modulation der gabaergen wirkung durch
TRANSCRIPT

Modulation der GABAergen Wirkung durch
Lebensmittelinhaltsstoffe
Der Naturwissenschaftlichen Fakultät
der Friedrich-Alexander-Universität Erlangen-Nürnberg
zur Erlangung des Doktorgrades
Dr. rer. nat.
vorgelegt von
Sümeyye Sahin
aus Giresun/Türkei

Als Dissertation genehmigt von der
Naturwissenschaften Fakultät der
Friedrich-Alexander-Universität Erlangen-Nürnberg
Tag der mündlichen Prüfung: 17.03.2016
Vorsitzender des Promotionsorgans: Prof. Dr. Jörn Wilms
Gutachterin: Prof. Dr. Monika Pischetsrieder
Prof. Dr. Carmen Villmann


Teile dieser Arbeit wurden bereits veröffentlicht:
Publikation
Sahin S., Eulenburg V., Villmann C.,Pischetsrieder M., Three-Step Test System for the
Identification of Novel GABAA Receptor Modulating Plant Foods (2016)-eingereicht
Tagungsbeitrag
Sahin S., Villmann C., Pischetsrieder, M., Modulation of Synaptic GABAA-Receptor
Activity by Food Extracts
7th International Conference and Exhibition on Nutraceuticals and Functional Foods,
14.-17. Oktober 2014 in Istanbul
Posterbeiträge
Sahin S., Villmann C., Pischetsrieder, M., Modulation of GABAergic Effect by Plant
Extracts
EUROFOODCHEM XVII, 7.-10. Mai 2013 in Istanbul
Sahin S., Villmann C., Pischetsrieder, M., Determination of γ-amino butyric acid in
several food extracts and investigation of its physiological effect
7th International Conference and Exhibition on Nutraceuticals and Functional Foods,
14.-17. Oktober 2014 in Istanbul

Danksagung
Allen voran möchte ich mich bei meiner Doktormutter Frau Prof. Dr. Monika Pischetsrieder für
die Aufnahme in ihre Arbeitsgruppe, die Überlassung des sehr interessanten Themas, die
freundliche und engagierte Unterstützung in den Jahren der Promotion und das mir entgegen
gebrachte Vertrauen sehr herzlich bedanken. Weiterhin bedanke ich mich besonders beim
Bildungsministerium in der Regierung der Republik Türkei für die finanzielle Unterstützung
während des Aufenthaltes in Deutschland. Ebenso danke ich sehr herzlich Frau Prof. Dr. Carmen
Villmann für die fachliche Betreuung und wertvollen Anregungen zu dieser Arbeit im Bereich der
elektrophysiologischen Untersuchungen sowie für die Erstellung des Zweigutachtens.
Des Weiteren möchte ich mich bei Frau Prof. Dr. Andrea Büttner für die Übernahme des
Prüfungsvorsitzes sowie bei Herrn Prof. Dr. Christian Alzheimer für die freundliche Bereitschaft
bedanken, sich als Zweitprüfer zur Verfügung zu stellen.
Mein Dank gilt weiterhin Herrn Dr. Volker Eulenberg für die Möglichkeit, die Voltage-Clamp
Messungen in seiner Arbeitsgruppe durchzuführen. Herrn Prof. Dr. med. Christoph Korbmacher
danke ich für die Bereitstellung der Xenopus Oocyten. Mein weiterer Dank richtet sich an die
Mitarbeiter des Institüts für Biochemie der Medizinischen Fakültät. Besonders danke ich Dr.
Christoph Kluck, Marina Wenzel und Ingmar Herz für ihre Unterstützung.
Mein herzlicher Dank gilt Frau Christiene Meißner für die stets freundliche Hilfsbereitschaft. Für
die Hilfe bei technischen Fragen möchte ich Alfred Zillich-Balthasar und Heribert Dittrich
danken.
Außerdem danke ich den Mitarbeitern des Arbeitskreises Büttner für die freundliche
Unterstützung. Ganz besonders bedanke ich mich bei Artur für seine Hilfe und Unterstützung in
allen Fragen zum Thema Elektrophysiologie. Weiterhin möchte ich Anja und Johannes für ihre
Hilfestellungen beim Messen an der GC-Massenspektrometrie danken.
Für sehr angenehmes Arbeitsklima, die Hilfsbereitschaft und die entstandenen Freundschaften
möchte ich mich bei meinen lieben jetzigen und ehemaligen Kollegen bedanken. Vielen Dank
dafür an Artur, Christiane, Isabel, Ingrid, Jasmin, Jennifer, Johannes, Judith, Júnia, Kerstin,
Leonie, Li, Lisa, Liu, Pelin, Sabrina, Sevim, Steffi, Sven, Tobias, Ulla, Waldemer, Wolfgang und die
Kollegen des AK Eichler und AK Heinrich. Mein besonderer Dank geht dabei an Lisa, Christiane
und Isabel für das Korrekturlesen. Mein großer Dank richtet sich auch an meine Freunde für ihre
Unterstützung.
Mein allergrößter Dank gilt meiner gesamten Familie insbesondere meinen Eltern und
Geschwistern, ohne deren Liebe und Unterstützung mein Studium und diese Arbeit nicht
möglich gewesen wären.


i
Inhaltsverzeichnis
Inhaltsverzeichnis .................................................................................................................................... i
Abkürzungsverzeichnis .......................................................................................................................... vi
Abbildungsverzeichnis ........................................................................................................................ viii
Tabellenverzeichnis ............................................................................................................................... xi
1 Einleitung .............................................................................................................................. 1
1.1. Neurotransmission ..................................................................................................................... 2
1.2. Ionenkanäle ................................................................................................................................ 3
1.3. Das GABAerge System ................................................................................................................ 5
1.3.1. GABA Rezeptoren ................................................................................................................... 7
1.3.1.1. GABAA-Rezeptoren .............................................................................................................. 7
1.3.1.2. GABAB- und GABAA-Rezeptoren .......................................................................................... 9
1.3.2. Funktionen des GABAergen Neurotransmittersystem ........................................................... 9
1.3.3. Modulation des GABAergen Neurotransmittersystems ....................................................... 10
1.4. Glycin-Rezeptoren .................................................................................................................... 12
1.5. Die Voltage-Clamp-Technik ...................................................................................................... 12
1.6. Test-Lebensmittel ..................................................................................................................... 14
1.6.1. Lavendel ................................................................................................................................ 14
1.6.2. Salbei ..................................................................................................................................... 14
1.6.3. Melisse (Zitronenmelisse) ..................................................................................................... 15
1.6.4. Sideritis ................................................................................................................................. 15
1.6.5. Grüner Tee ............................................................................................................................ 16
1.6.6. Schwarzer Pfeffer.................................................................................................................. 17
1.6.7. Kamille .................................................................................................................................. 17
1.6.8. Nelken ................................................................................................................................... 18

ii
1.6.9. Hopfen .................................................................................................................................. 19
1.6.10. Kakao .................................................................................................................................... 20
1.7. Ziele der Arbeit ......................................................................................................................... 20
2 Inhibition des GABA-Abbaus .............................................................................................. 23
2.1. Einleitung .................................................................................................................................. 23
2.2. Bestimmung der Aktivität von GABA-metabolisierenden Enzyme (GABA-AT und SSA-DH) .... 25
2.3. Einfluss von Vigabatrin auf die Aktivität der GABA-metabolisierenden Enzyme (GABA-AT
und SSA-DH) ......................................................................................................................................... 26
2.4. Einfluss der Lebensmittelextrakte auf die Aktivität der GABA-metabolisierenden Enzyme
(GABA-AT und SSA-DH) ........................................................................................................................ 28
2.5. Einfluss ausgewählter Lebensmittelinhaltsstoffe auf die Aktivität der GABA-
metabolisierenden Enzyme (GABA-AT und SSA-DH) ........................................................................... 31
2.6. Diskussion und Zusammenfassung ........................................................................................... 35
3 Modulation des GABAA-Rezeptors ...................................................................................... 39
3.1. Einleitung .................................................................................................................................. 39
3.2. Modulation der GABAA-Rezeptoren durch Lebensmittelextrakte ........................................... 40
3.3. Bestimmung der GABA Gehalte der Extrakte und Untersuchungen zu deren Wirkung auf
den GABAA Rezeptor ............................................................................................................................ 48
3.3.1. Quantifizierung von GABA in den Lebensmittelextrakten ................................................... 48
3.3.2. Direkte GABAA-Rezeptoraktivierung durch die untersuchten Lebensmittelextrakte .......... 51
3.4. Wirkung der Lebensmittelextrakte auf den Glycin-Rezeptor .................................................. 53
3.5. Diskussion und Zusammenfassung ........................................................................................... 57
4 Identifizierung der aktiven Inhaltsstoffe des wässrigen Nelkenextraktes .......................... 63
4.1. Einleitung .................................................................................................................................. 63
4.2. Aktivitätsgeleitete Fraktionierung des Nelkenextraktes .......................................................... 64
4.3. Identifizierung der neurophysiologisch aktiven Inhaltsstoffe von Nelken ............................... 66
4.3.1. Identifizierung und Quantifizierung von Eugenol im Nelkenextrakt und in der aktivsten
Nelken-Fraktion 11 ............................................................................................................................... 66

iii
4.3.2. Untersuchung der GABAA-Rezeptor modulierenden Wirkung von Eugenol ........................ 71
4.3.3. Identifizierung von anderen Inhaltsstoffe im Nelkenextrakt und in der aktivsten
Nelken-Fraktion 11 sowie Untersuchung zu deren GABAA-Rezeptor modulierenden Wirkung ......... 73
4.4. Diskussion und Zusammenfassung ........................................................................................... 75
5 Material und Methoden ....................................................................................................... 81
5.1. Geräte ....................................................................................................................................... 81
5.2. Verbrauchsmaterialien ............................................................................................................. 83
5.3. Chemikalien .............................................................................................................................. 84
5.4. Herkunft der Lebensmitteln und Herstellung der Lebensmittelextrakte................................. 87
5.4.1. Herkunft der Lebensmitteln ................................................................................................. 87
5.4.2. Herstellung der Lebensmittelextrakte .................................................................................. 87
5.4.2.1. Extraktionsmethode I ........................................................................................................ 87
5.4.2.2. Extraktionsmethode II ....................................................................................................... 88
5.5. Messung der Enzymaktivität mittels Enzymassay .................................................................... 88
5.5.1. Lösungen und Puffer ............................................................................................................. 88
5.5.2. Bestimmung der Enzymaktivität ........................................................................................... 89
5.5.3. Auswertung der Enzymaktivität ........................................................................................... 89
5.6. TEVC .......................................................................................................................................... 90
5.6.1. Lösungen, Puffer, Nährmedien und kommerziell erhältliche Kits ........................................ 90
5.6.2. Das Xenopus-Oocyten-Expressionssystem ........................................................................... 92
5.6.2.1. Herstellung der RNA der α1- und β2- Untereinheiten des humanen GABAA-Rezeptors . 92
5.6.2.1.1. Isolation der Plasmid DNA-Maxipräparation .................................................................... 92
5.6.2.1.2. Linearisierung der DNA ..................................................................................................... 93
5.6.2.1.3. Konzentrationsbestimmung der DNA/RNA ...................................................................... 94
5.6.2.1.4. Agarose-Gelelektrophorese .............................................................................................. 94
5.6.2.1.5. In vitro Transkription......................................................................................................... 94
5.6.2.2. Herstellung der RNA der α1- Untereinheit des humanen Glycin-Rezeptors .................... 95

iv
5.6.2.3. Isolation der Xenopus-Oocyten und Mikroinjektion der cRNA in Oocyten ..................... 95
5.6.3. TEVC-Technik ........................................................................................................................ 96
5.6.3.1. Versuchsdurchführung...................................................................................................... 96
5.6.3.2. Versuchsprotokoll für die Applikation der Liganden und Normalisierung der Ströme .... 97
5.6.3.3. Versuchsprotokoll für die Koapplikation der GABA mit den Lebensmittelextrakten
und Berechnung des relativen Stroms ................................................................................................. 98
5.6.3.4. Versuchsprotokoll für die Messung der direkten Rezeptoraktivierung durch
Lebensmittelextrakten und Berechnung des relativen Stroms ........................................................... 99
5.6.3.5. Versuchsprotokoll für die Koapplikation von Glycin mit den Lebensmittelextrakten
und Berechnung des relativen Stroms ................................................................................................. 99
5.6.3.6. Versuchsprotokoll für die Koapplikation der GABA mit den Fraktionen des wässrigen
Nelkenextraktes und Berechnung des relativen Stroms ................................................................... 100
5.6.3.7. Versuchsprotokoll für die Koapplikation der GABA mit den Reinsubstanzen Eugenol
und Acetyleugenol und Berechnung des relativen Stroms ............................................................... 101
5.7. Bestimmung der GABA mittels HPLC ...................................................................................... 103
5.7.1. Lösungen und Eluenten ...................................................................................................... 103
5.7.2. Derivatisierungsmethoden ................................................................................................. 103
5.7.2.1. 2-Hydroxynaphtaldehyd Derivatisierung ........................................................................ 103
5.7.2.2. Orthophtalaldehyd (OPA) Derivatisierung ...................................................................... 104
5.7.3. Analyse der GABA ............................................................................................................... 104
5.7.3.1. RP-HPLC-UV Parameter .................................................................................................. 104
5.7.3.2. RP-HPLC-FLD Parameter ................................................................................................. 105
5.7.3.3. Identifizierung und Quantifizierung der in den Lebensmitteln enthaltenen GABA ....... 105
5.7.3.3.1. Kalibrierung und Linearität ............................................................................................. 106
5.7.3.3.2. Nachweis- und Bestimmungsgrenze ............................................................................... 106
5.8. Fraktionierung des Extraktes mittels RP-HPLC ....................................................................... 107
5.9. Nachweis der Inhaltsstoffe von Nelken .................................................................................. 108
5.9.1. UHPLC-DAD Methode ......................................................................................................... 108

v
5.9.1.1. Herstellung der Referenzsubstanzen .............................................................................. 108
5.9.1.2. UHPLC-DAD Parameter ................................................................................................... 109
5.9.1.3. Kalibrierung und Linearität zur Quantifizierung des Eugenols im Nelkenextrakt bzw.
in seiner Fraktion 11 .......................................................................................................................... 109
5.9.1.4. Nachweis- und Bestimmungsgrenze ............................................................................... 110
5.9.2. GC-MS Methode ................................................................................................................. 110
5.10. Statistische Auswertung ..................................................................................................... 111
6 Zusammenfassung ........................................................................................................ 112
7 Abstract ........................................................................................................................ 117
A Anhang ............................................................................................................................................ 122
Literaturverzeichnis ........................................................................................................................... 125

vi
Abkürzungsverzeichnis
µ mikro
A Ampere
°C Grad Celsius
ANOVA Varinanzanalyse, aus dem Englischen: analysis of variances
AU Absorptionseinheiten
BSG Bestimmungsgrenze
bzw. Beziehungsweise
C Kohlenstoffatom
ca. circa
CI Konfidenzintervall, aus dem Englischen: confidence interval
Da Dalton
DAD Diodenarraydetektion
DCM Dichlormethan
DMSO Dimethylsulfoxid
DNA Desoxyribonukleinsäure
EC50 mittlere effektive Konzentration
EDTA Ethylendiamintetraessigsäure
et al. und andere
FLD Fluoreszenzdetektor
g gramm
GABA γ-Aminobuttersäure
GABA-AT GABA-Transaminase
GAD Glutamat-Decarboxylase
GC-MS Gaschromatographie mit Massenspektrometrie
H Wasserstoff
HEPES 2-(4-(2-Hydroxyethyl)-1-piperazinyl)-ethansulfonsäure
2-HN 2-Hydroxynaphthaldehyde
HPLC Hochleistungsflüssigchromatographie
IC50 mittlere Hemmung Konzentration
L Liter
M Molar
m milli
min Minuten
N Stickstoff
NAD+ Nicotinsäureamid-Adenin-Dinucleotid
NADH reduzierte Form von Nicotinsäureamid-Adenin-Dinucleotid

vii
NADP+ Nicotinsäureamid-Adenin-Dinucleotid-Phosphat
NADPH reduzierte Form von Nicotinsäureamid-Adenin-Dinucleotid-Phosphat
nd nicht detektierbar
nm Nanometer
ns nicht signifikant
NWG Nachweisgrenze
O Sauerstoff
OPA ortho-Phtalaldehyd
p Signifikanzniveau
RKI Robert Koch-Institut
RNA Ribonukleinsäure
RP Reversedphase
s Sekunden
S. arguta Sideritis arguta
S. condensata Sideritis condensata
SD Standardabweichung
SEM aus dem Englischen: standard error of the mean
SSA Succinatsemialdehyd
SSA-DH Succinatsemialdehyd-Dehydrogenase
SSA-R Succinatsemialdehyd-Reduktase
S. sipylea Sideritis sipylea
S. stricta Sideritis stricta
TEVC aus dem Englischen: Two electrode voltage clamp
UHPLC Ultrahochleistungsflüssigchromatographie
2xYT Medium 2- Hefeextrakt und Trypton
ZNS Zentralnervensystem

viii
Abbildungsverzeichnis
1.1 Schematische Darstellung der vier Transmembrandomänen der Cys-Loop Rezeptoren ............ 5
1.2 Schematische Abbildung des GABAergen System ........................................................................ 6
1.3 Schematische Darstellung der vier Transmembrandomänen der GABAA-Rezeptor- Untereinheit
und der aus α-, β-, und γ-Untereinheiten bestehende GABAA-Rezeptor. .................................... 7
1.4 Schematische Darstellung der allosterischen Modulation des GABAA-Rezeptors ..................... 11
1.5 Schematische Darstellung einer Voltage–Clamp Technik. ......................................................... 13
2.1 Schematische Überblick über Biosynthese und Abbau der Neurotransmitter GABA ................ 24
2.2 Konzentrationsabhängige Wirkung von Vigabatrin auf die Aktivität der GABA-
metabolisierenden Enzyme ........................................................................................................ 27
2.3 Einfluss unterschiedlicher Konzentrationen an schwarzer Pfeffer, Hopfen und Kakao auf die
Aktivität der GABA-metabolisierenden Enzyme ......................................................................... 28
2.4 Aktivität der GABA-metabolisierenden Enzyme in Abhängigkeit von der Konzentration der
Extrakte von (A) Nelken und verschiedenen Sideritis-Arten (S. condensata, S. arguta, S. stricta
und S. sipylea) (B) Grünem Tee und Salbei (C) Kamille (D) Lavendel und (E) Melisse. ............... 29
2.5 Strukturen von Vigabatrin, β-Damascenon, β-Ionon, (-)-Caryophyllenoxid und Myrcen. ......... 34
3.1 Wirkung von GABA auf den humanen α1β2 GABAA-Rezeptor in Xenopus leavis Oocyten. A)
Repräsentative Stromspuren für die direkte Rezeptoraktivierung durch den Liganden GABA.
GABA wurde in unterschiedlichen Konzentrationen zwischen 0,3 und 300 µM appliziert. B)
Dosis-Wirkungskurve für GABA .................................................................................................. 41
3.2 Die durch wässrigen Extrakt von S. condensata bei Abwesenheit und Anwesenheit von GABA
ausgelösten unspezifische Ströme bei der Negativkontrolle in nicht-transfizierten
Oocyten....................................................................................................................................... 42
3.3 Beispielstromkurven einer Messung für die Koapplikation von 1 µM GABA mit wässrigem
Lebensmittelextrakt an humanen α1β2 GABAA-Rezeptoren ..................................................... 43

ix
3.4 Potenzierende Effekte von wässrigen Lebensmittelextrakten in der Konzentration von 5 µg/mL
auf die durch 1 µM GABA induzierte Stromantworte des humanen α1β2 GABAA-
Rezeptors .................................................................................................................................... 47
3.5 RP-HPLC-FLD-Chromatogramme der 0,1 mg/mL GABA-Standardlösung und eines wässrigen
Kakao-Extraktes nach Derivatisierung mit OPA .......................................................................... 49
3.6 Beispielstromkurven einer Messung der direkten Aktivierung des humanen α1β2 GABAA-
Rezeptor durch wässrige Lebensmittelextrakte ......................................................................... 52
3.7 Wirkung von Glycin auf den humanen α1 Glycin-Rezeptor an Xenopus leavis Oocyten. A)
Repräsentative Stromspuren für die direkte Rezeptoraktivierung durch den Liganden Glycin,
der in unterschiedlichen Konzentrationen zwischen 10 und 3000 µM appliziert wurde. B) Die
Dosis-Wirkungskurve für den Agonisten Glycin. ........................................................................ 54
3.8 Exemplarische Stromkurven einer Messung der Modulation des humanen α1 Glycin-Rezeptors
durch wässrige Lebensmittelextrakte. ....................................................................................... 55
3.9 Effekte von wässrigen Lebensmittelextrakten in einer Konzentration von 5 µg/mL auf die
durch 30 µM Glycin induzierte Stromantwort des humanen α1-Glycin-Rezeptors ................... 56
4.1 RP-HPLC Chromatogramm des wässrigen Nelkenextrakt detektiert bei 254 nm ...................... 65
4.2 Modulation von humanen α1β2 GABAA-Rezeptoren durch Fraktionen des wässrigen
Nelkenextraktes in der Konzentration von 5 μg/mL .................................................................. 65
4.3 Repräsentative Stromkurven einer Messung der Koapplikation von 1 μM GABA mit Fraktion 11
des wässrigen Nelkenextraktes an humanen α1β2 GABAA-Rezeptoren. ................................... 66
4.4 UHPLC-DAD-Chromatogramm der Fraktion 11 von Nelken (A) und des wässrigen
Nelkenextraktes (B) .................................................................................................................... 67
4.5 Übereinandergelegte UHPLC-DAD-Chromatogramme der Eugenol-Standardlösung und der
Nelken-Fraktion 11. .................................................................................................................... 68
4.6 UV/Vis-Spektren der Eugenol-Standardlösung (A), der Nelken-Fraktion 11 (B) und des
Nelkenextraktes (C) bei einer Retentionszeit von 6,9 min. ........................................................ 68

x
4.7 Übereinandergelegte UHPLC-DAD-Chromatogramme der Eugenol-Standardlösung, der
Acetyleugenol-Standardlösung und des wässrigen Nelken-Extraktes ....................................... 69
4.8 TIC -Chromatogramm des wässrigen NelkenExtraktes detektiert mittels GC-MS (A).
Nebeneinanderliegende Massenspektren des bei 26,3 min eluierten Signal und des Eugenols
entnommen aus der NIST-Datenbank (B). Nebeneinanderliegende Massenspektren des Signals
mit Retentionszeit von 27,3 min und des Acetyleugenols, das mittels der NIST-Datenbank
ermittelt wurde (C). ................................................................................................................... 70
4.9 Repräsentative Stromkurven einer Messung der Koapplikation von 1 µM GABA mit Eugenol,
der Nelken-Fraktion 11 und dem wässrigen Nelkenextrakt an humanen α1β2 GABAA-
Rezeptoren. ................................................................................................................................ 72
4.10 UV/Vis-Spektren der Acetyleugenol-Standardlösung (A) und des Nelkenextraktes (B) bei einer
Retentionszeit von 7,9 min. ....................................................................................................... 73
4.11 Die Potenzierung der Aktivität von humanen α1β2 GABAA-Rezeptoren in den Xenopus leavis
Oocyten durch Fraktion 11, Eugenol und Nelken (A) und sowie durch Acetyleugenol (B) bei
der simultanen Applikation mit 1 μM GABA. ............................................................................. 74
4.12 Repräsentative Stromkurven einer Messung für die Koapplikation der 1 µM GABA mit
Acetyleugenol an humanen α1β2 GABAA-Rezeptoren. .............................................................. 75
4.13 Die Strukturformeln des Eugenols, Acetyleugenols, Metyleugenols, Thymols, Carvacrols und
Propofols ..................................................................................................................................... 78

xi
Tabellenverzeichnis
1.1 Die Superfamilien der ligandengesteuerten Ionenkanäle, deren Familien und strukturelle
Eigenschaften ........................................................................................................................... 4
2.1 IC50-Werte verschiedener Lebensmittelextrakten für GABA-metabolisierende Enzyme. Die
Werte sind angegeben als mg/mL mit 95%-Konfidenzintervall (CI). ...................................... 31
2. 2 Aktivität der GABA-metabolisierenden Enzyme (GABA-AT und SSA-DH) in Anwesenheit von
1 mM Testsubstanzen ............................................................................................................. 32
2.3 Konzentrationsabhängige Wirkung von β-Damascenon, Myrcen, β-Ionon und (-)-
Caryophyllenoxid auf die GABA-metabolisierende Enzyme (GABA-AT und SSA-DH). ........... 33
2.4 IC50-Werte verschiedener Testsubstanzen für GABA-metabolisierende Enzyme (GABA-AT
und SSA-DH). Die Werte sind angegeben als mM mit 95%-Konfidenzintervall (CI). ............. 34
3.1 Modulation des humanen α1β2 GABAA-Rezeptors durch wässrige Lebensmittelextrakte ... 45
3.2 Normalisierte Konzentration von GABA in den wässrigen Lebensmittelextrakten ............... 50
3.3 Relative Chloridionenströme der direkten Aktivierung des humanen α1β2 GABAA-Rezeptor
durch wässrige Lebensmittelextrakte. .................................................................................. 53
5.1 Übersicht über die untersuchten Lebensmittel...................................................................... 87

xii

1
Einleitung 1
Psychische Gesundheit, die als ein Zustand des Wohlbefindens definiert wird [1], ist wichtig für
Lebensqualität und Leistungsfähigkeit. Störungen der psychischen Gesundheit sind weltweit
verbreitet. In den europäischen Ländern ist etwa ein Viertel bis ein Drittel der Bevölkerung nach
Angaben der Weltgesundheitsorganisation (WHO) jährlich davon betroffen [1]. Die
Zusatzuntersuchung des Bundes-Gesundheitssurveys von 1997/ 1998 (BGS98), welche die erste
repräsentative gesamtdeutsche Untersuchung zum Gesundheitszustand der erwachsenen
Bevölkerung war, berichtete, dass die 12-Monats-Prävalenz von psychischen Störungen 31 % für
Erwachsene im Alter zwischen 18 und 65 Jahren betrug [2]. Nach den im Zusatzmodul der Studie
zur Gesundheit Erwachsener in Deutschland (DEGS1-MH von 2008/2011) des Robert Koch-
Instituts (RKI) ermittelten Daten ist die häufigste psychische Störungsgruppe die Angststörung,
deren 12-Monats-Prävalenz bei 15,3% der Erwachsenen im Alter von 18 bis 79 Jahren liegt,
gefolgt von der unipolaren Depressionen (7,7 %) und der Major-Depressionen (6%) [3].
Außerdem zeigt das RKI in den bevölkerungsbezogenen GEDA (Gesundheit in Deutschland
aktuell) – Studien in den vergangenen Jahren einen Anstieg der Auftretenshäufigkeit depressiver
Erkrankungen [4-6], welche gemäß ICD 10 durch Traurigkeit, Interesselosigkeit, Schuldgefühl,
Schlafstörungen, Müdigkeit, Störungen von Appetit, Konzentrationsschwächen, Verlust des
Selbstwertgefühls und suizidales Verhalten charakterisiert sind [7]. Diese Störungen belasten
nicht nur Patienten sondern auch Gesundheitssysteme durch Behandlungskosten und
Sozialsystem durch krankheitsbedingte Fehlzeiten. In einer Studie wurden die durch Depression
verursachten Krankheitskosten in Deutschland mit 21,5 Milliarden Euro pro Jahr veranschlagt
[8].
Eine wesentliche Ursache für die Entstehung einer Depression sind neurobiologische Störungen,
die mit Veränderungen der Neurotransmitterspiegeln im Gehirn assoziert sind [9]. Zur Therapie
der depressiven Krankheiten steht eine medikamentöse Behandlung zur Verfügung, wodurch
Stoffwechselstörungen im Gehirn wieder ins Gleichgewicht gebracht werden können. Zahlreiche
Medikamente wie antidepressive, angstlösende, schlafregulierende, und sedierende
Medikamente entfalten ihre Wirkungen über das GABAerge Neurotransmittersystem. Darüber
hinaus kann auch die Ernährung einen positiven Einfluss auf affektive Störungen ausüben, z. B.
über Lebensmittelinhaltsstoffe, die das GABAerge Neurotransmittersystem modulieren.

Einleitung
2
1.1. Neurotransmission
Neuronen (Nervenzellen), die Bausteine des Zentralnervensystems sind, bestehen aus dem
Zellkörper (Soma), dem Axon und den Dendriten. Die Informationsübertragung in Nervenzellen
erfolgt über elektrische Impulse, sogenannte Aktionspotentiale. Elektrische Impulse entstehen
dabei durch Konzentrationsunterschiede von Ionen zwischen Innenseite und Außenseite der
Nervenzellmembran. Das Aktionspotential von präsynaptischen Terminalen auf Dendriten wird
über Synapsen vermittelt, über die zahlreiche Neuronen miteinander verknüpft sind. Die Axone
leiten diese elektrischen Impulse an postsynaptische Neuronen oder Muskelzellen weiter. Die
Übertragung der Aktionspotentiale erfolgt über elektrische oder chemische Synapsen. Bei
elektrischen Synapsen wird das Aktionspotential direkt zwischen den Neuronen elektrisch
weiter geleitet, während die Informationsübertragung an chemischen Synapsen über
Botenstoffe, so genannte Neurotransmitter wie γ-Aminobuttersäure (GABA) oder Glycin, erfolgt.
Bei chemischen Synapsen liegt der synaptische Spalt zwischen der präsynaptischen
Axonterminale und der postsynaptische Membran.
Kommt ein Aktionspotential in der präsynaptischen Axonterminale an, werden spannungs-
aktivierte Calciumkanäle geöffnet und Calciumionen strömen in die Axonterminale ein. Durch
die Erhöhung der intrazellulären Calciumkonzentration (Depolarisation) werden die mit
Neurotransmitter gefüllten synaptischen Vesikel zur präsynaptischen Membran hinbewegt. Die
dann durch Exozytose freigesetzten Neurotransmitter [10, 11] diffundieren über den
synaptischen Spalt zur Membran der postsynaptischen Nervenzelle, wo sie als Liganden an ihren
spezifischen Rezeptoren, wie z.B. ligandaktivierten Ionenkanälen binden. Diese Interaktion
zwischen Ligand und seinem Rezeptor führt zu einer Konformationsänderung des Rezeptors,
wodurch sich eine Pore des Rezeptors öffnet, die selektiv für bestimmte Ionen ist. Der Einstrom
des Ions verursacht eine Änderung des Membranpotentials entweder in depolarisierender
Richtung oder in hyperpolarisierender Richtung abhängig von der Art des Rezeptors.
Beispielsweise führt der erregende (exzitatorische) Neurotransmitter Glutamat zu einer
Depolarisation der Neuronenmembran und damit zur Ausbildung eines so genannten
exzitatorischen postsynaptischen Potential (EPSP). Dies löst ab einem bestimmten
Schwellenwert ein Aktionspotential aus. Dagegen verursacht ein hemmender (inhibitorischer)
Neurotransmitter wie GABA oder Glycin eine Hyperpolarisation der Neuronenmembran, ein so
genanntes inhibitorisches postsynaptisches Potential (IPSP), womit das Auftreten eines
Aktionspotentials gehemmt wird. Die Wiederaufnahme des Neurotransmitters ins Axon oder
sein Abbau führt zu seiner Inaktivierung, wodurch die Signalübertragung beendet wird.

Einleitung
3
1.2. Ionenkanäle
Ionenkanäle sind für bestimmte Ionen durchlässige Poren in der Zellmembran, welche von
Proteinen gebildet werden. Sie besitzen einen Selektivitätsfilter und eine Schleuse. Die
Bestimmung der Ionenart, die durch die Membran passieren kann, erfolgt durch den
Selektivitätsfilter. Die Bedingungen für die Öffnung des Kanals werden von der Schleuse
festgelegt [12]. Ionenkanäle werden nach ihrem Aktivierungsmechanismus in zwei Klasse
eingeteilt: Ionenkanäle, die durch die Änderung des Membranpotentials aktiviert werden, so
genannte spannungsgesteuerte Ionenkanäle und Ionenkanäle, die durch Bindung des Liganden
aktiviert werden, so genannte ligandengesteuerte Ionenkanäle. Aufgrund ihrer
Aminosäuresequenz können die ligandengesteuerten Ionenkanäle in drei Superfamilien
unterteilt werden: Die ATP-gesteuerten Purin-Rezeptoren (P2X), die Superfamilie der Glutamat-
Rezeptoren und die Superfamilie der Cys-Loop-Rezeptoren. Bei einer Superfamilie handelt es
sich um mehrere Rezeptorfamilien, die trotz ihrer ähnlichen Sequenz und einiger
übereinstimmenden Strukturmerkmale verschiedene Funktionen und Wirkungen aufweisen [12-
15]. Tabelle 1.1 liefert einen Überblick über die Superfamilien von der ligandengesteuerten
Ionenkanäle, deren Familien und ausgewählte strukturelle Eigenschaften.

Einleitung
4
Tabelle 1.1 Die Superfamilien der ligandengesteuerten Ionenkanäle, deren Familien und strukturelle
Eigenschaften.
Superfamilie Familie Strukturelle Eigenschaften
ATP-gesteuerte Purin-
Rezeptoren (P2X)
kationen-spezifisch
Trimer 2-Transmembran-domänen
Superfamilie der Glutamat-
Rezeptoren
N-Methyl-D-Aspartat (NMDA)-Rezeptoren
kationen-spezifisch
Tetramer 3-Transmembran-domänen
Kainat-Rezeptoren
α-Amino-3-hydroxy-5-methyl - 4-isoxazol Propionsäure (AMPA)-Rezeptoren
Superfamilie der Cys-Loop-Rezeptoren
Nicotinische Acetylcholin-Rezeptoren
kationen-
spezifisch
Pentamer
4-Transmembran-
domänen
Einige Serotonin-Rezeptoren; 5-HT3-Rezeptoren
Zn2+-aktivierte Ionenkanäle
GABAA/C-Rezeptoren anionen-spezifisch
Glycin-Rezeptoren
Die kationenspezifischen ATP-gesteuerten Purin-Rezeptoren (P2X) bestehen aus Untereinheiten,
die aus zwei Transmembrandomänen bestehen, und bilden Trimere. Die Superfamilie der
kationenspezifischen Glutamat-Rezeptoren hat eine tetramere Struktur, die aus drei
Transmembrandomänen enthaltenden Untereinheiten aufgebaut ist [12-15].
Alle Rezeptoren der Cys-Loop-Rezeptor-Superfamilie bestehen aus fünf Untereinheiten, welche
radial um den inneren Ionenkanal angeordnet sind, was als pentamere Struktur bezeichnet wird.

Einleitung
5
Jede Untereinheit enthält vier Transmembrandomänen mit hydrophoben
Aminosäuresequenzen. Obwohl alle Rezeptoren der Cys-Loop-Rezeptor-Superfamilie eine
Homologie bei einer Sequenzidentität von etwa 30 % zeigen, haben sie eine größere Ähnlichkeit
in der Sekundär- und Tertiärstruktur [16]. Alle Mitglieder dieser Superfamilie besitzen ähnliche
Peptidschleifen, die durch Disulfidbrücken gebildet werden, ähnliche Glycosylierungsmuster und
sowie ähnlich lokalisierte Prolin- und Serin/Threonin-Reste (siehe Abbildung 1.1). Alle
Rezeptoren aus dieser Superfamilie können durch Zink-Ionen allosterisch moduliert werden
[13]. Neben dieser strukturellen Ähnlichkeit haben aus dieser Superfamilie nur GABAA/C- und
Glycin-Rezeptoren Anionen (Chloridionen)-spezifische Ionenporen (siehe Tabelle 1.1),
sogenannte Chloridionenkanäle [12-15]. Somit stellen ihre natürlichen Liganden GABA und
Glycin die wichtigsten hemmenden Neurotransmitter im Zentralnervensystem dar [17, 18].
Abbildung 1.1 Schematische Darstellung der vier Transmembrandomänen der Cys-Loop Rezeptoren
(modifiziert nach Hucho et al. und Collingridge [12, 14]).
1.3. Das GABAerge System
GABA (γ-Aminobuttersäure) ist der wichtigste inhibitorische Neurotransmitter im
Zentralnervensystem [13, 17-24] und konnte erstmals im Jahr 1950 im Gehirn von Säugetieren
identifiziert werden [25, 26], obwohl seine Anwesenheit im biologischen Gewebe schon im Jahr
1910 beschrieben wurde [23]. 40 % aller Neuronen im Zentralnervensystem von Säugetieren
sind GABAerge Neuronen [27], in denen der Neurotransmitter GABA synthetisiert, freigesetzt
und metabolisiert wird [22]. GABAerge Neuronen existieren in zahlreichen Hirnarealen: im
Großhirn (Frontallappen, Parietallappen, Temporallappen besonders in der Amygdala und im
Hippocampus, Okzipitallappen) in der Brücke/Pons, im Thalamus, im Hypothalamus, im Kleinhirn
(Cerebellum), im Nucleus caudatus, in der grauen Substanz und in der weißen Substanz [25, 28-
32].

Einleitung
6
Die Synthese von GABA im Gehirn erfolgt über die Decarboxylierung von Glutamat. Mit Hilfe der
Glutamat-Decarboxylase (GAD) wird Glutamat in GABA umgewandelt [25, 26, 33]. Beim
Menschen kommt GAD in zwei von unterschiedlichen Genen kodierten Isoformen GAD65 und
GAD67 vor [34, 35]. Während GAD67 (67 kDA mit 594 Aminosäureresten [34]) in der Zelle
weiter verbreitet ist und bevorzugt zytoplasmatisch GABA synthetisiert, tritt GAD65 (65 kDA mit
585 Aminosäureresten [34]) in der Zellmembran und in Nervenenden auf. Durch diese Enzyme
wird GABA vorzugsweise für die vesikuläre Freisetzung gebildet [35]. Durch vesikuläre GABA-
Transporter wird die synthetisierte GABA in die Vesikel transportiert und gespeichert [36, 37].
Abbildung 1.2 Schematische Abbildung des GABAergen System. GAD: Glutamat-Decarboxylase, SSA:
Succinatsemialdehyd, GABA-AT:GABA-Transaminase (modifiziert nach Treiman et al. [30]).
Erreicht ein Aktionspotenzial die präsynaptische GABAerge Nervenendung, öffnen sich
Calciumkanäle, wodurch Calciumionen einströmen. Aufgrund der erhöhten intrazellulären
Calciumkonzentration fusionieren die mit Neurotransmitter GABA gefüllten Vesikel mit der
präsynaptischen Membran. Dies bewirkt in den synaptischen Spalt die Freisetzung von GABA
[10, 11], die an GABA-Rezeptoren auf der Oberfläche von postsynaptischen GABAergen
Nervenzellen bindet (siehe Abbildung 1.2). Die Aktivierung der GABA-Rezeptoren führt zur
Hyperpolarisation der Plasmamembran. Durch Wiederaufnahme des Neurotransmitters GABA
mittels membranständiger GABA Transporter kann GABA aus dem synaptischen Spalt entfernt
werden, womit die synaptische Wirkung der GABA und somit die Signalübertragung beendet
wird [19, 30]. GABA wird nach ihrer Aufnahme enzymatisch abgebaut. Der Abbau von GABA

Einleitung
7
erfolgt durch Transaminierung, wobei GABA unter Katalyse der GABA-Transaminase (GABA-AT)
zu Succinatsemialdehyd umgesetzt wird, der dann von Succinatsemialdehyd-Dehydrogenasen zu
Succinat oxidiert wird oder über eine Nebenreaktion durch Succinatsemialdehyd-Reduktase zur
γ-Hydroxybuttersäure umgewandelt werden kann [38-40]. Beim Abbau von GABA wird durch
die GABA-AT gleichzeitig Glutamat aus α-Ketoglutarat regeneriert [41-44].
1.3.1. GABA Rezeptoren
Zur Auslösung der inhibitorischen Wirkung bindet GABA an einen von drei Rezeptortypen:
GABAA-, GABAB- und GABAC- Rezeptoren [45-47].
1.3.1.1. GABAA-Rezeptoren
Der GABAA-Rezeptor, welcher ein Mitglied der Cys-Loop-Rezeptorsuperfamilie der
ligandengesteuerten Ionenkanäle ist, besteht aus fünf heterologen Untereinheiten, von denen
jede aus vier- Transmembrandomänen (TM 1, TM 2, TM 3 und TM 4) aufgebaut ist [12-14, 46].
Die Transmembrandomänen sind durch intrazelluläre (zwischen TM 1 und TM 2; sowie zwischen
TM 3 und TM 4) oder extrazelluläre (zwischen TM 2 und TM 3) Schleifen verbunden. Die
intrazelluläre Schleife zwischen TM 3 und TM 4, welche Phosphorylierungsstellen enthält, ist an
der Modulation der Rezeptoraktivität durch Phosphorylierung beteiligt [48]. Durch die α-Helices
bildenden TM 2-Domänen der fünf Untereinheiten wird der zentrale Kanal gebildet (siehe
Abbildung 1.3), welcher eine Schleuse und den Selektivitätsfilter zur Passage der Chloridionen
enthält. Dabei umfasst jede Untereinheit einen großen extrazellulären hydrophilen N-Terminus
und einen relativ kurzen extrazellulären C-Terminus [16].
Abbildung 1.3 Schematische Darstellung der vier Transmembrandomänen der GABAA-Rezeptor-
Untereinheit und der aus α-, β-, und γ-Untereinheiten bestehende GABAA-Rezeptor
(modifiziert nach Bormann [46]).

Einleitung
8
Für die menschlichen GABAA-Rezeptoren wurden 19 verschiedene Untereinheiten aus acht
Untereinheitsfamilien identifiziert; sechs alpha- (α1–α6), drei beta- (β1–β3), drei gamma- (γ1–
γ3), 3 rho- (ρ1–ρ3), eine delta- (δ), eine epsilon- (ε), eine theta- (θ) und eine pi- (π) Untereinheit
[14, 16, 49]. Da es sich um viele verschiedene Untereinheiten der GABAA-Rezeptoren handelt, ist
eine große Zahl von Kombinationsmöglichkeit gegeben, um den vollständigen Rezeptor zu
bilden. Bei Erwachsenen wird allgemein der aus α1-, β2- und γ2- bestehende GABAA-
Rezeptorkomplex als Hauptisoform beschrieben [48]. Die pharmakologischen und
elektrophysiologischen Eigenschaften des Rezeptors hängen von den Untereinheiten ab, die den
Rezeptor bilden [16, 50-53]. In Tierversuchen wurde gezeigt, dass die sedativ, amnestische und
teilweise antikonvulsive Wirkung von Benzodiazepinen durch die α1- enthaltenden GABAA-
Rezeptoren vermittelt wird [16, 50, 51, 54-56]. Während diese α1- enthaltenden GABAA-
Rezeptoren keine Rolle bei der durch GABAA-Rezeptoren vermittelten angstlösenden Wirkung
spielen [55], entfalten die angstlösenden und muskelentspannenden Substanzen ihren Effekt
über GABAA-Rezeptoren, welche eine α2-, α3- oder α5-Untereinheit besitzen [16, 50, 51, 54, 56].
Darüber hinaus sind α2- und α3-Untereinheiten an den spinalen antihyperalgesischen Effekten
von Benzodiazepinen beteiligt [54]. Des Weiteren zeigen einige Substanzen wie beispielsweise
antikonvulsive Mefenaminsäure eine β-Untereinheit-spezifische Wirkung, so dass sie die β2
oder β3 enthaltenden GABAA-Rezeptoren potenzieren und auch direkt aktivieren [57]. Die
Bindungsstelle für GABA liegt an der Kontaktfläche zwischen den α-Untereinheiten und β-
Untereinheiten [58]. An den binären GABAA-Rezeptoren aus α- und β-Untereinheiten und an
den ternären GABAA-Rezeptoren aus α-, β- und γ2-Untereinheiten bestimmen die den Rezeptor
bildenden α-Untereinheiten die Wirksamkeit von GABA, wohingegen β2- oder β3-
Untereinheiten an den binären GABAA-Rezeptoren keinen Einfluss auf die GABA-Potenz haben
[52]. Damit zeigt jeder GABAA-Rezeptorkomplex unterschiedliche Affinität zu GABA aufgrund
extrasynaptischer oder synaptischer Lokalisation der den Rezeptor bildenden Untereinheiten
[52, 53, 59]. GABAA-Rezeptoren, welche die extrasynaptisch lokalisierten α4-/α6- oder δ-
Untereinheiten enthalten, sind die potentesten GABAA-Rezeptoren [52, 53, 59]. Die binären
GABAA-Rezeptoren weisen eine höhere Affinität zu GABA auf als ihre γ2-Untereinheit
enthaltenden ternären GABAA-Rezeptorkomplexe, sodass die aus α1 und β2 bestehenden
GABAA-Rezeptoren im Vergleich zu den α1β2γ2L GABAA-Rezeptoren einen kleineren EC50-Wert
für GABA haben [52].
Die durch synaptische GABA aktivierten GABAA-Rezeptoren, die auf der postsynaptischen Seite
im Gehirn liegen, verursachen schnell eine Hyperpolarisation der postsynaptischen Membran
durch den Einstrom von negativen Chloridionen, die so genannte phasische GABAerge
Inhibition. Dagegen können die auf der extrasynaptischen Seite lokalisierten GABAA-Rezeptoren
die tonische GABAerge Inhibition vermitteln, wobei es sich hier um eine langwierige Aktivierung
von GABAA-Rezeptoren durch die niedrige umgebende GABA Konzentration handelt [60-62]. Die

Einleitung
9
α1-, β2/3-, und γ2-Untereinheiten der GABAA-Rezeptoren finden sich vorwiegend in GABAergen
Synapsen [63, 64] in zahlreichen Gehirnregionen inklusive Hippocampus, Cerebellum und
Pallidum [65] und vermitteln die phasische GABAerge Inhibition [60]. Die α2- und α3-
Untereinheit enthaltenden GABAA-Rezeptoren (α2β2/3γ2 und α3β2/3γ2) verursachen auch eine
phasische neuronale Inhibition [60]. Dagegen sind δ-Untereinheit enthaltende GABAA-
Rezeptoren nur in extrasynaptischen somatischen und dendritischen Membranen in den
Granularzellen des Cerebellums lokalisiert und vermitteln eine tonische GABAerge Inhibition
[63, 66].
1.3.1.2. GABAB- und GABAA-Rezeptoren
Genauso wie heteropentamere GABAA-Rezeptoren gehören ionotrope homopentamere GABAC -
Rezeptoren zur Cys-Loop-Rezeptorsuperfamilie der ligandengesteuerten Ionenkanäle,
wohingegen es sich bei metabotropen GABAB-Rezeptoren um G-Protein-gekoppelte Rezeptoren
mit einer heterodimeren Struktur handelt. Im Vergleich zu GABAA-Rezeptoren werden GABAC -
Rezeptoren, die aus ρ- Untereinheiten (ρ1–ρ3) bestehen, durch Bicuculline nicht blockiert und
durch Steroide, Barbiturate, und Benzodiazepine nicht moduliert. Das GABA Analogon cis-4-
Aminocrotonsäure ist ein selektiver Agonist für GABAC -Rezeptoren, während (1, 2, 5, 6-
Tetrahydropyridine-4-yl) methyl phosphonensäure auf GABAC-Rezeptoren selektiv
antagonistisch wirkt. GABAC -Rezeptoren befinden sich überwiegend in der Retina von
Wirbeltieren [45-47].
GABAB-Rezeptoren, die Rezeptoren mit sieben Transmembrandomänen sind, vermitteln eine
neuronale Inhibition, indem sie über G-Proteine auf Ca+2- und K+- Ionenkanäle wirken. Diese
Rezeptoren können selektiv durch Baclofen aktiviert werden, während Phosphonobaclofen sie
selektiv blockiert [47].
1.3.2. Funktionen des GABAergen Neurotransmittersystem
Die Aktivierung der GABA-Rezeptoren führt zur Hyperpolarisation der Plasmamembran durch
Chloridionen-Einstrom. Auf diese Weise wird die Entstehung des Aktionspotentials gehemmt. Da
40 % aller Neuronen im Zentralnervensystem GABAerge Neuronen sind [27], spielt der
Neurotransmitter GABA durch seinen inhibitorischen Effekt auf Neurotransmission eine wichtige
Rolle bei der Regulation der Aktivität der exzitatorischen Neurotransmission [67]. Dabei haben
GABAerge Neuronen eine Schlüsselrolle für die Verhaltensregulation [68]. Das GABAerge
Neurotransmittersystem ist ein Zielort für zahlreiche Medikamente, welche zur Behandlung
vieler neurophysiologischer Krankheiten verwendet werden. Darunter zählt insbesondere
Epilepsie, wobei die GABAergen Neuronen eine wichtige Rolle bei der Entstehung und
Ausbreitung oder bei der Unterdrückung epileptischer Aktivität spielen [21, 30]. Neben

Einleitung
10
Antiepileptika (Antikonvulsiva) üben Sedativa, Hypnotika und Anxiolytica ihre Effekte durch
Verstärkung der GABAergen Inhibition über den GABAA-Rezeptoren aus [48, 61, 67-69]. Des
Weiteren regulieren viele Substanzen das Schlaf-Wach-Verhalten über die GABAA-Rezeptoren
[68-72]. Zudem wird die Verstärkung der GABAergen Inhibition zur Behandlung von
neuropathischen und chronischen Schmerzen verwendet [54, 69, 73, 74]. Weitere
physiologische Funktionen des GABAergen System sind als antidepressive und antihypertensive
Aktivität in der Literatur beschrieben [75].
1.3.3. Modulation des GABAergen Neurotransmittersystems
Zahlreiche pharmakologische Substanzen modulieren das GABAerge System auf verschiedene
Weise, wodurch die GABAerge Wirkung verstärkt werden kann. Dazu zählen die Erhöhung der
GABA-Synthese, Förderung der synaptischen GABA-Freisetzung, Inhibition des GABA-Abbaus,
Hemmung der GABA-Aufnahme, Verwendung von GABA-Agonisten und Anwendung von
positiven allosterischen Modulatoren des GABA-Rezeptors [21]. Dabei können z.B. Inhibitoren
der GABA-abbauenden Enzyme oder neue allosterische Modulatoren die besten
therapeutischen Möglichkeiten mit den geringsten Nebenwirkungen zur Behandlung von
Epilepsie anbieten [21].
Benzodiazepine, die positive allosterische Modulatoren der GABAA-Rezeptoren sind, binden an
eine Bindungsstelle, die an der Grenzfläche zwischen den α- und γ-Untereinheiten des
Rezeptors liegt [76], wodurch sie die Bindung des Neurotransmitters GABA an seinen Rezeptor
fördern [77]. Dies führt zu einer Erhöhung des Chloridionen-Einstroms [76], wodurch
Benzodiazepine ihre antikonvulsive, sedative-hypnotische und anxiolytische Wirkung vermitteln
[77]. Benzodiazepine können den Rezeptor nicht direkt aktivieren [76], sie können nur die
GABA-induzierte Öffnungsfrequenz des Rezeptors erhöhen [77] (siehe Abbildung 1.4).

