methylthioadenosine phosphorylase regulates … · the gene encoding methylthioadenosine...
TRANSCRIPT

Methylthioadenosine Phosphorylase Regulates Ornithine Decarboxylase by
Production of Downstream Metabolites*
Ahmad L. Subhi‡, Paula Diegelman§, Carl W. Porter§, Baiqing Tang‡, Zichun J. Lu¶,
George D. Markham¶, and Warren D. Kruger‡||
From the Divisions of ‡Population Science and ¶Basic Science, Fox Chase Cancer Center,
Philadelphia, PA 19111 and the §Roswell Park Cancer Institute, Pharmacology and
Therapeutics Department, Buffalo, NY 14263
* This work was supported by United States Army Grant DAMD17-97-1-7707, USPHS grant
CA-22153 and Core grant CA-06927 from the National Institutes of Health and by an
appropriation from the Commonwealth of Pennsylvania. We also acknowledge the work of the
Sequencing and Cell Culture Facilities at Fox Chase Cancer Center.
||To whom correspondence should be addressed: Fox Chase Cancer Center, 333 Cottman
Avenue, Philadelphia, PA 19111-2497. Tel: 215-728-3030; Fax: 215-214-1623;
E-mail: [email protected]
Running title: MTAP regulation of ODC
Copyright 2003 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on September 23, 2003 as Manuscript M308451200 by guest on June 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from

The abbreviations used are: MTA, methylthioadenosine; MTR1P, methylthioribose-1-
phosphate; MTAP, methylthioadenosine phosphorylase; ODC, ornithine decarboxylase ;
DFMO, α-difluoromethylornithine; SAM, S-adenosylmethionine, dSAM, decarboxylated S-
adenosylmethionine.
2
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from

SummaryThe gene encoding methylthioadenosine phosphorylase (MTAP), the initial enzyme in the
methionine salvage pathway, is deleted in a variety of human tumors and acts as a tumor
suppressor gene in cell culture (1-6). Overexpression of the polyamine biosynthetic enzyme
ornithine decarboxylase (ODC) is frequently observed in tumors and has been shown to be
tumorigenic in vitro and in vivo (7-9). In this paper, we demonstrate a novel regulatory pathway
in which the methionine salvage pathway products inhibit ODC activity. We show that in
Saccharomyces cerevisiae the MEU1 gene encodes MTAP and that Meu1∆ cells have an eight-
fold increase in ODC activity resulting in large elevations in polyamine pools. Mutations in
putative salvage pathway genes downstream of MTAP also cause elevated ODC activity and
elevated polyamines. Addition of the penultimate salvage pathway compound 4-methylthio-2-
oxobutanoic acid (MTOB) represses ODC levels in both MTAP deleted yeast and human tumor
cell lines, indicating that MTOB acts as a negative regulator of polyamine biosynthesis.
Expression of MTAP in MTAP deleted MCF-7 breast adenocarcinoma cells results in a
significant reduction of ODC activity and reduction in polyamine levels. Taken together, our
results show that products of the methionine salvage pathway regulate polyamine biosynthesis,
and suggest that MTAP deletion may lead to ODC activation in human tumors.
3
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from

INTRODUCTION
A quarter century ago, Toohey first recognized that certain murine malignant hematopoietic
cell lines lacked methylthioadenosine phosphorylase (MTAP) activity (10). MTAP is a key
enzyme in the methionine salvage pathway (see Figure 1). This pathway functions to salvage
methylthioadenosine (MTA), which is formed as a byproduct of polyamine metabolism.
Phosphorolysis of MTA by MTAP results in the conversion of MTA into adenine and
methylthioribose-1-phosphate (MTR1P). A series of reactions then salvages the methyl-thio
group from MTR1P to form methionine. This pathway has been most extensively studied in
Klebsiella pneumoniae (11-14), but has also been shown to exist in rat liver (15-17) and in
Saccharomyces cerevisiae (18,19).
Loss of MTAP activity through gene deletion is common in many kinds of human cancers
including non-small cell lung cancer, glioma, T-cell acute leukemia, bladder cancer,
osteosarcoma and endometrial cancer (3,5,20-22). Recently, we found that expression of MTAP
in a MTAP-deleted breast adenocarcinoma cell line (MCF-7) resulted in a dramatic inhibition
of tumorigenicity in vitro and in vivo, showing that MTAP can function as a tumor suppressor
gene (1). We also found that MTAP expression causes a significant decrease in intracellular
polyamine levels and alters the ratio of putrescine to total polyamines. Consistent with this
observation, the polyamine biosynthesis inhibitor α-difluoromethylornithine (DFMO) inhibits
the ability of MTAP deficient MCF-7 cells to form colonies in soft-agar, while addition of the
polyamine putrescine stimulates colony formation in MTAP expressing MCF-7 cells. These
4
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from

