· (mdr, rezisztencia legalább izoniazidra és rifampicinre) terjedése játszik kiemelked...
TRANSCRIPT

������������� �����������������
������������������
�
��������������� ��������� ������������������ �������������������������� �������������������������������
����
���� !�!�" #���$��"�!����� ������������%"��� ������������������ ������������������!������ ������
�����
��
�&�� ����������������!�!�!�����!����!���������������
�����'!�(�)**+�

Tartalomjegyzék
I. Bevezetés…………………………………………………………………………………….1
II. Irodalmi áttekintés……………………………………………………………………...…2
II.1. A tuberkulózis…………………………………………………………………………….2
II.1.1. A Mycobacterium tuberculosis baktérium…………………………………………...…5
II.1.2. Az antituberkulotikumok és a tuberkulózis kezelése…………………………….….….6
II.2. Kémiai módszerek leírása………………………………………………………….……..7
II.2.1. Szilárd fázisú peptidszintézis………………………………………………………..….7
II.2.2. Boc/Bzl módszer………………………………………………………………………..8
II.2.3. Fmoc/tBu módszer………………………………………………………………………9
II.2.4. Gyanták………………………………………………………………………………..10
II.2.5. Peptidkötés kialakítása a szintézis során………………………………………………11
II.2.6. Kapcsolási reakciók követése………………………………………………………….12
II.2.7. A peptid lehasítása a gyantáról és az oldallánc véd�csoportok eltávolítása…………..12
II.3. Hordozó molekulák és hatóanyag-konjugátumok alkalmazása…………………………13
II.3.1. Hordozóként alkalmazható tuftsin származékok………………………………………14
II.3.2. A M. tuberculosis immundomináns 16 kDa fehérje és a 91-106
(91SEFAYGSFVRTVSLPV106) T-sejt epitóppeptid…………………………………….……14
II.4. Antituberkulotikumok és konjugátumaik minimális gátló koncentrációjának
meghatározása folyékony táptalajon, telepszámának meghatározása szilárd táptalajon……..15
II.4.1. Baktériumok oltása és tenyésztése, a munka során használt tápközegek…….………..15
II.4.2. Baktériumok sejtszámának meghatározása……………………………………………17
II.4.3. Minimális gátló koncentráció (MIC) meghatározása………………………………….17
II.4.4. Telepszám (kolónia szám, colony forming unit, CFU) meghatározása……………….18
II.5. A sejtek életképességének vizsgálata antituberkulotikum (INH), hordozómolekulák és
antituberkulotikum-konjugátumok hatására, az in vitro citosztatikus hatás meghatározása
kolorimetriás tetrazólium (MTT) teszt alkalmazásával………………………………………19
III. Célkit�zések………………………………………………………………………….….21
IV. Kísérleti rész……………………………………………………………………………..22
IV.1. A szintetikus munka során használt vegyszerek,
m�szerek…………………………….22
IV.2. A peptidek szintézise…………………………………………………………….……..24

IV.2.1. A peptidek szintézise manuális Boc/Bzl stratégiával………………………………...24
IV.2.2. Peptidszintézis Fmoc/tBu stratégiával automata peptidszintetizátoron………………26
IV.3. A hatóanyag-konjugátumok szintézise…………………………………………………27
IV.3.1. 2-(izonikotinoil-hidrazino)ecetsav szintézise………………………………………...27
IV.3.2. 2-(izonikotinoil-hidrazino)ecetsav konjugálása peptidekhez………………………...28
IV.4. Tisztítási és analitikai módszerek………………………………………………………28
IV.4.1. A hatóanyag-konjugátumok/peptidszármazékok tisztítása…………………………...28
IV.4.2. Peptidek és konjugátumok analitikai jellemzése……………………………………..29
IV.4.2.1. Analitikai RP-HPLC vizsgálatok…………………………………………………..29
IV.4.2.2. Peptidek és konjugátumok aminosavanalízise……………………………………..29
IV.4.2.3. ESI-MS……………………………………………………………………………..30
IV.4.3. Elemanalízis…………………………………………………………………………..30
IV.4.4. Olvadáspont meghatározás……………………………………………………...……30
IV.5. Az INH, a hordozómolekulák és az INH-konjugátumok minimális inhibiciós
koncentrációjának (MIC) és telepszámának (CFU) meghatározása………………………….30
IV.6. A sejtek életképességének vizsgálata kolorimetriás tetrazólium (MTT) teszt
alkalmazásával………………………………………………………………………………..31
V. Eredmények……………………………………………………………………..………..33
V.1. Hordozó peptidek tervezése és szintézise……………………………………………….33
V.2. Hatóanyag-konjugátumok szintézise…………………………………………………....34
V.2.1. Az izoniazid származékok el�állítása távlságtartó „linker” beépítésével ……… ….34
V.2.2. Izoniazid származék konjugálása peptidekhez………………………………………..35
V.3. A hordozók, hatóanyagok és konjugátumok in vitro biológiai aktivitásának vizsgálata..37
V.3.1. Minimális inhibiciós koncentráció (MIC) és telepszám (CFU) meghatározása………37
V.3.2. Citosztatikus hatás vizsgálata………………………………………………………….38
VI. Összefoglalás………………………………………….…………………………………39
VII. Rövidítésjegyzék………….…………………………………………………………….40
VIII. Irodalomjegyzék............................................................................................................43
X. Köszönetnyilvánítás
IX. Függelék

I. Bevezetés
A makrofágok számos intracelluláris parazita köztes gazdái, ilyen a tuberkulózis
(tbc) megbetegedést okozó baktérium, a Mycobacterium tuberculosis. A tbc, amely
továbbra is komoly népegészségügyi kihívást jelent az egész világon, kialakulhat
bármelyik szervben, de a tüd�tbc a leggyakoribb megbetegedés. Ezek az obligát aerob,
lassan szaporodó baktériumok a makrofágok fagoszómáiban képesek kikerülni a szervezet
védekez� mechanizmusait. A betegség kezelésében alkalmazott vegyületek
(antituberkulotikumok) bejutása az érintett makrofágba els�sorban diffúzióval történhet,
meglehet�sen korlátozott mértékben. A hosszú kezelés során (1. fázis: 2-6 hónap, 2. fázis:
6-12 hónap) számolni kell mellékhatásaikkal, pl. a szervezet egészét érint� nem specifikus
toxicitásukkal, valamint gyors kiválasztódásukkal.
A peptidtípusú hordozók alkalmazása csökkentheti a terápiás szerek mellékhatásait,
valamint lehet�séget kínál a hatóanyagok szelektív bejuttatására a fert�zött makrofágokba.
A dolgozatban összefoglalt munka a terápiásan alkalmazott izoniazid antituberkulotikum
peptidtípusú hordozókhoz való kémiai konjugálását, valamint ezen új vegyületek kémiai
jellemzését és in vitro biológiai aktivitásának vizsgálatát foglalja össze.
Munkánk során hordozóként tuftsinszármazékokat (GTKPKG (T6), [TKPKG]4
(T20)) és az M. tuberculosis immundomináns 16 kDa fehérjének egy T-sejt epitóppeptidjét
(91SEFAYGSFVRTVSLPV106 (91-106)) választottunk. A felsorolt peptideket szilárdfázisú
szintézissel, Fmoc/tBu-, vagy Boc/Bzl-stratégiával, szintetizátoron illetve manuális
technikával állítottuk el�. Megterveztük, el�állítottuk és jellemeztük az izoniazid
kapcsolásához szükséges „távolságtartó” (linker) egységet is tartalmazó izoniazid
származékot ((2-(izonikotinoil-hidrazino)ecetsav)) és alkalmazásával sikeresen
végrehajtottuk a peptidekhez való konjugálást PyBOP/HOBt/DIEA kapcsolóreagensek
jelenlétében. A hordozókat és az új izoniazid konjugátumokat kémiailag jellemeztük RP-
HPLC, aminosavanalizis és tömegspektrometria alkalmazásával. Az új vegyületek
antituberkulotikus hatását, minimális inhibiciós koncentráció (MIC) értékének
meghatározását M. tuberculosis H37Rv törzs szuszpenzión vizsgáltuk Sula félszintetikus,
folyékony táptalajon. Meghatároztuk az új vegyületek citosztatikus hatását humán
hepatoma sejtvonalon (HepG2) kolorimetriás tetrazólium (MTT) teszt alkalmazásával.
Kísérleteink bizonyították, hogy a konjugátumokban az izoniazid meg�rizte in vitro
antituberkulotikus aktivitását és a vizsgált koncentrációkban a hordozók, konjugátumok
nem mutattak citosztatikus hatást HepG2 sejteken.

II. Irodalmi áttekintés II.1. A tuberkulózis
A tuberkulózis (tbc) fert�z� betegség, melyet a Mycobacterium tuberculosis baktérium
okoz. A baktérium els�sorban a tüd�t támadja meg (pulmonáris tbc. 2. ábra), de
megtámadhatja a központi idegrendszert (meningitisz), a nyirokrendszert és a keringési
rendszert (miliáris tbc), az ivarszerveket/hugyutakat, csontokat és izületeket is. A beteg
tüdejében rögök keletkeznek, innen a betegség magyar neve: güm�kór. Az M. tuberculosis
leggyakrabban csepp- és porfert�zést követ�en a tüd�be kerül, ahol az alveoláris makrofágok
bekebelezik. A baktériumok intracellulárisan szaporodnak, mivel gátolják a fagolizoszóma
kialakulását. A makrofágok reaktív oxigéngyökök, N-oxidok, lizozim, hidrolitikus enzimek
segítségével bontják le a baktériumokat savas vezikulumaikban. A fert�zött makrofágok
elpusztulnak, majd a kiszabaduló baktériumok újabbakat fert�znek (Gergely 1994; Frieden et
al., 2003). A fert�zött makrofágok eljuthatnak a környez� nyirokcsomókba, illetve a véráram
útján egyéb szövetekbe is (csontvel�, lép, vesék, központi idegrendszer). A legtöbb
Mycobacterium lassan szaporodik, 12-24 óra az osztódási id�, így ezek a fajok (pl. M.
tuberculosis, M. avium-intracellulare) 3-6 hét alatt képeznek telepeket (Ádám et al., 2003).
A tuberkulózis a történelem során az egyik legtöbb halálesetet okozó betegség,
napjainkban is több mint kétmilliárd ember fert�zött a baktériummal. A tuberkulózis ismételt
fellángolásában els�sorban gazdasági és szociális tényez�k, a HIV vírus és a tuberkulózis
közötti szinergizmus (világszerte az AIDS betegek 15%-a hal meg tuberkulózisban), és a XX.
század végén újabb komoly fenyegetésként megjelent multidrog rezisztens tuberkulózis
(MDR, rezisztencia legalább izoniazidra és rifampicinre) terjedése játszik kiemelked�
szerepet. Világszerte évente mintegy 8 millió ember betegszik meg újonnan és 2 millió ember
hal meg csak tuberkulózis következtében. A baktériummal fert�zöttek kb. 90%-a tünetmentes,
látens tbc-fert�zöttséggel él (LTBC (Kusner 2005)), 10% az esélye annak, hogy kifejl�dik
benne a betegség, amely, ha nem kezelik, 50%-os eséllyel vezet halálhoz. A tuberkulózis
tehet� felel�ssé az elkerülhet� feln�ttkori halálokok 25%-ért. A betegek 80%-a gyermekkorú
vagy a tanulás illetve munkavégzés szempontjából legproduktívabb 15-59 éves. Az
Egészségügyi Világszervezet el�rejelzése alapján 2000 és 2020 között közel 1 milliárd ember
válik újonnan fert�zötté és ezek közül várhatóan 200 millió egyénben ez a fert�zés
megbetegedéshez vezet majd és 35 millió beteg menthetetlenül meghal amennyiben a
tuberkulózis elleni védekezés nem er�södik számottev�en a jöv�ben. A HIV fert�zöttek

esetében ugyancsak a fert�zöttek 10%-nál várható tuberkulózis megbetegedés kialakulása,
mégpedig az expozíciót követ� egy éven belül. Évi mintegy 1,5 millió új megbetegedés a
szub-szaharai Afrikából származik, és ez a mutató a HIV/AIDS epidémiának köszönhet�en
évr�l évre drasztikusan emelkedik. Ázsiában (India, Kína) közel évi 3 millió új eset
jelentkezik, míg Kelet-Európában évente negyedmillió új megbetegedést regisztrálnak
(Kaufmann and Parida 2007) (1. ábra).
1. ábra. A tuberkulózis incidencia 2005-ben. A térkép megtalálható a http://gamapserver.who.int/mapLibrary/
honlapon
Hazai viszonyok mellett a feln�ttek között a legsúlyosabb kockázati tényez� a
szegénység, a hiányos táplálkozás, alkoholizmus. A kockázati tényez�k mellett azonban az
elmúlt évek statisztikája azt a meglep� megállapítást tette, hogy az új betegek több mint 40
százalékánál semmiféle kockázati tényez�t nem lehetett kimutatni (Jónás et al., 2005).
Viszonylag kiegyensúlyozott körülmények között él� egyének, akiknél a fert�z� forrást sem
lehetett kideríteni aktív tuberkulózisban betegedtek meg. Nem ritka, hogy csak a boncolás
fedezi fel a nem is sejtett güm�kórt. Számos megfigyelés azt bizonyítja, hogy akcidentális
fert�zés, zárt légtérben (repül�gép, rosszul szell�zött szállás, kocsma) kiinduló helye lehet a
megbetegedésnek.

� �� �
2. ábra: (A) Egy aktív tuberkulózisban szenved� beteg mellkas-röntgen felvétele, (a kép eredetije a www.mcd.hu_pubmed/2006/0609/183.htm honlapon található meg) (B) a tbc incidencia Magyarországon 2006-ban, forrás: www.koranyi.hu (Jónás et al., 2005)
Magyarországon javultak a megbetegedési adatok és összességében tovább javult a
tuberkulózis incidencia (2. ábra), de ez nem ad okot a megnyugvásra. Több megyénkben
emelkedett a megbetegedések száma (pl. Vas megye: 15%ooo – 25,2%ooo) és a
veszélyeztetett populációkban (pl. hajléktalanok) a helyzet vésztjóslóan súlyosabb.
Hazánkban a sz�rési fegyelem lazulása mellett az alkoholizmus, a romló szociális helyzet, a
munkanélküliség játszanak szerepet az incidencia emelkedésében az ország egyes területein.
A sz�r�vizsgálattal kiemeltek korcsoportos vizsgálata azt mutatja, hogy a legmagasabb arány
az új tbc-s esetek között a fiatal és középkorú korcsoportokban van. A megbetegedések
el�fordulása jelent�s földrajzi különbségeket is mutat (2. ábra), els�sorban a szociális helyzet,
a bevándorlás, illetve határokon keresztüli migráció, valamint a sz�rési fegyelem
meglazulásának következményeként. A pulmonáris tuberkulózis incidenciája tartósan a
legmagasabb Szabolcs-Szatmár-Bereg megyében (50,3 eset/100 000 lakos), de a szomorú
statisztikában kiemelkedik, Hajdú-Bihar (34,2 eset /100 000 lakos), Jász-Nagykun-Szolnok
(34,1 eset /100 000 lakos), Borsod-Abaúj-Zemplén (27,7 eset/100 000 lakos) megye illetve
Budapest (40,24 eset /100 000 lakos) is (Jónás et al., 2005).
Az idejében megkezdett antituberkulotikum kezelés célja a teljes gyógyulás elérése a
fert�z�képesség lehet� leggyorsabb megszüntetésével, a másodlagos gyógyszerrezisztencia
kialakulásának megakadályozásása, a teljes gyógyulás elérése.