Einleitung
11
Abbildung 1.4 Schematische Darstellung der allosterischen Modulation des GABAA-Rezeptors (modifiziert
nach Atack [76]).
Die Aktivierung der GABAA-Rezeptoren wird nicht nur durch Benzodiazepine sondern auch durch
zahlreiche Substanzen wie Neurosteroide, Barbiturate oder Anästhetika potenziert [18, 22, 46,
47, 78]. Neben diesen synthetischen Substanzen können verschiedene Lebensmittel bzw. ihre
Inhaltsstoffe den GABAA-Rezeptor modulieren [27, 79-96]. Des Weiteren können einige natürlich
vorkommende Substanzen wie Muscimol und auch synthetische Substanzen wie THIP (4, 5, 6, 7-
Tetrahydroisoxazolopyridin-3-ol) GABAA-Rezeptoren direkt aktivieren [22, 46, 47]. Einige
Lebensmittelinhaltsstoffe wie Terpenoide können eine direkte Rezeptoraktivierung auslösen
[87], während die Aktivität des Rezeptors durch manche Flavanoide wie Apigenin und
Epigallocatechingallat reduziert werden kann [96]. Biccuculline und Picrotoxin blockieren die
GABAA-Rezeptoren [22, 46, 47].

Einleitung
12
1.4. Glycin-Rezeptoren
Wie bereits erwähnt gehören die Glycin-Rezeptoren ebenfalls zu der Cys-Loop-Rezeptor-
Superfamilie [12-14]. Für Glycin-Rezeptoren wurden vier α-Untereinheiten (α1–α4) und eine β-
Untereinheit beschrieben [23, 97], welche aus vier Transmembrandomänen aufgebaut ist [12-
14]. Dabei können α-Untereinheiten entweder Homopentamere bilden oder mit der β-
Untereinheit Heteropentamere bilden[18].
Glycin wird aus Serin durch Serin-Hydroxy-Methyltransferasen synthetisiert und durch
vesikuläre inhibitorische Aminosäure-Transporter in die synaptischen Vesikel befördert. Das
durch Calcium-abhängige Exozytose freigesetzte Glycin interagiert mit seiner Bindestelle am
Glycin-Rezeptor, die von α-Untereinheiten gebildet wird. Durch die Bindung des Glycins öffnet
sich ein Chloridionen-Kanal. Hieraus resultiert eine Hyperpolarisation der Plasmamembran und
somit ein inhibitorisches postsynaptisches Potential. Die Neurotransmission wird durch
Wiederaufnahme des Glycins in Gliazellen und in der Präsynapse beendet [97].
Während GABAerge Neuronen in zahlreichen Gehirnrealen auftreten, sind glycinerge Neuronen
vor allem im Rückenmark und im Hirnstamm lokalisiert [17, 23, 97]. Glycin-Rezeptoren sind dort
bedeutene inhibitorische Rezeptoren [13]. Sie regulieren die Aktivität der Motorneuronen,
koordinieren Reflexe und verarbeiten sensorische Signale [23, 97]. Anästhetika, Alkohole,
Neurosteroide modulieren den Glycin-Rezeptor allosterisch [17, 18, 78, 97]. Strychnin blockiert
spezifisch den Glycin-Rezeptor [23, 97].
1.5. Die Voltage-Clamp-Technik
Zur Untersuchung der Wirkung von Lebensmittelextrakten bzw. Lebensmittelinhaltsstoffen auf
die GABAA- und Glycin-Rezeptoren wurde in dieser Arbeit die Voltage-Clamp (aus dem
Englischen: Two electrode voltage clamp; TEVC)-Technik, welche ein elektrophysiologisches
Messverfahren ist, verwendet. Durch diese elektrophysiologische Technik kann der
Chloridionenstrom, der über die exprimierten Rezeptoren bzw. Ionenkanäle vermittelt wird,
über das Membranpotential gemessen werden. Bei der TEVC-Technik werden Oocyten aus
Xenopus laevis als Expressionssystem für Ionenkanäle verwendet [98, 99]. Die Oocyten besitzen
alle für die Expression von Proteinen benötigten Komponenten, sodass die injizierte RNA
effizient translatiert werden kann [98].
Das TEVC-Messsystem besteht aus zwei intrazellulären Elektroden (Strom- und Spannungs-
elektrode), zwei extrazellulären Badelektroden (Erdungs- und Referenzelektrode), einem
Rückkopplungs-verstärker, einem Mikroskop, einer Messkammer und einem Perfusionssystem.
Alle Elektroden sind mit dem Verstärker verbunden und bestehen aus chloriertem Silberdraht.

Einleitung
13
Intrazelluläre Spannungs- und Stromelektroden befinden sich in mit 3 M KCl gefüllten
Glassmikropipetten, die aus Glasskapillaren mit Hilfe eines Mikropipettenziehgeräts hergestellt
werden.
Abbildung 1.5 Schematische Darstellung einer Voltage–Clamp Technik (modifiziert nach Wagner et al .[98]).
Die Badelektroden und die Oocyte werden in der Messkammer unter dem Mikroskop platziert.
Die applizierten Lösungen fließen aufgrund des hydrostatischen Drucks in die Kammer, wobei
das Perfusionssystem den schnellen Austausch der Lösungen bei der Messung ermöglicht. Die
Oocyte wird vor und nach der Applikation der Testsubstanzen kontinuierlich mit Puffer
überspült. Beide intrazelluläre Spannungs- und Stromelektroden werden zur Erfassung des
Membranpotentials in die Oocyte eingestochen (siehe Abbildung 1.5). Die Messung des
aktuellen Membranpotentials erfolgt durch die intrazelluläre Spannungselektrode.
Oocyte
Spannungselektrode
Stromelektrode
Badeelektrode
Verstärker

Einleitung
14
1.6. Test-Lebensmittel
Wie bereits in Abschnitt 1.3.3 beschrieben, kann das GABAerge System durch zahlreiche
Substanzen, darunter auch durch Lebensmittelinhaltsstoffe moduliert werden. Zur
Untersuchung der möglichen GABAergen Wirkung wurden in dieser Arbeit 13 Lebensmittel
(Salbei, Kamille, Hopfen, Melisse, Schwarzer Pfeffer, Nelken, Kakao, Grüner Tee, Lavendel und 4
Sideritis-Arten) ausgewählt, welche basierend auf untenstehenden Literaturdaten aus
Tierversuchen oder klinischen Studien eine relevante neurophysiologische Wirkung haben. Sie
besitzen beispielsweise beruhigende, angstlösende, sedative, antidepressive oder
antiepileptische Aktivität, die durch eine mögliche Modulation des GABAergen Systems
ausgelöst werden könnte.
1.6.1. Lavendel
Lavendel-Arten gehören zur die Gattung Lavendel (Lavandula) der Unterfamilie der Nepetoideae
in die Familie der Lippenblütler (Lamiaceae). In zahlreichen Tierversuchen sowie klinischen
Studien wurde die schlaffördernde, angstlösende und sedative Wirkung des ätherischen
Lavendelöls und des wässrigen sowie methanolischen Lavendel-Extraktes nachgewiesen [100-
102]. Des Weiteren wurde der GABAA-Rezeptor modulierende Effekt des Lavendelöls und sowie
des Dichlormethan-Extraktes von Lavendel in vitro ermittelt [80, 103]. Zudem wurde die
inhibierende Wirkung von Lavendelöl auf spannungsgesteuerte Calciumkanäle berichtet [104].
1.6.2. Salbei
Salbei gehört zur Gattung Salvia der Unterfamilie der Nepetoideae in der Familie der
Lippenblütler (Lamiaceae). Salbei-Arten sind weltweit besonders in Mittel- und Südwestasien
verbreitet [105]. Sie werden zur Behandlung von Atemwegserkrankungen, Magen-Darm-
Verdauungsstörungen, Herz-Kreislauf-Erkrankungen, Krebs, Entzündungen und Infektionen
verwendet. Des Weiteren wurde ihre neurophysiologische Aktivität beschrieben und gezeigt,
dass Salbei sedative, hypnotische, muskelrelaxierende, schlaffördernde und
gedächtnisfördernde Wirkung aufweist [106, 107]. In Tierexperimenten wurde herausgefunden,
dass der Wasser-Alkohol-Extrakt von Salvia sp. verticillata einen antidepressiven Effekt aufwies,
der durch den erzwungenen Schwimmtest (Forced Swimming Test) und den Tail-Suspension
Test gemessen wurde. Weiterhin zeigte der Extrakt eine antiepileptische Wirkung, die mittels
des Pentylentetrazol-Krampf-Tests und des maximalen Elektroschocks Beschlagnahme Tests
bestimmt wurde [105]. Darüber hinaus untersuchten Khayate-Nouri et al. die antikonvulsive
Wirkung von Salvia officinalis. Dabei stellten sie eine erhöhte Beschlagnahme-Schwelle bei den
Mäusen fest, denen ein hydroalkoholischer Extrakt von Salbei intraperitoneal appliziert wurde

Einleitung
15
[108]. Zudem wurde der angstlösende Effekt von Salbei untersucht. Dafür wurden der Elevated
Plus-Maze Test und der Hell/Dunkel (Light/Dark) Test an Mäusen durchgeführt, wobei keine
signifikanten Unterschiede zwischen den mit dem Salbeiextrakt behandelten Mäusen und der
Kontrollgruppe beobachtet werden konnten [105].
1.6.3. Melisse (Zitronenmelisse)
Melisse (Melissa officinalis) gehört zur Gattung der Melissen (Melissa) der Unterfamilie der
Nepetoideae der Familie der Lippenblütler (Lamiaceae). Melisse, die ursprünglich aus dem
östlichen Mittelmeerraum und Westasien stammt, wird als pflanzliches Arzneimittel traditionell
aufgrund ihrer krampflösenden, verdauungsfördernden, sedativ-hypnotischen, antidepressiven
und schmerzlindernden Wirkung verwendet [109]. Es wurde berichtet, dass der wässrige Extrakt
von Melisse einen starken inhibierenden Effekt auf die GABA-abbauenden Enzyme aufweist
[110].
In tierexperimentellen Studien wurde der antidepressive Effekt von Melisse untersucht. Dafür
wurden das ätherische Öl und ein wässriger Extrakt von Melisse in Mäuse intraperitoneal
injiziert. Um die antidepressive Wirkung zu bestimmen, wurde der erzwungene Schwimmtest
durchgeführt. Dabei wurde berichtet, dass sowohl das ätherische Öl von Melisse als auch ihr
wässriger Extrakt eine Erhöhung der Schwimmbewegungen hervorruft. Neben einer
antidepressiver Wirkung zeigt der wässrige Melissenextrakt einen sedativen Effekt, da er eine
dosisabhängige Verringerung der spontanen motorischen Aktivität verursacht [109]. Des
Weiteren zeigte die orale Applikation von 360 mg/kg Melissenextrakt bei Mäusen einen
anxiolytisch-ähnlichen Effekt, der durch Elevated Plus-Maze Test untersucht wurde [111].
1.6.4. Sideritis
Sideritis spp. gehört zur Gattung der Gliedkräuter (Sideritis) der Unterfamilie der Lamioideae der
Familie der Lippenblütler (Lamiaceae) und ist auch als Bergtee bekannt. Die Gattung Sideritis ist
im Mittelmeerraum weit verbreitet. Sideritis-Arten, die traditionell als Tee konsumiert werden,
werden in der Volksmedizin als entzündungshemmende, antimikrobielle, krampflösende,
antikonvulsive, analgetische, antiulcerative, antioxidative und verdauungsfördernde Mittel
verwendet [112]. In an Mäusen durchgeführten Studien wurde festgestellt, dass verschiedene
Sideritis-Arten Entzündungen hemmen und Schmerz lindern [113, 114].
Ein weiterer Tierversuch zeigte, dass die Applikation von 250 mg/kg Sideritis Extrakt zu einer
möglichen depressiven Wirkung führte, die durch Verringerung der Schwimmzeit von Mäusen
bestimmt wurde, während Sideritis–Extrakte bei einer hohen Konzentration von 500 mg/kg das

Einleitung
16
Zentralnervensystem stimulierten, sodass sie eine Erhöhung der Schwimmzeit verursachten
[115].
In einer tierexperimentellen Studie wurde die angstlösende Wirkung von Sideritis untersucht.
Zur Messung des Angstverhaltens wurde der Thigmotaxis Test durchgeführt, der die
Bewegungstendenz entlang des Randes bewertet. Dabei wurde berichtet, dass die Aufnahme
von Sideritis-Tee (4%(w/v)) bei den Mäusen eine Verringerung der thigmotaxischen Zeit und
eine Erhöhung der Anzahl der Besuche in der zentralen Zone verursacht, was als angstlösende
Wirkung interpretiert wird [116].
Des Weiteren wurde die Beeinflussung von GABAA-Rezeptoren durch Sideritis-Arten in vitro
untersucht. Dabei wurde festgestellt, dass die Dichlormethan-Extrakte von Sideritis-Arten die
Aktivität der GABAA-Rezeptoren potenzieren [103]. In einer kürzlich veröffentlichten Studie
wurden GABAerge Komponenten von Sideritis-Arten identifiziert, welche die GABAA-Rezeptor-
Aktivität allosterisch modulieren [87].
1.6.5. Grüner Tee
Grüner Tee (Camellia sinensis) gehört zur Gattung der Kamelien (Camellia) in der Familie
Teestrauchgewächse (Theaceae) und wird in der ganzen Welt zur Reduzierung von Stress und
Müdigkeit sowie zur Linderung von Neuralgie (Nervenschmerz) konsumiert [83]. Als
Hauptbestandteile von Tee wurden Flavanoide identifiziert, deren antioxidative, antitoxische,
antikarzinogene, antivirale, vaskuloprotektive und antispastische Wirkung bereits beschrieben
wurde [117]. Das Flavanoid Epigallocatechingallat, welches ein Hauptcatechin des grünen Tees
ist, verteilt sich nach dem Teekonsum im Gehirn [117]. An Küken durchgeführte Versuche
ergaben, dass Epigallocatechingallat im Gehirn sedativ und hypnotisch wirkt und damit die akute
Stressantwort moderiert [117]. Eine Studie an Mäusen zeigte, dass Epigallocatechingallat eine
angstlösende Wirkung hat, die durch den Elevated Plus-Maze Test gemessen wurde [118]. Des
Weiteren wurde die angstlösende Wirkung von L-Theanin, welches eine wichtige bioaktive
Komponente von grünem Tee ist, in einer Humanstudie beschrieben [119].
Die Wirkung des grünen Tees und seiner Inhaltsstoffe auf die GABAA-Rezeptoren wurde bereits
in zahlreichen Studien untersucht. Dabei wurde eine Verminderung der GABAA-Rezeptor-
Aktivität sowohl durch grünen Tee als auch durch einige seiner Inhaltsstoffe wie Koffein
berichtet [79, 83, 96]. Allerdings wurde auch die Potenzierung der Aktivierung von GABAA-
Rezeptoren durch manche natürliche Komponenten in grünem Tee wie beispielsweise Linalool
beschrieben [83, 95].

Einleitung
17
1.6.6. Schwarzer Pfeffer
Schwarzer Pfeffer (Piper nigrum) gehört zu der Familie der Pfeffergewächse (Piperaceae). In der
traditionellen Maya-Medizin (Q`eqchi Maya im Süden von Belize) werden die Pfeffer Spezies als
Heilpflanze zur Behandlung von Epilepsien verwendet [120]. Des Weiteren wird schwarzer
Pfeffer für eine analgetische, entzündungshemmende, antikonvulsive, antioxidative,
antidepressive Wirkung in der traditionalen Medizin in vielen Ländern eingesetzt [121]. In
tierexperimentellen Studien wurde der Effekt von Kava (Piper methysticum) auf Angstzustände
untersucht. Dabei wurde seine akute angstlösende Wirkung mittels Elevated Plus-Maze Tests
bei Ratten bestimmt [122]. Eine weitere Studie an Ratten zeigte, dass der methanolische Extrakt
von schwarzem Pfeffer die neurotoxische Beta-Amyloid (1-42)-induzierte Angst und
Depressionen durch Abschwächung des oxidativen Stresses lindert [121].
Es zeigte sich, dass die Piper-Arten die Aktivität der GABA-abbauenden Enzyme stark inhibieren
[120]. Des Weiteren wurde berichtet, dass der Ethylacetat- Extrakt von schwarzem Pfeffer den
GABA-vermittelten Strom über den GABAA-Rezeptor verstärkt [92].
1.6.7. Kamille
Die echte Kamille (Matricaria chamomilla L) gehört zur Familie der Korbblütler (Asteraceae). In
der Volksmedizin finden die getrockneten Blütenknöpfe von Kamille Anwendung zur Therapie
von vielfältigen Beschwerden wie Schlaflosigkeit, Muskelkrämpfe, Angstzustände, Magen-Darm-
Erkrankungen, Entzündungen und rheumatische Schmerzen [123, 124]. In Tierversuchen wurden
die neuropharmakologischen Effekte des Flavanoids Apigenin, das aus Kamillen isoliert wurde,
untersucht. Dabei wurde berichtet, dass das in Ratten intraperitoneal injizierte Apigenin eine
sedative Wirkung durch Reduzierung der lokomotorischen Aktivität zeigte. Dagegen wurde
keine muskelrelaxierende, angstlösende oder antikonvulsive Wirkung von Apigenin beobachtet
[124].
In einer klinischen Studie wurde berichtet, dass Kamille neben ihrer angstlösenden Aktivität eine
klinisch relevante antidepressive Wirkung auslösen kann [125].
Des Weiteren wurde die modulierende Wirkung von Kamille auf das GABAerge System
untersucht. Dabei wurde festgestellt, dass wässriger Extrakt von Kamille die GABA-abbauenden
Enzyme hemmt [110].

Einleitung
18
1.6.8. Nelken
Gewürznelken (Syzygium aromaticum, Synonyme: Eugenia aromatica, Eugenia caryophyllata)
gehören zur Gattung der Syzygium der Familie der Myrtengewächse (Myrtaceae) und wachsen
in Indonesien, Tansania, Indien, Malaysia, Madagaskar und Sri Lanka [126, 127]. In der Literatur
wurde beschrieben, dass Nelken, ihre Komponenten oder ihr ätherisches Öl antimikrobielle,
anti-fungale, muskelrelaxierende, antikarzinogene, analgetische und entzündungshemmende
Wirkung zeigen [127]. Das Nelkenöl wird traditionell in der Zahnpflege als ein analgetisches und
antiseptisches Mittel verwendet [126]. Des Weiteren werden die Knospen von Nelken in der
iranischen Volksmedizin als Antiepileptikum eingesetzt [127].
Die analgetische Wirkung des Nelkenöls auf Mäuse wurde in verschiedenen Testsystemen
untersucht. Dabei wurde berichtet, dass ein in Mäuse intraperitoneal injizierte Nelkenöl sowohl
im Hot-Plate-Test [127] als auch im Formalintest [126] analgetisch wirkt. Neben seiner
analgetischen Wirkung wurde sein lokalanästhetischer Effekt in einer klinischen Studie
nachgewiesen [128]. Des Weiteren wurde in einem Tierexperiment gezeigt, dass das Nelkenöl
eine angstlösende, hypnotische und antiepileptische Wirkung hat [129].
In der Literatur wurde Eugenol als Hauptbestandteil des Nelkenöls beschrieben [130-136].
Zahlreiche elektrophysiologische Untersuchungen zeigten, dass Eugenol spannungsgesteuerte
Natriumkanäle [137] und Calciumkanäle [138, 139] inhibiert, während es dagegen
ligandengesteuerte Chloridionenkanäle des GABAA-Rezeptor potenziert [80, 82, 89]. Diese
elektrophysiologisch aktive Substanz Eugenol zeigte in Tierversuchen anästhetische Wirkung auf
Ratten und Xenopus leavis Frösche [140, 141].
Des Weiteren wurde die analgetische, antikonvulsive, sedative und angstlösende Wirkung des
Eugenols in verschiedenen tierexperimentellen Modellen untersucht. Dabei wurde berichtet,
dass das in Mäuse bzw. Ratten intraperitoneal injizierte Eugenol eine analgetische Wirkung hat,
die durch den Hot-Plate-Test, den Tail-Flick-Test (Schwanzbewegungstest) und den Essigsäure-
Writhing-Test gemessen wurde. Eine antikonvulsive Wirkung des Eugenols zeigte sich sowohl im
Pentylentetrazol-Krampf-Test und im maximalen Elektroschock-Beschlagnahme-Test. Zudem
wirkte Eugenol sedativ durch Erhöhung der Schlafzeiten. Dagegen wurde keine signifikante
Wirkung des Eugenols auf Angstzustände im Open-Field Test und im Passiva Avoidance Test
beobachtet [142].
Bei einer Studie an Ratten wurde nach lumbaler intrathekaler Injektion die Penetration des
Eugenols ins ZNS (Zentral Nerven System) untersucht. Dabei wurde Eugenol im Gehirn und im
Rückenmark durch Flüssigchromatographie mit tandemmassenspektrometrischer Detektion (LC-

Einleitung
19
MS/MS) nachgewiesen. Zusätzlich zeigte sich, dass Eugenol neuropathische Schmerzen lindern
kann [143].
1.6.9. Hopfen
Echter Hopfen (Humulus lupulus) gehört zur Gattung der Hopfen (Humulus) der Familie
Hanfgewächse (Cannabaceae). Hopfen findet unter anderem wegen seines charakteristischen
Aromas Anwendung beim Bierbrauen. Darüber hinaus wird Hopfen nicht nur in der
traditionellen sondern auch in der modernen Phytotherapie wegen seines beruhigenden,
sedativen und milden hypnotischen Effekts verwendet [144-146].
In Tierexperimenten wurde seine sedative Wirkung untersucht. Dazu wurden Wachteln als
Versuchstiere verwendet, da sie einen ähnlichen Schlaf-Wach-Rhythmus aufweisen wie
Menschen. Dabei zeigte sich, dass die Aufnahme des Trockenextraktes aus Hopfen sowohl
während des Tages als auch während der Nacht die lokomotorischen Aktivität vermindert [144].
In klinischen Studien wurde berichtet, dass alkoholfreies Bier aufgrund seines Hopfen-Gehalt
eine sedative Wirkung auslöst [146]. Des Weiteren wurde in einer Studie an Ratten gezeigt, dass
die Fraktion aus Hopfen, die β-Säure enthält, im erzwungenen Schwimmtest ebenso wie
Imipramin (Antidepressiva) eine verringerte Immobilitätzeit verursacht und somit eine
antidepressive Wirkung vermuten lässt [145].
Des Weiteren wurde der Einfluss der wässrigen und ethonolischen Extrakte von Hopfen auf die
GABA-abbauenden Enzyme untersucht. Dabei wurde berichtet, dass sowohl ein wässriger als
auch ein ethonolischer Extrakt von Hopfen zur Inhibition der GABA-AT führt [110]. Zudem wurde
die modulierende Wirkung des Hopfens auf GABAA-Rezeptoren elektrophysiologisch getestet.
Dabei zeigte sich, dass ein ätherisches Hopfenöl die Aktivität der GABAA-Rezeptoren potenziert
[147].

Einleitung
20
1.6.10. Kakao
Der Kakaobaum (Theobroma cacao) gehört zur Gattung der Theobroma der Familie Malvenge-
wächse (Malvaceae). Die aus Kakao gewonnenen Produkte, die einen hohen Gehalt an
Flavanoiden haben, zeigen vor allem starke antioxidative Wirkung [148, 149]. Darüber hinaus
wurde in Tierexperimenten die antidepressive Wirkung von Kakao durch den erzwungenen
Schwimmtest untersucht. Dabei zeigte sich, dass die Aufnahme des polyphenolischen Extraktes
von Kakao bei Ratten einen antidepressivaähnlichen Effekt durch Reduzierung der Immobilitäts-
Dauer hervorruft [148]. Eine weitere Studie an Ratten zeigte, dass die Aufnahme des
polyphenolischen Extraktes von Kakao die kognitiven Leistungen verbessert und die
Lebensdauer erhöht. Dies lässt vermuten, dass Kakao altersbedingte Gehirnstörungen und
neurodegenerative Erkrankungen möglicherweise verzögert [149].
1.7. Ziele der Arbeit
Das GABAerge System spielt eine sehr wichtige Rolle bei der Behandlung vieler
neurophysiologischer Erkrankungen. Einige Medikamentengruppen wie Sedativa, Anxiolytika,
Muskelrelaxantien, Antidepressiva und Antiepileptika (Antikonvulsiva) entfalten ihre Wirkung
über die GABAerge Neurotransmission. Darüber hinaus besteht ein großes Interesse daran,
Naturstoffe und vor allem Lebensmittelinhaltsstoffe zu finden, welche über Interaktionen mit
der GABAerge Neurotransmission im Organismus neurophysiologische Wirkungen hervorrufen.
Aus diesen Gründen wurden 13 Lebensmittel ausgewählt, für die in Tierversuchen oder
klinischen Studien beruhigende, angstlösende, sedative, antidepressive oder antiepileptische
Wirkungen beschrieben wurden und deren neurophysiologischer Wirkungsmechanismus noch
nicht vollständig aufgeklärt ist. Daher sollte im Rahmen dieser Arbeit untersucht werden, ob die
aus diesen ausgewählten Lebensmitteln hergestellten wässrigen Extrakte das GABAerge System
beeinflussen. Hierzu sollten im ersten Teil der vorliegenden Arbeit ein möglicher hemmender
Effekt dieser Lebensmittelextrakte sowie verschiedener exemplarisch ausgewählten
Lebensmittelinhaltsstoffe (Terpenoide und Flavanoide) auf die GABA-abbauenden Enzyme
getestet werden.
Im zweiten Teil der vorliegenden Studie sollten die möglichen dosisabhängigen Effekte der
Lebensmittelextrakte auf die durch GABA über GABAA-Rezeptoren vermittelten
Chloridionenströme untersucht werden. Die Voltage-Clamp Technik, in der die rekombinanten
humanen GABAA-Rezeptoren mit der Untereinheitenkombination von α1 und β2 in Oocyten von
Xenopus leavis exprimiert werden konnten, wurde zur Messung der GABA-induzierten Ströme
eingesetzt. Da GABA in vielen Lebensmitteln vorkommt, sollte weiter untersucht werden, ob die
getesteten wässrigen Lebensmittelextrakte GABA enthalten und ob deren GABA-Gehalt einen

Einleitung
21
elektrophysiologisch bedeutsamen Einfluss auf die beobachtete Stromantworte ausüben kann.
Weiterhin sollte ein Versuch mit dem Glycin-Rezeptor, der ein Chloridionenkanal ist und zur
Cys-Loop Superfamilie von ligandengesteuerten Ionenkanälen gehört, durchgeführt werden, um
zu prüfen, ob die allosterisch positiv modulierenden Effekte der Lebensmittelextrakte auf die
von GABAA-Rezeptoren gebildeten Chloridionenkanäle spezifisch ist. Hierzu wurde die Voltage-
Clamp Technik an mit rekombinanten α1 humanen Glycin-Rezeptoren transfizierten Oocyten
durchgeführt.
Das Ziel des dritten Teils dieser vorliegenden Arbeit war die Identifizierung der GABAergen
aktiven Komponenten im Nelkenextrakt, der im ersten Teil der Arbeit die Aktivität der GABA-
abbauenden Enzyme am stärksten hemmte und im zweiten Teil der Arbeit der wirksamste
spezifischen allosterische Modulator der GABAA-Rezeptoren war.

Einleitung
22

23
Inhibition des GABA-Abbaus 2
2.1. Einleitung
Im ersten Teil der Arbeit sollte eine mögliche hemmende Wirkung von Lebensmitteln und
Lebensmittelinhaltsstoffen auf die GABA metabolisierende Enzyme getestet werden, um die
positive Modulation des GABAergen Neurotransmittersystems durch eine Inhibition des GABA-
Abbaus erfassen zu können.
Die Anwesenheit des Neurotransmitters GABA im Gehirn wurde erstmals im Jahr 1950
beschrieben [25, 26]. GABA wird aus Glutamat unter Katalyse der Glutamat–Decarboxylase
(GAD; EC 4.1.1.15), welche ein Pyridoxalphosphat abhängiges Enzym ist [44] und zwei Isoformen
(65-kDA GAD und 67-kDA) im menschlichen Organismus besitzt [34], im Gehirn synthetisiert [25,
26, 33]. Ein niedriger GABA-Spiegel im Gehirn wird mit einigen neurophysiologischen
Krankheiten wie Epilepsie, Depressionen, Panikstörungen, Angststörungen oder Schlafstörungen
in Verbindung mitgebracht [150-156]. Wood et al. berichten in ihrer Humanstudie, dass
epileptische Patienten signifikant niedrige GABA-Konzentration im Liquor cerebrospinalis
(Gehirn-Rückenmarks-Flüssigkeit) zeigten [154]. Bei depressiven Patienten wurde ebenso eine
signifikant erniedrigte GABA-Konzentration im Gehirn nachgewiesen [155]. Da die Permeabilität
der Blut-Hirn-Schranke für GABA sehr gering ist [21, 157], kann die GABA-Supplementierung
nicht zu einer Erhöhung der GABA-Konzentration im Gehirn führen. So konnte beispielsweise
durch intraperitoneale Injektion von GABA keine Wirkung auf die GABA-Konzentration im
Gehirn von Ratten nachgewiesen werden [158]. Die Erhöhung der Konzentration von GABA wird
jedoch durch die Inhibition des GABA-Abbaus ermöglicht [159-162].
Der Abbau von GABA erfolgt über eine Transaminierungsreaktion, welche durch die GABA-
Transaminase (GABA-AT; EC 2.6.1.19) katalysiert wird. Mit Hilfe des Cosubstrates α-
Ketoglutarat, das aus dem Citratzyklus kommt, wird GABA zu Succinatsemialdehyd
umgewandelt, wobei gleichzeitig α-Ketoglutarat in Glutamat übergeführt wird [41-44]. Bei der
Transaminierungsreaktion dient das Pyridoxalphosphat (Vitamin B6) als Cofaktor [44]. Der
entstehende Succinatsemialdehyd wird in NAD(P) abhängigen Reaktionen entweder durch die
Succinatsemialdehyd-Dehydrogenase (SSA-DH; EC 1.2.1.24) zum Succinat oxidiert oder durch die
Succinatsemialdehyd-Reduktase (SSA-R; EC 1.1.1.61) zur γ-Hydroxybuttersäure reduziert [38-40,
163]. Das mitochondriale Enzym GABA-AT [164] bildet mit SSA-DH einen stabilen

Inhibition des GABA-Abbaus
24
Enzymkomplex, welcher sich in der Mitochondrienmatrix befindet [165]. Der Metabolismus von
GABA ist in Abbildung 2.1 dargestellt.
Abbildung 2.1 Schematische Überblick über Biosynthese und Abbau der Neurotransmitter GABA. GAD:
Glutamat-Decarboxlase; GABA-AT: GABA-Transaminase; SSA-DH: Succinatsemialdehyd-
Dehydrogenase; SSA-R: Succinatsemialdehyd-Reduktase (modifiziert nach Jakoby et al., Pearl
et al., Tsukatani et al. und Yogeeswari et al. [38, 39, 42, 163]).
Die GABA-AT ist eine Zielstruktur für Antiepileptika [159], welche den GABA-Spiegel im Gehirn
durch Inhibition der GABA-AT erhöhen können [159-162], womit eine antiepileptische bzw.
antikonvulsive Wirkung ausgelöst wird [160, 161]. Schlechter et al. bewiesen eine lineare
Assoziation zwischen dem Anstieg der GABA-Konzentration im Gehirn und der Inhibition der
GABA-AT sowie der Verstärkung der ausgelösten antikonvulsive Wirkung [160].

Inhibition des GABA-Abbaus
25
GABA-AT kann durch einige synthetische GABA Analoga wie Vigabatrin gehemmt werden [39,
160, 166-170]. Neben diesen synthetisierten Substanzen können einige Lebensmittel sowie
deren Inhaltsstoffe wie Terpenoide und Flavanoide auf GABA-AT hemmend wirken [110, 120,
171-173]. Zum Beispiel konnte die hemmende Wirkung von Hopfen, Melisse und Kamille auf
GABA-AT nachgewiesen werden [110]. Zudem wurde die GABAergene Wirkung von 34 Pflanzen,
welche antiepileptische und angstlösende Eigenschaften haben, in einer Studie untersucht.
Dabei wurde festgestellt, dass die Familie der Pfeffergewächse die stärkste Inaktivierung der
GABA-AT zeigt [120].
Wie in Kapitel 1.6 beschrieben, wurden in zahlreichen in vivo Studien die neurophysiologischen
Wirkungen einiger Lebensmitteln gezeigt. Jedoch sind sehr wenige Studien vorhanden, die den
Wirkungsmechanismus untersuchen und die Wirkungen der Lebensmittel miteinander
vergleichen. Möglicherweise können sie ihre beruhigende, sedative, antiepileptische,
antidepressive oder angstlösende Wirkung über einen inhibierenden Einfluss auf die GABA-AT
ausüben. Daher sollte im Rahmen dieser Studie der Einfluss der 13 ausgewählten Lebensmittel,
für die bereits ein sedierender, angstlösender oder antikonvulsiver Effekt bekannt ist, auf GABA-
metabolisierende Enzyme (GABA-AT und SSA-DH) untersucht werden. Bei den Testextrakten
handelte es sich um Salbei, Nelken, Grüner Tee, Lavendel, Kamille, Melisse, Hopfen, Sideritis-
Arten, Kakao und Schwarzer Pfeffer und sowie 40 weitere Lebensmittelinhaltsstoffe. Dazu
wurde ein Enzymassay, welcher die Messung der Inaktivierung von GABA- metabolisierenden
Enzyme (GABA-AT und SSA-DH) durch Testsubstanzen erlaubt, verwendet.
2.2. Bestimmung der Aktivität von GABA-metabolisierenden Enzyme (GABA-AT und SSA-DH)
Um die Aktivität der GABA-metabolisierenden Enzyme zu bestimmen bzw. eine hemmende
Wirkung auf GABA-metabolisierende Enzyme nachzuweisen, wurden einige Methoden in der
Literatur beschrieben [42, 43, 110, 163, 166, 171, 174, 175]. In dieser vorliegenden Arbeit
erfolgte die Messung der hemmenden Wirkung von Testsubstanzen auf GABA-
metabolisierenden Enzymen (GABA-AT und SSA-DH) durch einen Enzymassay, der nach Jacoby
et al. modifiziert wurde [42]. Dieser Enzymassay basiert auf der photometrischen Bestimmung
der Umsetzung des Coenzyms NADP+ beim enzymatischen Abbau von GABA. Wie in Abbildung
2.1 dargestellt ist, wird Succinatsemialdehyd, der beim Abbau von GABA entsteht, von SSA-DH
zum Succinat oxidiert, wobei gleichzeitig NADP+ zu NADPH/H+ reduziert wird. Das entstehende
NADPH/H+ kann photometrisch bei 340 nm bestimmt werden [43]. Die Enzymaktivität kann
dann anhand der umgesetzten NADPH/H+ Menge berechnet werden [163]. In Anwesenheit
eines Inhibitors wird die umgesetzte NADPH/H+ Menge bzw. die gemessene UV/Vis-Absorption
vermindert.

Inhibition des GABA-Abbaus
26
Zur Erfassung einer inhibierenden Wirkung der Testsubstanzen wurden die verwendeten
Lösungsmittel (Wasser, Ethanol oder DMSO) bei jedem Versuch als Negativkontrolle eingesetzt.
Die Absorption zum Zeitpunkt von 0 min wurde von jedem Messwert der Absorption, welcher
nach 30-minütiger Inkubation ermittelt wurde, abgezogen. Die Abweichung der Absorption von
diesem Startwert entsprach einer Enzymaktivität von 100 % für die Negativkontrolle. Jede
Reduktion der Messwerte in Anwesenheit der Testsubstanzen bzw. Lebensmittelextrakte wurde
auf die jeweilige Negativkontrollwert (100 %) bezogen, und dadurch die inhibitorische Wirkung
ermittelt. Alle Versuche wurden als Triplikate durchgeführt. Die durchschnittliche inhibitorische
Wirkung jeder Testsubstanz wurde gegen die Konzentration aufgetragen, wodurch eine Dosis-
Wirkungskurve ermittelt wurde. Dabei wurde der IC50 Wert berechnet, der einer Konzentration
des Inhibitors entspricht, welche die Enzymaktivität um 50 % hemmt.
2.3. Einfluss von Vigabatrin auf die Aktivität der GABA-metabolisierenden Enzyme (GABA-AT und SSA-DH)
Vigabatrin (Abbildung 2.5), welches zur Erhöhung der GABA-Menge im ZNS entwickelt wurde
[39], bindet als Inhibitor selektiv [166] und irreversibel [39, 160, 166, 167] an der aktiven Stelle
der GABA-AT. Hier wurde Vigabatrin zur Bestimmung seiner hemmenden Wirkung auf GABA-
metabolisierenden Enzymen in den unterschiedlichen Konzentrationen von 0,09; 0,46; 0,92;
1,37; 1,83; und 2.29 mM im Enzymassay eingesetzt. Für die Negativkontrolle wurde Wasser
anstelle des Inhibitors eingesetzt. Die Absorption wurde nach einer Inkubationszeit von 30 und
60 Minuten bei 340 nm photometrisch gemessen. Abbildung 2.2 stellt die dosisabhängige
Wirkung von Vigabatrin auf die Enzymaktivität nach verschiedenen Inkubationszeiten dar.

Inhibition des GABA-Abbaus
27
Abbildung 2.2 Konzentrationsabhängige Wirkung von Vigabatrin auf die Aktivität der GABA-
metabolisierenden Enzyme. Die photometrische Messung erfolgte bei 340 nm nach einer
Inkubationszeit von 30 und 60 Minuten bei 37 °C. Die Daten weisen den Mittelwert ±
Standardabweichung aus drei unabhängigen Messungen auf. ns : nicht signifikanter
Unterschied zwischen Messwerten nach 30 –minütiger und 60 –minütiger Inkubationszeit;
**** (p< 0,0001) signifikanter Unterschied zwischen Negativkontrolle und Aktivität des
Vigabatrin nach einer Inkubationszeit von 30 Minuten bei 37 °C. **** (p< 0,0001)
signifikanter Unterschied zwischen Negativkontrolle und Aktivität des Vigabatrin nach einer
Inkubationszeit von 60 Minuten bei 37 °C, bestimmt durch einen zweifaktoriellen ANOVA
Holm-Sidak Vergleichstest.
Bei der niedrigsten eingesetzten Konzentration löste Vigabatrin einen Aktivitätsverlust des
Enzyms von ca. 30 % aus, während bei der Konzentration von 0,46 mM, ab welcher eine
schwächere Abnahme der Enzymaktivität festgestellt wurde, die GABA-AT nur noch zu 28,2 % ±
8,7 nach 30-minütiger Inkubation und 27,3 % ± 7,4 nach einer Inkubationszeit von 60 Minuten
aktiv war. Bis zu einer Vigabatrin-Konzentration von 1,83 mM nahm die Enzymaktivität mit
steigender Vigabatrin-Konzentration ab, so dass für diese Konzentration die größte
Enzyminhibition ermittelt werden konnte. Wurde die Konzentration weiter erhöht, fand keine
weitere Reduzierung der Enzymaktivität statt, sowohl nach der Inkubation von 30 Minuten als
auch nach 60 minütiger Inkubation. Dies deutet darauf hin, dass die GABA-AT vollständig
inhibiert wurde. Der IC50-Wert von Vigabatrin wurde als 0,192 (0,38-0,097) mM (entspricht
0,025 (0,049-0,013) mg/mL) (95%-Konfidenzintervall) berechnet.
Da kein signifikanter Unterschied zwischen 30- und 60-minütiger Inkubationszeit beim Verlauf
der Aktivitätsabnahme des Enzyms auftrat, war eine Inkubationszeit von 30 Minuten zur
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
80,00
0,00 0,50 1,00 1,50 2,00 2,50
Akt
ivit
ät [
%]
Konzentration von Vigabatrin [mM]
nach 30 min Inkubation nach 60 min Inkubation
ns **** ****
ns **** ****
ns **** **** ns
**** ****
ns **** ****
ns **** ****

Inhibition des GABA-Abbaus
28
weitergehenden Erfassung der Enzyminhibition ausreichend. Daher erfolgte bei den weiteren
Versuchen mit Lebensmittelextrakte und Lebensmittelinhaltsstoffe die Absorptionsmessung
nach einer 30-minütigen Inkubation.
2.4. Einfluss der Lebensmittelextrakte auf die Aktivität der GABA-metabolisierenden Enzyme (GABA-AT und SSA-DH)
Anschließend wurde die inhibitorische Wirkung der wässrigen Lebensmittelextrakte mit Hilfe
des Enzymassays bestimmt. Als Negativkontrolle wurde Wasser an Stelle des
Lebensmittelextraktes eingesetzt. Wegen seiner nachgewiesenen inhibitorischen Wirkung
diente Vigabatrin als Positivkontrolle. Mit Hilfe des Enzymassays konnte gezeigt werden, dass
sich die wässrigen Extrakte von Hopfen, Kakao und schwarzem Pfeffer nicht auf die
Enzymaktivität auswirkten (Abbildung 2.3). Die aus Nelken, grünem Tee, Salbei, und einigen
Sideritis-Arten (S. condensata,S. sipylea und S. stricta) gewonnenen Extrakte wurden in
Konzentrationen von 0,02; 0,04; 0,05; 0,07 und 0,09 mg/mL eingesetzt. Wegen seiner stark
hemmenden Wirkung konnte S.arguta zusätzlich in noch geringerer Konzentration (0,01 mg/mL)
verwendet werden. Die Extrakte von Melisse, Lavendel und Kamille wurden noch in größeren
Konzentrationen von mehr als 0.09 mg/mL eingesetzt, um eine Inhibierung der GABA
metabolisierende Enzyme von mehr als 50 % hervorzurufen. Die Abbildung 2.4 stellt die
Veränderung der Enzymaktivität durch Zugabe der Lebensmittelextrakte dar.
Abbildung 2.3 Einfluss unterschiedlicher Konzentrationen an schwarzer Pfeffer, Hopfen und Kakao auf die
Aktivität der GABA-metabolisierenden Enzyme. Die photometrische Messung erfolgte bei
340 nm nach einer 30-minutiger Inkubation bei 37 °C. Die Daten stellen den Mittelwert aus
zwei unabhängigen Messungen dar.
0,0
20,0
40,0
60,0
80,0
100,0
120,0
0,00 0,03 0,06 0,09 0,12 0,15 0,18
Akt
ivit
ät [
%]
Konzentration [mg/mL]
Schwarzer Pfeffer Hopfen Kakao

Inhibition des GABA-Abbaus
29
Abbildung 2.4 Aktivität der GABA-metabolisierenden Enzyme in Abhängigkeit von der Konzentration der
Extrakte von (A) Nelken und verschiedenen Sideritis-Arten (S. condensata, S. arguta, S. stricta
und S. sipylea) (B) Grünem Tee und Salbei (C) Kamille (D) Lavendel und (E) Melisse. Die
photometrische Messung erfolgte bei 340 nm nach einer 30-minutiger Inkubation bei 37 °C.
Die Daten stellen den Mittelwert ± Standardabweichung aus drei unabhängigen Messungen
dar.
0,0
20,0
40,0
60,0
80,0
100,0
0,00 0,02 0,04 0,06 0,08 0,10
Akt
ivit
ät [
%]
Konzentration[mg/mL]
S. arguta Nelken S.strictica S. sipylea S. condensataA)
0,0
20,0
40,0
60,0
80,0
100,0
0,00 0,02 0,04 0,06 0,08 0,10
Akt
ivit
ät [
%]
Konzentration[mg/mL]
Grüner tee SalbeiteeB)
0,0
20,0
40,0
60,0
80,0
100,0
0,00 0,05 0,10 0,15 0,20
Akt
ivit
ät [
%]
Konzentration[mg/mL]
KamilleC)
0,0
20,0
40,0
60,0
80,0
100,0
0,00 0,05 0,10 0,15 0,20
Akt
ivit
ät [
%]
Konzentration[mg/mL]
LavendelD)
0,0
20,0
40,0
60,0
80,0
100,0
0,00 0,05 0,10 0,15 0,20
Akt
ivit
ät [
%]
Konzentration[mg/mL]
MelissaE)

Inhibition des GABA-Abbaus
30
Es ist hier deutlich zu erkennen, dass die Aktivität der GABA-metabolisierenden Enzyme mit
steigender Konzentration der Extrakte abnahm. Die ermittelten Daten wurden durch
zweifaktorielle ANOVA mit Holm-Sidak Vergleichstest statistisch ausgewertet. Dabei wurde
festgestellt, dass Lebensmittelextrakte bei allen hier eingesetzten Konzentrationen die Aktivität
der GABA-abbauenden Enzyme im Vergleich zur Negativkontrolle signifikant (p<0,01) inhibierten
(Tabelle im Anhang A.1). Die höchste Hemmung der Enzyme konnte durch Nelkenextrakt bei
einer Konzentration von 0,09 mg/mL mit einer Restaktivität von lediglich 14,7 ± 8,9 % und durch
Extrakt von S.arguta bei einer Konzentration von 0,07 mg/mL mit einer Restaktivität 20,2 ± 3,5
% erreicht werden (Abildung 2.4.A und Tabelle im Anhang A.1). Dabei verursachten die Extrakte
von Nelken und S.arguta eine signifikant stärkere Enzymhemmung als die Extrakte von S. stricta,
S. condensata, S. sipylea, Salbei und grünem Tee bei den gleichen Konzentrationen (Abbildung
2.4.A und 2.4.B sowie Tabelle im Anhang A.1). Obwohl Melisse, Lavendel und Kamille in höheren
Konzentrationen als Nelken und S. arguta eingesetzt wurden, konnten sie die Enzymaktivität
nicht so stark inhibieren wie die beiden letztgenannten Extrakte. Außerdem unterschied sich die
durch Nelken ausgelöste Hemmung der Enzyme nicht signifikant von der durch S.arguta,
welches bei denselben Konzentrationen eingesetzt wurde (Abbildung 2.4.A und Tabelle im
Anhang A.1). Ebenso wurde kein signifikanter Unterschied zwischen der Inhibition des
Enzymsystems, welche durch Zugabe von drei Sideritis-Arten -S.strictica, S.condensata, und S.
sipylea -ausgelöst wurde, beobachtet (siehe Abbildung 2.4.A und Tabelle im Anhang A.1).
Ebenfalls war kein signifikanter Unterschied in der Hemmung der Enzymaktivität zwischen
grünem Tee und Salbei sowie S. stricta festzustellen (siehe Abbildung 2.4.B und Tabelle im
Anhang A.1). S. sipylea sowie S. condensata zeigten lediglich bei einer Einsatzkonzentration von
0,07 mg/mL eine signifikante (p<0,05) unterschiedliche Inhibition im Vergleich zu Salbei (siehe
Tabelle im Anhang A.1).
In Tabelle 2.1 sind die berechneten IC50-Werte der Lebensmittelextrakte mit Konfidenzintervall
(CI) zusammengestellt. Die benötigte Konzentration, bei der 50% der Aktivität des Enzyms
inhibiert werden kann, war für Nelken und S.arguta ähnlich klein wie die der Positivkontrolle
Vigabatrin (0,022 mg/mL für S.arguta; 0,023 mg/ mL für Nelken und 0,025 mg/mL für
Vigabatrin). Grüner Tee, Sideritis-Arten- S.stricta, S. sipylea und S.condensata zeigten IC50-Werte
zwischen 0,05 und 0,08 mg/mL. Dagegen lag der IC50-Wert für die Extrakte von Lavendel,
Melisse und Kamille über 0,10 mg/mL, wobei Kamille mit einen IC50-Wert von 0,166 mg/mL der
schwächste Inhibitor war.

