measuring, reversing, and modeling the mechanical changes due to the absence of fibulin-4 in mouse...
TRANSCRIPT

Biomech Model MechanobiolDOI 10.1007/s10237-014-0556-x
ORIGINAL PAPER
Measuring, reversing, and modeling the mechanical changes dueto the absence of Fibulin-4 in mouse arteries
Victoria P. Le · Yoshito Yamashiro ·Hiromi Yanagisawa · Jessica E. Wagenseil
Received: 20 November 2013 / Accepted: 27 January 2014© Springer-Verlag Berlin Heidelberg 2014
Abstract Mice with a smooth muscle cell (SMC)-specificdeletion of Fibulin-4 (SMKO) show decreased expression ofSMC contractile genes, decreased circumferential compli-ance, and develop aneurysms in the ascending aorta. Neona-tal administration of drugs that inhibit the angiotensin IIpathway encourages the expression of contractile genes andprevents aneurysm development, but does not increase com-pliance in SMKO aorta. We hypothesized that multidimen-sional mechanical changes in the aorta and/or other elasticarteries may contribute to aneurysm pathophysiology. Wefound that the SMKO ascending aorta and carotid arteryshowed mechanical changes in the axial direction. Thesechanges were not reversed by angiotensin II inhibitors, hencereversing the axial changes is not required for aneurysm pre-vention. Mechanical changes in the circumferential directionwere specific to the ascending aorta; therefore, mechanicalchanges in the carotid do not contribute to aortic aneurysmdevelopment. We also hypothesized that a published modelof postnatal aortic growth and remodeling could be usedto investigate mechanisms behind the changes in SMKOaorta and aneurysm development over time. Dimensions andmechanical behavior of adult SMKO aorta were reproducedby the model after modifying the initial component material
V. P. Le · J. E. WagenseilDepartment of Biomedical Engineering, Saint Louis University,St. Louis, MO, USA
Y. Yamashiro · H. YanagisawaDepartment of Molecular Biology, University of TexasSouthwestern Medical Center, Dallas, TX, USA
J. E. Wagenseil (B)Department of Mechanical Engineering and Materials Science,Washington University, One Brookings Dr., CB 1185,St. Louis, MO 63130, USAe-mail: [email protected]
constants and the aortic dilation with each postnatal time step.The model links biological observations to specific mechan-ical responses in aneurysm development and treatment.
Keywords Vascular mechanics · Extracellular matrix ·Constrained mixture model
1 Introduction
Fibulin-4 (FBLN4) is essential for elastic fiber assembly inthe skin, lungs, and arteries. FBLN4 mutations in humans areassociated with arterial tortuosity and aneurysms (Dasoukiet al. 2007). Mice that do not express Fibulin-4 (Fbln4−/−)die postnatally from lung and vascular defects (McLaugh-lin et al. 2006). Mice with a smooth muscle cell (SMC)-specific deletion of Fbln4 (SMKO) develop ascending aor-tic aneurysms. SMKO vascular SMCs exhibit hyperprolif-eration and loss of a contractile phenotype (Huang et al.2010). Local activation of angiotensin II (AngII) signalingis a primary cause of SMKO aneurysms. Aneurysms canbe prevented with neonatal administration of antihyperten-sive drugs that inhibit angiotensin-converting enzyme (ACE),such as captopril (CAP), or that block the angiotensin typeI receptor, such as losartan (LOS). Treatment with CAP orLOS encourages the expression of SMC contractile genesand reverses the enlarged diameter, but does not reversethe decreased circumferential compliance in SMKO aorta.Aneurysm prevention is not linked to blood pressure changesalone, because propranolol (PROP), an antihypertensive drugthat is a non-selective beta-adrenergic receptor blocker, doesnot prevent aneurysms in SMKO mice (Huang et al. 2013).
The goal of the current study was to further investigate themechanical behavior of SMKO arteries. We hypothesizedthat although preventative drug treatment did not reverse
123

V. P. Le et al.
the changes in circumferential compliance of the ascendingaorta, we may observe alterations in axial mechanical behav-ior of the ascending aorta and multidimensional mechanicalbehavior of other elastic arteries, such as the carotid artery,that would contribute to aneurysm development in SMKOmice. We also hypothesized that a previously published con-strained mixture model of aortic growth and remodeling(Wagenseil 2011) could provide insight into relationshipsbetween mechanically stimulated remodeling and aneurysmdevelopment in the growing mouse aorta.
2 Materials and methods
2.1 Mice
129SvEv/C57Bl6 male and female mice with an SMC-specific knockout of the Fibulin-4 gene (SMKO) (Huang et al.2010) and wild-type littermates (CTR) were killed at approx-imately 6 weeks of age. All protocols were approved by theInstitutional Animal Care and Use Committee.
2.2 Drug treatment protocols
LOS (0.6 g/L, provided by Merck Inc.), CAP (0.075 g/L,Sigma), and PROP (0.6 g/L, Sigma) were administered to themice in drinking water ad libitum from age 7 to 43 ± 2 days.Untreated (UNT) groups received plain water. Histology andWestern blot data were taken from mice on a different treat-ment protocol, where the mice were treated from age 7–90(histology) or age 7–30 (Western blot) days. Previous resultsshowed no differences between the treatment protocols, aslong as LOS was started by 7 days of age (Huang et al. 2013).
2.3 Arterial dissection and lengths
Small charcoal particles were placed on the left commoncarotid artery. The carotid was imaged, and the lengthsbetween particles were measured before (l) and after excision(L) to determine the in vivo axial stretch ratio (λiv
z = l/L).The ascending aorta was imaged and the in vivo lengthbetween the heart base and the innominate artery was mea-sured.
2.4 Mechanical testing and unloaded diameters
The left common carotid or the ascending aorta was mountedin a pressure myograph (Danish Myotechnology) formechanical testing (Le et al. 2011). The artery was immersedin physiologic saline solution at 37 ◦C and stretched to theapproximate in vivo length. The axial stretch ratio (λz) wascalculated by dividing the stretched length by the unstretchedlength. After preconditioning, the artery was inflated three
times from 0 to 175 mmHg in steps of 25 mmHg with 12 sbetween steps while pressure, outer diameter, and axial forcewere recorded. After testing, the artery was removed, placedin a dish of physiologic saline, cut into 2–3 rings, 0.2–0.3 mmthick, and imaged to measure the unloaded dimensions.
2.5 Data analysis
Compliance was calculated as the average change in outerdiameter per mmHg for each 25 mmHg pressure step. Theloaded inner radius for each artery, rin, was calculated assum-ing incompressibility (Faury et al. 1999). The average cir-cumferential stretch ratio, λθ , was calculated by
λθ = 1
2
(rin
Rin+ rout
Rout
), (1)
where Rin is the unloaded inner radius and rout and Rout arethe loaded and unloaded outer radii. The average circum-ferential wall stress, σθ , was calculated assuming negligibleshear:
σθ = prin
rout − rin, (2)
where p is the measured internal pressure. The incrementalelastic modulus in the circumferential direction was calcu-lated as the average change in circumferential stress dividedby the average change in circumferential stretch ratio foreach 25 mmHg pressure step. The average axial wall stress,σz , was calculated for a closed-end cylinder:
σz = f + pπr2in
π(r2out − r2
in), (3)
where f is the measured axial force.
2.6 Statistics
Data are mean ± SD, unless otherwise noted. ANOVA witha Bonferroni post hoc test between genotypes for each treat-ment group was used for all comparisons (SPSS). p < .05was considered significant.
2.7 Mechanical modeling
A published model (Wagenseil 2011) was used to inves-tigate possible mechanisms leading to the remodeling inUNT SMKO and LOS SMKO aorta compared to UNT CTR.These groups were chosen because the adult blood pres-sures are similar (Huang et al. 2013), which simplifies themodel assumptions. The model was previously used to pre-dict changes in dimensions and mechanical behavior dur-ing normal postnatal development of the mouse ascendingaorta from ages 3–30 days in discrete time steps of 4.5 days.The model is based on the observations that perturbations
123

Absence of Fibulin-4 in mouse arteries
in blood pressure, axial forces, or blood flow are counter-acted by respective changes in the aortic wall thickness, axiallength, or inner radius to maintain homeostatic wall stresses.The stresses include the circumferential and axial stresses(Eqs. 2, 3), as well as the wall shear stress (τ ). For steady,fully developed flow in a long, straight tube, the wall shearstress is
τ = 4Qμ
πr3in
, (4)
where Q blood flow, μ, blood viscosity (4 cP).The wall stresses at the current pressure (ph), deformed
length (lh), flow (Qh), deformed inner radius (rin,h), andthickness (hh) are considered the homeostatic values for eachdiscrete time step. To move to the next developmental time,the pressure, deformed axial length, and flow are increasedto new values determined from previously published exper-imental data
p = εp ph, (5a)
l = εl lh, (5b)
Q = εQ Qh, (5c)
where εP , εl , and εQ are constants for each time step.When pressure, length, and flow are increased, it is
assumed that first the aorta instantaneously dilates in anattempt to return to the homeostatic shear stress and thenthe remodeling of the wall precedes in an attempt to restorehomeostatic values of circumferential and axial stress. In pre-vious work (Wagenseil 2011), changes in blood flow had tobe decoupled from changes in the inner radius to predictpostnatal growth of the aorta. A constant 12 % increase ininner radius for each time step was assumed, regardless ofthe change in blood flow, and this assumption is includedhere. The decoupling of the inner radius from the blood flowimplies that the developing mouse aorta does not maintaina homeostatic shear stress and/or that Eq. 4 is insufficientto describe the shear stresses in the ascending aorta witha complex geometry and pulsatile flows (Van Doormaal etal. 2012). Despite these limitations, the model is able to pre-dict the dimensions and mechanical behavior of a 30-day-oldmouse aorta after growth and remodeling.
