGGTGGACGGAGTCGACTACATAGACGCAATG
GTAGAGACCAATGTGAAGGATGCTTCTGATATAGAAATCATCAGAGCCGGAGCATTCAT
CGCTGCTTCTACGCTCAAGATGTTCGCCAAGTCCTTCACTGGATGGACTCAAGCTTGGG
AACACAAGCATATCCAAAAAAGGTATGCTGATTTCTGTAAGACCGAGTATCCATTCAAG
GAATTCACGACAAACACCAAGTGCGCAGAGACCATGTATGAGGCCTACCAGGGTCAAAA
ACTGTATCAAGGAACTCTCGGCCGAATCCTCTATGCTCTGGGAGATGTGGCAGATCCAA
GGCAGACTGAGATGTTGTTCGATCAACATCTTGCCAGCACTGGCATGCACATCACTAGT
GAATTCACAAATGCTCAACTCTCGATCGGTGCCACTACGGCGGGGTTATTAAGTGCTCT
CAACTACGGACAGAACTTTGGCACTCTAATGCAGCTCAAGAAGCTCATCAATGAGAGTC
TCAGCAAACCACCGGGTCCAGACAACAGAGCAACCTGGAGATTTGCCAGGATCTTTGAT
CCATCTGTCTTTCAAACCCTTCAGACAAAATACTGTGCAGATACCGTTGCCATCTTAGC
CAACATCAATTCCATGGGAAAGCTATCTACAGAAACTAGCAATCCTCTGAACATTGCTG
TGCTAAAACAGATGGCTCCGGAGAGGAAGAGGTACACAAGACAGGTGGCGAAGAACATC
TACCATCACTTCATGGTGGTTGCCAGGGCCCTGAACAACGACATGTTCGACACAGACAA
ATACAAGTTTGTGGGATCCGACGATGAGGAGGAACATGTCGCGAATGAGGGAGAGACAC
CAGTAAAGGAG
AUSE5 GU220722
ATGGCACACCCCAAATTGAAGATGCTAGAGGCATTCAGTGACGTCGTGGAAATCACAGG
TAAGACTGCCGGGAAAGAATCCTGGGATGATGAAAGCACAATAGCTATGCCCTCCTACA
AACTTTCCGTACTGTCCGACGCCGATGCTGTTCGTGAGGTAAAGATCTTCCTGACAGGG
CTCTTTGTGAGGTCCTCTCCGAGGGCGATTGCTGCAGCTCTCATCATGACATGGAACAT
GAGGTCTGTTGATCCCGTGGCTGTCAGAATATTCCCCGCAAAGGACAAGGGGAAAGACA
CAGCGGATGTGGATGTCAAGAATCTGGAGGTGGACGGAGTCGACTACATAGACGCAATG
GTAGAGACCAATGTGAAGGATGCTTCTGATATAGAAATCATCAGAGCCGGAGCATTCAT
CGCTGCTTCTACGCTCAAGATGTTCGCCAAGTCCTTCACTGGATGGACTCAAGCTTGGG
AACACAAGCATATCCAAAAAAGGTATGCTGATTTCTGTAAGACCGAGTATCCATTCAAG
GAATTCACGACAAACACCAAGTGCGCAGAGACCATGTATGAGGCCTACCAGGGTCAAAA
ACTGTATCAAGGAACTCTCGGCCGAATCCTCTATGCTCTGGGAGATGTGGCAGATCCAA
GGCAGACTGAGATGTTGTTCGATCAACATCTTGCCAGCACTGGCATGCACATCACTAGT
GAATTCACAAATGCTCAACTCTCGATCGGTGCCACTACGGCGGGGTTATTAAGTGCTCT

168
CAACTACGGACAGAACTTTGGCACTCTAATGCAGCTCAAGAAGCTCATCAATGAGAGTC
TCAGCAAACCACCGGGTCCAGACAACAGAGCAACCTGGAGATTTGCCAGGATCTTTGAT
CCATCTGTCTTTCAAACCCTTCAGACAAAATACTGTGCAGATACCGTTGCCATCTTAGC
CAACATCAATTCCATGGGAAAGCTATCTACAGAAACTAGCAATCCTCTGAACATTGCTG
TGCTAAAACAGATGGCTCCGGAGAGGAAGAGGTACACAAGACAGGTGGCGAAGAACATC
TACCATCACTTCATGGTGGTTGCCAGGGCCCTGAACAACGACATGTTCGACACAGACAA
ATACAAGTTTGTGGAATCCGACGATGAGGAGGAACATGTCGCGAATGAGGGAGAGACAC
CAGTGAAGGAG
AUSA5 GU220723
ATGGCACACCCCAAATTGAAGATGCTAGAGGCATTCAGTGACGTCGTGGAAATCACAGG
TAAGACTGCCGGGAAAGAATCCTGGGATGATGAAAGCACAATAGCTATGCCCTCCTACA
AACTTTCCGTACTGTCCGACGCCGATGCTGTTCGTGAGGTAAAGATCTTCCTAACAGGG
CTCTTTGTGAGGTCCTCTCCGAGGGCGATTGCTGCAGCTCTCATCATGACATGGAACAT
GAGGTCTGTTGATCCCGTGGCTGTCAGAATATTCCCCGCAAAGGACAAGGGGAAAGACA
CAGCGGATGTGGATGTCAAGAATCTGGAGGTGGACGGAGTCGACTACATAGACGCAATG
GTAGAGACCAATGTGAAGGATGCTTCTGATATAGAAATCATCAGAGCCGGAGCATTCAT
CGCTGCTTCTACGCTCAAGATGTTCGCCAAGTCCTTCACTGGATGGACTCAAGCTTGGG
AACACAAGCATATCCAAAAAAGGTATGCTGATTTCTGTAAGACCGAGTATCCATTCAAG
GAATTCACGACAAACACCAAGTGCGCAGAGACCATGTATGAGGCCTACCAGGGTCAAAA
ACTGTATCAAGGAACTCTCGGCCGAATCCTCTATGCTCTGGGAGATGTGGCAGATCCAA
GGCAGACTGAGATGTTGTTCGATCAACATCTTGCCAGCACTGGCATGCACATCATCCCC
CAGTTCACAAATGCTCAACTCTCGATCGGTGCCACTACGGCGGGGTTATTAAGTGCTCT
CAACTACGGACAGAACTTTGGCACTCTAATGCAGCTCAAGAAGCTCATCAATGAGAGTC
TCAGCAAACCACCGGGTCCAGACAACAGAGCAACCTGGAGATTTGCCAGGATCTTTGAT
CCATCTGTCTTTCAAACCCTTCAGACAAAATACTGTGCAGATACCGTTGCCATCTTAGC
CAACATCAATTCCATGGGAAAGCTATCTACAGAAACTAGCAATCCTCTGAACATTGCTG
TGCTAAAACAGATGGCTCCGGAGAGGAAGAGGTACACAAGACAGGTGGCGAAGAACATC
TACCATCACTTCATGGTGGTTGCCAGGGCCCTGAACAACGACATGTTCGACACAGACAA
ATACAAGTTTGTGGAATCCGACGATGAGGAGGAACATGTCGCGAATGAGGGAGAGACAC
CAGTGAAGGAG
AUSB1 GU220724
ATGGCACACCCCAAATTGAAGATGCTAGAGGCATTCAGTGACGTCGTGGAAATCACAGG
TAAGACTGCCGGGAAAGAATCCTGGGATGATGAAAGCACAATAGCTATGCCCTCCTACA
AACTTTCCGTACTGTCCGACGCCGATGCTGTTCGTGAGGTAAAGATCTTCCTGACAGGG
CTCTTTGTGAGGTCCTCTCCGAGGGCGATTGCTGCAGCTCTCATCATGACATGGAACAT
GAGGTCTGTTGATCCCGTGGCTGTCAGAATATTCCCCGCAAAGGACAAGGGGAAAGACG
CAGCTGATGTGGATGTCAAGAATCTGGAGGTGGTCGGAGTCGACTACATAGACGCAATG
GTAGAGACCAATGTGAAGGATGCTTCTGATATAGAAATCATCAGAGCCGGAGCATTCAT

169
CGCTGCTTCTACGCTCAAGATGTTCGCCAAGTCCTTCACTGGATGGACTCAAGCTTGGG
AACACAAGCATATCCAAAAAAGGTATGCTGATTTCTGTAAGACCGAGTATCCATTCAAG
GAATTCACGACAAACACCAAGTGCGCAGAGACCATGTATGAGGCCTACCAGGGTCAAAA
ACTGTATCAAGGAACTCTCGGCCGAATCCTCTATGCTCTGGGAGATGTGGCAGATCCAA
GGCAGACTGAGATGTTGTTCGATCAACACCTTGCCAACACTGGAATGCACATCATCCCC
CAGTTCACAAATGCTCAACTCTCGATCGGTGCCACTACGGCGGGGTTATTAAGTGCTCT
CAACTACGGACAGAACTTTGGCACTCTAATGCAGCTCAAGAAGCTCATCAATGAGAGTC
TCAGCAAACCACCGGGTCCAGACAACAGAGCAACCTGGAGATTTGCCAGGATCTTTGAT
CCATCTGTCTTTCAAACCCTTCAGACAAAATACTGTGCAGATACCGTTGCCATCTTAGC
CAACATCAATTCCATGGGAAAGCTATCTACAGAAACTAGCAATCCTCTGAACATTGCTG
TGCTAAAACAGATGGCTCCGGAGAGGAAGAGGTACACAAGACAGGTGGCGAAGAACATC
TACCATCACTTTATGGTGGTTGCCAGGGCCCTGAACAACGACATGTTCGACACAGACAA
ATACAAGTTTGTGGAATCCGACGATGAGGAGGAACATGTCGCGAATGAGGGAGAGACAC
CAGAGAAGGAG
AUSB2 GU220725
ATGGCACACCCCAAATTGAAGATGCTAGAGGCATTCAGTGACGTCGTGGAAATCACAGG
TAAGACTGCCGGGAAAGAATCCTGGGATGATGAAAGCACAATAGCTATGCCCTCCTACA
AACTTTCCGTACTGTCCGACGCCGATGCTGTTCGTGAGGTAAAGATCTTCCTGACAGGG
CTCTTTGTGAGGTCCTCTCCGAGGGCGATTGCTGCAGCTCTCATCATGACATGGAACAT
GAGGTCTGTTGATCCCGTGGCTGTCAGAATATTCCCCGCAAAGGACAAGGGGAAAGACA
CAGCGGATGTGGATGTCAAGAATCTGGAGGTGGACGGAGTCGACTACATAGACGCAATG
GTAGAGACCAATGTGAAGGATGCTTCTGATATAGAAATCATCAGAGCCGGAGCATTCAT
CGCTGCTTCTACGCTCAAGATGTTCGCCAAGTCCTTCACTGGATGGACTCAAGCTTGGG
AACACAAGCATATCCAAAAAAGGTATGCTGATTTCTGTAAGACCGAGTATCCATTCAAG
GAATTCACGACAAACACCAAGTGCGCAGAGACCATGTACGAGGCCTATCAGGGTCAAAA
ACTGTATCAAGGAACTCTCGGCCGAATCCTCTATGCTCTGGGAGATGTGGCAGATCCAA
GGCAGACTGAGATGTTGTTCGATCAACATCTTGCCAACACTGGCATGCACATCATCCCC
CAGTTCACAAATGCTCAACTCTCGATCGGTGCCACTACGGCGGGGTTATTAAGTGCTCT
CAACTACGGACAGAACTTTGGCACTCTAATGCAGCTCAAAAAGCTCATCAATGAGAGTC
TCAGCAAACCACCGGGTCCAGACAACAGAGCAACCTGGAGATTTGCCAGGATCTTTGAT
CCATCTGTCTTTCAAACCCTTCAGACAAAATACTGTGCAGATACCGTTGCCATCTTAGC
CAACATCAATTCCATGGGAAAGCTATCTACAGAGACTAGTAATCCTCTGAACATTGCTG
TGCTAAAGCAGATGGCCCCGGAGAGGAAGAGGTACACAAGACAGGTGGCGAAGAACATC
TACCATCACTTCATGGTGGTTGCCAGGGCCCTGAACAACGACATGTTCGACACAGACAA
ATACAAGTTTGTGGAATCCGACGATGAGGAGGAACATGTCGCGAATGAGGGAGAGACAC
CAGTGAAGGAG

