mk-801–induced expression of fos protein family members in the rat retrosplenial granular cortex
TRANSCRIPT

MK-801–Induced Expression of Fos ProteinFamily Members in the Rat RetrosplenialGranular CortexX. Zhang,* X.-D. Fan, P. Mohapel, P.H. Yu, and A.A. BoultonNeuropsychiatry Research Unit, Department of Psychiatry, University of Saskatchewan, Saskatoon, Canada
The N-methyl-D-aspartate (NMDA) receptor antago-nist MK-801 has been shown to induce an acute Fosand Fos-related antigen (Fra) expression in the ratretrosplenial granular cortex (RSG), but the exactcomposition of the Fos protein family and theirindividual dynamic alterations are unknown. Weexamined this issue using immunocytochemistry andWestern blot analysis with two antibodies that recog-nize, respectively, Fos and all the identified membersof Fos protein family. Immunocytochemistry detecteda rapid and transient expression of Fos proteins insome RSG neurons and a delayed and prolongedexpression of Fra proteins in most RSG neuronsfollowing a single systemic MK-801 injection (1 mg/kg). Multiple MK-801 injections (i.e., ten consecutiveinjections once every other day; 1 mg/kg) produced amoderate Fra expression but failed to induce detect-able Fos expression. Western blot analysis furthershowed a transient expression of 72-kDa Fos proteinsfollowing a single MK-801 injection and a prolongedexpression of 46- and 43-kDa Fra proteins after eithera single or multiple MK-801 administration. Thedelayed onset and prolonged expression of these Fraproteins suggest that they may beDFosB. The possiblerelevance of these results to clinical psychotomimeticeffects of the NMDA receptor antagonists phencycli-dine and ketamine is discussed. J. Neurosci. Res.57:719–729, 1999. r 1999 Wiley-Liss, Inc.
Key words: Fos-related antigen; FosB; NMDA recep-tor; phencyclidine; schizophrenia
INTRODUCTIONTranscription products of thefosfamily of immedi-
ate-early genes consists of Fos and several identifiedFos-related antigen (Fra) proteins including Fra-1, Fra-2,and FosB/DFosB. It has been demonstrated that variousbrain stimuli can induce expression of an array ofdifferent fosproducts that vary in their time of onset andduration. In general, Fos and FosB proteins are inducedrapidly and transiently in the rat brain, whereas the
DFosB is induced more slowly and has a protractedexistence (Sonnenberg et al., 1989; Pennypacker et al.,1994, 1995; Doucet et al., 1996; Mandelzys et al., 1997;Hiroi et al., 1997, 1998). Whereas acute Fos expressionplays an important role in linking transient changes ofneuronal activity to both short- and long-term modifica-tions of structure and function in the mammalian brain(Heilig et al., 1993; Cirelli et al., 1995; Wollnik et al.,1995; Watanabe et al., 1996), the prolonged expression ofDFosB has been linked to molecular, electrophysiologi-cal, and behavioral adaptations to various brain insults(Hiroi et al., 1997, 1998; Mandelzys et al., 1997).
Recently, the non-competitiveN-methyl-D-aspar-tate (NMDA) receptor antagonist MK-801 has also beenshown to induce an acute expression of Fos and Fraproteins in the rat cerebral cortex, with prominent expres-sion in the retrosplenial granular cortex (RSG) (Sharp etal., 1991; Gass et al., 1993a,b; Hughes et al., 1993;Wedzony and Czyrak, 1996; Gao et al., 1998). However,it is not known what exactfosproducts are induced in therat RSG and what dynamic alterations occur for theindividual Fos protein family member following MK-801injection. This MK-801 phenomenom is of particularinterest because of its potential significance for theunderstanding of phencyclidine (PCP, ‘‘angel dust’’) andketamine-induced psychosis in humans. Both PCP andketamine are non-competitive NMDA receptor antago-nists that have been found to produce positive, negative,and cognitive schizophrenic-like symptoms in humans(Luby et al., 1959; Rosenbaum et al., 1959; Allen andYoung, 1978; Carlin et al., 1979; Pearlson, 1981; Younget al., 1987; Cosgrove and Newell, 1991; Javitt andZukin, 1991; Krystal et al., 1994; Malhotra et al., 1996,1997). It has been proposed that PCP- and ketamine-
Contract grant sponsor: Saskatchewan Health.
*Correspondence to: Dr. X. Zhang, Neuropsychiatry Research Unit,A114 Medical Research Bld., 103 Wiggins Road, University ofSaskatchewan, Saskatoon, SK, Canada S7N 5E4.E-mail: [email protected]
Received 13 January 1999; Revised 10 May 1999; Accepted 12 May1999
Journal of Neuroscience Research 57:719–729 (1999)
r 1999 Wiley-Liss, Inc.

induced psychosis is exclusively mediated through theirantagonistic effects on NMDA receptors (Javitt andZukin, 1991; Krystal et al., 1994; Malhotra et al., 1996,1997).
The aim of the present study was to plot the timecourse of the expression of Fos and different Fra proteinsin the rat RSG following a single or multiple systemicinjections of MK-801. Fos and Fra protein expressionwas detected using the narrow spectrum Fos and broad-spectrum Fos/Fra antibodies that recognize, respectively,Fos and all the identified members of the Fos proteinfamily. Quantitative Western blot analysis was under-taken, which was accompanied by cell counting of theRSG neurons stained with Fos and Fos/Fra immunocyto-chemistry and cresyl violet in order to provide semi-quantitative data about the percentage of Fos/Fra-immunoreactive neurons in the RSG. We focused on theRSG because it is an important nodal structure for thetransfer of information between those brain structures,i.e., the hippocampus and many areas of the neocortex(Wyss and Van Groen, 1992; Zilles and Wree, 1995),which have been reported to be associated with schizophre-nia (Bogerts, 1993; Port and Seybold, 1995; Bachus andKleinman, 1996; Velakoulis and Pantelis, 1996; O’Donnelland Grace, 1998). By characterizing the dynamic alter-ations of Fos and Fra expression in the RSG followingMK-801 injection, we hope to provide a rational forunderstanding how the Fos protein family may contributeto PCP- and ketamine-induced psychosis in humans.
MATERIALS AND METHODSAnimals
Eighty-seven female Sprague-Dawley rats (250–320 g) were used, and they were housed under tempera-ture- and light-controlled conditions (12:12 hr light:darkcycle with lights on at 07:00 AM), with food and wateravailable ad libitum. All animal use procedures were instrict accordance with the guidelines established by theCanadian Council on Animal Care as approved by theUniversity of Saskatchewan Animal Care Committee.
Systemic Administration of MK-801Subcutaneous administration of a moderate dose of
MK-801 (dizocilpine maleate; Research BiochemicalsInc., Natick, MA), i.e. 1 mg/kg, was chosen in order toinduce a massive Fos and Fra expression in the rat RSG(Sharp et al., 1991; Hughes et al., 1993) without produc-ing irreversible neuronal injury or neuronal death in situ(Olney et al., 1989). Both single and multiple administra-tion schedules were used for MK-801 injection. A singleinjection of MK-801 was undertaken according to theprevious finding that acute brain insults can evokesequential and differential Fos and Fra expression in
particular brain regions (Sonnenberg et al., 1989; Pen-nypacker et al., 1994, 1995; Hiroi et al., 1997, 1998). Themultiple injection study was designed to produce re-peated assaults by MK-801 on the rat brain, so that Fosand Fra expression could be further differentiated basedon the previous observation that multiple or chronicstimulations cause a protracted expression of Fra proteins(Hiroi et al., 1998), but no sustained increases in Fos,which undergoes a refractory period after repeated stimu-lations (Morgan and Curran, 1991).