Relevant model equations (Eqs. 6–18) are listed in the“Appendix.” The total stress in the aortic wall is the sumof the component stresses (Eq. 6). The components includeSMCs, elastin, and collagen. Each component is producedat a homeostatic stretch ratio (Eq. 7), and the mechanicalbehavior is described by a constitutive equation (Eqs. 8–17).Starting from the dimensions and mechanical behavior of a3-day-old mouse aorta, stepwise changes in blood flow, pres-sure, and axial length are applied. The arterial wall dilates12 % regardless of the change in blood flow, and then remod-eling proceeds as components turnover (Eq. 18) in an attempt
to maintain circumferential and axial stresses near the pre-vious homeostatic values. Remodeling is allowed to reachsteady state for each time step, and then the final stresses,loaded and unloaded dimensions, stretch ratios, and massfractions for newly produced components are calculated foruse as inputs for the next developmental time step. The solu-tion strategy, model input values, and additional equationsare in Wagenseil (2011).
The model is used to qualitatively reproduce the pressure–diameter and circumferential stretch–stress behavior of adultUNT CTR, UNT SMKO, and LOS SMKO aortae. The pre-viously published model parameters are used to reproducedata for UNT CTR aorta. Changes to the model parameters,motivated by experimental evidence in Huang et al. (2013),are investigated to determine which changes best reproducethe experimental data for UNT SMKO and LOS SMKO aor-tae. Starting model inputs are identical for all groups becauseSMKO aorta is histologically similar to CTR aorta at 1 dayold, with slight changes evident by 7 days old. LOS treatmentbegan at 7 days, which is the first time point where alter-ations to the model parameters have an effect. Adult SMKOaorta shows fragmentation and disorganization of the elas-tic fibers, with no obvious decrease in the amount of elasticfiber staining; hence, we assumed that the elastin contributionto the total wall stress is reduced through a decrease in theSMKO elastin material constant (b1, Eq. 8a). LOS treatmentpartially rescues the elastic fiber fragmentation in SMKOaorta, so LOS SMKO aorta may also have an altered elastinmaterial constant. We reasoned that the collagen contributionto the total wall stress may be increased to compensate forthe decreased elastin contribution. In mice lacking Fibulin-5,another protein necessary for elastic fiber assembly, the arter-ial collagen fibers show reduced undulation (Wan et al. 2010).Therefore, we investigated changes to the collagen materialparameters (b2, b3, b4, Eqs. 9–11) that would increase thecollagen stress contribution, as well as change the shape ofthe stress–stretch curve. Expression of SMC contractile pro-teins is reduced in SMKO aorta and is rescued by LOS treat-ment. Altered SMC phenotype may cause changes in thepassive SMC material parameters (b5, b6, b7, Eqs. 13–15),active SMC constants (λM , λ0, Eqs. 16, 17), basal SMC tone(TB , Eqs. 16, 17), or arterial dilation with each time step.Increased SMC proliferation is also evident in SMKO aortaand is rescued by LOS treatment. This may cause changes inthe SMC production constant (K g
m , Eq. 18) or total SMC massfraction (φm , Eq. 6). A summary of the altered model para-meters is listed in Table 1. For the parameter alterations thatbest reproduced changes in the dimensions and mechanicalbehavior of UNT and LOS SMKO aorta compared to UNTCTR, we calculated the physiologic shear stress, circumfer-ential and axial stress, circumferential and axial stretch ratio,the normalized unloaded dimensions, and the stress contri-bution of each wall component.
123
V. P. Le et al.
Table 1 Model parameters hypothesized to be affected by the loss of SMC-specific Fbln4 and subsequent treatment with LOS
Parameter name Parametersymbol (s)
Motivationfor change
Originalvalue (s)
Variation Results
Elastin materialconstant
b1 Fragmentation ofelastic fibers
35 kPa × 0.1–0.9 Shift press–diam curves left andmake stretch–stress morenonlinear
Collagen materialconstants
b2, b3, b4 Possiblecompensation bycollagen forfragmented elasticfibers
10 kPa,1.5, .01
× 1.1–1.8 allvalues
Shift press–diam curves left andmake stretch–stress morenonlinear
SMC passive materialconstants
b5, b6, b7 Reducedexpressionof SMCcontractileproteins
2 kPa,50, .01
× 1.1–1.8 allvalues
Almost no change inpress–diam or stretch–stressrelationships
SMC active materialconstants
λM, λ0 Reduced expressionof SMC contractileproteins
0.6, 2.8 × 0.5–2.0 each valueindependently
Almost no change inpress–diam or stretch–stressrelationships
SMC basal tone TB Reduced expressionof SMC contractileproteins
60 kPa × 0.5–2.0 Almost no change inpress–diam or stretch–stressrelationships
Arterial dilation witheach time step
None Reduced expressionof SMC contractileproteins
12 % 14–20 % Shift press–diam curves up,almost no change instretch–stress except morenonlinear at highest dilation
SMC productionconstant
K mg Increased SMC
proliferation6.9 × 1.5–4.5 Small shift left in press–diam
curves and slightly morenonlinear stretch–stresscurves
Total SMC massfraction
φm Increased SMCproliferation
Decreaseswith agefrom 17.3to 8.4 %
+1–5 %, elastin andcollagen reducedequally to accountfor extra SMCfraction
Almost no change inpress–diam, small shift to lessnonlinear stretch–stresscurves
The motivation for each parameter change is based on the data from Huang et al. (2013). Relevant model equations and parameter definitions aregiven in the “Appendix,” with additional equations and solution strategies in Wagenseil (2011)
2.8 Histology and Western blot analyses
For histology, at least four aortae in each group were har-vested, perfusion fixed in 4 % paraformaldehyde, and embed-ded in paraffin. 5µm sections were stained with hematoxylinand eosin (HE) for routine histology, Hart’s for visualizingelastic fibers, and Masson’s trichrome for the detection of col-lagen fibers. For Western blot analyses, three aortae in eachgroup were harvested and perivascular adipose tissues werethoroughly removed, then minced in liquid nitrogen by pes-tle, and dissolved into RIPA lysis buffer (Sigma) containing1 % protease inhibitor (Sigma). The lysates were mixed with3× SDS sample buffer containing 2-mercaptoethanol andboiled at 95 ◦C for 5 min and then subjected to SDS–PAGE.Proteins were transferred to a Western PVDF membrane(Millipore) and immunoblotted with anti-mouse tropoelastin(generous gift from Dr. R. Mecham, 1:1,000), anti-collagen I(1:500, Millipore), or anti-GAPDH (1:3,000, Cell Signaling)antibodies. Membranes were then incubated with anti-rabbit
HRP-conjugated secondary antibody (1:1,000, Bio-Rad) andvisualized with chemoluminescence kit (Santa Cruz Biotech-nology) or SuperSignal West Femto Maximum SensitivitySubstrate (Thermo Scientific).
3 Results
3.1 CAP and LOS reverse increases in SMKO aorticdiameter and thickness, but not length
The in vivo length of the ascending aorta in UNT SMKO miceis 93 % longer than UNT CTR (p < .001) (Fig. 1a). PROP(p = .015) and CAP (p = .003) SMKO aorta also haveincreased length, while LOS SMKO shows a trend towardincreased length. The unloaded outer diameters of UNT andPROP SMKO aortae are 62 and 84 % larger, respectively,than CTR (p < .001) (Fig. 1b). CAP and LOS SMKO aor-tic diameters are within 19 and 17 %, respectively, of CTR,although LOS SMKO diameter is still significantly larger
123

Absence of Fibulin-4 in mouse arteries
20
40
60
80
1.2
1.4
1.6
Car
axi
al s
tret
ch r
atio
A
D
0
2
4
6
Ao
rta
len
gth
(m
m) *
UN
T
PR
OP
LOS
CA
P
CTR
UN
T
PR
OP
LOS
CA
P
SMKO
0.6
1.4
1.8
Ao
rta
un
l dia
m (
mm
)
0.35
Car
un
l dia
m (
mm
)
0.45
0.40
0.30
1.0
1.0
UN
T
PR
OP
LOS
CA
P
CTRU
NT
PR
OP
LOS
CA
P
SMKO
100
150
200
Ao
rta
un
l th
ick
(µm
)
UN
T
PR
OP
LOS
CA
P
CTR
UN
T
PR
OP
LOS
CA
P
SMKO
Car
un
l th
ick
(µm
)
E F
* ** *
* **
*
*
UN
T
PR
OP
LOS
CA
P
CTR
UN
T
PR
OP
LOS
CA
P
SMKO
UN
T
PR
OP
LOS
CA
P
CTR
UN
T
PR
OP
LOS
CA
P
SMKO
UN
T
PR
OP
LOS
CA
P
CTR
UN
T
PR
OP
LOS
CA
P
SMKOB C
Fig. 1 SMKO mice show artery-specific changes in the mean lengthsand diameters compared to CTR. The in vivo length of SMKO ascend-ing aorta is longer than CTR and in general this is not reversed by thedrug treatments (a). UNT SMKO ascending aorta has a larger unloadeddiameter than CTR (b). The large diameter is not normalized by PROP,but is greatly reduced or made statistically insignificant by CAP or LOS.UNT SMKO aorta has an increased thickness compared to CTR, and
this is reversed with all drug treatments (c). The in vivo stretch ratioof the left common carotid artery is smaller in SMKO mice comparedto CTR, and in general, this is not reversed by the drug treatments (d).There are no differences between SMKO and CTR for the unloadeddiameter (e) or thickness (f) of the carotid. N = 6−8 for each group.∗ p < .05
than LOS CTR (p = .029). The unloaded thickness of UNTSMKO aorta is 43 % greater than UNT CTR (p = .005)(Fig. 1c). All drug treatments reverse the increased thicknessso that SMKO aortic thicknesses are not different from CTR.The in vivo stretch ratio of UNT SMKO left common carotidartery is reduced 18 % compared to UNT CTR (p < .001)
(Fig. 1d). A decreased stretch ratio may be caused by growthin the axial direction. PROP (p = .049) and CAP (p < .001)
SMKO carotid arteries also have decreased stretch ratios,while LOS SMKO carotid shows a trend in the same direc-tion. There are no differences in carotid unloaded dimen-sions between SMKO and CTR mice for any of the treat-ment groups (Fig. 1e, f). These results highlight SMKO arte-rial remodeling in the axial direction that has not previouslybeen reported. These results also confirm that expansion ofthe SMKO unloaded diameter is restricted to the ascendingaorta, with no change in the carotid.