II.1.1. A Mycobacterium tuberculosis baktérium
A Mycobacteriumok sejtfala komplexebb, mint a Gram-pozitív és Gram-negatív
baktériumoké. A sejtfalban a peptidoglikánvázhoz a D-arabinozon és a D-galaktózon
keresztül kovalens kötéssel mikolsav kapcsolódik, amely a legjelent�sebb lipid a
Mycobacteriumok sejtfalában (Brennan 2003). Egy másik sejtfal lipid, a 6,6’-
dimikoliltrehalóz, az ún. „cord-faktor” felel�s a baktériumok egymás mellé rendez�déséért,
amely a M. tuberculosis virulens törzseire jellemz�. A sejtfal küls� rétegeinek peptidláncai
képezik a sejtfal tömegének 15%-át és a biológiailag fontos antigének is itt találhatók. Ezek
felel�sek a celluláris immunválasz kiváltásáért. A baktériumok felszíne hidrofób, ezzel a
tulajdonságukkal függ össze, hogy ellenállóak a dezinficiensekkel szemben (Ádám et al.,
2003).
A M. tuberculosis mikroszkóp alatt festési jellegzetességei alapján azonosítható; savas
oldattal való kezelés után megtartja színezetét, ezért saválló baktériumként tartják nyilván. A
leggyakoribb festési eljárás, a Ziehl-Neelsen festés során élénkpirosra fest�dik (Ádám et al.,
2003), ami kit�n�en elválik a kék háttért�l (3.ábra). A saválló baktériumok fluoreszcens
mikroszkóppal is jól megfigyelhet�k (3.ábra).
� ��� ��
3. ábra: (A) A M. tuberculosis mikroszkópos képe Ziehl-Neelsen festést követ�en, forrás: www.kimicontrol.com/edu-e.html (B) M. tubercolosis baktérium fluoreszcens mikroszkóppal, forrás: www.tudobeteg.hu/news/061229tbc.html

II.1.2. Az antituberkulotikumok és a tuberkulózis kezelése
A M. tuberculosis szaporodását befolyásoló tényez�k a környezet oxigenizációja és
vegyhatása. A baktérium anyagcseréjének aktivitása és szaporodási frekvenciája is a
makrofágokon kívül, a tüd� kavernákban a legmagasabb, ahol magas az O2- tenzió és enyhén
lúgos pH uralkodik. A makrofágokban és hegszövetekben a baktérium számára el�nytelenek a
viszonyok (enyhén savas pH). A sajtos, nekrotikus gócok pedig pH semlegesek. A környezet
pH-ja és a baktérium szaporodási frekvenciája is dönt�en befolyásolja az
antituberkulotikumok aktivitását. Az izoniazid (INH), a rifampicin (RAMP) és a
sztreptomicin (SM) a leghatékonyabbak neutrális és enyhén lúgos közegben, gyorsan
szaporodó baktérium populációkkal szemben. A RAMP a semleges pH-jú sajtos gócokban
lassan, intermittálva szaporodó baktériumokkal szemben is kimagasló aktivitással
rendelkezik. A pirazinamid (PZA) intracellulárisan, savanyú közegben fejti ki a hatását. A
leggyakoribb antituberkulotikumok feltételezett hatását a különböz� baktériumpopulációkra a
4. ábrán tüntettem fel (Egerszegi 1995).
������������� ����������� ��������� �������� ��
���� ��� ������
�����
���������� ���������������������
�����
������������� ����� ��
������������� �������� �� ������������ ���������
�� ����� � � �
� ���
������������� ����������� ��������� �������� ��
���� ��� ������
�����
���������� ���������������������
�����
������������� ����� ��
������������� �������� �� ������������ ���������
�� ����� � � �
� ���
4. ábra. A legyakrabban alkalmazott antituberkulotikumok feltételezett hatása az egyes baktérium populációkra
(Egerszegi 1995)
A sokéves tapasztalat alapján mára az intenzív, 2 fázisból álló antituberkulotikus
kezelés vált ajánlottá (1. fázis: 2-6 hónap, 2. fázis: 6-12 hónap). Az els� két hónapban négyes

kombináció (INH-RAMP-PZA-ETB (etambutol) vagy SM), a második fázisban pedig négy
hónapig INH-RAMP kombináció adása ajánlott (Tripathi et al., 2005; Zhang 2005).
Az INH sok szempontból optimálisnak mondható szer, ezért használata széles körben
elterjedt. Hatása abban áll, hogy a mikobakteriális sejtfal fontos épít�elemének (a mikol-
savnak) a szintézisét gátolja, a membránpermeábilitás megváltozását eredményezve.
Metabolitként (INH NAD) viselkedve a M. tuberculosis metabolizmusát is megzavarja.
Baktericid hatású (intra- és extracelluláris baktériumokra is), a gyorsan szaporodó
baktériumpopulációkra a leghatékonyabb.
Az izoniazid egy prodrug, mely aktivációt igényel a bakteriális sejtben a
kataláz/peroxidáz enzim (KatG géntermék) által. A terápiásan inaktív formáját az N-
acetiltranszferáz (NAT) enzim hozza létre úgy, hogy acetilezi a molekulát (Sandy et al., 2005)
(5. ábra).
N
NHNH2
ON
NHNH
O
N
NHNH CH3
O
O
KatG
NAT
Aktív
Inaktív
Me
SCoA
O
CoASH
5. ábra. Az INH aktiválódásának és inaktiválódásának sematikus ábrája (Sandy et al., 2005)
II.2. Kémiai módszerek leírása
Munkám során lineáris peptidamidokat állítottam el�, ezért röviden összefoglalom az
általam használt szilárdfázisú szintézis stratégiák (Boc/Bzl; Fmoc/tBu) elméleti hátterét.
II.2.1. Szilárd fázisú peptidszintézis
A szilárd fázisú peptidszintézis technikájának bevezetése Merrifield nevéhez köthet�
(Merrifield 1963; Merrifield 1964). A módszer lényege, hogy a peptidláncot szilárd hordozón
α-aminosavszármazékok lépésenkénti összekapcsolásával építjük fel, C-terminális – N-

terminális irányban. A szilárd hordozó egy oldhatatlan polimer, melyhez az
aminosavszármazékokat egy reaktív csoporton keresztül kovalensen kapcsoljuk.
A szintézis során olyan aminosavszármazékokat használunk, amelyek nukleofil
oldalláncaikon állandó véd�csoportokkal, az α-NH2 csoportjaikon pedig átmeneti
véd�csoportokkal vannak ellátva. Egy adott poziciójú aminosavszármazék kapcsolása el�tt az
el�z�, gyantára kapcsolt aminosavszármazékról eltávolítjuk az átmeneti Nα-véd�csoportot és
a kapcsolni kívánt aminosavszármazék karboxil-terminálisát aktiváljuk. Ezen átmeneti
véd�csoport hasítási lépések és a kapcsolási reakciók sorozatos ismétlésével építjük fel a
peptidláncot. A szintézis végén a peptidet lehasítjuk a hordozóról és ezzel egy id�ben
távolítjuk el az állandó véd�csoportokat is.
A szilárdfázisú peptidszintézisnek két, széles körben elterjedt stratégiája a terc-
butiloxi-karbonil/benzil (Boc/Bzl) módszer és a 9-fluorenil-metiloxi-karbonil/terc-butil
(Fmoc/tBu) módszer. A két módszer alapelvének egyszer�sített sémája a 6. ábrán látható.
CH2OC
O
NHCH
CNH
O
H2CC
O
O
CH3
CH3
CH3
R
H
TFA
Piperidin
Fmoc
CH2O
COH2C
CHC
NH
O
RNHC
O
O
CH3
CH3
CH3HF
TFA
Boc
6. ábra. A Boc/Bzl és az Fmoc/tBu módszer összehasonlítása
II.2.2. Boc/Bzl módszer
Ez a módszer a különböz� er�sség� savakra érzékeny (átmeneti) és kevésbé érzékeny
(állandó) véd�csoportokon alapul. A terc-butiloxi-karbonil (Boc) véd�csoport lúgokkal és
nukleofil reagensekkel szemben stabil, de szervetlen vagy szerves savakkal eltávolítható
(Barany et al., 1987). Hasító elegyként leggyakrabban 30-50 % V/V TFA-t tartalmazó DCM
használatos, 20-30 perces reakció id�vel. Mivel az acidolízis során az Nα-aminocsoport
trifluor-acetát sója jön létre, ezért a hasítás után semlegesítenünk kell. Ekkor tercier-amin
hatására jön létre a szabad Nα-aminocsoport. A leggyakrabban használt semlegesít� elegy 5-
10 % V/V diizopropil-etil-amin (DIEA) DCM-os oldata, 3-5 perces reakció id�vel.

A Boc/Bzl módszernél az oldallánc véd�csoportok a benzil-alkohol éter-, észter- ill.
uretán- származékai (Grant 1992b). Ezek az oldallánc véd�csoportok csak er�s savak
jelenlétében hasíthatóak, tehát a Boc-csoport lehasítása során stabilak maradnak.
II.2.3. Fmoc/tBu módszer
Ezt a szintézis stratégiát két kutatócsoport egymástól függetlenül és párhuzamosan
fejlesztette ki, kiküszöbölend� a Boc/Bzl stratégia hátrányait: 1. az állandó és átmeneti
véd�csoportok is savérzékenyek, 2. a gyantáról való hasításhoz és az állandó véd�csoportok
hasításához a speciális készüléket igényl� HF alkalmazása (Carpino and Han 1972). Ezen
módszer esetében az aminosav-származékok átmeneti véd�csoportja (az általam szintetizált
peptidek esetében) a 9-fluorenil-metoxi-karbonil csoport (Fmoc), mely savakkal szemben
stabil, bázisokkal szemben viszont labilis. A hasítás történhet 20-55 % V/V piperidin/ DMF
vagy NMP elegyével 10-18 perces reakció id�vel (Atherton et al., 1978), (Albericio et al.,
1990), (Chang et al., 1980). Az Fmoc-csoport piperidinnel történ� eltávolításának reakciója a
7. ábrán látható. A hasításkor dibenzofulvén átmeneti termék keletkezik, mely reaktív, de a
piperidinnel stabil adduktot képez, ezáltal elkerülhet�ek az alkilez�dési mellékreakciók.
Bizonyos peptidszekvenciák esetében a kísérletek azt igazolták, hogy a 2% piperidint
és 2% 1,8-diazabiciklo[5.4.0]-undec-7-ént (DBU) tartalmazó eleggyel gyors, hatékony hasítás
érhet� el, ezen kívül csökkenthet� az enantiomerizáció valószín�sége és nagyobb az effektív
hasítás mértéke térbeli gátlás esetén (Newcomb et al., 1998), (Tickler et al., 2001), (Wade et
al., 1991). Az állandó oldallánc véd�csoportok (tBu, OtBu, Boc, Trt, stb) hasítása és a
peptidek gyantáról való eltávolítása általában TFA-val történik (Grant 1992b). Mivel a hasítás
során reaktív karbokationok keletkeznek, gyökfogók használata szükséges.

OO
NH
R'
R
O
NH
CH2
+ CO2 + R'NH2
R
O
NH
N
7. ábra. Az Fmoc-csoport piperidinnel történ� eltávolítása.
II.2.4. Gyanták
A hordozókkal, mint polimerekkel szemben elvárás, hogy jól duzzadjanak, optimális
legyen a sz�réssebességük, ne lépjenek reakcióba az oldószerekkel, a reagensekkel és az
épül� peptidlánccal.
A gyanta fontos jellemz�je a kapacitás. A ~1,00 mmol/g kapacitású gyanták a
viszonylag rövid peptidek szintézisére alkalmasak, míg az alacsonyabb szubsztitúciójú
gyantákat az elágazó ill. hosszú láncú peptidek szintézisénél használják. A Boc/Bzl
módszernél alkalmazott gyanták, melyekr�l savas hasítással szabad C-terminálisú peptidek
keletkeznek, nukleofilekkel történ� reakció után pedig peptidaminok, peptidhidrazinok, ill.
peptidészterek állíthatóak el�.
A peptidamidok el�állítására használt gyanták a benzhidril-amin (BHA) és a p-metil-
benzhidril-amin (MBHA) (Matsueda and Stewart 1981). Az Fmoc/tBu módszernél
leggyakrabban alkalmazott gyanták, melyek szabad karboxil-terminálisú peptideket
eredményeznek, a polisztirol alapú 4-benziloxi-benzilalkohol (Wang 1973) vagy a 4-
hidroximetil-fenoxi-ecetsav (Atherton et al., 1981) funkciós csoportot tartalmazóak. Ezen
kívül elterjedt még a 4-(2’,4’-dimetoxi-fenil-Fmoc-aminometil)-fenoxi (Rink-amid) gyanta,
amely esetében a hasítási reakció peptidamidot eredményez (Rink 1978).

II.2.5. Peptidkötés kialakítása a szintézis során
A peptidkötést a gyantához kötött szabad N-terminálisú aminosav vagy peptid és a
karboxilcsoportján aktivált és aminocsoportján védett aminosav-származék között alakítjuk
ki. Az aktivált aminosav-származék el�állítható a kapcsolási reakció el�tt, de in situ,
kapcsolóreagensek segítségével is. A kapcsoláshoz leggyakrabban használt reagensek a
karbodiimidek, pl. az N,N’-diciklohexil-karbodiimid (DCC) vagy az N,N’-diizopropil-
karbodiimid (DIC). A DCC rekciómechanizmusa a 8. ábrán látható. Ezek O-acil-izourea
származékot képeznek az aminosav-származék karboxilcsoportjával és ez acilezi a szabad
aminocsoportot, N,N’-diciklohexil-urea vagy N,N’-diizopropil-urea képz�dése mellett. In situ
aktív észter el�állításakor az N-acil-izourea képz�dés el�térbe kerülhet, ezért egyfajta
alternatív megoldásként 1-hidroxi-benzotriazollal (HOBt) állítjuk el� az aktív észtert. Ezáltal
gyors kapcsolás érhet� el és az aszparagin és glutamin esetében a dehidratáció megel�zhet�
(König 1970), (Mojsov et al., 1980).
H11C 6
N C N
C 6H 11
OH
O
Q
O
N
NH
O
Q
H 11C6
C 6H 11
NH2Q'
NH
O
Q
Q'
+O
NH
NH C 6H 11
C 6H 11
DCC
DCU amid(peptid)(karbam id)
8. ábra. A DCC reakciómechanizmusa
Ezen kívül foszfónium és urónium típusú kapcsolóreagensekkel is aktiválhatjuk az
aminosavakat. Ilyen kapcsolóreagensek például a benzotriazol-1-il-oxi-trisz-dimetil-amino-
foszfónium hexafluoro-foszfát (BOP) (Castro et al., 1975), a benzotriazol-1-il-oxi-trisz-

pirrolidino-foszfónium hexafluoro-foszfát (PyBOP) (Coste et al., 1990), a benzotriazol-1-il-
1,1,3,3,-tetrametil-utónium hexafluoro-foszfát (HBTU), a 7-aza-benzotriazol-1-il-1,1,3,3,-
tetrametil-urónium hexafluoro-foszfát (HATU) és a 7-aza-benzotriazol-1-il-1,1,3,3,-
tetrametil-urónium tetrafluoro-borát (TATU). Az aminosav benzotriazol-észtere, ill. aza-
benzotriazol-észtere az acilez� reagens a kapcsolási reakció során.
II.2.6. Kapcsolási reakciók követése
A kapcsolási/Nα- véd�csoport hasítási reakciók követése sokféleképpen történhet. A
legelterjedtebb módszerek a ninhidrin-(Kaiser et al., 1970), az izatin- (Kaiser et al., 1980) és a
brómfenolkék-próba (Krchnak et al., 1988), melyek szabad amino- ill. iminocsoportok
jelenlétét színreakcióval jelzik. Az izatin- és a ninhidrin-tesztnél lejátszódó reakció a 9. ábrán
látható.
NH
O
O
+NH
Opeptid-gyanta
R-COOH
N NO
peptid-gyantaO
+ H2O
+ -2H2O
O
O
N
COOH
Q
O
O
OH
OHNH2 COOH
Q
O
O
N
CH
Q
H2O+
O
O
NH2 + CHO
Q
ninhidrinO
O
N
O
O
O
OH
N
O
O
(ibolya szín�)
O
O
N
CH2 Q+ CO2
triketo-hidrindén-hidrát
ninhidrin
9. ábra. A ninhidrin- és az izatin-teszt reakciómechanizmusa
II.2.7. A peptid lehasítása a gyantáról és az oldallánc véd�csoportok eltávolítása
Az Fmoc/tBu módszernél a peptid gyantáról történ� hasítását és az oldallánc
véd�csoportok eltávolítását általában TFA-val végezzük. A hasítás során keletkez� reaktív

karbokationok miatt az 5% vizet tartalmazó TFA elegyet használjuk. Ha a peptid nukleofil
oldallánccal rendelkez� aminosavakat (Trp, Met, Tyr, Cys) tartalmaz, akkor a hasításhoz
célszer� 82,5% TFA, 5% fenol, 5% víz, 5% tioanizol, 2,5% etánditiol (V/m/V/V/V)
összetétel� elegyét alkalmaznunk.
A Boc/Bzl módszernél a peptid gyantáról való lehasítását és a véd�csoportok
eltávolítását hidrogén-fluoriddal (HF) végezzük teflon készülékben, -5°C-on. A hasítás a
peptidszekvenciától és oldallánc véd�csoport kombinációtól függ�en általában 1-1,5 órát vesz
igénybe. Ennél a módszernél is szükséges gyökfogókat (p-krezolt és tiokrezolt 1:1 (m/m)
arányban) alkalmazni.
II.3. Hordozó molekulák és hatóanyag-konjugátumok alkalmazása
A tbc kezelésében alkalmazott antituberkulotikumok bejutása a fert�zött makrofágba
els�sorban diffúzió révén történhet, meglehet�sen korlátozott mértékben. A hosszú kezelés
során (1. fázis: 2-6 hónap, 2. fázis: 6-12 hónap) számolni kell mellékhatásaikkal, pl. a
szervezet egészét érint� nem specifikus toxicitásukkal, valamint gyors kiválasztódásukkal.
A peptidtípusú hordozók alkalmazása csökkentheti a terápiás szerek mellékhatásait,
befolyásolják a biodisztribúciót, csökkentik a toxicitást (a hatóanyag fokozatos felszabadulása
által), segítenek a retard hatás elérésében és megakadályozzák a szervezetb�l való gyors
kiürülést. Ezen kívül talán a legfontosabb el�nyük, hogy lehet�séget kínálnak a hatóanyag
célsejtbe juttatására (esetünkben a fert�zött makrofágokba) receptor mediált endocitózissal,
pl. a makrofágok sejtmembránján megtalálható tuftsin receptoron keresztül (Gottlieb et al.,
1983; Bump et al., 1986; Bump et al., 1990; Fridkin et al., 2005; Gupta and Haq 2005).
Az antituberkulotikum hatásának kifejtése szempontjából fontos tényez� a hatóanyag
és hordozó közt lév� kötés kémiai jellege, ugyanis a hatóanyag tulajdonságai el�nytelenül
változhatnak, ha nem megfelel� kémiai szerkezettel szabadul fel. A konjugációnál
leggyakrabban az amid, észter, éter, hidrazon, és hidrazid jelleg� kötések alkalmazottak.
A terápiásan alkalmazott INH antituberkulotikum kémiai konjugálásához hordozóként
tuftsin származékokat (GTKPKG (T6), [TKPKG]4 (T20)) és az M. tuberculosis
immundomináns 16 kDa fehérjének egy T-sejt epitóppeptidjét (91SEFAYGSFVRTVSLPV106
(91-106)) választottuk.

