loss of abaxial leaf epicuticular wax in medicago truncatula irg1/palm1 mutants results in
TRANSCRIPT

Loss of Abaxial Leaf Epicuticular Wax in Medicago truncatulairg1/palm1 Mutants Results in Reduced Spore Differentiationof Anthracnose and Nonhost Rust Pathogens W
SrinivasaRaoUppalapati,a,1Yasuhiro Ishiga,a,1VanthanaDoraiswamy,aMohamedBedair,aShipraMittal,a Jianghua
Chen,a Jin Nakashima,a Yuhong Tang,a Million Tadege,a Pascal Ratet,b Rujin Chen,a Holger Schultheiss,c and
Kirankumar S. Mysorea,2
a Plant Biology Division, The Samuel Roberts Noble Foundation, Ardmore, Oklahoma 73401b Institut des Sciences du Vegetale, Centre National de la Recherche Scientifique, 91198 Gif sur Yvette, Francec BASF Plant Science Company GmbH, D-67117 Limburgerhof, Germany
To identify genes that confer nonhost resistance to biotrophic fungal pathogens, we did a forward-genetics screen using
Medicago truncatula Tnt1 retrotransposon insertion lines. From this screen, we identified an inhibitor of rust germ tube
differentation1 (irg1) mutant that failed to promote preinfection structure differentiation of two rust pathogens, Phakopsora
pachyrhizi and Puccinia emaculata, and one anthracnose pathogen, Colletotrichum trifolii, on the abaxial leaf surface.
Cytological and chemical analyses revealed that the inhibition of rust preinfection structures in irg1 mutants is due to
complete loss of the abaxial epicuticular wax crystals and reduced surface hydrophobicity. The composition of waxes on
abaxial leaf surface of irg1 mutants had >90% reduction of C30 primary alcohols and a preferential increase of C29 and C31
alkanes compared with the wild type. IRG1 encodes a Cys(2)His(2) zinc finger transcription factor, PALM1, which also
controls dissected leaf morphology in M. truncatula. Transcriptome analysis of irg1/palm1 mutants revealed down-
regulation of eceriferum4, an enzyme implicated in primary alcohol biosynthesis, and MYB96, a major transcription factor
that regulates wax biosynthesis. Our results demonstrate that PALM1 plays a role in regulating epicuticular wax metabolism
and transport and that epicuticular wax influences spore differentiation of host and nonhost fungal pathogens.
INTRODUCTION
Rusts are obligate biotrophic foliar pathogens that have evolved
specialized mechanisms of invasion (Heath, 1977). To initiate
rust infection, fungal urediniospores need to adhere to the leaf
surface and subsequently form germ tubes. Germ tubes typically
respond thigmotropically to host leaf surface features, such as
stomata, forming appressoria over these openings (Hoch et al.,
1987). Penetration pegs form at the appressoria-stomata inter-
face andmature into invasive hyphae that invademesophyll cells
eventually differentiating into specialized feeding structures
called haustoria (Heath, 1977; Hoch et al., 1987). Therefore, it
appears that rust pathogens require specific plant surface topo-
graphical and chemical signals to trigger the formation of
preinfection structures (Heath, 1977; Muller and Riederer,
2005). Traditionally, breeding for rust resistance in various crops
mainly relied on host resistance mediated by gene-for-gene
resistance (Ayliffe et al., 2008). However, in many cases, resis-
tance mediated by R genes has little durability in the field due to
the rapid evolution and emergence of new pathogen strains that
can escape recognition by R genes and R gene–mediated
downstream defenses. Alternatively, nonhost resistance (NHR)
is defined as a form of resistance exhibited by an entire plant
species to a particular microbial pathogen and is the most
common and durable form of resistance (Heath, 2000). There-
fore, identification and incorporation of traits that confer NHR to a
broad range of rust fungi is an attractive and durable alternative
to host resistance breeding. However, we know little about genes
that regulate NHR (Mysore and Ryu, 2004).
Asian soybean rust caused byPhakopsora pachyrhiziSydow is
a major concern for soybean (Glycine max) producers in Brazil
and the US (Goellner et al., 2010). Since most of the soybean
cultivars and other economically important legumes are suscep-
tible to soybean rust, there is an increasingly urgent demand for
identification of durable resistance to soybean rust (van de
Mortel et al., 2007). Four single resistance genes to P. pachyrhizi,
Rpp1-4, have been described (Hyten et al., 2007; Garcia et al.,
2008; Silva et al., 2008; Monteros et al., 2010). Interestingly, all
commercial cultivars that are cultivated in the US are susceptible
to rust, and no soybean varieties have been described as having
broad-spectrum resistance to all isolates of P. pachyrhizi
(Posada-Buitrago and Frederick, 2005). Similarly, rust disease
of switchgrass (Panicum virgatum), caused by Puccinia emacu-
lata, is a concern in Oklahoma and other parts of the US and
could become an important factor once switchgrass is grown in
monoculture over a long period of time (Bouton, 2007).
1 These authors contributed equally to this work.2 Address correspondence to [email protected] author responsible for distribution of materials integral to thefindings presented in this article in accordance with the policy describedin the Instructions for Authors (www.plantcell.org) is: Kirankumar S.Mysore ([email protected]).WOnline version contains Web-only data.www.plantcell.org/cgi/doi/10.1105/tpc.111.093104
The Plant Cell, Vol. 24: 353–370, January 2012, www.plantcell.org ã 2012 American Society of Plant Biologists. All rights reserved.

Therefore, there is an urgent need for identification of novel
genes that could be used for engineering broad-spectrum resis-
tance to soybean rust and/or switchgrass rust isolates.
Identifying novel sources of resistance through large-scale
forward or reverse genetics screens has the potential to improve
crop plant resistance (Heath, 2000; Mysore and Ryu, 2004).
Medicago truncatula is a rapidly emerging model plant species,
especially for legumes. Several genomic tools and resources are
already available for M. truncatula and include an extensive EST
database, genome sequence, gene expression, protein and
metabolite profiling tools, and a collection of insertion and fast
neutron bombardment mutants (Young and Udvardi, 2009). For
large-scale mutagenesis of theM. truncatula genome, a tobacco
(Nicotiana tabacum) retrotransposon, Tnt1, has been introduced
and been shown to efficiently transpose in the M. truncatula
genome during tissue culture, producing insertions that are
stable during seed-to-seed generations (d’Erfurth et al., 2003).
We have now generated ;20,000 Tnt1 insertion lines in M.
truncatulawith an average of 25 insertions per line (Tadege et al.,
2008). We set up a genetic screen to identify M. truncatula
mutants with altered interactions with P. emaculata and/or P.
pachyrhiziwith an aim of identifyingmutants exhibiting enhanced
susceptibility to P. emaculata or P. pachyrhizi. Characterization
of these susceptible mutants would lead to the identification of
target genes for genetic improvement of rust resistance in
switchgrass, soybeans, and other economically important crops,
including wheat (Triticum aestivum) and barley (Hordeum vul-
gare).
The high-throughput forward genetics screen ofM. truncatula
Tnt1 insertion lines performed in this study unexpectedly iden-
tified an inhibitor of rust germ tube differentiation1 (irg1) mutant
that failed to promote preinfection structure differentiation
against nonhost rust fungi, P. emaculata and P. pachyrhizi, and
also to the pathogenic anthracnose fungusColletotrichum trifolii.
Cytological, chemical, and transcriptome analyses revealed that
irg1 is defective in abaxial epicuticular wax deposition and/or
secretion. Flanking sequence tag sequencing of irg1 revealed
that IRG1 encoded a Cys(2)His(2) zinc finger transcription factor
(PALM1) that also controls dissected leaf morphology in M.
truncatula (Chen et al., 2010). Our results unraveled a role for
PALM1 in asymmetric epicuticular wax deposition onM. trunca-
tula leaves, which influences fungal spore differentiation.
RESULTS
Tnt1 Insertional Mutant Screening in
M. truncatula Identified irg1
Initial characterization of the M. truncatula–P. emaculata inter-
action showed that the urediniospores germinate and form germ
tubes on the leaf surface but fail to recognize and form appres-
soria on stomata, thereby precluding successful colonization of
the nonhost plant, M. truncatula (Figure 1A). These interactions
were significantly different from those observed on switchgrass.
On switchgrass leaves, P. emaculata spores adhered, germi-
nated, and formed appressoria possibly after thigmotrophic
signal-mediated oriented growth of the germ tubes to stomata.
The penetrating infection hyphae then formed infection sites that
eventually produced asexual urediniospores. Prehaustorial re-
sistance is a very common form ofNHR to parasitic rust fungi and
is usually mediated by the activation of plant defense responses
(Heath, 1977; Heath, 2000). Interestingly, the M. truncatula NHR
response to P. emaculata was not associated with major tran-
scriptional changes in the phenylpropanoid pathway or other
pathogenesis-related (PR) genes compared with the mock-
inoculated plants (see Supplemental Figure 1 online), suggesting
a passive resistancemechanism. However, it is important to note
that a subtle induction of Chitinase and PR-3 genes was ob-
served at 24 h after inoculation (HAI) when compared with the
wild-type plants (see Supplemental Figure 1 online). To identify
mutants that compromise this particular NHR, we screened 1200
Tnt1 lines or 14,400 independent R0 or R1 plants (12 plants per
each Tnt1 line) for loss of NHR to P. emaculata. Detached leaves
from ;12 plants of each Tnt1 line were challenged with P.
emaculata (see Methods and Supplemental Figure 2 online).
Micro- and macroscopy observations of disease development
were recorded at 8, 24, and 48 HAI and 5 d after inoculation to
identify mutants compromised in NHR.
Strikingly, four independent Tnt1 lines showed inhibition of
germ tube growth and differentiation on the abaxial leaf surface,
instead of the expected phenotype of enhanced susceptibility
(Figures 1A and 1B). We named these mutants irg1 and found
that all of the four identified independent mutants had Tnt1
insertions at different locations in the same exon of one gene.
Therefore, we treated them as null mutant alleles (irg1-1, irg1-2,
irg1-3, and irg1-4). Urediniospores were inoculated on the ab-
axial leaf surfaces (mimics natural infection on host plants), and
the formation of germ tubes and the preinfection structures was
visualized by the green fluorescence emitted by the wheat germ
agglutinin-Alexa Fluor 488 conjugate, a fluorescent lectin that
binds to N-acetyl-glucosamine in the cell walls and thus stains
fungal structures (Figures 1A and 1B). Unlike on the adaxial leaf
surface of the wild-type and irg1-1 mutants where;95% of the
spores adhered, germinated, and formed long germ tubes, on
the abaxial leaf surfaces of irg1-1 mutants, only ;60% of the
spores germinated (Figures 1G and 1H). By contrast, on the
abaxial leaf surface of wild-type plants ;95% of the spores
adhered, germinated, and differentiated into long germ tubes
(Figures 1A and 1H). Intriguingly, the spores that did germinate
on the abaxial leaf surface failed to undergo any further differ-
entiation on irg1 plants and did not showany further growth of the
germ tubes as they did on wild-type plants (Figures 1A, 1B, and
1H). Inhibition of preinfection structure formation on irg1-1 is
most likely due to alterations in surface signal(s) required for the
differentiation of preinfection structures of P. emaculata.
