methylation-associated silencing of the thrombospondin...
TRANSCRIPT

[CANCER RESEARCH 63, 6299–6310, October 1, 2003]
Methylation-associated Silencing of the Thrombospondin-1 Gene inHuman Neuroblastoma1
Qi-Wei Yang, Shuqing Liu, Yufeng Tian, Helen R. Salwen, Alexandre Chlenski, Joanna Weinstein, andSusan L. Cohn2
Department of Pediatrics [J. W., S. L. C.] and The Robert H. Lurie Comprehensive Cancer Center [Q-W. Y., S. L., Y. T., H. R. S., A. C.], Feinberg School of Medicine,Northwestern University, Chicago, Illinois 60614
ABSTRACT
Tumor angiogenesis, a major requirement for tumor outgrowth andmetastasis, is regulated by pro- and antiangiogenic factors. Methylation-associated inactivation of the angiogenesis inhibitor thrombospondin-1(TSP-1) has been observed recently in some adult tumors. To investigatethe role of TSP-1 in pediatric cancer, we examined its pattern of expres-sion and mechanisms of regulation in neuroblastoma (NB). TSP-1 wassilenced in a subset of undifferentiated, advanced-stage tumors and NBcell lines. In contrast, most localized tumors expressed this angiogenesisinhibitor, and a significant correlation between morphological evidence ofneuroblast differentiation and TSP-1 expression was observed. Luciferaseassays demonstrated the presence of nuclear factors required for TSP-1transcription in both TSP-1-positive and -negative cell lines, but no cor-relation between TSP-1 promoter activity and the level of TSP-1 mRNAexpression was seen. Our studies indicate that the transcriptional silencingof TSP-1 was caused by methylation. TSP-1 promoter methylation wasdetected in all of the NB cell lines lacking TSP-1 mRNA and in 37% of theNB clinical tumors analyzed. Furthermore, treatment with the demethy-lating agent, 5-Aza-2�-deoxycytidine (5-Aza-dC), restored TSP-1 expres-sion in NB cell lines. Disrupting methylation with 5-Aza-dC also led tosignificant inhibition of NB in vivo and re-expression of TSP-1 in a subsetof NB xenografts. These results suggest that 5-Aza-dC inhibits NB growthby augmenting the expression of TSP-1 along with other genes thatsuppress tumor growth. Demethylating agents may prove to be effectivecandidates for the treatment of children with NB.
INTRODUCTION
NB,3 the most common extracranial solid tumor of childhood, isremarkable for its broad spectrum of clinical behavior (1). Thisclinical diversity correlates closely with numerous clinical and bio-logical factors (1–3), although the biological basis underlying theclinical heterogeneity of NB tumors remains largely unknown. Sev-eral recent studies indicate that angiogenesis plays a critical role in theregulation of NB growth. Most clinically aggressive, advanced-stageNB tumors express high levels of angiogenic stimuli and are highlyvascular (4–7). In contrast, lower numbers of blood vessels are seenin tumors with a more benign behavior. We and others have reportedrecently that favorable histology tumors with abundant Schwannianstroma express high levels of angiogenesis inhibitors (8–10). Addi-tional laboratory studies have shown that changes in the balance of
angiogenic activators and inhibitors directly impacts NB tumorigenic-ity. Down-regulation of angiogenesis inhibitors is observed in highlymalignant NB cells with exogenous MYCN overexpression (11, 12),whereas angiogenic activators are down-regulated in Trk-A trans-fected NB cells that display impaired tumorigenicity (13). Further-more, administration of antiangiogenesis agents effectively inhibitsNB tumor growth in vivo (14–18).
TSP-1, a matrix-bound adhesive glycoprotein, is a potent inhibitorof angiogenesis (19). It is able to block endothelial cell migration inresponse to a wide variety of angiogenic stimuli, induce endothelialcell apoptosis, and prevent neovascularization in the rat cornea andpolyvinyl sponge assays (20–22). Transformed cells and tumor celllines usually express very low levels of TSP-1, and tumorigenicity issuppressed when TSP-1 is re-expressed in these cell lines (23–26).TSP-1 has also been shown to have antiangiogenic properties inbladder, melanoma, lung, and breast cancer (27, 28). In some cancers,expression of TSP-1 has been reported to inversely correlate withmalignant progression (28). Conversely, high plasma levels of TSP-1are associated with increased angiogenesis in colon cancer (29),suggesting that in some tumors TSP-1 does not have an inhibitoryeffect.
The mechanisms controlling the expression of TSP-1 are not com-pletely understood. A number of cis-acting transcription elements inthe TSP-1 promoter have been identified (30). Additional studiesindicate that the tumor suppressor p53 positively regulates the TSP-1gene (31). Alterations in DNA methylation also appear to play a rolein TSP-1 regulation, and promoter-associated CpG island methylationhas been implicated recently in the transcriptional inactivation ofTSP-1 in glioblastomas (32). Epigenetic mechanisms have similarlybeen shown to be responsible for the silencing of other tumor sup-pressor genes in a variety of human cancers (33). Chromatin structureis a dominant force in methylation-associated gene silencing (34). Themethyl-CpG-binding protein MeCP2 has been shown to recruit his-tone deacetylase activity to in vitro methylated promoters (34, 35)indicating that these two processes are linked. Thus, it may be nec-essary to simultaneously block both DNA methylation and histonedeacetylation to achieve maximal reactivation of genes silenced bymethylation (36).
In this study, we examined the level of TSP-1 expression and itsmechanisms of regulation in NB. TSP-1 was silenced in a subset ofNB cell lines, and the loss of expression was associated with aberrant5� CpG island methylation of the TSP-1 promoter. TSP-1 promotermethylation was also detected in 37% of the NB clinical tumorsanalyzed. Treatment with the demethylating agent 5-Aza-dC restoredTSP-1 transcription in the TSP-1 negative NB cell lines, confirmingthat the silencing of this gene in the cell lines was because ofmethylation. Furthermore, disrupting methylation with 5-Aza-dC re-sulted in inhibited tumor growth in vivo, and TSP-1 expression wasrestored in a subset of NB xenografts. These results demonstrate that5-Aza-dC augments the expression of TSP-1 and other genes thatimpair NB tumor growth. Demethylating agents may prove to beeffective candidates for the treatment of children with NB.
Received 4/11/03; revised 7/7/03; accepted 7/18/03.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1 Supported in part by the Neuroblastoma Children’s Cancer Society, Friends forSteven Pediatric Cancer Research Fund, the Elise Anderson Neuroblastoma ResearchFund, the North Suburban Medical Research Junior Board, and the Robert H. LurieComprehensive Cancer Center, NIH, National Cancer Institute Core Grant 5P30CA60553.
2 To whom requests for reprints should be addressed, at Children’s Memorial Hospital,Division of Hematology/Oncology, 2300 Children’s Plaza, Chicago, IL 60614. Phone:(773) 880-4562; Fax: (773) 880-3053; E-mail: [email protected].
3 The abbreviations used are: NB, neuroblastoma; TSP-1, thrombospondin-1; 5-Aza-dC, 5-Aza-2�-deoxycytidine; TSA, Trichostatin A; CM, conditioned medium; MSP,methylation-specific PCR; RT-PCR, reverse transcription-PCR; BrdUrd, 5-bromo-2�-deoxyuridine; RA, retinoic acid; �2-MG, �2-microglobulin; GNR, ganglioneuroma; GNB,ganglioneuroblastoma; MVD, microvascular density; PI, propidium iodide; N-type, neu-ronal; S-type, Schwannian.
6299
on May 6, 2018. © 2003 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

MATERIALS AND METHODS
Cell Culture and CM Collection. The cell lines used in this study havebeen described previously (37–40), and their biological characteristics aresummarized in Table 1 (38–40). Cells were grown at 5% CO2 in RPMI 1640(Invitrogen, Carlsbad, CA) supplemented with 10% heat-inactivated fetalbovine serum, L-glutamine, and antibiotics. In some experiments, 6 �M BrdUrd(Sigma, St. Louis, MO) or 10 �M RA (Sigma) was added to the medium. Cellswere harvested at times ranging from 3 h to 28 days of treatment with BrdUrdor RA. CM were collected and concentrated 20-fold using Centricon-3 con-centrators (Millipore, Bedford, MA) as described previously (9).
TSP-1 Expression in NB Cell Lines and CM. TSP-1 expression in NBcell lines was detected using RT-PCR. RNA, isolated from NB cell lines, andtreated with RQ1 DNase (Promega, Madison, WI; 1 unit/�l) for 1 h at 37°C.Total RNA (2.5 �g) was reverse-transcribed in a final volume of 20 �l, and 1�l of the diluted reaction mixture was subsequently amplified by PCR. Acoamplification reaction was performed with TSP-1 sense and antisense prim-ers, 5�-GTGACAACTGTCCCTACAACCAC-3� and 5�-CCATATCAGT-GTCTCTCTGGTCC-3�, respectively, and �2-MG sense and antisense prim-ers, 5�-ACCCCCACTGAAAAAGATGA-3� and 5�-ATCTTCAAACCTCC-ATGATG-3�, respectively, for 35 cycles. Each cycle consisted of denaturationat 94°C for 30 s, annealing at 55°C for 30 s, and extension at 72°C for 45 s.The PCR products were subjected to 2% agarose gel electrophoresis, and thelevel of �2-MG was used as an internal control. In some experiments, TSP-1mRNA expression was also analyzed by real-time quantitative RT-PCR usingmethods described previously (41). For these studies, the TSP-1 target primerpair 5�-TGGAACTATGGGCTTGAGAAAAC-3� and 5�-CACTGATGCA-AGCACAGAAAAGA-3� was used with the probe carboxyfluorescein-CCCAGGATCACTTCTCCTTGGCTTCC-QSY7. The level of TSP-1 proteinin CM was examined by Western blot analysis with a mouse monoclonalanti-TSP-1 antibody (1:1000 dilution; NeoMarkers, Fremont, CA) using meth-ods described previously (8).
