metabolic control of urea catabolism in chlamydomonas reinhardi
TRANSCRIPT

JOURNAL OF BACTERIOLOGY, Mar. 1975, p. 1022-1035Copyright 0 1975 American Society for Microbiology
Vol. 121, No. 3Printed in U.S.A.
Metabolic Control of Urea Catabolism in Chlamydomonasreinhardi and Chlorella pyrenoidosa
ROBERT C. HODSON,* STUART K. WILLIAMS II, AND WILLIAM R. DAVIDSON, JR.
Department of Biological Sciences, University of Delaware, Newark, Delaware 19711
Received for publication 6 December 1974
In the unicellular green alga Chlamydomonas reinhardi (strain y-1), synthesisof the enzymes required for urea hydrolysis is under substrate induction controlby urea and under end product repression control by ammonia. Hydrolysis ofurea is effected by the sequential action of the discrete enzymes urea carboxylaseand allophanate lyase, collectively called urea amidolyase. The carboxylaseconverts urea to allophanate in a reaction requiring biotin, adenosine 5'-triphos-phate, and Mg2+. The lyase hydrolyzes allophanate to ammonium ions andbicarbonate. Neither activity is present in more than trace amounts whencultures are grown with ammonia or urea plus ammonia, or when they are starvedfor nitrogen for 8 h. Urea in the absence of ammonia induces both activities 10 to100 times the basal levels. Addition of ammonia to an induced culture causescomplete cessation of carboxylase accumulation and an 80% depression of lyaseaccumulation. Ammonia does not reduce urea uptake by repressed cells, so itdoes not prevent induction by the mechanism of inducer exclusion. Theunicellular green alga Chlorella pyrenoidosa (strain 3 Emerson) also has discretecarboxylase and lyase enzymes, but only the carboxylase exhibits metaboliccontrol.
Urea arises in the metabolism of autotrophsand some heterotrophs as an intermediate inthe recycling of endogenous nitrogen. The char-acteristic reaction of urea catabolism in theseorganisms is hydrolytic and catalyzed by urease(EC 3.5.1.5), a highly specific enzyme with noknown activators or cofactors. Recently, it hasbeen shown with some species of yeast andgreen algae lacking urease that urea amidolyase(urea carboxylase [hydrolyzing], EC 6.3.4.6),which requires adenosine 5'-triphosphate(ATP), bicarbonate, Mg2+, biotin, and a univa-lent cation that may be K+, Na+, or NH,+ (37,39, 51, 52, 55), can serve as an adaptive systemfor urea hydrolysis.The urea amidolyases from the yeasts Can-
dida utilis (39) and Saccharomyces cerevisiae(56, 57) and the green alga Chlorella vulgarisvar. viridis (51) have been found to consist oftwo distinct enzymatic activities. These activi-ties are called urea carboxylase and allophanatelyase (51) (or allophanate hydrolase [57 ]), andthey catalyze the hydrolysis of urea to ammo-nium ion and bicarbonate as follows:
011 ~~~~~~~UreacarboxylaseH2N-C-NH2 + ATP + HCO3- Pa
Mg2+
Urea K +(Na+, NH,+)
O H O11 11
H2N-C-N--u--u- + ADP + PiAllophanate
(1)
O H O11 11 Allophanate
H2N-C-N--C-O- + 3H20 + H+ lyase-Allophanate
2NH4+ + 2HCO3- (2)The yeast urea carboxylase and allophanatelyase activities remain together through severalpurification steps (39, 56), whereas the sameenzyme activities from Chlorella are readilyseparated (51). Several other species of greenalgae have been shown to possess urea amidoly-ase (25), but further characterization of these
1022
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
18
Febr
uary
202
2 by
183
.104
.106
.140
.

UREA CATABOLISM IN CHLAMYDOMONAS AND CHLORELLA
enzymes has not been reported.Initial experiments in our laboratory (17) and
by Roon and Levenberg (39) on the biochemis-try of urea metabolism in the green algaChlamydomonas reinhardi indicated that itmight possess an inducible catabolic pathwayfor urea which is also under control by repres-sion. The evidence for this was that the amido-lyase could be detected only in cells grown onurea, but not in cells deprived of combinednitrogen for several days or grown on ammonia(17). Additionally, the amidolyase could beobtained in two separate fractions exhibitingthe properties of urea carboxylase and allopha-nate lyase (17). Thus, it appeared to be similarto the Chlorella activity but quite unlike that inyeast.
In contrast to bacteria and fungi, less atten-tion has been given to enzymes in algae subjectto induction (or derepression) or repression(e.g., 5, 13, 16, 20-23, 27, 28, 31, 34, 45-47, 49,52) and even less to the combined controls ofinduction (or derepression) and repressionoperating on a single pathway or enzyme (16,27, 31, 34, 45, 49). Furthermore, in many of theexisting algal studies, the focus has been ontemporal control of structural gene expressionduring the cell cycle (22, 23, 27, 31, 52), or thespecies studied does not possess a mechanism(so far as is known) for genetic recombination(5, 13, 23, 28, 31, 34, 49, 50).Chlamydomonas has a history of genetic
manipulation and many genetic markers areavailable (11, 26). Its possession of two ureacatabolic enzymes operating in sequence andthe potential for genetic recombination affordsthe opportunity to study the mechanism of dualcontrol over the expression of enzymes in asingle algal pathway by both biochemical andgenetic approaches.This paper reports the results of initial stud-
ies on the induction and repression of theChlamydomonas urea amidolyase system andsome biochemical properties of the enzymesinvolved. All studies were with a strain wild-type for urea catabolism and with nonsynchro-nous cultures of vegetative cells. The terms"induction" and "repression" are used in theiroperational sense (33). A few results from aChlorella system are included for comparison.Preliminary accounts of this work have ap-peared (17, 60).
MATERIALS AND METHODSOrganisms and growth conditions. C. reinhardi
Dangeard (strain y-1) and Chlorella pyrenoidosaChick (strain 3 Emerson) were maintained in Erlen-meyer flasks on a rotating shaker platform under 500
ft-c (ca. 5,000 lx) of cool white fluorescent illuminationand at 25 to 30 C. Chiamydomonas was grown in amineral salts medium (35) supplemented with 0.2%sodium acetate as the carbon source and with either 5mM NH4NO, or 3 mM urea as the nitrogen source.Nitrate is not metabolized by this strain. Chlorellawas grown in a mineral salts medium (40) with either10 mM NH,Cl or 3 mM urea, and with 1% glucose.
Preparation and partial purification of cell ex-tracts. Unless stated otherwise, 10 mM sodium tri-cine [N-tris(hydroxymethyl)methylglycine l, pH 8.0,plus 2 mM dithioerythritol was used in all proceduresrequiring buffer. All manipulations of enzymes wereat 0 to 4 C, and glass-distilled water was usedthroughout.
Chlamydomonas was harvested by centrifugationfor 10 min at 4,000 x g, washed once by suspension in10 mM sodium tricine, and finally suspended in 4volumes of 100 mM sodium tricine containing 2 mMdithioerythritol. Extraction was by either of twoessentially equivalent methods. For method A, thecell suspension was passed once through a chilledFrench pressure cell (American Instrument Com-pany) at 10,000 lb/in2 (ca. 7 x 10' kg/i2) andcentrifuged for 60 to 90 min at 275,000 x g. The clear,pale-yellow supernatant solution, comprising thecrude extract, was subjected to fractional precipita-tion sith solid ammonium sulfate. Those proteinsprecipitating between 35 and 45% of saturation weredissolved in buffer to 1/10 the original volume of crudeextract, giving a protein concentration between 10and 20 mg/ml. For method B, the cell suspension wasplaced in 50-ml polycarbonate centrifuge tubes, frozenin a dry ice-ethanol bath for 1 h, and thawed slowly incool water. The resulting suspension was centrifugedfor 10 min at 12,000 x g, the pellet was washed onceby suspension in 4 volumes of 100 mM buffer, and thecombined supematants were brought to 50% satura-tion with solid ammonium sulfate. The precipitatedprotein was dissolved in buffer as in method A.Extracts obtained either way could be stored forseveral weeks at - 20 C with retention of at least 90%of the initial urea amidolyase activity. Numerousthaw-freeze cycles over a period of 4 months decreasedthe urea carboxylase activity to 1% and the allopha-nate lyase activity to 50% of the initial values.