For immunocytochemistry, seven groups of threerats each were allowed to survive for 1 hr, 4 hr, 12 hr, 1day, 2 days, 4 days, and 1 week after a single MK-801injection; two control groups of three rats each wereinjected with equivalent volume of vehicle (physiologicalsaline) and killed at 4 hr and 1 day. Ten consecutiveinjections of MK-801 were given every 2nd day to threegroups of three rats each at 4-hr, 1-day, and 1-weeksurvival periods following the last injection; three vehicle-injected control rats were killed at day 1. For any one ofthe four Western blot studies, four groups of three ratseach were killed 4 hr and 1 day after a single MK-801, 4hr after a single vehicle injection, or 1 day following thetenth consecutive injection of MK-801.
ImmunocytochemistryRats were anesthetized with sodium pentobarbital
(50 mg/kg, i.p.) and perfused via the ascending aorta with150 ml of 0.1 M phosphate-buffered saline (PBS, pH 7.4),followed by 200 ml of freshly prepared 4% paraformalde-hyde in PBS. The rat brains were then immediatelyremoved, postfixed for 2 hr in the same fixative, andcryoprotected in 30% sucrose at 4°C for 1–2 days. Seriesof frozen coronal sections (30 µm) through the wholeretrosplenial cortex were cut on a cryostat and collectedin PBS.
Two antibodies were used in this study. The first one(Santa Cruz Biotechnology Inc., Santa Cruz, CA) wasraised in rabbit and directed against a synthetic peptidecorresponding to amino acids 3–16 mapping at the aminoterminus of human c-Fos p62. This narrow-spectrum Fosantibody displays reactivity only with Fos proteins with-out cross-reactivity to other known Fra proteins, ascharacterized by the manufacturer. We will refer to this asthe ‘‘Fos antibody.’’ The second antibody (a gift from M.Iadarola, NIH, Bethesda, MD) was raised in rabbit and iscapable of recognizing all the identified members of Fosprotein family in the rat brain (Young et al., 1991;Pennypacker et al., 1994; Doucet et al., 1996; Hiroi et al.,1998; M. Iadarola, personal communication). This broad-spectrum Fos antibody, referred to as the ‘‘Fos/Fraantibody,’’ was produced by conjugating the syntheticpeptide (Lys-Val-Glu-Gln-Leu-Ser-Pro-Glu-Glu-Glu-Glu-Lys-Arg-Arg-Ile-Arg-Arg-Glu-Arg-A sn-Lys-Met-Ala-Ala-Ala) to keyhole limpet hemocyanin that had been
720 Zhang et al.

exhaustively reacted with succinic anhydride as previ-ously specified (Young et al., 1991).
For rats receiving a single MK-801 or vehicleinjection, three series of adjacent sections were obtained,with the first and second series being stained with the Fosand Fos/Fra antibodies, and the third series stained withcresyl violet. For rats receiving multiple MK-801 injec-tions two series of sections were obtained, one for theFos/Fra antibody and the other for cresyl violet staining.Sections stained with cresyl violet were used to obtain thetotal neuronal counts for both layers III and IV in theRSG. For control purposes, some sections from MK-801–injected rats were processed in the same way but withoutthe first antibody in order to test for non-specific labeling.The designation and terminology to define the rat RSGcorrespond to Paxinos and Watson’s (1986) atlas of therat brain and its companion book (Zilles and Wree, 1995).
Analysis of Fos and Fra immunoreactivity wasperformed on free-floating sections using a conventionalavidin-biotin-immunoperoxidase technique as previouslydescribed in detail (Zhang et al., 1996, 1997a,b). Briefly,this procedure included pretreating sections for 30 mineach in 0.2% hydrogen peroxide and in 5% bovine serumalbumin. Sections were then incubated overnight with theprimary antiserum at dilutions of 1:1,000 (the Fos anti-body) and 1:5,000 (the Fos/Fra antibody) at room tempera-ture. The primary antibodies were localized using Vec-tastain Elite reagents (Vector Laboratories, Burlingame,CA). The reaction product was developed using a modi-fied nickel-enhanced diaminobenzidine (DAB) method(Shu et al., 1988), i.e., incubating the sections in 30 ml of2.5% ammonium nickel sulphate, 0.05% DAB, and 2 µlof 30% hydrogen peroxide for 5 min at room temperature.
Western Blot AnalysisRats were killed by decapitation at various time
points, and the brains were rapidly removed for dissectionof the bilateral RSG. As shown in Figure 1, the dissectedbrain tissue included the major portion of the RSG andthe underlying white matter, the corpus callosum. Owingto the difficulty in distinguishing the RSG from theretrosplenial agranular cortex (RSA), we suspect that asmall portion of the RSA was also included. The dis-sected tissues were stored at270°C until use.
Western blot analysis was performed using themodified procedure of Sonnenberg et al. (1989). Thetissues were homogenized in 4 vol (w/v) of a buffercontaining 0.25 M sucrose, 15 mM Tris-HCl (pH 7.9), 60mM KCl, 15 mM NaCl, 5 mM EDTA, 1 mM EGTA, andthe following protease inhibitors: 1 mM DTT, 0.1 mMphenylmethylsulfonyl fluoride (PMSF), 2 µg/ml leupep-tin, and 5 µg/ml aprotinin. Cells were collected bycentrifugation at 2,000g for 10 min and resuspended in 4vol of a buffer containing 10 mM HEPES (pH 7.9), 1.5mM MgCl2, 10 mM KCl, and the protease inhibitors.
After centrifugation at 4,000g for 10 min, the nuclei pelletwas resuspended in a buffer containing 50 mM HEPES(pH 7.9), 0.75 mM MgCl2, 0.5 mM EDTA, 0.5 M KCl,12.5% glycerol, and the protease inhibitors. Following
Fig. 1. Schematic illustration of the dissection cuts used toisolate the rat retrosplenial granular cortex (RSG).A and Bshow the medial and frontal views of the right half of the ratbrain, respectively. Four sequential cuts were used to dissect theRSG, which are represented by the four sequentially numberedsolid lines: (1) The brain was vertically cut through the anteriorhippocampus at about 7.0 mm before the caudal edge of thecerebral cortex; (2) the posterior part of the brain was thenvertically cut at about 2.0 mm before the caudal edge of thecerebral cortex; (3) the hippocampus (Hip) was pared off justbeneath the corpus callosum (cc); (4) the remaining tissue wasfinally cut between the RSG and the retrosplenial agranularcortex (RSA) from the inner top to the lateral base with a 45°angle to the horizontal plane.
MK-801–induced Fos Induction in Cortex 721

incubation on a rotator for 30 min, the mixture wascentrifugated at 14,000g for 30 min, and then thesupernatant was collected and stored at270°C. Theentire procedure was carried out at 4°C. Approximately100 µg of proteins from each sample were separated on10% sodium dodecyl sulfate (SDS)-polyacrylamide gels(Doucet et al., 1990) and then transferred onto polyvinyl-difluoride (PVDF) membranes (Bio-Rad [Richmond, CA]Trans-blot Transfer membrane 0.2 µm) (Towbin et al.,1979). After pre-blocking the non-specific binding sitesin PBS containing 5% skim milk for 30 min, blots wereincubated for 2 hr at room temperature in the Fos/Fraantibody (1:3,000). Labeling of the Fos and Fra proteinswas detected using anti-rabbit antiserum conjugated withhorseradish peroxidase (Amersham, IL; 1:1,000) for 1 hrat room temperature. The peroxidase activity on themembranes was revealed using the above nickel-enhanced DAB method.
