mapeamento de neurÔnios nitrÉrgicos no … · funcional. aprovado em: ... that inhabits rocky...
TRANSCRIPT

LUCIMÁRIO THIAGO FELIX DE ARAUJO
MAPEAMENTO DE NEURÔNIOS NITRÉRGICOS NO TRONCO
ENCEFÁLICO DO MOCÓ (Kerodon rupestris)
Dissertação de Mestrado apresentada ao Programa de Pós-graduação em Biologia Estrutural e Funcional da Universidade Federal do Rio Grande do Norte para obtenção do titulo de mestre.
Natal
2016

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE
DEPARTAMENTO DE MORFOLOGIA
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA ESTRUTURAL E FUNCIONAL
MAPEAMENTO DE NEURÔNIOS NITRÉRGICOS NO TRONCO
ENCEFÁLICO DO MOCÓ (Kerodon rupestris)
LUCIMÁRIO THIAGO FELIX DE ARAUJO
Orientador
Judney Cley Cavalcante
Natal
2016

Universidade Federal do Rio Grande do Norte - UFRN
Sistema de Bibliotecas - SISBI
Catalogação de Publicação na Fonte. UFRN - Biblioteca Setorial do Centro de Biociências - CB
Araujo, Lucimário Thiago Felix de.
Mapeamento de neurônios nitrégicos no tronco encefálico do
Mocó (Kerodon rupestris) / Lucimário Thiago Felix de Araujo. - Natal, 2016.
92 f.: il.
Dissertação (Mestrado) - Universidade Federal do Rio Grande do
Norte. Centro de Biociências. Programa de Pós-Graduação em Biologia Estrutural e Funcional.
Orientador: Prof. Dr. Judney Cley Cavalcante.
1. Óxido nítrico - Dissertação. 2. Kerodon rupestris -
Dissertação. 3. Tronco encefálico - Dissertação. I. Cavalcante, Judney Cley. II. Universidade Federal do Rio Grande do Norte.
III. Título.
RN/UF/BSE-CB CDU 546.172.6

ARAUJO, L. T. F. Mapeamento de neurônios nitrérgicos no tronco encefálico do
mocó (Kerodon rupestris). Dissertação apresentada à Universidade Federal do Rio
Grande do Norte para obtenção do título de Mestre em Biologia Estrutural e
Funcional.
Aprovado em:
Banca Examinadora
Prof. Dr. Judney Cley Cavalcante Instituição: UFRN
Julgamento: _________________________ Assinatura: ______________________
Prof. Dr. Vanessa de Paula Soares Rachetti Instituição: UFRN
Julgamento: _________________________ Assinatura: ______________________
Prof. Dr. José de Anchieta de Castro e Horta Júnior Instituição: UNESP
Julgamento:_________________________Assinatura:________________________

AGRADECIMENTOS
Agradeço a Deus por estar sempre me abençoando e me ajudando a alcançar
tantas coisas em minha vida, principalmente a acadêmica, toda honra e gloria a ele.
A toda minha família que sempre torceu junto nas minhas conquistas, meus irmãos
Lúcia, Luciana e Lucyelves, minha mãe Maria, e minha outra Lúcia, apesar de
pequena, não podia haver família melhor, amo a todos, principalmente minha vó
Maria (in memoriam) que nessa etapa não pode estar presente, mas sei que esta lá
em cima torcendo e me abençoando, obrigado “véia”, te amo, e a todos os amigos
de longas datas, do trabalho a todos meu muito obrigado.
A todos meus professores, os das disciplinas do mestrado, bem como os que
compõem o Labneuro (Expedito, Miriam, Ruthnaldo, Fernando, Aliandra), não posso
apontar qual destes foram os mais importantes, pois todos doaram um pouco do seu
saber, trabalho, tempo e paciência a mim, grato a todos.
Ao meu orientador Judney, que desde o inicio se prontificou em me orientar, sempre
com paciência e pensamento positivo que tudo daria certo, apesar dos percalços
que apareceram nessa caminhada, sempre solucionou os problemas e enfim, aqui
chegamos, sem sua valiosa ajuda, seria muito difícil, obrigado.
Aos meus grandes amigos (irmãos) de caminhada, se sem Profº Judney já seria
difícil, imagine sem Wilqui e Emanuela (Manu), seria impossível, Wilqui por toda sua
“sapiência” e Manu pela “paciência” em me orientar e ajudar nas mais diversas
dificuldades, do intelectual ao computacional, ambos nunca se negaram em ajudar e
sempre estiveram presente nos procedimentos, na elaboração e na descontração
que fez parte dessa caminhada acadêmica, agradeço e publicamente exponho que
os amo, contem comigo para o que precisar.
Pra finalizar, mas não menos importante a todos os amigos que fiz nessa jornada, os
de turma: Nayra, Jonas, Kadigna, Rodrigo, Dáfiny, Epifânio, Marília, Edson, Marden
e Natan, e os de Labneuro: Joacil, Melqui, Fladjany, Nelyane, Naryllenne, Nayana,
Hélder, Mayara, Brenna, Palôma, Ana, Daniel, e Paulo, cada um de vocês fazem
parte de quem sou agora, bem como Regina, sem ela, nem eu nem ninguém que
utiliza o Labneuro produziria nada, muito obrigado de coração a todos.

RESUMO
Óxido nítrico (NO) é uma molécula gasosa importante em processos intra e extracelular, está presente em vários tecidos, exercendo um papel importante no sistema imune, na regulação da pressão sanguínea, no relaxamento do músculo liso e na vasodilatação via nervos periféricos. Por ser um gás o NO atravessa livremente as membranas celulares. Portanto, na parte central do sistema nervoso age como um neurotransmissor em circuitos locais, distribuindo-se em curtas distâncias, difundindo-se de célula em célula participando de várias funções, como aprendizado e memória, podendo mediar respostas excitatórias a certos aminoácidos. O NO está presente em vários núcleos distribuídos por todo o sistema nervoso e tem sido descrito em diversas espécies animais através da expressão de sua enzima de síntese, a NOS. Utilizando as técnicas de imunoistoquímica contra NOS e de histoquímica para marcar a atividade enzimática da NADPH diaforase, a NADPHd, nós mapeamos a distribuição de NO no tronco encefálico de um roedor endêmico da caatinga brasileira que habita áreas rochosas e tem hábitos crepusculares, o mocó (Kerodon rupestris). Diversas regiões e núcleos do tronco encefálico apresentaram imunorreatividade ao NOS ou atividade NADPHd. O presente trabalho mostra que em mocó, entre outras regiões, NO está em núcleos do sistema visual primário como os núcleos pré-tectais e colículo superior; núcleos do sistema auditivo como o complexo olivar, colículo inferior e núcleo coclear; em diversas subdivisões da substância cinzenta periaquedutal, estrutura envolvida em comportamentos defensivos, nocicepção e ansiedade, assim como o locus cerúleos onde NO também foi encontrado; substância negra, onde NO está relacionado a encefalopatia hepática e com a doença de Parkinson em outros roedores. O grande número de núcleos nitrérgicos e seu padrão de distribuição no tronco encefálico de mocó foi semelhante ao observado em outras espécies de mamíferos, nos levando a concluir que a proximidade filogenética das espécies é mantida na distribuição de NO no tronco encefálico de mocó.
Palavras-chave: Óxido nítrico, Kerodon rupestris, Óxido nítrico sintase, Tronco
encefálico.

Title: Mapping the nitrergic neurons in the brainstem of the rock cavy (Kerodon
rupestris)
Nitric oxide (NO) is a gas very important in intra and extracellular process. It is present in many tissues, having an important role in the immunological system, regulation of blood pressure, relaxation of the smooth muscle and vasodilatation through peripheral nervous system. As a gas, NO can pass freely through cell membranes. Therefore, it acts as a neurotransmitter of local action in the central nervous system, reaching short distances, diffusing from cell to cell. It has many functions as learning and memory and modulation of excitatory responses to aminoacids. NO is present in many areas throughout the brain and has been described in many animal species by the expression of its enzyme of synthesis, the NOS. Using immunohistochemistry against NOS and NADPH diaphorase histochemistry we described the distribution of NO in the brainstem of a rodent typical from the Brazilian caatinga, that inhabits rocky areas and has crepuscular habits, the rock cavy (Kerodon rupestris). Several brainstem areas and nuclei presented immunorreactivity to NOS or NADPHd activity. Some of them showed a high density of immunostained neurons, for example the periaqueductal gray, the magnocellular nucleus of the posterior commissure and the interpeduncular nucleus. Others nuclei presented a moderated, as the posterior pretectal nucleus, olivary pretectal nucleus, layers of the superior colliculus and cuneate nucleus. Several showed a low density as the posterior commissure nuclei, paranigral nucleus external cortex of the inferior colliculus and the inferior oliva. Finally, there is nuclei with a very low density, for example the reticular part of the substantia nigra, the pararubral nucleus, the dorsal raphe nucleus, the gracile nucleus and interstitial nucleus of the medulla. The great number of nitrergic nuclei and its pattern of distribution in the rock cavy's brainstem is similar to the other mammals, leading us to conclude that the phylogenetic proximity between species is kept in the brainstem. The present work shows that among other regions the NO is in nuclei of the primary visual system as the pretectal nuclei and the superior coliculum; nuclei of the auditory system as the olivar complex, inferior coliculum and the coclear nuclei; it is in several divisions of the periaqueductal grey matter, strutucture involved in defensive behavior, nociception and anxiety, like the locus coeruleus, where NO also was find; the substantia nigra, where the NO has been related to hepatic encephalopatia and Parkinson's disease in other rodents.The great number of nitrergic nuclei and its pattern of distribution in the rock cavy's brainstem looked alike what is seen in other mammals, leading us to conclude that the philogenetic proximity of the species is mantained in the distribution of NO in the rock cavy's brainstem. Keywords: Nitric oxide, Kerodon rupestris, Nitric oxide sintase, Brainstem.

LISTA DE FIGURAS
Figura 1- Isoformas da óxido nítrico sintase..........................................................................24
Figura 2- Síntese do NO........................................................................................................25
Figura 3- Esquema representando a biossíntese do óxido nítrico catalisada pela NOS. sGC
(guanilato ciclase solúvel), guanosina monofosfato cíclico (GMPc) CaM (calmodulina), PKA,
PKC (proteína quinase A e C)................................................................................................26
Figura 4- O mocó (Kerodon rupestris) em ambiente natural.................................................31
Figura 5- Encéfalo do mocó em vista ventral (A), lateral (B) e dorsal (C). Escala de régua
em centímetros.......................................................................................................................35
Figura 6- Exemplos das diferentes intensidades de marcação celular das células marcadas
por imunoistoquímica para NOS dentro de cada núcleo........................................................38
Figura 7- Exemplos da diferente quantificação subjetiva da densidade das células marcadas
por imunoistoquímica para NOS.............................................................................................38
Figura 8- Reconstrução esquemática das secções coronais do encéfalo do mocó ilustrando
a distribuição dos neurônios NOS-IR do tronco encefálico e de fotomicrografias de cortes
submetidos à coloração de Nissl no nível correspondente, distando uma secção do seu
subsequente por 270 µm........................................................................................................45
Figura 9- Fotomicrografias em campo claro de secções coronais do encéfalo de mocó
submetidas à imunoistoquímica para NOS. (A) Menor aumento do mesencéfalo com alguns
núcleos apresentando neurônios NOS-IR. (B) Ampliação de caixa homônima em A com
área correspondente ao núcleo pré-comissural (PrC). (C) Ampliação de caixa homônima em
A com área correspondente a substância cinzenta periaquedutal (PAG). (D) Ampliação de
caixa homônima em A com área correspondente a parte dorsal do núcleo pré-tectal anterior
(APTD). (E) Ampliação de caixa homônima em A com área correspondente a parte ventral
do núcleo pré-tectal anterior (APTV). 3V, terceiro ventrículo. Barra: 500 µm em A, 100 µm
em B, C, D e E........................................................................................................................49

Figura 10- Fotomicrografias em campo claro de secções coronais do encéfalo de mocó
submetidas à imunoistoquímica para NOS. (A) Menor aumento do mesencéfalo com alguns
núcleos apresentando neurônios NOS-IR. (B) Ampliação de caixa homônima em A com
área correspondente a substância cinzenta periaquedutal dorsomedial (DMPAG). (C)
Ampliação de caixa homônima em A com área correspondente a substância cinzenta
periaquedutal dorsolateral (DLPAG). Aq, aqueduto; SuG, camada cinza superficial do
colículo superior; Op, camada de nervo óptico do colículo superior; InG, camada
intermediária cinza do colículo superior; DpG, camada cinza profunda do coliculo superior.
Barra: 500 µm em A, 100 µm em B e C..................................................................................50
Figura 11- Fotomicrografias em campo claro de secções coronais do encéfalo de mocó
submetidas à imunoistoquímica para NOS. (A) Menor aumento do mesencéfalo com alguns
núcleos apresentando neurônios NOS-IR. (B) Ampliação de caixa homônima em A com
área correspondente a parte lateral da substância negra (SNL). (C) Ampliação de caixa
homônima em A com área correspondente a parte reticulada da substância negra (SNR). cp,
pedúnculo cerebral. Barra: 500 µm em A, 100 µm em B e C.................................................52
Figura 12- Fotomicrografias em campo claro de secções coronais do encéfalo de mocó
submetidas à imunoistoquímica para NOS. (A) Menor aumento do mesencéfalo com alguns
núcleos apresentando neurônios NOS-IR. (B) Ampliação de caixa homônima em A com
área correspondente ao núcleo do trato óptico (OT). (C) Ampliação de caixa homônima em
A com área correspondente ao núcleo magnocelular da comissura posterior (MCPC) e o
núcleo retroparafascicular (RPF). (D) Ampliação de caixa homônima em A com área
correspondente ao núcleo de Darkschewitsch (DK). Aq, aqueduto; cp, pedúnculo cerebral.
Barra: 500 µm em A, 100 µm em B, C e D.............................................................................53
Figura 13- Fotomicrografias em campo claro de secções coronais do encéfalo de mocó
submetidas à imunoistoquímica para NOS. (A) Menor aumento do mesencéfalo com alguns
núcleos apresentando neurônios NOS-IR. (B) Ampliação de caixa homônima em A com
área correspondente ao campo retrorubral (RRF). (C) Ampliação de caixa homônima em A
com área correspondente ao núcleo interpeduncularrostral (IPR), caudal (IPC), intermediário
(IPI) e lateral (IPL). Barra: 500 µm em A, 100 µm em B e C..................................................55
Figura 14- Fotomicrografias em campo claro de secções coronais do encéfalo de mocó
submetidas à imunoistoquímica para NOS. (A) Menor aumento do mesencéfalo com alguns
núcleos apresentando neurônios NOS-IR. (B) Ampliação de caixa homônima em A com

área correspondente a parte lateral do núcleo dorsal da rafe (DRL), ao núcleo posterodorsal
da rafe (PDR), a parte dorsal do núcleo dorsal da rafe (DRD) e a parte ventral do núcleo
dorsal da rafe (DRV). (C) Ampliação de caixa homônima em A com área correspondente ao
núcleo retrorubral (RR). (D) Ampliação de caixa homônima em A com área correspondente
ao núcleo rabdoide (Rbd) e ao núcleo mediano da rafe (MnR). Barra: 500 µm em A, 100 µm
em B, C e D............................................................................................................................56
Figura 15- Fotomicrografias em campo claro de secções coronais do encéfalo de mocó
submetidas à imunoistoquímica para NOS. (A) Menor aumento da ponte com alguns núcleos
apresentando neurônios NOS-IR. (B) Ampliação de caixa homônima em A com área
correspondente ao córtex externo do colículo inferior (ECIC). (C) Ampliação de caixa
homônima em A com área correspondente a parte lateral do núcleo dorsal da rafe (DRL).
(D) Ampliação de caixa homônima em A com área correspondente ao núcleo tegmental
pedunculopontino (PTg). Barra: 500 µm em A, 100 µm em B, C e D.....................................58
Figura 16- Fotomicrografias em campo claro de secções coronais do encéfalo de mocó
submetidas à imunoistoquímica para NOS. (A) Menor aumento da ponte com alguns núcleos
apresentando neurônios NOS-IR. (B) Ampliação de caixa homônima em A com área
correspondente ao locus cerúleos (LC). (C) Ampliação de caixa homônima em A com área
correspondente ao núcleo trigêminal mesencefálico (Me5). (D) Ampliação de caixa
homônima em A com área correspondente ao núcleo subceruleus, parte ventral (SubCV).
Barra: 500 µm em A, 100 µm em B, C e D.............................................................................60
Figura 17- Fotomicrografias em campo claro de secções coronais do encéfalo de mocó
submetidas à imunoistoquímica para NOS. (A) Menor aumento do bulbo com alguns núcleos
apresentando neurônios NOS-IR. (B) Ampliação de caixa homônima em A com área
correspondente a parte parvocelular do núcleo vestibular medial (MVePC). (C) Ampliação de
caixa homônima em A com área correspondente ao núcleo preposito (Pr). (D) Ampliação de
caixa homônima em A com área correspondente ao núcleo dorsal paragigantocelular
(DPGi). Barra: 500 µm em A, 100 µm em B, C e D................................................................62
Figura 18- Fotomicrografias em campo claro de secções coronais do encéfalo de mocó
submetidas à imunoistoquímica para NOS. (A) Menor aumento do bulbo com alguns núcleos
apresentando neurônios NOS-IR. (B) Ampliação de caixa homônima em A com área
correspondente a parte lateral do núcleo cuneado externo (ECu). (C) Ampliação de caixa
homônima em A com área correspondente aos núcleos do trato solitário (Sol). (D)

Ampliação de caixa homônima em A com área correspondente ao núcleo preposito, parte
magnocelular (PrMc). (E) Ampliação de caixa homônima em A com área correspondente
aos núcleos olivares inferiores (IO). Barra: 500 µm em A, 100 µm em B, C, D e E...............63
Figura 19- Fotomicrografias em campo claro de secções coronais do encéfalo de mocó
submetidas à imunoistoquímica para NOS. (A) Menor aumento do bulbo com alguns núcleos
apresentando neurônios NOS-IR. (B) Ampliação de caixa homônima em A com área
correspondente ao núcleo grácil (Gr). (C) Ampliação de caixa homônima em A com área
correspondente a camada gelatinosa do núcleo trigêminal espinal caudal, (Ge5) e ao núcleo
trigêminal espinal, parte caudal (Sp5C). (D) Ampliação de caixa homônima em A com área
correspondente ao núcleo reticular lateral (LRt). Barra: 500 µm em A, 100 µm em B,C e
D.............................................................................................................................................64
Figura 20- Fotomicrografias em campo claro de secções coronais do encéfalo de mocó
submetidas à histoquímica. (A,C,G,I) Menor aumento de cortes do tronco encefálico com
alguns núcleos apresentando neurônios corados para NADPHd. (B) Ampliação de caixa
homônima em A area correspondente ao núcleo interpeduncular rostral (IPR), e suas
divisões: caudal (IPC), intermediário (IPI) e lateral (IPL), (D) Ampliação de caixa homônima
em C area correspondente ao a camada zonal (Zo) da camada cinza superficial do colículo
superior (SuG) e suas divisões seguintes: camada de nervo óptico (Op), camada
intermediária cinza (InG), (E) camada cinza profunda (DpG), (F) núcleo tegmental
microcelular (MiTg), (H) Ampliação de caixa homônima em G área correspondente ao Locus
cerúleos (LC), núcleo trigeminal mesencefálico (Me5), (J) Ampliação de caixa homônima em
I área correspondente ao camada gelatinosa do núcleo trigeminal espinhal caudal (Ge5),
núcleo trigeminal espinhal, parte caudal (Sp5C)....................................................................66