II.3.1. Hordozóként alkalmazható tuftsin származékok
A természetben el�forduló, immunstimuláló hatású tuftsin (Thr-Lys-Pro-Arg/Lys:
humán/kutya eredet�) (Fridkin et al., 1977; Fridkin and Gottlieb 1981; Najjar 1983; Fridkin
and Najjar 1989) szekvencián alapuló oligo-tuftsin hordozók jól definiált szerkezet�ek, a fenti
szekvencia a különböz� hosszúságú oligomerekben 4-, 6-, vagy 8-szor ismétl�dik (Mez� et
al., 2000). A natív humán tuftsin szekvencia, a TKPR az immunglobulinokban megtalálható
tetrapeptid, amely enzimatikus hasítás után felszabadulva jelent�s szerepet játszik az
immunfolyamatokban. A kutyában található tuftsin a C-terminálison lizint tartalmaz arginin
helyett. Így a monomerben két lizin van jelen, ami lehet�vé teszi a kés�bbiekben több
hatóanyag szelektív konjugálását. Egy glicin C-terminálisra való beépítésével pedig lehet�vé
válik oldat fázisban a nagyobb hordozó peptidek racemizációmentes felépítése,
fragmenskondenzációval. Ráadásul a TKPKG pentapeptid fagocitózist stimuláló hatása
nagyobb, mint a tetrapeptidé.
II.3.2. A M. tuberculosis immundomináns 16 kDa fehérje és a 91-106
(91SEFAYGSFVRTVSLPV106) T-sejt epitóppeptid
Az ELTE-MTA Peptidkémiai Kutatócsoportban 1995 óta foglalkoznak M.
tuberculosis immundomináns fehérjék epitópszerkezetének vizsgálatával és ezek közül is
külön figyelmet érdemel a 16 kDa (Hsp 16.3, acr) h�sokk fehérjék családjába tartozó
molekula (Verbon et al., 1992; Cunningham and Spreadbury 1998). A lecsökkent oxigén
tenzió arra készteti a baktériumot, hogy nyugalmi fázisba lépjen, melyet dormanciának
neveznek (Manabe and Bishae 2000). Az alacsony oxigén szinthez a baktérium többek között
vastagabb sejtfal kifejlesztésével alkalmazkodik. A 16 kDa fehérje fontos szerepet tölt be a
sejtfal vastagítás folyamatában (Cunningham and Spreadbury 1998). Ezenkívül molekuláris
chaperonként m�ködik, stabilizálja a sejt-struktúrákat a hosszú távú nyugalmi fázis során,
segítve ezzel a baktériumot a túlélésben.
Átlapoló szintetikus peptidek segítségével a kutatócsoport együttm�ködve Dr. Juraj
Ivanyival (Hammersmith Hospital, London, Anglia) és Dr. Francesco Dielivel (Department of
Biopathology, University of Palermo, Palermo, Olaszország) a fehérjében CD4+ T-sejt
epitópokat lokalizált (Wilkinson et al., 1999; Hudecz 2001; B�sze et al., 2004). Ezen T-sejt
epitóppeptidek képesek makrofágok felszínén kifejez�d� MHC (HLA) fehérjékhez köt�dni.

II.4. Antituberkulotikumok és konjugátumaik minimális gátló koncentrációjának
meghatározása folyékony táptalajon, telepszámának meghatározása szilárd táptalajon
Kísérleteinket az Országos Korányi TBC és Pulmonológiai Intézet területén található
Bakteriológiai Laboratóriumban (Corden International Magyarország Kft.), Dr. Szabó Nóra
irányitásával és Dávid Sándor közrem�ködésével végeztük.
II.4.1. Baktériumok oltása és tenyésztése, a munka során használt tápközegek
Az oltás a mikroorganizmusok tenyésztését megel�z� mikrobiológiai m�velet.
Lényege: steril oltóeszköz segítségével steril tápközegbe juttatjuk azokat a mikróbákat,
esetünkben baktériumokat, amelyeket szaporítani kívánunk. Az oltáshoz felhasznált mikróba
– sejttömeget inokulumnak nevezzük. A tenyésztés a baktériumok meghatározott
tápközegekben történ� mesterséges szaporítása. Laboratóriumi körülmények között a
tenyésztést megfelel� összetétel� tápközegben végezzük. Ezek a tápközegek a vizsgált
baktérium tápanyag-, energia-, pH-, vízaktivitás- és redoxpotenciál-igényeinek megfelel�ek.
A tápközegek tartalmazzák mindazokat az anyagokat, amelyekre a mikroorganizmusoknak
szüksége van (víz, redukált széntartalmú vegyületek, szerves és szervetlen
nitrogénvegyületek, ásványi anyagok, vitaminok) és amelyeket maguk szintetizálni nem
képesek. A szükséges h�mérséklet termosztátban állítható be.
A folyékony táptalajokra jellemz�, hogy a különböz� tápanyagokat szuszpenzióban,
valódi vagy kolloid oldat formájában tartalmazzák. A folyékony halmazállapotú tápközegek
közül a félszintetikus táptalajokat is “leveseknek” nevezzük. Félszintetikus tápközegek
összetev�inek egy része olyan természetes anyag, amelynek összetétele nem teljesen ismert,
vagy alkotóinak aránya változó. A szilárd táptalajok a folyékony táptalajokból
származtathatók szilárdító anyagok (agar-agar, zselatin, szilikagél) hozzáadásával. Régebben
a zselatin volt a legelterjedtebb szilárdító anyag, ma egyre inkább háttérbe szorul, ami
egyrészt igen alacsony olvadáspontjával (35°C) magyarázható, másrészt azzal a ténnyel, hogy
a zselatint számos baktériumfaj elfolyósítja zselatináz enzime révén. Éppen ezért ma már
inkább csak diagnosztikai céllal használják. Az agar kétkomponens� gélképz� poliszacharid-
keverék, amelyet egyes tengeri vörösalgákból vontak ki. Az agar nagy el�nye, hogy 38-40°C-
on megdermed, olvadáspontja azonban 98°C. Mivel az agar természetes anyag, összetétele
nem definiálható pontosan, csak természetes és félszintetikus táptalajokhoz használható. A

szilárd táptalaj megjelenési formája szerint lehet kémcs�ben elkészítve magas- vagy
ferdeagar, Petri-csészébe leöntve pedig agarlemez (10. ábra).
10. ábra. Baktérium tenyésztésére alkalmazott kémcs� (folyékony táptalaj) illetve Petri csésze oldal- és felülnézetben
A M. tuberculosis tenyésztésére a dolgozatban szerepl� kísérletek esetén a Sula-féle
félszintetikus, folyékony (Sula 1963; Sula 1963; Lányi 1980), valamint a Löwenstein-Jensen
féle szilárd táptalajt használtuk (Löwenstein 1931; Jensen 1932). Ezt a tojás
homogenizátumot tartalmazó szilárd tápközeget, amely a rezisztens baktériumok kimutatására
is használható Löwenstein javasolta (1931) és Jensen módosította (1932).
A Sula és a Löwenstein-Jensen táptalaj is tartalmaz malachitzöldet, amely gátolja a
vizsgálati anyagokban jelenlév� nem saválló baktériumok szaporodását. Ez fontos a M.
tuberculosis esetében a többhetes (4-6-8 hét) tenyésztési id� miatt. A legtöbb Mycobacterium
lassan szaporodik, 12-24 óra az osztódási id�. A lassan növ� fajok (pl. M. tuberculosis, M.
avium) 3-8 hét alatt képeznek telepeket. A táptalajon az M. tuberculosis és a közeli
rokonságban lév� fajok elkülöníthet�ek.

II.4.2. Baktériumok sejtszámának meghatározása
A legtöbb baktérium esetében a sejtek szaporodása során az utódsejtek elválnak
egymástól. A sejtszámmeghatározás módszerei közül a mikroszkópos számolást és a
turbidimetriát alkalmazzák legtöbbször a gyakorlatban: (a) Bürker-kamrás sejtszámolás:
nagyobb méret� sejteknél közvetlen sejtszámolás lehetséges Bürker-kamrával Ez a módszer
az összsejtszám megállapítására ad lehet�séget, gyors és egyszer� sejtszámlálási módszer. (b)
Turbidimetria: a meghatározás folyadéktenyészetek "zavarosságának" mérésén alapszik. A
turbidimetriás mérésnél a m�szeren mért "s�r�ség"-értékeket kalibrálni kell (közvetlen
sejtszámolással vagy közvetve, él�csíraszám megállapítással). A kalibrációs görbén egy-egy
turbiditás értéknek meghatározott sejtszám felel meg.
Turbidimetriában gyakran alkalmazott referenciák az úgynevezett “McFarland
standard” oldatok, amelyek segítségével (egy adott tartományon belül) egy baktérium
szuszpenzió “zavarossága”, vagyis sejtszáma állítható be (McFarland 1907; Bollela et al.,
1999). Az eredeti McFarland oldatok meghatározott koncentrációjú bárium-klorid és kénsav
elegyítésével készültek, ahol a keletkez� bárium-szulfát okozza a kívánt mérték�
“zavarosságot”, abszorbanciát. (Pl. a 0,5 McFarland (amely közelít�leg 1,5.108 sejtszámnak
felel meg) eredetileg 0,05 ml 1,175 %-os bárium-klorid dihidrát (BaCl2.2H2O) és 9,95 ml 1%-
os kénsav (H2SO4) keveréke, amely szuszpenzió abszorbanciáját 600 nm határozzák meg.
II.4.3. Minimális gátló koncentráció (MIC) meghatározása
Általános definíció: Minimális gátló koncentrációnak (minimum inhibitory
concentration = MIC) nevezzük azt a legkisebb antibiotikum mennyiséget, amely 1 ml
térfogatú oldatban (táptalajban) megakadályozza valamely baktérium szaporodását.
Hasonlóan informatív lehet az antibiotikumok minimális baktériumöl� koncentrációja
(minimum bactericid concentration = MBC), az a mennyiség, amely az inokulumként bevitt
baktériumok 99,9%-át elpusztítja. Esetünkben: a minimális gátló koncentráció (MIC) a
vizsgálandó vegyületnek az a legkisebb koncentrációja (mikrogramm/ml egységben
kifejezve), amely az adott mycobacterium törzs (a H37Rv) növekedését gátolja.
A minimális gátló koncentráció értéke függ az alkalmazott táptalajtól, a táptalaj pH
értékét�l, az inokulum kolónia számától, az inkubáció h�mérsékletét�l, atmoszférájától és
id�tartamától. A baktériumok esetében ez a jelenség a mikroszervezetek gyors adaptív
tulajdonságával magyarázható, nem örökletes, modifikatív sajátság.

A MIC értéket az általunk vizsgált antituberkulotikumok és a konjugátumaik esetében
„kvantitatív leves higításos módszer” alkalmazásával határoztuk meg. A módszer els� lépése,
hogy a vizsgálandó vegyületb�l - alkalmas oldószerrel – törzsoldatot, abból pedig megfelel�
léptékben, azonos térfogatú hígításokat készítünk. A kapott higítási sor oldataiból - a vegyület
végkoncentrációjának megfelel�en - egyforma mennyiségeket adunk a táptalajt tartalmazó
csövekhez. Ezután a csöveket a vizsgálandó mycobacterium törzs megfelel� higítású
szuszpenziójával beoltjuk. A minimális gátló szintet annak a cs�nek a vegyület koncentrációja
jelenti, amelyben a kell� id�tartamú inkubálás után szabad szemmel még nem észleltünk
növekedést.
II.4.4. Telepszám (kolónia szám, colony forming unit, CFU) meghatározása
A telepszámlálásos módszerek esetében a tenyésztést szilárd táptalajon végezzük, így
a kifejl�dött telepek közvetlenül megszámolhatók. A telepszámlálásos módszerek
alkalmazhatóságának alapfeltétele, hogy a táptalajokon kin�tt telepek egyetlen sejt
szaporodásából származzanak, és a telepek számolhatók legyenek, vagyis a cél olyan
lemeztenyészetek el�állítása, amelyen az egyedülálló (soliter) telepek száma 10-300 közötti.
A táptalajtól és a tenyésztési technikától függ�en - alkalmas mind az összcsíraszám; mind
pedig valamely tetsz�leges mikrobacsoport számának meghatározására. A CFU/ml érték
megadja, az adott minta esetében, az egységnyi térfogatban jelenlév� életképes sejtek közelít�
számát, amelyekb�l a számolható telepek kifejl�dtek.
Kísérleteinkben a MIC érték leolvasását követ�en (az inkubációs id� letelte után,
általában 28 nap) a növekedést szabad szemmel vizsgálva nem mutató csövekb�l, szilárd
Löwenstein-Jensen (LJ) táptalajokra oltottunk ki. Ezen a táptalajon 4 – 6 hét múlva a
kinövekedett kolóniák számát (CFU, colony forming units) határoztuk meg. A módszer
lényegét az 11. és a 12. ábra foglalja össze.

��!������" ��"����������� ����������#$% �������!& �'��� �������
������"���(�������������
)!*�'��
���������"��� +����������� � ��
��� ���'���,
��� ��'��� ���'����������� ��������
+� ����������������������"�������� ����,
� �
��!������" ��"����������� ����������#$% �������!& �'��� �������
������"���(�������������
)!*�'��
���������"��� +����������� � ��
��� ���'���,
��� ��'��� ���'����������� ��������
+� ����������������������"�������� ����,
��!������" ��"����������� ��!������"
��"����������� ����������#$% �������!& �'��� �������������"���(�������������
����������#$% �������!& �'��� �������������"���(�������������
)!*�'��
���������"��� +����������� � ��
��� ���'���,
���������"��� +����������� � ��
��� ���'���,
��� ��'��� ���'����������� ��������
+� ����������������������"�������� ����,
��� ��'��� ���'����������� ��������
+� ����������������������"�������� ����,
� �
11. ábra. (A) Kolónia szám (telepszám) meghatározásásának sematikus vázlata (B) M. tuberculosis kolóniák Löwenstein-Jensen szilárd táptalajon, nagyítva
�$������'����������������������������� ������$��� ����������'������ �������� � ���-�������
��������������������"���!.�/01+������ -��� ��� ���,
���������)�'�����!.�2 3/�4�5�γ
2 6����������� �789�+: ; //�<8<=),
�������������������������+)�'��������������,
2 3/�4������������������>���������������������������������"� ��� �������������� ��$��������������"�
�$������'����������������������������� ������$��� ����������'������ �������� � ���-�������
��������������������"���!.�/01+������ -��� ��� ���,
���������)�'�����!.�2 3/�4�5�γ
2 6����������� �789�+: ; //�<8<=),
�������������������������+)�'��������������,
2 3/�4������������������>���������������������������������"� ��� �������������� ��$��������������"�
12. ábra. Minimális gátló koncentráció (MIC) és telepszám (CFU) meghatározása M. tuberculosis H37Rv törzsön
II.5. A sejtek életképességének vizsgálata antituberkulotikum (INH), hordozómolekulák
és antituberkulotikum-konjugátumok hatására, az in vitro citosztatikus hatás
meghatározása kolorimetriás tetrazólium (MTT) teszt alkalmazásával
Citosztatikusnak nevezzük azokat az anyagokat (pl. kismolekulájú vegyületek,
hordozómolekulák), amelyekkel történ� kezelés hatására a sejtek elpusztulnak, illetve nem