Cell extracts of Chlorella were prepared as formethod A, except that the cells were disrupted in aFrench pressure cell operating at 20,000 lb/in2 (ca. 14x 106 kg/M2). To reduce the volume, urea amidolyaseactivity was precipitated with ammonium sulfatebetween 30 and 60% saturation.
Partial purification and separation ofChiamydomonas urea amidolyase activities wereachieved essentially by the method of Thompson andMuenster (51, 53). The protein obtained from precipi-tation with ammonium sulfate was diluted to 1 mg/mland placed on a column of brushite previously equili-brated with buffer, using a ratio of 2 mg of protein to 1or 2 ml of packed brushite. The column was elutedstepwise with 2 volumes of buffer containing 5 mMpotassium phosphate, followed by 2 volumes of bufferwith 20 mM potassium phosphate.
VOL. 121, 1975 1023
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
18
Febr
uary
202
2 by
183
.104
.106
.140
.

HODSON, WILLIAMS, AND DAVIDSON
Enzyme assays. Urea amidolyase activities wereassayed with radioactive substrates by proceduressimilar, in principle, to those published by others (38,55). All incubations were carried out in 25-ml Erlen-meyer flasks held in a water bath at 25 or 30 C andsealed with a rubber serum stopper. Each flask had acenter well containing 0.5 ml of 10% KOH. Theenzymatic reactions were always started by the addi-tion of "4C-labeled substrate and stopped with 0.5 mlof 6 N H2SO4. Radioactive carbon dioxide was allowedto diffuse into the KOH for at least 30 min withcontinuous shaking. A 0.2-ml sample from the centerwell was combined with 0.8 ml of water and 10 ml ofeither a standard Triton X-100 and toluene-basedscintillation mixture, or a 1:1 (vol/vol) mixture ofOmnifluor (New England Nuclear) in toluene and2-ethoxyethanol, and the radioactivity was deter-mined by liquid scintillation at room temperature.The counting efficiency of about 80% was checkedoccasionally with an internal standard of["C ]toluene. One unit of activity is equivalent to thatamount of extract liberating 1 gmol of carbon dioxideper min at pH 8.0 and the stated temperature.
["4C]urea uptake and intracellular ["C]urea. Anammonia-grown Chlamydomonas culture was har-vested by centrifugation, washed with sterile 10 mMpotassium phosphate (pH 6.5), and suspended infresh nitrogen-free medium to yield a density of 150Klett units. Portions of this suspension were placed in250-ml flasks either with or without 10 mM ammo-nium nitrate and returned to the growth shaker for 15min. ["4C]urea was added to a final concentration of0.3 mM and the flasks were returned to the shaker. Atintervals, duplicate 2.5-ml samples were filteredthrough 25-mm diameter fiber glass disks (Gelmantype E) and rinsed with 20 ml of ice-cold phosphatebuffer containing 10 mM urea. The filters were placedin 10 ml of a 2-ethoxyethanol and Omnifluor mixture,and the radioactivity was determined by liquid scin-tillation. Corrections were made for quenching bychlorophyll.
Nonspecific binding of ["4C ]urea to cells and filterwas estimated with similar suspensions previouslykilled by freezing to - 70 C. Corrections also had to bemade for ["4C ]urea enzymatically hydrolyzed to "4CO2and not assimilated by photosynthesis. To do this,immediately after addition of ["4C jurea, 1-ml portionsof suspension were sealed in 25-ml flasks possessing10% KOH in a center well. These flasks were incu-bated under the above conditions of light and agita-tion. At various times "4CO2 was liberated withsulfuric acid and trapped in the KOH, and theradioactivity was determined as for the urea amidoly-ase assay.
Intracellular ["C ]urea was estimated with filteredand washed cells. The filter disks were quickly placedin 4 ml of ice-cold 0.75 M sodium phosphate (pH 8)and frozen with liquid nitrogen. Upon thawing andcentrifugation, 1-ml samples of supernatant wereplaced in 25-ml center well flasks together with 1 ml ofjack bean urease (Sigma type IV dissolved in 0.02 Msodium phosphate [pH 8]; 1 U/ml) and incubated for30 min at 30 C. Radioactive carbon dioxide was
estimated by microdiffusion into KOH and liquidscintillation.
Kinetics of induction and repression. We do nothave a nonrepressing nitrogen source forChlamydomonas that will allow rapid growth rates,and so for induction experiments cells were grownwith ammonia. In the first experiments, cultureswere grown to about 50 Klett units with limitingammonia (0.5 mM) and then given inducer withoutchanging the medium. To provide more uniformmaterial, in later experiments cells were grown onexcess ammonia (5 mM), harvested during exponen-tial growth by low-speed centrifugation at roomtemperature, washed once by suspension in nitrogen-free medium, and resuspended in fresh nitrogen-freemedium to give an absorbance of 100 Klett units orless. Such cells gave rapid induction rates whether ornot they were preincubated to deplete endogenouspools. Cells grown with limiting ammonia exhibited alag in enzyme accumulation during the early minutesof induction.
Cells for repression experiments were grown withexcess urea in a single culture, which was divided intoseparate flasks just before making further additions.
Extracts were prepared from duplicate 15-ml sam-ples of culture. The cells were sedimented in 15-mlCorex tubes at 12,000 x g for 10 min at 0 to 4 C,washed once by suspension in 5 ml of 10 mM sodiumtricine (pH 8.0) containing freshly added 20 mMmercaptoethanol, and finally resuspended in 5 ml ofthe same buffer.
Disruption of cells held in an ice-salt bath by sonicoscillation with a Branson microtip (Heat Systems,Inc.) operating for 20 s at the lowest power settinggave at least 99% breakage and maximum recovery ofurea amidolyase activity. A few remaining cells andthe larger cellular debris were removed by centrifuga-tion for 10 min at 12,000 x g. The clear, greensupernatant fluid was used within 1 h, althoughactivity was stable for at least 6 h. Usually 1 ml ofextract was used for each enzyme assay.
Analytical procedures. Protein concentration wasdetermined by the Lowry procedure (24) or tur-bidometrically (24) after precipitation in 5% trichloro-acetic acid. Crystalline bovine serum albumin wasused as a standard. To remove thiol, which gives acolor reaction with the Folin reagent, protein inextracts was precipitated with 8% trichloroacetic acid,sedimented by centrifugation, and dissolved in 1 NNaOH with heating at 60 C for 10 min. This methodgave reproducible and quantitative extraction of pro-tein from whole cells as well.
Ammonia in culture media was determined bydirect Nesslerization. Growth of algal cultures wasmeasured as apparent absorbance with a Klett-Sum-merson colorimeter using a green filter whose trans-mission centered at 540 nm (18). Absorbance wasproportional to cellular dry weight in the range of 10to 200 Klett units.
Ascending paper chromatography employed What-man no. 1 filter paper and the following solvents: (i)methanol-pyridine-water (85:4:4) and (ii) 95%ethanol-water (7:3). High-voltage paper electrophore-
1024 J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
18
Febr
uary
202
2 by
183
.104
.106
.140
.

UREA CATABOLISM IN CHLAMYDOMONAS AND CHLORELLA
sis in a water-cooled apparatus (Savant Instruments,Inc.) was carried out with 0.05 M tris(2-amino-2-[hydroxymethyl ]-1,3-propanediol) acetate buffer (pH7.9), or 1% sodium tetraborate, and at 3 C in a
gradient of 80 V/cm. Ureido compounds were locatedon the paper with the usual Ehrlich reagent.
Reagents. "'C-labeled potassium allophanate was
prepared from, ["IC ]urea either enzymatically by theaction of Chiamydomonas urea carboxylase or chemi-cally. The enzyme reaction mixture contained theusual components of the urea carboxylase assay (seeFig. 3) but without allophanate lyase. The progress ofthe reaction was determined by passing a samplethrough a small column of quaternary amine resin inthe hydroxide form and counting the amount ofunreacted [4C ]urea in the effluent. When less than1% of unreacted urea remained, carrier allophanatewas added, the pH was adjusted to approximately 10with concentrated NaOH, and the mixture was
heated at 60 C for 3 min to destroy residual enzyme
activity. This preparation of [14C ]allophanate was
stored at -20 C and used without purification. Thechemical preparation of ["4C lallophanate followed theprocedure of Whitney and Cooper (57), except thatmore KOH was added to completely dissolve themethyl ester. The product was stored in a desiccatorat -20 C, and solutions were made up in 0.1 Msodium carbonate as needed. Unlabeled potassiumallophanate was obtained by the action of 1 N KOH in60% (vol/vol) aqueous ethanol on methyl allophanateand purified by repeated crystallization from aqueous
ethanol. Methyl allophanate was prepared by themethod of Dains and Wertheim (9).