LISTA DE QUADROS
Tabela 1- Densidade e intensidade dos núcleos imunorreativos a NOS...............................39

LISTA DE ABREVIATURAS
10N núcleo motor dorsal do vago
11N núcleo do nervo acessório
12N núcleo hipoglosso
3V terceiro ventrículo
4N núcleo troclear
4V quarto ventrículo
5Ma núcleo motor trigeminal, parte masseter
5N núcleo trigeminal
5Sol zona de transição trigeminal-solitário
5Te núcleo motor trigeminal, parte temporal
6N núcleo abducente
7N núcleo facial
7-NIO 7-nitroindazol
8vn raiz vestibular do nervo vestibulococlear
A1 A1células noradrenérgicas
A2 A2 células noradrenérgicas
AmbC núcleo ambíguo, parte compacta
APT núcleo pré-tectal anterior
APTD núcleo pré-tectal anterior, parte dorsal
APTV núcleo pré-tectal anterior, parte ventral
Aq aqueduto mesencefálico
BIC núcleo do braço do colículo inferior
bsc braço do colículo superior
Ca2+ ion cálcio
CaM calmodulina
CC canal central
CeCv núcleo cervical central da medula espinal
CEUA-UFRN Comitê de Ética em Uso Animal da Universidade Federal do RN
CGA cinza central, parte alfa
CGB cinza central, parte beta
CGG cinza central, parte gama
CGPn substancia cinzenta da ponte
ChAT colina acetil-transferase
cNOS óxido nítrico constitutiva

cp pedúnculo cerebral
Csc comissura do colículo superior
cu fascículo cuneado
Cu núcleo cuneado
CuR núcleo cuneado, parte rotunda
DAB diaminobenzidina
DCDp núcleo coclear dorsal, núcleo profundo
DCIC córtex dorsal do colículo inferior
dcs trato corticoespinal dorsal
Dk núcleo de Darkschewitsch
DLPAG substância cinzenta periaquedutal, porção dorsolateral
DMPAG substância cinzenta periaquedutal, porção dorsomedial
DMSp5 núcleo espinal trigeminal dorsomedial
DMTg área tegmental dorsomedial
DpG colículo superior, camada cinza profunda
DPGi núcleo paragigantocelular dorsal
DPO região periolivar dorsal
DR núcleo dorsal da rafe
DRD parte dorsal do núcleo dorsal da rafe
DRL parte lateral do núcleo dorsal da rafe
DRV parte ventral do núcleo dorsal da rafe
dsc/oc trato espinocerebelar dorsal e olivocerebelar
DT núcleo terminal dorsal do trato óptico acessório
ECIC córtex externo do colículo inferior
ECu núcleo cuneado externo, parte lateral
eNOS óxido nítrico endotelial
FAD dinucleótido de flavina-adenina
FMN mononucleótido de flavina
Fr fascículo retroflexo
FVe grupo de células do complexo vestibular
g7 genu do nervo facial
GC guanilato ciclase
GCs guanilato ciclase solúvel
Ge5 camada gelatinosa do núcleo trigeminal espinal caudal
Gi núcleo gigantocelular reticular
GiA núcleo gigantocelular reticular, parte alfa

GiV núcleo reticular gigantocelular, parte ventral
GMPc Monofosfato cíclico de guanosina
gr fascículo grácil
Gr núcleo grácil
GTP guanosina-5´-trifosfato
H2O2 peróxido de hidrogênio
HCT1026 [2-Fluoro-a-metil (1,1'-bifenil) -4-acético-4- (nitrooxi) butil éster]
I.C.V intracerebroventricular
IB núcleo intersticial da medula
icp pedúnculo cerebelar inferior
IF núcleo interfascicular
ILL núcleo intermediário do lemnisco lateral
InC núcleo intersticial de Cajal
InCSh concha do núcleo intersticial de Cajal
InG camada intermediária cinza do colículo superior
iNOS óxido nítrico induzível
IO oliva inferior
IOA oliva inferior, núcleo medial, subnúcleo A
IOB oliva inferior, núcleo medial, subnúcleo B
IOBe oliva inferior, subnúcleo beta
IOC oliva inferior, núcleo medial, subnúcleo C
IPC núcleo interpeduncular, subnúcleo caudal
IPDL núcleo interpeduncular, subnúcleo dorsolateral
IPI núcleo interpeduncular, subnúcleo intermediário
IPL núcleo interpeduncular, subnúcleo lateral
IPR núcleo interpeduncular, subnúcleo rostral
IRt núcleo reticular intermediário
IRtA núcleo reticular intermediário, parte alfa
IS núcleo salivatório inferior
isRt formação reticular ístmica
LC locus cerúleos
LDTg núcleo tegmental laterodorsal
LDTgV núcleo tegmental laterodorsal, parte ventral
lfp fascículo longitudinal da ponte
L-NOARG NG-nitro-L-arginina
LPAG substância cinzenta periaquedutal, porção lateral

LPB núcleo parabraquial lateral
LPBI núcleo parabraquial lateral, parte interna
LPBV núcleo parabraquial lateral, parte ventral
LPG núcleo paragigantocelular
LPGiA núcleo paragigantocelular lateral, parte alfa
LPGiE núcleo paragigantocelular lateral, parte externa
LRt núcleo reticular lateral
LRtS5 núcleo reticular lateral, parte subtrigeminal
LSO oliva superior lateral
LT núcleo terminal lateral do trato óptico acessório
Lth núcleo litóide
LVe núcleo vestibular lateral
LVPO núcleo periolivar lateroventral
MA3 núcleo medial acessório do oculomotor
macNOS óxido nítrico do macrófago
MCPC núcleo magnocelular da comissura posterior
MdD núcleo reticular medular, parte dorsal
MdV núcleo reticular medular, parte ventral
Me5 núcleo trigeminal mesencefálico
MHb núcleo habenular medial
MiTg núcleo tegmental microcelular
ml lemnisco medial
ML núcleo mamilar medial, parte lateral
Mlf fascículo media longitudinal
MnA núcleo acessório mediano da medula
MnR núcleo mediano da rafe
mp pedúnculo mamilar
MPB núcleo parabraquial medial
MPL núcleo medial paralemniscal
MPT núcleo pré-tectal medial
MPTP 1-metil-4-fenil-1,2,3,6-tetra-hidropiridina
MSO oliva superior medial
MT núcleo terminal medial do trato óptico acessório
MVeMC núcleo vestibular medial, parte magnocelular
MVePC núcleo vestibular medial, parte parvocelular
MVPO núcleo periolivar medioventral

Mx região matriz da medula
N2O óxido nitroso
NaCl cloreto de sódio
NADPH nicotinamida adenina dinucleótideo fosfato
NADPHd nicotinamida adenina dinucleótideo fosfato diaforase
NMDA N-metil D-aspartato
nNOS óxido nítrico neuronal
No núcleo olivar
NO óxido nítrico
NOS óxido nítrico sintase
NOS-ir oxido nítrico imunoreativo
NPLA Nω-propil-L-arginina
O2 oxigénio
ocb feixe olivococlear
Op camada do nervo óptico do colículo superior
OPT núcleo olivar pré-tectal
opt trato óptico
OT núcleo do trato óptico
P1Rt formação reticular
P5 zona peritrigeminal
Pa6 núcleo parabducente
PaC núcleo paracomissural da comissura posterior
PAG substância cinzenta periaquedutal
PaR núcleo pararubral
PB tampão fosfato 0,1M
PBG núcleo parabigeminal
PBP núcleo parabraquial pigmentado do VTA
PCom núcleo da comissura posterior
PCR Reação em cadeia da polimerase
PCRt núcleo reticular parvocelular
PCRtA núcleo reticular parvocelular, parte alfa
PDR núcleo posterodorsal da rafe
PDTg núcleo tegmental posterodorsal
PHOX NADPH oxidase
PKA proteína quinase A
PKC proteína quinase C

PKG I proteína quinase G I
PKG II proteína quinase G II
PLV núcleo perilemniscal , parte ventral
PMnR núcleo paramediano da rafe
PN núcleo paranigral do VTA
Pn núcleo pontino
PnC parte caudal do núcleo reticular pontino
PnO parte oral do núcleo reticular pontino
PnV núcleo reticular pontino, parte ventral
PP núcleo peripeduncular
PPT núcleo pré-tectal posterior
PR campo pré-rubral
Pr núcleo prepósito
Pr5DM parte dorsomedial do núcleo principal sensorial do trigêmeo
Pr5VL parte ventrolateral do núcleo principal sensorial do trigêmeo
PrBo complexo pre-Botzinger
PrC núcleo pré-comissural
PrCnF área pré-cuneiforme
PrEW núcleo pre-Edinger-Westphal
PrMc núcleo prepósito, parte magnocelular
PTg núcleo tegmental pedunculopontino
pyx decussação piramidal
RAmb núcleo retroambíguo
Rbd núcleo rabdóide
RIP núcleo interpósito da rafe
RLi núcleo linear rostral da rafe
RMC parte magnocelular do núcleo rubro
RMg núcleo magno da rafe
Ro núcleo de Roller
ROb núcleo obscuro da rafe
RPa núcleo pálido da rafe
RPC parte parvocelular do núcleo rubro
RPF núcleo retroparafascicular
RR núcleo retrorrubral
RRF campo retrorrubral
RtTg núcleo reticulotegmental da ponte

RtTgL núcleo reticulotegmental da ponte, parte lateral
SARA sistema de ativação reticular ascendente
scp pedúnculo cerebelar superior
SNCD substância negra, parte compacta, camada dorsal
SNCM substância negra, parte compacta, camada medial
SNL substância negra, parte lateral
SNR substância negra, parte reticulada
Sol trato solitário
SolDM núcleo trato solitário, parte dorsomedial
SolIM núcleo trato solitário, parte intermediaria
SolL núcleo trato solitário, parte lateral
SolM núcleo trato solitário, parte medial
SolV núcleo trato solitário, parte ventral
Sp5 núcleo espinal trigeminal
sp5 trato espinal trigeminal
Sp5C núcleo espinal trigeminal, parte caudal
Sp5I núcleo espinal trigeminal, parte interpolar
Sp5O núcleo espinal trigeminal, parte oral
SPO núcleo paraolivar superior
SPTg núcleo tegmental subpeduncular
SpVe núcleo vestibular espinal
Su3 supraoculomotor
Su3C supraoculomotor cap
SubB núcleo sub-braquial
SubCA núcleo subceruleus, parte alfa
SubCD núcleo subceruleus, parte dorsal
SubCV núcleo subceruleus, parte ventral
SuG camada cinza superficial do colículo superior
SuS núcleo salivatório superior
SuVe núcleo vestibular superior
TH tirosina hidroxilase
TH-ir tirosina hidroxilase imunorreativo
TrLL núcleo triangular do lemnisco lateral
Ts trato tectoespinal
tth trato trigeminotalâmico
TXPB tampão fosfato com tampão Triton X-100 a 0,4%

Tz núcleo do corpo trapezóide
un fascículo uncinado do cerebelo
VCA núcleo coclear ventral, parte anterior
VCP núcleo coclear ventral, parte posterior
verme trato vestibulomesencefalico
vesp trato vestibuloespinal
VG núcleo geniculado ventral
VLL núcleo ventral do lemnisco lateral
VLPAG substância cinzenta periaquedutal ventrolateral
VTAR área tegmental ventral, parte rostral
VTg núcleo tegmental ventral
X núcleo x
Zo camada zonal do colículo superior
β-NADP Nicotinamida adenina dinucleótido fosfato hidratado

SUMÁRIO
1. INTRODUÇÃO......................................................................................................................21
1.1 ÓXIDO NÍTRICO.................................................................................................................21
1.1.1 O NO como neurotransmissor......................................................................................22
1.1.2 Funções do NO...............................................................................................................23
1.1.3 Óxido nítrico sintase (NOS)..........................................................................................24
1.1.4 Síntese do NO.................................................................................................................25
1.2 TRONCO ENCEFÁLICO.....................................................................................................27
1.2.1 Mesencéfalo...................................................................................................................27
1.2.2 Ponte...............................................................................................................................28
1.2.3 Bulbo...............................................................................................................................28
1.3 MODELO EXPERIMENTAL................................................................................................29
2. OBJETIVOS..........................................................................................................................32
2.1 OBJETIVO GERAL ............................................................................................................32
2.2 OBJETIVOS ESPECÍFICOS...............................................................................................32
3. MATERIAL E MÉTODOS.....................................................................................................33
3.1 ANIMAIS.............................................................................................................................33
3.2 PROCEDIMENTOS............................................................................................................33
3.2.1 Perfusão..........................................................................................................................33
3.2.2 Remoção do encéfalo e microtomia.............................................................................34
3.2.3 Imunoistoquímica..........................................................................................................35
3.2.4 Método de Nissl.............................................................................................................36
3.2.5 Histoquímica.................................................................................................................36
3.2.6 Análise das imagens....................................................................................................37
4. RESULTADOS.....................................................................................................................39
4.1 IMUNOISTOQUÍMICA........................................................................................................48
4.2 HISTOQUÍMICA.................................................................................................................65
5. DISCUSSÃO......................................................................................................................67

5.1 ASPECTOS NEUROANATÔMICOS E FUNCIONAIS DO ÓXIDO NÍTRICO NO TRONCO
ENCEFÁLICO DO MOCÓ......................................................................................................68
6. CONCLUSÃO....................................................................................................................75
REFERÊNCIAS..................................................................................................................76
ANEXOS.............................................................................................................................89

21
1. INTRODUÇÃO
1.1 ÓXIDO NÍTRICO
Uma das menores moléculas produzidas nos organismos vivos, dos peixes
aos mamíferos (Alonso et al., 2000), o óxido nítrico (NO) é um gás altamente
lipofílico que se difunde através das membranas celulares e tem como
características químicas: Massa molar: 30,01 g/mol; Densidade: 1,34 kg/m³; Ponto
de ebulição: -152 °C; e Ponto de fusão: -164 °C.
Encontrado no ar atmosférico em quantidades muito pequenas, o NO é um
gás muito difusível e tóxico, pois possui sete elétrons na molécula de nitrogênio e
oito na de oxigênio, tendo um elétron desemparelhado que o torna altamente reativo
(Snyder; Bredt, 1992). Porém, também pode ser redutor dependendo do meio em
que ele está e é rapidamente destruído pelo oxigênio (Archer, 1993). Sua oxidação
produz como subproduto nitrito e nitrato (Kiechle et al., 1993).
O início dos estudos com NO se deu em pesquisas de diferentes grupos
independentes e em períodos próximos. Furchgott e Zawadzki (1980) investigavam
um fator vasodilatador associado ao endotélio vascular, devido a sua ação relaxante
vascular, promovendo vasodilatação pela ativação de guanilato ciclase, portanto
produzindo um aumento de Monofosfato cíclico de guanosina (GMPc). Em 1981
estudos liderados por Green e colaboradores demonstraram a produção de nitrato
em camundongos germ-free.
Posteriormente Stuehr e Marletta (1985) demonstraram que macrófagos
ativados por lipopossacarideos bacterianos eram capazes de produzir nitritos e
nitratos. Hibbs e colaboradores (1987) descobriram que a L-arginina era o substrato
para a produção do NO, e a L-citrulina era formada como co-produto, e no mesmo
ano, Ignarro e colaboradores (1987) confirmam ser o NO a molécula responsável
pelo fator relaxante derivado do endotélio. Trabalhos seguintes feitos por Hibbs e
colaboradores (1988) e Marletta e colaboradores (1988) identificaram que o NO era
um importante mediador citotóxico de células imunes ativadas, agindo na destruição
de patógenos e células tumorais.
Os estudos com o NO com ação neurotransmissora começaram com
Ferrendelli e colaboradores (1974) demonstraram que o glutamato aumentou o

22
GMPc na parte central do sistema nervoso. Miki e colaboradores (1977)
demonstraram a ativação da guanilato ciclase cerebral pelo NO. Mais de uma
década depois, Garthwaite e seus colaboradores (1989), afirmaram que o glutamato
é o mediador da liberação de NO por receptores N-metil-d-aspartato (NMDA). Bredt
e Snyder (1989) e Knowles e colaboradores (1989) confirmaram a produção de NO
no sistema nervoso. Enquanto Schmidt e colaboradores (1991) isolaram e
purificaram no cerebelo de ratos e porcos uma isoforma da enzima responsável pela
formação de NO, a óxido nítrico sintase (NOS).
O desfecho dessas linhas de pesquisa fez com que o NO passasse da
condição de molécula sem importância biológica e pouco estudada para a de um
dos mais importantes mediadores de processos intra e extracelulares.
1.1.1 O NO como neurotransmissor
Normalmente os neurotransmissores são moléculas fabricadas nos corpos
dos neurônios, empacotados em vesículas e liberados pelas terminações nervosas
de neurônios pré-sinápticos que tem por função promover a comunicação entre
células. Na célula pós-sináptica, o neurotransmissor ao alcançar seus receptores
estimula ou inibe essa segunda célula.
Atualmente a maioria dos neurotransmissores situa-se em uma de três
categorias: aminoácidos, aminas e peptídeos. Os neurotransmissores aminoácidos e
aminas são moléculas orgânicas pequenas com pelo menos um átomo de nitrogênio
e liberado pelas vesículas sinápticas. Os neurotransmissores peptídicos são grandes
moléculas formadas por múltiplos aminoácidos, comumente encontrados em
terminais axonais que contêm neurotransmissores aminas ou aminoácidos, e as
vezes dividem as mesmas vesículas (Bear, 2016).
O NO é diferente de todos os outros neurotransmissores clássicos, pois não
é armazenado em vesículas; não age em conjunto com mecanismos específicos de
liberação e captação; não necessita de receptores na membrana celular, pois
adentra a célula alvo por afinidade lipofílica; não há enzimas específicas para
metabolizá-lo, degradando-se espontaneamente por oxidação, por compostos tais
como superóxido ou oxihemoglobina (Ignarro et al., 1989).

