maturation rate of mouse neutrophilic granulocytes: acceleration by retardation of proliferation,...
TRANSCRIPT

Maturation Rate of Mouse Neutrophilic Granulocytes: Acceleration by Retardation of Proliferation, but No Detectable
Influence from G-CSF or Stromal Cells
XIULI WANG,a HEGE FJERDINGSTAD,b INGER STRØM -GUNDERSEN,a HAAKON B. BENESTADa
aDepartment of Physiology, Institute of Basic Medical Sciences, University of Oslo, Oslo, Norway;bNycomed Drug Research, Oslo, Norway
Key Words. Hematopoietic cells · Bone marrow stromal cells · Proliferation · Maturation · G-CSF
ABSTRACT
Our purpose was to examine the possible influenceof stromal and humoral mediators on granulocyticmaturation rates. Sorted immature murine progenitor(Lin –Sca-1+) cells were cultured in peritoneal diffusionchambers (DCs) with or without a confluent layer ofirradiated bone marrow stromal cells on one of themicropore membrane walls. In other experiments, 10µg/kg/d recombinant G-CSF (rhG-CSF) was adminis-tered continuously into DC host mice through s.c.implanted osmotic minipumps. Operationally, matura-tion rate was assessed as the ratio between the numberof polymorphonuclear cells (PMN) and proliferativegranulocytes (PG) in short-term cultures, based on thedifferential cell counts, and supported by flow cyto-metric measurement of a granulocytic differentiationmarker; and by the emergence time of PMN in the DCs,obtained by extrapolation. Also, increased maturationis associated with increased cell density, as reflected by
the positioning of the granulocytes during centrifuga-tion in a discontinuous Percoll gradient. This method,as well as the conversion rate of 3H-thymidine labeledPG into the heavier non-PG maturational stages, werealso used as indicators of maturation rate. After five,six, and seven days of culture in the peritoneal cavity,DC cells were harvested. Their proliferative status,based on measurement of incorporated bromod-eoxyuridine, was determined, and their maturationrates were evaluated. Proliferation of immature granu-locytic progenitor cells was apparently inhibited bydirect contact with bone marrow stromal cells, andstimulated by G-CSF during the early stage of cultur-ing. However, the subsequent maturation rate, whichcould be accelerated by increasing culture cellularity,thus decreasing PG proliferation rate, was notdetectably influenced by either stromal cells or G-CSF.Stem Cells1999;17:253-264
STEM CELLS 1999;17:253-264
Correspondence: Dr. Haakon B. Benestad, Department of Physiology, IMBA, UiO, P.O. Box 1103 Blindern, N-0317 Oslo,Norway. Accepted for publication July 31, 1999. ©AlphaMed Press 1066-5099/99/$5/0
INTRODUCTION
The control of hematopoietic cell production isachieved by balancing the rates of differentiation (i.e.,lineage commitment), proliferation, maturation (i.e., termi-nal differentiation), and death of parenchymal bone marrowcells. Regulation occurs at the level of the structuredmicroenvironment (stroma), via cell-cell interactions andspecific cytokines.
As a representative of late-acting, lineage-specificcytokines, G-CSF can stimulate precursor cell proliferation[1, 2] and allegedly increase the maturation process of these
cells [3], thereby leading to a rapid and amplified neu-trophil release to blood, when needed to kill invadingmicrobes. Moreover, G-CSF also promotes cell survival bysuppressing programed cell death (apoptosis) [4] and worksas an activator of mature neutrophil function [5].
Granulocytopoiesis occurs in bone marrow in close asso-ciation with stromal cells, and it has been shown that thesecells not only inhibit the proliferation of primitive progenitorcells [6, 7] but also facilitate differentiation and protect theprogenitors from programed cell death through, for exam-ple, integrin-mediated adhesion between hematopoietic and

Wang, Fjerdingstad, Strøm-Gundersen et al. 254
stromal cells [8-10]. A prevention of the maturation (termi-nal differentiation) of granulocytes mediated by CD34mucin, an adhesion molecule expressed on progenitors, hasalso been hypothesized [11, 12].
Our present purpose was to examine possible matura-tional effects in a new manner; by confining granulocy-topoiesis to a culture chamber in vivo, where thecomplicating loss of more or less mature granulocytes tothe circulation (i.e., mobilization) is avoided.
Consequently, the proliferative kinetics and maturationprocess of murine, multipotent hematopoietic (Lin–Sca-1+)progenitor cells were investigated under stroma-contact andstroma-noncontact situations with our established diffusionchamber (DC) in vivo technique [13-15]. G-CSF wasdelivered by s.c. implanted osmotic minipumps. A bromod-eoxyuridine (BrdUrd) incorporation method [16-18] wasused to investigate the proliferative behavior of the culturedLin–Sca-1+ cells. The relative maturation rate of granulo-cytes was operationally defined and determined in severalways, based on differential cell counting, Percoll densitygradient centrifugation, and flux of 3H-thymidine (3H-TdR)labeled cells into the nonproliferative compartment. Thefindings did not support the notion that G-CSF or stromaregulates the maturation rate of granulocytes in vivo.