Immunohistochemical Studies. Histological sections of human NB tumortissue and mouse xenografts were fixed in 10% buffered formalin, embeddedin paraffin, and immunostained using a mouse anti-TSP-1 monoclonal anti-body. Cell pellets, prepared from 1.5 � 107 cells from each NB cell lineanalyzed, were prepared similarly. Four-�m thick sections were rehydrated ingraded alcohols and rinsed in PBS. Antigen retrieval was performed with 0.01
M citrate buffer (pH 6.0) in a boiling steamer for 20 min. Sections wereincubated with a 1:100 dilution of primary antibody in a humidity chamberovernight at 4°C, and developed with peroxidase labeled-dextran polymerfollowed by diaminobenzidine (DAKO Envision Plus System; DAKO Corpo-ration, Carpinteria, CA). Sections were counterstained with Gill’s Hematoxylin(Fisher, Pittsburgh, PA). For negative controls, primary antibody was omitted.TSP-1 staining above the background in the tumor cytoplasm was scored aspositive. The correlation of TSP-1 expression with clinical data includingstage, MYCN amplification, and morphological evidence of differentiation wasstatistically evaluated using Fisher’s Exact Test. Differentiation was evaluatedusing the published criteria of the International Neuroblastoma PathologyClassification System (42). Tumors with �5% of the tumor cells showingsynchronous differentiation of the nucleus and conspicuous, eosinophilic, oramphophilic cytoplasm (i.e. showing differentiation toward ganglion cells)were classified as differentiating, whereas stroma-rich, intermixed tumors wereconsidered differentiated. Correlation with survival was analyzed with Kaplan-Meier curves. This study was approved by the Children’s Memorial MedicalCenter Institutional Review Board.
To highlight endothelial cells in the NB xenografts, immunohistochemicalstaining with an anti-PECAM-1 (CD31) antibody (Santa Cruz BiotechnologyInc., Santa Cruz, CA) was performed on formalin-fixed and paraffin-embeddedtumor sections. MVD was quantified by counting 10 consecutive fields at�600 magnification, and the average MVD counted in the 10 fields wasconverted into MVD/mm2. The Student t test was used to statistically comparethe MVD in the 5-Aza-dC-treated and control xenograft tumors.
Construction of Human TSP-1 Reporter Plasmids. Fragments of theTSP-1 promoter were amplified using genomic DNA isolated from SH-EPcells as the template. The PCR cycle included denaturation at 94°C for 2 minfollowed by 35 cycles of denaturation at 94°C for 30 s, annealing at 60°C for30 s, extension at 72°C for 45 s, and final extension at 72°C for 10 min.Products of the PCR reaction were digested with SacI/NcoI, low-meltingagarose gel purified, and cloned into the pGL3-Enhancer vector (Promega),which was digested with the same enzymes. The sense primer sequences usedto construct the reporter plasmids were as follows: pTSP-LUC-1.7: 5�-GGGT-GAGCTCTGTAAATAGCTGAAGACTCTGG-3�; pTSP-LUC-1.1: 5�-GGG-TGAGCTCTGTAAATAGCTGAAGACTCTGG-3�; pTSP-LUC-0.63: 5�-GG-CAGAGCTCTAACGAATGGCTCTCTTGG-3�; pTSP-LUC-0.42: 5�-CCCCG-
Table 1 Biological characteristics of NB cell lines
Cell line Subclone N or S type MYCN amplificationTumorigenic
in vivo TSP-1 mRNA TSP-1 protein expression TSP-1 protein secretion
SK-N-SH No / No � � �/� nda
SMS-KCNR No / Yes � � � �NMB No / Yes � � nd �IMR-5 No / Yes � � nd �NBL-S No / No � � �/� �SK-N-BE(1) No / Yes un � nd ndSK-N-BE(2) No / Yes � � � ndNGP No / Yes � � � ndNBL-W-N Yes N Yes � � � �NBL-W-S Yes S Yes � � � �LA1-55n Yes N Yes � � � �LA1-5s Yes S Yes � � nd �SH-SY5Y Yes N No � � � �SH-EP Yes S No � � � �a nd, not done; un, unknown.
Table 2 TSP-1 promoter constructs and its activity
Constructs Start End Intron1 Distal SP1 Proximal SP1 CAAT box TATA box Relative activity (%)
pTSP-LUC-1.7 �954 �783 � � � � � 92pTSP-LUC-1.1 �954 �147 � � � � � 93pTSP-LUC-0.63 �484 �147 � � � � � 85pTSP-LUC-0.42 �273 �147 � � � � � 83pTSP-LUC-0.3 �149 �147 � � � � � 100pTSP-LUC-0.24 �93 �147 � � � � � 52pTSP-LUC-0.2 �52 �147 � � � � � 11pTSP-LUC-0.13 �21 �147 � � � � � 5pTSP-LUC-SP1-Mut �134 �147 � � a � � 74pTSP-LUC-CAAT-Mut �75 �147 � � � a � 15
a Proximal SP1 binding site or CAAT box are mutated.
6300
METHYLATION OF TSP-1 IN NEUROBLASTOMA
on May 6, 2018. © 2003 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

AGCTCGCGTGGCGCAAGAGTAC-3�; pTSP-LUC-0.3: 5�-TTTCGAGCTCG-AAAGTTGCGCGCCAGGC-3�; pTSP-LUC-0.24: 5�-CCCCGAGCTCCCGC-TTCCTGCCCGGC-3�; pTSP-LUC-0.2: 5�-GCCGGAGCTCTCCCCAGGAAT-GCGAG-3�; pTSP-LUC-0.13: 5�-GCCCGAGCTCGCCTGCGAGTTCAG-3�;pTSP-LUC-SP1-Mut: 5�-TTGCGAGCTCGGCAGCGGGGTTCGGAGAGAG-3�; and pTSP-LUC-CAAT-Mut: 5�-TCCTGAGCTCCCGCCGCCCGGTGGC-CGGAG-3�. The antisense primer 5�-CTCCGGTACACCATGGTAAGTCG-CCC-3� was used for all of the constructs except pTSP-LUC-1.7 (5�-CTA-GCGTCCTGTTCCTGATGCAT-3�). The location of primers in the TSP-1 pro-moter region is shown in Fig. 2 and Table 2.
Luciferase Assays. Cells were seeded in a 24-well dish and grown to 90%confluence in growth medium. For each well, 0.5 �g of reporter construct wascotransfected with 25 ng of Renilla luciferase plasmid, pRL-CMV (Promega),into NB cell lines using Lipofectamine2000 (Invitrogen). Cells were harvested36 h after transfection, and reporter activity was measured using the DualLuciferase Assay (Promega) according to the manufacturer’s instructions.Experimental luciferase activities were normalized for efficiency against con-trol Renilla luciferase readings.
Analysis of TSP-1 Gene Hypermethylation by MSP and Bisulfite DNASequencing. Genomic DNA was isolated from NB cell lines and tumors bystandard techniques (38) and modified by sodium bisulfite using the CpGe-nome DNA Modification kit (Serologicals, Atlanta, GA). Briefly, 1 �g ofgenomic DNA was denatured with NaOH and modified by sodium bisulfite,which converts all of the unmethylated cytosines to uracils, whereas methyl-ated cytosines remain unchanged. The modified DNA was desulfonated withNaOH and purified. A pair of primers, 5�-GAATGTGAGTGTTTTTTTA-
AATGTG-3� and 5�-CCTAAACTCACAAACCAACTCAA-3�, was designedfor the unmethylated sequence of the TSP-1 promoter region around thetranscription start site. Another pair of primers, 5�-TGCGAGCGTTTTTTTA-AATGC-3� and 5�-TAAACTCGCAAACCAACTCG-3�, was used for the
Fig. 1. Expression of TSP-1 in NB cell lines and tumors. A, RT-PCR analysis of TSP-1 mRNA expression. Higher levels of expression were observed in nontumorigenic S-typeNB subclones than in tumorigenic N-type subclones. Expression of the housekeeping gene �2-MG was used as an internal control. B, Western blot analysis of TSP-1 protein expressionin NB cell line CM. TSP-1 protein was only detected in nontumorigenic S-type NB subclones. C, immunohistochemistry staining for TSP-1 in differentiated/differentiating andundifferentiated NB tumor samples. Top panel, GNB with TSP-1 expressed strongly in differentiated/differentiating NB cells. Middle panel, differentiating NB with TSP-1 expressionin NB cells with evidence of differentiation. Bottom panel, undifferentiated NB with undetectable TSP-1 expression (magnification, �400). D, real-time quantitative RT-PCR of TSP-1in SMS-KCNR NB cells induced to differentiate with RA or BrdUrd at indicated time points. Enhanced levels of TSP-1 mRNA were detected in cells induced to differentiate.