23
1.1.2 Funções do NO
As funções do NO são descritas por diversos autores como complexas e
antagônicas, pois as mesmas podem ser benéficas ou tóxicas de acordo com a
concentração ou depuração no tecido; por isso Schmidt e colaboradores (1991)
chamaram o NO de "faca de dois gumes". De uma forma geral, devido à sua entrada
na célula sem carreadores, os organismos utilizam o NO em funções fisiológicas em
que é necessária uma resposta rápida (Ignarro et al., 1987).
O NO também é considerado uma das primeiras defesas do organismo
contra infecções por bactérias, parasitas e vírus (Drapier et al., 1988; Gazzinelli et
al.,1992; Feng e Walker 1993; Oswald et al., 1994; Lowenstein et al.,1996), está
envolvido na função plaquetária, na regulação do tônus vascular e também
modulando o sistema imunológico (Snyder e Bredt, 1992).
Atuando em concentrações superiores à de simples mensageiros, o NO é
tóxico aos microrganismos invasores, porém é tênue o limite de concentração
tissular entre a não toxicidade às células do hospedeiro e a toxicidade necessária
para ação de injurias intra e extracelulares, e nos casos de doenças autoimunes e
situações de extremo ataque ao organismo o NO irá se encontrar em concentrações
tóxicas para as células, mesmo as saudáveis, atuando na maioria dos processos
inflamatórios (Snyder e Bredt, 1992).
No encéfalo, o NO demonstrou estar envolvido numa grande variedade de
efeitos biológicos, como no controle do fluxo sanguíneo (Phillips et al., 1997), atua
na mediação da diferenciação celular (Ogura et al.,1996) e regula a sobrevivência e
a morte celular (Sparrow, 1994; Wink e Mitchell, 1998; Ikonomidou et al., 2001).
Nelson e colaboradores (1997) demonstraram atuar como neurotransmissor
com função organogênica, pois acredita-se que seus efeitos sobre a liberação de
neurotransmissores, como o glutamato, juntamente com o monóxido de carbono
possa ter um importante papel na formação da memória, no aprendizado, na
agressividade e sexualidade. NO também tem sido envolvido em doenças
psiquiátricas como a esquizofrenia (Nasyrova et al., 2015) e motoras como o
Parkinson (Ayton et al., 2015).
No tronco encefálico o NO atua na regulação cardiovascular em ratos
(Tseng et al., 1996). Atenua a rápida adaptação do sistema arterial no controle

24
barorreflexo da atividade do nervo simpático em coelhos (Hironaga et al., 1998). Em
neurônios pré-ganglionares parassimpáticos em humano, NO está envolvido no
controle de lacrimação, salivação, secreção oral e nasofaríngea, e no controle da
dilatação de vasos sanguíneos extra e intracranianos (Gai e Blessing, 1996). NO
também participa da comunicação neuronal e da plasticidade sináptica, modulando a
resposta neuronal do sistema visual e durante o desenvolvimento embrionário
(Tenório et al., 1996).
1.1.3 Óxido nítrico sintase (NOS)
A NOS é a enzima de síntese de NO. Já foram isoladas três isoenzimas,
duas delas, chamadas de cNOS, são constitutivas, sendo elas: a nNOS (NOS
neuronal), isolado primeiramente nos cerebelos de porcos e camundongos (Schmidt
et al., 1991), está presente no encéfalo e leva a síntese do NO, que age como
neurotransmissor; eNOS (NOS endotelial), isolado de células endoteliais de
humano e bovinos, leva a síntese do NO, agindo basicamente como vasodilatador e
no controle de extravasamento tecidual (Pollock et al., 1991).
A terceira isoenzima é a iNOS, a única isoforma induzida, ou seja, só ocorre
nas células por fatores extrínsecos, e é representada pela macNOS (NOS do
macrófago) (Fig. 1). Descoberta no músculo liso (Teng et al., 1998) e hepatócitos de
camundongos a macNOS pode ser induzida em qualquer célula por meio de
lipopolisacarídeos bacterianos ou citocinas, sendo capaz de produzir o NO por um
longo período que o implica em processos patológicos (Stuehr et al., 1991). Como o
atual estudo é no sistema nervoso lidaremos apenas com nNOS e então o
chamaremos simplesmente de NOS.
Figura 1. Isoformas da óxido nítrico sintase (Adaptado de Dusse et al., 2003).

25
Diversos estudos demonstram a presença do NOS no encéfalo de diversas
espécies de vertebrados como peixes (Holmqvist et al., 2000; Masini et al., 2005),
salamandras (Moreno et al., 2002), lagartos (Smeets et al., 1997), pombos (Atoji et
al., 2001), camundongos ( Reuss e Riemann., 2000; Gotti et al., 2005), ratos (Hope
et al., 1991; Vincent e Kimura., 1992; Iwase et al., 1998; Freire et al., 2012), coelhos
(González-Soriano et al., 2002) e humanos (Egberongbe et al., 1994), sendo
particularmente abundante no tronco encefálico, com neurônios nitrérgicos
presentes desde o bulbo até a transição mesodiencefálica, o que denota uma
participação de NO em diversas funções bulbares, pontinas e mesencefálicas.
1.1.4 Síntese do NO
A produção do NO se dá quando a L-arginina é transformada em um
intermediário, a N-hidroxi-L-arginina, com a presença de nicotinamida-adenina
dinúcleotídeo-fostato-hidrogênio (NADPH) e Ca2+ sendo necessário mais NADPH e
O2, além de flavinas e biopterinas para a formação de L-citrulina e NO (Fig. 2).
Figura 2. Síntese do NO (Retirado de Dusse et al., 2003).
O mecanismo de ativação da produção do NO neuronal é iniciada pelo
glutamato, liberado de vesículas de neurônios pré-sinápticos, ligando-se a
receptores, principalmente do tipo NMDA, do neurônio pós-sináptico. Com isso
abrindo um canal fazendo com que o Ca2+ entre para o interior da célula, como
aumento na concentração desses íons no neurônio pós-sináptico ocorre a ativação
de uma proteína quinase dependente de cálcio, a calmodulina, uma proteína que

26
age como uma subunidade para muitas enzimas dependentes de Ca2+. O complexo
Ca2+/calmodulina liga-se a enzima NOS, que catalisa a oxidação de L-arginina em
L-citrulina e NO (Fig. 3) (Saavedra et al.,1997).
Figura 3. Esquema representando a biossíntese do óxido nítrico catalisada pela NOS. sGC
(guanilato ciclase solúvel), GMPc (guanosina monofosfato cíclico), CaM (calmodulina), PKA
e PKC (proteína quinase A e C). (Modificado de Salum et al., 2008).
Depois de produzido e difundido nos tecidos o efeito do NO ocorre a partir
da ativação da enzima guanilato ciclase (GC) que por sua vez, quando ativada,
produz guanosina-3´,5´-monofosfato cíclico (GMPc) a partir da guanosina-5´-
trifosfato (GTP). O GMPc é um segundo mensageiro que ativa proteínas quinase G I
(PKG I), que responde pelo controle da quantidade de Ca2+ intracelular, e proteínas
quinase G II (PKG II), uma proteína de membrana que passa a controlar o fluxo de
ânions, principalmente de íons cloreto.
Um dos alvos do NO sintetizado no elemento pós-sináptico é o próprio
neurônio pré-sináptico, promovendo a liberação de mais glutamato neste neurônio,
realizando assim a regulação de potenciais de longa duração e reiniciando o ciclo
(Moncada e Higgs, 1993).
O NO difunde-se nas terminações nervosas formando ligações covalentes
com alvos potenciais, o qual inclui a guanilato ciclase solúvel. A ação do NO é então
finalizada com a sua difusão para outras áreas, onde será convertido em nitrito/
nitrato (Dawson e Snyder, 1994).

27
1.2 TRONCO ENCEFÁLICO
O tronco encefálico situa-se entre a medula e o diencéfalo, localizando-se no
humano ventralmente ao cerebelo, conectando a medula espinal com outras
estruturas encefálicas superiormente. Embriologicamente ele surge do
rombencéfalo, dando origem a duas vesículas: Mielencéfalo, que dará origem ao
bulbo e o Metencéfalo, no qual dará origem a ponte e o cerebelo.
Ele se divide rostrocaudamente em: mesencéfalo, ponte e bulbo. Os corpos
de neurônios se agrupam em núcleos e fibras nervosas se agrupam em feixes
denominados tratos, fascículos e lemniscos. Muitos dos núcleos do tronco encefálico
recebem ou emitem fibras nervosas que entram na constituição dos nervos
cranianos. Dos 12 pares de nervos cranianos, 10 fazem conexão no tronco
encefálico (Machado, 2005). Uma lesão no tronco encefálico é frequentemente
devastadora e letal, devido ao fato de que no tronco ficam centros de controle
cardíacos e respiratórios (Gray e Standring, 2008).
1.2.1 Mesencéfalo
Interposto entre a ponte e o diencéfalo, é “atravessado” por um estreito
canal: o aqueduto mesencefálico. O teto do mesencéfalo é situado dorsalmente ao
aqueduto. Ventralmente ao aqueduto ficam os dois pedúnculos cerebrais, que
dividem-se em parte dorsal,o tegmento, e outra ventral, a base do pedúnculo (Gray e
Standring, 2011).
Nele temos os centros dos reflexos visuais. Os circuitos neurais da retina
levam informação de imagens em movimento e parado para os colículos superiores
e destes para o nervo oculomotor. Nos colículos também estão o centro responsável
pelos reflexos de controle dos movimentos da cabeça e tronco em resposta aos
estímulos visuais. Os colículos inferiores, são parte da via auditiva, retransmitindo
impulsos do núcleo coclear para o diencéfalo, bem como os centros dos reflexos de
sobressalto. Também encontramos dois pares de nervos cranianos: oculomotor e
troclear (Machado, 2005).
A substância negra é um grande núcleo onde parte dos neurônios produz
dopamina, estendendo-se até os núcleos da base. Axônios vindos do cerebelo e do

28
córtex cerebral formam sinapses nos núcleos rubros que ajudam a controlar
movimentos voluntários dos membros. Os neurônios dentro da formação reticular
têm funções ascendentes sensitivas e descendentes motoras, nela temos o sistema
de ativação reticular ascendente (SARA), axônios sensitivos que se projetam em
direção ao córtex cerebral, ajudando a manter a consciência, ativos durante o
despertar do sono, suas funções motoras vão ajudar a regular a postura e o tônus
muscular, o menor grau de contração nos músculos em repouso normal (Tortora e
Grabowski, 2010).
1.2.2 Ponte
Localizado entre o bulbo e o mesencéfalo, em humanos apresenta-se
ventralmente ao cerebelo e sobre a parte basilar do osso occipital e o dorso da sela
turca do osso esfenóide. A ponte apresenta na sua porção anteroinferior uma
estriação transversal devido a presença de numerosos feixes de fibras transversais
convergindo de cada lado para formar o pedúnculo cerebelar médio, que adentra no
hemisfério cerebelar correspondente (Tortora e Grabowski, 2010).
A ponte é formada por núcleos e tratos sensitivos e motores, contendo
núcleos vestibulares, componentes da via do equilíbrio, núcleos da área
pneumotáxica e a área apnêustica do centro respiratório, ajudando a controlar a
respiração e núcleos associados com os seguintes quatro pares de nervos
cranianos: trigêmeo, abducente, facial e vestibulococlear.
1.2.3 Bulbo
Também conhecido por medula oblonga, no homem tem o formato de cone
e sua extremidade inferior é continua com a medula espinal. Inferiormente o limite
entre medula e bulbo é o primeiro nervo cervical, ao nível do forame magno,
enquanto que superiormente o bulbo vai até o sulco bulbopontino (Gray e Standring,
2011).
No bulbo está o centro cardiovascular que regula a frequência e a
intensidade do batimento cardíaco e o diâmetro dos vasos; a área respiratória
rítmica, do centro respiratório, que ajusta o ritmo básico da respiração; o centro do

29
vômito, que provoca a expulsão forçada pela boca do que estiver na parte superior
do trato gastrointestinal. No bulbo também estão estruturas que promovem a
deglutição, o espirro, a tosse e o soluço.
Na substância branca estão todos os tratos sensitivos e motores, que se
estendem entre a medula espinal e as outras partes do encéfalo. Nele estão duas
protrusões chamadas de pirâmides (Tortora e Grabowski, 2010). É devido à
decussação das pirâmides que o hemisfério cerebral direito controla o lado esquerdo
do corpo e o mesmo acontece no sentido contrário (Gray e Standring, 2011).
No bulbo fica o núcleo olivar inferior que instrui o cerebelo para fazer ajustes
na atividade muscular, desenvolvendo habilidades motoras. A via nervosa do tato
epicrítico, pressão, vibração e propriocepção consciente passam pelos núcleos
grácil e cuneiforme. Com parte no bulbo e parte na ponte existe o núcleo do trato
solitário que faz parte da via gustativa e sensorial visceral; os núcleos cocleares, que
fazem parte da via auditiva recebendo influxo proveniente da cóclea na orelha
interna; e parte dos núcleos vestibulares, componentes da via vestibular, recebendo
informação sensitiva associada com o equilíbrio proveniente também da orelha
interna. No bulbo estão os núcleos associados aos pares de nervos cranianos
vestibulococlear, glossofaríngeo, vago, acessório e hipoglosso (Tortora e Grabowski,
2010).
1.3 ANIMAL EXPERIMENTAL
Os roedores formam uma ordem animal que inclui uma grande quantidade
de espécies adaptados aos mais numerosos habitats e distribuídos por todo o
planeta, desenvolvendo as mais variadas formas anatômicas e funcionais (Lima et
al., 2008).
O Kerodon rupestris, conhecido popularmente como mocó, ocorre em todos
os estados do Nordeste, especialmente na caatinga do semiárido nordestino ate o
norte do estado de Minas Gerais, sendo restritos a áreas rochosas, especialmente
serrotes e serras, mas algumas vezes podem ser encontrados em lajeiros de
planícies (Fig. 4) (Streilein, 1982).
Este roedor, pertencente à família dos cavídeos e da subfamilia caviinae,
possui habitat mais especializado em relação às outras formas de caviinaes e ao

30
gênero Kerodon. No Brasil é representada basicamente pelos gêneros Kerodon,
Cavia e Gálea (Lima et al., 2008).
As características morfológicas são: pernas longas propulsoras e unhas
rombas sobre coxins espessos para que subam em pedras e arvores, não possuem
clavículas, os pés possuem apenas 3 dedos e cauda atrofiada, adulto mede por
volta de 410mm, pesa em torno de 800gr (Santana et al., 2003) e possuem hábito
crepuscular (Sousa e Menezes, 2006).
O mocó habita locais rochosos com diversas fendas para abrigar-se dos
predadores. São facilmente adaptáveis às condições dos locais onde vivem como:
calor, escassez de água e de alimentos, especialmente nos períodos de grandes
secas do semiárido nordestino. Sua alimentação vem de galhos de árvores,
principalmente, pois são excelentes saltadores e escaladores, e em cativeiro comem
de frutas a raízes.
Apresentam uma coloração cinza clara com pelos pretos, amarelos ou
esbranquiçados no dorso, e castanho ferruginoso na região da cauda, um
acastanhado nas patas e branco na região cervical (Moojen, 1952).
Os mocós apresentam inúmeras características biológicas desejáveis à
domesticação, tais como a reprodução em cativeiro, podendo viver até 11 anos,
hábito gregário, poligamia, alta taxa de fecundidade, sociedade e docilidade
(Lacerda et al., 2006).
Apesar dos poucos estudos sobre o mocó, nos últimos anos o Laboratório
de Neuroanatomia da UFRN tem feito um esforço para transformar esta espécie de
roedor em um modelo para estudos neurocientíficos e diversos trabalhos têm sido
publicados (Nascimento Junior et al., 2008, 2010a e 2010b; Cavalcante et al., 2008;
Soares et al., 2012; Cavalcanti et al., 2014; Silva et al., 2014; Morais et al., 2014;
Oliveira et al., 2014). Este esforço tem se baseado principalmente no fato deste
roedor ser animal crepuscular, uma característica interessante e distinta dos
roedores de laboratório como ratos, camundongos e hamsters, além de ser um
animal regional, o que pode trazer características próprias de um animal adaptado a
vida na caatinga brasileira.
O NO é uma substância que vem sendo amplamente estudado já a algumas
décadas. Sua distribuição no sistema nervoso foi estudada em diversas espécies
mostrando semelhanças e diferenças que denotam prováveis diferenças funcionais

31
interespecíficas. Isto nos estimula a estudar a distribuição de NO (mesmo que de
forma indireta) no tronco encefálico de mais uma espécie, principalmente por tal
espécie ter características tão peculiares como seu hábitat e hábitos.
Figura 4.O mocó (Kerodon rupestris) em ambiente natural.Fonte: foto de internet, disponível
em:http://www.flickr.com/photos/21182099@N05/16075565604

32
2. OBJETIVOS
2.1 OBJETIVO GERAL
Mapear os neurônios produtores de NO no tronco encefálico do mocó
2.2 OBJETIVOS ESPECÍFICOS
Delimitar, por imunoistoquímica contra NOS, e por histoquímica para
NADPHd os locais onde estão os neurônios que sintetizam NO no tronco
encefálico de mocó;
Qualificar os núcleos pela densidade e intensidade de marcação dos
neurônios imunorreativos ao NOS.