MATERIALS AND METHODS
Chemicals Dynabeads M-450 coated with sheep anti-rat IgG
(Dynal; Oslo, Norway); rat gamma globulin (Pierce;Rockford, IL); fluorescein isothiocyanate (FITC)-labeledrat IgG2a kappa (Pharmingen; San Diego, CA); FITC anti-mouse Ly-6A/E (Pharmingen); Lin monoclonal cocktail:Gr-1mAb, L3T4mAb, Mac-1mAb, CD2mAb,CD45R/B220mAb, TER-119/erythroid cells (Pharmingen);antibody against BrdUrd incorporated into DNA (anti-BrdUrd, Becton Dickinson; San Jose, CA); goat anti-mouseIg FITC (GAM-FITC, Becton Dickinson); FITC-conjugatedrat anti-mouse Ly-6G (Gr-1) monoclonal antibody ([mAb],Pharmingen); BrdUrd (Boehringer Mannheim GmbH;Mannheim, Germany); propidium iodide (Calbiochem; LaJolla, CA); Pronase (Boehringer Mannheim GmbH); Tween20 (Sigma; St. Louis, MO); pepsin (Sigma); Percoll(Pharmacia; Uppsala, Sweden); 3H-TdR, (Amersham;Buckinghamshire, UK); recombinant human G-CSF ([rhG-CSF], Chugai Pharmaceutical Co. Ltd.; Tokyo, Japan; kindgift from Dr. K.J. Mori, Faculty of Science, NiigataUniversity, Japan); horse serum, Bio-Whittaker;Walkersville, MD); fetal bovine serum ([FBS], HycloneLaboratories Inc.; Logan, UT); fetal calf serum ([FCS],GIBCO BRL; Paisley, Scotland); bovine serum albumin
([BSA], Sigma); minimal essential medium (MEM), AlphaMedium (GIBCO BRL); Fisher’s medium (LifeTechnologies; Paisley, Scotland); RPMI 1640 (Bio-Whittaker); HEPES (Sigma); phosphate buffered saline([PBS], GIBCO BRL); ampicillin (Doctacillin™, STR;Södertälje, Sweden); sodium azide (Sigma); hydrocortisone(Sigma); hydrochloric acid ([HCl], May & Baker Ltd.;Dagenham, UK).
MiceAdult inbred C57BL/6J/OlaHsd females, 5 to 10 weeks
old, were used as donors of Lin–Sca-1+ bone marrow cellsand syngeneic stromal cells. NMRI (B & K Universal AS;Sollentuna, Sweden) or ICR/OlaHsd females, five to nineweeks old, of randomly bred strains, were employed ashosts of diffusion chambers. In some experiments, NMRImice were used as donors of bone marrow parenchymalcells and allogeneic bone marrow stromal cells. Theexperimental protocol was approved by the national animalexperimentation committee.
Establishment of Stromal Layers in DC Cultures In VitroFirst, 30 × 106 cells /ml from femora of C57BL/6J mice
were inoculated into micropore DCs (0.25 ml/DC), in the“Fisher-HEPES-antibiotics-horse serum” (FHabHS) medium[19], containing 1 µmol/l hydrocortisone as well. The DCswere incubated for 2 h to allow adherence of cells to one ofthe two micropore membrane walls of the DC. Then 6-mlmedium was pipetted into each 35-mm diameter culture dish,which supported two DCs. At the same time, 0.25 ml mediumwithout bone marrow cells was inoculated into control DCs.All the DCs were cultured under the same conditions (37°Cwith 5% carbon dioxide in air and saturated water vapor) fortwo weeks, and the medium in the dishes was replaced afterone week.
These established stromal DCs and stroma-free DCswere irradiated with 20 Gy from a Stabilipan-x-ray machine(Siemens; Erlangen, Germany) at a dose rate of 1 Gy perminute (220 kV, 20 mA, added filtration 0.5 mm Cu) beforeinoculation of the parenchymal hematopoietic cells. Insome experiments, host mice were exposed to 5 Gy x-rayirradiation before use.
Separation of Lin–Sca-1+ CellsLin–Sca-1+ cells were isolated from crushed femora, tib-
iae, and humeri of C57BL/6J mice according to the methodsdescribed [20]. Briefly, bone marrow cells (4 × 108/ml) wereincubated with an antibody cocktail, specifically reactive withthe various differentiated bone marrow cell lineages (Lin)(Gr-1; L3T4; Mac-1; CD2; CD45R/B220; TER-119/ery-throid cells). Then labeled cells (Lin+) were removed by

255 Maturation Rate of Mouse Neutrophilic Granulocytes
magnetic beads and Lin– cells obtained from the supernatant.Lin– cells were treated with FITC-conjugated antimouseLy6A/E antibody for 30 min. Finally, the positive “stem cellantigen” (Sca) cells were sorted with a Coulter Epics EliteCell Sorter (Coulter Electronics; Hialeah, FL).
Culture In Vivo Fresh Lin–Sca-1+ cells were plated at 1,500 to 2,500/DC
onto either the established and irradiated, once-washedsyngeneic bone marrow stromal layers, or into the stromalcell-free DCs. The DCs were then implanted i.p.. After five-,six-, and seven-day in vivo cultures, the DCs were removedfrom host mice and shaken in FHabHS medium containing0.35% Pronase at 37°C for 60 min. After washing out eachDC three times, the cell suspension pooled from one group offour DCs was weighed, its particle concentration was countedelectronically, and cell samples were retrieved for cytospinpreparation and differential cell counting. The numbers ofproliferative granulocytes (PG), nonproliferative granulocytes(NPG), polymorphonuclear cells (PMN), macrophage-likecells (MA), and lymphocytes (LY) were recorded accordingto morphological criteria established previously [21].