Data AnalysisImmunocytochemistry. Sections were quanti-
fied and photographed using an Olympus light micro-scope with brightfield illumination. Quantitative assess-ment was conducted by visually counting the neurons inlayers III and IV of the bilateral RSG from sections at thecoronal plains around Bregma25.8 (Paxinos and Wat-son, 1986). Three sections (120 µm apart) were countedfrom each rat for either immunocytochemistry or Nisslstaining. All the immunostained and Nissl-stained sec-tions counted were treated identically and cells werecounted in a predefined area of RSG. Under constantillumination conditions, using a blue filter to mask mostof the faintly immunostained non-specific backgroundcells and some specific cells, all Fos and Fos/Fra-immunopositive neurons with definite cell nucleus con-tours were counted using a 203 objective lens. Averagecell counts were obtained by averaging counts over bothlayers III and IV of the RSG, over both hemispheres, andover three sections of either immunolabeled or Nissl-stained cells.
Western blot analysis. Relative differences inFos and Fra protein expression were determined bydensitometric measurements of the optical densities ofFos- and Fra-immunoreactive bands in the films photo-graphed from PVDF membranes using an image analysissystem with NIH software.
RESULTSCell Counts of Nissl-Stained Sections
The average total number of Nissl-stained neuronsin layers III and IV of the unilateral RSG per section was1,5476 162, which was not significantly different among
either of the MK-801–treated or vehicle-injected controlgroups of rats. We conclude, therefore, that no apparent orsignificant loss of neurons occurred with 1 mg/kg ofeither a single or multiple MK-801 injections. This resultis in agreement with the previous finding that either asingle or repeated systemic administration of 1 mg/kg, orless, of MK-801 produces a reversible neuronal injury(i.e., cytoplasm vacuolation within the initial 24 hr) butno apparent neuronal death in the RSG (Olney et al.,1989). We have used the total mean number of RSGneurons (1,5476 162) in all rats as a baseline estimationfor the comparisons of the percentage of Fos andFos/Fra-immunoreactive neurons in the proceedingsection.
ImmunocytochemistryAll rats within the same group exhibited a consis-
tent pattern and intensity of immunostaining in the RSGof both hemispheres. No specific cellular immunoreactiv-ity could be detected throughout the RSG in the vehicle-injected control rats with the narrow-spectrum Fos anti-body (Fig. 2A), whereas a relatively faint immunolabelingover a large number of cell nuclei in layer II and a smallnumber in layer V in the RSG could be observed with thebroad-spectrum Fos/Fra antibody (Fig. 2F). After a singleor multiple administration of MK-801, immunocytochemi-cal localization of both Fos and Fos/Fra antibodies inadjacent series of sections revealed a significant increaseof labeling, primarily, in layers III and IV, with moreimmunoreactive cells in the posterior two-thirds than theanterior one-third of the RSG. The specific Fos or Fos/Fraimmunolabeling was observed exclusively within the cellnuclei and at least the majority of these immunostainedcell nuclei likely belongs to neurons instead of glia cellsas judged from their relative size. A small number ofimmunolabeled cell nuclei were also found in the RSAwith the broad-spectrum Fos/Fra antibody, whereas thecorpus callosum beneath the RSG exhibited no detectablespecific immunostaining with either of the antibodies. Weargue, therefore, that inclusion of the corpus callosum anda small portion of the RSA in the brain tissues for Westernblot analysis would not significantly affect our opticaldensities of the immunostained bands of proteins.
Immunopositive cell nuclei in the RSG were notice-able as early as 1 hr after a single MK-801 injection (Fig.2B,G), reached maximal levels at 4 hr (Fig. 2C, H),declined from 12 hr to 1 day (Fig. 2D,I), and completelydisappeared by 2 days using the Fos antibody or by 4–7days using the Fos/Fra antibody. Whereas the rats killedat 1 hr after a single MK-801 injection showed a similardensity of immunopositive cells with both Fos andFos/Fra antibodies (Fig. 2B,G), rats killed at 4 hr or 1 dayexhibited a much higher density of the stained cells withthe broad-spectrum Fos/Fra antibody than with the narrow-
722 Zhang et al.
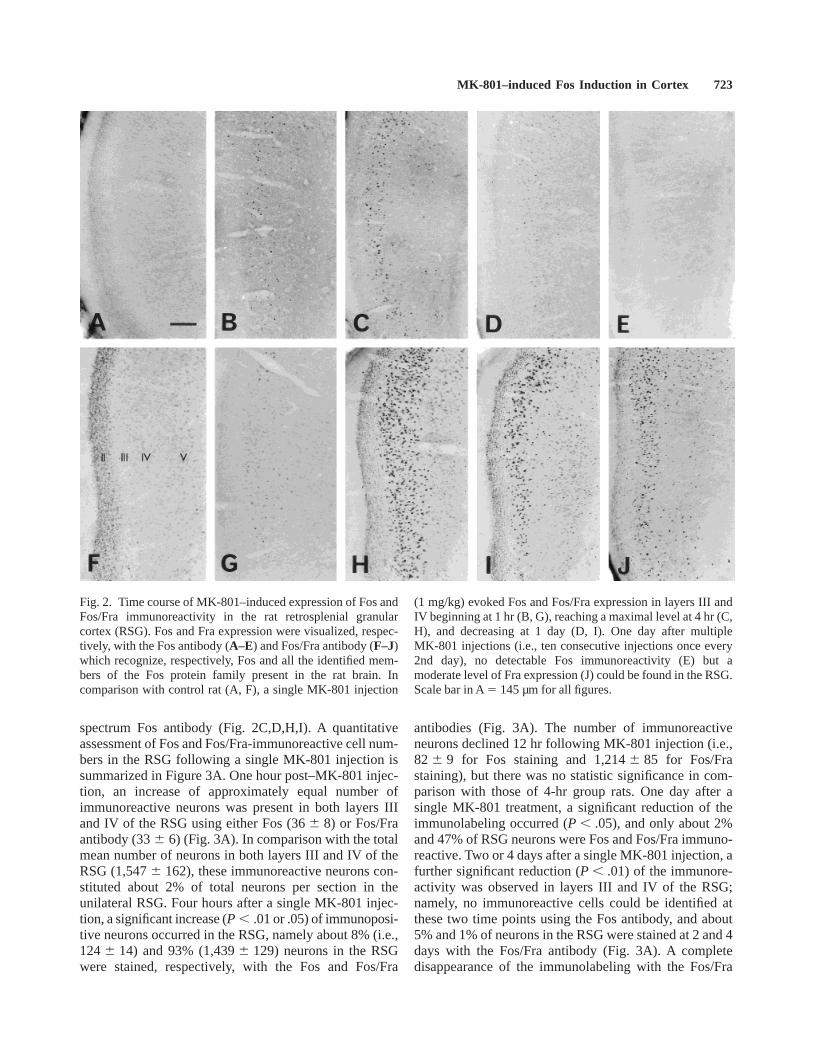
spectrum Fos antibody (Fig. 2C,D,H,I). A quantitativeassessment of Fos and Fos/Fra-immunoreactive cell num-bers in the RSG following a single MK-801 injection issummarized in Figure 3A. One hour post–MK-801 injec-tion, an increase of approximately equal number ofimmunoreactive neurons was present in both layers IIIand IV of the RSG using either Fos (366 8) or Fos/Fraantibody (336 6) (Fig. 3A). In comparison with the totalmean number of neurons in both layers III and IV of theRSG (1,5476 162), these immunoreactive neurons con-stituted about 2% of total neurons per section in theunilateral RSG. Four hours after a single MK-801 injec-tion, a significant increase (P , .01 or .05) of immunoposi-tive neurons occurred in the RSG, namely about 8% (i.e.,1246 14) and 93% (1,4396 129) neurons in the RSGwere stained, respectively, with the Fos and Fos/Fra
antibodies (Fig. 3A). The number of immunoreactiveneurons declined 12 hr following MK-801 injection (i.e.,82 6 9 for Fos staining and 1,2146 85 for Fos/Frastaining), but there was no statistic significance in com-parison with those of 4-hr group rats. One day after asingle MK-801 treatment, a significant reduction of theimmunolabeling occurred (P , .05), and only about 2%and 47% of RSG neurons were Fos and Fos/Fra immuno-reactive. Two or 4 days after a single MK-801 injection, afurther significant reduction (P , .01) of the immunore-activity was observed in layers III and IV of the RSG;namely, no immunoreactive cells could be identified atthese two time points using the Fos antibody, and about5% and 1% of neurons in the RSG were stained at 2 and 4days with the Fos/Fra antibody (Fig. 3A). A completedisappearance of the immunolabeling with the Fos/Fra
