majdi_2011_phytochem.pdf
DESCRIPTION
fitokimTRANSCRIPT

Phytochemistry 72 (2011) 1739–1750
Contents lists available at ScienceDirect
Phytochemistry
journal homepage: www.elsevier .com/locate /phytochem
Biosynthesis and localization of parthenolide in glandular trichomesof feverfew (Tanacetum parthenium L. Schulz Bip.)
Mohammad Majdi a,1, Qing Liu b,1, Ghasem Karimzadeh a, Mohammad Ali Malboobi c, Jules Beekwilder d,Katarina Cankar d, Ric de Vos d, Sladjana Todorovic e, Ana Simonovic e, Harro Bouwmeester b,⇑a Plant Breeding and Biotechnology Department, Faculty of Agriculture, Tarbiat Modares University, P.O. Box 14115-336, Tehran, Iranb Laboratory of Plant Physiology, Wageningen University, Droevendaalsesteeg 1, NL-6708 PB Wageningen, The Netherlandsc National Institute of Genetic Engineering and Biotechnology (NIGEB), P.O. Box 14155-6343, Tehran, Irand Plant Research International, P.O. Box 16, 6700 AA Wageningen, The Netherlandse Institute for Biological Research ‘‘Siniša Stankovic’’, University of Belgrade, 11000 Belgrade, Serbia
a r t i c l e i n f o
Article history:Received 7 March 2011Received in revised form 26 April 2011Available online 26 May 2011
Keywords:Tanacetum partheniumFeverfewGermacrene A synthaseParthenolideGlandular trichome
0031-9422/$ - see front matter � 2011 Elsevier Ltd. Adoi:10.1016/j.phytochem.2011.04.021
Abbreviations: FDP, farnesyl diphosphate; MVA, mrene A synthase; Tp, Tanacetum parthenium; CarS, carapid amplification of cDNA ends; RGE, relative gene4,11-diene synthase; SEM, scanning electron microscoacid.⇑ Corresponding author. Tel.: +31 317480528; fax:
E-mail address: [email protected] (H. Bo1 These authors contributed equally to this work.
a b s t r a c t
Feverfew (Tanacetum parthenium) is a perennial medicinal herb and is a rich source of sesquiterpene lac-tones. Parthenolide is the main sesquiterpene lactone in feverfew and has attracted attention because ofits medicinal potential for treatment of migraine and cancer. In the present work the parthenolide con-tent in different tissues and developmental stages of feverfew was analyzed to study the timing and local-ization of parthenolide biosynthesis. The strongest accumulating tissue was subsequently used to isolatesesquiterpene synthases with the goal to isolate the gene encoding the first dedicated step in partheno-lide biosynthesis. This led to the isolation and charachterization of a germacrene A synthase (TpGAS) andan (E)-b-caryophyllene synthase (TpCarS). Transcript level patterns of both sesquiterpene synthases wereanalyzed in different tissues and glandular trichomes. Although TpGAS was expressed in all aerial tissues,the highest expression was observed in tissues that contain high concentrations of parthenolide and inflowers the highest expression was observed in the biosynthetically most active stages of flower devel-opment. The high expression of TpGAS in glandular trichomes which also contain the highest concentra-tion of parthenolide, suggests that glandular trichomes are the secretory tissues where parthenolidebiosynthesis and accumulation occur.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction cal activities and uses for humans, including antimicrobial, anti-
Terpenoids are the largest class of plant secondary metaboliteswith over 20,000 compounds known (Davis and Croteau, 2000).Sesquiterpene lactones are a major class of terpenoids that arisefrom the assembly of a 15-carbon skeleton into bisabolane, cupara-ne, cadinane, humulane and germacrane backbones and the addi-tion of functional groups to these backbones (Chappell et al.,2010). They are particularly abundant in the Asteraceae familyand are biologically significant because of key roles in plant envi-ronment interaction, for example as chemical defense compoundsagainst pathogens and insects (Chappell et al., 2010; Cheng et al.,2007; Hristozov et al., 2007). Sesquiterpenes have various biologi-
ll rights reserved.
evalonic acid; GAS, germac-ryophyllene synthase; RACE,
expression; ADS, amorpha-py; DHA, dihydroartemisinic
+31 317418094.uwmeester).
cancer and anti-inflammatory properties (Li-Weber et al., 2002;Saranitzky et al., 2009; Trusheva et al., 2010). Feverfew(2n = 2x = 18) is a perennial herb belonging to the Asteraceae thathas been used traditionally as a fever-reducer (Palevitch et al.,1997). The most important constituent (up to 85% of total sesqui-terpenes) among the 30 sesquiterpene lactones identified in fever-few is parthenolide (Brown et al., 1997; Cretnik et al., 2005;Williams et al., 1995). Parthenolide (Fig. 1) is a germacranolide lac-tone which recently raised quite a lot of attention due to its medic-inal value and pharmacological activities, especially as a migraineprophylaxis agent and also for treatment of cancer (Knight, 1995;Lesiak et al., 2010; Li-Weber et al., 2002; Palevitch et al., 1997;Pfaffenrath et al., 2002; Vogler et al., 1998).
Considering these promising applications, the need for par-thenolide may be increasing in the near future. In order to in-crease the availability of parthenolide several approaches canbe envisaged, such as heterologous engineering into other plantspecies or microbial platforms. In order to be able to do soknowledge about the biosynthetic pathway of parthenolide ismandatory.

OPP
OH OCOOHOH
OO
4 6
15
3
2
1
10
9
8
712
1311
O 14
O
O
5
OHO
OO
OO
O
O
(+)-germacrene A
(+)-costunolideparthenolide germacranoic acid
germacranolides
eudesmanolides
guaianolides
1 2 3
5a
4
5b6
Mevalonate pathway
IPPIDI
DMAPP
FPPS
x 2
Farnesyl diphosphate (FPP)
Parthenolide
Fig. 1. Presumed biosynthetic pathway of parthenolide; step 1 is catalyzed by germacrene A synthase; steps 2, 3 and 4 are catalyzed by germacrene A oxidase; step 5a, 5b:costunolide synthase; step 6: costunolide epoxidase/parthenolide synthase.
1740 M. Majdi et al. / Phytochemistry 72 (2011) 1739–1750
Sesquiterpene lactones are mostly derived from the mevalonicacid (MVA) pathway (van Klink et al., 2003). They are classifiedon the basis of their carbon skeletons as germacranolides, guaia-niolides, pseudoguanolides and eudesmanolides (Fig. 1) (de Krakeret al., 1998). Parthenolide has a germacranolide backbone struc-ture and costunolide has been proposed to be the common precur-sor of all germacranolide-derived sesquiterpene lactones (Fig. 1)(de Kraker et al., 2002). The first committed step in the biosynthe-sis of costunolide is the cyclization of farnesyl diphosphate (FDP) togermacrene A by a germacrene A synthase (Bouwmeester et al.,2002; de Kraker et al., 1998). Recently two research groups showedthat the cytochrome P450, germacrene A oxidase, is responsible forthe conversion of germacrene A to germacrenoic acid (germacra-1(10),4,11(13)-trien-12oic acid) (Fig. 1) (Cankar et al., 2011;Nguyen et al., 2010).
To verify this pathway in feverfew and to identify the genesmissing in the parthenolide pathway, it is crucial to study the spa-tial and temporal regulation of the pathway. The biosynthesis andaccumulation of secondary metabolites in plants is closely associ-ated to the transcription level of the relevant genes in organs, tis-sues, as well as in different developmental stages. Biosynthesis andaccumulation of metabolites may occur at the same place (cells,tissues) or be spatially separated. For example, biosynthesis as wellas accumulation of artemisinin have been observed in the glandu-lar trichomes of Artemisia annua (Bertea et al., 2006; Teoh et al.,2006). In the Asteraceae, several different specialized accumula-
tion/biosynthesis structures for secondary metabolites have beenreported e.g. cavities in Solidago canadensis (Curtis and Lersten,1990; Cury and Appezzato-da-Gloria, 2009), ducts in Ambrosia tri-fida (Curtis, 1988), laticifers in Lactuca sativa (Esau, 1965; Sessaet al., 2000) and glandular trichomes in Artemisia annua (Kelseyand Shafizadeh, 1980).