In density separation experiments, normal bone marrowcells (300,000/DC) were inoculated on established and irra-diated (20 Gy x-rays) bone marrow stroma in DCs or in stan-dard DCs without stromal cells. The DCs were implantedinto hosts and cultured for five days. Each host mouse wasinjected i.p. with 10 µCi (37 kBq) 3H-TdR in 2 ml 0.9% NaCl24 h before the removal of DCs. Three and 18 h after injec-tion of 3H-TdR, 100 µg/kg rhG-CSF in 0.9% NaCl with 0.1%FCS was injected i.p. into the test host mice. The controlsreceived injections of 0.9% NaCl with 0.1% FCS. The har-vested DC cells (pooled from 10 DCs in each group) werepositioned into four different layers during centrifugation ina discontinuous Percoll gradient (see below). Cells from eachlayer were enumerated (see above), and their incorporatedradioactivity measured in a b counter.
Analysis of Proliferative StatusTo determine the proliferative status of the harvested
cells, 0.35% Pronase containing FHabHS medium withBrdUrd was used to harvest the DC cells. To ensure optimumBrdUrd labeling of DNA, 10–4 mol/l was adopted as the high-est nontoxic dose, after dose titration experiments. TheBrdUrd treated cells were resuspended in 1 ml PBS, fixed in9 ml absolute ethanol, and stored at –20°C until flow cyto-metric analysis could be performed. These fixed cells wereprocessed according to the method described [17, 18], butslightly modified as follows: the samples were washed thor-oughly with cold PBS, and 2 ml fresh pepsin-HCl solution(0.2% pepsin in 2N HCl) was added to each cell button. After
suspension, these fixed cells were incubated for 60 min in thedark at room temperature. After washing three times, a mix-ture of 30 µl 0.5% Tween 20 with 0.5% BSA solution in PBSand 20 µl anti-BrdUrd mAb was added to each tube. After a45-min incubation at room temperature and one washing,100 µl 1:40 GAM-FITC were pipetted into the tube. Thiswas followed by an incubation of 30 min, one washing, andthe addition of 1 ml propidium iodide (2.5 µg/ml) to eachtube. The samples were ready 50 min later to be analyzedwith flow cytometry (FACScan, Becton Dickinson).
Analysis of Gr-1 Antigen Expression on GranulocytesDC cells were collected after treatment with 0.35%
Pronase for 60 min. Cells were labeled by Gr-1 antibody asdescribed [22], but slightly modified. In short, 0.1 µg fluo-rescein-conjugated rat antimouse Gr-1/myeloid differentia-tion antibody was added to a 50 µl cell suspension andincubated for 45 min on ice. After washing twice with PBScontaining 1% FCS and 0.1% sodium azide, the cells wereresuspended in 0.5 ml PBS for fluorescence-activated cellscanning (FACS) analysis.
In Vivo Delivery of rhG-CSFOsmotic minipumps (Model 2002, Alzet, Alza; Palo
Alto, CA), loaded with rhG-CSF in Fisher’s medium withHEPES (10 mmol/l, pH ~ (7.35), 0.1% BSA and ampicillin(200 µg/ml), were implanted s.c. on the back of NMRI orICR host mice, so that 10 µg/kg rhG-CSF were deliveredevery day for the whole culture period of seven days. Thisdose has been proved effective in humans [23] and in mice[2] for stimulating myeloid proliferation in bone marrowand inducing neutrophilia in blood. Each experimentemployed two groups of host mice, one with minipumpsloaded with rhG-CSF, the other with minipumps deliveringsolvent solution only, or just control mice withoutminipumps implanted. Comparison between these two con-trols did not indicate any systematic differences concerningthe relevant variables. The DCs, containing Lin–Sca-1+
cells with a stromal layer, were implanted into the peri-toneal cavity, two chambers per host mouse. The DC cellswere harvested and analyzed as described above.
Assessment of Maturation Rate of Granulocyte Precursors byDensity Gradient Separation
A modification of the method described by RolstadandBenestad[24] was applied to the separation of DC cells accord-ing to cell densities. Percoll was adjusted to 285 mOsM/l with10 × PBS, and RPMI 1640 + 10% FCS to 290 mOsM/l withsterile water. Osmolarity-corrected Percoll and medium werethen mixed in different proportions to create four differentdensities: F1 containing 37.8% v/v Percoll (corresponding to a

Wang, Fjerdingstad, Strøm-Gundersen et al. 256
density relative to water of 1.053); F2: 47.9% (1.065); F3:56.3% (1.075), and F4: 62.2% (1.082). Three milliliters of eachfraction were layered carefully on top of each other as depictedin Figure 1, with 1 ml of osmolarity-corrected mediumcontaining 10 – 15 × 106 cells retrieved from DCs at the top, in15-ml Falcon centrifuge tubes, and spun at 400 g for 30 min atroom temperature. The DC cells in each layer were washed outof the Percoll before being used, and are hereafter termed DCcell fraction 1-4 (F1-F4), so that, for example, F2 cells are thecells just above the second density layer.
StatisticsA two-sided, nonparametric Wilcoxon pair test was
used in comparisons between the test and control DCcultures. Confidence intervals of 95% for the median valueswere calculated with a nonparametric method (MINITABcomputer program, Minitab; State College, PA).
RESULTS
Accelerated Granulocyte Maturation by Retardation ofProliferation Rate
After the density separation of the chamber (DC) cells,the majority of F1 (low density) cells (more than 80%) weremorphologically MA (modal diameter in isotonic solution 11µm), while the majority of F4 cells (about 80%) were NPG(modal diameter 7.3 µm). F3 contained the highest number ofcells and radioactivity (from 3H-TdR injected the day before).A representative differential count is 37% PG, 58% NPG, 4%MA, and 1% LY (modal diameter 9 µm). With increasedgranulocyte maturity the cells were distributed to the higher-density fractions (Figs. 1 and 2), as found earlier for rat bonemarrow cells [24]. DCs containing 300,000 bone marrowcells (“high cellularity DCs”) or 30,000 bone marrow cells(“low cellularity DCs”) were implanted i.p. into normal or 5 Gyirradiated host mice and harvested after five days. The cells
Figure 1. Sketch of Percoll density separated cells, granulocyte density increasing with increasing maturation. The majority (>80%) of F1cells are macrophage-like (MA) cells. Among the F4 cells, nonproliferative granulocytes (NPG) predominate. More granulocytes altogetherappear in F3 than F2. See text and Figure 2 for more details. PMN: polymorphonuclear cells; PG: proliferative granulocytes.