Brushite was made by the method of Siegelman etal. (43); ["C ]urea was purchased from Calbiochemand New England Nuclear. All other reagents were
obtained from commercial sources in the highestpurity available.
RESULTS
Properties of urea amidolyase fromChlamydomonas. Extracts from urea-growncells catalyze the hydrolytic decomposition of[14C ]urea to "CO2. Ammonia, the other productexpected, has been detected indirectly by cou-
pling its formation to the oxidation of reducednicotinamide adenine dinucleotide with bo-vine liver glutamate dehydrogenase (EC1.4.1.3). The hydrolytic activity requires ATPand Mg2+, and it is lost when extracts are
treated with heat or avidin prior to incubationwith substrate (Table 1). These characteristicsof the Chlamydomonas activity are similar tothe urea amidolyase system obtained from yeast(39, 55) and Chlorella (51, 53). This, togetherwith an earlier report (39), demonstrates thatChlamydomonas employs urea amidolyaserather than urease for catabolism of exogenousurea.The Chlamydomonas urea amidolyase sys-
TABLE 1. Requirements for urea amidolyase ofChlamydomonas in a cell extracta
System Activity (mU)
A. Complete system' 5.9-MgSO4 0.037-ATP 0.24Heated 100 C, 3 min 0.016
B. Native enzyme' 2.1+Avidinc 0.016
a Experiments A and B were with separate enzymepreparations. The extract was a 35 to 45% ammoniumsulfate fraction from a crude homogenate of urea-grown cells. (A) The extract was desalted through acolumn of Sephadex G-25; (B) the extract was notdesalted.
"The complete system had the following additionsto a final volume of 2.5 ml: KCl, 200 /mol; MgSO4, 20,umol; disodium ATP, 20 umol; sodium tricine (pH8.0), 500 Mmol; ,-mercaptoethanol, 50 Amol;NaHCOs, 20 Amol; [14C]urea, 2.1 Amol (1,610disintegrations/min per nmol); and extract equivalentto 2.3 ml of culture at 190 Klett units. Incubation wasat 25 C for 60 min. One milliunit, 1 nmol of "4CO2formed per min.
c200 ug of avidin was added 30 min before[14C jurea.
tem can be separated into two components withdistinctly different activities. Crude extractsubjected to fractional precipitation with am-monium sulfate followed by adsorption tobrushite in the presence of phosphate ions givestwo heat-labile components (Table 2), desig-nated for convenience fractions 1 and 2, whichmust be recombined to obtain 14CO from[14C]urea. The resolution of these two compo-nents varied from one experiment to the next,and complete separation has not been obtainedby this method. Fraction 1 is the componentwith less affinity for brushite in the presence of5 mM phosphate. It does not have appreciableactivity with [4C ]urea (Table 3 and Fig. 1B),but it hydrolyzes [14C ]allophanate to 14CO2(Table 3). Fraction 2, on the other hand, ad-sorbs to brushite in the presence of 5 mMphosphate and is eluted by 20 mM phosphate. Itis unable to hydrolyze either urea or allopha-nate (Table 3), but it converts ["C ]urea toanother radioactive substance (Fig. 1C). Thissubstance is identified as [4C ]allophanate byits lability to mineral acid and by its co-electro-phoresis and co-chromatography with authenticmaterial (Fig. 2). Based on these observations,fraction 1 contains the activity of allophanatelyase (allophanate hydrolyase, 56), whereasfraction 2 contains urea carboxylase activity.
VOL. 121, 1975 1025
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
18
Febr
uary
202
2 by
183
.104
.106
.140
.

HODSON, WILLIAMS, AND DAVIDSON
TABLE 2. Separation of Chlamydomonas ureaamidolyase activities by chromatography on brushite
Enzyme fraction Total activityaEnzymefraction(MU)Sample for columnb 91Column fractions:
1 (not adsorbed) 0.432 (adsorbed) 0.981+2 631 + 2 heated 0.471 heated + 2 0.17
a Urea amidolyase activity was assayed in thecomplete system listed in Table 1, except ["C lureaspecific activity was 1,524 disintegrations/min pernmol.
b A 35 to 45% ammonium sulfate fraction fromcrude homogenate of urea-grown cells. Two milli-grams of protein (equivalent to 171 ml of culture at200 Klett units) was applied to 2 ml of brushite. Heat-ing was at 100 C for 3 min.
Discrete enzymes with similar properties havebeen obtained from the green alga C. vulgarisvar. viridis (51). Thus, separate carboxylase andlyase enzymes may be a characteristic of greenalgae, in contrast to yeast systems in whichthese activities remain together through severalstages of purification (39, 56).Whether allophanate is a free or bound inter-
mediate in Chlamydomonas is uncertain. How-ever, cell extracts incubated with ["4C ]urea andthen immediately chilled to 0 C and subjectedto electrophoresis at an alkaline pH show anappreciable accumulation of [4C ]allophanate(Fig. IA). In addition, urea amidolyase activityexhibits in vitro a nonlinear relationship toextract concentration (Fig. 3A) which is absentfrom the individual urea carboxylase and allo-phanate lyase reactions (Fig. 3B, C). Thisnonlinearity may reflect the time it takes forfree allophanate to accumulate to a steady-stateconcentration. Both of these observations areconsistent with readily reversible binding ofallophanate to urea carboxylase or allophanatelyase, and they support the notion that allopha-nate is a free intermediate in cell extracts and,therefore, in the intact cell. Of course, it is alsopossible that urea carboxylase and allophanatelyase are weakly associated in the cell as acomplex, serving to channel allophanate, whichis very unstable at neutral and acidic pH.The non-linearity in the urea amidolyase
assay cited above has raised a technical prob-lem. It has been difficult to quantitate thisactivity from the usual fixed-timed assay. Animprovement is obtained if the rate is calcu-
lated from some time interval during the reac-tion when steady state is approached, for in-stance from 60 to 90 min, but even this has notbeen entirely satisfactory. For this reason, andalso for the sake of economy, a fixed time,usually 60 min, was employed for this work. Thevalues of urea amidolyase reported are thereforeminimum estimates. The urea carboxylase andallophanate lyase activities were proportional tothe incubation times and enzyme concentra-tions employed throughout this work.Regulation of urea amidolyase activities in
Chlamydomonas by urea and ammonia. Ureaamidolyase could sometimes be detected inextracts from ammonia-grown cells (see Table6), but the amounts were near the lower limit ofthe assay. Nevertheless, at least a 10- to 100-fold increase in urea amidolyase activity may beobtained if ammonia is replaced by urea in theculture medium (Fig. 4; see Table 6).The optimum concentration of exogenous
urea to obtain an elevated level of urea amidoly-ase is about 3 mM (Fig. 4). At this concentra-tion, induction was rapid and a high level ofactivity was maintained for several hours. Con-centrations less than optimum supported thesame rapid induction, but the specific activitythen declined. At a greater than optimumconcentration of urea, induction was slower and
TABLE 3. Activity of Chlamydomonas ureaamidolyase fractions incubated with [14C]urea and
["Cjallophanate as substrates
Total enzymaticactivitya (mU)
Extract fraction [l4C"1C jallo-14Cur phanate
0C2 "4Co02
35-45% ammonium sulfateb 731Brushite column fractions:
1 (not adsorbed) 31 1,0502 (adsorbed) 0 01+2 407
aUrea amidolyase ([4C Jurea to "4CO2) was assayedin the complete system listed in Table 1, except KClwas omitted and the incubation was for 30 min.["C lurea specific activity, 1,470 disintegrations/minper nmol. The allophanate lyase assay ("4C jallopha-nate to "4CO2) had the following additions in a totalvolume of 2.5 ml: sodium tricine (pH 8.0), 500,umol;fB-mercaptoethanol, 50 Mmol; NaHCO3, 20 umol;["4C jallophanate, 1.1 gmol (460 disintegrations/minper nmol); and extract. Incubation was at 25 C for 30min. One milliunit: 1 nmol of 4CO2 formed per min.
bPrepared from crude homogenate of urea-growncells and equivalent to 1,500 ml of culture at 178 Klettunits.
