mechanisms of dormancy, preharvest sprouting tolerance...
TRANSCRIPT

Mechanisms of dormancy, preharvest sprouting tolerance and how they are influenced by the environment
during grain filling and maturation in wheat (Triticum aestivum L.)
Thomas Benjamin Biddulph
This thesis is presented for the degree
of
Doctor of Philosophy
The University of Western Australia
School of Plant Biology
Faculty of Natural and Agricultural Science
2006



i
Summary Wheat is the main crop in Australia and there are stringent quality requirements. Preharvest
sprouting induced by rainfall between maturity and harvest lowers grain quality from
premium to feed grades and reduces yield. Wheat production has expanded into the
southern Western Australian region where preharvest sprouting occurs in ~1 in 4 seasons
and development of more preharvest sprouting tolerant genotypes is required. The main
mechanism for improving preharvest sprouting tolerance is grain dormancy. There is
genetic variation for dormancy based in the embryo and seed coat but dormancy is complex
and is influenced by environmental conditions during grain filling and maturation.
Screening and selecting for preharvest sprouting tolerance is problematic and the level of
tolerance needed for regions which differ in the level of dormancy they impose, requires
clarification. The research presented here aims to answer the underlying question for
breeders of how much dormancy is required for preharvest sprouting tolerance in
contrasting target environments of the central and coastal wheat belt regions of Western
Australia.
In the central and coastal wheat belt regions, field trials with modified environments were
used to determine the environmental influence on dormancy. Water supply (without
directly wetting the grain) and air temperature were modified during grain development in a
range of genotypes with different mechanisms of dormancy to determine the influence of
environment on dormancy. The grain produced was used to study the control of dormancy
during imbibition in the laboratory. Commercial and advanced breeding genotypes with a
range of preharvest sprouting tolerance were also used to quantify the level of dormancy
required to give adequate protection from preharvest rainfall at the contrasting sites.
Laboratory studies found the embryo component of dormancy was due, in part, to the
ability of intact dormant grains to maintain a higher embryo abscisic acid (ABA)
concentration during imbibition. The elevation of ABA concentration does not explain the
full embryo component and sensitivity to ABA still appears to be required for the dormant
phenotype. The seed coat component had an additive effect on the embryo component.

ii
In field experiments, modified environmental conditions during grain filling and maturation
influenced the level of dormancy. During grain filling consistently high temperatures and
moisture stress induced a dormant phenotype in a genotype, which is typically non-
dormant. Stressful environmental conditions during grain filling, which induced the most
dormancy, also induced the most sensitivity to applied ABA. During grain filling (15-30
dpa) stressful environments increased dormancy at maturity. In contrast to the grain filling
period, stressful environments during grain maturation decreased dormancy. High
temperatures shocks (> 12 days of maximum > 30°C) at 30-50 dpa, but not earlier, reduced
the level of dormancy in all genotypes. Excess water supply from rainfall or irrigation
(without wetting the heads) also reduced dormancy in all genotypes. Cleaving black point
and fungal infection of the grain was also observed under these conditions. It is proposed
that during grain maturation certain environments can reduce dormancy in all genotypes by
reducing the additive effect of the general seed coat effect.
The level of dormancy from the embryo component alone, in current advanced breeding
genotypes, was effective in the field and should consistently prevent preharvest sprouting
when introgressed into commercial genotypes. Genotypes with embryo dormancy were
consistently the most preharvest sprouting tolerant, even though this dormancy was
influenced by the environmental conditions in the different seasons. Pyramiding the embryo
component with the specific seed coat component and/or awnless head trait removed some
of the environmental variation in preharvest sprouting tolerance, but this was generally
considered excessive to the environmental requirements.
The methods developed here, of field imposed stresses may provide a valuable tool to
further understand the influence of environment on the regulation of dormancy, as different
phenotypes can be made with the same genotype. Moisture stress, sudden changes in water
supply or high temperatures during the late dough stages influenced dormancy phenotype
and should be considered and avoided if possible when selecting locations and running
trials for screening for genetic differences in preharvest sprouting tolerance. In the Western
Australian context, the embryo component of dormancy appeared to be sufficient and
should be adopted as the most important trait for breeding for preharvest sprouting
tolerance.

iii
Statement of original contribution
The research presented in this thesis is an original contribution to the field of cereal
dormancy and preharvest sprouting in wheat. The hypotheses and experiments presented
and discussed in this thesis are my own original ideas and writing.
Other people that made important contributions to this research are acknowledged in
Chapters 3-5.
• Julie Plummer, Tim Setter and Daryl Mares who were the supervisors of this
research project, guiding me through the process of forming hypotheses, designing
experiments and writing up material for submission.
• Andrew Poole and Frank Gubler provided technical support with the GC-MS
analysis and assistance with the ABA study.
• Peter Clarke and Katia Stefanova provided statistical support with trial design and
statistics.
The thesis has been completed during the course of enrolment in a PhD degree at the
University of Western Australia, and has not been used previously for a degree or diploma
at any other institution.
Thomas Benjamin Biddulph
July, 2007.

iv
Structure of thesis
This thesis includes a General Introduction, Literature Review, three Research Chapters presented
as papers which areaccepted, under review or in the process of submission and a General
Discussion.
Chapter 1 is a general introduction which outlines the rational for the project and the reasoning
behind the aims of the thesis.
Chapter 2 is a critique of the relevant literature, focusing on the current understanding of the
physiology of dormancy and how this mechanism, and hence preharvest sprouting tolerance, is
influenced by the environment during grain filling and maturation.
Chapter 3 is the first research chapter. It examines the ABA content of field grown, intact grains
during imbibition of dormant and non-dormant genotypes in order to better understand the
mechanism known as �embryo sensitivity.� This chapter is currently in preparation for submission
to Australian Journal of Agricultural Research.
Chapter 4 is the second research chapter, which includes three years of field trials at two sites
looking at quantifying the effect of temperature, water supply and their interaction on the embryo
sensitivity mechanism of dormancy. The particular stage during grain filling when temperature was
critical was determined. This chapter discusses the possible interaction of temperature and maturity
length and the implications for selecting genotypes differing in maturity for preharvest sprouting
tolerance based on dormancy. This chapter has been accepted and is in press for publication with
Field Crops Research.
Chapter 5 is the final research chapter and covers three years of field trials at two sites exploring
the relationship between dormancy and grain quality after natural weathering associated with water
supply during grain filling. It determines the level and mechanisms of dormancy required to
consistently give preharvest sprouting tolerance for environments which differ in their sprouting
risk. The implications in setting breeding objectives for different environments and preharvest
sprouting tolerance are discussed. This chapter has also been accepted by Field Crops Research.
Chapter 6 is the final chapter and contains the General Discussion. This chapter discusses the
interaction of results across chapters and their implications for hypotheses which should be
examined in future work on the regulation of dormancy by environment. Also discussed are the
implications for breeding for preharvest sprouting tolerance for environments which differ in their
susceptibility to preharvest sprouting.

v
Peer-reviewed publications arising from this thesis
1. Biddulph TB, Mares DJ, Gubler F, Poole AT, Plummer JA and Setter TL (2007)
Abscisic acid concentration of wheat (Triticum aestivum L.) embryos in
relation to expression of grain dormancy. (In preparation). (Chapter 3)
2. Biddulph TB, Plummer JA, Setter TL and Mares DJ (2007) Influence of high
temperature and terminal moisture stress on dormancy in wheat (Triticum
aestivum L.). Field Crops Research Doi:j.fcr.2007.05.005. (Chapter 4)
3. Biddulph TB, Mares DJ, Plummer JA and Setter TL (2007) Seasonal environmental
conditions influence dormancy and subsequent preharvest sprouting
tolerance in wheat (Triticum aestivum L.) in the field . Field Crops Research
Re-submitted (Chapter 5)
4. Biddulph TB, Mares DJ, Plummer JA and Setter TL (2005) Drought and high
temperature increases preharvest sprouting tolerance in a genotype without
grain dormancy. Euphytica 143, 277-283. Appendix A
Primary authored papers or abstracts presented or published in
unrefereed conference proceedings
1. Biddulph TB, (2004) Preharvest sprouting tolerance of wheat in Western
Australia. March 2004, Agribusiness Regional Crop Updates,
Ravensthorpe and Jerramungup, WA.
2. Biddulph TB, Mares, DJ, Setter, TL, and Plummer, JA. (2006) Environment is it
as important as variety in preharvest sprouting tolerance? Agribusiness
Crop Updates, March 2006, Burswood, Perth, WA.
3. Biddulph TB, Mares, DJ, Setter, TL, and Plummer, JA. (2006) Environment is it
as important as variety in preharvest sprouting tolerance? Agribusiness
Regional Crop Updates, March 2006, Esperance, WA.
4. Biddulph TB, Mares, DJ, Setter, TL, and Plummer, JA. (2006) Environment is it
as important as variety in preharvest sprouting tolerance? Fitzgerald
Biosphere Group Annual Trials meeting, March 2006, Jerramungup,
WA. Appendix B

vi
5. Biddulph TB, Plummer, JA, Mares, DJ and. Setter, TL. (2004) Drought and high
temperature increase preharvest sprouting tolerance in wheat without grain
dormancy. Combio, September 2004, Burswood, Perth, WA.
6. Biddulph TB, Plummer, JA, Mares, DJ and. Setter, TL. (2005) Preharvest sprouting
tolerance in the field. Agribusiness Crop Updates, February
2005, Burswood, Perth, WA. Appendix C
Extension presentations to interested grower groups and
researchers
1. August 2003, Sprouting in wheat. RAIN, AGM, Red Room, Ravensthorpe, WA.
2. February 2004, Preharvest sprouting tolerance of wheat in Western Australia; the
influence of environment and variety. Annual Plant Biology Postgraduate Retreat,
Rottnest, WA.
3. May 2004, Preharvest sprouting tolerance of wheat in Western Australia; the
influence of environment and variety. Plant Biology, University of Western
Australia Seminar Series, Perth, WA.
4. July 2004, Preharvest sprouting tolerance of wheat in Western Australia; the
influence of environment and variety. Crop Improvement, Department of Agriculture
Western Australia, Perth, WA.
5. October 2004, Preharvest sprouting tolerance of wheat in Western Australia.
Esperance Agricultural Centre Seminar Series, Esperance, WA.
6. October 2004, Preharvest sprouting tolerance of wheat in Western Australia. RAIN,
AGM, Red Room, Ravensthorpe, WA.
7. November 2004, Preharvest sprouting tolerance of wheat in Western Australia.
Chinese visitors delegation to WA, Department of Agriculture Western Australia,
Perth, WA.
8. February 2005, Preharvest sprouting tolerance of wheat in Western Australia; the
influence of environment and variety. Annual Plant Biology Postgraduate Retreat,
Rottnest, WA.
9. March 2005, Preharvest sprouting tolerance of wheat in Western Australia; the
influence of environment and variety. CSIRO, Seminar Series, Plant Industry, Black
Mountain, ACT.

vii
10. May 2005, Preharvest sprouting tolerance of wheat in Western Australia; the
influence of environment and variety. Annual Quality Defect Elimination Meeting
WAITE, University of Adelaide, Urbrrae, SA.
11. December 2005, Esperance Agricultural Centre Seminar Series, Esperance, WA.
12. February 2006, Abscisic acid in wheat embryos during expression of grain
dormancy. Annual Plant Biology Postgraduate Retreat, Rottnest, WA.
13. July 2006, Preharvest sprouting, dormancy, environment and falling number. Crop
Improvement, Department of Agriculture and Food Western Australia, Perth, WA.
14. August 2006, What happens to dormancy when you play with environment? Annual
Quality Defect Elimination Meeting WAITE, University of Adelaide, Urbrrae, SA.
15. August 2006, What happens to dormancy and preharvest sprouting tolerance when
you play with environment? Australian Grain Technology, Annual Breeders
Meeting, Horsham, Vic.
16. September 2006, Influence of temperature and terminal moisture stress on dormancy
and subsequent preharvest sprouting tolerance of wheat. Plant Biology Seminar
Series, University of Western Australia, Perth, WA.

viii
Definition of terms Aleurone-the outermost layer of cells of the endosperm responsible for production of
enzymes for reserve mobilisation at germination.
Black point- black point is a physiological or pathogenic defect in grain which
predominantly results in a dark discoloration of the embryo end of the grain. The exact
cause, whether physiological or pathnogenic has yet to be determined. High levels of black
point (>5 % of grains) results in downgrading of grain at receival (Australian Wheat Board
2003-2004).
Cleaving- the splitting of the seed coat during grain maturation usually at the beard end of
the dorsal side of the grain. Associated with a temperature shock early during grain filling
followed by optimum grain filling and maturation conditions which results in plump
overfilled grains and cleaving / splitting of the seed coat during dry down of the grain.
Dormancy- the state in which a grain will not germinate in a specific time period under
conditions which normally favour germination. In wheat a dormant grain is defined as one
that does not germinate in 7 days in the dark with adequate water and oxygen at 20°C.
Embryo- the rudimentarily plant in the grain composed of the primordial root and shoot
with the attached scutellum.
Embryo-half grain- the half of the grain containing the embryo after a transverse cut
across the grain.
Embryo ABA elevation- the component of dormancy associated with elevation of the
endogenous embryo free ABA concentration during imbibition, associated with no
germination in dormant grain.
Embryo sensitivity (E)- the component of dormancy associated with inhibition of
germination of isolated embryos or embryo-half grains by exogenous ABA. Genotypes
with out this dormancy are designated (e).

ix
Endosperm- the nutritional tissue which nourishes the embryo. It contains storage
reserves, mainly starch, which are absorbed after germination to fuel the growth of the
seedling.
Falling number- an estimate of α-amylase activity and hence the bread making ability of
the dough. It is the time taken, in seconds (s), for a weighted stirrer to �fall� a set distance
through a heated paste of flour and water. Falling number ranges from a minimum of 62 s
for flour with a high alpha-amylase activity to >500 s in sound grain.
Field environment- the soil, environment and agronomic practices typical of the regions in
which wheat is commercially produced.
Fungal staining / infection- grains which are infected with fungi and the fruiting bodies
are clearly visible with the naked eye. Dark discolouration of the beard end of the grain is
usually obsereved, predominantly with Alternaria sp. but can also include Ulocladium and
Stemphylium sp.
Germination- protrusion of the radicle through the pericarp, which is visible with the
naked eye.
Germination Index- a measure of dormancy for grains (GIseed) and embryo half-grains
(GIembryo). It is calculated using Eqn 1.
(7 x n1 + 6 x n2 + 5 x n3 + 4 x n4 + 3 x n5 + 2 x n6 + 1 x n7) / (total days of test x total grains) Eqn (1)
where n1, n2, � n7 are the number of grains or embryos that germinated on the first, second
and subsequent days until the seventh day, respectively. Seed viability in grain that fails to
germinate in 7 d is assessed by incubating the grain at 4°C for a further 3 d then a further 3
d at 20°C. The maximum GI representing non-dormant grains is 1.0, and the minimum
representing dormant grain is 0.0

x
Grain filling- the period of time from anthesis to maximum fresh weight of the grain,
typically spanning from anthesis until 30 dpa.
Grain maturation- the later stages of grain maturation after maximum fresh weight has
been obtained where the grain is drying down and losing water before maturity, typically
30-60 dpa.
Grain moisture- the percentage of the grain fresh weight taken up by water expressed on a
dry weight basis after oven drying for 24hrs at 70°C. eg fresh weight of 110g, dry weight of
100g after oven drying equals a grain moisture content of 10% on a dry weight basis.
Grain weight- the dry weight of the grain after oven drying for 24 hrs at 70°C.
Harvest-ripeness- the stage of maturity when the plant first reaches 12 % grain moisture
content. Corresponds approximately to the stage when a commercial grower in Australia
would asses the crop as machine harvestable, corresponds to Zadoks Z92 (Zadoks et al.
1974).
Intact grain- grain harvested in a manner to preserve the seed coat components of
dormancy. Typically gently hand threshed and cleaned.
Scutellum- the shield like structure, which represents the monocotyledon and functions as
an absorptive organ for the embryo from the endosperm between which it is sandwiched.
Seed coat effect, general (s)- grain with the seed coat broken (like embryo-half grains)
germinates more rapidly than intact grains. This occurs in all genotypes regardless of
dormancy, and hence has been termed a general seed coat effect.
Seed coat effect, specific (S)- inheritance studies (Mares 1998) have shown that there is a
specific effect that can be attributed to the seed coat in dormant but not non-dormant
genotypes that is greater than the general effect that enhances dormancy. The specific seed
coat has an additive effect on the embryo component, but does not appear to provide
substantial tolerance by itself and is thought to be epistatic.

xi
Sensitivity to ABA- the component of dormancy not fully described by elevation of
embryo ABA concentration. The results presented in Section 3.4 and 4.4 indicate that
sensitivity to ABA (for want of a better word) must be involved, as well, as high embryo
ABA concentrations during imbibition to result in dormancy.
Sprouting- preharvest sprouting.
Terminal drought/ moisture stress- a severe moisture stress at the end of the growing
season which is typical of a Mediterranean like climate and responsible for termination of
grain filling, often forcing a shortened maturity compared to optimum conditions.
Maturity- the stage during grain maturation when the plant first reaches ~20 % grain
moisture content on a dry weight basis. Typically assessed as when all green colour has
been lost from the stem and a thumb nail indent is held in representative grains from central
florets. Corresponds to Zadoks Z91.
Plant ABA concentration- general ABA concentration of the plant as a response to the
maternal environment, assumed to increase above the concentration under optimum
conditions when the plant is subject to temperature and moisture stress.
Preharvest sprouting- is the germination of grain in the ear following rainfall between
maturity and harvest.
Preharvest sprouting tolerance- the ability of a genotype to maintain a Hagberg falling
number greater than 300 seconds after being subjected to substantial rainfall typical of the
target environment.
Weather affect grains- the combined count of fungal stained and black point affected
grains. In terms of receival standards the maximum level is 5 %, or 20 grains out of 500 in
premium wheat grades of wheat delivered to the Australian Wheat Board (2003-2004).

xii
Wheat belt- regions in Western Australia or Australia where commercial cereal crops,
predominately wheat, are grown.
Acronyms and abbreviations
ABA Abscisic acid
E embryo dormancy
e no embryo dormancy
d days
dpa days post anthesis
DW dry weight
FN falling number
FW fresh weight
GA Giberellic acid
GC-MS-SIM Gas Chromatography coupled with a Mass Selective Detector for Selected
Ion Monitoring
GI Germination Index
GIseed Germination index of intact seeds
GIembryo Germination index of embryo-half grains
S specific seed coat factor
s no specific seed coat fator, just general seed coat factor
t ton
QTL Quantitative Trait Loci

xiii
List of Figures
Chapter 2 Fig. 1. The range in visible sprouting of a sample of grain with a falling number of 62 s. 5 Fig. 2. Relationship between falling number and the percentage visually
sprouted grain in eight different commercial wheat genotypes exposed
to preharvest sprouting. 6
Fig. 3. Generalised time course of change in abscisic acid concentration
in embryos of wheat grain during grain filling and grain
maturation under optimum environments. 14
Fig. 4. Model of the regulation of ABA metabolism in embryos of
imbibed grains from controlled environments following dormancy
release. 17
Fig. 5. The relationship between falling number after rainfall simulation
15 days post maturity and the total rainfall in the 20 days prior to
harvest. 22
Chapter 3 Fig. 1. At Katanning in 2003/04 (a) germination index at harvest ripeness of
grain in water (solid bars), embryo half grain in water (shaded bars)
and embryo half grain with 20 µM ABA (open bars). 40
Fig. 2. At Esperance in 2004/05 (a) germination index at maturity of grain in
water (solid bars), embryo half grain in water (shaded bars) and embryo
half grain with 20µM ABA (open bars). 41

xiv
Fig. 3. At Esperance in 2004/05 (a) embryo ABA concentration (ng g-1 FW)
and (b) germination index of irrigated DM 2001, DH 22 and
Cunderdin grain. 44
Fig. 4. At Esperance in 2004/05, embryo ABA concentration
(ng g-1 FW, solid lines) in DH 22 (a) and Cunderdin (b) grain. 45
Chapter 4
Fig. 1. A typical daily pattern of canopy air temperature (circles), relative
humidity (RH) (squares) and vapour pressure deficit (VPD)
(triangles) with the low (open symbols) and high (solid symbols)
temperature treatments. 58
Fig. 2. Average increase in canopy vapour pressure deficit (VPD; kPa)
during grain filling prior to maturity at Katanning in
2003/04 (open circles) and at Esperance in 2005/06 (closed squares). 60
Fig. 3. Grain dry weight (solid lines, solid symbols) and moisture content
(dashed lines, open symbols) during grain filling in Cunderdin (es)
and DM 2001(ES) for plants grown at Katanning in 2003/04 and at
Esperance in 2004/05. 61
Fig. 4. Germination index of developing whole grain and embryo half
grains in water (solid lines, solid symbols) and 20µM ABA (dashed
lines, open symbols) of Cunderdin (es) and DM 2001 (ES) at Katanning
in 2003/04. 63
Fig. 5. Germination index of grain from plants grown at Katanning in 2003/04. 64
Fig. 6. Germination index of grain from plants grown at Esperance in 2004/05. 64

xv
Fig. 7. Germination index of whole grain at maturity against the number of
days during the 30-50 day post anthesis period when the daily
maximum canopy temperature was greater than 30°C. 66
Chapter 5 Fig. 1. Germination index at maturity (a, c) and falling number (b, d) of irrigated
(a, b) or moisture stressed (c, d) plots at maturity (Harvest 1, solid bars,)
after 50 mm rain (Harvest 2, shaded bars) and 70 mm of rain
(Harvest 3, open bars) at Esperance in 2005/06. 84
Fig. 2. BLUPS for falling number at successive harvests at maturity,
Harvest 1 (solid bars, Harvest 2 (shaded bars) and Harvest 3,
(open bars) in Esperance (a) 2003/04, (b) 2004/05, (c) 2005/06,
or Katanning (d) 2003/04 and (e) 2005/06. 86
Fig. 3. Biplots for germination index at maturity (a) and falling number at
maturity (b) and the final harvest (c) for the set of 6 common genotypes. 87
Fig. 4. Relationships between germination index and falling number after
50 mm rainfall in Esperance 2005/06 (r = -0.68) for 26 Western
Australian breeding genotypes, 35 commercial genotypes common
to Western Australia and 10 sprouting tolerant check genotypes. 88
Fig. 5. Falling number (a), weather affected (black point + fungal stained) (b),
black point (c), fungal stained (d), and field mould (e), ratings according to
Australian Wheat Board 2003/04 delivery specifications. 89
Chapter 6 Fig. 1. Conceptual model of the control of dormancy in wheat by
environmental conditions during grain filling (a) influencing
dormancy at maturity (b) through ABA elevation,
sensitivity or seed coat integrity. 103

xvi
List of Tables
Chapter 4 Table 1. Details of pedigree and dormancy mechanisms of the different
genotypes used. 57
Table 2. Average daily maximum canopy air temperature (°C) at 10 day
intervals after anthesis at Katanning (Kt) and Esperance (Esp)
with the heat shock (HS). 59
Table 3. Germination index at maturity of whole grains (GIseed)
in water for genotypes: DM 2001, DH 22, DH 56 DH 45 and Cunderdin
grown under natural rainfed conditions and imposed treatments in
Katanning (Kt) 2003/04, Esperance (Esp) 2004/05 and 2005/06. 62
Table 4. Germination index at maturity of whole grains (GIseed) in water for
genotypes: DM 2001, DH 22, DH 56 DH 45 and Cunderdin with imposed
treatments in Katanning (Kt) 2003, Esperance (Esp) 2004 and 2005. 62
Table 5. Analysis of Variance for GIseed experiments under ambient conditions
at Katanning 2003, Katanning 2005, Esperance 2003, Esperance 2004
and Esperance 2005. 65
Table 6. Analysis of Variance for GIseed within each the water stressed and
temperature manipulated experiments at Katanning 2003,
Esperance 2004 and Esperance 2005. 65
Chapter 5 Table 1. Details of pedigree and dormancy mechanisms, embryo component
(present E, absent, e) and seed coat component (present, S, absent, s)
and unknown (?) of the different genotypes used in this study. 76
Table 2. Sowing dates and the range in days from sowing to anthesis or
maturity, and sowing to anthesis rainfall at Esperance and Katanning in
2003/04, 2004/05 and 2005/06. 79

xvii
Table 3. Cumulative rainfall after harvest dates at maturity (H 1) and after
significant rainfall events ~one month after maturity (H 2) and
~ two months after maturity (H 3) at Esperance and Katanning in
2003/04, 2004/05 and 2005/06. 79
Table 4. Analysis of variance tables for Bi plots of germination index at
maturity and falling number at maturity (H1) and the final harvest
(H3) for the set of 6 common genotypes. 82
Table 5. Analysis of variance table for germination index at maturity and
falling number with three harvests, with the water stress treatments
from Esperance 2005. 83
Chapter 6 Table 1. Effect of different mechanisms of embryo and seed coat components
of dormancy which influences the germination index and dormancy
phenotype at maturity under varying environmental conditions. 102

xviii
Table of Contents Summary i
Structure of thesis iv
Publications arising from this thesis v
Definition of terms viii
Acronyms and abbreviations xii
List of Figures xiii
List of Tables xvi
1 General Introduction 1
2 Literature Review: Grain dormancy, environment and the
expression of preharvest sprouting tolerance in wheat
(Triticum aestivum L.) 4
3 Embryo abscisic acid concentration during imbibition of intact
grain is associated with grain dormancy in field grown wheat
(Triticum aestivum L.) 29
4 Influence of high temperature and terminal moisture stress on dormancy
in wheat (Triticum aestivum L.) 54
5 Seasonal conditions influence dormancy and preharvest sprouting tolerance
in wheat (Triticum aestivum L.) 70
6 General Discussion 98
References Cited 110
Acknowledgements 122
Appendix A 125
Appendix B 132
Appendix C 133
Appendix D 135

1
1 Chapter 1 General Introduction Preharvest sprouting has become an increasing risk in Western Australia in particular and
Australia since the early 1980�s possibly due to a combination of factors; these include
(i) Cultivars with a lower level of preharvest sprouting tolerance.
(ii) The trend towards shorter season genotypes, which leads to crops maturing at a
time more prone to preharvest sprouting.
(iii) The adoption of minimal till and increased size of farm machinery, which
results in greater synchronisation of sowing so all the crop may be at the same
stage of development when preharvest sprouting may occur.
(iv) The wider recognition that yield losses result from delayed sowing, which
results in earlier sowing and increasing susceptibility of the crop.
(v) Widespread adoption of legume rotations, the return of canola and the
availability of better agronomic packages for grass weed control have all
allowed the expansion of wheat production into the more sprouting prone
coastal regions previously dominated by barley, and.
(vi) The increased reliance on cropping caused by declining terms of trade for wool
production, has also driven an increase in the area sown to wheat, particularly in
coastal and higher rainfall agricultural areas.
As a consequence in preharvest sprouting prone regions a larger proportion of cropping
program is now wheat and there is a greater risk of sprouting occurring over the whole of
an individual growers cropping program and the regions crop.
Management practices previously highlighted by Bolland (1984) involve harvesting the
crop quicker to minimise the risk of quality losses associated with rainfall, and these are
now widely practiced by farmers. These include the use of larger more efficient harvesting
machinery and harvesting grain at higher moisture contents, coupled with grain drying or
aerated silos to reduce the moisture to export receival standards (12.5 %; Metz and
Newman 2007). Growers have also been successful in lobbying grain handlers to install
aeration systems to allow delivery of high moisture grain (up to 13.5 %). These strategies
have enabled the growers to complete harvest quicker, reducing some of the risk of
preharvest sprouting by reducing the time the crop is in the field after maturity.

2
Development of more preharvest sprouting tolerant genotypes is another way to reduce the
risk of the crop from preharvest sprouting and wheat breeders are attempting to introgress
dormancy as a long-term strategy. Currently however, there are no high yielding, locally
adapted commercial wheat genotypes with preharvest sprouting tolerance in Western
Australia (Garlinge 2005), and unfortunately there is typically a 5-20% yield penalty with
growing moderately preharvest sprouting tolerant genotypes. Growers are reluctant to grow
these lower yielding genotypes and as a consequence sprouting is a problem in the
sprouting prone regions of the south and northwest coastal regions of the Western
Australian wheat belt.
Traditionally the Western Australian wheat-breeding program has concentrated on
producing genotypes for the central and northern wheat belt, which produces 80 % of
Western Australia�s grain. These regions typically experience hot (>35°C max) and dry
(<25 mm of rainfall) conditions during grain filling, maturation and harvest. Consequently,
preharvest sprouting is seldom a problem in these regions (~1 in 10 seasons) and it has not
been a high priority in the breeding program. However, because of improved agronomic
practices, previously highlighted wheat is now also grown in the coastal regions of Western
Australia (~20 %). These regions are prone to preharvest sprouting (~1 in 4 season) and
have cooler and milder grain filling, maturation and harvest conditions. As the breeding
program has been aimed at the majority of the wheat production areas, recently released
broadly adapted high yielding Western Australian genotypes generally lack preharvest
sprouting tolerance.
Initial work by the Department of Agriculture Western Australia and interstate breeders in
2001 obtained inconsistent dormancy measurements of grain from field trials between
locations within Western Australia and between Western Australia and Eastern Australia,
despite identical harvest methods and assays in the one laboratory (Setter et al. 2001). This
variation was attributed to different environmental conditions experienced during grain
filling and maturation. Determining the level of dormancy required for these contrasting
environments which differ in the preharvest sprouting risk, poses several questions.

3
• What genetic level of dormancy is required in genotypes for the cooler and milder
Southern Coastal regions of the wheat belt where preharvest sprouting is a common
problem (1in 4 seasons) compared to the hotter and drier central and northern
regions of the wheat belt where preharvest sprouting is rare ( <1 in 10 seasons)?
• Are water supply and temperature the key environmental conditions during grain
filling which are responsible for the differences in dormancy?
• Selection for preharvest sprouting tolerance is currently based on determination of
dormancy phenotype measured by germination index at maturity, yet in practice the
real measure of preharvest sprouting is the falling number after rainfall. What level
of dormancy is sufficient and are other traits also important?
• How is the dormancy influenced by the environment? Plant ABA concentrations
increased by addition of ABA to hydroponic solution have been shown to modulate
dormancy (Suzuki et al. 2000). Elevating ABA concentration by applying
exogenous ABA during the soft dough stages, increases dormancy. The high
temperature and in particular moisture stress experienced during grain filling in the
central and northern regions may lead to an increase in plant ABA, increasing
dormancy and hence preharvest sprouting tolerance, in these regions but not the
coastal areas.
The overall hypothesis of this work is that stressful environmental conditions, such
as high temperature and moisture stress, increase dormancy by increasing the
embryo component of dormancy, and this in turn may result in a higher level of
preharvest sprouting tolerance in the field.

4
2 Chapter 2 Literature Review: Grain dormancy, environment and the expression of preharvest sprouting tolerance in wheat (Triticum aestivum L.).
This review of the literature focuses on the main mechanism of tolerance to preharvest
sprouting, i.e. dormancy, and regulation of that dormancy by the environment during grain
maturation. The review has three sections; the first presents background information on
preharvest sprouting; why it is a problem, how it is measured and the mechanisms of
tolerance. The second section reviews the current understanding of the physiology of grain
dormancy. The final section of the review then investigates how temperature and rainfall
i.e. the environment during grain filling (anthesis to maximum grain fresh weight, FW) and
grain maturation (maximum FW to maturity) influences dormancy. While there is
significant literature on the effects of germination temperature on expression of dormancy,
this review focuses on the impact of environmental conditions on the development of
dormancy i.e. during grain filling and maturation, not the expression of that dormancy at
different temperatures i.e. germination termperature.
2.1 Background on preharvest sprouting
Preharvest sprouting in wheat is the precocious germination of grain in the ear before it is
harvested. Preharvest sprouting induces changes in the physical and chemical composition
of the grain. Physically, preharvest sprouting results in the rupturing of the pericarp by
growth of the embryo and radicle. Visually the pericarp of the grain is split at the very least
(Fig. 1, grain 2), or completely ruptured with protruding shoot and radicle visible (Fig. 1,
grains 4, 5, & 6). Chemically, sprouting results in the production of enzymes involved with
mobilisation of grain reserves required for growth. In terms of grain quality, α-amylase is a
particular problem, since the low levels of α-amylase in flour produced from sprouted grain
are substantial enough to cause degradation of starch during the bread making process and
reduce the ability of the flour to make dough (Belderok 1968; Hagberg 1960; Hagberg
1961). Once α-amylase is present the grain may no longer be suitable for many end uses,
and hence preharvest sprouting reduces the quality of the grain.

