mercury’s mitochondrial targeting with increasing age in scrobicularia plana inhabiting a...
TRANSCRIPT

Chemosphere 92 (2013) 1231–1237
Contents lists available at SciVerse ScienceDirect
Chemosphere
journal homepage: www.elsevier .com/locate /chemosphere
Mercury’s mitochondrial targeting with increasing age in Scrobiculariaplana inhabiting a contaminated lagoon: Damage-protection dichotomyand organ specificities
0045-6535/$ - see front matter � 2013 Elsevier Ltd. All rights reserved.http://dx.doi.org/10.1016/j.chemosphere.2013.04.054
⇑ Corresponding author at: Department of Chemistry & CESAM, University ofAveiro, 3810-193 Aveiro, Portugal. Tel.: +351 234401527x24907; fax: +351234370084.
E-mail address: [email protected] (I. Ahmad).
Iqbal Ahmad a,b,⇑, Iram Mohmood a, Mário Pacheco b, Maria A. Santos b, Armando C. Duarte a,Eduarda Pereira a
a Department of Chemistry & CESAM, University of Aveiro, 3810-193 Aveiro, Portugalb Department of Biology & CESAM, University of Aveiro, 3810-193 Aveiro, Portugal
h i g h l i g h t s
�Mitochondria proved a sensitive fraction for mercury effects in Scrobicularia plana.� S. plana age determined mercury induced damage vulnerability (younger > older).� A decreasing trend of TAC with increasing age was also observed at M.� Equal sensibility of animal or gill towards mitochondrial responses was observed.� Organs (gills and digestive glands) specificity was evident.
a r t i c l e i n f o
Article history:Received 2 August 2012Received in revised form 15 April 2013Accepted 20 April 2013Available online 25 May 2013
Keywords:MercuryMitochondriaScrobicularia planaAgeOrgan-specificitiesTotal antioxidants capacity
a b s t r a c t
This study aimed to understand bivalves’ (Scrobicularia plana) adaptive strategies, with emphasis onmitochondria as a target organelle, in response to mercury-contaminated environment. Inter-age andorgan-specific approaches were applied using different annual size classes (2+, 3+, 4+ and 5+ years old)and assessing specific organs (gill, digestive gland), respectively. Bivalves were collected from moderately(M) and highly (H) contaminated sites at Laranjo basin - Ria de Aveiro (Portugal), where a mercury gra-dient was identified, and compared with those from a reference (R) site. Total antioxidants capacity (TAC)was measured in mitochondria, whereas the lipid peroxidation was assessed as a marker of damage. S.plana age determined the clams’ vulnerability towards mercury induced mitochondrial peroxidativedamage depending upon the mercury accumulation: younger animals were more vulnerable than older.Clams showed a decreasing trend of TAC with increasing age. This decrease was found to be statisticallysignificant in 4+ and 5+ years at M; whereas, at H, with depleted TAC, remained same until they havegrown 5+ years. The organ specificity was evident for antioxidant response and peroxidative damage. Aclear pattern of overall TAC increase in digestive gland (at M) and decrease in gill (at H) was observed,while only gills were found to be susceptible to peroxidative damage. Overall, mitochondria proved tobe a sensitive fraction for the effects of mercury in S. plana inhabiting mercury contaminated area.
� 2013 Elsevier Ltd. All rights reserved.
1. Introduction cell death. Available informations in fish evidenced mitochondria
Oxidative stress endpoints are frequently assessed in aquaticorganisms as sensitive markers of exposure to environmentalstressors. However, most of the time, the investigation focusseson nonspecific cell fractions, overlooking the crucial and specificrole of mitochondria in reactive oxygen species (ROS) productionand adequate oxidative stress leading to organelle dysfunction or
‘a target organelle’ to be susceptible to environmental pollutantsincluding mercury. For example, dietary methyl mercury has beenshown to inhibit mitochondrial energy metabolism in skeletalmuscles of the Danio rerio (Cambier et al., 2009). Lethal effects ofinorganic mercury on cells and tissues of Trichomycterus brasiliensishave been shown to cause mitochondrial dysfunction (Ribeiroet al., 1996). In addition to an abundance of myelinoid bodies, his-tocytological responses of Dicentrarchus labrax following mercuryexposure exhibited alterations to the mitochondria (Giari et al.,2008). However, nothing is known about effects of mercury onmitochondrial ROS production in marine mollusks, which serveas a conduit for bottom-up energy transfer on the food webs. Also,

1232 I. Ahmad et al. / Chemosphere 92 (2013) 1231–1237
mitochondrial oxidant production with respect to mollusk’s ageand specific organ remains unexplored. In general, aging and lifespan shortening are associated with oxidative damage arising fromcorresponding changes in mitochondrial oxidant production. Con-sidering the organ specific response, antioxidant activities tend tobe higher in metabolically active tissues like liver versus gill in fish(Lemaire et al., 1993; Ansaldo et al., 2000), digestive gland versusbody wall in polychaetes (Buchner et al., 1996) and gill versusmantle tissue in cephalopods (Zielinski and Portner, 2000). Whenmitochondria become dysfunctional, for example, through long-term exposure to environmental toxicants, they produce less cellenergy and more ROS having its consequences at supra-cellular le-vel. To the author’s knowledge, so far no study has looked at theage and organ specific effects of metal on mitochondrial ROS pro-duction and damage in marine bivalve.