257 Maturation Rate of Mouse Neutrophilic Granulocytes
were separated into the four density fractions (F1-F4). Alarger portion of the 3H-TdR label and of the granulocytesappeared in F3 than in lighter fractions, when high-cellularitycultures were compared with low-cellularity cultures (Fig. 3).The relative expansion of the cell inocula (medians 30-, 25-,and 8-fold for 3 × 104 cells in 5 Gy irradiated hosts, 3 × 104
cells in normal hosts, and 30 × 104 cells in normal hosts,respectively) (Fig. 3) had been largest in the low-cellularityDCs. Accordingly, the incorporation of 3H-TdR (2.8, 1.4, and3.9 × 104 cpm/DC, respectively) had also been most intense,on a per cell basis, in the low-cellularity cultures.Consequently, proliferation had been most accelerated andmaturation—judged by the density distribution of both gran-ulocytes and 3H activity—most retarded when the DCs hadbeen carried by irradiated mice (Fig. 3).
Proliferation and Maturation of Lin –Sca-1+ Cells on Bone Marrow Stromal Cells
A good correlation was found between Gr-1+ cells mea-sured with flow cytometry and concentration of granulocytes
Figure 2. More mature cells move to higher densities during Percollgradient centrifugation. 300,000 bone marrow cells were cultured inin vivo diffusion chambers (DCs) with or without irradiated stromalcells (20 Gy x-rays) for five days. Each host mouse was injected i.p.with 10 µCi 3H-thymidine in 2 ml 0.9% NaCl 24 h before the removalof DCs. Harvested DC cells (pooled from 10 DCs in each group)were separated by density-gradient centrifugation. The distribution of granulocyte maturation stages and 3H activity to the four densityfractions is shown. Two replicate and independent experiments gavesimilar results. NPG-PMN are metamyelocytes and band-nucleatedgranulocytes.
Figure 3. Maturation rate of bone marrow cells in in vivo DCs withhigh- or low-culture cellularity. Relative population expansion: 3 × 104 cells in 5 Gy irradiated hosts > 3 × 104 cells in normal hosts> 30 × 104 cells in normal hosts. Concerning the maturation rate,assessed as the distribution of granulocytes or 3H activity to densityfractions F2 (with the less mature granulocytes) and F3 (with themore mature granulocytes), it increased with decreasing expansion ofthe cultured cell populations. A replicate experiment gave similarresults. See Figures 1 and 2 for further explanations.

Wang, Fjerdingstad, Strøm-Gundersen et al. 258
determined with differential counts, and between fluores-cence intensity of Gr-1 expression and concentration of PMN(Fig. 4).
Our DC harvesting procedure allowed virtually com-plete retrieval of cultured parenchymal cells from the DCs,leaving a substantial portion of the stromal cells adherent tothe micropore membranes.
The proliferative state of the irradiated (20 Gy x-ray)bone marrow stromal cells was also checked. DCs withstromal cells only, but without inoculated stem cells, wereharvested and analyzed. Only about 1.5% (mean value fromthree experiments) of bone marrow stromal cells wereapparently in S-phase of the cell cycle after five to sevendays’ in vivo culture.
To evaluate the proliferative kinetics of the culturedLin–Sca-1+ cells, the number of cells that incorporated
BrdUrd in stroma-contact and stroma-noncontact DCs wasrecorded on days 5, 6, and 7. Throughout the period ofobservation, 14% to 27% of S-phase cells (medians) wererecorded for stroma-contact and stroma-noncontact groupsof DCs. However, proliferating granulocytes appearedearlier in stroma-noncontact than in stroma-contact cultures(Table 1 versus Fig. 5).
Concerning granulocyte maturation rate, no detectabledifferences were found between the stroma-contact andstroma-noncontact cultures, as assessed with the PMN:PGratios (Fig. 5). The appearance of PMN in stroma-contactcultures was nonsignificantly delayed, 4.4 days versus 3.3 days for stroma-noncontact cultures (Fig. 6), the
Figure 4. Correlation between Gr-1 antigen expression measuredwith flow cytometry and granulocyte concentration (PG + NPG inupper panel, PMN in lower) determined with differential counting.The cell population retrieved after in vivo culture of Lin-Sca-1+ cellswith or without stroma was analyzed with both methods, the flowcytometry utilizing an FITC-labeled Gr-1 antibody. Regression lines,regression coefficients (r), and the probabilities that r ≠ 0 are given.
Table 1. Numbers of DNA-synthesizing cells and granulocytes (Gr-1+) harvested per Lin–Sca-1+ cell inoculated in diffusion chamberculture with or without a bone marrow stromal layer (SL)
Culture time (d) S-phase cells Gr-1+ cells
+ SL – SL + SL – SL
5 43 46 90 210
(22-88) (38-167) (62-667) (79-748)
6 84 68 300 333
(33-143) (29-122) (198-1,440) (113-971)
7 92 71 684 284
(43-165) (49-103) (157-1,908) (227-740)
Medians and their 95% confidence intervals (for n ≥ 6) and ranges (forn < 6) are given. n is the number of independent experiments, 11 forS-phase data and five for Gr-1 data.