1026 J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
18
Febr
uary
202
2 by
183
.104
.106
.140
.

UREA CATABOLISM IN CHLAMYDOMONAS AND CHLORELLA
5 10 15DISTANCE TRAVELLED (CM)
exogenous ammonia, induction was prevented(Table 4).Induction kinetics with Chlamydomonas.
The results of induction of urea amidolyase inammonia-grown cells during approximately two
300(
2404
804
0i
04
120
60(
180
120
60
20 25® 30C
- 15CFIG. 1. Electrophoretic pattern of products from
Chiamydomonas urea amidolyase components incu-bated with ["4CJurea. The components given in thefootnotes to Table 1, together with 5 MiCi of ["4C]urea(43 gCi/Iumol) in a total volume of 0.5 ml, wereincubated with enzyme at 25 C for 60 min. Heatingwas at 100 C for 3 min. The reactions were stoppedwith 4 volumes of ice-cold 95% ethanol, and 20-iAsamples were subjected to electrophoresis for 30 minat pH 7.9. The enzymes were as follows: (A, D) 35 to45% ammonium sulfate fraction, equivalent to 14 mlof culture at 81 Klett units; (B) nonadsorbed fraction,equivalent to 7 ml of culture; (C) adsorbed fraction,equivalent to 10 ml of culture. The incomplete recov-ery of radioactivity in (A) is due to "4CO2 production.
the maximum specific activity was also shortlived. This latter decline may have been causedby an accumulation of ammonia, which has arepressive action (see below). Indeed, if theculture medium had more than about 1 mM
u 6000a 4 DUC4
a 30C15C
ORIGIN0
05 10 15DISTANCE TRAVELLED (CM)
ORIGIN 5 10 15DISTANCE TRAVELLED (CM)
FIG. 2. Identification of ["4C]allophanate synthe-sized with Chlamydomonas extract by (A) co-electro-phoresis and (B) co-chromatography with authenticunlabeled potassium allophanate and unlabeled urea
before and after hydrolysis with mineral acid. Theradioactive material was the product obtained in theenzymatic synthesis of [4C ]allophanate. Hydrolysiswas with 1 N HCI for 5 min at ambient temperature.Electrophoresis was for 15 min at pH 7.9, andchromatography was in solvent i. The untreated andacid-hydrolyzed samples contained different amountsof radioactivity.
VOL. 121, 1975 1027
CRUME EXTRAC:T A
UREA/\ ~~~~~~ALLOPHANATE
FRA1:TION NOT ADSOWBD TO MIRUHITE (I) B
FRACTION ADSORBED TO BRUSHITE (2) C
HEATED CRUDEEXTACTD
200.
1000.
900-
600
400
200.
I.- woI--
X00
-4 400-a
200
800
600-
400-
200-
0-
ORIGIN
0E
BEFORE ACID ,a
ALLOPHANATE
0-
~~~0-~l
0-
10- I
AFTER ACID
0
10- UREA
o- r . .III
BEFORE ACID B
40-10 0
10 ALLOPHANATE
10 /\ URE A)O0
)0 AFTER ACID Q
10 UREA
10-
20
__Pj
:60Ci
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
18
Febr
uary
202
2 by
183
.104
.106
.140
.

1028 HODSON, WILLIAMS, AND DAVIDSON J. BACTERIOL.
A B C
4 2
cl) 140-
z
100-
2.-60-
w
z20w 2
0 0- 00 I2 02 46 0 223
AMOUNT OF EXTRACT (JL CULTURE -A)FIG. 3. Relationship between Chlamydomonas extract concentration and activities of (A) urea amidolyase,
(B) urea carboxylase, and (C) allophanate lyase. The activities were measured on separate occasions withdifferent extracts. All incubations were for 30 min at 25 C. Other assay conditions were as follows: (A) reactioncomponents given in the footnotes to Table 1, [14C ]urea (1,383 disintegrations/min per nmol); (B) as for (A) plus2 mUof Chlorella allophanate lyase (brushite fraction 1); (C) reaction components given in the legend to Table3, [l4Clallophanate (456 disintegrations/min per nmol).
i4- x generations are presented in Fig. 5A. For this4 *(. 3 MM) experiment, the culture was grown initially withw 12- 0.5 mM ammonia, a concentration that does not
-. prevent induction (Table 4). Enzyme activityJ-10- K per millititer of culture increased during a
period of exponential growth (about 16 h) andA / ^ "\then decreased rapidly as the culture ap-1 81 t/ \ < \ *- | proached the stationary phase. At about this
F ID\ / ^ \ ' same time, ammonia first appeared in thez culture medium (limit of detection: 0.05 mM)3 /// */\ \ \ SX UUREA- and began to accumulate. The near coincidence
GROWN between exogenous ammonia accumulation andH AD/\ \ * ( 10MblM) the decreasing rate of urea amidolyase accumu-> ll/ \0 \ lation suggests that the pathway is under con-
2 2- 1/ \ \ (o.3 M) trol by repression, with ammonia or a metabolic. A(O. M) product acting as corepressor. The subsequent
E x x xxK decline of enzyme activity may reflect inactiva-> (0) tion of urea amidolyase enzyme without con-z 0 4 8 12 16 20 24 comitant synthesis, or perhaps senescence and
TIME AFTER ADDING INDUCER (HRS) death of some cells not accompanied by adecrease in the culture absorbance. If the same
FIG. 4. Specific activity of Chlamydomonas urea data are normalized to unit culture absorbanceamidolyase from cultures induced with various urea (not shown), then the specific activity increasedconcentrations and for various times. ( ) Cultures for only the first 12 h as the culture absorbancegrown on 0.5 mM NH4NO3 and urea added to the increased about 2.4-fold. This relationship sug-concentration given after each curve. (---) Culture gests about potentis washipasug-grown on 10 mM urea. Urea amidolyase was assayed gests that the induction potential was realizedin the complete mixture of Table 1. [14C]urea specific in about one cell generation. Essentially theactivity: 1,798 disintegrations/min per nmol. Incuba- same temporal relationship among induction oftions were for 60 min at 25 C. urea amidolyase, growth, and ammonia excre-
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
18
Febr
uary
202
2 by
183
.104
.106
.140
.

UREA CATABOLISM IN CHLAMYDOMONAS AND CHLORELLA
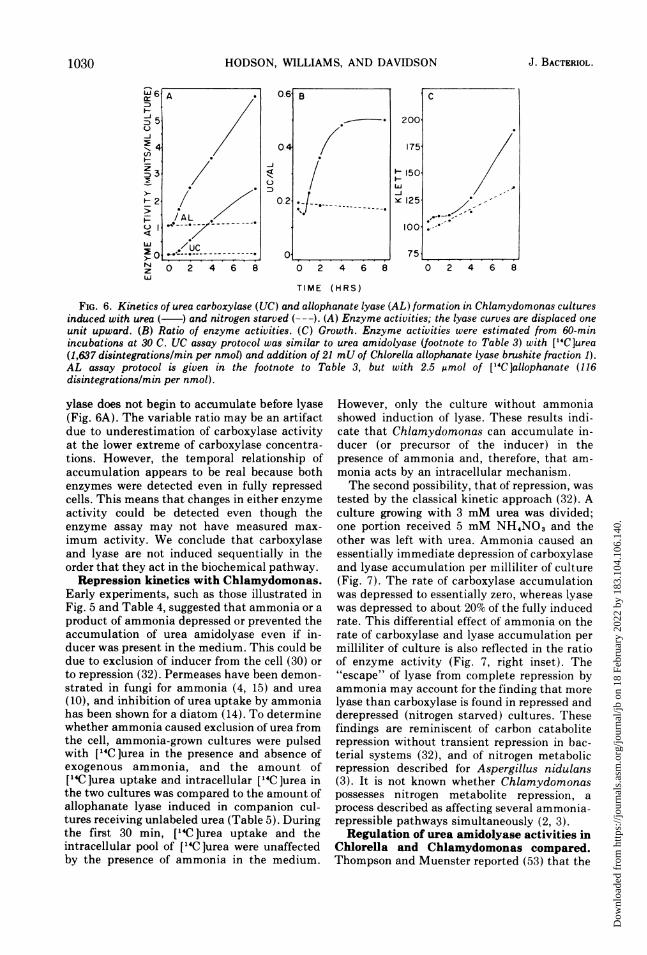
tion was obtained with a culture started in ureamedium (Fig. 5B).Since urea amidolyase embraces two en-
zymes acting in series, induction could be due toaccumulation of one or both enzyme activities.The kinetics of induction during one cell gen-eration in which urea carboxylase and allo-phanate lyase were assayed separately is shownin Fig. 6. For this experiment, Chlamydomonaswas grown with 5 mM ammonia, suspended innitrogen-free medium, and incubated for 2 h todeplete endogenous ammonia pools. Prior to
TABLE 4. Ability of urea to induce amidolyaseactivity in Chlamydomonas cultures grown with
various concentrations of ammoniaa
Density of Urea amido-Nitrogen source for cluea ys patNo. growth (initial culture at (lU/mla
concn,mM) assay" (A) of culture -A)
1 Urea (10) 70 232 NH4NO3 (0.25) 38 133 NH4NOs (0.50) 41 164 NH4NO3 (1.0) 53 0.0185 NH4NO3 (2.0) 39 0
aInduction of cultures 2 to 5 was for 6 h with 10 mMurea.
b A, Absorbance measured in Klett units.c Urea amidolyase was assayed in the complete
system listed in Table 1. Each incubation received theextract of 4 ml of culture. ["C jurea specific activity,1,763 disintegrations/min per nmol. One microunit: 1pmol of "CO2 formed per min at 25 C.