Infaunal bivalves cannot regulate metal uptake from the envi-ronment due to their close contact with water, sediment (Connelland Miller, 1984) and filter feeding nature, but can regulate its up-take in their tissue to an extent through metallothioneins, gran-ules, lipofuscin. As a result, bivalves are more likely to betargeted by the environmental mercury contamination. The obser-vations based on the post-mitochondrial supernatant (PMS) frac-tion in Peppery furrow shell Scrobicularia plana demonstrated thedevelopment of biochemical adaptations to environmental mer-cury exposure despite the weakening of their immune system (Ah-mad et al., 2011a,b, 2012a). In gill and digestive gland of S. plana(Ahmad et al., 2012a) environmentally exposed to mercury, or-gan-specific responses were evident to a pro-oxidant challengeposed by mercury. The differences were particularly evident forthiol compounds, since only gill displayed the potential to respondto moderate levels of mercury by increasing non-protein thiol andtotal glutathione contents. Under high contamination habitat, thedifferences between gill and digestive gland were mitigated, asboth organs were unable to increase thiol compounds, which werecompensated by the elevation of ascorbic acid content. In wholebody, the adaptive capacity to pro-oxidant challenge caused bymercury contamination, was dependent on its contamination ex-tent i.e. under moderate contamination, the intervention of the dif-ferent antioxidants took place harmoniously, evidencing anadjustment capacity with increasing age. Under higher contamina-tion, S. plana failed to cope with mercury exposure resulting in anage-dependent deterioration of the defence abilities (Ahmad et al.,2012a). Moreover, the comparison of whole-body and organs re-vealed that the oxidative stress responses were organ specificand whole-body analyses may be misleading in addressing and ex-actly revealing oxidative stress responses (Ahmad et al., 2011b).Thus, the main focus of the study was to provide an explanationof animal’s adjustment capacity to mercury exposure by assessingmitochondria as a target organelle of mercury. This study aimed to:(i) evaluate the clams adaptive strategy over time by assessing theage related variations on defence and damage responses; and (ii)assess organ-specificities by analyzing gill and digestive gland re-sponses to understand bivalves’ better adaptation under mercurystress conditions. Total antioxidants capacity (TAC) was assessedin mitochondria to overcome the uncertainty on single antioxidantdefences, whereas the lipid peroxidation (LPO) was assessed as amarker of damage. The study included S. plana, which is a widelydistributed, burrowing and deposit-feeding bivalve, inhabiting softbottoms of clay or mud in the inter-tidal zone of estuaries (Woot-ton and Pipe, 2003). The study area was Laranjo basin at Ria deAveiro (a coastal lagoon in north-western Portugal), where awell-defined mercury gradient was identified due to chlor-alkaliplant discharges. Due to the absence of other contaminant sources,this area has been regarded as a ‘field laboratory’ for assessingmercury toxicity under realistic environmental conditions (Guil-herme et al., 2008).
2. Materials and methods
2.1. Study area and sampling
Laranjo basin is the most mercury-contaminated area in the Riade Aveiro (Portugal), which received chlor-alkali plant dischargescontinuously for five decades, resulting in the generation of a mer-cury contamination gradient. Though in recent past (approxi-mately 15 years ago) this industry has stopped the effluentrelease, high mercury levels are still present in the sediments (Ah-mad et al., 2011a) and its progressive re-suspension, mainly duringthe periods of stronger tidal currents, are responsible for metalexportation and increased bioavailability (Pereira et al., 2009).
The sampling was carried out in July 2008 in three sites: twosites at Laranjo basin, termed as M (moderately contaminated)and H (highly contaminated), and a third site as reference (R), atVagueira, considered unpolluted including in terms of mercury(Fig. 1). This site belongs to an area, far from the main mercury pol-lution source (around 30 km), where neither industry in its sur-rounding, nor urban effluent outlets of noteworthy importance ispresent (Duarte et al., 2007; Pereira et al., 2009). The sites selectionwas based on a previous survey (Coelho et al., 2006), revealing theglobal mercury contamination status of Ria de Aveiro and pointingout some vulnerable areas. At each site, S. plana were collected dur-ing low tide conditions by digging out to a depth of 30 cm on themudflats and washed through 1 mm mesh bags. The collected ani-mals were divided into age groups, classified according to the shellyearly growth increments as proposed by Verdelhos et al. (2005):2+, 3+, 4+ and 5+ year old clams.
The present study encompassed two components: (i) inter-ageapproach – where all the parameters were assessed in the wholesoft tissues, addressing inter-age comparison and thus, using dif-ferent age classes (2+ to 5+ year old clams); (ii) organ-specific ap-proach – where all the parameters were assessed in specificorgans, viz. gill and digestive gland, in 4+ year old clams (the choiceof this class was based on animal’s availability and the body massin order to provide enough material to analyze). Accordingly, tenanimals destined for inter-age approach were dissected and thewhole soft tissues were immediately frozen in liquid nitrogenand stored at �80 �C for antioxidants assessment. Animals devotedto the organ-specific approach also included ten specimens of the4+ year old class. Animals were dissected and gill and digestivegland were collected for antioxidants assessment. The methodol-ogy in terms of animals handling and sampling processing wasthe same as previously described for inter-age approach.
2.2. Biochemical analyses
2.2.1. Isolation of mitochondriaMitochondria were isolated according to the method of Sayeed
et al. (2006). All procedures were carried out on ice or at 4 �C. Thepreviously collected whole animal or organs were dissected intoice-cold isolation medium (0.25 M sucrose and 1 mM EDTA, pH7.4), minced in fresh isolation medium and gently homogenizedmanually (10%, w/v in isolation medium) using a homogenizerwith a loose-fitting pestle. The homogenate was centrifuged at500g for 10 min. The supernatant fraction was retained, whereasthe pellet was washed with fresh isolation medium and recoveredby centrifugation at 500g for 10 min. The supernatant fraction fromthis wash was combined with the initial supernatant fraction andpooled fractions were centrifuged at 500g for 10 min. The superna-tant thus obtained was centrifuged at 5000g for 15 min to obtainthe mitochondrial pellet. The pellet was washed first with isolationmedium and then with the respiration reaction buffer (70 mM su-crose, 220 mM mannitol, 2 mM HEPES, 0.5 mM EDTA, 2.5 mM

Fig. 1. (A) Location of Ria de Aveiro (Portugal); (B) Location (j) of sampling sites at Ria de Aveiro: moderately (M) and highly (H) mercury contaminated sites at Laranjo basin,and a site at Vagueira assumed as reference (R).