170
AUSG1 GU220726
ATGGCACACCCCAAATTGAAGATGCCAGAGGCATTCAGTGACGTCGTGGAAATCACAGG
TAAGACTGCCGGGAAAGAATCCTGGGATGATGAAAGCACAATAGCTATGCCCTCCTACA
AACTTTCCGTACTGTCCGACGCCGATGCTGTTCGTGAGGTAAAGATCTTCCTGACAGGG
CTCTTTGTGAGGTCCTCTCCGAGGGCGATTGCTGCAGCTCTCATCATGACATGGAACAT
GAGGTCTGTTGATCCCGTGGCTGTCAGAATATTCCCCGCAAAGGACAAGGGGAAAGACG
CAGCTGATGTGGATGTCAAGAATCTGGAGGTGGTCGGAGTCGACTACATAGACGCAATG
GTAGAGACCAATGTGAAGGATGCTTCTGATATAGAAATCATCAGAGCCGGAGCATTCAT
CGCTGCTTCTACGCTCAAGATGTTCGCCAAGTCCTTCACTGGATGGACTCAAGCTTGGG
AACACAAGCATATCCAAAAAAGGTATGCTGATTTCTGTAAGACCGAGTATCCATTCAAG
GAATTCACGACAAACACCAAGTGCGCAGAGACCATGTACGAGGCCTATCAGGGTCAAAA
ACTGTATCAAGGAACTCTCGGCCGAATCCTCTATGCTCTGGGAGATGTGGCAGATCCAA
GGCAGACTGAGATGTTGTTCGATCAACATCTTGCCAGCACTGGCATGCACATCATCCCC
CAGTTCACAAATGCTCAACCCTCGATCGGTGCCACTACGGCGGGGTTATTAAGTGCTCT
CAACTACGGACAGAACTTTGGCACTCTAATGCAGCTCAAAAAGCTCATCAATGAGAGTC
TCAGCAAACCACCGGGTCCAGACAACAGAGCAACCTGGAGATTTGCCAGGATCTTTGAT
CCATCTGTCTTTCAAACCCTTCAGACAAAATACTGTGCAGATACCGTTGCCATCTTAGC
CAACATCAATTCCATGGGAAAGCTATCTACAGAGACTAGTAATCCTCTGAACATTGCTG
TGCTAAAGCAGATGGCCCCGGAGAGGAAGAGGTACACAAGACAGGTGGCGAAGAACATC
TATCATCACTTCATGGTGGTTGCCAGGGCCCTGAACAACGACATGTTCGACACAGACAA
ATACAAGTTTGTGGAATCCGACGATGAGGAGGAACATGTCGCGAATGAGGGAGAGACAC
CAGTGAAGGAG

171
AUSE6 GU220727
ATGGCACACCCCAAATTGAAGATGCTAGAGGCATTCAGTGACGTCGTGGAAATCACAGG
TAAGACTGCCGGGAAAGAATCCTGGGATGATGAAAGCACAATAGCTATGCCCTCCTACA
AACTTTCCGTACTGTCCGACGCCGATGCTGTTCGTGAGGTAAAGATCTTCCTGACAGGG
CTCTTTGTGAGGTCCTCTCCGAGGGCGATTGCTGCAGCTCTCATCATGACATGGAACAT
GAGGTCTGTTGATCCCGTGGCTGTCAGAATATTCCCCGCAAAGGACAAGGGAAAAGACG
CAGCTGATGTGGATGTCAAGAATCTGGAGGTGGTCGGAGTCGACTACATAGACGCAATG
GTAGAGACCAATGTGAAGGATGCTTCTGATATAGAAATCATCAGAGCCGGAGCATTCAT
CGCTGCTTCTACGCTCAAGATGTTCGCCAAGTCCTTCACTGGATGGACTCAAGCTTGGG
AACACAAGCATATCCAAAAAAGGTATGCTGATTTCTGTAAGACCGAGTATCCATTCAAG
GAATTCACGACAAACACCAAGTGCGCAGAGACCATGTACGAGGCCTATCAGGGTCAAAA
ACTGTATCAAGGAACTCTCGGCCGAATCCTCTATGCTCTGGGAGATGTGGCAGATCCAA
GGCAGACTGAGATGTTGTTCGATCAACATCTTGCCAGCACTGGCATGCACATCATCCCC
CAGTTCACAAATGCTCAACTCTCGATCGGTGCCACTACGGCGGGGTTATTAAGTGCTCT
CAACTACGGACAGAACTTTGGCACTCTAATGCAGCTCAAAAAGCTCATCAATGAGAGTC
TCAGCAAACCGCCGGGTCCAGACAACAGAGCAACCTGGAGATTTGCCAGGATCTTTGAT
CCATCTGTCTTTCAAACCCTTCAGACAAAATACTGTGCAGATACCGTTGCCATCTTAGC
CAACATCAATTCCATGGGAAAGCTATCTACAGAGACTAGTAATCCTCTGAACATTGCTG
TGCTAAAGCAGATGGCCCCGGAGAGGAAGAGGTACACAAGACAGGTGGCGAAGAACATC
TATCATCACTTCATGGTGGTTGCCAGGGCCCTGAACAACGACATGTTCGACACAGACAA
ATACAAGTTTGTGGAATCCGACGATGAGGAGGAACATGTCGCGAATGAGGGAGAGACAC
CAGTGAAGGAG
172
Appendix 6 – Locations where MLBVV isolates were collected
Western
AustraliaSouth
Australia
Queensland
Northern
Territory
New South Wales
Victoria
Tasmania
Australian
Capital
Territory
A
B
C
D
E
F G
Perth
Wanneroo
Indian
Ocean
20km
Inset 1
N
Indian Ocean
Antarctic Ocean
Pacific Ocean
Map of the Australian continent showing the location of the northern Perth region of
south-western Australia where MLBVV isolates were collected.
Inset 1 shows the locations of the farms sampled (red spots) and letters indicate
individual site codes.

173
Appendix 7 – Locations where LBVaV isolates were collected
Western
AustraliaSouth
Australia
Queensland
Northern
Territory
New South Wales
Victoria
Tasmania
Australian
Capital
Territory
N
Indian Ocean
Antarctic Ocean
Pacific Ocean
A
BE
G
Perth
Wanneroo
Indian
Ocean
20km
Inset 1
Map of the Australian continent showing the location of the northern Perth region of
south-western Australia where LBVaV isolates were collected.
Inset 1 shows the locations of the farms sampled (red spots) and letters indicate
individual site codes.