results indicate that MTAP’s tumor suppressor activity may be mediated by its effect on the
intracellular polyamine pools.
Polyamines are small, aliphatic amines involved in a variety of cellular processes including
transcription and apoptosis (8). The rate-limiting enzyme in the production of polyamines is
ornithine decarboxylase (ODC). Elevated ODC activity has been observed in a wide variety of
human and animal tumors, and overexpression of ODC in NIH/3T3 cells is sufficient to cause
transformation in vitro (7,23). Transgenic mice overexpressing ODC in skin develop skin
tumors at a high frequency (24). These observations show that overexpression of ODC is
tumorigenic.
Examination of the S. cerevisiae genome for MTAP homologues suggests that the MEU1
gene, with 35% identity and 53% similarity over 275 amino acids, may encode the yeast MTAP
homologue. Furthermore, recent studies show that cells lacking MEU1, in combination with a
mutation that allows yeast to take up methylthioadenosine, are unable to grow on medium
containing MTA as the sole sulfur source (25). MEU1 was initially identified in a screen for
genes that regulate expression of the yeast ADH2 gene (26). Overexpression of MEU1 increased
ADH2 expression while deletion of MEU1 resulted in reduced ADH2 expression. At the time
these experiments were published the MTAP gene had not yet been identified, so the relationship
between MEU1 and MTAP was unknown. These experiments demonstrate that MEU1 regulates
ADH2 expression and indicate a link between MEU1 and gene regulation.
In this paper we show that MEU1 encodes yeast MTAP, and we characterize the effect of
5
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from

loss of MTAP in Saccharomyces cerevisiae and human tumor cells. Our findings show that there
is a regulatory link between the methionine salvage pathway and the polyamine pathway.
Specifically, the penultimate metabolite, 4-methylthio-2-oxobutanoic acid (MTOB), acts to
repress ornithine decarboxylase, the rate-limiting enzyme in polyamine biosynthesis. These
results explain why MTAP deletion causes altered polyamine profiles, and suggest that MTAP’s
tumor suppressor activity may be due to its effect on ODC activity.
EXPERIMENTAL PROCEDURES
Plasmid pMEU1 construction – The yeast MEU1 ORF was PCR amplified from total yeast
DNA using primers MEU1-1F (5′- TCT GTG AAA CAT GTC) and MEU1-1R (5′ CAG TCC
CCA AGG GGG). The resulting 1.3 Kb product was the cloned into pCR2.1 (Invitrogen) and
designated pCR2.1::MEU1. PCR2.1::MEU1 was digested with BamH1 and NotI and the
resulting 1.3 Kb insert was cloned into pRS316 (27) digested with the same enzymes.
Yeast Strains – Yeast strains used are indicated in Table 1. Isogenic MEU1 and meu1∆ strains
were created by transformation of AS3-2a (a ura3∆ met15∆ meu1∆::LEU2 leu2∆) with pMEU1 or
empty vector pRS316. All other strains used were created by the S. cerevisiae deletion project (28).
Standard yeast growth media was used as described by Sherman (29). Polyamine free media was
prepared as described by Balasundaram (30).
Mammalian Cells – MTAP expressing and mutant expressing MCF-7 cells were created and
grown as previously described (1). MIA PaCa-2 pancreatic adenocarcinoma cells were obtained
6
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from

from the ATCC and grown as previously described (31).
Yeast Growth Studies – Yeast cells were grown to stationary phase and then diluted to an
OD600 of 0.05 in the indicated media. Cells were then grown in liquid media with shaking at 30oC
for 48 hours, at which time OD was determined.
Extract preparation – Yeast cultures (45 ml) were grown aerobically to an OD600 of 0.5-1.0,
pelleted, washed with 0.9ml of lysis buffer (50 mM Tris pH 7.5, 100 mM NaCl, 1 mM PMSF),
and resuspended in 750µl lysis buffer. Glass beads (0.5 mm, one-half weight of the cell pellet)
were then added to make up approximately 50-70% of total volume. Cells were lysed by using a
BeadBeater at 4°C with maximum speed (4500 rpm) for 1 minute three times and kept on ice
between pulses (Biospec Products). After lysis, the tubes were centrifuged for 4 minutes and the
supernatant was transferred to a new tube and glycerol was added to a final concentration of
15%. The protein concentration of yeast extracts was determined by the Coomassie Blue protein
assay reagent (Pierce) using bovine serum.
Mammalian cell lysates were prepared by 3 cycles of freeze-thawing at –80°C in 20 mM
KH2PO4, pH 7.4 containing 1 mM phenylmethylsulfonyl fluoride (Sigma) and 1 mM dithiothreitol
(Sigma).
Western blot analysis Forty µg of extract were run on pre-made 7% NuPage Tris-acetate
SDS gels (Invitrogen) and electrotransferred to PVDF membrane (Biorad) using a standard
7
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from

protocol (32). Samples were probed with a 1/10,000 dilution of yeast anti-ornithine
decarboxylase antiserum raised in rabbits (a generous gift from Martin Hoyt and Phil Cofino,
UCSF). Secondary antibody anti-rabbit HRP linked antibody was obtained from Amersham
Biosciences and used at a 1/3,000 dilution. The antibody-Antigen complex was visualized on
photographic film after treatment with SuperSignal West Dura Extended Duration Substrate
(Pierce biotechnology).
Measurement of ornithine decarboxylase, MTAP, and AdoMet decarboxylase activity –
Ornithine decarboxylase activity was assayed by measuring the 14CO2 formed by
decarboxylation of [1-14C] labeled ornithine in as previously described (33). L-carboxy 14C
ornithine (5mCi/mmol) was obtained from Moravek Biochemical (Brea, CA). 20 µg of yeast or
mammalian extract was used per reaction. Reactions were performed between three and seven
times per sample and results are presented as the average with the standard error. Adomet
decarboxylase activity was assayed by measuring the amount of 14CO2 released by
decarboxylation of [1-14C] labeled AdoMet as described (34).
MTAP activity in yeast extracts was determined using a spectrophotometric assay as
described previously (1). One unit of MTAP activity is defined as the amount that catalyzes the
formation of 1µmol of adenine/min.
Measurement of intracellular polyamine levels by using HPLC – Yeast cells grown in
synthetic complete media were collected, protein extracts were prepared, and then 100µl
8
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from