The irg1Mutants Also Showed Inhibited Preinfection
Structure Formation of P. pachyrhizi
To further test if irg1 mutants also inhibit preinfection structure
formation by a broad spectrum of rust pathogens, we inoculated
urediniospores of a direct-penetrating rust fungus, P. pachyrhizi,
on abaxial leaf surfaces of M. truncatula wild-type R108 and
irg1-1 mutants. M. truncatula is an incompatible host to P.
pachyrhizi (Figures 1C to 1F). Although P. pachyrhizi spores
354 The Plant Cell
germinated, formed appressoria, and penetrated the epidermal
cells causing visible necrosis, they failed to sporulate on wild-
typeM. truncatulawhen the inoculated plants weremaintained in
a dew chamber for 24 h with 100% humidity for spore germina-
tion and then incubated in a growth chamberwith low (30 to 40%)
humidity (Figure 1C). Very few necrotic lesions developed on irg1
compared with R108 plants (Figures 1C and 1D). On adaxial and
abaxial leaf surfaces of wild-type R108 plants and adaxial leaf
surface of irg1 mutants, urediniospores adhered, germinated,
and formed germ tubes, and most of the germ tubes underwent
differentiation to form appressoria and directly penetrated the
epidermal cells (Figures 1I to 1K). However, on the abaxial leaf
surface of irg1-1 mutants, the ability of the spores to adhere,
germinate, and form germ tubes with appressoria was severely
compromised, resulting in very low penetration of the epidermal
cells (Figures 1F and 1J). These results further confirmed that the
abaxial leaf surface of irg1 leaves does not promote or inhibit the
formation or differentiation of preinfection structures of at least
two rust pathogens tested.
InhibitionofRustGermTubeDifferentiation in irg1 IsLimited
to Abaxial Leaf Surface
We noticed that the abaxial but not the adaxial leaf surfaces of
irg1-1plantswere glossy in appearancewhen comparedwith the
wild-type R108 plants in which neither surface is glossy (Figures
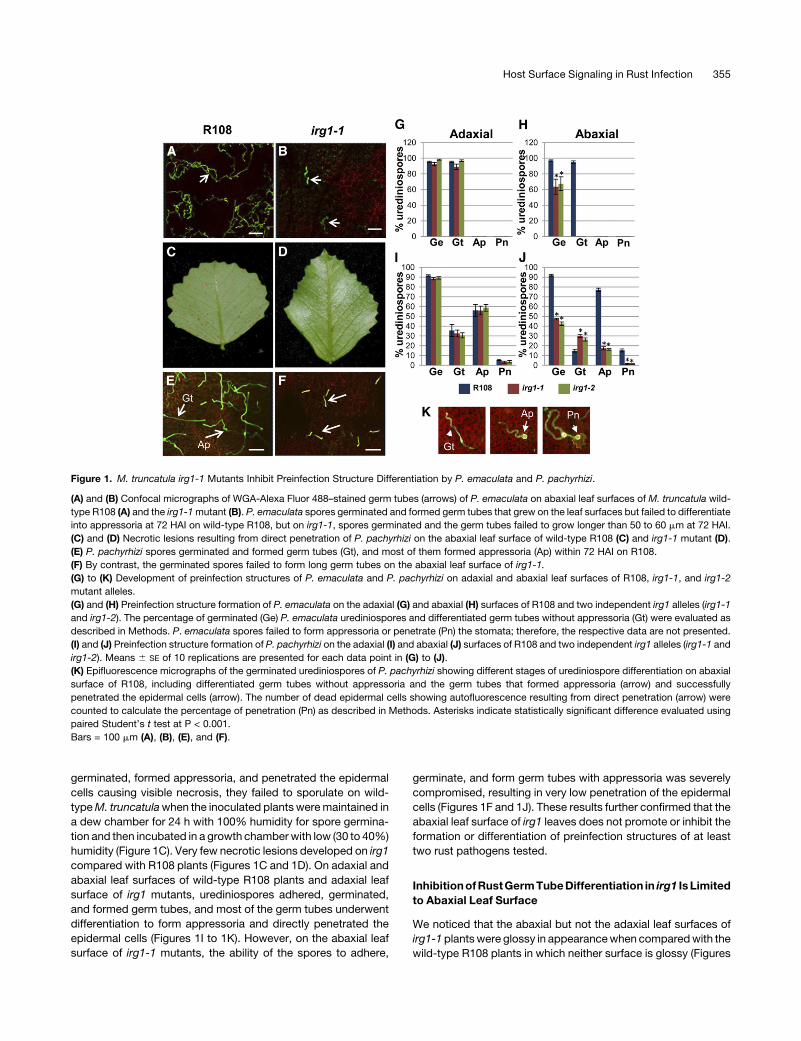
Figure 1. M. truncatula irg1-1 Mutants Inhibit Preinfection Structure Differentiation by P. emaculata and P. pachyrhizi.
(A) and (B) Confocal micrographs of WGA-Alexa Fluor 488–stained germ tubes (arrows) of P. emaculata on abaxial leaf surfaces of M. truncatula wild-
type R108 (A) and the irg1-1mutant (B). P. emaculata spores germinated and formed germ tubes that grew on the leaf surfaces but failed to differentiate
into appressoria at 72 HAI on wild-type R108, but on irg1-1, spores germinated and the germ tubes failed to grow longer than 50 to 60 mm at 72 HAI.
(C) and (D) Necrotic lesions resulting from direct penetration of P. pachyrhizi on the abaxial leaf surface of wild-type R108 (C) and irg1-1 mutant (D).
(E) P. pachyrhizi spores germinated and formed germ tubes (Gt), and most of them formed appressoria (Ap) within 72 HAI on R108.
(F) By contrast, the germinated spores failed to form long germ tubes on the abaxial leaf surface of irg1-1.
(G) to (K) Development of preinfection structures of P. emaculata and P. pachyrhizi on adaxial and abaxial leaf surfaces of R108, irg1-1, and irg1-2
mutant alleles.
(G) and (H) Preinfection structure formation of P. emaculata on the adaxial (G) and abaxial (H) surfaces of R108 and two independent irg1 alleles (irg1-1
and irg1-2). The percentage of germinated (Ge) P. emaculata urediniospores and differentiated germ tubes without appressoria (Gt) were evaluated as
described in Methods. P. emaculata spores failed to form appressoria or penetrate (Pn) the stomata; therefore, the respective data are not presented.
(I) and (J) Preinfection structure formation of P. pachyrhizi on the adaxial (I) and abaxial (J) surfaces of R108 and two independent irg1 alleles (irg1-1 and
irg1-2). Means 6 SE of 10 replications are presented for each data point in (G) to (J).
(K) Epifluorescence micrographs of the germinated urediniospores of P. pachyrhizi showing different stages of urediniospore differentiation on abaxial
surface of R108, including differentiated germ tubes without appressoria and the germ tubes that formed appressoria (arrow) and successfully
penetrated the epidermal cells (arrow). The number of dead epidermal cells showing autofluorescence resulting from direct penetration (arrow) were
counted to calculate the percentage of penetration (Pn) as described in Methods. Asterisks indicate statistically significant difference evaluated using
paired Student’s t test at P < 0.001.
Bars = 100 mm (A), (B), (E), and (F).
Host Surface Signaling in Rust Infection 355
1C and 1D). This suggested possible alterations in epicuticular
wax loading on the abaxial leaf surface of irg1. In plant interac-
tionswith host-specific biotrophic pathogens, includingErysiphe
pisi andBlumeria graminis, the components of abaxial leaf waxes
were shown to specifically promote the differentiation of pre-
infection structures of E. pisi and B. graminis (Gniwotta et al.,
2005; Hansjakob et al., 2010). To test if two different nonhost rust
pathogens with different preinfection processes require surface
cues for adherence or germ tube differentiation, we examined
the preinfection structure formation of P. emaculata and P.
pachyrhizi on the abaxial and adaxial leaf surfaces of the wild-
type and irg1 plants (Figures 1G to 1J). On the adaxial leaf
surfaces of both wild-type and irg1 plants, almost 90% of the
inoculated urediniospores of P. emaculata that germinated
formed germ tubes with no appressoria on stomata (Figure
1G). Similarly, no inhibition of urediniospore germination or germ
tube elongation was observed on the abaxial side of wild-type
plants (Figure 1H). However, only ;60% of the spores germi-
nated on the abaxial side of the irg1 mutants, and almost all the
germinated spores failed to undergo any further differentiation
(Figure 1H). These results further suggested the absence of
stimulatory signals for germ tube differentiation or presence of
inhibitory signals for growth of P. emaculata on the abaxial
surface of irg1 plants.
Unlike P. emaculata, P. pachyrhizi is a direct-penetrating
biotrophic rust fungus with a broad host range and, based on in
vitro assays conducted on artificial membranes, is suggested not
to require hydrophobic or chemical signals for preinfection struc-
ture formation (Koch and Hoppe, 1988; Goellner et al., 2010). Our
in vivo assays conducted on the adaxial leaf surface showed no
significant differences in germination, appressorium formation, or
epidermal penetration by P. pachyrhizi between wild-type R108
and two independent mutant alleles of irg1 (Figures 1I). Interest-
ingly, P. pachyrhizi showed slightly higher percentage of appres-
soria and penetration rate when inoculated on the abaxial leaf
surface compared with adaxial leaf surface of the wild-type R108
(Figure 1J). However, on the abaxial surface of irg1-1 mutants,
;50%of the spores failed to germinate (Figure 1J). Strikingly, only
;20% of the germinated spores formed appressoria on the
abaxial side of irg1-1 and irg1-2mutant alleles, whereas;75%of
the germinated spores formed appressoria on the abaxial side of
R108 (Figure 1J). Consistent with these results of reduced devel-
opment of infection structures, we observed no induction of any
PR and pathogen-inducible genes on the irg1-1 abaxial leaf
surface inoculated with P. pachyrhizi urediniospores, suggesting
that induced or constitutive defenses were not responsible for the
observed phenotype (see Supplemental Figure 3A online). Fur-
thermore, the total leaf proteins isolated from the irg1 mutants
showedno significant inhibitory effects on the germination or germ
tube differentiation of P. emaculata and P. pachyrhizi uredinio-
spores on plastic (hydrophobic) surfaces, suggesting no predom-
inant antimicrobial protein accumulation in these mutants that
could account for the phenotype we observed on the abaxial
leaf surface (see Supplemental Figures 3B and 3C online).