174
Appendix 8 – MLBVV amino acid and nucleotide distances
Percentage nucleotide (upper diagonal) and amino acid (lower diagonal) sequence identities of the coat protein (CP) encoding sequences of Mirafiori lettuce big-vein
virus. Numbers indicate MLBVV isolates; 1 (AUSG2), 2 (AUSE3), 3 (AUSE6), 4 (AUSF1), 5 (AUSB2), 6 (AUSA1), 7 (AUSE4), 8 (AUSG1), 9 (AUSE1), 10
(AUSD1), 11 (AUSA2), 12 (AUS1), 13 (AUSC1), 14 (GER1), 15 (LS301-0), 16 (HOL2), 17 (UK1), 18 (DEN1), 19 (JPN1), 20 (GER2), 21 (MUR1), 22 (ALM2),
23 (ITA1), 24 (GER3), 25 (AUSE2), 26 (ALM3), 27 (GAL1), 28 (ALM4), 29 (ALM1), 30 (SON1), 31 (SON4), 32 (SON3), 33 (ALM5) and 34 (SON2)
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34
1 **** 99.7 99.7 99.4 99.8 99.2 99.3 99.5 99.5 99.7 99.5 99.5 99.5 99.0 99.5 98.9 98.9 98.9 99.2 98.5 96.6 97.4 96.5 96.7 96.6 96.2 96.8 87.2 87.3 86.7 86.7 87.1 87.1 87.0
2 99.8 **** 99.8 99.5 99.9 99.3 99.5 99.7 99.7 99.8 99.6 99.7 99.6 99.2 99.7 99.1 99.1 99.1 99.4 98.6 96.7 97.5 96.7 96.9 96.7 96.4 97.0 87.3 87.4 86.9 86.9 87.2 87.2 87.1
3 99.5 99.8 **** 99.5 99.9 99.3 99.5 99.7 99.7 99.8 99.6 99.7 99.6 99.2 99.7 99.1 99.1 99.1 99.4 98.6 96.7 97.5 96.7 96.9 96.7 96.4 97.0 87.3 87.4 86.9 86.9 87.2 87.2 87.1
4 99.1 99.3 99.1 **** 99.6 99.0 99.2 99.4 99.4 99.5 99.3 99.4 99.3 98.8 99.4 98.8 98.8 98.8 99.1 98.3 96.6 97.3 96.5 96.7 96.6 96.2 96.8 86.9 87.0 86.5 86.5 86.9 86.9 86.8
5 99.8 100 99.8 99.3 **** 99.4 99.5 99.8 99.8 99.9 99.7 99.8 99.7 99.2 99.8 99.2 99.2 99.2 99.5 98.7 96.8 97.6 96.8 97.0 96.8 96.5 97.0 87.4 87.5 87.0 87.0 87.3 87.3 87.2
6 99.5 99.8 99.5 99.1 99.8 **** 99.7 99.2 99.2 99.3 99.1 99.2 99.1 98.6 99.2 98.5 98.5 98.5 98.8 98.1 96.1 96.9 96.1 96.3 96.3 95.8 96.4 86.9 87.0 86.6 86.6 87.0 87.0 86.9
7 99.8 100 99.8 99.3 100 99.8 **** 99.3 99.3 99.5 99.2 99.3 99.2 98.8 99.3 98.7 98.7 98.7 99.0 98.2 96.3 97.3 96.5 96.6 96.6 96.1 96.7 87.1 87.2 86.7 86.7 87.1 87.1 87.0
8 99.8 100 99.8 99.3 100 99.8 100 **** 99.5 99.7 99.5 99.5 99.5 99.0 99.5 98.9 98.9 98.9 99.2 98.5 96.6 97.3 96.6 96.7 96.6 96.2 96.8 87.3 87.4 86.9 86.9 87.2 87.2 87.1
9 99.5 99.8 99.5 99.1 99.8 99.5 99.8 99.8 **** 99.8 99.6 99.5 99.5 99.0 99.5 98.9 98.9 98.9 99.2 98.5 96.7 97.3 96.7 96.9 96.6 96.2 96.8 87.3 87.4 86.9 86.9 87.2 87.2 87.1
10 99.8 100 99.8 99.3 100 99.8 100 100 99.8 **** 99.8 99.7 99.6 99.2 99.7 99.1 99.1 99.1 99.4 98.6 96.9 97.5 96.7 96.9 96.7 96.4 97.0 87.3 87.4 86.9 86.9 87.2 87.2 87.1
11 99.5 99.8 99.5 99.1 99.8 99.5 99.8 99.8 99.5 99.8 **** 99.5 99.4 98.9 99.5 98.8 98.8 98.8 99.2 98.4 96.6 97.3 96.5 96.6 96.6 96.1 96.7 87.2 87.3 86.8 86.8 87.1 87.1 87.0
12 99.8 100 99.8 99.3 100 99.8 100 100 99.8 100 99.8 **** 99.5 99.0 99.5 98.9 98.9 98.9 99.2 98.6 96.7 97.3 96.5 96.7 96.6 96.2 96.8 87.3 87.4 86.9 86.9 87.2 87.2 87.1
13 99.8 100 99.8 99.3 100 99.8 100 100 99.8 100 99.8 100 **** 99.0 99.5 98.8 98.8 98.8 99.2 98.4 96.5 97.3 96.7 96.8 96.5 96.1 96.7 87.2 87.3 86.8 86.8 87.1 87.1 87.0
14 98.1 98.4 98.1 97.7 98.4 98.1 98.4 98.4 98.1 98.4 98.1 98.4 98.4 **** 99.3 98.7 98.8 98.7 99.0 98.2 96.3 97.1 96.5 96.5 96.3 96.0 96.5 86.8 86.9 86.4 86.4 86.7 86.7 86.6
15 99.5 99.8 99.5 99.1 99.8 99.5 99.8 99.8 99.5 99.8 99.5 99.8 99.8 98.1 **** 99.2 99.2 99.2 99.5 98.8 96.9 97.7 96.9 97.0 96.9 96.5 97.1 87.5 87.6 87.0 87.0 87.4 87.4 87.3
16 99.1 99.3 99.1 98.6 99.3 99.1 99.3 99.3 99.1 99.3 99.1 99.3 99.3 97.7 99.1 **** 98.9 98.9 99.0 98.1 96.4 97.0 96.2 96.4 96.2 95.9 96.5 87.0 87.1 86.5 86.5 86.9 86.9 86.8
17 98.6 98.8 98.6 98.1 98.8 98.6 98.8 98.8 98.6 98.8 98.6 98.8 98.8 97.7 98.6 98.6 **** 99.1 99.0 98.1 96.4 97.1 96.6 96.4 96.2 96.0 96.5 87.0 87.1 86.5 86.5 86.9 86.9 86.8
18 98.6 98.8 98.6 98.1 98.8 98.6 98.8 98.8 98.6 98.8 98.6 98.8 98.8 97.2 98.6 98.6 98.1 **** 99.0 98.1 96.4 97.1 96.2 96.4 96.2 95.9 96.5 87.2 87.3 86.6 86.6 87.0 87.0 86.9
19 99.5 99.8 99.5 99.1 99.8 99.5 99.8 99.8 99.5 99.8 99.5 99.8 99.8 98.1 99.5 99.1 98.6 98.6 **** 98.5 96.6 97.3 96.5 96.7 96.7 96.2 96.8 87.6 87.7 87.1 87.1 87.5 87.5 87.4
20 98.8 99.1 98.8 98.4 99.1 98.8 99.1 99.1 98.8 99.1 98.8 99.1 99.1 97.4 98.8 98.4 97.9 97.9 98.8 **** 96.2 96.9 96.4 96.7 96.2 96.0 96.5 86.8 86.9 86.6 86.6 86.9 86.9 86.8
21 98.4 98.6 98.4 97.9 98.6 98.4 98.6 98.6 98.4 98.6 98.4 98.6 98.6 97.0 98.4 97.9 97.4 97.4 98.4 97.7 **** 97.3 96.7 96.9 96.6 96.4 96.9 86.4 86.5 86.1 86.1 86.3 86.3 86.2
22 98.8 99.1 98.8 98.4 99.1 98.8 99.1 99.1 98.8 99.1 98.8 99.1 99.1 97.4 98.8 98.4 97.9 97.9 98.8 98.1 97.7 **** 97.7 97.8 97.7 97.4 98.0 87.1 87.2 86.7 86.7 87.0 87.0 86.9
23 98.4 98.6 98.4 97.9 98.6 98.4 98.6 98.6 98.4 98.6 98.4 98.6 98.6 97.4 98.4 97.9 97.9 97.4 98.4 97.7 97.2 98.1 **** 99.5 97.2 97.2 97.6 87.0 87.1 86.5 86.5 87.1 87.1 87.0
24 98.4 98.6 98.4 97.9 98.6 98.4 98.6 98.6 98.4 98.6 98.4 98.6 98.6 97.0 98.4 97.9 97.4 97.4 98.4 98.1 97.2 98.1 99.1 **** 97.4 97.3 97.7 87.2 87.3 86.8 86.8 87.3 87.3 87.2
25 99.3 99.5 99.3 98.8 99.5 99.3 99.5 99.5 99.3 99.5 99.3 99.5 99.5 97.9 99.3 98.8 98.4 98.4 99.3 98.6 98.1 99.1 99.1 99.1 **** 97.2 97.7 86.9 87.0 86.7 86.7 86.9 86.9 86.8
26 96.7 97.0 96.7 96.2 97.0 96.7 97.0 97.0 96.7 97.0 96.7 97.0 97.0 95.3 96.7 96.2 96.2 95.8 96.7 96.5 95.5 96.5 97.0 97.0 97.4 **** 98.8 85.9 86.0 85.4 85.4 85.8 85.8 85.7
27 98.8 99.1 98.8 98.4 99.1 98.8 99.1 99.1 98.8 99.1 98.8 99.1 99.1 97.4 98.8 98.4 97.9 97.9 98.8 98.1 97.7 98.6 98.6 98.6 99.5 97.4 **** 86.8 86.9 86.4 86.4 86.9 86.9 86.8
28 95.5 95.8 95.5 95.0 95.8 95.5 95.8 95.8 95.5 95.8 95.5 95.8 95.8 94.1 95.5 95.0 94.8 94.8 95.5 94.8 94.3 95.3 94.8 94.8 95.8 93.1 95.3 **** 99.8 98.8 98.8 98.5 98.5 98.5
29 95.3 95.5 95.3 94.8 95.5 95.3 95.5 95.5 95.3 95.5 95.3 95.5 95.5 93.8 95.3 94.8 94.5 94.5 95.3 94.5 94.1 95.0 94.5 94.5 95.5 92.8 95.0 99.8 **** 98.7 98.7 98.5 98.5 98.4
30 96.0 96.2 96.0 95.5 96.2 96.0 96.2 96.2 96.0 96.2 96.0 96.2 96.2 94.5 96.0 95.5 95.3 95.0 96.0 95.3 94.8 95.8 95.3 95.3 96.2 93.6 95.8 99.5 99.3 **** 100 98.8 98.8 98.7
31 96.0 96.2 96.0 95.5 96.2 96.0 96.2 96.2 96.0 96.2 96.0 96.2 96.2 94.5 96.0 95.5 95.3 95.0 96.0 95.3 94.8 95.8 95.3 95.3 96.2 93.6 95.8 99.5 99.3 100 **** 98.8 98.8 98.7
32 96.0 96.2 96.0 95.5 96.2 96.0 96.2 96.2 96.0 96.2 96.0 96.2 96.2 94.5 96.0 95.5 95.3 95.0 96.0 95.3 94.8 95.8 95.3 95.3 96.2 93.6 95.8 99.5 99.3 100 100 **** 100 99.9
33 96.0 96.2 96.0 95.5 96.2 96.0 96.2 96.2 96.0 96.2 96.0 96.2 96.2 94.5 96.0 95.5 95.3 95.0 96.0 95.3 94.8 95.8 95.3 95.3 96.2 93.6 95.8 99.5 99.3 100 100 100 **** 99.9
34 95.8 96.0 95.8 95.3 96.0 95.8 96.0 96.0 95.8 96.0 95.8 96.0 96.0 94.3 95.8 95.3 95.0 94.8 95.8 95.0 94.5 95.5 95.0 95.0 96.0 93.3 95.5 99.3 99.1 99.8 99.8 99.8 99.8 ****
Clade A Clade B