Figure 5. Maturation rate of Lin–Sca-1+ cells cultured with or with-out contact with irradiated bone marrow stroma in vivo. At eachtime point, the number of PG and PMN were determined, based ondifferential counting. Medians and their 95% confidence intervalsfrom 10 replicate experiments for both groups are given.

259 Maturation Rate of Mouse Neutrophilic Granulocytes
retardation possibly being caused by the delayed wave ofproliferating PG in the stromal DCs. Finally, the densitycentrifugation experiments did not reveal any maturationaleffect of the stroma on the later stages of granulocytedevelopment (Fig. 7).
Proliferation and Maturation of Lin –Sca-1+ Cells in Stroma-Contact Cultures Given Supplementary rhG-CSF
Continuously delivered rhG-CSF, from s.c. osmoticminipumps, gave rise to a higher percentage of Gr-1+ cellsin the bone marrow of test than control, solvent infusedmice (91% versus 82%). The proliferative activity in DCsbetween days 5 and 7 was high and was increased slightly
Stroma-noncontact Stroma-noncontact
3.3 days
10
5
0
3 4 5 6 7
800
600
400
200
0
5 6 7
10
5
0
4 5 6 7
10
5
0
4 5 6 7
800
600
400
200
0
5 6 7
800
600
400
200
0
5 6 7
4.0 days
4.4 days
Stroma-contactStroma-contact
Stroma-contact + G-CSF Stroma-contact + G-CSF
Culture time (d)
Nor
mal
ized
PM
N n
umbe
r
Gra
nulo
cyte
s re
trie
ved
per L
in– Sc
a-1+ c
ell i
nocu
late
d
Figure 6. Granulocytopoiesis from sorted Lin–Sca-1+ cells cultured under in vivo conditions. Absolute values (with their 95% confidenceintervals) of all granulocytes (PG + NPG) retrieved after five, six, and seven days’ culturing are given (left-hand panels). Normalized PMNvalues (values for stroma-contact group on day 5 = 1.0 in each experiment) are also presented (right-hand panels). Extended linear regressionlines (with their 95% confidence belts) gave the times of appearance of PMN in the cultures. n = 11 five to seven-day experiments for stroma-noncontact and stroma-contact groups. n = 6 for stroma-contact with or without G-CSF groups.
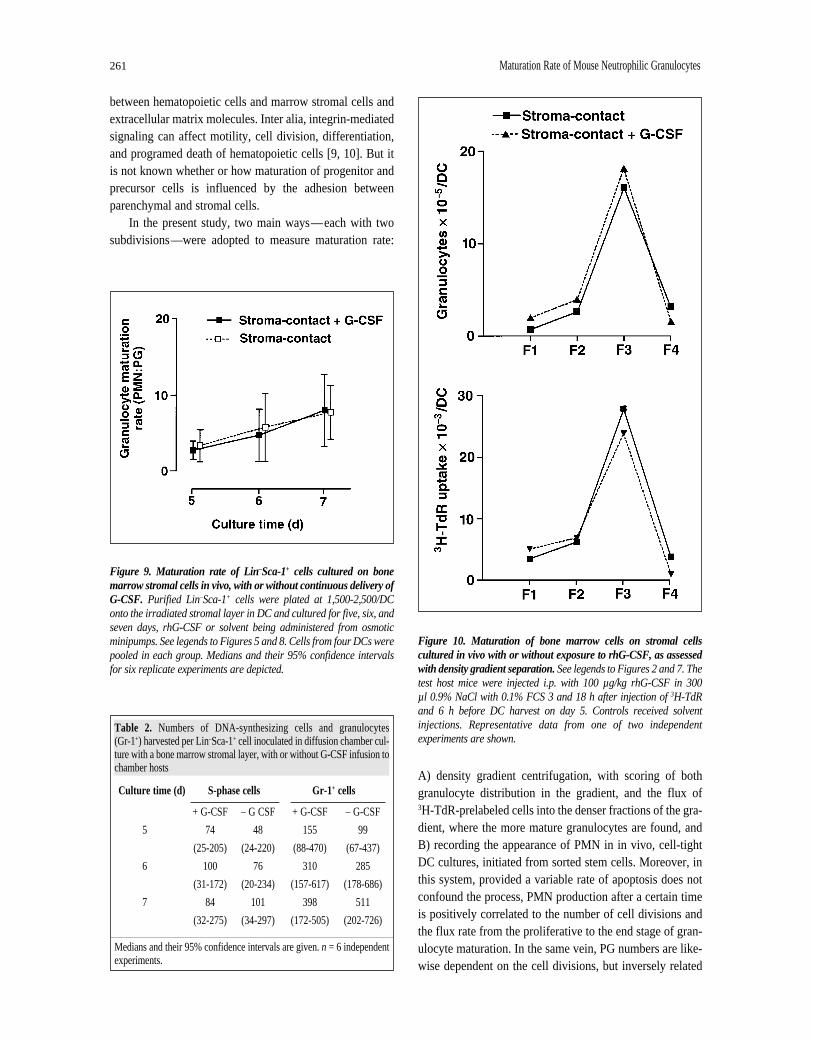
on day 5 by the rhG-CSF infusion (Table 2, Fig. 8). PMNappeared in the DC cultures at almost the same time in testas in control—4.0 days with G-CSF treatment versus 4.4days for the control (Fig. 6). The rhG-CSF had nodetectable influence on the maturation rate of granulocytesas judged by the occurrence of Gr-1+ cells and the PMN:PGratios either (Table 2, Fig. 9). Likewise, the density cen-trifugation approach could not substantiate any significantmaturational effect (Fig. 10).