Table 3 Clinical and biological characteristics of NB patientsa
CharacteristicCases
nTSP-1
positive n (%)No. TSP-1
negative n (%)
INSS Tumor Stageb
Localized (stages 1 and 2A) 9 7 (78) 2 (22)Regional/advanced (stages 2B, 3, and 4) 28 14 (50) 14 (50)
Age�1 year 13 8 (62) 5 (38)�1 year 24 13 (54) 11 (46)
MYCNAmplified 7 4 (57) 3 (43)Nonamplified 29 15 (52) 14 (48)Unknown 1 1 (100) 0 (0)
Neuroblast differentiationYes 21 18 (86) 3 (14)No 16 3 (19) 13 (81)
Outcomea
Alive (NED) 31 19 (61) 12 (39)Alive (with disease) 2 1 (50) 1 (50)Dead 4 1 (25) 3 (75)
Total 37 21 (57) 16 (43)a Median follow-up 69 months (range, 18–140 months).b INSS, International Neuroblastoma Staging System; NED, no evidence of disease.
6301
METHYLATION OF TSP-1 IN NEUROBLASTOMA
on May 6, 2018. © 2003 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
methylated sequence of the same region. Initial denaturation at 94°C for 3min was followed by 35 cycles of denaturation at 94°C for 30 s, annealingat 56°C for methylated or 54°C for the unmethylated reaction for 30 s,extension at 72°C for 45 s, and final extension at 72°C for 8 min. The PCRproducts were separated by electrophoresis on a 2.5% agarose gel andvisualized under UV illumination using ethidium bromide staining. Uni-versal Methylated DNA (Serologicals), which is enzymatically methylated
human genomic DNA, was used as a positive control for MSP. ForDNA sequencing, PCR products were gel-purified and cloned into thepCR2.1-TOPO vector (Invitrogen) according to the manufacturer’s proto-col. Plasmid DNA was purified with the QIA-prep Spin Mini Prep kit(Qiagen, Chatsworth, CA). Individual plasmids were then sequenced usingthe ABI PRISM 377 DNA sequencer (Applied Biosystems, FosterCity, CA).
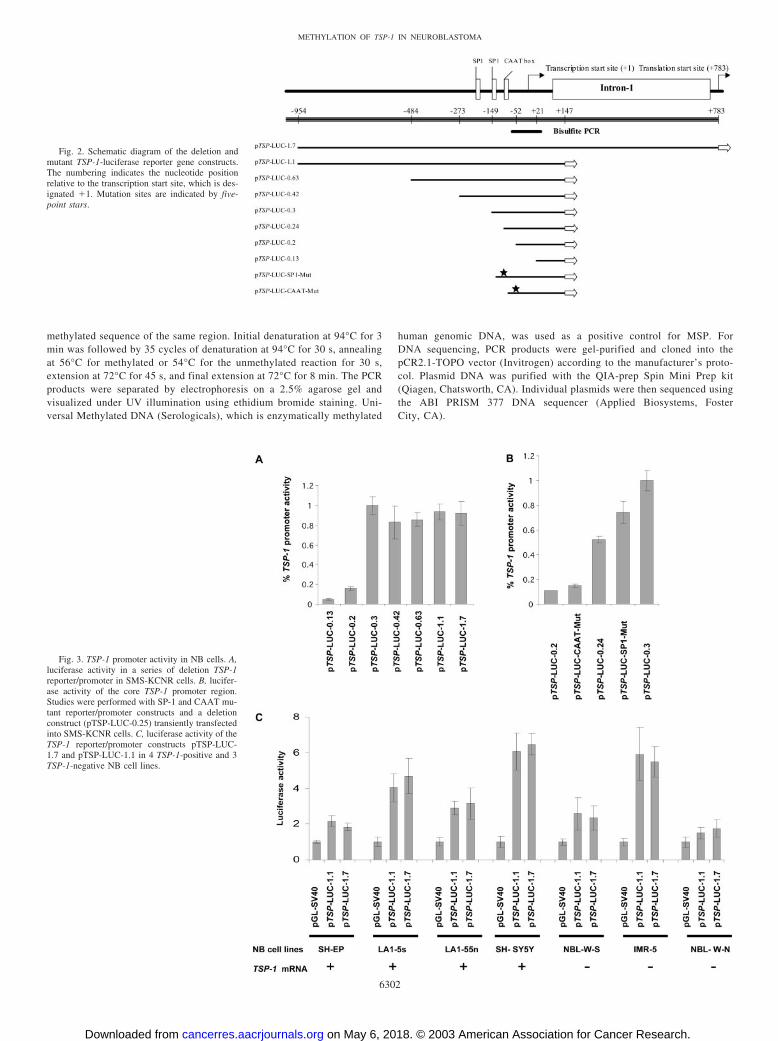
Fig. 2. Schematic diagram of the deletion andmutant TSP-1-luciferase reporter gene constructs.The numbering indicates the nucleotide positionrelative to the transcription start site, which is des-ignated �1. Mutation sites are indicated by five-point stars.
Fig. 3. TSP-1 promoter activity in NB cells. A,luciferase activity in a series of deletion TSP-1reporter/promoter in SMS-KCNR cells. B, lucifer-ase activity of the core TSP-1 promoter region.Studies were performed with SP-1 and CAAT mu-tant reporter/promoter constructs and a deletionconstruct (pTSP-LUC-0.25) transiently transfectedinto SMS-KCNR cells. C, luciferase activity of theTSP-1 reporter/promoter constructs pTSP-LUC-1.7 and pTSP-LUC-1.1 in 4 TSP-1-positive and 3TSP-1-negative NB cell lines.
6302
METHYLATION OF TSP-1 IN NEUROBLASTOMA
on May 6, 2018. © 2003 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

Treatment of NB Cells with 5-Aza-dC and TSA. Cells were seeded,allowed to attach over a 24-h period, and treated for times ranging from 6 h to5 days with the demethylating reagent 5-Aza-dC (Sigma) at a final concen-tration of 0.1, 1, or 5 �M, or a specific inhibitor of histone deacetylase TSA(Sigma) at 0.25 or 0.5 �M. For the combination 5-Aza-dC/TSA treatment, 1�M 5-Aza-dC was added for a range of times after which 0.25 or 0.5 �M TSAwas added for an additional 24 h. DMSO (Sigma) was used as a control fornonspecific solvent effects on cells. At the end of the treatment period, themedium was removed, and the RNA was extracted or the CM were prepared.
Proliferation Assay. NBL-W-S, NMB, and NBL-W-N cells were seededinto 96-well plates at a density of 5.0 � 103, 7.5 � 103, and 7.5 � 103,respectively. After 24 h, 5-Aza-dC was added at various concentrations to eachquadruplicate well. After 72-h treatment, 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxy-phenyl)-2-(4-sulfonyl)-2H-tetrazolium labeling mixture(Promega) was added, and cells were additionally incubated for 3 h. Theabsorbance of the samples was measured using a Bio-Kinetics MicroplateReader (Bio-Tek Instruments, Winooski, VT).
Measurement of Cell Cycle Phase Distribution. Cell cycle distributionwas determined by flow cytometric analysis. Briefly, NB cell lines NMB,
Fig. 4. Methylation status of the TSP-1 gene in NB cell lines. A,methylation status of TSP-1 was characterized in TSP-1-positiveand -negative NB cell lines using MSP. Universal methylated DNAwas used as a positive control. B, methylation status of plasmidclones bearing MSP products derived from the genomic DNA of 6NB cell lines was characterized. TSP-1-MSP products have 5 CpGsites (�18, �16, �7, �5, and �10 from transcription start site).Each row of circles represents a single sequenced plasmid contain-ing cloned MSP product of bisulfite-treated genomic DNA. Allcytosines in the 5 CpG islands were methylated in TSP-1 negativecell lines, and cytosines in the 5 CpG islands were unmethylated inTSP-1 positive cell lines. F indicates methylated cytosine; E in-dicates unmethylated cytosine. C, representative sequencing of theMSP product in NBL-W-N cells. Arrows indicate methylatedcytosines.
Fig. 5. Methylation status of TSP-1 in NB patient samples. A, MSP of TSP-1 inrepresentative samples of NB tumors. B, MSP of TSP-1 in representative samples of GNRand GNB tumors.