5
Fig. 1. The range in visible sprouting of a sample of grain with a falling number of 62 s. All but grain 1 are visually sprouted. Grain from sprouted samples at Esperance in 2001/02 season. Photo T.B. Biddulph.
2.1.1 Measuring preharvest sprouting damage
Preharvest sprouting damage is determined by measuring the α-amylase activity in
sprouted grain with the Hagberd falling number method (from herein referred to as falling
number; Hagberg 1960; Hagberg 1961). The falling number method measures the time
taken, in seconds, for a weighted stirrer to fall a set distance through a heated paste of flour
and water. There is a close relationship between rainfall, α-amylase activity and falling
number of sprouted grain (Mares 1993), for instance a sample with a greater proportion of
visual sprouting usually has a lower falling number i.e. < 150 s (Fig. 2). However, the same
falling number is possible from a sample with a high proportion of slightly sprouted grains
and a sample with a lower proportion of more severely sprouted grains. Australian Wheat
Board receival standards (2003/04 season) set 300-350 s as a minimum for Australian
premium grades, 300-150 s for various general purpose milling grades and <150 s, i.e.
severely sprouted grain, for feed grades (Australian Wheat Board 2003-2004).
1 2 3 4 5 6
6
0
10
20
30
40
50
60
70
50 100 150 200 250 300 350 400 450Falling number (s)
Vis
ually
spr
oute
d (%
) General purpose
milling
Feed Premium grades
0
10
20
30
40
50
60
70
50 100 150 200 250 300 350 400 450Falling number (s)
Vis
ually
spr
oute
d (%
) General purpose
milling
Feed Premium grades
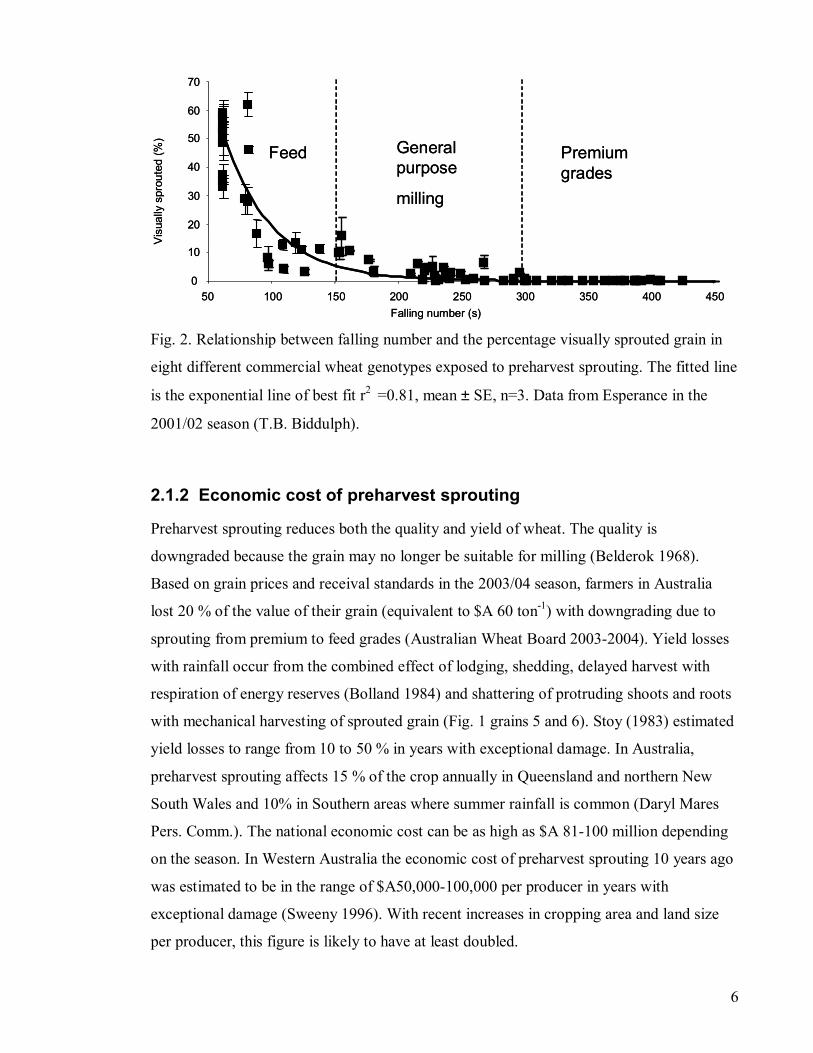
Fig. 2. Relationship between falling number and the percentage visually sprouted grain in
eight different commercial wheat genotypes exposed to preharvest sprouting. The fitted line
is the exponential line of best fit r2 =0.81, mean ± SE, n=3. Data from Esperance in the
2001/02 season (T.B. Biddulph).
2.1.2 Economic cost of preharvest sprouting
Preharvest sprouting reduces both the quality and yield of wheat. The quality is
downgraded because the grain may no longer be suitable for milling (Belderok 1968).
Based on grain prices and receival standards in the 2003/04 season, farmers in Australia
lost 20 % of the value of their grain (equivalent to $A 60 ton-1) with downgrading due to
sprouting from premium to feed grades (Australian Wheat Board 2003-2004). Yield losses
with rainfall occur from the combined effect of lodging, shedding, delayed harvest with
respiration of energy reserves (Bolland 1984) and shattering of protruding shoots and roots
with mechanical harvesting of sprouted grain (Fig. 1 grains 5 and 6). Stoy (1983) estimated
yield losses to range from 10 to 50 % in years with exceptional damage. In Australia,
preharvest sprouting affects 15 % of the crop annually in Queensland and northern New
South Wales and 10% in Southern areas where summer rainfall is common (Daryl Mares
Pers. Comm.). The national economic cost can be as high as $A 81-100 million depending
on the season. In Western Australia the economic cost of preharvest sprouting 10 years ago
was estimated to be in the range of $A50,000-100,000 per producer in years with
exceptional damage (Sweeny 1996). With recent increases in cropping area and land size
per producer, this figure is likely to have at least doubled.

7
2.1.3 Tolerance to preharvest sprouting
There are two mechanisms of tolerance to preharvest sprouting; (i) grain dormancy and (ii)
morphological characteristics of grains or ears, the latter reduces water uptake in the field.
A dormant grain will not germinate in a specific time period under conditions which
normally favour germination (Finch-Savage and Leubner-Metzger 2006). In wheat, a
dormant grain is defined as one that does not germinate in 7 days in the dark with adequate
water and oxygen at 20°C (Walker-Simmons 1987). Initial work comparing methods to
select preharvest sprouting tolerant genotypes determined a correlation (r = -0.56) between
a germination index of hand threshed grain and falling number after sprouting induced by
artificial wetting (Trethowan 1995).
Tolerance, determined as percent visual sprouting in ears in response to artificial wetting
had a low heritability compared with germination of isolated grain. Further work however
found a strong relationship between artificial wetting and falling number after natural
rainfall in the field (Trethowan et al. 1996). To overcome difficulties associated with
maturity and timing of rainfall this was carried out in an environment with consistent and
regular rainfall. Although an ideal environment to validate the relationship between
dormancy and preharvest sprouting tolerance, the results cannot be used to determine the
level of dormancy required for a target environment in Western Australia. Despite the
limitations, dormancy measured by germination index, remains the main criterion targeted
by breeders (Trethowan 1995; Trethowan et al. 1996; Xiao et al. 2002) and physiologists
(Gubler et al. 2005) in their efforts to improve tolerance of cereals to pre-harvest sprouting
in Australia.
Once concern with selection for dormancy however, is that dormancy is only transient, and
generally only lasts one month past maturity. After-ripening post maturity results in the
gradual loss of dormancy. Temperature greater than -10°C progressively reduces the level
of dormancy (Mares 1983b) and is more rapid at higher temperatures (Noda et al. 1994). In
the field, the level of dormancy required for preharvest sprouting tolerance is a function of
the dormancy present at maturity, the rate of loss of that dormancy, and the timing of a
particular rainfall event. If the level of dormancy is not high enough at maturity, the level
present at the rainfall event post maturity may not be high enough to confer preharvest

8
sprouting tolerance. Conversely, the duration the dormancy inhibits germination may be too
long, and hinder establishment of the crop in the following season. Hence a balance is
required between, the level of dormancy required for preharvest sprouting tolerance, and
the level that will not impact on the establishment of crop in the subsequent season. In the
sprouting prone regions of Western Australia, the level of dormancy required to meet these
considerations is unknown.
Morphological traits of the ear which reduce the duration the grain is wet with rainfall are
associated with minor levels of preharvest sprouting tolerance (King and Wettstein-
Knowles 2000). Ear characteristics such as awnlessness, surface waxes or glaucous lemma
and head nodding angle, (King 1984; King and Richards 1984; King and Wettstein-
Knowles 2000) reduce the duration the grain is wet during rainfall and hence reduce the
level of germination and preharvest sprouting. King and Richards (1984) found that
clubbed ears have 25 % greater water uptake than non-clubbed ears, and awned ears absorb
up to 30 % more water than awnless genotypes. In threshed grain there is also up to a two-
fold difference in the rate of grain water uptake that relates to the differences in physio-
chemical aspects of water imbibition into the grain (King 1984). No quantitative evidence
of the level of preharvest sprouting tolerance attributed to these morphological traits is
available.
In contrast, similar work by Mares (1983a) did not find significant genotypic differences in
water uptake by threshed grains, but did demonstrate variation between genotypes in water
uptake into grains in intact heads. The combined difference in water uptake caused by ear
and grain characteristics accounts for almost 20 % of the variation in sprouting in
genotypes with no dormancy (King and Richards 1984). The position of the ears in the crop
canopy and the susceptibility to lodging also influence the duration the grain is wet and
hence the preharvest sprouting tolerance of genotypes. These ear traits are not evaluated
with germination tests in isolated grain, and the level of additional protection they may
offer has not been quantified but is likely to be important (King and Wettstein-Knowles
2000). Genotypes, such as those with the awnless trait, that exhibit a slower water uptake
into intact heads or grain should have a higher falling number than other genotypes with a
similar level of dormancy after preharvest rainfall. Whether the level of tolerance from

9
morphological traits is sufficient for the preharvest sprouting prone regions of Western
Australia remains unknown.
2.1.4 Mechanisms of dormancy in wheat
Dormancy is typically found in genotypes expressing intermediate to tolerant preharvest
sprouting phenotypes. The dormancy is associated with inhibition of germination of
isolated embryos by exogenous ABA (Walker-Simmons 1987; Walker-Simmons and
Sesing 1990). Hence this component of dormancy is often termed �embryo sensitivity�.
The embryo component (E) on its own provides partial sprouting tolerance, whereas
complete tolerance appears to require combination with a specific seed coat factor (S). In
all genotypes a general seed coat component (s) is present and rupturing the seed coat
results in more rapid germination. However, inheritance studies (Mares 1998) have shown
that there is a specific effect that can be attributed to the seed coat in dormant but not non-
dormant genotypes. This specific seed coat component has an additive effect on the embryo
component, but it does not appear to provide substantial tolerance by itself, i.e. ES leads to
> dormancy than Es, but eS has no effect (Mares 1998). The exact mechanism of the
specific seed coat-based dormancy is not known, but current work focuses on a role of the
seed coat on physical parameters which influence germination. It may be due to inhibitory
compounds in the seed coat, or a restricted inflow of water or oxygen as in barley (Benech-
Arnold et al. 2006). Alternatively it may not be associated directly with the seed coat, but
may be associated with a wound response, which only occurs in grain with a damaged seed
coat.
Regardless of how the seed coat mechanism works, the seed coat is very susceptible to
mechanical damage. For example mechanical threshing removes the seed coat effect in
germination tests, and to avoid damage gentle hand threshing is required (Mares 1989). Red
grain wheat is typically more sprouting tolerant than white grain wheat because of the close
association of the red grain colour with the specific seed coat component (Warner et al.
2000). However, sprouting susceptible red grained wheat genotypes are relatively common
(Feurtado et al. 2004; Himi et al. 2002; Mares et al. 2005; Torada and Amano 2002;
Warner et al. 2000). The general and specific seed coat components have an additive effect
on the embryo component; hence a better understanding of embryo component is required

10
for a better understanding of preharvest sprouting tolerance and the possible role of the seed
coat.
2.1.5 Measuring dormancy Dormancy is typically measured by either a germination index (ranging from 0.0 to 1.0)
(Reddy et al., 1985) or germination resistance index (ranging from 0 to 50) calculated on
hand threshed grains or embryo half-grains imbibed on filter paper. The germination index
is a weighted index which gives maximum weight to grains which germinate early, and
progressively less weight to grains which germinate later (Reddy et al., 1985) while the
germination resistance index measures the relative rate of germination, by estimating the
time to 50 % germination (Gordon, 1971). There are several papers which detail the relative
merits of different assay methods, however most agree there is a significant correlation
between seed dormancy measured on grain from threshed heads and assays for sprouting
tolerance based on intact spikes (DePauw and McCaig 1991; DePauw et al., 1989; DePauw
et al., 1993).
One of the problems with the resistance method is that depending on ripening conditions
and the level of dormancy a significant number of genotypes may not reach 50 %
germination over 7 days requiring long observation times or extrapolation. Furthermore
estimating the time to 50 % requires you to graph out the data or generate a curve of best fit
which can be problematic. Practically the main difference between the two index�s is that in
genotypes which reach the same total germination percentage at 7 days the germination
index can differentiate better between genotypes which germinate rapidly during the first
three days compared to genotypes which germinate at a slower rate. This ability may give
an important differentiation in the field as given a relatively small rainfall event, the slower
germination rate, will result in a lower proportion of germinated grain, and hence greater
sprouting tolerance.
2.1.6 Molecular markers for dormancy and preharvest sprouting
Molecular markers have been developed for genes associated with the control of dormancy
in wheat and hence preharvest sprouting tolerance. The main markers associated with
dormancy are localised on the long arm of chromosome 4A (Flintham et al. 2002; Kato et

11
al. 2001; Lohwasser et al. 2005; Mares et al. 2005; Tan et al. 2006; Torada et al. 2005),
and appear to flank the gene associated with the embryo component (Mares and Mrva
2001). The markers Xgwm397, Xgwm269 and Xbarc170 appear to be closely linked to the
dormancy gene(s). They have been confirmed in three genotypes of diverse origin (Mares
et al. 2005), and thus appear to be indicative of an important source of dormancy in wheat.
These markers are generally within 10 cM from the gene(s), depending on the population.
Recent work has also found a marker for a minor QTL on the long arm of chromosome 5B
associated with dormancy (Tan et al. 2006).
There has also been research on genes for dormancy and preharvest sprouting in wheat
using the synteny between rice, wheat and barley (Li et al. 2004). The usefulness of
markers however, is only ever as good as the system employed to identify them. For
example a QTL reported for dormancy in barley (on 2H) (Li et al. 2003) was later found to
control maturity (Li, C. D. Pers. comm), possibly indicating that the environment during
grain filling and maturation was different in early compared to late maturing genotypes
when the population was phenotyped. Further discussion of molecular markers and their
use in breeding programs is outside the scope of this thesis, and further details on the
current molecular understanding of dormancy in wheat can be found in a review by Li and
Foley (1997) and recent papers by Flintham et al. (2002), Li et al. (2004), Torada et al.
(2005), Mares et al. (2005), and Tan et al. (2006).
2.1.7 Influence of Black point and fungal infection on dormancy and preharvest sprouting
Black point is a physiological or pathnogenic defect in grain which predominantly results in
a dark discoloration of the embryo end of the grain (Rees et al. 1984; ). High levels of black
point (> 5 %) result in downgrading of grain at receival (Australian Wheat Board 2003-
2004). Black point is thought to be associated with oxidation of phenolic compounds,
however the cause has not been determined.
Black point is induced by high temperature, humidity and nitrogen fertilizer during the soft
dough grain filling stages (Moschini et al. 2006) and hence is often associated with fungal
infection of the grain (predominantly Alternaria spp.) which also occurs under the same
conditions (Conner et al., 1992). Certain levels of nitrogen nutrition and foliar fungicide

12
application which were associated with an increase in grain weight also increased the
incidence of blackpoint and fungal infection of the grain (Gooding et al., 1993; Ruske et al.,
2003; Wang et al., 2002). Depite a clear association betwen resistance to fungal infection
and resistance to black point, there still remain conflicting views on whether the fungal
infection is the actual cause of black point (Conner and Davidson 1988; Lorenz 1986), or
just associated with the same conditions that induce black point (Williamson 1997).
Black point and fungal infected grains tend to germinate more rapidly than sound grain
from the same sample (Fernandez et al., 1998; Fox et al., 2003; Williamson 1997), and for
this reason they are omitted from dormancy germination tests (Mares 1989). However
despite omitting black point affected grains, recent work by Tan et al. (2006) still found an
association of a potential dormancy QTL with the same region as the black point QTL
reported by Lehmensiek et al. (2004). Work by Fox et al., (2003) also highlighted the
reduction in dormancy and hence sprouting tolerance with fungal infection in both dormant
and non-dormant genotypes. In Austrlaia the combined count of fungal and black point on
grain is termed weather affected grain and is another quality trait measured at receival
(Australian Wheat Board 2003-2004). Weather affected grains often result in downgrading.
As preharvest sprouting, black point and fungal infection are all associated with preharvest
rainfall and black point and fungal infection promote germination; developing preharvest
sprouting tolerance needs to occur in parallel with lower incidence of black point and
fungal staining.
2.2 Physiology of dormancy; role of Abscisic acid and other
compounds in dormancy
Abscisic acid is important for normal embryogenesis, and under controlled or in optimum
environments appears to be involved in the establishment and maintenance of grain
dormancy in cereals (Benech-Arnold et al. 2006; Benech-Arnold 2002; Gubler et al. 2005;
Jacobsen et al. 2002; Suzuki et al. 2000). Endogenous ABA concentrations during late
grain maturation are also strongly linked to the normal maturation pathway and inhibition
of precocious germination or vivipary before grain maturation. Mutants of Arabidopsis
(Karssen et al. 1983), maize (White et al. 2000) or wheat (Nakamura and Toyama 2001)
with low ABA, or reduced ABA sensitivity often lack grain dormancy and exhibit
precocious germination before grain maturation. Under controlled conditions ABA is

13
important in maintaining dormancy in mature wheat grain, and low ABA concentrations
caused by fluridone application during grain filling removes or prevents the development of
dormancy (Garello and Le Page-Degivry 1999; Rasmussen et al. 1997). Stressful
conditions during grain filling which are not optimum, may influence the concentration of
plant ABA and also influence the level of dormancy.
In wheat, during grain filling under well watered controlled conditions at 15/ 25°C or under
optimum environments, there are usually two peaks of endogenous ABA (King 1993;
Suzuki et al. 2000). The first peak is contributed by the plant tissues and occurs at around
25 days post anthesis (dpa) (Fig. 3). The second peak is attributed to the embryo and
reaches a maximum two weeks before maturity (typically at 0.9-2.5 µg g-1 dry weight
between 40-50 dpa), coinciding with maximum dry matter and fresh weight of the grain
(King 1976; Koshkin and Tararina 1990). At this development stage the ABA is produced
in the grain (King 1979), possibly to promote the transfer of photoassimulates to the grain
(Dewdney and McWha 1978). Following the second peak, the ABA concentration
decreases rapidly during grain drying / maturation to around 0.2-0.4 µg g-1 DW at maturity,
typically at 55-65 dpa (King 1979; Suzuki et al. 2000). In the field however, environmental
conditions are not ideal, temperature fluctuations are large and days to maturity can be
reduced by temperature and moisture stress during grain filling (Stone and Nicolas 1995).
Under field conditions the pattern of ABA accumulation and degree of dormancy may
differ from that under ideal controlled environments, as in King (1993), or optimum field
conditions reported by Suzuki et al. (2000), particularly in the Australian wheat belt which
is often characterized by terminal moisture stress and high temperatures (> 30°C).

14
0 10 20 30 40 50 60
Time during grain filling (dpa)
Rel
ativ
e A
BA
conc
entr
atio
n
Maternal driven
Embryo drivengrain drying
Maturity
0 10 20 30 40 50 60
Time during grain filling (dpa)
Rel
ativ
e A
BA
conc
entr
atio
n
Maternal driven
Embryo drivengrain drying
Maturity
Fig. 3. Generalised time course of change in abscisic acid concentration in embryos of
wheat grain during grain filling and grain maturation under optimum environments.
Adapted from King (1993) and Suzuki et al. (2000).
In wheat and barley, the ABA concentration in the husk, pericarp and embryo are deemed
important in maintaining dormancy, while that of the endosperm less important (King
1989). Investigation of ABA mutants of tobacco and Arabidopsis indicates that only ABA
produced in the embryo of the grain, and not ABA of plant origin, determines the degree of
dormancy (Frey et al. 2004; Karssen et al. 1983). However the surrounding tissues and
parent plant may play a role in encouraging and supporting the embryo to synthesise ABA
(Kermode 2005). Movement of ABA to the embryo from the husk and pericarp during
imbibition may also be important in maintaining dormancy (King 1989). Since it is the
embryo, scutellum and aleurone that control germination and reserve mobilisation of the
endosperm (King 1989) its here where the ABA concentrations are critical. The
concentration of ABA in the embryo is typically double that of the other grain parts
combined and hence the ABA concentration of embryos is measured in studies on
dormancy.
Between dormant and non-dormant wheat there are no significant differences in
endogenous embryo ABA concentrations which explain the difference in dormancy during
grain filling, grain maturation or maturity, in plants grown under controlled environments
(Walker-Simmons 1987) or optimum field conditions (Suzuki et al. 2000). Dormant

15
genotypes however tend to have a higher ABA concentration during grain filling, than non-
dormant genotypes, it is just not significant at P<0.05. These observations were
subsequently confirmed in wheat (Hagemann Wiedenhoeft et al. 1988; Himi et al. 2002;
Walker-Simmons and Sesing 1990) and barley (Millar et al. 2006; Romagosa et al. 2001).
Whilst there are no differences in endogenous ABA concentrations during grain filling,
grain maturation or at maturity, a role for ABA in expression of dormancy is possible as
exogenous ABA further reduces germination in dormant or partially dormant grains.
Walker-Simmons (1987) suggested that the embryo of dormant genotypes were more
sensitive to endogenous ABA, whilst Garello and LePage-Degivery (1999) proposed that
the embryo may vary in its capacity to synthesise ABA, however a combination of both is
likely.
2.2.1 Abscisic acid during imbibition
Abscisic acid concentration during imbibition controls dormancy in grain of other plants
which have been more closely studied and the current thinking is highlighted in Fig. 4. In
Arabidopsis, for example, dormant seeds maintain a high concentration of endogenous
ABA during imbibition and do not germinate, whereas non-dormant seeds, which have
been after-ripened, stratified or treated with the ABA inhibitor, fluridone, germinate when
the ABA concentration falls in the first few hours of imbibition (Ali-Rachedi et al. 2004).
Seeds of other plants such as tobacco (Grappin et al. 2000), pine (Feurtado et al. 2004) and
barley (Benech-Arnold et al. 2006; Jacobsen et al. 2002) show a similar response. Initial
controlled environment work looking at ABA content of imbibing isolated embryos of
wheat found dormant genotypes had higher ABA concentrations than non-dormant
genotypes 4 h after commencement of imbibition (Ried and Walker-Simmons 1990).
However by 18 h the isolated embryos of dormant genotypes had germinated and there
were no differences in ABA concentration compared to the embryos of non-dormant
genotypes which germinated 12 h earlier. One concern with this work is that germination of
isolated embryos does not mimic what happens when intact grains are imbibed. Isolated
embryos germinate much more rapidly (dormant embryos 18 hrs compared to 72 hrs in
dormant grain) as there is no additive effect of the seed coat. The brief period of differences
in embryo ABA concentration during imbibition in isolated embryos may be due to the
rapid germination of isolated embryos compared to intact grain. Further work is required in
wheat to confirm that dormant grains maintain a higher embryo ABA concentration

16
imbibition. Work with intact grains would also demonstrate that this is important for
dormancy in intact grains, making this knowledge more transferable to preharvest sprouting
tolerance.
In field grown plants under optimum conditions dormant embryos ABA concentration was
higher than non-dormant embryos for 8-12 hrs of imbibition, before the dormant embryos
started to germinate at 12 hrs (Suzuki et al. 2000). There was leakage of ABA out of the
isolated embryos as shown in the results of Suzuki et al. (2000) which may explain the
more rapid germination. However, only germination data for intact grains is present, not of
isolated embryos, so it is not clear if the concentration of ABA is responsible for the
differences in germination in whole grains. In contrast, in other field work there were no
differences in whole grain ABA between genotypes differing in dormancy (Tavakkol
Ahshari and Hucl 2001). However, the lack of difference in ABA between the genotypes
may be due to the small difference in dormancy between the genotypes, as they were all
relatively dormant, and/or the dilution of ABA caused by measuring the whole grain ABA,
not the embryo ABA. In wheat there are conflicting results in field grown plants and there
has been no detailed study during imbibition on the concentration of endogenous embryo
ABA isolated from intact grain of genotypes contrasting in dormancy. Yet germination of
intact grain at maturity is what is screened for preharvest sprouting tolerance (Trethowan et
al. 1996), and this is where and when the capacity to germinate or not impacts on
preharvest sprouting tolerance. If the hypothesis is that ABA is associated with grain
dormancy, then during imbibition embryos from intact dormant grain should maintain a
higher embryo ABA concentrations than germinating non-dormant grain for the period
during imbibition they maintain dormancy.
Maintaining a high ABA concentration during imbibition and therefore dormancy can be
due to either greater biosynthesis of ABA (Tavakkol Ahshari and Hucl 2001), reduced
catabolism of ABA (Garello and Le Page-Degivry 1999) or a combination of both (Fig. 4).
Alternatively there may be greater influx of ABA from other tissues. Recent expression
studies in barley and Arabidopsis have concluded that the lower ABA content of non-
dormant grain was due to catabolism of ABA by ABA 8�-hydroxylase exceeding
biosynthesis resulting in a drop in ABA content below the concentration required to
maintain dormancy (Chono et al. 2006; Millar et al. 2006). Husk-imposed dormancy in

17
barley is also due to the husk maintaining hypoxic conditions around the embryo
minimising degradation of endogenous ABA by oxidation (Benech-Arnold et al. 2006).
Work in beechnut (Barthe et al. 2000) found similar results and proposed that the low
concentration of oxygen was due to covering structures consuming oxygen, possibly by
peroxidase activity of phenolic compounds. Other work in rice has also shown that
phenolics can impose dormancy in this manner (Naversero et al. 1975). Regardless of how
the ABA concentration is elevated in dormant grain, confirmation that there are differences
in endogenous concentrations of ABA during imbibition of intact grains is required before
detailed studies on how this comes about are warranted.
After-ripening Stratification Seed coat damage
After-ripening Stratification Seed coat damage
After-ripening Stratification Seed coat damage
Fig. 4. Model of the regulation of ABA metabolism in embryos of imbibed grains from
controlled environments following dormancy release, indicating the synthesis of ABA from
Carotenoids and the breakdown of ABA to Phaesic acid (PA). Adapted from Gubler et al.
(2005).
2.2.2 Compounds which promote germination; Giberellic acid
Giberrellic Acid (GA) is often applied to grains to promote germination. Both ABA and
GA are important for development of the embryo, and ABA in maintaining dormancy and
preventing precocious germination (Ali-Rachedi et al. 2004). In wheat and barley aleurone
cells, ABA induces the production of a protein (PKABA1), which is part of the mechanism
involved in ABA inhibition of GA-induced α-amylase or proteinases (Gomez-Cadenas et
al. 1999; Johnson et al. 2002; Shen et al. 2001). In mature grain GA only induces

18
germination once endogenous embryo ABA concentration declines (Appleford and Lenton
1997). There is general agreement in the literature that GA is not responsible for dormancy
loss, but rather its primary function is in promoting moblisation of grain reserves to supply
the germinating embryo once ABA concentrations have declined (Lenton et al. 1994).
Recent reviews by Gubler et al. (2005) and Finch-Savage and Leubner-Metzger (2006)
provide more detail on these processes. The role of GA in promoting mobilisation of grain
reserves for seedling growth will not be investigated in this project, as it occurs once ABA
concentrations have declined and dormancy has been lost.
2.2.3 Other compounds which inhibit germination; Bran extracts, tryptophan and indoleacetic acid
There are several other compounds which are known to inhibit germination of wheat
including water soluble bran extracts, tryptophan and indoleacetic acid. Imbibing non-
dormant grains with bran from dormant red grain wheat results in inhibition of germination
(Himi et al. 2002). L-tryptophan, a precursor for the auxin, indoleacetic acid, has been
purified from water soluble bran extracts from dormant wheat (Morris et al. 1988). The
inhibition of germination by role of tryptophan and indoleacetic acid (precursors of auxin)
has since been confirmed by Ramaih et al. (2003). The level of germination inhibition of
auxin applied to excised dormant embryos (Ramaih et al. 2003) is similar to that reported
for ABA (Walker-Simmons 1987). Although auxin and its precursors can inhibit
germination, they appear to complement the role of abscisic acid in inhibiting germination
(Ramaih et al. 2003), hence they will not be considered further.
2.3 Regulation of dormancy by environment
During the later stages of grain filling and grain maturation the level of dormancy at
maturity develops and the environmental conditions during this time interacts with it.
Strand (1989a) speculated that 10-65 % of the non-genetic variation in grain dormancy can
be attributed to the weather conditions during grain maturation in the 20 days prior to
maturity. Correlations run on field trials have found that temperature and rainfall between
anthesis and maturity are two factors which have an influence on the expression of
dormancy (Mares 1993; Nielsen et al. 1984).