Glandular trichomes are specialized structures consisting ofusually 6–10 cells, that produce secondary metabolites whichare stored in a sub-cuticular cavity on top of the trichome,such that the phytotoxic secondary metabolites are stored awayfrom primary metabolism (Wagner, 1991). Glandular tric-homes have been used in several species of the Asteraceae toelucidate the biosynthesis of terpenes, using the fact that theyare the most active or even exclusive organ for biosynthesise.g. in A. annua (Bertea et al., 2006; Covello et al., 2007; Maeset al., 2011; Olsson et al., 2009) and Helianthus annuus (Gopfertet al., 2009).
The objectives of the present investigation were to determinethe developmental and spatial regulation of parthenolide biosyn-thesis in feverfew. This detailed analysis should provide the crucialknowledge necessary for further characterization of the partheno-lide biosynthetic pathway, for example to identify the right tissueand developmental stage to create an EST library that can be usedfor gene discovery. In addition, we set out to identify and charac-terize the sesquiterpene synthase that catalyses the first dedicatedstep in parthenolide biosynthesis.
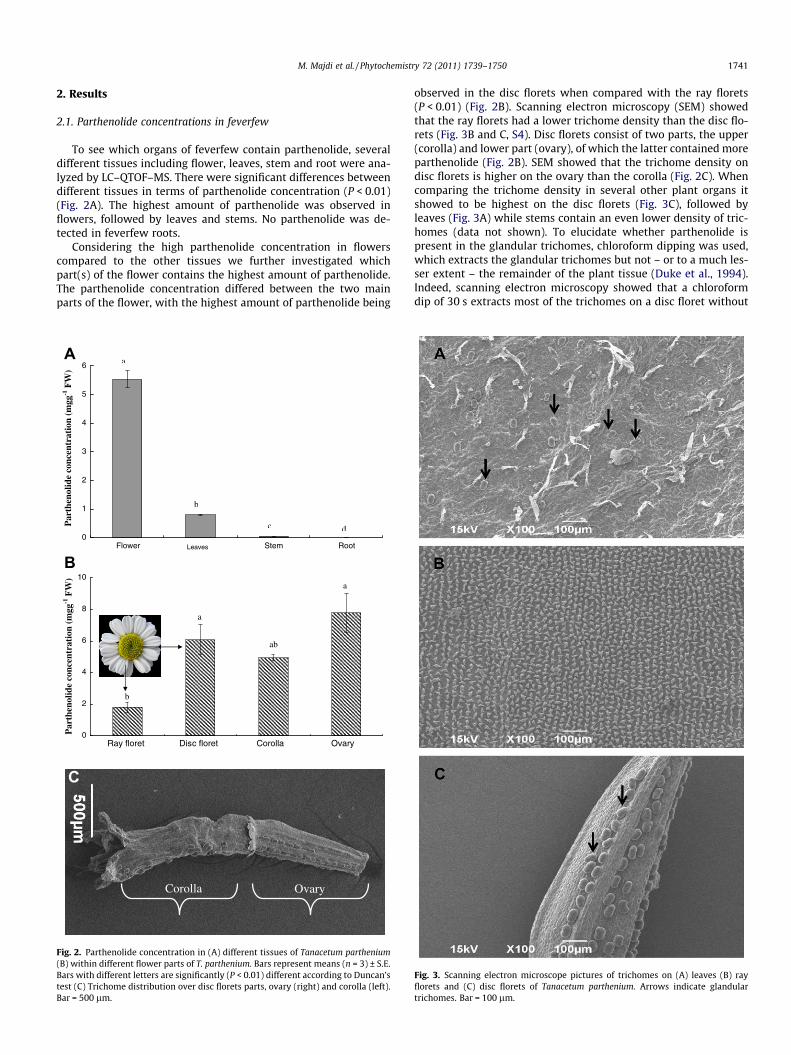
M. Majdi et al. / Phytochemistry 72 (2011) 1739–1750 1741
2. Results
2.1. Parthenolide concentrations in feverfew
To see which organs of feverfew contain parthenolide, severaldifferent tissues including flower, leaves, stem and root were ana-lyzed by LC–QTOF–MS. There were significant differences betweendifferent tissues in terms of parthenolide concentration (P < 0.01)(Fig. 2A). The highest amount of parthenolide was observed inflowers, followed by leaves and stems. No parthenolide was de-tected in feverfew roots.
Considering the high parthenolide concentration in flowerscompared to the other tissues we further investigated whichpart(s) of the flower contains the highest amount of parthenolide.The parthenolide concentration differed between the two mainparts of the flower, with the highest amount of parthenolide being
0
1
2
3
4
5
6
Flower Stem Root
a
b
c d
Par
then
olid
e co
ncen
trat
ion
(mgg
-1 F
W)
A
Par
then
olid
e co
ncen
trat
ion
(mgg
-1 F
W)
0
2
4
6
8
10
Ray floret Disc floret Corolla Ovary
b
a
ab
a
B
C
Corolla Ovary
Leaves
Fig. 2. Parthenolide concentration in (A) different tissues of Tanacetum parthenium(B) within different flower parts of T. parthenium. Bars represent means (n = 3) ± S.E.Bars with different letters are significantly (P < 0.01) different according to Duncan’stest (C) Trichome distribution over disc florets parts, ovary (right) and corolla (left).Bar = 500 lm.
observed in the disc florets when compared with the ray florets(P < 0.01) (Fig. 2B). Scanning electron microscopy (SEM) showedthat the ray florets had a lower trichome density than the disc flo-rets (Fig. 3B and C, S4). Disc florets consist of two parts, the upper(corolla) and lower part (ovary), of which the latter contained moreparthenolide (Fig. 2B). SEM showed that the trichome density ondisc florets is higher on the ovary than the corolla (Fig. 2C). Whencomparing the trichome density in several other plant organs itshowed to be highest on the disc florets (Fig. 3C), followed byleaves (Fig. 3A) while stems contain an even lower density of tric-homes (data not shown). To elucidate whether parthenolide ispresent in the glandular trichomes, chloroform dipping was used,which extracts the glandular trichomes but not – or to a much les-ser extent – the remainder of the plant tissue (Duke et al., 1994).Indeed, scanning electron microscopy showed that a chloroformdip of 30 s extracts most of the trichomes on a disc floret without
Fig. 3. Scanning electron microscope pictures of trichomes on (A) leaves (B) rayflorets and (C) disc florets of Tanacetum parthenium. Arrows indicate glandulartrichomes. Bar = 100 lm.

1742 M. Majdi et al. / Phytochemistry 72 (2011) 1739–1750
inducing visible damage to the epidermal cells (Fig. 4A). Chloro-form-dipped disc florets contained significantly (P < 0.01) less par-thenolide than non-dipped disc florets (Fig. 4B). More than 80% ofthe parthenolide was extracted from disc florets by the 30-s chlo-roform dip.