175
Appendix 9 – LBVaV amino acid and nucleotide distances
Percentage nucleotide (upper diagonal) and amino acid (lower diagonal) sequence identities of the coat protein (CP) encoding sequences of LBVaV and TStV.
Numbers indicate isolates; 1 (SON5), 2 (SON4), 3 (SON3), 4 (SON6), 5 (SON2), 6 (GRA1), 7 (ALM1), 8 (ALM3), 9 (MUR1), 10 (ALM4), 11 (ALM7), 12
(ALM6), 13 (ALM2), 14 (ALM5), 15 (UK2), 16 (MUR2), 17 (SON1), 18 (GAL1), 19 (AUSE4), 20 (AUSE5), 21 (AUSA5), 22 (AUSB1), 23 (AUSB2), 24
(AUSG1), 25 (AUSE6), 26 (Wa), 27 (Hy), 28 (LBVV-CP), 29 (A), 30 (Na), 31 (Kan), 32 (No88), 33 (No89) and 34 (No2)
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34
1 **** 98.6 98.6 98.5 98.6 98.6 98.6 98.7 98.6 98.6 98.5 98.5 98.1 96.7 95.5 96.3 96.7 95.9 96.1 96.6 96.3 97.0 96.6 96.6 94.6 94.7 94.7 94.6 94.9 94.7 94.6 94.7 94.5 98.6
2 97.7 **** 98.8 98.4 98.9 98.8 98.9 99.1 98.9 99.0 98.8 98.8 98.5 97.1 95.5 96.6 97.0 96.3 96.5 97.0 96.6 97.3 96.9 97.0 94.7 94.8 95.0 94.7 95.1 95.0 94.9 95.0 94.8 99.0
3 97.4 98.5 **** 98.9 99.3 99.2 99.1 99.2 99.1 99.2 99.0 99.0 98.6 97.3 95.6 96.7 97.2 96.6 96.7 97.3 96.9 97.6 97.2 97.3 95.4 95.5 95.7 95.4 95.2 95.3 95.2 95.3 95.1 99.2
4 97.4 97.2 97.7 **** 99.4 98.9 99.1 99.1 98.9 99.0 98.8 98.8 98.5 97.3 95.3 96.5 97.0 96.4 96.5 97.1 96.7 97.4 97.0 97.1 95.4 95.5 95.7 95.4 95.2 95.1 95.0 95.1 94.9 99.0
5 98.0 98.5 99.0 98.7 **** 99.4 99.4 99.6 99.4 99.5 99.3 99.3 99.0 97.8 95.8 97.0 97.5 96.9 97.1 97.6 97.3 98.0 97.5 97.6 95.8 95.8 96.0 95.8 95.8 95.7 95.6 95.7 95.5 99.5
6 97.7 98.2 98.5 98.0 99.0 **** 99.5 99.7 99.5 99.6 99.4 99.4 99.1 97.6 95.9 97.1 97.6 97.0 97.2 97.7 97.2 97.9 97.6 97.7 95.5 95.6 95.7 95.5 95.7 95.6 95.5 95.6 95.4 99.6
7 98.2 98.5 99.0 98.7 99.5 99.0 **** 99.8 99.7 99.7 99.6 99.6 99.2 97.8 96.0 97.3 97.8 97.0 97.2 97.7 97.3 98.0 97.6 97.7 95.5 95.6 95.7 95.5 95.9 95.8 95.7 95.8 95.6 99.7
8 98.2 98.7 99.2 98.5 99.7 99.2 99.7 **** 99.8 99.9 99.7 99.7 99.4 98.0 96.2 97.4 98.0 97.2 97.3 97.9 97.5 98.2 97.8 97.9 95.7 95.7 95.9 95.7 96.0 95.9 95.8 95.9 95.8 99.9
9 98.0 98.5 99.0 98.2 99.5 99.0 99.5 99.7 **** 99.7 99.6 99.6 99.2 97.8 96.0 97.3 97.8 97.0 97.2 97.7 97.3 98.0 97.6 97.7 95.5 95.6 95.7 95.5 95.8 95.8 95.7 95.8 95.6 99.7
10 98.0 98.5 99.0 98.2 99.5 99.0 99.5 99.7 99.5 **** 99.7 99.7 99.3 97.9 96.1 97.3 97.9 97.1 97.3 97.8 97.4 98.1 97.7 97.8 95.6 95.7 95.8 95.6 95.9 95.9 95.8 95.9 95.7 99.8
11 98.0 98.5 99.0 98.2 99.5 99.0 99.5 99.7 99.5 99.5 **** 99.5 99.2 97.7 95.9 97.2 97.7 96.9 97.1 97.6 97.3 98.0 97.5 97.6 95.4 95.5 95.7 95.4 95.8 95.7 95.6 95.7 95.5 99.7
12 98.2 98.7 99.2 98.5 99.7 99.2 99.7 100 99.7 99.7 99.7 **** 99.3 97.9 96.1 97.5 98.0 97.1 97.3 97.8 97.4 98.1 97.7 97.7 95.7 95.8 96.0 95.7 95.9 95.9 95.8 95.9 95.7 99.7
13 97.4 98.0 98.5 97.7 99.0 98.5 99.0 99.2 99.0 99.0 99.0 99.2 **** 97.5 95.9 97.2 97.7 96.9 97.1 97.6 97.3 98.0 97.5 97.5 95.4 95.5 95.7 95.4 95.8 95.7 95.6 95.7 95.5 99.3
14 96.9 97.4 98.0 97.2 98.5 98.0 98.5 98.7 98.5 98.5 98.5 98.7 98.0 **** 95.4 96.7 97.3 96.8 97.0 97.5 97.2 97.7 97.3 97.3 95.3 95.4 95.5 95.2 95.4 95.3 95.3 95.4 95.2 97.9
15 94.8 94.8 94.8 94.0 95.3 95.1 95.3 95.6 95.3 95.3 95.3 95.6 94.8 94.8 **** 96.0 96.5 95.4 95.6 95.7 95.4 96.1 95.7 95.7 93.7 93.8 93.6 93.9 93.7 93.6 93.8 93.9 93.7 96.1
16 96.9 97.2 97.7 96.9 98.2 97.7 98.2 98.5 98.2 98.2 98.2 98.5 97.7 98.2 95.6 **** 99.0 96.6 96.8 97.2 96.9 97.5 97.2 97.2 95.1 95.0 95.0 94.7 95.3 95.2 95.2 95.3 95.1 97.3
17 97.4 97.7 98.2 97.4 98.7 98.2 98.7 99.0 98.7 98.7 98.7 99.0 98.2 98.7 95.6 99.5 **** 97.3 97.5 97.9 97.5 98.2 97.8 97.8 95.3 95.4 95.4 95.3 95.9 95.8 95.8 95.9 95.7 97.9
18 96.1 96.4 96.9 96.1 97.4 97.4 97.4 97.7 97.4 97.4 97.4 97.7 96.9 97.4 94.3 97.7 98.2 **** 99.8 99.2 98.6 98.5 98.1 98.1 95.4 95.5 95.5 95.4 95.8 95.9 95.9 96.0 95.8 97.1
19 96.4 96.7 97.2 96.4 97.7 97.7 97.7 98.0 97.7 97.7 97.7 98.0 97.2 97.7 94.6 98.0 98.5 99.7 **** 99.3 98.7 98.6 98.3 98.3 95.6 95.7 95.7 95.6 96.0 96.1 96.1 96.2 96.0 97.3
20 97.2 97.4 98.0 97.2 98.5 98.5 98.5 98.7 98.5 98.5 98.5 98.7 98.0 98.5 95.3 98.7 99.2 99.0 99.2 **** 99.2 99.2 98.8 98.8 96.1 96.2 96.2 96.1 96.6 96.6 96.6 96.7 96.6 97.8
21 96.9 97.2 97.7 96.9 98.2 97.7 98.2 98.5 98.2 98.2 98.2 98.5 97.7 98.2 95.1 98.5 99.0 98.0 98.2 99.0 **** 98.7 98.7 98.7 95.7 95.8 95.8 95.7 96.2 96.3 96.3 96.4 96.2 97.4
22 97.4 97.7 98.2 97.4 98.7 98.2 98.7 99.0 98.7 98.7 98.7 99.0 98.2 98.7 95.6 99.0 99.5 98.7 99.0 99.7 99.2 **** 99.4 99.4 96.6 96.7 96.7 96.6 96.7 96.8 96.8 96.9 96.7 98.1
23 96.4 96.7 97.2 96.4 97.7 97.7 97.7 98.0 97.7 97.7 97.7 98.0 97.2 97.7 94.6 98.0 98.5 98.0 98.2 99.0 99.0 98.7 **** 99.7 96.2 96.3 96.3 96.2 96.3 96.4 96.4 96.5 96.3 97.7
24 96.9 97.2 97.7 96.9 98.2 98.2 98.2 98.5 98.2 98.2 98.2 98.5 97.7 98.2 95.1 98.5 99.0 98.5 98.7 99.5 99.5 99.2 99.5 **** 96.2 96.3 96.3 96.2 96.3 96.4 96.4 96.5 96.3 97.8
25 96.9 97.2 97.7 96.9 98.2 97.7 98.2 98.5 98.2 98.2 98.2 98.5 97.7 98.2 95.1 98.5 99.0 98.0 98.2 99.0 98.7 99.2 98.2 98.7 **** 99.7 99.4 99.2 95.6 95.7 95.8 96.3 95.9 95.6
26 96.7 96.9 97.4 96.7 98.0 97.4 98.0 98.2 98.0 98.0 98.0 98.2 97.4 98.0 94.8 98.2 98.7 97.7 98.0 98.7 98.5 99.0 98.0 98.5 99.7 **** 99.3 99.2 95.7 95.8 95.9 96.4 96.0 95.7
27 96.9 97.4 98.0 97.2 98.5 98.0 98.5 98.7 98.5 98.5 98.5 98.7 98.0 98.5 94.8 98.2 98.7 97.7 98.0 98.7 98.5 99.0 98.0 98.5 99.7 99.5 **** 99.1 95.5 95.6 95.8 96.2 95.8 95.8
28 96.9 97.2 97.7 96.9 98.2 97.7 98.2 98.5 98.2 98.2 98.2 98.5 97.7 98.2 95.1 98.5 99.0 98.0 98.2 99.0 98.7 99.2 98.2 98.7 100 99.7 99.7 **** 95.4 95.5 95.7 96.1 95.8 95.6
29 95.9 96.1 96.7 95.9 97.2 96.7 97.2 97.4 97.2 97.2 97.2 97.4 96.7 97.2 94.0 97.4 98.0 96.7 96.9 97.7 97.4 98.0 96.9 97.4 98.2 98.0 98.0 98.2 **** 99.6 99.2 99.2 99.0 95.9
30 96.4 96.7 97.2 96.4 97.7 97.2 97.7 98.0 97.7 97.7 97.7 98.0 97.2 97.7 94.6 98.0 98.5 97.2 97.4 98.2 98.0 98.5 97.4 98.0 98.7 98.5 98.5 98.7 99.5 **** 99.3 99.2 99.1 95.9
31 95.9 96.1 96.7 95.9 97.2 96.7 97.2 97.4 97.2 97.2 97.2 97.4 96.7 97.2 94.0 97.4 98.0 96.7 96.9 97.7 97.4 98.0 96.9 97.4 98.2 98.0 98.0 98.2 99.0 99.5 **** 99.4 99.2 95.8
32 96.4 96.7 97.2 96.4 97.7 97.2 97.7 98.0 97.7 97.7 97.7 98.0 97.2 97.7 94.6 98.0 98.5 97.2 97.4 98.2 98.0 98.5 97.4 98.0 98.7 98.5 98.5 98.7 99.5 100 99.5 **** 99.7 95.8
33 96.1 96.4 96.9 96.1 97.4 96.9 97.4 97.7 97.4 97.4 97.4 97.7 96.9 97.4 94.3 97.7 98.2 96.9 97.2 98.0 97.7 98.2 97.2 97.7 98.5 98.2 98.2 98.5 99.2 99.7 99.2 99.7 **** 95.7
34 98.0 98.5 99.0 98.2 99.5 99.0 99.5 99.7 99.5 99.5 99.5 99.7 99.0 98.5 95.6 98.2 98.7 97.4 97.7 98.5 98.2 98.7 97.7 98.2 98.2 98.0 98.5 98.2 97.2 97.7 97.2 97.7 97.4 ****
European isolates LBVaV Australian isolates LBVaV
Japanese
isolates LBVaV
Japanese
isolates TStV

176
References
Australian Bureau of Statistics. 2009a. 7121.0 Agricultural Commodities, Australia,
2007-08. Available World Wide Web. URL:
http://www.abs.gov.au/AUSSTATS/[email protected]/DetailsPage/7121.02007-
08?OpenDocument Accessed 17 November, 2009.
Australian Bureau of Statistics. 2009b. Value of Agricultural Commodities Produced,
Australia, 2007-08. Data cube: Excel spreadsheet cat. no. 75030DO001_200708.
Available World Wide Web. URL:
http://www.abs.gov.au/AUSSTATS/[email protected]/DetailsPage/7503.02007-
08?OpenDocument Accessed 17 November, 2009.
Barbetti, M.J. 1981. Effects of sowing date and oospore seed contamination on
subsequent crop incidence of white rust (Albugo candida) in rapeseed. Australas. Plant
Path. 10:44-46
Barr, D.J.S. 1988. Zoosporic plant parasites as fungal vectors of viruses: taxonomy and
lifecycle of species involved. In: J. I. Cooper and M.J.C. Asher, Editors, Viruses with
fungal vectors, Association of Applied Biologists, Wellesbourne, UK, pp 123-137
Borthwick, H.A. and Robbins, W.W. 1928. Lettuce seed and its germination. Hilgardia.
3:275-305
Bos, L. 1999. Plant viruses, unique and intriguing pathogens – a textbook of plant
virology. Backhuys Publishers, Leiden, pp. 37, 38, 226.
Bos, L. and Huijberts, N. 1990. Screening for resistance to big vein disease of lettuce.
Crop Prot. 9:446-452
Brittlebank, C.C. 1919. Tomato diseases. J. Agric. 17:231-235
Büchen-Osmond C (ed.) (2006) Index of Viruses – Varicosavirus. In: ICTVdB - The
Universal Virus Database, version 4. Columbia University, New York
Campbell, R.N. 1962. Relationship between the lettuce big-vein and its vector Olpidium
brassicae. Nature. 195:675-677
Campbell, R.N. 1965. Weeds as reservoir hosts of the Lettuce big-vein virus. Can. J.
Botany. 43:1141-1149
Campbell, R.N. 1985. Longevity of Olpidium brassicae in air-dried soil and the
persistence of the lettuce big-vein agent. Can. J. Botany. 63:2288-2289
Campbell, R.N. 1996. Fungal transmission of plant viruses. Annu. Rev. Phytopathol.
34:87-108

177
Campbell, R.N., Greathead, R.G. and Westerlund, F.V. 1980. Big-Vein of lettuce:
infection and methods of control. Phytopathology. 70:741-746
Campbell, R.N. and Grogan, R.G. 1963. Big-vein virus of lettuce and its transmission
by Olpidium brassicae. Phytopathology. 53:252-259
Campbell, R. N., and Grogan R. G. 1964. Aquisition and transmission of lettuce big-
vein virus by Olpidium brassicae. Phytopathology 54: 681-690
Campbell, R.N., Grogan, R.G. and Purcifull, D.E. 1961. Graft transmission of big-vein
of lettuce. Virology. 15:82-85
Campbell, R. N., and Lin M. T., 1976. Morphology and thermal death point of
Olpidium brassicae. Am. J. Bot. 63: 826-832
Campbell, R.N., Wipf-Scheibel, C. and Lecoq, H. 1996. Vector-assisted seed
transmission of Melon necrotic spot virus in melon. Phytopathology. 86:1294-1298
Cho, J.J., Mau, R.F.L., German, T.L., Hartman, R.W., Yudin, L.S., Gonsalves, D. and
Provvidenti, R. 1989. A multi-disciplinary approach to management of tomato spotted
wilt virus. Plant Dis. 73:375–83
Clark, M.F. and Adams, A.N. 1977. Characteristics of the microplate method of
Enzyme-Linked Immunosorbent Assay for the detection of plant viruses. J. Gen. Virol.
34:475-483
Colariccio, A., Chaves, A.L.R., Eivas, M., Chagas, C.M., Lenzi, R. and Roggero, P.
2003. Presence of lettuce big-vein and associated viruses in a sub-tropical area of
Brazil. New Disease Report. 7:1-2
Coutts, B.A., Thomas-Carroll, M.L. and Jones, R.A.C. 2004. Analysing spatial patterns
of spread of Lettuce necrotic yellows virus and lettuce big-vein disease in lettuce field
plantings. Ann. Appl. Biol. 145:339-343
Davison, E.M., Drenth, A., Kumar, S., Mack, S., Mackie, A.E. and McKirdy, S. 2006.
Pathogens associated with nursery plants imported into Western Australia. Australas.
Plant Path. 35:473-475
Fanigliulo, A., Pacella, A., Comes, S. And Crescenzi, A. 2007. Three years survey of
Tomato yellow leaf curl Sardinia virus reservoir weed hosts in southern Italy.
Communic. Agric. Appl. Biol. Sci. 72:1023-1028
Faruki, I. 1981. Epidemiological studies on Melon necrotic spot. Technical Bulletin 14.
Shizuoka Agricultural Experiment Station, Shizuokaken, Japan
Floyd, R. 2000. Diseases of lettuce. Farmnote 48/91. Department of Agriculture and
Food, Western Australia