Fig. 2. Time course of MK-801–induced expression of Fos andFos/Fra immunoreactivity in the rat retrosplenial granularcortex (RSG). Fos and Fra expression were visualized, respec-tively, with the Fos antibody (A–E) and Fos/Fra antibody (F–J)which recognize, respectively, Fos and all the identified mem-bers of the Fos protein family present in the rat brain. Incomparison with control rat (A, F), a single MK-801 injection
(1 mg/kg) evoked Fos and Fos/Fra expression in layers III andIV beginning at 1 hr (B, G), reaching a maximal level at 4 hr (C,H), and decreasing at 1 day (D, I). One day after multipleMK-801 injections (i.e., ten consecutive injections once every2nd day), no detectable Fos immunoreactivity (E) but amoderate level of Fra expression (J) could be found in the RSG.Scale bar in A5 145 µm for all figures.
MK-801–induced Fos Induction in Cortex 723

antibody occurred 7 days following a single dose ofMK-801.
In rats killed at 4 hr or 1 day following multipleMK-801 injections, the narrow-spectrum Fos antibodyfailed to detect any specific immunopositive cells in theRSG (Fig. 2E), whereas the broad-spectrum Fos/Fraantibody recognized a moderate density of immunoreac-tive cells (Fig. 2J). Seven days after the last (i.e., thetenth) MK-801 injection, the immunoreactivity declinedand only a low level of immunolabeled cell nucleiremained (results not shown). A quantitative assessmentof Fos and Fos/Fra-immunoreactive cell numbers in theRSG following multiple MK-801 injections was shown inFigure 3B. Four hours, 1 day, and 7 days followingmultiple MK-801 injections (i.e., ten consecutive injec-tions once every 2 days), about 7026 151, 6316 117,and 546 18 RSG neurons were immunoreactive for the
broad-spectrum Fos/Fra antibody, which are equal toapproximately 45%, 41%, and 3.5% neurons in theunilateral RSG. The numbers of Fos/Fra-positive neuronsin 4-hr and 1-day groups of rats were not significantlydifferent, whereas each of these numbers were signifi-cantly different from that of the 7-day group of rats(P , .01).
Western Blot AnalysisUsing quantitative Western blot analysis with the
broad-spectrum Fos/Fra antibody that recognizes all theidentified members of Fos protein family, we were able toassess the composition and time course of Fos andvarious Fra protein expression in the rat RSG followingMK-801 administration. Control rats exhibited variedlevels of the basal expression of 46-, 43-, and 28-kDaproteins (the lane C in Fig. 4A), with the average opticaldensities (O.D.) of 11.16 4.9, 4.06 2.9 (Fig. 4B), and95.56 10.7. Four hours after a single MK-801 injection,a significant increase of 72- (O.D.5 178.36 5.6), 46-(O.D. 5 174.76 6.2) and 43-kDa protein (O.D.5128.36 10.3) expression occurred (lane ‘‘4hr’’ in Fig.4A) (P , .01 in Fig. 4B) as compared to correspondingcontrol rats. One day after a single MK-801 injection, theexpression level of 72-kDa proteins (O.D.5 7.26 3.9)significantly decreased (P , .01), whereas the high expres-sion levels of both the 46- (O.D.5 181.46 18.9) and43-kDa proteins (O.D.5 139.26 14.8) remained un-changed (lane ‘‘S1d’’ in Fig. 4A) when compared to the4-hr group of rats (lane ‘‘4hr’’ in Fig. 4A). One day aftermultiple MK-801 injections, there was a trend for theexpression levels of 72-, 46-, and 43-kDa proteins(O.D. 5 4.16 3.2, 129.76 17.3, and 98.76 15.6) todecrease (lane ‘‘M1d’’ in Fig. 4A) in comparison withthose in rats killed at 1 day after a single MK-801injection. However, these seemingly differences were notsignificant (Fig. 4B;P 5 .09). The optical densities of28-kDa proteins remained unchanged (i.e., no significantdifference) across each time delay group of MK-801–treated rats in comparison with control rats.
DISCUSSIONA systemic administration of the non-competitive
NMDA receptor antagonist MK-801 has recently beenshown to induce an acute massive expression of Fos andFra immunoreactivity or mRNAs in neurons of the ratRSG (Sharp et al., 1991; Gass et al., 1993a,b; Hughes etal., 1993; Wedzony and Czyrak, 1996; Gao et al., 1998).These studies, however, failed to provide informationabout the differential dynamic alterations of the differentmembers of the Fos protein family in the rat RSGfollowing MK-801 administration. In this study, immuno-
Fig. 3. Quantitative analysis of the time course of MK-801–induced Fos and Fos/Fra immunoreactivity in the rat retrosple-nial granular cortex (RSG). Fos and Fos/Fra expression wereinduced by a single MK-801 (A) and multiple MK-801injections (B). The numbers to the right of each bar indicate theaverage group counts of immunoreactive cells (mean6 standarderror) per section per hemisphere in both layers III and IV of theRSG (n5 3 or 6). Statistical differences (performed usingone-way ANOVA with Newman-Keuls pairwise comparisons):P , .01, d or f or h to every other bar in A, b or d to a, c, e, f inB; P , .05, c to a or g, e to g, and d or f to h in A; no significantdifference between a and b, d and f or c and e in A or between band d in B.
724 Zhang et al.

cytochemical staining and Western blot analysis wereused to identify and characterize the composition andtime course of Fos protein family expression in the ratRSG in response to systemic MK-801 injection. Ourresults obtained with both immunocytochemical andWestern blot techniques demonstrate similar changes inFos protein family expression in the RSG over time
following a single or multiple MK-801 administration.Together, these results strongly support the view that Fosand Fra proteins are differentially expressed over time inthe RSG neurons in response to systemic MK-801injection.