supernatant added to 100µl 0.6N PCA and allowed to sit on ice for 5 minutes. The sample was
centrifuged at 15,000g for 10 min. 100µl of supernatant was dansylated and analyzed by HPLC
as described (35).
RESULTS
MEU1 encodes MTAP in S. cerevisiae – Early studies showed that yeast extracts contain
MTAP activity, but did not identify the gene encoding the activity (18,19). A BLAST search of
the S. cerevisiae genome with human MTAP (NP_002442) reveals two open reading frames,
YLR209C and YLR017W, with significant sequence similarity. Of the two, YLR017W (also
known as MEU1) has the most similarity with 35% identity over 275 amino acids to human
MTAP. To test the possibility that MEU1 encoded MTAP, we constructed isogenic MEU1 and
meu1∆ strains and measured MTAP activity in whole cell extracts (see Materials and Methods).
We found that MEU1 yeast extracts contained 0.38 units of MTAP activity, while extracts from
meu1∆ strain had no detectable activity (<0.01units). A second experiment to confirm our
hypothesis involved examining the growth of isogenic meu1∆ and MEU1 strains with
auxotrophic markers for adenine and methionine in media in which MTA had been substituted
for methionine and adenine. We found that wild-type but not meu1∆ yeast can grow using
methylthioadenosine (MTA) as a substitute for methionine and adenine (Figure 2). Taken
together, these observations show that MEU1 encodes yeast MTAP.
Effect of meu1∆ on polyamine pools and ODC activity Since MTA is a by-product of
9
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from

polyamine production, we suspected that cells lacking MTAP might have altered polyamine
pools due to end-product inhibition. Therefore, we measured the intracellular level of
polyamines (putrescine, spermine and spermidine) in an isogenic wild-type strain and in a
meu1∆ strains using HPLC (see Materials and Methods). We found that in synthetic complete media,
MEU1 cells had undetectable levels of putrescine, while meu1∆ cells contained at least a 20-fold
increase in putrescine (Figure 3a). Surprisingly, we also found that spermidine levels were
significantly increased and that the total polyamine content of meu1∆ cells was more than twice
that of isogenic wild-type yeast (62 vs. 28 nmoles/mg).
To explain the overall elevation in polyamine levels we examined ornithine decarboxylase
(ODC) activity in wild type and meu1∆ yeast. ODC is the rate-limiting enzyme in polyamine
production and is regulated at the post-transcriptional level (36). We found that cell extracts
from meu1∆ yeast had an eight-fold increase in ODC activity compared to isogenic wild-type
yeast (Figure 3b). This difference appeared to be due to elevated levels of ODC protein. We
were unable to detect ODC protein by immunoblot in extracts derived from MEU1 cells as has
been previously observed (37) and from ODC deficient cells (spe1∆), but we saw substantial
expression in meu1∆ extracts (Figure 4a, lanes 1-3). The increase in protein levels appeared to
be even greater then the measured activity, suggesting that not all of the protein was active (lanes
1 and 2). We also noticed that meu1∆ yeast formed significantly larger colonies compared to
isogenic MEU1 yeast on polyamine free media, suggesting that the elevated ODC is important
for growth under theses conditions (Figure 4b). These results show that MEU1 represses ODC
10
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from

expression and activity.
We also examined the effect of MEU1 on S-adenosylmethionine decarboxylase (SAMDC)
activity. We found that extracts from meu1∆ cells had twice the SAMDC activity of isogenic
MEU1 (7.7 ± 0.3 vs. 14.6 ± 0.8 nmol/mg/hr). This result is consistent with the idea that MEU1
acts to repress polyamine synthesis.
SPE2 and MEU1 regulate ODC through different mechanisms – Previous work has shown
that mutations in SPE2, encoding SAM decarboxylase, also cause a dramatic increase in ODC
protein levels (36) (also Figure 3a, lane 4). It was also shown that the ODC overexpression
observed in a spe2 mutant could be suppressed by the addition to spermidine to the media. We
compared ODC expression in spe2∆ and meu1∆ strains in the presence and absence of
spermidine. Meu1∆, spe2∆ and meu1∆/spe2∆ double mutants were grown in polyamine free
medium with or without the addition of exogenous spermidine. When we measured ODC
enzyme activity we found that addition of 0.1 mM spermidine to the media caused a 416%
decrease in ODC activity in spe2∆ cells, but only a 15% decrease in meu1∆ cells (Figure 5a).
The double mutant strain had an intermediate phenotype, with a 52% decrease. Differential
response to spermidine was also observed at the protein level as shown by immunoblot analysis
(Figure 5b). In the spe2∆ strain addition of spermidine resulted in undetectable levels of ODC,
while it had no detectable effect on meu1∆ strain. These results show that elevation of ODC
levels in meu1∆ strains and in spe2∆ strains are due to different regulatory systems.
MTA has no effect on ODC protein levels MTA, the substrate of MTAP, is known to be a
11
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from