178
Francki, R.I.B. and Randles, J.W. 1970. Descriptions of Plant Viruses: Lettuce necrotic
yellows virus. Assoc. Appl. Biol. 26. Available World Wide Web. URL:
http://www.dpvweb.net/dpv/showdpv.php?dpvno=26 Accessed 17 Nov, 2009
Fry, P.R. 1958. The relationship of Olpidium brassicae (Wor.) Dang. to big-vein
disease of lettuce. NZ. J. Agr. Res. 1:301-304
Fry, P.R. and Campbell, R.N. 1966 Transmission of Tobacco necrosis virus by
Olpidium brassicae. Virology 30:517-527
Fry, P.R., Close, R.C., Proctor, C.H. and Sunde, R. 1973. Lettuce necrotic yellows virus
in New Zealand. NZ. J. Agr. Res. 16:143-146
Funnekotter, B. 2006. Personal communication. 16th
March.
Garcia-Arenal F, Fraile A, Malpica JM (2001) Variability and genetic structure of plant
virus populations. Annu. Rev. Phytopathol. 39: 157-186
Germundsson, A., Sandgren, M., Barker, H., Savenkov, E.I. and Valkonen, J.P.T. 2002.
Initial infection of roots and leaves reveals different resistant phenotypes associated
with coat protein gene-mediated resistance to Potato mop top virus. J. Gen. Virol.
83:1201-1209
Hall, T. A. 1999. BioEdit: a user-friendly biological sequence alignment editor and
analysis program for Windows 95/98/NT. Nucl. Acid. Sym. Ser. 41: 95-98
Hardy, G.E. and Sivasithamparam, K. 1988. Phytophthora spp. associated with
container-grown plants in nurseries in Western Australia. Plant Dis. 72:435-437
Hartwright, L. M., Hunter, P. J., and Walsh J. A. 2009. A comparison of Olpidium
isolates from a range of host plants using internal transcribed spacer sequence analysis
and host range studies. Mycol. Res. Available online 1 October, 2009. doi:
10.1016/j.mycres.2009.09.008
Hayes, R.J., Wintermantel, W.M., Nicely, P.A. and Ryder, E.J. 2006. Host resistance to
Mirafiori lettuce big-vein virus and Lettuce big-vein associated virus and virus sequence
diversity and frequency in California. Plant Dis. 90:233-239
Herrera-Vasquez, J. A., Cebrian, M. C., Alfaro-Fernandez, A., Cordoba-Selles, M. C.,
and Jorda, C. (2009). Multiplex PCR assay for the simultaneous detection and
differentiation of Olpidium bornovanus, Olpidium brassicae and Olpidium virulentus.
Mycol. Res. 113:602-610
Huijberts, N., Blystad, D.R. and Bos, L. 1990. Lettuce big-vein virus: mechanical
transmission and relationships to tobacco stunt virus. Ann. Appl. Biol. 116:463-475
Jagger, I.C. and Chandler, N. 1934. Big-Vein, a disease of lettuce. Phyotpathology
24:1253-1256

179
James, T. Y., Letcher, P. M., Longcore, J. E., Mozley-Standridge, S. E., Porter, D.,
Powell, M. J., Griffith, G. W., and Vilgalys, R. (2006) A molecular phylogeny of
flagallated fungi (Chytridiomycota) and description of a new phylum (Blastidiomycota).
Mycologia 98:860-871
Jones, R. A. C., 2004. Using epidemiological information to develp effective integrated
virus disease management strategies. Virus Res. 100: 5-30
Jones, R.L. 1974. The structure of the lettuce endosperm. Planta. 121:133-146
Kawazu, Y., Sasaya, T., Morikawa, T., Sugiyama, K. and Natsuaki, T. 2003. Nucleotide
sequence of the coat protein gene of Mirafiori lettuce virus. J. Gen. Plant Pathol. 69:55–
60
Koganezawa, H., Inoue, H., and Sasaya, J. 2005. Host specificity and multiplication of
eight isolates of Olpidium brassicae sensu lato and its related Olpidium spp. Japanese
National Agricultural Research Centre for Western Region, Miscellaneous Bulletin, pp.
39-59
Koganezawa, H., Takayama, T. and Sasaya, T. 2004. Difference in resting spore
formation between crucifer and non-crucifer strains of Olpidium brassicae sensu lato
(In Japanese). Jpn. J. Phytopathol. 70:307-313
Kosakovsky-Pond, S.L. and Frost, S.D.W. 2005a. Datamonkey: rapid detection of
selective pressure on individual coding sites of codon alignments. Bioinformatics.
21:2531-2533
Kosakovsky-Pond, S.L. and Frost, S.D.W. 2005b. Not so different after all: a
comparison of methods for detecting amino acid sites under selection. Mol. Biol. And
Evol. 22:1208-1222
Kuwata, S., Kubo, S. 1981. Rod-shaped particles found in tobacco plants infected with
tobacco stunt agent. Ann. Phytopath. Soc. Japan 47: 264-268
Kuwata, S. and Kubo, S. 1986. Tobacco stunt virus. Commonwealth Mycological
Institute/ Association of Applied Biologists Descriptions of Plant Viruses. No. 313 pp 4
Kuwata, S., Kubo, S., Yamashita, S. and Doi, Y. 1983. Rod-shaped particles, a probable
entity of lettuce big-vein virus. Ann. Phytopath. Soc. Japan 49: 246-251
Latham, L.J. and Jones, R.A.C. 1997. Occurrence of Tomato spotted wilt tospovirus in
native flora, weeds and horticultural crops. Aust. J. Agr. Res. 48:359-369
Latham, L.J. and Jones, R.A.C. 2004. Deploying partially resistant genotypes and
plastic mulch on the soil surface to suppress spread of lettuce big-vein disease in lettuce.
Aust. J. Agr. Res. 55:131-138
Latham, L.J., Jones, R.A.C. and Mckirdy, S.J. 2004. Lettuce big-vein disease: sources,
patterns, and losses. Aust. J. Agr. Res. 55:125-130

180
Lindqvist K, 1960. On the origin of cultivated lettuce. Hereditas 46: 319-350
Lot, H., Campbell, R.N., Souche, S., Milne, R.G. and Roggero, P. 2002. Transmission
by Olpidium brassicae of Mirafiori lettuce virus and Lettuce big-vein virus, and their
roles in lettuce big-vein etiology. Phytopathology 92:288-293
McKirdy, S., Mackie, A.E. and Kumar, S. 2001. Apple scab successfully eradicated in
Western Australia. Australas. Plant. Path. 30:371
Mackie, A.E., McKirdy, S.J., Rodoni, B. And Kumar, S. 2002. Potato tuber spindle
viroid eradicated in Western Australia. Australas. Plant. Path. 31:311-312
Martin, S., Lopez, C., Garcia, M.L., Naum-Ongania, G, Grau, O., Flores, R., Moreno, P.
and Guerri, J. 2006. Genetic variation of populations of Citrus psorosis virus. J. Gen.
Virol. 87:3097-3102
Matthews, R.E.F. 1981. Plant Virology. 2nd
edition. Academic Press, New York. pp.
285-287
Mayo, M.A. 2000. Genus Varicosavirus. In: van Regenmortel MHV, Fauquet CM,
Bishop DHL, Carstens EB, Estes MK, Lemon SM, Maniloff J, Mayo MA, McGeoch
DJ, Pringle CR, Wickner RB (eds) Virus Taxonomy. Seventh Report of the
International Committee on Taxonomy of Viruses. Academic Press, San Diego,.
pp.521-523
Napier, T. 2004. Field lettuce production. Agfact H8.1.40. NSW Agriculture
Navarro, J.A., Botella, F., Maruhenda, A., Sastre, P., Sanchez-Pina, M.A. and Pallas, V.
2004. Comparative infection progress analysis of Lettuce big-vein virus and Mirafiori
lettuce virus in lettuce crops by developed molecular diagnostic techniques.
Phytopathology 94:470-477
Navarro, J.A., Torok, V.A., Vetten, H.J. and Pallas, V. 2005a. Genetic variability in the
coat protein genes of Lettuce big-vein associated virus and Mirafiori lettuce big-vein
virus. Arch. Virol. 150:681-694
Navarro, J.A., Botella, F., Maruhenda, A., Sastre, P., Sanchez-Pina, M.A. and Pallas, V.
2005b. Identification and partial characterisation of Lettuce big-vein associated virus
and Mirafiori lettuce big-vein virus in common weeds found amongst Spanish lettuce
crops and their role in lettuce big-vein disease transmission. Eur. J. Plant Pathol.
113:25-34
Nijsse, J., Erbe, E., Brantjes, N.B.M., Schel, J.H.N. and Wergin, W.P. 1998. Low-
temperature scanning electron microscopic observations on endosperm in imbibed and
germinated lettuce seeds. Can. J. Botany. 76:509-516
O‟Keefe, D.C., Berryman, D.I., Coutts, B.A. and Jones, R.A.C. 2007. Lack of seed coat
contamination with Cucumber mosaic virus in lupin permits reliable, large scale
detection of seed transmission in seed samples. Plant Dis. 91:504-508