These results suggested that, instead, the abaxial surface of
irg1 leaves may lack the surface chemical/physical cues re-
quired for the differentiation of preinfection structure formation
by P. pachyrhizi.
irg1 Showed Partial Resistance to C. trifolii but Not to
Phomamedicaginis
To further test if irg1 mutants also exhibit resistance to compat-
ible fungal pathogens, we challenged wild-type R108 and irg1-1
mutants with C. trifolii, a hemibiotrophic pathogen that forms
preinfection structures (Figure 2A), and a necrotrophic pathogen,
P. medicaginis, that directly penetrates without forming appres-
soria (Figure 2B). The percentage of C. trifolii spores that germi-
nated and formed preinfection structures (appressoria) was
significantly reduced on irg1-1mutants, as were the consequent
anthracnose symptoms developed on the abaxial leaf surface
when compared with wild-type R108 (Figure 2A). The percent-
ages of germination and appressoria formation were also slightly
reduced (by ;10%) on the adaxial surface of irg1-1 and irg1-2
mutant alleles compared with the wild-type R108 (Figure 2A).
However, no significant differences in symptom development or
in planta fungal growth of P. medicaginis were observed upon
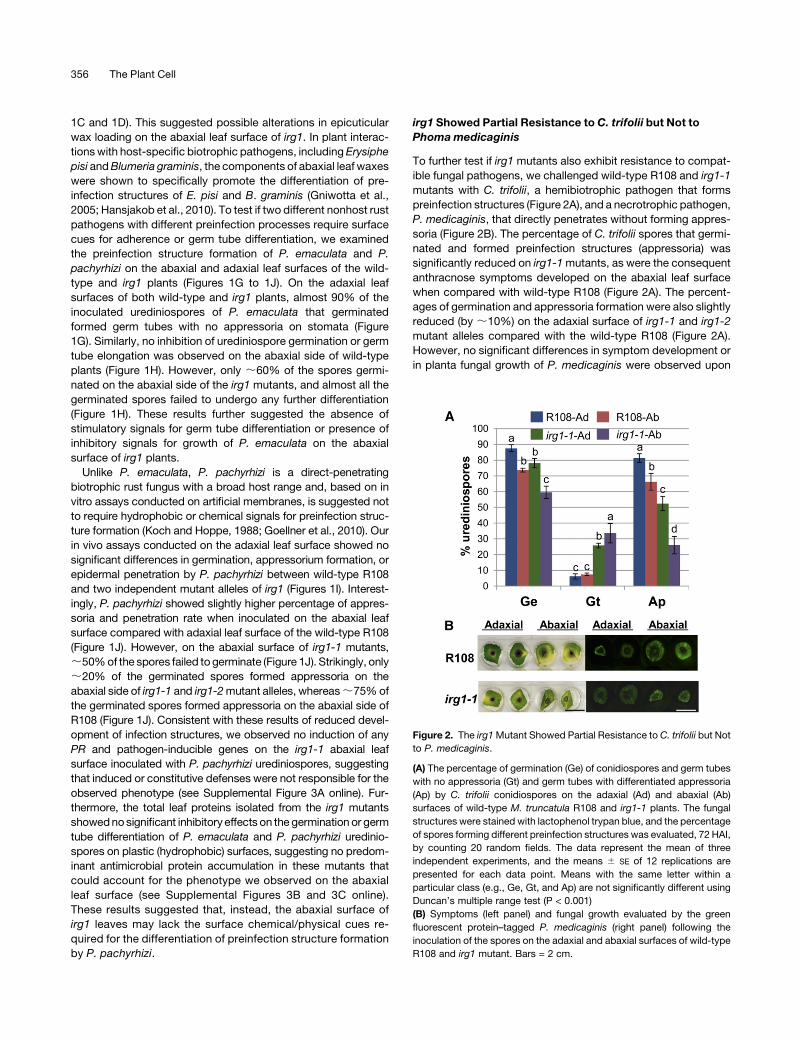
Figure 2. The irg1Mutant Showed Partial Resistance to C. trifolii but Not
to P. medicaginis.
(A) The percentage of germination (Ge) of conidiospores and germ tubes
with no appressoria (Gt) and germ tubes with differentiated appressoria
(Ap) by C. trifolii conidiospores on the adaxial (Ad) and abaxial (Ab)
surfaces of wild-type M. truncatula R108 and irg1-1 plants. The fungal
structures were stained with lactophenol trypan blue, and the percentage
of spores forming different preinfection structures was evaluated, 72 HAI,
by counting 20 random fields. The data represent the mean of three
independent experiments, and the means 6 SE of 12 replications are
presented for each data point. Means with the same letter within a
particular class (e.g., Ge, Gt, and Ap) are not significantly different using
Duncan’s multiple range test (P < 0.001)
(B) Symptoms (left panel) and fungal growth evaluated by the green
fluorescent protein–tagged P. medicaginis (right panel) following the
inoculation of the spores on the adaxial and abaxial surfaces of wild-type
R108 and irg1 mutant. Bars = 2 cm.
356 The Plant Cell
inoculation on either abaxial or adaxial leaf surfaces of R108 and
irg1-1 (Figure 2B). These results suggested that the effect of irg1
mutants may be limited to those fungi that form preinfection
structures (appressoria) in response to surface signals.
IRG1 Encodes a Cys(2)His(2) Zinc Finger Transcription
Factor and Is Allelic to PALM1
We recovered several Tnt1 flanking sequence tags from the irg1
mutant alleles, and the sequence of one of these had sequence
similarity to a gene that encodes a Cys(2)His(2) zinc finger
transcription factor. Chen et al. (2010) recently showed that the
Cys(2)His(2) zinc finger transcription factor PALM1 controls
trifoliate leaf development in M. truncatula. We decided to
investigate whether there was an association between the irg1
phenotype and the PALM1 mutation. We evaluated four inde-
pendent irg1 alleles identified in this study that have an insertion
in PALM1 and found that irg1-1 is palm1-5 and irg1-2 is palm1-4
(see Supplemental Figure 4 online). In addition, all irg1 mutants
as well as palm1 had five leaflets, unlike a typical M. truncatula
leaf, which has three leaflets (see Supplemental Figure 5 online).
To further confirm if IRG1 gene is allelic to PALM1, we
challenged a previously identified palm1 deletion mutant line
M469 of M. truncatula ecotype A17 (palm1-1) and another
independent Tnt1 line (line NF5022, palm1-6) that has an inser-
tion in PALM1 (Chen et al., 2010) with P. pachyrhizi and P.
emaculata. The formation of preinfection structures of P.
pachyrhizi and P. emaculata was severely impaired on the
abaxial leaf surface of all identified alleles of palm1, indicating
that the loss of function of PALM1 is responsible for the irg1
phenotype (see Supplemental Figures 4 online). Since thepalm1-
6 Tnt1 line and the palm1-1M469 deletion line also inhibited rust
germ tube differentiation, we referred to them as irg1-5/palm1-6
and irg1-6/palm1-1, respectively. We further tested if the irg
phenotype results from the loss of function of PALM1 by
complementing the inhibition of germ tube differentiation phe-
notype of irg1-1/palm1-5 (R108 background) and irg1-6/palm1-1
(A17 background) by expressing wild-type PALM1 under the
control of its native promoter in these mutants (Figure 3). The
complemented lines did not show any inhibition of the rust
preinfection structure formation when compared with their re-
spective mutant lines (Figure 3). Taken together, these results
showed that a gene involved in controlling leaf morphology also
contributes to fungal resistance; therefore, going forward, we
renamed our mutants and addressed them as irg1/palm1.
The irg1/palm1Mutant Affects Abaxial Leaf
Epicuticular Wax
Surface chemical and physical signals are known to affect
adhesion, germination, and differentiation of preinfection struc-
tures in pathogenic fungi (Podila et al., 1993; Kolattukudy et al.,
1995; Uppalapati and Fujita, 2000). Therefore, based on our
pathogen assays and our observation that the abaxial leaf
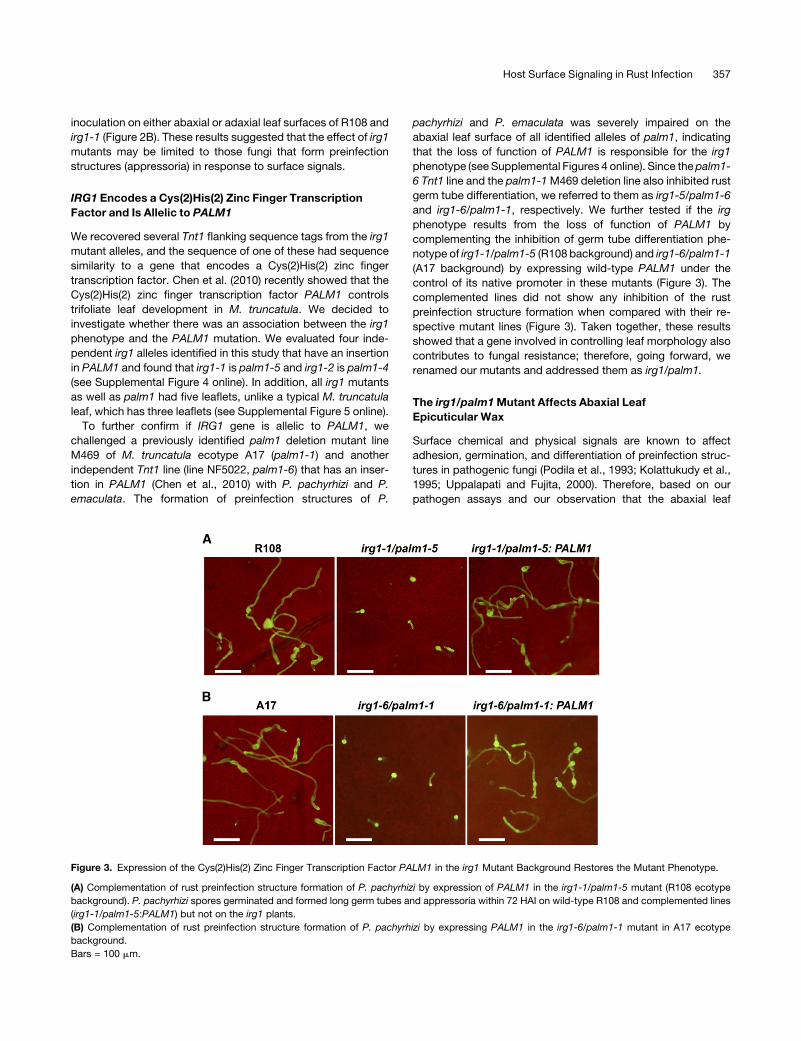
Figure 3. Expression of the Cys(2)His(2) Zinc Finger Transcription Factor PALM1 in the irg1 Mutant Background Restores the Mutant Phenotype.