Induction of Fos and Fra Expression in the RSG bySystemic MK-801
Thefosgene family consists of c-fosand severalfragenes, namely,fra-1, fra-2, andfosB genes (Curran et al.,1987; Morgan and Curran, 1991). The c-fosgene encodes55-kDa proteins, whereas thefra-1 and fra-2 genesencode 35- and 46-kDa Fra proteins, respectively (Cohenand Curran, 1988, 1989; Matsui et al., 1990; Nishina etal., 1990). A singlefosB gene generates two differentforms of fosB mRNAs by alternative splicing of thetranscript, and these mRNAs have been reported toencode 45- to 48-kDa FosB and 35- to 45-kDaDFosBproteins in different rat brain regions following differentstimuli (Zerial et al., 1989; Morgan and Curran, 1991;Nakabeppu and Nathans, 1991; Yen et al., 1991; Doucetet al., 1996; Hiroi et al., 1997, 1998).
The narrow-spectrum Fos antibody used in thisstudy recognizes Fos proteins only, as characterized bythe manufacturer (Santa Cruz Biotechnology Inc., SantaCruz, CA). Using this antibody for immunocytochemis-try we found a rapid expression of Fos proteins in theRSG neurons 1 hr after a single MK-801 injection. Withthe broad-spectrum Fos/Fra antibody that is capable ofdetecting both Fos and Fra proteins, similar numbers ofimmunolabeled RSG cells (i.e., about 2% RSG neurons)were detected in the same rat brains. Therefore, 1 hourafter MK-801 injection the expressed proteins in the Fosprotein family are most probably Fos rather than Fraproteins. Four hours after a single MK-801 injection, thenarrow-spectrum Fos antibody detected an increase ofFos-immunoreactive cells (about 8% RSG neurons),whereas the broad-spectrum Fos/Fra antibody visualizeda much more dramatic elevation of immunostained cells(about 93% RSG neurons), suggesting a massive expres-sion of Fra proteins at about this time point. Thereafter,both Fos and Fra expression gradually reduced over aperiod of 1–2 days and reached an undetectable level after2–4 days. This rapid and transient expression pattern ofFos proteins and the delayed and prolonged expressionpattern of Fra proteins following a single MK-801injection are comparable to the dynamic expressionpatterns of Fos and Fra proteins observed in the rat brainfollowing acute seizure attacks (Sonnenberg et al., 1989;Pennypacker et al., 1994; Hiroi et al., 1998). Similarly,we also demonstrated a long-term expression of Fraimmunoreactivity rather than Fos immunoreactivity inthe rat RSG neurons following multiple administration ofMK-801. These results are consistent with the previous
Fig. 4. Western blot analysis of the time course of MK-801–induced Fos/Fra expression in the rat retrosplenial granularcortex (RSG).A: Fos- and Fra-immunoreactive proteins innuclei isolated by centrifugation from control rats (lane C) andfrom rats 4 hr (lane 4hr) and 1 day (lane S1d) after a singleinjection of MK-801, and 1 day following multiple MK-801injections (lane M1d) n 5 3 for each lane. The approximatemolecular weights of four apparent immunoreactive bands arelabeled on the left side, and the different molecular weights ofprotein markers (lane Marker) are labeled on the right side (i.e.,110, 74, 45, and 26 kDa). Immunoreactivity was detected usinga polyclonal antibody capable of recognizing all the identifiedmembers of the Fos protein family.B: Optical densities ofFos/Fra bands. Each bar represents the mean and standard errorof measurement from three replications of three rats each.Statistical differences (performed using one-way ANOVA withNewman-Keuls pairwise comparisons.):P , .01, b to a, c, d,and e to f, g, h, or i to j, k, l; the other paired bars (c to d, f to g orh, and j to k or l) are not significantly different.
MK-801–induced Fos Induction in Cortex 725

observations that multiple or chronic brain insults cause aprotracted expression of Fra proteins (Hiroi et al., 1998)but not Fos proteins, which undergo a refractory periodafter a single stimulation (Morgan and Curran, 1991).
We further characterized the dynamic alterations ofFos protein family expression in the RSG after MK-801administration using Western blot analysis with thebroad-spectrum Fos/Fra antibody. Three prominent bandsof proteins with relative molecular weights of 72, 46, and43 kDa appeared at 4 hours following a single MK-801injection. The expression of 72-kDa proteins decreaseddramatically at 1 day after a single MK-801 injection andremained at this very low level following multipleMK-801 administration. Because of this transient expres-sion pattern of 72-kDa proteins after a single MK-801injection as well as their resistant induction nature inresponse to multiple administration of MK-801, wepresume these to be Fos proteins. Although our findingsof a massive expression of 72-kDa Fos-like proteins areconsistent with an unpublished observation (James I.Morgan, personal communication), they are inconsistentwith other published works that report Fos proteins with arelative molecular weight of 55 kDa. This discrepancymay likely not be produced by an incorrect performanceof the Western blot technique according to the followingresults: In another Western blot experiment in which boththe RSG from MK-801–treated rats and the hippocampusfrom kainic acid–treated rats (10 mg/kg, i.p.) were used torun through the the same gel and same membrane usingthe same broad-spectrum Fos/Fra antibody, we detected asimilar expression of 72-kDa proteins in the RSG as wellas a massive expression of 55-, 46-, and 35-kDa proteinsin the hippocampus 4 hr following kainic acid injection,thus confirming Pennypacker et al.’s findings (1994).These 72-kDa proteins are unlikely to represent theexpression of heat-shock protein-70 (HSP70) in the RSGbecause the time course of this 72-kDa protein expressionis entirely different from that of HSP70 expression:whereas the expression of these 72-kDa proteins reachedpeak levels at about 4 hr and almost disappeared at about1 day after a single MK-801 injection (Fig. 4), MK-801–induced HSP70 expression began at about 12 hr andpeaked at about 1 day post-injection (Sharp et al., 1991).We account for the discrepancy in molecular weight bythe fact that Fos proteins can undergo extensive post-translational modification whereby the 55-kDa Fos pro-teins can be converted into a discrete series of 57-, 60-,and 62-kDa products in the nucleus (Curran et al., 1984).Therefore, it is possible that the 72-kDa proteins that weobserved here may be yet another form of post-translationally modified 55-kDa Fos proteins.
Prolonged expression of the 46- and 43-kDa pro-teins were detected with Western blot analysis followingeither a single or multiple MK-801 administration. Al-
though the identity of these two bands of proteins ispresently unknown, the prolonged expression patternsuggests that these two bands most probably representFra proteins. We presume that the 43- and 46-kDaproteins are not Fra-2 and/or the post-translationallymodified form of Fra-2, even though Fra-2 proteins havea similar relative molecular weight, due to the fact thatthere have been no reports of protracted expression ofFra-2 proteins in the rat brain following any brain insults.It may be more likely that these proteins could be FosBand/or its post-translationally modified form, based onthe findings that a massive and relatively prolongedexpression of FosB or FosB-like proteins occurred in therat brain following acute brain insults (Sonnenberg et al.,1989; Pennypacker et al., 1994; Doucet et al., 1996; Hiroiet al., 1998). Some of these studies, however, also showeda resistance to the induction of FosB in response tomultiple or chronic brain insults (Doucet et al., 1996;Hiroi et al., 1998), which would argue against ourchronically induced 43- and 46-kDa proteins as represent-ing FosB.