2.2. Isolation of sesquiterpene synthases
The degenerate primer approach followed by 30- and 50-rapidamplification of cDNA ends (RACE), resulted in the isolation oftwo full-length feverfew sesquiterpene synthase cDNAs, TpGAS(Gene Bank Accession Number; JF819848) and TpCarS (Gene BankAccession Number; JF819849). The TpGAS ORF showed a lengthof 1677 bp encoding 559 amino acids. A Blast search in GenBankrevealed that TpGAS has high homology with other GASs of theAsteraceae, such as germacrene A synthase short form fromChichorium intybus (highest identity, 87%), germacrene A synthaseLTC1 from Lactuca sativa, germacrene A synthase 1 from H. annuus,germacrene A synthase from Crepidiastrum sonchifolium, germac-rene A synthase from A. annua and germacrene A synthase longform from C. intybus (lowest identity, 73%). The molecular weightof the TpGAS protein was calculated to be 64.5 kDa with an isoelec-tric point (pI) of 5.03. The deduced TpGAS protein alignment withother germacrene A synthases is shown in Fig. 5A. Phylogeneticanalysis using ClustalW (http://www.ebi.ac.uk/Tools/clustalw2/index.html) showed that TpGAS and A. annua GAS are in the samecluster and have the shortest evolutionary distance from eachother. In spite of the higher similarity between TpGAS and shortform GAS from chicory they have been categorized in differentclusters (Fig. 5B). The TpCarS ORF has a length of 1647 bp encodingfor 549 amino acids. Blast search in GenBank showed high identity(93%) with (E)-b-caryophyllene synthase from A. annua and 61%with GAS from Solidago canadensis. The molecular weight of thepredicted TpCarS protein was calculated to be 63.5 kDa withisoelectric point (pI) of 5.48. A protein sequence alignment and aphylogenetic tree of TpCarS with a number of other sesquiterpenesynthases are shown in Figs. S2 and S3.
2.3. Functional characterization of TpGAS and TpCarS
TpGAS and TpCarS were functionally characterized by cloning ofthe ORF in pACYCDuet™-1 vector and heterologous expression in
0
2
4
6
8
10
Disc florets Dipped florets
Par
then
olid
e co
ncen
trat
ion
(mgg
-1 F
W)
Fig. 4. Parthenolide concentration in disc florets and chloroform dipped disc floretsand corresponding Scanning Electron Microscope pictures. Bars represent means(n = 3) ± S.E. Asterisk symbol (⁄) indicates significant (P < 0.01) difference accordingto Student’s t-test.
E. coli. An enzyme assay with the recombinant TpGAS in the pres-ence of FDP, followed by GC–MS analysis showed a single peakwhich was identified as b-elemene by comparing its spectrumand retention index with library data (Fig. 6A and B). b-Elemeneis the Cope rearrangement product of germacrene A that is formedupon injection into the hot injection port of the GC–MS (de Krakeret al., 1998). A comparable enzyme assay with TpCarS generatedmostly b-caryophyllene and a trace of a-humulene (also calleda-caryophyllene) (Fig. 6C–E).
2.4. Expression of TpGAS and TpCarS
In order to assess the expression of TpGAS and TpCarS in differ-ent organs of feverfew, qRT-PCR experiments were carried outusing cDNA templates from whole flowers, leaves, stems and rootsas well as glandular tichomes. Comparison of RGE patterns forTpGAS and TpCarS revealed that the expression of TpCarS was high-er in leaves than in flowers and stems whereas the expression ofTpGAS in flowers was more than 4-fold higher than in leaves and25-fold higher than in stems (Fig. 7). Both genes were not ex-pressed in roots. The highest expression of TpGAS was observedin trichomes (RGE of more than 100), which is at least 10-fold high-er than the RGE of TpCarS in trichomes (P<0.001) (Fig. 8).
Analysis of variance showed significant differences in parthe-nolide concentration of ovaries of different developmental stagesof flowers (P < 0.001) (Fig. 9A). Mean comparison with Duncan’stest showed that the parthenolide concentration significantly in-creased with flower developmental stage, reaching a peak atstage 5 (2-fold higher than in stage 2) and subsequently declininggradually during stage 6 and 7 (Fig. 9A). The parthenolide con-centration in stage 7 was still higher than in stage 2 (Fig. 9A).Analysis of variance showed that also for TpGAS expression sig-nificant differences existed in the relative gene expression(RGE) in ovaries of different developmental stages (P < 0.001)(Fig. 9B). RGE increased significantly with flower developmentalstage, reaching a peak at stage 3 and 4 (about 2-fold increasecompared with stage 2) then strongly decreasing in stage 5(>4-fold lower than in stages 3 and 4). The lowest RGEs were ob-served for stage 6 and 7 ovaries. TpCarS displayed a stable, lowexpression pattern (RGE < 1) in all different developmental stages(Fig. 9B).
3. Discussion
3.1. Parthenolide is mainly accumulating in glandular trichomes
Our work show that the parthenolide present in feverfew is par-ticularly concentrated in the flowers, which is in accordance withthe results of others (Awang et al., 1991; Majdi et al., 2010). Withinthe flowers, the disc florets contain a higher concentration of par-thenolide than the ray florets which coincide with a higher glandu-lar trichome density on the disc florets. Within the disc florets theovary contains a higher concentration of parthenolide than the cor-olla and also here this coincides with a higher glandular trichomedensity on the former (Fig. 2B and C). All this suggests that theglandular trichomes are likely the place where parthenolide accu-mulation occurs. The mild chloroform dipping extraction method,which extracted about 80% of parthenolide from the disc floretswithout visible damage to the epidermis, further proved the pres-ence of parthenolide in the glandular trichomes (Fig. 4). The pres-ence of sesquiterpene lactones has usually been associated withother structures such as cavities, ducts and laticifers (Curtis,1988; Curtis and Lersten, 1990; Cury and Appezzato-da-Gloria,2009; Esau, 1965; Kelsey and Shafizadeh, 1980). On the other hand,the presence of sesquiterpenes, including oxidized derivatives, in

Fig. 5. (A) Multiple sequence alignment of TpGAS (Gene Bank Accession Number: JF819848) with other germacrene A synthases (GAS) using ClustalW (Bio Edit software) and(B) Phylogenetic tree analysis of TpGAS with other GASs from Asteraceae family; Aa [Artemisia annua] ABE03980.1; Ci.SF [Cichorium intybus short form] AF498000_1; Ci.LF[Cichorium intybus long form] AF497999_1; Ha [Helianthus annuus] AAY41421.2; Ls, [Lactuca sativa] AF489964_1; Sc [Solidago canadensis] CAC36896.1; Cs [Crepidiastrumsonchifolium] ABB00361.1; Pc [Pogostemon cablin] AAS86321.1; 5-Epi-aristolochene synthase [Nicotiana tabaccum], AAA19216.
M. Majdi et al. / Phytochemistry 72 (2011) 1739–1750 1743
glandular trichomes has been reported in other plants as well suchas A. annua (Bertea et al., 2006), Solanum habrochaites (Gianfagna
et al., 1992), Mentha � piperita (McCaskill and Croteau, 1995) andSolanum lycopersicum (Schilmiller et al., 2010).

Fig. 6. GC–MS analysis of the products of recombinantly expressed feverfew sesquiterpene synthases using FDP as substrate. (A) Product of TpGAS in vitro assay is b-elemene(a cope-rearrangement product of germacrene A). (B) Mass spectrum of b-elemene. (C) Products of TpCarS in vitro assay are b-caryophyllene and a-caryophyllene (a-humulene); (D) Mass spectrum of b-caryophyllene. (E) Mass spectrum of a-caryophyllene (a-humulene).
1744 M. Majdi et al. / Phytochemistry 72 (2011) 1739–1750
3.2. TpGAS expression reveals glandular trichomes as the site ofparthenolide biosynthesis
As discussed above, the parthenolide concentration is associ-ated with the distribution of glandular trichomes in several differ-ent organs. The highest density of glandular trichomes occurred inflowers (ovaries) followed by leaves and stems and the same pat-tern was observed for the parthenolide concentration (Figs. 2 and3) as well as the expression of TpGAS (Fig. 7). TpGAS is encodingthe enzyme that highly likely catalyses the first step in partheno-lide biosynthesis, germacrene A synthase (Fig. 1). The expression
of TpCarS – another feverfew sesquiterpene synthase – showed avery different profile and was for example highest in leaves(Fig. 7). Also parthenolide concentration and TpGAS expression intrichomes – isolated from different developmental stages of ova-ries – correlated, in contrast to TpCarS of which expression didnot correlate with parthenolide concentration (Fig. 9). The associa-tion of parthenolide with glandular trichomes is further substanti-ated by the fact that parthenolide was not detected in roots whichdo not have glandular trichomes (Figs. 2 and 7). Also closely relatedplant species belonging to the Asteraceae such as A. annua makeand store sesquiterpene lactones in the glandular trichomes

Rel
ativ
e ge
ne e
xpre
ssio
n (R
GE
)
0
0.2
0.4
0.6
0.8
1
1.2
1.4
Flower Leave Stem Root
TpGAS
TpCarS
Fig. 7. Relative gene expression (RGE) of TpGAS and TpCarS in different tissues ofTanacetum parthenum. Real-time qPCR was based on the Ct values (see Section 5).The Ct value for each sample was normalized using the housekeeing gene TpActin.