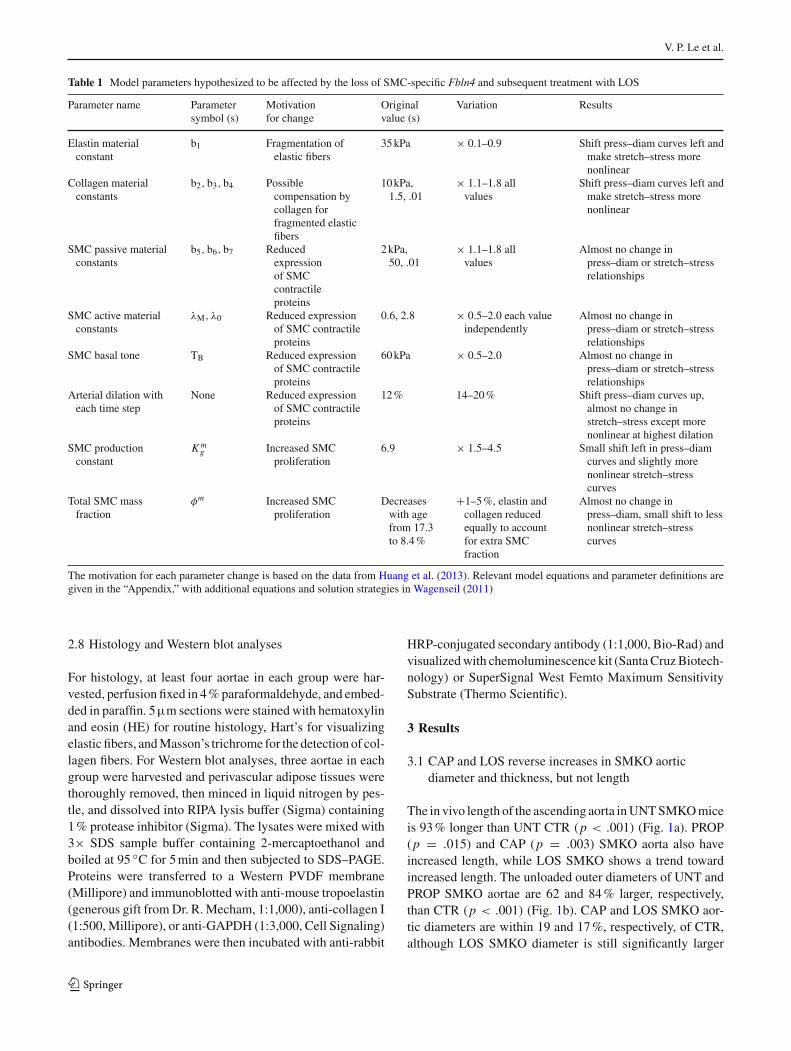
3.2 Mechanical behavior is altered in the SMKO ascendingaorta, but not in the carotid artery
The outer diameters of UNT and PROP SMKO aortae are 66–151 % larger than CTR, depending on the applied pressure(p < .002) (Fig. 2a). CAP and LOS SMKO aortic diam-
eters are not significantly different than CTR, although theshape of the pressure–diameter curve is altered. The axialforce in PROP CTR aorta is higher than PROP SMKO aorta(p < .006) (Fig. 2b), but there are no significant differencesbetween genotypes for any other groups. Compliance valuesfor UNT and PROP SMKO aortae are increased 324–577 %compared to CTR at 0–25 mmHg (p < .05), while compli-ance values for CAP and LOS SMKO aortae are comparableto CTR (Fig. 2c). At higher pressures (100–175 mmHg forUNT and PROP aortae; 100–150 mmHg for CAP and LOSaortae), SMKO aortic compliance is reduced 47–79 % com-pared to CTR values (p < .04). While the absolute diameterincreases in CAP and LOS SMKO aortae are mostly reversed,the decreased compliance values at physiologic pressures arenot. Additionally, at 150–175 mmHg, the compliance valuesof CAP and LOS CTR aortae are decreased 47–64 % com-pared to UNT CTR (p < .01), suggesting that CAP and LOStreatment affects the mechanical behavior of the aortic wall,regardless of Fbln4 deficiency.
In contrast to the large changes in mechanical behavior ofthe ascending aorta, the left common carotid artery shows nosignificant differences in the diameter (Fig. 2d), axial force(Fig. 2e), or compliance (Fig. 2f) between genotypes for anygroup. Although the left common carotid artery is one of
123
V. P. Le et al.
0 50 100 150−4
−2
0
2
4
6
0 50 100 1500
1
2
3
4
D
Ao
rta
dia
met
er (
mm
)
0 50 100 1500.5
1
1.5
2
2.5
3
0 50 100 150−10
0
10
20
pressure (mmHg)
Ao
rta
axia
l fo
rce
(mN
)
B0 50 100 150
0
5
10
15
Ao
rta
com
p (
µm
/mm
Hg
)
pressure (mmHg)C
Car
co
mp
(µ
m/m
mH
g)
pressure (mmHg)pressure (mmHg)
0 50 100 1500.3
0.4
0.5
0.6
Car
dia
met
er (
mm
)
pressure (mmHg) FE
CTR - UNTCTR - PROP
SMKO - UNTSMKO - PROP
CTR - CAPCTR - LOS
SMKO - CAPSMKO - LOS
*
** * * * * *
*
*
#
#$
$
$$
$$$$
* *
Car
axi
al f
orc
e (m
N)
A pressure (mmHg)
Fig. 2 Large differences are evident in the mean pressure–diameterand pressure–compliance behavior for SMKO aorta compared to CTR,with no differences for the carotid artery. UNT and PROP SMKO aor-tae have larger diameters than CTR at all pressures (a). The differencesbecome statistically insignificant after CAP or LOS treatment. Thepressure–force behavior is similar for SMKO and CTR aortae in mosttreatment groups (b). SMKO aortae have reduced compliance at phys-iologic pressure (100 mmHg) and above, regardless of drug treatment(c). CAP and LOS reduce the compliance of CTR aortae at high pres-
sures. There are no significant differences in the pressure–diameter (d),pressure–force (e), or pressure–compliance (f) behavior of the carotidartery regardless of genotype or drug treatment. Panel A includes a sub-set of the data from Fig. 4e in Huang et al. (2013). The subset includes allaortae for which unloaded dimensions were also measured. N = 6−8for each group. ∗ p < .05 for UNT/PROP SMKO compared to CTR.$ p < .05 for PROP SMKO compared to CTR. # p < .05 for all SMKOand CAP/LOS CTR compared to UNT CTR
the main branches off the ascending aorta, the mechanicalbehavior is not affected by Fbln4 deficiency. The mechani-cal behavior is also not affected by the drugs, showing thatCAP and LOS specifically target the ascending aorta in bothSMKO and CTR mice.
The circumferential stretch between 0–75 mmHg is 10–34% higher in UNT and PROP SMKO aorta compared to CTR(p < .03) (Fig. 3a). The circumferential stress between25–100 mmHg is 65–205 % higher in UNT and PROP SMKOaorta compared to CTR (p < .03) (Fig. 3b). At zero pres-sure, the axial stress in UNT and PROP SMKO aorta is75–80 % lower than CTR (p < .02) (Fig. 3c). For CAP andLOS SMKO aortae, there are no significant differences fromCTR in the circumferential stretch, circumferential stress, oraxial stress. There are also no significant differences betweenSMKO and CTR carotids in any of the groups (Fig. 3d–f).