19
Mares (1993) and Nielsen et al. (1984) found that large diurnal temperature fluctuations
reduce preharvest sprouting tolerance in the field. Mares (1993) also found in field work
that rainfall during grain maturation in the 20 days prior to maturity accounted for almost
85% of the variation in sprouting tolerance in a set of locally adapted commercial
genotypes. In the field however it is difficult to differentiate between the effect of rainfall
on directly wetting the heads and the effect of water supply to the plant. Furthermore in the
Australian wheat belt low temperatures are often associated with rainfall and high
temperatures (>30°C) with terminal moisture stress. Limited controlled environment work
has been carried out on the effect of high temperature and moisture stress (Auld and
Paulsen 2003), but is inconsistent with preliminary work under Australian conditions
(Mares 1993; Setter et al. 2001). The Australian wheat belt typically experiences terminal
moisture stress with high temperature and low relative humidity, and there is no published
field based work that quantifies the separate effects of temperature and water supply on
dormancy under these conditions.
2.3.1 Temperature during grain filling and grain maturation
The effect of temperature during grain filling and grain maturation on dormancy appears to
depend on the stage of grain development. In wheat, low temperatures during grain filling
generally increase dormancy (Lunn et al. 2002; Reddy et al. 1985; Walker-Simmons and
Sesing 1990), whereas once the grain is mature, low temperatures during imbibition reduce
expression of that dormancy (Mares 1984; Nyachiro et al. 2002; Reddy et al. 1985; Ueno
2002). This review focuses on the role of temperature during grain filling (anthesis to
maximum FW) and grain maturation (maximum FW to maturity) on the dormancy
expressed at maturity, not the effect of germination temperature.
In controlled environment room experiments, constant low temperatures increase dormancy
compared to constant high temperatures. Grain matured under constant 15ºC compared to
25-26ºC results in slower grain drying, heavier grain and more dormant grain (Reddy et al.
1985; Walker-Simmons and Sesing 1990). Work in controlled environments has found that
consistently low temperatures during grain filling result in more dormant grain, however
temperatures are never constant in the field and fluctuate hourly, diurnally and daily with
the weather patterns. Rodriguez et al. (2001) developed a model in barley based on
temperature sums above 5°C throughout grain filling and found that low temperatures,

20
during a window of sensitivity, increased dormancy. King (1993) proposed a similar
window of sensitivity in wheat at 40-50 dpa from controlled environment work where
humidity can influence dormancy. Similar models based on temperature were proposed for
wheat by Belderok (1968) and Lunn et al.(1998), however these were problematic and
never validated in the field (Lunn et al. 2002). In the field, correlation analysis or modelling
work has produced conflicting evidence for the influence of temperature on dormancy,
possibly because of the use of different sites and seasons to get the different temperature
treatments, and the association of low temperature with rainfall. Work needs to be done in
the field with controlled temperatures to elucidate the effect of temperature without the
confounding effect of rainfall and low temperature and different sites or seasons.
In the field, slight changes in dormancy have been reported with different temperature
patterns during grain filling, but not to the same extent as under controlled environments.
Mares (1993) for example observed that grain matured at lower temperatures (average max.
26ºC, min. 8ºC over a 20 d period prior to harvest) was slightly more preharvest sprouting
tolerant than at higher temperatures (average max. 34ºC, min. 17ºC during the 20 d period
prior to harvest). Nielsen et al. (1984) also found reduced preharvest sprouting tolerance
with high temperatures in the 2 weeks before maturity which is consistent with effects of
constant temperature on dormancy under controlled conditions, but the effect was several-
fold lower. In contrast in other field experiments there was no relationship between
preharvest sprouting tolerance and temperature once the year effects were removed by
analysis of covariance (Kettlewell and Cashman 1997). In the field trials reported by Mares
(1993), Nielsen et al. (1984) and Lunn et al. (2002), there were always the confounding
factors of site and season with the different temperature treatments. Furthermore, the
magnitude of differences in temperature in one environment tended to be smaller than
conditions in controlled conditions. In addition, field observations over 10 seasons by
Strand (1989b) led him to suggest that the effect of temperature may be genotype specific.
A partially dormant genotype may be sensitive to temperature, whereas a strongly dormant
genotype may be less sensitive. More work is required in the field to confirm the effect of
temperature during grain filling and grain maturation without the interaction and
confounding factor of site and season. Controlled conditions in the field might be able to
quantify more precisely the influence of temperature during grain filling and grain
maturation on dormancy, and confirm suggestions from previous work in controlled

21
conditions and correlation analysis of field trials that low temperature induces more
dormancy and high temperature induces less dormancy.
2.3.2 Rainfall / water supply
Rainfall before harvest reduces dormancy and hence the sprouting tolerance of the grain,
possibly through two slightly different effects on the plant. Rainfall during grain maturation
slows the grain drying rate and results in a higher grain moisture content for longer (Lunn
et al. 2002). Hence in trials where the rainfall was allowed to wet the heads during grain
maturation, rainfall accounted for almost 85 % of the variation in falling number after a
standard 15 day wetting treatment (Fig. 5; Mares 1993). Grain which received more rainfall
was less sprouting tolerant. Based on these observations it was recommended that, for
accurate characterisation of germplasm for preharvest sprouting tolerance, rainfall must be
excluded from sprouting nurseries during the last two weeks before maturity (Mares 1993;
Trethowan 1995). However this may affect the second way rainfall can influence
dormancy, through water supply. Without supplemental irrigation, covers to exclude
rainfall may impose a heat and/or moisture stress on plants and subsequently affect the
dormancy.
Rainfall may also influence preharvest sprouting tolerance, as a lack of rainfall is often
associated with moisture stress in a Mediterranean climate. In experiments conducted by
Mares (1993) without irrigation, the long season wheat genotype, Kleiber, experienced
severe moisture stress and was more dormant and hence more preharvest sprouting tolerant
with a four fold higher falling number after rain simulation than irrigated plants. In
controlled environments, a similar increase in dormancy was observed under low humidity
(35-40 %) compared to high relative humidity (90-100 %), in a glasshouse grown genotype
Suneca (King 1993). However this is not consistent with work in controlled environments
that found moisture stressed plants were less dormant than well watered controls (Auld and
Paulsen 2003). There are few if any field experiments which separate the effect of rainfall
into the component effects on (i) grain wetting and (ii) water supply. Research is required
to quantify the effects of moisture stress during grain filling in the field compared to well
irrigated controls in the absence of grain wetting.

22
y= 412-3.8x r2=0.84y= 412-3.8x r2=0.84
Fig. 5. The relationship between falling number after rainfall simulation 15 days post
maturity and the total rainfall in the 20 days prior to harvest. Data collected from samples
taken from 20 field trials over the period 1980 to 1986. Adapted from (Mares 1993).
2.3.3 Interaction of temperature, rainfall and moisture stress
In the field, the effect of, and interaction between, rainfall and temperature are difficult to
separate. In temperate and Mediterranean climates, low temperatures are commonly
associated with rainfall events, whereas high temperatures are often accompanied by low
rainfall and in some cases (in Mediterranean climates) moisture stress. Though the
influence of temperature and rainfall during grain filling has been widely studied in field
trials by correlation analysis in Mediterranean (Australia; Mares 1993), temperate (United
Kingdom; Kettlewell and Cashman 1997; Lunn et al. 1998; Lunn et al. 2002) and
continental (Kansas; Nielsen et al. 1984) climates, their effect and interaction are still
poorly understood. The interaction between temperature and rainfall and the confounding
factors of site and season with the different treatments makes attributing the sole influence
to temperature, rainfall or water supply in the field very difficult or impossible to separate
with correlation analysis. Future work on dormancy needs to confirm the effects of
temperature and water supply in the field without the interaction between low temperatures

23
and rainfall, high temperatures and moisture stress, the association between rainfall and
grain wetting and the possible confounding effects of different sites and seasons.
2.3.4 Influence of environment on dormancy, a role for ABA
Synthesis of ABA is also stimulated / enhanced when plants are subject to stress from
temperature extremes, moisture deficit, hard pans, salinity or pathogen infection (Leung
and Giraudat 1998; Rock 2000; Ross 1992). Abscisic acid typically causes responses that
help to protect the plant against these stresses. For example, ABA protects plants from
dehydration in response to extremes in high temperatures, moisture stress or salinity by
closing stomata and inhibiting stomata opening to reduce transpiration (Leung and Giraudat
1998; Ross 1992). One role of ABA is to prevent dehydration when the plant perceives the
water supply is limited or may become more difficult to obtain under moisture deficit.
As plant ABA concentrations are associated with the plant response to stress, it is likely
that plants grown under a high stress environment during grain filling may result in more
dormant grain than plants grown under optimum field conditions, particularly if the plant is
stressed enough to increase the plant driven peak in embryo ABA concentration at 25 dpa
(Fig. 3; Section 2.2). Previous observations from Mares (1993) with the long season
genotype, Kleiber, where terminal moisture stress in the field was associated with elevated
dormancy, support this hypothesis. Similarly, comparisons of sites differing in ripening
conditions (Mares 1993), as well as field observations from the Department of Agriculture
Western Australia in which material grown in the central wheat belt (Merredin) which
experiences terminal moisture stress was compared with the coastal wheat belt (Esperance)
which experiences more rainfall and little moisture stress (Setter et al. 2001), are consistent
with this hypothesis. By contrast in pot-based experiments, Auld and Paulsen (2003) found
that moisture stress reduced dormancy under a similar high temperature regimes. There are
conflicting observations between field grown plants and pot grown plants on the effect of
moistures stress during grain filling.
Experiments with pots however, are problematic and often have unrecognised artifacts, see
recent review by Passioura (2006). In pots there may be a background response
concentration of ABA generated by roots of plants encountering an impenetrable layer such
as a pot wall. When cereal roots hit compacted soil, the xylem sap ABA concentrations

24
increase four-fold, possibly increasing flag leaf ABA. Higher flag leaf ABA is apparent in
moisture stressed plants (Westgate et al. 1996). ABA in the flag leaf is transported into
grain (Goldbach et al. 1977) so root restriction by pots might increase the ABA content in
grain in the same manner that moisture stress increases the ABA content in floral organs of
wheat (Westgate et al. 1996). The sudden imposition of moisture stress in millet can lead to
a faster and higher concentration of ABA in leaves (Henson 1985). The smaller soil volume
associated with pots can lead to a more sudden and severe moisture stress and higher ABA
concentrations in leaves than in field grown plants under similar moisture stress conditions
(Henson 1985). Put simply, the sudden drying of the small soil volume in a pot may impose
moisture stress more severe than in the field where soil volume is larger and contribute to
higher ABA accumulation in plants grown in pots than those grown in the field.
There are several possible causes for the observed differences in dormancy in plants
exposed to moisture stress between pot and field grown plants. In moisture stress
experiments, pots typically hold 3 kg of potting mix, are 16 cm in diameter with a depth of
15 cm, and often have 5 plants per pot. This equates to a planting density of ~450 plants m-
2, a rooting depth of little more than 15 cm and a 20-fold reduction in soil per plant.
Commercial crops in Western Australia are sown at an optimum plant density of 120-150
plants m-2 (Del Cima et al. 2004) with a rooting depth of around 1 m. Hence even in well
watered plants in pots, the root binding and restricted rooting volume may influence the
concentration of plant ABA and hence dormancy. Furthermore, imposition of moisture
stress and field capacity in pots is restricted by the lack of soil pores in potted soil or the
abundance of large pores in potting mix (Passioura 2006). One indication of the potential
problems associated with moisture stress based pot experiments is the lack of yield
reduction in moisture stressed pots compared to �field capacity� controls in pot-based work
(Auld and Paulsen 2003). Field work is required where rooting depth is unrestricted to
ensure well watered controls have limited plant ABA at the first peak of embryo ABA
concentration, and moisture stressed treatments are realistic and mimic conditions that
normally occur in the field and the concentration of ABA accumulation and hence
dormancy that is representative of the field environment.

25
2.3.5 Predicting risk of preharvest sprouting based on environmental factors
Several studies have tried to predict the risk and likelihood of preharvest sprouting using
temperature during grain filling to predict the level of dormancy of the crop at maturity.
Belderok (1968) developed a model based on temperature sums above 12.5°C after
systematic studies in controlled environments to predict the level of dormancy. This
correlation has not been validated in the field and other studies in Europe have reported
weak correlations between the single factor of temperature and dormancy (Lunn et al.
2002; Strand 1983; Strand 1989b; Strand 1990). In Norway, Strand (1989b) in particular
found the effects of temperature and rainfall were genotype specific, in some genotypes
temperature was the predominant factor influencing dormancy, while in others it appeared
to be rainfall. Strand (1983) also noted that in warm seasons maximal dormancy is obtained
earlier during grain filling compared with cooler seasons. This may explain the poor
correlations between temperature and dormancy across seasons that differ in temperature
and rainfall. As originally proposed by Strand (1989b), a warning system needs to be based
on actual dormancy measurements at maturity to indicate the likelihood of preharvest
sprouting and the falling number at harvest-ripeness. Development of a risk prediction
system could be problematic as specific genotypes respond in different ways to temperature
and rainfall. Further work is required to confirm the influence of temperature and water
supply, and determine if genotypes with different mechanisms of dormancy respond in
different ways before a risk prediction system can be fully developed.
2.4 Research directions for this thesis
Dormancy is the major trait being targeted by breeders (Trethowan 1995; Xiao et al. 2002)
and physiologists in an effort to improve crop tolerance to pre-harvest sprouting but the
mechanisms involved and the environmental influence on them is not well understood.
Abscisic acid concentration and/or embryo sensitivity to ABA appear to be involved in the
embryo component of dormancy, and a better understanding of the role of ABA in inducing
and maintaining dormancy in intact grains during imbibition is important. Much of the
research on ABA has been in model systems or for plants grown under controlled
environments, and the information is not necessarily transferable to whole plants growing
in a range of field environments where preharvest sprouting is the problem.

26
Environmental conditions during grain filling can have a large effect on expression of
dormancy. Results obtained in controlled environment responses do not correlate strongly
with field trials and explanations for this difference may include several factors. Firstly
field trials, even with the use of different sites or seasons to get various temperature and
rainfall combinations, generally do not involve temperature differences as great as those in
controlled environments. In addition, field temperatures during grain filling follow a
diurnal fluctuation over day and night, whereas controlled environment experiments are
usually carried out at a constant temperature, e.g. 15 or 25-26ºC (Reddy et al. 1985;
Walker-Simmons and Sesing 1990). Secondly, other environmental factors such as rainfall,
which wets the grain and affects water supply are not factors in controlled environment
work. Furthermore in the field, water supply, wetting of the grain and rainfall can not be
separated from each other, and usually coincide with low temperature. In all of the recent
field work on temperature and rainfall by Lunn et al. (2002), Strand (1989b) and Mares
(1993) they have used different sites and seasons to get different environmental conditions
and then tried to correlate these with the different temperatures and rainfall events in the
different sites and seasons. As a result, site and season are often confounding factors in the
analysis. This combination of confounding factors makes correlating a response to either
temperature, rainfall (water supply or grain wetting) or the interaction between temperature
and water supply difficult. There is a clear need for work to be carried out in the field under
conditions where different temperature and water supply during grain filling are closely
monitored, and possibly controlled, and where grain wetting is excluded.
Controlled conditions in the field may be able to quantify more precisely the influence of
temperature during grain filling on dormancy, and confirm the previous work in controlled
conditions that low temperatures induce more dormancy and high temperatures induce less
dormancy. Further work is also required to determine if the differences in dormancy due to
the different environmental conditions in the target environment result in different levels of
preharvest sprouting tolerance (falling number) in response to preharvest sprouting rainfall.
This work needs to be carried out in the target environment to answer the primary
questions; What level of dormancy is required for preharvest sprouting prone regions in
Western Australia? Is the effect of temperature and water supply during grain filling and
maturation consistent between genotypes and worth worrying about? Then how does the
dormancy control germination and provide preharvest sprouting tolerance?

27
2.5 Thesis aims
This thesis presents original research on the embryo component of dormancy of wheat in
the field, the interaction of temperature and water supply during grain filling and grain
maturation on expression of dormancy, and the ability of that dormancy to provide
tolerance in the target environment. Preharvest sprouting is a problem in 1 in 4 years in
affected areas of the Western Australian wheat belt. However, the environmental conditions
experienced differ greatly between the cooler and milder coastal areas where preharvest
sprouting is more prevalent and the hotter and drier central regions, which are less prone to
preharvest sprouting. This difference must be considered during development of protocols
for screening genotypes for dormancy and preharvest sprouting tolerance to ensure
sprouting tolerance is robust. It is unlikely that the level of dormancy at harvest-ripeness in
the central and northern regions is equivalent to preharvest sprouting tolerance in the cooler
southern coastal regions. The key objectives of this research project were to therefore test
the following hypotheses:
1. Dormant grain will maintain a higher endogenous embryo ABA concentration
during imbibition of intact grain compared to non-dormant genotypes or after-
ripened grain for the time period where their germination is different, (Chapter 3).
2. During grain filling, conditions which increase the plant concentration of ABA (high
temperature and moisture stress), may also increase the level of dormancy in the
grain at maturity by increasing embryo sensitivity to ABA (Chapter 4).
3. Seasons, which favour development of greater dormancy will provide better
tolerance to preharvest sprouting leading to better grain quality in all genotypes than
seasons which do not (Chapter 5).
4. Protection against preharvest sprouting will be directly related to the level of
dormancy, with strongly dormant genotypes maintaining a higher falling number,
than partially dormant and non-dormant genotypes (Chapter 5).
These hypotheses are each considered in more detail in the designated chapters.

28
2.6 Limitations
Quarantine embargos, as a result of Wheat Streak Mosaic virus outbreaks in Eastern
Australia in 2003 and early 2004, prohibited the importation of any wheat grain into
Western Australia. As a result, the range of germplasm available to initiate this project was
limited to germplasm that was already available within breeder�s collections in Western
Australia. This unfortunately meant that an important set of doubled haploid genotypes
differing in preharvest sprouting tolerance from the AUS 1408/ Cascades doubled haploid
population were not included in the first year�s experiments although they were available in
subsequent years.
ABA measurements were only able to be carried out on one set of data, due to technical
problems with instruments and the transfer of technical staff. Insufficient money was
available for analysis elsewhere. This meant, samples taken throughout grain filling in
2003/04 and 2004/05 were not measured as planned in a subsequent trip to CSIRO Plant
Industry. Furthermore the initial ABA work was not able to be confirmed in grain samples
with different environmentally induced dormancy in grain from the 2005/06 season.
2.7 Field methods A photo outlining the imposition of water stress and temperature treatments used in the series of experiments is presented in Appendix D, and an overview of the treatments, split plots and harvests of the same experimental plots is also included.

29
3 Chapter 3
Embryo abscisic acid concentration during imbibition of intact grain is associated with grain dormancy in field grown wheat (Triticum aestivum L.) These experiments have been prepared as a thesis chapter only at this
stage, but a manuscript from it will be submitted to Australian Journal
of Agricultural Research.

30
Embryo abscisic acid concentration during imbibition of intact grain is associated with grain dormancy in field grown wheat (Triticum aestivum L.) Biddulph, Thomas B1,*., Plummer, Julie A1., Setter, Tim L2., Gubler, Frank3., Poole,
Andrew T3., and Mares, Daryl J4. 1. Plant Biology, MO84, Faculty of Natural and Agricultural Science, University of Western Australia, 35 Stirling
Highway, Crawley, WA, 6009 Australia.
2. Crop Improvement, Department of Agriculture and Food WA, 3 Baron-Hay Court, South Perth, WA, 6151 Australia.
3. Plant Industry, CSIRO, Black Mountain, Canberra, ACT, 2601 Australia.
4. School of Agriculture, Food and Wine, University of Adelaide, Waite Campus, Glen Osmond, SA, 5064 Australia.
* Author for correspondence, address line 2, Email: [email protected]
3.1 Abstract
Physiological dormancy is common in cereals but the mechanism is not well understood,
yet dormancy is the primary trait used to protect wheat crops from germination prior to
harvest (preharvest sprouting). Dormant (DM 2001, DH 22) and non-dormant (Cunderdin)
wheat genotypes were grown in the field and grain samples collected at maturity.
Dormancy was assayed along with concentrations of endogenous embryo ABA in grain
with varying levels of dormancy from different genotypes or physiological stages of
maturity; mature, harvest ripe (same as preharvest sprouting phenotyping) and after-ripened
grain. DM 2001 and DH 22 exhibited strong dormancy (Germination Index, GI generally <
0.20) at maturity whereas Cunderdin was non-dormant (GI > 0.70). Over seasons 2003/04
and 2004, despite being grown under a range of stressful conditions which gave a range of
dormancy levels for each genotype, there were no differences in embryonic ABA content or
concentration at maturity that would explain the differences in dormancy phenotype. There
were also no differences in embryonic ABA concentrations between genotypes at maturity,
harvest-ripeness or following after-ripening, despite the gradual loss of dormancy over this
time period. However, during imbibition from 12 to 72 h, dormant harvest-ripe grain of DH
22 maintained a two-fold higher concentration of ABA in the embryo than non-dormant
after-ripened grain. These observations are consistent with an important role for ABA in
dormancy of wheat grains. The capacity of intact wheat grains to maintain dormancy
appears to be related, at least in part, to the concentration of ABA that is maintained in the

31
embryo during the early stages of imbibition, but not the ABA concentration prior to
imbibition.
Key words: Physiological dormancy, endogenous embryo ABA, germination index,
harvest-ripe, after-ripening, seed dormancy, preharvest sprouting.
3.2 Introduction
Preharvest sprouting (germination of the grain in the ear prior to harvest) is a major defect
which results in downgrading of grain from milling to feed grades in all major wheat
producing countries (Derera 1982). Dormancy (Finch-Savage and Leubner-Metzger 2006)
is the main mechanism targeted by breeders (DePauw and McCraig 1991; Hucl 1995;
Trethowan 1995; Xiao et al. 2002) and physiologists in an effort to improve crop tolerance
to pre-harvest sprouting. Under controlled conditions abscisic acid (ABA) is important for
normal embryo development and appears to be involved in the establishment and
maintenance of seed dormancy in cereals (Benech-Arnold et al. 2006; Chono et al. 2006;
Jacobsen et al. 2002; Millar et al. 2006; Suzuki et al. 2000). A better understanding of this
mechanism of dormancy is an important step in improving preharvest sprouting tolerance in
wheat.
Endogenous ABA concentrations during late seed maturation are strongly linked to the
normal maturation pathway and inhibition of precocious germination or vivipary before
grain maturation under controlled conditions (Finkelstein et al. 2002; King 1989; Ross
1992). Mutants of Arabidopsis (Karssen et al. 1983) and maize (White et al. 2000) with
low ABA, or reduced ABA sensitivity often lack seed dormancy and exhibit precocious
germination before grain maturation. Application of ABA during imbibition inhibits wheat
germination while addition of fluridone, an ABA biosynthesis inhibitor reduces dormancy
(Garello et al. 1997). Under optimal growing conditions with abundant water and ideal
temperatures, 15/ 25°C, there are usually two peaks of endogenous ABA during grain
filling in wheat (King 1976). The first peak is contributed by the plant tissues of the grain
and occurs at around 25 days post anthesis (dpa), whereas the second peak is attributed to
the embryo and appears 2 weeks before maturity (typically at 0.9-2.5 µg g-1 DW),
coinciding with maximum dry matter and fresh weight of the grain (King 1976; Koshkin

32
and Tararina 1990). Following the second peak, the ABA concentration decreases rapidly
to around 0.2-0.4 µg g-1 DW at maturity, (King 1976). Under these controlled environments
(Walker-Simmons and Sesing 1990) or with optimum field conditions (Suzuki et al. 2000)
work has shown an association between these peaks and the level of dormancy and embryo
sensitivity to ABA.
Under optimum growth conditions in a number of plants, seed dormancy has also been
shown to correlate with changes in ABA content during imbibition. During imbibition
dormant barley grains retain a higher ABA content in embryos compared to non-dormant
seed (Benech-Arnold et al. 2006; Jacobsen et al. 2002; Millar et al. 2006). Seeds of other
plants such as Arabidopsis (Ali-Rachedi et al. 2004; Millar et al. 2006), tobacco (Grappin
et al. 2004), pine (Feurtado et al. 2004) and preliminary work with isolated wheat embryos
(Ried and Walker-Simmons 1990; Walker-Simmons and Sesing 1990) exhibit a similar
phenomenon. In wheat from the field under growth optimum conditions, with imbibition of
isolated embryos, dormant embryos are able to maintain a higher ABA concentration than
non-dormant embryos for the first 12 hours. However, in the field preharvest sprouting
occurs in intact grain, after harvest-ripeness and the germination of isolated embryos at
maturity is different to that of whole grains at harvest ripeness, which is when preharvest
sprouting is the problem. Furthermore other work on wheat with intact grains from the field
did not find significant differences in ABA of whole grains during imbibition (Tavakkol
Ahshari and Hucl 2001). However, a limitation of this work is that whole seed ABA was
measured not embryo ABA concentration during imbibition. Hence further work is required
in the field to determine if the elevation of embryo ABA during imbibition of isolated
embryos also occurs in the embryos from intact grains during imbibition.
Environmental conditions are rarely ideal in the field, particularly in the Australian wheat
belt which is often characterized by terminal moisture stress and high (>30°C)
temperatures. Temperature fluctuations are also large and days to maturity can be reduced
by temperature and moisture stress during grain filling (Stone and Nicolas 1995). As plant
ABA concentrations are associated with the plant response to stress, it is likely that plants
grown under a high stress environment may result in more dormant grain than plants grown
under optimum field conditions. Previous observations from Mares (1993) with the long

33
season genotype, Kleiber, where terminal moisture stress in the field, was associated with
elevated dormancy, support this hypothesis. Similarly, field observations from the
Department of Agriculture Western Australia in which material grown in the central wheat
belt (Merredin) which experiences terminal moisture stress was compared with the coastal
wheat belt (Esperance) which experiences more rainfall and little moisture stress (Setter et
al. 2001) are consistent with this hypothesis. By contrast in pot based experiments, Auld
and Paulsen (2003) found that moisture stress reduced dormancy under similar high
temperature regimes.
There are several possible causes for the observed differences in dormancy in plants
exposed to moisture stress between pot and field grown plants. Experiments with pots are
problematic and often have unrecognised artifacts (Passioura 2006). For example there may
be a background response concentration of ABA generated by roots of plants encountering
an impenetrable layer such as a pot wall. When cereal roots hit compacted soil, the xylem
sap ABA concentrations increase 4-fold, possibly increasing flag leaf ABA (Westgate et al.
1996). Imposition of moisture stress and field capacity in pots is restricted by the lack of
soil pores in potted soil or the abundance of large pores in potting mix (Passioura 2006).
One indication of the potential problems associated with moisture stress based pot
experiments is the lack of yield reduction in moisture stressed pots compared to �field
capacity� controls (Auld and Paulsen 2003). Field work is required, where rooting depth is
unrestricted to ensure well watered controls have limited plant ABA at the first peak of
embryo ABA concentration, and moisture stressed treatments are realistic and mimic
conditions that normally occur in the field and the level of ABA accumulation and hence
dormancy that is representative of the field environment. There is little if any work on
plants grown under stressful conditions of higher temperature and limited water supply
during grain filling in the field. Under conditions where the plant is stressed there may be
differences in embryo ABA at maturity associated with these differences in dormancy.
In this investigation the role of ABA on dormancy of intact grains of field grown wheat is
explored under contrasting environmental conditions. Concentration of ABA in the
embryos of grain from genotypes with different dormancy, grown under varying
environmental conditions and sampled at maturity, harvest-ripeness and following after-
ripened were determined and compared with the dormancy phenotype at maturity.

34
Furthermore the concentration of embryo ABA during imbibition of intact grains was also
examined.
3.3 Materials and methods
3.3.1 Plant material
Hard white spring wheat (Triticum aestivum L.) genotypes, strongly dormant breeding
genotypes DM 2001 (embryo and seed coat ES; derived from AUS 1408) and DH 22,
(embryo and seed coat ES; Cascades x AUS 1408 doubled haploid) and a non-dormant
Western Australian genotype, Cunderdin (non dormant es), were sown at Katanning and
Esperance Western Australia on the 29th May 2003 and 24th May 2004. These genotypes
were selected on the basis of presence or absence of the AUS 1408 allele at the 4A
dormancy QTL (Mares and Mrva 2001; Mares et al. 2005), consistent dormancy phenotype
over several seasons, tolerance to black point and similar maturity.
The Katanning site (Lat 33°42� S, Long 117°36� E, elevation 320 m) consisted of a grey to
brown duplex soil, while the Esperance site, (Lat 33°60� S, Long 121°78� E, elevation 143
m) was yellow duplex sand. Seed was treated before sowing with Jockey® (167 g L-1
fluquinconazole), at 4.5 mL kg-1 to prevent root and foliar disease during early growth.
Plots, 80 cm long single rows, were sown with a Wintersteiger horticultural single plot
seeder at a depth of 3 cm, to achieve a plant density of 250 plants m-2. Rows were 21 cm
apart, leaving a 20 cm space between plots and 80 cm between watering treatments.
3.3.2 Agronomic management
A knock down herbicide of 500 mL ha-1 Roundup (360g L-1 glyphosate) was applied to
control weeds before sowing. The soil was cultivated with 21 cm row spacing to place
fertiliser (Agstar Plus�; CSBP, Perth) at 150 kg ha-1 at a depth of 5 cm. This equates to
23.0, 19.4, 17.1, 0.3, 0.15, 0.03 kg ha-1 N, P, S, Cu, Zn and Mo respectively. The fertiliser
was treated with Impact-In-Furrow (flutriafol) at 2.7 mL kg-1 to prevent leaf and root
disease during early growth. Fungicide and pesticide applications of 290 mL ha-1 Folicur
430EW (tebuconizole, 250 g L-1) and 195 mL ha-1 Fastac (alpha-cypermethrin, 250g L-
1) or equivalent fungicides, were applied at tillering, booting and full flag leaf emergence as

35
a preventative measure against foliar or root diseases and to control aphids. No visible
symptoms of foliar and root disease or insect damage were evident.
3.3.3 Temperature, moisture stress and irrigation treatments
Between sowing and anthesis there was 272 mm and 262 mm of rain in 2003/04 and 2004
respectively. Spikes, ~ 50 per plot, were tagged every alternate day during anthesis (24th
September to 2nd October 2003 and 17th to 23rd September 2004). The trial was enclosed in
a rainout tunnel, 20 x 9 x 3 m high centre covered with a UV stable translucent cover
(Solarweave Natural , diffuse light transmission of 82 % Jaylon, Perth) in order to exclude
rain and moisture stress and irrigated treatments began at anthesis. Plants in the moisture
stress treatments had to rely on residual soil moisture from anthesis to maturity. For the
irrigation treatment, water was supplied without wetting the spikes using ground water fed
through T-Tape (International Model 515-20-250), at a rate that was equivalent to 116 and
130 mm rainfall spread over five weeks and averaged 60 ± 4 % of actual Epan at Esperance
in 2004/05. Irrigation was carried out weekly until the moisture stressed plots reached
maturity (Zadoks Z91 approximately 20-30% grain moisture content, on a dry weight basis
(Zadoks et al. 1974), thumb nail indent held).
At Katanning in 2003/04 at anthesis, low and high temperature treatments, were
superimposed on the trial with an air conditioning unit (Fujitsu 3.0 hp, Japan, Tokyo) that
was placed inside a small tent (Hyticlear co-extruded greenhouse film, Jaylon, Perth WA)
within the larger rainout tunnel. The air-conditioned plants inside the tent were the low
temperature treatment and non-air conditioned plots outside the tent were the high
temperature treatment.
At maturity 3 replicates of 10 spikes in 2003/04 or 200 main stem ears / spikes in 2004/05
that had all reached anthesis over the same 3 day window were harvested. In 2004/05 more
grain was collected to enable the comparison of samples with different physiological
stages, maturity, harvest ripe, after-ripened and then during the imbibition of harvest ripe
and after-ripened samples. The top and bottom florets were discarded and the primary and
secondary florets gently thrashed by hand to ensure the grain and, in particular, the seed
coat was not damaged. Within 2 h of threshing 10 embryos from each replicate were

36
isolated with forceps, placed in pre-weighed 2.5 mL cryovials and frozen on dry ice before
storage at -80 °C. Embryo ABA content, was later determined on this grain. In 2004/05, grain dormancy was determined within two hours of harvest on mature grain
(Walker-Simmons 1987) using 30 grains or embryo half grains incubated at 20 °C for 7 d
in 90 mm Petri dishes with filter paper (3 x 70 mm, No. 2 Advantec, Toyo Roshi Kaisha,
Ltd. Japan) moistened with either 6 mL DI water or 6 mL 20 µM ABA ((±)-cis,trans-
Abscisic Acid, Cat No. A-1049, Sigma-Aldrich Co., St Louis USA). Germinated grains or
embryo half-grains were counted daily. Germination was defined as rupture of the pericarp
overlying the embryo in both grains and embryo half-grains. The germination index for
grains (GIseed) and embryo half-grains (GIembryo) was calculated using Equation 1 (Walker-
Simmons 1987).
(7 x n1 + 6 x n2 + 5 x n3 + 4 x n4 + 3 x n5 + 2 x n6 + 1 x n7) / (7 x total grains) (1)
where n1, n2, � n7 are the number of grains or embryos that germinated on the first, second
and subsequent days until the seventh day, respectively. Seed viability of grain that failed to
germinate in 7 d, was assessed by incubating the grain at 4 °C for a further 3 d and then for
a further 3 d at 20 °C and the total number of grains adjusted if necessary. The maximum
GI representing non-dormant grains is 1.0 and the minimum is 0 (Walker-Simmons 1987).
The remainder of the harvested grain was dried at room temperature (23 ± 5 °C) for 5 d to
reach a moisture content of < 12 %, equivalent to harvest-ripeness, before storage at -20 °C
to preserve dormancy (Mares 1983b).
In 2003/04, grain dormancy and ABA responsiveness was determined on harvest-ripe
samples only, with grain harvested from the field one week after maturity.
In 2004/05, after 14 weeks storage at -20 °C, grain samples were divided into two groups.
Half were returned to -20 °C immediately to maintain the level of dormancy present at
harvest-ripeness and the other half were after-ripened at 37 ± 2 °C. During this after-
ripening treatment dormancy was monitored weekly and when the GI in DM 2001 and DH
22 had increased from 0.2 to >0.70, after 6 weeks, the grain was classified as after-ripened
and returned to -20 °C to preserve the dormancy status. Responsiveness of embryos to

37
ABA was not carried out on harvest-ripe or after-ripened grain in 2004/05 due to
insufficient grain.
The embryo ABA concentration for moisture stressed and irrigated samples in 2004/05 was
determined on mature, harvest-ripe and after-ripened grain (3 replicates). For irrigated DH
22 and Cunderdin, embryo ABA was also determined after 0, 6, 12, 18, 24, 36, 48 and 72 h
imbibition of harvest-ripe and after-ripened grain. Grain was placed crease side down (3
reps of 30 grains) in each of 8 (one for each sampling time) 90 mm Petri dishes fitted with
filter paper as above and moistened with 6 mL DI water. At each sampling time the
embryos and attached scutella were excised, from 10 out of the 30 grains, removing as
much of the adhering endosperm, pericarp and seed coat tissue as possible without
physically damaging the embryo. Isolated embryos were then frozen on dry ice and stored
at -80 °C for later determination of ABA.
3.3.4 ABA extraction and derivitisation
The endogenous embryonic ABA concentration was determined according to the methods
of Green et al. (1997) and Jacobsen et al. (2002). Isolated embryos were transferred to 15
mL plastic centrifuge tubes (Falcon) containing 0.5 mL of chilled 80 v/v MeOH and the
sample ground with a Teflon tipped rod. An internal standard, 20 µL/10 ng [2H6] ABA (a
gift from Dr Sue Abrams, Plant Biotechnology Institute, National Research Council of
Canada, Saskatoon, Saskatchewan, Canada), was added, samples were made up to 10 mL
with 80 v/v MeOH and mixed on a spinning rotator/ inverter for ~16 h at 4 °C. Tubes were
then centrifuged at 4500 rpm for 5 minutes, the supernatant collected, the pellet rinsed with
5 mL 80 v/v MeOH, centrifuged again and the supernatant collected and pooled with the
first supernatant.
Extracts were reduced in volume to ≤3 mL by SpeedVak (Savant Instruments, Farmingdale,
NY) and the pH adjusted to 2.75 with 20 mM HCl. Samples were then partitioned with
water-saturated ethyl acetate (EtOAc) (3 x 10 mL) and the pooled EtOAc fraction
evaporated to dryness. Residues were dissolved in 200 µL 100 v/v MeOH + 1800 µL 0.4
v/v acetic acid, then the total 2 mL loaded onto pre-conditioned C18 Sep-Pak columns
(Waters, Massachusetts). Columns were washed twice with 2 mL 0.4 v/v acetic acid.