0
20
40
60
80
100
120
Rel
ativ
e ge
ne e
xpre
ssio
n (R
GE
)
*
Isolated glandular trichomes
TpGAS TpCarS
Fig. 8. Relative gene expression (RGE) of TpGAS and TpCarS in purified glandulartrichomes (see insert). Bars represent means (n = 3) ± S.E. ⁄Indicates significant(P < 0.01) difference according to Student’s t-test. Real-time qPCR was based on theCt values (see Section 5). The Ct value for each sample was normalized using thehousekeeing gene TpActin.
M. Majdi et al. / Phytochemistry 72 (2011) 1739–1750 1745
(Bertea et al., 2006; Covello et al., 2007). Consistent with our re-sults, the relationship between the density of glandular trichomesand the essential oil content has also been reported in other plantspecies (Bertea et al., 2006; Maffei et al., 1989; McCaskill andCroteau, 1999). In feverfew, TpGAS is only expressed in the aerialparts in which also parthenolide exists, while the expression ofGAS in roots of other Asteraceae – that produce sesquiterpenelactones in their roots – has been reported e.g. C. intybus andH. annuus (Bouwmeester et al., 2002; Göpfert et al., 2009). In thelatter case, not only has different regulation of transcription ofbiosynthetic genes allowed for biosynthesis of sesquiterpenelactones in the roots but also storage of the sesquiterpene lactoneswas adapted to accommodate for the absence of trichomes on theroots (Göpfert et al., 2009).
The much higher expression of TpGAS in trichomes when com-pared with other tissues (Fig. 7) shows that TpGAS is a glandulartrichome specific gene. In consistence with our work there havebeen many reports on different plant species which havedemonstrated a strong correlation between terpene amount (oremission) and the level of the corresponding mRNA, indicating thatterpenoid biosynthesis is mainly regulated at the transcript level(Nagegowda, 2010). Also gene expression analysis in different celltypes of glandular trichomes of A. annua including apical, sub-apical and mesophyl cells showed that the expression of threeenzymes specific to the artemisinin biosynthetic pathway areactive only in the apical cells (Olsson et al., 2009). These apical cells
are likely the ones in which artemisinin biosynthesis occurs, closeto the subcuticular cavity where artemisinin and/or its precursorsare stored. In contrast, FDP synthase which is not specific only forartemisinin biosynthesis was expressed in all the glandulartrichomes cell types (Olsson et al., 2009). All this suggests that alsofor uncovering the regulation and site of parthenolide biosynthesis,analysis of TpGAS expression and its localisation is a reliableapproach.
3.3. Developmental changes in TpGAS expression and parthenolideconcentration
Analysis of parthenolide concentration and TpGAS expression indifferent developmental stages of flowers showed that partheno-lide accumulation and TpGAS expression closely correlate.Although TpCarS gene expression was observed in the trichomesin several flower developmental stages, both the level of transcriptand the pattern of transcript change do not show any relationshipwith parthenolide biosynthesis, excluding that TpCarS is involvedin parthenolide biosynthesis. Parthenolide accumulation is devel-opmentally regulated and displays three distinct phases (Fig. 9).The first phase (stages 1–4) can be considered as ‘‘parthenolideaccumulation phase’’ in which there is a direct relationship be-tween the increase in TpGAS expression and the increase in parthe-nolide concentration (Fig. 9). The second phase (stage 5) can beconsidered as ‘‘parthenolide saturation phase’’ in which the par-thenolide concentration reaches its maximum while the expres-sion of TpGAS is down-regulated compared with the previousphase. This may indicate a feed back regulatory mechanism or adevelopmental program controlling parthenolide accumulationby down-regulating the expression of the gene(s) in the biosyn-thetic pathway of parthenolide, in this case TpGAS. Also feedbackinhibition of artemisinin biosynthesis by repression of ADS and CY-P71AV1 expression by artemisinic acid and artemisinin has re-cently been suggested (Arsenault et al., 2010). Considering therelationship between flower development and glandular trichomedevelopment (Göpfert et al., 2005; Werker, 2000), this phase mayrepresent physiological maturity of the glandular trichomes whichcoincides with the end of cell expansion in floret development. Thethird phase (stages 6 and 7) can be considered as ‘‘parthenolidedecreasing phase’’, in which a decrease in parthenolide concentra-tion was observed, along with the strongly reduced expression ofTpGAS (Fig. 9). The TpGAS expression pattern resembles that ofthe H. annuus germacrene A synthase. In this species, up-regulationof the germacrene A synthase during the secretory stage anddown-regulation during the post-active secretory phase has beenreported confirming developmental stage-specific expression inglandular trichomes (Gopfert et al., 2009). The high expression ofTpGAS during the parthenolide accumulation phase and the contin-ued expression of TpGAS during the parthenolide decreasing phaseshow the necessity of active and continuous transcription of TpGAS(and likely other genes downstream of TpGAS) for parthenolideaccumulation. This is consistent with the high positive correlationbetween the expression of ADS and CYP71AV1 and the accumula-tion of the artemisinin precursor dihydroartemisinic acid in A. an-nua (Arsenault et al., 2010). Also in peppermint a high rate ofmonoterpene biosynthesis and high enzyme activities were ob-served during early leaf development (Gershenzon, 1994). Thestrong developmental changes in parthenolide concentration thatwe find (Fig. 9) are supported by the literature. For example, Omi-dbaigi et al. (2007) has reported developmental changes in fever-few essential oil content and composition in flowers at variousharvesting times. Developmental regulation appears to be animportant factor in parthenolide biosynthesis and this is consistentwith other reports on terpene biosynthesis (Arsenault et al., 2010;Kim et al., 2008; Lommen et al., 2006; McConkey et al., 2000). The

0
2
4
6
8
10
12
Ovary1 Ovary2 Ovary3 Ovary4 Ovary5 Ovary6 Ovary7
Ovary1 Ovary2 Ovary3 Ovary4 Ovary5 Ovary6 Ovary7
d
bcbc
a
b
Not determined
cP
arth
enol
ide
conc
entr
atio
n (m
gg-1 F
W)
0
1
2
3
4
5
6
Rel
ativ
e ge
ne e
xpre
ssio
n (R
GE
)
cc
Not determined
ab
a
a
bc
aab
ab b bab
A
B
TpGAS
TpCarS
Fig. 9. (A) Parthenolide concentration and (B) relative gene expression of TpGAS and TpCarS in ovaries from different developmental stages of Tanacetum parthenium flowers.Bars represent means (n = 3) ± S.E. Bars with different letters are significantly (P < 0.05) different according to Duncan’s test.