szaporodnak, proliferálnak, életképességük megváltozik (antiproliferatív hatás, életképesség
megváltoztatása, ~ citosztatikus hatás). A citosztatikus hatás a vizsgált vegyületre jellemz�
adat, az adott sejttípuson. A citosztatikusság ténye önmagában nem jellemzi a vegyület által
kiváltott sejthalál mechanizmusát.
A citosztatikus hatás mértékének meghatározása, különböz� sejttípusokon egy gyors
és viszonylag költségkímél� módszer, azoknak a vegyületeknek a kiküszöbölésére – a
fejlesztések korai szakaszában – amelyek a sejtek halálát okozzák már kis koncentrációban.
Ezek a tesztek nagymértékben segítik a vegyületek optimalizálását az in vitro kísérletek során.
Leggyakrabban a nem radiaktív, kolorimetriás, funkcionális teszteket alkalmazzák
kémiai vegyületek toxicitásának és citosztatikus hatásának mérésére, mert használhatók 3-24-
96 órás kezelési protokolokban, in vitro sejtkultúráknál. Érzékenyek, jól reprodukálhatók,
egyszer�en, gyorsan kivitelezhet�ek, viszonylag kis mennyiség� sejtet igényelnek
(sejttípustól függ�en: 5.103 –1.105 sejt/mélyedés/szövettenyészt� lemez).
A dolgozatban összefoglalt munkában a vegyületek (szabad antituberkulotikumok,
hordozó molekulák, antituberkulotikum-konjugátumok) in vitro citosztatikus hatását
tetrazólium (MTT) teszt segítségével határoztuk meg (Slater et al., 1963; Mosmann 1983;
Gerlier and Thomasset 1986). Az MTT egy tetrazólium só (3-(4,5-dimetiltiazol-2-il)-2,5-
difeniltetrazolium bromid), amelyet az él�, metabolikusan aktív sejtek mitokondriális
dehidrogenáz enzimje képes formazán származékká alakítani. Ez a származék mutatható ki
kolorimetriásan, az él� sejtek számával korreláltatható mértékben.
A citosztázis mértékét (%) λ=540 nm-en mért abszorbancia értékek alapján a
következ� képlettel határoztuk meg:
��
�
�
��
�
�−⋅=
sejtekkontroll
sejtekkezelt
A
Ascitosztázitáscitotoxici 1100[%]/
ahol Akezelt a kezelt sejtek, míg Akontroll a negatív kontroll sejtek esetében mért abszorbanciát
(λ=540 nm) jelenti.
A citosztázis mértékét (%) a koncentráció függvényében ábrázolva a kapott görbék
segítségével meghatározhatóak az IC50 értékek. Az IC50 érték azt a koncentrációértéket jelenti,
amely a vizsgált sejtkultúrában a sejtek 50%-ának pusztulását okozza.
A vegyületek citosztatikus hatását a meghatározott MIC értékek alatti, valamint annál
magasabb koncentrációtartományban vizsgáltuk humán sejtvonalon (HepG2 – májból izolált,
hepatocelluláris karcinoma sejtvonal (Knowles et al., 1980; Knowles and Aden 1983)).

III. Célkit�zések
A dolgozatban összefoglalt munka célja a terápiásan alkalmazott INH
antituberkulotikum fert�zött makrofágokba való szelektív célbajuttatására alkalmas
hatóanyag-hordozó konjugátumok el�állítása volt.
E célunk megvalósítása érdekében a kísérleti munka során hordozóként
tuftsinszármazékokat és az M. tuberculosis immundomináns 16 kDa fehérjének egy T-sejt
epitóppeptidjét választottunk.
A hordozó peptideket szilárdfázisú szintézissel állítottuk el�. El�állítottuk az
izoniazid kapcsolásához szükséges távolságtartó „linker” molekulát és alkalmazásával INH-
hordozó konjugátumokat állítottunk el�. Az új vegyületek antituberkulotikus hatását,
minimális inhibiciós koncentrációjának (MIC) meghatározását M. tuberculosis in vitro
tenyészeten hasonlítottuk össze a szabad INH molekula hatásával.
3? �3? �
3? � �����"�����������
������'��������
3? � �����"�����������
������'��������
�����"�
9��������� ����� ����������� ��������������"�����
0�����������������--�������"������������� �����@�-�������
3? �3? �
3? � �����"�����������
������'��������
3? � �����"�����������
������'��������
�����"�
9��������� ����� ����������� ��������������"�����
0�����������������--�������"������������� �����@�-�������
13. ábra. Az INH fert�zött makrofágokba történ� szelektív célbajuttatásához választott stratégia sematikus
összefoglalása

IV. Kísérleti rész
IV.1. A szintetikus munka során használt vegyszerek, m�szerek Aminosavszármazékok, gyanták Elnevezés, kapacitás Rövidítés, jelzés Gyártó cég terc-butiloxi-karbonil- (Boc), Reanal és 9-fluorenil-metiloxi-karbonil- (Budapest) (Fmoc) aminosavszármazékok Fluka (Buchs, Svájc)
Nova Biochem (Läufelfingen, Svájc) 4-metil-benzhidrilamin-gyanta MBHA Nova Biochem (1,1 mmol/g) (Läufelfingen, Svájc) 4-(2’, 4’-dimetoxifenil-Fmoc- Rink Amide MBHA Nova Biochem -aminometil)-fenoxiacetamido- (Läufelfingen, Svájc) -norleucil-MBHA-gyanta (0,69 mmol/g)
Oldószerek, reagensek Elnevezés, kapacitás Rövidítés, jelzés Gyártó cég N, N-dimetil-formamid DMF Molar (Budapest) N, N’-diizopropil-karbodiimid DIC Fluka
(Buchs, Svájc) 1,2-etánditiol, piperidin, ninhidrin diklór-metán DCM Molar (Budapest) 1-hidroxi-benztriazol HOBt Fluka
(Buchs, Svájc) trifluor-ecetsav TFA Reanal (Budapest) etilén-diizopropil-amin DIEA izoniazid INH Fluka
(Buchs, Svájc) glioxilsav-monohidrát Gli Merck
(Darmstadt, Németország) nátrium-ciano-borohidrid NaCNBH3 Merck
(Darmstadt, Németország) benzotriazol-1-il-oxi-trisz-pirrolidino- PyBOB Fluka -foszfónium hexafluorofoszfát (Buchs, Svájc) ecetsav, jégecet, metanol Reanal acetonitril (Budapest)

szövettenyészt� médium RPMI 1640 Gibco magzati (foetális) borjú savó FCS (Paisly, UK) (foetal calf serum) gentamicin 3-(4,5-dimetiltiazol-2-il) MTT Sigma -2,5-difeniltetrazolium bromid (St.Louis, MO, USA) 96 lyukú szövettenyészt� lemez Greiner (Frickeneusen, Németország) M�szerek Elnevezés Gyártó cég Knauer HPLC rendszer Knauer (pumpa, UV detektor, kézi injektor) (Bad Homburg, Németország) VARIO EL III elemanalizátor Elementar Analysensysteme GmbH (Németország) SYKAM 4300 SYKAM automata aminosav analizátor (Eresing, Németország) Bruker Daltonics Esquire 3000+ Bruker tömegspektrométer (Bremen, Németország) HF teflon készülék Peninsula Laboratories, INC. (Belmont, CA, USA) SYRO MultiSyntech automata peptidszintetizátor (Witten, Németország) BlockTerm 656 termosztát (Magyarország) Büchi 530 olvadáspontmér� Büchi Labortechnik AG (Flawil, Svájc) Labsystem iEMS, MF Labsystem mikrotitrációs lemez leolvasó (Helsinki, Finnország) spektrofotométer
HPLC oszlopok, töltetek
Elnevezés Töltet, méret, jellemz�k Gyártó cég Phenomenex fordított fázisú Phenomenex módosított szilika töltet (Torrance, CA, USA)
-félpreparatív Jupiter (C18 10 x 250 mm) 10µm, 300 Å Eurospher
-analitikai Eurospher (C18 4 x 250mm) 5µm, 100 Å

IV.2. A peptidek szintézise
A szintetikus munka során a következ� szekvenciájú peptidamidokat állítottuk el�:
GTKPKG (T6), [TKPKG]4 (T20), 91SEFAYGSFVRTVSLPV106 (91-106).
IV.2.1. A peptidek szintézise manuális Boc/Bzl stratégiával
A T6 és T20 peptideket manuális szilárdfázisú szintézissel állítottuk el� Boc/Bzl
stratégia alkalmazásával.
A peptidek szintéziséhez 4-metil-benzhidrilamin (MBHA) (Matsueda and Stewart
1981) gyantát használtunk. Valamennyi aminosavat Nα-Boc-származék (Merrifield 1963;
Merrifield 1964) formájában kapcsoltuk. A Boc-aminosavak oldalláncai a 1. táblázatban
felsorolt véd�csoportokat tartalmazták.
1. táblázat. A Boc/Bzl stratégia szerint az MBHA gyantákon felhasznált Boc-aminosavszármazékok oldallánc véd�csoportjai.
Aminosav Funkciós csoport Véd�csoport Rövidítés
Lys amino 9-fluorenil-metiloxi-karbonil Fmoc
Thr hidroxil benzil Bzl
Arg guanidino 4-toluol-szulfonil Tos
ββββ-Ala, Gly, Pro oldallánc véd�csoport nélkül
Az MBHA gyanta (kapacitás:1,1 mmol/g) hidroklorid formában kerül
kereskedelmi forgalomba, ezért a gyantát a duzzasztást követ�en 33% TFA / 66% DCM V/V
hasítóelegyével kezeltük, 2+20 percig, majd a DCM-mel való ötszöri mosást követ�en,
semlegesítettük 10% DIEA / 90% DCM V/V elegyével 4 x 1 percig, majd újból mostuk
háromszor DCM-mel. Az MBHA gyantához az el�kezelés után kapcsoltuk az els� Boc-
aminosavszármazékot. A gyantakapacitásra számolva háromszoros moláris feleslegben adtuk
az aminosavszármazékot és az in situ aktív észter kialakításához szükséges kapcsolószereket
(DIC, HOBt) (König 1970; Mojsov et al., 1980) DCM-ben oldva. A kapcsolási id� egy óra
volt. A mosási lépést (DCM) követ�en ninhidrin-, illetve izatin- (Kaiser)-próbával (Kaiser et
al., 1970; Kaiser et al., 1980) ellen�riztük a kapcsolás végbemenetelét. Pozitív Kaiser-próba
esetén a kapcsolást megismételtük.

A Boc-véd�csoportot minden esetben a gyanta el�kezelésénél leírt hasítóeleggyel
(33% TFA / 66% DCM V/V) távolítottuk el, majd a DCM-mel való mosást követ�en,
semlegesítettük (10% DIEA / 90% DCM V/V).
A Boc/Bzl stratégiával végzett szintézis egy ciklusának összefoglalása a 2.
táblázatban látható.
2. táblázat. A Boc/Bzl stratégia szerint végzett peptidszintézis egy ciklusának menete M�velet Reagens, oldószer Id�tartam
(perc)gyanta mosása 3 x 2-5 ml DCM 3 x 1
Boc-véd�csoport eltávolítása 2 x 2-4 ml hasítóelegy 2+2033% TFA/ DCM (V/V)
gyanta mosása 5 x 2-5 ml DCM 5 x 1semlegesítés 4 x 2-4 ml elegy 4 x 1
10% DIEA/DCM (V/V)gyanta mosása 3 x 2-5 ml DCM 3 x 1
3 ekvivalens* Boc-aminosav/DCMkapcsolás ekvimoláris HOBt/DCM 60
ekvimoláris DICelegy végtérfogat 4-5 ml
gyanta mosása 2 x 2-5 ml DMF 2 x 12 x 2-5 ml DCM 2 x 1
kapcsolási reakció követése izatin*-, ninhidrin (Kaiser)-próba 5*gyantakapacitásra számított mennyiség
*Pro-aminosavszármazékhoz való kapcsolást követ�en
A kapcsolás/ véd�csoport eltávolítás végbemenetelét ninhidrin-próba segítségével
ellen�riztük. A próbához szükséges oldatok összetétele a következ� volt: 1. ninhidrin/etanol
10:1 (m/V), 2. fenol/etanol 4:1 (m/V), 3. piridin/víz 98:2 (V/V) elegy, amely 4 µmol/dm3
KCN-ot tartalmazott. Egy üveg reakciócs�be kevés gyantaszemet raktunk és erre a három
reagensb�l 1.-3. sorrendben 2-2 cseppet adtunk, majd a csövet BlockTerm-ben 105°C-on
tartottuk 5 percig. Prolinhoz való kapcsolás ellen�rzéséhez használt elegy összetétele 3%
izatin, 5% Boc-Phe-OH / benzilalkohol volt, melyet a fent leírt oldatok el�tt csepegtettünk a
gyantaszemekhez.
Sikertelen kapcsolás esetén újabb adag Boc-aminosavszármazékkal és in situ aktiv
észter (DIC, HOBt) képzés segítségével megismételtük a kapcsolást.
A szilárd hordozóról a peptideket/peptidkonjugátumokat és az oldallánc
véd�csoportokat hidrogén-fluorid (HF) segítségével távolítottuk el (Grant 1992b). A hasitást
teflon edényben kevertetve végeztük, -5°C-on 1,5 óráig, gyökfogók (p-krezol, tiokrezol)
jelenlétében. 0,5-1,00 g peptidre/peptidkonjugátumra átlagosan 10 ml HF-ot számolva 0,5 g
p-krezolra és 0,5 g tiokrezolra volt szükség. Hasítás után a HF-ot a gyanta