(A) Complementation of rust preinfection structure formation of P. pachyrhizi by expression of PALM1 in the irg1-1/palm1-5 mutant (R108 ecotype
background). P. pachyrhizi spores germinated and formed long germ tubes and appressoria within 72 HAI on wild-type R108 and complemented lines
(irg1-1/palm1-5:PALM1) but not on the irg1 plants.
(B) Complementation of rust preinfection structure formation of P. pachyrhizi by expressing PALM1 in the irg1-6/palm1-1 mutant in A17 ecotype
background.
Bars = 100 mm.
Host Surface Signaling in Rust Infection 357
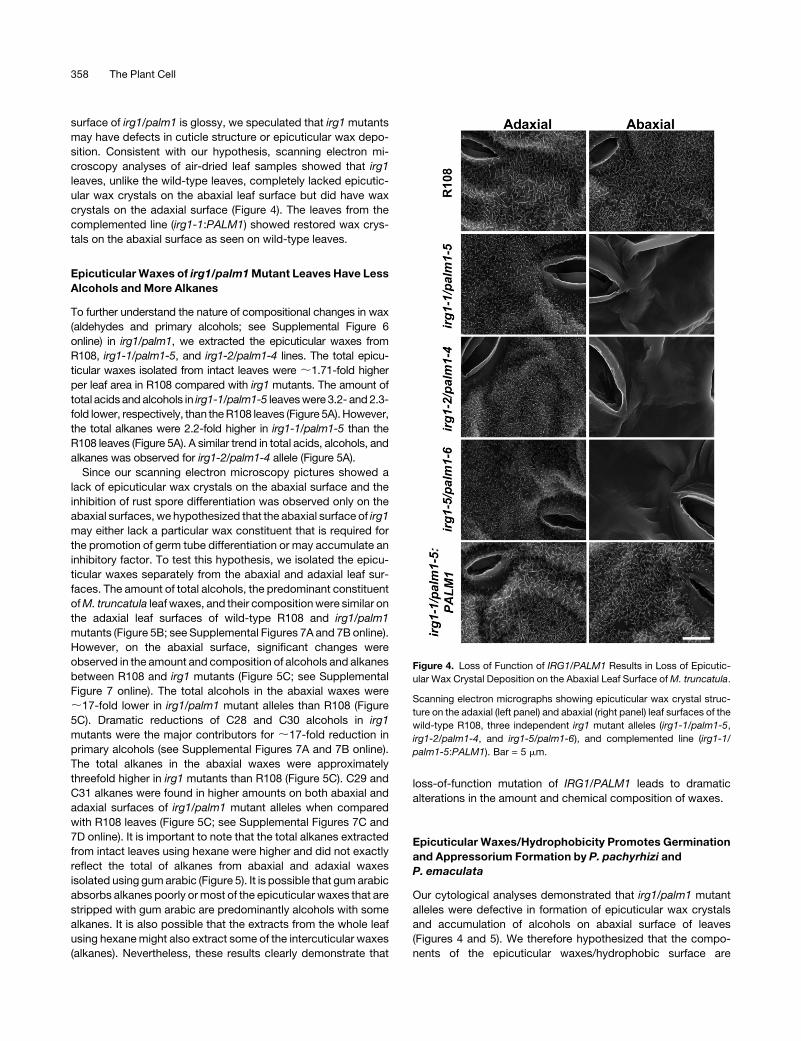
surface of irg1/palm1 is glossy, we speculated that irg1mutants
may have defects in cuticle structure or epicuticular wax depo-
sition. Consistent with our hypothesis, scanning electron mi-
croscopy analyses of air-dried leaf samples showed that irg1
leaves, unlike the wild-type leaves, completely lacked epicutic-
ular wax crystals on the abaxial leaf surface but did have wax
crystals on the adaxial surface (Figure 4). The leaves from the
complemented line (irg1-1:PALM1) showed restored wax crys-
tals on the abaxial surface as seen on wild-type leaves.
Epicuticular Waxes of irg1/palm1Mutant Leaves Have Less
Alcohols and More Alkanes
To further understand the nature of compositional changes in wax
(aldehydes and primary alcohols; see Supplemental Figure 6
online) in irg1/palm1, we extracted the epicuticular waxes from
R108, irg1-1/palm1-5, and irg1-2/palm1-4 lines. The total epicu-
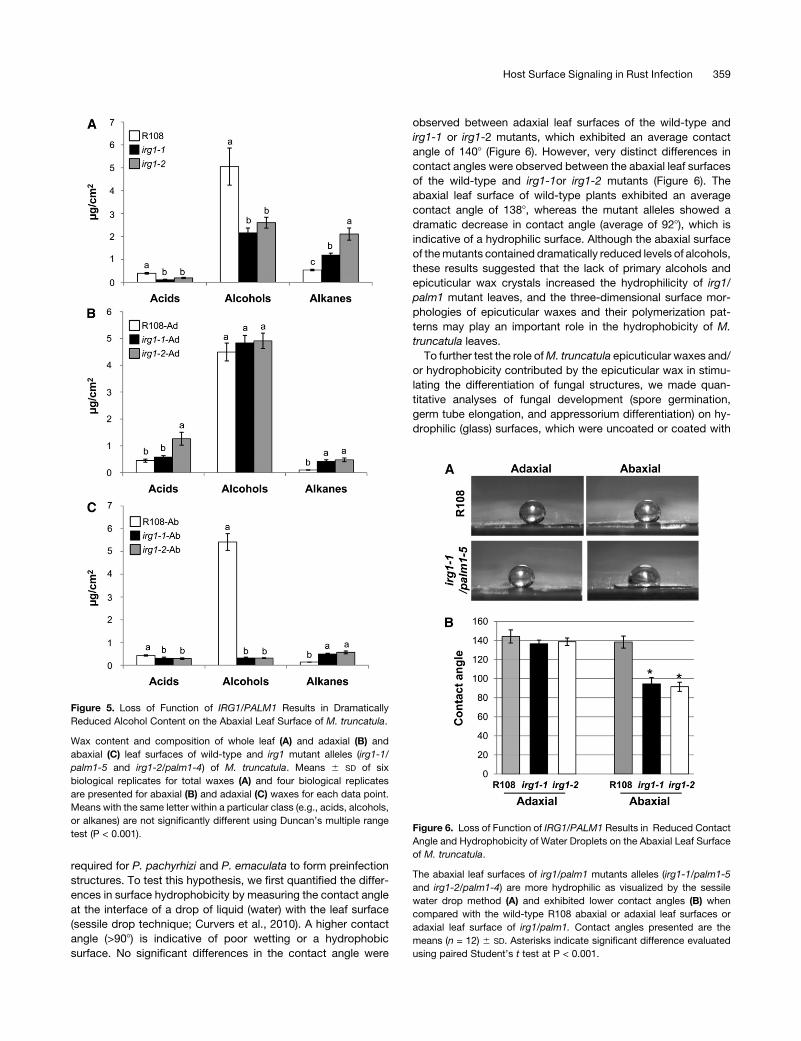
ticular waxes isolated from intact leaves were ;1.71-fold higher
per leaf area in R108 compared with irg1 mutants. The amount of
total acids and alcohols in irg1-1/palm1-5 leaveswere 3.2- and2.3-
fold lower, respectively, than theR108 leaves (Figure 5A).However,
the total alkanes were 2.2-fold higher in irg1-1/palm1-5 than the
R108 leaves (Figure 5A). A similar trend in total acids, alcohols, and
alkanes was observed for irg1-2/palm1-4 allele (Figure 5A).
Since our scanning electron microscopy pictures showed a
lack of epicuticular wax crystals on the abaxial surface and the
inhibition of rust spore differentiation was observed only on the
abaxial surfaces, we hypothesized that the abaxial surface of irg1
may either lack a particular wax constituent that is required for
the promotion of germ tube differentiation or may accumulate an
inhibitory factor. To test this hypothesis, we isolated the epicu-
ticular waxes separately from the abaxial and adaxial leaf sur-
faces. The amount of total alcohols, the predominant constituent
ofM. truncatula leafwaxes, and their compositionwere similar on
the adaxial leaf surfaces of wild-type R108 and irg1/palm1
mutants (Figure 5B; see Supplemental Figures 7A and 7B online).
However, on the abaxial surface, significant changes were
observed in the amount and composition of alcohols and alkanes
between R108 and irg1 mutants (Figure 5C; see Supplemental
Figure 7 online). The total alcohols in the abaxial waxes were
;17-fold lower in irg1/palm1 mutant alleles than R108 (Figure
5C). Dramatic reductions of C28 and C30 alcohols in irg1
mutants were the major contributors for ;17-fold reduction in
primary alcohols (see Supplemental Figures 7A and 7B online).
The total alkanes in the abaxial waxes were approximately
threefold higher in irg1 mutants than R108 (Figure 5C). C29 and
C31 alkanes were found in higher amounts on both abaxial and
adaxial surfaces of irg1/palm1 mutant alleles when compared
with R108 leaves (Figure 5C; see Supplemental Figures 7C and
7D online). It is important to note that the total alkanes extracted
from intact leaves using hexane were higher and did not exactly
reflect the total of alkanes from abaxial and adaxial waxes
isolated using gumarabic (Figure 5). It is possible that gumarabic
absorbs alkanes poorly ormost of the epicuticular waxes that are
stripped with gum arabic are predominantly alcohols with some
alkanes. It is also possible that the extracts from the whole leaf
using hexanemight also extract some of the intercuticular waxes
(alkanes). Nevertheless, these results clearly demonstrate that
loss-of-function mutation of IRG1/PALM1 leads to dramatic
alterations in the amount and chemical composition of waxes.
Epicuticular Waxes/Hydrophobicity Promotes Germination
and Appressorium Formation by P. pachyrhizi and
P. emaculata
Our cytological analyses demonstrated that irg1/palm1 mutant
alleles were defective in formation of epicuticular wax crystals
and accumulation of alcohols on abaxial surface of leaves
(Figures 4 and 5). We therefore hypothesized that the compo-
nents of the epicuticular waxes/hydrophobic surface are
Figure 4. Loss of Function of IRG1/PALM1 Results in Loss of Epicutic-
ular Wax Crystal Deposition on the Abaxial Leaf Surface ofM. truncatula.
Scanning electron micrographs showing epicuticular wax crystal struc-
ture on the adaxial (left panel) and abaxial (right panel) leaf surfaces of the
wild-type R108, three independent irg1 mutant alleles (irg1-1/palm1-5,
irg1-2/palm1-4, and irg1-5/palm1-6), and complemented line (irg1-1/
palm1-5:PALM1). Bar = 5 mm.