38
Material (ABA) retained on the column was eluted with 5 mL of 50 v/v MeOH in 0.4 v/v
acetic acid.
Following drying in vacuo by Speed Vac the samples were methylated by dissolving in 200
µL 100 v/v MeOH, 20 drops of ethereal diazomethane added and the mixture incubated for
15 min before re-drying. The residue was then transferred into 200 µL GC-MS vials using
50 µL pure EtOAc and rinsed twice with 50 µL, to a total of 150 µL, before reducing to
dryness.
3.3.5 ABA analysis by GC-MS-SIM
GC-MS-SIM conditions were similar to Jacobsen et al. (2002) and Green et al. (1997). GC-
SIM was performed using a gas chromatograph (model HP5890 series II, Hewlett-Packard)
coupled to a mass-selective detector (model 5971, Hewlett Packard). Helium was used as
the carrier gas, with an initial pressure pulse of 207 kPa, followed by a constant column
flow of 0.7 mL min-1 (initial head pressure of 80 kPa). Samples were dissolved in 5 µL
EtOAc and 1µL was injected in the splitless mode into a 25 m x 0.22 mm i.d. 0.25 µm film
thickness BPX-5 fused silica column (SGE, Ringwood) with the voltage boosted to a total
of 2576 volts. The initial oven temperature (60°C) was held for 1.5 min then increased to
200°C at 25°C min-1, then increased to 300°C at 5°C min-1 and held for 2 min. Three ions
distinctive fo ABA methyl ester (190,162,134) and three ions for [2H6] ABA methyl ester
(194, 166, 138) were monitored using selected ion monitoring over the period 11-12 min
when ABA is know to elute. Concentrations of endogenous ABA were calculated using the
peak area ratio of two major ions (190/194) and calibration curves constructed using
authentic standards.
3.3.6 Experimental Design and Statistical Analysis
The 2003/04 field trial was grown as a spilt-split-plot design with the main plot factor
moisture stress the subplot factor temperature and the sub-subplot factor genotype and the
block rep; genotype was randomised within temperature. The main effects of moisture
stress, temperature or genotype and their interaction were determined for the GI and
embryonic ABA concentrations at harvest-ripeness by ANOVA in Genstat 7.1 (Lawes
Agricultural Trust, Rothamsted), for a spilt-split-plot design, n = 3. The 2004/05 field trial
was grown as a split-plot design with the main plot factor moisture stress, the subplot factor

39
genotype and the block rep; genotype was randomised within water supply. The main
effects of moisture stress or genotype and there interaction were determined on the GI and
embryonic ABA concentrations at maturity by ANOVA, for a split-plot design, n = 3.
Fischers Least Significant Differences (LSD0.05) were calculated for comparisons of means
between treatments and genotypes, where analysis by ANOVA found significant
differences (P<0.05).
The imbibition experiment was laid out in a split- spilt plot design with the main plot factor
after-ripening, sub-plot factor sampling time, the sub-sub plot factor genotype and the block
week. The main effects and interaction of after-ripening, sampling time and genotype were
determined for the GI by ANOVA. For embryo ABA the main effects of after-ripening,
sampling time and genotype were analysed by Regression Analysis in Genstat 7.1 (Lawes
Agricultural Trust, Rothamsted) with a Generalised Linear Model because not all samples
were evenly replicated. Approximately ¼ of one replicate of the imbibition samples were
not successfully analysed for ABA, either because of milling problems during sample
preparation or low peak areas which were a results of low ABA recoveries in some samples
or low GC-MS-SIM sensitivity.
3.4 Results
3.4.1 Influence of season and environment on dormancy
In 2003/04 harvest-ripe grain of DM 2001 was dormant (low GI) and Cunderdin was non-
dormant (high GI) (Fig. 1a). By contrast, when embryo half grains were germinated in
water there was usually no difference in GI between DM 2001 and Cunderdin (Fig. 1a
shaded bars). However, when ABA was included in the imbibition medium, the GI of DM
2001 embryo half grains was reduced to values similar to intact grains in water, but was
there was no reduction in the GI of Cunderdin embryos (Fig. 1a open bars). Moisture
stressed Cunderdin grain at high temperature had a lower GI than the corresponding
treatment at low temperature (Fig. 1a). In contrast the GI of irrigated DM 2001 grain was
increased by high temperature (Fig. 1a). Despite these quantitative changes in dormancy,
the relative ranking of genotypes was always consistent, DM 2001 was always more
dormant, i.e. had a lower GI, than Cunderdin.

40
Embr
yoFW
(µg)
b) Embryo FW
c) Embryo ABA concentration
d) ABA per embryo
Embr
yo A
BA C
once
ntra
tion
(ng
g-1FW
) AB
A p
er E
mbr
yo
(pg
embr
yo -1
)
a) Germination index
0.0
0.2
0.4
0.6
0.8
1.0
Ger
min
atio
n in
dex
0
1
2
3
4
050
100150
200250
300
0.0
0.1
0.2
0.3
low high low high low high low high
moisture stressed
irrigated irrigated
DM 2001ES
Cunderdines
moisture stressed
5
Fig. 1. At Katanning in 2003/04 (a) germination index at harvest ripeness of grain in water
(solid bars), embryo half grain in water (shaded bars) and embryo half grain with 20 µM
ABA (open bars). (b) embryo FW, (c) embryo ABA concentration (ng g-1 FW) and (d)
ABA per embryo, (pg embryo-1) in DM 2001 and Cunderdin grain sampled at harvest-
ripeness, under irrigated or moisture stress with low or high temperature during grain
filling. Means presented, n=3, vertical bars represent LSD0.05 for comparisons between
treatments and genotypes.

41
0.0
0.2
0.4
0.6
0.8
1.0
0
100
200
300
0
1
2
3
4
5
Embr
yoFW
(µg)
0.00
0.10
0.20
0.30
moisture stressed
irrigated irrigated irrigated
DM 2001ES
DH 22ES
Cunderdines
b) Embryo FW
c) Embryo ABA concentration
d) ABA per embryo
Embr
yo A
BA C
once
ntra
tion
(ng
g-1FW
) AB
A p
er E
mbr
yo
(ng
embr
yo -1
)
a) Germination index
Ger
min
atio
n in
dex
moisture stressed
moisture stressed
Fig. 2. At Esperance in 2004/05 (a) germination index at maturity of grain in water (solid
bars), embryo half grain in water (shaded bars) and embryo half grain with 20µM ABA
(open bars). (b) embryo FW, (c) embryo ABA concentration (ng g-1 FW) and (d) ABA per
embryo, (pg embryo-1) in DM 2001, DH 22 and Cunderdin under irrigation and moisture
stress. Means presented, n=3, vertical bars represent LSD0.05 for comparisons between
treatments and genotypes.

42
The embryo FW, 20 µg-1 embryo (Fig. 1b), and the endogenous embryo ABA
concentration at maturity, 125 ng g-1 FW (Fig. 1c) or 0.28 pg-1 embryo (Fig. 1d), was the
same in DM 2001 and Cunderdin across a range of environmental conditions during grain
filling. There were no difference in endogenous ABA content of embryos between DM
2001 and Cunderdin in 2003/04 that were associated with the differences in GI (Fig. 1a)
regardless of whether the ABA content was expressed on an embryo FW basis or per
embryo basis.
In 2004/05 at maturity DM 2001 and DH 22 were dormant (GI generally <0.20) and
Cunderdin was non-dormant (GI >0.70) (Fig. 2a). These results for DM 2001 and
Cunderdin at maturity at Esperance (Fig. 2a) were similar to dormant (GI) at harvest
ripeness at Katanning (Fig. 1a). At Esperance the GI of DM 2001, irrigated DH 22 and
Cunderdin embryo half grains incubated in water were similar, >0.70 (Fig. 2a), with the
exception of moisture stressed DH 22 embryos that were significantly lower (GI = 0.30).
When ABA was applied, the GI of embryos was reduced in DM 2001 and DH 22 but was
not in Cunderdin (open bars, Fig. 2a) and in each case to GI values similar to intact grain
imbibed with water.
Moisture stress did not effect the GI of intact grain of DM 2001, DH 22 or Cunderdin (solid
bars, Fig. 2a), but there were differences in embryo FW (Fig. 2b). Fresh weight of
moisture-stressed DM 2001 embryos was only one third that of embryos from irrigated
plants (Fig. 2b). This was associated with a higher endogenous embryo ABA concentration
of moisture stressed DM 2001, more than three times that of irrigated DM 2001 and the
other genotypes (Fig. 2c). However, when ABA concentrations were expressed on an
embryo basis (Fig. 2d) there was no difference in ABA concentrations between watering
treatments in DM 2001, or the other genotypes except irrigated Cunderdin, which had only
a third the ABA per embryo. A similar picture was obtained when ABA amounts were
expressed on a DW basis, (data not shown) reflecting the fact that all embryos had similar
moisture contents at maturity (mean of 13 ± 1.8 %).

43
3.4.2 ABA in mature, harvest-ripe and after-ripened grain
Dormancy gradually declined, as indicated by the increase in GI from maturity (grain
moisture ~20 %) through harvest-ripeness (maturity plus one week) to fully after-ripe
(maturity plus six weeks) grain that was non-dormant (Fig. 3a). Changes were less marked
in Cunderdin but nevertheless there was an increase in GI from maturity to after-ripened
grain (Fig. 3a). The GI of both DM2001 and DH22 of the after-ripened grain are not
significantly different from one another but are still significantly less than that of
Cunderdin (Fig 3a).
The differences in GI between mature, harvest-ripe and after-ripened grain were not
consistently reflected in differences in endogenous embryo ABA concentrations (Fig. 3a &
Fig. 3b). All genotypes and stages of development had similar ABA concentrations of
approximately 100 ng g-1 FW, except for harvest-ripe Cunderdin, which was three fold
higher (Fig. 3b).
3.4.3 ABA concentration during imbibition, DH 22 and Cunderdin
During imbibition the changes in endogenous embryo ABA concentration differed between
harvest ripe and after-ripened grain of DH 22, whilst the initial embryo ABA concentration
(before imbibition) was similar in harvest-ripe and after-ripened grains at 100 ng g-1FW
(Fig. 4a). During the first 12 h of imbibition the ABA concentration fell to 20 ng g-1FW in
after-ripe grain but only 40 ng g-1 FW in harvest-ripe grain. The germination of harvest-ripe
grain of DH 22 was slower than after-ripened grain, starting at 48 h and reaching only 32 %
by 72 h (Fig. 4a dashed lines,). From 12 to 72 h of imbibition the endogenous embryo ABA
concentration of harvest-ripe grain of DH 22 was double that of after-ripened grain
embryos except for a brief dip at 24 h.
The pattern in Cunderdin for both harvest-ripe and after-ripened was similar to after-
ripened DH 22 up to 12 h after imbibition. However, by 24 h the Cunderdin grain started to
germinate and had reached 80 % germination by 48 h (Fig. 4b dashed lines). Samples for
DM2001 were lost during milling of the embryos, and sufficient samples were not left to
enable a representative concentration of ABA to be presented. Only irrigated grain was
used for these imbibition studies as the moisture stress lead to a small size and quantity of
grain.

44
a) Germination index
b) Embryo ABA concentration
0.0
0.2
0.4
0.6
0.8
1.0
Ger
min
atio
n In
dex
0
100
200
300
DM 2001ES
DH 22ES
Cunderdines
Embr
yo A
BA C
once
ntra
tion
(ng
g-1FW
)
Fig. 3. At Esperance in 2004/05 (a) germination index and (b) embryo ABA concentration
(ng g-1 FW) of irrigated DM 2001, DH 22 and Cunderdin grain in water at maturity (solid
bars), harvest-ripe (shaded bars) or after-ripened (open bars). Means presented, n=3,
vertical bars represents LSD0.05 for comparisons between treatments and genotypes.

45
Time of imbibition (hours)
0
50
100
150
200
250
300
0 6 12 18 24 30 36 42 48 54 60 66 72
Embr
yo A
BA
conc
entra
tion
(ng
ABA
/ g F
W)
0
20
40
60
80
100
Ger
min
atio
n (%
)
Embr
yo A
BA
conc
entra
tion
(ng
ABA
/ g
FW)
0
50
100
150
0
20
40
60
80
100
Ger
min
atio
n (%
)
a) DH 22 ES
b) Cunderdin es
Time of imbibition (hours)
0
50
100
150
200
250
300
0 6 12 18 24 30 36 42 48 54 60 66 72
Embr
yo A
BA
conc
entra
tion
(ng
ABA
/ g F
W)
0
20
40
60
80
100
Ger
min
atio
n (%
)
Embr
yo A
BA
conc
entra
tion
(ng
ABA
/ g
FW)
0
50
100
150
0
20
40
60
80
100
Ger
min
atio
n (%
)
a) DH 22 ES
b) Cunderdin es
Fig. 4. At Esperance in 2004/05, embryo ABA concentration (ng g-1 FW, solid lines) in DH
22 (a) and Cunderdin (b) grain which was harvest-ripe (solid squares) or after-ripened
(solid circles) and the percentage of grain germinated (dashed lines) in harvest-ripe (open
squares) or after-ripened (open circles). Predicted mean ± SE for ABA from Generalised
Linear Model, n=3 for DH 22 or n= 1 to 2 for Cunderdin. Means ± SE for germination, n=
3.

46
3.5 Discussion
Moisture stress and temperature treatments during grain filling affected the dormancy
phenotype in 2003/04, but not in 2004/05. Moisture stress combined with consistent high
temperature and low humidity during grain filling was associated with an apparent induced
embryo sensitivity to ABA in Cunderdin, a non-dormant genotype, but no additional
dormancy in DM 2001, a dormant genotype. This was not however associated with a higher
embryo ABA concentration, nor was the reduction in dormancy from mature to harvest-ripe
to after-ripened grain. This confirms earlier work under optimum conditions that failed to
find differences in endogenous ABA at maturity between dormant and non-dormant
genotypes (Walker-Simmons 1987; Walker-Simmons and Sesing 1990), or harvest-ripe and
after-ripened grain (Ried and Walker-Simmons 1990; Suzuki et al. 2000; Tavakkol Ahshari
and Hucl 2001). However, there were two differences in endogenous embryo ABA
concentration, which require explanation.
One difference in ABA occurred in moisture stressed DM 2001 embryos, when expressed
on a FW basis (ng g-1 FW), at maturity. The ABA concentration was almost double that of
the other samples (Fig. 2c), however the embryos were around one third the size and when
expressed as a weight per embryo (pg embryo-1) there were no differences in ABA between
genotypes. Secondly in Cunderdin, the harvest-ripe grain had a higher endogenous embryo
ABA concentration than mature or after-ripened grain (Fig. 3b). A cyclic increase and then
decrease in embryo ABA concentration over eight weeks of after-ripening has been
previously reported in wheat (Tavakkol Ahshari and Hucl 2001). The significance of this is
not understood, however both dormant and non-dormant genotypes appear to react in the
same manner (Tavakkol Ahshari and Hucl 2001). Whilst it seems clear that ABA
concentration in ripening or ripe grain per se in optimum or stressed environments cannot
explain the observed differences in dormancy between genotypes or treatments, the
possibility remains that differences in embryo ABA concentration during imbibition or the
embryo�s sensitivity to the ABA that is present may be important.
Significant differences in endogenous embryo ABA concentrations were found during
imbibition of dormant harvest-ripe and non-dormant after-ripened intact grain of DH 22.
During imbibition, following the initial rapid loss of ABA, embryos of harvest-ripe DH 22

47
grain maintained an endogenous ABA concentration double that of embryos from after-
ripened DH 22 grain. This supports earlier investigations on imbibition of isolated
embryos, from plants grown under optimum conditions (Ried and Walker-Simmons 1990;
Suzuki et al. 2000). However the time scale is substantially shorter in imbibing isolated
embryos with germination by 8 and 18 h (Suzuki et al. 2000) compared with the 24 and 48
h in non-dormant and dormant grain in this study. The more rapid germination of isolated
embryos compared to intact grain, (18 vs 48 h) may occur as a result of the endogenous
ABA concentration in imbibed dormant isolated embryos falling more quickly than in the
embryos from intact dormant grain observed in this investigation.
The seed coat of intact grain of DH 22 may have increased the capacity of dormant grain to
maintain higher concentrations of endogenous ABA compared to work with isolated
embryos (Ried and Walker-Simmons 1990; Suzuki et al. 2000). Benech-Arnold et al.
(2006) demonstrated that hull-imposed dormancy in barley was due the glumellae
depriving the embryo of oxygen during imbibition inhibiting ABA degradation by ABA 8�-
hydroxylase which requires oxygen. A similar role of the seed coat in wheat is unlikely, as
previous studies have shown no relationship between oxygen consumption during
imbibition and dormancy (Miyamoto et al. 1961). However, the fact remains that the seed
coat did have an effect, possibility by enhancing the embryo�s ability to maintain a higher
endogenous ABA concentration during imbibition in intact dormant grain. After-ripening
or damaging the seed coat, somehow reduces dormancy, possibly by reducing the ability of
the embryo to maintain a high ABA concentration. The dormancy in wheat previously
described as embryo sensitivity displayed by DH 22, may in fact be related to the ability of
the grain to maintain a higher embryo ABA concentration during imbibition and the seed
coat may have an additive effect on this. Further work is required to confirm the elevation
of ABA and the additive effect of the seed coat in a wider range of genotypes,
environmental conditions and at different stages throughout development.
The data presented here for intact dormant grain is consistent with the hypothesis that the
ability to maintain ABA at a higher concentration results in dormant grain. However,
applied ABA had little or no effect on germination of harvest-ripe embryo half-grains of
non-dormant Cunderdin, yet it could be assumed that there was sufficient exogenous ABA
to counter losses due to degradation or diffusion as in the case of embryo half-grains of DH

48
22. Similar results are also found in barley (Benech-Arnold et al. 1999; Wang et al. 1995).
A combination of high embryo ABA concentration and embryo sensitivity to ABA appears
to be required for dormancy. Further work needs to determine if ABA sensitivity is also
required in combination with a high ABA concentration to achieve a dormant phenotype
Once germination started in after-ripened DH 22 grain the endogenous embryo ABA
concentration remained the same for the next 24 hours. This contrasts with Cunderdin and
other work which has demonstrated a several-fold increase in ABA associated with the
transition from germination to post-germination growth in wheat (Tavakkol Ahshari and
Hucl 2001), barley (Yamada 1984), rice (Qin 1990), chick-peas (Iglesias and Babiano
1997) and pine (Feurtado et al. 2004). However, similar results have also been reported for
wheat (Ried and Walker-Simmons 1990; Suzuki et al. 2000), barley (Benech-Arnold et al.
2006; Jacobsen et al. 2002) and Arabidopsis (Ali-Rachedi et al. 2004). The reason and
significance of these inconsistent observations remains unclear and is an area, which
requires further clarification.
3.6 Acknowledgments
We thank the Grains Research and Development Corporation in support of TBB through a
GRDC Grains Research Scholarship GRS66 and CSIRO Plant Industry for a studentship,
the Department of Agriculture and Food Western Australia for running of field trials and
study leave awarded to TBB, the Ravensthorpe Agricultural Initiative Network for a grant
to TBB. Thank you also to Colin Norwood, Colin Boyd and the technical staff at Esperance
Downs Research Station for excellent advice and assistance with these trials.

49
3.7 References
Ali-Rachedi S, Bouinot D, Wagner M, Bonnet M, Sotta B, Grappin P and Jullien M (2004)
Changes in endogenous abscisic acid levels during dormancy release and maintenance of
mature seeds: studies with the Cape Verde islands ecotype, the dormant model of
Arabidopsis thaliana. Planta 219: 479-488.
Auld AS and Paulsen GM (2003) Effects of drought and high temperature during
maturation on preharvest sprouting tolerance of hard white winter wheat. Cereal Research
Communications 31, 169-176.
Benech-Arnold RL, Giallorenzi MC, Frank J and Rodriguez MV (1999) Termination of
hull-imposed dormancy in developing barley grains is correlated with changes in
embryonic ABA levels and sensitivity. Seed Science Research 9: 39-47.
Benech-Arnold RL, Gualano N, Leymarie J, Come D and Corbineau F (2006) Hypoxia
interferes with ABA metabolism and increases ABA sensitivity in embryos of dormant
barley grains. Journal of Experimental Botany 57: 1423-1430.
Chono M, Honda I, Shinoda S, Kushiro T, Kamiya Y, Nambara E, Kawakami N, Kaneko S
and Wantabe Y (2006) Field studies on the regulation of abscisic acid content and
germinability during grain development of barley: molecular and chemical analysis of pre-
harvest sprouting. Journal of Experimental Botany 57: 2421-2434.
Derera NF (1982) The harmful harvest rain: Farrer memorial oration, 1981. The Journal of the Australian Institute of Agricultural Science 1982, 67-75.
DePauw RM and McCraig TN (1991) Components of variation, heritabilities and
correlations for indices of sprouting tolerance and seed dormancy in Triticum spp.
Euphytica 52: 221-229.

50
Feurtado JA, Ambrose SJ, Cutler AJ, Ross ARS, Abrams SR and Kermode AR (2004)
Dormancy termination of western white pine (Pinus monticola Dougl. Ex D. Don) seeds is
associated with changes in abscisic acid metabolism. Planta 218: 630-639.
Finkelstein RR, Gampala SSL and Rock CR (2002) Abscisic acid signaling in seeds and
seedlings. The Plant Cell Supplement: s15-s45.
Finch-Savage WE and Leubner-Metzger G (2006) Seed dormancy and the control of
germination. New Phytologist 171: 501-523.
Garello G, Holappa LD, Abrams GD and Abrams SR (1997) ABA metabolites induce
group 3 LEA mRNA and inhibit germination in wheat. Physiologia Plantarum 100: 474-
480.
Grappin P, Bouinot D, Sotta B, Miginiac E and Jullien M (2004) Control of seed dormancy
in Pinus monticola: post-imbibition abscisic acid metabolism. Planta 218: 630-639.
Green LS, Faergestad EM, Poole AT and Chandler PM (1997) Grain development mutants
of barley. Plant Physiology 114: 203-212.
Hucl P (1995) Divergent selection for sprouting resistance in spring wheat. Plant Breeding
114: 199-204.
Iglesias RG and Babiano MJ (1997) Endogenous abscisic acid during the germination of
chick-pea seeds. Physiologia Plantarum 100: 500-504.
Jacobsen JV, Pearce DW, Poole AT, Pharis RP and Mander LN (2002) Abscisic acid,
phaseic acid and giberellin contents associated with dormancy and germination in barley.
Physiologia Plantarum 115: 428-441.

51
Karssen CM, Brinkhorst-van der Swan DLC, Breekland DE and Koornneef M (1983)
Induction of dormancy during seed development by endogenous abscisic acid: studies on
abscisic acid deficient genotypes of Arabidopsis thaliana (L.) Heynh. Planta 157: 158-165.
King RW (1976) Abscisic acid in developing wheat grains and its relationship to grain
growth and maturation. Planta 132: 43-51.
King RW (1989) Physiology of Sprouting Resistance. In 'Preharvest Field Sprouting in
Cereals' pp. 27-60. (CRC Press: Boca Raton, Florida)
Koshkin EI and Tararina VV (1990) Differences in source-sink ratios in wheat and their
relationship to grain yield and content of abscisic acid. Plant Physiological Biochemistry
28: 609-616.
Mares DJ (1983) Preservation of dormancy in freshly harvested wheat grain. Australian
Journal of Agricultural Research 34: 33-38.
Mares DJ (1993) Pre-harvest sprouting in wheat. I. influence of cultivar, rainfall and
temperature during grain ripening. Australian Journal of Agricultural Research 44, 1259-
1272.
Mares DJ and Mrva K (2001) Mapping quantitative trait loci associated with variation in
grain dormancy in Australian wheat. Australian Journal of Agricultural Research 52:
1257-1265.
Mares DJ, Mrva K, Cheong J, Williams K, Watson B, Storlie E, Sutherland M and Zou Y
(2005) A QTL located on chromosome 4A associated with dormancy in white- and red-
grained wheats of diverse origin. Theoretical and Applied Genetics 111: 1357-1364.
Millar AA, Jacobsen JV, Ross JJ, Helliwell CA, Poole AT, Scofield G, Reid JB and Gubler
F (2006) Seed dormancy and ABA metabolism in Arabidopsis and barley: the role of ABA
8'-hydroxylase. The Plant Journal 45: 942-954.

52
Miyamoto T, Tolbert NE, Everson EH (1961) Germination inhibitors related to dormancy in wheat seeds. Plant Physiology 36, 739-746.
Passioura JB (2006) The perils of pot experiments. Functional Plant Biology 33, 1075-
1079.
Qin ZZ (1990) Changes and endogenous abscisic acid levels in rice embryo and endosperm
and association with development and germination. Acta Bot. Sin. 32: 448-455.
Ried JL and Walker-Simmons M (1990) Synthesis of abscisic acid-responsive, heat -stable
proteins in embryonic axes of dormant wheat grain. Plant Physiology 93: 662-667.
Ross CW (1992) Hormones and Growth Regulators: Cytokinins, Ethylene, Abscisic Acid,
and Other Compounds. In 'Plant Physiology'. (Eds FB Salisbury and CW Ross) pp. 382-
406. (Wadsworth Publishing Company: Belmont, California)
Setter TL, Waters I, Mares DJ and Wilson R (2001) Sprouting tolerance of AGWEST
wheat breeding genotypes: Combined and comparative measurements from Crop
Improvement Institute, AGWEST (AGWEST) and I.A. Watson Wheat Research Centre,
PBI, Narrabri, NSW (PBI). Pages. 1-18. (Agriculture Western Australia: Perth, Western
Australia)
Stone PJ and Nicolas ME (1995) Effect of timing of heat stress during grain filling on two
wheat varieties differing in heat tolerance. I. Grain growth. Australian Journal of Plant
Physiology 22, 927-934.
Suzuki T, Matsuura T, Kawakami N and Noda K (2000) Accumulation and leakage of
abscisic acid during embryo development and seed dormancy in wheat. Plant Growth
Regulation 30: 253-260.
Tavakkol Ahshari R and Hucl P (2001) Temporal changes in abscisic acid concentration in
dormant and non-dormant seed of wheat (Triticum spp.) genotypes. Journal of Agricultural
Science and Technology 3: 9-18.

53
Trethowan RM (1995) Evaluation and selection of Bread Wheat (Triticum aestivum L.) for
Preharvest Sprouting Tolerance. Australian Journal of Agricultural Research 46: 463-474.
Walker-Simmons M (1987). ABA levels and sensitivity in developing wheat embryos of
sprouting resistant and susceptible cultivars. Plant Physiology 84: 61-66.
Walker-Simmons M and Sesing J (1990). Temperature effects on embryonic abscisic acid
levels during development of wheat grain dormancy. Journal of Plant Growth Regulation
9: 51-56.
Wang M, Heimovaara-Dijkstra S and Van Duijn B (1995) Modulation of germination of
embryos isolated from dormant and nondormant barley grains by manipulation of
endogenous abscisic acid. Planta 195, 586-592.
Westgate ME, Passioura JB and Munns R (1996) Water status and ABA content of floral
organs in drought-stressed wheat. Australian Journal of Plant Physiology 23, 763-772.
White CN, Proebsting WM, Hedden P and Rivin CJ (2000) Gibberellins and seed
development in maize. I. Evidence that giberellin/ abscisic acid balance governs
germination versus maturation pathways. Plant Physiology 122: 1081-1088.
Xiao S-H, Zhang X-Y, Yan C-S and Lin H (2002) Germplasm improvement for preharvest
sprouting resistance in Chinese white-grained wheat: An overview of the current strategy.
Euphytica 126: 35-38.
Yamada K (1984) Changes in the level of endogenous abscisic acid in barley during
germination and use of abscisic acid in malting. J. Am. Soc. Brew. Chem. 42: 79-84.
Zadoks JC, Chang TT and Konzak CF (1974) A decimal code for the growth stages of
cereals. Weed Research 14: 415-421.

54
4 Chapter 4
Influence of high temperature and terminal moisture stress on dormancy in wheat (Triticum aestivum L.)
This manuscript was accepted for publication in Field Crops Research
in June, 2007.
Biddulph, T.B., Plummer, J.A., Setter, T.L. Mares, D.J., 2007. Influence of high
temperature and terminal moisture stress on dormancy in wheat (Triticum aestivum
L.). Field Crop. Res. Doi:j.fcr.2007.05.005.