DISCUSSION
Long-term stroma-based culturing [25, 26] has shownthat sustained hematopoiesis depends on close proximity

Wang, Fjerdingstad, Strøm-Gundersen et al. 260
Figure 7. Maturation of bone marrow cells cultured in vivo with or without stromal cells.300,000 bone marrow cells were cultured in eachDC on micropore membrane or on a layer of irradiated (20 Gy x-rays) bone marrow stromal cells for five days. Harvested cells, which had beenlabeled with 3H-thymidine one day before retrieval, were subjected to Percoll density gradient centrifugation. See legend to Figure 2. Tworeplicate experiments gave similar results.
Figure 8. BrdUrd incorporation profiles of five-day cultures. Freshly sorted Lin–Sca-1+ cells were cultured on bone marrow stromal cells in DCs in the peritoneal cavity of host mice. An osmotic minipump loaded with rhG-CSF was implanted s.c. into each host, so that 10 µg/kg rhG-CSF (A) or solvent (B) was delivered every day. DC cells were labeled with BrdUrd in vitro before cell retrieval from the chambers, and the cells which incorporated BrdUrd (enclosed in rectangles), were considered S-phase cells. Five independent, replicate experiments gavesimilar results.
261 Maturation Rate of Mouse Neutrophilic Granulocytes
between hematopoietic cells and marrow stromal cells andextracellular matrix molecules. Inter alia, integrin-mediatedsignaling can affect motility, cell division, differentiation,and programed death of hematopoietic cells [9, 10]. But itis not known whether or how maturation of progenitor andprecursor cells is influenced by the adhesion betweenparenchymal and stromal cells.
In the present study, two main ways—each with twosubdivisions—were adopted to measure maturation rate:
Table 2. Numbers of DNA-synthesizing cells and granulocytes (Gr-1+) harvested per Lin–Sca-1+ cell inoculated in diffusion chamber cul-ture with a bone marrow stromal layer, with or without G-CSF infusion tochamber hosts
Culture time (d) S-phase cells Gr-1+ cells
+ G-CSF – G CSF + G-CSF – G-CSF
5 74 48 155 99
(25-205) (24-220) (88-470) (67-437)
6 100 76 310 285
(31-172) (20-234) (157-617) (178-686)
7 84 101 398 511
(32-275) (34-297) (172-505) (202-726)
Medians and their 95% confidence intervals are given. n = 6 independentexperiments.
A) density gradient centrifugation, with scoring of bothgranulocyte distribution in the gradient, and the flux of 3H-TdR-prelabeled cells into the denser fractions of the gra-dient, where the more mature granulocytes are found, andB) recording the appearance of PMN in in vivo, cell-tightDC cultures, initiated from sorted stem cells. Moreover, inthis system, provided a variable rate of apoptosis does notconfound the process, PMN production after a certain timeis positively correlated to the number of cell divisions andthe flux rate from the proliferative to the end stage of gran-ulocyte maturation. In the same vein, PG numbers are like-wise dependent on the cell divisions, but inversely related
Figure 9. Maturation rate of Lin–Sca-1+ cells cultured on bonemarrow stromal cells in vivo, with or without continuous delivery ofG-CSF. Purified Lin–Sca-1+ cells were plated at 1,500-2,500/DConto the irradiated stromal layer in DC and cultured for five, six, andseven days, rhG-CSF or solvent being administered from osmoticminipumps. See legends to Figures 5 and 8. Cells from four DCs werepooled in each group. Medians and their 95% confidence intervalsfor six replicate experiments are depicted.
Figure 10. Maturation of bone marrow cells on stromal cellscultured in vivo with or without exposure to rhG-CSF, as assessedwith density gradient separation. See legends to Figures 2 and 7. Thetest host mice were injected i.p. with 100 µg/kg rhG-CSF in 300 µl 0.9% NaCl with 0.1% FCS 3 and 18 h after injection of 3H-TdRand 6 h before DC harvest on day 5. Controls received solventinjections. Representative data from one of two independentexperiments are shown.

Wang, Fjerdingstad, Strøm-Gundersen et al. 262
to the loss by further maturation. Consequently, we alsochose the PMN:PG ratio as an indicator of maturation rate.An even increase of the PMN:PG ratios from day 5 to day7, with no detectable differences between test and controlvalues, would strongly suggest both similar maturation anddeath (apoptosis) rates in the tests and controls.
Our present studies suggested that stroma-contact cul-turing retarded the initiation of development from purifiedLin–Sca-1+ cells toward PMN, perhaps for one day, com-pared with stroma-noncontact cells. But when it started, thematuration of granulocytes proceeded at approximately thesame rate with and without stromal contact.
G -CSF, initially identified as a differentiation-induc-ing factor for a leukemia cell line [27], is certainly a gran-ulopoietin with an important stimulatory role for baselinegranulocytopoiesis, as demonstrated with gene knock-outmice [28, 29]. Its granulocyte differentiation function(via a STAT-3 signal pathway) can apparently occur atlower concentrations of G-CSF than needed to accelerateproliferation [30]. G-CSF can increase the production ofmature granulocytes [23, 31] and effect mobilization toblood of granulocytes and clonogenic hematopoieticprogenitors [1, 31, 32]. Such findings suggest that G-CSFmight also accelerate—or at least affect—maturation ofgranulocytes. However, the present experiments couldnot support this hypothesis. When the high proliferationrate of granulocyte precursors on a stromal cell layer inperitoneal DC was marginally increased by a continuouss.c. infusion of rhG-CSF, we could not find any changedmaturation rate. This was in contrast to the accelerationof maturation rate that occurred when proliferation ratewas slowed down by utilizing high-cellularity DC cul-tures in nonstimulated host mice. These latter results arein line with our previously published results from per-turbations of proliferation rate of mouse granulocyte pre-cursors [13]. A possible mechanism is that markedproliferation inhibition could result in one (or more)skipped cell divisions, and the cells enter the nonprolif-erative compartment instead. This would increase thematuration rate.