358 The Plant Cell
required for P. pachyrhizi and P. emaculata to form preinfection
structures. To test this hypothesis, we first quantified the differ-
ences in surface hydrophobicity by measuring the contact angle
at the interface of a drop of liquid (water) with the leaf surface
(sessile drop technique; Curvers et al., 2010). A higher contact
angle (>908) is indicative of poor wetting or a hydrophobic
surface. No significant differences in the contact angle were
observed between adaxial leaf surfaces of the wild-type and
irg1-1 or irg1-2 mutants, which exhibited an average contact
angle of 1408 (Figure 6). However, very distinct differences in
contact angles were observed between the abaxial leaf surfaces
of the wild-type and irg1-1or irg1-2 mutants (Figure 6). The
abaxial leaf surface of wild-type plants exhibited an average
contact angle of 1388, whereas the mutant alleles showed a
dramatic decrease in contact angle (average of 928), which is
indicative of a hydrophilic surface. Although the abaxial surface
of themutants contained dramatically reduced levels of alcohols,
these results suggested that the lack of primary alcohols and
epicuticular wax crystals increased the hydrophilicity of irg1/
palm1 mutant leaves, and the three-dimensional surface mor-
phologies of epicuticular waxes and their polymerization pat-
terns may play an important role in the hydrophobicity of M.
truncatula leaves.
To further test the role ofM. truncatula epicuticular waxes and/
or hydrophobicity contributed by the epicuticular wax in stimu-
lating the differentiation of fungal structures, we made quan-
titative analyses of fungal development (spore germination,
germ tube elongation, and appressorium differentiation) on hy-
drophilic (glass) surfaces, which were uncoated or coated with
Figure 5. Loss of Function of IRG1/PALM1 Results in Dramatically
Reduced Alcohol Content on the Abaxial Leaf Surface of M. truncatula.
Wax content and composition of whole leaf (A) and adaxial (B) and
abaxial (C) leaf surfaces of wild-type and irg1 mutant alleles (irg1-1/
palm1-5 and irg1-2/palm1-4) of M. truncatula. Means 6 SD of six
biological replicates for total waxes (A) and four biological replicates
are presented for abaxial (B) and adaxial (C) waxes for each data point.
Means with the same letter within a particular class (e.g., acids, alcohols,
or alkanes) are not significantly different using Duncan’s multiple range
test (P < 0.001).Figure 6. Loss of Function of IRG1/PALM1 Results in Reduced Contact
Angle and Hydrophobicity of Water Droplets on the Abaxial Leaf Surface
of M. truncatula.
The abaxial leaf surfaces of irg1/palm1 mutants alleles (irg1-1/palm1-5
and irg1-2/palm1-4) are more hydrophilic as visualized by the sessile
water drop method (A) and exhibited lower contact angles (B) when
compared with the wild-type R108 abaxial or adaxial leaf surfaces or
adaxial leaf surface of irg1/palm1. Contact angles presented are the
means (n = 12) 6 SD. Asterisks indicate significant difference evaluated
using paired Student’s t test at P < 0.001.
Host Surface Signaling in Rust Infection 359
epicuticular waxes isolated from soybean, switchgrass, wild-
type M. truncatula, and irg1 mutants. We also studied the effect
of C30 alcohol, the predominant alcohol that was absent on the
abaxial leaf surfaces of irg1/palm1mutants, by quantifying fungal
development on slides coated with C30 alcohol. In addition, we
also studied fungal development on host leaf surfaces where the
waxes were manually removed.
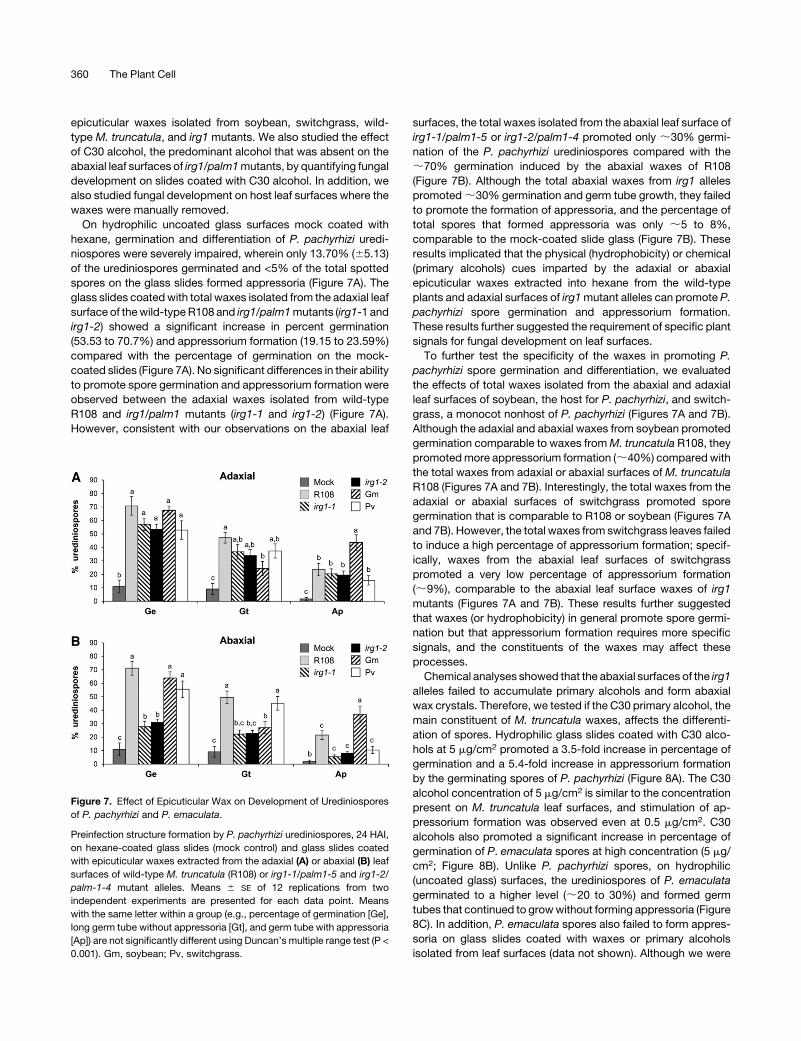
On hydrophilic uncoated glass surfaces mock coated with
hexane, germination and differentiation of P. pachyrhizi uredi-
niospores were severely impaired, wherein only 13.70% (65.13)
of the urediniospores germinated and <5% of the total spotted
spores on the glass slides formed appressoria (Figure 7A). The
glass slides coatedwith total waxes isolated from the adaxial leaf
surface of thewild-typeR108 and irg1/palm1mutants (irg1-1 and
irg1-2) showed a significant increase in percent germination
(53.53 to 70.7%) and appressorium formation (19.15 to 23.59%)
compared with the percentage of germination on the mock-
coated slides (Figure 7A). No significant differences in their ability
to promote spore germination and appressorium formation were
observed between the adaxial waxes isolated from wild-type
R108 and irg1/palm1 mutants (irg1-1 and irg1-2) (Figure 7A).
However, consistent with our observations on the abaxial leaf
surfaces, the total waxes isolated from the abaxial leaf surface of
irg1-1/palm1-5 or irg1-2/palm1-4 promoted only ;30% germi-
nation of the P. pachyrhizi urediniospores compared with the
;70% germination induced by the abaxial waxes of R108
(Figure 7B). Although the total abaxial waxes from irg1 alleles
promoted;30% germination and germ tube growth, they failed
to promote the formation of appressoria, and the percentage of
total spores that formed appressoria was only ;5 to 8%,
comparable to the mock-coated slide glass (Figure 7B). These
results implicated that the physical (hydrophobicity) or chemical
(primary alcohols) cues imparted by the adaxial or abaxial
epicuticular waxes extracted into hexane from the wild-type
plants and adaxial surfaces of irg1mutant alleles can promote P.
pachyrhizi spore germination and appressorium formation.
These results further suggested the requirement of specific plant
signals for fungal development on leaf surfaces.
To further test the specificity of the waxes in promoting P.
pachyrhizi spore germination and differentiation, we evaluated
the effects of total waxes isolated from the abaxial and adaxial
leaf surfaces of soybean, the host for P. pachyrhizi, and switch-
grass, a monocot nonhost of P. pachyrhizi (Figures 7A and 7B).
Although the adaxial and abaxial waxes from soybean promoted
germination comparable to waxes fromM. truncatula R108, they
promotedmore appressorium formation (;40%) comparedwith
the total waxes from adaxial or abaxial surfaces ofM. truncatula
R108 (Figures 7A and 7B). Interestingly, the total waxes from the
adaxial or abaxial surfaces of switchgrass promoted spore
germination that is comparable to R108 or soybean (Figures 7A
and 7B). However, the total waxes from switchgrass leaves failed
to induce a high percentage of appressorium formation; specif-
ically, waxes from the abaxial leaf surfaces of switchgrass
promoted a very low percentage of appressorium formation
(;9%), comparable to the abaxial leaf surface waxes of irg1
mutants (Figures 7A and 7B). These results further suggested
that waxes (or hydrophobicity) in general promote spore germi-
nation but that appressorium formation requires more specific
signals, and the constituents of the waxes may affect these
processes.