strong competitive inhibitor of spermine synthase, SAH hydrolase, and other methyltransferases
(38-40). We tested the possibility that elevated MTA levels could cause the elevation of ODC
levels we observed in meu1∆ yeast. We grew cultures of MEU1, meu1∆, and spe2∆ yeast with
and without added 5 mM MTA to media, and examined ODC protein by immunoblot (Figure
5c). We saw no significant increase in ODC levels of any of the three strains by the addition of
exogenous MTA. This result indicates that build-up of MTA is not responsible for the elevated
ODC levels observed in meu1∆ yeast.
Mutations in E1 and E2 homologues effect ODC and polyamine levels – Given the result above,
we reasoned that a salvage pathway metabolite downstream of MTA might play a key role in
repressing ODC levels. To test this hypothesis, we examined ODC levels in yeast strains deleted
for genes similar to two known salvage pathway enzymes from Klebsiella oxytoca. The E1 protein
from Klebsiella catalyzes the conversion of 2,3-diketo-5-methylthio-1-phosphopentene to 1,2-
dihydroxy-3-keto-5-methylthiopentene, while the E2 protein catalyzes the subsequent formation
of 2-keto-4-methylthiobutyrate (see Figure 1)(6,41-43). We searched the S. cerevisiae genome
for ORFs similar to these proteins and found one for each gene. Yeast YEL038W encodes a protein
that is 37% identical to Klebsiella E1, while yeast YMR009W encodes a protein with 26% identity
to E2. We obtained deletions of each of these genes from the Saccharomyces Genome Deletion
Project (28). Deletion strains for each of these proteins were viable and whole cell extracts were
prepared and examined for polyamine levels, ODC activity, and protein levels. E1 and E2 deletion
strains had elevated ODC activity and protein levels compared to the MEU1 control strain (see
12
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from

Figure 3b and Figure 4a). However, neither strain had quite the same level of induction as observed
in the meu1∆ strain. The polyamine profiles also showed an intermediate effect (Figure 3a).
Spermidine levels were clearly elevated in the E1 and E2 mutants, but putrescine was still below the
limit of our detection. These results show that mutations in enzymes downstream of MTAP cause
elevated ODC levels and elevated levels of spermidine.
MTOB repression of ODC in meu1∆ Based on the findings above and our knowledge of the
biochemical pathway, we reasoned that perhaps MTOB was a negative regulator of ODC. We
tested this hypothesis by growing the various mutants in media in which 1 mM MTOB was
substituted for methionine. Yeast grown in MTOB had identical doubling times as yeast grown in
methionine (data not shown). However, immunoblot analysis of ODC clearly shows that the
addition of MTOB dramatically abolishes the elevated ODC levels observed in meu1, E1, and E2
mutants (Figure 6). This finding shows that MTOB represses ODC protein levels.
MTAP and MTOB repress ODC in human tumor cells – Previously we have described the
creation of isogenic MTAP positive and MTAP negative MCF-7 breast adenocarcinoma cell lines,
and showed that MTAP caused a three-fold decrease in total polyamine levels (see Table 3, ref 1).
To see if this repression might be related to ODC repression, we measured ODC activity in two
MTAP expressing and two MTAP non-expressing clones (Figure 7a). Depending on the clones
compared, there is a three to nine-fold drop in ODC activity in the MTAP expressing compared to
the non-expressing cells. These results show that MTAP represses ODC in mammalian cells as
13
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from

well as yeast.
We also examined if exposure to MTOB could repress ODC activity in mammalian cells. For
these experiments we used MCF-7 cells and MIAPaca-2 cells, a pancreatic adenocarcinoma cell
line deleted for MTAP (31). Cells were grown in regular methionine containing media and then
placed in either media with methionine or lacking methionine and supplemented with either 0.2 or 2
mM MTOB. Previously we have shown that MCF-7 cells can utilize MTOB as a source of fixed
sulfur and have identical doubling times on either methionine or MTOB (44). We found similar
growth results with MIAPaca-2 cells (data not shown). After twenty-four hours cells were
harvested and ODC activity was determined (Figure 7b). Cells from both lines treated with either
concentration of MTOB had significant decreases in ODC activity compared to cells growing in 100
µM methionine. These results indicate that MTOB acts as a negative regulator of ODC activity in
mammalian cells.
DISCUSSION
In this report we demonstrate that the deletion of MTAP causes activation of ODC resulting in
elevated polyamine pools. Our data indicates that a specific downstream salvage pathway
metabolite, MTOB, may be a key player in this process. In yeast, mutations in putative salvage
pathway genes located between MTAP and MTOB caused elevated ODC protein and activity
levels. In both MTAP-deleted yeast and human cells, addition of MTOB to the media causes a
substantial reduction in ODC activity. Our studies cannot determine if MTOB is a direct regulator
14
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from