Inhibition des GABA-Abbaus
31
Tabelle 2.1 IC50-Werte verschiedener Lebensmittelextrakten für GABA-metabolisierende Enzyme. Die
Werte sind angegeben als mg/mL mit 95%-Konfidenzintervall (CI).
Lebensmittel IC50 [mg/mL] (95% CI)
Sideritis arguta 0,022 (0,024-0,020)
Nelken 0,023 (0,034-0,016)
Grüner Tee 0,051 (0,056-0,039)
Siderititis stricta 0,06 (0,075-0,048)
Siderititis sipylea 0,065 (0,072-0,056)
Salbei 0,066 (0,084-0,051)
Sideritis condensata 0,082 (0,105-0,063)
Lavendel 0,107 (0,125-0,092)
Melisse 0,130 (0,163-0,105)
Kamille 0,166 (0,21-0,132)
Kontrolle; Vigabatrin 0,025 (0,049-0,013)
2.5. Einfluss ausgewählter Lebensmittelinhaltsstoffe auf die Aktivität der GABA-metabolisierenden Enzyme (GABA-AT und SSA-DH)
Einige terpenoide Inhaltsstoffe von Lebensmitteln wurden als Inhibitor der GABA-Abbauenzyme
nachgewiesen [172]. Neben Terpenoiden konnten auch Flavanoide die Aktivität der GABA
abbauenden Enzyme hemmen [171]. Daher wurde ein möglicher Effekt verschiedener
exemplarisch ausgewählter Terpenoide und Flavanoide auf das Enzymsystem getestet. Nur 4-
Hdroxybenzaldehyd konnte im Wasser gelöst werden. β-Damascenon, p-Coumarinsäure und
Ferulasäure wurden in Ethanol verdünnt, während die anderen Stoffe in DMSO gelöst wurden.
Zunächst wurden die organischen Lösungsmittel DMSO und Ethanol in verschiedenen
Konzentrationen eingesetzt, um einen Effekt der Lösungsmittel ausschließen zu können. Die
verwendeten Endkonzentrationen betrugen 0,1 %; 0,5 %; 1 % und 2 %, wobei durch die
Lösungsmittel kein Effekt auf die GABA abbauende Enzyme festgestellt wurde. Anschließend
wurde ein Vorversuch durchgeführt, bei dem alle Substanzen in einer höchsten Konzentration
von 1 mM eingesetzt wurden. Bei jedem Versuch wurden die verwendeten Lösungsmittel
(Wasser, Ethanol oder DMSO) als Negativkontrolle eingesetzt. In der folgenden Tabelle ist die
Aktivität der Enzyme in Anwesenheit der Lebensmittelinhaltsstoffe in einer Konzentration von 1
mM zusammengestellt.

Inhibition des GABA-Abbaus
32
Tabelle 2. 2 Aktivität der GABA-metabolisierenden Enzyme (GABA-AT und SSA-DH) in Anwesenheit von 1
mM Testsubstanzen. Die photometrische Messung erfolgte bei 340 nm nach einer
Inkubationszeit von 30 Minuten bei 37 °C.
Komponente [1 mM] Aktivität [%] Komponente [1 mM] Aktivität [%]
β-Caryophyllen 73,5 Theaspiran 61,0
Sabinen 72,3 β-Ionon 46,8
(-)-Caryophyllenoxid 45,3 α-Ionon 85,2
R -(+)-Limonen 92,2 Isopulegol 71,8
Citral 98,9 α-Thujon 75,1
Myrcen 43,6 Pinocarveol 85,6
Menthol 97,5 β-Damascenon 28,2
4-Ethylguajacol 97,5 Nerolidol 87,3
Eucalyptol 95,3 2-cis,4-trans-Abscisinsäure 73,7
α-Bisabolol 95,1 Cedrol 72,7
α-Santonin 101,6 Nootkaton 91,1
Ocimen 69,9 2-Methyl-3-buten-2-ol 89,2
β -Citronellol 93,9 3-Methyl-2-buten-1-ol 67,5
Terpinolen 73,2 Rosenoxid 80,1
Carveol 101,5 R- Carvon 94,6
Perillylalkohol 97,6 S- Carvon 87,0
Myrtenol 72,0 Ferulasäure 96,9
Borneol 99,4 109,7
4-Hdroxybenzaldehyd Apigenin
74,9
1-R-Fenchon 96,4
β -Farnesen 89,7 p-Coumarinsäure 99,5
Kontrolle; Vigabatrin [0,25 mM] 33,6
Wie Tabelle 2.2 zu entnehmen ist, bewirkte β-Damascenon die stärkste Inaktivierung der
Enzyme. Neben β-Damascenon verursachten β-Ionon, Myrcen und (-)-Caryophyllenoxid einen
Aktivitätsverlust der Enzyme von mehr als 50%. Daraufhin wurden diese Komponenten in
verschiedenen Konzentrationen getestet, um zu untersuchen, ob es sich um einen
konzentrationsabhängigen Effekt handelt. Die Messergebnisse werden in der nachstehenden
Tabelle dargestellt.

Inhibition des GABA-Abbaus
33
Tabelle 2.3 Konzentrationsabhängige Wirkung von β-Damascenon, Myrcen, β-Ionon und (-)-
Caryophyllenoxid auf die GABA-metabolisierende Enzyme (GABA-AT und SSA-DH). Die
photometrische Messung erfolgte bei 340 nm nach einer 30-minutiger Inkubation bei 37 °C.
Die Daten weisen den Mittelwert ± Standartabweichung aus drei unabhängigen Messungen
auf.
β-Damas-
cenon
Konzentration [mM] 0,20 0,40 0,60 0,80 1,0
Aktivität [%] 74,6 ± 3,7 72,2 ± 4,1 59,2 ± 2,4 40,5 ± 9,7 27,1 ± 6,3
Signifikanzwert **** **** **** **** ****
Myrcen
Konzentration [mM] 0,04 0,1 0,5 0,75 1,0
Aktivität [%] 89,9 ± 4,3 80,8 ± 5,9 64,3 ± 2,1 51,1 ± 3,1 43,6 ± 8,5
Signifikanzwert ns ** **** **** ****
β-Ionon
Konzentration [mM] 0,1 0,25 0,5 0,75 1,0
Aktivität [%] 66,0 ± 9,6 65,3 ± 4,8 55,7 ± 6,7 51,4 ± 3,7 46,8 ± 4,6
Signifikanzwert **** **** **** **** ****
(-)-
Caryophyl-
lenoxid
Konzentration [mM] 0,04 0,1 0,5 0,75 1,0
Aktivität [%] 75,3 ± 7,1 73,2 ± 3,3 65,8 ± 10 57,3 ± 10,2 47,5 ± 7,8
Signifikanzwert *** **** **** **** ****
ns : nicht signifikanter Unterschied, ** p< 0,01, **** p< 0,0001 signifikanter Unterschied im Vergleich zu
Negativkontrolle; zweifaktorieller ANOVA Holm-Sidak Vergleichstest.
Wie aus Tabelle 2.3 deutlich wird, verursachten die getesteten Komponenten eine
konzentrationsabhängige Hemmung der GABA-metabolisierenden Enzyme, wobei sich die
Enzymaktivität mit steigenden Konzentrationen der Substanzen verringerte. Bei der höchsten
Konzentration von 1 mM zeigte β-Damascenon die stärkste Hemmung der Enzyme, welche sich
mit einer Restaktivität von 27,1 ± 6,3 % hoch signifikant (p< 0,0001) von der Negativkontrolle
unterschied. Myrcen zeigte eine signifikante Abnahme der Aktivität der GABA-
metabolisierenden Enzyme erst bei Konzentrationen ≥ 0,1 mM. Die Enzyminaktivierung war in
Anwesenheit von β-Damascenon und Myrcen stark konzentrationsabhängig. Dagegen führten β-
Ionon und (-)-Caryophyllenoxid zu einem leichten konzentrationsabhängigen Abfall der Aktivität
der GABA-metabolisierenden Enzyme in den Konzentrationen kleiner als 0,5 mM.
Die Tabelle 2.4 stellt die IC50 -Werte der getesteten Komponenten dar. β-Damascenon hemmte
am stärksten, da es einen kleineren IC50-Wert von 0,573 mM als die anderen Komponenten

Inhibition des GABA-Abbaus
34
aufwies. Allerdings war der IC50-Wert fast 3-fach größer im Vergleich zu dem von Vigabatrin
(0,192 mM). Myrcen zeigte einen ähnlichen IC50-Wert wie β-Ionen, während für (-)-
Caryophyllenoxid ein höherer IC50-Wert berechnet wurde.
Tabelle 2.4 IC50-Werte verschiedener Testsubstanzen für GABA-metabolisierende Enzyme (GABA-AT und
SSA-DH). Die Werte sind angegeben als mM mit 95%-Konfidenzintervall (CI).
Komponente IC50 [mM] (95% CI)
β-Damascenon 0,573 (0,681-0,466)
Myrcen 0,819 (1,067-0,570)
β-Ionon 0,820 (0,940-0,705)
(-)-Caryophyllenoxid 0,977 (1,271 - 0, 683)
Kontrolle; Vigabatrin 0,192 (0,38-0,097)
Abbildung 2.5 Strukturen von Vigabatrin, β-Damascenon, β-Ionon, (-)-Caryophyllenoxid und Myrcen.

Inhibition des GABA-Abbaus
35
2.6. Diskussion und Zusammenfassung
Vigabatrin, welches zur Erhöhung der GABA-Menge im ZNS entwickelt wurde [39], wird als
Medikament zur Behandlung von Epilepsie verwendet [161, 162]. Vigabatrin bindet selektiv
[166] und irreversibel [39, 160, 166, 167] an der aktiven Stelle der GABA-AT und hemmt somit
deren Aktivität [39, 159-162, 166, 167] , wodurch das GABA-Spiel im Gehirn erhöht wird [161,
162].
Vigabatrin, das hier als Positivkontrolle verwendet wurde, ist in der Lage die Konzentration von
GABA im Gehirn zu erhöhen [161, 162], da Vigabatrin als ein Strukturanalog der GABA die Blut-
Hirn-Schranke überwinden und die GABA-AT inhibieren kann [174]. Im Tierversuch untersuchte
Coelha et al. die antiepileptogene Wirkung des Vigabatrins. Dabei stellten sie fest, dass
Vigabatrin das Auftreten der Konvulsionen vermindert [176]. Bei Ratten zeigte Vigabatrin in der
Konzentration von 50 mg/kg einen angstlösenden Effekt, welcher mittels des elevated plus maze
Test bestimmt wurde [167]. Auch in Humanstudien wurde die Wirkung von Vigabatrin auf
epileptische Anfälle untersucht. Dabei wurde eine Reduktion der Anfallshäufigkeit (um 50 %
oder mehr) bei Epilepsiepatienten durch Gabe von Vigabatrin beobachtet [177]. Allerdings
können visuelle Nebenwirkungen bei der Anwendung des Vigabatrins auftreten, welche im
Tierversuch durch die Abnahme der Potentiale im Elektroretinogramm identifiziert wurden
[178]. Darüber hinaus verursachte Vigabatrin eine Verringerung der Sehschärfe und abnormales
Farbsehen in Humanstudien [179]. Aufgrund dieser Nebenwirkungen von Vigabatrin wurden
bereits einige GABA-Analoge zur Inaktivierung der GABA-AT synthetisiert [168]. Zum Beispiel hat
das GABA-Analog (2S, 4S)-4-Aminotetrahydrothiophen-2-carbonsäure Hydrochlorid eine 8-fach
höhere hemmende Wirkung auf GABA-AT als Vigabatrin [168]. Ebenso zeigte die synthetische
(1S,3S)-3-Amino-4-difluoremethylenyl-1-cyclopentansäure im Vergleich zu Vigabatrin eine
höhere Inaktivierung der GABA-AT [169]. Zudem zeigten weitere Vigabatrin-Analoga - cis-3-
Aminocyclohex-4-ene-1-carbonsäure und cis-2-Aminocyclohex-3-ene-1- carbonsäure - eine
irreversible konzentrationsabhängige Inaktivierung der GABA-AT [170].
Obwohl es synthetisierte Substanzen gibt, die eine Inhibition auf GABA-AT hervorrufen, gibt es
auch ein großes Interesse an Naturprodukten bzw. Lebensmitteln, welche in der Lage sind, die
Aktivität des GABA-abbauenden Enzyms zu hemmen. In der vorliegenden Arbeit wurde die
enzymhemmende Wirkung von Lebensmittelextrakten untersucht. Es wurde festgestellt, dass
alle getesteten Lebensmittelextrakte außer Hopfen, Kakao und schwarzem Pfeffer inhibitorisch
wirkten. Die Enzyminaktivierung erfolgte dabei konzentrationsabhängig. Extrakte von Nelken
und S.arguta hemmten am stärksten, wobei die Extrakte einen ähnlichen IC50-Wert zeigten wie
Vigabatrin. Zudem wurde eine ähnlich stärke enzymhemmende Wirkung nicht nur für drei

Inhibition des GABA-Abbaus
36
Sideritis-Arten (S. stricta, S. sipylea und S.condensata), sondern auch für Salbei und Grüner Tee
gefunden.
Awad et al. untersuchten den enzymhemmenden Effekt einiger Pflanzen, welche aufgrund
angstlösender Wirkung in der Volksmedizin verwendet werden und ermittelten den wässrigen
Melisse-Extrakt als den stärksten Inhibitor der GABA- AT mit einem IC50-Wert von 0,35 mg/mL
[110]. Andererseits zeigte der wässrige Melisse-Extrakt in dieser Arbeit eine schwächere
inhibierende Wirkung mit einem IC50-Wert von 0,130 (0,163-0,105) mg/mL (95% CI) (siehe
Tabelle 2.1) auf die GABA-Abbauenzyme im Vergleich zu Sideritis Arten, Grüner Tee, Nelken,
Lavendel, und Salbeitee. Somit könnten diese Lebensmittelextrakte stärkere Inhibitoren der
GABA-AT als die in der Literatur beschriebenen natürlichen Inhibitoren sein. In einer weiteren
Studie wurde ein IC50-Wert des wässrigen Melisse-Extraktes von 0,85 mg/mL ermittelt [172]. Der
Unterschied zwischen diesen in der Literatur angegebenen IC50-Werte und dem in dieser Arbeit
ermittelten IC50-Wert kann möglicherweise an den Extraktionsmethode liegen, da die aktivsten
Komponenten in Abhängigkeit von verschiedenen Parametern wie beispielsweise
Extraktionszeit oder -temperatur in unterschiedlichen Mengen extrahiert werden. Als aktivste
Komponente von Melisse wurden schon die Hauptkomponente Rosmarinsäure sowie die
Triterpenoide Ursolsäure und Oleanolsäure identifiziert [172]. Rosmarinsäure wurde auch in
Salbei und Lavendel nachgewiesen [180]. In einem Kindling-Epilepsiemodell an Mäusen wurde
die Wirkung der Rosmarinsäure auf die durch die elektrische Stimulation verursachte
epileptische Anfälle untersucht. Dabei wurde festgestellt, dass Rosmarinsäure die Häufigkeit der
klonischen Anfälle verringerte und die Latenzzeit für die erste klonische Konvulsionen erhöhte
[176].
In der Literatur wurde die enzymhemmende Wirkung von wässrigem Kamillenextrakt im
Vergleich zu der von Melisse als schwächer eingestuft, da für Kamille ein größerer IC50-Wert als
für Melisse ermittelt wurde [172]. Dies stimmt mit der in dieser Arbeit ermittelte Ergebnisse
überein, da der wässrige Extrakt von Kamille die schwächste Inhibition der GABA-AT hervorrief.
Der wässrige Extrakt von Hopfen wurde von Awad et al. als ein Inhibitor identifiziert [110].
Allerdings zeigte der wässrige Hopfenextrakt, welcher bis 0,2 mg/mL eingesetzt wurde, in der
vorliegenden Arbeit keine hemmende Wirkung auf GABA-AT. In der Literatur wurde für Hopfen
ein IC50-Wert von 1,62 mg/mL beschrieben [110], so dass man erst bei höheren als der hier
eingesetzte Konzentrationen eine Wirkung erwarten würde. Zudem zeigte ein ethanolischer
Extrakt eine stärkere Inaktivierung der GABA-AT als der wässrige Hopfenextrakt [110].
In dieser Arbeit inhibierte der wässrige Extrakt von schwarzem Pfeffer das Enzymsystem nicht.
Der wässrige Extrakt von Kava (Piper methysticum), einer Pflanzenart, die ebenso zur Familie der
Pfeffergewächse gehört wie der schwarze Pfeffer, wurde bereits als ein schwacher Inhibitor der

Inhibition des GABA-Abbaus
37
GABA-AT identifiziert, der erst bei einer hohen Konzentration ( > 4 mg/mL) eine hemmende
Wirkung hervorruft [110]. Dagegen zeigte eine Studie, bei der die ethanolischen Extrakte von 34
Pflanzen mit antiepileptischen und angstlösenden Eigenschaften untersucht wurden, die
stärkste Inaktivierung der GABA-AT durch Vertreter der Familie der Pfeffergewächse [120].
Neben den Extrakten von Hopfen und schwarzem Pfeffer zeigte auch der wässrige Kakaoextrakt
keinen inhibierenden Effekt auf die GABA-AT. Dies kann darauf hindeuten, dass die Aktivität der
GABA-AT durch Kakao nicht gehemmt werden kann oder dass die eingesetzte Konzentrationen
zu niedrig waren. Möglicherweise sind aktive Komponenten nicht im wässrigen Extrakt löslich.
Da mittels der in dieser Arbeit verwendeten Methode nur das bei der Metabolisierung von
GABA-und Succinatsemialdehyd umgesetzte NADPH/H+ gemessen werden kann und dabei nicht
zwischen der Hemmung von GABA-AT und der Inaktivierung von SSA-DH unterschieden werden
kann, stellt sich die Frage nach den genauen Wirkungsmechanismus der untersuchten
Lebensmittel. Neben GABA-AT könnten diese Lebensmittelextrakte vielleicht auch die SSA-DH
hemmen. Es wurde schon gezeigt, dass einige Pflanzenextrakte wie Erigeron breviscapus beide
Enzyme inhibierten [171].
In einer in-vivo Studie wurde der schlaffördernde Effekt der ätherischen Öle verschiedener
Pflanzen sowie ihr Wirkungsmechanismus untersucht. Dabei verursachten Baldrian (Valeriana
officinalis) und Rosen (Rosa damascena) bei Ratten eine längere Schlafdauer. Zudem zeigte
Baldrian eine Inhibition von GABA-AT, wohingegen keine Veränderung der Enzymaktivität bei
der Applikation des ätherischen Öls von Rosen ermittelt wurde [181]. Acorus gramineus,
welches aufgrund seiner sedativen und schmerzlindernden Effekte im koreanischen Arzneibuch
gelistet ist, zeigte eine Erhöhung des GABA-Spiegels und eine Verminderung des Glutamat-
Spiegels im Gehirn der Mäuse durch die Inhibition der GABA-AT und zusätzlich antiepileptische
und sedative Wirkung [182]. Das ätherische Öl von SuHeXiang Wan, das zur Behandlung von
Epilepsie in der chinesischen Volksmedizin verwendet wird, inhibierte ebenso die GABA-AT,
erhöhte damit den GABA-Spiegel und verminderte den Glutamat-Spiegel im Mäusegehirn und
wirkte dadurch sedativ und antikonvulsiv [183]. Dieser antiepileptische Effekt kann durch
Inhibition der GABA-AT verursacht werden, da die Konvulsion teilweise von der GABA-
Konzentration im Gehirn abhängt [182, 183]. Aufgrund dieses Zusammenhangs zwischen
antiepileptischem Effekt und Inhibition der GABA-AT könnten die Extrakte, welche sich in dieser
Arbeit als Inhibitor der GABA-AT erwiesen, einen antiepileptischen Effekt verursachen, sofern
die aktiven Komponenten dieser Extrakte die Blut-Hirn-Schranke überwinden können.
Weiterhin wurde bereits die Modulation der GABAerge Wirkung durch definierte
Lebensmittelinhaltsstoffe untersucht. Beispielsweise wurden die Triterpenoide Ursolsäure und
Oleanolsäure als Inhibitor der GABA-Abbauenzyme identifiziert [172]. Ebenso wurde bereits

Inhibition des GABA-Abbaus
38
festgestellt, dass einige Flavanoide wie Baicalein, Scutellarein, Scutellarin, Quercetin,
Kaempferol, Rutin sowohl die GABA-AT und als auch die SSA-DH inhibieren konnten [171]. Daher
wurden in dieser Arbeit die Wirkung verschiedener, exemplarisch ausgewählter Terpenoide und
einiger Flavanoide auf das Enzymsystem getestet. Dabei wurden β-Damascenon, Myrcen, β-
Ionon und (-)-Caryophyllenoxid als Inhibitoren identifiziert. 4-Hydroxybenzaldehyd hemmte bei
der höchsten eingesetzten Konzentration von 1 mM lediglich 25% der Aktivität der GABA-
metabolisierende Enzyme (GABA-AT und SSA-DH). Allerdings wurde von Tao et al. festgestellt,
dass 4-Hydroxybenzaldehyd und auch sein Strukturanalog 4-Hydroxybenzylamine kompetitive
Inhibitoren sowohl der GABA-AT als auch der SSA-DH waren. Ihre ähnliche Molekularstruktur,
der Benzolring, könnte diese kompetitive Inhibition verursachen, da der Benzolring von aktivem
Zentrum der Enzyme akzeptiert werden könnte [173].
In dieser Arbeit wurde β-Damascenon als aktivster Inhibitor des Enzymsystems bestimmt mit
einen IC50 -Wert von 0,573 (0,681-0,466) mM (Tabelle 2.4). Dagegen erwies sich β-Damascenon
nur als ein schwacher Inhibitor im Vergleich zu Vigabatrin, da sein IC50-Wert fast 3-fach höher als
der von Vigabatrin (IC50-Wert Vigabatrin: 0,192 (0,38-0,097) mM) war. Neben β-Damascenon
zeigten auch seine Strukturanaloga β-Ionon, Myrcen und (-)-Caryophyllenoxid eine deutliche
Inaktivierung mit einem IC50-Wert von ca. 1 mM. α-Ionon verursachte einen niedrigen
Aktivitätsverlust von ca. nur 15 % im Gegensatz zu seinem Isomer β-Ionon, das einen
Aktivitätsverlust von ca. 55 % hervorrief.
Waye et al. untersuchten die GABAerge Wirkung der verschiedenen kanadischen Koniferen wie
Abies balsamea L., Pinus strobus L., Picea glauca (Moench) Voss und vom 16 unterschiedlichen
Phytochemikalien. Dabei verursachten diese Pflanzen und deren Phytochemikalien wie Linalool,
Vanilin, Isoeugenol keine Veränderung der Aktivität der GABA-AT, während sie eine neuroaktive
Wirkung über eine Modulierung der Monoaminooxidase und der Glutamat-Decarboxylase
zeigten [184]. Dies spricht dafür, dass Lebensmittel und ihre aktiven Inhaltsstoffe wie
Terpenoide, welche keine Inhibition der GABA-AT in dieser Arbeit zeigten, über andere
Mechanismen neuroaktiv bzw. GABAerg wirken könnten. So wurde bereits festgestellt, dass
terpenoide Inhaltsstoffe von Lebensmitteln wie beispielsweise ß-Caryophyllen, Isopulegol,
Pinocarveol, Myrtenol die Aktivität der GABAA-Rezeptoren erhöhen konnten [87].
Zusammenfassend konnte in dieser Arbeit eine konzentrationsabhängige, signifikant hemmende
Wirkung der Lebensmittel (Salbei, Nelken, grünem Tee, Lavendel, Kamille, Melisse, und Sideritis-
Arten) und Lebensmittelinhaltsstoffe (β-Damascenon, Myrcen, β-Ionon und (-)-
Caryophyllenoxid) auf GABA-metabolisierende Enzyme nachgewiesen werden. Dies deutet
darauf hin, dass diese Lebensmittel über eine Inhibition des GABA-Abbaus das GABAerge
Neurotransmittersystem positiv modulieren können.

Modulation des GABAA-Rezeptors
39
Modulation des GABAA-Rezeptors 3
3.1. Einleitung
In Kapitel 2 wurde der hemmende Effekt wässriger Extrakte von 13 verschiedenen
Lebensmitteln auf GABA-Abbauenzyme untersucht. Hierbei wurden Lebensmittel ausgewählt,
deren neurophysiologische Wirkungen, insbesondere eine beruhigende, sedative, angstlösende,
schlaffördernde, antiepileptische, und antidepressive Wirkung, in in vivo Studien gezeigt wurde
(siehe Kapitel 1.6). Mehrere endogene und synthetische Substanzen entfalten ihre Wirkung auf
das Schlaf-Wach-Verhalten über GABAA-Rezeptoren [68-70, 72, 185]. Zudem sind GABAA-
Rezeptoren das Target sedativer, hypnotischer und angstlösender Medikamente [48, 61, 67-69].
Deshalb befasst sich der folgende Abschnitt der Arbeit nun mit der modulierenden Wirkung der
Extrakte dieser 13 Lebensmitteln - Grüner Tee, Hopfen, Kamille, Kakao, Lavendel, Melisse,
Nelken, Salbei, Schwarzer Pfeffer sowie 4 Sideritis Arten (S. condensata, S. stricta, S. arguta und
S. sipylea) - auf den humanen GABAA-Rezeptor.
Heteropentamere GABAA-Rezeptoren gehören zur der Cys-Loop-Rezeptorsuperfamilie der
ligandengesteuerten Ionenkanäle und bestehen aus verschiedenen Untereinheiten (α1–α6, β1–
β3, γ1–γ3, ρ1–ρ3, δ, ε, θ und π), die miteinander kombinieren können, um die
Chloridionenkanäle zu bilden [12-16, 49]. Da nur die α- und β- Untereinheiten die Bindungstelle
für den Liganden GABA bilden [58], werden in der experimentellen pharmakologischen
Forschung diese binären GABAA-Rezeptoren eingesetzt um die Anzahl der zu transfizierenden
Untereinheiten zu begrenzen [18]. Weiterhin besitzen die binären GABAA-Rezeptoren eine
höhere Affinität zu GABA als ihre γ2-Untereinheit enthaltenden ternären GABAA-
Rezeptorkomplexe [52]. Daher wurde der binäre GABAA-Rezeptor aus α1- und β2 –
Untereinheiten in der vorliegenden Arbeit verwendet. Hierzu wurde der humane α1β2 GABAA-
Rezeptor rekombinant in Oocyten von Xenopus leavis nach Injektion entsprechender cRNA
exprimiert und seine Modulation durch Lebensmittelextrakte elektrophysiologisch mittels TEVC
(aus dem Englischen: Two electrode voltage clamp)-Technik untersucht.
GABA ist bekanntlich ein natürlicher Inhaltsstoff von zahlreichen Lebensmitteln, insbesondere
von verschiedenen Teesorten [186-193]. Daher sollte aufgeklärt werden, ob diese
Lebensmittelextrakte endogene GABA als einen natürlichen Inhaltsstoff enthalten und ob diese
eine Änderung der mittels TEVC gemessenen Ströme hervorrufen kann. Zudem wurde eine
direkte Rezeptoraktivierung durch die Lebensmittelextrakte in Abwesenheit der Zugabe des
Agonisten GABA untersucht. Weiterhin wurde ein Kontrollversuch unter Verwendung von
Glycin-Rezeptoren durchgeführt, die ebenfalls zur Cys-Loop Superfamilie der

Modulation des GABAA-Rezeptors
40
ligandengesteuerten Ionenkanälen zählen und selektiv für Chloridionen sind [12-15]. Dadurch
sollte bestimmt werden, ob der modulierende Effekt der Lebensmittelextrakte GABA-Rezeptor
spezifisch ist.
3.2. Modulation der GABAA-Rezeptoren durch Lebensmittelextrakte
Die Untersuchung der Wirkung von Lebensmittelextrakten auf GABAA-Rezeptoren erfolgte durch
die elektrophysiologische Messung der Stromamplituden des GABA-induzierten
Chloridionenstromes mit Hilfe der TEVC-Technik. Die Versuche wurden an Xenopus leavis
Oocyten durchgeführt, in denen der humane α1β2 GABAA-Rezeptor rekombinant exprimiert
wurde.
Im ersten Schritt wurde die erfolgreiche GABAA-Rezeptorexpression überprüft und die
dosisabhängige Wirkung des Agonisten GABA bestimmt. Dazu wurde GABA in verschiedenen
Konzentrationen (0,3 µM, 1 µM, 3 µM, 10 µM, 30 µM, 100 µM und 300 µM) appliziert. Die
Negativkontrolle erfolgte durch Applikation von GABA auf nicht-transfizierten Oocyten, wobei
kein Effekt beobachtet wurde. In Abbildung 3.1.A ist der durch GABAA-Rezeptorkanäle
vermittelte Chloridionenstrom während und nach der Applikation von GABA dargestellt.
Während der Applikation des Liganden GABA stieg der Ioneneinstrom stetig an und erreichte
nach ein paar Sekunden einen Maximalwert. Nach Beendigung der GABA-Applikation kehrte
der Ioneneinwärtsstrom durch Auswaschen mit Puffer auf das Niveau des Ruhestroms (die
vorhergehende Grundlinie des Stromflusses) zurück, wodurch die Deaktivierung der
Chloridionenkanäle erkennbar wird. Anschließend blieb der Strom im Ruhezustand der Oocyte
konstant bis zur erneuten Applikation von GABA und der damit einhergehenden Aktivierung der
Ionenkanäle. Die Differenz zwischen dem Ruhestrom und dem durch Applikation von GABA
ermittelten Chloridionenstrom ergab den GABA-induzierten Strom, der ein Maß für die
Aktivierung der Ionenkanäle ist und mit ansteigenden Konzentrationen von GABA kontinuierlich
zunahm. Die GABA-induzierten Ströme wurden auf den maximalen Strom normiert, den GABA in
der höchsten applizierten Konzentration von 300 µM auslöste. Die normierten GABA-
induzierten Stromantworten wurden gegen die applizierten Konzentrationen von GABA
aufgetragen. Hierbei wurde ein konzentrationsabhängiger Anstieg der Kurve für den Agonisten-
GABA ermittelt. Diese GABA-Dosis-Wirkungskurve wurde mit der Hill-Gleichung angepasst,
wobei der EC5, EC15 und EC50 Wert für GABA berechnet wurde. Die Konzentration für eine
halbmaximale Aktivierung des Rezeptors (EC50) betrug 7,47 µM. Da berichtet wird, dass der
Effekt von Modulatoren am sensitivsten bei einer niedrigen Konzentration des Liganden-GABA
vermessen werden kann [83, 84], wurde die GABA-Konzentration von 1 µM GABA, welche im

Modulation des GABAA-Rezeptors
41
EC5-15 Bereich lag (EC5: 0,4 µM und EC15: 1,3 µM), für die Ko-Applikation von Modulatoren
gewählt. Abbildung 3.1.B zeigt die Stromantwort in Abhängigkeit der GABA Konzentration.
A) B)
Abbildung 3.1 Wirkung von GABA auf den humanen α1β2 GABAA-Rezeptor in Xenopus leavis Oocyten. A)
Repräsentative Stromspuren für die direkte Rezeptoraktivierung durch den Liganden GABA.
GABA wurde in unterschiedlichen Konzentrationen zwischen 0,3 und 300 µM appliziert. B)
Dosis-Wirkungskurve für GABA. Die Stromamplituden wurden auf die höchste GABA-
induzierte Stromantwort normalisiert; 1 = Stromantwort, ausgelöst durch 300 µM GABA. Die
Anpassung erfolgte mittels der Hill-Funktion mit einem Hill-Koefizienten von 0,99. Mittelwert
± SEM aus 6 Oocyten von drei unabhängig transfizierten Chargen.
Im nächsten Schritt wurde der mögliche modulierende Effekt von 13 Lebensmittelextrakten -
Nelken, Grüner Tee, Salbei, 4 Sideritis-Arten, Lavendel, Melisse, Kamille, Hopfen, Kakao und
Schwarzer Pfeffer- auf den α1β2 GABAA-Rezeptoren untersucht. Als Vorversuch wurde jeder
Lebensmittelextrakt in einer Konzentration von 10 µg/mL in Abwesenheit von GABA getestet
und anschließend mit einer Applikation von 1 µM GABA bzw. einer Koapplikation (eine
Mischung aus Lebensmittelextrakt und GABA) verglichen. Bei der Negativkontrolle zeigten
einige wässrige Lebensmittelextrakte einen unspezifischen Strom in nicht transfizierten
Oocyten, wie in Abbildung 3.2 dargestellt ist. Dies steht im Einklang mit der bereits publizierten
Beobachtung, dass wässrige Tee-Extrakte aufgrund ihres hohen Gehaltes an Polyphenolen
Membranen schädigen können, wodurch eine Stromamplitude in nicht-transfizierten Oocyten
erzeugt werden kann [194].
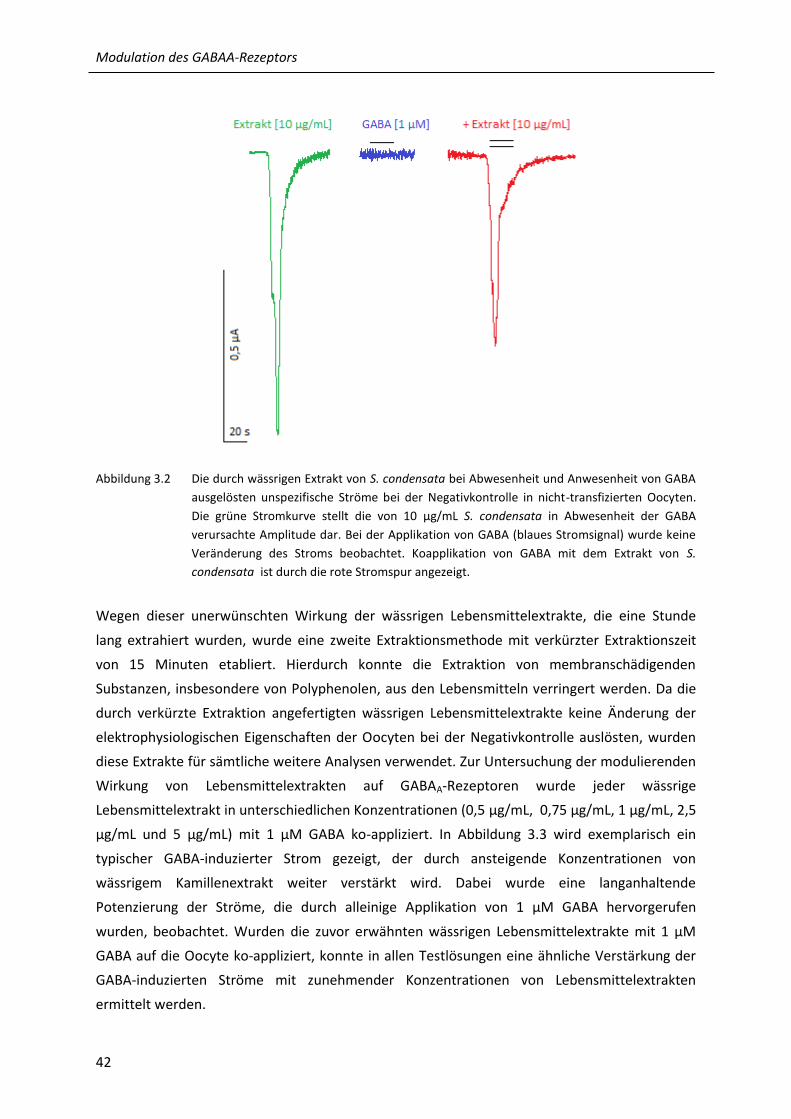
Modulation des GABAA-Rezeptors
42
Abbildung 3.2 Die durch wässrigen Extrakt von S. condensata bei Abwesenheit und Anwesenheit von GABA
ausgelösten unspezifische Ströme bei der Negativkontrolle in nicht-transfizierten Oocyten.
Die grüne Stromkurve stellt die von 10 µg/mL S. condensata in Abwesenheit der GABA
verursachte Amplitude dar. Bei der Applikation von GABA (blaues Stromsignal) wurde keine
Veränderung des Stroms beobachtet. Koapplikation von GABA mit dem Extrakt von S.
condensata ist durch die rote Stromspur angezeigt.
Wegen dieser unerwünschten Wirkung der wässrigen Lebensmittelextrakte, die eine Stunde
lang extrahiert wurden, wurde eine zweite Extraktionsmethode mit verkürzter Extraktionszeit
von 15 Minuten etabliert. Hierdurch konnte die Extraktion von membranschädigenden
Substanzen, insbesondere von Polyphenolen, aus den Lebensmitteln verringert werden. Da die
durch verkürzte Extraktion angefertigten wässrigen Lebensmittelextrakte keine Änderung der
elektrophysiologischen Eigenschaften der Oocyten bei der Negativkontrolle auslösten, wurden
diese Extrakte für sämtliche weitere Analysen verwendet. Zur Untersuchung der modulierenden
Wirkung von Lebensmittelextrakten auf GABAA-Rezeptoren wurde jeder wässrige
Lebensmittelextrakt in unterschiedlichen Konzentrationen (0,5 µg/mL, 0,75 µg/mL, 1 µg/mL, 2,5
µg/mL und 5 µg/mL) mit 1 µM GABA ko-appliziert. In Abbildung 3.3 wird exemplarisch ein
typischer GABA-induzierter Strom gezeigt, der durch ansteigende Konzentrationen von
wässrigem Kamillenextrakt weiter verstärkt wird. Dabei wurde eine langanhaltende
Potenzierung der Ströme, die durch alleinige Applikation von 1 µM GABA hervorgerufen
wurden, beobachtet. Wurden die zuvor erwähnten wässrigen Lebensmittelextrakte mit 1 µM
GABA auf die Oocyte ko-appliziert, konnte in allen Testlösungen eine ähnliche Verstärkung der
GABA-induzierten Ströme mit zunehmender Konzentrationen von Lebensmittelextrakten
ermittelt werden.

Modulation des GABAA-Rezeptors
43
Abbildung 3.3 Beispielstromkurven einer Messung für die Koapplikation von 1 µM GABA mit wässrigem
Lebensmittelextrakt an humanen α1β2 GABAA-Rezeptoren. Die blauen Stromspuren zeigen
die durch 1 µM GABA erzeugten Amplituden. Die Potenzierung der GABA-induzierten Ströme
durch wässrigen Kamillenextrakt in unterschiedlichen Konzentrationen (0,5-5 µg/mL) bei der
simultanen Applikation mit 1 µM GABA ist durch die roten Stromkurven dargestellt.
Der Mittelwert der Ströme, die durch alleinige Applikation von 1 µM GABA vor und nach der
jeweiligen Koapplikation von GABA und Lebensmittelextrakt vermittelt wurde, wurde gleich
100% gesetzt. Hierauf wurde die Verstärkung der GABA-induzierte Ströme durch
Lebensmittelextrakte bezogen und als prozentualer Anteil berechnet, um die aktivierende
Wirkung verschiedener Extrakte miteinander zu vergleichen. Die ermittelten Potenzierungen der
GABA-vermittelten Ströme durch wässrige Lebensmittelextrakte sind in Tabelle 3.1
zusammengefasst. Alle getesteten Lebensmittelextrakte führten zu einer
konzentrationsabhängigen Steigerung der GABA-induzierten Ströme. Dabei zeigten Salbei,
Kakao, Lavendel, Schwarzer Pfeffer, S. sipylea Hopfen und Nelken in allen eingesetzten

Modulation des GABAA-Rezeptors
44
Konzentrationen eine signifikante Verstärkung der Aktivität des GABAA Rezeptors, während
S.condensata, S. arguta, Grüner Tee, S. stricta und Melisse erst ab einer Konzentration von 0,75
µg/mL die GABA-induzierten Ströme signifikant potenzierten. Im Fall des Kamillenextraktes trat
erst bei einer Konzentration von 1 µg/mL eine signifikante Verstärkung der GABAA-
Rezeptorantwort auf (siehe Tabelle 3.1). Bei der höchsten applizierten Konzentration riefen alle
untersuchten Lebensmittelextrakte eine höchst signifikante Potenzierung der GABA-
vermittelten Ströme hervor, wie in Abbildung 3.4 aufgezeigt wird (p < 0.0001). Dabei erwies sich
Nelkenextrakt als der potenteste Modulator des GABAA-Rezeptors, der eine Erhöhung des
GABA-vermittelten Stroms auf 427 % im Vergleich zur GABA-Kontrolle bewirkte. Melisse und S.
stricta potenzierten die Aktivität des GABAA-Rezeptorkanals um bis zu 360 %. Durch Extrakte
von Hopfen, grünem Tee und S. arguta wurde der GABA-induzierten Strom um ca. 340-348 %
gesteigert, während andere Extrakte die durch GABA induzierte Amplitude nur um 216-286 %
verstärkten (siehe Tabelle 3.1).