peptid/peptidkonjugátum elegyr�l ledesztilláltuk, majd a reakcióelegyet felöntöttük hideg
éterrel, ezután sz�rtük. A sz�rés után kevés A eluens (0,1% TFA/víz (V/V)) segítségével
kioldottuk a peptidet/peptidkonjugátumot a sz�r�n a gyantaszemek közül. Az oldatot
lefagyasztottuk, majd liofilizáltuk.
IV.2.2. Peptidszintézis Fmoc/tBu-stratégiával automata peptidszintetizátoron
A 91-106 peptidet SYRO (MultiSyntech) automata peptidszintetizátor segítségével
állítottuk el�, Fmoc/tBu-stratégia alakalmazásával (Carpino and Han 1972). A szintézishez
Rink Amide MBHA gyantát (Rink 1978) (kapacitás: 0,69 mmol/g) és Nα-Fmoc-
aminosavszármazékokat használtunk. Az Fmoc-aminosavak oldalláncai a 3. táblázatban
felsorolt véd�csoportokat tartalmazták.
Az Fmoc-véd�csoport hasításához piperidin/DMF 2:8 (V/V) és 4:6 (V/V) elegyét
használtuk. Az amidkötések kialakítása DIC/HOBt in situ aktív észterek kialakításával történt.
Az aminosavszármazékokat és reagenseket négyszeres moláris feleslegben alkalmaztuk a
szintézis során. Az aminosavszármazékok oldása a HOBt megfelel� koncentrációjú, NMP-nal
készült oldatában történt, amelyben a HOBt koncentrációja szintézist�l függ�en 0,50-0,69
mol/l tartományban volt. A peptidszintetizátor egy ciklusának protokollja a 4. táblázatban
olvasható.
3. táblázat. A Fmoc/tBu stratégia szerint a Rink Amide MBHA gyantákon felhasznált Fmoc-aminosavszármazékok oldallánc véd�csoportjai
Aminosav Funkciós csoport Véd�csoport RövidítésGlu karboxil terc -butil t BuThr hidroxil tritil Trt
Arg guanidino2,2,4,6,7-pentametil-dihidro-benzofurán-5-
szulfonilPbf
Ser, Tyr hidroxil terc -butil t BuAla, Gly, Leu, Pro,
Phe, Valoldallánc
véd�csoport nélkül
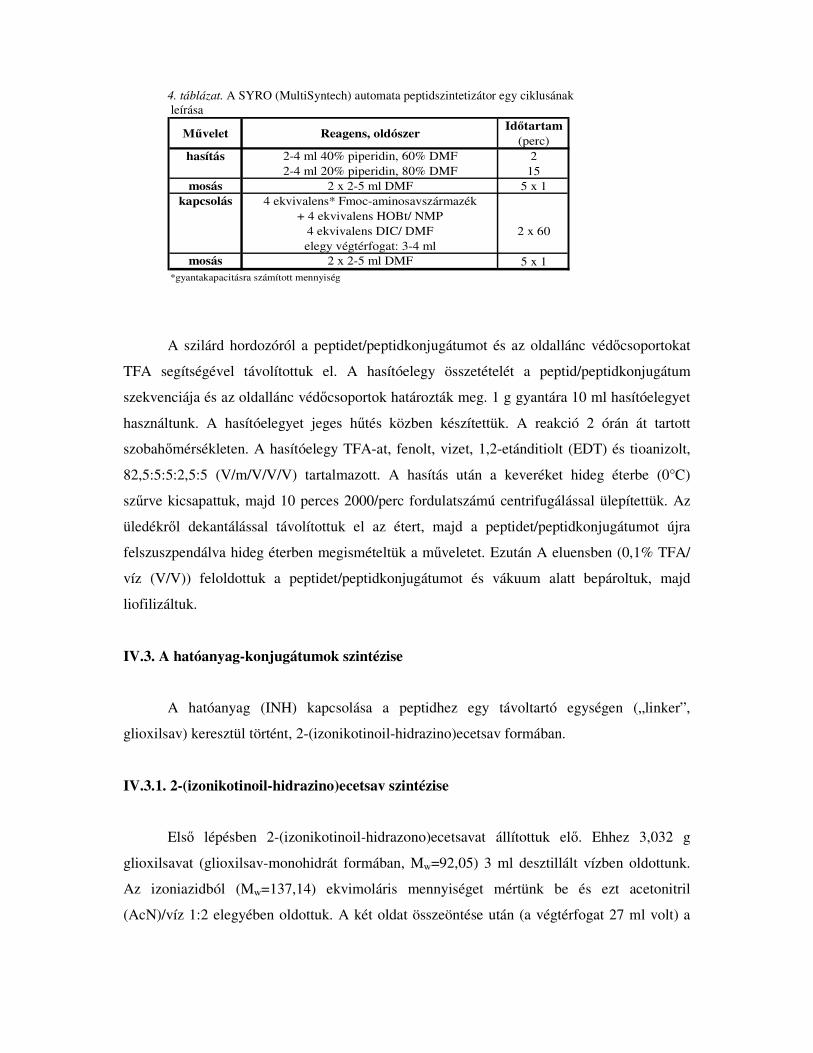
4. táblázat. A SYRO (MultiSyntech) automata peptidszintetizátor egy ciklusának leírása
M�velet Reagens, oldószer Id�tartam (perc)
hasítás 2-4 ml 40% piperidin, 60% DMF 22-4 ml 20% piperidin, 80% DMF 15
mosás 2 x 2-5 ml DMF 5 x 1kapcsolás 4 ekvivalens* Fmoc-aminosavszármazék
+ 4 ekvivalens HOBt/ NMP4 ekvivalens DIC/ DMF 2 x 60elegy végtérfogat: 3-4 ml
mosás 2 x 2-5 ml DMF 5 x 1*gyantakapacitásra számított mennyiség
A szilárd hordozóról a peptidet/peptidkonjugátumot és az oldallánc véd�csoportokat
TFA segítségével távolítottuk el. A hasítóelegy összetételét a peptid/peptidkonjugátum
szekvenciája és az oldallánc véd�csoportok határozták meg. 1 g gyantára 10 ml hasítóelegyet
használtunk. A hasítóelegyet jeges h�tés közben készítettük. A reakció 2 órán át tartott
szobah�mérsékleten. A hasítóelegy TFA-at, fenolt, vizet, 1,2-etánditiolt (EDT) és tioanizolt,
82,5:5:5:2,5:5 (V/m/V/V/V) tartalmazott. A hasítás után a keveréket hideg éterbe (0°C)
sz�rve kicsapattuk, majd 10 perces 2000/perc fordulatszámú centrifugálással ülepítettük. Az
üledékr�l dekantálással távolítottuk el az étert, majd a peptidet/peptidkonjugátumot újra
felszuszpendálva hideg éterben megismételtük a m�veletet. Ezután A eluensben (0,1% TFA/
víz (V/V)) feloldottuk a peptidet/peptidkonjugátumot és vákuum alatt bepároltuk, majd
liofilizáltuk.
IV.3. A hatóanyag-konjugátumok szintézise
A hatóanyag (INH) kapcsolása a peptidhez egy távoltartó egységen („linker”,
glioxilsav) keresztül történt, 2-(izonikotinoil-hidrazino)ecetsav formában.
IV.3.1. 2-(izonikotinoil-hidrazino)ecetsav szintézise
Els� lépésben 2-(izonikotinoil-hidrazono)ecetsavat állítottuk el�. Ehhez 3,032 g
glioxilsavat (glioxilsav-monohidrát formában, Mw=92,05) 3 ml desztillált vízben oldottunk.
Az izoniazidból (Mw=137,14) ekvimoláris mennyiséget mértünk be és ezt acetonitril
(AcN)/víz 1:2 elegyében oldottuk. A két oldat összeöntése után (a végtérfogat 27 ml volt) a

reakcióelegyet egy órán át kevertettük szobah�mérsékleten, majd a kivált csapadékot sz�rtük,
mostuk vízzel és AcN/víz 1:4 elegyével, majd exszikkátorban P2O5 fölött szárítottuk.
A második lépés a 2-(izonikotinoil-hidrazono)ecetsav (INH-gli(ox)) redukálása volt.
Ehhez 3,114 g INH-gli(ox)-at (Mw=194,19) 40 ml absz. metanolban oldottunk és 15-20
percenként egy órán belül kis mennyiségekben adagoltuk hozzá az ekvimoláris mennyiség�
nátrium-ciano-borohidridet (1,009 g, Mw=62,84). Az elegyet 12 órán át kevertettük
szobah�mérsékleten, majd sz�rtük és mostuk metanollal. A sz�rletet vákuumban bepároltuk
60°C-on, majd hexán hozzáadásával újra bepároltuk. A termék (2-(izonikotinoil-
hidrazino)ecetsav) sárga kristályok formájában vált ki. A nyers terméket 1 M HCl-ból
átkristályosítottuk. A kivált fehér kristályokat sz�rtük, hideg vízzel mostuk, majd
exszikkátorban szárítottuk.
IV.3.2. 2-(izonikotinoil-hidrazino)ecetsav konjugálása peptidekhez
A T20 és a T6 peptidek esetében a lizinek oldallánc véd�csoportjait (Fmoc), a 91-106
peptid esetében pedig az N- terminális véd�csoportot (Fmoc) eltávolítottuk 2% piperidin, 2%
DBU, 96% DMF elegyével, 2+2+5+10 perc alatt. A konjugálást a szabad aminocsoportokra
számolva 5 ekvivalens PyBOP, 5 ekvivalens HOBt, 10 ekvivalens DIEA jelenlétében
végeztük 5 ekvivalens (2-izonikoninoil-hidrazino)ecetsavval. A kapcsolási id� minden
esetben egy óra volt.
IV.4. Tisztítási és analitikai módszerek
A lehasított peptideket, peptidszármazékokat RP-HPLC segítségével választottuk el a
szennyezésekt�l. A nyers termékek homogenitását minden esetben analitikai RP-HPLC
vizsgálattal ellen�riztük. A célvegyületet a frakciók azonosítása (aminosavanalízis,
tömegspektrometria) után a retenciós id� meghatározásával (RP-HPLC) is jellemeztük. A
peptidek analitikai jellemzését az 5-7. táblázatok tartalmazzák.
IV.4.1. A hatóanyag-konjugátumok/peptidszármazékok tisztítása
Az el�zetesen feldolgozott peptidszármazékokat RP-HPLC alkalmazásával
tisztítottuk. Knauer készülék, Phenomenex fordított fázisú, C18 oszlopok felhasználásával,
félpreparatív eljárással választottuk el a nyers terméket a szennyezésekt�l. Az oszlopok

mérete 10 x 250 mm, töltetük 10 µm szilika (300 Å) volt. Az oszlop ekvilibrálásához és az
elúcióhoz alkalmazott elegyek a következ�ek voltak: A eluens: 0,1% TFA/víz (V/V), B
eluens: 0,1% TFA/acetonitril-víz (80:20 V/V). Minden peptidszármazék esetében lineáris
gradiens elúciót alkalmaztunk, a gradienst a peptidszármazék hidrofilitása határozta meg. A
detektálás abszorbancia mérésével történt, λ=220 nm-en. Szobah�mérsékleten végeztük az
elválasztásokat, a folyássebesség 4 ml/perc volt. A tisztítandó mintát A eluensben oldottuk
(estenként kevés B eluens hozzáadásával) 2-3 mg/ml töménységben. Az oszlopra egyszerre 2
ml-t injektáltunk. A f�komponenst tartalmazó elegyet vákuum alatt bepároltuk, a bepárlási
maradékot lefagyasztottuk, majd liofilizáltuk.
IV.4.2. Peptidek és konjugátumok analitikai jellemzése
A peptidek és peptidszármazékok analitikai jellemzését analitikai RP-HPLC
vizsgálattal, aminosavanalízissel és tömegspektrometriai mérésekkel végeztük.
IV.4.2.1. Analitikai RP-HPLC vizsgálatok
Az analitikai RP-HPLC vizsgálatok KNAUER rendszeren a IV.4.1. fejezetben
ismertetett eluensek alkalmazásával történtek. Az analitikai oszlopon (Eurospher C18 250 x
4mm, 5µm szilika, 100 Å) lineáris gradiens elúciót alkalmaztunk. Az A eluensben oldott 0,5-
1,0 mg/ml töménység� mintákból az oszlopra 20 µl mennyiségeket injektáltunk. Az
elválasztásokat szobah�mérsékleten végeztük, a folyási sebesség 1 ml/perc volt. A detektálás
a félpreparatív tisztítási eljárásnál alkalmazottal megegyez�en abszorbancia mérésével
(λ=220nm) történt.
IV.4.2.2. Peptidek és konjugátumok aminosavanalízise
Az aminosavanalízis során a tisztított peptidek 100-250 µg-nyi mennyiségét 6 M HCl-
dal, 110oC-on, 24-36 óra alatt hidrolizáltuk. A hidrolizátumból az aminosavösszetételt
SYKAM 4300 automata aminosav analizátor segítségével, ninhidrin módszerrel határoztuk
meg. Az aminosavanalíziseket az MTA-ELTE Peptidkémiai Kutatócsoportban dr.
Medzihradszky Schweiger Hedvig végezte.

IV.4.2.3. ESI-MS
A mérések Bruker Esquire 3000+ típusú ioncsapdás tömegspektrométer készüléken
történtek. A mintákat 0,1 % AcOH-t tartalmazó ACN/víz 1:1 (V/V) oldószerelegyben
oldottuk. A mintákat 10 µl/perc áramlási sebességgel injektáltuk. A minták ionizációja
elektrospray forrással történt. A spektrumokat pozitív módban vettük fel 50-3000 m/z
tartományban, 13,000 m/z/sec mintavételi sebességgel. A tömegspektrometriai méréseket az
MTA-ELTE Peptidkémiai Kutatócsoportban Horváti Kata (PhD hallgató) végezte.
IV.4.3. Elemanalízis
A 2-(izonikotinoil-hidrazino)ecetsav, valamint a 2-(izonikotinoil-hidrazono)ecetsav
vegyületek elemanalízise VARIO EL III automata elemanalizátoron készült. A méréseket Dr.
Medzihradszky Schweiger Hedvig végezte.
IV.4.4. Olvadáspont meghatározás
A 2-(izonikotinoil-hidrazino)ecetsav, valamint a 2-(izonikotinoil-hidrazono)ecetsav
vegyületeket achátmozsárban való elporítás után egyik végén leforrasztott üvegkapillárisba
töltöttük. Az olvadáspontot Büchi 530 készülék segítségével határoztuk meg 0,5oC/perc f�tési
sebesség mellett.
IV.5. Az INH, a hordozómolekulák és az INH-konjugátumok minimális inhibiciós
koncentrációjának (MIC) és telepszámának (CFU) meghatározása
Kísérleteinket az Országos Korányi TBC és Pulmonológiai Intézet területén található
Bakteriológiai Laboratóriumban (Corden International Magyarország Kft.), Dr. Szabó Nóra
irányitásával és Dávid Sándor közrem�ködésével végeztük.
A vizsgálatokhoz folyékony, félszintetikus Sula táptalajt használtuk, a kémcsövek 5
ml steril tápfolyadékot tartalmaztak (Sula 1963; Sula 1963; Lányi 1980). Els� lépésként a
táptalajt tartalmazó csövekbe a vizsgálandó vegyület (INH, hordozó, INH-konjugátum)
megfelel� koncentrációjú oldatából egyforma mennyiségeket, 100 µl-t pipettáztuk. A
végkoncentráció INH-ra nézve a következ� sorozatnak megfelel� volt (γ [µg/ml]): 0,05; 0,1;
0,14; 0,16; 0,18; 0,20; 0,22; 0,24; 0,30; 0,40.

A vegyületekb�l a törzsoldatokat (INH tartalomra nézve 0,031 mg/ml) és a megfelel�
higításokat steril körülmények között készítettük. A vegyületek különböz� koncentrációit 2
párhuzamosban teszteltük. Minden vegyületet legalább 2 független kísérletben vizsgáltunk.
Az inokulum M. tuberculosis (H37RV, ATCC 27294) 4 hetes, friss tenyészetéb�l
készült, röviden: a H37RV törzsb�l golyós lombikban megfelel� denzitású szuszpenziót
készítettünk (0,5 Mcfarland, 1,5.108 CFU/ml(McFarland 1907)) Sauton táptalajjal. A
vizsgálandó vegyületet tartalmazó és a kontroll csöveket a baktériumszuszpenzió 103–104
hígításainak 100 – 100 µl-vel fert�ztük be. A mintákat 28 napig, 37oC-os thermosztátban
inkubáltuk.
A 28 nap elteltével meghatároztuk a vizsgált vegyületnek azt a legkisebb
koncentrációját, amely szabad szemmel vizsgálva teljes gátlást okozott, a kontrol csövekben
tapasztalható zavarosodáshoz képest (baktériumok szaporodása). A növekedést szabad
szemmel vizsgálva nem mutató csövekb�l szilárd Löwenstein-Jensen (LJ) táptalajokra
oltottunk ki, és 4 – 6 hét múlva megszámláltuk a kinövekedett kolóniák számát
(CFU)(Löwenstein 1931; Jensen 1932). A kolónia vagy telepszámnak a megfelel�
baktériumszuszpenzió hígítással való szorzatát hasonlítottuk össze a kiindulásként
alkalmazott 0,5 Mcfarland szuszpenzió kolónia számával (1,5.108 CFU/ml).
IV.6. A sejtek életképességének vizsgálata kolorimetriás tetrazólium (MTT) teszt alkalmazásával
Az INH, a hordozómolekulák és az INH-konjugátumok citosztatikus hatását a sejtek
életképességének vizsgálatával, tetrazólium (MTT) módszer (Slater et al., 1963; Mosmann
1983; Gerlier and Thomasset 1986) segítségével in vitro határoztuk meg humán HepG2 –
májból izolált, hepatocelluláris karcinoma sejtvonalon.
A sejtek kultúrában tartása, növesztése 150 ml térfogatú szövettenyészt� flaskákban
történt (37oC, 5% CO2 atmoszféra), RPMI 1640 médiumban. A teljes (komplett) médium
10% FCS-t (magzati borjú szérum, foetal calf serum), L-glutamint (2 µmol/ml), nátrium-
piruvátot (1 µmol/ml) és gentamicint (160 µg/ml) tartalmazott.
A kísérlet els� napján a sejteket egy 96 lyukú szövettenyészt� lemezre osztottuk 100
µl szérumtartalmú, teljes médiumban (HepG2 sejtek esetében: 5.103 sejt/lyuk). Következ� nap
50 µl médium eltávolítását követ�en, a negatív kontroll sejtekre 150 µl szérummentes
médiumot, míg a kezelt sejtekre 50 µl szérummentes médiumot és 100 µl szérummentes
médiumban oldott különböz� koncentrációjú szabad hatóanyagot (INH), hordozómolekulát,

hatóanyag-konjugátumot pipettáztunk. A kezeléseket az INH esetében c = 3,6*10-2 – 5,5*10-6
Μ, a hordozók és a konjugátumok esetében c = 5,0*10-4 – 7,6*10-8 Μ,
koncentrációtartományban végeztük el. Három óra kezelési id� elteltével a sejteket 135-135
µl szérummentes médiummal mostuk 3-szor, majd lyukanként ugyanennyi teljes médiumot
adtunk a sejtekhez.
Három nap inkubálás után 45 µl MTT-oldatot (c=2 mg/ml, szérummentes médiumban
oldva, 0,22 µm filterrel sz�rve) adtunk a sejtekhez lyukanként, majd a lemezeket sötétben, 3,5
órát inkubáltuk. Az inkubálási id� letelte után a sejtekr�l a folyadékot G30 t� segítségével
óvatosan eltávolítottuk és a lila kristályokat 100 µl DMSO-ban oldottuk ki. A kioldódott
kristályok okozta színintenzitást mikrotitrációs lemez leolvasó spektrométer segítségével
(λ=540 nm) mértük (Elisa Reader (Labsystems, Finnország)). A mélyedésekben található
sejtek növekedésének illetve túlélésének mértékét a mért abszorbanciák segítségével
számítottuk ki.
A citosztázis mértékét (%) a koncentráció függvényében ábrázoltuk és a kapott görbék
segítségével határoztuk meg az IC50 értékeket.