I. Ahmad et al. / Chemosphere 92 (2013) 1231–1237 1233
MgCl2, 0.5 mM KH2PO4, and 2 mM K2HPO4, pH 7.4), and thepurified mitochondria were obtained by centrifugation at 12,000gfor 10 min. The final mitochondrial pellet was re-suspended inrespiration reaction buffer (1 mL g�1 tissue) to produce a suspen-sion containing 25–40 mg of mitochondrial protein mL�1. Onemitochondrial suspension per animal/tissue was obtained. Usingthree replicates each of 4 mg mL�1 of mitochondrial suspensionfrom each individual animal for lipid peroxidation assay, rest mito-chondrial suspension was kept for antioxidant evaluation. Prior tothe assay, different dilutions of the test samples with 1� assaybuffer were made to bring the antioxidants level within the rangeof standard curve (0.015–0.42 mM). Trolox dilutions with 1� assaybuffer were used to create a standard curve. Then, 10 lL of testsamples from each replicate were processed for TAC assay follow-ing the instruction given on leaflet. Assays were performed induplicate using ultra pure water. Due to lack of samples for theprocessing of mitochondrial verifications, the obtained fractionby differential centrifugation was directly used for variousbiochemical parameters as adopted by Rigobello et al. (1995), Kris-tal et al. (1997), Jang et al. (2000), Tabassum et al. (2010) and An-jum et al. (2011). Total protein contents were determinedaccording to the Biuret method (Gornall et al., 1949), using bovineserum albumin as a standard.
2.2.2. Mitochondrial total antioxidants capacityMitochondrial total antioxidants capacity using Antioxidant As-
say Kit (CS0790-1KT, Sigma) following instruction provided on theleaflet. The principle of the antioxidants assay was the formation ofa ferryl myoglobin radical from metmyoglobin and hydrogen per-oxide, which oxidizes the ABTS (2,20-azino-bis (3-ethylbenzthiazo-line-6-sulfonic acid) to produce a radical cation, ABTS�+, a soluble
chromogen that is green in color and the absorbance of the con-tents was recorded at 405 nm using a standard 96-well plate read-er (UV/VIS, Spectramax 384). Trolox, a water-soluble vitamin Eanalog, was used as a standard or control antioxidant in this assay.Results were expressed as mM antioxidant concentration mg�1
mitochondrial protein.
2.2.3. Mitochondrial lipid peroxidationThiobarbituric acid reactive substances (TBARS) were measured
spectrophotometrically as described by Buege and Aust (1978).Briefly, mitochondrial preparation (4 mg mL�1) was mixed withan equal volume of Buege and Aust reagent (trichloroacetic acid,15% w/v in 0.25 M HCl and thiobarbituric acid, 0.37% w/v in0.25 M HCl) and heated for 15 min in boiling water. After cooling,the precipitate was removed by centrifugation at 1000g for10 min at room temperature. The absorbance of the supernatantwas measured at 532 nm. The concentration of TBARS was deter-mined using an extinction coefficient of 1.56 � 105 and resultswere expressed as nmol of TBARS mg�1 of mitochondrial protein.
One absorbance measurement from each of the three replicatesof an animal tissue was made. Standard quality assurance/qualitycontrol (QA/QC) procedures included the use of method blank, ref-erence standards and calibration verifications.
2.3. Data analyses
SPSS (PASW statistics 18) for Windows was used for statisticalanalyses of data. Descriptive statistics was initially performed onthe data and homogeneity of variance was tested. ANOVA analysisfollowed by the Tukey’s test was performed in order to compare re-sults between sites (within the same class) and between classes
1234 I. Ahmad et al. / Chemosphere 92 (2013) 1231–1237
(within the same site), while correlations between variables weretested through Pearson’s correlation. All the results significancewas ascertained at a = 0.05.
3. Results
Inter-age and organ-specific approach is presented below in de-tail. Besides the results comparison with reference site (R), themitochondrial responses relationships between and within the or-gans (gill and digestive gland) and whole body versus organs rela-tionship for 4+ year clams have also been analyzed statistically. Theresults have been depicted as examining each studied parameter inseparate paragraphs starting from between-site analysis followedby the between-age analysis.