It has been shown that acute seizure attacks or drugadministration produce a delayed and prolonged expres-sion of DFosB proteins (Sonnenberg et al., 1989; Pen-nypacker et al., 1994; Doucet et al., 1996; Mandelzys etal., 1997; Hiroi et al., 1998), and that multiple or chronicseizure attacks or other brain insults induce a relativelyhigh level expression ofDFosB (Doucet et al., 1996;Hiroi et al., 1998). Whereas some of these studiesdetectedDFosB with relative molecular weights of 35–37kDa in the rat hippocampus or the cerebral cortexfollowing seizure attacks (Sonnenberg et al., 1989; Pen-nypacker et al., 1994; Mandelzys et al., 1997; Hiroi et al.,1998), one study showedDFosB with relative molecularweights of 43 and 45 kDa in the rat striatum followingdestruction of the nigrostriatal pathway by 6-hydroxydo-pamine (Doucet et al., 1996). These results suggest thatdifferent forms ofDFosB proteins may be expressed indifferent brain regions following different brain stimula-tions, possibly as a result of the regulation ofDFosBpost-translational modification process by some unknownfactors. The present findings of a delayed and prolongedexpression of the 43- and 46-kDa proteins in the RSGfollowing an acute MK-801 injection as well as theirprotracted expression following multiple MK-801 injec-tions strongly suggest that these proteins representDFosB.It is apparent that this suggestion needs to be furtherconfirmed by employing a specific antibody that recog-nizeDFosB proteins.
In addition, we also detected a steady expression of28-kDa proteins in the RSG after vehicle or MK-801injection(s). This observation is in agreement with Doucetet al. (1996) who, using the same Fos/Fra antibody thatwe used here, demonstrated that a lesion to the medial
726 Zhang et al.

forebrain bundle evokes a substantial expression ofDFosB without obvious effects on the basal expression of28-kDa proteins in the striatum. The identity of these28-kDa proteins is not known.
Possible Relevance to Psychotomimetic Effectsof PCP and Ketamine
The RSG is an important nodal structure for thetransfer of information between the hippocampus andmany areas of the neocortex (Wyss and Van Groen, 1992;Zilles and Wree, 1995) and is critically involved in someabnormal behavioral alterations in rats induced by MK-801, such as ataxic movements and horizontal headwavings (Reperant et al., 1987; Markowska et al., 1989;Murray et al., 1989; Taube, 1992). It has been proposedthat c-fos gene expression plays an important role inlinking transient changes of neuronal activity to bothshort- and long-term modifications of structure andfunction in the mammalian brain (Heilig et al., 1993;Cirelli et al., 1995; Wollnik et al., 1995; Watanabe et al.,1996). The prolonged expression ofDFosB has also beenfound to be critical for long-term molecular, electrophysi-ological, and behavioral adaptations to individual braininsults (Hiroi et al., 1997, 1998; Mandelzys et al., 1997).Given that about 93% of neurons in the RSG have beenobserved in this study to express Fos and most probablyDFosB 4 hr following a single MK-801 injection and thatabout 45% of neurons exhibitDFosB-like immunoreactiv-ity 4 hr following the last multiple MK-801 injection, it ishighly probably that MK-801–induced massive Fos andDFosB-like expression in the RSG may play a critical rolein MK-801–induced pathophysiological changes in therat brain.
Similar to our results with MK-801, systemic PCPor ketamine injection produces a massive expression ofFos/Fra in the rat RSG (Nakki et al., 1996; Duncan et al.,1998). Acute or chronic digestion of PCP has long beenknown to produce and reactivate acute (Luby et al., 1959;Rosenbaum et al., 1959; Allen and Young, 1978; Javittand Zukin, 1991) or enduring (Carlin et al., 1979;Pearlson, 1981; Young et al., 1987; Cosgrove and Newell,1991) positive, negative, and cognitive schizophrenicsymptoms in humans, which is seen only after pubertyand thus is consistent with the development history ofschizophrenia (Coyle, 1996; Hirsch et al., 1997). Re-cently, ketamine has also been found to produce similarsymptoms in humans (Krystal et al., 1994; Malhotra etal., 1996, 1997). As many of these schizophrenic symp-toms of PCP and ketamine have also been shown to occurin rodents in varying degrees (Wesierska et al., 1990;Ogawa et al., 1994; Sams-Dodd, 1996; Gallo et al., 1998;Jentsch et al., 1998a,b; Pie Traszek and Ossowska, 1998;Turgeon et al., 1998), these two drugs are routinely used
in many laboratories as both clinical and animal modelsof schizophrenia (for reviews, see Javitt and Zukin, 1991;Halberstadt, 1995; Coyle, 1996; Goff and Wine, 1997;HerescoLevy and Javitt, 1998; O’Donnell and Grace,1998). We propose that MK-801 may provide an addi-tional tool for the study of PCP/ketamine-induced psycho-sis or schizophrenia based on the following arguments: 1)Although the potent and selective NMDAreceptor antago-nist MK-801 was not approved for human use, PCP- andketamine-induced psychosis in humans has been associ-ated exclusively with their antagonistic effects on NMDAreceptors (Javitt and Zukin, 1991; Krystal et al., 1994;Malhotra et al., 1996, 1997); 2) MK-801–induced behav-ioral changes in rats highly resemble those of PCP (Koeket al., 1988; Willets et al., 1990); and 3) MK-801, PCPand ketamine can induce similar expression of Fos/Fra inthe rat RSG. Thus, given that the RSG is tightlyassociated with those brain structures, including thehippocampus and many areas of the neocortex, whichhave been found to be associated with schizophrenia(Bogerts, 1993; Port and Seybold, 1995; Bachus andKleinman, 1996; Velakoulis and Pantelis, 1996; O’Donnelland Grace, 1998), all the above lines of evidence suggestthat further studies on the exact role(s) of MK-801–induced Fos and Fra protein expression in the rat RSGmay provide additional insight into understanding thepathophysiological mechanisms of schizophrenia.
REFERENCES
Allen RM, Young SJ. 1978. Phencyclidine-induced psychosis. Am JPsychiatry 135:1081–1084.
Bachus SE, Kleinman JE. 1996. The neuropathology of schizophrenia.J Clin Psychiatr 57(Suppl 11):72–83.
Bogerts B. 1993. Recent advances in the neuropathology of schizophre-nia. Schiz Bull 19:401–445.
Carlin AS, Grant I, Adams KM, Reed R. 1979. Is phencyclidine (PCP)abuse associated with organic mental impairment? Am J DrugAlcohol Abuse 6:273–281.
Cirelli C, Pompeiano M, Arrighi P, Tononi G. 1995. Sleep-wakingchanges after c-fos antisense injection in the medial preopticarea. Neuroreport 6:801–805.
Cohen DR, Curran T. 1988.fra-1: a serum-inducible, cellular immediate-early gene that encodes a Fos-related antigen. Mol Cell Biol8:2063–2069.
Cohen DR, Curran T. 1989. The structure and function of thefosproto-oncogene. Crit Rev Oncogene 1:65–88.
Cosgrove J, Newell TG. 1991. Recovery of neuropsychologicalfunctions during reduction in use of phencyclidine. J ClinPsychol 47:159–169.