6303
METHYLATION OF TSP-1 IN NEUROBLASTOMA
on May 6, 2018. © 2003 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

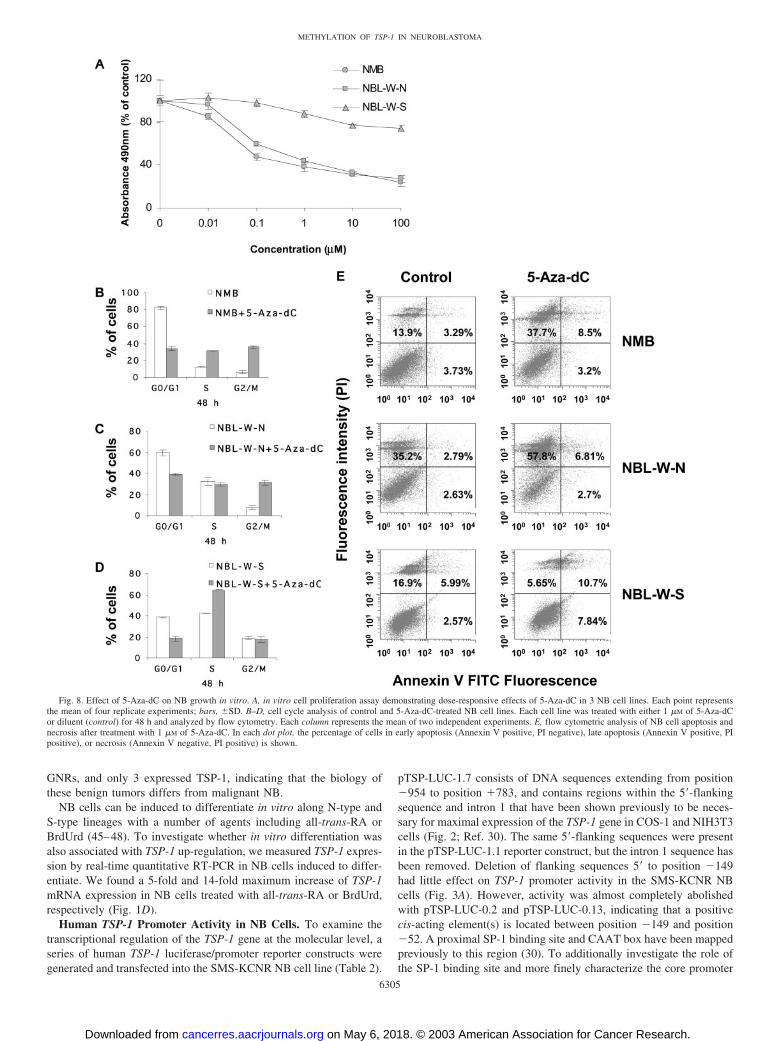
NBL-W-N, and NBL-W-S were cultured in RPMI 1640 containing 1 �M of5-Aza-dC for 48 h. Control cells were cultured in medium lacking 5-Aza-dC.Cells were then washed with PBS, fixed in 70% ethanol, and hypotonicallylysed in 1 ml of DNA staining solution [0.05 mg/ml PI (Sigma) and 0.1%Triton X-100]. The cells were incubated, while protected from light, at 4°Covernight before analysis. The cell cycle data were analyzed with an EpicsXL-MCL flow cytometer (Beckman Coulter, Miami, FL), with System II(version 3.0) software (Beckman Coulter). Additional analysis of cell cycledistribution was determined by using Modfit LT (Verity Software House,Topsham, ME).
Annexin V/PI Staining. To investigate whether treatment with 5-Aza-dCinduced NB cells to undergo apoptosis and/or necrosis, two-color flow cyto-metric analysis using fluorescein-labeled Annexin V and PI was performed(43). Cells were fluorescein-labeled using methods provided by the manufac-tures of the Annexin V-FITC Apoptosis Detection kit (Immunotech, Marseille,France). In brief, cultured NB cell lines were treated with 1 �M 5-Aza-dC forthe indicated times. Cells were washed twice with PBS, and resuspended at1 � 106 cells/ml in binding buffer with FITC-conjugated Annexin V and PI.Samples were analyzed using the Epics XL-MCL flow cytometer.
NB Xenograft Studies. Female 4–6 week old homozygous athymic nudemice (Harlan, Madison, WI) were inoculated s.c. into the right flank with1 � 107 cells from the NBL-W-S, NBL-W-N, or NMB NB cell lines. Oncetumors were palpable, mice were treated with three doses of 5-Aza-dC (5mg/kg/dose) in 1 day with 3-h intervals. For the NBL-W-S cell line, 15 micewere used in the control group, and 25 mice were used for treatment. For theNBL-W-N cell line, 15 mice were used as controls, and 16 mice were used fortreatment. For the NMB cell line, 15 mice were used for the control group, and15 mice were used for treatment. Tumor volumes were measured twice a weekfor up to 12 days after 5-Aza-dC administration and calculated using theformula: tumor volume � (length � width)2/2 (44). Mice were sacrificed at 6,9, or 12 days after 5-Aza-dC treatment to analyze TSP-1 re-expression andMVD at various time points post-treatment. The Student t test was used tocompare tumor size and MVD in the control and treatment groups. Animalswere treated according to NIH guidelines for animal care and use, andprotocols were approved by the Animal Care and Use Committee at North-western University.
RESULTS
Analysis of TSP-1 Expression in NB Cell Lines and ClinicalTumors. TSP-1 was silenced in 5 of 14 NB cell lines analyzed byRT-PCR (Fig. 1A; Table 1). Three pairs of NB subclones wereexamined, and higher levels of TSP-1 were seen in the nontumori-
genic S-type LA1-5s and SH-EP cells compared with tumorigenicN-type LA1-55n and SH-SY5Y cells. However, TSP-1 was silencedin both NBL-W-N and NBL-W-S cells, N-and S-type subclones of theNBL-W NB cell line (37). Despite sharing morphological and bio-chemical features of other S-type NB subclones, NBL-W-S are capa-ble of forming tumors in nude mice (see below). TSP-1 protein wasdetected in the cytoplasm of SH-EP cells by immunohistochemistry.NBL-S and SK-N-SH had weak staining for TSP-1, whereas no TSP-1protein expression was observed in the other 7 NB cell lines tested(Table 1). To investigate whether NB cells secrete TSP-1 protein, CMwere collected from 10 NB cell lines and subjected to Western blotanalysis. As shown in Fig. 1B, TSP-1 was not secreted by most of theNB cell lines. High levels of TSP-1 protein were only present inmedium collected from the S-type SH-EP cells, whereas low levels ofprotein were secreted by LA1-5s S-type subclone. Thus, TSP-1 ap-pears to be transcriptionally silenced in some of the cell lines, whereasin others TSP-1 seems to be post-transcriptionally regulated. The lackof TSP-1 in medium collected from the NBL-S and SK-N-SH cells,cell lines with low levels of cytoplasmic TSP-1 protein, suggests thatthe level of protein secretion may be below the level of detection byWestern blot or that protein secretion is blocked.
To investigate whether TSP-1 was also silenced in NB tumors,histological sections from 37 NB clinical samples were stained withantibody against human TSP-1. The frequency of TSP-1 expressionwas somewhat higher in low-stage tumors (Table 3). Seven of 9localized tumors expressed TSP-1 compared with 14 of 28 regional-and advanced-stage tumors (P � 0.20). No correlation was detectedbetween TSP-1 expression and MYCN amplification. Four of the 7MYCN-amplified tumors expressed TSP-1, compared with 15 of the30 tumors with single copy MYCN (P � 0.99). In addition, TSP-1expression was not associated with survival in this small cohort. Of 4deceased patients, 1 had a TSP-1-positive tumor, and 3 had TSP-1-negative tumors. However, TSP-1 expression did correlate with tumordifferentiation. Of the 21 tumors with morphological evidence ofneuroblast differentiation, TSP-1 was detected in 18 (86%). In con-trast, only 3 of 16 (19%) undifferentiated NB tumors expressed TSP-1(P � 0.01; Fig. 1C). TSP-1 expression was also analyzed in 12 mature
Fig. 6. Restoration of TSP-1 gene expression after treatment with 5-Aza-dC. A, cellswere treated with DMSO (vehicle), or 1 or 5 �M of 5-Aza-dC for 60 h. Re-expressedTSP-1 was detected by RT-PCR analysis. B, time-dependent re-expression of TSP-1 inNBL-W-S cells exposed to 1 �M of 5-Aza-dC at indicated time points. C, Western blotanalysis of TSP-1 expression in CM collected from NBL-W-S after treatment with5-Aza-dC at the indicated times.
Fig. 7. Re-expression of TSP-1 mRNA after treatment with TSA and 5-Aza-dC in NBcells. A, expression of TSP-1 mRNA was measured by RT-PCR in NBL-W-N, NMB, andIMR-5 cell lines after treatment with TSA alone or cotreatment with TSA and 5-Aza-dCat the indicated time points. B, RT-PCR analysis of TSP-1 mRNA expression in NBL-W-Scells after treatment with TSA with or without 5-Aza-dC for the indicated times.
6304
METHYLATION OF TSP-1 IN NEUROBLASTOMA
on May 6, 2018. © 2003 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
GNRs, and only 3 expressed TSP-1, indicating that the biology ofthese benign tumors differs from malignant NB.
NB cells can be induced to differentiate in vitro along N-type andS-type lineages with a number of agents including all-trans-RA orBrdUrd (45–48). To investigate whether in vitro differentiation wasalso associated with TSP-1 up-regulation, we measured TSP-1 expres-sion by real-time quantitative RT-PCR in NB cells induced to differ-entiate. We found a 5-fold and 14-fold maximum increase of TSP-1mRNA expression in NB cells treated with all-trans-RA or BrdUrd,respectively (Fig. 1D).