of ODC or whether some metabolite of MTOB is involved. Studies using BAF3 murine lymphoid
cells have demonstrated that MTOB can be converted to malondialdehyde and methional in cell
extracts (45). Interestingly, MTOB and methional at high concentrations can induce apoptosis in
BAF3 cells. It has been shown that drugs that cause depletion of polyamines can sometimes also
cause apoptosis in tumor derived cell lines (46). Thus it is possible that in BAF3 cells, MTOB may
cause apoptosis by depletion of polyamine pools.
We found in S. cerevisiae that mutations in the homologues from Klebsiella E1 and E2 had
more modest effects on ODC activation than MTAP deletion. If MTOB were the only inhibitor of
ODC, one would expect levels to be identical. There are two possible explanations. First, it may be
that there are alternate enzymatic pathways in yeast that can convert methylthioribose-1-phosphate
to MTOB, and thus the E1 and E2 mutants may have some low level of MTOB present. Consistent
with this idea, Dibner et al. reported that methylthioribose-1-phosphate could be converted to 2-
hydroxy-4-methylthiobutanoic acid which could then be converted to MTOB (47). Alternatively,
other metabolites in the pathway may also inhibit ODC levels. For example, MTOB and
methylthioribose-1-phosphate may both be repressors of ODC.
In this paper we show several lines of evidence that the yeast MEU1 gene encodes MTAP,
including amino acid similarity, requirement of MEU1 for enzyme activity, and growth on MTA
containing media. The MEU1 gene was initially identified in a genetic screen designed to isolate
mutations affecting transcriptional regulation of ADH2 (26). Why did MTAP come up in such a
15
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from

screen? Our finding that MTAP deletion alters polyamine homeostasis by overexpression of ODC
suggests a possible explanation. Recent evidence indicates that there is a significant link between
polyamines, enzymes involved in chromatin remodeling, and transcriptional regulation. In yeast it
has been found that mutations that reduce polyamine levels are able to bypass the need for the core
histone deacetylase, GCN5, for the expression of certain genes (48). In ODC over-expressing
transgenic mice both intrinsic histone acetyltransferase (HAT) and deacetylase activities (HDAC )
are elevated (49,50). Alterations in HAT and HDAC activities would be expected to have effects on
a large number of genes (51). Consistent with this hypothesis, microarray analysis of 15,000 ESTs
in isogenic MTAP+ and MTAP- MCF-7 cells indicate that over 200 genes are either two-fold
induced or repressed by MTAP expression (B. Tang and W.D. Kruger, unpublished data).
ODC levels in yeast and mammalian cells are regulated post-transcriptionally. In higher cells,
the antizyme protein targets ODC for degradation by the proteosome (52). Antizyme production
requires a frame-shifting event that is greatly stimulated when polyamines are absent. S. cerevisiae
lacks a canonical antizyme, but appears to have a similar post-transcriptional mechanism for ODC
regulation in response to polyamine depletion (36). Our work here shows that the salvage pathway
products also negatively regulate ODC. Thus ODC is feed-back regulated by both of its
downstream pathways. We did not observe a “hyper” increase in the ODC levels in the double
mutant suggesting that both feedback processes may be affecting the same underlying mechanism,
i.e. disruption of either feedback loop results in elevated ODC.
16
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from

Our findings also support our earlier hypothesis that MTAP acts as a tumor suppressor by
altering polyamine pools. Previously, our lab has shown that expression of MTAP in MTAP
deleted MCF-7 cells suppresses anchorage-independent growth, tumor formation in SCID mice,
and causes significant reduction in polyamine levels (1). This reduction in polyamines is easily
explained by inhibition of ODC by MTAP. We also demonstrated in our previous work that
addition of putrescine could partially restore anchorage-independent growth to MTAP expressing
cells, indicating that at least part of MTAP’s ability to suppress tumorigenicity is due to its affect on
polyamine production. However, it is possible that MTAP deletion in tumor cells may be affecting
other pathways in addition to polyamine biosynthesis that contribute to tumorigenicity. Future
experiments will focus on answering this question.
Acknowledgements – We thank Martin Hoyt and Phil Cofino for the use of their yeast ODC
anti-serum. We also thank Randy Strich, Elizabeth Henske, and Eric Moss for their critical reading
of the manuscript.
17
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from

REFERENCES
1. Christopher, S. A., Diegelman, P., Porter, C. W., and Kruger, W. D. (2002) Cancer Res 62,
6639-6644
2. Della Ragione, F., Russo, G., Oliva, A., Mastropietro, S., Mancini, A., Borrelli, A., Casero,
R. A., Iolascon, A., and Zappia, V. (1995) Oncogene 10, 827-833.
3. Batova, A., Diccianni, M. B., Nobori, T., Vu, T., Yu, J., Bridgeman, L., and Yu, A. L.
(1996) Blood 88, 3083-3090.
4. Nobori, T., Takabayashi, K., Tran, P., Orvis, L., Batova, A., Yu, A. L., and Carson, D. A.
(1996) Proc Natl Acad Sci U S A 93, 6203-6208.
5. Garcia-Castellano, J. M., Villanueva, A., Healey, J. H., Sowers, R., Cordon-Cardo, C.,
Huvos, A., Bertino, J. R., Meyers, P., and Gorlick, R. (2002) Clin Cancer Res 8, 782-787.
6. Wray, J. W., and Abeles, R. H. (1995) J Biol Chem 270, 3147-3153
7. Auvinen, M., Paasinen, A., Andersson, L. C., and Holtta, E. (1992) Nature 360, 355-358
8. Pegg, A. E. (1988) Cancer Res 48, 759-774
9. O’Brien, T. G., Megosh, L. C., Gilliard, G., and Soler, A. P. (1997) Cancer Res 57, 2630-
2637.
10. Toohey, J. I. (1978) Biochem Biophys Res Commun 83, 27-35.
11. Trackman, P. C., and Abeles, R. H. (1981) Biochem Biophys Res Commun 103, 1238-1244
12. Trackman, P. C., and Abeles, R. H. (1983) J Biol Chem 258, 6717-6720
13. Furfine, E. S., and Abeles, R. H. (1988) J Biol Chem 263, 9598-9606
18
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from