Modulation des GABAA-Rezeptors
45
Tabelle 3.1 Modulation des humanen α1β2 GABAA-Rezeptors durch wässrige Lebensmittelextrakte. Die
unterschiedlichen Konzentrationen (0,5–5 µg/mL) der Extrakte wurden mit 1 µM GABA
simultan appliziert. Die Daten stellen die prozentualen mittleren Stromantworten (Irel )
bezogen auf die GABA-Kontrolle (100%) ± SEM aus mindestens drei unabhängig transfizierten
Oocyten-Chargen dar. Signifikante Unterschiede wurden durch zweifaktorielle-ANOVA mit
Holm-Sidak Vergleichstest ermittelt (ns: nicht signifikant; * p < 0,05; ** p < 0, 01; *** p <
0,001; **** p < 0,0001 versus GABA-Kontrolle). N bezeichnet die Anzahl der untersuchten
Oocyten.
Lebensmittelextrakt Konzentration [µg/mL] Irel [%] Signifikanzwert N
Salbei 0,5 154 ± 14 **** 7 0,75 174 ± 10 **** 9 1 188 ± 10 **** 6 2,5 199 ± 13 **** 6 5 216 ± 8 **** 8
Kakao 0,5 160 ± 8 **** 8 0,75 190 ± 10 **** 6 1 213 ± 10 **** 9 2,5 231 ± 12 **** 13 5 248 ± 11 **** 8
Lavendel 0,5 156 ± 7 * 8 0,75 183 ± 20 ** 6 1 204 ± 25 **** 9 2,5 214 ± 17 **** 10 5 264 ± 27 **** 9
S. condensata 0,5 168 ± 20 ns 6 0,75 192 ± 21 * 6 1 226 ± 33 ** 7 2,5 267 ± 56 **** 7 5 271 ± 35 **** 8
Schwarzer Pfeffer 0,5 173 ± 30 ** 5 0,75 215 ± 25 **** 5 1 222 ± 16 **** 7 2,5 249 ± 23 **** 6 5 274 ± 33 **** 5
Kamille 0,5 141 ± 10 ns 9 0,75 179 ± 24 ns 5 1 210 ± 36 ** 6 2,5 213 ± 23 ** 8 5 286 ± 59 **** 8
S. sipylea 0,5 173 ± 18 ** 7 0,75 200 ± 14 **** 10 1 238 ± 20 **** 9 2,5 264 ± 22 **** 10 5 291 ± 25 **** 9
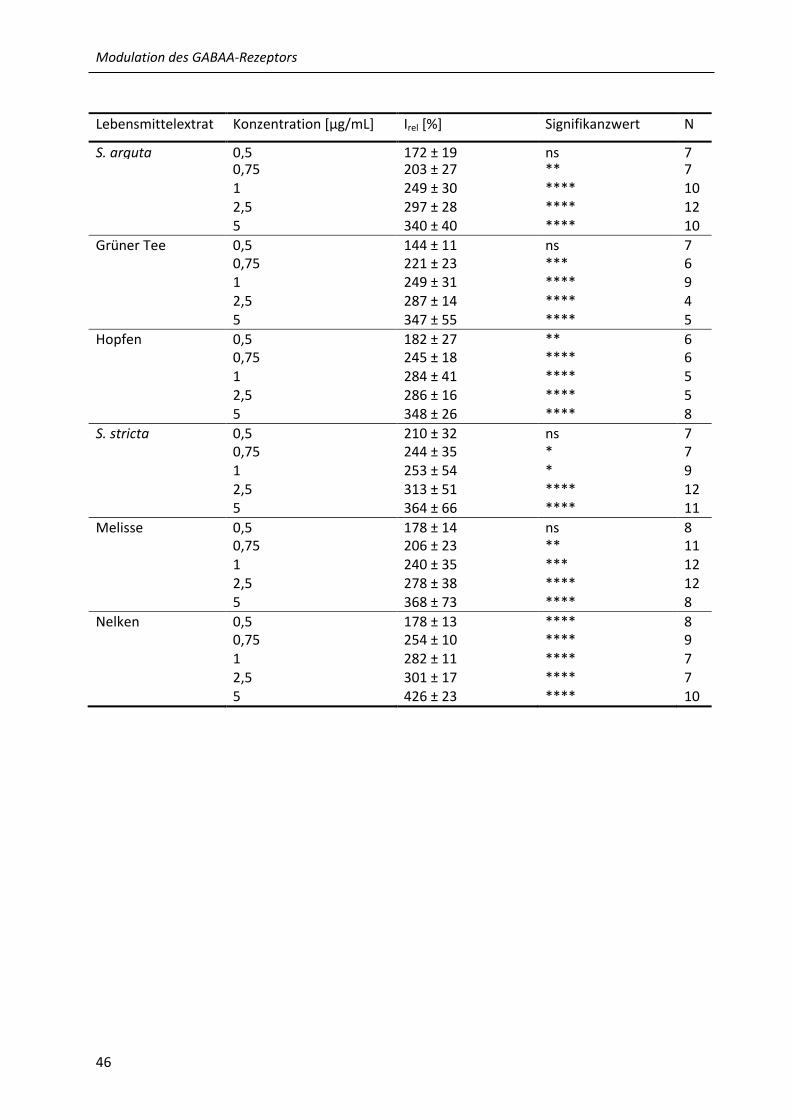
Modulation des GABAA-Rezeptors
46
Lebensmittelextrat Konzentration [µg/mL] Irel [%] Signifikanzwert N
S. arguta 0,5 172 ± 19 ns 7 0,75 203 ± 27 ** 7 1 249 ± 30 **** 10 2,5 297 ± 28 **** 12 5 340 ± 40 **** 10
Grüner Tee 0,5 144 ± 11 ns 7 0,75 221 ± 23 *** 6 1 249 ± 31 **** 9 2,5 287 ± 14 **** 4 5 347 ± 55 **** 5
Hopfen 0,5 182 ± 27 ** 6 0,75 245 ± 18 **** 6 1 284 ± 41 **** 5 2,5 286 ± 16 **** 5 5 348 ± 26 **** 8
S. stricta 0,5 210 ± 32 ns 7 0,75 244 ± 35 * 7 1 253 ± 54 * 9 2,5 313 ± 51 **** 12 5 364 ± 66 **** 11
Melisse 0,5 178 ± 14 ns 8 0,75 206 ± 23 ** 11 1 240 ± 35 *** 12 2,5 278 ± 38 **** 12 5 368 ± 73 **** 8
Nelken 0,5 178 ± 13 **** 8 0,75 254 ± 10 **** 9 1 282 ± 11 **** 7 2,5 301 ± 17 **** 7 5 426 ± 23 **** 10

Modulation des GABAA-Rezeptors
47
Abbildung 3.4 Potenzierende Effekte von wässrigen Lebensmittelextrakten in der Konzentration von 5
µg/mL auf die durch 1 µM GABA induzierte Stromantworte des humanen α1β2 GABAA-
Rezeptors. Die prozentualen Angaben beziehen sich jeweils auf die GABA-Kontrolle (100%).
Die Daten zeigen den Mittelwerten ± SEM aus 5-11 Oocyten (siehe Tabelle 3.1) von drei
unabhängig transfizierten Chargen. Signifikante Unterschiede wurden durch zweifaktorielle-
ANOVA mit Holm-Sidak Vergleichstest ermittelt (**** p < 0,0001 versus GABA-Kontrolle).
****
****
****
****
****
****
****
****
****
****
****
****
****
0 100 200 300 400 500
Nelken
Melisse
S. stricta
Hopfen
Grüner Tee
S. arguta
S. sipylea
Kamille
Schwarzer Pfeffer
S. condensata
Lavendel
Kakao
Salbei
Kontrolle
Relativer Strom [%]

Modulation des GABAA-Rezeptors
48
3.3. Bestimmung der GABA Gehalte der Extrakte und Untersuchungen zu deren Wirkung auf den GABAA Rezeptor
Um zu überprüfen, ob die untersuchten Lebensmittelextrakte GABA enthalten und ob die
beobachtete Potenzierung der GABAA-Rezeptoraktivierung aus endogener GABA dieser
Lebensmittel resultiert, wurde zunächst der natürliche GABA-Gehalt der Lebensmittel mit einer
HPLC-Methode gemessen. Anschließend wurde die elektrophysiologische Wirkung der in den
Lebensmittelextrakten enthaltenen GABA mittels TEVC-Technik untersucht.
3.3.1. Quantifizierung von GABA in den Lebensmittelextrakten
Zur Bestimmung der GABA-Gehalte in den Lebensmittelextrakten wurden zwei verschiedene
Derivatisierungsmethoden bzw. verschiedene HPLC-Methoden angewendet. Bei der ersten
Methode, modifiziert nach Khuhawar et al. [195] wurde GABA mit 2-Hydroxynaphthaldehyde (2-
HN) derivatisiert, mittels HPLC an einer RP (RP, aus dem Englischen: reversed phase;
Umkerphase)- Säule aufgetrennt und anschließend durch UV-Detektion bei 330 nm bestimmt
(RP-HPLC-UV-Methode). Da bei der Messung mit dieser Methode ein unsymmetrischer GABA-
Peak auftrat, sollte eine weitere Methode, welche nach Zhao et al. modifiziert wurde [189],
getestet werden. Hierbei wurde GABA mit ortho-Phtalaldehyd (OPA) derivatisiert. Nach der
Derivatisierung mit OPA wurde das entstandene fluoreszierende GABA-Derivat mittels HPLC an
einer RP- Säule aufgetrennt und mittels Fluoreszenzdetektor (FLD) mit einer
Anregungswellenlänge von 230 nm und einer Emissionswellenlänge von 450 nm nachgewiesen
(RP-HPLC-FLD-Methode). Ein RP-HPLC-FLD-Chromatogramm einer OPA-derivatisierten GABA-
Standardlösung (0,1 mg/mL in Reinstwasser), die als Referenzsubstanz diente, ist in Abbildung
3.5 dargestellt. Da mit Hilfe dieser Methode eine gute Trennleistung erreicht werden konnte,
wurde sie für alle darauffolgenden Bestimmungen von GABA in Lebensmittelextrakten
verwendet. Dabei wurden die wässrigen Lebensmittelextrakte in unterschiedlichen
Konzentrationen (5-40 mg/mL) hergestellt und dreimal unabhängig voneinander mit OPA
derivatisiert. Anschließend wurde das erhaltene GABA-Derivat mit Hilfe RP-HPLC-FLD-Methode
analysiert. Die Identifizierung von GABA in den Lebensmittelextrakten erfolgte durch den
Vergleich der Retentionszeiten und Fluoreszenzspektren der Signale der Lebensmittelextrakte
und der Referenzsubstanz. Ein Beispielchromatogramm eines Lebensmittelextraktes nach
Derivatisierung der GABA mit OPA ist in Abbildung 3.5 gezeigt.

Modulation des GABAA-Rezeptors
49
Abbildung 3.5 RP-HPLC-FLD-Chromatogramme der 0,1 mg/mL GABA-Standardlösung (pink) und eines
wässrigen Kakao-Extraktes (blau) nach Derivatisierung mit OPA. Das GABA-Signal ist
zusätzlich in einer vergrößerten Ansicht dargestellt. Anregungswellenlänge: 230 nm;
Emissionswellenlänge: 450 nm.
Zur Quantifizierung der GABA in den Lebensmittelextrakten wurde eine externe Kalibrierung
durchgeführt. Dazu wurden GABA-Standardlösungen in einem Konzentrationsbereich zwischen
1 µg/mL und 25 µg/mL mit OPA derivatisiert und anschließend mittels RP-HPLC-FLD vermessen.
Durch Auftragung der gemessenen Peakflächen gegen die eingesetzte GABA-Konzentration
wurde eine Kalibriergerade erstellt (siehe Abbildung im Anhang A.2). Diese wurde auf Linearität
geprüft und zur Ermittlung der Nachweisgrenze (NWG) sowie der Bestimmungsgrenze (BSG)
herangezogen. Die Kalibrierung zeigte eine gute lineare Regression mit einem
Determinationskoeffizient (R2) von 0,9988. Die NWG und BSG für GABA betrugen 0,6 µg/mL und
1,7 µg/mL. Mit Hilfe der externen Kalibrierkurve konnten anschließend die GABA-Konzentration
der Lebensmittelextrakte berechnet werden. Um diese ermittelten Konzentrationswerte
miteinander vergleichen zu können, wurden die Werte auf einen Lebensmittelextrakt mit einer
Konzentration von 1 mg/mL normalisiert. Auf diese Weise konnte GABA in einem
Konzentrationsbereich zwischen 0,14 und 0,81 µg/mL in allen Lebensmittelextrakten mit
Ausnahme des Nelkenextraktes identifiziert und quantifiziert werden. Beim Nelkenextrakt lag
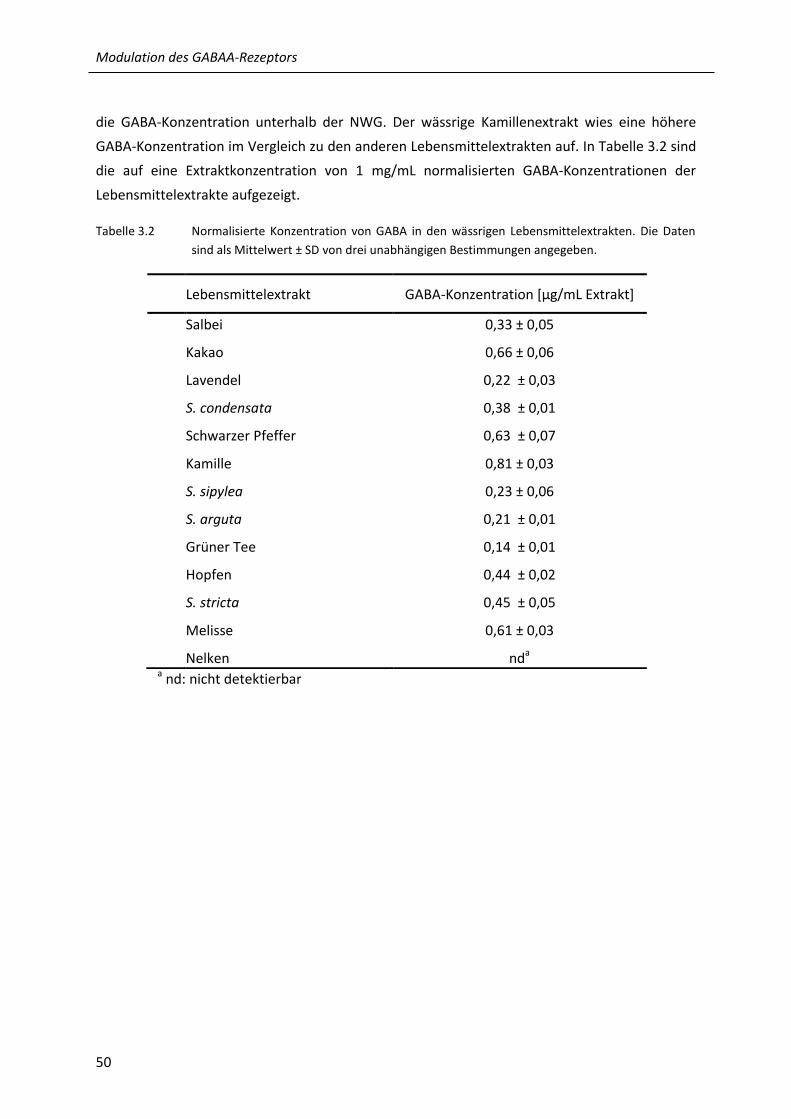
Modulation des GABAA-Rezeptors
50
die GABA-Konzentration unterhalb der NWG. Der wässrige Kamillenextrakt wies eine höhere
GABA-Konzentration im Vergleich zu den anderen Lebensmittelextrakten auf. In Tabelle 3.2 sind
die auf eine Extraktkonzentration von 1 mg/mL normalisierten GABA-Konzentrationen der
Lebensmittelextrakte aufgezeigt.
Tabelle 3.2 Normalisierte Konzentration von GABA in den wässrigen Lebensmittelextrakten. Die Daten
sind als Mittelwert ± SD von drei unabhängigen Bestimmungen angegeben.
a nd: nicht detektierbar
Lebensmittelextrakt GABA-Konzentration [µg/mL Extrakt]
Salbei 0,33 ± 0,05
Kakao 0,66 ± 0,06
Lavendel 0,22 ± 0,03
S. condensata 0,38 ± 0,01
Schwarzer Pfeffer 0,63 ± 0,07
Kamille 0,81 ± 0,03
S. sipylea 0,23 ± 0,06
S. arguta 0,21 ± 0,01
Grüner Tee 0,14 ± 0,01
Hopfen 0,44 ± 0,02
S. stricta 0,45 ± 0,05
Melisse 0,61 ± 0,03
Nelken nda

Modulation des GABAA-Rezeptors
51
3.3.2. Direkte GABAA-Rezeptoraktivierung durch die untersuchten Lebensmittelextrakte
Die RP-HPLC-FLD Ergebnisse ergaben, dass die untersuchten Lebensmittelextrakte GABA in nur
sehr geringen Konzentrationen enthalten. Aufgrund dieser Ergebnisse ist zu erwarten, dass
diese GABA-Konzentrationen nicht ausreichen, um den GABAA Rezeptor als Agonisten
elektrophysiologisch zu stimulieren. Neben GABA könnten diese Lebensmittelextrakte jedoch
zusätzliche weitere Substanzen enthalten, die einen agonistischen Effekt auf den α1β2 GABAA
Rezeptor haben können und die beobachteten Potenzierung der GABA-induzierten Ströme
verursachen. Daher wurde überprüft, ob die in den Lebensmittelextrakten enthaltene GABA
oder mögliche andere Agonististen einen elektrophysiologischen Effekt auf den in Oocyten
exprimierten GABAA Rezeptor evozieren können. Dazu wurde jeder Lebensmittelextrakt in einer
Konzentration von 7,5 µg/mL ohne Zugabe von GABA in die transfizierten Oocyten appliziert.
Diese Konzentration war höher als die maximale Extraktkonzentration, welche bei der
Koapplikation mit einer 1 µM GABA-Lösung getestet wurde (siehe Kapitel 3.2). In Abwesenheit
von GABA erzeugten die getesteten Lebensmittelextrakte nur einen sehr kleinen Ionenfluss
durch die Membran der transfizierten Oocyten. Ein Beispielapplikation von Kamillenextrakt, in
dem die höchste GABA-Konzentration gemessen wurde (siehe Tabelle 3.2), ist in Abbildung 3.6
dargestellt.

Modulation des GABAA-Rezeptors
52
Abbildung 3.6 Beispielstromkurven einer Messung der direkten Aktivierung des humanen α1β2 GABAA-
Rezeptor durch wässrige Lebensmittelextrakte. Die grüne Stromspur stellt die von 7,5 µg/mL
Kamille in Abwesenheit von GABA verursachte Amplitude dar. Der durch 1 µM GABA
induzierte Strom ist in blau angezeigt. Die rote Stromkurve zeigt die Potenzierung des GABA-
induzierten Stroms durch den wässrigen Kamillenextrakt in einer Konzentration von 7,5
µg/mL bei der simultanen Applikation von 1 µM GABA.
Bei jedem Versuch wurde 1 µM GABA als Kontrolle mitbestimmt und diese Stromamplitude auf
100 % gesetzt. Bei der alleinigen Applikation der Lebensmittelextrakte betrug die
Stromamplitude 1-19 % bezogen auf diesen durch 1 µM GABA-induzierten Stromfluss (siehe
Tabelle 3.4). Sollte die beobachtete Potenzierung der GABA-induzierte Ströme (siehe Kapitel
3.2) allein durch agonistische Substanzen in den Lebensmittelextrakten ausgelöst werden, so
wäre eine Stromamplitude von nahezu 100 % zu erwarten. Diese Ergebnisse deuten demnach
darauf hin, dass der GABAA-Rezeptor durch Lebensmittelextrakte in Abwesenheit von
zugesetzter GABA nicht aktiviert wird. Somit kann die Wirkung der Lebensmittelextrakte auf den
GABAA-Rezeptor als eine positive allosterische Modulation charakterisiert werden.

Modulation des GABAA-Rezeptors
53
Tabelle 3.3 Relative Chloridionenströme der direkten Aktivierung des humanen α1β2 Rezeptors durch
wässrige Lebensmittelextrakte in der Konzentration von 7,5 µg/mL in Abwesenheit von
GABA. Die Daten stellen die prozentualen mittleren Stromantworten (Irel) bezogen auf die
GABA-Kontrolle (100%) ± SEM aus mindestens zwei unabhängig transfizierten Oocyten-
Chargen dar. N ist die Anzahl der untersuchten Oocyten.
Lebensmittelextrakt Irel [%] N
Salbei 6 ± 1 7
Kakao 5 ± 1 9
Lavendel 3 ± 1 6
S. condensata 12 ± 2 6
Schwarzer Pfeffer 15 ± 2 6
Kamille 15 ± 1 4
S. sipylea 7 ± 2 5
S. arguta 3 ± 1 7
Grüner Tee 1 ± 0,2 5
Hopfen 19 ± 4 5
S. stricta 7 ± 1 6
Melisse 12 ± 2 5
Nelken 6 ± 2 4
3.4. Wirkung der Lebensmittelextrakte auf den Glycin-Rezeptor
Im Folgenden wurde der Einfluss der Lebensmittelextrakte auf Glycin-Rezeptoren untersucht,
die, wie auch die GABA-Rezeptoren, zur Cys-Loop-Rezeptor Superfamilie der
ligandengesteuerten Ionenkanäle gehören [12-15]. Hierdurch sollte überprüft werden, ob die
Lebensmittelextrakte spezifisch den GABAA-Rezeptor adressieren, oder ob sie eher unselektiv
wirken. Die Messung der Glycin-vermittelten Chloridionenströme erfolgte mittels TEVC-Technik
an Oocyten, in die cRNA der humanen α1 Untereinheit des Glycin-Rezeptors injiziert wurde.
Hierbei wurde zunächst die Glycin-Rezeptorexpression kontrolliert und die dosisabhängige
Wirkung des Agonisten Glycin bestimmt. Dafür wurde Glycin in unterschiedlichen
Konzentrationen (10 µM, 30 µM, 50 µM, 100 µM, 300 µM, 1000 µM und 3000 µM) den Oocyten
zugeführt. Wie in Abbildung 3.7.A dargestellt, stiegen die Stromamplituden erwartungsgemäß
mit zunehmender Glycin-Konzentration an. Die Stromantworten wurden auf den maximalen
Chloridionenstrom normiert. Zur Erstellung der Dosis-Wirkungskurve für Glycin wurde die Hill-
Funktion verwendet. Dabei wurde der EC5, EC15 und EC50 Wert für den Agonisten Glycin
ermittelt, wobei der EC50 Wert 132,9 µM betrug. Der Effekt der Lebensmittelextrakte auf den
α1-Glycin-Rezeptor wurde bei einer Glycin-Konzentration von 30 µM, die im EC5-15-Bereich (24,3
– 49,1 µM) lag, untersucht, um eine optimale Sensitivität zu erzielen. In Abbildung 3.7.B ist die
Dosis-Wirkungskurve für Glycin angezeigt. Während der Applikation von Glycin auf nicht-
transfizierte Oocyten wurde keine elektrophysiologische Änderung registriert. Dies diente als
Negativkontrolle.

Modulation des GABAA-Rezeptors
54
A) B)
Abbildung 3.7 Wirkung von Glycin auf den humanen α1 Glycin-Rezeptor an Xenopus leavis Oocyten. A)
Repräsentative Stromspuren für die direkte Rezeptoraktivierung durch den Liganden Glycin,
der in unterschiedlichen Konzentrationen zwischen 10 und 3000 µM appliziert wurde. B) Die
Dosis-Wirkungskurve für den Agonisten Glycin. Die Stromamplituden wurden auf die höchste
durch Glycin induzierte Stromantwort normalisiert; 1 = durch 3000 µM Glycin ausgelöste
Stromantwort. Die Anpassung erfolgte mittels der Hill-Funktion mit einem Hill-Koeffizienten
von 1,73. Mittelwert ± SEM aus 7 Oocyten von drei unabhängig transfizierten Chargen.
Zur Erfassung der modulierenden Wirkung der Lebensmittelextrakte auf den Glycin-Rezeptor
wurde jeder Lebensmittelextrakt in einer Konzentration von 5 µg/mL getestet, bei der die
Lebensmittelextrakte die höchste Potenzierung der GABAA Rezeptor-Aktivität zeigten (siehe
Kapitel 3.2). Die Lebensmittelextrakte wurden hierbei mit 30 µM Glycin auf die mit dem α1
Glycin-Rezeptor transfizierten Oocyten ko-appliziert. Wurden die Lebensmittelextrakte in
Abwesenheit von Glycin appliziert, konnte keiner der Lebensmittelextrakte eine Stromamplitude
in den Oocyten induzieren. Als ein Beispiel stellt Abbildung 3.8 die Wirkung des wässrigen
grünen Tee-Extraktes auf die Glycin-vermittelten Ströme dar. Vor und nach jeder Messung
wurde 30 µM Glycin appliziert, deren Stromantwort auf 100 % gesetzt wurde. Im Gegensatz zur
Potenzierung des GABAA-Rezeptors lösten die untersuchten Lebensmittelextrakte - außer dem
Extrakt von S. sipylea - eine Verminderung der Glycin-induzierten Ströme aus, wie in Abbildung
3.9 gezeigt. Dabei war dieser Effekt für S. sipylea, S. arguta, Lavendel, Hopfen und Kakao nicht
signifikant, während die Extrakte von Kamille (p<0,05), schwarzem Pfeffer (p<0,05), Melisse
(p<0,01), Nelken (p<0,01), S. stricta (p<0,01), grünem Tee(p<0,001), S. condensata (p<0,001),
und Salbei (p<0,0001) kleine, aber signifikante Reduktionen der Glycin-vermittelten Ströme
verursachten. Durch Salbei und S. condensata wurde in einer Konzentration von 5 µg/mL eine

Modulation des GABAA-Rezeptors
55
Reduktion der Stromantwort auf 56% erreicht. Bei den negativen Kontrollen, die durch
Applikation der Extrakte auf nicht-transfizierte Oocyten bei jedem Versuch erfolgten, wurde
keine Stromantwort gemessen.
Abbildung 3.8 Exemplarische Stromkurven einer Messung der Modulation des humanen α1 Glycin-
Rezeptors durch wässrige Lebensmittelextrakte. Bei der Applikation des grünen Tee-
Extraktes in Abwesenheit von Glycin (Grünes Stromsignal) wurde keine Veränderung des
Stroms beobachtet. Die blauen Stromspuren stellen die durch 30 µM Glycin induzierten
Stromantworten dar. Die rote Stromkurve zeigt die Hemmung des Glycin-vermittelten
Stroms durch grünen Tee-Extrakt in einer Konzentration von 5 µg/mL bei simultaner
Applikation mit 30 µM Glycin.

Modulation des GABAA-Rezeptors
56
Abbildung 3.9 Effekte von wässrigen Lebensmittelextrakten in einer Konzentration von 5 µg/mL auf die
durch 30 µM Glycin induzierte Stromantwort des humanen α1-Glycin-Rezeptors. Die
prozentualen Angaben beziehen sich jeweils auf die Glycin-Kontrolle (100%). Die Daten
zeigen den Mittelwerten ± SEM aus 4-7 Oocyten von mindestens zwei unabhängig
transfizierten Chargen. Signifikante Unterschiede wurden durch zwei faktorielle-ANOVA mit
Holm-Sidak Vergleichstest ermittelt (ns: nicht signifikant; * p < 0,05; ** p < 0, 01; *** p <
0,001; **** p < 0,0001 versus Glycin-Kontrolle).
** **
**
ns
***
ns
ns
*
*
***
ns
ns
****
0 20 40 60 80 100 120 140
Nelken
Melisse
S. stricta
Hopfen
Grüner Tee
S. arguta
S. sipylea
Kamille
Schwarzer Pfeffer
S. condensata
Lavendel
Kakao
Salbei
Kontrolle
Relativer Strom [%]

Modulation des GABAA-Rezeptors
57
3.5. Diskussion und Zusammenfassung
Zur Behandlung verschiedener neurologischer Krankheiten wie beispielsweise chronischer
Schmerzen, Schlafstörungen, Angststörungen und Epilepsie werden Wirkstoffe eingesetzt, die
eine Verstärkung der GABAergen Inhibition verursachen [69]. Eine Möglichkeit zur Erhöhung der
GABAergen Inhibition besteht in der Potenzierung der Aktivität der GABAA-Rezeptoren [21],
welche die schnellsten synaptischen Inhibition im Säugetiergehirn vermitteln [61, 196]. Viele
Medikamente, die sedative, angstlösende und hypnotische Wirkungen haben, adressieren
spezifisch die GABAA-Rezeptoren [48, 61, 67, 68]. In diesem Kapitel der vorliegenden Arbeit
wurde die Wirkung von 13 verschiedenen wässrigen Lebensmittelextrakten - Grüner Tee,
Hopfen, Kamille, Kakao, Lavendel, Melisse, Nelken, Salbei, Schwarzer Pfeffer sowie 4 Sideritis-
Arten (S. condensata, S. stricta, S. arguta und S. sipylea), die aufgrund ihres bekanntermaßen
sedativen, antidepressiven oder lokalanästhetischen Effektes für die Untersuchung ausgewählt
wurden, auf den humanen α1β2 GABAA-Rezeptor elektrophysiologisch mittels TEVC-Technik
untersucht. Ziel war hierbei die Mechanismen zu identifizieren, über die die ausgewählten
Lebensmittel ihre genannten neurophysiologischen Effekte entfalten.
Im Verlauf dieser Arbeit zeigten mehrere wässrige Lebensmittelextrakte, bei deren Herstellung
die Extraktionsdauer eine Stunde betrug, einen unspezifischen Strom in den nicht transfizierten
Oocyten. Dieses Ergebnis steht im Einklang mit vorherigen Studien, die den Einfluss von
wässrigen Tee-Extrakten sowie von deren polyphenolischen Inhaltsstoffen auf GABAA
Rezeptoren untersuchten [194, 197]. Dabei wurde gezeigt, dass Polyphenole durch eine direkte
starke Bindung an Membranen oder durch die Produktion von Wasserstoffperoxid die
Membranstruktur stören können [197]. Dementsprechend schädigten wässrige Tee-Extrakte
aufgrund ihres hohen Gehaltes an Polyphenolen die Oocytemembran [194]. Infolge dieser
Membranschädigung kann eine Stromamplitude in nicht-injizierten Oocyte verursacht werden
[194, 197]. Um das Auftreten dieser unerwünschten Wirkung auszuschließen, wurde eine zweite
Extraktionsmethode mit verkürzter Extraktionszeit (15 Minuten) etabliert, wodurch vermutlich
weniger membranschädigende Substanzen, insbesondere Polyphenole, aus den Lebensmitteln
gelöst wurden. Diese wässrigen Lebensmittelextrakte veränderten die elektrophysiologischen
Eigenschaften der Oocyten nicht.
Es zeigte sich, dass alle untersuchten 13 Lebensmittelextrakte auf den humanen GABAA-
Rezeptor wirkten und die Aktivität des Rezeptors bereits in einem niedrigen
Konzentrationsbereich (0,5 - 5 µg/mL) signifikant potenzierten. In der Literatur wurde zuvor
berichtet, dass organische Extrakte einiger Lebensmitteln wie Sideritis-Arten, schwarzer Pfeffer,
Kaffee, und Whisky die Aktivität des GABAA-Rezeptors verstärkten [84, 85, 90-92, 103], während
die wässrigen Lebensmittelextrakte wie Kaffee, Grüner Tee, Schwarz Tee den GABAA-Rezeptor

Modulation des GABAA-Rezeptors
58
inhibierten [83, 85]. In der vorliegenden Arbeit wurde erstmals die Potenzierung der Aktivität
des GABAA-Rezeptors durch wässrige Extrakte verschiedener Lebensmittel nachgewiesen. Da die
Auswahl der untersuchten Lebensmittel aufgrund ihrer in der Literatur beschriebenen
neurophysiologischen Eigenschaften wie der sedativen, antidepressiven oder
lokalanästhetischen Wirkung erfolgte, kann vermutet werden, dass diese ausgewählten
Lebensmittel ihre Wirkung über einen gemeinsamen Mechanismus, d.h. die Potenzierung der
GABAA-Rezeptoren, ausüben könnten.
Es konnte gezeigt werden, dass wässriger Nelkenextrakt, der potenteste GABAA-Rezeptor-
Modulator war und einen Anstieg des GABA-induzierten Stroms auf 426 % im Vergleich zur
GABA-Kontrolle auslöste. Dies deutet darauf hin, dass wässriger Nelkenextrakt eine oder
mehrere aktive GABAerge Komponenten enthalten könnte, die einzeln oder synergetisch
miteinander auf GABAA-Rezeptoren wirken. Die angstlösende, hypnotische und antiepileptische
Wirkung von Nelkenöl wurde bereits bei Mäusen beobachtet [129]. Zudem wurde ein
analgetischer Effekt durch Nelkenextrakt nicht nur an Mäusen [126, 127] sondern auch am
Menschen nachgewiesen [128]. Da die Potenzierung des GABAA-Rezeptors zur Behandlung der
neuropathischen und chronischen Schmerzen genutzt wird [54, 73], könnte die positive GABA-
modulatorische Wirkung von Nelken, wie sie in der vorliegenden Arbeit beobachtet wurde, den
in vivo gefundenen analgetischen Effekt erklären.
Bei einer Konzentration von 5 µg/mL verstärkten die wässrigen Extrakte von Melisse, Hopfen,
grünem Tee und verschiedener Sideritis-Arten die GABA-induzierten Ströme um mehr als 340 %
im Vergleich zur GABA-Kontrolle. Es wurde schon früher die antidepressive und sedative
Wirkung der Melisse an Mäusen anhand eines erzwungenen Schwimmtests untersucht. Hierbei
wurde demonstriert, dass der wässrige Extrakt von Melisse eine antidepressive Wirkung hat, die
sich durch eine Erhöhung der Schwimmbewegungen äußerte und zum anderen sedierend wirkt,
was durch eine verminderte spontane motorische Aktivität nachgewiesen wurde. Das ätherische
Öl der Melisse zeigte dagegen keinen sedativen Effekt, sondern ausschließlich einen
antidepressiven Effekt [109]. Diese Unterschiede deuten darauf hin, dass allein ein wässriger
Melissenextrakt die eine Sedierung auslösenden Komponenten in einer wirksamen
Konzentration enthält. Neben Melisse wurde auch für Hopfen eine sedative Wirkung im
Tierversuch nachgewiesen. Franco et al. berichtete, dass ein Trockenextrakt aus Hopfen eine
Verminderung der lokomotorischen Aktivität bei Wachteln sowohl tagsüber, als auch nachts
verursacht, wobei Wachteln einen dem Menschen ähnlichen Schlaf-Wach-Rhythmus aufweisen
[144]. Zudem wurde in einer Humanstudie gezeigt, dass alkoholfreies Bier aufgrund seines
Hopfen-Gehaltes sedativ wirkt [146]. Die in der vorliegenden Arbeit beschriebene Verstärkung
der α1β2 GABAA-Rezeptor-Aktivität durch wässrige Melissen- und Hopfen-Extrakte könnte beim

Modulation des GABAA-Rezeptors
59
Auftreten dieser sedierenden Wirkung eine Rolle spielen, da die α1-Untereinheit des GABAA-
Rezeptors an der Vermittlung sedativer neuronaler Signale beteiligt ist [16, 50, 51, 55, 76].
Obwohl bisher kaum in vivo Studien zur sedativen Wirkung von Sideritis-Arten durchgeführt
wurden, liegen bereits Hinweise auf einen analgetischen Effekt vor [112-114]. Ein wässriger
Sideritis-Extrakt zeigte bei einem Writhing-Test (Krümmtest) bei Mäusen eine analgetische
Wirkung [114]. Neben diesem analgetischen Effekt wurde eine angstlösende Wirkung wässriger
Sideritis-Extrakte bei Mäusen nachgewiesen [116]. In einer kürzlich veröffentlichten Studie von
Kessler et al. wurde die modulierende Wirkung von Dichlormethan-Extrakten verschiedener
Sideritis-Arten (S. condensata, S. stricta, S. arguta und S. sipylea) auf GABAA-Rezeptoren mit
Hilfe zwei verschiedener elektropyhsiologischer Methoden untersucht. Dabei wurde eine
Potenzierung der GABA-vermittelten Ströme durch die flüchtigen Inhaltsstoffe von Sideritis
sowohl mittels einer Patch-Clamp-Technik an mit dem rekombinanten α1β2γ2L-GABAA-Rezeptor
der Ratte transfizierten HEK293-Zellen, als auch mittels einer TEVC-Technik an mit dem α1β2-
GABAA-Rezeptor der Ratte transfizierten Oocyten nachgewiesen [103]. Zusätzlich wurden
mehrere Terpenoide in den Sideritis-Arten identifiziert, bei denen es sich um positive
Modulatoren des GABAA-Rezeptors handelt [87]. In der vorliegenden Arbeit erfolgte die
Verstärkung der Aktivität des rekombinant in Xenopus leavis Oocyten exprimierten humanen
α1β2-GABAA-Rezeptors durch wässrige Extrakte der Sideritis-Arten S. condensata, S. stricta, S.
arguta und S. sipylea. Da die GABA-potenzierende Wirkung bereits in sehr geringen
Konzentrationen (teilweise bei 0,5 µg/mL) beobachtet wurde, ist naheliegend, dass auch neben
den Terpenoiden nicht-flüchtige Komponenten der Sideritis-Arten wichtig für ihre modulierende
Wirkung auf GABAA-Rezeptoren sind.
Hossain et al. zeigten, dass der wässrige Extrakt des grünen Tees die GABAA-Rezeptor-
Stromantwort inhibieren kann [83]. Im Gegensatz zu dieser Beobachtung wurde in der
vorliegenden Arbeit eine signifikant verstärkte Aktivität der GABAA-Rezeptoren um 347 %
(versus GABA-Kontrolle) durch wässrigen Extrakt grünen Tees in einer Konzentration von 5
µg/mL gefunden. Diese Diskrepanz könnte sich unter anderem durch die unterschiedlichen
Extraktionsverfahren erklären. Im Gegensatz zu den vorliegenden Experimenten extrahierten
Hossain et al. den grünen Tee mit der sogenannten Ringerlösung (115 mM NaCl, 1 mM KCl, und
1,8 mM CaCl2 in einem Tris-Puffer), wodurch vermutlich die aktive Komponente aus dem Tee
nicht in ausreichenden Konzentration gelöst wurden. Grüner Tee enthält verschiedene
physiologisch aktive Komponenten mit gegenläufigen Effekten (Agonist oder Antagonist) auf die
Aktivität von GABAA-Rezeptoren. Zum Beispiel wurde durch die Applikation von Koffein [79, 83]
sowie Epigallocatechingallat [96] eine Verminderung der GABA-induzierten Chloridionenströme
beobachtet. Eine Potenzierung der Aktivierung von GABAA-Rezeptoren wurde hingegen durch
die im grünen Tee enthaltenen Alkohole, wie beispielsweise Blätteralkohol oder Linalool,

Modulation des GABAA-Rezeptors
60
nachgewiesen [83]. Es ist anzunehmen, dass die Extraktionsverfahren auf die
Konzentrationsprofile der hemmenden oder aktivierenden Substanzen im Extrakt einen
entscheidenden Einfluss haben können. Des Weiteren unterscheiden sich die untersuchten
GABAA-Rezeptoren in der Zusammensetzung ihrer Untereinheiten; so exprimierten Hossain et
al. den α1β1 Rinder-GABAA-Rezeptor in Xenopus leavis Oocyten [83], während in der
vorliegender Arbeit mit humanen α1 und β2 Untereinheiten des GABAA-Rezeptors injizierten
Oocyten verwendet wurden. Es wurde berichtet, dass manche Komponenten eine Untereinheit-
spezifische Modulation der GABAA-Rezeptoren zeigen. Beispielweise inhibiert (2-
Hydroxybenzyliden)salicylohydrazid die Aktivität der β1 enthaltenen GABAA-Rezeptoren, nicht
jedoch derjenigen GABAA-Rezeptoren, die aus β2 oder β3 Untereinheiten bestehen [198].
Ebenso konnte von Halliwell et al. gezeigt werden, dass die antikonvulsive Mefenaminsäure
GABAA-Rezeptoren mit einer β1 Untereinheit hemmte, wohingegen sie sowohl eine
Potenzierung als auch eine direkte Aktivierung von GABAA-Rezeptoren aus β2 oder β3
Untereinheiten verursachte [57].
In dieser Arbeit wurde eine Potenzierung der GABA-vermittelten Stromantwort von 200 – 300 %
durch wässrige Extrakte von Kamille, Salbei, Lavendel, Kakao und schwarzem Pfeffer
beobachtet. In zahlreichen Tierversuchen wurde die sedative, schlaffördernde und angstlösende
Wirkung von Kamille und von Flavanoiden, die aus Kamille isoliert wurden, demonstriert [123,
124, 199]. Eine klinische Studie zeigte, dass Kamille eine antidepressive Wirkung ausüben kann
[125]. Messaoudi et al. wiesen in ihrer Studie an Ratten nach, dass der polyphenolische Extrakt
aus Kakao ebenfalls eine antidepressive Wirkung aufwies [148]. Ebenso berichteten Naderi et
al., dass der hydroalkoholische Extrakt von Salvia (Salbei) sp. verticillata einen antidepressiven
und darüber hinaus antiepileptischen Effekt bei Mäusen auflöste. Dabei konnten sie keine
angstlösende Wirkung von Salbei feststellen [105]. Des Weiteren zeigte Salvia officinalis eine
antikonvulsive Wirkung, die zu einer erhöhte Beschlagnahme-Schwelle bei Mäusen führte [108].
In einer elektrophysiologischen Studie wurde von Zaugg et al. festgestellt, dass der Ethylacetat-
Extrakt von schwarzem Pfeffer (Piper nigrum L.) bei der Konzentration von 100 µg/mL zu einer
Verstärkung von GABA-vermittelter Stromantwort der GABAA-Rezeptoren auf 169,1 ± 2,4 %
führte. Die Untersuchungen wurden hierbei an GABAA-Rezeptoren durchgeführt, die mittels
Injektion der cRNA von α1-, β2- und γ2s- GABAA-Rezeptor-Untereinheiten der Ratte in Xenopus
leavis Oocyten exprimiert wurden [92]. In der vorliegenden Arbeit wurde ermittelt, dass der
wässrige Extrakt von schwarzem Pfeffer die Aktivität des α1β2 GABAA-Rezeptors
konzentrationsabhängig verstärkte. Bei der höchsten Konzentration von 5 µg/mL erreichte diese
Wirkung eine 274 % -ige Potenzierung der GABA-vermittelten Ströme. Diese Beobachtung stützt
die Annahme, dass der modulierende Effekt von schwarzem Pfeffer auf GABAA-Rezeptoren
unabhängig von der γ2-Untereinheit erfolgt. Im Vergleich zum Ethylacetat-Extrakt aus

Modulation des GABAA-Rezeptors
61
schwarzem Pfeffer [92] könnte der wässrige schwarze Pfeffer-Extrakt die GABAA-Rezeptoren
eventuell noch stärker potenzieren.
Die schlaffördernde und angstlösende Wirkung von ätherischem Lavendelöl wurde in Human-
und Tierversuchen demonstriert [100], wobei diese Effekte mit der Potenzierung der GABAA-
Rezeptoren in Verbindung gebracht wurden [80, 103]. Obwohl die Wirkungen von Lavendel
bereits in klinischen Studien gezeigt wurden, wurde in dieser Arbeit beobachtet, dass
Lavendelextrakt die durch GABAA-Rezeptoren vermittelten Ströme zwar signifikant potenzierte,
jedoch einige im Vergleich zu den anderen getesteten Lebensmittelextrakten deutlich geringere
Aktivität aufwies. Die starke in vitro ermittelte GABAA-Rezeptor-modulierende Wirkung einiger
anderer Lebensmittelextrakte könnte daher auf ein bedeutendes Potential für eine klinische
Applikation hindeuten. Allerdings zeigte Lavendel auch eine Wirkung auf die
spannungsgesteuerten Calciumkanäle, was seinen GABAA-Rezeptor-vermittelten Effekt weiter
vergrößern könnte [104].
Es wurde schon früher berichtet, dass der Neurotransmitter GABA, welcher der natürliche
Agonist des GABAA-Rezeptors ist, in verschiedenen Lebensmitteln enthalten sein kann [190-
193], unter anderem in schwarzem [187, 189] und grünem Tee [186-189]. Mit Ausnahme von
Nelkenextrakt, bei dem die Konzentration der GABA unterhalb der NWG lag, konnte GABA in
allen untersuchten wässrigen Lebensmittelextrakten in dieser Arbeit detektiert werden. Als eine
bioaktive Komponente in Lebensmitteln kann GABA im menschlichen Körper mehrere wichtige
physiologische Funktionen beeinflussen [200]. Zum Beispiel kann GABA in Lebensmitteln eine
antidepressive [75] und stressreduzierende [201] Wirkung auslösen. Zudem wirkt Maoyecha
Tee, der einen hohen Gehalt an GABA aufweist, schlaffördernd, wahrscheinlich über die
Verstärkung der GABAergen Neurotransmission [202]. Darüber hinaus wird in der Literatur
berichtet, dass wässrige Tee-Extrakte wie grüner Tee, schwarzer Tee und Oolong Tee in
Abwesenheit von GABA die durch GABAA-Rezeptoren vermittelten Chloridionenströme
induzierten, wahrscheinlich aufgrund von GABA oder GABA-ähnlichen Verbindung(en) in den
Tee-Extrakten [83]. Die hier dargestellten Versuche zeigten jedoch, dass die gemessenen GABAA-
Rezeptor-vermittelten Ströme in Abwesenheit und Anwesenheit von GABA nicht durch die sehr
geringe Menge an endogener GABA in den Lebensmittelextrakten erzeugt wurden. Die
Verstärkung der GABA-induzierten Ströme ist höchstwahrscheinlich auf allosterische
Modulatoren des GABAA-Rezeptors, die in den Lebensmittelextrakten vorhanden sind,
zurückzuführen.
Um festzustellen, ob die beobachtete neurophysiologische Aktivität tatsächlich spezifisch für
den GABAA-Rezeptor ist, wurde auch die Wirkung der Lebensmittelextrakte auf den Glycin-
Rezeptor untersucht, der ebenfalls zur Cys-Loop-Rezeptorsuperfamilie der ligandengesteuerten
Ionenkanäle gehört [13, 14]. Neben GABAA- und Glycin-Rezeptoren zählen auch die

Modulation des GABAA-Rezeptors
62
nikotinischen Acetylcholinrezeptoren, Serotonin Typ3 Rezeptoren und Zink-aktivierte
Ionenkanäle zur Unterfamilien der Cys-Loop-Rezeptorsuperfamilie. Alle Mitglieder dieser
Superfamilie haben eine ähnliche pentamere Struktur. Dabei sind GABAA- und Glycin-
Rezeptoren als Chloridionenkanäle selektiv für Anionen, wohingegen die anderen Rezeptoren
dieser Superfamilie selektive Kationenkanäle sind [13, 14]. Es wurde in dem durchgeführten
Versuch beobachtet, dass keiner der getesteten Lebensmittelextrakte die Glycin-induzierten
Ströme über den homomerischen α1-Glycin-Rezeptor verstärkte. Auch bei der höchsten Extrakt-
Konzentration von 5 µg/mL wurden die Glycin-vermittelten Stromantworten entweder nicht
moduliert oder sogar auf bis zu 56 - 109 % der Kontrolle reduziert. Im Gegensatz dazu
verstärkten die Lebensmittelextrakte in ähnlicher Konzentration die GABAA-Rezeptorantwort auf
bis zu 216-426%. Ähnliche Ergebnisse wurden für das Steroid-Anästhetikum 5β-Pregnan-3α-ol-
20-on beobachtet, das die Aktivität des α1β2γ2L GABAA-Rezeptor um bis zu 722 % potenzierte,
aber die Glycin-vermittelte Stromantwort der α1-Glycin-Rezeptoren auf 57% inhibierte [78].
Andere Steroid Anästhetika wie Alphaxalon, Minaxolon, ORG 20599 und 5α-Pregnan-3α-ol-20-
on verstärkten dagegen die Aktivität sowohl der GABAA-Rezeptoren als auch der Glycin-
Rezeptoren [78]. Ebenfalls zeigten einige Alkohole (Ethanol, Butanol, Hexanol, Octanol, Decanol
sowie Monoterpen-Alkohole Menthol und Borneol) eine konzentrationsabhängige Potenzierung
der α1β1- oder α1β2γ2s-GABAA-Rezeptoren [82, 203] und verstärkten darüber hinaus die
Aktivierung homomerischer α1- oder α2-Glycin-Rezeptoren [17, 203, 204]. Zusammenfassend
deuten die Ergebnisse der vorliegenden Untersuchung darauf hin, dass die getesteten
Lebensmittelextrakte eine spezifische positive allosterische modulierende Wirkung auf den
GABAA-Rezeptor haben und entweder keine oder eine inhibitorische Modulation der Glycin-
Rezeptoren verursachen.

63
Identifizierung der aktiven Inhaltsstoffe des 4
wässrigen Nelkenextraktes
4.1. Einleitung
Nelken, die unter anderem zum Würzen von Speisen verwendet werden, zeigen eine
lokalanästhetische Wirkung auf Menschen [128] sowie einen analgetischen und angstlösenden
Effekt auf verschiedene Tierarten [127, 129]. In Kapitel 2 wurde die inhibitorische Wirkung von
13 ausgewählten Lebensmitteln- Grüner Tee, Hopfen, Kamille, Kakao, Lavendel, Melisse, Nelken,
Salbei, Schwarzer Pfeffer sowie 4 Sideritis-Arten (S. condensata, S. stricta, S. arguta und S.
sipylea)- auf GABA-Abbauenzyme untersucht. Dabei wurde festgestellt, dass der wässrige
Nelkenextrakt eine potenteste Hemmung der GABA-Abbauenzyme hervorrief. In Kapitel 3
wurde die modulierende Wirkung von diesen Lebensmitteln auf GABAA-Rezeptoren untersucht.
Dabei zeigte sich ebenfalls der wässrige Nelkenextrakt als potentester Modulator. Da der
wässrige Nelkenextrakt die stärkste Potenzierung des GABAA-Rezeptors verursachte, ist zu
erwarten, dass er eine oder mehrere aktive Komponenten enthält, welche die GABA-induzierten
Ströme alleine oder durch additive bzw. synergistische Effekte potenzieren.
In diesem Teil der Arbeit sollte aufgeklärt werden, welche Inhaltsstoffe des wässrigen
Nelkenextraktes für die GABAA-Rezeptor modulierenden Effekt verantwortlich sind. Dazu wurde
zunächst der wässrige Nelkenextrakt mittels HPLC fraktioniert und die GABAerge Aktivität jeder
aufgefangenen Fraktion mittels TEVC elektrophysiologisch gemessen. Um die
neurophysiologisch aktiven Inhaltsstoffe von Nelken zu identifizieren, wurden die aktivste
Fraktion von Nelken, welches die stärkste Modulation des GABA-Rezeptors bewirkte, sowie der
Nelkenextrakt mittels UHPLC-DAD (Ultrahochleistungsflüssigchromatographie mit
Diodenarraydetektor) untersucht. Zur Bestimmung der nicht UV-aktiven Substanzen sowie zur
weiteren Identifizierung aktiver Inhaltsstoffe wurde der wässrigen Nelkenextrakt mit Hilfe GC-
MS (Gaschromatographie mit Massenspektrometrie) analysiert. Anschließend wurde die
neurophysiologische Aktivität der Inhaltsstoffe durch die TEVC-Technik untersucht.