1746 M. Majdi et al. / Phytochemistry 72 (2011) 1739–1750
decrease in parthenolide concentration during the parthenolidedecreasing phase is more difficult to explain. Volatilization andleaching have been postulated as possible mechanisms that causea loss of sesquiterpene lactones (Gershenzon, 1994). However, par-thenolide is not volatile and leaching is not likely to occur in agreenhouse or climate room. Possibly, parthenolide decrease inthe decreasing phase is caused by a biodegradation mechanismor further conversion to another compound(s). Further conversionof parthenolide to epoxyparthenolide and hydroxyparthenolide inAnvillea radiata a member of the Asteraceae has been reported(Hassany et al., 2004). Biodegradation or further conversion couldalso explain the diurnal changes in parthenolide content (decreaseduring the night) in feverfew leaves (Fonseca et al., 2005, 2006).The decrease in parthenolide concentration in phase 3 could alsoindicate trichome collapse, as has been reported for A. annua(Arsenault et al., 2010; Lommen et al., 2006). Lommen et al.(2006) have proposed that trichome collapse in A. annua is respon-sible for the decrease in artemisinin content during the later stagesof leaf development.
4. Conclusions
Cloning of TpGAS and the study of its transcript patterns in dif-ferent tissues and developmental stages of flowers in feverfewshows that the expression of TpGAS is closely associated with par-thenolide biosynthesis. The high expression of TpGAS in glandulartrichomes and the localization of parthenolide in the trichomesshow that the trichomes are the specific site of parthenolide bio-synthesis and accumulation. Hence, isolation of feverfew glandulartrichomes will be a powerful tool to generate an EST library, whichcan be used for high throughput transcript sequencing and identi-fication of additional genes involved in the biosynthetic pathway ofparthenolide. This approach has been successful in other plant spe-cies and has lead to the identification of genes involved in the bio-synthesis of several secondary metabolites (Lange et al., 2000;McConkey et al., 2000; Iijima et al., 2006; Bertea et al., 2006; Nagelet al., 2008). Transcript profiling in combination with metaboliteprofiling (untargeted large scale metabolomics) of glandular tric-homes isolated from flowers in different developmental stages

M. Majdi et al. / Phytochemistry 72 (2011) 1739–1750 1747
could also provide insight into the regulation of accumulation andloss of parthenolide in Tanacetum parthenium.
5. Experimental
5.1. Plant growth condition
Tanacetum parthenium L. plants were grown under controlledconditions in a climate room with a 16 h day length supplied witha photosynthetic photon flux density of 320 lmol m�2 s�1 withday/night temperature of 24/18 �C. Samples for phytochemicaland gene expression analysis were immediately frozen in liquidnitrogen and then stored at �80 �C until further use.
5.2. Developmental stages of flower determination
Seven developmental stages of feverfew flower developmentwere defined based on a scale developed for Chrysanthemum ciner-ariaefolium (Head, 1966). Stage 1 (well developed closed buds),stage 2 (ray florets vertical), stage 3 (ray florets horizontal and firstrow of disc florets open), stage 4 (approximately three rows of discflorets open), stage 5 (all disc florets open, fully mature), stage 6(early senescence; color of disc florets diminishing but ray floretsstill intact), stage 7 (late senescence; disc florets dried out) (Head,1966). Ovaries were isolated from all these stages except stage 1.
5.3. Parthenolide measurement
Plant materials (flower, leave, stem, root and ovary) wereground to a fine powder using a mortar and pestle in liquid nitro-gen. Non-volatile compounds were analyzed using a protocol foruntargeted metabolomics of plant tissues (de Vos et al., 2007).
Three biological replicates were used for each sample. In brief,100 mg of sample for each replicate (leaves from one month oldplants in the vegetative stage, whole stage 3 flowers, ovaries fromdifferent flower developmental stages, stems and roots) wereground in liquid nitrogen and extracted with 0.3 mL methanol:for-mic acid (1000:1, v/v). The extracts were briefly vortexed and thensonicated for 15 min. Subsequently, the extracts were centrifugedand filtered through a 0.2 lm inorganic membrane filter (RC4, Sar-torius, Germany). LC-PDA-MS analysis was performed using aWaters Alliance 2795 HPLC connected to a Waters 2996 PDA detec-tor and a QTOF Ultima V4.00.00 mass spectrometer (Waters, MSTechnologies, UK) operating in positive ionization mode. The col-umn used was an analytical column (2.0 � 150 mm; Phenomenex,USA) with a C18 pre-column (2.0 � 4 mm; Phenomenex, USA). De-gassed eluent A (ultra pure water:formic acid; 1000:1, v/v) andeluent B (acetonitril:formic acid; 1000:1, v/v) were pumped at0.19 mL min�1 into the HPLC system. The gradient started at 5%B and increased linearly to 35% B in 45 min. Then the columnwas washed and equilibrated for 15 min before the next injection.
An authentic standard of parthenolide (Sigma–Aldrich, USA)was used to make a calibration curve by injecting different concen-trations of parthenolide for quantification (Fig. S1).
5.4. Chloroform dipping
To 100 mg of ovaries from feverfew in a 2 mL Eppendorf vial,1 mL of chloroform was added followed by vortexing for 30 s andshort centrifugation. The chloroform phase was removed and thechloroform evaporated in a fume hood for 3 min. Then the parthe-nolide content of dipped and non-dipped ovaries was measured byLC–MS as described above.
5.5. Isolation of glandular trichomes and RNA extraction
Ovaries (about 300 flowers) were separated from the rest of theflower for trichome isolation. About 3 g of ovaries were placed in a50 mL pre-cooled Greiner tube and 10 mL liquid nitrogen wasadded. The tube was covered using a 20 lm mesh followed by vor-texing for 1 min and addition of another 10 mL liquid nitrogen. Thevortexing was repeated 6–10 times. The isolated trichomes wereseparated from the other cells and debris by passing through a120 lm mesh and collection on a 20 lm mesh. RNA was isolatedfrom the glandular trichomes using the Spectrum™ Plant TotalRNA isolation kit (Sigma, USA).
5.6. RNA extraction and cDNA synthesis for sesquiterpene synthasesisolation
Flowers of Tanacetum parthenium were used for RNA extraction.Tripure isolation reagent (Roche, Mannheim, Germany) was usedfor extraction of total RNA according to the manufacturer’s instruc-tions with minor modifications. Isolated RNA was subsequentlytreated with DNAse I (Invitrogen, USA) and then purified througha silica column using the RNeasy RNA clean up kit (Qiagen#74104, USA). The quantity and quality of RNA was determinedusing a NanoDrop (NanoDrop Technologies, USA) and agarose gelelectrophoresis. The reverse transcription reaction was carriedout with the Taqman Reverse Transcription Reagent kit (AppliedBiosystems, USA) using 1 lg of total RNA and 50 lM oligo dTaccording to the manufacturer’s instructions.
To clone sesquiterpene synthases, a homology-based PCR clon-ing strategy was used. Sequence alignments of published Astera-ceae sesquiterpene synthase cDNAs in public databases wereused to design degenerate oligonucleotide primers. Primer pairswere designed corresponding to the conserved regions, avoidingdegeneracy at the 30 end: forward primer 50-TATWCNGTNCAYCGTCTYGG 30 and reverse primer 50-AYYKCATATCRTTCCACCAC30. The temperature program for PCR was initial denaturation at94 �C for 5 min, followed by 30 cycles of 94 �C for 30 s, 50 �C for30 s, 72 �C for 1 min and the final elongation at 72 �C for 5 min.
A second set of degenerate primers (set 2 degenerate primers)consisted of: forward primer 50-GAY GAR AAY GGI AAR TTY AARGA-30 and reverse primer 50-CCR TAI GCR TCR AAI GTR TCR TC-30
(Wallaart et al., 2001). The temperature program for PCR was ini-tial denaturation at 94 �C for 3 min, followed by 35 cycles of94 �C for 30 s, 48 �C for 1 min, 72 �C for 1 min and the final elonga-tion at 72 �C for 5 min. The resulting purified fragments werecloned into pGEM-T Easy Vector (Promega, USA) and sequenced.