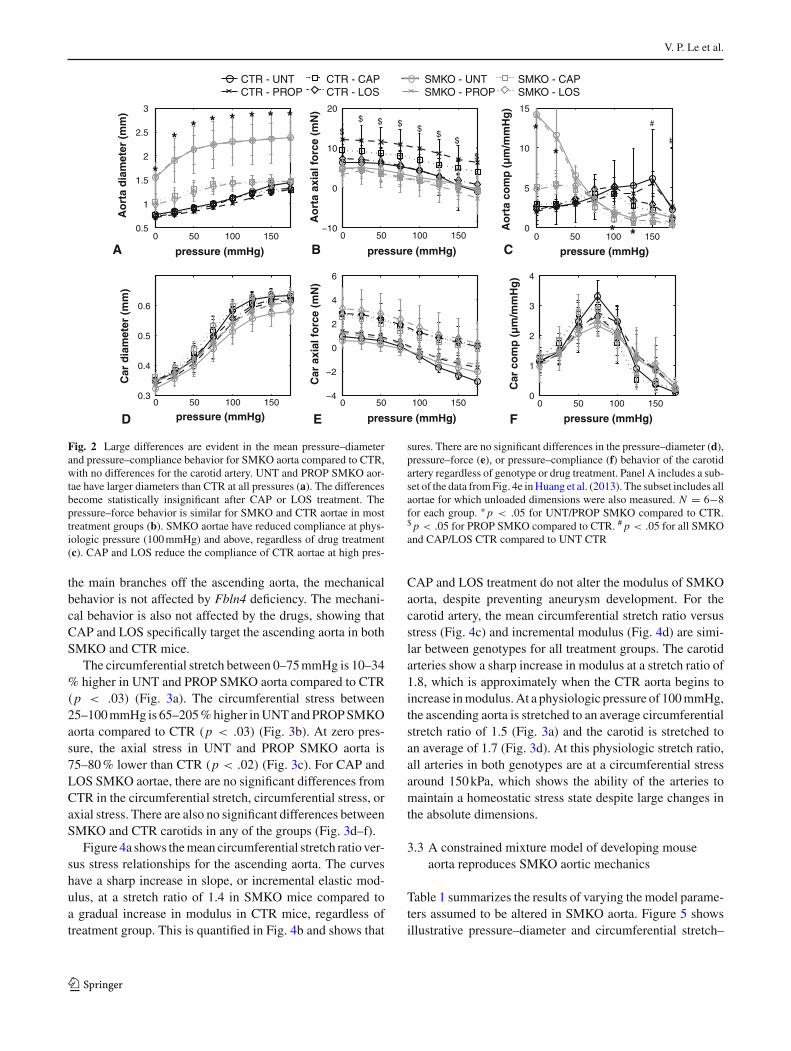
Figure 4a shows the mean circumferential stretch ratio ver-sus stress relationships for the ascending aorta. The curveshave a sharp increase in slope, or incremental elastic mod-ulus, at a stretch ratio of 1.4 in SMKO mice compared toa gradual increase in modulus in CTR mice, regardless oftreatment group. This is quantified in Fig. 4b and shows that
CAP and LOS treatment do not alter the modulus of SMKOaorta, despite preventing aneurysm development. For thecarotid artery, the mean circumferential stretch ratio versusstress (Fig. 4c) and incremental modulus (Fig. 4d) are simi-lar between genotypes for all treatment groups. The carotidarteries show a sharp increase in modulus at a stretch ratio of1.8, which is approximately when the CTR aorta begins toincrease in modulus. At a physiologic pressure of 100 mmHg,the ascending aorta is stretched to an average circumferentialstretch ratio of 1.5 (Fig. 3a) and the carotid is stretched toan average of 1.7 (Fig. 3d). At this physiologic stretch ratio,all arteries in both genotypes are at a circumferential stressaround 150 kPa, which shows the ability of the arteries tomaintain a homeostatic stress state despite large changes inthe absolute dimensions.
3.3 A constrained mixture model of developing mouseaorta reproduces SMKO aortic mechanics
Table 1 summarizes the results of varying the model parame-ters assumed to be altered in SMKO aorta. Figure 5 showsillustrative pressure–diameter and circumferential stretch–
123

Absence of Fibulin-4 in mouse arteries
0 50 100 1500.8
1.2
1.6
2
0 50 100 1500.8
1.2
1.6
2
0 50 100 1500
50
100
150
200
0 50 100 1500
100
200
300
400
0 50 100 1500
50
100
150
200
0 50 100 1500
100
200
300
400
A
D
Ao
rta
circ
str
etch
pressure (mmHg) pressure (mmHg)
Ao
rta
circ
str
ess
(kP
a)
B
Ao
rta
axia
l str
ess
(kP
a)
pressure (mmHg)C
Car
axi
al s
tres
s (k
Pa)
pressure (mmHg)pressure (mmHg)pressure (mmHg) FE
CTR - UNTCTR - PROP
SMKO - UNTSMKO - PROP
CTR - CAPCTR - LOS
SMKO - CAPSMKO - LOS
Car
cir
c st
retc
h
Car
cir
c st
ress
(kP
a)
**
* *
**
**
*
Fig. 3 Mean stretch ratios and stresses are altered in SMKO aorta, butnot in SMKO carotid. The circumferential stretch ratio of SMKO aor-tae is larger than CTR in the UNT and PROP groups at low pressures(a). This difference becomes statistically insignificant between SMKOand CTR after CAP and LOS treatment. The circumferential stress inSMKO aortae is larger than CTR in the UNT and PROP groups atlow pressures (b). This difference becomes statistically insignificant
between SMKO and CTR with CAP and LOS treatment. At zero pres-sures, the axial stress in SMKO aorta is lower than CTR in the UNTand PROP groups (c). There are no significant differences between thecircumferential stretch (d), circumferential stress (e), or axial stress (f)for the carotid artery regardless of genotype or drug treatment. Errorbars are not shown for clarity. N = 6−8 for each group. ∗ p < .05 forUNT/PROP SMKO compared to CTR
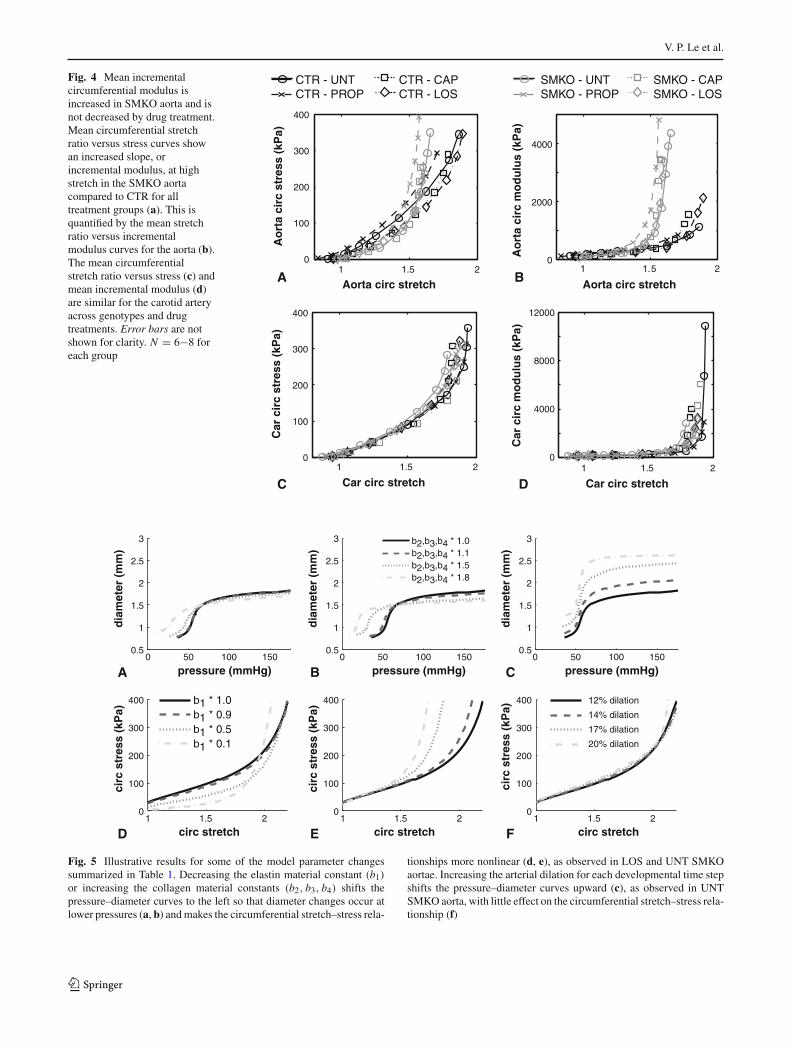
stress behavior for remodeled adult aortae with a reducedelastin material constant (b1) (a, d), increased collagen mater-ial constants (b2, b3, b4) (b, e), and increased arterial dilationwith each time step (c, f). Results for the other options are notshown because the changes are minor compared to the exper-imental data. Reductions in elastin or increases in collagenmaterial constants shift the pressure–diameter curves to theleft, so that diameter increases occur at lower pressures, asseen in SMKO aorta (Fig. 2a). Alterations in the elastin orcollagen material constants reproduce the increased nonlin-earity of the circumferential stretch–stress curves for SMKOaorta (Fig. 4b). Increases in the arterial dilation with eachtime step reproduce the increased diameter for UNT SMKOaorta, with little change in the circumferential stretch–stressbehavior. Changing the arterial dilation with each time stepis the only parameter variation in Table 1 that results in alarge diameter increase.
If collagen stress contributions are altered to compen-sate for the fragmented elastic fibers, changes in the elastinand collagen material constants would likely occur concur-rently in SMKO aorta. We examined permutations of theillustrative elastin and collagen parameter variations andcompared the resulting pressure–diameter, circumferential
stretch–stress behavior and physiologic stress, stretch, andunloaded dimensions. We determined that a 50 % reduc-tion in the elastin material constant and 10 % increasesin the collagen material constants qualitatively reproducethe behavior of LOS SMKO aorta (Fig. 6). Compared toUNT CTR calculated with the original model parameters(Wagenseil 2011), this combination of changes maintainsshear stress, decreases circumferential and axial stresses by30 %, decreases circumferential and axial stretch ratios by4 %, and reproduces the trends in the unloaded dimensionchanges (Table 2). Further decreases in the elastin materialconstant included in Fig. 5 decrease circumferential and axialstresses at physiologic pressure by 70 % and further increasesin the collagen parameters increase the circumferential andaxial stresses by 65 %, which do not match the experimentalresults.