H
--
0
-i
C,)H-z
H
w
adding inducer, there was a small but signifi-cant amount of each enzyme (Fig. 6A), and thiswas increased only slightly during a further 6 hin nitrogen-free medium. Upon addition of urea,both enzyme activities increased with biphasicrate curves; an initial exponential rate gave wayto an approximately linear one. Ammonia didnot accumulate in the medium, so it is assumedthat the culture was under control only byinducer. These kinetics are similar to the induc-tion rate curve for nitrate reductase in Aspergil-lus nidulans (6) and, as pointed out by Cove (7),they contrast with most Escherichia coli sys-tems in which the rate of induction is linear andproportional to cell mass.Some enzyme systems in fungi "induced" by
substrate have turned out to be derepressed bythe unintentional removal of corepressor (33).Induction, rather than derepression, of ureacarboxylase and allophanate lyase inChlamydomonas is demonstrated by the data inFig. 6A. Incubation in nitrogen-limited mediumfor 8 h "derepressed" the carboxylase only1.6-fold and derepressed the lyase only 1.8-fold.With 3 mM urea in the medium of a companionculture, the carboxylase was induced 79-foldand the lyase was induced 26-fold over thederepressed levels of these enzymes. Growthwith urea, therefore, involves induction of bothenzymes over the basal (derepressed) levels.The data in Fig. 6 indicate also that the ratio
of carboxylase to lyase during induction doesnot remain constant (Fig. 6B), and that carbox-
0 8 16 24 32 0 8 16 24 32 40TIME (HRS)
P1r-
240 >H
200 rmC)
160 c
120 I
80 r-rr-4q
40 --c
0 ZCn
FIG. 5. Urea amidolyase activity and extracellular ammonia concentration during growth ofChlamydomonas with urea. (A) Culture started on 0.5 mMNH4NO, and induced with 3 mM urea at time zero.(B) Culture started on 3 mM urea and no more urea added. The enzyme assay components were as given in thefootnote to Table 3; [14C]urea specific activity was 1,738 disintegrations/min per nmol. Incubation was for 60min at 25 C.
VOL. 121, 1975 1029
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
18
Febr
uary
202
2 by
183
.104
.106
.140
.
HODSON, WILLIAMS, AND DAVIDSON
I-,,6 Aa:
Z~4
2 / /u /u> .
LU
D 3
> ......1~~~~hJ Z
06f B
04
-0
0.2
o0
0260- - --- o
O 2 4 6 8
200
175
H 150
-JY 125
100
75
c
0 - 4 6 8
O 2 4 6 8
TIME (HRS)
FIG. 6. Kinetics of urea carboxylase (UC) and allophanate lyase (AL) formation in Chlamydomonas culturesinduced with urea ( ) and nitrogen starved (---). (A) Enzyme activities; the lyase curves are displaced oneunit upward. (B) Ratio of enzyme activities. (C) Growth. Enzyme activities were estimated from 60-minincubations at 30 C. UC assay protocol was similar to urea amidolyase (footnote to Table 3) with ["4C]urea(1,637 disintegrations/min per nmol) and addition of 21 mU of Chlorella allopharnate lyase brushite fraction 1).AL assay protocol is given in the footnote to Table 3, but with 2.5 Mmol of ["4C]allophanate (116disintegrations/min per nmol).
ylase does not begin to accumulate before lyase(Fig. 6A). The variable ratio may be an artifactdue to underestimation of carboxylase activityat the lower extreme of carboxylase concentra-tions. However, the temporal relationship ofaccumulation appears to be real because bothenzymes were detected even in fully repressedcells. This means that changes in either enzymeactivity could be detected even though theenzyme assay may not have measured max-imum activity. We conclude that carboxylaseand lyase are not induced sequentially in theorder that they act in the biochemical pathway.Repression kinetics with Chlamydomonas.
Early experiments, such as those illustrated inFig. 5 and Table 4, suggested that ammonia or aproduct of ammonia depressed or prevented theaccumulation of urea amidolyase even if in-ducer was present in the medium. This could bedue to exclusion of inducer from the cell (30) orto repression (32). Permeases have been demon-strated in fungi for ammonia (4, 15) and urea(10), and inhibition of urea uptake by ammoniahas been shown for a diatom (14). To determinewhether ammonia caused exclusion of urea fromthe cell, ammonia-grown cultures were pulsedwith [4C ]urea in the presence and absence ofexogenous ammonia, and the amount of["C]urea uptake and intracellular ["4C]urea inthe two cultures was compared to the amount ofallophanate lyase induced in companion cul-tures receiving unlabeled urea (Table 5). Duringthe first 30 min, [4C ]urea uptake and theintracellular pool of ["4C]urea were unaffectedby the presence of ammonia in the medium.
However, only the culture without ammoniashowed induction of lyase. These results indi-cate that Chlamydomonas can accumulate in-ducer (or precursor of the inducer) in thepresence of ammonia and, therefore, that am-monia acts by an intracellular mechanism.The second possibility, that of repression, was
tested by the classical kinetic approach (32). Aculture growing with 3 mM urea was divided;one portion received 5 mM NH4NO3 and theother was left with urea. Ammonia caused anessentially immediate depression of carboxylaseand lyase accumulation per milliliter of culture(Fig. 7). The rate of carboxylase accumulationwas depressed to essentially zero, whereas lyasewas depressed to about 20% of the fully inducedrate. This differential effect of ammonia on therate of carboxylase and lyase accumulation permilliliter of culture is also reflected in the ratioof enzyme activity (Fig. 7, right inset). The"escape" of lyase from complete repression byammonia may account for the finding that morelyase than carboxylase is found in repressed andderepressed (nitrogen starved) cultures. Thesefindings are reminiscent of carbon cataboliterepression without transient repression in bac-terial systems (32), and of nitrogen metabolicrepression described for Aspergillus nidulans(3). It is not known whether Chlamydomonaspossesses nitrogen metabolite repression, aprocess described as affecting several ammonia-repressible pathways simultaneously (2, 3).Regulation of urea amidolyase activities in
Chlorella and Chlamydomonas compared.Thompson and Muenster reported (53) that the
1030 J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
18
Febr
uary
202
2 by
183
.104
.106
.140
.

UREA CATABOLISM IN CHLAMYDOMONAS AND CHLORELLA 1031
TABLE 5. Comparison of urea uptake and allophanate lyase induction of ammonia-grown Chlamydomonas inthe absence and presence of exogenous ammoniaa
[I"C jurea uptake"° Intracellular ["4C J urea""d Allophanate lyase activityeIncubation time (nmollml of suspension) (nmol/ml of suspension) (mU/ml of suspension)
(min)-NH,, +NH,+ -NH4+ +NH4+ -NH,+ +NH,+
0 0 0 0 0 0.020 0.02030 0.013 0.014 0.014 0.013 0.047 0.02060 0.312 0.021 0.080 0.021 0.440 0.020
'Culture was grown with 5 mM NH4NO, to 180 Klett units, washed, and suspended to 150 Klett units innitrogen-free medium (-NH4+ treatment) or 10 mM NH4NO, medium (+NH4+ treatment). Preincubated 15min.