55

56

57

58

59

60

61

62

63

64

65

66

67

68

69
5

70
Chapter 5
Seasonal conditions influence dormancy and preharvest sprouting tolerance in wheat (Triticum aestivum L.) in the field
This manuscript was re-submitted following acceptance for publications
with major corrections to Field Crops Research July, 2006.

71
Seasonal conditions influence dormancy and preharvest sprouting tolerance of wheat (Triticum aestivum L.) in the field T.B. Biddulpha,b*, J.A. Plummera, T.L. Setterb, D.J. Mares c, a, Plant Biology, MO84, Faculty of Natural and Agricultural Sciences, University of Western Australia, 35 Stirling Highway, Crawley, WA, 6009 Australia. b Crop Improvement, Department of Agriculture and Food Western Australia, 3 Baron-Hay Court, South Perth, WA, 6151 Australia. c School of Agriculture and Wine, University of Adelaide, WAITE Campus, Glen Osmond, SA, 5064 Australia. * Corresponding author at Crop Improvement, Department of Agriculture and Food Western Australia, 3 Baron-Hay Court, South Perth, WA, 6151 Australia. Tel.: +61 8 9368 3333. Fax. +61 8 9368 2958 Email: [email protected].
5.1 Abstract Preharvest sprouting occurs following rainfall after maturity and reduces grain quality and
value. Dormancy at maturity is a trait frequently used by wheat breeders to improve
tolerance to sprouting. To determine the environmental influence on the predicative
relationship between dormancy at maturity and improved preharvest sprouting tolerance,
dormancy (germination index) at maturity and grain quality (falling number), after rainfall,
was measured over three seasons. Based on the results it was possible to draw three main
conclusions. Firstly, genotypes with strong dormancy (germination index <0.20) which
have the embryo and seed coat component of dormancy maintained a falling number >300
s at all sites and seasons for the two month period after maturity despite receiving up to 122
mm of rain. Adequate preharvest sprouting tolerance also occurred in dormant genotypes,
with just the embryo component; in all but the most sever conditions. Secondly, though the
effect of environment and interaction of genotype and environment was significant, the G
by E interaction did not account for a large proportion of the variation (<6%) on sprouting
tolerance (measured by falling number after rainfall) or change the relative rankings of
preharvest sprouting tolerance. Finally, other defects associated with rainfall during grain
filling such as black point and fungal staining may slightly reduce dormancy estimates and
preharvest sprouting tolerance, and for this reason need to be improved in parallel with
preharvest sprouting tolerance. In conclusion, dormancy provides a reliable source of
preharvest sprouting tolerance in the field.
Keywords: Preharvest sprouting, dormancy, germination index, falling number, fungal staining, black point.

72
5.2 1. Introduction
Preharvest sprouting refers to germination of grain in the ear prior to harvest. Preharvest
sprouting is a problem in all major white wheat producing regions of the world including
Australia, South Africa, Canada, Central Asia and Europe when rainfall leads to high
moisture conditions before the crop is harvested. Sprouting is a problem, in white wheat
producing regions such as Western Australia and globally because locally adapted, high
yielding genotypes lack sprouting tolerance and there are yield penalties associated with
growing sprouting tolerant genotypes. Preharvest sprouting leads to a reduction in both
grain yield and quality. Yield losses from 10 to 50 % occur in years with exceptional
damage in wheat (Stoy, 1983). Wheat quality losses result in downgrading at receival. In
2003/04 farmers in southern regions of the Australian wheat belt lost 20 % of the value of
their grain because of downgrading due to sprouting when the quality (falling number) of
delivered grain fell below the Australian Wheat Board minimum for premium grades
(Australian-Wheat-Board, 2003-2004). Falling number is a measure of suitability of grain
for milling and is determined by the Hagberg falling number method (Hagberg, 1960;
Hagberg, 1961). The minimum receivable standard for falling number in Australia is
currently set at 300 s for premium grades (Australian-Wheat-Board, 2003-2004). Grain
receival points world wide have similar grading systems based on grain quality, of which
falling number is one of a suite of standards that payment grades are based on. Preharvest
sprouting resulting in a falling number lower than 300 s occurs in approximately 1 in 4
years in high risk areas of the Western Australian wheat belt (personal observations), and
hence results in lower payment grades in those years.
Dormancy is typically measured by either a germination index (ranging from 0.0 to 1.0)
(Reddy et al., 1985) or germination resistance index (ranging from 0 to 50) calculated on
hand threshed grains imbibed on filter paper. The germination index is a weighted index
which gives maximum weight to grains which germinate early, and progressively less
weight to grains which germinate later (Reddy et al., 1985) while the germination
resistance index measures the relative rate of germination, by estimating the time to 50 %
germination (Gordon, 1971). There are several papers which detail the relative merits of
different assay methods, however most agree there is a significant correlation between seed
dormancy measured on grain from threshed heads and assays for sprouting tolerance based

73
on intact spikes (DePauw and McCaig 1991; DePauw et al., 1989; DePauw et al., 1993).
Initial work comparing methods to select preharvest sprouting tolerant genotypes
determined a correlation (r = -0.56) between germination of hand threshed seed and falling
number (Trethowan, 1995; Trethowan et al., 1996). Practically the main difference between
the germination index and resistance index is that in genotypes which reach the same total
germination percentage at 7 days the germination index can differentiate better between
genotypes which germinate rapidly during the first three days compared to genotypes which
germinate at a slower rate. This ability may give an important differentiation in the field as
given a relatively small rainfall event, the slower germination rate, will result in a lower
proportion of germinated grain, and hence greater sprouting tolerance. As a result
dormancy, measured by germination index, remains one of the main mechanism targeted by
breeders (Trethowan, 1995; Xiao et al., 2002) and physiologists (Gubler et al., 2005) in
their efforts to improve tolerance of cereals to pre-harvest sprouting.
However in this initial research no genotype-by-year interactions occurred as the seasons
were similar (Trethowan, 1995) and in a subsequent field evaluation only one site and
season was used (Trethowan et al., 1996). Dormancy however can be influenced by the
different environmental conditions experienced at different sites, seasons or in genotypes
with different maturity (Auld and Paulsen, 2003; Biddulph et al., 2005; Fenner, 1991;
Hagemann and Ciha, 1987; King, 1993; Lunn et al., 2002; Mares, 1993; Nielsen et al.,
1984; Reddy et al., 1985; Strand, 1989), particularly in partially dormant genotypes
(Biddulph et al., 2007). Other traits, such as ear characteristics, are also associated with
some level of preharvest sprouting tolerance but may not be influenced by the environment.
Ear characteristics, such as awnless ears, surface waxes, glaucous lemma and head nodding
angle (King, 1984; King and Richards, 1984), are not evaluated with germination tests, and
the level of additional protection they may offer has not been quantified but is likely to be
important (King and Wettstein-Knowles, 2000).
Given that dormancy changes with environment, and that the timing, intensity and duration
of rain and grain wetting associated with preharvest sprouting also changes with the
environment, there are several questions which arise from a breeding perspective. Firstly,
does this environmentally induced dormancy lead to differences in preharvest sprouting
tolerance in the field with natural rainfall? Furthermore, what level of dormancy is required

74
for environments where the probability of preharvest sprouting occurring is low, as in these
environments the level of dormancy is potentially higher (Biddulph et al., 2007). For
example the probability of at least 20 mm of rainfall occurring in the two months post
maturity is 3 out of 4 seasons in Esperance and 2 out of 4 seasons in Katanning, Western
Australia (Bureau-of-Meteorology, 1956-2006), however, the experience of farmers is that
preharvest sprouting occurs in 1 out of 4 seasons in Esperance and 1 out of 10 seasons in
Katanning. Finally, given that grain and ear characteristics which influence water uptake
are unlikely to be influenced by the environmental conditions, what level of protection
compared to dormancy do these characteristics give against preharvest rainfall?
These experiments were run with the aim of determining the level of dormancy required to
give adequate protection from preharvest sprouting under seasonal conditions which
induced different levels of dormancy and evaluate the relative contribution of the main
mechanisms (embryo and seed coat based seed dormancy) of sprouting tolerance compared
to current commercial genotypes.
5.3 2. Materials and methods
A set of nine hard white spring wheat (Triticum aestivum L.) genotypes with different
levels of dormancy and ear characteristics were sown at Katanning (Lat 33°40� S, Long
117°36� E, elevation 320 m) and Esperance (Lat 33°36� S, Long 121°47� E, elevation 143
m) Western Australia over three successive seasons in 2003/04, 2004/05 and 2005/06. In
addition to this main trial in 2003/04 three of the nine genotypes (Wylkatchem, Camm and
Janz) were also sown in Ravensthorpe (Lat 33°42� S, Long 119°41� E, elevation 150 m). At
Esperance in 2004/05 and 2005/06 the trial was run a moisture stress trial to try and
replicate different grain filling conditions, one with abundant water, and one with terminal
drought and in 2005/06, 30 commercial genotypes, 26 Western Australian breeding
genotypes, 10 sprouting tolerant check genotypes and two Canadian genotypes were sown
at Esperance as part of the Department of Agriculture and Food Western Australia�s
preharvest sprouting nursery.
5.3.1 2.1. Plant material
The original set of nine genotypes included the strongly dormant genotypes AUS1408, DM
2001(strongly dormant, embryo and seed coat dormancy, ES) and locally adapted, non-

75
dormant Western Australian commercial cultivars; EGA Eagle Rock (awnless), Camm,
Cascades, EGA Castle Rock, Janz, Cunderdin and Hartog. Further details of the parentage
of the genotypes sown are shown in Table 1. Cunderdin was excluded in 2003/04 and EGA
Castle Rock in 2004/05 due to sampling errors. In addition at Ravensthorpe in 2003/04
Wyalkatchem, Camm and Janz were sown. For the manipulated environment trials in
Esperance 2004/05 and 2005/06, AUS1408, DM2001, three doubled haploids (DH) from
Cascades/AUS1408, with different levels of grain dormancy: DH 22 (strongly dormant,
embryo and seed coat dormancy, ES), DH 56 (dormant, embryo dormancy only, Es) and
DH 45 (partially dormant, seed coat dormancy only, eS), Cascades and Cunderdin were
sown in addition to the original set of nine lines. The DH genotypes were selected on the
basis of presence or absence of the AUS1408 allele at the 4A dormancy QTL (Mares and
Mrva, 2001), consistent dormancy phenotype over several seasons, tolerance to black-point
and similar maturity to DM 2001.
5.3.2 2.2. Agronomic management
Trials were sown in Katanning, Western Australia on 29 May 2003, 28 May 2004 and 2
June 2005; in Esperance on 6 June 2003, 24 May 2004 and 31 May 2005; and in
Ravensthorpe on 2 June 2003. The Katanning site consisted of a grey to brown duplex soil,
the Esperance site was yellow duplex sand and the Ravensthorpe site was a yellow duplex
soil. Similar agronomic management was carried out for all trials; typically a knock down
herbicide of 500 mL ha-1 Roundup (360 g L-1 glyphosate) was applied to control weeds
before sowing. The soil was cultivated with 21 cm row spacing to place fertiliser (Agstar
Plus�; CSBP, Perth) at 150 kg ha-1 at a depth of 5 cm. This equates to 23.0, 19.4, 17.1,
0.3, 0.15, 0.03 kg ha-1 N, P, S, Cu, Zn and Mo respectively. The fertiliser was treated with
Impact-In-Furrow (flutriafol) at 2.7 mL kg-1 to prevent leaf and root disease during early
growth. Seed was also treated before sowing with Jockey® (167 g L-1 fluquinconazole) at
4.5 mL kg-1 to prevent root and foliar disease during early growth. Three replicated plots,
80 cm long single rows, were sown with a Wintersteiger horticultural single plot seeder at a
depth of 3 cm, to achieve a plant density of 250 plants m-2. Rows were 21 cm apart, leaving
a 20 cm space between plots and 80 cm between watering treatments. Fungicide and
pesticide applications of 290 mL ha-1 Folicur 430EW (tebuconizole, 250 g L-1) and 195
mL ha-1 Fastac (alpha-cypermethrin, 250g L-1), were applied at tillering, booting and full

76
flag leaf emergence as a preventative measure against foliar or root diseases and to control
aphids. No visible symptoms of disease or insect damage were evident.
Table 1. Details of pedigree and dormancy mechanisms, embryo component (present E,
absent, e) and seed coat component (present, S, absent, s) and unknown (?) of the different
genotypes used in this study.
Genotype Pedigree Seed coat
Dormancy mechanisms
Canadian Snowbird RL4137*6//Thatcher/Poso48/3/AC Domain white ? - awnless Kanata RL4137*6//Thatcher/Poso48/3/AC Domain white ? - awnless AUS1408 AUS1408 Land race from Transvaal region of S. Africa white ES DH 22 Cascades/AUS1408 white ES DM2001 Hartog/Vasco//AUS1408/Hartog white ES DH 56 Cascades/AUS1408 white Es CIYMMT 7HRWSN58 Asio/3/F6.74/Bunting//Siskin red ? Cascades QT7475 AUS1408/3*Janz/Cunningham white ? Calingiri Chino/Kulin//Reeves white es Cadoux Centrifen/Gamenya*2/3/Jacup white es Cascades Aroona*3//Tadorna/Inia66 white es EGA Castle Rock 3Ag/3*Cascades white es Binnu Arrino/Y89-4034(Eradu*4/VPM1) white es Spear types Clearfield STL Veranopolis/3*RAC177//3*Spear/3/Dagger. white es Spear Sabre/MEC-3b(RAC111)//Insignia white es Camm VPM1.5*Cook/4*Spear white es WAWHT 2730 synCPI133899/1308-26 white es Stiletto Veranopolis/3*RAC177/2/3*Spear/3/Dagger white es EGA Eagle Rock Sunelg/2*Blade white es - awnless Frame Molineux/3*Dagger white es WAWHT2884 Sunelg/2*Westonia white es WAWHT2894 Sun239R/2*Ajana white es Blade Snr-64//TznsPntsPrcz/Yq-54/3/Kt/RAC177 white es Non tolerant
Cunderdin Cranbrook sib/Sunfield sib white es
checks Hartog Vcm-71//Cn"S"/StCrrs/3/Klynsn/Blbrd. white es Janz 3Ag3/4*Condor//Cook white es Wyalkatchem Machete/ W84-129c*504 white es a WAWHT 2030 =IW725/Hyden b MEC-3=Sonora 64/Tezanos Pitos Precoz//Yaqui 54 c Gutha..Jacup*2.11th ISEPTON 135.
5.3.3 2.3. Trial design and treatments
The 2003/04 trials in Katanning, Esperance and Ravensthorpe, and the 2005/06 trial in
Katanning were laid out as split plot designs with harvest randomised within each

77
genotypes for the three reps. The moisture stress trials in Esperance in 2004/05 and 2005/06
were laid out as a split split-split-plot design, with the main plot factor water supply and the
subplot factor genotype and the subsubplot factor harvest. Essentialy this trial was the same
design as the others except two trials were sown, and one was irrigated and one moisture
stressed. The genotypes were randomised within water supply and harvest within genotype
with three reps. In these trials at Esperance in 2004/05 and 2005/06 a rainout tunnel, 20m x
9m with 3m high centres was placed over the entire trial and covered with a UV stable
translucent cover (Solarweave Natural , diffuse light transmission of 82 % Jaylon, Perth)
at anthesis (Zadoks Z69), and moisture stressed or irrigated treatments began with the
terminal moisture stress imposed by the rainout tunnel preventing any further rainfall, i.e.
plants in the moisture stress treatments had to rely on residual soil moisture from anthesis
to maturity. The irrigated plots were watered weekly to approximately 70 % field capacity
that was estimated as 70 % of historical Pan Evaporation figures, until the moisture stressed
plots all reached maturity. Irrigation was supplied by ground water using T-Tape
(International Model 515-20-250), without wetting the ears, at a rate that was equivalent to
130 mm rainfall spread over five weeks in 2004/05 and 195 mm spread over seven weeks
in 2005/06, and this averaged to 60 ± 4 % in 2004/05 and 90 ± 11 % in 2005/06 of actual
Pan Evaporation.
The clear solar weave cover reduced the daily solar radiation sum by 13 % from an average
of 23,299 ± 1,244 kj/m2 to 20,197 ± 1,244 kj m-2 in Esperance 2004/05. Previous
unpublished comparisons of germination indexes (2 reps, 4 shelters) with a set of check
lines under sealed white, sealed clear and clear only when rain was imminent covers has
shown that clear covers can lead to a slightly higher temperatures, higher humidity and
lower GI compared to white covers or plots only covered with a clear cover when rain was
imminent (Mares, unpublished). As clear covers were required to impose the moisture
stress from anthesis to maturity, and for this reason clear covers of the same material of the
same age were sourced from the same manufacturer and used over all trials. To minimise
the temperature and humidity build up associated with the clear cover an open ended tunnel
design was also used, and temperature and humidity monitored spatially throughout the
tunnel. No significant spatial variation could be detected with the 0.5°C accuracy of the

78
temperature sensors used. Hence any slight effect of the covers is likely to be the same
across this series of trials, as all trials were covered to the same extent.
5.3.4 2.4. Sampling
To prevent rainfall near maturity from inducing changes in dormancy in genotypes
differing in maturity, or preharvest sprouting in early maturing genotypes while later
genotypes were still green, clear rainout tunnels were placed over plots not already covered
at Zadoks Z85 (soft dough stages). The covers were removed after all genotypes had
reached maturity, Zadoks Z91 (~15-20 % grain moisture content, dry weight basis, thumb
nail indent held) for in-field estimation of preharvest sprouting tolerance.
In all trials ten ears from primary tillers were collected at maturity (Harvest 1) for each
replicate. Within 5 days of harvest the ears were gently threshed by hand and allowed to
dry on the bench to a grain moisture content of <12 % before storage at -20°C according to
the methods of Mares (1983). Grain dormancy was later determined on these samples. The
ears remaining in each plot (~70) were also harvested at the same time, (Harvest 1) allowed
to dry down to harvest maturity on the bench, then cleaned and threshed with a small plot
thresher for determination of falling number. For harvest 2 which was ~one month after
maturity after the next significant rainfall event and for harvest 3 which was ~two months
after maturity and after the next significant rainfall event each plot was hand harvested,
(~80 ears), threshed and cleaned with a small plot thresher for determination of falling
number. Some genotypes were not included at every harvest due to insufficient room under
the rainout tunnel. After maturity all the trials in Esperance and Katanning received at least
20 mm of rain between harvest 1 and 2 (Table 3). In 2003/04 and 2004/05 essentially no
more rain fell after harvest 2, however, in 2005/06 the rainfall after harvest 1 was much
higher than in previous seasons in both Katanning and Esperance (Table 3). Each harvest
was on a separate split plot, designed for the time series harvest.

79
Table 2. Sowing dates and the range in days from sowing to anthesis or maturity, and
sowing to anthesis rainfall at Esperance and Katanning in 2003/04, 2004/05 and 2005/06.
Sowing date anthesis maturity
earliest latest earliest latest
Days range in maturity
Rainfall sowing to anthesis
ESP 03 6/06/03 117 130 173 185 12 350 KT 03 29/05/03 118 132 175 180 5 272 ESP 04 24/05/04 112 129 169 179 10 262 ESP 05 31/05/05 107 126 174 188 12 294 KT 05 2/06/05 119 128 165 170 7 197 ESP 05 Stage 4 31/05/05 107 137 178 198 20 294
Table 3. Cumulative rainfall after harvest dates at maturity (H 1) and after significant
rainfall events ~one month after maturity (H 2) and ~ two months after maturity (H 3) at
Esperance and Katanning in 2003/04, 2004/05 and 2005/06.
Season Harvest
Esperance D Rainfall (mm)
N°RD Katanning
D Rainfall (mm)
N°RD
2003/04 1 10 Dec 03 0 8 Dec 03 0 2 7 Jan 04 28 23 8 3 4 Feb 04 56 25 9 5 Feb 04 59 29 5 2004/05 1 20 Nov 04 0 2 6 Jan 05 47 21 9 3 31 Jan 05 72 22 10 2005/06 1 30 Nov 05 0 21 Nov 03 0 2 9 Jan 06 40 50 10 10 Jan 06 50 75 18 3 31 Jan 06 62 70 23 2 Feb 06 73 122 24 D= number of days since maturity at harvest 1.
N°RD= number of rainy days during the time period between each harvest.
Grain dormancy was estimated by the methods of Walker-Simmons (1987). Fifty grains or
proximal (embryo) half-grains were incubated in a Petri dish with filter paper (3 x 70 mm,
No. 2 Advantec, Toyo Roshi Kaisha, Ltd. Japan) moistened with 6 mL deionised water.
Petri dishes were incubated at 20°C for 7 days and germinated grains or embryo half-grains
were counted daily. Germination was defined as pericarp rupture over the embryo. The
germination index for grains (GI) was calculated using Eqn 1.
(7 x n1 + 6 x n2 + 5 x n3 + 4 x n4 + 3 x n5 + 2 x n6 + 1 x n7) / (total days of test x total grains) (1)

80
where n1, n2, � n7 are the number of grains that germinated on the first, second and
subsequent days until the seventh day, respectively. The maximum germination index
representing non-dormant grains is 1.0 and the minimum is 0.0 representing fully dormant
grains (Walker-Simmons, 1987). Seed viability of grain that failed to germinate in seven
days was assessed by incubating the grain at 4°C for a further 3 days then a further 3 days
at 20°C.
Hagberg falling number (falling number) measurements were made on the grain samples at
maturity (Harvest 1), one month after maturity (Harvest 2) and two months after maturity
(Harvest 3) (Hagberg, 1960; Hagberg, 1961). Approximately 100 g of grain was milled
with a Perten 1800 falling number mill (Huddinge, Sweden). The moisture content of the
whole meal flour was then determined with a Marconi Instruments TF933C moisture meter
(St Albans, England). The falling number was determined with 25 ± 0.2 mL distilled water
and 7 ± 0.05 g of flour adjusted for 15 % moisture content in a Perten 1700 falling number
machine (Huddinge, Sweden). Control samples were run every 25 samples and analysis
continued as long as the value was ± 20 s of the mean.
5.3.5 2.5. Statistical Analysis
For the manipulated environment experiments in 2004/05 for GI and falling number in
Figure 1 the main effects and interaction of water supply and genotype were determined by
ANOVA in Genstat 9.1 for a split-plot design with the treatment water
supply*genotype*harvest, n = 3. Maturity scores were not used as a co-variate in this
analysis because it is confounded by the water supply treatments. Fischer�s LSD0.05 was
calculated for comparisons between means of treatments and cultivars. ANOVA tables
from this analysis are presented in Table 5.
For a combined analysis of germination index and falling number in the trials across the 5
sites and season combinations a linear mixed model was used. In each model environment
(site) were considered as fixed, environment.genotype and environment.rep as random. Two
additional terms maturity score and temperature were included as covariates. Where
maturity score was the days from seeding to maturity and temperature the sum of
maximum daily temperatures during the 30-10d period prior to maturity previously found
to influence dormancy (Biddulph et al, 2007). The irrigated plots from Esperance in 2004

81
and 2005 were included in this analysis as that year, as the irrigation only had a small effect
on falling number (2% Table 5). Also a diagonal covariance structure was fitted for
environment.genotype term in order to accommodate the heterogeneity of genetic variance
across environments. The model was fitted using REML procedure in Genstat Release 9.1.
BLUPS (Best Linear Unbiased Predictors) for each genotype at each site were obtained
from the model and presented in Figure 2.
Bi plots were constructed to visually depict the genotype, environment main effects and
interaction. A subset of the data analysed by REML which constituted a balanced design
for a common set of 6 genotypes were analysed by ANOVA for a randomised block design,
for germination index and falling number at maturity and harvest 3. With the treatment
structure genotype*environment. The terms environment (site) were considered as fixed,
environment.genotype and environment.rep as random.
For the analysis of the stage 4�s in Fig. 3. the germination index and falling number were
analysed by ANOVA in Genstat 9.1 for a randomised block design. Maturity scores were
used as a co-variate in this analysis. Fischer�s LSD0.05 was calculated for comparisons
between means for GI and falling number. For the analysis of the germination index,
weather affected grain, black point, fungal staining and field mould were analysed by
ANOVA in Genstat 9.1 for randomised block designs. Maturity scores were used as a co-
variate in this analysis. Fischer�s LSD0.05 was calculated for comparisons between the
means of three reps.
5.4 3. Results
In 2005/06 at Esperance the germination index at maturity was the same under both
irrigated and moisture stress; DM 2001, AUS 1408, DH 22 and DH 56 were strongly
dormant (GI <0.02), DH 45 was partially dormant, (GI 0.30) and Cascades and Cunderdin
non-dormant, (GI > 0.60; Fig. 1 a, c). There was no significant effect of the irrigation
treatment on germination index (Table 5). Maturity scores were not used as a co-variate on
dormancy and falling number because drought reduced time to maturity and irrigation, but
within droughted plants or irrigated plants maturity was not a significant co-variate, only
between droughted and irrigated plants. At maturity falling number was significantly
influenced by genotype, water supply and their interaction (Table 5). Moisture stressed

82
plants had a higher falling number than irrigated, particularily the partially and non-
dormant genotypes (Fig. 1b, d). After rainfall however at harvest 2 and 3 the falling number
under both water stressed and irrigated conditions was the same (Table 5) with the strongly
dormant genotypes maintaining >300 s despite up to 70 mm of rain (Fig. 1 b, d). By
comparison the falling number of DH 45, Cascades and Cunderdin declined to <150 s (Fig.
1 b, d). Most of the variation in falling number after 50 and 70 mm of rainfall at Harvest 2
and 3 was due to genotype (90 and 95% respectively) and there was no genotype by
irrigation interaction (Table 5). Samples from Esperance in 2004/05 had a similar
germination index to 2005/06, and there was no decline in falling number following 25 mm
of rain (Table 3).
Table 4. Analysis of variance tables for Bi plots of germination index at maturity and
falling number at maturity (H1) and the final harvest (H3) for the set of 6 common
genotypes (AUS1408, DM2001, Camm, Cascades, EGA Eagle Rock and Janz) over 5
common sites.
Source of variation deg. of freedom
Mean squares
Variance ratio
F prob. % Variance
Germination Index Site (S) 4 0.097 24.91 <.001 7Block 10 0.004 0.9 0Genotype (G) 5 1.228 282.96 <.001 91S.G 20 0.023 5.34 <.001 2Residual 47 0.004 0Total 86 Falling Number H1 Site (S) 4 20539.7 36.62 <.001 63Block 10 560.9 1.13 2Genotype (G) 5 9084.1 18.37 <.001 28S.G 20 1864.6 3.77 <.001 6Residual 49 494.5 Total 88 Falling Number H3 Site (S) 4 289395 371.15 <.001 74Block 10 780 0.75 Genotype (G) 5 88020 84.47 <.001 23S.G 20 10312 9.9 <.001 3Residual 50 1042 Total 89

83
Table 5. Analysis of variance table for germination index at maturity and falling number
with three harvests, with the water stress treatments from Esperance 2005.
Source of variation deg. of freedom
Mean squares
Variance ratio
F prob. % Variance
Germination Index Block 2 0.023 3.99 5Genotype (G) 6 0.413 72.08 <.001 92Irrigation (I) 1 0.008 1.44 0.241 2G.I 6 0.001 0.17 0.983 0Residual 26 0.006 0Total 41 Falling Number H1 Block 2 604.6 0.81 4Genotype (G) 6 5155.3 6.91 <.001 31Irrigation (I) 1 7360.4 9.87 0.004 45G.I 6 2656.9 3.56 0.01 16Residual 26 745.6 5Total 41 Falling Number H2 Block 2 254 0.12 0Genotype (G) 6 61802 28.76 <.001 90Irrigation (I) 1 3829 1.78 0.194 6G.I 6 527 0.25 0.957 1Residual 26 2149 3Total 41 Falling Number H3 Block 2 1685 1.4 2Genotype (G) 6 70888 58.86 <.001 95Irrigation (I) 1 315 0.26 0.613 0G.I 6 289 0.24 0.959 0Residual 26 1204 2Total 41
Between Katanning and Esperance over three seasons, levels of dormancy were
significantly different. The bi-plots (Fig. 3 a) and ANOVA analysis (Table 4) indicate that
in terms of dormancy different environments produced different levels of dormancy. For
example Esperance 04 and 05 grouped together as did Katanning 05 and Esperance 03 (Fig.
3a). However 91 % of the variation in GI was attributed to genotype and only 7 % to site
and 2 % to the site by genotype interaction (Table 4). Hence all the bi plot really indicates
is that AUS1408 and DM2001 responded in a similar fashion to the environments
compared to the other genotypes (Fig. 3a). In terms of falling number, after rainfall at
harvest 2 and 3 most of the variation was due to site (63 and74% respectively; Table 4) and

84
genotype (28 and 23% respectively; Table 4). Though the site by genotype interaction was
significant (Table 4) it did not account for much of the variation in falling number after
rainfall (6 and 3 % at H2 and 3). Esperance and Katanning 05 which had the most rainfall
(Table 3), also had reduced falling number at maturity and harvest 3 (Fig. 2 c, e). In terms
of the environments at the different sites, the biplots in Figure 3 indicate that Katanning
2005, was distinct from the rest. The main point is that the relationship between genotypes
changes from falling number at maturity to falling number at harvest (Fig. 3 b vs Fig. 3 c).
Indicating that at harvest 3 following rainfall regardless of environment the falling number
of AUS1408 and DM 2001 are similar while that of Janz, Camm and Cascades fall into a
different group and EGA Eagle Rock was somewhere in between.
DM 2001ES
Fallin
g nu
mbe
r (s)
Ger
min
atio
n In
dex
b) Irrigated falling number
a) Irrigated germination index c) Moisture stressed germination index
d) Moisture stressed falling number
0.0
0.2
0.4
0.6
0.8
1.0
0
100
200
300
400
500
DM 2001ES
AUS 1408ES
DH22ES
DH56Es
DH45eS
Cascadeses
Cunderdines
Dormant Partially dormant None
AUS 1408ES
DH22ES
DH56Es
DH45eS
Cascadeses
Cunderdines
Dormant NonePartially dormant Fig. 1. Germination index at maturity (a, c) and falling number (b, d) of irrigated (a, b) or
moisture stressed (c, d) plots at maturity (Harvest 1, solid bars,) after 50 mm rain (Harvest
2, shaded bars) and 70 mm of rain (Harvest 3, open bars) at Esperance in 2005/06. In
genotypes with strong dormancy with the embryo and seed coat components, strong
dormancy with just the embryo component, partial dormancy, with just the seed coat
component or in non-dormant genotypes. The dashed lines in b and d represent the
minimum falling number for premium grades. Means of 3 reps, vertical bars represent
LSD0.05.

85
A similar response of falling number to rainfall as presented in the biplots can be observed
in the REML estimates presented in Figure 2. Adding either maturity score, measured as
days from seeding to maturity or the sum of maximum temperature during the 30-10d
period prior to maturity improved the REML analysis, however adding both did not.
Indicating hat they were accounting for the same proportion of variation, and as a result
maturity was used. In Esperance 03, 25 mm of rainfall resulted in a decline in falling
number for Cascades, EGA Castle Rock, Janz and Hartog (Fig. 2 a) but not AUS 1408, DM
2001, EGA Eagle Rock or Camm in 2003/04. In 2004/05 however, 22 mm of rainfall had
no effect on falling number (Fig. 2 b). In 2005/06 50 mm of rainfall in the first month
reduced falling number in Cascades and Cunderdin but not AUS 1408 and DM 2001 (Fig. 2
c). Whilst a further 20 mm in the second month reduced falling number in all genotypes
except AUS 1408 (Fig. 2c). By contrast a decline in falling number was only observed in
Katanning in 2005/06 2 months after maturity with a total of 122 mm of rainfall. In
Katanning, 75 mm (Fig. 2f) in the first month in 2005/06 or 29 mm (Fig. 2d) in 2003/04
had no significant effect, indicating that more rainfall was required in Kataning to reduce
the falling number that Esperance. Across sites and seasons after rainfall the falling
numbers for the strongly dormant genotypes AUS 1408 and DM 2001 remained the same
as before, Camm and EGA Eagle Rock declined 50-100 s, while EGA Castle Rock,
Cascades, Janz, Cunderdin and Hartog declined >100 s.