For UNT SMKO aorta, we used the elastin and colla-gen material constants determined for LOS SMKO aorta andincluded a 20 % aortic dilation with each time step. Themodel results for the pressure–diameter and circumferen-tial stretch–stress behavior qualitatively match the experi-mental data (Fig. 6). Compared to UNT CTR, this com-bination of parameter changes reduces the shear stress by
123
V. P. Le et al.
Fig. 4 Mean incrementalcircumferential modulus isincreased in SMKO aorta and isnot decreased by drug treatment.Mean circumferential stretchratio versus stress curves showan increased slope, orincremental modulus, at highstretch in the SMKO aortacompared to CTR for alltreatment groups (a). This isquantified by the mean stretchratio versus incrementalmodulus curves for the aorta (b).The mean circumferentialstretch ratio versus stress (c) andmean incremental modulus (d)are similar for the carotid arteryacross genotypes and drugtreatments. Error bars are notshown for clarity. N = 6−8 foreach group
1 1.5 20
100
200
300
400
1 1.5 20
4000
8000
12000
1 1.5 20
2000
4000
1 1.5 20
100
200
300
400
A
C
Ao
rta
circ
str
ess
(kP
a)Aorta circ stretch Aorta circ stretch
Ao
rta
circ
mo
du
lus
(kP
a)
B
Car circ stretchCar circ stretch D
CTR - UNTCTR - PROP
SMKO - UNTSMKO - PROP
CTR - CAPCTR - LOS
SMKO - CAPSMKO - LOS
Car
cir
c st
ress
(kP
a)
Car
cir
c m
od
ulu
s (k
Pa)
0 50 100 1500.5
1
1.5
2
2.5
3
dia
met
er (
mm
)
pressure (mmHg)
1 1.5 20
100
200
300
400
circ
str
ess
(kP
a)
b1 * 1.0b1 * 0.9b1 * 0.5b1 * 0.1
1 1.5 20
100
200
300
400 12% dilation
14% dilation
17% dilation
20% dilation
0 50 100 1500.5
1
1.5
2
2.5
3
dia
met
er (
mm
)
pressure (mmHg)
b2,b3,b4 * 1.0b2,b3,b4 * 1.1b2,b3,b4 * 1.5b2,b3,b4 * 1.8
A B
D F1 1.5 2
0
100
200
300
400
circ
str
ess
(kP
a)
E
0 50 100 1500.5
1
1.5
2
2.5
3
dia
met
er (
mm
)
pressure (mmHg)C
circ
str
ess
(kP
a)
circ stretch circ stretch circ stretch
Fig. 5 Illustrative results for some of the model parameter changessummarized in Table 1. Decreasing the elastin material constant (b1)
or increasing the collagen material constants (b2, b3, b4) shifts thepressure–diameter curves to the left so that diameter changes occur atlower pressures (a, b) and makes the circumferential stretch–stress rela-
tionships more nonlinear (d, e), as observed in LOS and UNT SMKOaortae. Increasing the arterial dilation for each developmental time stepshifts the pressure–diameter curves upward (c), as observed in UNTSMKO aorta, with little effect on the circumferential stretch–stress rela-tionship (f)
123

Absence of Fibulin-4 in mouse arteries
A
B1 1.5 2
0
100
200
300
400
circ stretch
circ
str
ess
(kP
a) CTR − UNT
SMKO − LOS
SMKO − UNT
0 50 100 1500.5
1
1.5
2
2.5
3
dia
met
er (
mm
)
pressure (mmHg)
1 1.5 20
100
200
300
400
circ
str
ess
(kP
a)
0 50 100 1500.5
1
1.5
2
2.5
3
dia
met
er (
mm
)
pressure (mmHg)
circ stretch
C
D
CTR − UNT
SMKO − LOS
SMKO − UNT
Fig. 6 Mechanical changes in SMKO aorta can be reproduced by aconstrained mixture model of growth and remodeling. Starting fromidentical inputs for a 3 day-old mouse aorta, a previously publishedmodel (Wagenseil 2011) was used to reproduce pressure–diameter (a)and circumferential stretch ratio versus stress (b) relationships for a 30-day-old mouse aorta after growth and remodeling. Experimental datafrom Fig. 2a and Fig. 4a are replotted in c and d for comparison to the
model results. Original model parameters were used to calculate therelationships for the UNT CTR aorta. For both LOS and UNT SMKOaortae, the elastin material constant was reduced by 50 % and the colla-gen material constants were increased by 10 %. Additionally, for UNTSMKO aorta, the arterial dilation for each developmental time stepwas increased from 12 to 20 %. These illustrative changes qualitativelyreproduce the behavior of SMKO aorta
Table 2 Physiologic values calculated in the model using published parameter values for UNT CTR aorta and parameter variations that reproducethe pressure–diameter and circumferential stretch–stress behavior of LOS and UNT SMKO aortae
Shear stress(Pa)
Circ stress(kPa)
Axial stress(kPa)
Circ stretchratio
Axial stretchratio
Unl inner diam(ND)
Unl thickness(ND)
Unl length(ND)
UNT CTR 2.8 188 66 1.93 1.14 2.35 1.92 1.68
SMKO–LOS 2.8 125 46 1.84 1.10 2.45 2.00 2.21
SMKO–UNT 0.8 141 45 1.90 1.07 3.60 2.04 3.06
SMKO aorta values were obtained by applying a 50 % decrease in the elastin material constant and 10 % increases in the collagen material constants.For UNT SMKO aorta, the arterial dilation with each developmental time step was increased from 12 to 20 % compared to UNT CTR and LOSSMKO. All results start from the same 3 day-old mouse aorta and calculate growth and remodeling based on step changes in pressure and axialstretch for developmental time steps of 4.5 days up to a final age of 30 days. Relevant equations are in the “Appendix,” with additional informationin Wagenseil (2011). ND non-dimensional, final dimensions at 30 day olds/starting dimensions at 3 day olds
71 %, reduces the physiologic circumferential and axialstresses by 25 and 32 %, respectively, reduces the circum-ferential and axial stretch by 1 and 6 %, respectively, andreproduces the trends in the unloaded dimension changes(Table 2).
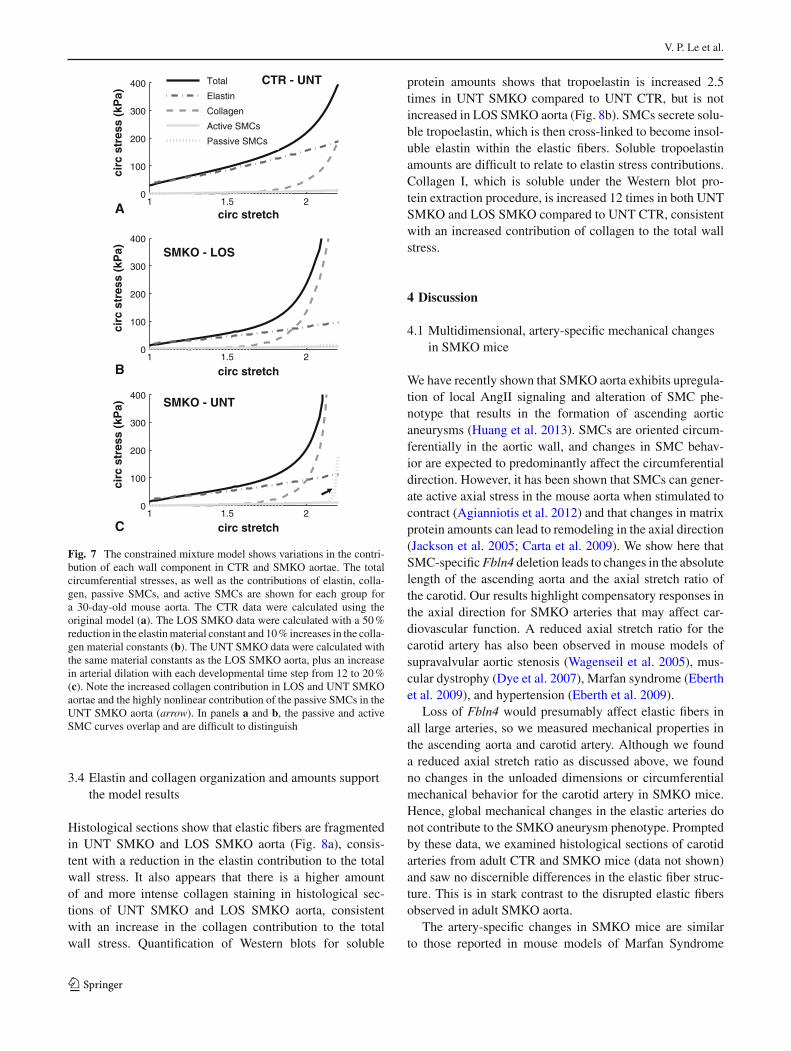
Figure 7 shows the circumferential stress contributionsof each component in the model for adult UNT CTR, LOSSMKO, and UNT SMKO aortae. For UNT CTR aorta,elastin contributes primarily at low stretch, collagen con-tributes at high stretch, and passive and active SMCs con-tribute very little to the total stress (Fig. 7a). This is consis-tent with expectations for large elastic arteries (Faury et al.