V. Eredmények
Munkánk során el�állítottuk az 5. táblázatban szerepl� tuftsin származékokat és 16
kDa fehérje 91-106 szekvenciájának megfelel� peptidamidot. A szintetikus peptideket
kémiailag jellemeztük. El�állítottuk a 6. táblázatban szerepl� izoniazid származékokat és
elvégeztük a peptidekkel való konjugálásukat (7. táblázat). A felsorolt vegyületeket az INH
szelektív, receptorközvetített célbajuttatása céljából terveztük, annak érdekében, hogy els�
lépésben ezen vegyületek segítségével tanulmányozzuk a kémiai átalakítások hatását az INH
in vitro antituberkulotikus aktivitására.
V.1. Hordozó peptidek tervezése és szintézise
A peptideket Boc/ Bzl, és Fmoc/tBu stratégiával állítottuk el�. A GTKPKG (T6) és a
[TKPKG]4 (T20) peptidek esetében az el�bbit, a 91SEFAYGSFVRTVSLPV106 (91-106)
peptid esetében pedig az utóbbi módszert alkalmaztuk.
A T6 és T20 peptidek szintéziséhez MBHA gyantát alkalmaztunk. Valamennyi
aminosavat Nα-Boc-származék formájában kapcsoltuk a szintézis során (a IV.2.1. fejezetnek
megfelel�en).
A szilárd hordozóról a peptideket/peptidkonjugátumokat, illetve az oldallánc
véd�csoportokat hidrogén-fluorid (HF) segítségével távolítottuk el a IV.2.1. fejezetnek
megfelel�en. A nyers termékeket liofilizálással izoláltuk, majd tisztítottuk RP-HPLC
rendszeren (IV.4.1. fejezet). A tisztított peptidek analitikai adatait az 5. táblázatban foglaltuk
össze. Az ESI-MS és az aminosavanalízis adatok a várt értékekkel egyezést mutatnak.
Az el�állított peptidek analitikai RP-HPLC kromatogramjai és tömegspektrumai a
Függelékben találhatóak (Függelék 1-2. ábra).
A 91-106 peptidet SYRO (MultiSyntech) automata peptidszintetizátor segítségével
állítottuk el�, Fmoc/tBu-stratégia alakalmazásával. A szintézishez Rink Amide MBHA
gyantát használtunk. Valamennyi aminosavat Nα-Fmoc-származék formájában alkalmaztuk a
szintézis során (a IV.2.2. fejezetnek megfelel�en).
A szilárd hordozóról a peptideket/petidkonjugátumokat, illetve az oldallánc
véd�csoportokat TFA segítségével távolítottuk el a IV.2.2. fejezetnek megfelel�en. A nyers

terméket liofilizálással izoláltuk, majd RP-HPLC rendszer segítségével tisztítottuk (IV.4.1.
fejezet).
A nyers termék, illetve a tisztított termék homogenitását analitikai RP-HPLC
alkalmazásával vizsgáltuk. Az analitikai RP-HPLC vizsgálatokhoz a IV.4.2.1. fejezetben
ismertetett készülékeket és eluenseket használtuk. A gradiens megegyezett a tisztítás során
használt gradienssel. A tisztított peptid analitikai adatai az 5. táblázatban találhatók meg. Az
ESI-MS és az aminosavanalízis adatok a várt értékekkel egyezést mutatnak.
Az el�állított, tisztított 91-106 peptid analitikai RP-HPLC kromatogramja és
tömegspektruma a Függelékben található (Függelék 3. ábra).
.
5. táblázat. Az el�állított hordozó peptidek analitikai jellemzése
T 0.94 [1]; S 2.78 [3]; E 1.08 [1]; P 1.11 [1]; G 1.08 [1]; A 1.11 [1]; V 2.97 [3]; L 1.05 [1]; Y 0.90 [1];
F 2.03 [2]; R 1.05 [1]
T 3.63 [4]; P 3.51 [4]; G 4.82 [4]; K 8.04 [8];
T 1.01 [1]; P 1.07 [1]; G 1.94 [2]; K 1.98 [2]
Aminosavanalízis c
32,01756,91756,991SEFAYGSFVRTVSLPV106
91-106
14,62062,52062,3[TKPKG]4T20
4,0585,3585,3GTKPKGT6
Rt b
(perc)Mmo
a
mértMmo
számítottszekvenciakód
T 0.94 [1]; S 2.78 [3]; E 1.08 [1]; P 1.11 [1]; G 1.08 [1]; A 1.11 [1]; V 2.97 [3]; L 1.05 [1]; Y 0.90 [1];
F 2.03 [2]; R 1.05 [1]
T 3.63 [4]; P 3.51 [4]; G 4.82 [4]; K 8.04 [8];
T 1.01 [1]; P 1.07 [1]; G 1.94 [2]; K 1.98 [2]
Aminosavanalízis c
32,01756,91756,991SEFAYGSFVRTVSLPV106
91-106
14,62062,52062,3[TKPKG]4T20
4,0585,3585,3GTKPKGT6
Rt b
(perc)Mmo
a
mértMmo
számítottszekvenciakód
a ESI-MS, Bruker Esquire 3000+b RP-HPLC, Knauer, Eurospher-100 C18, 5µm, 250x4mm oszlop, λ=214nm, gradiens: 5-60B% 35perc. A eluest: H2O+0,1 v/v% TFA, B eluens: AcN: H2O =80:20 (v/v) +0,1 v/v% TFA C hidrolízis: 6N HCl, 24 h, 110oC
V.2. Hatóanyag-konjugátumok szintézise
Az izoniazidot a peptidekhez minden esetben egy távoltartó egységen („linker”)
keresztül kapcsoltuk. A 2-(izonikotinoil-hidrazino)ecetsav (INH-gli(red)) INH-származékban
a távoltartó egység a „beépített” glioxilsav részlet.
V.2.1. Az izoniazid származékok el�állítása távlságtartó „linker” beépítésével
A 2-(izonikotinoil-hidrazono)ecetsavat INH és glioxilsav reakciójával állítottuk el� a
IV.3.1. fejezetben leírtaknak megfelel�en, 98%-os termeléssel. A termék analitikai adatait a 6.
táblázat tartalmazza, tömegspektruma a Függelékben található (Függelék 4. ábra). A mosott,
szárított 2-(izonikotinoil-hidrazono)ecetsavat NaCNBH3-del redukáltuk a IV.3.1. fejezetnek
megfelel�en. A sz�rletet (INH-gli(red)) vákuumban bepároltuk, majd a nyers terméket
átkristályosítottuk. A csapadékot sz�rtük, mostuk, majd vákuumexszikkátorban szárítottuk
(kitermelés: 95%). A termék tömegspektruma a Függelékben (Függelek 4. ábra), az analitikai
adatai pedig szintén a 6. táblázatban találhatóak. A 2-(izonikotinoil-hidrazino)ecetsav
szintézisének reakcióegyenlete a 14. ábrán látható.
N
O
NH
NH2
+CHO
COOH1 h
AcN:H2O1:4
N
O
NHN
CHCOOH
NaCNBH3red.
abs. MeOH
N
O
NHNH
CH2
COOH
izoniazid glioxilsav
14. ábra. A 2-(izonikotinoil-hidrazino)ecetsav szintézise
6. táblázat. Az el�állított hatóanyagszármazékok analitikai elemzése
95N%: (21,54); 20,96C%: (49,23); 47,79H%: (4,62); 4,33
192,5195,0195,12-(izonikotinoil-hidrazino)ecetsav
INH-gli(red)
98N%: (21,75); 22,15C%: (49,72); 49,76H%: (3,63); 3,53
204,5 193,0193,12-(izonikotinoil-hidrazono)ecetsav
INH-gli(ox)
Kitermelés(%)
ElemanalíziscOlvadáspontb
(t/°C)Mmo
a
mértMmoszámított
Teljes névKód
95N%: (21,54); 20,96C%: (49,23); 47,79H%: (4,62); 4,33
192,5195,0195,12-(izonikotinoil-hidrazino)ecetsav
INH-gli(red)
98N%: (21,75); 22,15C%: (49,72); 49,76H%: (3,63); 3,53
204,5 193,0193,12-(izonikotinoil-hidrazono)ecetsav
INH-gli(ox)
Kitermelés(%)
ElemanalíziscOlvadáspontb
(t/°C)Mmo
a
mértMmoszámított
Teljes névKód
a ESI-MS, Bruker Esquire 3000+b Büchi 530 olvadáspontmér�c VARIO EL III elemanalizátor
V.2.2. Izoniazid származék konjugálása peptidekhez
A hatóanyag konjugálása el�tt a peptidekr�l eltávolítottuk az Fmoc-
véd�csoportokat a IV.3.2. fejezetben leírtaknak megfelel�en. A konjugálást
PyBOP/HOBt/DIEA kapcsolóreagensek jelenlétében végeztük. A 2-(izonikotinoil-
hidrazino)ecetsav peptidhez történ� konjugálásának reakcióegyenlete a 15. ábrán található. A
kész hatóanyag-konjugátumokat a T6(INH) és a T20(4INH) esetében HF-dal, az INH-gli-91-

106 esetében pedig TFA-val távolítottuk el a gyantáról a IV.2.1. és IV.2.2. fejezetekben
leírtaknak megfelel�en. A nyers termékeket liofilizálással izoláltuk.
N
O
NHNH COOH
+ NH2
R
C
O
peptidgyanta
N
O
NHNH
O
NH
R
Cpeptidgyanta
O
15. ábra. A 2-(izonikotinoil-hidrazino)ecetsav peptidhez történ� konjugálásának reakcióegyenlete
A szintézis után a peptidkonjugátumok tisztítását RP-HPLC alkalmazásával végeztük
(IV.4.1. fejezet).
A nyers termék, illetve a tisztított termék homogenitását analitikai RP-HPLC
alkalmazásával vizsgáltuk. Az analitikai RP-HPLC vizsgálatokat a IV.4.2.1. fejezetben
ismertetettük. Az el�állított petidkonjugátumok analitikai RP-HPLC kromatogramjai és
tömegspektrumai a Függelékben találhatóak (Függelék 5-7. ábrák). A 7. táblázatban
összefoglalt ESI-MS és aminosavanalízis adatok a várt értékekkel egyezést mutatnak. Az
el�állított konjugátumok sematikus szerkezete a 16. ábrán látható.
7. táblázat. Az el�állított peptidkonjugátumok analitikai jellemzése
T 1.01 [1]; P 1.07 [1]; G 1.94 [2]; K 1.98 [2]14,8762,4762,4GTKPK(INH)GT6(INH)
T 0.94 [1]; S 2.78 [3]; E 1.08 [1]; P 1.11 [1]; G 1.08 [1]; A 1.11 [1]; V 2.97 [3]; L 1.05 [1]; Y 0.90 [1];
F 2.03 [2]; R 1.05 [1]
T 3.63 [4]; P 3.51 [4]; G 4.82 [4]; K 8.04 [8];
Aminosavanalízis c
18,52770,52770,7[TKPK(INH)G]4T20(4INH)
30,51934,01934,0INH-gli-91SEFAYGSFVRTVSLPV106INH-gli-91-106
Rt b
(perc)Mmo
a
mértMmo
számítottszekvenciakód
T 1.01 [1]; P 1.07 [1]; G 1.94 [2]; K 1.98 [2]14,8762,4762,4GTKPK(INH)GT6(INH)
T 0.94 [1]; S 2.78 [3]; E 1.08 [1]; P 1.11 [1]; G 1.08 [1]; A 1.11 [1]; V 2.97 [3]; L 1.05 [1]; Y 0.90 [1];
F 2.03 [2]; R 1.05 [1]
T 3.63 [4]; P 3.51 [4]; G 4.82 [4]; K 8.04 [8];
Aminosavanalízis c
18,52770,52770,7[TKPK(INH)G]4T20(4INH)
30,51934,01934,0INH-gli-91SEFAYGSFVRTVSLPV106INH-gli-91-106
Rt b
(perc)Mmo
a
mértMmo
számítottszekvenciakód
a ESI-MS, Bruker Esquire 3000+b RP-HPLC, Knauer, Eurospher-100 C18, 5µm, 250x4mm oszlop, λ=214nm, gradiens: 5-60B% 35perc. A eluest: H2O+0,1 v/v% TFA, B eluens: AcN: H2O =80:20 (v/v) +0,1 v/v% TFA C hidrolízis: 6N HCl, 24 h, 110oC
TKPKGTKPKGTKPKGTKPKGO
N
O NH
NH
O
N
O NH
NH
O
N
O NH
NH
O
N
O NH
NH
O
N
O
NH NH=ASEFAYGSFVRTVSLPVABC
GTKPKGO
N
O NH
NH
T6(INH) T20(4INH) INH-gli-91-106
TKPKGTKPKGTKPKGTKPKGO
N
O NH
NH
O
N
O NH
NH
O
N
O NH
NH
O
N
O NH
NH
O
N
O
NH NH=ASEFAYGSFVRTVSLPVABC
GTKPKGO
N
O NH
NH
T6(INH) T20(4INH) INH-gli-91-106
TKPKGTKPKGTKPKGTKPKGO
N
O NH
NH
O
N
O NH
NH
O
N
O NH
NH
O
N
O NH
NH
TKPKGTKPKGTKPKGTKPKGO
N
O NH
NH
O
N
O NH
NH
O
N
O NH
NH
O
N
O NH
NH
O
N
O
NH NH=ASEFAYGSFVRTVSLPVABC
O
N
O
NH NH=ASEFAYGSFVRTVSLPVABC
GTKPKGO
N
O NH
NH
GTKPKGO
N
O NH
NH
T6(INH) T20(4INH) INH-gli-91-106
16. ábra. Az el�állított konjugátumok szerkezete

V.3. A hordozók, hatóanyagok és konjugátumok in vitro biológiai aktivitásának
vizsgálata
V.3.1. Minimális inhibiciós koncentráció (MIC) és telepszám (CFU) meghatározása
A kísérleteket az ún. „higitásos leves” módszerrel végeztük M. tuberculosis (H37RV)
tenyészeten. A vizsgálatokhoz folyékony, félszintetikus Sula táptalajt használtunk. Minden
vegyületet két független kísérletben vizsgáltunk.
28 nap elteltével meghatároztuk a vizsgált vegyületnek azt a legkisebb
koncentrációját, amely szabad szemmel vizsgálva teljes gátlást okozott, a kontroll csövekben
tapasztalható zavarosodáshoz képest, amit a baktériumok szaporodása okozott. A növekedést
nem mutató csövekb�l szilárd Löwenstein-Jensen (LJ) táptalajokra oltottunk ki, és
meghatároztuk a kolóniák számát (CFU). A meghatározott MIC és CFU értékek a 8.
táblázatban foglaltuk össze.
8. táblázat. A szabad INH és az INH-hordozó konjugátumok esetében Sula táptalajon meghatározott MIC és CFU értékek
---Nincs hatásGTKPKG
1,12x10-62,2620,16INH-91SEFAYGSFVRTVSLPV106
2,26x10-66,47500,32[TKPK(INH)G]4
1,31x10-61,00200,18GTKPK(INH)G
-
-
6
12
CFU
-
-
0,57
0,16
konjugátumraszámolva γγγγ (µµµµg/ml)
2,07x10-60,40INH-gli(red)
1,17x10-60,16INH
Nincs hatás
Nincs hatás
MIC*γγγγ (µµµµg/ml)
-[TKPKG]4
-91SEFAYGSFVRTVSLPV106
konjugátumraszámolva c(mol/dm3)
Hatóanyag / konjugátum / hordozó
---Nincs hatásGTKPKG
1,12x10-62,2620,16INH-91SEFAYGSFVRTVSLPV106
2,26x10-66,47500,32[TKPK(INH)G]4
1,31x10-61,00200,18GTKPK(INH)G
-
-
6
12
CFU
-
-
0,57
0,16
konjugátumraszámolva γγγγ (µµµµg/ml)
2,07x10-60,40INH-gli(red)
1,17x10-60,16INH
Nincs hatás
Nincs hatás
MIC*γγγγ (µµµµg/ml)
-[TKPKG]4
-91SEFAYGSFVRTVSLPV106
konjugátumraszámolva c(mol/dm3)
Hatóanyag / konjugátum / hordozó
*INH tartalomra számolva Vizsgált tartomány: c = 0,02 – 0,5 µg/mlIrodalmi adatok alapján MIC (INH): 0,1-0,2 µg/ml
A kapott eredmények alapján megállapíthatjuk, hogy a konjugátumok MIC értéke jó egyezést
mutat a szabad izoniazid MIC értékével. A T20(4INH) esetében nem kaptuk meg a
négyszeres mennyiség� izoniazodnak megfeleltethet� gátló koncentrációt. Tervezzük a
konjugátumokból és az INH-származékokból az INH-felszabadulás mechanizmusának,
valamint a konjugátumok sejtbejutási mechanizmusának vizsgálatát makrofág modelleken.