86
d) Katanning 2003/04a) Esperance 2003/04
c) Esperance 2005/06
b) Esperance 2004/05
Falli
ng n
umbe
r (s)
e) Katanning 2005/06
0
100
200
300
400
500
0 mm
20 mm
25 mm
0 mm
20 mm
25 mm
0
100
200
300
400
500
0
100
200
300
400
500
AUS 1408
DM 2001
EGA Eag
le Roc
kCam
mCas
cade
s
EGA Cas
tle R
ock
Janz
Cunde
rdin
Hartog
0 mm
21 mm
22 mm
0 mm
21 mm
22 mm
0 mm
50 mm
70 mm
0 mm
50 mm
70 mm
0
100
200
300
400
500
0
AUS 1408
DM 2001
EGA Eag
le Roc
kCam
mCas
cade
s
EGA Cas
tle R
ock
Janz
Cunde
rdin
Hartog
0 mm
75 mm
122 mm
0 mm
75 mm
122 mm
0 mm
29 mm
0 mm
29 mm
100
200
300
400
500
Fig. 2. BLUPS for falling number at successive harvests at maturity, Harvest 1 (solid bars,
Harvest 2 (shaded bars) and Harvest 3, (open bars) in Esperance (a) 2003/04, (b) 2004/05,
(c) 2005/06, or Katanning (d) 2003/04 and (e) 2005/06 with different amounts of rainfall
detailed in Table 3. The dashed lines represent the minimum falling number for premium
grades. Means of 3 reps, vertical bars represent LSD0.05.
At Esperance in 2005/06 when diverse germplasm was screened for dormancy at maturity
(germination index) and grain quality (falling number) after 1 month and 50 mm of rainfall
there was a correlation (r = -0.68) between germination index and falling number (Fig. 4).
The genotypes were separated into three groups on the basis of germination index into
strongly dormant genotypes (germination index <0.20), partially dormant genotypes
(germination index 0.20-0.50) and non-dormant genotypes (germination index >0.50) (Fig.
4). When genotypes with strong dormancy were removed from the analysis the strength of
the correlation was greatly reduced (r = -0.44). The genotypes with strong dormancy
included three types; those with dormancy from AUS 1408 (AUS 1408, DM 2001, DH 22
and DH 56), which all had a falling number around 400 s, a second group of Canadian
origin (Snowbird and Kanata) which had a falling number around 500 s (Fig. 4) and
7HRWSN58 a red grained CIMMYT genotype with a falling number of 311 s. The

87
partially dormant material QT7475, Calingiri, Cadoux, Cascades, EGA Castle Rock and
Binnu had a germination index of ~0.40 but lower falling number at ~250 s (Fig. 3). The
non-dormant material consisted of two types with similar germination index but separate,
distinct falling numbers. The first type, mainly of Spear derived genotypes; (Stiletto,
Clearfield STL and Camm) plus EGA Eagle Rock, Frame Blade and three breeding
genotypes (Table 1) had a slightly higher falling number (250-380 s) than the �others�
group, which included 22 named genotypes and 23 breeding genotypes whose falling
number ranged from 70-250 s (Fig. 4). These Spear derived genotypes were distinguishable
from the others in two out of the four seasons in Esperance and Katanning using
germination index, but not in 2005/06
c) FN H3
PC-1 environments
a) GI maturity
AE5
EAJ
CA
E4
E3
K3
K5
CS
D
-0.8
-0.6
-1.0
-0.41.0
-0.8
0.0
-0.6
0.6
0.2
-0.21.0
0.8
1.0
0.6
-1.0
0.4
-0.6
0.2
0.0 0.4
0.0
0.0
-1.00.8
-0.2
0.4
0.8
0.2
-0.4
1.0
0.2
0.4
-0.8
-1.0
-0.6
PC-1 genotypes
-0.4
-0.2
0.6
-0.4
-0.8 0.6
-0.2
0.8
PC
-2 environmentsP
C-2
gen
otyp
es
b) FN H1
CS
CA
EA
E5
J
E4DA
E3K3
K5
10
15
5
-15
20
-5
15
0
1050
15
-5
5
-10
15
-15
-5
-20
-15
PC-1 environments
PC-1 genotypes
-10-20
-15
20
-10
5
0
-20
-5
0
10
-10
-20 10 20
20
PC
-2 environments
PC
-2 g
enot
ypes
CSCA
EA
E5E4
D
A
E3
K3
K5
10
15
5
-15
20
-5
15
0
1050
15
-5
5
-10
15
-15
-5
-20
-15
PC-1 environments
PC-1 genotypes-10
-20
-15
20
-10
5
0
-20
-5
0
10
-10
-20 10 20
20
PC
-2 environmentPC
-2 g
enot
ypes
J
Fig. 3. Biplots for germination index at maturity (a) and falling number at maturity (b) and
the final harvest (c) for the set of 6 common genotypes (A = AUS1408, D = DM2001, CA
= Camm, CS = Cascades, E = EGA Eagle Rock and J = Janz) over 5 common sites (E3 =
Esperance 2003/04, E4 = Esperance 2004/05, E5 = Esperance 2005/06, K3 = Katanning
2003/04 and K5 = Katanning 2005/06).

88
Staining of the grain due to black point and various fungal pathogens including
Alternarium, Ulocladium and Stemphylium sp. occurred in Esperance and Ravensthorpe in
the grain sampled from 2003/04, but not to the same extent in subsequent sites or seasons.
Ravensthorpe received 47 mm, twice as much rainfall after maturity than Esperance and
had a several-fold higher count of fungal staining and field mould affected grains, but lower
levels of black point (Fig. 4). At both Esperance and Ravensthorpe genotypic differences in
weather affected (Fig. 4b), black point (Fig. 4c) and field mould (Fig. 4e) affected grains
were evident. In Esperance the majority (~80 %) of the weather affected count (black point
+ fungal stained) was made up of black point affected grains (Fig. 4), while in
Ravensthorpe the majority was fungal stained grain.
0.0
0.2
0.4
0.6
0.8
1.0
0 100 200 300 400 500
Falling number (s)
Ger
min
atio
n In
dex
Spear types
Cascades
AUS 1408
Others
Strongly dormant
Partially dormant
Non dormant
CIYMMT
Fig. 4. Relationships between germination index and falling number after 50 mm rainfall in
Esperance 2005/06 (r = -0.68) for 26 Western Australian breeding genotypes, 35
commercial genotypes common to Western Australia and 10 sprouting tolerant check
genotypes. Three groups of dormancy are shown; non-dormant, partially dormant and
strongly dormant, with five plant types (circles) informally allocated based on similar
parentage detailed in Table 1 and similar falling number after rainfall. The dashed line
represents the minimum falling number for premium grades. Values are the means of 3 reps
for germination index and falling number.

89
b) Weather affected = black point + fungal stained
e) Field mould
0100200300400500
Falli
ng n
umbe
r (s
)0
102030405060
0102030405060
0102030405060
0102030405060
Casca
des
EGA Eagle
RockCam
m
Hartog
EGA Cas
tle R
ock
Wya
lkatch
em
DM 2001
Janz
AUS 1408
Fiel
d M
ould
(g
rain
s / 5
00 g
rain
s)Fu
ngal
sta
ined
(g
rain
s / 5
00 g
rain
s)Bl
ack
poin
t (g
rain
s / 5
00 g
rain
s)W
eath
er a
ffect
ed
(gra
ins
/ 500
gra
ins)
a) Falling number
c) Black point
d) Fungal stained
Fig. 5. Falling number (a), weather affected (black point + fungal stained) (b), black point
(c), fungal stained (d), and field mould (e), ratings according to Australian Wheat Board
2003/04 delivery specifications. Grain was grown under natural rainfall conditions in
Esperance (solid bars) or Ravensthorpe (open bars) 2003/04, harvested on 20 January 2004
after 23 mm and 47 mm post maturity. Plots were covered with a rainout shelter at Zadoks
Z85 of earliest genotype, to minimise rainfall by maturity interactions in Esperance but not
in Ravensthorpe. The dashed lines represent the minimum falling number or the maximum
count for weather affect grains for premium grades. Means, 3 reps, vertical bars represent
LSD0.05 for comparisons between genotypes

90
5.5 4. Discussion
In Australia the Australian wheat Board sets minimum standards for receiving grain into
different payment grades. Down-grading from premium to feed grades results in a 20 %
lower grain price, hence the ability of a genotype to maintain a falling number above the
minimum 300 s for premium grades is important in determining the returns to growers and
forms the basis of what is defined as preharvest sprouting tolerant. At sites and seasons
when preharvest sprouting occurred the strongly dormant (germination index <0.20) AUS
1408 derived genotypes of DM 2001, DH 22, and DH 56 had falling numbers greater than
the Australian Wheat Board premium grade cut-off for falling number of 300 s (Australian-
Wheat-Board, 2003-2004) (Fig 1b,d and Fig. 2). The partially dormant DH 45 and
Cascades (germination index 0.20-0.50) generally had falling numbers less than the 300 s
cut off, but still higher than 250 s. All other non dormant genotypes (germination index
>0.50) except the Spear types showed little tolerance and had a falling number below 250 s
and would have been downgraded to feed grades with an associated 20 % loss in value
(Australian-Wheat-Board, 2003-2004). Genotypes with strong dormancy (embryo and seed
coat or embryo mechanism) were consistently tolerant to preharvest sprouting, genotypes
with partial dormancy were intermediate and genotypes without dormancy were generally
susceptible.
Selection for genotypes with a partially dormant phenotype (germination index 0.20-0.50)
does not give effective preharvest sprouting tolerance in severe conditions. The partially
dormant genotypes (germination index 0.20-0.50) generally had falling numbers less than
the 300 s cut off, but still higher than 250 s. The higher falling number of these genotypes
means partial dormancy does give greater tolerance than non-dormant genotypes as all non
dormant genotypes (germination index >0.50) except the Spear types showed little
tolerance and had a falling number below 250 s. An alternative to partial dormancy for
environments, such as Katanning, which are less prone to preharvest sprouting, may be to
use traits, other than dormancy. For example the Spear types (Spear, Camm, Stiletto,
Clearfield STL) and EGA Eagle Rock, Frame and Blade had some tolerance to preharvest
sprouting in the field despite having similar dormancy to the other genotypes that were
susceptible. These genotypes appeared to have a greater level of preharvest sprouting
tolerance than could be explained by dormancy alone. These genotypes were among the last

91
to mature and all had high falling number >350 at maturity. King (1984) also identified ear
characteristics which reduce the duration the grain was wet and suggested that these could
reduce sprouting; further work is required to determine if these Spear types contain such
traits. Despite this the high falling number at maturity and matching maturity to
environment gave a minor level of preharvest sprouting tolerance when falling number was
measured.
Although there was a significant G by E effect on dormancy, 91% of the variation in
dormancy was due to genotype and only 7 % due to environment and 2% to environment
by genotype interactions (Table 4). Falling number after rainfall at harvest 2 and 3 however
was more strongly influenced by the environment (63 and 74%) and genotype (28 and
23%), with a small G by E interaction (6, 3%).The larger influence of environment on
falling number after rainfall is expected as sites differed substantially in the duration,
amount and timing of the rainfall events and hence severity of sprouting. Highlighting the
major limitation of using falling number as out of the five environments, only two seasons
gave reasonable discrimination for sprouting tolerance on a falling number basis.
Selection for preharvest sprouting tolerance in some wheat breeding programs is currently
based on the germination index at maturity and an important validation of this strategy is to
compare this trait with the response to weathering in the field (falling number after natural
rainfall). However, the main limitation of the falling number approach to characterise grain
for preharvest sprouting is the variation due to differences in time from maturity to a
specific rainfall event. Furthermore the rainfall, although similar in amount between some
sites and seasons, i.e. 20-29 mm in the first month after maturity, rainfall presumably varies
in the extent it wets and hence germinates the grain. In the 2003/04 season at Katanning for
example, there was no preharvest sprouting or decline in falling number following 20 mm
of rain in contrast to Esperance. The same response of falling number to rainfall occurred
in 2005/06, with 122 mm of rainfall in Katanning being roughly equivalent to 70 mm of
rainfall in Esperance in 2005/06 (Fig 2 c,e). Other studies have also found a poor
correlation with direct evaluation of sprouting tolerance by falling number with only one in
three years correlated (Guta and Bichonski 2007). Esperance 2005/06 gave the best
genotype discrimination in terms of falling number. However according to Humphreys and
Noll (2002) falling number after natural rainfall or artificial weathering is a more reliable

92
procedure for screening for preharvest sprouting than dormancy. The data presented here
appears to be consistent with this suggestion. Selection for preharvest sprouting tolerance in
the field requires selection for genotypes with a germination index <0.20, and/or selecting
for genotypes with a high falling number after preharvest sprouting inducing rainfall.
Although falling number after rainfall is the fundamental test of sprouting tolerance, it has
limitations and should be used in conjunction with dormancy estimates of sprouting
tolerance.
In Esperance 2004/05 and Katanning 2003/04 the rainfall was not enough to induce a
significant level of sprouting, i.e. less than 1% of grains were visually sprouted in the
susceptible genotypes. Under these conditions without sprouting, the falling number
increased. The increase in falling number is a common phenomenon observed under high
temperatures post maturity (Lambe, W. Pers.comm). Alpha-amylase is degraded by the
constant exposure to high temperatures in the field. High temperature (>30°C) degrades
cereal alpha amylase (Lim et al., 2003), and in the case of flour this results in higher falling
numbers. This occurs routinely in the laboratory and for this reason our grain samples for
alpha amylase are kept in cold storage where it still degrades, albeit at a slower rate. Hence
the increase in falling number in the field with daily maximum temperatures >30°C in the
absence of sprouting may be due to gradual degradation of alpha amylase.
In preharvest sprouting prone areas, avoiding genotypes susceptible to other weather
associated defects such as black point and fungal infection may also be beneficial. It was
observed here and elsewhere (Mares and Mrva, 2001; Williamson, 1997) that black point
affected grains germinate rapidly regardless of the typical germination pattern of the
genotype. A similar increase in germination has also been reported with fungal infection by
Pyrenophora sp. during the germination assay (Fernandez et al, 1998; Fox et al, 2003).
Genotypes with lower levels of black point also have lower levels of fungal infection on the
grain. This is consistent with Williamson (1997) who found an association between levels
of black point and Alternarium sp. infection. Genotypic tolerance for black point is well
documented (Lehmensiek et al., 2004), and data presented here supports other work in
wheat (Fernandez et al, 1998) and barley (Young and Loughman, 2001) which also found
genotypic differences in fungal staining of grain are associated with the differences in black
point. Further work is required to confirm if the association between black point and fungal

93
staining of grain is due to the similar environmental conditions which induce both, or if
black point occurs as a response to the fungal infection.
The environmental conditions during grain filling may be influencing preharvest sprouting
tolerance through their known effect on black point and fungal infection. Even though
visually black point and mould affected grains were excluded from the germination tests,
the possibility remains that non-visible symptoms may still be influencing the germination
indexes presented here. Furthermore, more severe black point infection has been found to
be associated with increased alpha-amylase activity and lower falling numbers (Lorenz
1986). Hence a lower level of black point and possibly fungal infection will also lead to
less preharvest sprouting for any given level of dormancy. In addition to the fact that black
point and fungal infection are also important defects associated with weather damage,
selecting for preharvest sprouting tolerance requires selecting tolerance to black point and
fungal infection in addition to dormancy , to ensure that dormancy is not eroded by black
point or fungal infection and falling numbers are not reduced by sever black point.
5. Conclusion The combination of strong dormancy (germination index <0.20), matching maturity to
environment and high initial falling number may lead to more durable preharvest sprouting
tolerance than is currently available in white grain wheat. Screening for preharvest
sprouting tolerance based on falling number after rainfall has limitations but can highlight
minor levels of tolerance in addition to dormancy, but should only be done to complement
dormancy estimates of sprouting tolerance. Other defects associated with rainfall during
grain filling such as black point and fungal staining may slightly reduce dormancy
estimates/ preharvest sprouting tolerance, and for this reason need to be improved in
parallel with preharvest sprouting tolerance.
5.6 Acknowledgments
This work was funded by the Grains Research and Development Corporation in support of
TBB through a GRDC Grains Research Scholarship GRS66, the Department of Agriculture
and Food Western Australia who assisted with the running of field trials and study leave
awarded to TBB and the Ravensthorpe Agricultural Initiative Network through a grant to

94
TBB. Thank you also to Colin Norwood, Colin Boyd and the technical staff at Esperance
Downs Research Station for excellent assistance and advice with running these trials.
5.7 References
Auld, A.S., Paulsen, G.M., 2003. Effects of drought and high temperature during
maturation on preharvest sprouting tolerance of hard white winter wheat. Cereal
Res. 31, 169-176.
Australian-Wheat-Board, 2003-2004. AWB wheat receival standards 2003-2004.
(Australian Wheat Board Limited)
Biddulph, T.B., Mares, D.J., Plummer, J.A. Setter, T.L., 2005. Drought and high
temperature increases preharvest sprouting tolerance in a genotype without grain
dormancy. Euphytica 143, 277-283.
Biddulph, T.B., Plummer, J.A., Setter, T.L. Mares, D.J., 2007. Influence of high
temperature and terminal moisture stress on dormancy in wheat (Triticum aestivum
L.). Field Crop. Res. Doi:j.fcr.2007.05.005.
Bureau of Meteorology, 1956-2006. Historical rainfall records, Esperance Downs Research
Station and Great Southern Agricultural Research Institute. (Bureau of
Meteorology, Perth).
DePauw, R.M., McCaig, T.N., Mares, D.J., Brennan, P., Henry, R.J., King, R., McEwan,
J.M., Gordon, I. 1989. Interrelationships among assays for germination of
kernels from threshed spikes, unthreshed spikes and alpha-amylase in wheat.
In '5th International Symposium on Preharvest Sprouting in Cereals'. USA.
(Eds K Ringlund, E Mosleth and DJ Mares) 195-205. (Westview Press Inc.
Boulder Co)
DePauw, R.M., McCraig, T.N., (1991) Components of variation, heritabilities and
correlations for indices of sprouting tolerance and seed dormancy in Triticum spp.
Euphytica 52, 221-229.
DePauw, R.M., McCaig, T.N., Baker, R.J., Clarke, J.M., 1993 Constructing a sprouting
tolerance index. In 'Sixth International Symposium on Pre-harvest Sprouting in
Cereals'. USA. (Eds MK Walker-Simmons, JL Ried) 47�53. (Westview Press,
Boulder, Co).
Fenner, M., 1991. The effects of the parent environment on seed germinability. Seed Sci.
Res. 1, 75-84.

95
Fernandez, M.R., DePauw, R.M., Clarke, F.R., Fox , S.L., 1998 Discoloration of wheat
kernels by Pyrenophora tritici-repentis. Can. J. of Plant Path. 20, 380-383.
Fox, S.L., Fernandez, M.R., DePauw, R.M., 2003. Red Smudge infection modifies
sprouting response in four wheat lines. Can. J. of Plant Sci. 83, 163-169.
Gordon, A.G., 1971. The germination resistance test-A new test for measuring germination
quality of cereals. Can. J. Plant Sci. 51, 181-183.
Gubler, F., Millar, A.A., Jacobsen, J.V., 2005. Dormancy release, ABA and pre-harvest
sprouting. Curr. Opin. Plant Biol. 8, 183-187.
Gut, M., Bichonski, A., 2007 Technological quality and yield's components of winter
wheat lines under Polish climatic conditions. Cereal Res. Comm. 35, 151-161.
Hagberg, S., 1960. A rapid method for determining alpha-amylase activity. Cereal Chem.
37, 218-222.
Hagberg, S., 1961. Note on a simple rapid method for determining alpha-amylase activity.
Cereal Chem. 38, 202-203.
Hagemann, M.G., Ciha, A.J., 1987. Environmental x genotype effects on seed dormancy in
wheat. Agron. J. 79, 192-196.
Humphreys, D.G., Noll, J., 2002. Methods for characterization of preharvest sprouting
resistance in a wheat breeding program. Euphytica 126, 61-65.
King, R.W., 1984. Water uptake in relation to pre-harvest sprouting damage in wheat: grain
characteristics. Aust. J. Agric. Res. 35, 337-345.
King, R.W., 1993. Manipulation of grain dormancy in wheat. J. Exp. Bot. 44, 1059-1066.
King, R.W., Richards, R.A., 1984. Water uptake in relation to pre-harvest sprouting
damage in wheat: ear characteristics. uptake in relation to pre-harvest sprouting
damage in wheat: grain characteristics. Aust. J. Agric. Res. 35, 327-336.
King, R.W., Wettstein-Knowles, P.V., 2000. Epicuticular waxes and regulation of ear-
wetting and pre-harvest sprouting in barley and wheat. Euphytica 112, 157-166.
Lehmensiek, A., Campbell, A.W., Williamson, P.M., Michalowwitz, M., Sutherland,
M.W., Daggard, G.E., 2004. QTLs for black-point resistance in wheat and the
identification of potential markers for use in breeding programmes. Plant Breed.
123, 410-416.
Lim, L.H., Macdonald, D.G., Hill, G.A., 2003. Hydrolysis of starch particles using
immobilized barley a-amylase. Biochem. Eng. J. 13, 53-62.

96
Lorenz, K., 1986 Effects of blackpoint on grain composition and baking quality of New
Zealand wheat. N. Z. J. of Agric. Res. 29, 711-718.
Lunn, G.D., Kettlewell, P.S., Major, B.J., Scott, R.K., 2002. Variation in dormancy
duration in the U.K. wheat cultivar Hornet due to environmental conditions during
grain development. Euphytica 126, 89-97.
Mares, D.J., 1983. Preservation of dormancy in freshly harvested wheat grain. uptake in
relation to pre-harvest sprouting damage in wheat: grain characteristics. Aust. J.
Agric. Res. 34, 33-38.
Mares, D.J., 1993. Pre-harvest sprouting in wheat. I. influence of cultivar, rainfall and
temperature during grain ripening. uptake in relation to pre-harvest sprouting
damage in wheat: grain characteristics. Aust. J. Agric. Res. 44, 1259-1272.
Mares, D.J., Mrva, K., 2001. Mapping quantitative trait loci associated with variation in
grain dormancy in Australian wheat. Aust. J. Agric. Res. 52, 1257-1265.
Nielsen, M.T., McCrate, A.J., Heyne, E.G., Paulsen, G.M., 1984. Effect of weather
variables during maturation on preharvest sprouting of hard white winter wheat.
Crop Sci. 24, 779-782.
Reddy, L.V., Metzger, R.J., Ching, T.M., 1985. Effect of temperature on seed dormancy of
wheat. Crop Sci. 25, 455-458.
Stoy, V., 1983. Progress and prospect in sprouting research. In 'Third International
Symposium on Pre-Harvest Sprouting in Cereals.' Canada. (Eds J. E. Kruger and D.
E. LaBerge) pp. 3-7. (Westview Press, Inc.)
Strand, E., 1989. Studies on seed dormancy in small grain species. II. wheat. Norwegian J.
Ag. Sci. 3, 101-115.
Trethowan, R.M., 1995. Evaluation and selection of Bread Wheat (Triticum aestivum L.)
for Preharvest Sprouting Tolerance. Aust. J. Agric. Res. 46, 463-474.
Trethowan, R.M., Rajaram, S. Ellison, F.W., 1996. Pre-harvest sprouting tolerance of
wheat in the field and under rain simulation. Aust. J. Agric. Res. 47, 705-716.
Walker-Simmons, M., 1987. ABA levels and sensitivity in developing wheat embryos of
sprouting resistant and susceptible cultivars. Plant Physiol. 84, 61-66.
Williamson, P.M., 1997. Black point of wheat: in vitro production of symptoms, enzymes
involved, and association with Alternaria alternata. Aust. J. Agric. Res. 48, 13-19.

97
Xiao, S.-H., Zhang, X.-Y., Yan, C.-S., Lin, H., 2002. Germplasm improvement for
preharvest sprouting resistance in Chinese white-grained wheat: An overview of the
current strategy. Euphytica 126, 35-38.
Young, K.J., Loughman, R., 2001. Fungal associations with weather stained barley in
Western Australia. In '10th Australian Barley Technical Symposium'. Canberra. pp.
1-7. (ABT Ltd)

98
6 Chapter 6 General Discussion Preharvest sprouting tolerance for the purpose of this thesis has been defined as the level of
dormancy required to achieve sound grain with a falling number >300 s despite rain, in
genotype with regional averages, between maturity and harvest. Breeding and selecting for
preharvest sprouting tolerance is difficult and problematic because the main adaptive trait,
dormancy at maturity, is influenced by the environmental conditions during grain
maturation. There are three main factors which influence the level of preharvest sprouting
tolerance; (i) dormancy, (ii) grain filling and maturation environment and (iii)
morphological traits. The first factor, (i) dormancy, appears to be controlled by three
mechanisms, (a) embryo ABA elevation, (b) sensitivity and (c) the seed coat. The second
factor, (ii) environmental conditions during grain filling and maturation, can influence the
level of dormancy, possibly by changing one or all of the three dormancy mechanisms. The
last factor, (iii) morphological traits of the head that influence grain wetting in the field,
appears to have an additional effect on preharvest sprouting independent of dormancy. The
combination of these three factors, dormancy, environment and morphological traits, then
represents the level of preharvest sprouting tolerance, which will occur in a particular
genotype in the field in a particular season; these are each considered separately below.
6.1 Control of dormancy; elevation, sensitivity and seed coat
6.1.1 Embryo elevation of ABA
Results presented in Chapter 3 are the first evidence that the embryo component of
preharvest sprouting tolerance is related to the endogenous embryo ABA concentration
during imbibition under preharvest sprouting assay conditions. Hence, higher
concentrations of endogenous embryo ABA for longer time is associated with preharvest
sprouting tolerance. The greater dormancy in intact grain compared to isolated embryos or
embryo-half grains (Chapter 3) is due to a longer elevation of ABA concentration
compared to work using isolated embryos of wheat (Suzuki et al. 2000). The difference
between Cunderdin and DH 22 is the embryo ABA concentrations during the 6 to 18 hours
of imbibition period. The control of dormancy in wheat by the embryo component appears
to be similar to the dormancy previously reported in barley and Arabidopsis (Ali-Rachedi
et al. 2004; Benech-Arnold et al. 2006; Chono et al. 2006; Jacobsen et al. 2002; Millar et

99
al. 2006). Elevation of embryo ABA concentration during imbibition therefore appears to
explain, at least in part, the dormancy and hence preharvest sprouting tolerance originally
described as embryo dormancy. Further work is required to confirm the elevation of
embryo ABA during imbibition to see if the relationship holds for a wider range of
situations including genotypes with a range of dormancy and environments throughout
grain maturation.
In barley and Arabidopsis, a low ABA concentration in non dormant germinating grain is
due to catabolism of ABA by 8�-ABA hydroxylase. (Ali-Rachedi et al. 2004; Benech-
Arnold et al. 2006; Chono et al. 2006; Millar et al. 2006). The degradation of ABA by this
enzyme requires oxygen. The hull of barley (Benech-Arnold et al. 2006) and beechnut
(Barthe et al. 2000) can inhibit this degradation by maintaining anoxic conditions in the
embryo, preventing ABA catabolism and hence maintain dormancy. Oxygen consumption
by the seed coat due to the oxidation of phenolic compounds has been suggested in
beechnut to restrict oxygen diffusion into the embryo (Barthe et al. 2000). The hull in
barley has been proposed to play a similar role (Benech-Arnold et al. 2006). Without a
tightly attached lemma and palea in wheat, a similar function of the seed coat in wheat is
less likely, and other work has shown no difference in oxygen consumption during the first
critical 12 hours of imbibition between dormant and non-dormant genotypes of wheat
(Miyamoto et al. 1961). This however requires further confirmation using better estimates
of oxygen and an examination of phenolic contents of wheat seed coats.
In wheat, synthesis of free ABA by the embryo during imbibition has been proposed to
counteract the loss of ABA by leaching from isolated embryos during imbibition (Suzuki et
al. 2000). However, whether this is synthesis from ABA precursors or re-synthesis from the
conjugated pool of ABA has not been examined in the work presented in Section 3.4, or the
work of Suzuki et al. (2000). Recent work in barley leaves has also highlighted the possible
role of conjugated ABA as a source of ABA (Dietz et al. 2000), not actual synthesis.
Whether the synthesis of ABA is from ABA precursors or whether it is reactivation by
hydrolysis of conjugated ABA to free ABA during imbibition requires clarification. Once
ABA elevation has been confirmed, further work is required to clarify if the higher
concentration of ABA is due to reduced catabolism, increased synthesis, or a balance
between re-activation and de-activation of ABA to conjugated ABA.

100
6.1.2 Sensitivity to ABA
Elevation of embryo ABA concentration during imbibition appears to explain, at least in
part, embryo dormancy and hence preharvest sprouting tolerance found here. In contrast to
dormant intact grains, embryo-half grains germinate rapidly i.e. within 48 hrs (Section 3.4
and 4.4; Flintham et al. 1998; Mares 1998; Suzuki et al. 2000; Walker-Simmons 1987),
possibly because they do not have or are not able to maintain a high ABA concentration
during imbibition. In dormant half grains of wheat application of exogenous ABA restores
the dormant phenotype probably by saturating the embryo with ABA. However, application
of ABA to embryo-half grains of non-dormant genotypes or after-ripened grain should also
saturate the embryo with ABA, but it does not result in a dormant phenotype and embryos
germinate. Hence, in addition to elevation of embryo ABA concentration, the results
presented in Section 3.4 and 4.4 indicate that ABA sensitivity must be involved as well as
high embryo ABA concentrations at 6 to 18 hrs during imbibition to result in dormancy. A
lack of germination inhibition of non-dormant grain with applied ABA has also been
reported in barley (Wang et al. 1995), but this has not been investigated in recent studies on
ABA regulation and control of dormancy in barley (Benech-Arnold et al. 2006; Jacobsen et
al. 2002; Millar et al. 2006) or Arabidopsis (Ali-Rachedi et al. 2004; Gubler et al. 2005;
Millar et al. 2006). Further work is required in wheat, barley and Arabidopsis to determine
the embryo ABA concentration in non-dormant grain to confirm that applied ABA leads to
higher embryo ABA during imbibition. If the ABA concentration in non-dormant embryos
with applied ABA is high and the grain still germinates, this would confirm that ABA
sensitivity is a component of dormancy that requires further work.
6.1.3 Seed coat
In wheat, there is a general and a specific seed coat effect on dormancy and hence
preharvest sprouting tolerance. The general seed coat effect occurs in all genotypes
regardless of dormancy, i.e. grain with the seed coat broken (like embryo-half grains or
isolated embryos) germinates more rapidly than intact grains. However in dormant
genotypes, inheritance studies have shown (Flintham 2000; Mares 1998) that there is
another specific component of the seed coat, which has a greater effect on dormancy than
the general seed coat effect, but this is only evident in genotypes with the embryo
dormancy component. One way to measure the specific seed coat effect is to compare the
germination of genotypes with and without the specific seed coat component (S with; s

101
without) and the embryo component (E with; e without). The ES genotype (e.g. DH 22) has
a germination index which is 0.15-0.10 lower (Table 1; Esperance 2004/05) than the Es
genotype (e.g. DH 56; Table 1); and this is typical of the additive effect the specific seed
coat has on the embryo component (Mares 1998). How the general or specific seed coat
component improves dormancy is unknown. Previous hypotheses have focused around the
physical restraint of water, oxygen and/or inhibitors but this work in wheat has largely been
inconclusive (King 1989; Mares 1998; Rathjen, J. Pers. Comm.). An alternative hypothesis
could be that the general and specific seed coat components in some way influences the
ABA elevation and / or sensitivity to ABA with the specific component having an effect for
longer than the general component. Further work on separating out the influences of the
embryo and the specific seed coat component requires the use of better genetic material,
possibly near isogenic genotypes for ES, Es, eS and es. Work with these genotypes could
then confirm if the additive effect of the general and specific seed coat components is due
to elevation of embryo ABA and /or ABA sensitivity.