33
3. MATERIAL E MÉTODOS
3.1 ANIMAIS
Os animais utilizados neste projeto são os mesmos utilizados em outros
projetos previamente aprovados pelo IBAMA: licença SISBIO no. 42960-1 de
10/03/2014 e pelo Comitê de Ética em Uso Animal (CEUA-UFRN): protocolo no
004/2014 (Anexo).
Foram utilizados sete mocós adultos quatro machos e três fêmeas, que pela
pouca idade, não foi visualizado dimorfismo sexual, sendo feitas analises
histológicas, para posterior identificação dos sexos, foram capturados em cidades do
interior do Rio Grande do Norte e mantidos em quarentena na fazenda SAMISA de
propriedade da UFRN no município de Extremoz-RN. O espaço onde os animais
foram colocados consiste de uma sala ampla, com porta e janelas gradeadas,
permitindo o acesso a iluminação e ventilação natural. O piso é revestido com
vegetação rasteira e pedras, simulando o habitat natural do animal, com agua a
vontade, sendo alimentados com frutas e verduras.
Os encéfalos aqui estudados, bem como as demais partes dos animais,
serão também utilizados em outras pesquisas e no ensino.
3.2 PROCEDIMENTOS
Os animais foram submetidos às etapas de perfusão, remoção e microtomia
do encéfalo, além das técnicas de Nissl, imunoistoquímica e histoquímica.
3.2.1 Perfusão
Os sete animais utilizados neste trabalho foram anestesiados com ketamina
e xilazina via intramuscular na dosagem de 20mg/kg e 2mg/kg respectivamente. Ao
atingir o plano anestésico, cada animal foi submetido à perfusão transcardíaca, que
compreende os seguintes passos:

34
1. Posicionamento do animal em decúbito dorsal sobre uma tela de arame com
um ponto de água.
2. Toracotomia, com incisão de pele, músculos e arco costal, sendo estes
removidos em bloco, para exposição do coração.
3. Cardiopunção no ventrículo esquerdo, utilizando uma agulha de 17 mm x 1,5
mm, a qual é direcionada para a aorta, seguindo-se uma incisão no átrio
direito. A agulha é conectada a uma bomba peristáltica (Cole-Parmer),
passando-se 300 ml de solução salina a 0,9% em tampão fosfato 0,1M, pH 7,4
com heparina (Parinex, Hipolabor, 2ml/1000 ml de solução salina), seguido de
700 ml de solução paraformaldeído 4% em tampão fosfato 0,1M, pH 7.4.
3.2.2 Remoção do encéfalo e microtomia
Após concluída a perfusão, os animais foram posicionados no aparelho
estereotáxico para roedores. Depois de fazer uma incisão longitudinal na pele e
rebatê-la lateralmente, foi feita a limpeza da superfície óssea, facilitando a
visualização do bregma e do lambda, os quais foram nivelados na mesma altura,
ajustando-se a barra dos incisivos, padronizando assim o plano de corte coronal
para todos os animais. Após anotação das coordenadas do bregma e lambda, o
osso da calota craniana foi removido com o uso de uma broca expondo-se o
encéfalo. Ainda no aparelho estereotáxico, o encéfalo foi seccionado em três blocos,
através de duas secções coronais: uma no nível do bregma e outra no nível do
lambda. Os blocos foram retirados delicadamente para evitar danos (Fig. 5) e em
seguida armazenados em uma solução de sacarose a 30% em tampão fosfato 0,1M,
pH 7,4 a 4°C até serem submetidos a microtomia. Os encéfalos foram congelados
por gelo seco e seccionados em micrótomo de deslizamento, obtendo-se secções
coronais de 30 µm. Estas foram coletadas em um meio líquido de tampão fosfato
0,1M, pH 7,4 distribuídas sequencialmente em 6 compartimentos, de maneira cíclica
e sequenciada, de modo a manter a distância de aproximadamente 180 µm entre
uma secção e a outra imediatamente seguinte de um mesmo compartimento. Os
cortes de um compartimento foram imediatamente montados em lâminas de vidro
gelatinizadas e submetidas à coloração pelo método de Nissl para permitir uma

35
melhor demarcação das estruturas. Os cortes dos demais compartimentos foram
transferidos para solução anticongelante e conservados a -20°C para utilização.
Figura 5. Encéfalo do mocó em vista ventral (A), lateral (B) e dorsal (C). Escala de régua em
centímetros. Fonte: arquivo pessoal
3.2.3 Imunoistoquímica
A técnica consistiu de uma imunoperoxidase padrão pelo método ABC, onde
após quatro lavagens com duração de dez minutos cada em tampão fosfato 0,1M
(PB), pH 7,4, os cortes free floating foram submetidos a um pré-tratamento com uma
solução de peróxido de hidrogênio a 0,03% em PB 0,1M durante 20 minutos em
agitador orbital para inativação da peroxidase endógena. Após mais quatro lavagens
de dez minutos, os cortes então foram incubados em tampão fosfato contendo
tampão Triton X-100 a 0,4% (TXPB) com anticorpo primário anti-NOS feito em
camundongo (1:1000, Sigma) e soro normal de cabra a 2% (Jackson Laboratories)
permanecendo em incubação por 16 a 24 horas em agitador orbital. No dia seguinte,
os cortes foram lavados em PB por quatro vezes e incubados em anticorpo
secundário biotilinado anti-camundongo (Jackson Laboratories) feito em cabra
diluído a 1:1000 e TXPB por 2 horas em agitador orbital. Os cortes foram novamente

36
lavados em PB e incubados em solução avidina-biotina-peroxidase (kit ABC, Vector,
1:100) diluídos em TXPB a 0,4% contendo cloreto de sódio (NaCl) por noventa
minutos. Em seguida, os cortes foram lavados e para a visualização da reação, as
secções foram postas em meio contendo PB e H2O2 0,03% como substrato e o
cromógeno diaminobenzidina (DAB) 0,05% por aproximadamente 10 minutos. A
reação foi parada com lavagens em PB em agitador orbital. Os cortes finalmente
foram montados em lâminas gelatinizadas, mergulhados em tetróxido de ósmio
0,05% para intensificar a reação, desidratadas em uma sequência crescente de
alcoóis e deslipidicados em xilol e cobertos com ERV-Mount e lamínula.
3.2.4 Método de Nissl
A coloração pelo método de Nissl é constituída por uma série de etapas que
se iniciam com a desidratação do tecido passando-o em concentrações crescentes
de alcoóis etílicos (70% - 1 vez, por 1h, 95% - 2 vezes, 3 minutos cada, 100% - 2
vezes, 3 minutos cada). Posteriormente são deslipidificados em dois recipientes com
xilol, por 3 e 30 minutos, nessa ordem. Na sequência, o tecido é reidratado em
concentrações decrescentes de alcoóis (mesmo processo anterior, mas no sentido
inverso), submergido em água destilada por 2 minutos, colocado 1 minuto na
solução tionina a 0,25%. Por fim os cortes foram rapidamente submergidos e
emergidos por 1 minuto em água destilada e novamente desidratados e
deslipidificados como descrito anteriormente, sendo acrescentada mais uma etapa
em álcool 95%. Os cortes foram deslipidificados em xilol em dois recipientes (2 e 4
minutos, respectivamente) e finalmente cobertos com lamínula utilizando como meio
de montagem o ERV-Mount.
3.2.5 Histoquímica
Foi utilizado o método histoquímico para marcação de NADPH-diaforase.
Esse processo iniciou-se diluindo 45 mg de azul de nitrotetrazólio em 1,5 ml de
dimetil sulfóxido, formando a solução I. Em seguida acrescentou-se 900 mg de ácido
málico em um béquer contendo 150 ml de tampão TRIS 0,05M, ajustando o pH da
solução resultante para 8,0. Após o ajuste do pH foi acrescentado 150 mg do

37
composto β-NADP e 60 mg de cloreto de manganês, formando a solução II. Após a
homogeinização desta última, as duas soluções foram misturadas, acrescentando
em seguida 4,5 ml de Triton-X100. Uma vez que a solução resultante é fotossensível
ela foi sempre protegida da luz. As secções foram imersas nessa solução e
colocadas para reagir sob agitação manual constante a 39°C em banho maria e
protegidos da luz. A duração tem uma alta variabilidade que depende da
temperatura e da iluminação passando aproximadamente quatro horas em reação.
No entanto, passado um período de 90 minutos os cortes foram observados em
microscópio óptico para verificação do grau de marcação em intervalos de 10
minutos. Após a reação foi feito 3 lavagens sucessivas em PB de 3 minutos cada e
por fim as lâminas foram cobertas com lamínula usando ERV-Mount.
3.2.6 Análise das imagens
As secções do tronco encefálico coradas pelo método de Nissl e as
submetidas à imunoistoquímica e histoquímica foram examinadas ao microscópio
óptico (Nikon eclipse Ci-L) em campo claro. As áreas que contem neurônios
imunorreativos a NOS foram mapeadas e classificadas qualitativamente em
densidade de neurônios imunorreativos e nível de intensidade de marcação de
acordo com as figuras 6 e 7 utilizando como base o atlas de Paxinos e Watson
(2007) de rato, Paxinos e colaboradores (2012) de sagui e Franklin e Paxinos (2008)
de camundongo para a correlação anatômica das estruturas do tronco encefálico do
mocó.
Imagens digitais foram obtidas de secções representativas usando uma
videocâmera digital (Nikon DS-Ri1). As imagens foram analisadas, corrigidas
minimamente para brilho, contraste e documentadas em fotomicrografias, charters
construídos utilizando Canvas (versão 12.0; ACD Systems of America, Inc.; 2010).

38
Figura 6. Exemplos das diferentes intensidades de marcação das células imunomarcadas
para NOS.
Figura 7. Exemplos de quantificação subjetiva da densidade das células imunomarcadas
para NOS dentro de cada núcleo.

39
4. RESULTADOS
No presente estudo a imunoistoquímica para NOS e a histoquímica para a
NADPH-diaforase (NADPHd) foram utilizadas. Encontramos neurônios
imunorreativos para NOS (NOS-IR) e marcadas com NADPHd ao longo de todo
tronco encefálico. A imunoistoquímica apresentou uma coloração castanha bem
definida onde neurônios NOS-IR apresentaram uma marcação que definiu bem o
citoplasma, assim como a NADPHd, que apresentou uma coloração azul com
intensidade que variaram dependendo da atividade da enzima.
O padrão de distribuição dos neurônios marcados pelas técnicas de
imunoistoquímica e histoquímica foi semelhante em todos os animais, como exposto
na seção 4.2 histoquímica. A descrição da distribuição baseia-se principalmente na
imunoistoquímica e como listada no quadro 1.
Quadro 1. Densidade e intensidade dos núcleos imunorreativos a NOS.
Núcleos e Regiões Densidade Intensidade
Núcleo pré-comissural +++ +++
Substância negra
- parte reticulada +/- +
- parte compacta, camada dorsal + +++
- parte lateral ++ ++
Núcleo magnocelular da comissura posterior +++ +++
Núcleo da comissura posterior + ++
Núcleo paracomissural da comissura posterior + +++
Núcleo retroparafascicular +++ +++
Núcleo intersticial de Cajal +/- ++
Substância cinzenta periaquedutal
- parte pleomórfica ++ ++
- lateral +++ ++
- ventrolateral +++ ++
- dorsolateral +++ +++
- dorsomedial ++ +++
- supraoculomotor +/- +++

40
- supraoculomotor cap +/- +++
Núcleo litóide + +++
Área tegmental ventral
- parte rostral ++ +++
- núcleo parabraquial pigmentado + +++
- núcleo paranigral + ++
Núcleo pré-tectal medial + +++
Núcleo pré-tectal anterior
- parte dorsal + ++
- parte ventral + ++
Núcleo pré-tectal posterior ++ ++
Núcleo pré-tectal olivar ++ ++
Núcleo do trato óptico + ++
Núcleo de Darkschewitsch ++ +++
Trato óptico acessório
- núcleo terminal medial +/- +++
- núcleo terminal dorsal + +++
- núcleo terminal lateral +/- +++
Campo pré-rubral + +++
Núcleo pararubral -/+ ++
Campo retrorubral + ++
Núcleo interfascicular + +++
Núcleo pré-Edinger-Westphal +/- ++
Núcleo acessório medial do nervo oculomotor +/- +++
Colículo superior
- camada zonal do colículo superior ++ +++
- camada intermediária cinza ++ ++
- camada do nervo óptico do colículo superior ++ ++
- camada cinza superficial ++ +++
- camada cinza profunda ++ +++
Núcleo peripeduncular +/- +++
Núcleo rubro

41
- parte parvocelular +/- ++
- parte magnocelular +/- ++
Núcleo interpeduncular
- subnúcleo rostral +++ +++
- subnúcleo caudal ++ +++
- subnúcleo intermediário ++ +++
- subnúcleo lateral ++ +++
- subnúcleo dorsolateral + ++
Núcleo sub-braquial +/- +++
Núcleo do braço do colículo inferior +++ ++
Córtex externo do colículo inferior + ++
Córtex dorsal do colículo inferior + ++
Núcleo linear rostral da rafe ++ +++
Núcleo dorsal da rafe +/- ++
- parte dorsal +/- ++
- parte ventral +/- ++
- parte lateral +++ +++
Núcleo mediano da rafe +/- ++
Núcleo posterodorsal da rafe ++ +++
Núcleo paramediano da rafe + +++
Núcleo magno da rafe + ++
Núcleo obscuro da rafe ++ +++
Núcleo troclear ++ +++
Núcleo tegmental pedunculopontino +++ +++
Núcleo retrorubral ++ ++
Núcleo rabdóide + +++
Núcleo pontino + ++
Núcleo reticular pontino, parte oral + ++
Núcleo reticular pontino, parte caudal + ++
Núcleo reticular pontino, parte ventral + ++
Núcleo parabigeminal + ++
Área pré-cuneiforme + +++

42
Núcleo reticulotegmental da ponte + ++
- parte lateral + +++
Núcleo tegmental subpeduncular + +++
Núcleo tegmental microcelular + +++
Núcleo tegmental posterodorsal + +
Núcleo triangular do lemnisco lateral + +
Núcleo intermediário do lemnisco lateral + +
Núcleo ventral do lemnisco lateral + ++
Núcleo medial paralemniscal + +++
Núcleo perilemniscal, parte ventral +/- ++
Núcleo parabraquial medial + ++
Núcleo parabraquial lateral + ++
- parte interna + ++
- parte ventral + ++
Núcleo periolivar
- medioventral +/- ++
- lateroventral +/- ++
Núcleo tegmental laterodorsal + +++
- parte ventral + +++
Área tegmental dorsomedial + ++
Núcleo principal sensorial do trigêmeo
- parte ventrolateral + ++
- parte dorsomedial + ++
Núcleo do corpo trapezóide +/- ++
Núcleo subcerúleus
- parte alfa + ++
- parte dorsal + ++
- parte ventral + ++
Núcleo coclear ventral
- parte anterior +/- ++
- parte posterior + +++
Locus cerúleos ++ +++

43
Núcleo trigêminal mesencefálico + +++
Substancia cinzenta central
- parte alfa + ++
- parte beta + ++
Núcleo vestibular superior + +++
Substância cinzenta da ponte +/- +
Núcleo parabducente ++ +++
Núcleo abducente ++ +++
Núcleo gigantocelular reticular + ++
- parte alfa ++ +++
Núcleo reticular intermediário + +++
- parte alfa + +++
Núcleo vestibular medial
- parte magnocelular + +++
- parte parvocelular + +++
Núcleo reticular parvocelular + ++
- parte alfa + ++
Núcleo prepósito ++ +++
-parte magnocelular ++ +++
Núcleo paragigantocelular dorsal ++ ++
Núcleo paragigantocelular lateral
- parte alfa ++ +++
- parte externa ++ +++
Núcleo vestibular lateral ++ ++
Núcleo vestibular espinal + +++
Núcleo X ++ +++
Núcleo salivatório inferior + ++
Região matriz da medula + +++
Núcleo espinal trigeminal
- parte dorsomedial + ++
- parte oral + ++
- parte interpolar + ++

44
- parte caudal ++ +++
- camada gelatinosa +++ +++
Zona de transição trigeminal-solitário + ++
Núcleo do trato solitário
- parte intermediaria + ++
- parte lateral + ++
- parte medial + +++
- parte ventral + ++
- parte dorsomedial + +++
Núcleo motor dorsal do vago +/- ++
Núcleo reticular lateral ++ +++
Núcleo reticular medular
- parte dorsal + +++
- parte ventral + +++
Núcleo cuneado ++ +++
- parte rotunda ++ +++
- parte externa ++ +++
Núcleo grácil +/- +++
Oliva inferior, núcleo medial
- subnúcleo A + +++
- subnúcleo B + +++
- subnúcleo C + +++
Núcleo retroambíguo + +++
Núcleo intersticial da medula +/- +++
Regiões marcadas com NOS-IR. Nível da densidade dos neurônios nos núcleos: +++ Alta,
++ Moderada, + Baixa, +/- Muito baixa; e da intensidade da marcação:+++ Forte, ++ Média,
+ Fraca (Baseado em padronização subjetiva de densidade e intensidade de neurônios
corados pelos métodos de imunoistoquímica em cada núcleo. Ver figuras 5 e 6).

45
Figura 08. Reconstrução esquemática das secções coronais do encéfalo do mocó ilustrando
a distribuição dos neurônios NOS-IR do tronco encefálico e de fotomicrografias de cortes
submetidos à coloração de Nissl no nível correspondente, distando uma secção do seu
subsequente por 1620 µm. Representação do nível mais rostral em (A) e do nível mais
caudal em (M). Cada ponto representa um neurônio. Para abreviações ver lista de
abreviações.

46
Figura 08. Continuação

47
Figura 08. Continuação

48
4.1 IMUNOISTOQUÍMICA
Cranialmente os primeiros neurônios NOS-IR surgiram no nível dos núcleos
mamilares mediais e lateral. Pela localização e relações, identificamos esse
grupamento como o núcleo pré-comissural (PrC) que apresentou neurônios
marcados em toda sua extensão, apresentando densidade alta e marcação forte
com uma neurópila pronunciada (Fig. 8 A; 9 A e B).
O núcleo pré-tectal medial (MPT) apresentou neurônios NOS-IR com
densidade baixa e forte marcação. O núcleo pré-tectal anterior (APT) e suas partes:
dorsal (APTD) e parte ventral (APTV) apresentaram neurônios NOS-IR com
densidade baixa das células e de marcação média (Fig. 8 A e B; 9 A,D-E). O núcleo
pré-tectal posterior (PPT) e o núcleo pré-tectal olivar (OPT) apresentaram densidade
de neurônios NOS-IR moderada e média marcação (Fig. 8 B).
A substância cinzenta periaquedutal, parte pleomórfica (PAG), apresentou
uma densidade moderada de neurônios NOS-IR e marcação média em todas as
células, que ficaram concentradas na região medial da PAG (Fig. 8 A; 9 A e C). Sua
parte lateral (LPAG) e ventrolateral (VLPAG) com densidade de neurônios NOS-IR
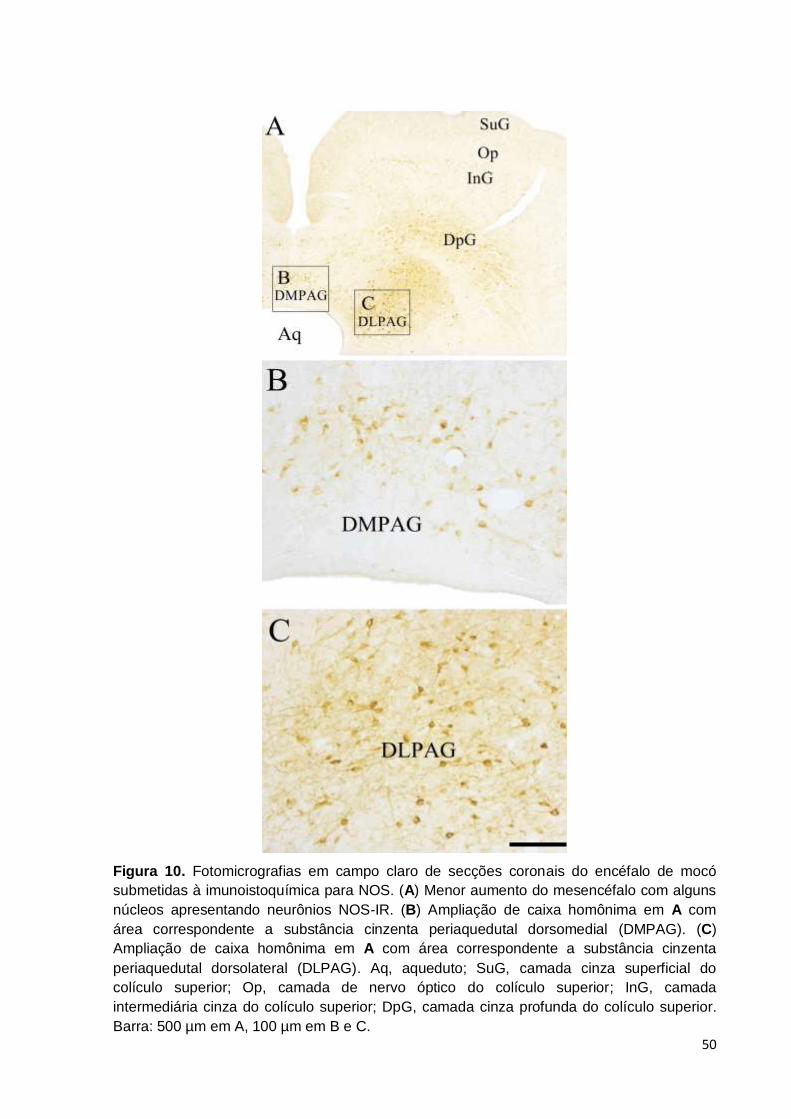
alta com marcação média (Fig. 8 B-D). A partedorsomedial (DMPAG) e dorsolateral
(DLPAG) teve densidade alta e marcação forte (Fig. 8 B-D; 10 A-C). As partes
supraoculomotor (Su3) e supraoculomotor cap (Su3C) exibiram neurônios NOS-IR
com muito baixa densidade e marcação forte.
No colículo superior, neurônios NOS-IR foram encontrados em diversas
regiões. Sendo divididas em 7 camadas, foram observados neurônios NOS-IR na:
camada zonal do colículo superior (Zo) que demonstrou uma densidade moderada e
uma intensidade forte de marcação. A camada cinza superficial (SuG) apresentou
neurônios NOS-IR de densidade moderada e com marcação média dessas células
(Fig. 8 B-D; 10 A).
A camada do nervo óptico do colículo superior (Op) mostrou um padrão de
densidade moderado e uma coloração média em todos os níveis visualizados. A
camada intermediária cinza (InG), camada extensa onde os neurônios NOS-IR
tiveram densidade moderada, com intensidade de coloração média. A camada cinza
profunda (DpG) apresentou densidade moderada de neurônios NOS-IR com
marcação forte com intensa neurópila (Fig. 8 C e D; 10 A).