V.3.2. Citosztatikus hatás vizsgálata
Az INH, a hordozómolekulák és a INH-konjugátumok citosztatikus hatását a sejtek
életképességének vizsgálatával, a tetrazólium (MTT) módszer segítségével in vitro határoztuk
meg humán HepG2 – májból izolált, hepatocelluláris karcinoma sejtvonalon.
A sejteket teljes médiumban növesztettük, majd 96 lyukú szövettenyészt� lemezeken
kezeltük a vegyületekkel (szabad hatóanyagok, hordozómolekulák, hatóanyag-konjugátumok)
megfelel� koncentrációtartományban (általában az oldhatóságtól függ�en 10-3 – 10-7 M
koncentrációban), 3 óra id�tartamig. A kezelést követ�en a hatóanyagokat mosással
eltávolítottuk a sejtekr�l. A citosztatikus vizsgálatok esetén, a sejteket a kezelést követ�en 3
napig kultúrában tartottuk, teljes médiumban, az MTT-oldat hozzáadása el�tt, hogy
meghatározhassuk a vegyület proliferáció gátló hatását az adott sejteken.
Minden vegyület esetében három független kísérletben, 6-8 párhuzamos méréssel
határoztuk meg a citosztatikus hatás mértéket. A kapott eredmények a 17. ábrán foglaltuk
össze, a T20(4INH) és az INH-gli-91-106 esetében a citosztatikus hatás meghatározása
folyamatban van.
1E-5 1E-4 1E-3 0.01
0
20
40
60
80
100
cyto
stas
is %
lg (c/M)
INH
INHIC50 > 3,6. 10-2MMIC =1,17.10-6 M
1E-7 1E-6 1E-5 1E-4
0
20
40
60
80
100
cyto
stas
is %
lg (c/M)
T6IC50 > 5,0. 10-4 M
1E-7 1E-6 1E-5 1E-4
0
20
40
60
80
100
cyto
stas
is %
lg (c/M)
T6(INH) konjugátumIC50 > 5,0. 10-4 MMIC =1,31 . 10-6 M
Cito
sztá
zis
%
Cito
sztá
zis
%
Cito
sztá
zis
%
17. ábra. Az INH és az INH-hordozó konjugátumok citosztatikus hatása a koncentráció függvényében
A grafikonok alapján megállapíthatjuk, hogy a vizsgált hatóanyagnak, hordozónak és
hatóanyag-konjugátumnak a legkisebb gátló koncentrációban nincs citosztatikus hatása.

VI. Összefoglalás
• El�állítottuk a GTKPKG (T6), [TKPKG]4 (T20) és a 91SEFAYGSFVRTVSLPV106
(91-106) peptidamidokat. A peptidek homogenitását RP-HPLC segítségével
jellemeztük, az el�állított, tisztított peptidek >95%-os tisztaságúak voltak. A
peptidamidok els�dleges szerkezetetét tömegspektrometriával és aminosavanalízissel
igazoltuk.
• El�állítottuk a 2-(izonikotinoil-hidrazono)ecetsav (INH-gli(ox)) és a 2-(izonikotinoil-
hidrazino)ecetsav (INH-gli(red)) INH-származékokat >95%-os kitermeléssel.
Meghatároztuk az új vegyületek olvadáspontját és elemi összetételét.
• Bizonyítottuk, hogy a 2-(izonikotinoil-hidrazino)ecetsav INH-származék alkalmas
reakciópartner szilárd fázison mind Boc/Bzl, mind pedig Fmoc/tBu stratégia
alkalmazása esetén peptidekhez való konjugáláshoz. A 2-(izonikotinoil-
hidrazino)ecetsav alkalmazásával állítottuk el� a GTKPK(INH)G, [TKPK(INH)G]4 és
a INH-gli-91SEFAYGSFVRTVSLPV106 hatóanyag-konjugátumokat. Az analitikai
vizsgálatok során a várt homogenitású és összetétel� vegyületeket kaptuk.
• Meghatároztuk az INH, a hordozómolekulák és az INH-konjugátumok minimális
inhibiciós koncentrációját (MIC) és telepszámát (CFU) in vitro M. tuberculosis H37Rv
törzsön Sula félszintetikus, folyékony táptalajon. A kapott eredmények alapján
megállapíthatjuk, hogy a konjugátumok MIC értéke jó egyezést mutat a szabad INH
MIC értékével. A T20(4INH) vegyület esetében nem kaptunk meg a 4 izoniazidnak
megfeleltethet� gátló koncentrációt, ebben a konstrukcióban is a többi konjugátummal
megfelel� egyezést mutató MIC értéket kaptunk. Tervezzük az INH felszabadulási,
valamint a konjugátumok sejtbejutási mechanizmusámak vizsgálatát humán makrofág
típusú sejtvonalakon.
• Meghatároztuk az INH-konjugátumok citosztatikus hatását kolorimetriás tetrazólium
(MTT) teszt alkalmazásával. Az eredményeink igazolták, hogy in vitro HepG2
sejteken az INH önmagában és az INH-konjugátumok a vizsgált koncentráció
tartományban nem citosztatikus hatásúak.
• Az INH, a hordozómolekulák és az INH-konjugátumok membránhoz való affinitását
monomolekulás lipid film modellen, Langmuir-mérleggel Dr. Kiss Éva irányításával
Pénzes Csanád Botond végezte el a Fizikai Kémia Tanszék Határfelületi és
Nanoszerkezetek Laboratóriumában. Munkája eredményeit tudományos diákköri
dolgozatban foglalta össze.

VII. Rövidítésjegyzék
A dolgozatban el�forduló rövidítések, jelölések jegyzéke
AcN acetonitril
Boc terc-butiloxi-karbonil
Bzl benzil
CFU kolónia szám, telepszám
(colony forming unit)
DBU 1,8-diazabiciklo[5.4.0]undec-7-én
DCC N,N’-diciklohexil-karbodiimid
DCM diklór-metán
DIC N,N’-diizopropil-karbodiimid
DIEA N-etil-diizopropil-amin
DMF N,N’-dimetil-formamid
EDT 1,2-etánditiol
ESI-MS electrospray ionizációs tömegspektrométer
Fmoc 9-fluorenil-metiloxi-karbonil
FCS magzati (foetális) borjú savó
(foetal calf serum) Gli glioxilsav-monohidrát
HepG2 májból izolált, hepatocelluláris karcinoma
sejtvonal
HF hidrogén-fluorid
HCl hidrogén-klorid
HOBt 1-hidroxi-benztriazol
HPLC high performance liquid chromatography
INH izoniazid
INH-gli(ox) (2-izonikotinoil-hidrazono)ecetsav
INH-gli(red) (2-izonikotinoil-hidrazino)ecetsav
MBHA 4-metil-benzhidrilamin
MeOH metanol
MIC minimális inhibiciós koncentráció
(minimum inhibitory concentration)

MS mass spectrometry
MTT (3-(4,5-dimetil-tiazol-2-il)
-2,5-difenil-tetrazolium) bromid
NaCNBH3 nátrium-ciano-borohidrid
PyBOB benzotriazol-1-il-oxi-trisz-pirrolidino-
-foszfónium hexafluorofoszfát
PZA pirazinamid
P2O5 foszfor-pentoxid
RAMP rifampicin
Rink Amide MBHA 4-(2’, 4’-dimetoxifenil-Fmoc-
-aminometil)-fenoxiacetamido-
-norleucil-MBHA
RP-HPLC revers phase high performance liquid
chromatography
RPMI 1640 szövettenyészt� médium
SM sztreptomicin
tBu terc-butil
TFA trifluor-ecetsav

VIII. Irodalomjegyzék
Ádám, É., F. D. Tóth, L. Em�dy, L. Gergely, É. Gönczöl, E. Nagy, T. Pál, R. Pusztai, F.
Rozgonyi and B. Szabó (2003). Orvosi mikrobiológia, Alliter Kiadói és Oktatásfejlesztõ Alapítvány.
Albericio, F., R. van Abel and G. Barany (1990). "Solid-phase synthesis of peptides with C-terminal asparagine or glutamine. An effective, mild procedure based on N alpha-fluorenylmethyloxycarbonyl (Fmoc) protection and side-chain anchoring to a tris(alkoxy)benzylamide (PAL) handle." Int J Pept Protein Res 35(3): 284-286.
Atherton, E., H. Fox, D. Harkiss, C. J. Logan, R. L. Sheppard and B. J. Williams (1978). "A mild procedure for solid phase peptide synthesis: Use fluorenylmethyloxy-carbonylamino acids." J Chem Soc Chem Commun: 537-540.
Atherton, E., W. Hubscher, R. C. Sheppard and V. Woolley (1981). "Synthesis of a 21-residue fragment of human proinsulin by the polyamide solid phase method." Hoppe Seylers Z Physiol Chem 362(7): 833-839.
Barany, G., N. Kneib-Cordonier and D. G. Mullen (1987). "Solid-phase peptide synthesis: a silver anniversary report." Int J Pept Protein Res 30(6): 705-739.
Bollela, V. R., D. N. Sato and B. A. Fonseca (1999). "McFarland nephelometer as a simple method to estimate the sensitivity of the polymerase chain reaction using Mycobacterium tuberculosis as a research tool." Braz J Med Biol Res 32(9): 1073-1076.
B�sze, S., N. Caccamo, Z. Majer, G. Mez�, F. Dieli and F. Hudecz (2004). "In vitro T-Cell Immunogenicity of Oligopeptides Derived from the Region 92-110 of the 16 kDa Protein of Mycobacterium tuberculosis." Biopolymers 76: 467-476.
Brennan, P. J. (2003). "Structure, function, and biogenesis of the cell wall of Mycobacterium tuberculosis." Tuberculosis (Edinb) 83(1-3): 91-97.
Bump, N. J., J. Lee, M. Wleklik, J. Reichler and V. A. Najjar (1986). "Isolation and subunit composition of tuftsin receptor." Proc Natl Acad Sci U S A 83(19): 7187-7191.
Bump, N. J., V. A. Najjar and J. Reichler (1990). "The characteristics of purified HL60 tuftsin receptors." Mol Cell Biochem 92(1): 77-84.
Carpino, L. A. and G. Y. Han (1972). "The fluorenylmethoxycarbonyl amino-protecting group." Journal of Organic Chemistry 37: 3404-3409.
Castro, B., J. R. Dormoy, G. Ervin and C. Selve (1975). "Reactifs de couplage peptidique IV (1) L'hexafluorophophate de benzotriazolyl N-oxitrisdimethylamino phosphonium (B.O.P)." Tetrahedron Letters 14: 1219-1222.
Chang, C. D., A. M. Felix, M. H. Jimenez and J. Meienhofer (1980). "Solid-phase peptide synthesis of somatostatin using mild base cleavage of N alpha-9-fluorenylmethyloxycarbonylamino acids." Int J Pept Protein Res 15(5): 485-494.
Coste, J., D. Lenguyen and B. Castro (1990). "PyBOP - A new peptide coupling reagent devoid of toxic by-product." Tetrahedron Letters 31: 205-208.
Cunningham, A. F. and C. L. Spreadbury (1998). "Mycobacterial stationary phase induced by low oxygen tension: cell wall thickening and localization of the 16-kilodalton alpha-crystallin homolog." J Bacteriol 180(4): 801-808.
Egerszegi, S. (1995). "A tuberculosis kezelésének aktuális problémái." Magyar Infektológiai Társaság tudományos és továbbképzõ folyóirata 2. évf. 1. sz.: 20-26.
Fridkin, M. and P. Gottlieb (1981). "Tuftsin, Thr-Lys-Pro-Arg. Anatomy of an immunologically active peptide." Mol Cell Biochem 41: 73-97.
Fridkin, M. and V. A. Najjar (1989). "Tuftsin: its chemistry, biology, and clinical potential." Crit Rev Biochem Mol Biol 24(1): 1-40.

Fridkin, M., Y. Stabinsky, V. Zakuth and Z. Spirer (1977). "Tuftsin and some analogs: synthesis and interaction with human polymorphonuclear leukocytes." Biochim Biophys Acta 496(1): 203-211.
Fridkin, M., H. Tsubery, E. Tzehoval, A. Vonsover, L. Biondi, F. Filira and R. Rocchi (2005). "Tuftsin-AZT conjugate: potential macrophage targeting for AIDS therapy." J Pept Sci 11(1): 37-44.
Frieden, T. R., T. R. Sterling, S. S. Munsiff, C. J. Watt and C. Dye (2003). "Tuberculosis." Lancet 362(9387): 887-899.
Gergely, P. (1994). "A tuberculosis az immunológus szemével." Lege Artis Medicinae 4: 402-411.
Gerlier, D. and N. Thomasset (1986). "Use of MTT colorimetric assay to measure cell activation." J Immunol Methods 94(1-2): 57-63.
Gottlieb, P., E. Tzehoval, M. Feldman, S. Segal and M. Fridkin (1983). "Tuftsin binding to various macrophage hybridomas." Ann N Y Acad Sci 419: 107-113.
Grant, A. G. (1992b). "Principles and Practice of Solid-Phase Peptide Synthesis, in:." Synthetic peptides A User's Guide, Oxford, University Press, UK: pp. 88-96.
Gupta, C. M. and W. Haq (2005). "Tuftsin-bearing liposomes as antibiotic carriers in treatment of macrophage infections." Methods Enzymol 391: 291-304.
Hudecz, F. (2001). "Manipulation of epitope function by modification of peptide structure: A minireview." Biologicals 29: 197-207.
Jensen, K. A. (1932). "Rinzuchtung und Typenbestimmung von Tuberkelbazillentamen. Zentralb." Bakteriol Parasitenkd. infektionskr. hyg. Agt. I Orig. 125: 222.
Jónás, J., K. Barsiné Fodor, K. P., M. Péterfiné Tüfgyei and N. L. (2005). A pulmonológiai Intézmények 2004. évi epidemiológiai és mûködési adatai, Országos Korányi TBC és Pulmonológiai Intézet.
Kaiser, E., R. L. Colescott, C. D. Bossinger and P. I. Cook (1970). "Color test for detection of free terminal amino groups in the solid-phase synthesis of peptides." Anal.Biochem 34: 595-598.
Kaiser, E., R. L. Colescott, C. D. Bossinger and D. B. Olser (1980). "Color test for terminal prolyl residues in the solid-phase synthesis of peptides." Anal.Chem.Acta 118: 149-151.
Kaufmann, S. H. E. and S. K. Parida (2007). "Changing funding patterns in tuberculosis." Nature Medicine 13: 299-303.
Knowles, B. and D. Aden (1983). Human hepatoma derived cell line, process for preparation thereof, and uses therefor. USA.
Knowles, B. B., C. C. Howe and D. P. Aden (1980). "Human hepatocellular carcinoma cell lines secrete the major plasma proteins and hepatitis B surface antigen." Science 209(4455): 497-499.
König, W. (1970). "Eine neue Methode zur Synthese von Peptiden: Aktivierung der Carboxylgruppe mit mit Dicyclohexylcarbodiimid unter Zusatz von 1-hydroxy-benzotriazolen." Chem. Ber. 103: 788-798.
Krchnak, V., J. Vagner and M. Lebl (1988). "Noninvasive continuous monitoring of solid-phase peptide synthesis by acid-base indicator." Int J Pept Protein Res 32(5): 415-416.
Kusner, D. J. (2005). "Mechanisms of mycobacterial persistence in tuberculosis." Clin Immunol 114(3): 239-247.
Lányi, B. (1980). Folyékony mycobacterium tenyésztésre alkalmas standard táptalaj,. Járványügyi és Klinikai Bakteriológia, Módszertani Útmutató. B. Lányi. Budapest: 488.
Löwenstein, E. (1931). "Die Zachtung der Tuberkelbazillen aus dem stramenden Blute. Zentralb." Bakteriol Parasitenkd. infektionskr. hyg. Abt. I orig. 120: 127.