181
Paulson, R.E. and Srivastava, L.M. 1968. The fine structure of the embryo of Lactuca
sativa. I. Dry embryo. Can. J. Botany. 46:1437-1445
Peters, D. 1998. An updated list of plant species susceptible totospoviruses. In
Proceedings of the Fourth International Symposium on Tospoviruses and Thrips
in Floral and Vegetable Crops, pp. 107-110, Eds D Peters and R Peters D. 1998
Goldbach.Wageningen, The Netherlands
Randles, J.W. and Carver, M. 1971. Epidemiology of Lettuce necrotic yellows virus in
South Australia. Aust. J. Agr. Res. 22:231-237
Rochon, D‟A., Kakani, K., Robbins, M., and Reade, R. 2004. Molecular aspects of plant
virus transmission by Olpidium and Plasmodiophorid vectors. Ann. Rev. Phytopathol.
42: 211-241
Roggero, P., Ciufffo, M., Vaira, A.M., Accotto, G.P., Masenga, V. and Milne, R.G.
2000. An ophiovirus isolated from lettuce with big-vein symptoms. Arch. Virol.
145:2629-2642
Roggero, P., Lot, H., Souche, S., Lenzi, R. and Milne, R.G. 2003. Occurrence of
Mirafiori lettuce virus and Lettuce big-vein virus in relation to development of big-vein
symptoms in lettuce crops. Eur. J. Plant Pathol. 109:261-267
Ryder, E.J. 1973. Seed transmission of LMV in mosaic resistant lettuce. J. Am. Soc.
Hort. Sci. 95:610-614
Sahtiyanci, S. 1962. Studien über einige wurzelparasitäre Olpidiaceen. Arch. Microbiol.
41: 187–228
Sambrook, J., and Russell, D. W. 2001. Molecular cloning: A laboratory manual. New
York: Cold Spring Harbour Laboratory Press
Sampson, K. 1939. Olpidium brassicae (Wor.) Dangeard and its connection with
Asterocystis vadicis de Wildeman. Trans. Br. Mycol. Soc. 23:199-205
Sanches, M.M. 2006. Molecular and biological charatcterization of lettuce big-vein
associated virus and Mirafiori lettuce big-vein virus and study of the occurrence during
the year and on lettuce plantings showing symptoms from the Sao Paulo state. Thesis.
FCA-UNESP Botucatu, Sao Paulo, Brazil.
Sasaya, T., Fujii, H., Ishikawa, K. and Koganezawa, H. 2006. Further evidence of
Mirafiori lettuce big-vein virus but not of Lettuce big-vein associated virus with big-
vein disease in lettuce. Phytopathology. 98:464-468
Sasaya T, Ishikawa K, Koganezawa H (2001) Nucleotide sequence of the coat protein
gene of Lettuce big-vein virus. J. Gen. Virol. 82: 1509-1515
Sasaya T, Ishikawa K, Kuwata S, Koganezawa H (2005) Molecular analysis of coat
protein coding region of tobacco stunt virus shows that it is a strain of Lettuce big-vein
virus in the genus Varicosavirus. Arch. Virol. 150: 1013-1021

182
Sasaya, T., and Koganezawa, H. 2006. Molecular analysis and virus transmission tests
place Olpidium virulentus, a vector of Mirafiori lettuce big-vein virus and Tobacco stunt
virus, as a distinct species rather than a strain of Olpidium brassicae. J. Gen. Plant
Pathol. 72: 20-25
Schenk, P.M., Antoniw, J.F., Batista., M.D., Jacobi, V., Adams, M.J. and Steinbiss,
H.H. 1995. Movement of Barley mild mosaic and Barley yellow mosaic viruses in
leaves and roots of barley. Ann. Appl. Biol. 126:291-305
Sivasithamparam, K. and Goss, O.M. 1981. „Jarrah dieback‟... a threat to horticulture. J.
Ag. WA. 21:88-91
Stubbs, L.L. and Grogan, R.G. 1963. Necrotic yellows: A newly recognised virus
disease of lettuce. Aust. J. Agr. Res. 14:439-459
Tamura K., Dudley J., Nei M. and Kumar S. 2007. MEGA4: molecular evolutionary
genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 24:1596-1599
Teakle, D. S. 1960. Association of Olpidium brassicae and tobacco necrosis virus.
Nature 188: 431-432
Teakle, D.S. and Thomas, B.J. 1985. Effect of heat on zoospore motility and
multiplication of Olpidium radicale and O. brassicae. Ann. Appl. Biol. 107:11-15
Temmink, J.H.M., Campbell, R.N. and Smith, P.R. 1970. Specificity and site of in vitro
acquisition of tobacco necrosis virus by zoospores of Olpidium brassicae. J. Gen. Virol.
9:201-213
Thompson, J.D., Higgins, D.G. and Gibson, T.J. 1994. ClustalW: Improving the
sensitivity of progressive multiple sequence alignment through sequence weighting,
position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 22:4673-
4680
Tomlinson, J.A. 1988. Chemical control of Spongospora and Olpidium in hydroponic
systems and in soil. In: Cooper, J.I. and Asher, M.J.C. (Eds.), Developments in Applied
Biology 2: Viruses with fungal vectors. Association of Applied Biologists. Warwick,
UK. pp. 293-303
Ufer, T., Beltz, H., Brand, T., Kaminski, K., Luttman, R., Posner, M., Wagner, S.,
Werres, S. and Wessels, H-P. 2005. Introduction of filtration systems in container
nurseries for nonchemical elimination of Phytophthora spp. from irrigation water.
Proceedings of the sudden oak death second symposium: the state of our knowledge. Pg
287-289
Vetten, H.J., Lesemann, D.E. and Dalchow, J. 1987. Electron microscopical and
serological detection of virus-like particles associated with lettuce big vein disease. J.
Phytopathol. 120: 53-59
Walsh, J.A. 1994 Effects of some biotic and abiotic factors on symptom expression of
lettuce big-vein virus in lettuce (Lactuca sativa). J. Hort. Sci. 69:21-28

183
Webster, C.G., Coutts, B.A., Jones, R.A.C., Jones, M.G.K. and Wylie, S.J. 2007. Virus
impact at the interface of an ancient ecosystem and a recent agroecosystem: studies on
three legume-infecting potyviruses in southwest Australian floristic region. Plant Pathol.
56:729-742
Westerlund, F. V., Campbell, R. N., and Grogan, R. G. 1978. Effect of temperature on
transmission, translocation, and persistence of the lettuce big-vein agent and big-vein
symptom expression. Phytopathology 68:921-926
Whitaker, T.W. 1969. Salads for everyone – A look at the lettuce plant. Econ. Bot. 23:
261-264
White, T. J., Bruns, T., Lee, S., and Taylor, J. W. 1990. Amplification and direct
sequencing of fungal ribosomal RNA genes for polygenetics, pp. 315-322 In: M. A.
Inns, D. H. Gelfand, J. J. Sninsky and T. J. White, Editors PCR Protocols: A Guide to
Methods and Applications. Academic Press, Inc., New York
Wilson, C. R. 1998. Incidence of weed reservoirs and vectors of Tomato spotted wilt
tospovirus on South Tasmanian lettuce farms. Plant. Pathol. 47:171-176
Woods, W.M., Trend, W.J., Thwaite, G., Monzu, N., Portman, A., Power, G.J. and
Davis, P.R. 2001. The history of coddling moth eradication from WA. Good Fruit and
Vegetables. 12:79-81
van der Wilk, F., Dullemans, A. M., Verbeek, M. and van den Heuvel, J. F. 2002.
Nucleotide sequence and genomic organization of an ophiovirus associated with lettuce
big-vein disease. J. Gen. Virol. 83:2869–2877
Zitter, T.A. and Daughtrey, M.L. 1989. Tomato spotted wilt virus. Fact Sheet
Page:735.90. Cornell University. Available World Wide Web. URL:
http://vegetablemdonline.ppath.cornell.edu/factsheets/Virus_SpottedWilt.htm Accessed
17 Nov, 2009

BRIEF REPORT
Comparison of the coat protein genes of Lettuce big-vein associatedvirus isolates from Australia with those of isolates from othercontinents
Linda D. Maccarone • Martin J. Barbetti •
Krishnapillai Sivasithamparam • Roger A. C. Jones
Received: 30 November 2009 / Accepted: 17 February 2010
� Springer-Verlag 2010
Abstract The complete coat protein (CP) nucleotide
sequences of seven Lettuce big-vein associated virus
(LBVaV) isolates from Australia were compared to those
of 22 other LBVaV and five tobacco stunt virus (TStV)
isolates. On phylogenetic analysis, clade I contained only
LBVaV isolates from Europe, sub-clade IIa only Australian
LBVaV isolates, IIb only Japanese LBVaV isolates, and IIc
only TStV isolates from Japan. In the amino acid sequences
deduced, the central region of the gene was most divergent.
Mean Dn/Ds ratios were 0.283 and 0.124 for clades I and
II, respectively. The suggestion that TStV is a strain of
LBVaV was supported.
Lettuce big-vein associated virus (LBVaV) and tobacco
stunt virus (TStV) belong to the genus Varicosavirus of
which LBVaV is type species [1]. LBVaV was formerly
considered the cause of lettuce big-vein disease (LBVD)
[16] until another virus occurring in mixed infection,
Mirafiori lettuce big-vein virus (MLBVV; genus Ophiovi-
rus), was found responsible for the symptoms [5, 7]. TStV
infects tobacco causing severe symptoms [4]. LBVaV and
TStV have rod shaped particles 320–360 (LBVaV) or 300–
340 (TStV) nm in length [4, 16]. Both have a CP 1,194
nucleotides long that codes for 397 amino acids [12] and
are serologically related [4, 16]. Both were considered
different species despite their similar morphology, serology
and vector relationships because TStV does not to infect
lettuce but infects tobacco whilst LBVaV does the opposite
[4]. Subsequently, Sasaya et al. [12] suggested that TStV is
a tobacco infecting strain of LBVaV. TStV and MLBVV
are transmitted by the chytrid Olpidium virulentus [13].
Sasaya et al. [12] reported that the complete nucleotide and
amino acid CP sequence identities between eight LBVaV
and five TStV isolates were 95.6–96.5% and 97.2–98.7%,
respectively. When Navarro et al. [9] analysed the CP
sequences of 17 LBVaV isolates, they found identities of
95.2–99.8% (nucleotides) and 98.0–100% (amino acids),
and when they subjected 13 complete and four partial
LBVaV CP sequences to phylogenetic analysis, three iso-
late groupings were found. This paper compares the CPs of
seven previously unreported isolates from Australia with
those of 30 LBVaV and five TStV isolates from different
continents.
Lettuce leaves with LBVD symptoms were sampled
from four farms (A, B, E and G) in the northern Perth
Metropolitan region, south-west Australia. Seven samples
were collected, one each from A and G, two from B and
three from E. On the collection day, each sample was
ground in liquid nitrogen before storage at -20�C before
RNA extraction. Total RNA was extracted from each
sample using an UltraCleanTM Plant RNA Isolation Kit
(MoBio Laboratories, Inc.) according to manufacturer’s
instructions. RNA extracts were stored at -80�C. Primer
pairs specific for the CP of LBVaV (fwd L1-21 50ATG
GCA CCC CAA ATT GAA G30, rev L700-679 50GTG
CAT GCC AGT GCT GGC AAG 30 and fwd L511-5 31
50AGA CCG AGT ATC CAT TCA AG 30, rev L1210-1185
50TCA CTC CTT CAC TGG TGT CTC TCC C30) were
L. D. Maccarone � M. J. Barbetti � K. Sivasithamparam �R. A. C. Jones (&)
Faculty of Natural and Agricultural Sciences,
School of Plant Biology, University of Western Australia,
Stirling Highway, Crawley, WA 6009, Australia
e-mail: [email protected]
R. A. C. Jones
Department of Agriculture and Food Western Australia,
Agricultural Research Western Australia, Locked Bag No. 4,
Bentley Delivery Centre, Perth, WA 6983, Australia
123
Arch Virol
DOI 10.1007/s00705-010-0641-0