Identifizierung der aktiven Inhaltsstoffe des wässrigen Nelkenextraktes
64
4.2. Aktivitätsgeleitete Fraktionierung des Nelkenextraktes
Um die GABAergen Substanzen zu isolieren, wurde der wässrige Nelkenextrakt mittels HPLC
fraktioniert. Die Auftrennung erfolgte an einer semipräparativen RP C18ec-Säule. In Abbildung
4.1 ist das erhaltene Chromatogramm dargestellt. 13 Fraktionen wurden mit Hilfe eines
automatischen Fraktionensammlers zu definierten Zeiten aufgefangen. Die Entfernung des
Lösungmittels aus den Fraktionen erfolgte durch Vakuumdestillation und anschließender
Gefriertrocknung. Zur Analyse wurden die getrockneten Fraktionen in Reinstwasser gelöst.
Um die Wirkung der Nelkenfraktionen auf die Aktivität der GABAA-Rezeptoren zu bestimmen,
wurde jede Fraktion des Nelkenextraktes in einer Konzentration von 5 µg/mL, in welcher der
wässrige Nelkenextrakt die höchste Potenzierung des GABA-induzierten Stroms auslöste (siehe
Tabelle 3.1 in Kapitel 3.2), mittels TEVC-Technik elektrophysiologisch gemessen. Hierzu wurden
die einzelnen Fraktionen in Anwesenheit des zugesetzten GABA (1 µM) auf die Xenopus leavis
Oocyte, in welcher rekombinante menschlichen α1β2 GABAA-Rezeptoren exprimiert wurden,
appliziert. Der von 1 µM GABA verursachte Strom wurde als Kontrolle auf 100 % gesetzt. Die
Potenzierung der GABA-induzierte Ströme durch die einzelnen Fraktionen wurde prozentual in
Bezug auf die Kontrolle angegeben. Die Verstärkung der GABA-vermittelten Ströme durch die
wässrigen Fraktionen ist in Abbildung 4.2 graphisch dargestellt. Alle Fraktionen potenzierten die
Aktivität von GABAA-Rezeptoren. Dabei konnte kein signifikanter Effekt der Fraktion 2, Fraktion
8, Fraktion 10 und Fraktion 12 festgestellt werden, während die Fraktionen 1, 3, 4, 5, 6, 7, 9, 11
und 13 zu einer signifikanten Verstärkung der GABAA-Rezeptor-vermittelten Ströme führten
(siehe Abbildung 4.2). Die geringste signifikante GABAerge Aktivität wurde bei der Ko-
Applikation von 1 µM GABA mit Fraktion 3, welche die GABA-induzierte Ströme um 195 %
potenzierte, beobachtet. Der maximale signifikant potenzierende Effekt wurde durch eine
simultane Applikation von Fraktion 11 mit 1 µM GABA erreicht, wobei Fraktion 11 eine
Verstärkung des GABA-vermittelten Stroms auf 780 % im Vergleich zur GABA-Kontrolle
verursachte (siehe Abbildung 4.2). Als ein Beispiel zeigt die Abbildung 4.3 die repräsentativen
Stromspuren, welche durch Applikation der Fraktion 11 mit GABA ermittelt wurden.

Identifizierung der aktiven Inhaltsstoffe des wässrigen Nelkenextraktes
65
Abbildung 4.1 RP-HPLC Chromatogramm des wässrigen Nelkenextraktes detektiert bei 254 nm. Die mittels
Fraktionensammler aufgefangenen Fraktionen sind rot markiert. Die Zahlen (1-13) zeigen die
Fraktionen-Nummer.
Abbildung 4.2 Modulation von humanen α1β2 GABAA-Rezeptoren durch Fraktionen des wässrigen Nelken-
extraktes in der Konzentration von 5 μg/mL. Die prozentualen Angaben beziehen sich jeweils
auf die GABA-Kontrolle (100%). Die Daten zeigen den Mittelwert ± SEM aus 4-10 Oocyten von
zwei unabhängig transfizierten Chargen. Signifikante Unterschiede wurden durch
zweifaktorielle-ANOVA mit Holm-Sidak Vergleichstest ermittelt (ns: nicht signifikant; * p <
0,05; ** p < 0, 01; *** p < 0,001; **** p < 0,0001 versus GABA-Kontrolle).
**
ns *
** *
****
** ns
****
ns
****
ns
****
0
200
400
600
800
1000
1200
Rel
ativ
er S
tro
m [
%]
1 2 3 4 5 6 7 8 9 10 11 12 13

Identifizierung der aktiven Inhaltsstoffe des wässrigen Nelkenextraktes
66
Abbildung 4.3 Repräsentative Stromkurven einer Messung der Koapplikation von 1 μM GABA mit Fraktion
11 des wässrigen Nelkenextraktes an humanen α1β2 GABAA-Rezeptoren.
4.3. Identifizierung der neurophysiologisch aktiven Inhaltsstoffe von Nelken
Um die aktiven Inhaltsstoffe von Nelken zu identifizieren, wurden Fraktion 11 des wässrigen
Nelkenextraktes, welche die stärkste potenzierende Modulation des GABAA-Rezeptors aufzeigt,
sowie der Nelkenextrakt mittels zweier chromatographischer Trennmethoden untersucht. Zur
Identifizierung der UV-aktiven Substanzen wurde eine UHPLC-DAD Methode durchgeführt. Die
Untersuchung der nicht UV-aktiven Substanzen sowie die Identifizierung der Wirkstoffe
erfolgten durch GC-MS Analyse.
4.3.1. Identifizierung und Quantifizierung von Eugenol im Nelkenextrakt und in der aktivsten Nelken-Fraktion 11
Zur Identifizierung der Inhaltsstoffe wurde der wässrige Nelkenextrakt, sowie die aktivste
Fraktion des wässrigen Nelkenextraktes mittels UHPLC (Ultrahochleistungsflüssigchro-
matographie; aus dem Englischen: ultra high-performance liquid chromatography) an einer C18
Säule aufgetrennt. Die Detektion erfolgte mittels Diodenarraydetektor. Abbildung 4.4.A zeigt
das UHPLC-DAD-Chromatogramm der Nelken-Fraktion 11 bei 282 nm. In der Fraktion 11 von
Nelken trat ein großes Signal bei einer Retentionszeit von 6,9 Minuten auf. Auf Grund des
UV/Vis-Spektrums könnte dieses Hauptsignal Eugenol oder ein Eugenol-Derivat sein. Daher
wurde die authentische Referenzsubstanz Eugenol unter Verwendung gleicher UHPLC-
Parameter analysiert. Anschließend wurden die Retentionszeit und das UV/Vis-Spektrum des
Referenzstandards Eugenol mit denen der unbekannten Verbindung verglichen, um die Struktur
dieser unbekannten Verbindung aufzuklären.

Identifizierung der aktiven Inhaltsstoffe des wässrigen Nelkenextraktes
67
Abbildung 4.4 UHPLC-DAD-Chromatogramm der Fraktion 11 von Nelken (A) und des wässrigen
Nelkenextraktes (B), detektiert bei 282 nm.
Dabei zeigte Eugenol exakt die gleiche Retentionszeit (siehe Abbildung 4.5) und identisches
UV/Vis- Spektrum (siehe Abbildung 4.6) wie unbekannte Verbindung. Auf diese Weise wurde
das Hauptsignal in der Fraktion 11 von Nelken als Eugenol identifiziert. Ebenfalls wurde dasselbe
Signal mit einer Retentionszeit von 6,9 Minuten im unfraktionierten Nelkenextrakt beobachtet,
wie in Abbildung 4.4.B aufgezeichnet ist. Da sowohl die Retentionszeit (siehe Abbildung 4.7) als
auch UV/Vis-Spektren (siehe Abbildung 4.6) des Hauptsignals und der Eugenol-Standardlösung

Identifizierung der aktiven Inhaltsstoffe des wässrigen Nelkenextraktes
68
identisch waren, konnte die Anwesenheit von Eugenol auch im unfraktionierten Nelkenextrakt
bestätigt werden.
Abbildung 4.5 Übereinandergelegte UHPLC-DAD-Chromatogramme der Eugenol-Standardlösung (rot) und
der Nelken-Fraktion 11 (blau). Detektion erfolgte bei 282 nm.
Abbildung 4.6 UV/Vis-Spektren der Eugenol-Standardlösung (A), der Nelken-Fraktion 11 (B) und des
Nelkenextraktes (C) bei einer Retentionszeit von 6,9 min.

Identifizierung der aktiven Inhaltsstoffe des wässrigen Nelkenextraktes
69
Abbildung 4.7 Übereinandergelegte UHPLC-DAD-Chromatogramme der Eugenol-Standardlösung (rot), der
Acetyleugenol-Standardlösung (orange) und des wässrigen Nelkenextraktes (blau). Detektion
erfolgte bei 282 nm.
Um die Identität des Eugenols im Nelkenextrakt weiter zu verifizieren, wurde eine GC-MS
(Gaschromatographie mit Massenspektrometrie) Analyse durchgeführt. Dabei wurde ein sehr
intensives Signal bei einer Retentionszeit von 26,3 Minuten im Nelkenextrakt detektiert (TIC-
Chromatogramm siehe Abbildung 4.8.A). Massenspektrometrische Untersuchung lieferte für
dieses Signal ein Molekulargewicht von 164 g/mol. Massenspektren des Signals wurden mit der
Spektren der NIST-Datenbank verglichen, wodurch dies Hauptsignal als Eugenol identifiziert
wurde (siehe Abbildung 4.8.B). Folglich konnte die Anwesenheit von Eugenol im Nelkenextrakt
eindeutig bestätigt werden.

Identifizierung der aktiven Inhaltsstoffe des wässrigen Nelkenextraktes
70
A)
B)
C)
Abbildung 4.8 TIC -Chromatogramm des wässrigen NelkenExtraktes detektiert mittels GC-MS. Im Fokus sind
Massenspektren des großen Signals bei einer Retentionszeit von 26,3 min (link) und des
kleinen Signals bei einer Retentionszeit von 27,3 min (recht) dargestellt (A).
Nebeneinanderliegende Massenspektren des bei 26,3 min eluierten Signal (blau) und des
Eugenols entnommen aus der NIST-Datenbank (rot) (B). Nebeneinanderliegende
Massenspektren des Signals mit Retentionszeit von 27,3 min (blau) und des Acetyleugenols,
das mittels der NIST-Datenbank ermittelt wurde (rot) (C).
Im unfraktionierten Nelkenextrakt sowie in der Fraktion 11 wurde Eugenol mittels UHPLC-DAD
quantifiziert. Die Quantifizierung von Eugenol erfolgte durch eine externe Kalibrierung anhand

Identifizierung der aktiven Inhaltsstoffe des wässrigen Nelkenextraktes
71
des Kalibrierungsverfahrens. Dazu wurde Eugenol-Standardlösung in einem
Konzentrationsbereich zwischen 0,5 µg/mL und 50 µg/mL gemessen. Die Linearität,
Nachweisgrenze (NWG) sowie Bestimmungsgrenze (BSG) konnten über eine Kalibriergerade
bestimmt werden, welche aus der Korrelation zwischen gemessene Peakflächen und
eingesetzten Konzentrationen von Eugenol ermittelt wurde. Die lineare Regression für die
Kalibrierungskurve war y=0,3553x-0,0103 mit einem Determinationskoeffizient (R2) von 0,9996.
Als NWG wurden 0,53 µg/mL und als BSG 1,6 µg/mL berechnet. Die Berechnung der Eugenol-
Konzentrationen des Nelkenextraktes und seiner Fraktion 11 erfolgte über die externe
Kalibrierkurve (siehe Abbildung im Anhang A.3). Für den Nelkenextrakt (0,25 mg/mL) wurde
eine Eugenol-Konzentration von 42,6 ± 0,5 µg/mL und für die Nelken-Fraktion 11 (1 mg/mL) eine
Eugenol-Konzentration von 22,8 ± 0,1 µg/mL bestimmt.
4.3.2. Untersuchung der GABAA-Rezeptor modulierenden Wirkung von Eugenol
Als Hauptkomponente wurde Eugenol sowohl im unfraktionierten Nelkenextrakt als auch in der
aktivsten Nelken-Fraktion 11 nachgewiesen und quantifiziert (siehe Kapitel 4.3.1). Dadurch
wurde eine Eugenol-Konzentration von 0,1 µg/mL in der Nelken-Fraktion 11 (5 µg/mL) und von 1
µg/mL im Nelkenextrakt (5 µg/mL) berechnet. Um den Beitrag von Eugenol zur gesamten
GABAA-Rezeptor-modulierenden Aktivität des NelkenExtraktes zu bestimmen, wurden diese
Konzentrationen von Eugenol in Anwesenheit von 1 µM GABA an den mit rekombinanten
humanen α1β2 GABAA-Rezeptoren transfizierten Oocyten von Xenopus leavis mittels TEVC-
Technik getestet. Parallel dazu wurde Nelken-Fraktion 11 und Nelkenextrakt mit 1 µM GABA
koappliziert. Abbildung 4.9 stellt repräsentative GABA-induzierte Stromspuren dar, die durch Ko-
Applikation von Eugenol, Nelkenextrakt und Nelken-Fraktion 11 mit 1 µM GABA verstärkt
wurden.

Identifizierung der aktiven Inhaltsstoffe des wässrigen Nelkenextraktes
72
Abbildung 4.9 Repräsentative Stromkurven einer Messung der Koapplikation von 1 µM GABA mit Eugenol,
der Nelken-Fraktion 11 und dem wässrigen Nelkenextrakt an humanen α1β2 GABAA-
Rezeptoren. Die blauen Stromspuren zeigen die durch 1 µM GABA erzeugten Amplituden.
Zur Berechnung der prozentualen Potenzierung wurde der Mittelwert der vor und nach
Koapplikation durch 1 µM GABA ausgelöste Strom als GABA-Kontrolle auf 100 % gesetzt. Die
Potenzierungen der GABAA-Rezeptor-vermittelten Ströme durch Eugenol, Nelkenextrakt und
seine Fraktion 11 sind in Abbildung 4.11.A graphisch gezeigt. Der durch 0,1 µg/mL Eugenol
hervorgerufene Strom betrug 152 ± 18 % bezogen auf die GABA-Kontrolle, während Fraktion 11
von Nelken eine Verstärkung um 188 ± 34 % verursachte. Dabei zeigte die zweifaktorielle-
ANOVA mit Holm-Sidak Vergleichstest, dass keine signifikanten Unterschiede zwischen den
potenzierenden Effekten von 0,1 µg/mL Eugenol und von Nelken-Fraktion 11 auf GABAA-
Rezeptoren bestanden (p=0,61, zweifaktorielle ANOVA mit Holm-Sidak Vergleichstest). Der
Einsatz von Eugenol in höheren Konzentrationen (1 µg/mL) führte zu einer signifikanten
Verstärkung des GABA-induzierten Stroms um ca. 230 % im Vergleich zur GABA-Kontrolle. Dabei
zeigten sich keine signifikante Unterschiede zwischen relativen Stromantworten von 1 µg/mL
Eugenol und des Nelkenextraktes (p=0,35, zweifaktorielle ANOVA mit Holm-Sidak
Vergleichstest). Daraus kann geschlussfolgert werden, dass Eugenol die wichtigste Determinante
der GABAA-Rezeptor modulierende Wirkung des Nelkenextraktes darstellt. Es kann jedoch nicht

Identifizierung der aktiven Inhaltsstoffe des wässrigen Nelkenextraktes
73
ausgeschlossen werden, dass andere Komponenten zu einem geringen Teil ebenfalls zur
Aktivität beitragen.
4.3.3. Identifizierung von anderen Inhaltsstoffe im Nelkenextrakt und in der aktivsten Nelken-Fraktion 11 sowie Untersuchung zu deren GABAA-Rezeptor modulierenden Wirkung
Im wässrigen Nelkenextrakt wurde ein kleiner Peak bei einer Retentionszeit von 7,9 Minuten
detektiert. Dieser nach dem Eugenol-Signal auftretende Peak könnte auf Grund seines UV/Vis
Spektrums ein Derivat von Eugenol sein. Um diese Hypothese belegen zu können, wurde eine
Standardlösung von Acetyleuenol in gleicher Weise mittels UHPLC/DAD analysiert. Da der
Acetyleuenol-Referenzstandard identische Retentionszeit (siehe Abbildung 4.7) sowie UV/Vis-
Spektren (siehe Abbildung 4.10) zeigte wie unbekannte Verbindung, konnte die Anwesenheit
von Acetyleugenol in unfraktionierten Nelkenextrakt belegt werden. Weiterhin konnte mittels
GC-MS die Anwesenheit von Acetyleugenol im Nelkenextrakt eindeutig verifiziert werden (siehe
Abbildung 4.8). In der Nelken-Fraktion 11 konnte auch ein Signal mit einer Retentionszeit von
8,2 Minuten nach der Elution von Eugenol detektiert (siehe Abbildung 4.4.A). Jedoch handelte
es sich bei diesem Signal nicht um Acetyleugenol.
Abbildung 4.10 UV/Vis-Spektren der Acetyleugenol-Standardlösung (A) und des Nelkenextraktes (B) bei einer
Retentionszeit von 7,9 min.
Aufgrund der geringeren Konzentrationen konnte das Acetyleugenol im Nelkenextrakt nicht
quantifiziert werden. Um die GABAA-Rezeptor modulierende Wirkung von Acetyleugenol mit der
Wirkung von Eugenol zu vergleichen, wurde Acetyleugenol in einer Konzentration von 1 µg/mL
in Anwesenheit von 1 µM GABA mittels TEVC-Technik den Oocyten, die mit rekombinanten
humanen α1β2 GABAA-Rezeptoren transfiziert wurden, wie Eugenol koappliziert. In der
Applikationskonzentration von 1 µg/mL löste Acetyleugenol eine Verstärkung der GABA-
vermittelten Ströme auf 308 ± 26 % im Vergleich zur GABA-Kontrolle aus (siehe Abbildung
4.11.B). Dabei war die durch Acetyleugenol hervorgerufene Steigerung des GABA-vermittelten

Identifizierung der aktiven Inhaltsstoffe des wässrigen Nelkenextraktes
74
Stroms signifikant (p < 0,05) höher war als der durch Eugenol ausgelöste Effekt (234 ± 29 %). Die
repräsentativen Ergebnisse einer Applikation des Acetyleugenol sind in Abbildung 4.12
dargestellt.
Abbildung 4.11 Die Potenzierung der Aktivität von humanen α1β2 GABAA-Rezeptoren in den Xenopus leavis
Oocyten durch Fraktion 11, Eugenol und Nelken (A) und sowie durch Acetyleugenol (B) bei
der simultanen Applikation mit 1 μM GABA. Die prozentualen Angaben beziehen sich jeweils
auf die GABA-Kontrolle (100%). Die Daten zeigen den Mittelwert ± SEM aus 5-10 Oocyten
von 2-3 unabhängig transfizierten Chargen. Signifikante Unterschiede wurden durch
zweifaktorielle-ANOVA mit Holm-Sidak Vergleichstest ermittelt (ns: nicht signifikanter
Unterschied,*p<0.05, **p<0.01, ***p<0.001, und **** p < 0,0001).
Obwohl sich Acetyleugenol als ein stärkerer Modulator des GABAA-Rezeptors erwies, ist sein
Beitrag zur Aktivität des wässrigen Nelkenextraktes eher gering, da es nur in Spuren vorhanden
ist. Im Gegensatz dazu wurde berichtet, dass Nelkenknospenöl bis zu 85 % Eugenol und bis zu 21
% Acetyleugenol enthält [135, 136]. Daher wurde die durch 0,25 mg/mL Acetyleugenol
ausgelöste Potenzierung der GABA-vermittelten Ströme mit der durch 1 µg/mL Eugenol
hervorgerufene Verstärkung der Ströme verglichen. In dieser Konzentration potenzierte
Acetyleugenols die Aktivität der GABAA-Rezeptoren um 222 ± 32 % bezogen auf GABA-Kontrolle
(siehe Abbildung 4.11.B). Dabei zeigten sich keine signifikanten Unterschiede zwischen den
GABAA-Rezeptor-potenzierenden Effekt von 0,25 µg/mL Acetyleugenol und von 1 µg/mL
Eugenol (p = 0,93). Daher kann davon ausgegangen werden, dass im Gegensatz zum wässrigen
Nelkenextrakt Acetyleugenol im Nelkenknospenöl eine ähnliche Bedeutung für die GABAA-
Rezeptor-modulatorische Aktivität haben kann wie Eugenol. Aufgrund seiner geringen
Wasserlöslichkeit kann es jedoch im wässrigen Extrakt nicht erfasst werden.
* **
***
****
0
100
200
300
400
500
Rel
ativ
er S
tro
m [
%]
A ns
ns
** ****
****
0
100
200
300
400
500
Rel
ativ
er S
tro
m [
%]
B *

Identifizierung der aktiven Inhaltsstoffe des wässrigen Nelkenextraktes
75
Abbildung 4.12 Repräsentative Stromkurven einer Messung für die Koapplikation der 1 µM GABA mit
Acetyleugenol an humanen α1β2 GABAA-Rezeptoren.
Darüber hinaus wurde β-Caryophyllen schon im ätherischen Öl der Nelkenknospen identifiziert
[131-136]. Da β-Caryophyllen bei hohen Konzentrationen eine GABAA-Rezeptor-modulierende
Aktivität hat [87], wurde seine Anwesenheit im Nelkenextrakt mittels GC-MS analysiert.
Allerdings konnte β-Caryophyllen im wässrigen Nelkenextrakt nicht detektiert werden. Dies
deutet darauf hin, dass es nicht genügend wasserlöslich ist.
4.4. Diskussion und Zusammenfassung
Die GABAerge Neurotransmission kann durch verschiedene Lebensmittel wie Tee, Gewürze und
Getränke [83-85, 90-93, 103], sowie durch Lebensmittelinhaltsstoffe wie Terpenoide und
Flavanoide moduliert werden [27, 80-96]. In Kapitel 3 wurde für den wässrigen Nelkenextrakt
die stärkste GABAA- Rezeptor modulierende Wirkung von insgesamt 13 getesteten wässrigen
Lebensmittelextrakten gemessen. Daher sollten in diesem Teil der Arbeit die Inhaltsstoffe im
Nelkenextrakt identifiziert werden, welche für dessen GABAA-Rezeptor modulierenden Effekt
verantwortlich sind. Hierzu wurde eine HPLC-basierte Methode zur Profilierung der GABAA-
Rezeptor potenzierenden Aktivität des Nelkenextraktes etabliert, wodurch die bioaktiven
Inhaltsstoffe identifiziert werden konnten.
Trotz zahlreicher Studien, in denen die Inhaltsstoffe des Nelkenknospenöls beschrieben
wurden, ist bisher wenig Literatur zur Identifizierung der Inhaltsstoffe wässriger Nelkenextrakte
bekannt. Eugenol wurde als Hauptbestandteil des Nelkenknospenöls identifiziert [130-136].
Neben Eugenol enthält das ätherische Öl der Nelkenknospen Acetyleugenol [130-132, 134-136]
und β-Caryophyllen [131-136]. β-Caryophyllen, das die Aktivität des GABAA-Rezeptors verstärkt
[87], wurde in dieser Arbeit im wässrigen Extrakt nicht detektiert. Dagegen konnten Eugenol, als

Identifizierung der aktiven Inhaltsstoffe des wässrigen Nelkenextraktes
76
Hauptbestandteil, und Acetyleugenol, als eine Minorkomponente, im wässrigen Extrakt der
Nelkenknospen anhand der UV/Vis-Spektren, die mittels UHPLC-DAD ermittelt wurden,
identifiziert werden. Dies stimmte mit dem Ergebnis der GC-MS-Analyse überein, sodass
Eugenol und Acetyleugenol anhand der Massenspektren nach gaschromatographischer
Auftrennung als Haupt- bzw. Nebenkomponenten des wässrigen Extraktes von Nelken
identifiziert wurden. Zudem konnte Eugenol in Fraktion 11, die GABA-aktivste Fraktion des
wässrigen Nelkenextraktes, mittels UHPLC-DAD identifiziert sowie quantifiziert werden. Die
Quantifizierung ergab einen Eugenolgehalt von 1 µg/mL im wässrigen Nelkenextrakt und einen
Eugenolgehalt von 0,1 µg/mL in der Fraktion 11. Wurde Eugenol in den gleichen
Konzentrationen wie im Nelkenextrakt vorhandene Konzentration appliziert, war die GABAerge
Aktivität nur leicht, aber nicht signifikant geringer als die potenzierende Wirkung des
Nelkenextraktes. Ebenso zeigten sich keine signifikanten Unterschiede zwischen den GABAA-
Rezeptor potenzierenden Effekten von Eugenol und von Nelken-Fraktion 11, wenn Eugenol in
den gleichen Konzentrationen wie in der Nelken-Fraktion vorhandene Konzentration
elektrophysiologisch getestet wurde. Diese Ergebnisse deuten darauf hin, dass Eugenol als die
Hauptkomponente des wässrigen Nelkenextraktes bzw. der Nelken-Fraktion 11 für ihren
positiven, GABAA-Rezeptor modulierenden Effekten verantwortlich ist. Allerdings ist ein Trend
zu beobachten, das Eugenol eine niedrige Aktivität zeigt als der Gesamtaktivität bzw. die
Fraktion 11. Deshalb kann ein geringer Effekt von anderen Inhaltsstoffen nicht ausgeschlossen
werden.
Eugenol, das zur Stoffgruppe der Phenylpropanoide gehört [132], ist schwach löslich in Wasser
[134] und kann die Blut-Hirn-Schranke überwinden [205]. Aoshima et al. untersuchten die
GABAerge modulierende Wirkung von Eugenol durch TEVC Technik an Oocyten, in denen der
GABAA-Rezeptor durch die Mikroinjektion der mRNA aus Rattenhirn rekombinant exprimiert
wurde. Dabei stellten sie fest, dass Eugenol in der Konzentration von 650 µM (entspricht 106,7
µg/mL) in Abhängigkeit der applizierten Konzentrationen von GABA eine Steigerung der GABA-
induzierten Chloridionenströme hervorrief, wodurch sich möglicherweise die Affinität von GABA
zum Rezeptor erhöhte [80]. Zudem wurde Eugenol als allosterischer Modulator des GABAA-
Rezeptors identifiziert, sodass es eine konzentrationsabhängige Verstärkung der 2 µM GABA-
induzierten Chloridionenströme mit einem EC50-Wert von 532 µM (entspricht 87 µg/mL) in
Primärkulturen von kortikalen Neuronen aus Mäusen verursachte [89]. Des Weiteren wurde
berichtet, dass Eugenol die Aktivität des α1β1 GABAA-Rezeptors konzentrationsabhängig
potenzierte, welcher aus Rinderhirn rekombinant in Oocyten von Xenopus leavis exprimiert
wurde [82]. In dieser Arbeit wurde die Wirkung von Eugenol auf mit rekombinanten humanen
α1β2 GABAA-Rezeptoren transfizierten Oocyten von Xenopus leavis untersucht. Dabei zeigte
sich, dass Eugenol bereits in deutlich niedrigeren Konzentrationen (0,1 µg/mL und 1 µg/mL) die
1 µM GABA-vermittelten Ströme verstärkte. Dies deutet darauf hin, dass die potenzierende

Identifizierung der aktiven Inhaltsstoffe des wässrigen Nelkenextraktes
77
Wirkung von Eugenol auf GABAA-Rezeptoren vom Typ der β-Untereinheit unabhängig ist.
Obwohl die Potenzierung des GABAA-Rezeptors durch Eugenol in oben beschriebene Studien
und auch in dieser Arbeit nachgewiesen werden konnte, wurde in einer kürzlich veröffentlichten
elektrophysiologischen Studie von Lee et al. berichtet, dass Eugenol eine
konzentrationsabhängige reversible Inhibition der 20 µM GABA-induzierten Ströme mit einem
IC50-Wert von 369,2 µM (entspricht 60,6 µg/mL) in HEK-293-Zellen, die mit α1 und β2
Untereinheiten aus Ratten-GABAA-Rezeptoren sowie humanen γ2L-Untereinheit des GABAA-
Rezeptors exprimiert wurden, auslöste. Dabei inhibierte 1 mM Eugenol reversibel den 500 µM
GABA-induzierten Strom in Neuronen des Trigeminusganglions von Ratten [206]. Diese
Diskrepanz erklärt sich unter anderem durch die unterschiedlichen Expressionssysteme bzw.
Neuronen. Die positiv modulierende Wirkung von Eugenol konnte auf den GABAA-Rezeptor,
welcher in Xenopus leavis Oocyten transfiziert wurde oder in Primärkulturen von kortikalen
Neuronen aus Mäusen vorhanden war, festgestellt werden. Die durch Eugenol ausgelöste
Inhibition wurde dagegen in HEK-Zellen oder in Neuronen Ratten-Trigeminusganglions
beobachtet. Des Weiteren kann diese gemessene Inhibition aufgrund hoher GABA-
Konzentration (20 µM für HEK-Zellen und 500 µM für Neuronen [206]) auftreten, da sich die
Potenzierung der GABA-induzierten Ströme durch Eugenol mit steigender Konzentration von
GABA reduziert [80]. Eine weitere Ursache der Inhibition kann die extrem hohe Konzentration
von Eugenol (1 mM; entspricht 164 µg/mL) sein.
In der vorliegenden Arbeit zeigte Acetyleugenol eine noch stärkere GABAA-Rezeptor
modulierende Wirkung als Eugenol. Aufgrund seiner sehr geringen Konzentrationen im
wässrigen Extrakt ist allerdings sein Beitrag zur Gesamtaktivität eher gering. Dieses Ergebnis
unterstützt die Hypothese, dass der Nelkeninhaltsstoff Eugenol einen sehr wichtigen Beitrag zur
durch Nelken ausgelösten Potenzierung der GABA-vermittelter Ströme liefert. Allerdings ist
Acetyleugenol, auf Grund seiner stark potenzierenden Effekte im Vergleich zum Eugenol, bei
Verwendung gleicher Konzentrationen, der potentere Modulator des ionotropen GABAA-
Rezeptors. Dies deutet darauf hin, dass die Acetylierung der Hydroxylgruppe zu einer weiteren
Aktivitätssteigerung führt (Strukturformeln des Eugenols und Acetyleugenols siehe Abbildung
4.13). Im Gegensatz zum wässrigen Nelkenextrakt kann Acetyleugenol aufgrund seiner stärker
GABAA-Rezeptor modulierenden Aktivität eine relevante Komponente des Nelkenknospenöls
oder eines alkoholischen Extraktes sein.

Identifizierung der aktiven Inhaltsstoffe des wässrigen Nelkenextraktes
78
Abbildung 4.13 Die Strukturformeln des Eugenols, Acetyleugenols, Metyleugenols, Thymols, Carvacrols und
Propofols
Bisher liegen keine Studien vor, die die GABAA-Rezeptor potenzierende Wirkung von
Acetyleugenol untersuchten. Ding et al. identifizierten kürzlich Methyleugenol (Abbildung 4.13),
welches ein orthoalkyliertes Derivat von Eugenol ist, als neuartigen Agonist des GABAA-
Rezeptors. Sie berichten, dass Methyleugenol in den Konzentrationen zwischen 1 und 30 µM
(0,17 - 5 µg/mL) die 0,3 µM GABA-vermittelten Ströme bei hippocampalen Neuronen von
Mäusen verstärkten sowie die in HEK 293 Zellen transfizierten α1β2γ2 oder α5β2γ2 GABAA-
Rezeptoren aus Ratten mit einem EC50-Wert von 290 µM (entspricht 52 µg/mL, für den α1β2γ2-
GABAA-Rezeptor) bzw. 198 µM (entspricht 35 µg/mL, für den α5β2γ2- GABAA-Rezeptor) direkt
aktivierten [88]. In dieser vorliegender Arbeit zeigten Eugenol und sein Derivat Acetyleugenol in
sehr geringen Konzentrationen (0,1 - 1 µg/mL) eine signifikante Potenzierung der α1β2- GABAA-
Rezeptor Aktivität. Diese Ergebnisse deuten darauf hin, dass sich diese GABAerge modulierende

Identifizierung der aktiven Inhaltsstoffe des wässrigen Nelkenextraktes
79
Wirkung von Eugenol und seiner Derivaten (Acetyl- und Methyleugenol) auf ihrer strukturellen
Ähnlichkeit beruht. Bei den drei Verbindungen handelt es sich um 4-Allylbenzene [207], die zur
Stoffgruppe der Phenylpropanoide gezählt werden [88, 132, 208]. Die phenolische Struktur ist
offensichtlich ein wichtiger Faktor für die Interaktion mit dem GABAergen System. Zum Beispiel
bewirkte auch Carvacrol eine positive allosterische Modulation der nativen GABAA-Rezeptoren
in Primärkulturen von kortikalen Neuronen, die aus Mäusen gewonnen wurden [89]. Kessler et
al. untersuchten die GABAerge aktiven Inhaltsstoffe des Sideritis-Tees (S. stricta) und stellten
fest, dass Carvacrol ein Modulator der menschlichen, in Xenopus leavis Oocyten transfizierten
α1β2γ2 GABAA-Rezeptoren ist. Dabei wurde diese allosterische modulierende Wirkung von
Carvacrol ab einer Konzentration von 3 µM (entspricht 0,5 µg/mL) beobachtet [87]. Da Eugenol
hier bereits in einerer Konzentration von 0,1 µg/mL die Aktivität des GABAA-Rezeptors
signifikant potenzierte, ist davon auszugehen, dass Eugenol im Vergleich zu Carvacrol ein noch
stärkerer allosterische GABAA-Rezeptor-Modulator sein kann. Diese Hypothese unterstützen
auch die Ergebnisse, die zeigten dass der Nelkenextrakt, dessen Hauptbestandteil Eugenol ist,
eine stärkere Potenzierung der GABA-vermittelten Ströme auslöste als die Extrakte von Sideritis-
Arten (S. stricta, S. arguta, S. condensata, S. sipylea), die vermutlich als Inhaltsstoff das Carvacrol
enthalten (siehe Kapitel 3.2).
Thymol führte auch zu einer positive allosterische Modulation der nativen GABAA-Rezeptoren in
Primärkulturen von kortikalen Neuronen der Mäusen wie sein Isomer Carvacrol [94]. Zudem
zeigte Thymol eine Steigerung der Aktivität von menschlichen α1β1γ2s-, α1β3γ2s- und α6β3γ2s
GABAA-Rezeptoren, die in Xenopus leavis Oocyten transfiziert wurden [86]. Zusätzlich zu seiner
positive allosterische modulierende Wirkung löste Thymol eine direkte Aktivierung der GABAA-
Rezeptoren aus [81, 86, 94]. Ebenfalls aktivierte das Anästhetikum Propofol, eine orthoalkylierte
Phenolverbindung mit einer ähnlichen Struktur wie das Thymol (Abbildung 4.13), direkt den
GABAA-Rezeptor, wobei dieser agonistische Effekt zur seiner sedativ- hypnotischen Aktivität
beitragen kann [81, 86]. Trotz struktureller Ähnlichkeiten konnten Eugenol und Carvacrol keinen
über GABAA-Rezeptor vermittelten Chloridionenstrom in Abwesenheit von GABA hervorrufen
[89]. In der vorliegenden Arbeit wurde auch festgestellt, dass der wässrige Nelkenextrakt, der
als Hauptkomponente Eugenol enthält, keinen Chloridionenstrom über direkte Aktivierung des
GABAA-Rezeptors auslöste. Allerdings konnte der orthoalkylierte Eugenol-Derivat
(Methyleugenol) den GABAA-Rezeptor direkt aktivieren [88]. Dies steht im Einklang damit, dass
die orthoalkylierte Phenolverbindung zu einer direkte Aktivierung des GABAA-Rezeptors führen
[81]. Da Eugenol bzw. Carvacrol keine aliphatische Gruppe in ortho-Position zur Hydroxygruppe
des Benzolringes enthalten, zeigen sie keine agonistische Wirkung gegenüber den GABAA-
Rezeptor.

Identifizierung der aktiven Inhaltsstoffe des wässrigen Nelkenextraktes
80
Auf Grund seiner anästhetischen und analgetischen Wirkung ist der Einsatz von Eugenol in der
Zahnmedizin weit verbreitet. Der lokalanästhetische Effekt des Eugenol auf Menschen konnte
bereits nachgewiesen werden, so dass Eugenol in hausgemachten Nelkengel eine signifikante
Abschwächung der Schmerzen auslöste [128]. Zudem zeigten einige Tierversuche die
anästhetische Wirkung des Eugenols an männlichen Sprague-Dawley Ratten [140] und an
Xenopus leavis Fröschen [141]. Des Weiteren wurde eine analgetische und antikonvulsive
Wirkung von Eugenol an Mäusen und Ratten beobachtet [142]. Darüber hinaus zeigten
Untersuchungen an neokortikalen und hippokampalen Gewebe der Ratten, dass Eugenol
epileptiforme Feldpotentiale sowie die kortikale Streudepolarisierung (cortical spreading
depression) unterdrückt, die der Migräneaura und den nachfolgenden Kopfschmerzen zugrunde
liegt [209]. Es wurde auch beobachtet, dass Eugenol eine antidepressivaähnliche Wirkung auf
Mäuse hat [210]. Da die Steigerung der GABAergen Inhibition zur Behandlung von
neuropathischen und chronischen Schmerzen, von Depression sowie von Epilepsie verwendet
wird [21, 30, 54, 69, 73, 74, 211], kann Eugenol seine analgetische, antiepileptische und
antidepressive Wirkung teilweise über Potenzierung der GABAA-Rezeptor Aktivität vermitteln.
Zusammenfassend lässt sich festhalten, dass Eugenol und sein Derivat Acetyleugenol positive
Modulatoren des GABAA-Rezeptors sind und Eugenol als Hauptkomponente im wässrigen
Nelkenextrakt für den positive allosterische modulierenden Effekt von Nelkenextrakt
verantwortlich ist.

Material und Methoden
81
Material und Methoden 5
5.1. Geräte
Analytische GC Agillent 6890
Autosampler MPS2 Gerstel Multi Purpose
Massenspektrometer, hp Hewlett Packard 5973
Analytische UHPLC Dionex Ultimate 3000 RS
Autosampler, WPS-3000TSL
Degasser, Ultimate 3000
Pumpe, HPG-3400
Säulen Kompartiment, TCC- 3000
Diodenarraydetektor (DAD) -3000
Software, Dionex Chromeleon 6.80
Analysenwaage Mettler Toledo
Sartorius
Autoklav Systec V-65
Elektrophorese Power Pac basic BioRad
Fraktionensammler Advantec CHF 122 SB
GC-Kapillarsäule DB-FFAP, J&W Scientific, Phenomenex, 0,32 mm x 30 m,
Filmdicke 0,25 µM
Geldokumentationssystem Bio Doc Analyze, Biometra
HPLC-Vorsäule Macherey-Nagel Nucleodur C18ec, 10x8 mm,
Partikelgröße 5 µM, Porengröße 110 Å
HPLC-Säulen Macherey-Nagel Nucleodur C18ec, 10x250 mm,
Partikelgröße 5 µM, Porengröße 110 Å
Agillent,Zorbax Eclipse XDB-C8, 4,6 x 150 mm,
Partikelgröße 5 µM, Porengröße 80 Å
Kolbenhubpipetten, Abimed und Discovery von HTL Lab Solutions,
diverse Größen Brand, Eppendorf PhysioCare, Biozym
Lyophylle Savant Novalyphe NL150
Varian DS 602 Pumpe
Magnetrührer IKAMAG RET/RCT

Material und Methoden
82
Mikropipettenziergerät P97-Micropipette Puller Sutter Instrument
Mikroplattenphotometer µQuant BioTek
Software, Gen 5, Version 1.04.5
Mikroplatte Schüttler VWR
Mikro-Zentrifuge neoLab
Nano Injektor Nanoject II Drummond Scientific
Nano-Spektrophotometer NanoDrop 2000/2000c, Thermo
pH Meter Metrohm PM 400
Metrohm PM 654
HI9321 Microprocessor HANNA
Pipettierhilfe Pipetus, Hirschmann
Rotationsverdampfer Heidolph
CVC 2, vacuubrand Vacuum Controller
Semi-präparative HPLC (I) Jasco
Autosampler, Jasco AS-1555 Plus
3-line Degasser, Jasco DG-1580-53
Pumpe, Jasco PU-1580
Fluoreszenzdetektor, Jasco FP-920
Multiwavelength-Detektor, Jasco MD-1510
Software, Borwin
Semi-präparative HPLC (II) Jasco
Autosampler, Jasco AS-2057 Plus
3-line Degasser, Jasco DG-980-50
Binäre Pumpe, Jasco PU-2087 Plus
Mischkammer 3000 µL
4 λ UV/Vis-Detektor, Jasco UV-2077 Plus
Software, Chrompass
Thermomixer Eppendorf comfort
Trockenschrank Binder BD-53
UHPLC-Säule YMC-Ultra HT Pro C18, 2x50 mm,
Partikelgröße 2 µM, Porengröße 120 Å
Ultraschallbad Bandelin Sonorex Super RK 255 H
Ultratiefkühlgerät New Brunswick Scientific, Modell U 101
Zentrifugen Rotina 420R, HETTICH
Sorvall RC6, Thermo
Biofuge Primo, Heraeus
Mikro 200R, HETTICH

Material und Methoden
83
Voltage-Clamp Verstärker, npi TURBO TEC 03X
Mikroskop, Reichert-Jung
Kaltlichtlampe, Carl Zeiss KL1500
Pumpe, KNF Laboport
Software, cell works lite Version 5.5.1
Vortex-Schüttler VWR
Wasseraufbereitung Millipore, Synergy 185
Wasserbad GFL 1002
5.2. Verbrauchsmaterialien
Einmalhandschuhe Kimberly-Clark
Einmalspritzen Braun
Faltenfilter MN 615 ¼, Macherey-Nagel
Injektionskapillare 3,5``, Drummond Scientific
Mikroplatten Greiner bio-one 96 Well UV Star Microplate
Pipettenspitzen Brand, Eppendorf
Reaktionsgefäße Eppendorf
Schraubdeckel für Vials VWR
Septen VWR
Spritzenfilter Rotilabor®, PVDF steril, 0,22 µM und 0,45 µM; Roth
Vials VWR
Voltage-Clamp Glaskapillare GB 150F-10, Science Products
Voltage-Clamp Schlauch Silikon, Innendurchmesser 1mm, Außendurchmesser 3mm, VWR
PTFE, Innendurchmesser 1mm, Außendurchmesser 1,6 mm, VWR

Material und Methoden
84
5.3. Chemikalien
2-cis-4-trans-Abscisinsäure, 99 % Alfa-Aesar
Acetonitril (HPLC-Grade) Fisher-Scientific
Acetonitril (LC-MS Grade) für UHPLC Fluka
Acetyleugenol, Rotichrom, GC CarlRoth
Agarose, Low electroendosmosis Biozym
Alpha-Ketoglutarat Natriumsalz ≥ 98 % Sigma
Ameisensäure Fluka
Ampicillin Natriumsalz Carl Roth
Apigenin, approx 95% Sigma
α-Bisabolol ≥ 95 % Sigma-Aldrich
Borax Riedel-de-Haen
Borneol ≥ 95 % Fluka
Borsäue p.A. Carl Roth
Bromphenolblau Natriumsalz Carl Roth
Calciumchlorid Dihydrat p.A. Carl Roth
(-)-Caryophyllene oxide, 95% Sigma
β-Caryophyllen ≥ 80 % Sigma
(-)-Caryophyllenoxid ≥ 95 % Sigma
L-Carveol, mixture of cis and trans ≥ 95 % Sigma
R- Carvon 98 % Alfa-Aesar
S- Carvon 96 % Alfa-Aesar
Cedrol ≥ 99 % Sigma
Citral 95 % Sigma-Aldrich
β –Citronellol ≥ 95 % Sigma
Chloroform/Isoamylalkohol, Carl-Roth
zur Extraktion von Nukleinsäuren
p-Coumarinsäure ≥ 98 % Sigma-Aldrich
β-Damascenon, sum of isomers ≥ 95 % Aldrich
Dichlormethan Acros
Dimethylsulfoxid (DMSO), wasserfrei, ≥ 99,9 % Acros
EDTA Dinatriumsalz Dihydrat ≥ 99 % Carl Roth
Essigsäue Merck
Ethanol Merck
Ethidiumbromid Roth

Material und Methoden
85
4-Ethylguajacol ≥ 98 % Sigma
Eucalyptol ≥ 99 % Sigma
Eugenol, 99% AlfaAesar
β –Farnesen Fluka
1-R-Fenchon ≥ 98 % Sigma-Aldrich
Ferulasäure 99 % Sigma-Aldrich
GABA ≥ 99 % Sigma
GABase from Pseudomonas fluorescens, ≥ 30 % Sigma
Gentamicin Sigma
Glycerin p.A. ≥ 86 % Carl Roth
Glycin ≥ 99 % Sigma-Aldrich
Hefeextrakt für die Bakteriologie Carl-Roth
HEPES Pufferan Carl Roth
4-Hdroxybenzaldehyd,analytical standard Sigma-Aldrich
2-Hydroxy-1-naphthaldehyde, 98% Alfa-Aesar
β-Ionon ≥ 96 % Sigma
α-Ionon ≥ 90 % Fluka
Isoeugenol, cis + trans, 98% + AlfaAesar
Isopulegol ≥ 98 % Sigma
R -(+)-Limonen (Dipenten) mixture of isomers Sigma
Kaliumchlorid p.A. Carl Roth
Kaliumpyrophospat ≥ 97 % Aldrich
Magnesiumchlorid Hexahydrat p.A. Carl Roth
Menthol ≥ 99 % Sigma
Methanol für HPLC-Analytik Fisher-Scientific
2-Methyl-3-buten-2-ol Sigma
3-Methyl-2-buten-1-ol Sigma
Myrcen ≥ 90 % Sigma
Myrtenol ≥ 95 % Sigma
NADP Natriumsalz AppliChem
Natriumacetat, wasserfrei Riedel-de-Haen
Natriumchlorid p.A. Carl Roth
Natriumhydroxid ≥ 98 % Sigma-Aldrich
Natriumsulfat, wasserfreie VWR
Nerolidol, mixture of cis and trans ≥ 98 % Sigma
Nootkaton ≥ 99 % Sigma
Nuclease-freies Wasser Ambion

Material und Methoden
86
Ocimen, mixture of isomers ≥ 90 % Sigma
Orthophtalaldehyd (OPA) Sigma
Perillylalkohol ≥ 90 % Sigma
Phenol/Chloroform/Isoamylalkohol Carl-Roth
zur Extraktion von Nukleinsäuren
Pinocarveol, trans ≥ 96 % Sigma
Rosenoxid Sigma
Sabinen ≥ 98,5 % Carl Roth
α-Santonin ≥ 99 % Sigma
Terpinolen ≥ 90 % Sigma
TE-Puffer 100X AppliChem
Theaspiran ≥ 90 % Sigma
α-Thujon ≥ 96 % Sigma
Tris BASE Pufferan Carl Roth
Trypton (Pepton aus casein) AppliChem
Vigabatrin British Pharmacopeie Commision
Laborotry
Xylencyanol Carl Roth

Material und Methoden
87
5.4. Herkunft der Lebensmitteln und Herstellung der
Lebensmittelextrakte
5.4.1. Herkunft der Lebensmitteln
Die getrockneten Bergtee-Proben S. argura, S. condensata, S. strictica und S. sipylea wurden
von AG Büttner zur Verfügung gestellt. Kakaopulver, Gewürznelken und Schwarzer Pfeffer
(gemahlen) wurden auf dem lokalen Markt in Erlangen gekauft. Die zerkleinerten Pflanzenteile
von Melisse, Kamille, Hopfen, Salbei, grünem Tee und Lavendelblüten wurden in der örtlichen
Apotheke erworben. In der unten stehenden Tabelle sind die Hersteller der verwendeten
Lebensmittel aufgelistet.
Tabelle 5.1 Übersicht über die untersuchten Lebensmittel
Lebensmittel Hersteller
Grüner Tee Sidroga
Hopfenzapfen Heinrich Klenk
Kakao Pulver Dr.Oetker
Kamillenblüten Müller
Lavendelblüten Aurica
Melisse Sidroga
Nelken Ganz HES
Salbeiblätter Heinrich Klenk
Schwarzer Pfeffer (gemahlen) Ostmann
5.4.2. Herstellung der Lebensmittelextrakte
Vor der Extraktion wurden Bergtee und Gewürznelken grob zerkleinert. Die anderen Pflanzen
wurden direkt für die Extraktion verwendet.
5.4.2.1. Extraktionsmethode I
Zur Extraktion der Lebensmittel wurden 2,5 g Lebensmittel eingewogen und mit 100 ml
kochendem Reinstwasser versetzt. Der Ansatz wurde für 1h auf dem Magnetrührer bei 90 °C
gerührt. Der Extrakt wurde durch einen Faltenfilter abfiltriert, anschließend das Filtrat auf
Raumtemperatur gekühlt und aliquotiert. Die erhaltenen Aliquots wurden tiefgefroren und dann
lyophilisiert. Durch Wägung wurden die Trockenmassen der gefriergetrockneten Extrakte
bestimmt. Zuletzt wurden die lyophilisierten Extrakte mit Reinstwasser gelöst und auf eine

Material und Methoden
88
Konzentration von 10 mg/mL für die Versuche verdünnt. Die Lagerung der Extrakte erfolgte bei
-20°C.
5.4.2.2. Extraktionsmethode II
Da die mittels Extraktionsmethode I hergestellten Extrakte einen unspezifischen Strom in der
Membran von Oocyten, in die keine RNA injiziert wurde, erzeugten, wurde die angewendete
Extraktionsmethode modifiziert. Dazu wurden 2,5 g des Lebensmittels mit 100 ml kochendem
heißem Reinstwasser versetzt, für 15 Minuten auf dem Magnetrührer gerührt und anschließend
abfiltriert. Das Filtrat wurde aliquotiert, tiefgefroren und lyophilisiert. Nach der Bestimmung der
Trockenmasse wurden die gefriergetrockneten Extrakte mit Reinstwasser gelöst und auf die
entsprechende Konzentrationen verdünnt um eine Extrakt-Stammlösung für Versuche
herzustellen (10 mg/mL für die elektro-physiologischen Untersuchungen; 40 mg/mL für die
HPLC-Versuche zur GABA-Bestimmung; 20 mg/mL für die HPLC-Fraktionierung). Alle
Konzentrationswerte der Extrakte für die Versuche sind auf die Trockenmasse bezogen. Die
hergestellten Extrakte wurden bei -20°C gelagert und für sämtliche weitere Analysen
verwendet.
5.5. Messung der Enzymaktivität mittels Enzymassay
5.5.1. Lösungen und Puffer
Für das Enzymassay wurden folgende Reagenzien verwendet.
228,5 mM α- Ketoglutarat
873,0 mM GABA
26,1 mM NADP+
25 mM Vigabatrin
1,5-1,7 units/mg GABase
Alle Lösungen wurden in Reinstwasser angesetzt.