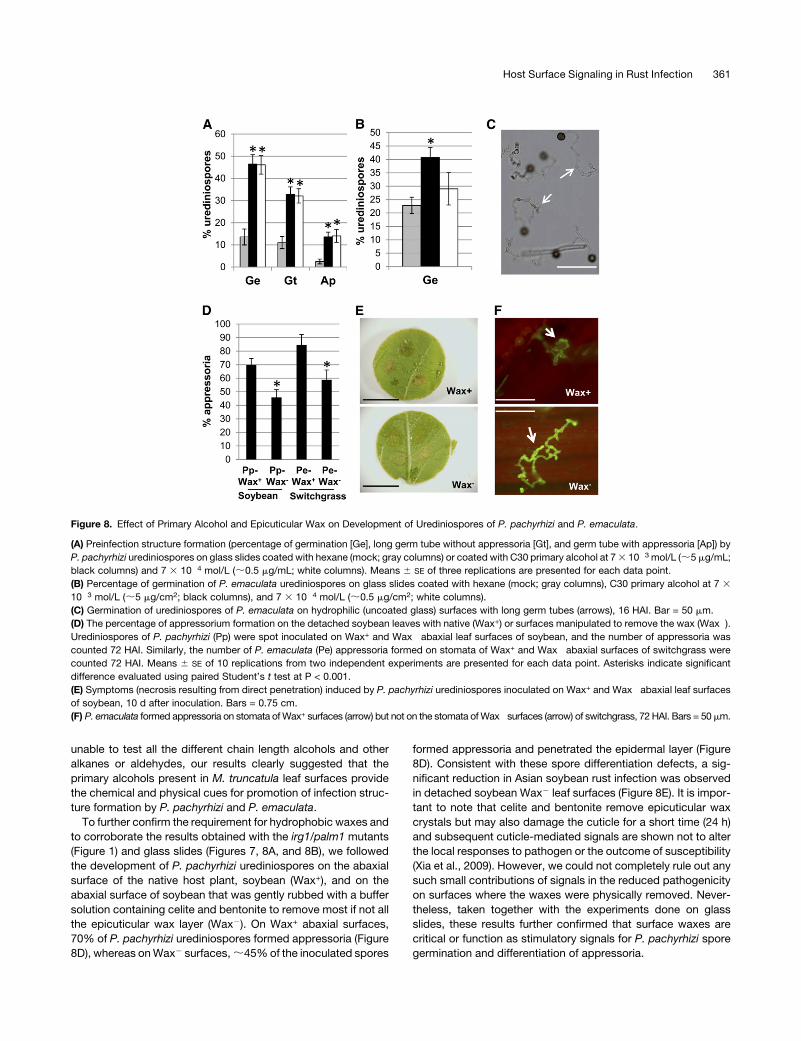
Chemical analyses showed that the abaxial surfaces of the irg1
alleles failed to accumulate primary alcohols and form abaxial
wax crystals. Therefore, we tested if the C30 primary alcohol, the
main constituent of M. truncatula waxes, affects the differenti-
ation of spores. Hydrophilic glass slides coated with C30 alco-
hols at 5 mg/cm2 promoted a 3.5-fold increase in percentage of
germination and a 5.4-fold increase in appressorium formation
by the germinating spores of P. pachyrhizi (Figure 8A). The C30
alcohol concentration of 5 mg/cm2 is similar to the concentration
present on M. truncatula leaf surfaces, and stimulation of ap-
pressorium formation was observed even at 0.5 mg/cm2. C30
alcohols also promoted a significant increase in percentage of
germination of P. emaculata spores at high concentration (5 mg/
cm2; Figure 8B). Unlike P. pachyrhizi spores, on hydrophilic
(uncoated glass) surfaces, the urediniospores of P. emaculata
germinated to a higher level (;20 to 30%) and formed germ
tubes that continued to growwithout forming appressoria (Figure
8C). In addition, P. emaculata spores also failed to form appres-
soria on glass slides coated with waxes or primary alcohols
isolated from leaf surfaces (data not shown). Although we were
Figure 7. Effect of Epicuticular Wax on Development of Urediniospores
of P. pachyrhizi and P. emaculata.
Preinfection structure formation by P. pachyrhizi urediniospores, 24 HAI,
on hexane-coated glass slides (mock control) and glass slides coated
with epicuticular waxes extracted from the adaxial (A) or abaxial (B) leaf
surfaces of wild-type M. truncatula (R108) or irg1-1/palm1-5 and irg1-2/
palm-1-4 mutant alleles. Means 6 SE of 12 replications from two
independent experiments are presented for each data point. Means
with the same letter within a group (e.g., percentage of germination [Ge],
long germ tube without appressoria [Gt], and germ tube with appressoria
[Ap]) are not significantly different using Duncan’s multiple range test (P <
0.001). Gm, soybean; Pv, switchgrass.
360 The Plant Cell
unable to test all the different chain length alcohols and other
alkanes or aldehydes, our results clearly suggested that the
primary alcohols present in M. truncatula leaf surfaces provide
the chemical and physical cues for promotion of infection struc-
ture formation by P. pachyrhizi and P. emaculata.
To further confirm the requirement for hydrophobic waxes and
to corroborate the results obtained with the irg1/palm1 mutants
(Figure 1) and glass slides (Figures 7, 8A, and 8B), we followed
the development of P. pachyrhizi urediniospores on the abaxial
surface of the native host plant, soybean (Wax+), and on the
abaxial surface of soybean that was gently rubbed with a buffer
solution containing celite and bentonite to remove most if not all
the epicuticular wax layer (Wax2). On Wax+ abaxial surfaces,
70% of P. pachyrhizi urediniospores formed appressoria (Figure
8D), whereas onWax2 surfaces,;45%of the inoculated spores
formed appressoria and penetrated the epidermal layer (Figure
8D). Consistent with these spore differentiation defects, a sig-
nificant reduction in Asian soybean rust infection was observed
in detached soybean Wax2 leaf surfaces (Figure 8E). It is impor-
tant to note that celite and bentonite remove epicuticular wax
crystals but may also damage the cuticle for a short time (24 h)
and subsequent cuticle-mediated signals are shown not to alter
the local responses to pathogen or the outcome of susceptibility
(Xia et al., 2009). However, we could not completely rule out any
such small contributions of signals in the reduced pathogenicity
on surfaces where the waxes were physically removed. Never-
theless, taken together with the experiments done on glass
slides, these results further confirmed that surface waxes are
critical or function as stimulatory signals for P. pachyrhizi spore
germination and differentiation of appressoria.
Figure 8. Effect of Primary Alcohol and Epicuticular Wax on Development of Urediniospores of P. pachyrhizi and P. emaculata.
(A) Preinfection structure formation (percentage of germination [Ge], long germ tube without appressoria [Gt], and germ tube with appressoria [Ap]) by
P. pachyrhizi urediniospores on glass slides coated with hexane (mock; gray columns) or coated with C30 primary alcohol at 73 10�3 mol/L (;5 mg/mL;
black columns) and 7 3 10�4 mol/L (;0.5 mg/mL; white columns). Means 6 SE of three replications are presented for each data point.
(B) Percentage of germination of P. emaculata urediniospores on glass slides coated with hexane (mock; gray columns), C30 primary alcohol at 7 3
10�3 mol/L (;5 mg/cm2; black columns), and 7 3 10�4 mol/L (;0.5 mg/cm2; white columns).
(C) Germination of urediniospores of P. emaculata on hydrophilic (uncoated glass) surfaces with long germ tubes (arrows), 16 HAI. Bar = 50 mm.
(D) The percentage of appressorium formation on the detached soybean leaves with native (Wax+) or surfaces manipulated to remove the wax (Wax�).Urediniospores of P. pachyrhizi (Pp) were spot inoculated on Wax+ and Wax� abaxial leaf surfaces of soybean, and the number of appressoria was
counted 72 HAI. Similarly, the number of P. emaculata (Pe) appressoria formed on stomata of Wax+ and Wax� abaxial surfaces of switchgrass were
counted 72 HAI. Means 6 SE of 10 replications from two independent experiments are presented for each data point. Asterisks indicate significant
difference evaluated using paired Student’s t test at P < 0.001.
(E) Symptoms (necrosis resulting from direct penetration) induced by P. pachyrhizi urediniospores inoculated on Wax+ and Wax� abaxial leaf surfaces
of soybean, 10 d after inoculation. Bars = 0.75 cm.
(F) P. emaculata formed appressoria on stomata ofWax+ surfaces (arrow) but not on the stomata ofWax� surfaces (arrow) of switchgrass, 72 HAI. Bars = 50mm.
Host Surface Signaling in Rust Infection 361
We further tested the requirement for surface waxes (hydro-
phobicity) for prepenetration development (i.e., germination,
germ tube elongation, and appressorium differentiation) of P.
emaculata urediniospores on Wax+ and Wax2 abaxial surfaces
of the host plant switchgrass. A 35 to 40% reduction in appres-
soria (appressoria formation on stomata) was observed on
Wax2 abaxial surfaces (Figure 8F). On Wax+ switchgrass, the
germinated spores formed appressoria over the stomatal open-
ings (Figure 8F, top panel). On the Wax2 surfaces, although the
germinated spores oriented to recognize the stomata, a signif-
icant number of them failed to form appressoria on the stomata
(Figure 8F, bottom panel). These results along with data about
germination on slides coated with primary alcohols suggested
that P. emaculata spores require waxy surface signals for ap-
pressorium formation and for enhanced germination but not for
initial germ tube growth.
Transcript Profiling Identifies a Role for IRG1/PALM1 in
Regulating Expression of Genes Involved in Long-Chain
Fatty Acid Biosynthesis and Transport
One of the findings of our study is that the loss-of-function
mutation of a transcription factor involved in leaf morphogenesis
impacts epicuticular wax loading in M. truncatula, which in turn
affects germination and differentiation of fungal spores (Figures 4
and 5). To understand this phenomenon at a molecular level, we
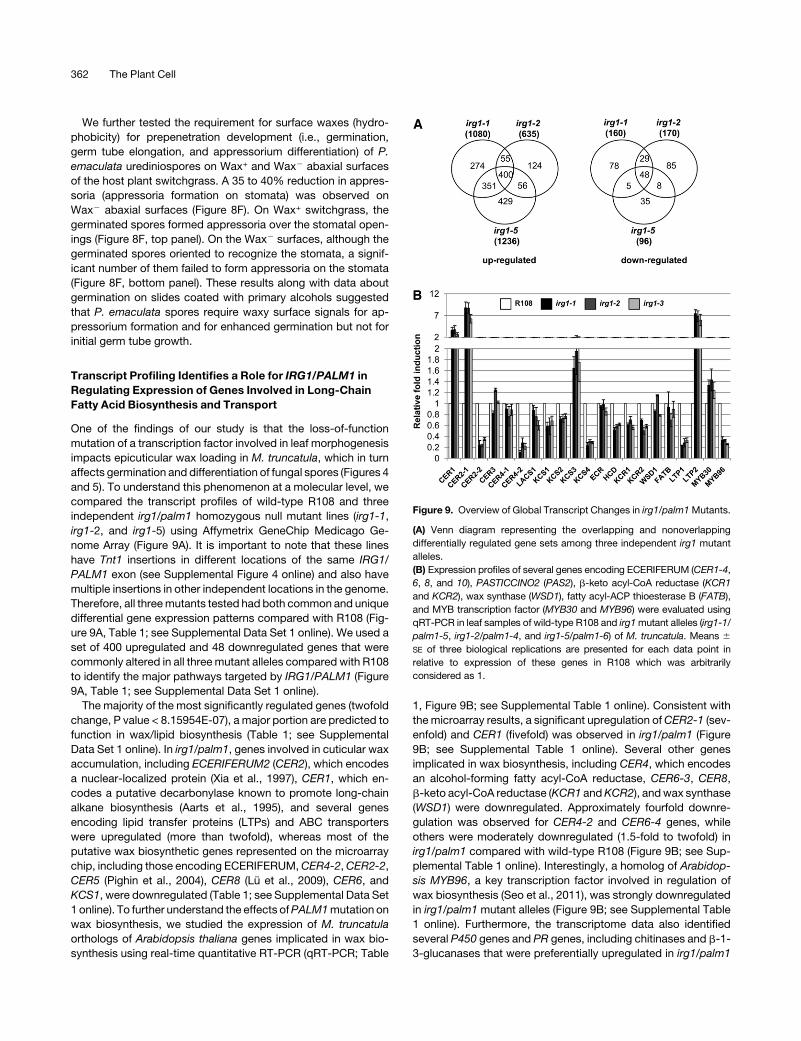
compared the transcript profiles of wild-type R108 and three
independent irg1/palm1 homozygous null mutant lines (irg1-1,
irg1-2, and irg1-5) using Affymetrix GeneChip Medicago Ge-
nome Array (Figure 9A). It is important to note that these lines
have Tnt1 insertions in different locations of the same IRG1/
PALM1 exon (see Supplemental Figure 4 online) and also have
multiple insertions in other independent locations in the genome.