49
Figura 9. Fotomicrografias em campo claro de secções coronais do encéfalo de mocó
submetidas à imunoistoquímica para NOS. (A) Menor aumento do mesencéfalo com alguns
núcleos apresentando neurônios NOS-IR. (B) Ampliação de caixa homônima em A com
área correspondente ao núcleo pré-comissural (PrC). (C) Ampliação de caixa homônima em
A com área correspondente a substância cinzenta periaquedutal (PAG). (D) Ampliação de
caixa homônima em A com área correspondente a parte dorsal do núcleo pré-tectal anterior
(APTD). (E) Ampliação de caixa homônima em A com área correspondente a parte ventral
do núcleo pré-tectal anterior (APTV). 3V, terceiro ventrículo. Barra: 500 µm em A, 100 µm
em B, C, D e E.
50
Figura 10. Fotomicrografias em campo claro de secções coronais do encéfalo de mocó
submetidas à imunoistoquímica para NOS. (A) Menor aumento do mesencéfalo com alguns
núcleos apresentando neurônios NOS-IR. (B) Ampliação de caixa homônima em A com
área correspondente a substância cinzenta periaquedutal dorsomedial (DMPAG). (C)
Ampliação de caixa homônima em A com área correspondente a substância cinzenta
periaquedutal dorsolateral (DLPAG). Aq, aqueduto; SuG, camada cinza superficial do
colículo superior; Op, camada de nervo óptico do colículo superior; InG, camada
intermediária cinza do colículo superior; DpG, camada cinza profunda do colículo superior.
Barra: 500 µm em A, 100 µm em B e C.

51
A parte reticulada da substância negra (SNR) apresentou neurônios NOS-IR
com densidade muito baixa em todos os níveis observados, sua marcação foi fraca
(Fig. 8 A-C; 11 A e C). A parte lateral da substância negra (SNL) apresentou
densidade moderada e intensidade média (Fig. 8 A-C; 11 A e B). A parte compacta,
camada dorsal (SNCD), mostrou uma densidade baixa e uma forte marcação (Fig. 8
B e C).
O núcleo do trato óptico (OT) mostrou densidade baixa de neurônios NOS-IR
e marcação média (Fig. 12 A e B). O núcleo magnocelular da comissura posterior
(MCPC) apresentou neurônios NOS-IR com alta densidade e com uma marcação
intensa desses neurônios (Fig. 12 A e C). As células NOS-IR do núcleo da
comissura posterior (PCom) demonstraram uma densidade baixa de neurônios NOS-
IR e de marcação média. O núcleo paracomissural da comissura posterior (PaC)
apresentou neurônios NOS-IR com densidade baixa e marcação forte.
O núcleo retroparafascicular (RPF) apresentou neurônios NOS-IR com
densidade alta e coloração forte (Fig. 12 A e C), O núcleo intersticial de Cajal (InC)
demonstrou uma densidade muito baixa de neurônios NOS-IR com marcação média
(Fig. 8 B e C). O núcleo litoide (Lth) apresentou neurônios NOS-IR com densidade
baixa e com forte marcação.
Na área tegmental ventral (VTA) encontramos neurônios NOS-IR no núcleo
parabraquial pigmentado (PBP) de densidade baixa e com coloração forte (Fig. 8 A-
B e E). Os núcleos: núcleo parabraquial medial (MPB); núcleo parabraquial lateral
(LPB); parte interna (LPBI) e parte ventral (LPBV) demonstraram o mesmo padrão
de densidade baixa e intensidade média. Na parte rostral (VTAR) com densidade
moderada e forte marcação das células. E o núcleo paranigral (PN) apresentou
neurônios com densidades baixas e com uma marcação média (Fig. 8 C).
O núcleo de Darkschewitsch (DK) apresentou neurônios NOS-IR com
densidade moderada, e com marcação forte das células (Fig. 8 B e C; 12 A e D). O
núcleo terminal medial do trato óptico acessório (MT) e terminal lateral do trato
óptico acessório (LT) apresentaram neurônios NOS-IR com densidade muito baixa
em toda sua extensão, com forte marcação celular (Fig. 8 A e B). O núcleo terminal
dorsal do trato óptico acessório (DT) apresentou neurônios NOS-IR de densidade
alta e marcação forte.

52
Figura 11. Fotomicrografias em campo claro de secções coronais do encéfalo de mocó
submetidas à imunoistoquímica para NOS. (A) Menor aumento do mesencéfalo com alguns
núcleos apresentando neurônios NOS-IR. (B) Ampliação de caixa homônima em A com
área correspondente a parte lateral da substância negra (SNL). (C) Ampliação de caixa
homônima em A com área correspondente a parte reticulada da substância negra (SNR).
cp, pedúnculo cerebral. Barra: 500 µm em A, 100 µm em B e C.

53
Figura 12. Fotomicrografias em campo claro de secções coronais do encéfalo de mocó
submetidas à imunoistoquímica para NOS. (A) Menor aumento do mesencéfalo com alguns
núcleos apresentando neurônios NOS-IR. (B) Ampliação de caixa homônima em A com
área correspondente ao núcleo do trato óptico (OT). (C) Ampliação de caixa homônima em
A com área correspondente ao núcleo magnocelular da comissura posterior (MCPC) e o
núcleo retroparafascicular (RPF). (D) Ampliação de caixa homônima em A com área
correspondente ao núcleo de Darkschewitsch (DK). Aq, aqueduto; cp, pedúnculo cerebral.
Barra: 500 µm em A, 100 µm em B, C e D.
O campo pré-rubral (PR), apresentou neurônios NOS-IR com densidade
baixa em toda a extensão do núcleo e com forte marcação celular (Fig. 8 G e H). O
núcleo pararubral (PaR) e o campo retrorubral (RRF) apresentaram intensidade
média e densidade muito baixa no PaR e baixa no RRF (Fig. 8 C; 13 A e B). O
núcleo interfascicular (IF) apresentou neurônios NOS-IR de densidade baixa com
marcação forte dessas células (Fig. 8 C). O núcleo Pré-Edinger-Westphal (PrEW)
demonstrou neurônios NOS-IR de densidade muito baixa e de marcação média. O
núcleo acessório medial do nervo oculomotor (MA3), demonstrou neurônios NOS-IR
de densidade muito baixa e de marcação forte (Fig. 8 C).

54
O núcleo peripeduncular (PP) demonstrou neurônios NOS-IR com densidade
muito baixa e uma marcação forte desses neurônios (Fig. 8 C). A parte parvocelular
do núcleo rubro (RPC) e a parte magnocelular (RMC) apresentaram neurônios NOS-
IR de densidade muito baixa em todas suas porções e intensidade média.O núcleo
interpeduncular (IP) em seu subnúcleo rostral (IPR) teve a maior densidade de
neurônios NOS-IR entre os núcleos mesencefálicos e todos com marcação alta. Já o
subnúcleo caudal (IPC) mostrou densidade moderada, permanecendo com o mesmo
padrão de coloração do subnúcleo rostral (Fig. 8 C). Os subnúcleos intermediário
(IPI) e lateral (IPL) mostraram densidade moderada e forte marcação (Fig. 8 C; 13 A
e C) já o subnúcleo dorsolateral (IPDL), o menor dos subnúcleos, demonstrou uma
baixa densidade e uma coloração média, nos encéfalos observados.
O núcleo sub-braquial (SubB) apresentou neurônios NOS-IR de densidade
muito baixa na porção cranial, com marcação forte. No núcleo do braço do colículo
inferior (BIC) foram encontrados neurônios NOS-IR com densidades altas e
coloração média (Fig. 8 D).
Neurônios NOS-IR foram encontrados em diversos núcleos da rafe. O
núcleo linear rostral da rafe (RLi) apresentou neurônios NOS-IR com densidade
moderada e com forte marcação das células (Fig. 8 A). O núcleo dorsal da rafe (DR)
demonstrou uma densidade muito baixa de neurônios NOS-IR com marcação média.
A parte lateral do núcleo dorsal da rafe (DRL) apresentou marcação forte
em todos os níveis e com alta densidade (Fig. 8 D; 14 A e B; 15 A e C). Assim como
o núcleo posterodorsal da rafe (PDR) (Fig. 8 D; 14 A e B). A parte dorsal do núcleo
dorsal da rafe (DRD) e a parte ventral do núcleo dorsal da rafe (DRV) mostraram
neurônios NOS-IR com densidade muito baixa e marcação média (Fig. 8 D; 14 A e
B).

55
Figura 13. Fotomicrografias em campo claro de secções coronais do encéfalo de mocó
submetidas à imunoistoquímica para NOS. (A) Menor aumento do mesencéfalo com alguns
núcleos apresentando neurônios NOS-IR. (B) Ampliação de caixa homônima em A com
área correspondente ao campo retrorubral (RRF). (C) Ampliação de caixa homônima em A
com área correspondente ao núcleo interpeduncular rostral (IPR), caudal (IPC),
intermediário (IPI) e lateral (IPL). Barra: 500 µm em A, 100 µm em B e C.

56
Figura 14. Fotomicrografias em campo claro de secções coronais do encéfalo de mocó
submetidas à imunoistoquímica para NOS. (A) Menor aumento do mesencéfalo com alguns
núcleos apresentando neurônios NOS-IR. (B) Ampliação de caixa homônima em A com
área correspondente a parte lateral do núcleo dorsal da rafe (DRL), ao núcleo posterodorsal
da rafe (PDR), a parte dorsal do núcleo dorsal da rafe (DRD) e a parte ventral do núcleo
dorsal da rafe (DRV). (C) Ampliação de caixa homônima em A com área correspondente ao
núcleo retrorubral (RR). (D) Ampliação de caixa homônima em A com área correspondente
ao núcleo rabdóide (Rbd) e ao núcleo mediano da rafe (MnR). Barra: 500 µm em A, 100 µm
em B, C e D.
O núcleo mediano da rafe (MnR) exibiu densidade de neurônios NOS-IR
muito baixa e marcação média (Fig. 8 D; 14 A e D). O núcleo paramediano da rafe
(PMnR) apresentou densidade de neurônios NOS-IR baixa e marcação forte. Por fim
os dois núcleos mais caudais da rafe, núcleo magno da rafe (RMg), apresentou
baixa densidade e uma marcação média (Fig. 8 H e E), enquanto o núcleo obscuro
da rafe (ROb) mostrou uma moderada densidade e uma forte marcação (Fig. 8 I e J).
O córtex externo (ECIC) e o córtex dorsal (DCIC) do colículo inferior
apresentaram neurônios NOS-IR com densidade baixa e com marcação de média
(Fig. 8 D; 15 A e B). O núcleo tegmental pedunculopontino (PTg) apresentou

57
densidade alta e um forte marcação (Fig. 8 D; 15 A e D). O núcleo retrorubral (RR)
com densidade moderada e marcação média (Fig. 15 A e C). O núcleo rabdóide
(Rbd) e o núcleo troclear (4N) mostraram densidade baixa e marcação forte (Fig. 8
D; 15 A e D).
O núcleo pontino (Pn), bem como o núcleo reticular pontino e suas divisões:
parte oral (PnO), parte ventral (PnV) e parte caudal (PnC), apresentaram uma
densidade baixa e uma intensidade média de marcação (Fig. 8 C-E). O núcleo
parabigeminal (PBG) apresentou baixa densidade e média marcação (Fig. 8 D), já a
área pré-cuneiforme (PrCnF) apresentou a mesma densidade do núcleo anterior
com uma forte intensidade de marcação (Fig. 8 D). O núcleo reticulotegmental da
ponte (RtTg) mostrou uma baixa densidade e uma média intensidade, a sua parte
lateral (RtTgL) mostrou o mesmo padrão de densidade, porém com uma intensidade
de marcação forte.
Os núcleos tegmental subpeduncular (SPTg) e o tegmental microcelular
(MiTg) apresentaram baixa densidade de neurônios e intensidade de marcação forte
(Fig. 8 D), diferindo do núcleo tegmental posterodorsal (PDTg) quanto a intensidade
que neste núcleo é fraca (Fig. 8 E). Densidade baixa e uma intensidade fraca de
marcação foi observada nos núcleos triangular do lemnisco lateral (TrLL) e
intermediário do lemnisco lateral (ILL). No núcleo ventral do lemnisco lateral (VLL) a
densidade foi idêntica aos anteriores com uma intensidade de marcação média. O
núcleo medial paralemniscal (MPL) apresentou a mesma densidade dos três núcleos
anteriores e uma intensidade de marcação forte. Por fim, o núcleo perilemniscal,
parte ventral (PLV) apareceu com uma densidade de neurônios NOS-IR muito baixa
e intensidade média.
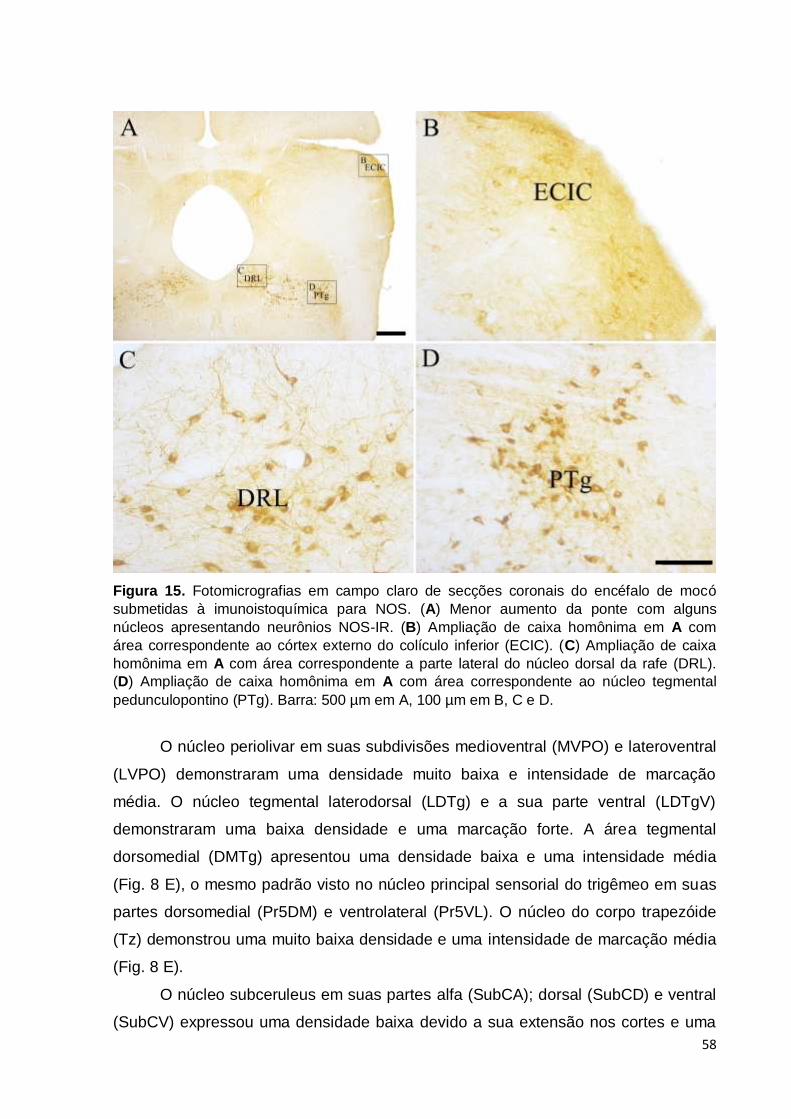
58
Figura 15. Fotomicrografias em campo claro de secções coronais do encéfalo de mocó
submetidas à imunoistoquímica para NOS. (A) Menor aumento da ponte com alguns
núcleos apresentando neurônios NOS-IR. (B) Ampliação de caixa homônima em A com
área correspondente ao córtex externo do colículo inferior (ECIC). (C) Ampliação de caixa
homônima em A com área correspondente a parte lateral do núcleo dorsal da rafe (DRL).
(D) Ampliação de caixa homônima em A com área correspondente ao núcleo tegmental
pedunculopontino (PTg). Barra: 500 µm em A, 100 µm em B, C e D.
O núcleo periolivar em suas subdivisões medioventral (MVPO) e lateroventral
(LVPO) demonstraram uma densidade muito baixa e intensidade de marcação
média. O núcleo tegmental laterodorsal (LDTg) e a sua parte ventral (LDTgV)
demonstraram uma baixa densidade e uma marcação forte. A área tegmental
dorsomedial (DMTg) apresentou uma densidade baixa e uma intensidade média
(Fig. 8 E), o mesmo padrão visto no núcleo principal sensorial do trigêmeo em suas
partes dorsomedial (Pr5DM) e ventrolateral (Pr5VL). O núcleo do corpo trapezóide
(Tz) demonstrou uma muito baixa densidade e uma intensidade de marcação média
(Fig. 8 E).
O núcleo subceruleus em suas partes alfa (SubCA); dorsal (SubCD) e ventral
(SubCV) expressou uma densidade baixa devido a sua extensão nos cortes e uma

59
marcação média a forte (Fig. 8 E). Já o núcleo coclear ventral na parte anterior
(VCA), de densidade muito baixa e intensidade média, e a parte posterior (VCP),
com densidade baixa e intensidade de marcação forte. O locus cerúleos (LC)
mostrou uma densidade moderada, pois é um núcleo pequeno, com uma
intensidade forte. O núcleo do trato mesencefálico do trigêmeo (Me5) demonstrou
uma densidade baixa e uma intensidade forte (Fig. 8 E).
As partes alfa (CGA) e beta (CGB) da substância cinzenta central tiveram
densidade baixa e marcação média (Fig. 8 E). O núcleo vestibular superior (SuVe)
apresentou densidade baixa e intensidade de marcação forte (Fig. 8 F). A substância
cinzenta da ponte (CGPn) teve uma densidade muito baixa e uma marcação fraca
(Fig. 8 E). Os núcleos parabducente (Pa6) e abducente (6N) expressaram uma
densidade moderada e uma intensidade de marcação forte. Núcleo gigantocelular
reticular (Gi) apresentou um conjunto grande de neurônios espalhados em sua
extensão com marcação média no geral e marcação forte em sua parte alfa (GiA)
(Fig. 8 F - H e J).
O núcleo reticular intermediário (IRt) e sua parte alfa (IRtA), demonstraram
uma baixa densidade de neurônios e uma forte marcação (Fig. 8 F - J e L). O núcleo
vestibular medial em suas partes magnocelular (MVeMC) e parvocelular (MVePC),
com densidade baixa e uma marcação forte (Fig. 8 F - I; 17 A e B). O núcleo reticular
parvocelular (PCRt) bem como sua parte alpha (PCRtA), demonstraram densidade
baixa e uma intensidade média (Fig. 8 F - J). O núcleo preposito (Pr), bem como sua
parte magnocelular (PrMC) demonstrou uma densidade moderada e uma forte
intensidade de marcação (Fig. 8 F - I; 17 A e C; 18 A e D) respectivamente.