5.7. cDNA extension using 50 and 30-RACE
50-RACE and 30-RACE were performed using the SMART-RACEcDNA amplification kit (Clontech, USA) according to the manufac-turer’s protocol. The first strand cDNA for 50 and 30 RACE were syn-thesized from 1 lg of total RNA extracted from ovaries accordingto the protocol (Clontech, USA). According to the partial sequenceof the TpGAS fragment (550 bp) amplified using degenerate prim-ers set 1, high annealing temperature (70 �C) primers were de-signed for both 50 and 30 RACE. Nested primers were designedaccording to the protocol suggestions. The Gsp1-GAS (Gene specificprimer 50) and NGsp1-GAS (Nested gene specific primer 50) for 50-RACE were 50-GCATCCCTTGATGGAATGGTCTCCTT-30 and 50-CCCAACATACCCCTCACATCACTCG-30, respectively. The Gsp2-GAS (Genespecific primer 30) and NGsp2-GAS (Nested gene specific primer30) for 30-RACE were 50-GTGATGTGAGGGGTATGTTGGGCTTA-30
and 50-TGCACAGTTGAGAATAAGAGG-30, respectively.Two additional sesquiterpene synthase like sequences were
generated using the second set of degenerate primers. For one of

1748 M. Majdi et al. / Phytochemistry 72 (2011) 1739–1750
them the 50 and 30 RACE did not produce the expected cDNA frag-ments, while the other did. The Gsp1–CarS (Gene specific primer50) and NGsp1–CarS (Nested gene specific primer 50) for 50-RACEwere 50CTTCGCCTTGCACCCCCAAGTATGT-30 and 50-CAAGCAGGCCTTCTACATCGTTGGTG-30, respectively. The Gsp2–CarS (Gene spe-cific primer 30) and NGsp2–CarS (Nested gene specific primer 30)for 30-RACE were 50-GGCCTGCTTGAGCTGTATGAGGCAAC-30 and50-TTCGCTGGCCACTGTTCTTGACGATA-30, respectively. 50-RACEand 30-RACE products were cloned into the pGEM-T easy vector(Promega) and sequenced. Assembly of the sequences from the30 RACE and 50 RACE products using SeqMan software yielded thefull-length cDNA sequences of TpGAS and TpCarS.
5.8. Heterologous expression of TpGAS and TpCarS in Escherichia coli
According to the 50 and 30 ends new primers were designedintroducing BamHI and NotI restriction sites to clone the fragmentinto pACYC-DUET-1 (Novagen). A full length of TpGAS was ampli-fied using 50 Race cDNA as a template, using primers forward 50-ACTACGGATCCGGCAGCGGTTCAAGCTACTAC-30 and reverse 50-ACCACGCGGCCGCTTACACGGGTAGAGAATCCACAA-30 (restrictionsites for BamHI and NotI underlined). cDNA amplification for theconstruction of expression plasmids was performed using the Phu-sion DNA polymerase. The full length of TpCarS was also clonedinto pACYC-DUET-1 (Novagen) using primers forward 50-ACTACG-GATCCGTCTGCTAAAGAAGAGAAAGTA-30 and reverse 50-ACCACGCGGCCGCTTATATAGGTATAGGATGAACGAG-30. For func-tional characterization the two sesquiterpene synthase ORFs[cloned into the pACYC-DUET-1 (Novagen) expression vector withan aminoterminal histidine tag] were expressed in Escherichia coliBL-21 under an isopropylthio-b-galactoside and arabinose induc-ible promoter. For the functional assay, 1 mL of transformed bacte-ria was cultured overnight at 250 rpm at 37 �C in LB supplementedwith chloramphenicol. Subsequently, 500 lL of the bacteria wereadded to 50 ml 2xYT supplemented with chloramphenicol for1.5 h on shaker incubator at 250 rpm and 37 �C. Then 50 lL 20%L-arabinose was added to induce the bacteria and the culture wasincubated overnight on a shaker at 18 �C at 250 rpm. The bacteriawere centrifuged for 10 min at 2500 rpm. The resulting pellet wasresuspend in 1 mL buffer A (50 mM Tris, pH 8; 300 mM NaCl,1.4 mM b-mercaptoethanol) followed by sonication on ice. Thesample was centrifuged for 10 min at 13,000rpm at 4 �C and thesupernatant transferred to a Qiagen Nickel column which was thencentrifuged for 2 min at 3000 rpm. The column was washed twotimes with 600 lL buffer A and then eluted with 200 lL buffer Acontaining 175 mM imidazole. One hundred microliter of this puri-fied enzyme was diluted into 800 lL assay buffer (15 mM MOPSO,pH 7.5; 12.5% (v/v) glycerol, 1 mM ascorbic acid, 0.001% (v/v)Tween-20, 1 mM MgCl2, 2 mM DTT). To the assay a final concentra-tion of 50 lM farnesyl diphosphate was added after which the as-say was overlayed with 1 mL of pentane and incubated at 30 �Cwith mild agitation for 2 h. Subsequently, the pentane phase wascollected and the assay extracted with 1 mL ethylacetate. The eth-ylacetate and pentane phases were combined, centrifuged at1200rpm and then dried using anhydrous Na2SO4 packed in a smallglass column and used for GC–MS analysis.
5.9. GC–MS analysis
Volatiles produced in the enzyme assays were analyzed by anAgilent Technologies 7890A GC, equipped with a 5975C inertMSD with Triple Axis Detector using helium as carrier gas at a flowrate of 1 ml min�1. The injector was used in splitless mode with theinlet temperature set to 250 �C. The initial temperature of 45 �Cwas gradually increased after 1 min to 300 �C by a ramp of10 �C min�1 held for 5 min at 300 �C. Peaks were identified by com-
parison of mass spectra to the Wiley mass spectra library. Kovatsindices (KI) were calculated for each peak according to the reten-tion time relative to alkane standards (Sekiwa-Iijima et al., 2001).
5.10. Gene expression analysis
First-strand cDNA was synthesized using the iScript cDNA Syn-thesis kit (BioRad) according to the manufacturer’s instructions.Gene expression analysis was done by real-time quantitative PCR(qPCR) with the iCycler iQ5 system (BioRad) (Spinsanti et al.,2006) using the iQ™ SYBR� Green Supermix master mix (Biorad)in three independent biological and two technical replicates. Actinwas used as a housekeeping gene. The DCt was calculated asfollows: DCt = Ct(TpGAS) – Ct(TpActin) and the RGE as: RGE =POWER(2;-dCt) (Livak and Schmittgen, 2001). A two step programwas used as follows: 3 min at 95 �C; 40 cycles of 10 s at 95 �C, 30 sat 55 �C followed by melting curve analysis. The following primerpairs were used: forward actin 50-CCTCTTAATCCTAAGGCTAATC-30: reverse actin 50-CCAGGAATCCAGCACAATACC-30; forward TpGAS50-TTCTCCTCTTATTCTCAACTGTGG-30; reverse TpGAS 50-TGCTATCTCGGGTACTTTCAAGG-30; forward TpCarS 50-GCATCCAGTGTTGAAAGTTAC-30; reverse TpCarS 50-GGTCATCAGAGGCATCGG-30.
5.11. Data analysis
Sequence comparison was performed by Blast Search in Gen-Bank (http://www.ncbi.nih.gov). Phylogenetic tree analysis andsesquiterpene alignment analysis were performed using ClustalW(http://www.ebi.ac.uk/Tools/clustalw2/index.html) and BioEditsoftware, respectively.
A completely randomized design (CRD) with 3 replications wasused for the comparison of the parthenolide concentration and RGEin different tissues, within flower parts and in different develop-mental stages of the flower. Mean comparison was conductedusing Duncan’s test. Student’s t-test was conducted to compareparthenolide concentration in disc florets and chloroform dippeddisc florets and for the comparison of the RGE for TpGAS and TpCarSin glandular trichomes. All the data were statistically analyzedusing SPSS statistical software.
Acknowledgements
MM acknowledges the Ministry of Sciences, Research andTechnology of Iran for awarding a fellowship. LQ, RdV, ST, AS andHB acknowledge funding by the European Commission (EU-projectTerpMed, 227448). JB and KC were funded by IBOS-ACTS053.63.305. We acknowledge Francel Verstappen, BenyaminHoushyani, Aldana Ramirez, Miriam Goedbloed, Liping Gao, andJos Molthoff for their helpful suggestions.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.phytochem.2011.04.021.