Therefore, all threemutants tested had both common and unique
differential gene expression patterns compared with R108 (Fig-
ure 9A, Table 1; see Supplemental Data Set 1 online). We used a
set of 400 upregulated and 48 downregulated genes that were
commonly altered in all threemutant alleles compared with R108
to identify the major pathways targeted by IRG1/PALM1 (Figure
9A, Table 1; see Supplemental Data Set 1 online).
The majority of the most significantly regulated genes (twofold
change, P value < 8.15954E-07), a major portion are predicted to
function in wax/lipid biosynthesis (Table 1; see Supplemental
Data Set 1 online). In irg1/palm1, genes involved in cuticular wax
accumulation, including ECERIFERUM2 (CER2), which encodes
a nuclear-localized protein (Xia et al., 1997), CER1, which en-
codes a putative decarbonylase known to promote long-chain
alkane biosynthesis (Aarts et al., 1995), and several genes
encoding lipid transfer proteins (LTPs) and ABC transporters
were upregulated (more than twofold), whereas most of the
putative wax biosynthetic genes represented on the microarray
chip, including those encoding ECERIFERUM,CER4-2,CER2-2,
CER5 (Pighin et al., 2004), CER8 (Lu et al., 2009), CER6, and
KCS1, were downregulated (Table 1; see Supplemental Data Set
1 online). To further understand the effects ofPALM1mutation on
wax biosynthesis, we studied the expression of M. truncatula
orthologs of Arabidopsis thaliana genes implicated in wax bio-
synthesis using real-time quantitative RT-PCR (qRT-PCR; Table
1, Figure 9B; see Supplemental Table 1 online). Consistent with
themicroarray results, a significant upregulation ofCER2-1 (sev-
enfold) and CER1 (fivefold) was observed in irg1/palm1 (Figure
9B; see Supplemental Table 1 online). Several other genes
implicated in wax biosynthesis, including CER4, which encodes
an alcohol-forming fatty acyl-CoA reductase, CER6-3, CER8,
b-keto acyl-CoA reductase (KCR1 andKCR2), andwax synthase
(WSD1) were downregulated. Approximately fourfold downre-
gulation was observed for CER4-2 and CER6-4 genes, while
others were moderately downregulated (1.5-fold to twofold) in
irg1/palm1 compared with wild-type R108 (Figure 9B; see Sup-
plemental Table 1 online). Interestingly, a homolog of Arabidop-
sis MYB96, a key transcription factor involved in regulation of
wax biosynthesis (Seo et al., 2011), was strongly downregulated
in irg1/palm1 mutant alleles (Figure 9B; see Supplemental Table
1 online). Furthermore, the transcriptome data also identified
several P450 genes and PR genes, including chitinases and b-1-
3-glucanases that were preferentially upregulated in irg1/palm1
Figure 9. Overview of Global Transcript Changes in irg1/palm1Mutants.
(A) Venn diagram representing the overlapping and nonoverlapping
differentially regulated gene sets among three independent irg1 mutant
alleles.
(B) Expression profiles of several genes encoding ECERIFERUM (CER1-4,
6, 8, and 10), PASTICCINO2 (PAS2), b-keto acyl-CoA reductase (KCR1
and KCR2), wax synthase (WSD1), fatty acyl-ACP thioesterase B (FATB),
and MYB transcription factor (MYB30 and MYB96) were evaluated using
qRT-PCR in leaf samples of wild-type R108 and irg1mutant alleles (irg1-1/
palm1-5, irg1-2/palm1-4, and irg1-5/palm1-6) of M. truncatula. Means 6
SE of three biological replications are presented for each data point in
relative to expression of these genes in R108 which was arbitrarily
considered as 1.
362 The Plant Cell
mutants (see Supplemental Data Set 1 online). In summary, the
transcript profiles of the three independent irg1/palm1 alleles
showed alterations in expression of several genes involved in
wax biosynthesis and transport. Functional analysis of the trans-
porters (ABC/LTPs) and overexpression of CER4, CER2, and
MYB96 in M. truncatula will help us to better understand the
mechanism of wax biosynthesis and asymmetric epicuticular
wax loading in irg1 mutants.
DISCUSSION
In this study, we identified and characterized an epicuticular wax
mutant (irg1) ofM. truncatula in a gene encoding aCys(2)His(2)zinc
finger transcription factor and showed that a loss-of-function
mutation of IRG1 causes major changes in epicuticular wax con-
tent and composition on abaxial leaf surfaces. Most intriguingly,
these changes in the irg1 mutant impart resistance to certain
biotrophic fungal pathogens by inhibiting the differentiation of
preinfection structures. Our results showed that irg1/palm1 mu-
tants were completely devoid of wax crystals on the abaxial leaf
surface and suggested a possible role for signals at the cuticle
interface, especially the components of the leaf wax in promotion
or inhibition of rust preinfection structure differentiation on the
abaxial leaf surface of the irg1mutants (Figures 1 and 2). Consis-
tent with our results, two other previous studies have shown
normal development of fungal preinfection structures on the
adaxial surface and short germ tubes with few appressoria on
abaxial leaf surfaces during host-specific pathogenic interactions
involving wild-type pea (Pisum sativum)–E. pisi and wild-type
ryegrass (Lolium spp)–Erysiphe graminis interactions (Carver et al.,
1990; Gniwotta et al., 2005). Variations in wax crystal composition
between adaxial and abaxial leaf surfaceswere implicated in pea–
E. pisi interactions (Gniwotta et al., 2005). However, the genes or
mechanism(s) responsible for this variation in wax crystals be-
tween adaxial and abaxial surfaces were not identified. Further-
more, our results also provided evidence for requirement of waxes
at contact surfaces for appressorium differentiation by P. pachyr-
hizi and further strengthened the earlier hypothesis that surface
composition and hydrophobicity play important roles in appres-
sorium formation and penetration by direct penetrating fungi (Lee
and Dean, 1994).
Interestingly, hydrophobicity was not very critical for germina-
tion or germ tube elongation or preinfection structure develop-
ment by P. emaculata urediniospores. Although primary alcohols
enhanced the percentage germination (Figure 8B), P. emaculata
urediniospores germinated quite efficiently (20 to 25%) and
formed long germ tubes on the hydrophilic (glass) surface (Figure
8C) and on host leaf surfaces where epicuticular waxes were
removed (Figures 8F, bottom panel). However, on the abaxial
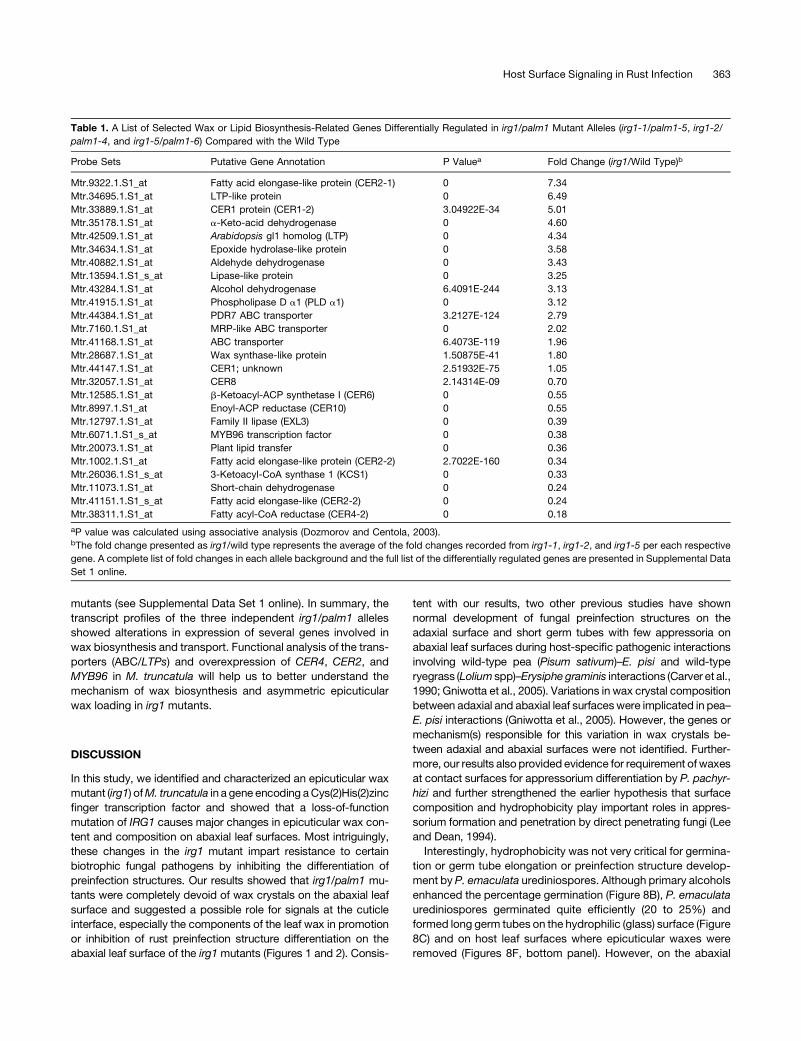
Table 1. A List of Selected Wax or Lipid Biosynthesis-Related Genes Differentially Regulated in irg1/palm1 Mutant Alleles (irg1-1/palm1-5, irg1-2/
palm1-4, and irg1-5/palm1-6) Compared with the Wild Type
Probe Sets Putative Gene Annotation P Valuea Fold Change (irg1/Wild Type)b
Mtr.9322.1.S1_at Fatty acid elongase-like protein (CER2-1) 0 7.34
Mtr.34695.1.S1_at LTP-like protein 0 6.49
Mtr.33889.1.S1_at CER1 protein (CER1-2) 3.04922E-34 5.01
Mtr.35178.1.S1_at a-Keto-acid dehydrogenase 0 4.60
Mtr.42509.1.S1_at Arabidopsis gl1 homolog (LTP) 0 4.34
Mtr.34634.1.S1_at Epoxide hydrolase-like protein 0 3.58
Mtr.40882.1.S1_at Aldehyde dehydrogenase 0 3.43
Mtr.13594.1.S1_s_at Lipase-like protein 0 3.25
Mtr.43284.1.S1_at Alcohol dehydrogenase 6.4091E-244 3.13
Mtr.41915.1.S1_at Phospholipase D a1 (PLD a1) 0 3.12
Mtr.44384.1.S1_at PDR7 ABC transporter 3.2127E-124 2.79
Mtr.7160.1.S1_at MRP-like ABC transporter 0 2.02
Mtr.41168.1.S1_at ABC transporter 6.4073E-119 1.96
Mtr.28687.1.S1_at Wax synthase-like protein 1.50875E-41 1.80
Mtr.44147.1.S1_at CER1; unknown 2.51932E-75 1.05
Mtr.32057.1.S1_at CER8 2.14314E-09 0.70
Mtr.12585.1.S1_at b-Ketoacyl-ACP synthetase I (CER6) 0 0.55
Mtr.8997.1.S1_at Enoyl-ACP reductase (CER10) 0 0.55
Mtr.12797.1.S1_at Family II lipase (EXL3) 0 0.39
Mtr.6071.1.S1_s_at MYB96 transcription factor 0 0.38
Mtr.20073.1.S1_at Plant lipid transfer 0 0.36
Mtr.1002.1.S1_at Fatty acid elongase-like protein (CER2-2) 2.7022E-160 0.34
Mtr.26036.1.S1_s_at 3-Ketoacyl-CoA synthase 1 (KCS1) 0 0.33
Mtr.11073.1.S1_at Short-chain dehydrogenase 0 0.24
Mtr.41151.1.S1_s_at Fatty acid elongase-like (CER2-2) 0 0.24
Mtr.38311.1.S1_at Fatty acyl-CoA reductase (CER4-2) 0 0.18
aP value was calculated using associative analysis (Dozmorov and Centola, 2003).bThe fold change presented as irg1/wild type represents the average of the fold changes recorded from irg1-1, irg1-2, and irg1-5 per each respective
gene. A complete list of fold changes in each allele background and the full list of the differentially regulated genes are presented in Supplemental Data
Set 1 online.