The discrepancies between the results of Lord et al.[1, 3] and ours are not easily explained. In their experi-ments, after labeling with 3H-TdR, labeled neutrophilsappeared in the circulation of control mice after 24 h andpeaked (about 15% labeling index) after 72 to 96 h. In G-CSF-treated mice, label appeared in blood after about 12 hand peaked (about 25% labeling index) after 24 h. This ear-lier appearance could be due to acceleration of maturation,release to blood (mobilization), or both. In our experience,it may not be easy to distinguish between segmented andband-nucleated rodent neutrophils in liquid emulsion
radioautographs. Moreover, it has been shown that G-CSFcan certainly mobilize neutrophils from the bone marrow[3, 33], whereas an effect on maturation rate per se has notbeen unequivocally demonstrated by others. Lord et al.used a higher dose of rhG-CSF (250 versus 10 µg/kg × d),given as two daily injections, whereas we provided a con-stant infusion rate with osmotic minipumps. Their dose waschosen to give a maximal blood leukocytosis, but theyreported that 10 µg/kg × d gave a similar degree of marrowstimulation as the higher dose. The high G-CSF dose woulddecrease marrow cellularity by about 50% after 70 h, due toa redistribution of hematopoiesis between the bone marrowand the spleen [34]. They studied granulocytopoiesis insitu, whereas granulocyte formation in our experimentstook place in a confined, ectopic, and artificial microenvi-ronment. It may also be necessary to take into account thatwe have used different strains of mice, which may react dif-ferently to G-CSF.
One further finding by Lord et al.does indeed suggestthat G-CSF accelerates maturation through the myelocytestage—that is, the lack of increase in the proportion ofmyelocytes, morphologically defined, in the marrow—despite their more rapid proliferation after G-CSF treat-ment. However, this might also be explained by anincreased rate of apoptosis, since there are instances whereenhanced cell cycling is associated with enhancedapoptosis [35, 36]. In any case, more experiments need tobe done to clarify this matter.
Our results are also in line with the observation that G-CSF receptor signals are not required for terminal differ-entiation, that is, maturation, since no accumulation ofimmature granulocytic cells was observed in the bone mar-row of G-CSF receptor-deficient mice [37]. It would appearthat a rather constant maturation rate of granulocytes ismaintained, possibly at a genetically fixed level, under mostphysiological circumstances [38], since actual maturationprograms are not dependent on the particular hematopoieticregulator used as the proliferative stimulus [39]. Incombination with possibilities to regulate, for example,proliferative rate, mobilization to blood, and apoptosis, afixed maturation rate would still provide enough flexibilityto allow granulocytopoietic adaptation to the challenges ofordinary inflammatory states.
ACKNOWLEDGMENTS
We thank Arne Bøyumand Dagfinn Løvhaugfor criti-cally reading and Anne Berit Morrowfor reviewing themanuscript. This research was supported by grants fromAnders Jahre’s Foundation for the Promotion of Science,The Norwegian Cancer Society, and the NorwegianResearch Council.

263 Maturation Rate of Mouse Neutrophilic Granulocytes
1 Lord BI, Molineux G, Pojda Z et al. Myeloid cell kinetics inmice treated with recombinant interleukin-3, granulocytecolony-stimulating factor (CSF), or granulocyte-macrophageCSF in vivo. Blood 1991;77:2154-2159.
2 Pojda Z, Molineux G, Dexter TM. Hemopoietic effects ofshort-term in vivo treatment of mice with various doses ofrhG-CSF. Exp Hematol 1990;18:27-31.
3 Lord BI. Myeloid cell kinetics in response to haemopoieticgrowth factors. Baillieres Clin Haematol 1992;5:533-550.
4 Williams GT, Smith CA, Spooncer E et al. Haemopoieticcolony stimulating factors promote cell survival by suppressingapoptosis. Nature 1990;343:76-79.
5 Nicola NA. Granulocyte colony-stimulating factor. ImmunolSer 1990;49:77-109.
6 Verfaillie CM. Direct contact between human primitivehematopoietic progenitors and bone marrow stroma is notrequired for long-term in vitro hematopoiesis. Blood1992;79:2821-2826.
7 Hurley RW, McCarthy JB, Verfaillie CM. Direct adhesion tobone marrow stroma via fibronectin receptors inhibitshematopoietic progenitor proliferation. J Clin Invest1995;96:511-519.
8 Foley KP, McArthur GA, Queva C et al. Targeted disruptionof the MYC antagonist MAD1 inhibits cell cycle exit duringgranulocyte differentiation. EMBO J 1998;17:774-785.
9 Howe A, Aplin AE, Alahari SK et al. Integrin signaling andcell growth control. Curr Opin Cell Biol 1998;10:220-231.
10 Giancotti FG. Integrin signaling: specificity and control ofcell survival and cell cycle progression. Curr Opin Cell Biol1997;9:691-700.
11 Cheng J, Baumhueter S, Cacalano G et al. Hematopoieticdefects in mice lacking the sialomucin CD34. Blood1996;87:479-490.
12 Fackler MJ, Krause DS, Smith OM et al. Full-length but nottruncated CD34 inhibits hematopoietic cell differentiation ofM1 cells. Blood 1995;85:3040-3047.