Human TSP-1 Promoter Activity in NB Cells. To examine thetranscriptional regulation of the TSP-1 gene at the molecular level, aseries of human TSP-1 luciferase/promoter reporter constructs weregenerated and transfected into the SMS-KCNR NB cell line (Table 2).
pTSP-LUC-1.7 consists of DNA sequences extending from position�954 to position �783, and contains regions within the 5�-flankingsequence and intron 1 that have been shown previously to be neces-sary for maximal expression of the TSP-1 gene in COS-1 and NIH3T3cells (Fig. 2; Ref. 30). The same 5�-flanking sequences were presentin the pTSP-LUC-1.1 reporter construct, but the intron 1 sequence hasbeen removed. Deletion of flanking sequences 5� to position �149had little effect on TSP-1 promoter activity in the SMS-KCNR NBcells (Fig. 3A). However, activity was almost completely abolishedwith pTSP-LUC-0.2 and pTSP-LUC-0.13, indicating that a positivecis-acting element(s) is located between position �149 and position�52. A proximal SP-1 binding site and CAAT box have been mappedpreviously to this region (30). To additionally investigate the role ofthe SP-1 binding site and more finely characterize the core promoter
Fig. 8. Effect of 5-Aza-dC on NB growth in vitro. A, in vitro cell proliferation assay demonstrating dose-responsive effects of 5-Aza-dC in 3 NB cell lines. Each point representsthe mean of four replicate experiments; bars, �SD. B–D, cell cycle analysis of control and 5-Aza-dC-treated NB cell lines. Each cell line was treated with either 1 �M of 5-Aza-dCor diluent (control) for 48 h and analyzed by flow cytometry. Each column represents the mean of two independent experiments. E, flow cytometric analysis of NB cell apoptosis andnecrosis after treatment with 1 �M of 5-Aza-dC. In each dot plot, the percentage of cells in early apoptosis (Annexin V positive, PI negative), late apoptosis (Annexin V positive, PIpositive), or necrosis (Annexin V negative, PI positive) is shown.
6305
METHYLATION OF TSP-1 IN NEUROBLASTOMA
on May 6, 2018. © 2003 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

elements, an additional deletion construct, pTSP-LUC-0.24, wasgenerated that included the CAAT box but lacked the SP-1 bindingsite. In addition, two constructs, pTSP-LUC-SP1-Mut and pTSP-LUC-CAAT-Mut containing mutations in the SP1 binding site orCAAT box, respectively, were generated and transfected into SMS-KCNR NB cells. Activity was decreased 50% with construct pTSP-LUC-0.24 compared with pTSP-LUC-0.3. Mutation of the SP-1 bind-ing site was associated with a 25% decrease promoter activitycompared with pTSP-LUC-0.3, whereas mutation of the CAAT boxabolished promoter activity (Fig. 3B). Thus, the CAAT box is requiredfor basal transcription of TSP-1, whereas the adjacent SP-1 site hasonly a moderate effect on TSP-1 expression in NB cells.
To determine whether the disparity in the levels of TSP-1 expres-sion in the NB cell lines was consequent to differences in activity ofthe TSP-1 promoter, TSP-1 reporter constructs were transfected into 7NB cell lines expressing a range of TSP-1 mRNA levels, and lucif-erase assays were performed. We found no correlation between pro-moter activity and TSP-1 mRNA expression in the 7 NB cell linesanalyzed (Fig. 3C). Relatively low levels of TSP-1 promoter activitywere seen in SH-EP cells, which express high levels of TSP-1 mRNA,and high levels of TSP-1 promoter activity were seen in IMR-5 cells,
which express very low to undetectable levels of TSP-1 mRNA. Foreach cell line examined, similar levels of promoter activity were seenwith pTSP-LUC-1.7 and pTSP-LUC-1.1 indicating that cis-actingelements within intron-1 do not play a major regulatory role in thepromoter activity in NB cells (Fig. 3C).
Methylation and Silencing of the TSP-1 Promoter in NB CellLines and Tumors. The luciferase assays demonstrated that nuclearfactors required for TSP-1 transcription were present in both TSP-1-positive and -negative cell lines, indicating that the silencing of TSP-1in NB was not because of an absence of positive trans-acting factors.Therefore, we next examined the methylation status of the TSP-1promoter. Methylation of promoter-associated CpG islands has beenimplicated in the transcriptional inactivation of a number of genesincluding TSP-1 in various types of cancer (32). Genomic DNA wasisolated from 4 TSP-1 negative NB cell lines (IMR-5, NMB, NBL-W-S, and NBL-W-N) and 2 NB cell lines that express TSP-1 (LA1-5sand SH-EP). Bisulfite-PCR analysis indicated that the TSP-1 CpGislands were methylated in all 4 of the TSP-1 negative cell lines. Incontrast, TSP-1 was completely unmethylated in the TSP-1 positiveNB cell lines (Fig. 4A).
To confirm that the hypermethylation occurred around the tran-scription start site, MSP PCR products were purified, cloned into thepCR2.1-TOPO vector, and sequenced. The results show that all 5 ofthe CpG sites around the transcription start site were completelymethylated in the 4 NB cell lines with silenced TSP-1. On the otherhand, CpG sites are completely unmethylated in the 2 TSP-1-positiveNB cell lines (Fig. 4B). Cytosines at non-CpG sites were converted tothymine, excluding the possibility that successful amplification couldbe attributable to incomplete bisulfite conversion (Fig. 4C).
To investigate whether aberrant methylation also occurred in vivo,the methylation status of the TSP-1 promoter was analyzed in 60 NBtumor samples using MSP. Methylation of the TSP-1 promoter wasdetected in 37% of the NB tumor samples (Fig. 5, A and B), but wasnot observed in 5 benign GNRs. Only 1 of 5 GNBs had TSP-1promoter methylation. Evaluation of TSP-1 protein expression in 12NB cases by immunohistochemistry did not show a correlation be-tween methylation status of the promoter and TSP-1 protein expres-sion. Methylated promoters were detected in 2 of 4 TSP-1 positivetumors and 1 of 8 TSP-1 negative NB tumors. The methylation studieswere performed with whole tumor DNA, whereas only a subset oftumor cells express TSP-1. Thus, the lack of association betweenpromoter methylation and TSP-1 expression in these studies mayreflect the heterogeneous nature of NB tumor tissues.
Restoration of TSP-1 Expression in TSP-1 Negative Cell Linesby 5-Aza-dC Alone and in Combination with TSA. The demethy-lating agent 5-Aza-dC, a methyltransferase inhibitor, was used toinvestigate whether TSP-1 expression could be restored in TSP-1negative cell lines. Four TSP-1 negative cell lines were treated with 1�M and 5 �M 5-Aza-dC for 60 h, and then TSP-1 mRNA expressionlevels were analyzed. Although treatment with 5-Aza-dC had little orno effect on TSP-1 expression in NBL-W-N, NMB, and IMR-5 celllines at this time point (Fig. 6A), expression was induced after longerexposure to 5-Aza-dC in all 3 of the cell lines (Fig. 7A). The NBL-W-S cells responded more rapidly to treatment. In these cells, TSP-1was up-regulated within 24 h of treatment with the demethylatingagent, and maximal levels were observed between 72 h and 96 h oftreatment (Fig. 6B). Furthermore, Western blot analysis revealed thepresence of TSP-1 protein in CM collected from NBL-W-S cells after96 h and 114 h of treatment with 5-Aza-dC (Fig. 6C).
To examine the role histone deacetylase activity plays in the si-lencing of the TSP-1 gene in NB, cells were treated with the histonedeacetylase inhibitor TSA alone and in combination with 5-Aza-dC.As shown in Fig. 7A, treatment with TSA alone failed to reactivate
Fig. 9. Effect of 5-Aza-dC on NB growth in vivo. A–C, rate of in vivo growth of NMB,NBL-W-N, and NBL-W-S xenografts after treatment with 5-Aza-dC. Five-point andeight-point stars indicate a statistical significance at P � 0.01 and P � 0.05, respectively,versus control by the Student’s t test.
6306
METHYLATION OF TSP-1 IN NEUROBLASTOMA
on May 6, 2018. © 2003 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

transcription of TSP-1 in any of the NB cell lines. Furthermore,treatment with TSA after 5-Aza-dC treatment did not result in en-hanced levels of TSP-1 mRNA expression in 3 of the cell lines.Although higher levels of TSP-1 mRNA were detected with thecombination treatment in NBL-W-S cells (Fig. 7B), histone deacety-lation does not appear to play a significant role in the silencing ofTSP-1 in NB.