developed by aligning known CP sequences from GenBank
in Bioedit. To amplify the entire CP sequence, both primer
sets were used on each sample. Reverse transcription PCR
(RT-PCR) was done using a OneStep RT-PCR kit (Qiagen,
Australia) according to manufacturer’s instructions. PCR
amplification conditions consisted of 50�C for 30 min,
95�C for 15 min, followed by 30 cycles of 94�C for 1 min,
57�C for 1 min, 72�C for 1 min, then a final extension at
72�C for 10 min before being held at 14�C. PCR amplicons
were visualised by electrophoresis on 1% agarose gels.
Table 1 Isolates of Lettuce big-
vein associated virus (LBVaV)
and tobacco stunt virus (TStV)
used in sequence comparisons
a Complete LBVaV CP
nucleotide sequences
AB050272 (isolate LBVaV-CP)
and AB114138 (isolate Ka),
both from Kagawa in Japan,
were identicalb Partial LBVaV CP sequencesc TStV isolates
Virus isolate Accession number Geographic origin Species isolated from Reference
LBVaV-CPa AB050272a Kagawa, Japan Lettuce [9, 11, 12]
No2c AB190521 Takamatsu, Japan Tobacco [12]
No89c AB190522 Chunan, Japan Tobacco [12]
No88c AB190523 Chunan, Japan Tobacco [12]
Kanc AB190524 Kannonji, Japan Tobacco [12]
Nac AB190525 Fukue, Japan Tobacco [12]
A AB190526 Kannonji, Japan Lettuce [12]
Hy AB190527 Nandan, Japan Lettuce [12]
Wa AB190528 Hiochigawa, Japan Lettuce [12]
MUR1 AY366411 Murcia, Spain Lettuce [9]
GAL1 AY366412 Galicia, Spain Lettuce [9]
ALM1 AY366413 Pulpi, Almeria, Spain Lettuce [9]
ALM2 AY366414 Pulpi, Almeria, Spain Lettuce [9]
USAb AY496053 Salinas, CA, USA Lettuce [9]
UKb AY496054 Wellesbourne, UK Lettuce [9]
AUSb AY496055 Perth, south-west Australia Lettuce [9]
NLb AY496056 De Lier, The Netherlands Lettuce [9]
ALM3 AY581683 Pulpi, Almeria, Spain Lettuce [9]
ALM4 AY581685 Pulpi, Almeria, Spain Lettuce [9]
ALM5 AY581686 Pulpi, Almeria, Spain Lettuce [9]
ALM6 AY581687 Pulpi, Almeria, Spain Lettuce [9]
ALM7 AY581688 Pulpi, Almeria, Spain Lettuce [9]
GRA1 AY581689 Granada, Spain Lettuce [9]
UK2 AY581690 UK Lettuce [9]
MUR2 AY581691 Aguilas, Murcia, Spain Lettuce [9]
SON1 AY581681 Almeria, Spain Sowthistle [8]
SON2 AY839619 Almeria, Spain Sowthistle [8]
SON3 AY839620 Almeria, Spain Sowthistle [8]
SON4 AY839621 Almeria, Spain Sowthistle [8]
SON5 AY839622 Granada, Spain Sowthistle [8]
SON6 AY839623 Granada, Spain Sowthistle [8]
107b DQ530352 Bauru, Brazil Lettuce Unpublished
104b DQ530353 Campinas, Brazil Lettuce Unpublished
105b DQ530354 Bauru, Brazil Lettuce Unpublished
106b DQ530355 Mogi das Cruzes, Brazil Lettuce Unpublished
AUSE4 GU220721 Perth, south-west Australia Lettuce This study
AUSE5 GU220722 Perth, south-west Australia Lettuce This study
AUSA5 GU220723 Perth, south-west Australia Lettuce This study
AUSB1 GU220724 Perth, south-west Australia Lettuce This study
AUSB2 GU220725 Perth, south-west Australia Lettuce This study
AUSG1 GU220726 Perth, south-west Australia Lettuce This study
AUSE6 GU220727 Perth, south-west Australia Lettuce This study
L. D. Maccarone et al.
123

Tris-boric acid-EDTA (TBE) electrophoresis buffer was
used [10].
PCR amplicons were purified by ethanol precipitation
and quantified using a Nanodrop UV–vis spectrophotom-
eter following the manufacturer’s protocol. Standard liga-
tion and transformation reactions were done into JM109
competent cells using a pGEM�-T Easy Vector (Promega
Corp, Madison, Wisconsin) as recommended by the man-
ufacturer. Screening for recombinant plasmids used M13
primers: M13 fwd (50CGT CAG GCT TTT CCC AGT
CAC GAC30) and M13 rev (50TCA CAC AGG AAA CAG
CTA TGA C30). Amplification conditions were 1 min
denaturation at 94�C followed by 25 cycles of 94�C for
30 s, 55�C for 30 s, 72�C for 30 s before being held at
14�C. Recombinant plasmids were inoculated into 5 mL of
Luria–Bertani (LB) broth with ampicillin (100 mg/L) and
incubated on a shaker at 37�C overnight. The plasmid was
then purified from the culture using an AurumTM Bio-Rad
Plasmid Extraction Kit following the manufacturer’s pro-
tocol. Presence and size of inserts was confirmed with a
restriction digest of purified plasmid DNA by EcoR1
(10 lL of purified plasmid DNA, 2 lL of 109 EcoR1
buffer, 1 lL of EcoR1 and 7 lL of water). Dideoxy-ter-
mination sequencing was done at the State Agricultural
Biotechnology Centre (SABC), Murdoch University using
BigDye terminator V3.1 chemistry and an Applied Bio-
systems/Hitachi 37030 DNA Analyser. The complete CP
sequences of each isolate were submitted to GenBank
(Table 1). One clone was sequenced per isolate.
The complete CP genes of the seven new Australian
LBVaV isolates contained one ORF 1,194 bp in length that
coded for 397 amino acids. In addition, 22 complete and
eight partial CP sequences of other LBVaV isolates and
complete CP sequences of five TStV isolates were
retrieved from GenBank (Table 1). The partial sequences
were four from Brazil [DQ530352 (889 bp), DQ530353
(882 bp), DQ530354 (866 bp) and DQ530355 (876 bp)]
and one each from USA (AY496053), UK (AY496054),
Netherlands (AY496056) and Australia (AY496055) (all
1,010 bp in length). Complete CP nucleotide sequences
AB050272 (isolate LBVaV-CP) and AB114138 (isolate
Ka), both from Kagawa Japan, were identical so only the
former was included. A partial LBVaV sequence from
Salinas, CA (DQ182566) was excluded as it was only
302 bp [2]. The 34 complete LBVaV and TStV CP
sequences were assembled and analysed using Molecular
Evolutionary Genetics Analysis (MEGA) program Version
4.1 with Clustal W [15] and bootstrapped with 1,000
re-samplings [14]. These sequences were analysed in direct
pairwise comparisons to establish percentage identities.
Evolutionary history was inferred by the Neighbour-Join-
ing method. Evolutionary distances between nucleotide
sequences were computed using the Maximum Composite
Likelihood method. This was done first with the 29 com-
plete CP sequences of LBVaV and five of TStV, and then
repeated to include the eight partial sequences after trim-
ming complete sequences to reflect the length of the partial
sequences. Evolutionary distances of deduced amino acid
sequences were computed for the complete CP sequences
using the Poisson correction method in MEGA4.1. Any
positions containing gaps and missing data were elimi-
nated. Nucleotide substitutions for the CP region were
inferred using the SLAC model [3]. Next, the mean number
of non-silent substitutions (Dn) and silent substitutions
(Ds), and Dn/Ds ratios were determined to assess the
selective constraints on the CP gene [3]. Dn/Ds ratios for
the sequences of complete CP genes were estimated first
for the seven Australian isolates alone and then for all
complete nucleotide sequences.
The sequence identities of the seven new Australian
isolates were 95.6–99.7% (nucleotides) and 98.2–99.7%
(amino acids). They formed a geographically distinct group
most closely related to each other and to LBVaV isolates
from Japan. The complete sequence identities of all 29
isolates of LBVaV and five of TStV ranged from 93.6 to
99.7% (nucleotides) and 94.3 to 100% (amino acids).
These values for both nucleotide and amino acid sequence
identity were the same when comparing the 29 LBVaV
isolates alone. The sequence identities of the five TStV
isolates alone were 95.7–99.7% (nucleotides) and 97.2–
100% (amino acids). LBVaV isolate UK2 was most
divergent with 93.6% identities to Japanese LBVaV Hy
and TStV Na isolates. These sequence identities resemble
those reported previously [9, 12]. Thus, our data showing
that the CP of LBVaV and TStV is highly conserved
confirmed data from previous work with fewer isolates and
supported TStV being a strain of LBVaV [12].
When the complete CP nucleotide sequences (29 of
LBVaV and five of TStV) were subjected to phylogenetic
analysis, all were within two clades (I and II), II being
resolved into three distinct sub-clades (IIa–c) (Fig. 1a). All
clades and sub-clades were geographically distinct as
regards the origins of the isolates within them. Clade I
contained 12 lettuce and six sowthistle isolates of LBVaV,
all from Europe. Within clade II, sub-clade IIa only con-
tained the seven new Australian LBVaV isolates, IIb the
four LBVaV isolates, and IIc the five Japanese TStV iso-
lates. Sub-clades IIb and IIc were closer to each other than
to sub-clade IIa containing Australian isolates. The
groupings obtained not only agreed with the three group-
ings of Sasaya et al. [12], (a) five TStV isolates, (b) four
Japanese LBVaV isolates and (c) four Spanish LBVaV
isolates, but also added 16 extra European isolates to (c)
and revealed a new grouping containing only Australian
isolates. Our findings also agree with those of Navarro
et al. [9] in which, apart from one Japanese LBVaV isolate,
Comparison of the coat protein genes
123

the only complete CP sequences examined were 12 from
Europe. Moreover, in our study, inclusion together in one
analysis of the complete CP sequences of seven additional
isolates with those of all other 27 isolates now available on
GenBank allowed greater resolution revealing presence of
two clades, and three sub-clades containing isolates from
geographically distant regions (Europe, Japan and
Australia). In addition, although lettuce isolates were
present in both clades and two of the three sub-clades,
sowthistle isolates (SON1-5) were all together in one clade
(I), while tobacco isolates were isolated within a sub-clade
of their own (IIc). When the eight partial CP sequences
were added to the 34 complete sequences and aligned, the
single sequences of Brazilian isolate 106 and those from
Australia and the Netherlands all grouped with the Aus-
tralian isolates within sub-clade IIa (Fig. 1b). The other
three Brazilian sequences and the UK and USA sequences
were in clade I, the latter being most closely related to
UK2, but the former to Spanish isolates SON1 and GAL1.
None grouped with sub-clades IIb or IIc.
Assuming that LBVaV evolved with lettuce in its centre
of domestication in the Mediterranean region, greatest
genetic diversity would be expected there. This was
reflected in the results with the 17 Spanish isolates.
Occurrence of all Australian sequences within the same
sub-clade (IIa) suggests only one introduction of LBVaV
SON5
SON4
SON3
SON6
SON2
GRA1
ALM1
ALM3
MUR1
ALM4
ALM7
ALM6
ALM2
ALM5
UK2
MUR2
SON1
GAL1
I
AUSE4
AUSE5
AUSA5
AUSB1
AUSB2
AUSG1
AUSE6
IIa
Wa
Hy
LBVaV CP
A
IIb
Na
Kan
No88
No89
No2
IIc
II87
50
100
93
81
55
100
100
99
99
94
98
85
62
96
86
96
74
62
64
66
98
63
65
50
52
0.005
SYNV
0.1
ASYNV
0.2
AUSE4
AUSE5
AUSA5
AUSB1
AUS
NL
106
AUSB2
AUSG1
AUSE6
IIa
Wa
Hy
A
LBVaV CP
IIb
Na
KanNo88
No89
No2
IIc
II
UK2
UK
USA
ALM2
ALM5
ALM4
ALM6
MUR1
ALM7
ALM3
ALM1
GRA1
SON6
SON2
SON3
SON5
SON4
105
107
SON1
104
GAL1
MUR2
85
49
100
65
46
54
100
100
73
94
98
97
86
51
54
56
83
92
73
69
99
78
84
83
77
64
69
68
44
70
69
93
47
29
62
45
0.005
I
B
Fig. 1 Phylogenetic relationships among complete coat protein (CP)
nucleotide sequences of 29 Lettuce big-vein associated virus(LBVaV) isolates from Europe, Australia and Japan, and five tobacco
stunt virus (TStV) isolates from Japan (a), and these same sequences
with eight partial LBVaV CP sequences included (b). Trees generated
with MEGA 4.1 using default parameters. Tree branches were
bootstrapped with 1,000 replications. Numbers at nodes indicate
bootstrap scores[50% (a) or all scores (b). The scale bar represents a
genetic distance of 0.005 for horizontal branch lengths. For isolate
designations, see Table 1. Inset shows isolates rooted with sonchus
yellow net virus (SYNV), accession number M17210
L. D. Maccarone et al.
123