10
2
Tabl
e 1.
Effe
cts o
f diff
eren
t mec
hani
sms o
f em
bryo
and
seed
coa
t com
pone
nts o
f dor
man
cy w
hich
influ
ence
the
germ
inat
ion
inde
x, i.
e.
dorm
ancy
phe
noty
pe a
t mat
urity
und
er v
aryi
ng e
nviro
nmen
tal c
ondi
tions
in w
heat
. A g
erm
inat
ion
inde
x of
1.0
repr
esen
ts n
on-d
orm
ant
and
0.0
dorm
ant.
E eq
uals
embr
yo se
nsiti
vity
alle
le, e
is n
o em
bryo
sens
itivi
ty a
llele
, S is
seed
coa
t alle
le, s
is n
o se
ed c
oat a
llele
.
Kat
anni
ng
Espe
ranc
e
2003
/04
cons
isten
t low
tem
pera
ture
2003
/04
cons
isten
t hig
h
tem
pera
ture
2004
/05
cons
isten
t hig
h
tem
pera
ture
2005
/06
cons
isten
t low
tem
pera
ture
2005
/06
cons
isten
t low
plu
s
40-5
0dpa
hig
h
tem
pera
ture
shoc
k
moi
stur
e
stre
ss
irrig
ated
m
oist
ure
stre
ss
irrig
ated
m
oist
ure
stre
ss
irrig
ated
m
oist
ure
stre
ss
irrig
ated
m
oist
ure
stre
ss
irrig
ated
DM
2001
E
S 0.
09
0.07
0.
06
0.35
0.
13
0.09
0.
02
0.03
0.
14
0.45
DH
22
E S
0.02
0.
13
0.04
0.
05
0.43
0.
45
DH
56
E s
0.17
0.
34
0.06
0.
02
0.41
0.
54
DH
45
e S
0.33
0.
35
0.22
0.
23
0.49
0.
67
Cun
derd
in
e s
0.52
0.
86
0.41
0.
66
0.65
0.
70
0.80
0.
66
0.84
0.
91
Pool
ed L
SD0.
050.
12

10
3
Relative embryo ABA concentration (%)
Dor
man
cy in
crea
sed
Con
sist
ent s
tres
s
ABA
ele
vatio
n
Dor
man
cy d
ecre
ased
Sudd
en s
tres
s
sens
itivi
ty ?
seed
coa
t int
egrit
y ?
Tim
e (d
ays
post
ant
hesi
s)
010
2030
4050
+
Gen
otyp
e +
En
viro
nmen
t
=
Phe
noty
pe a
t mat
urity
Con
ditio
ns d
urin
g gr
ain
fillin
g
Res
pons
e at
mat
urity
dur
ing
imbi
bitio
n
100
Non
-dor
man
t
Dor
man
t
Dor
man
t no
germ
inat
ion
06
1218
2430
3642
48
Tim
e (h
ours
of i
mbi
bitio
n)
Non
-dor
man
t ger
min
atio
n
AB
A el
evat
ion
+
Sens
itivi
ty
+
Seed
coa
t
Relative embryo ABA concentration (%)
Dor
man
cy in
crea
sed
Con
sist
ent s
tres
s
ABA
ele
vatio
n
Dor
man
cy d
ecre
ased
Sudd
en s
tres
s
sens
itivi
ty ?
seed
coa
t int
egrit
y ?
Tim
e (d
ays
post
ant
hesi
s)
010
2030
4050
+
Gen
otyp
e +
En
viro
nmen
t
=
Phe
noty
pe a
t mat
urity
Con
ditio
ns d
urin
g gr
ain
fillin
g
Res
pons
e at
mat
urity
dur
ing
imbi
bitio
n
100
Non
-dor
man
t
Dor
man
t
Dor
man
t no
germ
inat
ion
06
1218
2430
3642
48
Tim
e (h
ours
of i
mbi
bitio
n)
Non
-dor
man
t ger
min
atio
n
AB
A el
evat
ion
+
Sens
itivi
ty
+
Seed
coa
t
mat
erna
l driv
enem
bryo
driv
en
ES Es es
+G
E
Relative embryo ABA concentration (%)
Dor
man
cy in
crea
sed
Con
sist
ent s
tres
s
ABA
ele
vatio
n
Dor
man
cy d
ecre
ased
Sudd
en s
tres
s
sens
itivi
ty ?
seed
coa
t int
egrit
y ?
Tim
e (d
ays
post
ant
hesi
s)
010
2030
4050
+
Gen
otyp
e +
En
viro
nmen
t
=
Phe
noty
pe a
t mat
urity
Con
ditio
ns d
urin
g gr
ain
fillin
g
Res
pons
e at
mat
urity
dur
ing
imbi
bitio
n
100
Non
-dor
man
t
Dor
man
t
Dor
man
t no
germ
inat
ion
06
1218
2430
3642
48
Tim
e (h
ours
of i
mbi
bitio
n)
Non
-dor
man
t ger
min
atio
n
AB
A el
evat
ion
+
Sens
itivi
ty
+
Seed
coa
t
Relative embryo ABA concentration (%)
Dor
man
cy in
crea
sed
Con
sist
ent s
tres
s
ABA
ele
vatio
n
Dor
man
cy d
ecre
ased
Sudd
en s
tres
s
sens
itivi
ty ?
seed
coa
t int
egrit
y ?
Tim
e (d
ays
post
ant
hesi
s)
010
2030
4050
+
Gen
otyp
e +
En
viro
nmen
t
=
Phe
noty
pe a
t mat
urity
Con
ditio
ns d
urin
g gr
ain
fillin
g
Res
pons
e at
mat
urity
dur
ing
imbi
bitio
n
100
Non
-dor
man
t
Dor
man
t
Dor
man
t no
germ
inat
ion
06
1218
2430
3642
48
Tim
e (h
ours
of i
mbi
bitio
n)
Non
-dor
man
t ger
min
atio
n
AB
A el
evat
ion
+
Sens
itivi
ty
+
Seed
coa
t
mat
erna
l driv
enem
bryo
driv
en
ES Es es
+G
E
Fig.
1. C
once
ptua
l mod
el o
f the
con
trol o
f dor
man
cy in
whe
at b
y en
viro
nmen
tal c
ondi
tions
dur
ing
grai
n fil
ling
(a) i
nflu
enci
ng
dorm
ancy
at m
atur
ity (b
) thr
ough
AB
A e
leva
tion,
sens
itivi
ty o
r see
d co
at in
tegr
ity
. Th
e tim
ing
of th
e tra
nsiti
on fr
om th
ese
stag
es c
an b
e m
oved
by
the
envi
ronm
enta
l con
ditio
ns. S
hade
d ar
eas r
epre
sent
the
perio
d du
ring
embr
yo d
evel
opm
ent w
hen
envi
ronm
ent c
an in
fluen
ce d
orm
ancy
. Dor
man
cy in
crea
sed
by c
onsi
sten
t stre
ss a
t Kat
anni
ng in
200
3 an
d by
app
lied
AB
A
(Suz
uki e
t al.
2000
) at 1
5-30
dpa
.
Dor
man
cy d
ecre
ased
at 3
0-50
dpa
poss
ibly
by
redu
ced
embr
yo se
nsiti
vity
or s
eed
coat
inte
grity
cau
sed
by h
eat s
hock
s at E
sper
ance
in 2
005,
or b
y cl
eavi
ng w
ith e
xces
s wat
er a
t Esp
eran
ce in
200
3, o
r at
Kat
anni
ng in
200
3. G
ener
alis
ed ti
me
cour
se o
f AB
A a
ccum
ulat
ion
unde
r opt
imum
con
ditio
ns a
dapt
ed fr
om K
ing
(199
3) a
nd
Suzu
ki e
t al.
(200
0) c
ompa
red
to F
ig. 3
Sec
tion
2.2.

104
6.2 Environment
The environmental conditions during grain filling and maturation have a significant though
contrasting effects on the level of dormancy at maturity. During grain filling consistent high
temperatures and moisture stress induced a dormant phenotype in a genotype, which was
typically non-dormant. In contrasts, during grain maturation high temperatures at 30-50
days post anthesis reduced the level of dormancy in all genotypes, confirming previous
proposals from controlled environment work (King 1993) and correlation analyses of field
trials and weather variables over several seasons (Strand 1989b; Strand 1990). In addition,
during grain maturation under both controlled and field conditions with excess water supply
from rainfall or irrigation (without wetting the heads), a reduction in dormancy of all
genotypes also occured. Figure 1 presents a hypothetical model of the time periods during
grain filling when the environment appears to influence dormancy. The influence of
environment on dormancy may be through one or all of the three aspects, which control
dormancy; (a) embryo ABA elevation, (b) sensitivity or (c) the factors associated with the
integrity of the seed coat.
During grain filling ABA sensitivity appears to be affected by the environment. The
dormancy induced in the normally non�dormant Cunderdin by the consistent high
temperature and moisture stress during grain filling is associated with an increase in the
sensitivity of the embryo to germination inhibition by applied ABA. Sensitivity of embryos
to applied ABA in dormant genotypes is highest under conditions that also produced the
most dormancy, suggesting that the effect of environment on sensitivity appears to occur
during grain filling (15-30 dpa; Fig. 1).
Walker-Simmons and Sesing (1990) were among the first of several authors who also
observed a strong correlation between grain filling conditions which induced high levels of
dormancy and increased sensitivity to applied ABA. Several studies with different wheat
genotypes have found that moisture stress during the early stages of grain filling can
increase embryo ABA concentrations (Bhaglal et al. 1999; Goldbach et al. 1977; Haeder
and Beringer 1981; Westgate et al. 1996). In addition Suzuki et al. (2000) found in wheat
with isolated stems and heads in nutrient solution that plants fed with ABA early in grain
filling (at 15-50 and 20-50 dpa) but not later during grain filling (25-50 dpa) have more

105
dormant grain at maturity with increased sensitivity to applied ABA. So either synthesized
or applied ABA at this stage may increase dormancy. In barley, ABA applied during grain
filling also increases dormancy (Takahashi 1980). A similar response may have occurred
here in Cunderdin with moisture stress and consistent high temperature throughout grain
filling inducing an increase in whole plant concentrations of ABA similar to applied ABA.
The maternal or plant ABA may then be transferred to the developing embryo and
influence dormancy. The plant ABA during grain maturation may represent an important
mechanism by which the dormancy phenotype can be affected by the environment during
grain filling at 15-30 dpa.
Further work is required to confirm if embryo ABA elevation and/or sensitivity, induced in
a non-dormant genotype such as Cunderdin, is related to the level of stress during grain
filling. If it is, the environmental influence on dormancy could be an ideal model in which
to study the control of dormancy in identical genetic backgrounds because the same
genotype could be manipulated to have two phenotypes. This may overcome some of the
problems associated with comparing mature and after-ripened grain or different genotypes
with contrasting dormant and non-dormant grain.
In contrast, during grain maturation heat shocks reduce dormancy, and one possible
hypothesis is that when these occur at 30-50 dpa they may reduce endogenous ABA
concentrations during imbibition. Work in barley has shown that the ability of the embryo
to maintain a high ABA concentration during imbibition is related to the ability of the seed
coat to maintain an oxygen deficiency (Benech-Arnold et al. 2006) since oxygen is required
for the breakdown of ABA. In wheat, it is less likely that oxygen deficiency imposed by the
seed coat is causing elevation of ABA, as outlined previously (Section 6.1.3). Regardless of
how ABA concentrations remain high during imbibition, the general seed coat has an
additive effect in all genotypes, and this additive effect may be what is compromised by the
heat shocks at 30-50 dpa. As all genotypes whether, dormant partially dormant or non-
dormant were affected (Table 1), it is likely the general seed coat effect, not the specific
was responsible. Symons et al. (1983) also found pre-mature drying of grain promoted
germination similar to the effect of removing the pericarp, i.e. removal of the additive
effect of the general seed coat component. A dry finish can lead to cracks and fissures in
the grain which can result in different rates of water uptake and loss (Pool and Paterson

106
1958a). Over twenty years ago Woodbury and Wiebe (1983) originally proposed a role for
cracks in the seed coat in the control of germination. An extension of this concept is that the
presence of cracks in the seed coat may be modulated by the environment during grain
maturation. The cracks and fissures in the seed coat may then lead to a reduced additive
effect of the seed coat, although further work is required to confirm this hypothesis.
Other conditions during grain maturation, such as excess water supply, which lead to a
decrease in dormancy may have also lead to cracks and fissures in the seed coat. Cleaving,
black point and fungal infection was observed under consistent high temperature and
irrigation and under ambient conditions with 50 mm of water supply (without wetting the
heads) in the late grain filling stages (Section 4.4), and was associated with a loss of
dormancy in the more dormant genotypes. Visually cleaved, black point and fungal infected
grains were omitted from germination tests, however the possibility remains that lower
levels of these symptoms were present and not avoided. With cleaving for example smaller
fissures may have still been present in the crease. Cleaving of grain also produces cracks
and fissures in the seed coat and has been reported after a period of high temperature
(36/31°C) at 6-10 dpa (Tashiro and Wardlaw 1990) where the number of seed coat cells is
set, after this; excess grain filling then splits the seed coat. Previous work has determined
that water movement during imbibition into the grain can occur through cracks in the beard
end of the pericarp (Woodbury and Wiebe 1983). Other unpublished recent work with
Magnetic Resonance Micro-Imaging has shown that water movement into the grain is
through the micropyle, with hydration of the embryo within 2 hours, the crease and
aleurone within 6 hours, and the seed coat after that, with the endosperm remaining un-
hydrated for at least 18 hours (Rathjen, J. Pers.comm.). Disruption of the seed coat may
result in a different hydration pattern and hence a different transfer of solutes to or from the
embryo. Further work is required during grain maturation to determine if (i) the differences
in dormancy due to excess water resulting in cleaving, or heat shocks during the 30-50 dpa
period lead to different concentrations of ABA during imbibition or differences in
sensitivity, and then (ii) how the concentrations of ABA are affected, possibly by changes
in the seed coat integrity through cracks and fissures.

107
6.3 Morphological traits
Screening for preharvest sprouting tolerance based on falling number after a rainfall event
has limitations, since the time from maturity to a particular rainfall event is not the same
between genotypes differing in maturity this means the after-ripening period is not
consistent between genotypes differing in maturity when this method of screening is used.
Despite this limitation, this method highlighted tolerance to preharvest sprouting in
genotypes without dormancy. For example the awnless, EGA Eagle Rock non-dormant (es,
germination index typically 0.70-0.80) had a falling number > 250 s, similar to the dormant
genotype DH 56 (Es, germination index typically <0.20) after 70 mm of rainfall (Section
5.4 Fig. 2). Under the same conditions other awned genotypes with a similar dormancy
level as EGA Eagle Rock had a falling number <100 s.
The ability of the awnless traits and other head characteristics to protect a crop from
preharvest sprouting has been identified earlier (King and Richards 1984) and proposed to
be associated with lower water uptake and hence lower sprouting (King and Richards 1984;
Pool and Paterson 1958b). Work presented in Section 5.4 indicates there is a measureable
level of preharvest sprouting tolerance from morphological traits compared to embryo and
seed coat imposed dormancy in this target environment. However, it has still not been
determined if the level of tolerance is due to the presence of awns or some other trait
associated with them (King and Richards 1984). Spear types, for example also tended to
have a higher falling number (> 300 s) after preharvest sprouting than other genotypes with
a similar level of dormancy (falling number <250 s; Section 5.4, Fig. 3). These spear types
and awnless genotypes were more difficult to thresh, mechanically and by hand, than
typically awned genotypes, with a greater proportion of un-threshed grains. This may be
correlated with the ability of the grain to imbibe water during rainfall events. Further work
is required to clarify (i) the exact means by which awnless types have a lower water uptake
rate and (ii) the source of preharvest sprouting tolerance reported in the awned Spear types.
It remains unclear whether this is due to tighter glumes, less gaping lemma and palea or
other additional factors such as germination inhibitors in bran and husk extracts as
previously highlighted in wheat (Himi et al. 2002).

108
Using the awnless trait for preharvest sprouting tolerance has a cost with slower grain
drying rates, reduced threshability and reduced adaptation to grain filling under stress. Once
the head is wet, awnless genotypes dry slower than awned genotypes (King and Richards
1984; Pool and Paterson 1958b). Awnless genotypes are also usually harder to thresh than
awned genotypes, which may also reduce the harvesting time available per day. Yield
losses occur with delayed harvest at a rate of 0.50 % per day at Esperance in Western
Australia (Bolland 1984). A slower grain drying rate and reduced harvestability may mean
the crop remains in the paddock for longer and hence is at greater risk of preharvest
sprouting and reduced yield. Under optimum growth conditions, there is usually no yield
penalty associated with awnlessness, however under stressful conditions awns can improve
yield and grain size compared to awnless genotypes (Atkins and Norris 1955; Motzo and
Giunta 2002). Hence, breeding preharvest sprouting tolerant awnless genotypes may result
in lower yield in some seasons. While many areas of Western Australia are associated with
high temperatures and moisture stress during grain filling, preharvest sprouting in Western
Australia is not usually associated with areas of the wheat belt subject to the extremes of
high temperature and moisture stress. These concerns could be alleviated by finding (i) an
awnless genotype, which is free threshing but still maintains the preharvest sprouting
tolerance, or (ii) an awned genotype, which is inherently free threshing, with the awnless
mechanism of tolerance.
6.4 Implications for screening for preharvest sprouting tolerance
with dormancy
Wheat genotypes with strong dormancy (ES or Es) were consistently preharvest sprouting
tolerant, i.e. falling number > 300 s; genotypes with partial dormancy (eS or es) were
intermediately sprouting tolerant, i.e. falling number <300 s but >150 s; and genotypes
without dormancy (es) were generally preharvest sprouting susceptible i.e. falling number
<150 s except for genotypes with specific morphological traits (Chapter 5, Fig. 3). As the
level of dormancy in all genotypes was influenced by the environmental conditions during
grain filling, preharvest sprouting tolerance was also influenced by the environmental
conditions during grain filling.
In terms of the initial problem concerning the level of dormancy required for regions
differing in preharvest sprouting risk (Chapter 2 Section 2.5). Strong dormancy (ES or Es)

109
is required for coastal regions such as Esperance prone to late season rainfall. However the
embryo component alone (Es) is probably sufficient for the majority of situations in this
environment. The addition of the specific seed coat component with the embryo component
(i.e. ES) removes an even larger part of the seasonal fluctuations in dormancy, but the
specific seed coat component is only required for the most severe sprouting seasons. Partial
dormancy is not sufficient for sprouting prone coastal regions as it is not stable between
seasons and did not give complete tolerance (falling number >300 s; Chapter 5 Fig. 2). It
was however substantially better than non-dormant genotypes, and for this reason partial
dormancy may be sufficient for preharvest sprouting tolerance in regions which are not
prone to preharvest sprouting, such as the central wheat belt of Western Australia.
Combining morpholigial traits such as awnlessness with dormancy may be a sound strategy
to improve preharvest sprouting tolerance in some situations despite the limitations of the
awnless trait. The extra tolerance from awnlessness in genotypes with partial dormancy, for
example, may remove the seasonal variation in preharvest sprouting tolerance in these
genotypes when awned. Further work is required to clarify if the seasonal variation in
partial dormancy preharvest sprouting tolerance is still present in combination with
awnlessness. The knowledge that the embryo component alone, (Es) or the possible
addition of awnlessness with partial dormancy should be sufficient for this target
environment provides a confident strategy to improve the preharvest sprouting tolerance of
commercial wheat genotypes targeted for the small but productive Western Australian
sprouting prone regions (40%, of Western Australia�s current season wheat production
came from the Esperance Port Zone in 2006/07 season due to severe drought in the rest of
Western Australia).

110
References Cited Ali-Rachedi S, Bouinot D, Wagner M, Bonnet M, Sotta B, Grappin P and Jullien M (2004) Changes in endogenous abscisic acid levels during dormancy release and maintenance of mature seeds: studies with the Cape Verde islands ecotype, the dormant model of Arabidopsis thaliana. Planta 219, 479-488. Appleford NEJ and Lenton JR (1997) Hormonal regulation of α-amylase gene expression in germinating wheat (Triticum aestivum) grains. Physiologia Plantarum 100, 534-542. Atkins ID and Norris MJ (1955) The influence of awns on yield and certain morphological characters of wheat. Agronomy Journal 47, 218-220. Auld AS and Paulsen GM (2003) Effects of drought and high temperature during maturation on preharvest sprouting tolerance of hard white winter wheat. Cereal Research Communications 31, 169-176. Australian Wheat Board (2003-2004) AWB wheat receival standards 2003-2004. (Australian Wheat Board Melbourne) Barthe P, Garello G, Bianco-Trichant J and Le Page-Degivry MT (2000) Oxygen availability and ABA metabolism in Fagus sylvatica seeds. Plant Growth Regulation 30, 185-191. Belderok B (1968) Seed dormancy problems in cereals. Field Crop Abstracts 21, 203-211. Benech-Arnold RL, Giallorenzi MC, Frank J and Rodriguez MV (1999) Termination of hull-imposed dormancy in developing barley grains is correlated with changes in embryonic ABA levels and sensitivity. Seed Science Research 9: 39-47. Benech-Arnold RL, Gualano N, Leymarie J, Come D and Corbineau F (2006) Hypoxia interferes with ABA metabolism and increases ABA sensitivity in embryos of dormant barley grains. Journal of Experimental Botany 57: 1423-1430. Benech-Arnold RL (2002) Bases of preharvest sprouting resistance in barley: Physiology, molecular biology and environmental control of dormancy in the barely grain. In 'Barely Science. Recent advances from molecular biology to agronomy of yield and quality. (Ed. GA Slater) pp. 481-502. (Food Products Press: New York) Bhaglal P, Singh P and Bhullar SS (1999) Drought-induced increase in wheat germ agglutinin (WGA) accumulation in developing wheat embryos appears to be independent of ABA. Australian Journal of Plant Physiology 26, 787-791.

111
Biddulph TB, Mares DJ, Plummer JA and Setter TL (2005) Drought and high temperature increases preharvest sprouting tolerance in a genotype without grain dormancy. Euphytica 143, 277-283. Biddulph TB, Gubler F, Poole AT, Plummer JA, Setter TL and Mares DJ (2007a) Abscisic acid concentration of wheat (Triticum aestivum L.) embryos in relation to expression of grain dormancy. In submission. Biddulph TB, Plummer JA, Setter TL and Mares DJ (2007b) Seasonal environmental conditions influence dormancy and subsequent preharvest sprouting tolerance in wheat (Triticum aestivum L.). Field Crops Research In review. Biddulph TB, Plummer JA, Setter TL and Mares DJ (2007c) Influence of high temperature and terminal moisture stress on dormancy in wheat (Triticum aestivum L.). Field Crops Research In press. Bolland MDA (1984) Grain losses due to delayed harvesting of barley and wheat. Australian Journal of Experimental Agriculture and Animal Husbandry 24, 391-393. Bureau of Meteorology (1956-2006) Historical rainfall records, Esperance Downs Research Station and Great Southern Agricultural Research Institute. (Bureau of Meteorology, Perth) Conner RL, Carefoot JM, Bole JB, Kozub GC (1992) The effect of nitrogen fertilizer and irrigation on black point incidence in soft white spring wheat. Plant and Soil 140, 41-47. Conner RL, Davidson JGN (1988) Resistance in wheat to black point caused by Alternaria alternata and Cochlibolus sativus. Canadian Journal of Plant Science 68, 351-359. Chono M, Honda I, Shinoda S, Kushiro T, Kamiya Y, Nambara E, Kawakami N, Kaneko S and Wantabe Y (2006) Field studies on the regulation of abscisic acid content and germinability during grain development of barley: molecular and chemical analysis of pre-harvest sprouting. Journal of Experimental Botany 57, 2421-2434. Del Cima R, D'Antuono MFD and Anderson WK (2004) The effects of soil type and seasonal rainfall on the optimum seed rate for wheat in Western Australia. Australian Journal of Experimental Agriculture 44, 585-594.
DePauw RM, McCaig TN, Mares DJ, Brennan P, Henry RJ, King R, McEwan JM and Gordon I (1989) Genetic variation for length of dormancy period, sprouting resistance and α-amylase content in wheat. In '5th International Symposium on Preharvest Sprouting in Cereals'. USA. (Eds K Ringlund, E Mosleth and DJ Mares) pp. 206-212. (Westview Press Inc. Boulder Co) DePauw RM, McCaig TN, Mares DJ, Brennan P, Henry RJ, King R, McEwan JM and Gordon I (1989) Interrelationships among assays for germination of kernels from threshed spikes, unthreshed spikes and alpha-amylase in wheat. In '5th International Symposium on

112
Preharvest Sprouting in Cereals'. USA. (Eds K Ringlund, E Mosleth and DJ Mares) pp. 195-205. (Westview Press Inc. Boulder Co) DePauw RM and McCraig TN (1991) Components of variation, heritabilities and correlations for indices of sprouting tolerance and seed dormancy in Triticum spp. Euphytica 52: 221-229. DePauw RM, McCaig TN, Baker RJ, Clarke JM (1993) Constructing a Sprouting Tolerance Index. In 'Sixth International Symposium on Pre-harvest Sprouting in Cereals'. USA. (Eds MK Walker-Simmons, JL Ried) pp. 47�53. (Westview Press, Boulder, Co). Derera NF (1982) The harmful harvest rain: Farrer memorial oration, 1981. The Journal of the Australian Institute of Agricultural Science 1982, 67-75. Dewdney SJ and McWha JA (1978) Abscisic acid and the movement of photosynthetic assimilates towards developing wheat (Triticum aestivum L.) grains. Z. Pflanzenphysiol 92, 183-186. Dietz K-J, Sauter A, Wichert K, Mesdaghi D and Hartung W (2000) Extracellular B-glucosidase acitivity in barley involved in the hydrolysis of ABA glucose conjugate in leaves. Journal of Experimental Botany 51, 937-944.
Fenner M (1991) The effects of the parent environment on seed germinability. Seed Science Research 1, 75-84. Feurtado JA, Ambrose SJ, Cutler AJ, Ross ARS, Abrams SR and Kermode AR (2004) Dormancy termination of western white pine (Pinus monticola Dougl. Ex D. Don) seeds is associated with changes in abscisic acid metabolism. Planta 218, 630-639. Fernandez MR, DePauw RM, Clarke FR, Fox SL (1998) Discoloration of wheat kernels by Pyrenophora tritici-repentis. Canadian Journal of Plant Pathology 20, 380-383. Finch-Savage WE and Leubner-Metzger G (2006) Seed dormancy and the control of germination. New Phytologist 171, 501-523.
Finkelstein RR, Gampala SSL and Rock CR (2002) Abscisic acid signaling in seeds and seedlings. The Plant Cell Supplement: s15-s45. Flintham JE (2000) Different genetic components control coat-imposed and embryo-imposed dormancy in wheat. Seed Science Research 10, 43-50. Flintham JE, Adlam R, Bassoi M, Holdsworth M and Gale MD (2002) Mapping genes for resistance to sprouting damage in wheat. Euphytica 126, 39-45. Flintham JE, Adlam R and Gale MD (1998) Seedcoat and embryo dormancy in wheat. In 'Eighth International Symposium on Preharvest Sprouting in Cereals.' Detmold, Germany. (Ed. D Weipert) pp. 67-76. (Association for Cereal Science Research)

113
Fox SL, Fernandez MR, DePauw RM (2003) Red Smudge infection modifies sprouting response in four wheat lines. Canadian Journal of Plant Science 83, 163-169. Frey A, Godin B, Bonnet M, Sotta B and Marion-Poll A (2004) Maternal synthesis of abscisic acid controls seed development and yield in Nicotiana plumbaginifolia. Planta 218, 958-964. Garello G, Holappa LD, Abrams GD and Abrams SR (1997) ABA metabolites induce group 3 LEA mRNA and inhibit germination in wheat. Physiologia Plantarum 100: 474-480. Garello G and Le Page-Degivry MT (1999) Evidence for the role of abscisic acid in the genetic and environmental control of dormancy in wheat (Triticum aestivum L.). Seed Science Research 9, 219-226. Garlinge J (2005) '2005 crop variety sowing guide for Western Australia.' (Department of Agriculture Western Australia: South Perth) Goldbach H, Goldbach E and Michael G (1977) Transport of abscisic acid from leaves to grains in wheat and barley plants. Naturwissenschaften 64, 488-489. Gomez-Cadenas A, Verhey SD, Holappa LD, Shen Q, Ho DTH and Walker-Simmons MK (1999) An abscisic acid-induced protein kinase, PKABA1, mediates abscisic acid-suppressed gene expression in barley aleurone layers. Proceedings National Acadamey Science USA 96, 1767-1772. Gooding MJ, Thompson AJ, Collingborn FMB, Smith SP, Davies WP (1993) Blackpoint on wheat grain: Influences of cultivar, managment and season on symptom severity. Aspects of Applied Biology 36, 391-396. Gordon AG (1971) The germination resistance test-A new test for measuring germination quality of cereals. Canadian Journal of Plant Science 51, 181-183. Gordon IL, Balaam LN and Derera NF (1979) Selection against sprouting damage in wheat. II Harvest ripeness, grain maturity and germinability. Australian Journal of Agricultural Research 30, 1-17. Grappin P, Bouinot D, Sotta B, Miginiac E and Jullien M (2000) Control of seed dormancy in Nicotiana plumbaginifolia: post-imbibition abscisic acid synthesis imposes dormancy maintenance. Planta 210, 279-285.
Grappin P, Bouinot D, Sotta B, Miginiac E and Jullien M (2004) Control of seed dormancy in Pinus monticola: post-imbibition abscisic acid metabolism. Planta 218: 630-639. Gubler F, Millar AA and Jacobsen JV (2005) Dormancy release, ABA and pre-harvest sprouting. Current Opinion in Plant Biology 8, 183-187.

114
Green LS, Faergestad EM, Poole AT and Chandler PM (1997) Grain development mutants of barley. Plant Physiology 114, 203-212. Haeder HE and Beringer H (1981) Influence of potassium nutrition and water stress on the content of abscisic acid in grains and flag leaves of wheat during grain development. Journal of the Science of Food and Agriculture 32, 552-556. Hagberg S (1960) A rapid method for determining alpha-amylase activity. Cereal Chemistry 37, 218-222. Hagberg S (1961) Note on a simple rapid method for determining alpha-amylase activity. Cereal Chemistry 38, 202-203. Hagemann MG and Ciha AJ (1987) Environmental x genotype effects on seed dormancy in wheat. Agronomy Journal 79, 192-196. Hagemann Wiedenhoeft M, Chevalier P and Walker-Simmons M (1988) Field studies on abscisic acid and embryonic germinability in winter wheat. Field Crops Research 18, 271-278. Hendershot KL (1992) Induction temperature of heat-shock protein synthesis in wheat. Crop Science 32, 256-261. Henson IE (1985) Dependence of abscisic acid accumulation in leaves of pearl millet (Pennistum americanum [L] Leeke) on rate of development of water stress. Journal of Experimental Botany 36, 1232-1239. Himi E, Mares DJ, Yanagisawa A and Noda K (2002) Effect of grain colour gene (R) on grain dormancy and sensitivity of the embryo to abscisic acid (ABA) in wheat. Journal of Experimental Botany 53, 1569-1574. Hucl P (1995) Divergent selection for sprouting resistance in spring wheat. Plant Breeding 114, 199-204. Humphreys DG and Noll J (2002) Methods for characterization of preharvest sprouting resistance in a wheat breeding program. Euphytica 126, 61-65. Iglesias RG and Babiano MJ (1997) Endogenous abscisic acid during the germination of chick-pea seeds. Physiologia Plantarum 100, 500-504. Jacobsen JV, Pearce DW, Poole AT, Pharis RP and Mander LN (2002) Abscisic acid, phaseic acid and giberellin contents associated with dormancy and germination in barley. Physiologia Plantarum 115, 428-441. Johnson RR, Wagner RL, Verhey SD and Walker-Simmons M (2002) The abscisic acid-responsive kinase PKABA1 interacts with a seed-specific abscisic acid response element- binding factor, TaABF, and phosphorylates TaABF peptide sequences. Plant Physiology 130, 837-846.