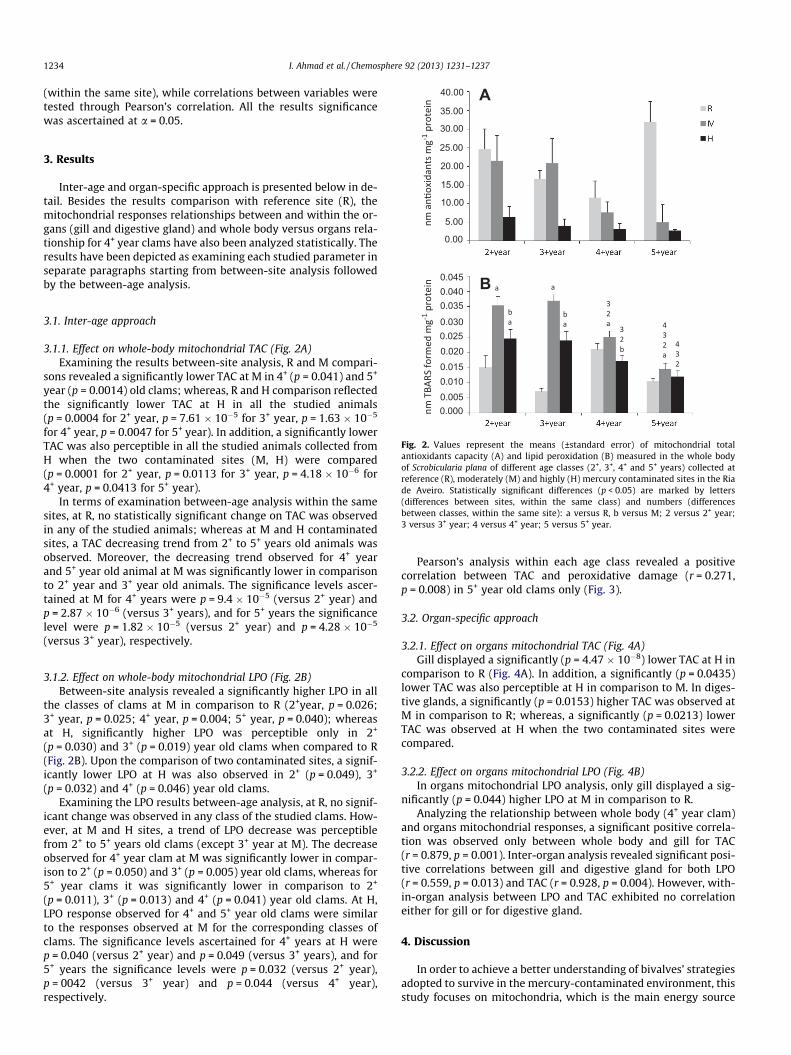
Fig. 2. Values represent the means (±standard error) of mitochondrial totalantioxidants capacity (A) and lipid peroxidation (B) measured in the whole bodyof Scrobicularia plana of different age classes (2+, 3+, 4+ and 5+ years) collected atreference (R), moderately (M) and highly (H) mercury contaminated sites in the Riade Aveiro. Statistically significant differences (p < 0.05) are marked by letters(differences between sites, within the same class) and numbers (differencesbetween classes, within the same site): a versus R, b versus M; 2 versus 2+ year;3 versus 3+ year; 4 versus 4+ year; 5 versus 5+ year.
3.1. Inter-age approach
3.1.1. Effect on whole-body mitochondrial TAC (Fig. 2A)Examining the results between-site analysis, R and M compari-
sons revealed a significantly lower TAC at M in 4+ (p = 0.041) and 5+
year (p = 0.0014) old clams; whereas, R and H comparison reflectedthe significantly lower TAC at H in all the studied animals(p = 0.0004 for 2+ year, p = 7.61 � 10�5 for 3+ year, p = 1.63 � 10�5
for 4+ year, p = 0.0047 for 5+ year). In addition, a significantly lowerTAC was also perceptible in all the studied animals collected fromH when the two contaminated sites (M, H) were compared(p = 0.0001 for 2+ year, p = 0.0113 for 3+ year, p = 4.18 � 10�6 for4+ year, p = 0.0413 for 5+ year).
In terms of examination between-age analysis within the samesites, at R, no statistically significant change on TAC was observedin any of the studied animals; whereas at M and H contaminatedsites, a TAC decreasing trend from 2+ to 5+ years old animals wasobserved. Moreover, the decreasing trend observed for 4+ yearand 5+ year old animal at M was significantly lower in comparisonto 2+ year and 3+ year old animals. The significance levels ascer-tained at M for 4+ years were p = 9.4 � 10�5 (versus 2+ year) andp = 2.87 � 10�6 (versus 3+ years), and for 5+ years the significancelevel were p = 1.82 � 10�5 (versus 2+ year) and p = 4.28 � 10�5
(versus 3+ year), respectively.
3.1.2. Effect on whole-body mitochondrial LPO (Fig. 2B)Between-site analysis revealed a significantly higher LPO in all
the classes of clams at M in comparison to R (2+year, p = 0.026;3+ year, p = 0.025; 4+ year, p = 0.004; 5+ year, p = 0.040); whereasat H, significantly higher LPO was perceptible only in 2+
(p = 0.030) and 3+ (p = 0.019) year old clams when compared to R(Fig. 2B). Upon the comparison of two contaminated sites, a signif-icantly lower LPO at H was also observed in 2+ (p = 0.049), 3+
(p = 0.032) and 4+ (p = 0.046) year old clams.Examining the LPO results between-age analysis, at R, no signif-
icant change was observed in any class of the studied clams. How-ever, at M and H sites, a trend of LPO decrease was perceptiblefrom 2+ to 5+ years old clams (except 3+ year at M). The decreaseobserved for 4+ year clam at M was significantly lower in compar-ison to 2+ (p = 0.050) and 3+ (p = 0.005) year old clams, whereas for5+ year clams it was significantly lower in comparison to 2+
(p = 0.011), 3+ (p = 0.013) and 4+ (p = 0.041) year old clams. At H,LPO response observed for 4+ and 5+ year old clams were similarto the responses observed at M for the corresponding classes ofclams. The significance levels ascertained for 4+ years at H werep = 0.040 (versus 2+ year) and p = 0.049 (versus 3+ years), and for5+ years the significance levels were p = 0.032 (versus 2+ year),p = 0042 (versus 3+ year) and p = 0.044 (versus 4+ year),respectively.