60
Figura 16. Fotomicrografias em campo claro de secções coronais do encéfalo de mocó
submetidas à imunoistoquímica para NOS. (A) Menor aumento da ponte com alguns
núcleos apresentando neurônios NOS-IR. (B) Ampliação de caixa homônima em A com
área correspondente ao locus cerúleos (LC). (C) Ampliação de caixa homônima em A com
área correspondente ao núcleo trigeminal mesencefálico (Me5). (D) Ampliação de caixa
homônima em A com área correspondente ao núcleo subceruleos, parte ventral (SubCV).
Barra: 500 µm em A, 100 µm em B, C e D.
O núcleo paragigantocelular dorsal (DPGi), com densidade moderada e
intensidade média (Fig. 8 G e H; 17 A e D), é seguido pelo núcleo paragigantocelular
lateral (LPGi) e sua parte alfa (LPGiA) e externa (LPGiE) onde ambas monstram
uma densidade moderada e uma forte intensidade de marcação (Fig. 8 I). O núcleo
vestibular lateral (LVe) apresentou densidade moderada e média marcação (Fig. 8
G). Núcleo vestibular espinal (SpVe), com densidade baixa e forte marcação (Fig. 8
G-I). O núcleo X (X), com densidade moderada e marcação forte (Fig. 8 H e I). O
núcleo salivatório inferior (IS) mostrou densidade baixa e marcação média enquanto
a região matriz da medula (Mx) mostrou uma densidade idêntica ao núcleo anterior e
marcação forte (Fig. 8 H - K).

61
Núcleo espinal trigeminal (Sp5), e suas partes: dorsomedial (DMSp5), oral
(Sp5O) e parte interpolar (Sp5I), com características idênticas de densidade baixa e
média intensidade, variando apenas na sua parte caudal (Sp5C), com densidade
moderada e forte marcação (Fig. 8 F - M; 19 A e C). O núcleo do trato solitário (Sol)
é dividido em muitas partes: intermediária (SolIM), lateral (SolL), medial (SolM),
ventral (SolV) e dorsomedial (SolDM). Todas elas com densidades baixas e
marcação média, porém nas divisões mediais a intensidade alta (Fig. 8 H - M; 18 A e
C). Mesmo padrão para a zona de transição trigeminal-solitário (5Sol). Por fim a
camada gelatinosa do núcleo trigeminal espinal caudal (Ge5), com densidade alta e
forte marcação (Fig. 8 J - M; 19 A e C). O núcleo motor dorsal do vago (10N)
apresentou densidade baixa e coloração media (Fig. 8 J).
O núcleo reticular lateral (LRt), apresentou uma densidade moderada e uma
marcação forte (Fig. 8 J e K; 18 A e D). Núcleo reticular medular e suas partes dorsal
(MdD) e ventral (MdV), demonstraram uma densidade baixa e uma intensidade forte
(Fig. 8 K - M). Núcleo cuneado (Cu), e a sua divisão externa (ECu), expressaram
uma densidade moderada e uma forte intensidade de marcação (Fig. 8 I - M; 18 A e
B). Assim como sua parte rotunda (CuR). O núcleo grácil (Gr) mostrou uma
densidade muito baixa e uma forte marcação (Fig. 8 J - M; 19 A e B). Oliva inferior,
núcleo medial, e seus subnúcleos A (IOA), B (IOB) e C (IOC), com densidade de
marcação baixa e forte marcação (Fig. 8 J e K; 19 A e E). O núcleo retroambíguo
(RAmb), com densidade baixa e forte marcação (Fig. 8 K e L) e o núcleo intersticial
da medula (IB), com densidade muito baixa e marcação forte (Fig. 8 K e L).

62
Figura 17. Fotomicrografias em campo claro de secções coronais do encéfalo de mocó
submetidas à imunoistoquímica para NOS. (A) Menor aumento do bulbo com alguns núcleos
apresentando neurônios NOS-IR. (B) Ampliação de caixa homônima em A com área
correspondente a parte parvocelular do núcleo vestibular medial (MVePC). (C) Ampliação de
caixa homônima em A com área correspondente ao núcleo prepósito (Pr). (D) Ampliação de
caixa homônima em A com área correspondente ao núcleo dorsal paragigantocelular
(DPGi). Barra: 500 µm em A, 100 µm em B, C e D.

63
Figura 18. Fotomicrografias em campo claro de secções coronais do encéfalo de mocó
submetidas à imunoistoquímica para NOS. (A) Menor aumento do bulbo com alguns núcleos
apresentando neurônios NOS-IR. (B) Ampliação de caixa homônima em A com área
correspondente a parte lateral do núcleo cuneado externo (ECu). (C) Ampliação de caixa
homônima em A com área correspondente aos núcleos do trato solitário (Sol). (D)
Ampliação de caixa homônima em A com área correspondente ao núcleo prepósito, parte
magnocelular (PrMc). (E) Ampliação de caixa homônima em A com área correspondente
aos núcleos olivares inferiores (IO). Barra: 500 µm em A, 100 µm em B, C, D e E.

64
Figura 19. Fotomicrografias em campo claro de secções coronais do encéfalo de mocó
submetidas à imunoistoquímica para NOS. (A) Menor aumento do bulbo com alguns núcleos
apresentando neurônios NOS-IR. (B) Ampliação de caixa homônima em A com área
correspondente ao núcleo grácil (Gr). (C) Ampliação de caixa homônima em A com área
correspondente a camada gelatinosa do núcleo trigeminal espinal caudal (Ge5) e ao núcleo
trigeminal espinal, parte caudal (Sp5C). (D) Ampliação de caixa homônima em A com área
correspondente ao núcleo reticular lateral (LRt). Barra: 500 µm em A, 100 µm em B, C e D.

65
4.2 HISTOQUÍMICA
Com técnica de NADPHd vimos padrões de marcações semelhantes ao
apresentados na técnica de imunoistoquímica, como nos diversos subnúcleos do
núcleo interpeduncular (IP) (Fig. 20 A e B).
O núcleo tegmental microcelular (MiTg), com neurônios densos e bem
esparsos, foi facilmente visualizado (Fig. 20 D e F). A parte lateral do núcleo dorsal
da rafe (DRL) e o núcleo posterodorsal da rafe (PDR) diferiram um pouco da
imunoistoquímica pela densidade de neurônios, porém tamanho e formato foram o
mesmo. O locus cerúleos (LC) e o núcleo trigêminal mesencéfalico (Me5)
apresentaram uma marcação NADPHd bem destacada e com neurônios grandes
(Fig. 20 G e H). Outros núcleos com marcação bem visível foram o núcleo grácil (Gr)
com neurônios pequenos e bem claros, a camada gelatinosa do núcleo trigêminal
espinal caudal (Ge5), bem delineado, e o núcleo espinal trigêminal, parte caudal
(Sp5C) com neurônios bem pequenos e mal visualizados devido ao Ge5 (Fig. 20 I e
J).
Em algumas áreas a reação enzimática da NADPH apresentou somente
uma marcação residual mais escuro que a marcação de fundo (background), como
uma neurópila marcada onde não se distinguem citoplasmas. Estas áreas
correspondem a núcleos cujos neurônios tem uma fraca marcação imunoistoquímica
contra NOS, como a parte reticulada da substância negra (SNR).

66
Figura 20. Fotomicrografias em campo claro de secções coronais do encéfalo de mocó
submetidas à histoquímica. (A, C, G, I) Menor aumento de cortes do tronco encefálico com
alguns núcleos apresentando neurônios corados para NADPHd. (B) Ampliação de caixa
homônima em A área correspondente ao núcleo interpeduncular rostral (IPR), e suas
divisões: caudal (IPC), intermediário (IPI) e lateral (IPL), (D) Ampliação de caixa homônima
em C área correspondente a camada zonal (Zo) da camada cinza superficial do colículo
superior (SuG) e suas divisões seguintes: camada de nervo óptico (Op), camada
intermediária cinza (InG), (E) camada cinza profunda (DpG), (F) núcleo tegmental
microcelular (MiTg), (H) Ampliação de caixa homônima em G área correspondente ao locus
cerúleos (LC), núcleo trigeminal mesencefálico (Me5), (J) Ampliação de caixa homônima em
I área correspondente a camada gelatinosa do núcleo trigeminal espinhal caudal (Ge5),
núcleo trigeminal espinhal, parte caudal (Sp5C). Barra: 500 µm em A-G e I-J, 166 µm em H.

67
5. DISCUSSÃO
A molécula do NO é sintetizada pela enzima NOS a partir do substrato L-
arginina, num processo que envolve transferências de elétrons do NADPH através
de flavinas FAD e FMN, para que no domínio redutase carboxi-terminal e para o
heme no domínio oxigenasse amino-terminal, possa transformar o substrato em L-
citrulina e a molécula NO, por meio de oxidação (Andrew e Mayer, 1999).
Com isso realizamos as duas técnicas de marcação (imunoistoquímica e
histoquímica) para que ambas uma complementasse a outra, possa corroborar os
achados. Porém, vale destacar que para a última técnica a marcação dos neurônios
dentro dos núcleos não apresentou a nitidez da imunoistoquímica, pois a marcação
positiva ocorre apenas em neurônios que estejam sintetizando o NO durante o
momento da perfusão. Enquanto que na imunoistoquímica irá ser positivo para todos
os neurónios que contêm NOS, de acordo com Hope e colaboradores (1991).
Além disso, Matsumoto e colaboradores (1993) sugerem que o pré-
tratamento dos tecidos com paraformaldeído dependente da dose (0.06 a 4%) altera
o padrão de distribuição da marcação com o NADPH, pois o mesmo relata que na
porção reticulada dos tecidos não houve qualquer atividade. Já na parte citosólica o
padrão de atividade da enzima permaneceu inalterada entre 40% a 60%, com o
resultado obtido por meio da mensuração de moléculas específicas para cada parte
celular e com resultados diferentes de acordo com o tecido investigado. Com isso
ele mostra que a maior parte da ação enzimática do NADPH se perde.
Além disso, Zhou e Zhu (2009), em seu artigo de revisão explicitam que a
localização subcelular do NOS na célula pode contribuir para a sua diversidade de
funções. De qualquer forma, a marcação por NADPHd serviu pra confirmar os
achados corroborando ainda mais as pequenas discrepâncias achadas entre as
técnicas realizadas aqui.
Foi possível analisar a distribuição dos núcleos com marcação positiva para o
NOS comparando o material analisado com seu análogo corado com a técnica de
Nissl. A partir daí foi possível verificar que a organização topográfica dos núcleos
cerebrais do mocó é muito semelhante a outros mamíferos. Em especial ao rato, do
qual foi utilizado em maior proporção para a comparação e identificação desses

68
núcleos, por meio do atlas de Paxinos e Watson (2007), sem esquecer também dos
demais atlas já citados no iten: 3.2.6 Análise das imagens.
5.1 ASPECTOS NEUROANATÔMICOS E FUNCIONAIS DO OXIDO NÍTRICO NO
TRONCO ENCEFÁLICO DO MOCÓ
O sistema visual primário é formado basicamente pelos núcleos pré-tectais,
colículo superior e o geniculado lateral no tálamo. Os núcleos pré-tectais estão
envolvidos nos reflexos pupilares e movimentos dos olhos (Scalia e Arango, 1979;
Klooster et al.,1995). Os núcleos pré-tectais no mocó, localizados no mesencéfalo
seguiu o mesmo padrão morfológico de rato e camundongo (Silva, 2009).
Os núcleos pré-tectais também estão envolvidos com a analgesia em ratos. L-
arginina quando injetado via intracerebroventricular (I.C.V.) no pré-tectal provocou
aumento da produção do NO, que por sua vez ativou o sistema de analgesia de
longa duração, envolvendo o GMP- cíclico (Kumar et al., 1993).
O colículo superior do mocó está subdividido em 7 camadas (Silva, 2009).
O colículo superior é responsável pela integralização de diversas informações
sensoriais como a visão e os sinais para que os olhos se movimentem (Werner et
al., 1997). A expressão de NOS por células coliculares aumentou a partir do final da
primeira semana pós-natal e atingindo um pico de expressão máximo por volta do
15º dia, e como o NO tem sido implicado na plasticidade das projeções durante o
desenvolvimento, o mesmo sugeriu que neurônios NO-IR estão envolvidos no
refinamento de projeções, retino-colicular e córtico-tectal (Tenório et al., 1996).
Corroborando ainda mais essa função organogênica do NO e sistema visual, a
síntese de NO ocorre em células microgliais como uma resposta à lesão do axônio,
enquanto que a expressão em astrócitos parece estar associada tanto a processos
normais em condições fisiológicas como a fase regenerativa após a lesão do nervo
optico em peixes tenca (Clemente et al., 2002). Chacur e colaboradores (2006)
demonstraram por meio de diversos métodos como imunoistoquímica e PCR em
tempo real, que a expressão da NOS no colículo superior do rato fica elevada após a
enucleação unilateral. Cramer e colaboradores (1998) fez uma grande revisão, sobre
a sua função, no desenvolvimento do sistema visual e suas interações em diferentes
áreas cerebrais.

69
Mocó é um roedor de hábitos crepusculares (Sousa e Menezes, 2006) e usam
bem sua visão. O olho é lateralizado, abrigado em uma órbita óssea, equipada por
duas dobras da pele superior e inferior. No pólo anterior do olho, a íris preta e a
pupila em forma de fenda vertical. O globo ocular tem forma elipsoidal (Oliveira et al.,
2014). Com todas as funções que NO tem no sistema visual faz sentido que NO
esteja de forma abundante no sistema visual de mocó.
Relacionando o NO com o sistema auditivo, as funções do complexo olivar
superior incluem a detecção de diferentes intensidades de som e o mapeamento
espacial, o controle de feedback dos mecanismos cocleares e o processamento de
sinais cocleares através de vias auditivas ascendente, sendo o colículo inferior o
primeiro local dessas conexões (Helfert et al., 1988). O NO pode estar envolvido no
processamento e transmissão da entrada acústica para o córtex auditivo, uma vez
que o tratamento de ratos com um inibidor de NOS reduziu a amplitude das
respostas auditivas de latência média (Grassi et al., 1995) e da dessincronização
eletrocortical evocada pelo som (Iannone et al., 1996).
Em estudos conduzidos, por Schaeffer e colaboradores (2003) no complexo
olivar superior, formado pela oliva superior lateral (LSO), núcleo paraolivar (SPO) e a
oliva superior medial (MSO), verificou que projeções ascendentes para o colículo
inferior eram nitrégicos, bem como é sabido que subpopulações de neurônios do
complexo olivar superior produz NOS (Riemann e Reuss, 1999). Alvarado e
colaboradores (2016) mostraram que um condicionamento apropriado à exposição
ao ruído pode reduzir os danos causados nas próximas exposições a ruídos
semelhantes, esse efeito protetor era sabido, mas não era ainda muito bem
compreendido, então eles mostraram que a exposição de ratos a sons de 118
decibéis por uma hora a cada 72 horas por 4 vezes, aumentou a concentração de
neurônios nitrérgicos no núcleo coclear, se estendendo a outros componentes do
sistema auditivo central. Este fenômeno pode estar em parte relacionado a uma
interação entre as vias de sinalização do núcleo coclear com o NO devido a um
aumento da capacidade tamponadora do cálcio citosólico induzido pelo protocolo de
condicionamento de ruído.
O habitat do mocó (caatinga) é um ambiente aberto de pouca vegetação,
beneficiando a detecção de sons a longa distância. A presença de NO no sistema
auditivo central de mocó mostra um papel importante dele na audição e outras

70
funções deste sistema, podendo beneficiar esta espécie na detecção de predadores
neste ambiente.
A substância cinzenta periaquedutal (PAG) e suas divisões são grandes
grupos de núcleos que circunda o aqueduto mesencefálico. É um importante sítio
neural para inúmeras funções incluindo a regulação cardiovascular, a modulação da
dor e atividades comportamentais (Xing et al, 2012). Quando a DPAG de ratos ou
camundongos é estimulada química ou eletricamente produz saltos e fugas, além de
comportamento de congelamento motor (Schenberg et al., 2005 e Miguel e Nunes-
de-Souza, 2006), comportamento bem semelhante aquele de fuga de predadores
naturais (Blanchard et al., 2001), o que também leva a um estado de antinocicepção.
Diversos neurotransmissores como a serotonina o ácido gama-aminobutírico
(GABA) e glutamato, quando aí injetados alteram esses comportamentos (Graeff,
2004 e Engin e Treit, 2008). Este último é expresso em grande quantidade em todo
a PAG e consequentemente sua ativação desencadeia a síntese do NO (Garthwaite
et al., 1989) e injeções de doadores de NO na DPAG provoca reações de luta e fuga
(Braga et al., 2009). Já os inibidores de NO, quando injetados na PAG, provocam
efeitos ansiolíticos em ratos, semelhantes aos expostos ao labirinto em cruz elevado
quando um potente inibidor do NOS, o Nω-propil-L-arginina (NPLA), é injetado
I.C.V., atenuando a defesa de ratos em testes de presa-predador (Carvalho-Netto,
2009). No camundongo a injeção de NMDA inverte o comportamento nociceptivo e
defensivo quando confrontado com um predador (Miguel e Nunes-de-Souza, 2006).
Emmanouil e colaboradores (2008) demonstraram que a antinocicepção
induzida pelo óxido nitroso (N2O) ocorria na PAG em camundongos que passavam
por teste de constrição abdominal, sugerindo que o N2O atua no PAG através de um
receptor mediado por receptores opióides dependentes de NO para induzir
antinocicepção.
Xing e colaboradores (2012) acharam alta co-localização da angiotensina e
NOS no DLPAG. A angiotensina é considerada um membro biologicamente ativo do
sistema renina-angiotensina, com propriedades vasodilatadoras e anti-proliferativas,
sendo assim foi visto que a angiotensina, desempenha um papel inibitório, através
de uma sinalização dependente de NO, o que pode oferecer uma base fisiológica
para entender a regulação de diversas funções na parte central do sistema nervoso
por parte da NO.