1999). For LOS SMKO aorta, collagen provides more of thestress contribution, with reduced contributions from elastin(Fig. 7b). This is consistent with our reasoning that frag-mented elastic fibers in SMKO aorta have reduced stress con-tributions, which are partially compensated for by increasedcollagen stress contributions. For UNT SMKO aorta, col-lagen and elastin look similar to LOS SMKO aorta, butthe passive SMC stress contribution becomes highly non-linear (Fig. 7c). This behavior does not occur at 18 %dilation or lower, which highlights how remodeling overtime can compound small changes in the model parame-ters.
123
V. P. Le et al.
1 1.5 20
100
200
300
400
circ stretch
circ
str
ess
(kP
a)
1 1.5 20
100
200
300
400
circ stretch
circ
str
ess
(kP
a)
1 1.5 20
100
200
300
400
circ
str
ess
(kP
a)
Total
Elastin
Collagen
Active SMCs
Passive SMCs
A
B
C
circ stretch
CTR - UNT
SMKO - UNT
SMKO - LOS
Fig. 7 The constrained mixture model shows variations in the contri-bution of each wall component in CTR and SMKO aortae. The totalcircumferential stresses, as well as the contributions of elastin, colla-gen, passive SMCs, and active SMCs are shown for each group fora 30-day-old mouse aorta. The CTR data were calculated using theoriginal model (a). The LOS SMKO data were calculated with a 50 %reduction in the elastin material constant and 10 % increases in the colla-gen material constants (b). The UNT SMKO data were calculated withthe same material constants as the LOS SMKO aorta, plus an increasein arterial dilation with each developmental time step from 12 to 20 %(c). Note the increased collagen contribution in LOS and UNT SMKOaortae and the highly nonlinear contribution of the passive SMCs in theUNT SMKO aorta (arrow). In panels a and b, the passive and activeSMC curves overlap and are difficult to distinguish
3.4 Elastin and collagen organization and amounts supportthe model results
Histological sections show that elastic fibers are fragmentedin UNT SMKO and LOS SMKO aorta (Fig. 8a), consis-tent with a reduction in the elastin contribution to the totalwall stress. It also appears that there is a higher amountof and more intense collagen staining in histological sec-tions of UNT SMKO and LOS SMKO aorta, consistentwith an increase in the collagen contribution to the totalwall stress. Quantification of Western blots for soluble
protein amounts shows that tropoelastin is increased 2.5times in UNT SMKO compared to UNT CTR, but is notincreased in LOS SMKO aorta (Fig. 8b). SMCs secrete solu-ble tropoelastin, which is then cross-linked to become insol-uble elastin within the elastic fibers. Soluble tropoelastinamounts are difficult to relate to elastin stress contributions.Collagen I, which is soluble under the Western blot pro-tein extraction procedure, is increased 12 times in both UNTSMKO and LOS SMKO compared to UNT CTR, consistentwith an increased contribution of collagen to the total wallstress.
4 Discussion
4.1 Multidimensional, artery-specific mechanical changesin SMKO mice
We have recently shown that SMKO aorta exhibits upregula-tion of local AngII signaling and alteration of SMC phe-notype that results in the formation of ascending aorticaneurysms (Huang et al. 2013). SMCs are oriented circum-ferentially in the aortic wall, and changes in SMC behav-ior are expected to predominantly affect the circumferentialdirection. However, it has been shown that SMCs can gener-ate active axial stress in the mouse aorta when stimulated tocontract (Agianniotis et al. 2012) and that changes in matrixprotein amounts can lead to remodeling in the axial direction(Jackson et al. 2005; Carta et al. 2009). We show here thatSMC-specific Fbln4 deletion leads to changes in the absolutelength of the ascending aorta and the axial stretch ratio ofthe carotid. Our results highlight compensatory responses inthe axial direction for SMKO arteries that may affect car-diovascular function. A reduced axial stretch ratio for thecarotid artery has also been observed in mouse models ofsupravalvular aortic stenosis (Wagenseil et al. 2005), mus-cular dystrophy (Dye et al. 2007), Marfan syndrome (Eberthet al. 2009), and hypertension (Eberth et al. 2009).
Loss of Fbln4 would presumably affect elastic fibers inall large arteries, so we measured mechanical properties inthe ascending aorta and carotid artery. Although we founda reduced axial stretch ratio as discussed above, we foundno changes in the unloaded dimensions or circumferentialmechanical behavior for the carotid artery in SMKO mice.Hence, global mechanical changes in the elastic arteries donot contribute to the SMKO aneurysm phenotype. Promptedby these data, we examined histological sections of carotidarteries from adult CTR and SMKO mice (data not shown)and saw no discernible differences in the elastic fiber struc-ture. This is in stark contrast to the disrupted elastic fibersobserved in adult SMKO aorta.
The artery-specific changes in SMKO mice are similarto those reported in mouse models of Marfan Syndrome
123

Absence of Fibulin-4 in mouse arteries
Fig. 8 Histological and biochemical data support the model assump-tions. Representative histological sections of UNT CTR, UNT SMKO,and LOS SMKO aorta stained with HE (for general tissue organization),Hart’s (for elastin), and Masson-Trichrome (for collagen) (a). Elasticfibers are thinner and fragmented in SMKO aorta, even after LOS treat-
ment. Bars are 50µm. Western blot analyses of the ascending aortashowing tropoelastin (Tropo E), collagen I (Col I), and GAPDH (loadingcontrol) (b). Right graphs show quantification. Bars are mean±SEM.∗ p < 0.05. NS not significant
that also develop ascending aortic aneurysms (Bunton et al.2001; Eberth et al. 2009). It has been suggested that dif-ferent embryonic origins of SMCs may account for localizedaneurysm development. SMCs in the ascending aorta and thecarotid arteries derive from the neural crest (Majesky 2007),so this cannot explain the differences we observed. How-ever, SMCs at the base of the ascending aorta near the aorticvalve derive from the secondary heart field, and these SMCsmay respond differently than neural crest-derived SMCs togenetic, chemical, or mechanical signals and propagate thesechanges to the rest of the ascending aorta. Alternatively, ithas been shown that shear stress, which varies with locationin the arterial tree, is an important determinant of arterialfunction and gene expression (Reneman and Hoeks 2008).Hence, shear stress may contribute to localized aneurysmdevelopment.
4.2 Artery and direction-specific effects of CAP and LOStreatment
Treatment for SMKO mice with CAP or LOS preventsaneurysm development. Although neither of the drugsreverse the decrease in aortic circumferential compliance,decreased compliance may still contribute to aneurysmpathophysiology. For example, in mouse models of Mar-fan syndrome, combined therapy with LOS and doxycyclineis more effective at preventing and treating aneurysms thaneither drug alone (Yang et al. 2010; Xiong et al. 2012). Doxy-cycline is a non-specific matrix metalloproteinase inhibitor.The combination therapy significantly improves elastic fiberorganization and decreases the number of elastic fiber breaks,which may increase aortic compliance. In CTR aorta, wefound that CAP and LOS treatments actually decrease the
123

V. P. Le et al.
aortic compliance compared to UNT CTR at high pressuresand the peak compliance shifts to lower pressures. CAP andLOS treatments have no effect on the compliance of the CTRcarotid artery. Since CAP and LOS reduce systolic bloodpressures in CTR mice (Huang et al. 2013), these drug-treatedaortae may have remodeled in response to lower physio-logic blood pressure. The peak compliance of the ascendingaorta occurs just below the physiologic pressure in develop-ing mice, even if the compliance and physiologic pressureare altered by reduced elastin levels (Le et al. 2011). Aorticremodeling may be an important consideration in the choiceof antihypertensive treatments, as reduced compliance wouldnot be a desired outcome (Cecelja and Chowienczyk 2009).
CAP and LOS treatment for SMKO arteries does notreverse the mechanical changes in the axial direction. Thisshows that altering the axial mechanics is not requiredfor aneurysm prevention, but does not exclude relation-ships between axial mechanics and aneurysm pathophysiol-ogy. Doxycycline prevents axial remodeling in arteries withdecreased axial stretch (Jackson et al. 2005). Affecting axialmechanics, increasing compliance, and inhibiting AngII maybe mechanisms through which combined doxycycline andLOS therapy improves aneurysm treatment and preventionin Marfan mouse arteries (Xiong et al. 2012; Yang et al.2010). Persistent increases in axial length and decreases inaxial stretch ratio can lead to arterial tortuosity (Jackson et al.2005). Tortuosity has been linked to flow abnormalities, clotformation, and stroke (Chesnutt and Han 2011) and coulddevelop in individuals where aneurysm formation has beenprevented, but arterial axial mechanical behavior has not beenaddressed.
4.3 Models of developmental arterial growth andremodeling
Mechanical models of arterial growth and remodeling havebeen used to predict general trends in development (Alford etal. 2008; Wagenseil 2011) and in adult arteries with alteredpressure or flow (Gleason et al. 2004; Taber and Eggers1996). To our knowledge, this is the first application of amodel to reproduce changes with a specific genetic defectand drug treatment. Our results show that increasing the arte-rial dilation for each time step, decreasing the elastin materialconstant, and increasing the collagen material constants qual-itatively reproduce the differences in pressure–diameter andcircumferential stretch–stress behavior between UNT CTRand UNT SMKO aorta. Behavior of LOS SMKO aorta isreproduced by rescuing the arterial dilation with each timestep, but maintaining the changes in elastin and collagen con-stants. The model provides rationale for how the mechanicalproperties (determined by the change in elastin and collagenmaterial constants) can be separated from the SMC pheno-type and aneurysm development (determined by the altered
arterial dilation response) to reproduce the observed differ-ences between UNT SMKO and LOS SMKO aorta.