Pearson’s analysis within each age class revealed a positivecorrelation between TAC and peroxidative damage (r = 0.271,p = 0.008) in 5+ year old clams only (Fig. 3).
3.2. Organ-specific approach
3.2.1. Effect on organs mitochondrial TAC (Fig. 4A)Gill displayed a significantly (p = 4.47 � 10�8) lower TAC at H in
comparison to R (Fig. 4A). In addition, a significantly (p = 0.0435)lower TAC was also perceptible at H in comparison to M. In diges-tive glands, a significantly (p = 0.0153) higher TAC was observed atM in comparison to R; whereas, a significantly (p = 0.0213) lowerTAC was observed at H when the two contaminated sites werecompared.
3.2.2. Effect on organs mitochondrial LPO (Fig. 4B)In organs mitochondrial LPO analysis, only gill displayed a sig-
nificantly (p = 0.044) higher LPO at M in comparison to R.Analyzing the relationship between whole body (4+ year clam)
and organs mitochondrial responses, a significant positive correla-tion was observed only between whole body and gill for TAC(r = 0.879, p = 0.001). Inter-organ analysis revealed significant posi-tive correlations between gill and digestive gland for both LPO(r = 0.559, p = 0.013) and TAC (r = 0.928, p = 0.004). However, with-in-organ analysis between LPO and TAC exhibited no correlationeither for gill or for digestive gland.
4. Discussion
In order to achieve a better understanding of bivalves’ strategiesadopted to survive in the mercury-contaminated environment, thisstudy focuses on mitochondria, which is the main energy source

Fig. 3. Pearson’s correlation coefficients (r) between the mitochondrial lipid peroxidation (LPO) and total antioxidants in the whole body (5+ year old) of Scrobicularia plana(p < 0.05). Nonsignificant correlations are not presented.
Fig. 4. Values represent the means (±standard error) of mitochondrial totalantioxidants (A) and lipid peroxidation (B) measured in gill and digestive gland ofScrobicularia plana (4+ year old specimens) collected at reference (R), moderately(M) and highly (H) mercury contaminated sites in the Ria de Aveiro. Statisticallysignificant differences (p < 0.05) are marked by letters (between sites) are: a versusR, b versus M.
I. Ahmad et al. / Chemosphere 92 (2013) 1231–1237 1235
and the primary target of mercury in fish and mammals. In addi-tion, this paper links to the previous paper where S. plana collectedfrom the mercury contaminated areas reflected higher haemo-lymph mercury concentrations capable to induce immunosuppres-sion (Ahmad et al., 2011a). It was also observed that the adaptivecapacity to mercury pro-oxidant challenge, expressed as antioxi-dant induction and lesser vulnerability to enzyme inhibition, in-creased with clam’s age (Ahmad et al., 2011b). Age-relatedvariations on non-enzymatic antioxidant responses demonstratedthat the evolution of the adaptive skills of S. plana over time de-pends on the contamination extent (Ahmad et al., 2012a). Hence,under a moderate contamination habitat, the intervention of thedifferent antioxidants (measured in whole-body) took place
harmoniously, evidencing an adjustment capacity increasing withthe age. In opposition, under a higher contamination degree, S. pla-na failed to efficiently cope with mercury threat, showing an age-dependent deterioration of the non-enzymatic defence abilities.Moreover, age related variation in accumulation of total and organ-ic mercury was also observed. It was revealed that all age classesaccumulated total mercury according to the mercury gradient inthe environment, whereas for organic mercury only 5+ year oldanimal showed the accumulation according to the mercury gradi-ent in the environment. The specific analyses of gill and digestivegland revealed that both organs were able to mirror external levelsof exposure in the accumulation of total and organic mercury.However, only digestive gland seems more appropriate usage ableto identify the moderate level of contamination on the basis of to-tal mercury accumulation, whereas gill displayed higher accumu-lation of organic forms. Thus, using two approaches viz. inter-ageand organ-specific, the mitochondrial responses in the currentstudy have been related with previously published mercury bioac-cumulation data as reported by Ahmad et al. (2011b).
4.1. Inter-age approach
In the current study, mitochondrial antioxidants defence sys-tem seems to be dependent on mercury accumulation and age overtime, since clams showed a decreasing trend of TAC with increas-ing age statistically significant in 4+ and 5+ years at M; whereasat H, once the TAC was depleted, remained same until they havegrown 5+ years. At M, in the beginning (2+, 3+ year), clams wereable to cope with mercury accumulation, but with increasing age(4+, 5+ years) antioxidants breakdown was perceptible that maybe due to the mercury accumulation increase beyond its thresholdlimit accountable for antioxidants depletion over time as proposedby Black et al. (1996) for metal, regarding DNA damage versus re-pair requiring sufficient toxicant for its induction or activation.Interestingly, mercury accumulation pattern reflected that theaccumulation at M after a 3 years period was less than the accumu-lation at H after 2 years in case of total mercury (Ahmad et al.,2011b). However, this explanation supporting the TAC increasewas perceptible only in 5+ year old clams due to significant in-crease in total mercury accumulation (0.22 ± 0.03), since 4+ yearclass showed a further decrease (0.18 ± 0.03) in comparison to 2+
(0.13 ± 0.02) and 3+ year (0.2 ± 0.02) old animals. In addition,absurdities of TAC responses over time were also observed in

1236 I. Ahmad et al. / Chemosphere 92 (2013) 1231–1237
animals collected from R despite a smooth linearity of mercuryaccumulation; however, statistically insignificant. Variations inmetal accumulation and size-dependence in metal concentrationshave often been observed in invertebrates, particularly in bivalvesinhabiting contaminated sites, and the storage of metal into metalconcretions (i.e. metal rich granules, metallothioneins) has been ci-ted as a possible mechanism to explain these observations (Wal-lace et al., 2003). The current findings clarified the previousstudy where a contamination dependent antioxidants modulationalong the clam’s age could be difficult to summarize into a singleindex especially when the same organisms exhibit an increase ofcertain antioxidants concomitant to a decrease of others (Ahmadet al., 2012a,b). This difference in observations between mitochon-dria and post-mitochondrial supernatant (PMS) confirms that themitochondria in bivalves can also be the target organelle of mer-cury as it is the known primary target of mercury (48%) followedby nuclei (38%), cytosol (8%) and microsomes (7%) in mammals(Konigsberg et al., 2001).