5-Aza-dC Inhibits NB Growth in Vitro. We next examined theeffect of 5-Aza-dC treatment on NB growth in vitro. As shown inFig. 8A, growth of NMB and NBL-W-N cells was significantlysuppressed, with concentrations of 5-Aza-dC as low as 0.1 �M. Flowcytometric analysis revealed a marked accumulation of NMB andNBL-W-N cells in the G2-M phase of the cell cycle after 5-Aza-dCtreatment (Fig. 8, B and C). Treatment with 5-Aza-dC also inducedNB cell apoptosis and necrosis. As shown in Fig. 8E, double stainingfor Annexin-V and PI revealed that a large fraction of NMB andNBL-W-N cells progressed to late apoptosis/secondary necrosis (pos-itive for Annexin-V and positive for PI) or necrosis (negative forAnnexin-V and positive for PI) after 48 h of treatment. In contrast,even at high concentrations of the demethylating agent (100 �M), littlechange in proliferation of NBL-W-S cells was seen (Fig. 8A). Fur-thermore, G2-M phase arrest was not detected in these cells aftertreatment (Fig. 8D). A low percentage of NBL-W-S cells also under-went apoptosis after treatment with 5-Aza-dC, although an increase in
necrosis was not observed. Thus, although 5-Aza-dC restored TSP-1expression in the NBL-W-S cells, this drug had limited effect on thecell cycle, cell viability, and in vitro proliferation.
5-Aza-dC Suppresses Growth of NB Xenografts in Vivo andRestores TSP-1 Expression in a Subset of NB Xenografts. Toinvestigate whether NB growth in vivo could be suppressed with5-Aza-dC treatment, we generated xenografts using NMB, NBL-W-N, and NBL-W-S cell lines in nude mice. Treatment with 5-Aza-dC resulted in significant inhibition in the growth of the NB xe-nografts derived from the cell lines that also displayed suppression ofin vitro growth with treatment (NMB and NBL-W-N; P � 0.01;Fig. 9, A and B). Interestingly, growth of the NBL-W-S xenograftswas also significantly inhibited (P � 0.01; Fig. 9C), although in vitroproliferation of these cells was largely not effected by treatment.
Histological analyses of the treated NBL-W-S xenografts revealedmorphological evidence of neuroblast differentiation with increasedamounts of cytoplasm and vesicular nuclei. Immunohistochemicalstudies demonstrated that 5-Aza-dC restored TSP-1 expression inmany of the treated tumors (Fig. 10, A–F). We found that the per-centage of TSP-1 positive tumors increased with prolonged treatment.Tumors from 5 of 10 animals sacrificed 6 days after treatment showedrestoration of TSP-1. All of the 7 tumors that were harvested 9 daysafter treatment re-expressed TSP-1, and 5 of 7 tumors harvested 12days after treatment were TSP-1 positive. To investigate whether the
Fig. 10. Effect of 5-Aza-dC on TSP-1 expres-sion in NB xenografts. Immunohistochemistrystaining of untreated NBL-W-S xenografts withanti-TSP-1 antibody. No TSP-1 expression wasdetected in the untreated xenografts (A, C, and E).TSP-1 expression was detected in the cytoplasm ofthe NBL-W-S xenografts harvested 6, 9, and 12days after 5-Aza-dC treatment (B, D, and F). (Mag-nification, �400).
6307
METHYLATION OF TSP-1 IN NEUROBLASTOMA
on May 6, 2018. © 2003 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

restoration of TSP-1 expression in the treated tumors was associatedwith inhibited angiogenesis, tumor vascularity was examined in thecontrol and 5-Aza-dC-treated xenografts by immunohistochemistry(Fig. 11, A–F). The MVD was significantly lower in the treatedxenografts (82 � 34) compared with controls (120 � 33; P � 0.003;Fig. 11G). Although 5-Aza-dC treatment also inhibited the growth ofall of the NB xenografts, TSP-1 was detected in only 1 of 8 NBL-W-Nand 0 of 11 NMB 5-Aza-dC-treated-xenografts. Thus, in these NBxenografts, the impaired tumor growth does not appear to be becauseof re-expression of TSP-1.
DISCUSSION
TSP-1 is a potent inhibitor of angiogenesis, and several studies haveindicated that this glycoprotein has tumor suppressive properties invivo (25, 49). In this study, we examined TSP-1 expression and its
mechanisms of regulation in NB. We found that TSP-1 was silencedin a subset of NB tumors and cell lines. The frequency of TSP-1expression was higher in localized tumors compared with regional andwidely disseminated NBs, and a strong correlation between morpho-logical evidence of differentiation and TSP-1 expression was ob-served in the NB tumors. However, only 3 of 12 mature GNRsexpressed TSP-1, indicating that whereas this protein may play a rolein inhibiting angiogenesis in a subset of NB tumors, other factors arelikely to be responsible for the low vascularity observed in benignGNRs (4). Schwann cells, which are present in GNRs, also produce aspectrum of angiogenesis inhibitors, including tissue inhibitor ofmetalloproteinase-2, pigment epithelium-derived factor, and secretedprotein acidic and rich in cysteine, which appear to contribute to themore benign nature of tumors with abundant Schwannian stroma(8–10).
Fig. 11. Immunohistochemistry staining ofNBL-W-S xenograft tumors with anti-CD31 anti-body in control (A, C, and E) and 5-Aza-dC-treated(B, D, and F) NBL-W-S xenografts (magnification,�600). Treated tumors, harvested 6, 9, and 12 daysafter treatment, exhibited decreased vascular den-sity compared with controls. G, bar graph showingthe mean MVD measured in the treated- anduntreated-NBL-W-S xenografts (P � 0.003);bars, �SD.
6308
METHYLATION OF TSP-1 IN NEUROBLASTOMA
on May 6, 2018. © 2003 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

Enhanced levels of TSP-1 expression were seen in NB cell linesinduced to differentiate with either RA or BrdUrd. Furthermore,consistent with the tumor-suppressive activity of TSP-1 reported inother types of cancer cell lines, both of the NB subclones that were notcapable of forming tumors in nude mice expressed TSP-1 mRNA andsecreted TSP-1 protein. In contrast, Western blot analysis did notreveal TSP-1 protein secretion into CM collected from any of thetumorigenic NB cell lines analyzed, although TSP-1 mRNA andprotein expression were detected in a subset of these cell lines. Theseobservations emphasize the complex nature of TSP-1 regulation, andindicate that transcriptional and post-transcriptional mechanisms areinvolved.
Luciferase assays, performed with TSP-1 promoter-reporter con-structs, demonstrated no correlation between TSP-1 promoter activityand the level of TSP-1 expression in NB cell lines. Therefore, weexamined the methylation status of CpG residues in the TSP-1 pro-moter to determine whether epigenetic mechanisms were responsiblefor the silencing of TSP-1 in NB. TSP-1 was detected in NB cell linesthat lacked promoter methylation, whereas loss of TSP-1 transcriptionwas associated with hypermethylation of the promoter. Treatmentwith the demethylating agent 5-Aza-dC restored TSP-1 expression inTSP-1 negative cell lines, confirming that the transcriptional silencingof TSP-1 was caused by methylation. Although DNA methylation andhistone deacetylation have been reported to synergistically silencegenes in cancer (36), combinations of the histone deacetylase inhibitorTSA with 5-Aza-dC only increased TSP-1 expression levels in 1 ofthe 4 NB cell lines studied.
TSP-1 CpG island methylation also occurred in 37% of the clinicalNB tumors studied. A few of these NB tumors were available forTSP-1 expression studies, and no clear correlation between methyla-tion of the TSP-1 promoter and silencing of this gene was seen. Yanet al. (50) have examined recently the mechanisms of regulation ofCD44, an adhesion receptor that is silenced in highly malignant NBwith MYCN amplification. Hypermethylation of the CD44 gene pro-moter was seen in CD44-negative NB cell lines, whereas CD44-expressing cell lines were unmethylated. However, similar to ourresults, the level of expression of CD44 did not correspond to themethylation status of the promoter in NB tumors. These resultsemphasize the difficulty of studying gene regulation in clinical tumorsthat contain heterogeneous cell populations. However, it is also pos-sible that whereas the status of promoter methylation is a criticaldeterminant of TSP-1 and CD44 expression in NB cell lines, silencingof these genes in NB tumors may involve methylation-dependent aswell as methylation-independent mechanisms.
Two independent groups have reported recently that the promoterregion of RASSF1A, a newly described 3p21.3 tumor suppressor gene,is methylated in NB tumors and cell lines (51, 52). AlthoughRASSF1A expression was not evaluated in primary NB tumors ineither study, treatment with 5-Aza-dC restored RASSF1A mRNA incell lines. CASP8, a gene encoding a component of the Fas apoptoticpathway, has also been shown to be inactivated in NB tumors and celllines by DNA methylation (53). Interestingly, Astuti et al. (52) dem-onstrated a strong association between RASSFIA and CASP8 methy-lation in NB, indicating that �1 gene is likely to be aberrantlymethylated in the same tumor. Thus, the phenotype of an individualNB tumor is likely to be influenced by the silencing of several genes.
5-Aza-dC treatment significantly inhibited the growth of the NBL-W-N and NMB NB cell lines both in vitro and in vivo. In addition toarresting cell cycle, tumor cell apoptosis and necrosis were induced by5-Aza-dC. In contrast, in vitro growth of the NBL-W-S cell line waslargely unaffected by 5-Aza-dC. Nevertheless, the in vivo growth ofNBL-W-S xenografts was significantly impaired after treatment withthis demethylating agent. Restored TSP-1 expression and decreased
tumor vascularity was seen in almost all of the NBL-W-S xenograftsafter 5-Aza-dC treatment, suggesting that the impaired growth wasattributable, at least in part, to angiogenesis inhibition. Other mech-anisms are likely to be involved in the inhibition of tumor growthobserved in the NBL-W-N and NMB-xenografts after treatment, asTSP-1 re-expression was only rarely detected in these tumors. It ispossible that other growth-suppressive genes that are known to beaberrantly hypermethylated in cancer (54, 55) may be restored in thesecells after treatment, or that 5-Aza-dC may directly inhibit tumorgrowth by arresting cell cycle or by inducing apoptosis and necrosis.