Material und Methoden
89
Kaliumpyrophosphatpuffer (150 mM, pH= 8,0)
1,24 g Kaliumpyrophosphat wurde in einen 25 ml Messkolben eingewogen und in Reinstwasser
gelöst. Anschließend wurde der pH-Wert mit einer 1 M Salzsäure auf 8,0 eingestellt und mit
Reinstwasser bis zur Marke aufgefüllt. Die Lösung wurde bei 4°C gelagert.
5.5.2. Bestimmung der Enzymaktivität
Um die Enzymaktivität bzw. inhibitorische Wirkung von Substanzen auf GABA abbauende
Enzyme zu vermessen, wurde eine nach Jacoby et al. [42] modifizierte Methode durchgeführt.
Dazu wurden 220 µL der Inkubationslösung, welches aus 214 µL des Kaliumpyrophosphatpuffer
(150 mM, pH= 8,0) und 6 µL der 228,5 mM α- Ketoglutarat-Lösung besteht, 14 µL GABA-Lösung
(873,0 mM) und unterschiedliche Konzentrationen von Substanzen oder von Extrakten in die
Vertiefungen einer Mikrotiterplatte gegeben. Eine Dreifachbestimmung wurde pro eingesetzte
Konzentration der zu testenden Substanzen bzw. Extrakte durchgeführt. Als Negativkontrolle
wurde Reinstwasser, Ethanol oder DMSO, welche als Lösungsmittel für Substanzen bzw.
Extrakten verwendet wurden, statt Inhibitor eingesetzt. Die positive Kontrolle für die
Bestimmung hemmender Wirkung von Substanzen wurde mit Vigabatrin durchgeführt.
Anschließend wurde 4 µL GABase-Lösung (1,5-1,7 units/mg) in jede Vertiefung hinzu pipettiert.
Nach der Zugabe der Enzymlösung wurde die Mikrotiterplatte 2 Minuten bei Raumtemperatur
geschüttelt (500 U/min) und für 30 Minuten bei 37°C vorinkubiert, bevor sie mit NADP+
inkubiert wurde. Nach der Vorinkubation wurde 10 µL NADP+ -Lösung (26,1 mM) in jede
Vertiefung pipettiert und Platte wurde für 30 Sekunden geschüttelt (500 U/min). Danach
erfolgte die erste photometrische Messung bei 340 nm. Dieser erste Messwert wurde als
Startwert verwendet. Nach der ersten Messung wurde die Mikrotiterplatte für 30 Minuten bei
37 °C inkubiert und dann erneut gemessen. Der Differenzwert zwischen Startwert und nach 30-
minütiger Inkubation ermitteltem Messwert wurde zur Berechnung der Enzymaktivität
verwendet. Zuletzt wurde die Mikrotiterplatte erneut für 30 Minuten bei 37°C inkubiert und
anschließend bei 340 nm vermessen. Die letzte photometrische Messung, welche nach
insgesamt 60-minütiger Inkubation erfolgte, wurde durchgeführt um zu prüfen, ob die Reaktion
weiterlaufen kann.
5.5.3. Auswertung der Enzymaktivität
Um die Enzymaktivität zu berechnen, wurde der Absorption-Startwert von jedem Messwert der
Absorption, welcher nach 30-minütiger Inkubation ermittelt wurde, abgezogen. Der
Absorptionswert der Negativkontrolle, d.h. Wasser-, Ethanol- oder DMSO-Kontrolle, wurde auf

Material und Methoden
90
eine Aktivität von 100 % festgelegt. Die Enzymaktivität jedes Messwertes wurde mit Hilfe
folgender Formel bestimmt:
Aktivität (%)=Messwert-Startwert
Negativkontrolle-Startwert ×100
Um die inhibitorischen Wirkungen von Substanzen bzw. Lebensmittelextrakten miteinander
vergleichen zu können, wurden ihre IC50-Werte mittels Origin® (Origin® 2015G von OriginLab
Corporation, Northampton, MA 01060 USA) berechnet. Dazu wurde eine nicht-lineare
Kurvenanpassung mit Hilfe einer Dose Response-Funktion in der Kategorie Growth/Sigmaidol
für Vigabatrin durchgeführt.
Für Lebensmittelextrakte und Substanzen wurde eine lineare Kurvenanpassung in Origin erstellt,
womit ihre IC50-Werte bestimmt werden konnten. Die IC50-Werte wurden mit einem
Konfidenzintervall (CI) von 95% ermittelt.
5.6. TEVC
5.6.1. Lösungen, Puffer, Nährmedien und kommerziell erhältliche Kits
Alle Lösungen wurden, soweit nicht anders angegeben in Reinstwasser angesetzt.
GABA Stammlösung (0,1 M)
1,03 g GABA wurde in 100 mL ND 96 aufgelöst und anschließend aliquotiert (à 1,0 mL) und bei
-20 °C gelagert. Die Herstellung der GABA-Standardlösungen erfolgte durch Verdünnung der
GABA-Stammlösung mit ND 96 Pufferlösung.
Gentamicin (50 mg/mL)
0,5 g Gentamicin, ein Antibiotikum, wurde in einen 10 mL Messkolben eingewogen und bis zur
Marke mit Reinstwasser aufgefüllt. Diese Lösung wurde bei 4 °C gelagert und vor der
Verwendung mit ND 96 Pufferlösung 1:1000 verdünnt.

Material und Methoden
91
Glycin Stammlösung (0,1 M)
188 mg Glycin wurde in 25 mL ND 96 gelöst, aliquotiert (á 1,0 mL) und bei -20 °C gelagert. Für
die Herstellung der Glycin-Standardlösungen wurde diese Stammlösung mit ND 96 Pufferlösung
verdünnt.
ND 96 Puffer (10-fach konzertriert)
Für die Herstellung der 10-fachen konzentrierten ND 96 Pufferlösung wurden 56,1 g NaCl (960
mM), 1,49 g KCl (20 mM), 2,03 g MgCl2.6H2O (10 mM), 1,74 g CaCl2.2H2O (10 mM) und 11,92 g
HEPES (50 mM) in ca 950 mL Reinstwasser gelöst. Der pH der Lösung wurde mit Natrounlage auf
7,4 eingestellt. Anschließend wurde die Lösung auf 1 L mit Reinstwasser aufgefüllt und
membranfiltriert. Diese konzentrierte Lösung wurde als Stocklösung bei 4 °C gelagert. Vor der
Verwendung wurde diese Stocklösung mit Reinstwasser 1:10 verdünnt.
TBE (Tris-Borat-EDTA)Puffer (10-fach konzertriert)
890 mM Tris
890 mM Borsäure
20 mM EDTA
DNA-Elektrophoreseladepuffer (10-fach konzertriert)
0,05% (w/v) Bromphenolblau
0,05% (w/v) Xylencyanol
0,30% (v/v)Glycerin
200 mM EDTA
2xYT Medium (1L)
15 g Trypton (Pepton aus casein)
10 g Hefeextrakt
5 g NaCl
Kommerziell erhältliche Kits
Jetstar Plasmid Kits JETSTAR 2.0, Genomed
mMessage mMachine T7 Polymerase Kit Ambion, Life Technologies
Restriktionsenzyme New England Biolabs (NEB)

Material und Methoden
92
5.6.2. Das Xenopus-Oocyten-Expressionssystem
Oocyten aus Xenopus laevis wurden als Expressionssystem für die TEVC-Technik verwendet.
Dank ihres großen Durchmessers von 1,1-1,3 mm ermöglichen Oocyten eine einfache
Handhabung, Injektion und weitere elektrophysiologische Messung. Zudem kann die injizierte
RNA effizient translatiert werden und die Aktivität der synthetisierten Ionenkanäle kann
bestimmt werden [98].
Die Expression des GABAA-und Glycin- Rezeptoren erfolgte durch Injektion der entsprechender
cRNA in Oocyten aus Xenopus laevis.
5.6.2.1. Herstellung der RNA der α1- und β2- Untereinheiten des humanen GABAA-
Rezeptors
Die cDNA der α1- und β2- Untereinheiten der humanen GABAA-Rezeptoren wurde in den von
der Firma Lundbeck (Valby) zur Verfügung gestellten pcDNA3.1-Vektor subkloniert. Zunächst
wurde der Vektor mit Phosphatase zum Einbringen bestimmter DNA-Sequenz behandelt
(Dephosphorylierung). Die Verknüpfung geschnittener DNA-Fragmente erfolgte mittels Quick
Ligation. Der Ligationsansatz wurde in E. coli-Zellen (NEB5α) durch Hitzeschock zur
Vervielfältigung der Plasmid-DNA eingebracht (Transformation). Diese Plasmid-DNA wurde von
AG Büttner zur Verfügung gestellt.
Für die Isolierung der größeren Plasmid-DNA Konzentrationen wurde ein Maxi-Prep Kit
verwendet. Anschließend wurde die Plasmid-DNA mit dem Restriktionsenzym linearisiert. Die
Synthese der cRNA erfolgte durch in vitro Transkription der linearisierten DNA mit Hilfe des
mMessage mMachine T7 Polymerase Kits.
5.6.2.1.1. Isolation der Plasmid DNA-Maxipräparation
Die Maxipräparation erfolgte gemäß dem Jetstar Plasmid Protokoll und unter Einsatz des Jetstar
Plasmid Kits und der mitgelieferten Puffer. 10 µL Bakterienkultur (α1- oder β2- Untereinheit des
humanen GABAA-Rezeptors) wurden in 200 mL mit 40 µL des Antibiotikums Ampicillin
(Endkonzentration 100 µg/mL) versetzten 2 YT-Mediums angeimpft und über Nacht bei 37 °C
und 200 rpm im Schüttler inkubiert. Nach Inkubation wurden die Bakterien für 20 Minuten bei
400 rpm abzentrifugiert und anschließend wurde das Pellet in 10 mL des mit RNAse versetzten
Resuspensierungspuffers resuspendiert. 10 mL Lysispuffer wurde hinzugefügt und für 3 Minuten
bei Raumtemperatur inkubiert. Anschließend wurden 10 mL Neutralisierungpuffer
(Präzipitationspuffer) hinzugegeben und für 15 Minuten bei 20°C und 15000 g zentrifugiert.

Material und Methoden
93
Dann wurde der Überstand auf eine Säule gegeben. Nach Spülen der Säule mit 60 mL
Waschpuffer wurden 15 mL Elutiospuffer auf die Säule gegeben. Anschließend wurden 3 mL
Isopropanol für die Präzipitation der DNA hinzugefügt und bei 4 °C, 12000 g für 1 Stunde
abzentrifugiert. Der Überstand wurde abgegossen. Das Pellet wurde mit 5 mL Ethanol (70%)
durch Zentrifugation (bei 4 °C und 12000 g für 15) gewaschen. Dann wurde das entstandene
DNA-Pellet ca. 30 Minuten bei Raumtemperatur getrocknet. Anschließend wurde das
getrocknete Pellet in 200-400 µL TE-Puffer gelöst. Die Lagerung der Plasmid-DNA erfolgte bei
-20°C.
5.6.2.1.2. Linearisierung der DNA
Für die Linearisierung der Plasmid-DNA wurde ein Restriktionsverdau entsprechend der
Herstellerangaben (NEB) mittels des im Kit enthaltenen Bgl II Restriktionsenzyms, BSA
(Rinderserumalbumin, aus dem Englischen: bovine serum albumin) und dem entsprechenden
Puffer durchgeführt. Dazu wurden 50 µL des folgenden Verdauansatzes verwendet:
Plasmid-DNA 20-30 µL (variabel nach DNA-Konzentration)
BSA (10x) 5 µL
P3 Puffer 5 µL
Bgl II Restriktionsenzym 5 µL
Nuklease-freies Wasser 5-15 µL (variabel nach eingesetzte DNA-Volume)
Nach dem Pipettieren wurde der Verdauansatz bei 37 °C für 6 Stunden inkubiert. Anschließend
erfolgte die Aufreinigung der linearisierten DNA durch eine Phenol/Chloroform Extraktions-
Methode. Nach Zugabe von 50 µL einer Phenol-Chloroform-Isoamylalkohol (25:24:1) Mischung
wurde der Ansatz bei 14000 rpm für 5 Minuten bei Raumtemperatur abzentrifugiert. Nach der
Phasentrennung wurde die wässrige Oberphase in neues Eppendorfgefäß überführt und 50 µL
Chloroform/Isoamylalkohol (24:1) dazu pipettiert. Nach einer fünfminütiger Zentrifugation bei
14000 rpm wurde die oberste Phase in ein neues Eppendorfgefäß überführt und mit 0,25 µL
Ethylendiamintetraessigsäure(EDTA) (0,5 M), 110µL Ethanol (98,8%) und 5µL Natriumacetat (0,3
M) versetzt. Anschließend wurde bei -20°C über Nacht inkubiert. Dann wurde die linearisierte
DNA bei 4°C und 14000 rpm für 30 Minuten gefällt. Das DNA-Pellet wurde mit 1 mL Ethanol
(70%) durch eine zwanzigminütige Zentrifugation mit 14000 rpm bei 4°C gewaschen. Nach dem
Trocknen wurde die linearisierte DNA in 10 µL Nuklease-freiem Wasser gelöst. Nach
photometrischer Konzentrationsbestimmung wurde die vollständige Linearisierung der DNA
mittels Agarose-Gelelektrophorese überprüft. Diese linearisierte DNA wurde bis zur
Verwendung bei -20 °C gelagert.

Material und Methoden
94
5.6.2.1.3. Konzentrationsbestimmung der DNA/RNA
Die Konzentration der isolierten DNA/RNA wurde mittels Nano-Spektophotometer bei 260 nm
bestimmt.
5.6.2.1.4. Agarose-Gelelektrophorese
Die Auftrennung der DNA-Fragmente erfolgte durch 1%iges Agarose-Gelelektrophorese. Dazu
wurde 0,6 g Agarose auf 60 g mit dem 1xTBE (Tris-Borat-EDTA) Puffer aufgefüllt und
anschließend in der Mikrowelle erhitzt, um die Agarose komplett auszulösen. Nach Zugabe von
6 µL Ethidiumbromid wurde das Gel in die Gelkammer gegossen und gewartet, bis es kühl und
fest war. Anschließend wurde das abgekühlte Gel in die Laufkammer, die mit 1xTBE Puffer
gefüllt wurde, gelegt. Nach der Färbung mit Ladepuffer wurden die DNA-Proben und der DNA-
Längenstandard in die Geltaschen aufgetragen. Bei der Spannung von 120 mV wurden die DNA-
Fragmente elektrophoretisch aufgetrennt und anschließend unter UV-Licht fotodokumentiert.
5.6.2.1.5. In vitro Transkription
Zur Synthese der cRNA wurde eine in vitro Transkription mittels mMessage mMachine T7
Polymerase Kit durchgeführt. Die Transkription erfolgte gemäß den Angaben des Herstellers
und unter Verwendung der mitgelieferten Puffer. Dazu wurde der folgende Ansatz pipettiert.
NTP/CAP (2x) 5 µL
Reaktionspuffer (10x) 1µL
Linearisierte DNA 2 µL
Enzym Mix 1 µL
Nuklease-freies Wasser 1µL
Der Ansatz wurde bei 37°C für 2 Stunden inkubiert. Anschließend wurden dem Ansatz 0,5 µL
DNase hinzugefügt, um die verbliebene DNA zu entfernen. Nach einer 20 minütiger Inkubation
bei 37°C erfolgte die Aufreinigung der RNA. Dazu wurden 15 µL Lithiumchlorid und 15 µL
Nuklease-freies Wasser hinzugefügt und bei -20°C über Nacht inkubiert. Anschließend wurde
diese RNA-Lösung bei 4°C und 14000 rpm für 30 Minuten zentrifugiert. Das RNA-Pellet wurde
mit 1 mL Ethanol (70%) durch eine zwanzigminütige Zentrifugation mit 14000 rpm bei 4°C
gewaschen. Der Überstand wurde verworfen und das entstandene RNA-Pellet wurde bei
Raumtemperatur getrocknet. Anschließend wurde die RNA in 5-10 µL Nuklease-freiem Wasser
gelöst. Nach photometrischer Konzentrationsbestimmung wurde die RNA der α1- Untereinheit
des humanen GABAA-Rezeptors mit derselben Menge an β2-Untereinheit des humanen GABAA-

Material und Methoden
95
Rezeptors gemischt und aliquotiert (à 1,5 µL). Die RNA wurde bis zur Mikroinjektion bei - 80 °C
gelagert.
5.6.2.2. Herstellung der RNA der α1- Untereinheit des humanen Glycin-Rezeptors
Die cDNA der α1 Untereinheit des menschlichen Glycin-Rezeptors wurde in den pNKS 2-Vektor
subkloniert. Die Plasmid-DNA wurde mit Hilfe einer Restriktionsendonuklease Hind II linearisiert.
Anschließend wurde die cRNA aus dieser linearisierten DNA mittels mMessage mMachine SP6
Polymerase Kit in vitro synthetisiert. Diese cRNA des humanen α1 Glycin-Rezeptors wurde von
Dr. Volker Eulenberg zur Verfügung gestellt.
5.6.2.3. Isolation der Xenopus-Oocyten und Mikroinjektion der cRNA in Oocyten
Die Isolation der Oocyten aus Xenopus laevis erfolgte durch chirurgische Oocytenentnahme und
anschließende Defollikulierung der Oocyten. Diese defollikulierten Oocyten wurden vom
Institut für zelluläre und molekulare Physiologie in Erlangen zur Verfügung gestellt. Zur Injektion
wurden die defollikulierten Oocyten der letzten Entwicklungsstadien unter dem Mikroskop
ausgewählt.
Um die untersuchten GABAA- oder Glycin-Rezeptoren zu exprimieren, wurden die
entsprechenden cRNA in selektierte Xenopus-Oocyten injiziert. Die Mikroinjektion erfolgte
durch einen Mikroinjektor (Nanoject II). Hierfür wurde die aus einer Glasskapillare mittels eines
Mikropipettenziehgeräts hergestellte Injektionskapillere mit Mineralöl luftblasenfrei gefüllt und
über die Nadel des Mikroinjektors befestigt. Anschließend wurde die Spitze der
Injektionskapillare unter dem Mikroskop auf einem Durchmesser von ca. 10-20 µm
abgebrochen. Die hergestellte cRNA-Lösung humaner α1β2 (1:1) GABAA-Rezeptoruntereinheiten
oder des humanen α1-Glycin-Rezeptors wurde mit Hilfe des Mikroinjektors in die
Injektionskapillare eingezogen und anschließend in die Oocyten intrazellulär injiziert. Die
Injektion fand in einer mit ND 96 befüllten Injektionskammer statt. Das Injektionsvolume betrug
41,4 nL; womit ca. 18-28 ng cRNA des α1β2 (1:1) GABAA-Rezeptors oder ca. 0,5 pg cRNA des α1-
Glycin-Rezeptoren in die Oocyte injiziert wurden. Nach der Injektion wurden die Oocyten in
einer mit Gentamicin zugesetzte ND 96 Lösung bei 18 °C inkubiert. Zwei Tagen nach der
Injektion wurden die Oocyten zur Messung verwendet.

Material und Methoden
96
5.6.3. TEVC-Technik
5.6.3.1. Versuchsdurchführung
Der Strom, der durch die in der Oocyte exprimierten Ionenkanäle vermittelt wurde, wurde in
Abhängigkeit des Membranpotentials mittels TEVC-Technik gemessen. Die
elektrophysiologische Messung erfolgte zwei Tagen nach der Injektion. Dazu wurde eine Oocyte
in der Vertiefung der Messkammer, wo zwei mit dem Verstärker verbundene Badelektrode
(Erdungs- und Referenzelektrode) positioniert wurden, platziert. Die aus einer Glasskapillare mit
Hilfe eines Mikropipettenziehgeräts ausgezogene Glassmikropipetten wurden mit 3 M KCl
luftblasenfrei gefüllt und hatten einen Widerstand von 0,2-3 MΩ. Anschließend wurde ein
chlorierter Silberdraht, über den ein elektrischer Kontakt zum Verstärker hergestellt wurde, in
diese gefüllte Glassmikropipette eingetaucht und als Strom- oder Spannungselektrode
verwendet. Zunächst wurden die mit dem Verstärker verbundenen Strom- und
Spannungselektroden in die Badlösung getaucht, um einen Nullabgleich der
Potentialdifferenzen an den Elektroden in Bezug auf die Badelektrode vorzunehmen.
Anschließend wurde die Strom- und Spannungselektrode unter dem Mikroskop in die Oocyte
eingestochen. Durch die intrazelluläre Spannungselektrode und die extrazelluläre
Referenzelektrode wurde das aktuelle Membranpotential gemessen. Im Ruhezustand
(Umspülung der Oocyten mit ND 96 Pufferlösung) entspricht das Membranpotential dem
vorgegebenen Haltepotential. Wurde ein Agonist des Rezeptors appliziert, wich das
Membranpotential vom Haltepotential aufgrund des Chloridionenfluss in das Zellinnere ab. Um
das Membranpotential mit dem Haltepotential auszugleichen, injizierte der Verstärker über die
intrazelluläre Stromelektrode den entsprechenden Ausgleichstrom in die Oocyte. Da dieser
Ausgleichstrom dem Chloridionenfluss entspricht, konnte sie als die vom Agonisten ausgelöste
Stromantwort quantifiziert werden. Über die Erdungselektrode konnte dieser Strom abfließen.
Die Stromspuren wurden bei 200 Hz mit Hilfe der Software (cell works lite Version 5.5.1)
digitalisiert, aufgenommen und analysiert. Alle Versuche wurden bei einem Haltepotential von
-50 mV durchgeführt. Die Applikation der Substanzen bzw. Lebensmittelextrakte erfolgte durch
in einem Perfusionssystem. Nach der Applikation wurden die Lösungen mit einer Pumpe
abgesaugt. Die Versuche wurden bei Raumtemperatur durchgeführt.

Material und Methoden
97
5.6.3.2. Versuchsprotokoll für die Applikation der Liganden und Normalisierung der
Ströme
Zur Bestimmung der Liganden-Aktivität wurden die Agonisten GABA oder Glycin in
verschiedenen Konzentrationen auf die Oocyten, die mit der entsprechenden cRNA des
humanen α1β2-GABAA-Rezeptors oder des humanen α1-Glycin-Rezeptors injiziert wurden,
appliziert. Dazu wurden GABA-Standardlösungen im untersuchten Konzentrationsbereich
zwischen 0,3 und 300 µM oder Glycin-Standardlösungen im untersuchten
Konzentrationsbereich zwischen 10 und 3000 µM appliziert. Vor der ersten Liganden-
Applikation wurden die Oocyten jeweils 90 Sekunden mit dem Puffer ND 96 umspült. Jede
GABA- oder Glycin-Konzentration wurde der Oocyte 20 Sekunden lang zugeführt. Nach jeder
Applikation wurden die Oocyten 180-320 Sekunden mit dem Puffer ND 96 gewaschen, um den
restlichen Liganden GABA oder Glycin aus der Oocytemembran zu entfernen. Dieses
Versuchsprotokoll wurde auch für die Negativkontrolle angewendet, bei der nicht injizierte
Oocyten getestet wurden.
Die Stromamplituden wurden in der Einheit „µA“ gemessen. Diese Stromamplituden wurden auf
die maximale Stromantwort, die durch GABA oder Glycin in der höchsten applizierten
Konzentration ausgelöst wurde, normiert, um die Dosis-Wirkungskurven der Liganden GABA und
Glycin zu ermitteln. Die Normalisierung der Stromamplituden erfolgte anhand folgender Formel:
INormiert(x) = IMesswert (x)
IMaximum
mit
INormiert(x) = Normierter Chloridionenstrom für x Konzentration des Liganden GABA oder Glycin
IMesswert(x) =Gemessene Stromantwort, die durch x Konzentration des Liganden GABA oder Glycin
ausgelöst wurde [µA]
IMaximum = Die maximale durch die höchste Konzentration des Liganden GABA oder Glycin
ausgelöste Stromantwort [µA]
x = Die applizierte Ligand-Konzentration; x für GABA: 0,3 µM, 1 µM, 3 µM, 10 µM, 30 µM, 100
µM oder 300 µM; x für Glycin: 10 µM, 30 µM, 50 µM, 100 µM, 300 µM, 1000 µM oder 3000
µM
Die Hill-Funktion wurde zur Erstellung der Dosis-Wirkungskurve für Agonisten GABA oder Glycin
verwendet. Dabei wurde der EC5, EC15 und EC50 Wert für den Agonisten GABA oder Glycin
mittels Origin® ermittelt.

Material und Methoden
98
5.6.3.3. Versuchsprotokoll für die Koapplikation der GABA mit den
Lebensmittelextrakten und Berechnung des relativen Stroms
Jeder Lebensmittelextrakt wurde in einem Konzentrationsbereich zwischen 0,5 und 5 µg/mL mit
1 µM GABA simultan appliziert, um seine konzentrationsabhängige potenzierende Wirkung auf
den humanen α1β2 GABAA-Rezeptor zu untersuchen, der in den Oocyte exprimiert wurde. Vor
und nach jeder Koapplikation der GABA mit dem Extrakt wurde eine Kontrollapplikation
durchgeführt. Dazu wurde der Oocyte 1 µM GABA zugeführt. Jede Applikation dauerte 20
Sekunden. Die Oocyten wurden vor der erster Applikation 90 Sekunden und nach jeder
Applikation 180-320 Sekunden mit ND 96 Puffer umspült. Die Negativkontrolle erfolgte durch
Applikation von GABA oder GABA in Verbindung mit Lebensmittelextrakt an Oocyten, die nicht
mit cRNA injiziert wurden.
Um die relativen Stromantworten zu bestimmen, die ein Maß für die Modulation der GABA-
induzierte Ströme durch Lebensmittelextrakte darstellen, wurde die mittlere Stromantwort der
1 µM GABA Kontrollapplikationen, die vor und nach jeder Koapplikation durchgeführt wurden,
auf 100% gesetzt. Dann wurde der gemessene Strom der Koapplikation auf diese ermittelten
prozentualen GABA-Kontrollströme normalisiert. Die relative Stromantwort durch die jeweiligen
Lebensmittelextrakte wurde folgendermaßen berechnet:
IRelativ(x) = IMesswert der Koapplikation x
(IGABA(vor der Koapplikation x) + IGABA(nach der Koapplikation x)) 2⁄× 100%
mit
IRelativ(x) = Relativer Chloridionenstrom für x Konzentration des Extraktes [%]
IMesswert der Koapplikation x = Der gemessene Chloridionenstrom, der durch Koapplikation der 1 µM
GABA mit x Konzentration des Extraktes ausgelöst wurde [µA]
IGABA(vor der Koapplikation x) =Stromantwort durch alleinige Applikation der 1 µM GABA, die vor der
Koapplikation der 1 µM GABA mit x Konzentration des Extraktes appliziert wurde [µA]
IGABA(nach der Koapplikation x) = Stromantwort durch alleinige Applikation der 1 µM GABA, die nach der
Koapplikation der 1 µM GABA mit x Konzentration des Extraktes appliziert wurde [µA]
x = Die mit 1 µM GABA applizierte Extrakt-Konzentration; 0,5 µg/mL, 0,75 µg/mL, 1 µg/mL, 2,5
µg/mL und 5 µg/mL

Material und Methoden
99
5.6.3.4. Versuchsprotokoll für die Messung der direkten Rezeptoraktivierung durch
Lebensmittelextrakten und Berechnung des relativen Stroms
Zur Erfassung der direkten Rezeptoraktivierung durch Lebensmittelextrakte wurde jeder Extrakt
in einer höchsten Konzentration von 7,5 µg/mL in Abwesenheit von GABA auf die mit dem
humanen α1β2-GABAA-Rezeptor transfizierten Oocyten appliziert. Nach dieser ersten
Applikation erfolgte eine Kontrollapplikation durch 1 µM GABA. Als letzte Applikation wurde
jeder Extrakt bei der Konzentration von 7,5 µg/mL mit 1 µM GABA simultan der Oocyte
zugegeben. Jede Applikation dauerte 20 Sekunden. Die Oocyten wurden vor erster Applikation
90 Sekunden und nach jeder Applikation 240 Sekunden mit ND 96 Puffer umspült. Dieses
Versuchsprotokoll wurde auch für die Negativkontrolle angewandt, bei der nicht transfizierte
Oocyten getestet wurden. Die Berechnung der relativen Stromantwort für den GABAA-Rezeptor
durch Lebensmittelextrakte ohne Zugabe von GABA erfolgte anhand folgender Formel:
IRelativ = IMesswert des Extraktes
IGABA× 100%
mit
IRelativ = Relativer Chloridionenstrom für 7,5 µg/mL Extrakt in Abwesenheit von GABA[%]
IMesswert des Extraktes = Die gemessene, durch alleinige Applikation von 7,5 µg/mL Extrakt ohne
Zugabe von GABA ausgelöste Stromantwort [µA]
IGABA = Die durch 1 µM GABA induzierte Stromantwort [µA]
5.6.3.5. Versuchsprotokoll für die Koapplikation von Glycin mit den
Lebensmittelextrakten und Berechnung des relativen Stroms
Zur Bestimmung der Spezifität der GABAA-Rezeptor modulierenden Wirkung der
Lebensmittelextrakte wurde ein Kontrollversuch mit dem Glycin-Rezeptor durchgeführt. Dazu
wurde jeder Lebensmittelextrakt bei der höchsten Konzentration von 5 µg/mL in Abwesenheit
und Anwesenheit von 30 µM Glycin auf die mit dem humanen α1-Glycin-Rezeptor transfizierten
Oocyten appliziert. Die erste Applikation erfolgte durch 5 µg/mL Extrakt in Abwesenheit von
Glycin. Dann wurde Glycin allein in einer Konzentration von 30 µM appliziert. Diese Applikation
galt als Kontrollapplikation. Die letzte Applikation erfolgte durch eine Koapplikation aus 30 µM
Glycin mit 5 µg/mL Extrakt. Jede Applikation dauerte 20 Sekunden. Die Oocyten wurden vor
erster Applikation 90 Sekunden und nach jeder Applikation 240 Sekunden mit der ND 96
Pufferlösung umspült. Die Negativkontrolle erfolgte durch alleinige Applikation von Glycin,
Koapplikation von Glycin in Verbindung mit dem jeweiligen Extrakt oder durch alleinige

Material und Methoden
100
Applikation des Extraktes anhand dieses Versuchsprotokolls auf Oocyten, die nicht mit cRNA
injiziert wurden.
Die relative Stromantwort des Glycin-Rezeptors durch Lebensmittelextrakte wurde
folgendermaßen berechnet:
IRelativ = IMesswert des Extraktes
IGlycin× 100%
mit
IRelativ = Relativer Chloridionenstrom für 5 µg/mL des Extraktes [%]
IMesswert des Extraktes = Der gemessener Chloridionenstrom, der durch Koapplikation von 30 µM
Glycin mit 5 µg/mL Extrakt ausgelöst wurde [µA]
IGlycin = Die durch 30 µM Glycin induzierte Stromantwort [µA]
5.6.3.6. Versuchsprotokoll für die Koapplikation der GABA mit den Fraktionen des
wässrigen Nelkenextraktes und Berechnung des relativen Stroms
Jede Fraktion, welche mittels in Kapitel 5.8 vorgestellter HPLC-Methode erhalten wurde, wurde
in der Konzentration von 5 µg/mL der mit den humanen α1β2 GABAA-Rezeptoren exprimierten
Oocyten mit 1 µM GABA simultan appliziert. Vor Koapplikation der GABA mit der Fraktion
wurde eine Applikation der GABA bei der Konzentration von 1 µM als positive Kontrolle
durchgeführt. Die Applikationszeit war 20 Sekunden. Die Oocyten wurden vor der ersten
Applikation 90 Sekunden und nach jeder Applikation 240 Sekunden mit ND 96 Puffer umspült.
Die Negativkontrolle erfolgte durch alleinige Applikation von GABA und Koapplikation der GABA
in Verbindung mit der Fraktion auf Oocyten, die nicht mit cRNA injiziert wurden.
Um die relative Stromantwort zu bestimmen, wurde die durch 1 µM GABA induzierte
Stromantwort auf 100 % gesetzt. Dann wurde der durch Koapplikation der Fraktion mit 1 µM
GABA ausgelöste Strom auf diese GABA-Kontrollströme normalisiert. Die Berechnung der
relativen Stromantwort durch die jeweiligen Fraktionen erfolgte anhand folgender Formel:
IRelativ = IMesswert der Koapplikation der Fraktion
IGABA× 100%
mit
IRelativ = Relativer Chloridionenstrom der Fraktion [%]
IMesswert der Koapplikation der Fraktion = Der gemessene Chloridionenstrom, der durch Koapplikation der 1
µM GABA mit 5 µg/mL der Fraktion ausgelöst wurde [µA]
IGABA = Die durch 1 µM GABA induzierte Stromantwort [µA]

Material und Methoden
101
5.6.3.7. Versuchsprotokoll für die Koapplikation der GABA mit den Reinsubstanzen
Eugenol und Acetyleugenol und Berechnung des relativen Stroms
Um die für GABAA-Rezeptor modulierende Wirkung des Nelkenextraktes verantwortliche
Substanz zu bestimmen, wurden die mit Hilfe der chromatographischen Trennmethoden
identifizierten Nelkeninhaltsstoffe Eugenol und Acetyleugenol auf die mit der humanen α1β2
GABAA-Rezeptoren transfizierten Oocyten appliziert. Hierzu wurden die in Kapitel 5.9.1.1
hergestellten Standardlösungen des Eugenols bzw. Acetyleugenols mit ND 96 Puffer gemäß ihrer
Applikationskonzentration verdünnt.
Die Applikation-Konzentrationen von Eugenol wurden anhand der Ergebnisse der UHPLC-DAD-
Untersuchungen ermittelt, so dass das Eugenol-Gehalt in der 1 mg/mL Nelken-Fraktion 11 als 23
µg/mL und im 0,25 mg/mL Nelkenextrakt als 43 µg/mL bestimmt wurde (siehe Kapitel 4.3.1).
Basierend auf diesen Daten wurde die Eugenol-Konzentration in der 5 µg/mL Nelken-Fraktion
11 als 0,1 µg/mL und im 5 µg/mL Nelkenextrakt als 1 µg/mL berechnet. Als erste Koapplikation
wurde 0,1 µg/mL Eugenol mit 1 µM GABA der Oocyte zugeführt. Dann wurden 5 µg/mL Nelken-
Fraktion 11, Eugenol in der Konzentration von 1 µg/mL sowie 5 µg/mL Nelkenextrakt simultan
mit 1 µM GABA koappliziert.
Zur Erfassung der modulierenden Wirkung wurde 0,25 µg/mL des Acetyleugenols durch eine
simultane Applikation mit 1 µM GABA der Oocyte zugeführt. Nach der Kontrollapplikation
wurde Acetyleugenol bei der Konzentration von 1 µg/mL in Anwesenheit von 1 µM GABA der
Oocyte appliziert wie Eugenol, um seine modulierende Wirkung auf GABAA-Rezeptoren mit der
durch Eugenol hervorgerufenen Wirkung zu vergleichen.
Vor und nach jeder Koapplikation der Reinsubstanzen bzw. der Nelken-Fraktion 11 oder des
Nelkenextraktes wurde eine Applikation der 1 µM GABA durchgeführt. Dies diente als
Kontrollapplikation. Jede Applikation dauerte 20 Sekunden. Die Oocyten wurden vor der ersten
Applikation 90 Sekunden und nach jeder Applikation 180 - 240 Sekunden mit ND 96 Puffer
umspült. Bei der Negativkontrolle wurden die nicht-transfizierten Oocyten anhand dieses
Versuchsprotokolls getestet.
Zur Berechnung der relativen Stromantworten wurde der mittlere Strom der 1 µM GABA
Kontrollapplikationen, die vor und nach jeder Koapplikation durchgeführt wurden, auf 100 %
gesetzt. Anschließend wurde der gemessene Strom der Koapplikation von Substanzen, Fraktion
oder Extrakt auf diese ermittelte prozentualen GABA-Kontrollströme normalisiert. Die
Berechnung der relativen Stromantwort für den GABAA-Rezeptor erfolgte anhand folgender
Formel:

Material und Methoden
102
IRelativ = IMesswert der Koapplikation
(IGABA(vor der Koapplikation ) + IGABA(nach der Koapplikation )) 2⁄× 100%
mit
IRelativ = Relativer Chloridionenstrom des Eugenols bzw. Acetyleugenols, des Nelkenextraktes
oder seiner Fraktion 11 [%]
IMesswert der Koapplikation = Der gemessene Chloridionenstrom, der durch Koapplikation der 1 µM
GABA mit dem Eugenol bzw. Acetyleugenol, dem Nelkenextraktes oder seiner Fraktion 11
ausgelöst wurde [µA]
IGABA(vor der Koapplikation ) =Stromantwort durch alleinige Applikation von 1 µM GABA, die vor der
Koapplikation der 1 µM GABA mit dem Eugenol bzw. Acetyleugenol, dem Nelkenextraktes oder
seiner Fraktion 11 appliziert wurde [µA]
IGABA(nach der Koapplikation ) = Stromantwort durch alleinige Applikation von 1 µM GABA, die nach der
Koapplikation der 1 µM GABA mit dem Eugenol bzw. Acetyleugenol, dem Nelkenextraktes oder
seiner Fraktion 11 appliziert wurde [µA]

Material und Methoden
103
5.7. Bestimmung der GABA mittels HPLC
5.7.1. Lösungen und Eluenten
Derivatisierungsreagenzen
Für das 0,3 % igen 2-Hydroxynaphtaldehyd -Derivatisierungsreagenz wurden 0,3 mg 2-Hydroxy-
naphtaldehyd in einen 100 mL Messkolben eingewogen und mit Methanol bis zur Marke
aufgefüllt. Für die Herstellung des Orthophtalaldehyd -Derivatisierungsreagenzes wurden 100
mg Orthophtalaldehyd eingewogen und mit dem Methanol auf 10 mL aufgefüllt. Beide
Derivatisierungsreagenzen wurden arbeitstäglich frisch hergestellt.
0,1 M Borax-Puffer, pH: 8
3,814 g Borax wurden in ca. 90 mL Reinstwasser gelöst. Anschließend wurde der pH Wert mit
Salzsäure auf 8,0 eingestellt und die Lösung wurde mit Reinstwasser auf 100 mL aufgefüllt. Die
Lagerung erfolgte bei 4 °C.
90 mM Natriumacetatpuffer in 7 % Acetonitril, pH: 6,5
Für die Herstellung des Natriumacetatpuffers als Elutionsmittel wurden 7, 38 g Natriumacetat in
ca. 900 mL 7%iger wässrige Acetonitril-Lösung gelöst. Der pH Wert wurde mit Essigsäure auf 6.5
eingestellt und die Lösung wurde anschließend auf 1 L mit der 7%igen Acetonitril-Lösung
aufgefüllt.
5.7.2. Derivatisierungsmethoden
5.7.2.1. 2-Hydroxynaphtaldehyd Derivatisierung
Die 2-Hydroxynaphtaldehyd Derivatisierungsmethode wurde nach Khuhawar et al. [195]
modifiziert. Dazu wurden 1 mL der GABA-Standardlösung [100 µg/mL] bzw. 1 mL Reinstwasser
(Blindwert) mit 1 mL des 2-Hydroxynaphtaldehyd Derivatisierungsreagenzes [0,3 % in
Methanol] sowie 0,6 mL des Borax-Puffers (0,1 M; pH: 8,0) versetzt, gemischt und 10 Minuten
im Wasserbad bei 80 °C inkubiert. Dieser Reaktionsansatz wurde nach Abkühlung auf
Raumtemperatur mit 2,4 mL Methanol gemischt, membranfiltriert, und anschließend mittels
RP-HPLC-UV analysiert.

Material und Methoden
104
5.7.2.2. Orthophtalaldehyd (OPA) Derivatisierung
Orthophtalaldehyd (OPA) reagiert mit Aminosäuren und biogenen Aminen im alkalischen Milieu
und bildet fluoreszierende Verbindungen. In der vorliegenden Arbeit wurde eine von Zhao et al.
beschriebene OPA Derivatisierung und anschließende RP-HPLC-FLD Messung [189] mit
mehreren Modifikationen durchgeführt. Dazu wurden 0,4 mL der Probe (GABA-Standardlösung
oder Lebensmittelextrakt) mit 0,02 mL OPA [10 mg/mL in Methanol]-Derivatisierungsreagenz
und 0,2 mL NaOH [1 N] versetzt. Nach gründlicher Durchmischung wurde der Reaktionsansatz 2
Minuten bei Raumtemperatur zur Inkubation stehen gelassen. Anschließend wurden 0,4 mL
Salzsäure [0,7 N] zugegeben, um die Derivatisierungsreaktion zu stoppen. Nach
Membranfiltration erfolgte die Messung mittels RP-HPLC-FLD. Als Negativkontrolle wurden 0,4
mL Reinstwasser anhand dieser OPA-Derivatisierung-Methode derivatisiert und anschließend
vermessen.
5.7.3. Analyse der GABA
Für die Bestimmung der GABA wurden die nachfolgenden HPLC-Parameter verwendet. Dabei
wurden alle Elutionsmittel vor der Verwendung membranfiltriert.
5.7.3.1. RP-HPLC-UV Parameter
HPLC: Jasco, Semi-präparative HPLC (I)
Säule: Zorbax Eclipse XDB-C8 (4,6 x 150 mm, 5 µM)
Flussrate: 0,7 mL/min
Einspritzvolumen: 50 µL
Detektion: Multiwavelength-Detektor mit der Anregungswellenlänge von 330 nm
Elutionsmittel: Isokratisch, Reinstwasser : Methanol (35 : 65)

Material und Methoden
105
5.7.3.2. RP-HPLC-FLD Parameter
HPLC: Jasco, Semi-präparative HPLC (I)
Säule: Zorbax Eclipse XDB-C8 (4,6 x 150 mm, 5 µM)
Flussrate: 1 mL/min
Einspritzvolumen: 50 µL
Detektion: Fluoreszenzdetektor mit der Anregungswellenlänge von 230 nm und
Emissionswellenlänge von 450 nm
Elutionsmittel A: 90 mM Natriumacetatpuffer in 7 % Acetonitril (pH : 6,5)
Elutionsmittel B: 80 % Acetonitril
Gradient: Zeit [min] A [%] B [%]
0 100 0
10 100 0
15 85 15
16 5 95
21 5 95
22 100 0
25 100 0
5.7.3.3. Identifizierung und Quantifizierung der in den Lebensmitteln enthaltenen GABA
Zur Bestimmung der GABA-Gehalte der Lebensmittelextrakte wurde eine OPA-Derivatisierung
mit anschließender RP-HPLC-FLD Messung durchgeführt. Hierzu wurden die wässrigen
Lebensmittelextrakte in unterschiedlichen Konzentrationen (5-40 mg/mL) hergestellt und
dreimal unabhängig voneinander anhand der in Kapitel 5.7.2.2 vorgestellten OPA-
Derivatisierungsmethode derivatisiert. Anschließend wurde das GABA-Derivat mittels RP-HPLC-
FLD (siehe Kapitel 5.7.3.2) analysiert. Um die in den Lebensmitteln enthaltene GABA zu
identifizieren, wurden die Retentionszeit und das Fluorenzspektrum der Lebensmittelextrakte
mit der Referenzsubstanz GABA [0,1 mg/mL] verglichen, die bei jedem Versuch vor oder nach
dem Extrakt in die HPLC eingespritzt wurde. Zur Quantifizierung der GABA wurde eine externe
Kalibrierung mit GABA-Standardlösungen verwendet.