Mitochondrial LPO was observed higher in all the studiedorganisms at M whereas, at H, it was perceptible only in 2+ and3+ year old animals. These findings confirm that like other metals(Stohs and Bagchi, 1995; Ercal et al., 2001; Wang et al., 2004; Mas-so et al., 2007), mercury also promotes the generation of ROS as re-flected in mitochondrial peroxidative damage in the current study.Some studies on mammals and on fish pinpoint the mitochondrionas the primary target for the effects of mercury, largely through theproduction of ROS and consequent damage to the mitochondrialmembrane (Pourahmad and O’Brien, 2000; Belyaeva et al., 2002,2004). In the current study, it is clearly evident that previously ob-served mercury influence is not only limited to fish and mammalsbut also includes bivalves in the mercury contaminated benthicenvironment. The age of clams strongly defined clams’ susceptibil-ity towards mercury toxicity and its detoxification overtime sincethe propensity for LPO increase relative to reference decreasedwith age. Moreover, it cannot be overlooked that the most resistant4+ and 5+ year old clams reflected an unaltered LPO relative to R,which was not evident using fraction other than mitochondriawhere peroxidation of membrane lipids displayed vulnerabilityof 5+ year old clams towards mercury accumulation (Ahmadet al., 2011b).
Concomitant to increased LPO, no change in mitochondrial TACat initial stage as well as increased use of TAC incapable to preventperoxidative damage is in agreement with Cossu et al. (2000), whostated that LPO increase cannot be predicted only on the basis ofantioxidant depletion. Furthermore, Ferreira-Cravo et al. (2007)demonstrated that the low antioxidant competence against HO�
radicals was not concomitant with high LPO values in estuarinespecies sampled from a contaminated site, suggesting that thehigher ROS levels could affect other macromolecules. Since theantioxidants including macro- and micro-molecules and enzymesrepresenting altogether the total antioxidant activity of the systemplay a central role in prevention of oxidative stress, it can be seenclearly in the current study that the TAC is being used to maintainthe redox status in the animal cells where its increased use to scav-enge ROS was perceptible with progressive decreases in LPO withage. Inter-age comparison also revealed that 2+ and 3+ years oldclams possess same level of TBARS; however, this level startsdecreasing from 4+ to 5+ years at M; whereas, at H, it became thesame as in animals collected from R. If the study is compared withthe previous studies on PMS fraction where per-oxidative damage(at H) was perceptible even at the induction of enzymes (catalase,ascorbate peroxidise, glutathione peroxidise and glutathione sulfo-transferase) (Ahmad et al., 2011b) and balances between enzy-matic and non-enzymatic antioxidants was attempted by theorganisms for their adaptation to environmental mercury, the cur-rent study strongly suggests that the damage prevention is brought
at the cost of increased TAC use and therefore, inclusion of mito-chondrial overall antioxidants as well can be recommended inthe assessment for future field investigations.
4.2. Organ-specific approach
In gill, mitochondrial TAC remained unaffected despite signifi-cant mercury accumulation indicating that there may be a bal-anced ROS production at M; however, LPO increase in animalscollected from the same site does not strengthen the previousstatement and pointed out the fact that there may be a possiblerole of mercury induced direct mitochondrial oxidative burst activ-ity contributing to overall ROS production leading up to peroxida-tive damage. Mitochondria’s role in ROS production and itscontribution in redox signaling from organelle to cytosol and nu-cleus is undisputable in mammals (Konigsberg et al., 2001). We re-ported earlier that mercury accumulation caused overall ROSproduction increase in non-phagocytes circulatory cells corrobo-rating the peroxidative damage in animals collected from the samesite (Ahmad et al., 2011a). At H, no LPO induction but significantlydecreased TAC clearly indicates its increased use due to highermercury accumulation, providing a better protection to exposedbivalves in comparison to those collected from R. With regards todigestive gland, a clear induction on TAC observed at M in the cur-rent study implies that additional synthesis of TAC in response tothe moderate ROS level production may account for preventingLPO increase. No change in TAC and corroborating LPO absenceat H indicate the effectiveness of the overall antioxidant mecha-nisms preventing peroxidative damage. Analyzing the relationshipbetween gill and digestive gland, mercury accumulation depen-dent organs’ resistance is perceptible in the current study. Clamscollected from M site reflected that digestive gland is more respon-sive in terms of TAC, whereas at H, gill is more responsive. How-ever, in terms of LPO, gill is more responsive since there is asignificant LPO increase instead of an incremental trend (digestivegland) and thus, both the organs should be considered for monitor-ing purpose in order to better assess the hazard of mercury expo-sure. Analyzing the significant positive correlation between wholebody and gill (TAC), it can be suggested that the assessment ofeither whole body or gill specific mitochondrial oxidative stress re-sponses is a valid approach for mercury impacted areas bio-moni-toring programme.
5. Conclusions
– Mitochondria proved to be a sensitive fraction for the effects ofmercury in S. plana inhabiting mercury contaminated area.