71
Ainda na PAG, Barbaresi e colaboradores (2013) identificaram calbindina em
neurônios NADPHd. A calbidina desempenha um papel na modulação da
excitabilidade neuronal e tem um efeito profundo na amplitude de picos transientes
de cálcio (Ca2+). Esta co-localização foi de 42% a 66%, sendo mais elevada nessa
região do tronco encefálico, o que pode sugerir uma elevada expressão por uma alta
demanda do NO. No nosso trabalho mostramos que na PAG como um todo os
neurônios NOS-IR são expressos em grande quantidade e intensamente corados,
corroborando achados prévios em roedores e humanos (Leigh et al., 1990; Vicent e
Kimura, 1992; Egberongbe et al., 1994; Gotti et al., 2005), e reforçando ainda mais
a intença relação da NO com a PAG nos mamíferos e em especial no mocó.
O locus cerúleos (LC) é um pequeno núcleo em forma de “vírgula” presente
de cada lado na porção superior da ponte, margeando o 4º ventrículo. É o principal
núcleo noradrenérgico do encéfalo e desempenha importantes papeis na parte
central do sistema nervoso como a regulação da excitação, vigília e o controle
autonômico (Samuels e Szabadi, 2008). Agamatina estimula a taxa de disparo dos
neurônios no LC através de um mecanismo dependente da síntese de NO (Ruiz-
Durantez et al., 2002). A agamatina é uma substância que produz inúmeros efeitos
na parte central do sistema nervoso, entre elas, potencializa a analgesia da morfina,
bloqueia o desenvolvimento de tolerância a opiáceos (Kolesnikov et al., 1996;
Aricioglu-Kartal e Uzbay, 1997) e estimula a liberação de hormônio pituitário (Kalra
et al., 1995). Estando NO também presente no LC de mocó, podemos inferir que o
mesmo deve influenciar as ações catecolaminérgicas do LC nesta espécie.
Karolewicz e colaboradores (2004) em pacientes humanos diagnosticados
com depressão visualizou uma baixa concentração de NOS no LC, com isso
refletindo uma transmissão excitatória alterada promovida pelo glutamato,
neurotransmissor este que tem o NO como mediador intracelular da ativação do seu
receptor. Com isso reforçando o argumento que desordens depressivas ocorrem por
interrupções da sinalização glutamatérgica.
NO também está envolvido diretamente em mecanismo patológicos cerebrais.
A cirrose hepática pode progredir para outros órgãos inclusive o sistema nervoso,
causando a encefalopatia hepática (Chen et al., 2007). Ela provoca alterações
motoras incluindo rigidez e tremor, refletindo em distúrbios dopaminérgicos
estriatais. Como a substância negra (SN) é uma importante produtora de dopamina,

72
os pesquisadores observaram que a concentração de NO está correlacionada com a
liberação endógena da dopamina. Então ratos com cirrose hepática apresentaram
uma quantidade reduzida de neurônios NADPHd na SN, seu soma neuronal
encolheu, bem como a quantidade de fibras. Reforçando o papel de NO na SN.
Matthews e colaboradores (1997) examinaram os efeitos da injeção estriatal de 1-
metil-4-fenilpiridinio (MPP+) na degeneração da SN em camundongos “knockout”
para o gene NOS com parkingson induzido por (MPTP). Tanto o volume da lesão
estriatal quanto a degeneração da SN foram significativamente atenuados nos
camundongos mutantes para NOS. L’Episcopo e colaboradores (2010)
demonstraram que um doador de NO, derivado do flurbiprofeno, o [2-Fluoro-a-metil
(1,1'-bifenil) -4-acético-4- (nitrooxi) butil éster], ou HCT1026, quando administrada
oralmente a ratos que apresentaram a doença de Parkinson induzida por MPTP, fez
com que houvesse uma diminuição significativa na expressão de iNOS (NOS
induzida), antígeno-1 do macrófago (Mac-1) e NADPH oxidase (PHOX) em células
microgliais dentro do estriado e do mesencéfalo ventral, diminuindo os efeitos
inflamatórios e neurotóxicos a essas células neuronais e prevenindo o aparecimento
de sintomas motores da doença.
Nosso trabalho mostra que o mocó apresenta neurônios nitrérgicos nos três
núcleos dopaminérgicos mesencefálicos (substância negra, área tegmental ventral e
região retrorubral), e mesmo sem um trabalho de co-localização podemos inferir uma
estreita relação do NO com a dopamina também em mocó. O NO comprovadamente
esta relacionado à modulação da atividade motora e em comportamentos
relacionados a ansiedade. Del Bel e colaboradores (2002) mostraram a redução da
atividade locomotora por inibidores de NOS o NG-nitro-L-arginina (L-NOARG) e o 7-
nitroindazol (7-NIO) em ratos expostos a campo aberto. Corroborando a modulação,
Carletti e colaboradores (2009) com testes em ratos vivos com inibidores e doadores
de NOS, além de testes eletrofisiológicos demonstraram a existência de um sistema
neuromodulatório intrínseco da ação exercida pelo NO em neurônios da parte
reticulada da SN, atraves de uma via de sinalização pelo cGMP.
A área tegmental ventral (VTA) possui neurônios dopaminérgicos que estão
dentro de núcleos como o interfascicular (IF), parabraquial (PBP) e linear rostral da
rafe (RLi). Estes neurônios estão envolvidos na regulação motora, comportamentos
motivacionais, revelam uma elevada plasticidade neuronal, colocalizam com NOS e

73
parecem interagir uns com outros pela presença de terminações NOS em neurônios
TH e vice-versa, com isso sugerindo que o NO está envolvido na modulação de
transmissão dopaminérgica (Klejbor et al., 2004). Em nossos achados, os 3 núcleos
apresentaram uma marcação forte, mostrando neurônios com uma grande produção
de NO apesar de uma densidade moderada, sugerindo sobre uma interação entre
NO e dopamina em mocó.
Com foco no desenvolvimento de terapias que previna ou retarde o
aparecimento de sintomas motores e cognitivos, Balez e Ooi (2015) apresentam um
amplo artigo de revisão, corroborando o papel neuroprotetor do NO protegendo as
sinapses e aumentando a excitabilidade neuronal. E o mecanismo potencial para
esse aumento da excitabilidade por NO é via modulação da atividade dos canais de
potássio dependentes de voltagem.
Volke e colaboradores (1997) mostraram uma clara ação do tipo ansiolítico do
inibidor da NOS (7-NIO) em quatro testes de ansiedades diferentes (campo aberto,
compartimento claro e escuro teste de interação social e labirinto em cruz elevado)
feitos em ratos e camundongos. Navabi e colaboradores (2016) demonstraram que a
administração aguda de cloreto de zinco leva a um comportamento ansiolítico com
uma possível via do NO, através da administração da L-arginina, um precursor do
NO, em ratos expostos a paradigmas de ansiedade, demonstrando o quão
importante é o envolvimento de NO nesses comportamentos, assim também deve
ser no mocó.
Os núcleos tegmental laterodorsal (LDT) e tegmental pedunculopontino (PPT)
apresentaram neurônios NOS-IR em mocó. Estes são núcleos classicamente
colinérgicos e em ratos há uma co-localização de aproximadamente 100% entre
ChAT, enzima de síntese de acetilcolina, e NOS (Veleanu et al., 2016). Estes
núcleos são componentes de suma importância para o sistema ativador reticular
ascendente (Moruzzi e Magoun, 1949), e apesar de não sabermos se em mocó há
co-localização entre NOS e ChAT, podemos pressupor um papel de NOS no
controle da ativação cortical também em mocó.
O papel regulatório do NO aparece ainda em outros núcleos do tronco
encefálico. Beltrán e colaboradores (1999) mostraram que neurônios nitrérgicos no
núcleo motor dorsal do vago inibem a liberação de ácido gástrico no estômago de
ratos, o que pode indicar ser esse mecanismo o responsável por alguns distúrbios

74
de secreção gástrica. No trabalho atual vimos alguns neurônios NOS-IR neste
núcleo, o que ainda assim pode indicar um papel de NO na secreção gástrica do
mocó.

75
6. CONCLUSÃO
Nossos achados sobre a distribuição de NOS no tronco encefálico de mocó
monstraram uma grande semelhança com os achados em outros mamíferos,
principalmente outros roedores. Através da imunistoquímica contra NOS vimos uma
grande distribuição de neurônios nitrérgicos por todas as estruturas que compõe o
tronco encefálico. Tal distribuição foi mais branda quando utilizamos a NADPHd,
demonstrando que a técnica de NADPHd utilizada aqui não é confiável para o
mapeamento de neurônios nitrérgicos em mocó, mas ao mesmo tempo revela a
atividade da enzima de síntese.
Esta semelhança entre a distribuição de NOS no mocó e outros mamíferos
mostra que a neuroanatomia do NO é filogeneticamente bem conservada e assim
também devem ser suas funções, não excluindo aqui a recomendação que mais
estudos, que estejam ligados ao comportamento devem ser realizados. Portanto,
acreditamos que no mocó NO tem um papel fundamental em diversas funções vitais
realizadas pelo tronco encefálico.

76
REFERÊNCIAS1
ALONSO, J. R. et al. Chapter III comparative and developmental neuroanatomical aspects of the NO system. Handbook of chemical neuroanatomy, v. 17, p. 51-109, 2000. ALVARADO, J. C. et al. Noise-Induced “toughening” effect in wistar rats: enhanced auditory brainstem responses are related to calretinin and nitric oxide synthase Upregulation. Frontiers in neuroanatomy, v. 10, 2016. ANDREW, P. J.; MAYER, B. Enzymatic function of nitric oxide synthases. Cardiovascular research, v. 43, n. 3, p. 521-531,1999. ARCHER, S. Measurement of nitric oxide in biological models. The FASEB journal,
v. 7, n. 2, p. 349-360, 1993. ARICIOGLU-KARTAL, F.; UZBAY, I. T. Inhibitory effect of agmatine on naloxone-precipitated abstinence syndrome in morphine dependent rats. Life sciences, v. 61, n. 18, p. 1775-1781, 1997. ATOJI, Y.; YAMAMOTO, Y.; SUZUKI, Y. Distribution of NADPH diaphorase-containing neurons in the pigeon central nervous system. Journal of chemical neuroanatomy, v. 21, n. 1, p. 1-22, 2001.
AYTON, S. et al. Parkinson's disease iron deposition caused by nitric oxide-induced loss of β-amyloid precursor protein. The journal of neuroscience, v. 35, n. 8, p.
3591-3597, 2015. BALEZ, R.; OOI, L. Getting to NO Alzheimer’s disease: neuroprotection versus neurotoxicity mediated by nitric oxide. Oxidative medicine and cellular longevity, v. 2016, 2015. BARBARESI, P. et al. Differential distribution of parvalbumin-and calbindin-D28K-immunoreactive neurons in the rat periaqueductal gray matter and their colocalization with enzymes producing nitric oxide. Brain research bulletin, v. 99, p. 48-62, 2013.
1 De acordo com a Associação Brasileira de Normas Técnicas. NBR 6023.

77
BEAR, M. F.; CONNORS, B. W.; PARADISO, M. A. Neurociências: desvendando o sistema nervoso. Artmed Editora, 2016. BELTRÁN, B. et al. Synthesis of nitric oxide in the dorsal motor nucleus of the vagus mediates the inhibition of gastric acid secretion by central bombesin. British journal of pharmacology, v. 127, n. 7, p. 1603-1610, 1999.
BLANCHARD, D. C. et al. Human defensive behaviors to threat scenarios show parallels to fear-and anxiety-related defense patterns of non-human mammals. Neuroscience & biobehavioral Reviews, v. 25, n. 7, p. 761-770, 2001. BÖHME, G. A. et al. Possible involvement of nitric oxide in long-term potentiation. European journal of pharmacology, v. 199, n. 3, p. 379-381, 1991. BÖHME, G. A. et al. Altered synaptic plasticity and memory formation in nitric oxide synthase inhibitor-treated rats. Proceedings of the national academy of sciences, v. 90, n. 19, p. 9191-9194, 1993. BRAGA, A. A.; AGUIAR, D. C.; GUIMARÃES, F. S. NOC-9, a selective nitric oxide donor, induces flight reactions in the dorsolateral periaqueductal gray of rats by activating soluble guanylate cyclase. Neuroscience letters, v. 459, n. 2, p. 79-83, 2009. BREDT, D. S.; SNYDER, S. H. Nitric oxide mediates glutamate-linked enhancement of cGMP levels in the cerebellum. Proceedings of the national academy of sciences, v. 86, n. 22, p. 9030-9033, 1989. BREDT, D. S. et al. Cloned and expressed nitric oxide synthase structurally resembles cytochrome P-450 reductase. Nature, 351, p. 714 – 718, 1991. CARLETTI, F. et al. Nitric oxide-and cGMP-active compounds affect the discharge of substantia nigra pars reticulata neurons: in vivo evidences in the rat. Journal of neural transmission, v. 116, n. 5, p. 539-549, 2009.
CARVALHO-NETTO, E. F. et al. Role of glutamate NMDA receptors and nitric oxide located within the periaqueductal gray on defensive behaviors in mice confronted by predator. Psychopharmacology, v. 204, n. 4, p. 617-625, 2009.

78
CAVALCANTE, J. S. et al. Calcium-binding proteins in the circadian centers of the common marmoset (Callithrix jacchus) and the rock cavy (Kerodon rupestris) brains. Brain research bulletin, v. 76, n. 4, p. 354-360, 2008.
CAVALCANTI, J. R. L. P. et al. A cytoarchitectonic and TH-immunohistochemistry characterization of the dopamine cell groups in the substantia nigra, ventral tegmental area and retrorubral field in the rock cavy (Kerodon rupestris). Journal of chemical neuroanatomy, v. 55, p. 58-66, 2014.
CHACUR, M. et al. Differential regulation of the neuronal isoform of nitric oxide synthase in the superior colliculus and dorsal lateral geniculate nucleus of the adult rat brain following eye enucleation. International journal of developmental neuroscience, v. 24, n. 7, p. 461-468, 2006.
CHEN, Y. et al. Distribution of nitric oxide synthase positive neurons in the substantia nigra of rats with liver cirrhosis. Neural regeneration research, v. 2, n. 7, p. 399-
402, 2007. CLEMENTE, D. et al. Effects of axotomy on the expression of NADPH-diaphorase in the visual pathway of the tench. Brain research, v. 925, n. 2, p. 183-194, 2002. CRAMER, K. S.; LEAMEY, C. A.; SUR, M. Nitric oxide as a signaling molecule in visual system development. Progress in brain research, v. 118, p. 101-114, 1998. DARWINKEL, A. et al. Distribution of orexin-1 receptor-green fluorescent protein-(OX 1-GFP) expressing neurons in the mouse brain stem and pons: co-localization with tyrosine hydroxylase and neuronal nitric oxide synthase. Neuroscience, v. 278, p.
253-264, 2014. DAWSON, T. M.; SNYDER, S. H. Gases as biological messengers: nitric oxide and carbon monoxide in the brain. The journal of neuroscience, v. 14, n. 9, p. 5147-5159, 1994. DEL BEL, E. et al. Motor effects of acute and chronic inhibition of nitric oxide synthesis in mice. Psychopharmacology, v. 161, n. 1, p. 32-37, 2002.
DRAPIER, J.; WIETZERBIN, J.; HIBBS, J B. Interferon‐γ and tumor necrosis factor induce the L‐arginine‐dependent cytotoxic effector mechanism in murine macrophages. European journal of immunology, v. 18, n. 10, p. 1587-1592, 1988.

79
DUSSE, L. M. S.; VIEIRA, L. M.; CARVALHO, M. G. Revisão sobre óxido nítrico. Jornal brasileiro de patologia e medicina laboratorial, v. 39, n. 4, p. 435-50, 2003. EGBERONGBE, Y. I. et al. The distribution of nitric oxide synthase immunoreactivity in the human brain. Neuroscience, v. 59, n. 3, p. 561-578, 1994.
EMMANOUIL, D. E. et al. Nitrous oxide-antinociception is mediated by opioid receptors and nitric oxide in the periaqueductal gray region of the midbrain. European neuropsychopharmacology, v. 18, n. 3, p. 194-199, 2008. ENGIN, E.; TREIT, D. The effects of intra-cerebral drug infusions on animals unconditioned fear reactions: a systematic review. Progress in neuro-psychopharmacology and biological psychiatry, v. 32, n. 6, p. 1399-1419, 2008.
FENG, H. M.; WALKER, D. H. Interferon-gamma and tumor necrosis factor-alpha exert their antirickettsial effect via induction of synthesis of nitric oxide. The american journal of pathology, v. 143, n. 4, p. 1016, 1993. FERRENDELLI, J. A.; CHANG, M. M.; KINSCHERF, D. A. Elevation of cyclic gmp levels in central nervous system by excitatory and inhibitory amino acids. Journal of neurochemistry, v. 22, n. 4, p. 535-540, 1974.
FREIRE, M. A. M. et al. Morphometric variability of nicotinamide adenine dinucleotide phosphate diaphorase neurons in the primary sensory areas of the rat. Neuroscience, v. 205, p. 140-153, 2012. FURCHGOTT, R. F. et al. Endothelial cells as mediators of vasodilation of arteries. Journal of cardiovascular pharmacology, v. 6, p. 336-343, 1984. FURCHGOTT, R. F. et al. The obligatory role of endothelial cells in the relaxation of arterial smooth muscle by acetylcholine. Nature, v. 288, n. 5789, p. 373-376, 1980. GAI, W. P.; BLESSING, W. W. Human brainstem preganglionic parasympathetic neurons localized by markers for nitric oxide synthesis. Brain, v. 119, n. 4, p. 1145-1152, 1996. GARTHWAITE, J. et al. NMDA receptor activation induces nitric oxide synthesis from arginine in rat brain slices. European journal of pharmacology: molecular pharmacology, v. 172, n. 4, p. 413-416, 1989.