Material und Methoden
106
5.7.3.3.1. Kalibrierung und Linearität
Die Kalibrierung der Methode erfolgte durch Standardlösungen von GABA. Dazu wurde 1 mg
GABA in einen 10 mL Messkolben eingewogen und der Messkolben mit Reinstwasser bis zur
Marke aufgefüllt. Anschließend wurde diese Standardlösung mit Reinstwasser auf die folgenden
GABA-Konzentrationen eingestellt: 1 µg/mL, 2,5 µg/mL, 5 µg/mL, 10 µg/mL, 15 µg/mL, 20 µg/mL
und 25 µg/mL. Jede Konzentrationsstufe wurde dreimal unabhängig voneinander mit OPA
anhand der in Kapitel 5.7.2.2 beschriebenen Methode derivatisiert und anschließend mittels RP-
HPLC-FLD vermessen. Durch Auftragung der Peakflächen gegen die zugehörige GABA-
Konzentration wurde eine Kalibriergerade (Abbildung im Anhang A.2) erstellt. Die Prüfung der
Kalibrierung erfolgte durch Berechnung des Bestimmtheitsmaßes (R2) der linearen Regression.
Mit Hilfe der Kalibrierfunktion (lineare Regression; y = 6033,9x + 2,3312) konnten aus den
gemessenen Peakflächen der Lebensmittelextrakte die Konzentrationswerte der GABA
berechnet werden.
5.7.3.3.2. Nachweis- und Bestimmungsgrenze
Die NWG bzw. BSG wurden mittels Regressionrechnung aus der externen GABA- Kalibrierung
ermittelt. Dazu wurden zunächst die Steigung (b), die Reststandardabweichung (Sy,x) und die
Verfahrensstandartabweichung (SxO) wie folgt berechnet:
Steigung (b)=∑ (x-x̅)(y-y̅)
∑ (x-x̅)2
Reststandardabweichung (Sy,x)=√
∑ (y-y̅)2 -[ ∑ (x-x̅)(y-y̅)]
2
∑ (x-x̅)2
n-2
Verfahrensstandartabweichung (SxO)=Sy,x
b
NWG= SxO . tf,a√1
n+
1
m+
x̅2
∑ (x-x̅)2
BSG=k . SxO . tf,a√1
n+
1
m+
(k.NWG-x̅)2
∑ (x-x̅)2
mit
x = Die Konzentration der GABA-Standardlösung
x̅ = Arithmetischer Mittelwert für die Konzentration
y = Die gemessene Peakfläche der GABA-Standardlösung
y̅ = Arithmetischer Mittelwert der Peakflächen

Material und Methoden
107
n = Die Zahl der verwendeten Kalibrationsstandards
m = Die Zahl der Messungen
tf,a= t-Wert aus t-Tabelle für zweiseitigen Test; f : Freiheitsgrade (f = n-2) und α: Irrtumswahr-
scheinlichkeit (α=0,05)
k = 3; 1/k : Relative Ergebnisunsicherheit (33 %)
5.8. Fraktionierung des Extraktes mittels RP-HPLC
Für die Fraktionierung des wässrigen Nelkenextraktes wurden nachfolgende HPLC-Parameter
verwendet. Dabei wurden alle Elutionsmittel vor der Verwendung membranfiltriert.
HPLC: Jasco, Semi-präparative HPLC (II)
Säule: VP 250/10 Nucleodur 100-5 C18ec (10 x 250 mm, 5 µM)
Vorsäule: VP 10/8 Nucleodur 100-5 C18ec (10 x 8 mm, 5 µM)
Flussrate: 3 mL/min
Einspritzvolumen: 500 µL
Detektion: 4 λ UV/Vis-Detektor mit der Anregungswellenlänge von 254 nm
Elutionsmittel A: Reinstwasser
Elutionsmittel B: Methanol
Gradient: Zeit [min] A [%] B [%]
0 80 20
25 5 95
30 5 95
35 80 20
40 80 20

Material und Methoden
108
Fraktionierungszeit: Zeit [min] Fraktionen
0,0 - 3,0 Fraktion 1
3,0 - 6,8 Fraktion 2
6,8 - 8,4 Fraktion 3
8,4 - 10,8 Fraktion 4
10,8 - 11,7 Fraktion 5
11,7 - 13,3 Fraktion 6
13,3 - 16,8 Fraktion 7
16,8 - 18,7 Fraktion 8
18,7 - 20,7 Fraktion 9
20,7 - 23,4 Fraktion 10
23,4 - 24,9 Fraktion 11
24,9 - 34 Fraktion 12
34 - 40 Fraktion 13
Mittels Fraktionensammler wurden Fraktionen nach oben definierten Zeitintervallen in 50 mL-
Falcons gesammelt. Die aufgefangenen Fraktionen wurden mittels Vakuumdestillation
eingeengt und anschließend bei -80 °C eingefroren und an der Lyophylle getrocknet. Die
getrockneten Fraktionen wurden in einem entsprechenden Volumen Reinstwasser gelöst, so
dass die Endkonzentrationen 1 mg/mL betrugen. Die Fraktionen wurden bei – 20°C gelagert und
vor der Verwendung für TEVC-Analyse mit ND 96; für UHPLC-DAD Analyse mit Reinstwasser
verdünnt.
5.9. Nachweis der Inhaltsstoffe von Nelken
5.9.1. UHPLC-DAD Methode
5.9.1.1. Herstellung der Referenzsubstanzen
Eugenol Standardlösung (0,5 mg/mL)
2,5 mg Eugenol wurden eingewogen und mit Reinstwasser auf 5 mL aufgefüllt. Die Lagerung
erfolgte bei -20 °C.
Acetyleugenol Standardlösung (0,06 mg/mL)
0,6 mg Acetyleugenol wurden in einen 10 mL Messkolben eingewogen und der Messkolben mit
Reinstwasser bis zur Marke aufgefüllt. Diese Lösung wurde bis zur Verwendung bei -20 °C
gelagert.
Isoeugenol Standardlösung (1 %)
Isoeugenol wurde im Verhältnis 1: 100 mit Reinstwasser verdünnt.

Material und Methoden
109
β-Caryophyllen Standardlösung (0,1%)
β-Caryophyllen wurde im Verhältnis 1: 1000 mit Reinstwasser verdünnt.
5.9.1.2. UHPLC-DAD Parameter
Zur Identifizierung der Inhaltsstoffe von Nelken wurden die nachfolgenden UHPLC-Parameter
verwendet.
Säule: YMC-UltraHT Pro C18 mit Inlinefilter (2 x 50 mm, 2 µM)
Säulentemperatur: 30 °C
Flussrate: 0,3 mL/min
Einspritzvolumen: 10 µL
Detektion: Diodenarraydetektor mit λ = 190 – 500 nm
Elutionsmittel A: Reinstwasser
Elutionsmittel B: Acetonitril
Gradient: Zeit [min] A [%] B [%]
-5 95 5
0 95 5
10 5 95
11 5 95
14 95 5
15 95 5
Alle Elutionsmittel und Proben wurden vor der Verwendung membranfiltriert. Neben
Referenzsubstanzen wurden der wässrige Nelkenextrakt in der Konzentration von 0,25 mg/mL
und seine Fraktion 11 in der Konzentration 1 mg/mL in die UHPLC dreimal unabhängig
miteinander eingespritzt. Um die Substanzen in Nelken und seiner Fraktion 11 zu identifizieren
wurden die Retentionszeit und das UV-Vis Spektrum von Nelken und seiner Fraktion 11 mit
Referenzsubstanzen verglichen.
5.9.1.3. Kalibrierung und Linearität zur Quantifizierung des Eugenols im Nelkenextrakt
bzw. in seiner Fraktion 11
Zur Quantifizierung von Eugenol wurde eine externe Kalibrierung mit Eugenol-Standardlösungen
durchgeführt. Hierzu wurde die Standardlösung des Eugenols (0,5 mg/mL) mit Reinstwasser auf
die folgenden Eugenol-Konzentrationen eingestellt: 0,5 µg/mL, 0,75 µg/mL, 1 µg/mL, 2,5 µg/mL,
5 µg/mL, 7,5 µg/mL, 10 µg/mL, 20 µg/mL, 25 µg/mL, 35 µg/mL, 45 µg/mL und 50 µg/mL. Jede
Konzentrationsstufe des Eugenols wurde dreimal unabhängig voneinander vermessen. Eine
Kalibriergerade (Abbildung im Anhang A.3) wurde erstellt, indem die Peakflächen gegen die

Material und Methoden
110
zugehörigen Eugenol-Konzentrationen aufgetragen wurden. Die Bestimmtheitsmaßes (R2) der
linearen Regression wurde zur Prüfung der Kalibrierung berechnet. Die Konzentrationswerte des
Eugenols konnten aus den gemessenen Peakflächen des Nelkenextraktes und seiner Fraktion 11
mittels der Kalibrierfunktion (lineare Regression; y = 355,26x -0,0103) berechnet werden.
5.9.1.4. Nachweis- und Bestimmungsgrenze
Die Bestimmung des NWG bzw. BSG erfolgte durch Regressionrechnung aus der externen
Eugenol-Kalibrierung. Hierzu wurden die in Kapitel 5.7.3.3 beschriebenen Formeln verwendet.
5.9.2. GC-MS Methode
Mit Hilfe GC-MS wurde der wässrige Nelkenextrakt zur weiteren Identifizierung aktiver
Inhaltsstoffe analysiert. Dazu wurde 100 µL des wässrigen Nelkenextraktes (1 mg/mL) mit 200
µL DCM (Dichlormethan) versetzt und zur Überführung der Inhaltsstoffen aus des
Nelkenextraktes in die DCM-Phase bei Raumtemperatur für 10 Minuten bei 14000 rpm
geschüttelt. Nach der Separation wurde die DCM-Phase abpipettiert und mit wasserfreiem
Natriumsulfat getrocknet. Anschließend wurden die angefertigten Proben zur GC-MS Analyse in
Vials überführt. Referenzsubstanz, β-Caryophyllen, wurde vor der Injektion mit DCM verdünnt.
Zur Identifizierung der Inhaltsstoffe von Nelken wurden die nachfolgenden GC-MS-Parameter
verwendet.
Säule: DB - FFAP (0,32 mm x 30 m, 0,25 µM)
Einspritzvolumen: 2 µL
Trägergas: Helium (1 mL/min)
Detektion: Massenspektrometer
Ionenquelle: 200 °C, Elektronenstoßionisation (EI) mit einer Ionisierungsenergie von
70 eV
Scan-Modus: Full-Scan, m/z 30-300
Temperaturprogram: 40°C/7 min//8°C /min//240°C /15 min

Material und Methoden
111
5.10. Statistische Auswertung
Die statistischen Auswertung erfolgte mit Microsoft® Office Excel 2010, Origin® (Origin® 2015G
von OriginLab Corporation, Northampton, MA 01060 USA) und GraphPad Prism® (Version 6.04,
GraphPad Software Inc., La Jolla, CA 92037 USA). Ergebnisse von Enzymassay, RP-HPLC-FLD und
UHPLC-DAD sind als Mittelwert ± Standardabweichung dargestellt. Für die Daten der
elektrophysiologischen Messungen (TEVC) wurde Mittelwert ± SEM (Standardfehler) erstellt. Die
Berechnung des EC5-, EC15- und EC50- sowie IC50-Wertes erfolgte mittels der Software Origin®.
Zur Duchführung von ANOVA (Varinanzanalyse, aus dem Englischen: analysis of variances)
wurde die Software GraphPad Prism® verwendet. Die ANOVA wurde mit dem Holm-Sidak
Vergleichstest durchgeführt um signifikante Unterschiede zwischen den entsprechenden
Gruppen zu ermitteln. Die zweifaktorielle ANOVA wurde für die Auswertung der TEVC-Daten
eingesetzt. Dadurch wurden Signifikanzniveaus auf diese Weise bestimmt: * p < 0,05, ** p <
0,01, *** p < 0,001 und **** p < 0,0001.

Zusammenfassung
112
Zusammenfassung 6
Lebensmittel bzw. Lebensmittelinhaltsstoffe können die neuronale Aktivität beeinflussen und
somit eine neurophysiologische Wirkung auslösen. γ-Aminobuttersäure (GABA), bekannt als der
wichtigste inhibitorische Neurotransmitter, spielt eine wichtige Rolle bei der Regulation der
Aktivität der exzitatorischen Neurotransmission. Aus diesem Grund ist das GABAerge System
von großer Bedeutung für die Behandlung von neurophysiologischen Störungen und ein Target
für verschiedene Medikamentengruppen, unter anderem für Sedativa, Anxiolytika,
Muskelrelaxantien, Antidepressiva und Antiepileptika (Antikonvulsiva). Das GABAerge System
kann durch zahlreiche Substanzen und auf verschiedene Weise moduliert werden. Aufgrund
unerwünschter Nebenwirkungen der Medikamente gibt es ein großes Interesse an Naturstoffen,
deren Interaktion mit der GABAergen Neurotransmission im Organismus die entsprechenden
neurophysiologischen Wirkungen hervorrufen kann. Auf natürlicher Basis sind verschiedene
Lebensmittel bzw. Lebensmittelinhaltsstoffe in der Lage, das GABAerge System zu beeinflussen.
Obwohl die beruhigende, angstlösende, sedative, antidepressive oder antiepileptische Wirkung
mancher Lebensmittel wie Salbei, Kamille, Hopfen, Melisse, Schwarzer Pfeffer, Nelken, Kakao,
Grüner Tee, Sideritis und Lavendel in verschiedenen tierexperimentellen Modellen oder in
klinischen Studien bereits bewiesen wurde, ist der neurophysiologische Wirkungsmechanismus
von diesen Lebensmitteln noch nicht vollständig aufgeklärt. Daher sollte im Rahmen dieser
Arbeit untersucht werden, ob die aus diesen Lebensmitteln hergestellten wässrigen Extrakte das
GABAerge System modulieren.
Ziel des ersten Teils dieser Arbeit war es, den Einfluss wässriger Extrakte von ausgewählten
Lebensmitteln auf die GABA-abbauenden Enzyme (GABA-AT und SSA-DH) zu untersuchen. Dazu
wurde ein Enzymassay durchgeführt. Dabei wurde festgestellt, dass alle getesteten
Lebensmittelextrakte, mit Ausnahme von Hopfen, Kakao und schwarzem Pfeffer, eine
konzentrationsabhängige Inhibition der Enzymaktivität hervorriefen. Extrakte von Nelken und
S.arguta hemmten die Enzymaktivität am stärksten und wiesen sehr niedrige IC50-Werte
(halbmaximale inhibitorische Konzentration) auf, die mit denen des Antiepileptikums Vigabatrin,
einem bekannten Inhibitor der GABA-Transaminase, vergleichbar waren (IC50-Wert mit 95 %
Konfidenzintervall für Vigabatrin: 0,025 (0,049-0,013) mg/mL, für Nelken 0,023 (0,034-0,016)
mg/mL und für S.arguta 0,022 (0,024-0,020) mg/mL). Des Weiteren wurden insgesamt 40
verschiedene exemplarisch ausgewählte terpenoide und flavanoide Lebensmittelinhaltsstoffe
bei einer Konzentration von 1 mM getestet. Einige dieser Inhaltsstoffe sind auch in den
untersuchten Lebensmittelextrakten enthalten. Es zeigte sich, dass β-Damascenon, Myrcen, β-
Ionon und (-)-Caryophyllenoxid auf die Enzymaktivität hemmend wirkten und diese Inhibition

Zusammenfassung
113
von ihrer eingesetzten Konzentration abhing. Hierbei löste β-Damascenon die stärkste
Hemmung der untersuchten Enzyme mit einem IC50-Wert von 0,573 (0,681-0,466) mM aus.
Das zweite Ziel der vorliegenden Dissertation bestand darin, die Wirkung der wässrigen
Lebensmittelextrakte auf GABAA-Rezeptoren elektrophysiologisch zu untersuchen. Hierfür
wurden die GABA-vermittelten Chloridionenströme über die humanen α1β2- GABAA-Rezeptoren
bestimmt, der rekombinant in Xenopus leavis Oocyten exprimiert wurde. Die
elektrophysiologische Messung erfolgte durch die Voltage-Clamp (TEVC) Technik. Die
Lebensmittelextrakte wurden in einem niedrigen Konzentrationsbereich (0,5-5 μg/mL) mit 1 µM
GABA simultan appliziert. Es zeigte sich, dass alle untersuchten 13 Lebensmittelextrakte den
durch 1 µM GABA induzierten Strom signifikant verstärkten. Dabei wurde nachgewiesen, dass
die durch Lebensmittelextrakte ausgelösten Potenzierungen des GABAA-Rezeptors
konzentrationsabhängig waren und mit steigender Konzentration der Extrakte zunahmen.
S.condensata, S. arguta, S. stricta, Grüner Tee und Melisse riefen eine signifikante Potenzierung
ab einer Konzentration von 0,75 μg/mL hervor. Der erste signifikant verstärkende Effekt bei
Applikation von Kamille trat bei der Konzentration von 1 μg/mL auf, während die anderen
Lebensmittelextrakte bei allen eingesetzten Konzentrationen die GABA-induzierten Ströme
signifikant potenzierten. Bei der höchsten applizierten Konzentration führten alle untersuchten
Lebensmittelextrakte zur einer höchst signifikanten Verstärkung der GABA-vermittelten
Stromantwort (p < 0.0001). Die stärkste Potenzierung wurde durch den Nelkenextrakt ausgelöst,
der den GABA-vermittelten Strom um bis zu 426 ± 23 % verstärkten.
Da GABA in vielen Lebensmitteln vorkommt, sollte im nächsten Schritt aufgeklärt werden, ob die
untersuchten wässrigen Lebensmittelextrakte endogene GABA als natürlichen Inhaltsstoff
enthalten und ob diese GABA-Gehalte einen messbaren Einfluss auf die Chloridionenströme
ausüben. Die Bestimmung der GABA in den Lebensmittelextrakten erfolgte nach Derivatisierung
mit ortho-Phtalaldehyd (OPA) mittels Umkehrphasen-Hochleistungsflüssigchromatographie (RP-
HPLC) mit Fluoreszenzdetektor (FLD). Durch den Vergleich der Retentionszeiten und
Fluoreszenzspektren der entsprechenden Signale und der GABA-Referenzsubstanz konnte GABA
in den Lebensmittelextrakten eindeutig identifiziert werden. Die Quantifizierung erfolgte über
eine externe Kalibrierung. Mit Ausnahme von Nelkenextrakt, bei dem die Konzentration der
GABA unterhalb der Nachweisgrenze (NWG) lag, wurde GABA als ein Inhaltsstoff in allen
untersuchten wässrigen Lebensmittelextrakten in einem niedrigen Konzentrationsbereich
quantifiziert (0,14-0,81 μg/mL). Zur Erfassung eines möglichen agonistischen Effekts dieser in
den Lebensmittelextrakten enthaltenen endogenen GABA oder anderer natürlicher Substanzen
wurden die Lebensmittelextrakte ohne Zugabe des Liganden GABA elektrophysiologisch
untersucht. Bei der alleinigen Applikation der Lebensmittelextrakte wurde ein Ionenstrom mit
einer sehr kleinen Stromamplitude beobachtet, die 1-19 % des durch 1 μM GABA-induzierten

Zusammenfassung
114
Stroms betrug. Da die GABAA-Rezeptoren durch Lebensmittelextrakte direkt, also in
Abwesenheit von zugesetzter GABA, nicht aktiviert wurden, konnte die beobachtete
konzentrationsabhängige Potenzierung der GABAA-Rezeptoren bei der simultanen Applikation
der Lebensmittelextrakte mit GABA als eine positive allosterische Modulation charakterisiert
werden.
Anschließend wurde ein Kontrollversuch durchgeführt, um zu prüfen, ob die allosterische
positive modulierende Wirkung der Lebensmittelextrakte spezifisch für die von GABAA-
Rezeptoren gebildeten Chloridionenkanäle ist. Dazu wurde die Wirkung auf Glycin-Rezeptoren
untersucht, da diese Chloridionenkanäle darstellen, die ebenfalls zur Cys-Loop Superfamilie der
ligandengesteuerten Ionenkanäle zählen und eine große Homologie zum GABAA-Rezeptor
aufweisen. Mittels der TEVC-Technik erfolgte die Messung der Glycin-vermittelten
Chloridionenströme an mit rekombinanten humanen α1 Glycin-Rezeptoren transfizierten
Oocyten. Bei der Applikation der Lebensmittelextrakte in Abwesenheit von Glycin wurden keine
Ionenströme, d.h. keine direkte Rezeptoraktivierung beobachtet. Darüber hinaus zeigte sich,
dass keiner der getesteten Lebensmittelextrakte bei einer simultanen Applikation mit 30 μM
Glycin die Glycin-induzierten Ströme potenzierte. Dies lässt darauf schließen, dass die
allosterische positive modulierende Wirkung der untersuchten Lebensmittelextrakte auf die
GABAA-Rezeptoren spezifisch ist. Allerdings riefen die Extrakte von Kamille, schwarzem Pfeffer,
Melisse, Nelken, S. stricta, grünem Tee, S. condensata und Salbei eine signifikante Verringerung
der Glycin-induzierten Ströme hervor, wobei es sich um eine spezifische allosterische negative
Modulation der Glycin-Rezeptoren handeln könnte.
Da Nelkenextrakt im ersten Teil der vorliegenden Arbeit die stärkste Hemmung der GABA-
abbauenden Enzyme verursachte und im zweiten Teil als der potenteste spezifische
allosterische Modulator der GABAA-Rezeptoren identifiziert wurde, war das Ziel des dritten Teils
dieser Dissertation Identifizierung der GABAergen aktiven Komponenten im Nelkenextrakt. Dazu
wurde der wässrige Nelkenextrakt mittels RP-HPLC mit UV/Vis Detektor fraktioniert. Schließlich
wurden die Fraktionen elektrophysiologisch auf eine mögliche GABAA-Rezeptor-modulierende
Wirkung hin untersucht. Dabei führte eine der gewonnenen Fraktionen zur mit Abstand
stärksten Modulation des GABAA-Rezeptors. Zur weiteren Charakterisierung wurden die
Inhaltsstoffe in dieser aus dem Nelkenextrakt isolierten aktivsten Fraktion und im
unfraktionierten Nelkenextrakt mittels Ultrahochleistungsflüssigchromatographie-
Diodenarraydetektor (UHPLC-DAD) sowie mittels Gaschromatographie-Massenspektrometrie
(GC-MS) näher untersucht. Anhand der UV/Vis- und Massenspektren wurde Eugenol als
Hauptbestandteil nicht nur in der aktivsten Fraktion, sondern auch im unfraktionierten
Nelkenextrakt identifiziert. Neben Eugenol wurde Acetyleugenol im Nelkenextrakt detektiert.
Die Quantifizierung von Eugenol erfolgte mittels externer Kalibrierung durch UHPLC-DAD.

Zusammenfassung
115
Aufgrund seiner niedrigen Konzentration konnte Acetyleugenol nicht quantifiziert werden.
Abschließend wurde die Wirkung der beiden identifizieren Komponenten auf den GABAA-
Rezeptor elektrophysiologisch untersucht. Dabei wurde zum ersten Mal eine GABAerge Wirkung
von Acetyleugenol gezeigt. Bei der gleichen Konzentration zeigte das Acetyleugenol eine
stärkere Potenzierung des GABAA-Rezeptors als Eugenol. Aufgrung der sehr geringen
Konzentrationen trägt aber Acetyleugenol nicht wesentlich zur GABAergen Wirkung des
Nelkenextraktes bei. Im Gegensatz hierzu unterschied sich die Verstärkung des GABAA-
Rezeptors durch den im Extrakt enthaltenen Eugenolgehalt nicht signifikant vom
potenzierenden Effekt des Nelkenextraktes. Daraus kann man schließen, dass Eugenol als die
Hauptkomponente des wässrigen Nelkenextraktes im Wesentlichen für den allosterischen
positiven modulatorischen Effekt auf GABAA-Rezeptoren verantwortlich ist. Da Eugenol die Blut-
Hirn-Schranke überwinden kann, ist zu vermuten, dass seine GABAA-Rezeptor modulierende
Wirkung wesentlich an der Auslösung seiner in vivo Studien nachgewiesenen
neurophysiologischen Wirkung wie der analgetischen, antikonvulsiven, sedativen und
angstlösenden Wirkung im Organismus beteiligt sein kann.
Zusammenfassend belegen die Ergebnisse dieser Dissertation, dass die ausgewählten 13
Lebensmittel bereits in niedrigen Konzentrationen den GABA-vermittelten Chloridionenstrom
über GABAA-Rezeptoren allosterisch potenzieren können. Daraus lässt sich schließen, dass diese
Lebensmittelextrakte ihre in der Literatur beschriebene neurophysiologische Wirkung (unter
anderem beruhigend, angstlösend, sedativ, antidepressiv oder antiepileptisch) über denselben
Mechanismus, also durch Modulation der GABAergen neuronalen Aktivität, auslösen können.
Zusätzlich zeigt die vorliegende Arbeit, dass einige Lebensmittelextrakte in der Lage sind, den
Abbau von GABA zu hemmen, indem sie eine Inhibition der GABA-abbauenden Enzyme
hervorrufen. Diese inhibierende Wirkung von Lebensmittelextrakten auf GABA-abbauende
Enzyme könnte zur Auslösung ihrer neurophysiologischen Effekte beitragen. Außerdem zeigte
sich, dass die GABAerge modulierende Wirkung der wässrigen Lebensmittelextrakte
möglicherweise von ihrer besonders in Wasser gut löslichen Bestandteile abhängt und dass
Eugenol als der Hauptbestandteil des Nelkenextraktes für dessen positiven GABAA-Rezeptor
modulierenden Effekt verantwortlich ist. Zur weiteren Charakterisierung, über welche
Mechanismen Eugenol den GABAA-Rezeptor moduliert und ob dieser Effekt spezifisch ist für die
hier untersuchten Rezeptoruntereinheiten, sind weitere elektrophysiologische Experimente
notwendig. Darüber hinaus ist die Untersuchung der GABAergen aktiven Komponenten der
anderen untersuchten wässrigen Lebensmittelextrakte in weiterführenden Studien angestrebt.

Zusammenfassung
116

Abstract
117
Abstract 7
Foods and food ingredients can influence the neural activity and thus could have a
neurophysiological effect. γ-Amino butyric acid (GABA), known as the most important inhibitory
neurotransmitter, plays an important role in regulating activity of the excitatory
neurotransmission. For this reason, the GABaergic system is of great importance in the
treatment of neurophysiological disorders and a target for various groups of medications,
among others for sedatives, anxiolytics, muscle relaxants, antidepressants and antiepileptics
(anticonvulsants). The GABaergic system can be modulated by several substances and in
different ways. Because of adverse effects associated with some of these drugs there is a great
interest in natural products which can exibit the corresponding neurophysiological effect in the
organism by interacting with the GABaergic neurotransmission. Thus, several foods or food
ingredients are able to affect the GABAergic system.
Although the calming, anxiolytic, sedative, antidepressant or antiepileptic effects of some foods
such as Sideritis species, sage, cocoa, lavender, black pepper, chamomile, green tea, hops,
lemon balm, and clove were already demonstrated in various animal models or clinical studies,
the molecular mechanisms of these foods in the central nervous system are not fully elucidated
yet. Therefore, the modulation of the GABAergic system by aqueous extracts from these foods
was investigated in the present study.
The aim of the first part of this work was to examine the effect of aqueous extracts of selected
foods on the GABA-degrading enzymes (GABA-AT and SSA-DH). For this purpose, an enzyme
assay was performed. It was shown that all tested food extracts, except hop, cocoa and black
pepper which exibited no inhibitory effect caused a dose-dependent inhibition of GABA-
degrading enzymes. Extracts of clove and S. arguta showed the most potent inhibition with very
low IC50-values (half maximal inhibitory concentration) which were comparable with the
inhibitory concentration of the antiepileptic drug vigabatrin, a known inhibitor of GABA-AT (IC50-
values with 95 % confidence interval for vigabatrin: 0.025 (0.049-0.013) mg/mL, for clove 0.023
(0.034-0.016) mg/mL and for S.arguta 0.022 (0.024-0.020) mg/mL). Furthermore, 40 selected
food components with terpenoid and flavonoid structure were tested at a concentration of 1
mM. Some of these compounds were also components of the tested food extracts. Among
those, β-damescenone, β-ionone, myrcene and (-)-caryophyllene oxide inhibited enzyme activity
in a concentration- dependent manner. β-Damescenone exerted the strongest inhibitory effect
with an IC50-value of 0.573 (0.681-0.466) mM.
The second purpose of this project was to analyze electrophysiologically the effect of the
aqueous food extracts on GABAA receptors. Therefore, GABA-mediated chloride ion currents via

Abstract
118
the human GABAA α1β2 heteropentameric receptor expressed in Xenopus leavis oocytes were
measured using two-electrode voltage-clamp (TEVC). The food extracts were applied at low
concentrations (0.5-5 µg/mL) simultaneously with 1 µM GABA. It was shown that all thirteen
tested aqueous plant food extracts exerted a significant potentiation of the 1 uM GABA-induced
current. The potentiating effects of food extracts on the GABAA receptor response were
concentration dependent. S.condensata, S. arguta, S. stricta, green tea and lemon balm exibited
significant enhancement of GABAA receptor activity at concentration as low as 0.75 µg/mL. The
first significant enhancement effect of chamomile was observed at the concentration of 1
µg/mL, while the other tested food extracts significantly potentiated the GABA-evoked currents
at all applied concentrations. At the highest concentration, all tested extracts led to a highly
significant enhancement of GABA-mediated current response (p < 0.0001). The highest
potentiation was caused by the clove extract that elicited a potentiation of GABA-evoked
current by 426 ± 23 %.
Since GABA is found in many foods, the next step evaluated whether the test extracts
themselves contain endogenous GABA and whether this GABA content exerts a measurable
influence on the chloride ion currents. The GABA concentrations in food extracts were analysed
by reversed phase high performance liquid chromatography (RP-HPLC) with fluorescence
detector (FLD) after derivatization with OPA. GABA from food extracts was identified
unequivocally by comparing the retention times and fluorescence spectra of the unknown peaks
with those of a GABA standard. Quantification of GABA was carried out by external calibration
with the GABA standard. With the exception of clove extract, in which the concentration of
GABA was below the detection limit (LOD), GABA was detected as an ingredient in all analysed
aqueous food extracts, but in very low concentrations (0.14 and 0.81 μg/mL). In order to
examine a possible agonistic effect of the endogenous GABA or other natural substances
contained in the food extracts, all aqueous food extracts were electrophysiologically
investigated without the addition of ligand GABA. The currents resulting from the perfusion by
pure food extracts were very small and only reached 1-19% of the current evoked by 1 μM
GABA. Because the GABAA receptors were not activated by the food extracts directly in the
absence of added GABA, the active components in the extracts function most likely as positive
allosteric modulators.
Subsequently, a control experiment was performed to check whether the positive allosteric
modulatory effect of food extracts is specific for chloride ion channels formed by GABAA
receptors. For this purpose, the food extracts were also tested for their effects on the glycine
receptor, since it belongs, similar to the GABAA receptors, to the Cys-loop superfamily of ligand-
gated ion channels, shows high homology and also forms a chloride ion channel. By means of
TEVC, the glycine-mediated chloride ion currents via human α1 glycine-receptors expressed in

Abstract
119
Xenopus oocytes were measured. When the test extracts were applied in the absence of glycine,
no substance-induced current indicating a direct activation of receptors could be observed.
Moreover, it was shown that none of the tested food extracts enhanced the glycine-evoked
responses at the homomeric α1-glycine receptors during co-application with 30 µM glycine.
These results indicate that the tested food extracts show specific positive modulatory activity
towards the GABAA receptors. In contrast, the extracts of chamomile, black pepper, lemon balm,
clove, S. stricta, green tea, S. condensata and sage caused a significant decrease of glycine-
mediated currents indicating rather allosteric negative modulation of the glycine receptors.
Since clove extract caused in the first part of this work the strongest inhibition of GABA-
degrading enzymes and was identified in the second part as the most potent specific allosteric
modulator, the aim of the third part of this thesis was the identification of GABAergic active
compounds in the clove extract. For this purpose, the aqueous clove extract was fractionated by
RP-HPLC with UV/Vis detector and obtained fractions were examined electrophysiologically for
possible GABAA receptor-modulatory effects. One of the collected fractions evoked by far the
strongest modulation of GABAA receptor. To further characterize the components in the most
active fraction, ultra-high performance liquid chromatographic with diode array detector
(UHPLC-DAD) and gas chromatography-mass spectrometry (GC-MS) were applied. Based on
UV/Vis- and mass spectra, eugenol was identified as the principal component not only in the
most active fraction, but also in non-fractionated clove extract. Besides of eugenol,
acetyleugenol could be detected in the aqueous clove extracts. Quantification of eugenol was
performed by UHPLC-DAD using external calibration. Concentrations of acetyleugenol were
below the limit of quantification. Finally, the GABAA receptor-modulatory effect of both
components identified in the clove extract was electrophysiologically investigated. It was shown
for the first time that acetyleugenol exerts GABAergic activity. Compared to the application of
eugenol in the same concentration, acetyleugenol exhibited a significantly higher potentiation
of GABAA receptor activity. However, due to its very low concentrations in the aqueous extracts,
its contribution to the GABAergic activity was negligible. In contrast, the potentiating effect of
eugenol applied at the concentration contained in the clove extract on the response of GABAA
receptor function was not significantly different from that of the clove extract itself. Thus, it can
be concluded that eugenol is the principal component of clove responsible for the positive
allosteric modulation of the GABAA receptor. As eugenol can cross the blood-brain barrier, it can
be assumed that its GABAA receptor-modulatory effect may be involved substantially in
triggering the neurophysiological effects of eugenol such as analgesic, anticonsulvant, sedative
and anxiolytic effects demonstrated in vivo studies.
In summary, the results of this thesis demonstrate that the selected 13 foods can potentiate
already at low concentration allosterically the GABA-mediated chloride ion current via GABAA

Abstract
120
receptor. It can thus be concluded that these food extracts may exert their neurophysiological
activities, such as calming, anxiolytic, sedative, antidepressant or antiepileptic effects, via the
same mechanism, by modulating the GABAergic neuronal activity. In addition, the present work
shows that some food extracts may be able to increase GABA concentration by inhibition of
GABA-degrading enzymes. This inhibitory effect of food extracts on GABA-degrading enzymes
could additionally contribute to their neurophysiological effects. It was also shown that the
GABAA receptor modulating activity of plant foods is mainly mediated by their water soluble
components. Eugenol was further identified as the component of clove extract, which is mainly
responsible for its positive modulatory effect on GABAA receptor activity. Further
electrophysiological experiments are required to investigate the mechanism how eugenol
modulates the GABAA receptor and whether this effect is specific for the applied receptor
subunits. Further studies should also determine the active components of the other tested
aqueous food extracts.

121

A Anhang
122
A Anhang A1 Konzentrationsabhängige Wirkung der Lebensmittelextrakten auf die Aktivität GABA-
AT und SSA-DH Tabelle A.1 Konzentrationsabhängige Wirkung von wässriger Lebensmittelextrakten auf die Aktivität der
GABA-metabolisierenden Enzyme (GABA-AT und SSA-DH). Die photometrische Messung
erfolgte bei 340 nm nach einer 30-minutiger Inkubation bei 37 °C. Die Daten weisen den
Mittelwert ± Standardabweichung aus drei unabhängigen Messungen auf.
Nelken
Konzentration [mg/mL] 0,02 0,04 0,05 0,07 0,09
Aktivität [%] 62,1 ± 7,7 35,4 ± 3,8 31,2 ± 5,9 21,0 ± 5,2 14,7 ± 8,9
SNW vs. Neg. Kontr. **** **** **** **** ****
Sideritis
arguta
Konzentration [mg/mL] 0,01 0,02 0,04 0,05 0,07
Aktivität [%] 70,8 ± 4,6 55,8 ± 7,4 40,0 ± 5,9 26,9 ± 5,2 20,2 ± 3,5
SNW
vs.
Neg.Kontr **** **** **** **** ****
Nelken nt ns ns ns ns
Sideritis
stricta
Konzentration [mg/mL] 0,02 0,04 0,05 0,07 0,09
Aktivität [%] 80,6 ± 2,9 61,8 ± 8,7 50,3 ± 3,7 49,8 ± 10,0 44,8 ± 7,6
SNW
vs. Neg.Kontr. **** **** **** **** ****
Nelken *** **** *** **** ****
S.arguta nt **** **** **** ****
Sideritis
sipylea
Konzentration [mg/mL] 0,02 0,04 0,05 0,07 0,09
Aktivität [%] 88,2 ± 5,1 71,4 ± 5,9 61,4 ± 7,0 52,7 ± 4,4 38,9 ± 0,8
Signifi-
kanz
wert
vs.
Neg.Kontr. ** **** **** **** ****
Nelken **** **** **** **** ****
S.arguta nt **** **** **** ****
S.stricta ns ns ns ns ns
Sideritis
condensata
Konzentration [mg/mL] 0,02 0,04 0,05 0,07 0,09
Aktivität [%] 80,9 ± 4,2 68,7 ± 3,8 59,2 ± 8,7 53,8 ± 5,0 43,6 ± 7,7
SNW
vs.
Neg.Kontr. **** **** **** **** ****
Nelken *** **** **** **** ****
S. arguta nt **** **** **** ****
S. stricta ns ns ns ns ns
S. sipylea ns ns ns ns ns

123
Grüner Tee
Konzentration [mg/mL] 0,02 0,04 0,05 0,07 0,09
Aktivität [%] 80,5 ± 5,8 57,6 ± 5,7 46,3 ± 4,7 38,7 ± 2,0 33,9 ± 2,9
SNW
vs.
Neg.Kontr. **** **** **** **** ****
Nelken *** **** ** *** ***
S. arguta **** **** **** **** nt
S. stricta ns ns ns ns ns
S. sipylea ns * ** * ns
S. condensata ns ns * ** ns
Salbeitee
Konzentration [mg/mL] 0,02 0,04 0,05 0,07 0,09
Aktivität [%] 82,9 ± 3,1 65,5 ± 1,9 57,7 ± 1,3 40,6 ± 3,8 34,4 ± 3,7
SNW
vs.
Neg.Kontr. *** **** **** **** ****
Nelken **** **** **** *** ***
S.arguta **** **** **** **** nt
S. stricta ns ns ns ns ns
S. sipylea ns ns ns * ns
S. condensata ns ns ns * ns
Grünertee ns ns ns ns ns
Melisse
Konzentration [mg/mL] 0,03 0,05 0,09 0,12 0,15
Aktivität [%] 75,2 ± 0,7 68,4 ± 5,7 61,0 ± 9,4 52,7 ± 9,6 44,4 ± 8,9
SNW vs. Neg. Kontr. **** **** **** **** ****
Kamille
Konzentration [mg/mL] 0,03 0,05 0,08 0,12 0,17
Aktivität [%] 74,7±10,7 68,6 ± 6,4 61,9 ± 6,3 57,0 ± 5,5 48,1 ± 4,1
SNW vs. Neg. Kontr. **** **** **** **** ****
Lavendel
Konzentration [mg/mL] 0,03 0,06 0,10 0,13 0,16
Aktivität [%] 85,8 ± 3,1 69,0 ± 8,1 48,8 ± 9,4 46,1 ± 2,8 41,9 ± 4,9
SNW vs. Neg. Kontr. ** **** **** **** ****
SNW: Signifikanzwert
nt: nicht statistisch getestet; da die gleiche Konzentration nicht eingesetzt wurde.
ns: nicht signifikant ; * : p<0,05; **: p<0,01; ***: p<0,001; und ***: p<0,0001; zweifaktorielle ANOVA Holm-
Sidak Vergleichstest

A Anhang
124
A2 Kalibrationskurve zur Quantifizierung der GABA
Abbildung A1 Kalibrationskurve zur Quantifizierung der GABA. GABA-Standards wurden in einem
Konzentartionsbereich 0, 001 -0,025 mg/mL analysiert. Jede Konzentratiosstufe wurde
dreimal unabhängig voneinander mit OPA derivatisiert und anschließend mittels RP-HPLC-
FLD vermessen.
A3 Kalibrationskurve zur Quantifizierung des Eugenols
Abbildung A2 Kalibrationskurve zur die Quantifizierung des Eugenols. Eugenol-Standards wurden in einem
Konzentartionsbereich 0, 5 -50 µg/mL analysiert. Jede Konzentratiosstufe wurde dreimal
unabhängig voneinander mittels UHPLC-DAD vermessen.
0
20
40
60
80
100
120
140
160
180
0 0,005 0,01 0,015 0,02 0,025 0,03
Pe
akfl
äch
e [m
V*s
]
Konzentration [mg/mL]
0
5
10
15
20
0 15 30 45 60
Pea
kflä
che
[mA
U*m
in]
Konzentration [µg/mL]

125
Literaturverzeichnis
1. Weltgesundheitsorganization. Faktenblatt zu psychisher Gesundheit. WHO-Regionalkomitee
für Europa, 63.Tagung, abgerufen am 10.11.2015; Available from:
http://www.euro.who.int/__data/assets/pdf_file/0012/216210/RC63-Fact-sheet-MNH-
Ger.pdf?ua=1.
2. Jacobi, F., Wittchen, H. U., Hölting, C., Höfler, M., Pfister, H., Müller, N., Lieb, R., Prevalence,
co-morbidity and correlates of mental disorders in the general population: results from the
German Health Interview and Examination Survey (GHS). Psychological Medicine, 2004.
34(4): p. 597-611.
3. Jacobi, F., Höfler, M., Strehle, J., Mack, S., Gerschler, A., Scholl, L., Busch, M.A., Maske, U.,
Hapke, U., Gaebel, W., Maier, W., Wagner, M., Zielasek, J., Wittchen, H.U., Psychische
Störungen in der Allgemeinbevölkerung. Studie zur Gesundheit Erwachsener in Deutschland
und ihr Zusatzmodul Psychische Gesundheit (DEGS1-MH). Nervenarzt, 2014. 85(1): p. 77-87.
4. RKI, Robert Koch-Institut, Daten und Fakten: Ergebnisse der Studie Gesundheit in
Deutschland aktuell 2009. Beiträge zur Gesundheitsberichterstattung des Bundes, 2011, RKI:
Berlin.
5. RKI, Robert Koch-Institut, Daten und Fakten: Ergebnisse der Studie Gesundheit in
Deutschland aktuell 2010. Beiträge zur Gesundheitsberichterstattung des Bundes, 2012, RKI:
Berlin.
6. RKI, Robert Koch-Institut, Depression. Faktenblatt zu GEDA 2012: Ergebnisse der Studie
"Gesundheit in Deutschland aktuell 2012", 2014, RKI: Berlin.
7. DIMDI. Deutsches Institut für Medizinische Dokumentation und Information, ICD-10- WHO
Version 2016, Kapitel V Psychische und Verhaltensstörungen (F00-F99), F 32 Depressive
Episode. abgerufen am 12.11.2015; Available from:
https://www.dimdi.de/static/de/klassi/icd-10-
who/kodesuche/onlinefassungen/htmlamtl2016/block-f30-f39.htm.
8. Stahmeyer, J.T., Gensichen, J., Walelu, O. A., Petersen, J. .J., Gerlach, F. M., Krauth, C.,
Krankheitskosten von Depressionen – nicht nur die Patienten leiden! Gesundheitswesen,
2011. 73 p. A319.
9. Wittchen, H.U., Jacobi, F., Klose, M., Ryl, L., Depressive Erkrankungen, in
Gesundheitsberichterstattung des Bundes. Heft 512010, Robert Koch-Institut Statistisches
Bundesamt: Berlin.
10. Miledi, R., Trasmitter release induced by injection of calcium ions into nerve terminals. Proc.
R.Soc. Lond.B., 1973. 183: p. 421-425.

Literaturverzeichnis
126
11. Li, L., Chin, L. S., The molecular machinery of synaptic vesicle exocytosis. Cell Mol Life Sci,
2003. 60(5): p. 942-960.
12. Hucho, F., Weise, C., Ligandengesteuerte Ionenkanäle Angewandte Chemie, 2001. 113: p.
3194-3211.
13. Connolly, C.N., Wafford, K. A., The Cys-loop superfamily of ligand-gated ion channels: the
impact of receptor structure on function. Biochemical SocietyTransactions, 2004. 32(Part 3):
p. 529-534.
14. Collingridge, G.L., Olsen, R. W., Peters, J., Spedding, M., A nomenclature for ligand-gated ion
channels. Neuropharmacology, 2009. 56(1): p. 2-5.
15. Jarvis, M.F., Khakh, B. S., ATP-gated P2X cation-channels. Neuropharmacology, 2009. 56(1):
p. 208-215.
16. Olsen, R.W., Sieghart, W., International Union of Pharmacology. LXX. Subtypes of g-
aminobutyric acid(A) receptors: classification on the basis of subunit composition,
pharmacology, and function. Update. Pharmacol Rev, 2008. 60(3): p. 243-260.
17. Mascia, M.P., Machu, T. K., Harris, R. A., Enhancement of homomeric glycine receptor
function by long-chain alcohols and anaesthetics. British Journal of Pharmacology, 1996. 119:
p. 1331-1336.
18. Krasowski, M.D., Harrison, N. L., The actions of ether, alcohol and alkane general
anaesthetics on GABAA and glycine receptors and the effects of TM2 and TM3 mutations.
British Journal of Pharmacology, 2000. 129 p. 731-743.
19. Borden, L.A., GABA Transporter Heterogeneity: Pharmacology And Cellular Localization.
Neurochemistry International, 1996. 29(4): p. 335–356.
20. Soudijn, W., van Wijngaarden, I., The GABA transporter and its inhibitors. Curr Med Chem,
2000. 7(10): p. 1063-1079.
21. Meldrum, B.S., GABAergic mechanisms in the pathogenesis and treatment of epilepsy. British
Journal of Clinical Pharmacology, 1989. 27: p. 3S-11S.
22. Macdonald, R.L., Olsen R. W., GABAA Receptor Channels. Annu. Rev. Neurosci, 1994. 17: p.
569-602.
23. Bowery, N.G., Smart, T. G., GABA and glycine as neurotransmitters: a brief history. Br J
Pharmacol, 2006. 147 (Suppl 1): p. S109-S119.
24. Johnston, G., A., R., , GABAA Receptor Channel Pharmacology. Current Pharmaceutical
Design, 2005. 11(15): p. 1867-1885.

127
25. Awapara, J., Landua, A. J., Fuerst, R., Seale, B., Free γ-aminobutyric acid in brain. Journal of
Biological Chemistry 1950. 187: p. 35-39.
26. Roberts, E., Frankel, S., γ-Aminobutyric acid in brain: Its formation from glutamic acid.
Journal of Biological Chemistry, 1950. 187: p. 55-63.
27. Granger, R.E., Campbell, E. L., Johnston, G. A. R., (+)- And (-)-borneol: efficacious positive
modulators of GABA action at human recombinant a1b2g2L GABAA receptors. Biochem
Pharmacol, 2005. 69(7): p. 1101-1111.
28. Pitkanen, A., Amaral, D.G. , The distribution of GABAergic cells, fibers, and terminals in the
monkey amygdaloid complex: an immunohistochemical and in situ hybridization study. The
Journal of Neuroscience, 1994. 14(4): p. 2200-2224.
29. Isaacson, J.S., Solís, J. M., Nicoll, R. A. , Local and diffuse synaptic actions of GABA in the
hippocampus. Neuron, 1993. 10(2): p. 165-175.
30. Treiman, D.M., GABAergic Mechanisms in Epilepsy. Epilepsia, 2001. 42(3): p. 8-12.
31. Arcelli, P., Frassoni, C., Regondi, M. C., De Biasi, S., Spreafico, R., GABAergic neurons in
mammalian thalamus a marker of thalamic complexity? Brain Res Bull, 1997. 42(1): p. 27-37.
32. Bäckberg, M., Ultenius, C., Fritschy, J. M., Meister, B., Cellular Localization of GABAA
Receptor a Subunit Immunoreactivity in the Rat Hypothalamus: Relationship With Neurones
Containing Orexigenic or Anorexigenic Peptides. Journal of Neuroendocrinology, 2004. 16: p.
589–604.
33. Wingo, W.J., Awapara J., Decarboxylation of L-glutamic acid by brain. Journal of Biological
Chemistry, 1950. 187: p. 267-271.
34. Bu, D.F., Erlander, M. G., Hitz, B. C., Tillakaratne, N. J. K., Kaufman, D. L., Wagner-
McPherson, C. B., Evans, G. A.,Tobin, A. J. , Two human glutamate decarboxylases, 65-kDa
GAD and 67-kDa GAD, are each encoded by a single gene. Proc. Natl. Acad. Sci. USA, 1992.
89: p. 2115-2119.
35. Soghomonian, J.J., Martin, D. L., Two isoforms of glutamate decarboxylase: why? Trends
Pharmacol Sci. , 1998. 19(12): p. 500-505.
36. McIntire, S.L., Reimer, R. J., Schuske, K., Edwards, R. H., Jorgensen, E. M., Identification and
characterization of the vesicular GABA transporter. Nature, 1997. 389(6653): p. 870-876.
37. Jin, H., Wu, H., Osterhaus, G., Wei, J., Davis, K., Sha, D., Floor, E., Hsu, C. C., Kopke, R. D.,Wu,
J. Y., Demonstration of functional coupling between γ-aminobutyric acid (GABA) synthesis
and vesicular GABA transport into synaptic vesicles. Proc Natl Acad Sci U S A, 2003. 100(7): p.
4293-4298.