– S. plana age determined the clams’ vulnerability towards mer-cury induced mitochondrial peroxidative damage: younger ani-mals were more vulnerable than older.
– A decreasing trend of TAC with increasing age statistically sig-nificant in 4+ and 5+ years was also observed at M; whereas,at H, once the TAC was depleted, remained same until they havegrown 5+ years.
– The organ specificity was evident where a clear pattern ofhigher TAC was perceptible in digestive gland (at M) and lowerTAC in gill (at H); whereas, towards peroxidative damage, onlygill was susceptible.
– A positive correlation between whole animal and organ (gill)clearly revealed that the use of whole body and individual organ(gill) to be equally valid approach in addressing mercuryinduced mitochondrial oxidative stress responses.
– The results from the present study strongly suggests and rec-ommends the use of either S. plana whole body or gill organfor the bio-monitoring of mercury impacted areas.

I. Ahmad et al. / Chemosphere 92 (2013) 1231–1237 1237
Acknowledgements
Financial support provided by the Portuguese FCT (Foundationfor Science and Technology) to CESAM (Centre for Environmentaland Marine Studies) is gratefully acknowledged. The authors arethankful to J.P. Coelho, Department of Chemistry, University ofAveiro for his valuable help during field campaign.
References
Ahmad, I., Mohmood, I., Coelho, J.P., Pacheco, M., Santos, M.A., Duarte, A.C., Pereira,E., 2012a. Role of non-enzymatic antioxidants on the bivalves’ adaptation toenvironmental mercury: organ-specificities and age effect in Scrobicularia planainhabiting a contaminated lagoon. Environ. Pollut. 163, 218–225.
Ahmad, I., Coelho, J.P., Mohmood, I., Anjum, N.A., Pacheco, M., Santos, M.A., Duarte,A.C., Pereira, E., 2012b. Mercury contaminated systems under recovery canrepresent an increased risk to seafood human consumers – a paradox depictedin bivalves’ body burdens. Food Chem. 133, 665–670.
Ahmad, I., Coelho, J.P., Mohmood, I., Pacheco, M., Santos, M.A., Duarte, A.C., Pereira,E., 2011a. Immunosuppression in the infaunal bivalve Scrobicularia planaenvironmentally exposed to mercury and association with its accumulation.Chemosphere 82, 1541–1546.
Ahmad, I., Mohmood, I., Mieiro, C.L., Coelho, J.P., Pacheco, M., Santos, M.A., Duarte,A.C., Pereira, E., 2011b. Lipid peroxidation versus antioxidant modulation in thebivalve Scrobicularia plana in response to environmental mercury–organspecificities and age effect. Aquat. Toxicol. 103, 150–158.
Anjum, S., Rahman, S., Kaur, M., Ahmad, F., Rashid, H., Ansari, R.A., Raisuddin, S.,2011. Melatonin ameliorates bisphenol A-induced biochemical toxicity intesticular mitochondria of mouse. Food Chem. Toxicol. 49, 2849–2854.
Ansaldo, M., Luquet, C.M., Evelson, P.A., Polo, J.M., Llesuy, S., 2000. Antioxidantlevels from different Antarctic fish caught around South Georgia and Shag rocks.Polar Biol. 23, 160–165.
Belyaeva, E.A., Glazunov, V.V., Korotkov, S.M., 2002. Cyclosporin A-sensitive perme-ability transition pore is involved in Cd2+-induced dysfunction of isolated ratliver mitochondria: doubts no more. Arch. Biochem. Biophys. 405, 252–264.
Belyaeva, E.A., Glazunov, V.V., Korotkov, S.M., 2004. Cd2+-promoted mitochondrialpermeability transition: a comparison with other heavy metals. Acta Biochim.Pol. 51, 545–551.
Black, M.C., Ferrell, J.R., Horning, R.C., Martin, L.K., 1996. DNA strand breakage infreshwater mussels (Anodonta grandis) exposed to lead in the laboratory andfield. Environ. Toxicol. Chem. 15, 802–808.
Buchner, T., Abele-Oeschger, D., Theede, H., 1996. Aspects of antioxidant status inthe polychaete Arenicola marina: tissue and subcellular distribution, andreaction to environ- mental hydrogen peroxide and elevated temperatures.Mar. Ecol. Prog. Ser. 143, 141–150.
Buege, J.A., Aust, S.D., 1978. Microsomal lipid peroxidation. Methods Enzymol. 52,302–310.
Cambier, S., Benard, G., Mesmer-Dudons, N., Gonzalez, P., Rossignol, R., Brethes, D.,Bourdineaud, J.-P., 2009. At environmental doses, dietary methylmercuryinhibits mitochondrial energy metabolism in skeletal muscles of the zebrafish (Danio rerio). Int. J. Biochem. Cell Biol. 4, 791–799.
Coelho, J.P., Rosa, M., Pereira, M.E., Duarte, A.C., Pardal, M.A., 2006. Pattern andannual rates of Scrobicularia plana mercury bioaccumulation in a humaninduced mercury gradient (Ria de Aveiro, Portugal). Estuar. Coast. Shelf Sci. 69,629–635.
Connell, D.W., Miller, G.J., 1984. Chemistry and Ecotoxicology of Pollution. JohnWiley & Sons, New York.
Cossu, C., Doyotte, A., Babut, M., Exinger, A., Vasseur, P., 2000. Antioxidantbiomarkers in freshwater bivalves, Unio tumidus, in response to differentcontamination profiles of aquatic sediments. Ecotoxicol. Environ. Saf. 45, 106–121.
Duarte, A., Rodrigues, S., Pato, P., Coelho, C., Pereira, M.E., 2007. A review on studiesof mercury contamination in the coastal lagoon ‘Ria de Aveiro’ Portugal. LaHouille Blanche 4, 35–39.