Coyle JT. 1996. The glutamatergic dysfunction hypothesis for schizo-phrenia. Harvard Rev Psychiatry 3:241–253.
Curran T, Gordon MB, Rubino KR, Sambucetti LC. 1987. Isolation andcharacterization of the c-fos (rat) cDNA and analysis ofpost-translational modification in vitro. Oncogene 2:79–84.
Curran T, Miller AD, Zokas L, Verma IM. 1984. Viral and cellular Fosproteins: a comparative analysis. Cell 36:259–268.
MK-801–induced Fos Induction in Cortex 727

Doucet JP, Murphy BJ, Tuna BS. 1990. Modification of a discontinuousand highly porous sodium dodecyl sulfate-polyacrylamide gelsystem for minigel electrophoresis. Anal Biochem 190:209–211.
Doucet JP, Nakabeppu Y, Bedard PJ, Hope BT, Nestler EJ, Jasmin BJ,Chen JS, Iadarola MJ, St-Jean M, Wigle N, Blanchet P, GrondinR, Roberson GS. 1996. Chronic alterations in dopaminergicneurotransmission produce a persistent elevation of (FosB-likeprotein(s) in both the rodent and primate striatum. Eur JNeurosci 8:365–381.
Duncan GE, Moy SS, Knapp DJ, Mueller RA, Breese GR. 1998.Metabolic mapping of the rat brain after subanesthetic doses ofketamine: potential relevance to schizophrenia. Brain Res787:181–190.
Gallo M, Bielavska E, Roldan G, Bures J. 1998. Tetrodotoxininactivation of the gustatory cortex disrupts the effect of theN-methyl-D-aspartate antagonist ketamine on latent inhibitionof conditioned taste aversion in rats. Neurosci Lett 240:61–64.
Gao XM, Hashimoto T, Tamminga CA. 1998. Phencyclidine (PCP) anddizocilpine (MK801) exert time-dependent effects on the expres-sion of immediate early genes in rat brain. Synapse 29:14–28.
Gass P, Herdegen T, Bravo R, Kiessling M. 1993a. Induction andsuppression of immediate-early genes in specific rat brainregions by the non-competitiveN-methyl-D-aspartate receptorantagonist MK-801. Neuroscience 53:749–758.
Gass P, Herdegen T, Bravo R, Kiessling M. 1993b. Spatiotemporalinduction of immediate early genes in rat brain after limbicseizures: effects of NMDA receptor antagonist MK-801. Eur JNeurosci 53:749–758.
Goff DC, Wine L. 1997. Glutamate in schizophrenia: clinical andresearch implications. Schizophr Res 27:157–168.
Halberstadt AL. 1995. The phencyclidine-glutamate model of schizo-phrenia. Clin Neuropharmacol 18:237–249.
Heilig M, Engel JA, Soderpalm B. 1993. C-fos antisense in the nucleusaccumbens blocks the locomotor stimulant action of cocaine.Eur J Pharmacol 236:339–340.
HerescoLevy U, Javitt DC. 1998. The role ofN-methyl-D-aspartate(NMDA) receptor-mediated neurotransmission in the pathophysi-ology and therapeutics of psychiatric syndromes. Eur Neuropsy-chopharmacol 8:141–152.
Hiroi N, Brown JR, Haile CN, Ye H, Greenberg ME, Nestler EJ. 1997.FosB mutant mice: loss of chronic cocaine induction ofFos-related proteins and heightened sensitivity to cocaine’spsychomotor and rewarding effects. Proc Natl Acad Sci USA94:10397–10402.
Hiroi N, Marek GJ, Brown JR, Ye H, Saudou F, Vaidya VA, Duman RS,Greenberg ME, Nestler EJ. 1998. Essential role of the FosBgene in molecular, cellular, and behavioral actions of chronicelectroconvulsive seizures. J Neurosci 18:6952–6962.
Hirsch SR, Das I, Garey LJ, De Belleroche J. 1997. A pivotal role forglutamate in the pathogenesis of schizophrenia, and its cogni-tive dysfunction. Pharmacol Biochem Behav 56:797–802.
Hughes P, Dragunow M, Beilharz E, Lawlor P, Gluckman P. 1993.MK801 induces immediate-early gene proteins and BDNFmRNA in rat cerebrocortical neurons. Neuroreport 4:183–186.
Javitt DC, Zukin SR. 1991. Recent advances in the phencyclidinemodel of schizophrenia. Am J Psychiatry 138:1301–1308.
Jentsch JD, Elsworth JD, Taylor JR, Redmond DE, Roth RH. 1998a.Dysregulation of mesoprefrontal dopamine neurons induced byacute and repeated phencyclidine administration in the nonhu-man primate: implications for schizophrenia. Adv Pharmacol42:810–814.
Jentsch JD, Taylor JR, Roth RH. 1998b. Subchronic phencyclidineadministration increases mesolimbic dopaminergic system re-
sponsitivity and augments stress- and psychostimulant-inducedhyperlocomotion. Neuropsychopharmacol 19:105–113.
Koek W, Woods J, Winger G. 1988. MK-801, a proposed non-competitive antagonist of excitatory amino acid neurotransmis-sion, produced phencyclidine-like behavioral effects in pigeons,rats and rhesus monkeys. J Pharmacol Exp Ther 245:969–974.
Krystal JH, Karper LP, Seibyl JP, Freeman GK, Delaney R, BremnerJD, Heninger GR, Bowers MBJ, Charney DS. 1994. Subanes-thetic doses of the non-competitive NMDA antagonist, ket-amine, in humans. Psychotomimetic, perceptual, cognitive andneuroendocrine responses. Arch Gen Psychiatry 51:199–214.
Luby E, Cohen B, Rosenbaum F, Gottlieb J, Kelley R. 1959. Study of anew schizophrenomimetic drug, Sernyl. Arch Neurol Psychiatry81:363–369.
Malhotra AK, Pinals DA, Weingartner H, Sirocco K, Missar CD,Pickar D, Breier A. 1996. NMDA receptor function and humancognition: the effects of ketamine in healthy volunteers. Neuro-psychopharmacol 14:301–307.
Malhotra AK, Pinals DA, Adler CM, Elman I, Clifton A, Pickar D,Breier A. 1997. Ketamine-induced exacerbation of psychoticsymptoms and cognitive impairment in neuroleptic-free schizo-phrenics. Neuropsychopharmacol 17:141–150.
Mandelzys A, Gruda MA, Bravo R, Morgan JI. 1997. Absence of apersistently elevated 37 kDa Fos-related antigen and AP-1-likeDNA-binding activity in the brains of kainic acid-treatedfosBnull mice. J Neurosci 17:5407–5415.
Markowska AL, Olton DS, Murray EA, Gaffan D. 1989. A comparativeanalysis of the role of fornix and cingulate cortex in memory:rats. Exp Brain Res 74:187–201.
Matsui M, Tokuhara M, Konuma Y, Nomura N, Ishizaki R. 1990.Isolation of humanfos-related genes and their expression duringmonocyte-macrophage differentiation. Oncogene 5:249–255.
Morgan JI, Curran T. 1991. Stimulus-transcription coupling in thenervous system: involvement of the inducible proto-oncogenesfosandjun.Annu Rev Neurosci 14:421–451.
Murray EA, Davidson M, Gaffan D, Olton DS, Suomi S. 1989. Effectsof fornix transaction and cingulate cortical ablation on spatialmemory in monkeys. Exp Brain Res 74:173–186.