Manabe, Y. C. and W. Bishae (2000). "Latent Mycobacterium tuberculosis- persistance, patience, and winning by waiting." Nature Medicine 6: 1327-1329.
Matsueda, G. R. and J. M. Stewart (1981). "A p-methylbenzhydrylamine resin for improved solid-phase synthesis of peptide amides." Peptides 2(1): 45-50.
McFarland, J. (1907). "Nephelometer: an instrument for estimating the number of bacteria in suspensions used for calculating the opsonic index and for vaccines." Journal of the American Medical Association 14: 1176-1178.
Merrifield, R. B. (1963). "Solid phase peptide synthesis. I. The synthesis of a tetrapeptide." J. Am. Chem. Soc. 85: 2149-2154.
Merrifield, R. B. (1964). "Solid phase peptide synthesis. III. An improved snthesis of bradykinin." Biochemistry 3: 1385-1390.
Mez�, G., A. Kalászi, R. J., N. Mihala, Z. Majer, Á. Hilbert, O. Láng, L. Kõhidai, K. Barna and D. Gaál (2000). "Synthesis of new carrier molecules based on repeated tuftsin sequences." Peptides 2000.
Mojsov, S., A. R. Mitchell and R. B. Merrifield (1980). "A quantitative evaluation of methods for coupling asparagine." J. Org. Chem. 45: 555-560.
Mosmann, T. (1983). "Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays." J Immunol Methods 65(1-2): 55-63.
Najjar, V. A. (1983). "Tuftsin, a natural activator of phagocyte cells: an overview." Ann N Y Acad Sci 419: 1-11.
Newcomb, R., B. Szoke, A. Palma, G. Wang, X. Chen, W. Hopkins, R. Cong, J. Miller, L. Urge, K. Tarczy-Hornoch, J. A. Loo, D. J. Dooley, L. Nadasdi, R. W. Tsien, J. Lemos and G. Miljanich (1998). "Selective peptide antagonist of the class E calcium channel from the venom of the tarantula Hysterocrates gigas." Biochemistry 37(44): 15353-15362.
Rink, H. (1978). "Solid-phase synthesis of protected peptide fragments using a trialkoxy-diphenyl-methylester resin." Tetrahedron Letters 28: 3787-3790.
Sandy, J., S. Holton, E. Fullam, E. Sim and M. Noble (2005). "Binding of the anti-tubercular drug isoniazid to the arylamine N-acetyltransferase protein from Mycobacterium smegmatis." Protein Sci 14(3): 775-782.
Slater, T. F., B. Sawyer and U. Straeuli (1963). "Studies on Succinate-Tetrazolium Reductase Systems. Iii. Points of Coupling of Four Different Tetrazolium Salts." Biochim Biophys Acta 77: 383-393.
Sula, L. (1963). "WHO cooperative studies on a simple culture techniques for the isolation of mycobacteria: 1. preparation, lyophilization and reconstitution of a simple semi-synthetic concentrated liquid medium; culture technique, growth pattern of different mycobacteria." Bull World Health Organ 29(5): 589-606.
Sula, L. (1963). "WHO cooperative studies on a simple culture techniques for the isolation of mycobacteria: 2. Comparison of the efficacy of lyophilized liquid medium with that of Lowenstein-Jensen (L-J) medium." Bull World Health Organ 29(5): 607-625.
Tickler, A. K., C. J. Barrow and J. D. Wade (2001). "Improved preparation of amyloid-beta peptides using DBU as Nalpha-Fmoc deprotection reagent." J Pept Sci 7(9): 488-494.
Tripathi, R. P., N. Tewari, N. Dwivedi and V. K. Tiwari (2005). "Fighting tuberculosis: an old disease with new challenges." Med Res Rev 25(1): 93-131.
Verbon, A., R. A. Hartskeerl, A. Schuitema, A. H. Kolk, D. B. Young and R. Lathigra (1992). "The 14,000-molecular-weight antigen of Mycobacterium tuberculosis is related to the alpha-crystallin family of low-molecular-weight heat shock proteins." J Bacteriol 174(4): 1352-1359.

Wade, J. D., J. Bedford, R. C. Sheppard and G. W. Tregear (1991). "DBU as an N alpha-deprotecting reagent for the fluorenylmethoxycarbonyl group in continuous flow solid-phase peptide synthesis." Pept Res 4(3): 194-199.
Wang, S. S. (1973). "p-alkoxybenzyl alcohol resin and p-alkoxybenzyloxycarbonylhydrazide resin for solid phase synthesis of protected peptide fragments." J Am Chem Soc 95(4): 1328-1333.
Wilkinson, K. A., F. Hudecz, H. M. Vordermeier, J. Ivanyi and R. Wilkinson (1999). " Enhancement of the T cell response to a mycobacterial peptide by conjugation to synthetic branched polupeptide." Eur. J. Immunol. 29: 2788-2796.
Zhang, Y. (2005). "The magic bullets and tuberculosis drug targets." Annu Rev Pharmacol Toxicol 45: 529-564.

X. Köszönetnyilvánítás
Köszönetet mondok Dr. Perczel András tanszékvezet� egyetemi tanárnak, aki lehet�vé
tette, hogy az ELTE Szerves Kémiai Tanszéken dolgozhassak.
Dr. Hudecz Ferenc rektornak, az MTA-ELTE Peptidkémiai Kutatócsoport
vezet�jének, hogy lehet�vé tette, hogy a kutatócsoportban dolgozhassak.
Nagyon köszönöm Dr. B�sze Szilvia tudományos f�munkatársnak és Horváti Kata
tudományos segédmunkatársnak, témavezet�imnek, hogy segítségemre voltak a téma
kidolgozásában és a TDK dolgozat elkészítésében.
Dr. Szabó Nórának és Dávid Sándornak, hogy az Országos Korányi TBC és
Pulmonológiai Intézet területén található Bakteriológiai Laboratóriumban (Corden
International Magyarország Kft.) kísérleteink elvégzésében közrem�ködtek.
Köszönöm Dr. Medzihradszky Schweiger Hedvig tudományos f�munkatárs segítségét
a mikroanalitikai feladatok megoldásában.

Függelék

Függelék 1. ábra. a. A GTKPKG (T6) peptid tömegspektruma, b. a tisztított peptid analitikai RP-HPLC kromatogramja.
586.3
500 600 700 m/z
[M+H]+
Mmo (számított) =585,3
a.
0 5 10 15 20 25 30 35 40
0
50
100
150
200
250
A
t / min
b.
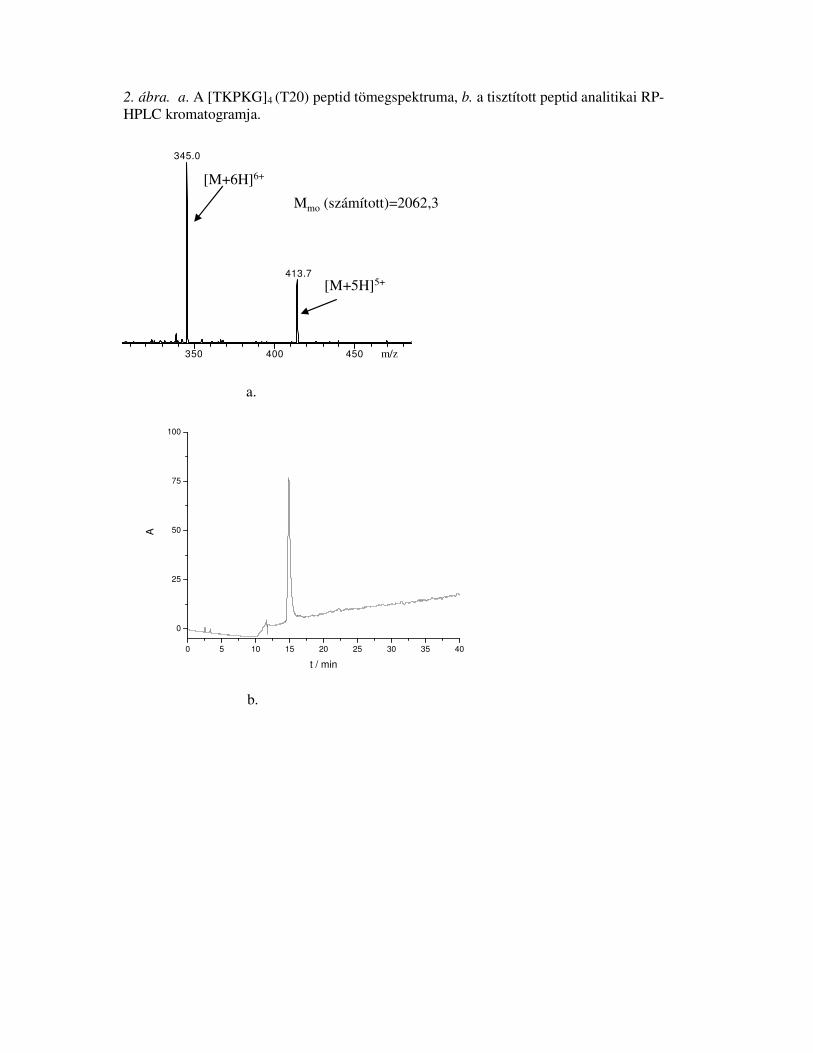
2. ábra. a. A [TKPKG]4 (T20) peptid tömegspektruma, b. a tisztított peptid analitikai RP-HPLC kromatogramja.
345.0
413.7
350 400 450 m/z
[M+6H]6+
[M+5H]5+
Mmo (számított)=2062,3
a.
0 5 10 15 20 25 30 35 40
0
25
50
75
100
A
t / min
b.

3. ábra. a. A 91SEFAYGSFVRTVSLPV106 (91-106) peptid tömegspektruma, b. a tisztított peptid analitikai RP-HPLC kromatogramja.
586.9
879.8
500 1000 1500 m/z
Mmo(számított) =1756,9
[M+3H]3+
[M+2H]2+
a.
5 10 15 20 25 30 35 400
50
100
150
200
250
300
350
400
A
t / min
b.

4.ábra. a. A 2-(izonikotinoil-hidrazono)ecetsav (INH-gli(ox)) és b. a 2-(izonikotinoil-hidrazino)ecetsav (INH-gli(red)) tömegspektruma.
180 190 200 210
194.0
195.0
m/z180 190 200 210
194.0
195.0
180 190 200 210180 190 200 210180 190 200 210
194.0
195.0
m/z
[M+H]+
Mmo (számított) =193,1
a.
180 190 200 210180 190 200 210
196.0
197.0
m/z180 190 200 210180 190 200 210
196.0
197.0
m/z
[M+H]+
Mmo (számított) = 195,1
b.
180 190 200 210180 190 200 210
196.0
197.0
m/z180 190 200 210180 190 200 210
196.0
197.0
m/z
[M+H]+
Mmo (számított) = 195,1
180 190 200 210180 190 200 210
196.0
197.0
m/z180 190 200 210180 190 200 210
196.0
197.0
m/z
[M+H]+
Mmo (számított) = 195,1
b.
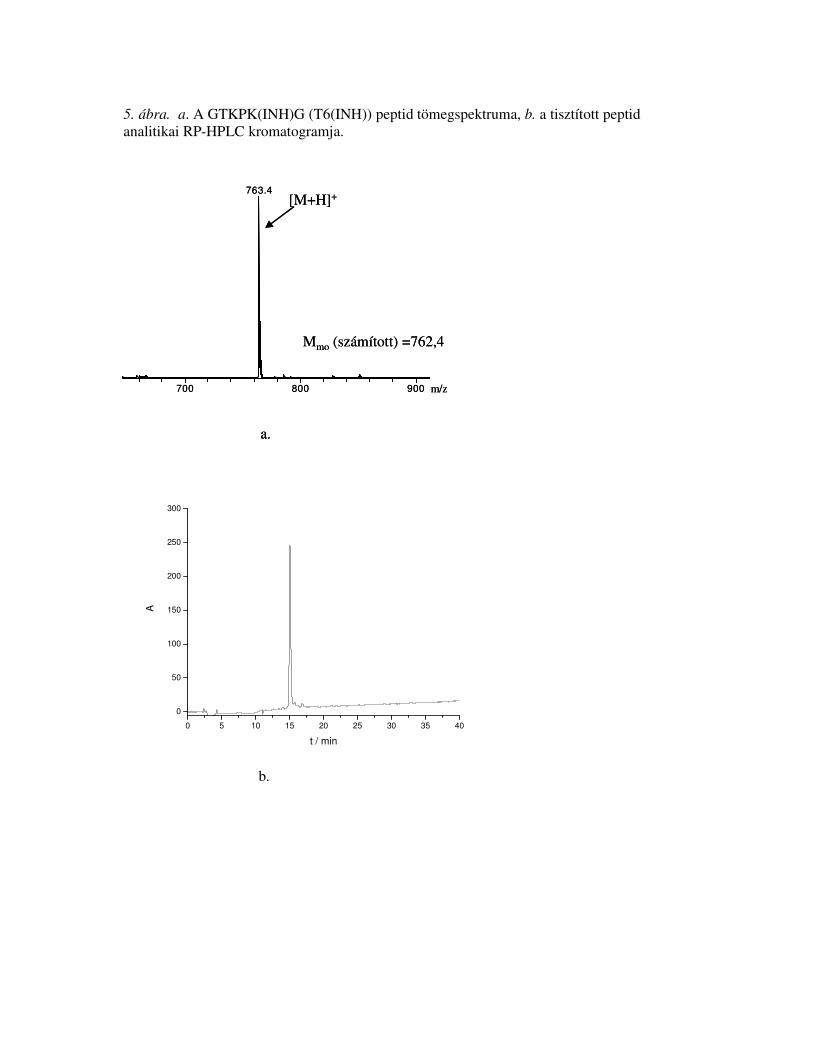
5. ábra. a. A GTKPK(INH)G (T6(INH)) peptid tömegspektruma, b. a tisztított peptid analitikai RP-HPLC kromatogramja.
763.4
700 800 900 m/z
[M+H]+
Mmo (számított) =762,4
a.
763.4
700 800 900 m/z
[M+H]+
Mmo (számított) =762,4
a.
0 5 10 15 20 25 30 35 40
0
50
100
150
200
250
300
A
t / min
b.
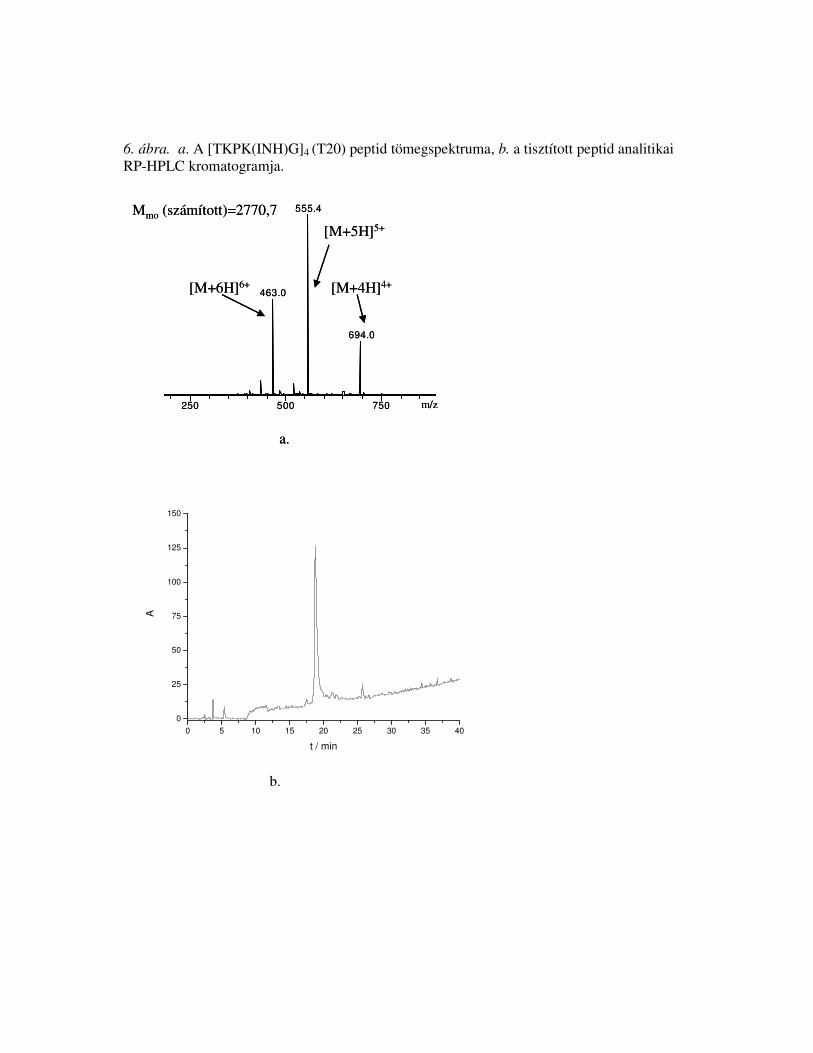
6. ábra. a. A [TKPK(INH)G]4 (T20) peptid tömegspektruma, b. a tisztított peptid analitikai RP-HPLC kromatogramja.
463.0
555.4
694.0
250 500 750 m/z
[M+5H]5+
[M+4H]4+[M+6H]6+
Mmo (számított)=2770,7
a.
463.0
555.4
694.0
250 500 750 m/z
[M+5H]5+
[M+4H]4+[M+6H]6+
Mmo (számított)=2770,7
a.
0 5 10 15 20 25 30 35 400
25
50
75
100
125
150
A
t / min
b.

7. ábra. a. Az INH-gli-91SEFAYGSFVRTVSLPV106 (INH-gli-91-106) peptid tömegspektruma, b. a tisztított peptid analitikai RP-HPLC kromatogramja.
645.9
968.4
600 700 800 900 1000
[M+2H]2+
[M+3H]3+
645.9
968.4
600 700 800 900 1000
[M+2H]2+
[M+3H]3+
Mmo(számított) = 1934,0
m/z
a.
645.9
968.4
600 700 800 900 1000
[M+2H]2+
[M+3H]3+
645.9
968.4
600 700 800 900 1000
[M+2H]2+
[M+3H]3+
Mmo(számított) = 1934,0
m/z
a.
5 10 15 20 25 30 35 400
50
100
150
200
250
300
350
A
t / min
b.