From the experimental behavior of the composite aorticwall, one may assume that LOS does not change the circum-ferential stretch–stress behavior of SMKO aorta and conse-quently that rescuing the altered mechanical properties ofUNT SMKO aorta plays no role in aneurysm prevention.However, the contribution of each wall component in themodel shows that the passive SMC stress is different forUNT SMKO compared to LOS SMKO. The distribution ofstresses among wall components and increased SMC stressmay play a role in aneurysm development. The model resultslead to new hypotheses on the interactions between mechani-cal behavior and aneurysmal disease that cannot be generatedfrom the experimental data alone.
4.4 Relating model results to biological signaling
PROP, an antihypertensive medication that does not actthrough the AngII pathway, does not prevent aneurysms inSMKO mice (Huang et al. 2013). ACE, which converts AngIto AngII, is upregulated in SMKO ascending aorta and theACE inhibitor CAP prevents SMKO aneurysms. Treatmentwith the AngII receptor type I blocker LOS increases theexpression of SMC contractile genes and prevents aneurysmsin SMKO mice. Our model results suggest that increasedlocal AngII signaling and the resultant decrease in SMC con-tractile genes are directly related to arterial dilation with eachdevelopmental time step. Increased arterial dilation may becaused by a dysregulated response to flow changes in SMKOaorta. In vitro, endothelial cells and SMCs respond to flowby upregulating ACE (Gosgnach et al. 2000), which com-bined with the already increased ACE expression in SMKOaorta may exaggerate SMKO arterial dilation. AngII is apotent vasoconstrictor, so it is counterintuitive that increasesin AngII would increase arterial dilation. However, the vas-cular response to AngII depends on the secondary pathway.Specifically, hydrogen peroxide (H2O2) is induced by AngIIand can exert either a contraction or dilation response depend-ing on the vascular bed, contractile state, and cellular source(Nguyen Dinh Cat and Touyz 2011).
Remodeling of LOS SMKO aorta can be reproducedby rescuing the arterial dilation with no changes in elastinand collagen material constants compared to UNT SMKO,implying that LOS blocks local AngII signaling, but cannotrepair the elastic fibers. Because elastic fibers are assem-bled during late embryonic and early postnatal development,it is reasonable that postnatal LOS treatment cannot preventSMKO elastic fiber fragmentation. The elastic fiber fragmen-tation and subsequently reduced elastin material constant forUNT and LOS SMKO aorta are supported by histologicalevidence. Although our model shows that the elastin mater-ial constants can be the same, it is likely that there are some
123

Absence of Fibulin-4 in mouse arteries
differences between LOS and UNT SMKO aorta. Westernblot analyses show that there are differences between solu-ble tropoelastin amounts in UNT SMKO and LOS SMKOaorta. H2O2 solubilizes elastin (Umeda et al. 2001), whichmay increase soluble tropoelastin in UNT SMKO aorta.This may be prevented in LOS SMKO aorta by blockingAngII-induced H2O2 generation. Quantification of insolubleelastin amounts, elastin cross-linking, and fiber fragmenta-tion would add support to the model assumption of a reducedelastin stress contribution in SMKO aorta.
It was assumed that the collagen material constants wouldincrease to compensate for the reduced elastin stress con-tribution in SMKO aorta. This would increase the collagenstress contribution, as well as the nonlinearity of the stretch–stress relationship. Although difficult to quantify, histolog-ical evidence supports an increased amount of collagen inUNT and LOS SMKO aorta. Western blot analyses show adramatic increase in collagen I amounts in UNT and LOSSMKO compared to UNT CTR. Because of the nonlinearbehavior of collagen in the model, stress contribution cannotbe directly related to collagen amounts. Physical character-istics of the collagen fibers such as size, orientation, undu-lation, and cross-link density will influence the mechanicalbehavior. Changes in any of these factors in SMKO aortacould increase the nonlinearity of the collagen stretch–stressrelationship. The current model results provide new avenuesfor further research on the mechanical and biological factorsin SMKO aneurysm development.
4.5 Limitations
We use two models in the current work: a mouse model ofhuman disease and a mechanical model of growth and remod-eling in the aortic wall. Mice offer a convenient and well-controlled model where genetic and phenotypic changes canbe linked. However, they often do not recapitulate many fac-tors of the related human diseases and may not respond todrug treatments in the same manner as humans. Mechani-cal models offer a convenient and well-controlled methodfor investigating factors, such as the contribution of differ-ent wall components, that cannot easily be separated in theexperimental data. One limitation is the number of assump-tions and parameters that must be included in the model equa-tions. In this work, we did not try to fit model parameters tothe experimental data or make changes to the previous modelassumptions (Wagenseil 2011). The previous model has sev-eral limitations, including the separation of the radial growthfrom the blood flow and step changes in pressure and length,that must be addressed in future work. Our goal in this workwas to use the previous model for UNT CTR aorta and toinvestigate experimentally motivated changes in the modelparameters to provide illustrative examples that qualitativelyreproduce experimental data for UNT and LOS SMKO aorta.
Lastly, we provide histological images and protein quantifi-cation as supporting evidence for the model assumptions, butmore work is needed to relate the changes in stress contribu-tions to physical changes in elastin and collagen amount andorganization.
5 Conclusions
SMKO mice show axial remodeling in the ascending aortaand carotid artery, which is not reversed by AngII inhibitorsthat prevent aneurysms. However, the axial remodeling maybe linked to additional cardiovascular complications, suchas tortuosity. SMKO mice show circumferential changes inthe pressure–diameter behavior and stretch–stress behaviorthat are specific to the ascending aorta. Hence, circumfer-ential mechanical changes in the carotid artery do not con-tribute to aortic aneurysm development. A previously pub-lished constrained mixture model for developing mouse aortaqualitatively reproduces the pressure–diameter and circum-ferential stretch–stress relationships after growth and remod-eling for the adult CTR aorta. Increasing the arterial dilationwith each developmental time step from 12 to 20 %, alongwith a 50 % decrease in the elastin material constant and a10 % increase in the collagen material constants in the modelreproduce the behavior of UNT SMKO aorta. The behaviorof LOS SMKO aorta can be reproduced by rescuing the arter-ial dilation with each time step, while maintaining the alteredelastin and collagen material constants. Histological and bio-chemical data support the model assumptions. The modelresults link biological observations to mechanical responsesand suggest new hypotheses on the relationships betweenmechanical behavior and aneurysm development.
Acknowledgments This work was supported by NIH R01HL115560(JEW), R01HL105314 (JEW), R01HL106305 (HY), grants from theAmerican Heart Association (Grant-In-Aid, 0855200F, HY), and TheNational Marfan Foundation (HY426g). HY is a recipient of the Estab-lished Investigator Award from the American Heart Association. Wethank Jianbin Huang for his assistance in the drug treatment experi-ments.
Appendix
The aorta is considered a constrained mixture of wall com-ponents (k) where the total mean Cauchy stress (σ ) in thecircumferential (θ) and axial (z) direction is the sum of thestresses in each component (σ k
θ , σ kz ) multiplied by the mass
fraction (φk) of each component at time, s:
σθ (λθ , λz) =∑
k
φk(s)σ kθ (λk
θ , λkz ), (6a)
σz(λθ , λz) =∑
k
φk(s)σ kz (λk
θ , λkz ), (6b)
123

V. P. Le et al.
where λθ , λz are the stretch ratios of the mixture and λkθ , λ
kz
are the stretch ratios of each component. The componentshave individual homeostatic stretch ratios (λk
θh, λkzh) at which
they are produced and these values increase 3 % in the cir-cumferential direction and decrease 3 % in the axial direc-tion with each developmental time step (Wagenseil 2011).The unloaded stretch ratios of the mixture when the compo-nents are produced are λθu, λzu . The different stretch ratiosare related by
λkθ = λθλ
kθh
λθu, (7a)
λkz = λzλ
kzh
λzu. (7b)
The component stresses are defined by constitutive equationsfor elastin (e), collagen (c), and SMCs (m). SMCs have bothpassive (pas) and active (act) stress contributions (Gleasonand Humphrey 2004; Gleason et al. 2004):Elastin:
σ eθ (λe
θ , λez) = 2λe
θb1
(1 − 1
λe4
θ λe2z
), (8a)
σ ez (λe
θ , λez) = 2λe
zb1
(1 − 1
λe2
θ λe4z
), (8b)
Collagen:
σ cθ (λc
θ , λcz) = 2λc
θb2b3
(1 − 1
λc4
θ λc2z
)exp(Qc(λc
θ , λcz)),
(9)
σ cz (λc
θ , λcz) = 2λc
θb2
[b3
(1 − 1
λc2
θ λc4z
)+ 2b4
(λc2
z − 1)]
× exp(Qc(λcθ , λ
cz)), (10)
with
Qc(λcθ , λ
cz) = b3
(λc2
θ + λc2
z + 1
λc2
θ λc2z
− 3
)
+ b4
(λc2
z − 1)2
, (11)
SMCs:
σmθ = σm
θ,pas + σmθ,act , (12a)
σmz = σm
z,pas, (12b)
Passive SMCs:
σmθ,pas(λ
mθ , λm
z )
= 2λm2
θ
[b5
(1 − 1
λm4
θ λm2z
)+ 2b6b7
(λm2
θ − 1)]
× exp(Qm(λmθ )), (13)
with
Qm(λmθ ) = b7
(λm2
θ − 1)2
, (14)
σmz,pas(λ
mθ , λm
z ) = 2λm2
z b5
(1 − 1
λm2
θ λm4z
), (15)
Active SMCs:
σmθ,act (λ
mθ ) = Tact f̂ (λm
θ ),
with
f̂ (λmθ )=λm
θ
[1 −
(λM − λm
θ
λM − λ0
)2]
, Tact =TB −TQ, (16)
where b1−7 are passive material constants that increase 8 %with each developmental time step (Wagenseil 2011). λM , λ0
are active SMC material constants, TB = basal SMC toneconstant and TQ = SMC activation caused by changes inflow. TQ can be calculated according to (Gleason et al. 2004):
TQ = 1
φm f̂ (ε1/3Q λm
θ (0))(σm
θ,pas(sv) − σmθ,pas(0)d)
+ TB
(1 − f̂ (λm
θ (0)d)
f̂ (ε1/3Q λm
θ (0))
), (17)
where the SMC stretch ratios and stresses are functions of thetime elapsed since each step change in pressure, length, andflow. At time=0, the aorta is at its homeostatic state beforethe step change occurs and at time= sv , the instantaneousdilation response occurs. Additionally, d = ε
1/3Q ho/h(sv),
where ho = initial wall thickness at time=0.The components are continually produced with each
developmental time step. SMCs and collagen are also contin-ually degraded, but elastin is not because of its long half-life.Kinetic functions for the production (g) and the degradation(q) of each component are (Gleason and Humphrey 2004;Gleason et al. 2004):
gk(s) = 1 − exp[−K kg s/sh], (18a)
qk(s) = exp[−K kq s/sh], (18b)
where K kg and K k
q are the associated rate constants for eachcomponent and sh is the homeostatic time at which remodel-ing is complete. A rate constant of 6.9 allows almost completeturnover with about 0.1 % of the original component remain-ing. Total mass fractions (original+new components) at eachtime step are determined from previously published experi-mental data (Wagenseil 2011).