[['ICJurea (16,990 disintegrations/min per nmol) added at 0 min to 0.3 mM. Cultures shaken at 27 to 28 Cwith 500 ft-c (ca. 5,000 lx) of white fluorescent light.
c At times indicated, 2.5 ml of suspension was filtered and cell radioactivity was counted. Data are means ofduplicates, adjusted for nonspecific binding of [14C Jurea to freeze-killed cells and for 14C lost from the cells as14CO2.
d At times indicated, 2.5 ml of suspension was filtered, the cells were lysed by freeze-thawing in phosphatebuffer, and the [14C Jurea content of lysates was determined from the "4CO2 released by jack bean urease. Dataare means of duplicates, adjusted for "4CO2 present in minus urease controls.
e Urea added (to 0.3 mM) at 0 min. Allophanate lyase activity assayed in the reaction system listed in Table3. Extract from 2 ml of cell suspension and 1 mM ["4C jallophanate (150 disintegrations/min per nmol) wereincubated 60 min at 30 C. Data are means of four assays.
cr7 uc AL
-J
74xo z l160
ZI HOHR
F- I- w
Sz 4,s**,,-
-2 0 2 4 6>T3HHOURS
2-~ ~ -7
o .64-'--A 4~~~~ 02
Z HOURS
-2 0 2 4 6 -2 0 2 4 6
TIME (HRS)FiG. 7. Repression kinetics of urea carboxylase (UC, left) and allophanate lyase (AL, right) upon addition of
ammonia to one-half of a Chlamydomonas culture growing with 3 mM urea. Symbols: 0, no addition; A,addition of 5 mM NH4NOs at the arrow. Left, Growth. Right, Ratio of enzyme activities. UC was assayed withthe reaction components given for urea amidolyase in the legend to Table 3, but with ["4C urea (1,549disintegrations/min per nmol) and 3 mU of Chlamydomonas allophanate lyase (brushite fraction 1). AL wasassayed with the reaction components given in the legend to Table 3, but with 2.5 Mlmol of ["4C allophanate (121disintegrations/min per nmol). Incubations were for 45 min at 30 C.
VOL. 121, 1975
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
18
Febr
uary
202
2 by
183
.104
.106
.140
.

HODSON, WILLIAMS, AND DAVIDSON
specific activity of urea carboxylase in C. vul-garis var. viridis was greater in urea-growncompared to ammonia-grown cells, whereas thespecific activity of allophanate lyase remainedunchanged. This response to nitrogen source byChlorella is dramatically different from theChlamydomonas system, and it prompted acomparison between representatives of the twospecies.
Cultures of Chlorella and Chlamydomonaswere grown organophototrophically to mid-exponential phase, with urea and ammonia asseparate nitrogen sources. Since cultures of thetwo species have different apparent absorban-cies, specific activities are expressed on thebasis of trichloroacetic acid-precipitable proteinleft in the extracts after centrifugation. Bothspecies had depressed urea amidolyase activitywhen they were grown with ammonia, andgrowth with urea induced this activity (Table6). The induction potential for Chlamydomonaswas about 200-fold greater than that in Chlorel-la. This difference is due both to the greaterlevel of urea amidolyase in repressed Chlorellaand to the greater level of induced activity inChlamydomonas. A further difference in regula-tion between the two species is found in theindividual enzyme activities. Urea amidolyasein Chlorella is inducible by virtue of only thecarboxylase; the lyase appears to behave as a
constitutive enzyme. In Chlamydomonas, as
shown above, both enzymes are inducible.These results with Chlorella confirm a previ-
ous observation (53) and indicate that regula-tion of urea catabolism in Chlorella may be
fundamentally different from that inChlamydomonas. It is indeed unfortunate thatgenetic recombination has not been demon-strated with Chlorella, so that the genetic basisfor these differences in regulation can be ex-
plored only with Chlamydomonas at this time.
DISCUSSIONThere are two potential endogenous sources of
urea in algal metabolism (29). One comes fromthe action of arginase (EC 3.5.3.1) in the argi-nine catabolic pathway, and the other comes
from allantoicase (EC 3.5.3.4) in purine degra-dation. Sussenbach and Strijkert (47) foundthat Chlamydomonas does not possess arginaseand, instead, degrades arginine to citrulline andammonia. The enzyme involved, arginine dei-midase (or desimidase, EC 3.5.3.6), was alsodescribed earlier as being responsible for argi-nine catabolism in Chlorella (42). Thus, tworepresentatives of different major taxons withinthe green algae are not able to form urea fromarginine. It remains to be seen if this is a generalfeature of chlorophycean algae. With respect tourea from purines, Amman and Lynch (1)showed in nutritional studies that a strain of C.pyrenoidosa could utilize adenine, hypoxan-thine, xanthine, and uric acid as sole nitrogensources for growth. We have observed in prelim-inary work (60) that Chlamydomonas can uti-lize xanthine but not adenine; hypoxanthineand uric acid were not tested by us. The usualpathway of uric acid to urea involves thesequential action of uricase (EC 1.7.3.3), allan-toinase (EC 3.5.2.5), and allantoicase (EC
TABLE 6. Specific activities of enzymes of urea hydrolysis pathway from Chlamydomonas and Chlorella grownwith urea or ammoniaa
Enzymatic activityc (gU/mg of protein)
Alga Nitrogen sourcefor growthb Urea amidolyase Urea carboxylase Allophanate
lyase
Chlamydomonas reinhardi (y-1) Urea 11,000 20,000 67,000NH4NO3 15 (2) 3.4 (6.5) 260 (20)
Chlorella pyrenoidosa (3E) Urea 1,400 4,000 (100) 21,000 (1,000)NH4Cl 370 (10) 1,200 (100) 25,000
a The amounts of culture assayed were: Chlamydomonas, 1 ml at 115 Klett units (urea grown) and 24 ml at119 Klett units (NH4+ grown); Chlorella, 8 ml at 234 Klett units (urea grown) and 8 ml at 166 Klett units (NH4+grown). Crude homogenates of Chlamydomonas were prepared by sonic treatment; those of Chlorella wereprepared by using a French pressure cell. All incubations were for 60 min at 25 C. Urea amidolyase and allopha-nate lyase were assayed with the reaction components given in footnote a to Table 3. Urea carboxylase wasassayed as for amidolyase plus either 6 mU (for Chlorella extract) or 9 mU (for Chlamydomonas extract) ofallophanate lyase. Substrate specific activities: [C4C]urea, 1,417 disintegrations/min per nmol; [4C ]allo-phanate, 456 disintegrations/min per nmol. One microunit; 1 pmol of "CO2 formed per min.
" Concentrations: Urea, 3 mM; NH4NO3, 5 mM; NH4Cl, 10 mM.c Numbers in parentheses are standard errors of the mean; they are omitted where smaller than the last
significant digit.
1032 J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
18
Febr
uary
202
2 by
183
.104
.106
.140
.

UREA CATABOLISM IN CHLAMYDOMONAS AND CHLORELLA
3.5.3.4). Villeret detected allantoinase activityin toluenized cells of Chlamydomonas andChlorella, but did not detect allantoicase ineither strain (54). Until the absence of allantoi-case is confirmed with other enzyme extractionmethods, it remains possible that Chlorella andChlamydomonas have the usual pathway forpurine catabolism and that the urea amidolyasesystem plays a role in recycling endogenouspurine nitrogen. Substrates for urea amidolyaseother than urea have been sought in yeast andChlorella, but none has been found (38, 53, 58).
If urea amidolyase is necessary, or important,in the recovery of nitrogen from endogenousurea, then it is curious that we have notobserved conditions other than culturing on
urea that give rise to significant levels of theenzymatic activity in Chlamydomonas, i.e.,autoinduction. A common ploy with Chlorellato obtain rapid urea catabolism is to starve theorganism of nitrogen for several days (19). SinceChlamydomonas is capable of a reproductivecycle involving gametic differentiation and zy-
gote formation in addition to the usual vegeta-tive cycle shared with Chlorelle, it is conceiv-able that urea amidolyase in Chlamydomonasmight be confined to events surrounding sexualdifferentiation. Nitrogen starvation promotesgametogenesis, and sexual competency isachieved in asychronous cultures during theperiod of starvation employed here (5 to 8 h[31]), so it is likely that we have alreadyincluded the early period of the reproductivecycle in our studies. It is also possible that urea
amidolyase is significant when Chlamydomonasis grown on a nitrogen source other than urea or
ammonia, such as nitrate. Unfortunately, they-1 strain is unable to utilize nitrate.The urea amidolyase system in
Chlamydomonas, as in Chlorella (51), is shownto consist of two discrete enzymatic activitiesthat carboxylate urea and then hydrolyze it toammonium ion and bicarbonate. The enzymes,urea carboxylase and allophanate lyase, are
readily separated and may be assayed individ-ually in the presence of each other. Urea carbox-ylase probably requires biotin because it isinhibited by egg white avidin, and it alsorequires ATP and Mg2+. These requirementsare consistent with an energy-dependent car-
boxylation of urea. Allophanate lyase does notappear to have cofactor requirements. Theseproperties of amidolyase systems from two dif-ferent unicellular algae are similar to those ofyeast, except that yeast carboxylase and lyasehave not been physically separated from eachother with retention of enzymatic activity.