Nakabeppu Y, Nathans D. 1991. A naturally occurring truncated formof FosB that inhibits Fos/Jun transcriptional activity. Cell64:751–759.
Nakki R, Sharp FR, Sagar SM, Honkaniemi J. 1996. Effects ofphencyclidine on immediate early gene expression in the brain.J Neurosci Res 45:13–27.
Nishina H, Sato H, Suzuki T, Sato M, Iba H. 1990. Isolation andcharacterization offra-2, an additional member of thefosgenefamily. Proc Natl Acad Sci USA 87:3619–3623.
O’Donnell P, Grace AA. 1998. Dysfunctions in multiple interrelatedsystems as the neurobiological bases of schizophrenic symptomclusters. Schiz Bull 24:267–283.
Ogawa S, Okuyama S, Araki H, Nakazato A, Otomo S. 1994. A ratmodel of phencyclidine psychosis. Life Sci 55:1605–1610.
Olney JW, Labruyere J, Price MT. 1989. Pathological changes inducedin cerebrocortical neurons by phencyclidine and related drugs.Science 244:1360–1362.
Paxinos G, Watson C. 1986. The rat brain in stereotaxic coordinates.Sydney: Academic.
Pearlson GD. 1981. Psychiatric and medical syndromes associatedwith phencyclidine (PCP) abuse. Johns Hopkins Med J 148:25–33.
Pennypacker KR, Thai L, Hong JS, McMillian MK. 1994. Prolongedexpression of AP-1 transcription factors in the rat hippocampusafter systemic kainate treatment. J Neurosci 14:3998–4006.
Pennypacker KR, Hong JS, McMillian MK. 1995. Implications ofFos-related antigens. Trends Pharmacol Sci 16:317–321.
728 Zhang et al.

Pie Traszek M, Ossowska K. 1998. Chronic treatment with haloperidoldiminishes the phencyclidine-induced sensorimotor gating defi-cit in rats. Naunyn Schmiedebergs Arch Pharmacol 357:466–471.
Port RL, Seybold KS. 1995. Hippocampal synaptic plasticity as abiological substrate underlying spisodic psychosis. Biol Psychia-try 37:318–324.
Reperant J, Weidner C, Pakhomova A, Desroches AM, Vesseklin N,Lemire M. 1987. A retino-thalamo-hippocampal pathway in therat. CR Acad Sci 305:601–604.
Rosenbaum G, Cohen BD, Luby ED, Gottlieb JS, Yelen D. 1959.Comparison of phencyclidine hydrochloride (Sernyl) with otherdrugs: simulation of schizophrenic performance with Sernyl,LSD-25, and amobarbital (Amytal) sodium. I. Attention, motorfunction, and proprioception. Arch Gen Psychiatry 1:651–656.
Sams-Dodd F. 1996. Phencyclidine-induced stereotyped behaviour andsocial isolation in the rat: a possible animal model of schizophre-nia. Behav Pharmacol 7:3–23.
Sharp FR, Jasper P, Hall J, Noble L, Sagar SM. 1991. MK-801 andketamine induce heat shock protein HSP72 in injured neurons inposterior cingulate and retrosplenial cortex. Ann Neurol 30:801–809.
Shu S, Ju G, Fan L. 1988. The glucose oxidase-DAB-nickel method inperoxidase histochemistry of the nervous system. Neurosci Lett85:169–171.
Sonnenberg JL, Macgregor-Leon PF, Curran T, Morgan JI. 1989.Dynamic alterations occur in the levels and composition oftranscription factor AP-1 complexes after seizure. Neuron3:359–365.
Taube JS. 1992. Quantitative analysis of head-direction cells recordedin the rat anterior thalamus. Soc Neurosci Abstr 18:708.
Towbin H, Staehelin T, Gordon J. 1979. Electrophoretic transfer ofproteins from polyacrylamide gels to nitrocellulose sheets:procedure and some applications. Proc Natl Acad Sci USA76:4350–4354.
Turgeon SM, Auerbach EA, Heller MA. 1998. The delayed effects ofphencyclidine (PCP) disrupt latent inhibition in a conditionedtaste aversion paradigm. Pharmacol Biochem Behav 60:553–558.
Velakoulis D, Pantelis C. 1996. What have we learned from functionalimaging studies in schizophrenia? The role of frontal, striataland temporal areas. Aust N Z JPsychiatr 30:195–209.
Watanabe Y, Johnson RS, Butler LS, Binder DK, Spiegelman BM,Papaioannou VE, McNamara JO. 1996. Null mutation of c-fos
impairs structural and functional plasticities in the kindlingmodel of epilepsy. J Neurosci 16:3827–3836.
Wedzony K, Czyrak A. 1996. Competitive and non-competitiveNMDA receptor antagonists induce c-Fos expression in the ratanterior cingulate cortex. J Physiol Pharmacol 47:525–533.
Wesierska M, Macias-Gonzalez R, Bures J. 1990. Differential effect ofketamine on the reference and working memory versions of theMorris water maze task. Behav Neurosci 104:74–83.
Willets J, Balster RL, Leander D. 1990. The behavioral pharmacologyof NMDA receptor antagonists. Trends Pharmacol Sci 11:423–428.
Wollnik F, Brysch W, Uhlmann E, Gillardon F, Bravo R, ZimmermannM, Schlingensiepen KH, Herdegen T. 1995. Block of c-fosandjunB expression by antisense oligonucleotides inhibits lightinduced phase shifts of the mammalian clock. Eur J Neurosci7:388–393.
Wyss JM, Van Groen T. 1992. Connections between the retrosplenialcortex and the hippocampal formation in the rat: a review.Hippocampus 2:1–12.
Yen J, Wisdom RM, Tratner I, Verma IM. 1991. An alternative splicedform of FosB is a negative regulator of transcriptional activationand transformation by Fos proteins. Proc Natl Acad Sci USA88:5077–5081.
Young ST, Porrino LT, Iadarola MJ. 1991. Cocaine induces striatalc-Fos-immunoreactive proteins via dopaminergic D1 receptors.Proc Natl Acad Sci USA 88:1291–1195.
Young T, Lawson GW, Gacono CB. 1987. Clinical aspects of phency-clidine (PCP). Int J Addict 22:1–15.
Zerial M, Toschi L, Ryseck RP, Schuermann M, Muller R, Bravo R.1989. The product of a novel growth fact activated gene,fosB,interacts with JUN proteins enhancing their DNA bindingactivity. EMBO J 8:805–813.
Zhang X, Boulton AA, Yu PH. 1996. Expression of heat shockprotein-70 and limbic seizure-induced neuronal death in the ratbrain. Eur J Neurosci 8:1432–1440.
Zhang X, Gelowitz DL, Lai CT, Boulton AA, Yu PH. 1997a. Gradationof kainic acid-induced rat limbic seizures and expression ofhippocampal heat shock protein-70. Eur J Neurosci 9:760–769.
Zhang X, Le Gal La Salle G, Ridoux V, Yu PH, Ju G. 1997b. Preventionof kainic acid-induced limbic seizures and Fos expression byGABA-A receptor agonist muscimol. Eur J Neurosci 9:29–40.
Zilles K, Wree A. 1995. Cortex: areal and laminar structure. In: PaxinosG, editor. The rat nervous system, 2nd ed. San Diego: AcademicPress. p 649–685.
MK-801–induced Fos Induction in Cortex 729