References
Agianniotis A, Rachev A, Stergiopulos N (2012) Active axial stress inmouse aorta. J Biomech 45(11):1924–1927. doi:10.1016/j.jbiomech.2012.05.025
123

Absence of Fibulin-4 in mouse arteries
Alford PW, Humphrey JD, Taber LA (2008) Growth and remodelingin a thick-walled artery model: effects of spatial variations in wallconstituents. Biomech Model Mechanobiol 7(4):245–262. doi:10.1007/s10237-007-0101-2
Bunton TE, Biery NJ, Myers L, Gayraud B, Ramirez F, Dietz HC (2001)Phenotypic alteration of vascular smooth muscle cells precedes elas-tolysis in a mouse model of Marfan syndrome. Circ Res 88(1):37–43
Carta L, Wagenseil JE, Knutsen RH, Mariko B, Faury G, Davis EC,Starcher B, Mecham RP, Ramirez F (2009) Discrete contributionsof elastic fiber components to arterial development and mechani-cal compliance. Arterioscler Thromb Vasc Biol 29(12):2083–2089.doi:10.1161/ATVBAHA.109.193227
Cecelja M, Chowienczyk P (2009) Dissociation of aortic pulse wavevelocity with risk factors for cardiovascular disease other than hyper-tension: a systematic review. Hypertension 54(6):1328–1336. doi:10.1161/HYPERTENSIONAHA.109.137653
Chesnutt JK, Han HC (2011) Tortuosity triggers platelet activa-tion and thrombus formation in microvessels. J Biomech Eng133(12):121004. doi:10.1115/1.4005478
Dasouki M, Markova D, Garola R, Sasaki T, Charbonneau N,Sakai L, Chu M (2007) Compound heterozygous mutations infibulin-4 causing neonatal lethal pulmonary artery occlusion, aor-tic aneurysm, arachnodactyly, and mild cutis laxa. Am J Med GenetA 143(22):2635–2641. doi:10.1002/ajmg.a.31980
Dye WW, Gleason RL, Wilson E, Humphrey JD (2007) Altered bio-mechanical properties of carotid arteries in two mouse models ofmuscular dystrophy. J Appl Physiol 103(2):664–672
Eberth JF, Gresham VC, Reddy AK, Popovic N, Wilson E, HumphreyJD (2009a) Importance of pulsatility in hypertensive carotid arterygrowth and remodeling. J Hypertens 27(10):2010–2021. doi:10.1097/HJH.0b013e32832e8dc8
Eberth JF, Taucer AI, Wilson E, Humphrey JD (2009b) Mechanics ofcarotid arteries in a mouse model of Marfan Syndrome. Ann BiomedEng 37(6):1093–1104. doi:10.1007/s10439-009-9686-1
Faury G, Maher GM, Li DY, Keating MT, Mecham RP, Boyle WA(1999) Relation between outer and luminal diameter in cannulatedarteries. Am J Physiol 277(5 Pt 2):H1745–1753
Gleason RL, Humphrey JD (2004) A mixture model of arterial growthand remodeling in hypertension: altered muscle tone and tissueturnover. J Vasc Res 41(4):352–363
Gleason RL, Taber LA, Humphrey JD (2004) A 2-D Model of flow-induced alterations in the geometry, structure and properties ofcarotid arteries. J Biomech Eng 126:371–381
Gosgnach W, Challah M, Coulet F, Michel JB, Battle T (2000) Shearstress induces angiotensin converting enzyme expression in culturedsmooth muscle cells: possible involvement of bFGF. Cardiovasc Res45(2):486–492
Huang J, Davis EC, Chapman SL, Budatha M, Marmorstein LY, WordRA, Yanagisawa H (2010) Fibulin-4 deficiency results in ascendingaortic aneurysms: a potential link between abnormal smooth musclecell phenotype and aneurysm progression. Circ Res 106(3):583–592.doi:10.1161/CIRCRESAHA.109.207852
Huang J, Yamashiro Y, Papke CL, Ikeda Y, Lin Y, Patel M, Inagami T,Le VP, Wagenseil JE, Yanagisawa H (2013) Angiotensin-convertingenzyme-induced activation of local Angiotensin signaling is requiredfor ascending aortic aneurysms in fibulin-4-deficient mice. ScienceTransl Med 5(183):183ra158. doi:10.1126/scitranslmed.3005025
Jackson ZS, Dajnowiec D, Gotlieb AI, Langille BL (2005) Partialoff-loading of longitudinal tension induces arterial tortuosity. Arte-rioscler Thromb Vasc Biol 25(5):957–962
Le VP, Knutsen RH, Mecham RP, Wagenseil JE (2011) Decreased aorticdiameter and compliance precedes blood pressure increases in post-natal development of elastin-insufficient mice. Am J Physiol HeartCirc Physiol 301(1):H221–229. doi:10.1152/ajpheart.00119.2011
Majesky MW (2007) Developmental basis of vascular smooth musclediversity. Arterioscler Thromb Vasc Biol 27(6):1248–1258. doi:10.1161/ATVBAHA.107.141069
McLaughlin PJ, Chen Q, Horiguchi M, Starcher BC, Stanton JB,Broekelmann TJ, Marmorstein AD, McKay B, Mecham R, Naka-mura T, Marmorstein LY (2006) Targeted disruption of fibulin-4abolishes elastogenesis and causes perinatal lethality in mice. MolCell Biol 26(5):1700–1709
Nguyen Dinh Cat A, Touyz RM (2011) Cell signaling of angiotensin IIon vascular tone: novel mechanisms. Curr Hypertens Rep 13(2):122–128. doi:10.1007/s11906-011-0187-x
Reneman RS, Hoeks AP (2008) Wall shear stress as measured in vivo:consequences for the design of the arterial system. Med Biol EngComput 46(5):499–507. doi:10.1007/s11517-008-0330-2
Taber LA, Eggers DW (1996) Theoretical study of stress-modulatedgrowth in the aorta. J Theor Biol 180:343–357
Umeda H, Nakamura F, Suyama K (2001) Oxodesmosine and isoox-odesmosine, candidates of oxidative metabolic intermediates of pyri-dinium cross-links in elastin. Arch Biochem Biophys 385(1):209–219. doi:10.1006/abbi.2000.2145
Van Doormaal MA, Kazakidi A, Wylezinska M, Hunt A, TremoledaJL, Protti A, Bohraus Y, Gsell W, Weinberg PD, Ethier CR (2012)Haemodynamics in the mouse aortic arch computed from MRI-derived velocities at the aortic root. J R Soc Interf 9(76):2834–2844.doi:10.1098/rsif.2012.0295
Wagenseil JE (2011) A constrained mixture model for developingmouse aorta. Biomech Model Mechanobiol 10(5):671–687. doi:10.1007/s10237-010-0265-z
Wagenseil JE, Nerurkar NL, Knutsen RH, Okamoto RJ, Li DY, MechamRP (2005) Effects of elastin haploinsufficiency on the mechani-cal behavior of mouse arteries. Am J Physiol Heart Circ Physiol289(3):H1209–1217. doi:10.1152/ajpheart.00046.2005
Wan W, Yanagisawa H, Gleason RL Jr (2010) Biomechanical andmicrostructural properties of common carotid arteries from fibulin-5 null mice. Ann Biomed Eng 38(12):3605–3617. doi:10.1007/s10439-010-0114-3
Xiong W, Meisinger T, Knispel R, Worth JM, Baxter BT (2012) MMP-2regulates Erk1/2 phosphorylation and aortic dilatation in Marfan syn-drome. Circ Res 110(12):e92–e101. doi:10.1161/CIRCRESAHA.112.268268
Yang HH, Kim JM, Chum E, van Breemen C, Chung AW (2010) Effec-tiveness of combination of losartan potassium and doxycycline ver-sus single-drug treatments in the secondary prevention of thoracicaortic aneurysm in Marfan syndrome. J Thorac Cardiovasc Surg140(2):305–312. doi:10.1016/j.jtcvs.2009.10.039
123