All of the amidolyase systems examined todate are metabolically regulated. Of these,Chlamydomonas seems to have one of thelargest potentials for response to nutritionalconditions. Growth on ammonia allows only atrace of carboxylase and lyase activity, andderepression by nitrogen starvation increasesboth by less than a factor of two. But in thepresence of urea, both enzymatic activities areinduced to levels as much as 100 times over thebasal or derepressed levels. In addition, am-monia overrides urea when the two are pre-sented to Chlamydomonas at the same time.Thus, urea amidolyase is inducible rather thanderepressible, and is also subject to repression.This type of dual control has been shown forurea amidolyase in Candida (39) and Sac-charomyces (59), and for nitrite reductase inNeurospora (12) as well as for other enzymesystems in eukaryotes, and it has inspired aninteresting model (8).One of the perhaps less interesting ways in
which apparent repression can occur is byinducer exclusion (30, 32). In our case this hasbeen made unlikely by showing that [4C ]ureauptake by repressed cells is not reduced byammonia. A more likely means for repressioncontrol is found in recent work on Aspergillusnidulans (3, 4), where several ammonia-produc-ing enzyme systems are simultaneously re-pressed by ammonia, aptly called nitrogen met-abolic repression. It may be significant for thisargument that, in Chlamydomonas, ammoniarepresses nitrate and nitrite reductases (16) aswell as arginine deimidase (45). It remains to beseen whether there is evidence for a commonmechanism, perhaps a single kind of mac-romolecule, coupling these various repressions.The true inducer may not be urea but a
metabolic product of urea. Whitney and Cooper(59) have proposed that yeast urea amidolyaseis induced by allophanate, the product of thefirst enzyme activity.
Control mechanisms operating at the enzymelevel have been considered here only briefly. Wehave no evidence for feedback inhibition of ureacarboxylase by ammonia or for inactivation byammonia such as described by Herrera et al.(16) for nitrate reductase. Ammonium sulfateup to 0.1 M has not been inhibitory to ureacarboxylase or allophanate lyase in cell-freesystems (Hodson, unpublished observations). Itis possible that ATP or the "energy charge"may regulate carboxylase activity. Roon andLevenberg (39) reported that adenosine 5'-diphosphate inhibited the carboxylase in Can-dida, but this has not been investigated further.
VOL. 121, 1975 1033
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
18
Febr
uary
202
2 by
183
.104
.106
.140
.

HODSON, WILLIAMS, AND DAVIDSON
One of the questions that has intrigued inves-tigators of controlled systems in eukaryotes iswhether the genes specifying those systemsoccur in operons (33). The kinetic evidence wenow have on repression of urea carboxylase andallophanate lyase in Chlamydomonas does notfit the bacterial model (36). Repressionquenches the accumulation of carboxylase permilliliter of culture, whereas the lyase continuesto accumulate, albeit at a much reduced rate.On the other hand, induction is not obviouslysequential in the order that the enzymes func-tion in the pathway, as has been suggested fornitrate and nitrite reductases in the duckweedLemna minor (44).
ACKNOWLEDGMENTS
We wish to thank John F. Thompson, Cornell University,and R. J. Ellis, Bucknell University, for helpful suggestions,and David E. Sheppard, University of Delaware, for criticaldiscussions and reading of the manuscript. The assistance ofNancy Lomax, Teresa Della-Volpe, Susan Wilson, and AnnSiberski is gratefully acknowledged.
This work was supported by grants from the University ofDelaware Research Foundation.
LITERATURE CITED
1. Amman, E. C. B., and V. H. Lynch. 1964. Purinemetabolism by unicellular algae. II. Adenine, hypoxan-thine and xanthine degradation by Chlorella pyrenoi-dosa. Biochim. Biophys. Acta 87:370-379.
2. Arst, H. N., Jr., and D. J. Cove. 1969. Methylammoniumresistance in Aspergillus nidulans. J. Bacteriol.98:1284-1293.
3. Arst, H. N., Jr., and D. J. Cove. 1973. Nitrogen metabo-lite repression in Aspergillus nidulans. Mol. Gen.Genet. 126:111-141.
4. Arst, H. N., Jr., and M. M. Page. Mutants of Aspergillusnidulans altered in the transport of methylammoniumand ammonium. Mol. Gen. Genet. 121:239-245.
5. Cook, J. R., and M. Carver. 1966. Partial photo-repres-sion of the glyoxylate by-pass in Euglena. Plant CellPhysiol. 7:377-383.
6. Cove, D. J. 1966. The induction and repression of nitratereductase in the fungus Aspergillus nidulans. Biochim.Biophys. Acta 113:51-56.
7. Cove, D. J. 1970. Control of gene action in Aspergillusnidulans. Proc. Roy. Soc. London Ser. B 176:267-275.
8. Cove, D. J., and J. A. Pateman. 1969. Autoregulation ofthe synthesis of nitrate reductase in Aspergillus nidu-lans. J. Bacteriol. 97:1374-1378.
9. Dains, F. B., and E. Wertheim. 1920. The action ofammonia and amines on the substituted ureas andurethanes. II. Allophanic ester. J. Biol. Chem.42:2303-2309.
10. Dunn, E., and J. A. Pateman. 1972. Urea and thioureauptake in Aspergillus nidulans. Heredity 29:129.
11. Ebersold, W. T., R. P. Levine, E. E. Levine, and M. A.Olmsted. 1962. Linkage maps in Chlamydomonas rein-hardi. Genetics 47:531-543.
12. Garrett, R. H. 1972. The induction of nitrite reductase inNeurospora crassa. Biochim. Biophys. Acta264:481-489.
13. Goulding, K. H. and M. J. Merrett. 1966. The photome-tabolism of acetate by Chlorella pyrenoidosa. J. Exp.Bot. 17:678-689.
14. Grant, B. R., J. Madgwick, and G. Dal Pont. 1967.
Growth of Cylindrotheca closterium var. Californica(Mereschk.) Reimann and Lewin on nitrate, ammonia,and urea. Aust. J. Mar. Freshwater Res. 18:129-136.
15. Hackette, S. L., G. E. Skye, C. Burton, and I. H. Segel.1970. Characterization of an ammonium transportsystem in filamentous fungi with methylammonium-IC as the substrate. J. Biol. Chem. 245:4241-4250.
16. Herrera, J. A. Paneque, J. M. Maldonado, J. L. Barea,and M. Losada. 1972. Regulation by ammonia ofnitrate reductase synthesis and activity inChlamydomonas reinhardi. Biochem. Biophys. Res.Commun. 48:996-1003.
17. Hodson, R. C. 1973. Nutritional control of urea metabo-lism in Chiamydomonas reinhardi (strain y-1). PlantPhysiol. 51(Suppl.):55.
18. Hodson, R. C., J. A. Schiff, and A. J. Scarsella. 1968.Studies of sulfate utilization by algae. 7. In vivometabolism of thiosulfate by Chlorella. Plant Physiol.43:570-577.
19. Hodson, R. C., and J. F. Thompson. 1969. Metabolism ofurea by Chlorella vulgaris. Plant Physiol. 44:691-696.
20. Holden, J., and I. Morris. 1970. Regulation of argininebiosynthesis in Chlamydomonas reinhardi: studies invivo and of ornithine transcarbamylase and ar-gininosuccinate lyase activities. Arch. Mikrobiol.74:58-68.
21. Hudock, G. A. 1963. Repression of argininosuccinase inChlamydomonas reinhardi. Biochem. Biophys. Res.Commun. 10:133-138.