14. Myers, R. W., and Abeles, R. H. (1990) J Biol Chem 265, 16913-16921
15. Backlund, P. S., Jr., and Smith, R. A. (1981) J Biol Chem 256, 1533-1535
16. Backlund, P. S., Jr., Chang, C. P., and Smith, R. A. (1982) J Biol Chem 257, 4196-4202
17. Backlund, P. S., Jr., and Smith, R. A. (1982) Biochem Biophys Res Commun 108, 687-695
18. Cone, M. C., Marchitto, K., Zehfus, B., and Ferro, A. J. (1982) J Bacteriol 151, 510-515
19. Marchitto, K. S., and Ferro, A. J. (1985) J Gen Microbiol 131, 2153-2164
20. Schmid, M., Malicki, D., Nobori, T., Rosenbach, M. D., Campbell, K., Carson, D. A., and
Carrera, C. J. (1998) Oncogene 17, 2669-2675.
21. Stadler, W. M., and Olopade, O. I. (1996) Urol Res 24, 239-244
22. Nobori, T., Karras, J. G., Della Ragione, F., Waltz, T. A., Chen, P. P., and Carson, D. A.
(1991) Cancer Res 51, 3193-3197.
23. Cohen, S. S. (1998) in A guide to the polyamines, pp. 296-319, Oxford University Press,
Oxford
24. Megosh, L., Gilmour, S. K., Rosson, D., Soler, A. P., Blessing, M., Sawicki, J. A., and
O’Brien, T. G. (1995) Cancer Res 55, 4205-4209.
25. Thomas, D., Becker, A., and Surdin-Kerjan, Y. (2000) J Biol Chem 275, 40718-40724.
26. Donoviel, M. S., and Young, E. T. (1996) Genetics 143, 1137-1148.
27. Sikorski, R. S., and Hieter, P. (1989) Genetics 122, 19-27
28. Giaever, G., Chu, A. M., Ni, L., Connelly, C., Riles, L., Veronneau, S., Dow, S., Lucau-
Danila, A., Anderson, K., Andre, B., Arkin, A. P., Astromoff, A., El-Bakkoury, M.,
19
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from

Bangham, R., Benito, R., Brachat, S., Campanaro, S., Curtiss, M., Davis, K., Deutschbauer,
A., Entian, K. D., Flaherty, P., Foury, F., Garfinkel, D. J., Gerstein, M., Gotte, D., Guldener,
U., Hegemann, J. H., Hempel, S., Herman, Z., Jaramillo, D. F., Kelly, D. E., Kelly, S. L.,
Kotter, P., LaBonte, D., Lamb, D. C., Lan, N., Liang, H., Liao, H., Liu, L., Luo, C., Lussier,
M., Mao, R., Menard, P., Ooi, S. L., Revuelta, J. L., Roberts, C. J., Rose, M., Ross-
Macdonald, P., Scherens, B., Schimmack, G., Shafer, B., Shoemaker, D. D., Sookhai-
Mahadeo, S., Storms, R. K., Strathern, J. N., Valle, G., Voet, M., Volckaert, G., Wang, C.
Y., Ward, T. R., Wilhelmy, J., Winzeler, E. A., Yang, Y., Yen, G., Youngman, E., Yu, K.,
Bussey, H., Boeke, J. D., Snyder, M., Philippsen, P., Davis, R. W., and Johnston, M. (2002)
Nature 418, 387-391.
29. Sherman, F. (1991) Methods Enzymol 194, 3-21
30. Balasundaram, D., Tabor, C. W., and Tabor, H. (1991) Proc Natl Acad Sci U S A 88, 5872-
5876
31. Chen, Z. H., Olopade, O. I., and Savarese, T. M. (1997) Mol Pharmacol 52, 903-911.
32. Harlow, E., and Lane, D. (1988) Antibodies: a laboratory manual, Cold Spring Harbor
Laboratory, Cold Spring Harbor, NY
33. Tyagi, A. K., Tabor, C. W., and Tabor, H. (1983) Methods Enzymol 94, 135-139.
34. Cohn, M. S., Tabor, C. W., and Tabor, H. (1983) Methods Enzymol 94, 231-234
35. Vujcic, S., Halmekyto, M., Diegelman, P., Gan, G., Kramer, D. L., Janne, J., and Porter, C.
W. (2000) J Biol Chem 275, 38319-38328.
20
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from