References
Arsenault, P.R., Vail, D., Wobbe, K.K., Erickson, K., Weathers, P.J., 2010. Reproductivedevelopment modulates gene expression and metabolite levels with possiblefeedback inhibition of artemisinin in Artemisia annua. Plant Physiol. 154, 958–968.
Awang, D.V.C., Dawson, B.A., Kindack, D.G., Crompton, C.W., Heptinstall, S., 1991.Parthenolide content of feverfew (Tanacetum parthenium) assessed by HPLC and1H NMR spectroscopy. J. Nat. Prod. 54, 1516–1521.
Bertea, C.M., Voster, A., Verstappen, F.W.A., Maffei, M., Beekwilder, J., Bouwmeester,H.J., 2006. Isoprenoid biosynthesis in Artemisia annua: cloning and heterologousexpression of a germacrene A synthase from a glandular trichome cDNA library.Arch. Biochem. Biophys. 448, 3–12.

M. Majdi et al. / Phytochemistry 72 (2011) 1739–1750 1749
Bouwmeester, H.J., Kodde, J., Verstappen, F.W.A., Altug, I.G., de Kraker, J.W.,Wallaart, T.E., 2002. Isolation and characterization of two germacrene Asynthase cDNA clones from chicory. Plant Physiol. 129, 134–144.
Brown, A.M.G., Edwards, C.M., Davey, M.R., Power, J.B., Lowe, K.C., 1997. Effects ofextracts of Tanacetum species on human polymorphonuclear leucocyte activityin vitro. Phytother. Res. 11, 479–484.
Cankar, K., Houwelingen, A., Bosch, D., Sonke, T., Bouwmeester, H., Beekwilder, J.,2011. A chicory cytochrome P450 mono-oxygenase CYP71AV8 for the oxidationof (+)-valencene. FEBS Lett. 585, 178–182.
Chappell, J., Coates, R.M., Lew, M., Hung-Wen, L., 2010. Sesquiterpenes.Comprehensive Natural Products II. Elsevier, Oxford, pp. 609–641.
Cheng, A.X., Lou, Y.G., Mao, Y.B., Lu, S., Wang, L.J., Chen, X.Y., 2007. Plant terpenoids:biosynthesis and ecological functions. J. Integr. Plant Biol. 49, 179–186.
Covello, P.S., Teoh, K.H., Polichuk, D.R., Reed, D.W., Nowak, G., 2007. Functionalgenomics and the biosynthesis of artemisinin. Phytochemistry 68, 1864–1871.
Cretnik, L., Skerget, M., Knez, Z., 2005. Separation of parthenolide from feverfew:performance of conventional and high-pressure extraction techniques. Sep.Purif. Technol. 41, 13–20.
Curtis, J.D., 1988. Secretory reservoirs (ducts) of two kinds in giant ragweed,Ambrosia trifida (Asteraceae). Am. J. Bot. 75, 1313–1323.
Curtis, J.D., Lersten, N.R., 1990. Oil-reservoirs in stem, rhizome, and root of Solidago-canadensis (Asteraceae, tribe Astereae). Nord. J. Bot. 10, 443–449.
Cury, G., Appezzato-da-Gloria, B., 2009. Internal secretory spaces in thickenedunderground systems of Asteraceae species. Aust. J. Bot. 57, 229–239.
Davis, E.M., Croteau, R., 2000. Cyclization enzymes in the biosynthesis ofmonoterpenes, sesquiterpenes, and diterpenes. Top Curr. Chem. 209, 53–95.
de Kraker, J.W., Franssen, M.C.R., de Groot, A., Konig, W.A., Bouwmeester, H.J., 1998.(+)-Germacrene A biosynthesis – the committed step in the biosynthesis ofbitter sesquiterpene lactones in chicory. Plant Physiol. 117, 1381–1392.
de Kraker, J.W., Franssen, M.C.R., Joerink, M., de Groot, A., Bouwmeester, H.J., 2002.Biosynthesis of costunolide, dihydrocostunolide, and leucodin. Demonstrationof cytochrome P450-catalyzed formation of the lactone ring present insesquiterpene lactones of chicory. Plant Physiol. 129, 257–268.
de Vos, R.C.H., Moco, S., Lommen, A., Keurentjes, J.J.B., Bino, R.J., Hall, R.D., 2007.Untargeted large-scale plant metabolomics using liquid chromatographycoupled to mass spectrometry. Nat. Protoc. 2, 778–791.
Duke, M.V., Paul, R.N., Elsohly, H.N., Sturtz, G., Duke, S.O., 1994. Localization ofartemisinin and artemisitene in foliar tissues of glanded and glandless biotypesof Artemisia annua L.. Int. J. Plant Sci. 155, 365–372.
Esau, K., 1965. Plant Anatomy. John Wiley and Sons, New York.Fonseca, J.M., Rushing, J.W., Rajapakse, N.C., Thomas, R.L., Riley, M.B., 2005.
Parthenolide and abscisic acid synthesis in feverfew are associated butenvironmental factors affect them dissimilarly. J. Plant Physiol. 162, 485–494.
Fonseca, J.M., Rushing, J.W., Rajapakse, N.C., Thomas, R.L., Riley, M.B., 2006. Potentialimplications of medicinal plant production in controlled environments: Thecase of feverfew (Tanacetum parthenium). Hortscience 41, 531–535.
Gershenzon, J., 1994. Metabolic costs of terpenoid accumulation in higher-plants. J.Chem. Ecol. 20, 1281–1328.
Gianfagna, T.J., Carter, C.D., Sacalis, J.N., 1992. Temperature and photoperiodinfluence trichome density and sesquiterpene content of Lycopersicon hirsutum.Plant Physiol. 100, 1403–1405.
Göpfert, J.C., Heil, N., Conrad, J., Spring, O., 2005. Cytological development andsesquiterpene lactone secretion in capitate glandular trichomes of sunflower.Plant Biol. 7, 148–155.
Göpfert, J.C., MacNevin, G., Ro, D.K., Spring, O., 2009. Identification, functionalcharacterization and developmental regulation of sesquiterpene synthases fromsunflower capitate glandular trichomes. BMC Plant Biol. 9, 86.
Hassany, B.E.l., Hanbali, F.E.l., Akssira, M., Mellouki, F., Haidour, A., Barrero, A.F.,2004. Germacranolides from Anvillea radiate. Fitoterapia 75, 573–576.
Head, S.W., 1966. A study of the insecticidal constituents in Chrysanthemumcinerariaefolium. (1) Their development in the flower head. (2) Their distributionin the plant. Pyrthrum Post 8, 32–37.
Hristozov, D., Da Costa, F.B., Gasteiger, J., 2007. Sesquiterpene lactones-basedclassification of the family Asteraceae using neural networks and k-nearestneighbors. J. Chem. Inf. Model. 47, 9–19.
Kelsey, R.G., Shafizadeh, F., 1980. Glandular trichomes and sesquiterpene lactonesof Artemisian ova (Asteraceae). Biochem. Syst. Ecol. 8, 371–377.
Kim, S.H., Chang, Y.J., Kim, S.U., 2008. Tissue specificity and developmental patternof amorpha-4, 11-diene synthase (ADS) proved by ADS promoter-driven GUSexpression in the heterologous plant, Arabidopsis thaliana. Planta Med. 74, 188–193.
Knight, D.W., 1995. Feverfew – chemistry and biological activity. Nat. Pro. Rep. 12,271–276.
Lange, B.M., Wildung, M.R., Stauber, E.J., Sanchez, C., Pouchnik, D., Croteau, R., 2000.Probing essential oil biosynthesis and secretion by functional evaluation ofexpressed sequence tags from mint glandular trichomes. Proc. Natl. Acad. Sci.USA 97, 2934–2939.