115
Karssen CM, Brinkhorst-van der Swan DLC, Breekland DE and Koornneef M (1983) Induction of dormancy during seed development by endogenous abscisic acid: studies on abscisic acid deficient genotypes of Arabidopsis thaliana (L.) Heynh. Planta 157, 158-165. Kato K, Nakamura W, Tabiki T, Miura H and Sawada S (2001) Detection of loci controlling seed dormancy on group 4 chromosomes of wheat and comparative mapping with rice and barley genomes. Theoretical and Applied Genetics 102, 980-985. Kermode AR (2005) Role of abscisic acid in seed dormancy. Journal of Plant Growth Regulation 24, 319-344. Kettlewell PS and Cashman MM (1997) Alpha-amylase activity of wheat grain from crops differing in grain drying rate. Journal of Agricultural Science Cambridge 128, 127-134. King RW (1976) Abscisic acid in developing wheat grains and its relationship to grain growth and maturation. Planta 132, 43-51. King RW (1979) Abscisic acid synthesis and metabolism in wheat ears. Australian Journal of Plant Physiology 6, 99-108. King RW (1984) Water uptake in relation to pre-harvest sprouting damage in wheat: grain characteristics. Australian Journal of Agricultural Research 35, 337-345. King RW (1989) Physiology of Sprouting Resistance. In 'Preharvest Field Sprouting in Cereals' pp. 27-60. (CRC Press: Boca Raton, Florida) King RW (1992) Genotypic and environmental effects on dormancy and in-ear sprouting in wheat. In 'Pre-harvest Sprouting in Cereals 1992'. Washington. (Eds MK Walker-Simmons and JL Ried) pp. 319-328. (American Association of Cereal Chemists) King RW (1993) Manipulation of grain dormancy in wheat. Journal of Experimental Botany 44, 1059-1066. King RW and Richards RA (1984) Water uptake in relation to pre-harvest sprouting damage in wheat: ear characteristics. Australian Journal of Agricultural Research 35, 327-336. King RW and Wettstein-Knowles PV (2000) Epicuticular waxes and regulation of ear-wetting and pre-harvest sprouting in barley and wheat. Euphytica 112, 157-166. Koshkin EI and Tararina VV (1990) Differences in source-sink ratios in wheat and their relationship to grain yield and content of abscisic acid. Plant Physiological Biochemistry 28, 609-616. Lehmensiek A, Campbell AW, Williamson PM, Michalowwitz M, Sutherland MW and Daggard GE (2004) QTLs for black-point resistance in wheat and the identification of potential markers for use in breeding programmes. Plant Breeding 123, 410-416.

116
Lenton JR, Appleford NEJ and Croker SJ (1994) Gibberellins and α-amylase gene expression in germinating wheat grains. Plant Growth Regulation 15, 261-270. Leung J and Giraudat J (1998) Abscisic acid signal transduction. Annual Review of Plant Physiology and Plant Molecular Biology 49, 199-222. Li B and Foley ME (1997) Genetic and molecular control of seed dormancy. Trends in Plant Science 2, 384-389. Li CD, Tarr A, Lance RC Harasymow S, Uhlmann J, Wescot S, Young KJ, Grime CR, Cakir M, Broughton S and Appels R (2003) A major QTL controlling seed dormancy and pre-harvest sprouting/grain α-amylase in two-rowed barley (Hordeum vulgare L.). Australian Journal of Agricultural Research 54, 1303-1313. Lohwasser U, Roder MS and Borner A (2005) QTL mapping of the domestication traits pre-harvest sprouting and dormancy in wheat (Triticum aestivum L.). Euphytica 143, 247-249. Lorenz K (1986) Effects of blackpoint on grain composition and baking quality of New Zealand wheat. New Zealand Journal of Agricultural Research 29, 711-718. Lunn GD, Kettlewell PS, Major BJ and Scott RK (2002) Variation in dormancy duration in the U.K. wheat cultivar Hornet due to environmental conditions during grain development. Euphytica 126, 89-97. Lunn GD, Scott RK, Kettlewell PS and Major BJ (1998) Evaluation of the Belderok temperature-dormancy relationship in the UK. In 'Eighth International Symposium on Preharvest Sprouting in Cereals.' Detmold, Germany. (Ed. D Weipert) pp. 190-198. (Association for Cereal Science Research Detmold) Mares DJ (1983a) Investigation of the pre-harvest sprouting damage resistance mechanisms in some Australian white wheats. In 'Third International Symposium on Pre-Harvest Sprouting in Cereals'. Canada. (Eds JE Kruger and DE LaBerge) pp. 59-65. (Westview Press Boulder USA) Mares DJ (1983b) Preservation of dormancy in freshly harvested wheat grain. Australian Journal of Agricultural Research 34, 33-38. Mares DJ (1984) Temperature dependence of germinability of wheat (Triticum aestivum L.) grain in relation to pre-harvest sprouting. Australian Journal of Agricultural Research 35, 115-128. Mares DJ (1989) Preharvest sprouting damage and sprouting tolerance: assay methods and instrumentation. In 'Preharvest field sprouting in cereals'. (Ed. NF Derera) pp. 129-170. (CRC Press: Boca Raton, Florida) Mares DJ (1993) Pre-harvest sprouting in wheat. I. Influence of cultivar, rainfall and temperature during grain ripening. Australian Journal of Agricultural Research 44, 1259-1272.

117
Mares DJ (1996) Genetic studies of sprouting tolerance in red and white wheats. In 'Seventh International Symposium on Pre-Harvest Sprouting in Cereals 1995'. Osaka, Japan. (Eds K Noda and DJ Mares) pp. 21-29. (Centre for Academic Societies Japan) Mares DJ (1998) The seed coat and dormancy in wheat grains. In 'Eighth International Symposium on Preharvest Sprouting in Cereals'. Detmold, Germany. (Ed. D Weipert) pp. 77-81. (Association for Cereal Science Research) Mares DJ and Mrva K (2001) Mapping quantitative trait loci associated with variation in grain dormancy in Australian wheat. Australian Journal of Agricultural Research 52, 1257-1265. Mares DJ, Mrva K, Cheong J, Williams K, Watson B, Storlie E, Sutherland M and Zou Y (2005) A QTL located on chromosome 4A associated with dormancy in white- and red-grained wheats of diverse origin. Theoretical and Applied Genetics 111, 1357-1364. Metz N and Newman C (2006) 'The WA guide to high moisture grain managment ' (Department of Agriculture and Food Western Australia: Perth), In press. Millar AA, Jacobsen JV, Ross JJ, Helliwell CA, Poole AT, Scofield G, Reid JB and Gubler F (2006) Seed dormancy and ABA metabolism in Arabidopsis and barley: the role of ABA 8'-hydroxylase. The Plant Journal 45, 942-954. Miyamoto T, Tolbert NE and Everson EH (1961) Germination inhibitors related to dormancy in wheat seeds. Plant Physiology 36, 739-746. Morris CF, Mueller DD, Faubion JM and Paulsen GM (1988) Identification of L-Tryptophan as an endogenous inhibitor of embryo germination in white wheat. Plant Physiology 88, 435-440. Moschini RC, Sisterna MN and Carmona MA (2006) Modelling of wheat black point incidence based on meteorological variables in the southern Argentinean Pampas region. Australian Journal of Agricultural Research 57, 1151-1156. Motzo R and Giunta F (2002) Awnedness affects grain yield and kernal weight in near-isogenic lines of durum wheat. Australian Journal of Agricultural Research 53, 1285-1293. Nakamura S and Toyama T (2001) Isolation of a VP1 homologue from wheat and analysis of its expression in embryos of dormant and non-dormant cultivars. Journal of Experimental Botany 52, 875-876. Naversero EP, Baun LC and Juliano BO (1975) Grain dormancy, peroxidase activity and oxygen uptake in Oryza sativa. Phytochemistry 14, 1899-1902. Nielsen MT, McCrate AJ, Heyne EG and Paulsen GM (1984) Effect of weather variables during maturation on preharvest sprouting of hard white winter wheat. Crop Science 24, 779-782.

118
Noda K, Kawabata C and Kawakami N (1994) Response of wheat grain to ABA and imbibition at low temperature. Plant Breeding 113, 53-57. Nyachiro JM, Clarke FR, DePauw RM, Knox RE and Armstrong KC (2002) Temperature effects on seed germination and expression of seed dormancy in wheat. Euphytica 126, 123-127. Passioura JB (2006) The perils of pot experiments. Functional Plant Biology 33, 1075-1079. Pool M and Paterson FL (1958a) Moisture relations in soft red winter wheats I. Varietal differences and delayed harvest effects. Agronomy Journal 50, 153-157. Pool M and Paterson FL (1958b) Moisture relations in soft red winter wheats II. Awned versus awnless and waxy versus nonwaxy glumes. Agronomy Journal 50, 158-160. Ramaih S, Guedira M and Paulsen GM (2003) Relationship of indoleacetic acid and tryptophan to dormancy and preharvest sprouting of wheat. Functional Plant Biology 30, 939-945. Rasmussen RD, Hole D, Hess JR and Carman JG (1997) Wheat kernel dormancy and + abscisic acid level following exposure to fluridone. Journal of Plant Physiology 150, 440-445. Reddy LV, Metzger RJ and Ching TM (1985) Effect of temperature on seed dormancy of wheat. Crop Science 25, 455-458. Rees RG, Martin DJ and Law DP (1984) Black point in bread wheat: effects on quality and germination, and fungal associations. Australian Journal of Experimental Agriculture and Animal Husbandry 24, 601-605. Ried JL and Walker-Simmons M (1990) Synthesis of abscisic acid-responsive, heat -stable proteins in embryonic axes of dormant wheat grain. Plant Physiology 93, 662-667. Rock CR (2000) Pathways to abscisic acid-regulated gene expression. New Phytologist 148, 357-396. Rodriguez MV, Margineda M, F. G-MJ, Insausti P and Benech-Arnold RL (2001) Predicting preharvest sprouting suscetibility in barley: A model based on temperature during grain filling. Agronomy Journal 93, 1071-1079. Romagosa I, Prada D, Moralejo MA, Munoz P, Casas AM, Swanston JS and Molina-Cano JL (2001) Dormancy, ABA content and sensitivity of a barley mutant to ABA application during seed development and after ripening. Journal of Experimental Botany 52, 1499-1506. Ross CW (1992) Hormones and growth regulators: cytokinins, ethylene, abscisic acid and other compounds. In 'Plant Physiology'. (Eds FB Salisbury and CW Ross) pp. 382-406. (Wadsworth Publishing Company: Belmont, California)

119
Qin ZZ (1990) Changes and endogenous abscisic acid levels in rice embryo and endosperm and association with development and germination. Acta Bot. Sin. 32, 448-455. Setter TL, Waters I, Mares DJ and Wilson R (2001) Sprouting tolerance of AGWEST wheat breeding lines: Combined and comparative measurements from Crop Improvement Institute, Agriculture Western Australia and I.A. Watson Wheat Research Centre, Plant Breeding Institute, Narrabri, NSW. pp. 1-18. (Agriculture Western Australia: Perth) Shen Q, Gomez-Cadenas A, Zhang P, Walker-Simmons M, Sheen J and Ho THD (2001) Dissection of abscisic acid signal transduction pathways in barley aleurone layers. Plant Molecular Biology 47, 437-448. Stone PJ and Nicolas ME (1995) Effect of timing of heat stress during grain filling on two wheat varieties differing in heat tolerance. I. Grain growth. Australian Journal of Plant Physiology 22 927-934. Stone PJ, Savin R, Wardlaw IF and Nicolas ME (1995) The influence of recovery temperature on the effects of a brief heat shock on wheat I. Grain growth. Australian Journal of Plant Physiology 22, 945-954. Stoy V (1983) Progress and prospect in sprouting research. In 'Third International Symposium on Pre-Harvest Sprouting in Cereals.' Canada. (Eds JE Kruger and DE LaBerge) pp. 3-7. (Westview Press, Boulder USA) Strand E (1983) Effects of temperature and rainfall on seed dormancy of small grain cultivars. In 'Third International Symposium on Preharvest Sprouting in Cereals'. Canada. (Eds JE Kruger and DE LaBerge) pp. 260-266. (Westview Press, Boulder USA) Strand E (1989a) Studies on seed dormancy in small grain species. I. barley. Norwegian Journal of Agricultural Sciences 3, 85-99. Strand E (1989b) Studies on seed dormancy in small grain species. II. wheat. Norwegian Journal of Agricultural Sciences 3, 101-115. Strand E (1990) Effects of environmental factors on seed dormancy in cultivars of wheat, barley and oats. In 'Fifth International Symposium on Preharvest Sprouting in Cereals' pp. 15-26. (Westview Press, Boulder Colorado) Suzuki T, Matsuura T, Kawakami N and Noda K (2000) Accumulation and leakage of abscisic acid during embryo development and seed dormancy in wheat. Plant Growth Regulation 30, 253-260. Sweeny T (1996) Salvaging the south coast harvest. In 'Western Australian Cereal Update Meeting for Advisers and Consultants'. Perth, Western Australia. (Ed. S Trevenen) pp. 9-13. (Agriculture Western Australia, Perth)

120
Symons SJ, Angold RE, Black M and Chapman JM (1983) Changes in the growth capacity of the developing wheat embryo I. The influences of the enveloping tissues and premature drying. Journal of Experimental Botany 34, 1541-1550. Takahashi N (1980) Effect of environmental factors during seed formation on pre-harvest sprouting. Cereal Research Communications 8, 175-183. Tan M-K, Sharp PJ, Lu M-Q and Howes N (2006) Genetics of grain dormancy in a white wheat. Australian Journal of Agricultural Research 57, 1157-1165. Tashiro T and Wardlaw IF (1990) The response to high temperature shock and humidity changes prior to and during the early stages of grain development in wheat. Australian Journal of Plant Physiology 17, 551-561. Tavakkol Ahshari R and Hucl P (2001) Temporal changes in abscisic acid concentration in dormant and non-dormant seed of wheat (Triticum spp.) genotypes. Journal of Agricultural Science and Technology 3, 9-18. Torada A and Amano Y (2002) Effect of seed coat color on seed dormancy in different environments. Euphytica 126, 99-105. Torada A, Ikeguchi S and Koike M (2005) Mapping and validation of PCR-based markers associated with a major QTL for seed dormancy in wheat. Euphytica 143, 251-255. Trethowan RM (1995) Evaluation and selection of bread wheat (Triticum aestivum L.) for preharvest sprouting tolerance. Australian Journal of Agricultural Research 46, 463-474. Trethowan RM, Rajaram S and Ellison FW (1996) Pre-harvest sprouting tolerance of wheat in the field and under rain simulation. Australian Journal of Agricultural Research 47, 705-716. Ueno K (2002) Effects of desiccation and a change in temperature on germination of immature grains of wheat (Triticum aestivum L.). Euphytica 126, 107-113. Walker-Simmons M (1987) ABA levels and sensitivity in developing wheat embryos of sprouting resistant and susceptible cultivars. Plant Physiology 84, 61-66. Walker-Simmons M and Sesing J (1990) Temperature effects on embryonic abscisic acid levels during development of wheat grain dormancy. Journal of Plant Growth Regulation 9, 51-56. Wang E, Smith CJ, Bond WJ and Verburg K (2004) Estimates of vapour pressure deficit and crop water demand in APSIM and their implications for prediction of crop yield, water use, and deep drainage. Australian Journal of Agricultural Research 55, 1227-1240. Wang H, Fernandez MR, Clarke FR, DePauw RM, Clarke JM (2002) Effects of foliar fungicides on kernel blackpoint of wheat in southern Saskatchewan. Canadian Journal of Plant Pathology 24, 287-293.

121
Wang M, Heimovaara-Dijkstra S and Van Duijn B (1995) Modulation of germination of embryos isolated from dormant and nondormant barley grains by manipulation of endogenous abscisic acid. Planta 195, 586-592. Wardlaw IF, Blumenthal C, Larroque O and Wrigley CW (2002) Contrasting effects of chronic heat stress and heat shock on kernel weight and flour quality in wheat. Functional Plant Biology 29, 25-34. Warner RL, Kundrna DA, Spaeth SC and Jones SS (2000) Dormancy in white-grain mutants of Chinese spring wheat (Triticum aestivum L.). Seed Science Research 10, 51-60. Westgate ME, Passioura JB and Munns R (1996) Water status and ABA content of floral organs in drought-stressed wheat. Australian Journal of Plant Physiology 23, 763-772. White CN, Proebsting WM, Hedden P and Rivin CJ (2000) Gibberellins and seed development in maize. I. Evidence that giberellin / abscisic acid balance governs germination versus maturation pathways. Plant Physiology 122, 1081-1088. Williamson PM (1997) Black point of wheat: in vitro production of symptoms, enzymes involved, and association with Alternaria alternata. Australian Journal of Agricultural Research 48, 13-19. Woodbury W and Wiebe TJ (1983) A possible role for the pericarp in control of germination and dormancy of wheat. In 'Third International Preharvest Sprouting in Cereals Symposium'. Manitoba, Canada. (Eds JE Kruger and DE LaBerge) pp. 51-58. (Westview Press, Inc.) Xiao S-H, Zhang X-Y, Yan C-S and Lin H (2002) Germplasm improvement for preharvest sprouting resistance in Chinese white-grained wheat: An overview of the current strategy. Euphytica 126, 35-38. Yamada K (1984) Changes in the level of endogenous abscisic acid in barley during germination and use of abscisic acid in malting. J. Am. Soc. Brew. Chem. 42, 79-84. Young KJ and Loughman R (2001) Fungal associations with weather stained barley in Western Australia. In '10th Australian Barley Technical Symposium'. Canberra pp. 1-7. (ABT Ltd, Canberra) Zadoks JC, Chang TT and Konzak CF (1974) A decimal code for the growth stages of cereals. Weed Research 14, 415-421.

122
Acknowledgements My PhD started with an idea to investigate the main problem limiting the consistent
production of marketable grain in the region in which I grew up. Over the last 20 years I
have been involved with the anguish, heartbreak and financial devastation that preharvest
sprouting can have on the livelihood and profitability of growers in the Southern coastal
regions of the Western Australian wheat belt. Preharvest sprouting can be one of the most
demoralising receival standards from a grower�s perspective, often described as a kick in
the teeth when you are already down, by many old growers. After a season, often filled
with uncertainty, anguish and stress, the difference between profit and loss rides on the
whims of the weather in the last few weeks of the season and a 5 minute test (falling
number) carried out in a tin shed.
My deepest thanks to my industry supervisor Tim Setter, for being a fantastic supervisor
and mentor; Tim you have not only encouraged my and mentored me in many aspects of
this project, but also in my pursuit of a career in applied, field based, research. Thanks for
always running a practicality check ensuring my research never lost its relevance. Further
more, thank you in helping to solve the often daunting logistical task of running controlled
temperature experiments in the field, 750 km from home. Thank you Tim.
Thank you also to Daryl Mares, for being a constant guide and wealth of knowledge on
quality traits and the science behind preharvest sprouting tolerance. Daryl you are always
on the other end of the phone line, available to discuss experimental designs, technical
constraints, data interpretation, the odd mistake and anything else that came up. Thank you
Daryl for your guidance, direction and consideration.
Thank you to Julie Plummer, for keeping a watchful eye over the whole project, ensuring
no piece of data was left unturned, and always being available on Monday morning at 10:00
am to discuss the latest problem. Thank you Julie for being considerate and patient in
understanding the time constraints imposed by industry partners.
Thank you also to the people at CSIRO Plant Industry who helped with the analytical
experiments. In particular Frank Gubler, Andrew Poole, Peter Chandler and Rod King, for

123
technical assistance, advice and support, without your support the ABA aspect of this
project would not have been possible.
Thank you to my fellow colleagues at the Department of Agricultural Western Australia for
advice, support and friendship throughout this thesis thanks to; Keith Alcock for approved
study leave. The Cereal Physiology team, Tim Setter, Irene Waters, Glenn McDonald and
Hossein Saberi. The Cereal Pathology team including Rob Loughman and John Majewski.
The Breeders, Robin Wilson, Robyn McLean and Iain Barclay. The Statisticians including
Peter Clarke and Mario D�Antuono and Katia Stefanova. The Cereal Chemistry staff Bill
Lambe and colleagues and the Cereal Agronomy team in Esperance, Ben Curtis,
Mohammad Amjad and Jeremy Lemon. You have supported me in the PhD journey and
allowed me to disrupt your normal routine and helped out in many ways.
The friendship and encouragement of fellow PhD Students Leida Williams, Lindsay Bell,
Eleftheria Dalmaris, Cameron Beck, Chris Jones, Craig Scanlan, Judith Rathjen, Claire
Farrell, Chris Szota and Al Grigg was invaluable and will never be forgotten.
My gratitude goes to the various people who have assisted with my harvests carrying out
mundane tasks, with often little reward, including; Belinda Boyd, Michelle Boyd, Colin
Boyd, Glenn McDonald, Kirsten Frost, Darren Dixon, Vince Lambert and Colin Norwood.
Your willingness to help and lend a hand was sincerely appreciated.
My deepest thanks to the colleagues, friends and family who allowed my to roll a swag out
in their spare rooms, on my three month stints at Esperance, Katanning and Canberra,
including; Matt and Fiona Ryan, Mick and Wendy Robinson, Suzanne Hill, Mary, Julia,
Gabrielle and Graham Collins, Jacinta Falconer, Tania and Daryl Wisewould, Patricia and
Daniel Hill, John-Paul Collins, Glenn and Kathi McDonald and Phillip and Taryn Blight.
Thank you also to the industry funding bodies that have supported this work. In particular
the Grains Research and Development Corporation through a Grains Research Scholarship
GRS66, the Department of Agriculture and Food Western Australia who assisted with the
running of field trials and study leave, Ravensthorpe Agricultural Initiative Network and
CSIRO Plant Industry for a studentship to carry out the ABA measurements. Thank you to

124
all of these financial contributors, who entrusted me to carry out relevant research for their
stakeholders, the growers.
Thank you to my wife, for your love, unconditional support and lots of pre-cooked meals.
To my immediate family, Rachael, Daniel, Belinda, Mum, Dad, MIL, Dave and Jaydon,
thank you for your love, patience and moral support.
My last thanks are to the many enthusiastic growers whom I have met over the past years of
this PhD. You have asked the most difficult questions, yet provide the most valuable
inspiration and encouragement, Thank you.

125
Appendix A

126

127

128

129

130

131

132
Appendix B Details of four oral presentations made throughout the candidature of this PhD at regional
and national conferences which had papers as part of the proceedings.
• Biddulph TB, (2004) Preharvest sprouting tolerance of wheat in Western Australia.
March 2004, Agribusiness Regional Crop Updates, Ravensthorpe and Jerramungup,
WA.
• Biddulph TB, Mares, DJ, Setter, TL, and Plummer, JA. (2006) Environment is it as
important as variety in preharvest sprouting tolerance? Agribusiness Crop Updates,
March 2006, Burswood, Perth, WA.
• Biddulph TB, Mares, DJ, Setter, TL, and Plummer, JA. (2006) Environment is it as
important as variety in preharvest sprouting tolerance? Agribusiness Regional Crop
Updates, March 2006, Esperance, WA.
• Biddulph TB, Mares, DJ, Setter, TL, and Plummer, JA. (2006) Environment is it as
important as variety in preharvest sprouting tolerance? Fitzgerald Biosphere Group
Annual Trials meeting, March 2006, Jerramungup, WA.

133
Appendix C Details of two poster presentations made throughout the candidature of this PhD at national
conferences.
• Biddulph TB, Plummer, JA, Mares, DJ and. Setter, TL. (2004) Drought and high
temperature increase preharvest sprouting tolerance in wheat without grain
dormancy. Combio, September 2004, Burswood, Perth, WA.
DROUGHT AND HIGH TEMPERATURE INCREASES PREHARVEST
SPROUTING TOLERANCE IN WHEAT WITHOUT GRAIN DORMANCY
Biddulph T.B.1, Mares D.J.2, Plummer J.A.1 and Setter T.L.3 1Plant Biology, FNAS University of Western Australia, 35 Stirling Hwy Crawley
WA 6009. 2School of Agriculture & Wine, WAITE Campus, University of
Adelaide, Urrbrae SA 5064. 3Department of Agriculture WA, 3 Baron Hay Ct,
South Perth WA 6151.
Preharvest sprouting is a common problem in cereals without grain dormancy. It
occurs when grain is exposed to rainfall or high moisture conditions. Environmental
conditions during grain filling have a substantial impact on the expression of
sprouting tolerance, however how much is uncertain. Dormant and non-dormant
hard white winter wheat lines were exposed to moisture stress or irrigated
conditions and either low or high temperatures during grain filling in a controlled
field experiment. Moisture stress increased dormancy and this overrode the impact
of low temperatures on increasing dormancy. Embryo sensitivity was induced in a
non-dormant line. This has implications for selection of lines in breeding programs.

134
• Biddulph TB, Plummer, JA, Mares, DJ and. Setter, TL. (2005) Preharvest sprouting
resistance in the field. Agribusiness Crop Updates, February 2005, Burswood,
Perth, WA.
Preharvest Sprouting Resistance
of Wheat in the Field
Biddulph, T. B1,2., Plummer, J. A.,2 Mares, D.J3 & Setter, T. L4.1. Email: [email protected]. 2. Plant Biology, FNAS, University of Western Australia. 3. School of Agriculture and Wine, WAITE Campus, University of Adelaide. 4. Crop Improvement, Department of Agriculture WA, 3 South Perth.
BACKGROUND
� In coastal portions of the Western Australian wheat belt , downgrading due to sprouting can result in up to a 20% price reduction for producers. End-use quality losses due to rainfall include a reduction in weight, dough strength (Derera, 1989). Sprouting is a problem in Western Australia because locally adapted high yielding varieties lack sprouting tolerance and there is a yield penalty associated with growing sprouting tolerant varieties.
� There is little published evidence that seed dormancy, the main mechanism targeted by breeders, equals sprouting tolerance in the field. The objectives of this study were to determine if seed dormancy results in lower Preharvest Sprouting tolerance with natural rainfall.
MATERIAL & METHODS
� Nine white seeded spring wheat varieties from Western Australia and four lines commonly used as preharvest sprouting standards were evaluated in field tests in Katanning (low probability of rainfall) and Esperance (high probability of rainfall) Western Australia.
� Seed dormancy was evaluated at physiological maturity using germination tests (Figure 1A) and a weighted germination index was calculated (Mares and Mrva, 2001).
GI = (7 x n1 + 6 x n2 + 5 x n3 + 4 x n4 + 3 x n5 + 2 x n6 + 1 x n7)(7 x 20)
Ravensthorpe Agricultural
Initiative Network
Ravensthorpe Agricultural
Initiative Network
Ravensthorpe Agricultural
Initiative Network
� Preharvest sprouting resistance was evaluated using the falling number test (AACC Method 56-81B) on the grain samples after exposure to natural field weathering (Figure 1B).
RESULTS & DISCUSSION� It rained in Esperance but not in Katanning
� Falling number declined in some varieties but not others when exposed to rainfall after maturity (Table 1)
� Dormant varieties had a higher falling number for longer than non-dormant varieties, (Figure 2 and Table 1)
� Weather affected grains (mould + black point) varied between genotypes with some on average below the 20 grains out of 500 maximum for premium grades
CONCLUSIONS� Breeding for seed dormancy should result in usable sprouting
tolerance from a farmers perspective at the paddock level
41
520
492
516
519
514
470
461
535
495
463
518
Katanning
70
460
439
432
410
359
289
259
498
436
240
193
Esperance
Falling Number 5th
February 2004
190.67Cascades
290.51Sunlin
Esperance
150.28LSD0.05
190.75EGA Eagle Rock
190.63Clearfield Stiletto
310.66Stiletto
270.75Camm
380.73EGA Castle Rock
440.39AUS1408
520.39DM2001
370.73Janz
280.76Hartog
Weather affected
GIVariety
41
520
492
516
519
514
470
461
535
495
463
518
Katanning
70
460
439
432
410
359
289
259
498
436
240
193
Esperance
Falling Number 5th
February 2004
190.67Cascades
290.51Sunlin
Esperance
150.28LSD0.05
190.75EGA Eagle Rock
190.63Clearfield Stiletto
310.66Stiletto
270.75Camm
380.73EGA Castle Rock
440.39AUS1408
520.39DM2001
370.73Janz
280.76Hartog
Weather affected
GIVariety
Table 1. Mean germination index, falling number and number of weather affected grains (mould + black point) for eleven genotypes grown at Esperance and Katanning.
REFERENCESDerera, N. F. 1989. The effects of preharvest rain In: Derera (ED.), Preharvest Sprouting In Cereals, pp. 2-14. CRC Press Inc., Boca Raton, USA.Mares , D. J. and Mrva, K. 2001. Mapping Quantitative trait loci associated with variation in grain dormancy in Australian wheat. Australian Journal of Agricultural Research. 44: 1259-1272.
0
100
200
300
400
500
10/12/2003 24/12/2003 7/01/2004 21/01/2004 4/02/2004
Falli
ng N
umbe
r (se
cond
s)
0
2
4
6
8
10
12
14
16
18
20
Rai
nfal
l (m
m) DM2001
JanzHartogASW minGeneral Purpose min
Figure 2. Decline in Falling Number for sprouting tolerant checks over time when grown at Esperance. Means, n=3, LSD0.05
0
100
200
300
400
500
10/12/2003 24/12/2003 7/01/2004 21/01/2004 4/02/2004
Falli
ng N
umbe
r (se
cond
s)
0
2
4
6
8
10
12
14
16
18
20
Rai
nfal
l (m
m) DM2001
JanzHartogASW minGeneral Purpose min
Figure 2. Decline in Falling Number for sprouting tolerant checks over time when grown at Esperance. Means, n=3, LSD0.05
Figure 1 A Seed dormancy evaluation using germination tests.
B genotypes naturally weathered in Esperance.
A B
Figure 1 A Seed dormancy evaluation using germination tests.
B genotypes naturally weathered in Esperance.
A B
ACKNOWLEDGEMENTS�We acknowledge the financial support from the Grains Research and Development Council, Department of Agriculture Western Australia and the Ravensthorpe Agricultural Initiative Network
�We are indebted to Glenn McDonald, Department of Agriculture, Katanning, for assistance and advice in managing the trial

135
Appendix D
IR MS IR MS IR
In the foreground is my controlled temperature and water stress experiment at Esperance on
the 11th October 2005 (Chapter 4). The square poly shelter, is linked to a 3.5 hp Fujitsu
reverse-cycle air-conditioner. It was moved at ten day intervals from left to right, and
would hold six rows of wheat. Its current position was during the 30-40 dpa heat shocking
treatment, straddling both moisture stressed (MS) and irrigated (IR) plots. The cups indicate
the position of temperature sensors at head height, used to measure any potential variation
in temperature through the length or width of the both the inner and outer poly shelters. No
significant differences were detected spatially, within either shelter with the 0.5°C accuracy
of the thermistors used
In the background, is the experiment for Chapter 5. It is covered at the moment as is the
foreground for the moisture stress and irrigation treatments. Beyond that under the open
frame is the Stage 4 Department of Agriculture and Food Western Australia, preharvest
screening nursery, samples of which are presented in Chapter 5. The image below provides
a bird�s eye view of the plot layout, with the top of the page representing the forground of
the picture.

136