22. Kates, J. R., and R. F. Jones. 1967. Periodic increases inenzyme activity in synchronized cultures ofChlamydomonas reinhardi. Biochim. Biophys. Acta145:153-158.
23. Knutsen, G. 1965. Induction of nitrite reductase insynchronized cultures of Chlorella pyrenoidosa. Bio-chim. Biophys. Acta 103:495-502.
24. Layne, E. 1957. Spectrophotometric and turbidometricmethods for measuring proteins, p. 447-454. In S. P.Colowick and N. 0. Kaplan (ed.), Methods in enzymol-ogy, vol. 3. Academic Press Inc., New York.
25. Leftley, J. W., and P. J. Syrett. 1973. Urease andATP:urea amidolyase activity in unicellular algae. J.Gen. Microbiol. 77:109-115.
26. Levine, R. P., and W. T. Ebersold. 1960. The genetics andcytology of Chlamydomonas. Annu. Rev. Microbiol.14:197-216.
27. Lien, T., and G. Knutsen. 1973. Synchronous cultures ofChlamydomonas reinhardi: properties and regulationof repressible phosphatases. Physiol. Plant.28:291-298.
28. Losada, M., A. Paneque, P. J. Aparicio, J. M. Vega, J.Cardenas, and J. Herrera. 1970. Inactivation andrepression by ammonium of the nitrate reducing sys-
tem in Chlorella. Biochem. Biophys. Res. Commun.38:1009-1015.
29. Lui, N. S. T., and 0. A. Roels. 1970. Nitrogen metabolismof aquatic organisms. 1. The assimilation and forma-tion of urea in Ochromonas malhamensis. Arch. Bio-chem. Biophys. 139:269-277.
30. McCarthy, J. J., and R. W. Eppley. 1972. A comparisonof chemical, isotopic and enzymatic methods for mea-
suring nitrogen assimilation of marine phytoplankton.Limnol. Oceanogr. 17:371-382.
31. McCullough, W., and P. C. L. John. 1972. Control of denovo isocitrate lyase synthesis in Chlorella. Nature(London) 239:402-405.
32. Magasanik, B. 1970. Glucose effects: inducer exclusionand repression, p. 189-220. In J. R. Beckwith and D.Zipser (ed.), The lactose operon. Cold Spring HarborLaboratory, Cold Spring Harbor.
33. Metzenberg, R. L. 1972. Genetic regulatory systems inNeurospora. Annu. Rev. Genet. 6:111-132.
1034 J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
18
Febr
uary
202
2 by
183
.104
.106
.140
.

VOL. 121, 1975 UREA CATABOLISM IN CHLAMYDOMONAS AND CHLORELLA
34. Morris, T., and P. J. Syrett. 1963. The development ofnitrate reductase in Chlorella and its repression byammonium. Arch. Mikrobiol. 47:32-41.
35. Orth, G. M., N. E. Tolbert, and E. Jimenez. 1966. Rate ofglycolate formation during photosynthesis at high pH.Plant Physiol. 41:143-147.
36. Richmond, M. H. 1968. Enzymatic adaptation in bacte-ria: its biochemical and genetic basis, p. 105-154. In P.N. Campbell and G. D. Greville (ed.), Essays inbiochemistry, vol. 4. Academic Press Inc., New York.
37. Roon, R. J., J. Hampshire, and B. Levenberg. 1972. Ureaamidolyase. II. The involvement of biotin in ureacleavage. J. Biol. Chem. 247:7539-7545.
38. Roon, R. J., and B. Levenberg. 1970. ATP:urea amidoly-ase (ADP) (Candida utilis), p. 317-324. In S. P.Colowick and N. 0. Kaplan (ed.), Methods in enzymol-ogy, vol. 17. Academic Press Inc., New York. X
39. Roon, R. J., and B. Levenberg. 1972. Urea amidolyase. I.Properties of the enzyme from Candida utilis. J. Biol.Chem. 247:4107-4113.
40. Schiff, J. A., and M. Levinthal. 1968. Studies of sulfateutilization by algae. 4. Properties of a cell-free sulfate-reducing system from Chlorella. Plant Physiol.43:547-554.
41. Schmeisser, E. T., D. M. Baumgartel, and S. H. Howell.1973. Gametic differentiation in Chlamydomonas rein-hardi: cell cycle dependency and rates in attainment ofmating competency. Develop. Biol. 31:31-37.
42. Shafer, J., Jr., and J. F. Thompson. 1968. Argininedesimidase in Chlorella. Phytochemistry 7:391-399.
43. Siegelman, H. W., G. A. Wieczarek, and B. C. Turner.1965. Preparation of calcium phosphate for proteinchromatography. Anal. Biochem. 13:402-404.
44. Stewart, G. R. 1968. The effect of cycloheximide on theinduction of nitrate and nitrite reductase in Lemnaminor L. Phytochemistry 7:1139-1142.
45. Strijkert, P. J., R. Loppes, and J. S. Sussenbach. 1971.Arginine metabolism in Chlamydomonas reinhardi.Regulation of uptake and breakdown. FEBS Lett.14:329-332.
46. Strijkert, P. J., and J. S. Sussenbach. 1969. Argininemetabolism in Chlamydomonas reinhardi. Evidencefor a specific regulatory mechanism of the biosynthesis.Eur. J. Biochem. 8:408-412.
47. Sussenbach, J. S., and P. J. Strijkert. 1969. Argininemetabolism in Chlamydomonas reinhardi. Argininedeiminase: the first enzyme of the catabolic pathway.FEBS Lett. 3:166-168.
48. Sussenbach, J. S., and P. J. Strijkert. 1969. Argininemetabolism in Chlamydomonas reinhardi. On the
regulation of the arginine biosynthesis. Eur. J. Bio-chem. 8:403-407.
49. Syrett, P. J. 1966. The kinetics of isocitrate lyase forma-tion in Chlorella: evidence for the promotion of enzymesynthesis by photophosphorylation. J. Exp. Bot.17:641-654.
50. Talley, D. J., L. H. White, and R. R. Schmidt. 1972.Evidence for NADH- and NADPH-specific isozymes ofglutamate dehydrogenase and the continuous induci-bility of the NADPH-specific isozyme throughout thecell cycle of the eucaryote Chlorella. J. Biol. Chem.247:7927-7935.
51. Thompson, J. F., and A.-M. E. Muenster. 1971. Separa-tion of the Chlorella ATP:urea amidolyase into twocomponents. Biochem. Biophys. Res. Commun.43:1049-1055.
52. Thompson, J. F., and A.-M. E. Muenster. 1972.ATP-dependent urease of Chlorella. Plant Physiol.49(Suppl.):35.
53. Thompson, J. F., and A.-M. E. Muenster. 1974. ATP-dependent urease: characterization of and control inChlorella; the search for it in higher plants. Bull. Roy.Soc. New Zealand. 12:91-97.
54. Villeret, M. S. 1955. Sur la presence des enzymes desureides glyoxyliques chez les Algues d'eau douce. C. R.Acad. Sci. 241:90-92.
55. Whitney, P. A., and T. G. Cooper. 1970. Requirement forHCO,- by ATP:urea amido-lyase in yeast. Biochem.Biophys. Res. Commun. 40:814-819.
56. Whitney, P. A., and T. G. Cooper. 1972. Urea carboxylaseand allophanate hydrolase; two components of a mul-tienzyme complex in Saccharomyces cerevisiae. Bio-chem. Biophys. Res. Commun. 49:45-51.
57. Whitney, P. A., and T. G. Cooper. 1972. Urea carboxylaseand allophanate hydrolase. Two components of adeno-sine triphosphate: urea amido-lyase in Saccharomycescerevisiae J. Biol. Chem. 247:1349-1353.
58. Whitney, P. A., and T. Cooper. 1973. Urea carboxylasefrom Saccharomyces cerevisiae. Evidence for a mini-mum two-step reaction sequence. J. Biol. Chem.248:325-330.
59. Whitney, P. A., T. G. Cooper, and B. Magasanik. 1973.The induction of urea carboxylase and allophanatehydrolyase in Saccharomyces cerevisiae. J. Biol. Chem.248:6203-6209.
60. Williams, S. K., W. R. Davidson, and R. C. Hodson. 1974.Regulation of enzymes catalyzing urea hydrolysis inChiamydomonas reinhardi (strain y-1). Plant Physiol.Suppl., p. 13.
1035
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
18
Febr
uary
202
2 by
183
.104
.106
.140
.