into Australia since European settlement approximately
200 years ago. Presence of a partial CP sequence from the
Netherlands in this sub-clade might reflect its introduction
from Europe. Occurrence of only Japanese LBVaV
sequences in sub-clade IIb and Japanese TStV sequences in
sub-clade IIc suggests two introductions to Japan, but does
not indicate their origin. Cultivated tobacco is from the
Americas but TStV has not been found there as yet. Further
complete CP sequences of LBVaV from North America,
South America, Asia (outside Japan) and Europe (outside
Spain) would help identify the geographical origins of
isolates found in different parts of the world. Identification
and sequencing of TStV isolates from countries other than
Japan would do the same for the tobacco form of LBVaV.
Further sequencing might also permit resolution of clade I
into 2–3 sub-clades (Fig. 1b).
When the 34 complete amino acid sequences were
compared, the central region of the CP gene was most
divergent with few amino acid substitutions towards the N
and C terminus regions (Table 2). When the partial
sequences were included, there were 79 amino acid sub-
stitutions across the entire CP of 397 nucleotides, the
partial sequences contributing an additional four. Forty-
six of these substitutions occurred between amino acid
positions 200 and 300. Among the isolates in clade I,
there were 55 amino acid substitutions over the entire CP
Table 2 Variable amino acid
positions in the coat protein
(CP) encoding region of 37
Lettuce big-vein associatedvirus (LBaV) isolates and five
tobacco stunt virus isolates
1111111112 2222222222 2222222222 2222222222 2222233333 33333 1566899 0112267890 0000001111 1112222233 3444456667 7788956678 99999 7890379379 2066935391 2357890123 4790378956 7358904690 6734927999 12345 ALM2 KMLEVRIDKT VADVIQPTYG TLRLYALGDV TRTEFLANIP QLITTATMKK LSDNFKNMED GETPVSON5 .......... .......... ....FR..N. Q.IK...... .......... .......... ..... SON4 .......... .......... I...LRF... .....FP... .......... .......... ..... SON3 .......... .....P.... ...F...... .....F.... .......... .......... ..... SON6 .......... ........F. ..G....E.. ..I.L..... .......... .......... ..... SON2 .......... .......... A......E.. .......... .......... .......... ..... GRA1 .......... .......... ......V... ..S....S.. .......... .......... ..... ALM1 .......... .......... A......... .......... .......... .......... ..... ALM3 .......... .......... .......... .......... .......... .......... .....MUR1 .......... .......... .......... .......... ..L....... .......... ..... ALM4 .......... .......... .......... .......... .......... .......... .Q... ALM7 .......... .......... .......... .......... ......A... .......... ..... ALM6 .......... .......... .......... .G........ .......... .......... ..... ALM5 .......... ......S..R .......... .......... .......... .......... A.... UK2 .I........ L.NA...... .......... .......... .......... .........N ..... UK -----..... ...A...... .......... .......... ....A..... .....----- -----USA -----..... ...A...... .......... .......... .......... .....----- ----- MUR2 .......... .......... ......F.NG A..G...... ..SA.G.VIE RNV.L.S..N ..... 104 -----.M... ...A...... .......... A......... .......... .....----- ----- 105 -----GM... ...A...... .......... A......... .......... .....----- ----- 107 -----....A ...A...... .......... A......... .......... .....----- -----SON1 R......... ...A...... .......... A......... ...S...... V........N ..... GAL1 R......... ...A...... .......... A......... .......... .........N ..... AUSE4 .......... .D.A...... .......... A......STS E......... ........GN ..... AUSE5 .......... .D.A...... .......... A......STS E......... .........N ..... AUSA5 .......... .D.A...... .......... A......S.. .......... .........N ..... AUSB1 .........A .V.A...... .......... A......... .......... .........N ....E AUS -----..... .D.A...... .......... A......... .......... .....----- ----- NL -----..G.. .D.A...... .......... A......... .......... .....----- ----- 106 -----GM... .D.A...... .......... A......... .......... .....----- ----- AUSB2 .......... .D.A...... .......... A......... .......... .........N ..... AUSG1 ..P......A .V.A...... .......... A......S.. .P........ .........N ..... AUSE6 ...D.....A .V.A...... .......... A......S.. .......... .........N ..... Wa ...D...... .P.A...N.. .......... A......... .......... .........N ..... Hy ...D...... .P.A...N.. .......... A......... .......... .........N ...S. A ...D...... .P.A...N.. .......... .......... .......... .........N ..... LBVaV-CP...D...... .P.A...N.. .......... A......... .......... .........N ..... Na ...DA...R. ...A...A.. .F........ A......... .......... ...T.....N ..A.. Kan ...DA...R. ...A...A.. .......... A......... .......... .........N ..A.. No88 ...DA...R. ...A...A.. .......... A......... .......... .....E.V.N ..A.. No89 ...DA...R. ...A...A.. .......... A......... .......... .........N ..A.. No2 ...DA...R. ...AT..A.. .......... A......... .......... .........N ..A..
Amino acid substitution sites *
I
IIa
IIb
IIc
* Numbers vertically positioned above the sequence alignment indicate positions of the LBVaV CP and TStV CP amino acid residues. The consensus sequence is shown by isolate ALM2 and amino acid residues that are different from the consensus are indicated, whereas amino acid residues that are identical to the consensus aremarked with dots. Partial sequences for which there is no amino acid sequence information available are markedwith dashes.
Comparison of the coat protein genes
123

gene sequence and 48 of them were unique. There were
39 substitutions between positions 200 and 300. There
were 31 amino acid substitutions in clade II which were
relatively evenly spread throughout the CP gene except
for a cluster at the C terminus. The Australian LBVaV
isolates in sub-clade IIa showed 13 amino acid substitu-
tions, 10 of which were found only in sub-clade IIa; 6
substitutions were in a cluster between positions 214 and
243. The Japanese LBVaV isolates in sub-clade IIb
showed seven amino acid substitutions, those at positions
110, 183 and 394 being unique. In sub-clade IIc, there
were 13 amino acid substitutions 9 of which were unique,
those at positions 53, 97, 183 and 393 being present in all
isolates.
Navarro et al. [9] reported a mean Dn/Ds ratio of 0.132
for LBVaV CP gene sequences. In our study, a mean Dn/
Ds ratio of 0.187 was obtained for all 34 complete CP
sequences indicating that the majority of the nucleotide
substitutions were silent. The mean Dn/Ds ratios for clades
I and II were 0.283 and 0.124, respectively, while those for
sub-clades IIa, IIb and IIc were 0.274, 0.060 and 0.156,
respectively. Mean Dn/Ds ratios \1 suggest selection for
amino acid conservation. Genome conservation is expected
as genetic stability is commonly seen in natural plant virus
populations so the low mean Dn/Ds ratios in clades I and II
reflect selection against nucleotide changes causing amino
acid substitutions, especially in sub-clades IIb and IIc.
There were 74 amino acid substitutions in the complete CP
sequences, 52 were in clade I with 47 of them being
unique. This greater level of amino acid substitution could
also explain the higher Dn/Ds ratio for clade I. Similarly,
the greater Dn/Ds ratio for sub-clade IIa might be
explained by 10 of the 13 amino acid substitutions in the
complete CP sequences being unique to this sub-clade.
Only three unique amino acid substitutions were found in
sub-clade IIb which had the lowest Dn/Ds ratio. The CPs of
vector-borne plant viruses are subject to greater stabilising
selection than those of viruses transmitted otherwise
because interaction between the CP and cellular receptors
on vectors places greater selective constraint on the amino
acid sequence than host-virus interactions. For example, in
Melon necrotic spot virus vectored by Olpidium bornov-
anus just one amino acid substitution from Ile to Phe at
position 300 resulted in loss of specific binding and fungal
transmission as the CP acts as a ligand to the vector zoo-
spore [6]. A similar scenario might also occur within
LBVaV, explaining its high degree of CP sequence
conservation.
Acknowledgments We thank Stephen Wylie, Craig Webster and
Denis Phillips for advice, the SABC for laboratory facilities and local
farmers for allowing sampling. Linda Maccarone received an
Australian Postgraduate Award from the Australian Research
Council.
References
1. Brunt AA, Milne RG, Sasaya T, Verbeek M, Vetten HJ, Walsh
JA (2005) Varicosavirus. In: Fauquet CM, Mayo MA, Maniloff J,
Desselberger U, Ball LA (eds) Virus taxonomy. Eighth Report of
the International Committee on Taxonomy of Viruses. Elsevier/
Academic Press, London, pp 669–671
2. Hayes RJ, Wintermantel WM, Nicely PA, Ryder EJ (2006) Host
resistance to Mirafiori lettuce big-vein virus and Lettuce big-veinassociated virus and virus sequence diversity and frequency in
California. Plant Dis 90:233–239
3. Kosakovsky-Pond SL, Frost SDW (2005) Datamonkey: rapid
detection of selective pressure on individual sites of codon
alignments. Bioinformatics 21:2531–2533
4. Kuwata S, Kubo S (1986) Tobacco stunt virus, Descriptions of
plant viruses, no. 313. Association of Applied Biologists,
Wellesbourne, UK, 4 p
5. Lot H, Campbell RN, Souche S, Milne RG, Roggero P (2002)
Transmission by Olpidium brassicae of Mirafiori lettuce virusand Lettuce big-vein virus, and their roles in lettuce big-vein
etiology. Phytopathology 92:288–293
6. Mochizuki T, Ohnishi J, Ohki T, Kanda A, Tsuda S (2008)
Amino acid substitution in the coat protein of Melon necrotic spotvirus causes loss of binding to the surface of Olpidium bornov-anus zoospores. J Gen Plant Pathol 74:176–181
7. Navarro JA, Botella F, Maruhenda A, Sastre P, Sanchez-Pina
MA, Pallas V (2004) Comparative infection progress analysis of
Lettuce big-vein virus and Mirafiori lettuce virus in lettuce crops
by developed molecular diagnosis techniques. Phytopathology
94:470–477
8. Navarro JA, Botella F, Marhuenda A, Sastre P, Sanchez-Pina
MA, Pallas V (2005) Identification and partial characterisation of
Lettuce big-vein associated virus and Mirafiori lettuce big-veinvirus in common weeds found amongst Spanish lettuce crops and
their role in lettuce big-vein disease transmission. Eur J Plant
Pathol 113:25–34
9. Navarro JA, Torok VA, Vetten HJ, Pallas V (2005) Genetic vari-
ability in the coat protein genes of Lettuce big-vein associated virusand Mirafiori lettuce big-vein virus. Arch Virol 150:681–694
10. Sambrook J, Russell DW (2001) Molecular cloning: a laboratory
manual. Cold Spring Harbour Laboratory Press, New York
11. Sasaya T, Ishikawa K, Koganezawa H (2001) Nucleotide
sequence of the coat protein gene of Lettuce big-vein virus. J Gen
Virol 82:1509–1515
12. Sasaya T, Ishikawa K, Kuwata S, Koganezawa H (2005)
Molecular analysis of coat protein coding region of tobacco stunt
virus shows that it is a strain of Lettuce big-vein virus in the genus
Varicosavirus. Arch Virol 150:1013–1021
13. Sasaya T, Koganezawa H (2006) Molecular analysis and virus
transmission tests place Olpidium virulentus, a vector of Mirafiorilettuce big-vein virus and Tobacco stunt virus, as a distinct spe-
cies rather than a strain of Olpidium brassicae. J Gen Plant Pathol
72:20–25
14. Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4: Molec-
ular Evolutionary Genetics Analysis (MEGA) software version
4.0. Mol Biol Evol 24:1596–1599
15. Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W:
improving the sensitivity of progressive multiple sequence
alignment through sequence weighting, position-specific gap
penalties and weight matrix choice. Nucleic Acids Res 22:4673–
4680
16. Vetten HJ, Lesemann DE, Dalchow J (1987) Electron micro-
scopical and serological detection of virus-like particles associ-
ated with lettuce big vein disease. J Phytopathol 120:53–59
L. D. Maccarone et al.
123