80
GAZZINELLI, R. T. et al. The microbicidal activity of interferon‐γ‐treated
macrophages against Trypanosoma cruzi involves an l‐arginine‐dependent, nitrogen
oxide‐mediated mechanism inhibitable by interleukin‐10 and transforming growth factor‐β. European journal of immunology, v. 22, n. 10, p. 2501-2506, 1992.
GONZÁLEZ‐SORIANO, J. et al. NADPH‐diaphorase distribution in the rabbit superior colliculus and co‐localization with calcium‐binding proteins. Journal of anatomy, v.
200, n. 3, p. 297-308, 2002. GOTTI, S. et al. Distribution of nitric oxide synthase immunoreactivity in the mouse brain. Microscopy research and technique, v. 68, n. 1, p. 13-35, 2005.
GRAEFF, F. G. Serotonin, the periaqueductal gray and panic. Neuroscience & biobehavioral Reviews, v. 28, n. 3, p. 239-259, 2004.
GRASSI, C. et al. Possible modulation of auditory middle latency responses by nitric oxide in the inferior colliculus of anaesthetized rats. Neuroscience letters, v. 196, n.
2, p. 213-217, 1995. GRAY, H.; STANDRING, S. Gray’s anatomy: the anatomical basis of clinical practice, 40th edition. Elsevier. 2008.
GREEN, L. C. et al. Nitrate synthesis in the germfree and conventional rat. Science, v. 212, n. 4490, p. 56-58, 1981. HELFERT, R. H.; SNEAD, C. R.; ALTSCHULER, R. A. The ascending auditory pathways. Neurobiology of hearing: The central auditory system, p. 1-25, 1991. HIBBS, J. B. et al. Nitric oxide: a cytotoxic activated macrophage effector molecule. Biochemical and biophysical research communications, v. 157, n. 1,
p. 87-94, 1988. HIBBS, J. B.; TAINTOR, R. R.; VAVRIN, Z. Macrophage cytotoxicity: role for L-arginine deiminase and imino nitrogen oxidation to nitrite. Science, v. 235, n. 4787, p. 473-476, 1987. HIRONAGA, K. et al. Role of endogenous nitric oxide in the brain stem on the rapid adaptation of baroreflex. Hypertension, v. 31, n. 1, p. 27-31, 1998.
HOLMQVIST, B. et al. Identification and distribution of nitric oxide synthase in the brain of adult zebrafish. Neuroscience letters, v. 292, n. 2, p. 119-122, 2000.

81
HOPE, B. T. et al. Neuronal NADPH diaphorase is a nitric oxide synthase. Proceedings of the national academy of sciences, v. 88, n. 7, p. 2811-2814, 1991. IANNONE, M. et al. Sound-evoked electrocortical desynchronization is inhibited by N ω-nitro-l-arginine methyl ester microinfused into the inferior colliculi in rats. Electroencephalography and clinical neurophysiology, v. 99, n. 1, p. 57-62, 1996. IGNARRO, L. J. et al. Endothelium-derived relaxing factor produced and released from artery and vein is nitric oxide. Proceedings of the national academy of sciences, v. 84, n. 24, p. 9265-9269, 1987. IGNARRO, L. J. et al. Basic polyamino acids rich in arginine, lysine, or ornithine cause both enhancement of and refractoriness to formation of endothelium-derived nitric oxide in pulmonary artery and vein. Circulation research, v. 64, n. 2, p. 315-
329, 1989. IKONOMIDOU, C. et al. Neurotransmitters and apoptosis in the developing brain. Biochemical pharmacology, v. 62, n. 4, p. 401-405, 2001. INUTSUKA, A.; YAMANAKA, A. The physiological role of orexin/hypocretin neurons in the regulation of sleep/wakefulness and neuroendocrine functions. Frontiers in endocrinology, v.4. p.1-10. 2013.
IWASE, K. et al. Precise distribution of neuronal nitric oxide synthase mRNA in the rat brain revealed by non-radioisotopic in situ hybridization. Molecular brain research, v. 53, n. 1, p. 1-12, 1998. KALRA, S. P. et al. Agmatine, a novel hypothalamic amine, stimulates pituitary luteinizing hormone release in vivo and hypothalamic luteinizing hormone-releasing hormone release in vitro. Neuroscience letters, v. 194, n. 3, p. 165-168, 1995.
KAROLEWICZ, B. et al. Low nNOS protein in the locus coeruleus in major depression. Journal of neurochemistry, v. 91, n. 5, p. 1057-1066, 2004.
KAY, K. et al. Hindbrain orexin 1 receptors influence palatable food intake, operant responding for food, and food-conditioned place preference in rats. Psychopharmacology, v. 231, n. 2, p. 419-427, 2014.

82
KIECHLE, F. L.; MALINSKI, T. Nitric oxide: biochemistry, pathophysiology, and detection. American journal of clinical pathology, v. 100, n. 5, p. 567-575, 1993. KLEJBOR, I. et al. The relationships between neurons containing dopamine and nitric oxide synthase in the ventral tegmental area. Folia histochemica et cytobiologica, v. 42, n. 2, p. 83-88, 2004.
KLOOSTER, J. et al. Efferent projections of the olivary pretectal nucleus in the albino rat subserving the pupillary light reflex and related reflexes a light microscopic tracing study. Brain research, v. 688, n. 1, p. 34-46, 1995. KNOWLES, R. G. et al. Formation of nitric oxide from L-arginine in the central nervous system: a transduction mechanism for stimulation of the soluble guanylate cyclase. Proceedings of the national academy of sciences, v. 86, n. 13, p. 5159-
5162, 1989. KOLESNIKOV, Y.; JAIN, S.; PASTERNAK, G. W. Modulation of opioid analgesia by agmatine. European journal of pharmacology, v. 296, n. 1, p. 17-22, 1996. KUMAR, A. et al. Evidence for involvement of nitric oxide in pretectal analgesia in rat. Neuroreport, v. 4, n. 6, p. 706-708, 1993. LACERDA, P. M. O. et al. Origem do plexo lombossacral de mocó (Kerondo rupestris). Brazilian journal of veterinary research and animal science, v. 43, n. 5, p. 620-628, 2006. LEIGH, P. N.; CONNICK, J. H.; STONE, T. W. Distribution of NADPH-diaphorase positive cells in the rat brain. Comparative biochemistry and physiology part C: comparative pharmacology, v. 97, n. 2, p. 259-264, 1990. L'EPISCOPO, F. et al. Combining nitric oxide release with anti-inflammatory activity preserves nigrostriatal dopaminergic innervation and prevents motor impairment in a 1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine model of Parkinson's disease. Journal of neuroinflammation, v. 7, n. 1, p. 1, 2010. LIMA, M. C. et al. Glândula mamária do mocó (Kerodon rupestris-Wied Neuwied, 1820): aspectos morfológicos. Brazilian journal of veterinary research and animal science, v. 45, n. supl., p. 88-93, 2008.

83
LOWENSTEIN, C. J. et al. Nitric oxide inhibits viral replication in murine myocarditis. Journal of clinical investigation, v. 97, n. 8, p. 1837, 1996. MACHADO, A. B. M. Neuroanatomia funcional. Atheneu, 2005.
MARLETTA, M. A. et al. Macrophage oxidation of L-arginine to nitrite and nitrate: nitric oxide is an intermediate. Biochemistry, v. 27, n. 24, p. 8706-8711, 1988.
MASINI, M. A. et al. Identification and distribution of nitric oxide synthase in the brain of adult Antarctic teleosts. Polar biology, v. 29, n. 1, p. 23-26, 2005.
MATSUMOTO, T. et al. A correlation between soluble brain nitric oxide synthase and NADPH-diaphorase activity is only seen after exposure of the tissue to fixative. Neuroscience letters, v. 155, n. 1, p. 61-64, 1993. MATTHEWS, R. T. et al. MPP+ induced substantia nigra degeneration is attenuated in nNOS knockout mice. Neurobiology of disease, v. 4, n. 2, p. 114-121, 1997. MIGUEL, T. T.; NUNES-DE-SOUZA, R. L. Defensive-like behaviors and antinociception induced by NMDA injection into the periaqueductal gray of mice depend on nitric oxide synthesis. Brain research, v. 1076, n. 1, p. 42-48, 2006. MIKI, N.; KAWABE, Y.; KURIYAMA, K. Activation of cerebral guanylate cyclase by nitric oxide. Biochemical and biophysical research communications, v. 75, n. 4, p. 851-856, 1977. MONCADA, S.; HIGGS, A. The L-arginine-nitric oxide pathway. New england journal of medicine, v. 329, n. 27, p. 2002-2012, 1993.
MOOJEN, J. Os roedores do brasil. Rio de Janeiro: Instituto Nacional do Livro, 1952. MORENO, N. et al. Development of NADPH-diaphorase/nitric oxide synthase in the brain of the urodele amphibian Pleurodeles waltl. Journal of chemical neuroanatomy, v. 23, n. 2, p. 105-121, 2002. MORUZZI, G.; MAGOUN, H. W. Brain stem reticular formation and activation of the EEG. Electroencephalography and clinical neurophysiology, v. 1, n. 1, p. 455-473, 1949.

84
MORAIS, P. L. A. G. et al. Retinal projections into the zona Incerta of the rock cavy (Kerodon rupestris): A CTb study. Neuroscience research, v. 89, p. 75-80, 2014.
NASYROVA, R. F. et al. Role of nitric oxide and related molecules in schizophrenia pathogenesis: biochemical, genetic and clinical aspects. Frontiers in physiology, v.
6, 2015. NAVABI, S. P. et al. Involvement of l-arginine-nitric oxide pathway in anxiolytic-like effects of zinc chloride in rats. Biomedicine & pharmacotherapy, v. 83, p. 521-525, 2016. NASCIMENTO JUNIOR, E. S. et al. Retinal projections to the thalamic paraventricular nucleus in the rock cavy (Kerodon rupestris). Brain research, v.
1241, p. 56-61, 2008. NASCIMENTO JUNIOR, E. S. et al. Retinal afferents to the thalamic mediodorsal nucleus in the rock cavy (Kerodon rupestris)(a). Neuroscience letters, v. 475, n. 1, p. 38-43, 2010. NASCIMENTO JUNIOR, E. S. et al. The suprachiasmatic nucleus and the intergeniculate leaflet in the rock cavy (Kerodon rupestris): retinal projections and immunohistochemical characterization (b). Brain research, v. 1320, p. 34-46, 2010. NELSON, R. J. et al. Effects of nitric oxide on neuroendocrine function and behavior. Frontiers in neuroendocrinology, v. 18, n. 4, p. 463-491, 1997. OGURA, T. et al. Neuronal nitric oxide synthase expression in neuronal cell differentiation. Neuroscience letters, v. 204, n. 1, p. 89-92, 1996. OLIVEIRA, F. G. et al. The eye of the crepuscular rodent rock cavy. Journal of morphology, v. 31, n. 2, p. 89-97, 2014. OSWALD, I. P. et al. Endothelial cells are activated by cytokine treatment to kill an intravascular parasite, Schistosoma mansoni, through the production of nitric oxide. Proceedings of the national academy of sciences, v. 91, n. 3, p. 999-1003,
1994. PAAKKARI, I.; LINDSBERG, P. Nitric oxide in the central nervous system. Annals of medicine, v. 27, n. 3, p. 369-377, 1995.

85
PAXINOS, G. F. K. B. J.; FRANKLIN, K. B. J. The mouse brain in stereotaxic coordinates. 2008, 2nd edition. San Diego: Academic Press. PAXINOS, G. F. K. B.J.; WATSON, C. The rat brain in stereotaxic coordinates.
2007, 6th edition. Academic press. PAXINOS, G. F. K. B. J. et al. The marmoset brain in stereotaxic coordinates.
2012, 1st edition. Academic Press. PHILLIPS, D. E. et al. The development of the blood-brain barrier in alcohol-exposed rats. Alcohol, v. 14, n. 4, p. 333-343, 1997.
POLLOCK, J. S. et al. Purification and characterization of particulate endothelium-derived relaxing factor synthase from cultured and native bovine aortic endothelial cells. Proceedings of the national academy of sciences, v. 88, n. 23, p. 10480-10484, 1991. REUSS, S.; RIEMANN, R. Distribution and projections of nitric oxide synthase neurons in the rodent superior olivary complex. Microscopy research and technique, v. 51, n. 4, p. 318-329, 2000. RUIZ‐DURÁNTEZ, E. et al. Effect of agmatine on locus coeruleus neuron activity: possible involvement of nitric oxide. British journal of pharmacology, v. 135, n. 5, p. 1152-1158, 2002. SAAVEDRA, E. E. El óxido nítrico en medicina (I). aspectos básicos. Medicina integral: medicina preventiva y asistencial en atención primaria de la salud, v.
29, n. 5, p. 199-208, 1997. SALUM, C.; PEREIRA, A. C. C I.; GUIMARÃES, E. A. D. B. Dopamine, nitric oxide and their interactions in models for the study of schizophrenia. Psicologia: reflexão e crítica, v. 21, n. 2, p. 186-194, 2008.
SAMUELS, E. R.; SZABADI, E. Functional neuroanatomy of the noradrenergic locus coeruleus: its roles in the regulation of arousal and autonomic function part I: principles of functional organisation. Current neuropharmacology, v. 6, n. 3, p. 235-253, 2008. SANTANA, J. J. et al. Origem do plexo braquial de mocós (Kerodon rupestris Wied, 1820). Brazilian journal of veterinary research and animal science, v. 40, p. 6,
2003.

86
SCALIA, F.; ARANGO, V. Topographic organization of the projections of the retina to the pretectal region in the rat. Journal of comparative neurology, v. 186, n. 2, p. 271-292, 1979. SCHAEFFER, D. F. et al. A nitrergic projection from the superior olivary complex to the inferior colliculus of the rat. Hearing research, v. 183, n. 1, p. 67-72, 2003.
SCHENBERG, L. C. et al. Functional specializations within the tectum defense systems of the rat. Neuroscience & biobehavioral Reviews, v. 29, n. 8, p. 1279-
1298, 2005. SCHMIDT, H. H. et al. Purification of a soluble isoform of guanylyl cyclase-activating-factor synthase. Proceedings of the national academy of sciences, v. 88, n. 2, p. 365-369, 1991. SCHMIDT, H. H. H. W.; WALTER, U. NO at work. Cell, v. 78, n. 6, p. 919-926, 1994. SILVA, S. F. Centros subcorticais dos sistemas visual primário e óptico acessório no mocó (kerodon rupestris): caracterização pela projeção retiniana e citoarquitetura. Natal: UFRN, 2009. 90 pag. Tese (doutorado)- Programa de Pos-
Graduação em Psicobiologia, Universidade Federal do Rio Grande do Norte. 2009. SILVA, A. M. et al. Serotonergic fibers distribution in the midline and intralaminar thalamic nuclei in the rock cavy (Kerodon rupestris). Brain research, v. 1586, p. 99-108, 2014. SMEETS, W. J. A. J.; ALONSO, J. R.; GONZÁLEZ, A. Distribution of NADPH‐diaphorase and nitric oxide synthase in relation to catecholaminergic neuronal structures in the brain of the lizard Gekko gecko. Journal of comparative neurology, v. 377, n. 1, p. 121-141, 1997.
SNYDER, S. H.; BREDT, D. S. Biological roles of nitric oxide. Scientific american, v. 266, n. 5, p. 68-71, 1992. SOARES, J. G. et al. Nuclear organization of the serotonergic system in the brain of the rock cavy (Kerodon rupestris). Journal of chemical neuroanatomy, v. 43, n. 2, p. 112-119, 2012. SOUSA, R. A.; MENEZES, A. A. L. Circadian rhythm of motor activity of the Brazilian rock cavy (Kerodon rupestris) under artificial photoperiod. Biological rhythm research, v. 37, n. 5, p. 443-450, 2006.

87
SPARROW, J. R. Inducible nitric oxide synthase in the central nervous system. Journal of molecular neuroscience, v. 5, n. 4, p. 219-229, 1994. STREILEIN, K. E. Ecology of small mammals in the semiarid brazilian caatinga.
Carnegie Museum of Natural History, 1982. STUEHR, D. J.; MARLETTA, M. A. Mammalian nitrate biosynthesis: mouse macrophages produce nitrite and nitrate in response to Escherichia coli lipopolysaccharide. Proceedings of the national academy of sciences, v. 82, n.
22, p. 7738-7742, 1985. STUEHR, D. J. et al. Purification and characterization of the cytokine-induced macrophage nitric oxide synthase: an FAD-and FMN-containing flavoprotein. Proceedings of the national academy of sciences, v. 88, n. 17, p.
7773-7777, 1991. TENG, B. Q. et al. Expression of endothelial nitric oxide synthase in human and rabbit gastrointestinal smooth muscle cells. American journal of physiology-gastrointestinal and liver physiology, v. 275, n. 2, p. G342-G351, 1998.
TENÓRIO, F. et al. Morphology of NADPH-diaphorase-positive cells in the retinoceptive layers of the developing rat superior colliculus. International journal of developmental neuroscience, v. 14, n. 1, p. 1-10, 1996. TORTORA, G. J.; GRABOWSKI, S. R. Princípios de anatomia e fisiologia. 12
edição, Guanabara koogan, 2010. TSENG, C. et al. Cardiovascular effects of nitric oxide in the brain stem nuclei of rats. Hypertension, v. 27, n. 1, p. 36-42, 1996. VELEANU, M. et al. Comparison of bNOS and chat immunohistochemistry in the laterodorsal tegmentum (LDT) and the pedunculopontine tegmentum (PPT) of the mouse from brain slices prepared for electrophysiology. Journal of neuroscience methods, v. 263, p. 23-35, 2016. VINCENT, S. R.; KIMURA, H. Histochemical mapping of nitric oxide synthase in the rat brain. Neuroscience, v. 46, n. 4, p. 755-784, 1992.

88
VOLKE, V. et al. 7-Nitroindazole, a nitric oxide synthase inhibitor, has anxiolytic-like properties in exploratory models of anxiety. Psychopharmacology, v. 131, n. 4, p. 399-405, 1997. WERNER, W.; DANNENBERG, S.; HOFFMANN, K. P. Arm-movement-related neurons in the primate superior colliculus and underlying reticular formation: comparison of neuronal activity with EMGs of muscles of the shoulder, arm and trunk during reaching. Experimental brain research, v. 115, n. 2, p. 191-205, 1997.
WINK, D. A.; MITCHELL, J. B. Chemical biology of nitric oxide: insights into regulatory, cytotoxic, and cytoprotective mechanisms of nitric oxide. Free radical biology and medicine, v. 25, n. 4, p. 434-456, 1998. XING, J. et al. Angiotensin-(1-7) inhibits neuronal activity of dorsolateral periaqueductal gray via a nitric oxide pathway. Neuroscience letters, v. 522, n. 2, p. 156-161, 2012. ZHOU, L.; ZHU, D. Neuronal nitric oxide synthase: structure, subcellular localization, regulation, and clinical implications. Nitric oxide, v. 20, n. 4, p. 223-230, 2009.
ZIELASEK, J.; HARTUNG, H. Molecular mechanisms of microglial activation. Advances in neuroimmunology, v. 6, n. 2, p. 191-222, 1996.

89

90