36. Gupta, R., Hamasaki-Katagiri, N., White Tabor, C., and Tabor, H. (2001) Proc Natl Acad
Sci U S A 98, 10620-10623.
37. Toth, C., and Coffino, P. (1999) J Biol Chem 274, 25921-25926
38. Williams-Ashman, H. G., Seidenfeld, J., and Galletti, P. (1982) Biochem Pharmacol 31,
277-288.
39. Pegg, A. E., Borchardt, R. T., and Coward, J. K. (1981) Biochem J 194, 79-89.
40. Ferro, A. J., Vandenbark, A. A., and MacDonald, M. R. (1981) Biochem Biophys Res
Commun 100, 523-531.
41. Myers, R. W., Wray, J. W., Fish, S., and Abeles, R. H. (1993) J Biol Chem 268, 24785-
24791
42. Balakrishnan, R., Frohlich, M., Rahaim, P. T., Backman, K., and Yocum, R. R. (1993) J Biol
Chem 268, 24792-24795
43. Dai, Y., Wensink, P. C., and Abeles, R. H. (1999) J Biol Chem 274, 1193-1195
44. Tang, B., Li, Y. N., and Kruger, W. D. (2000) Cancer Res 60, 5543-5547.
45. Quash, G., Roch, A. M., Chantepie, J., Michal, Y., Fournet, G., and Dumontet, C. (1995)
Biochem J 305, 1017-1025.
46. Schipper, R. G., Penning, L. C., and Verhofstad, A. A. (2000) Semin Cancer Biol 10, 55-68
47. Dibner, J. J., Durley, R. C., Kostelc, J. G., and Ivey, F. J. (1990) J Nutr 120, 553-560
48. Pollard, K. J., Samuels, M. L., Crowley, K. A., Hansen, J. C., and Peterson, C. L. (1999)
Embo J 18, 5622-5633.
21
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from

49. Hobbs, C. A., and Gilmour, S. K. (2000) J Cell Biochem 77, 345-360.
50. Hobbs, C. A., Paul, B. A., and Gilmour, S. K. (2002) Cancer Res 62, 67-74.
51. Kuo, M. H., and Allis, C. D. (1998) Bioessays 20, 615-626.
52. Coffino, P. (2001) Nat Rev Mol Cell Biol 2, 188-194
53. Brachmann, C. B., Davies, A., Cost, G. J., Caputo, E., Li, J., Hieter, P., and Boeke, J. D.
(1998) Yeast 14, 115-132
22
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from
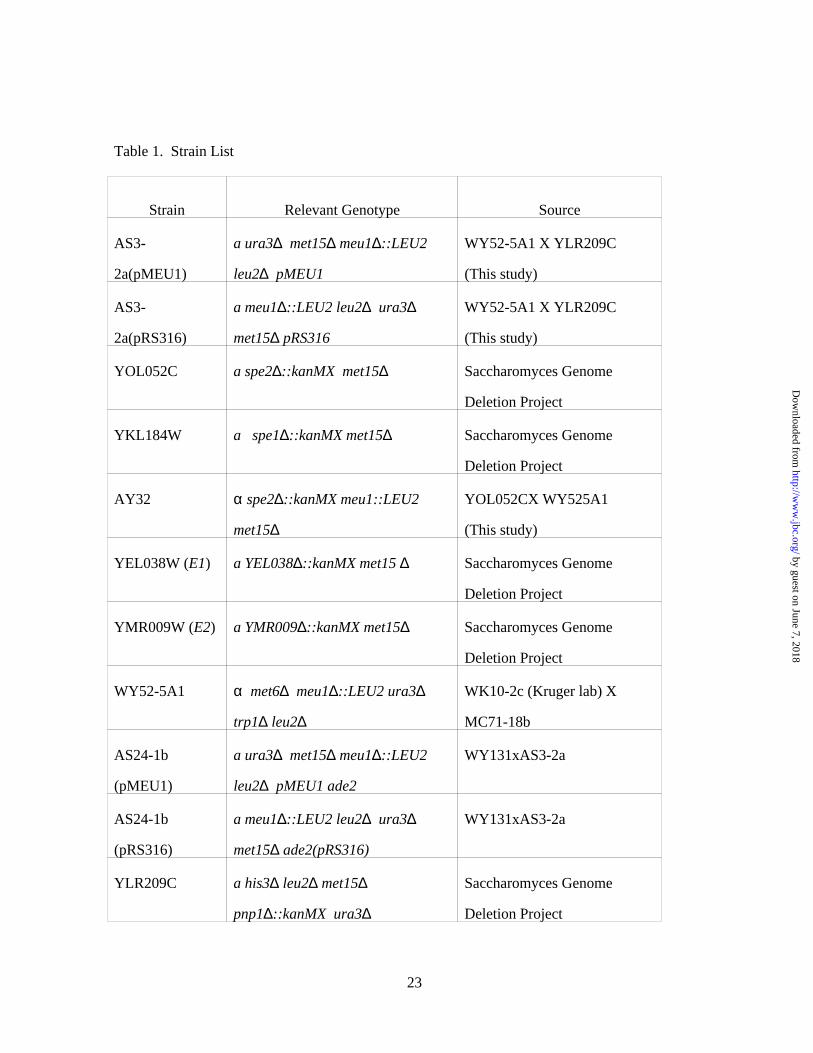
Table 1. Strain List
Strain Relevant Genotype Source
AS3-
2a(pMEU1)
a ura3∆ met15∆ meu1∆::LEU2
leu2∆ pMEU1
WY52-5A1 X YLR209C
(This study)
AS3-
2a(pRS316)
a meu1∆::LEU2 leu2∆ ura3∆
met15∆ pRS316
WY52-5A1 X YLR209C
(This study)
YOL052C a spe2∆::kanMX met15∆ Saccharomyces Genome
Deletion Project
YKL184W a spe1∆::kanMX met15∆ Saccharomyces Genome
Deletion Project
AY32 α spe2∆::kanMX meu1::LEU2
met15∆
YOL052CX WY525A1
(This study)
YEL038W (E1) a YEL038∆::kanMX met15 ∆ Saccharomyces Genome
Deletion Project
YMR009W (E2) a YMR009∆::kanMX met15∆ Saccharomyces Genome
Deletion Project
WY52-5A1 α met6∆ meu1∆::LEU2 ura3∆
trp1∆ leu2∆
WK10-2c (Kruger lab) X
MC71-18b
AS24-1b
(pMEU1)
a ura3∆ met15∆ meu1∆::LEU2
leu2∆ pMEU1 ade2
WY131xAS3-2a
AS24-1b
(pRS316)
a meu1∆::LEU2 leu2∆ ura3∆
met15∆ ade2(pRS316)
WY131xAS3-2a
YLR209C a his3∆ leu2∆ met15∆
pnp1∆::kanMX ura3∆
Saccharomyces Genome
Deletion Project
23
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from