Ercal, N., Gurer-Orhan, H., Aykin-Burns, N., 2001. Toxic metals and oxidative stress.Part I: Mechanisms involved in metal-induced oxidative damage. Curr. Top.Med. Chem. 1, 529–539.
Ferreira-Cravo, M., Piedras, F.R., Moraes, T.B., Ferreira, J.L.R., Freitas, D., Machado, M.,Geracitano, L., Monserrat, J.M., 2007. Antioxidant responses and reactive oxygenspecies generation in different body regions of the estuarine polychaetaLaeonereis acuta (Nereididae). Chemosphere 66, 1367–1374.
Giari, L., Simoni, E., Manera, M., Dezfuli, B.S., 2008. Histo-cytological responses ofDicentrarchus labrax (L.) following mercury exposure. Ecotoxicol. Environ. Saf.70, 400–410.
Gornall, A.G., Bardawill, C.J., David, M.M., 1949. Determination of serum proteins bymeans of the Biuret reaction. J. Biol. Chem. 177, 751–766.
Guilherme, S., Válega, M., Pereira, M.E., Santos, M.A., Pacheco, M., 2008. Antioxidantand biotransformation responses in Liza aurata under environmental mercuryexposure - relationship with mercury accumulation and implications for publichealth. Mar Pollut. Bull. 56, 845–859.
Jang, Y.Y., Song, J.H., Shin, Y.K., Han, E.S., Lee, C.S., 2000. Protective effect of boldineon oxidative mitochondrial damage in streptozotocin-induced diabetic rats.Pharmacol. Res. 42, 361–371.
Konigsberg, M., López-Díazguerrero, N.E., Bucio, L., Gutiérrez-Ruiz, M.C., 2001.Uncoupling effect of mercuric chloride on mitochondria isolated from anhepatic cell line. J. Appl. Toxicol. 21, 323–329.
Kristal, B.S., Jackson, C.T., Chung, H.Y., Matsuda, M., Nguyen, H.D., Yu, B.P., 1997.Defects at center p underlie diabetes-associated mitochondrial dysfunction.Free Rad. Biol. Med. 22, 823–833.
Lemaire, P., Viarengo, A., Canesi, L., Livingstone, D.R., 1993. Pro-oxidant andantioxidant processes in gas gland and other tissues of cod (Gadus morhua). J.Comp. Physiol. 163B, 477–486.
Masso, E.L., Corredor, L., Antonio, M.T., 2007. Oxidative damage in liver after peri-natal intoxication with lead and/or cadmium. J. Trace Elem. Med. Biol. 21, 210–216.
Pereira, M.E., Lillebø, A.I., Pato, P., Válega, M., Coelho, J.P., Lopes, C., Rodrigues, S.,Cachada, A., Otero, M., Pardal, M.A., Duarte, A.C., 2009. Mercury pollution in Riade Aveiro (Portugal): a review of the system assessment. Environ. Monit. Assess.155, 39–49.
Pourahmad, J., O’Brien, P.J., 2000. A comparison of hepatocyte cytotoxicmechanisms for Cu2+ and Cd2+. Toxicology 143, 263–273.
Ribeiro, C.A., Oliveira, F.E., Turcatt, N.M., Cardoso, R.J., Carvalho, C.S., 1996. Lethaleffects of inorganic mercury on cells and tissues of Trichomycterus brasiliensis(Pisces: Siluroidei). Ecotoxicol. Environ. Saf. 33, 160–166.
Rigobello, M.P., Barzon, E., Marin, O., Bindoli, A., 1995. Effect of polycation peptideson mitochondrial permeability transition. Biochem. Biophys. Res. Commun.217, 144–149.
Sayeed, I., Parvez, S., Winkler-Stuck, K., Seitz, G., Trieu, I., Wallesch, C.W., Schonfeld,P., Siemen, D., 2006. Patch clamp reveals powerful blockade of themitochondrial permeability transition pore by the D2-receptor agonistpramipexole. FASEB J. 20, 556–558.
Stohs, S.J., Bagchi, D., 1995. Oxidative mechanisms in the toxicity of metal ions. FreeRadic. Biol. Med. 18, 321–336.
Tabassum, H., Parvez, S., Pasha, S.T., Banerjee, B.D., Raisuddin, S., 2010. Protectiveeffect of lipoic acid against methotrexate-induced oxidative stress in livermitochondria. Food. Chem. Toxicol. 48, 973–979.
Verdelhos, T., Neto, J.M., Marques, J.C., Pardal, M.A., 2005. The effect ofeutrophication abatement on the bivalve Scrobicularia plana. Estuar. Coast.Shelf Sci. 63, 261–268.
Wallace, W.G., Lee, B.-G., Luoma, S.N., 2003. Subcellular compartmentalization of Cdand Zn in two bivalves. I. Significance of metal-sensitive fractions (MSF) andbiologically detoxified metal (BDM). Mar. Ecol. Prog. Ser. 249, 183–197.
Wang, Y., Fang, J., Leonard, S.S., Rao, K.M.K., 2004. Cadmium inhibits the electrontransfer chain and induces reactive oxygen species. Free Radic. Biol. Med. 36,1434–1443.
Wootton, E.C., Pipe, R.K., 2003. Structural and functional characterisation of theblood cells of the bivalve mollusc, Scrobicularia plana. Fish Shellfish Immunol.15, 249–262.
Zielinski, S., Portner, H.O., 2000. Oxidative stress and antioxidative defence incephalopods: a function of metabolic rate or age? Comp. Biochem. Physiol.125B, 147–160.