Lesiak, K., Zalesna, I., Nejc, D., Düchler, M., Czyz, M., 2010. Parthenolide, asesquiterpene lactone from the medical herb feverfew, shows anticanceractivity against human melanoma cells in vitro. Melanoma Re. 20, 21–34.
Maes, L., van Nieuwerburgh, F.C.W., Zhang, Y., Reed, D.W., Pollier, J., Vande Casteele,S.R.F., Inze, D., Covello, P.S., Deforce, D.L.D., Goossens, A., 2011. Dissection of thephytohormonal regulation of trichome formation and biosynthesis of the
antimalarial compound artemisinin in Artemisia annua plants. New Phytol. 189,176–189.
Livak, K.J., Schmittgen, T.D., 2001. Analysis of relative gene expression data usingreal-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 25,402–408.
Lommen, W.J.M., Schenk, E., Bouwmeester, H.J., Verstappen, F.W.A., 2006. Trichomedynamics and artemisinin accumulation during development and senescence ofArtemisia annua leaves. Planta Med. 72, 336–345.
Iijima, Y., Wang, G., Fridman, E., Pichersky, E., 2006. Analysis of the enzymaticformation of citral in the glands of sweet basil. Arch. Biochem. Biophys. 448,141–149.
Maffei, M., Chialva, F., Sacco, T., 1989. Glandular trichomes and essential oils indeveloping peppermint leaves. New Phytol. 111, 707–716.
Majdi, M., Karimzadeh, G., Malboobi, M.A., Omidbaigi, R., Mirzaghaderi, G., 2010.Induction of tetraploidy to feverfew (Tanacetum parthenium Schulz-Bip.):morphological, physiological, cytological, and phytochemical changes.Hortscience 45, 16–21.
McCaskill, D., Croteau, R., 1995. Monoterpene and sesquiterpene biosynthesis inglandular trichomes of peppermint (mentha x piperita) rely exclusively onplastid-derived isopentenyl diphosphate. Planta 197, 49–56.
McCaskill, D., Croteau, R., 1999. Strategies for bioengineering the development andmetabolism of glandular tissues in plants. Nat. Biotechnol. 17, 31–36.
McConkey, M.E., Gershenzon, J., Croteau, R.B., 2000. Developmental regulation ofmonoterpene biosynthesis in the glandular trichomes of peppermint. PlantPhysiol. 122, 215–223.
Li-Weber, M., Giaisi, M., Treiber, M.K., Krammer, P.H., 2002. The anti-inflammatorysesquiterpene lactone parthenolide suppresses IL-4 gene expression inperipheral blood T cells. Eur. J. Immunol. 32, 3587–3597.
Nagegowda, D.A., 2010. Plant volatile terpenoid metabolism: biosynthetic genes,transcriptional regulation and subcellular compartmentation. FEBS Lett. 584,2965–2973.
Nagel, J., Culley, L.K., Lu, Y.P., Liu, E.W., Matthews, P.D., Stevens, J.F., Page, J.E.,2008. EST analysis of hop glandular trichomes identifies an O-methyltransferase that catalyzes the biosynthesis of xanthohumol. Plant Cell20, 186–200.
Nguyen, D.T., Göpfert, J.C., Ikezawa, N., MacNevin, G., Kathiresan, M., Conrad, J.,Spring, O., Ro, D.K., 2010. Biochemical conservation and evolution ofgermacrene a oxidase in Asteraceae. J. Biol. Chem. 285, 16588–16598.
Olsson, M.E., Olofsson, L.M., Lindahl, A.L., Lundgren, A., Brodelius, M., Brodelius, P.E.,2009. Localization of enzymes of artemisinin biosynthesis to the apical cells ofglandular secretory trichomes of Artemisia annua L.. Phytochemistry 70, 1123–1128.
Omidbaigi, R., Dadman, B., Yavari, S., 2007. Various harvest times of Tanacetumperthenium cv. Zardband flowers affecting its oil content and compositions. J.Essent. Oil Bear. Plants. 10, 287–291.
Palevitch, D., Earon, G., Carasso, R., 1997. Feverfew (Tanacetum parthenium) as aprophylactic treatment for migraine: a double-blind placebo-controlled study.Phytother. Res. 11, 508–511.
Pfaffenrath, V., Diener, H.C., Fischer, M., Friede, M., Henneicke-von Zepelin, H.H.,2002. The efficacy and safety of Tanacetum parthenium (feverfew) in migraineprophylaxis – a double-blind, multicentre, randomized placebo-controlleddose-response study. Cephalalgia 22, 523–532.
Saranitzky, E., White, M., Baker, E.L., Baker, W.L., Coleman, C.I., 2009.Feverfew for migraine prophylaxis: a systematic review. J. Dietary Suppl.6, 91–103.
Schilmiller, A.L., Miner, D.P., Larson, M., McDowell, E., Gang, D.R., Wilkerson, C.,Last, R.L., 2010. Studies of a biochemical factory: tomato trichome deepexpressed sequence tag sequencing and proteomics. Plant Physiol. 153,1212–1223.
Sekiwa-Iijima, Y., Aizawa, Y., Kubota, K., 2001. Geraniol dehydrogenase activityrelated to aroma formation in Ginger (Zingiber officinale Roscoe). J. Agri. FoodChem. 49, 5902–5906.
Sessa, R.A., Bennett, M.H., Lewis, M.J., Mansfield, J.W., Beale, M.H., 2000. Metaboliteprofiling of sesquiterpene lactones from Lactuca species - Major latexcomponents are novel oxalate and sulfate conjugates of lactucin and itsderivatives. J. Biol. Chem. 275, 26877–26884.
Spinsanti, G.P.C., Lazzeri, E., Marsili, L., Casini, S., Frati, F., Fossi, C.M., 2006. Selectionof reference genes for quantitative RT-PCR studies in striped dolphin (Stenellacoeruleoalba) skin biopsies. BMC Mol. Biol. 7, 32.
Teoh, K.H., Polichuk, D.R., Reed, D.W., Nowak, G., Covello, P.S., 2006. Artemisia annuaL. (Asteraceae) trichome-specific cDNAs reveal CYP71AV1, a cytochrome P450with a key role in the biosynthesis of the antimalarial sesquiterpene lactoneartemisinin. FEBS Lett. 580, 1411–1416.
Trusheva, B., Todorov, I., Ninova, M., Najdenski, H., Daneshmand, A., Bankova, V.,2010. Antibacterial mono- and sesquiterpene esters of benzoic acids fromIranian propolis. Chem. Cent. J. 4, 8.
van Klink, J., Becker, H., Andersson, S., Boland, W., 2003. Biosynthesis ofanthecotuloide, an irregular sesquiterpene lactone from Anthemis cotula L.(Asteraceae) via a non-farnesyl diphosphate route. Org. Biomol. Chem. 1, 1503–1508.
Vogler, B.K., Pittler, M.H., Ernst, E., 1998. Feverfew as a preventive treatment formigraine: a systematic review. Cephalalgia 18, 704–708.
Wagner, G.J., 1991. Secreting glandular trichomes: more than just hairs. PlantPhysiol. 96, 675–679.
Wallaart, T.E., Bouwmeester, H.J., Hille, J., Poppinga, L., Maijers, N.C.A., 2001.Amorpha-4, 11-diene synthase: cloning and functional expression of a key

1750 M. Majdi et al. / Phytochemistry 72 (2011) 1739–1750
enzyme in the biosynthetic pathway of the novel antimalarial drug artemisinin.Planta 212, 460–465.
Werker, E., 2000. Trichome diversity and development. Adv. Bot. Res. Incorporat.Adv. Plant Pathol. 31, 1–35.
Williams, C.A., Hoult, J.R.S., Harborne, J.B., Greenham, J., Eagles, J., 1995. Biologically-active lipophilic flavonol from Tanacetum-parthenium. Phytochemistry 38, 267–270.