WY131
(ATCC 200869)
α ade2 leu2∆ lys2∆ met15∆ ura3∆ ATCC (Isogenic to strains used
in Genome Deletion Project
see (53)
24
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from

Figure Legends
Figure 1. Methionine salvage and polyamine pathways. Enzymes are shown in italics. Yeast
genes are shown in parenthesis.
Figure 2. Growth of MTAP+ and MTAP- yeast cells on MTA. Yeast strains AS24-1b (MEU1)
and AS24-1b (meu1∆) were diluted to OD600=0.05 in synthetic complete media supplemented
with 5mM MTA and the standard amino acids and nucleotides (SC) except for the ones indicated
at the bottom of the graph. Cells were grown 48 hours and OD600 was determined.
Figure 3. Polyamine and ODC activity in wild-type and salvage pathway mutants.
A) Strains of the indicated genotype were grown in synthetic complete media to an OD of 1.0,
extracted with PCA and polyamines were measured as described in Methods. Putrescine was at
levels below detection limit (<0.5 nmoles/pg) in three of the samples. Error bars indicate
standard error of two measurements. B) ODC activity of the indicated cells were measured in
whole cell extracts as described in Methods. Error bar indicated standard error of four
measurements.
Figure 4. Steady-state ODC protein levels in salvage pathway mutants. A) ODC protein levels
were examined in total extracts made from yeast of the indicated genotype by immunoblot (see
25
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from

Materials and Methods). Yeast ODC runs at 53kD and is indicated by the arrow. The additional
bands below and slightly above ODC are presumably ODC degradation products, as they are
absent in both the wild-type and spe1 controls. B) Comparison of colony size of isogenic wild-
type and meu1∆ yeast grown on polyamine free media after three days of growth.
Figure 5. ODC activity and protein levels in response to spermidine and MTA. A) Yeast strain
AS3-2a (meu1∆), YOL052C (spe2∆), and AY32 (meu1∆ spe2∆) were grown in synthetic
complete polyamine free media in the presence or absence of 0.1 mM spermidine for 24 hours.
Extracts were prepared and analyzed for ODC activity. Error bars indicate standard error of three
measurements. B) Same strains as (A) analyzed for ODC protein levels by immunoblot. C)
Yeast strains AS3-2a (meu1∆), AS3-2a (MEU1), and YOL052C (spe2∆) were grown on
synthetic complete polyamine free media in the presence or absence of 5 mM MTA for 24 hours.
Extracts were prepared and analyzed as above.
Figure 6. Steady-state ODC protein levels in response to MTOB. Yeast strains AS3-2a
(MEU1), AS3-2a (meu1∆), YEL038W (E1∆), and YMR009W (E2∆) were grown in SC media in the
presence or absence or 0.1 mM MTOB for 24 hours. Extracts were prepared and immunoblot
analysis of ODC was preformed as described in Materials and Methods.
Figure 7. ODC activity in mammalian cells. A) MCF-7 cells were stably transfected with
26
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from

either an MTAP expressing construct (MTAP4 and MTAP8) or with a construct that expresses a
mutant non-functional MTAP (D220A4 and D220A6) (1). Cells were grown to 80%
confluency, whole cell extracts were prepared, and ODC activity was measured as described in
Methods. Error bars indicate standard error of three measurements. B) MTAP-deleted MIA
PaCa-2 pancreatic adenocarcinoma cells were plated as equal density and grown for 24 hours in
media containing the indicated levels of methionine or MTOB. Cells were then harvested,
extracts prepared, and ODC activity was measured as described in Methods. Error bars indicate
standard error of three measurements.
27
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from








Markham and Warren D. KrugerAhmad L. Subhi, Paula Diegelman, Carl W. Porter, Baiqing Tang, Zichun J. Lu, George D.
of downstream metabolitesMethylthioadenosine phosphorylase regulates ornithine decarboxylase by production
published online September 23, 2003J. Biol. Chem.
10.1074/jbc.M308451200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on June 7, 2018http://w
ww
.jbc.org/D
ownloaded from