13 Benestad HB, Rytömaa T. Regulation of maturation rate ofmouse granulocytes. Cell Tissue Kinet 1977;10:461-468.
14 Benestad HB, Reikvam A. Diffusion chamber culturing ofhaematopoietic cells: methodological investigation andimprovement of the technique. Exp Hematol 1975;3:249-260.
15 Benestad HB, Toogood EO. Diffusion chamber (DC)culturing of haemopoietic cells: a reliable assay system forregulators of proliferation? Exp Hematol 1982;10:161-171.
16 Dolbeare F, Gratzner H, Pallavicini MG et al. Flow cytometricmeasurement of total DNA content and incorporated bromod-eoxyuridine. Proc Natl Acad Sci USA 1983;80:5573-5577.
17 van Erp PEJ, Brons PPT, Boezeman JBM et al. A rapid flowcytometric method for bivariate bromodeoxyuridine/DNAanalysis using simultaneous proteolytic enzyme digestion andacid denaturation. Cytometry 1988;9:627-630.
REFERENCES
18 Kirkhus B, Fraas Clausen OP. Cell kinetics in mouse epider-mis studied by bivariate DNA/bromodeoxyuridine andDNA/keratin flow cytometry. Cytometry 1990;11:253-260.
19 Breivik H. Haematopoietic stem cell content of murine bonemarrow, spleen, and blood. Limiting dilution analysis ofdiffusion chamber cultures. J Cell Physiol 1971;78:73-78.
20 Spangrude GJ, Scollay R. A simplified method for enrich-ment of mouse hematopoietic stem cells. Exp Hematol1990;18:920-926.
21 Benestad HB. Formation of granulocytes and macrophages indiffusion chambers of mouse blood leucocytes. Scand J Haematol 1970;7:279-288.
22 Tapley P, Shevde NK, Schweitzer PA et al. Increased G-CSFresponsiveness of bone marrow cells from hematopoietic cellphosphatase deficient viable motheaten mice. Exp Hematol1997;25:122-131.
23 Campbell LJ, Maher DW, Tay DLM et al. Marrow prolifera-tion and the appearance of giant neutrophils in response torecombinant human granulocyte colony stimulating factor(rhG-CSF). Br J Haematol 1992;80:298-304.
24 Rolstad B, Benestad HB. Spontaneous alloreactivity ofnatural killer (NK) and lymphokine-activated killer (LAK)cells from athymic rats against normal haemic cells. NK cellsstimulate syngeneic but inhibit allogeneic haemopoiesis.Immunology 1991;74:86-93.
25 Klein G. The extracellular matrix of the hematopoieticmicroenvironment. Experientia 1995;51:914-926.
26 Dexter TM, Coutinho LH, Spooncer E et al. Stromal cells inhaemopoiesis. Ciba Found Symp 1990;148:76-86.
27 Nicola NA, Metcalf D, Matsumoto M et al. Purification of afactor inducing differentiation in murine myelomonocyticleukemia cells. Identification as granulocyte colony-stimulat-ing factor. J Biol Chem 1983;258:9017-9023.
28 Lieschke GJ, Grail D, Hodgson G et al. Mice lacking granu-locyte colony-stimulating factor have chronic neutropenia,granulocyte and macrophage progenitor cell deficiency, andimpaired neutrophil mobilization. Blood 1994;84:1737-1746.
29 Liu F, Wu HY, Wesselschmidt R et al. Impaired productionand increased apoptosis of neutrophils in granulocyte colony-stimulating factor receptor-deficient mice. Immunity1996;5:491-501.
30 Ward AC, Hermans MHA, van Aesch YM et al. Tyrosine-dependent and -independent mechanisms of STAT3 activa-tion by the human granulocyte colony-stimulating factor(G-CSF) receptor are differentially utilized depending on G-CSF concentration. Blood 1999;93;113-124.
31 Bungart B, Loeffler M, Goris H et al. Differential effects ofrecombinant human colony stimulating factor (rhG-CSF) onstem cells in marrow, spleen and peripheral blood in mice. Br J Haematol 1990;76:174-179.
32 Lord BI, Woolford LB, Wood LM et al. Mobilization of earlyhematopoietic progenitor cells with BB-10010: a genetically

Wang, Fjerdingstad, Strøm-Gundersen et al. 264
engineered variant of human macrophage inflammatoryprotein-1a. Blood 1995;85:3412-3415.
33 Price TH, Chatta GS, Dale DC. Effect of recombinant granu-locyte colony-stimulating factor on neutrophil kinetics innormal young and elderly humans. Blood 1996;88:335-340.
34 Molineux G, Pojda Z, Dexter TM. A comparison ofhematopoiesis in normal and splenectomized mice treatedwith granulocyte colony-stimulating factor. Blood1990;75:563-569.
35 Long MW, Dixit VM. Thrombospondin functions as acytoadhesion molecule for human hematopoietic progenitorcells. Blood 1990;75:2311-2318.
36 Traycoff CM, Orazi A, Ladd AC et al. Proliferation-induceddecline of primitive hematopoietic progenitor cell activity is
coupled with an increase in apoptosis of ex vivo expandedCD34+ cells. Exp Hematol 1998;26:53-62.
37 Jacob J, Haug JS, Raptis S et al. Specific signals generated by the cytoplasmic domain of the granulocyte colony-stimulating factor (G-CSF) receptor are not required for G-CSF-dependent granulocytic differentiation. Blood1998;92:353-361.
38 Ogawa M. Hemopoietic stem cells: stochastic differentiationand humoral control of proliferation. Environ Health Perspect1989;80:199-207.
39 Metcalf D. Lineage commitment and maturation inhematopoietic cells: the case for extrinsic regulation. Blood1998;92:345-347.