Our studies demonstrate that 5-Aza-dC treatment results in signif-icant inhibition of NB growth. In a subset of NB xenografts, TSP-1expression was restored, and these tumors displayed inhibited angio-genesis. However, 5-Aza-dC is known to globally affect gene expres-sion, and the level of expression of other genes that are capable ofsuppressing NB tumor growth is also likely to be affected by thisdemethylating agent. We are currently conducting microarray studiesin an effort to identify additional genes that are up-regulated in NBafter treatment with 5-Aza-dC. Hopefully, these experiments willenhance our understanding of the molecular mechanisms by whichdemethylating agents inhibit tumor growth and provide insight intohow this treatment strategy can most effectively be used to treatchildren with NB.
REFERENCES
1. Brodeur, G. M., and Maris, J. M. Neuroblastoma. In: P. A. Pizzo and D. G. Poplack(eds.), Principles and Practice of Pediatric Oncology, 4 ed., pp. 895–937. Philadel-phia: Lippincott-Raven, 2001.
2. Brodeur, G. M., Pritchard, J., Berthold, F., Carlsen, N. L., Castle, V., Castelberry,R. P., De Bernardi, B., Evans, A. E., Favrot, M., Hedborg, F., Kaneko, M., Kemshead,J., Lampert, F., Lee, R. E. J., Look, A. T., Pearson, A. D. J., Philip, T., Roald, B.,Sawada, T., Seeger, R. C., Tsuchida, Y., and Voute, P. A. Revisions of the interna-tional criteria for neuroblastoma diagnosis, staging, and response to treatment. J. Clin.Oncol., 11: 1466–1477, 1993.
3. Maris, J. M., and Matthay, K. K. Molecular biology of neuroblastoma. J. Clin. Oncol.,17: 2226–2279, 1999.
4. Meitar, D., Crawford, S. E., Rademaker, A. W., and Cohn, S. L. Tumor angiogenesiscorrelates with metastatic disease, N-myc amplification, and poor outcome in humanneuroblastoma. J. Clin. Oncol., 14: 405–414, 1996.
5. Eggert, A., Ikegaki, N., Kwiatkowski, J., Zhao, H., Brodeur, G. M., and Himelstein,B. P. High-level expression of angiogenic factors is associated with advanced tumorstage in human neuroblastomas. Clin. Cancer Res., 6: 1900–1908, 2000.
6. Erdreich-Epstein, A., Shimada, H., Groshen, S., Liu, M., Metelitsa, L. S., Kim, K. S.,Stins, M. F., Seeger, R. C., and Durden, D. L. Integrins �v�3 and �v�5 are expressedby endothelium of high-risk neuroblastoma and their inhibition is associated withincreased endogenous ceramide. Cancer Res., 60: 712–721, 2000.
7. Ribatti, D., Surico, G., Vacca, A., De Leonardis, F., Lastilla, G., Montaldo, P. G.,Rigillo, N., and Ponzoni, M. Angiogenesis extent and expression of matrix metallo-proteinase-2 and -9 correlate with progression in human neuroblastoma. Life Sci., 68:1161–1168, 2001.
8. Huang, D., Rutkowski, J. L., Brodeur, G. M., Chou, P. M., Kwiatkowski, J. L., Babbo,A., and Cohn, S. L. Schwann cell-conditioned medium inhibits angiogenesis. CancerRes., 60: 5966–5971, 2000.
9. Chlenski, A., Liu, S., Crawford, S. E., Volpert, O. V., DeVries, G. H., Evangelista, A.,Yang, Q., Salwen, H. R., Farrer, R., Bray, J., and Cohn, S. L. SPARC is a keySchwannian-derived inhibitor controlling neuroblastoma tumor angiogenesis. CancerRes., 62: 7357–7363, 2002.
10. Crawford, S. E., Stellmach, V., Ranalli, M., Huang, X., Huang, L., Volpert, O.,De Vries, G. H., Abramson, L. P., and Bouck, N. Pigment epithelium-derived factor(PEDF) in neuroblastoma: a multifunctional mediator of Schwann cell antitumoractivity. J. Cell Sci., 114: 4421–4428, 2001.
11. Breit, S., Ashman, K., Wilting, J., Rossler, J., Hatzi, E., Fotsis, T., and Schweigerer,L. The N-myc oncogene in human neuroblastoma cells: down-regulation of anangiogenesis inhibitor identified as Activin A. Cancer Res., 60: 4596–4601, 2000.
12. Hatzi, E., Murphy, C., Zoephel, A., Rasmussen, H., Morbidelli, L., Ahorn, H.,Kunisada, K., Tontsch, U., Klenk, M., Yamauchi-Takihara, K., Ziche, M., Rofstad,E. K., Schweigerer, L., and Fotsis, T. N-myc oncogene overexpression down-regulates IL-6; evidence that IL-6 inhibits angiogenesis and suppresses neuroblastomatumor growth. Oncogene, 21: 3552–3561, 2002.
13. Eggert, A., Grotzer, M. A., Ikegaki, N., Liu, X. G., Evans, A. E., and Brodeur, G. M.Expression of the neurotrophin receptor TrkA down-regulates expression and func-tion of angiogenic stimulators in SH-SY5Y neuroblastoma cells. Cancer Res., 62:1802–1808, 2002.
14. Davidoff, A. M., Leary, M. A., Ng, C. Y., and Vanin, E. F. Gene therapy-mediatedexpression by tumor cells of the angiogenesis inhibitor flk-1 results in inhibition ofneuroblastoma growth in vivo. J. Pediatr. Surg., 36: 30–36, 2001.
6309
METHYLATION OF TSP-1 IN NEUROBLASTOMA
on May 6, 2018. © 2003 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

15. Katzenstein, H. M., Rademaker, A. W., Senger, C., Salwen, H., Nguyen, N., andCohn, S. L. Effectiveness of the angiogenesis inhibitor TNP-470 in reducing thegrowth of human neuroblastoma in nude mice inversely correlates with tumor burden.Clin. Cancer Res., 5: 4273–4278, 1999.
16. Nagabuchi, E., VanderKolk, W. E., Une, Y., and Ziegler, M. M. TNP-470 antian-giogenic therapy for advanced murine neuroblastoma. J. Pediatr. Surg., 32: 287–293,1997.
17. Wassberg, E., Pahlman, S., Westlin, J. E., and Christofferson, R. The angiogenesisinhibitor TNP-470 reduces the growth rate of human neuroblastoma in nude rats.Pediatr. Res., 41: 327–333, 1997.
18. Shusterman, S., Grupp, S. A., Barr, R., Carpentieri, D., Zhao, H., and Maris, J. M. Theangiogenesis inhibitor TNP-470 effectively inhibits human neuroblastoma xenograftgrowth, especially in the setting of subclinical disease. Clin. Cancer Res., 7: 977–984,2001.
19. Bornstein, P. Diversity of function is inherent in matricellular proteins: an appraisalof thrombospondin 1. J. Cell Biol., 130: 503–506, 1995.
20. Good, D. J., Polverini, P. J., Rastinejad, F., Le Beau, M. M., Lemons, R. S., Frazier,W. A., and Bouck, N. P. A tumor suppressor-dependent inhibitor of angiogenesis isimmunologically and functionally indistinguishable from a fragment of throm-bospondin. Proc. Natl. Acad. Sci. USA, 87: 6624–6628, 1990.
21. Jimenez, B., Volpert, O. V., Crawford, S. E., Febbraio, M., Silverstein, R. L., andBouck, N. Signals leading to apoptosis-dependent inhibition of neovascularization bythrombospondin-1. Nat. Med., 6: 41–48, 2000.
22. Tolsma, S. S., Volpert, O. V., Good, D. J., Frazier, W. A., Polverini, P. J., and Bouck,N. Peptides derived from two separate domains of the matrix protein throm-bospondin-1 have anti-angiogenic activity. J. Cell Biol., 122: 497–511, 1993.
23. Mettouchi, A., Cabon, F., Montreau, N., Vernier, P., Mercier, G., Blangy, D.,Tricoire, H., Vigier, P., and Binetruy, B. SPARC and thrombospondin genes arerepressed by the c-jun oncogene in rat embryo fibroblasts. EMBO J., 13: 5668–5678,1994.
24. Sheibani, N., and Frazier, W. A. Repression of thrombospondin-1 expression, anatural inhibitor of angiogenesis, in polyoma middle T transformed NIH3T3 cells.Cancer Lett., 107: 45–52, 1996.
25. Sheibani, N., and Frazier, W. A. Thrombospondin 1 expression in transformedendothelial cells restores a normal phenotype and suppresses their tumorigenesis.Proc. Natl. Acad. Sci. USA, 92: 6788–6792, 1995.