Literaturverzeichnis
128
38. Pearl, P.L., Novotny, E. J., Acosta, M. T., Jakobs, C., Gibson, K. M., Succinic semialdehyde
dehydrogenase deficiency in children and adults. Ann Neurol, 2003. 54 (Suppl 6): p. S73-S80.
39. Yogeeswari, P., Sriram, D., Vaigundaragavendran, J., The GABA shunt: an attractive and
potential therapeutic target in the treatment of epileptic disorders. Curr Drug Metab., 2005.
6(2): p. 127-139.
40. Beleboni, R.O., Carolino, R. O. G, Pizzo, A. B., Castellan-Baldan, L., Coutinho-Netto, J., dos
Santos, W. F., Coimbra, N. C., Pharmacological and Biochemical Aspects of GABAergic
Neurotransmission: Pathological and Neuropsychobiological Relationships. Cellular and
Molecular Neurobiology, 2004. 24(6): p. 707-728.
41. Bessman, S.P., Rossen, J., Layne, E. C., γ-Aminobutyric Acid-Glutamic Acid Transamination In
Brain. Journal of Biological Chemistry, 1953. 201: p. 385-391.
42. Jakoby, W.B., Enzymes of γ-Aminobutyrate Metabolism (Bacterial). Methods in Enzymology,
ed. S.P. Colowick, Kaplan, N.O. Vol. V. 1962, New York: Academic Press. 765-778.
43. Scott, E.M., .Jakoby, W. B., Soluble γ-Aminobutyric-Glutamic Transaminase from
Pseudomonas fluorescens. Journal of Biological Chemistry, 1959. 234: p. 932-936.
44. Baxter, C.F., Roberts, E. , The γ-Aminobutyric Acid-α-Ketoglutaric Acid Transaminase of Beef
Brain. Journal of Biological Chemistry, 1958. 233(5): p. 1135-1139.
45. Bormann, J., Feigenspan, A.,, GABAC receptors. Trends in Neurosciences, 1995. 18(12): p.
515-519.
46. Bormann, J., The ‘ABC’ of GABA receptors. Trends in Pharmacological Sciences, 2000. 21(1):
p. 16-19.
47. Chebib, M., Johnston, G. A. R., The ‘ABC’ of GABA Receptors: A Brief Review Clinical and
Experimental Pharmacology and Physiology, 1999. 26(11): p. 937-940.
48. Sigel, E., Steinmann, M. E., Structure, function, and modulation of GABAA receptors. J Biol
Chem, 2012. 287(48): p. 40224-40231.
49. Simon, J., Wakimoto, H., Fujita, N., Lalande, M., Barnard, E. A., Analysis of the set of GABAA
receptor genes in the human genome. The Journal Of Biological Chemistry, 2004. 279(40): p.
41422-41435.
50. Rudolph, U., Crestani, F., Möhler H., GABAA receptor subtypes: dissecting their
pharmacological functions. Trends Pharmacol Sci, 2001. 22(4): p. 188-194.
51. Sieghart W., S.G., Subunit composition, distribution and function of GABAA receptor subtypes.
Current Topics in Medicinal Chemistry, 2002. 2: p. 795-816.

129
52. Karim, N., Wellendorph, P., Absalom, N., Johnston, G. A. R., Hanrahan, J. R., Chebib, M.,
Potency of GABA at human recombinant GABAA receptors expressed in Xenopus oocytes: a
mini review. Amino Acids, 2013. 44(4): p. 1139-1149.
53. Karim, N., Wellendorph, P., Absalom, N., Bang, L. H., Jensen, M. L., Hansen, M. M., Lee, H. J.,
Johnston, G. A. R., Hanrahan, J. R., Chebib, M., Low nanomolar GABA effects at extrasynaptic
a4b1/b3d GABAA receptor subtypes indicate a different binding mode for GABA at these
receptors. Biochemical Pharmacology, 2012. 84(4): p. 549-557.
54. Li, J.X., Zhang, Y., Emerging drug targets for pain treatment. Eur J Pharmacol, 2012. 681(1-3):
p. 1-5.
55. McKernan, R.M., Rosahl, T. W., Reynolds, D. S., Sur, C., Wafford, K. A., Atack, J. R., Farrar, S.,
Myers, J., Cook, G., Ferris, P., Garrett, L., Bristow, L., Marshall, G., Macaulay, A., Brown, N.,
Howell, O., Moore, K. W., Carling, R. W., Street, L. J., Castro, J. L., Ragan, C. I., Dawson, G. R.,
Whiting, P. J., Sedative but not anxiolytic properties of benzodiazepines are mediated by the
GABAA receptor α1 subtype. Nat Neurosci, 2000. 3(6): p. 587-592.
56. Rudolph, U., Crestani, F., Benke, D., Brünig, I., Benson, J. A., Fritschy, J. M., Martin, J. R.,
Bluethmann, H., Möhler, H., Benzodiazepine actions mediated by specific γ-aminobutyric
acidA receptor subtypes. Nature, 1999. 401(6755): p. 796-800.
57. Halliwell, R.F., Thomas, P., Patten, D., James, C. H., Martinez-Torres, A., Miledi, R.,Smart, T.
G., Subunit-selective modulation of GABAA receptors by the non-steroidal anti-inflammatory
agent, mefenamic acid. The European Journal of Neuroscience, 1999. 11: p. 2897-2905.
58. Smith, G.B., Olsen, R. W., Functional domains of GABAA receptors. Trends in Pharmacological
Sciences, 1995. 16(5): p. 162–168.
59. Mortensen, M., Patel, B., Smart, T. G., GABA Potency at GABAA Receptors Found in Synaptic
and Extrasynaptic Zones. Frontiers in Cellular Neuroscience, 2011. 6(1): p. 1-10.
60. Farrant, M., Nusser, Z., Variations on an inhibitory theme: phasic and tonic activation of
GABAA receptors. Nat Rev Neurosci., 2005 6(3): p. 215-229.
61. Vithlani, M., Terunuma, M., Moss, S. J., The Dynamic Modulation of GABAA Receptor
Trafficking and Its Role in Regulating the Plasticity of Inhibitory Synapses. Physiol Rev, 2011.
91 p. 1009-1022.
62. Belelli, D., Harrison, N. L., Maguire, J., Macdonald, R. L., Walker, M. C., Cope, D. W.,
Extrasynaptic GABAA Receptors: Form, Pharmacology, and Function. The Journal of
Neuroscience, 2009. 29(41): p. 12757-12763.
63. Nusser, Z., Sieghart, W., Somogyi, P.,, Segregation of Different GABAA Receptors to Synaptic
and Extrasynaptic Membranes of Cerebellar Granule Cells. The Journal of Neuroscience,
1998. 18(5): p. 1693-1703.

Literaturverzeichnis
130
64. Nusser, Z., Roberts, J. D. B., Baude, A., Richards, J. G.,Somogyi, P., Relative Densities of
Synaptic and Extrasynaptic GABAA Receptors on Cerebellar Granuale Cells as Determined by a
Quantitative Immunogold Method. The Journal of Neuroscience, 1995. 15(4): p. 2948-2960.
65. Somogyi, P., Fritschy, J. M., Benke, D., Roberts, J.D., Sieghart, W., The gamma 2 subunit of
the GABAA receptor is concentrated in synaptic junctions containing the alpha 1 and beta
2/3 subunits in hippocampus, cerebellum and globus pallidus. Neuropharmacology, 1996.
35(9-10): p. 1425-1444.
66. Richardson, B.D., Ling, L. L., Uteshev, V. V., Caspary, D. M., Extrasynaptic GABAA Receptors
and Tonic Inhibition in Rat Auditory Thalamus. Plos One, 2011. 6(1): p. e16508.
67. Lydiard, R.B., The Role of GABA in Anxiety Disorders. J Clin Psychiatry 2003. 64(3): p. 21-27.
68. Möhler, H., GABAA receptors in central nervous system disease: anxiety, epilepsy, and
insomnia. J Recept Signal Transduct Res, 2006. 26(5-6): p. 731-740.
69. Roth, F.C., Draguhn, A., GABA metabolism and transport: effects on synaptic efficacy. Neural
Plast, 2012. 2012: p. 805830.
70. Steiger, A., Sleep and its modulation by substances that affect GABAA receptor function.
GABA and sleep -molecular, functional and clinical aspects, ed. J.M. Monti, Pandi-Perumal, S.
R., Möhler, H. . 2010, Basel: Springer.
71. Gottesmann, C., GABA Mechanisms and Sleep. Neuroscience 2002 111(2): p. 231-239.
72. Lancel, M., Steiger, A., Sleep and Its Modulation by Drugs That Affect GABAA Receptor
Function. Angewandte Chemie, 1999. 111: p. 2852-2864.
73. Malan. T. P., M., H. P., Porreca, F., Spinal GABAA and GABAB receptor pharmacology in a rat
model of neuropathic pain. Anesthesiology, 2002. 96(5): p. 1161-1167.
74. Paul, J., Yevenes, G. E., Benke, D., Di Lio, A., Ralvenius, W. T., Witschi, R., Scheurer, L., Cook,
J. M., Rudolph, U., Fritschy, J. M., Zeilhofer, H. U., Antihyperalgesia by a2-GABAA receptors
occurs via a genuine spinal action and does not involve supraspinal sites.
Neuropsychopharmacology, 2014. 39(2): p. 477-487.
75. Chuang, C.Y.S., Y. C. You, H. P. Lo, Y. H. Pan, T. M., Antidepressant effect of GABA-rich
monascus-fermented product on forced swimming rat model. J Agric Food Chem, 2011.
59(7): p. 3027-34.
76. Atack, J.R., The benzodiazepine binding site of GABAA receptors as a target for the
development of novel anxiolytics. Expert Opin Investig Drugs, 2005. 14(5): p. 601-618.
77. Wasowski, C., Marder, M., Flavonoids as GABAA receptor ligands:The whole story? Journal of
Experimental Pharmacology, 2012. 4 p. 9-24.

131
78. Weir, C.J., Ling, A. T. Y., Belelli, D., Wildsmith, J. A. W., Peters, J. A., Lambert, J. J. , The
interaction of anaesthetic steroids with recombinant glycine and GABAA receptors. British
Journal of Anaesthesia, 2004. 92(5): p. 704-711.
79. Aoshima, H., Tenpaku, Y, Modulation of GABA Receptors Expressed in Xenopus Oocytes by
13-L-Hydroxylinoleic Acid And Food Additives. Bioscience, Biotechnology and Biochemistry,
1997. 61(12): p. 2051-2057.
80. Aoshima, H., Hamamoto, K. , Potentiation of GABAA Receptors Expressed in Xenopus Oocytes
by Perfume and Phytoncid. Bioscience, Biotechnology, and Biochemistry, 1999. 63(4): p. 743-
748.
81. Mohammadi, B., Haeseler, G., Leuwer, M., Dengler, R., Krampfl, K., Bufler, J. , Structural
requirements of phenol derivatives for direct activation of chloride currents via GABAA
receptors. European Journal of Pharmacology, 2001. 421(2): p. 85-91.
82. Aoshima, H., Hossain, S. J., Hamamoto, K., Yokoyama, T., Yamada, M., Shingai, R., Kinetic
analyses of alcohol-induced potentiation of the response of GABAA receptors composed of α1
and β1 subunits. Journal of Biochemistry, 2001. 130: p. 703-709.
83. Hossain, S.J., Hamamoto, K., Aoshima, H., Hara, Y.,, Effects of tea components on the
response of GABAA receptors expressed in Xenopus Oocytes. J. Agric. Food Chem., 2002. 50:
p. 3954–3960.
84. Hossain, S.J., Aoshima, H., Koda, H., Kiso, Y., Potentiation of the ionotropic GABA receptor
response by whiskey fragrance. J. Agric. Food Chem. , 2002 50: p. 6828–6834.
85. Hossain, S.J., Aoshima, H., Koda, H., Kiso, Y., Effects of coffee components on the response of
GABAA receptors expressed in Xenopus oocytes. Journal of Agricultural and Food Chemistry,
2003. 51: p. 7568-7575.
86. Priestley, C.M., Williamson, E. M., Wafford, K. A., Sattelle, D. B., Thymol, a constituent of
thyme essential oil, is a positive allosteric modulator of human GABAA receptors and a homo-
oligomeric GABA receptor from Drosophila melanogaster. British Journal of Pharmacology,
2003. 140(8): p. 1363-1372.
87. Kessler, A., Sahin-Nadeem, H., Lummis, S. C., Weigel, I., Pischetsrieder, M., Buettner, A.,
Villmann, C., GABAA receptor modulation by terpenoids from Sideritis extracts. Mol Nutr
Food Res, 2014. 58(4): p. 851-862.
88. Ding, J., Huang, C., Peng, Z., Xie, Y., Deng, S., Nie, Y. Z., Xu, T. L., Ge, W. H., Li, W. G., Li, F.,
Electrophysiological characterization of methyleugenol: a novel agonist of GABA(A)
receptors. ACS Chem Neurosci, 2014. 5(9): p. 803-811.
89. Reiner, G.N., Delgado-Marin, L., Olguin, N., Sanchez-Redondo, S., Sanchez-Borzone, M.,
Rodriguez-Farre, E., Sunol, C., Garcia, D. A., GABAergic pharmacological activity of propofol

Literaturverzeichnis
132
related compounds as possible enhancers of general anesthetics and interaction with
membranes. Cell Biochem Biophys, 2013. 67(2): p. 515-525.
90. Zaugg, J., Eickmeier, E., Ebrahimi, S. N., Baburin, I., Hering, S., Hamburger, M., Positive GABAA
receptor modulators from Acorus calamus and structural analysis of (+)-dioxosarcoguaiacol
by 1D and 2D NMR and molecular modeling. J Nat Prod, 2011. 74(6): p. 1437-1443.
91. Zaugg, J., Ebrahimi, S. N., Smiesko, M., Baburin, I., Hering, S., Hamburger, M., Identification
of GABA A receptor modulators in Kadsura longipedunculata and assignment of absolute
configurations by quantum-chemical ECD calculations. Phytochemistry, 2011. 72(18): p.
2385-2395.
92. Zaugg, J., Baburin, I., Strommer, B., Kim, H. J., Hering, S., Hamburger, M., HPLC-Based Activity
Profiling: Discovery of Piperine as a Positive GABAA Receptor Modulator Targeting a
Benzodiazepine-Independent Binding Site. In J. Nat. Prod. , 2010. 73(2): p. 185–191.
93. Kim, H.J., Baburin, I., Khom, S., Hering, S.,Hamburger, M., HPLC-based activity profiling
approach for the discovery of GABAA receptor ligands using an automated two
microelectrode voltage clamp assay on Xenopus oocytes. Planta Med, 2008. 74(5): p. 521-
526.
94. Garcia, D.A., Bujons, J., Vale, C., Sunol, C., Allosteric positive interaction of thymol with the
GABAA receptor in primary cultures of mouse cortical neurons. Neuropharmacology, 2006.
50(1): p. 25-35.
95. Hossain, S.J., Aoshima, H., Koda, H., Kiso, Y., Fragrances in oolong tea that enhance the
response of GABAA receptors. Bioscience, Biotechnology, and Biochemistry, 2004. 68(9): p.
1842-1848.
96. Campbell, E.L., Chebib, M., Johnston, G. A. R., The dietary flavonoids apigenin and (-)-
epigallocatechin gallate enhance the positive modulation by diazepam of the activation by
GABA of recombinant GABAA receptors. Biochemical Pharmacology, 2004. 68(8): p. 1631-
1638.
97. Laube, B., Maksay, G., Schemm, R., Betz, H., Modulation of glycine receptor function:a novel
approach for therapeutic intervention at inhibitory synapses? Trends in Pharmacological
Sciences, 2002. 23(11): p. 519-527.
98. Wagner, C.A., Friedrich, B., Setiawan, I., Lang, F., Bröer, S., The Use of Xenopus laevis Oocytes
for the Functional Characterization of Heterologously Expressed Membrane Proteins. Cellular
Physiolgy and Biochemmistry 2000. 10: p. 1-12.
99. Baumgartner, W., Islas, L., Sigworth, F. J. , Two-Microelectrode Voltage Clamp of Xenopus
Oocytes: Voltage Errors and Compensation for Local Current Flow. Biophysical Journal 1999.
77 (1980–1991).

133
100. Woronuk, G., Demissie, Z., Rheault, M., Mahmoud, S., Biosynthesis and therapeutic
properties of Lavandula essential oil constituents. Planta Med, 2011. 77(1): p. 7-15.
101. Alnamer, R., Alaoui, K., Bouidida el, H., Benjouad, A., Cherrah, Y., Sedative and Hypnotic
Activities of the Methanolic and Aqueous Extracts of Lavandula officinalis from Morocco. Adv
Pharmacol Sci, 2012. 2012: p. 270824.
102. Shaw, D., Annett, J. M., Doherty, B., Leslie, J. C. , Anxiolytic effects of lavender oil inhalation
on open-field behaviour in rats. Phytomedicine, 2007. 14(9): p. 613-620.
103. Kessler, A., Villmann, C., Sahin-Nadeem, H., Pischetsrieder, M., Buettner, A., GABAA receptor
modulation by the volatile fractions of Sideritis species used as ‘Greek’ or ‘Turkish’ mountain
tea. Flavour and Fragrance Journal, 2012. 27(4): p. 297-303.
104. Schuwald, A.M., Nöldner, M., Wilmes, T., Klugbauer, N.,Leuner, K.,Muller, W. E., Lavender
oil-potent anxiolytic properties via modulating voltage dependent calcium channels. PLoS
One, 2013. 8(4): p. e59998.
105. Naderi, N., Akhavan, N., Aziz Ahari, F., Zamani, N., Kamalinejad, M., Shokrzadeh, M.,
Ahangar, N., Motamedi, F. , Effects of hydroalcoholic extract from Salvia verticillata on
pharmacological models of seizure, anxiety and depression in mice. Iranian Journal of
Pharmaceutical Research, 2011. 10(3): p. 535-545.
106. Imanshahidi, M., Hosseinzadeh, H., The pharmacological effects of Salvia species on the
central nervous system. Phytother Res, 2006. 20(6): p. 427-437.
107. Viola, H., Wasowski, C., Marder, M., Wolfman, C., Paladini, A. C., Medina J. H. , Sedative and
hypnotic properties of Salvia guaranitica St. Hil. and of its active principle, Cirsiliol.
Phytomedicine, 1977. 4(1): p. 47-51.
108. Khayate-Nouri, M.H., Namvaran-Abbasabad, A., Tavakkoli, F. , Salvia officinalis and cisplatin
effects on pentylenetetrazole induced seizure threshold. Zahedan Journal of Research in
Medical Sciences, 2013. 15(11): p. 1-3.
109. Emamghoreishi, M., Talebianpour, M. S. , Antidepressant effect of Melissa officinalis in the
forced swimming test. DARU Journal of Pharmaceutical Sciences, 2009. 17(1): p. 42-47.
110. Awad, R., Levac, D., Cybulska, P., Merali, Z., Trudeau, V. L., Arnason, J. T., Effects of
traditionally used anxiolytic botanicals on enzymes of the g-aminobutyric acid (GABA)
system. Can. J. Physiol. Pharmacol, 2007. 85(9): p. 933-942.
111. Ibarra, A., Feuillere, N., Roller, M., Lesburgere, E, Beracochea, D. , Effects of chronic
administration of Melissa officinalis L. extract on anxiety-like reactivity and on circadian and
exploratory activities in mice. Phytomedicine, 2010. 17(6): p. 397-403.

Literaturverzeichnis
134
112. Gonzalez-Burgos, E., Carretero, M. E., Gomez-Serranillos, M. P. , Sideritis spp.: Uses, chemical
composition and pharmacological activities - A review. Journal of Ethnopharmacology, 2011.
135 (2): p. 209-225.
113. Hernandez-Perez, M., Rabanal, R. M. , Evaluation of the antinflammatory and analgesic
activity of Sideritis canariensis var. pannosa in mice. Journal of Ethnopharmacology, 2002.
81: p. 43-47.
114. Hernandez-Perez, M., Sanchez-Mateo, C. C., Montalbetti-Moreno, Y., Rabanal, R. M., Studies
on the analgesic and anti-inflammatory effects of Sideritis candicans Ait. var. eriocephala
Webb aerial part. Journal of Ethnopharmacology, 2004. 93(2-3): p. 279-284.
115. Öztürk, Y., Aydin, S., Öztürk, N., Başer, K. H. C.,, Effects of extracts from certain Sideritis
species on swimming performance in mice. Phytotherapy Research, 1996. 10(1): p. 70-73.
116. Vasilopoulou, C.G., Kontogianni, V. G., Linardaki, Z. I., Iatrou, G., Lamari, F. N., Nerantzaki, A.
A., Gerothanassis, I. P., Tzakos, A. G., Margarity, M., Phytochemical composition of
"mountain tea" from Sideritis clandestina subsp. clandestina and evaluation of its behavioral
and oxidant/antioxidant effects on adult mice. Eur J Nutr, 2013. 52(1): p. 107-116.
117. Adachi, N., Tomonaga, S., Tachibana, T., Denbow, D. M., Furuse, M., (-)-Epigallocatechin
gallate attenuates acute stress responses through GABAergic system in the brain. European
Journal of Pharmacology, 2006. 531(1-3): p. 171-175.
118. Vignes, M., Maurice, T., Lante, F., Nedjar, M., Thethi, K., Guiramand, J., Recasens, M.,
Anxiolytic properties of green tea polyphenol (-)-epigallocatechin gallate (EGCG). Brain
Research, 2006. 1110(1): p. 102-115.
119. Higashiyama, A., Htay, H. H., Ozeki, M., Juneja, L. R., Kapoor, M. P., Effects of L-theanine on
attention and reaction time response. Journal of Functional Foods, 2011. 3(3): p. 171-178.
120. Awad, R., Ahmed, F., Bourbonnais-Spear, N., Mullally, M., Ta, C. A., Tang, A., Merali, Z.,
Maquin, P., Caal, F., Cal, V., Poveda, L., Vindas, P. S., Trudeau, V. L., Arnason, J. T.,
Ethnopharmacology of Q’eqchi’ Maya antiepileptic and anxiolytic plants: Effects on the
GABAergic system. Journal of Ethnopharmacology, 2009. 125(2): p. 257-264.
121. Hritcu, L., Noumedem, J. A., Cioanca, O., Hancianu, M., Postu, P., Mihasan, M., Anxiolytic and
antidepressant profile of the methanolic extract of Piper nigrum fruits in beta-amyloid (1-42)
rat model of Alzheimer's disease. Behavioral and Brain Functions, 2015. 11: p. 13.
122. Rex, A., Morgenstern, E., Fink, H., Anxiolytic like effects of Kava-Kava in the elevated plus
maze test a comparison with diazepam. Progress in Neuro-Psychopharmacology & Biological
Psychiatry 2002. 26 p. 855–860.
123. Srivastava, J.K., Shankar, E., Gupta, S., Chamomile: A herbal medicine of the past with bright
future. Mol Med Rep, 2010. 3(6): p. 895-901.

135
124. Avallone, R., Zanoli, P., Puia, G., Kleinschnitz, M., Schreier, P., Baraldi, M. , Pharmacological
profile of apigenin, a flavonoid isolated from Matricaria chamomilla. Biochemical
Pharmacology, 2000. 59: p. 1387-1394.
125. Amsterdam, J.D., Shults, J., Soeller, I., Mao, J. J., Rockwell, K., Newberg, A. B., Chamomile
(Matricaria recutita) May Have Antidepressant Activity in Anxious Depressed Humans - An
Exploratory Study. Altern Ther Health Med, 2012. 18 (5): p. 44–49.
126. Halder, S., Mehta, A. K., Mediratta, P. K., Sharma, K. K. , Acute effect of essential oil of
Eugenia caryophyllata on cognition and pain in mice. Naunyn-Schmiedeberg's Archives of
Pharmacology, 2012. 385: p. 587-593.
127. Hosseini, M., Kamkar-Asl, M., Rakhshandeh, H. , Analgesic effect of clove essential oil in mice.
Avicenna Journal of Phytomedicine, 2011. 1(1): p. 1-6.
128. Alqareer, A., Alyahya, A., Andersson, L., The effect of clove and benzocaine versus placebo as
topical anesthetics. Journal of Dentistry, 2006. 34(10): p. 747-750.
129. Galal, A.A.A., Abdellatief, S. A., Neuropharmacological studies on Syzygium aromaticum
(clove) essential oil. International Journal of Pharma Sciences, 2015. 5(2): p. 1013-1018.
130. Lee, K.G., Shibamoto, T. , Antioxidant property of aroma extract isolated from clove buds
[Syzygium aromaticum (L.) Merr. et Perry]. Food Chemistry 2001. 74 (4): p. 443–448.
131. Pino, J.A., Marbot, R., Agüero, J., Fuentes, V., Essential Oil from Buds and Leaves of Clove
(Syzygium aromaticum (L.) Merr. et Perry) Grown in Cuba. Journal of essential oil research,
2001. 13(4): p. 278-279.
132. Chaieb, K., Hajlaoui, H., Zmantar, T., Kahla-Nakbi, A. B., Rouabhia, M., Mahdouani, K.,
Bakhrouf, A., The chemical composition and biological activity of clove essential oil, Eugenia
caryophyllata (Syzigium aromaticum L. Myrtaceae): a short review. Phytotheraphy Research,
2007. 21(6): p. 501-506.
133. Bhuiyan, M.N.I., Begum, J., Nandi, N. C., Akter, F. , Constituents of the essential oil from
leaves and buds of clove (Syzigium caryophyllatum (L.) Alston). African Journal of Plant
Science, 2010. 4(11): p. 451–454.
134. Rahimi, A.A., Ashnagar, A., Hamideh, N. , Isolation and characterization of 4-allyl-
2methoxyphenol (eugenol) from clove buds marketed in Tehran city of Iran. International
Journal of ChemTech Research 2012. 4(1): p. 105-108.
135. Mittal, M., Gupta, N., Parashar, P., Mehra, V., Khatri, M., Phytochemical evaluation and
pharmacological activity of Syzygium aromaticum: a comprehensive review. International
Journal of Pharmacy and Pharmaceutical Sciences 2014. 6(8): p. 67-72.
136. Razafimamonjison, G., Jahiel, M., Duclos, T., Ramanoelina, P., Fawbush, F., Danthu, P., Bud,
leaf and stem essential oil composition of Syzygium aromaticum from Madagascar,

Literaturverzeichnis
136
Indonesia and Zanzibar. International Journal of Basic and Applied Sciences, 2014. 3(3): p.
224-233.
137. Park, C.K., Li, H. Y., Yeon, K. Y., Jung, S. J., Choi, S. Y., Lee, S. J., Lee, S., Park, K., Kim, J. S., Oh,
S. B., Eugenol Inhibits Sodium Currents in Dental Afferent Neurons. Journal of Dental
Research, 2006. 85(10): p. 900-904.
138. Lee, M.H., Yeon, K. Y., Park, C. K., Li, H. Y., Fang, Z., Kim, M. S., Choi, S. Y., Lee, S. J., Lee, S.,
Park, K., Lee, J. H., Kim, J. S., Oh, S. B., Eugenol Inhibits Calcium Currents in Dental Afferent
Neurons. Journal of Dental Research 2005. 84(9): p. 848–851.
139. Chung, G., Rhee, J. N., Jung, S. J., Kim, J. S., Oh, S. B., Modulation of CaV2.3 Calcium Channel
Currents by Eugenol. Journal of Dental Research, 2008. 87(2): p. 137-141.
140. Guenetta, S.A., Beaudry, F., Marier, J. F., Vachon, P. , Pharmacokinetics and anesthetic
activity of eugenol in male Sprague-Dawleys rats. J.vet. Pharmacol. Therap. , 2006. 29(4): p.
265–270.
141. Goulet, F., Hélie, P., Vachon, P., Eugenol Anesthesia in African Clawed Frogs (Xenopus laevis)
of Different Body Weights. Journal of the American Association for Laboratory Animal
Science 2010. 49 (4): p. 460–463.
142. Sharma, M., Rauniar, G. P., Das, B. P. , Experimental study of various central nervous system
effects of eugenol in mice and rats. Health Renaissance, 2012. 10(3): p. 208–214.
143. Lionnet, L., Beaudry, F., Vachon, P., Intrathecal eugenol administration alleviates
neuropathic pain in male Sprague-Dawley rats. Phytotherapy Research, 2010. 24(11): p.
1645-1653.
144. Franco, L., Sanchez, C., Bravo, R., Rodriguez, A., Barriga, C., Juanez, J. C., The sedative effects
of hops (Humulus lupulus), a component of beer, on the activity/rest rhythm. Acta Physiol
Hung, 2012. 99(2): p. 133-139.
145. Zanoli, P., Zavatti, M., Rivasi, M., Brusiani, F., Losi, G., Puia, G., Avallone, R., Baraldi, M.,
Evidence that the b-acids fraction of hops reduces central GABAergic neurotransmission.
Journal of Ethnopharmacology, 2007. 109(1): p. 87-92.
146. Franco, L., Sánchez, C., Bravo, R., Rodríguez, A. B., Barriga, C., Romero, E., Cubero, J. , The
Sedative Effect of Non-Alcoholic Beer in Healthy Female Nurses. PLoS One, 2012. 7(7): p.
e37290.
147. Aoshima, H., Takeda, K., Okita, Y., Hossain, S.J., Koda, H., Kiso, Y., Effects of beer and hop on
ionotropic g-aminobutyric acid receptors. J Agric Food Chem, 2006. 54(7): p. 2514-2519.
148. Messaoudi, M., Bisson, J. F., Nejdi, A., Rozan, P., Javelot, H., Antidepressant-like effects of a
cocoa polyphenolic extract in Wistar-Unilever rats. Nutr Neurosci, 2008. 11(6): p. 269-276.

137
149. Bisson, J.F., Nejdi, A., Rozan, P., Hidalgo, S., Lalonde, R., Messaoudi, M., Effects of long-term
administration of a cocoa polyphenolic extract (Acticoa powder) on cognitive performances
in aged rats. British Journal of Nutrition 2008. 100(1): p. 94-101.
150. Nitz, D., Siegel, J. M. , GABA release in the locus coeruleus as a function of sleep/wake state.
Neuroscience, 1977. 78(3): p. 795–801.
151. Gottesmann, C., Brain inhibitory mechanisms involved in basic and higher integrated sleep
processes. Brain Res Brain Res Rev, 2004. 45(3): p. 230-249.
152. Streeter, C.C., Whitfield, T. H., Owen, L., Rein, T., Karri, S. K., Yakhkind, A., Perlmutter, R.,
Prescot, A., Renshaw, P. F., Ciraulo, D. A., Jensen, J. E., Effects of yoga versus walking on
mood, anxiety, and brain GABA levels: a randomized controlled MRS study. The Journal of
Alternative and Complementary Medicine, 2010. 16(11): p. 1145-1152.
153. Chang, L., Cloak, C. C., Ernst, T., Magnetic resonance spectroscopy studies of GABA in
neuropsychiatric disorders. The Journal of Clinical Psychiatry 2003. 64 (3): p. 7-14.
154. Wood, J.H., Hare, T. A., Glaeser, B. S., Ballenger, J. C., Post, R. M., Low cerebrospinal fluid γ-
aminobutyric acid content in seizure patients. Neurology, 1979. 29(9, Part 1): p. 1203-1208.
155. Sanacora, G., Mason, G. F., Rothman, D. L, Behar, K. L, Hyder, F., Petroff, O. A., Berman, R.
M., Charney, D. S., Krystal, J. H., Reduced cortical γ-aminobutyric acid levels in depressed
patients determined by proton magnetic resonance spectroscopy. Arch Gen Psychiatry, 1999
56(11): p. 1043-1047.
156. Rimón, R., Lepola, U., Jolkkonen, J., Halonen, T., Riekkinen, P., Cerebrospinal fluid gamma-
aminobutyric acid in patients with panic disorder. Biol Psychiatry 1995. 38(11): p. 737-741.
157. Knudsen, G.M., Schmidt, J., Almdal,T., Paulson, O. B., Vilstrup, H, Passage of Amino Acids and
Glucose Across the Blood-brain Barrier in Patients with Hepatic Encephalopathy. Hepatology,
1993. 17(6): p. 987-992.
158. van Gelder, N.M., Elliott, K. A. C. , Disposition of γ-Aminobutyric Acid Administered to
Mammals. Journal of Neurochemisrry, 1958. 3: p. 139-143.
159. Storici, P., De Biase, D., Bossa, F., Bruno, S., Mozzarelli, A., Peneff, C., Silverman, R. B.,
Schirmer, T., Structures of g-aminobutyric acid (GABA) aminotransferase, a pyridoxal 5'-
phosphate, and [2Fe-2S] cluster-containing enzyme, complexed with g-ethynyl-GABA and
with the antiepilepsy drug vigabatrin. J Biol Chem, 2004. 279(1): p. 363-373.
160. Schechter, P.J., Tranier, Y., Jung, M. J., Böhlen, P. , Audiogenic seizure protection by elevated
brain GABA concentration in mice: effects of γ-acetylenic GABA and γ-vinyl GABA, two
irreversible GABA-T inhibitors. European Journal of Pharmacology, 1977. 45 p. 319-328.
161. Ben-Menachem, E., Mechanism of action of vigabatrin: correcting misperceptions. Acta
Neurol Scand Suppl, 2011. 124(Suppl. 192): p. 5-15.

Literaturverzeichnis
138
162. Waterhouse, E.J., Mims, K. N., Gowda, S. N. , Treatment of refractory complex partial
seizures: role of vigabatrin. Neuropsychiatric Disease and Treatment, 2009. 5 p. 505–515.
163. Tsukatani, T., Higuchi, T., Matsumoto, K., Enzyme-based microtiter plate assay for γ-
aminobutyric acid: Application to the screening of γ-aminobutyric acid-producing lactic acid
bacteria. Analytica Chimica Acta, 2005. 540(2): p. 293-297.
164. Waksman, A., Rubinstein, M. K., Kuriyama, K., Roberts, E., Localization of γ-Aminobutyric-α-
Oxoglutaric Acid Transaminase In Mouse Brain. Journal of Neurochemistry, 1968. 15: p. 351–
357.
165. Hearl, W.G., Churchich, J. E. , Interactions between 4-Aminobutyrate Aminotransferase and
Succinic Semialdehyde Dehydrogenase, Two Mitochondrial Enzymes. Journal of Biological
Chemistry, 1984. 259(18): p. 11459-11463.
166. Lippert B., M., B. W., Jung, M. J., Casara, P., 4-Amino-hex-5-enoic Acid, a Selective Catalytic
Inhibitor of 4-Aminobutyric-Acid Aminotransferase in Mammalian Brain. Eur. J. Biochem.,
1977. 74: p. 441-445.
167. Sherif, F., Oreland, E., Effects of chronic treatment with the GABA-transaminase inhibitor
vigabatrin on exploratory behaviour in rats. Behavioural Brain Research 1994. 63 p. 11-15.
168. Le, H.V., Hawker, D. D., Wu, R., Doud, E., Widom, J., Sanishvili, R., Liu, D., Kelleher, N. L.,
Silverman, R. B., Design and Mechanism of Tetrahydrothiophene-Based g-Aminobutyric Acid
Aminotransferase Inactivators. J Am Chem Soc, 2015. 137(13): p. 4525-4533.
169. Pan, Y., Qiu, J., Silverman, R. B., Design, synthesis and biological activity of a difluoro-
substituted, conformationally-rigid vigabatrin analogue as a potent γ-aminobutyric acid
aminotransferase inhibitor. J. Med. Chem., 2003. 46: p. 5292-5293.
170. Silverman, S.C.a.R.B., Inactivation and Inhibition of γ-Aminobutyric Acid Aminotransferase by
Conformationally Restricted Vigabatrin Analogues. J. Med. Chem., 2002. 45: p. 4531-4539.
171. Tao, Y.H., Jiang, D.Y., Xu, H. B., Yang, X.L. , Inhibitory effect of Erigeron breviscapus extract
and its flavonoid components on GABA shunt enzymes. Phytomedicine, 2008. 15(1-2): p. 92-
97.
172. Awad, R., Muhammad, A., Durst, T., Trudeau, V. L., Arnason, J. T., Bioassay-guided
Fractionation of Lemon Balm (Melissa officinalis L.) using an In Vitro Measure of GABA
Transaminase Activity. Phytotherapy Research, 2009. 23(8): p. 1075–1081.
173. Tao, Y.H., Yuan, Z., Tang, X. Q., Xu, H. B., Yang, X. L., Inhibition of GABA shunt enzymes`
activity by 4-hydroxybenzaldehyde derivatives. Bioorganic & Medicinal Chemistry Letters
2006. 16: p. 592–595.
174. Jung, M.J., Lippert, B., Metcalf, B. W., Schechter, P. J., Böhlen, P., Sjoerdsma, A., The effect of
4-amino hex-5-ynoic acid (gamma-acetylenic GABA, gammma-ethynyl GABA) a catalytic

139
inhibitor of GABA transaminase, on brain GABA metabolism in vivo. J Neurochem, 1977
28(4): p. 717-723.
175. Qiu, J., Pingsterhaus, J. M., Silverman, R. B. , Inhibition and Substrate Activity of
Conformationally Rigid Vigabatrin Analogues with γ-Aminobutyric Acid Aminotransferase. J.
Med. Chem., 1999. 42 p. 4725-4728.
176. Coelho, V.R., Vieira, C. G., De Souza, L. P., Moyses, F., Basso, C., Papke, D. K., Pires, T. R.,
Siqueira, I. R., Picada, J. N., Pereira, P., Antiepileptogenic, antioxidant and genotoxic
evaluation of rosmarinic acid and its metabolite caffeic acid in mice. Life Sciences, 2015. 122:
p. 65-71.
177. Hemming, K., Maguire, M. J., Hutton, J. L., Marson, A. G., Vigabatrin for refractory partial
epilepsy. Cochrane Database Syst Rev, 2013. 1: p. CD007302.
178. Pan, Y., Gerasimov, M. R., Kvist, T., Wellendorph, P., Madsen, K. K., Pera, E., Lee, H.,
Schousboe, A., Chebib, M., Brauner-Osborne, H.,Craft, C. M., Brodie, J. D., Schiffer, W. K.,
Dewey, S. L., Miller, S. R., Silverman, R. B., (1S, 3S)-3-amino-4-difluoromethylenyl-1-
cyclopentanoic acid (CPP-115), a potent g-aminobutyric acid aminotransferase inactivator
for the treatment of cocaine addiction. J Med Chem, 2012. 55(1): p. 357-366.
179. Miller, N.R., Johnson, M. A., Paul, S. R., Girkin, C. A., Perry, J. D., Endres, M., Krauss, G. L. ,
Visual dysfunction in patients receiving vigabatrin : Clinical and electrophysiologic findings.
Neurology, 1999. 53(9): p. 2082-2087.
180. Wang, H., Provan, G. J., Helliwell, K., Determination of rosmarinic acid and caffeic acid in
aromatic herbs by HPLC. Food Chemistry, 2004. 87(2): p. 307-311.
181. Teruhisa Komori, T.M., Eishi Motomura and Takashi Shiroyama, The Sleep-Enhancing Effect
of Valerian Inhalation and Sleep-Shortening Effect of Lemon Inhalation. Chem. Senses, 2006.
31: p. 731-737.
182. Koo, B.S., Park, K. S., Ha, J. H., Park, J. H., Lim, J. C., Lee, D. U., Inhibitory Effects of the
Fragrance Inhalation of Essential Oil from Acorus gramineus on Central Nervous System. Biol.
Pharm. Bull., 2003. 26(7): p. 978—982.
183. Koo, B.S., Lee, S. I., Ha, J. H., Lee, D. U., Inhibitory Effects of the Essential Oil from SuHeXiang
Wan on the Central Nervous System after Inhalation. Biol. Pharm. Bull. , 2004. 27(4): p. 515-
519.
184. Waye, A., Annal, M., Tang, A., Picard, G., Harnois, F., Guerrero-Analco, J. A., Saleem, A.,
Hewitt, L. M., Milestone, C. B., MacLatchy, D. L., Trudeau, V. L., Arnason, J. T., Canadian
boreal pulp and paper feedstocks contain neuroactive substances that interact in vitro with
GABA and dopaminergic systems in the brain. Science of the Total Environment 2014. 468–
469: p. 315–325.

Literaturverzeichnis
140
185. Gottesmann, C., GABA Mechanisms and Sleep. Neuroscience, 2002. 111(2): p. 231-239.
186. Wang, H.F., Tsai, Y. S., Lin, M. L., Ou, A. S., Comparison of bioactive components in GABA tea
and green tea produced in Taiwan. Food Chemistry, 2006. 96(4): p. 648-653.
187. Syu, K.Y., Lin, C. L., Huang, H. C., Lin, J. K. , Determination of theanine, GABA, and other
amino acids in green, oolong, black, and Pu-erh teas with dabsylation and high-performance
liquid chromatography. Journal of Agricultural and Food Chemistry, 2008. 56(17): p. 7637-
7643.
188. Su, Y.S., Lin, Y. P., Cheng, F. C., Jen, J. F., In-capillary derivatization and stacking
electrophoretic analysis of g-aminobutyric acid and alanine in tea samples to redeem the
detection after dilution to decrease matrix interference. J Agric Food Chem, 2010. 58(1): p.
120-126.
189. Zhao, M., Ma, Y., Wei, Z. Z., Yuan, W. X., Li, Y. L., Zhang, C. H., Xue, X. T., Zhou, H. J.,
Determination and comparison of g-aminobutyric acid (GABA) content in pu-erh and other
types of Chinese tea. J Agric Food Chem, 2011. 59(8): p. 3641-3648.
190. Limon, A., Gallegos-Perez, J. L., Reyes-Ruiz, J. M., Aljohi, M. A., Alshanqeeti, A. S., Miledi, R.,
The endogenous GABA bioactivity of camel, bovine, goat and human milks. Food Chemistry,
2014. 145: p. 481-487.
191. Kim, H.S., Lee, E. J., Lim, S. T.,Han, J. A., Self-enhancement of GABA in rice bran using various
stress treatments. Food Chem, 2015. 172: p. 657-662.
192. Peñas, E., Limón, R. I., Martínez-Villaluenga, C., Restani, P., Pihlanto, A., Frias, J., Impact of
elicitation on antioxidant and potential antihypertensive properties of lentil sprouts. Plant
Foods Hum Nutr, 2015. 70: p. 401–407.
193. Peñas, E., Diana, M., Frias, J., Quílez, J., Martínez-Villaluenga, C., A multistrategic approach in
the development of sourdough bread targeted towards blood pressure reduction. Plant
Foods Hum Nutr 2015. 70: p. 97–103.
194. Hossain, S.J., Kato, H., Aoshima, H., Yokoyama, T., Yamada, M., Hara, Y., Polyphenol-Induced
Inhibition of the Response of Na+/Glucose Cotransporter Expressed in Xenopus Oocytes. J.
Agric. Food Chem., 2002. 50(18): p. 5215–5219.
195. Khuhawar, M.Y.R., A. D., Liquid chromatographic determination of γ-aminobutyric acid in
cerebrospinal fluid using 2-hydroxynaphthaldehyde as derivatizing reagent. Journal of
Chromatography B, 2003. 788(2): p. 413-418.
196. Jacob, T.C., Moss, S. J., Jurd, R., GABAA receptor trafficking and its role in the dynamic
modulation of neuronal inhibition. Nature Reviews Neuroscience, 2008. 9: p. 331-343.
197. Aoshima, H., Okita, Y., Hossain, S. J., Fukue, K., Mito, M., Orihara, Y., Yokoyama, T., Yamada,
M., Kumagai, A., Nagaoka, Y., Uesato, S., Hara, Y., Effect of 3-O-Octanoyl-(+)-catechin on the

141
Responses of GABAA Receptors and Na+/Glucose Cotransporters Expressed in Xenopus
Oocytes and on the Oocyte Membrane Potential. Journal of Agricultural and Food Chemistry,
2005. 53(6): p. 1955–1959.
198. Thompson, S.A., Wheat, L., Brown, N. A., Wingrove, P. B., Pillai, G. V., Whiting, P. J., Adkins,
C., Woodward, C. H., Smith, A. J., Simpson, P. B., Collins, I., Wafford, K. A., Salicylidene
salicylhydrazide, a selective inhibitor of b1-containing GABAA receptors. British Journal of
Pharmacology, 2004. 142(1): p. 97-106.
199. Zanoli, P., Avallone, R., Baraldi, M. , Behavioral characterisation of the flavonoids apigenin
and chrysin. Fitoterapia 2000. 71: p. S117 S123.
200. Diana, M., Quílez, J., Rafecas, M., Gamma-aminobutyric acid as a bioactive compound in
foods: a review. Journal of Functional Foods, 2014. 10: p. 407-420.
201. Nakamura, H., Takishima, T., Kometani, T., Yokogoshi, H., Psychological stress-reducing effect
of chocolate enriched with g-aminobutyric acid (GABA) in humans: assessment of stress using
heart rate variability and salivary chromogranin A. International Journal of Food Sciences
and Nutrition, 2009. 60 (Suppl 5): p. 106-113.
202. Wu, C., Huang, Y., Lai, X., Lai, R., Zhao, W., Zhang, M., Zhao, W., Study on quality components
and sleep-promoting effect of GABA Maoyecha tea. Journal of Functional Foods, 2014. 7: p.
180-190.
203. Hall, A.C., Turcotte, C. M., Betts, B. A., Yeung, W. Y., Agyeman, A. S., Burk, L. A., Modulation
of human GABAA and glycine receptor currents by menthol and related monoterpenoids.
European Journal of Pharmacology, 2004. 506(1): p. 9-16.
204. Mascia, M.P., Wick, M. J., Martinez, L. D., Harris, R. A. , Enhancement of glycine receptor
function by ethanol: role of phosphorylation. British Journal of Pharmacology, 1998 125: p.
263-270.
205. Liang, Z.H., Cheng, X. H., Ruan, Z. G., Wang, H., Li, S. S., Liu, J., Li, G. Y., Tian, S. M., Protective
effects of components of the Chinese herb grassleaf sweetflag rhizome on PC12 cells
incubated with amyloid-beta42. Neural Regen Res, 2015. 10(8): p. 1292-1297.
206. Lee, S.H., Moon, J. Y., Jung, S. J., Kang, J. G., Choi, S. P., Jang, J. H., Eugenol inhibits the GABAA
current in trigeminal ganglion neurons. PLoS One, 2015. 10(1): p. 1-12.
207. Wang, L.H., Sung, W. C., Rapid evaluation and quantitative analysis of eugenol derivatives in
essential oils and cosmetic formulations on human skin using attenuated total reflectance–
infrared spectroscopy. Spectroscopy, 2011. 26(1): p. 43-52.
208. Da Silva, R.R., Da Câmara, C. A. G., Almeida, A. V., Ramos, C. S. , Biotic and Abiotic Stress-
Induced Phenylpropanoids in Leaves of the Mango (Mangifera indica L., Anacardiaceae). J.
Braz. Chem. Soc., 2012. 23(2): p. 206–211.

Literaturverzeichnis
142
209. Müller, M., Page, H. C., Speckmann, E. J., Gorji, A., Effect of eugenol on spreading depression
and epileptiform discharges in rat neocortical and hippocampal tissues. Neuroscience, 2006.
140(2): p. 743-751.
210. Irie, Y., Itokazu, N., Anjiki, N., Ishige, A., Watanabe, K., Keung, W. M., Eugenol exhibits
antidepressant-like activity in mice and induces expression of metallothionein-III in the
hippocampus. Brain Res, 2004. 1011(2): p. 243-246.
211. Chuang, C.Y., Shi, Y. C., You, H. P., Lo, Y. H., Pan, T. M., Antidepressant effect of GABA-rich
Monascus-fermented product on forced swimming rat model. J Agric Food Chem, 2011.
59(7): p. 3027-3034.