26. Weinstat-Saslow, D. L., Zabrenetzky, V. S., VanHoutte, K., Frazier, W. A., Roberts,D. D., and Steeg, P. S. Transfection of thrombospondin 1 complementary DNA intoa human breast carcinoma cell line reduces primary tumor growth, metastatic poten-tial, and angiogenesis. Cancer Res., 54: 6504–6511, 1994.
27. Grossfeld, G. D., Ginsberg, D. A., Stein, J. P., Bochner, B. H., Esrig, D., Groshen, S.,Dunn, M., Nichols, P. W., Taylor, C. R., Skinner, D. G., and Cote, R. J. Throm-bospondin-1 expression in bladder cancer: association with p53 alterations, tumorangiogenesis, and tumor progression. J. Natl. Cancer Inst., 89: 219–227, 1997.
28. Zabrenetzky, V., Harris, C. C., Steeg, P. S., and Roberts, D. D. Expression of theextracellular matrix molecule thrombospondin inversely correlates with malignantprogression in melanoma, lung and breast carcinoma cell lines. Int. J. Cancer, 59:191–195, 1994.
29. Yamashita, Y., Kurohiji, T., Tuszynski, G. P., Sakai, T., and Shirakusa, T. Plasmathrombospondin levels in patients with colorectal carcinoma. Cancer (Phila.), 82:632–638, 1998.
30. Laherty, C. D., Gierman, T. M., and Dixit, V. M. Characterization of the promoterregion of the human thrombospondin gene. DNA sequences within the first intronincrease transcription. J. Biol. Chem., 264: 11222–11227, 1989.
31. Dameron, K. M., Volpert, O. V., Tainsky, M. A., and Bouck, N. Control of angio-genesis in fibroblasts by p53 regulation of thrombospondin-1. Science (Wash. DC),265: 1582–1584, 1994.
32. Li, Q., Ahuja, N., Burger, P. C., and Issa, J. P. Methylation and silencing of theThrombospondin-1 promoter in human cancer. Oncogene, 18: 3284–3289, 1999.
33. Laird, P. W., and Jaenisch, R. DNA methylation and cancer. Hum. Mol. Genet., 3:1487–1495, 1994.
34. Nan, X., Ng, H. H., Johnson, C. A., Laherty, C. D., Turner, B. M., Eisenman, R. N.,and Bird, A. Transcriptional repression by the methyl-CpG-binding protein MeCP2involves a histone deacetylase complex. Nature (Lond.), 393: 386–389, 1998.
35. Jones, P. L., Veenstra, G. J., Wade, P. A., Vermaak, D., Kass, S. U., Landsberger, N.,Strouboulis, J., and Wolffe, A. P. Methylated DNA and MeCP2 recruit histonedeacetylase to repress transcription. Nat. Genet., 19: 187–191, 1998.
36. Cameron, E. E., Bachman, K. E., Myohanen, S., Herman, J. G., and Baylin, S. B.Synergy of demethylation and histone deacetylase inhibition in the re-expression ofgenes silenced in cancer. Nat. Genet., 21: 103–107, 1999.
37. Foley, J., Cohn, S. L., Salwen, H. R., Chagnovich, D., Cowan, J., Mason, K. L., andParysek, L. M. Differential expression of N-myc in phenotypically distinct subclonesof a human neuroblastoma cell line. Cancer Res., 51: 6338–6345, 1991.
38. Cohn, S. L., Salwen, H., Quasney, M. W., Ikegaki, N., Cowan, J. M., Herst, C. V.,Kennett, R. H., Rosen, S. T., DiGiuseppe, J. A., and Brodeur, G. M. Prolonged N-mycprotein half-life in a neuroblastoma cell line lacking N-myc amplification. Oncogene,5: 1821–1827, 1990.
39. Ciccarone, V., Spengler, B. A., Meyers, M. B., Biedler, J. L., and Ross, R. A.Phenotypic diversification in human neuroblastoma cells: expression of distinctneural crest lineages. Cancer Res., 49: 219–225, 1989.
40. Reynolds, C. P., Tomayko, M. M., Donner, L., Helson, L., Seeger, R. C., Triche, T. J.,and Brodeur, G. M. Biological classification of cell lines derived from humanextra-cranial neural tumors. Prog. Clin. Biol. Res., 271: 291–306, 1988.
41. Manohar, C. F., Short, M. L., Nguyen, A., Nguyen, N. N., Chagnovich, D., Yang, Q.,and Cohn, S. L. HuD, a neuronal-specific RNA-binding protein, increases the in vivostability of MYCN RNA. J. Biol. Chem., 277: 1967–1973, 2002.
42. Shimada, H., Ambros, I. M., Dehner, L. P., Hata, J., Joshi, V. V., and Roald, B.Terminology and morphologic criteria of neuroblastic tumors: recommendations bythe International Neuroblastoma Pathology Committee. Cancer (Phila.), 86: 349–362,1999.
43. Vermes, I., Haanen, C., Steffens-Nakken, H., and Reutelingsperger, C. A novel assayfor apoptosis. Flow cytometric detection of phosphatidylserine expression on earlyapoptotic cells using fluorescein labelled Annexin V. J. Immunol. Methods, 184:39–51, 1995.
44. Chen, S. Y., Yang, A. G., Chen, J. D., Kute, T., King, C. R., Collier, J., Cong, Y.,Yao, C., and Huang, X. F. Potent antitumour activity of a new class of tumour-specific killer cells. Nature (Lond.), 385: 78–80, 1997.
45. Thiele, C. J., Reynolds, C. P., and Israel, M. A. Decreased expression of N-mycprecedes retinoic acid-induced morphological differentiation of human neuroblas-toma. Nature (Lond.), 313: 404–406, 1985.
46. Schubert, D., and Jacob, F. 5-Bromodeoxyuridine-induced differentiation of a neu-roblastoma. Proc. Natl. Acad. Sci. USA, 67: 247–254, 1970.
47. Reynolds, C. P., and Maples, J. Modulation of cell surface antigens accompaniesmorphological differentiation of human neuroblastoma cell lines. Prog. Clin. Biol.Res., 175: 13–37, 1985.
48. Sidell, N. Retinoic acid-induced growth inhibition and morphologic differentiation ofhuman neuroblastoma cells in vitro. J. Natl. Cancer Inst., 68: 589–596, 1982.
49. Roberts, D. D. Regulation of tumor growth and metastasis by thrombospondin-1.FASEB J., 10: 1183–1191, 1996.
50. Yan, P., Muhlethaler, A., Bourloud, K. B., Beck, M. N., and Gross, N. Hypermethy-lation-mediated regulation of CD44 gene expression in human neuroblastoma. GenesChromosomes Cancer, 36: 129–138, 2003.
51. Harada, K., Toyooka, S., Maitra, A., Maruyama, R., Toyooka, K. O., Timmons, C. F.,Tomlinson, G. E., Mastrangelo, D., Hay, R. J., Minna, J. D., and Gazdar, A. F.Aberrant promoter methylation and silencing of the RASSF1A gene in pediatrictumors and cell lines. Oncogene, 21: 4345–4349, 2002.
52. Astuti, D., Agathanggelou, A., Honorio, S., Dallol, A., Martinsson, T., Kogner, P.,Cummins, C., Neumann, H. P., Voutilainen, R., Dahia, P., Eng, C., Maher, E. R., andLatif, F. RASSF1A promoter region CpG island hypermethylation in phaeochromo-cytomas and neuroblastoma tumours. Oncogene, 20: 7573–7577, 2001.
53. Teitz, T., Wei, T., Valentine, M. B., Vanin, E. F., Grenet, J., Valentine, V. A., Behm,F. G., Look, A. T., Lahti, J. M., and Kidd, V. J. Caspase 8 is deleted or silencedpreferentially in childhood neuroblastomas with amplification of MYCN. Nat. Med.,6: 529–535, 2000.
54. Herman, J. G., Merlo, A., Mao, L., Lapidus, R. G., Issa, J. P., Davidson, N. E.,Sidransky, D., and Baylin, S. B. Inactivation of the CDKN2/p16/MTS1 gene isfrequently associated with aberrant DNA methylation in all common human cancers.Cancer Res., 55: 4525–4530, 1995.
55. Herman, J. G., Jen, J., Merlo, A., and Baylin, S. B. Hypermethylation-associatedinactivation indicates a tumor suppressor role for p15INK4B. Cancer Res., 56:722–727, 1996.
6310
METHYLATION OF TSP-1 IN NEUROBLASTOMA
on May 6, 2018. © 2003 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

2003;63:6299-6310. Cancer Res Qi-Wei Yang, Shuqing Liu, Yufeng Tian, et al. Gene in Human Neuroblastoma
Thrombospondin-1Methylation-associated Silencing of the
Updated version
http://cancerres.aacrjournals.org/content/63/19/6299
Access the most recent version of this article at:
Cited articles
http://cancerres.aacrjournals.org/content/63/19/6299.full#ref-list-1
This article cites 50 articles, 24 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/63/19/6299.full#related-urls
This article has been cited by 16 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/63/19/6299To request permission to re-use all or part of this article, use this link
on May 6, 2018. © 2003 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from