Host Surface Signaling in Rust Infection 363

surface of irg1 mutants, although the spores germinated, they
failed to elongate and undergo any further morphogenesis.
These results suggested a possible role for unknown chemical
signals on the abaxial leaf surface of irg1 in inhibiting P.
emaculata germ tube elongation. Plants with defective cuticle
structure and hydrophobicity (Bessire et al., 2007; Chassot et al.,
2007, 2008; Curvers et al., 2010) have strong resistance to
Botrytis cinerea, possibly through increased release of antimi-
crobial compounds from more permeable epidermal cells. Very-
long-chain aldehydes have been shown to promote preinfection
structure formation of B. graminis (Hansjakob et al., 2010) and
Puccinia graminis f. sp tritici (Reisige et al., 2006). Chemical
analyses of irg1 mutants showed increased accumulation of
alkanes in comparison to wild-type R108 (Figure 5; see Supple-
mental Figure 7 online), indicating a possible role in inhibiting the
growth of P. emaculata germ tubes. It is possible that the effects
of other inhibitory signals may be more conspicuous in the
absence of alcohols that show some stimulatory effects on
overall percentage of germination. We tried to isolate any pos-
sible surface antimicrobials (proteins) from leaves using the
methods described for tomato (Solanum lycopersicum) fruit
surfaces (Yeats et al., 2010) but failed to obtain sufficient protein
concentration to conduct spore germination assays. However,
our results using total leaf proteins isolated from irg1/palm1
mutant did not show any inhibitory activity.
The asymmetric distribution of leaf epicuticular waxes to the
abaxial side in the irg1/palm1 mutants is intriguing. Complete
absence of wax crystals on abaxial and adaxial leaf surfaces of
wild-type Arabidopsis (Jenks et al., 1995) and absence of wax
crystals on abaxial but not adaxial surfaces of wild-type Lolium
perenne (Ringelmann et al., 2009) have been reported. Several
studies have reported complex changes in wax composition in
epicuticular wax mutants, including cer1-cer6, resulting from
increased flux of precursors into other metabolic pathways in
Arabidopsis stems (Aarts et al., 1995; Jenks et al., 1995; Rowland
et al., 2006; Kunst and Samuels, 2009). Our chemical analyses
further demonstrated that loss of function of a Cys(2)His(2) zinc
finger transcription factor (PALM1) also results in changes in the
distribution of different classes of waxes among adaxial and
abaxial surfaces. These complex changes in composition and
distribution of waxes in irg1 can be attributed to the increased
flux of metabolites into the decarbonylation pathway (see Sup-
plemental Figure 6 online).
Our transcriptome analysis revealed that IRG1/PALM1 regu-
lates several target genes involved in lipid metabolism and
transport. Based on the complete absence of wax crystals
phenotype on the abaxial side and 50% reduction in primary
alcohols, the major wax component, we expected a dramatic
downregulation of target genes involved in wax biosynthesis.
However, only homologs of CER4 and CER6 showed significant
downregulation (less than or equal to twofold) in irg1 plants
compared with the wild type (Figure 9B, Table 1). Interestingly,
homologs of Arabidopsis CER2 were upregulated in irg1 plants
(Table 1, Figure 9B). CER6/CUT1 encodes a putative KCS that is
potentially involved in elongation of fatty acyl-CoAs longer than
C22, and deletion of CER6/CUT1 results in 93 to 94% reduction
of total wax loading and almost inactive decarbonylation path-
way in Arabidopsis (Millar et al., 1999; Fiebig et al., 2000). CER4
encodes fatty acyl-CoA reductase, which is responsible for
primary alcohol formation in Arabidopsis (Rowland et al., 2006).
Consistent with our chemical analyses, the transcriptome anal-
ysis provided the genetic evidence that the downregulation of a
CER4 was responsible for the reduced alcohols on the abaxial
surface of irg1/palm1mutant leaves. Furthermore,CER1, amajor
enzyme that promotes long-chain alkane biosynthesis (Aarts
et al., 1995; Bourdenx et al., 2011), and homologs of CER1,
including gl1/cer3/wax2 (Rowland et al., 2007; Mao et al., 2012),
were upregulated in the microarray analyses. Taken together
with the chemical analyses, our results also further suggested
that the homologs of Arabidopsis CER4 and CER1 function to
promote alcohol and alkane biosynthesis, respectively, in M.
truncatula. Based on these findings and our results, it is tempting
to speculate that downregulation of CER4 and concomitant
upregulation ofCER1 result in a reduced accumulation of primary
alcohols and increased flux of precursors into the decarbonylation
pathway, resulting in the accumulation of alkanes in irg1mutants of
M. truncatula (Table 1, Figure 5; see Supplemental Figure 6 online).
Therefore, the irg1/palm1mutant may be helpful in elucidating the
biochemical function of CER1 in C30 alcohol biosynthesis in M.
truncatula and other crop legumes. Primary alcohols appear to be
the predominant form of very-long-chain fatty acids in the epicu-
ticular waxes of fabaceae, including M. truncatula (Zhang et al.,
2005, 2007) and pea (Gniwotta et al., 2005). Furthermore, our
results also showed that the plate-type wax morphologies in M.
truncatulamainly contained primary alcohols andwere required for
three-dimensional structure formation of surface waxes and sur-
face hydrophobicity. Our results also suggested that the physical
(hydrophobicity) and chemical surface cues promote spore
germination and germ tube elongation of several biotrophic
fungi. In addition, the physical and chemical surface cues also
promoted appressorium formation by P. pachyrhizi (Figures 7
and 8). Altered wax composition impacts the initial events of
pathogenesis and spore differentiation during compatible plant–
fungal interactions (Kolattukudy et al., 1995; Gniwotta et al.,
2005; Zabka et al., 2008; Hansjakob et al., 2010, 2011). Due to
the absence of the long-chain aldehydes from the leaf cuticular
wax, the glossy11 mutant of maize (Zea mays) doesn’t support
appressorium formation and subsequent prepenetration by B.
graminis and thus is resistant to this fungus (Hansjakob et al.,
2010, 2011). Future studies involving overexpression of CER1
and Myb96 in wild-type and irg1/palm1 mutants may shed new
light on the role of different classes of very-long-chain fatty acids
and their quantities in formation of epicuticular wax structures
and their contributions to hydrophobicity/fungal differentiation in
M. truncatula. The abaxial leaf surface of irg1/palm1 mutants
may also provide a natural leaf surface with altered wax content
and composition that would allow us to test new hypotheses for
the role of epicuticular waxes in fungal spore germination and
germ tube differentiation.
In addition to CER1, CER2 and two genes encoding LTP-like
proteins were upregulated in irg1 mutants when compared with
wild-type R108 (Table 1). Based on the EST information, we
could not confirm if the LTP genes encoded glycosylphospha-
tidylinositol-anchored LTPs. However, one of the LTPs (Affy ID,
Mtr.13293.1.S1_at) showed high similarity to nonspecific LTPs
(ns-LTPs). Some LTPs have been shown to be secreted and
364 The Plant Cell

accumulate extracellularly where they can play a role in diverse
functions, including cuticular wax transport and defense
against pathogens (Segura et al., 1993; Kader, 1996; Kunst
and Samuels, 2003). Loss of function or reduced expression of
a glycosylphosphatidylinositol-anchored LTP results in reduced
alkane accumulation at the plant surface and plays a role in lipid
export (Debono et al., 2009; Lee et al., 2009). Therefore, it is
tempting to speculate that upregulation of the LTP (Mtr.13293.1.
S1_at) could be one of the reasons for increased alkane accu-
mulation in irg1 mutants.
Unlike the contributions of the constituents and morphologies
of the epicuticular waxes, the role of cuticle and cuticular lipids
has been well studied in plant–pathogen interactions. Studies
done with cuticle mutants, including gpat4/gpat8 (Li et al., 2007),
lacs2 and att1 (Xiao et al., 2004; Tang et al., 2007; Lee et al.,
2009), and gl1 (Xia et al., 2010) have shown that these mutants
are more susceptible to pathogens due to a range of alterations,
including stomatal or substomatal spaces or cuticle-derived
active signaling. By contrast, enhanced resistance of att1 and
lacs2 mutants to B. cinerea was reported either via enhanced
perception of the fungal elicitors because of the permeable
surfaces of these mutants leading to the accumulation of anti-
microbials or enhanced upregulation of defense-related genes
(Bessire et al., 2007). Increased susceptibility to the biotrophic
pathogen Erysiphe cichoracearum and resistance to necrotrophic
fungal pathogens B. cinerea and Alternaria brassicicola were
demonstrated in an Arabidopsis rst1 mutant. Interestingly, rst1
was shown to be a cuticular wax mutant with 59.1% reduction in
waxes (and wax crystal deposition) on stem but 43% increase of
waxes in the leaves (Chen et al., 2005). RST1 was shown to
influence plant defense responses by altering the interactions with
jasmonic acid– and salicylic acid–mediated pathways (Mang et al.,
2009). However in our study, irg1/palm1 showed cuticular wax
defects but did not show any alteration in the expression of genes
involved inSApathway or other phytoalexin-mediatedpathways in
mock- or pathogen-inoculated leaves comparedwith thewild type,
suggesting a predominant role of altered abaxial leaf surface
properties (hydrophobicity or wax constituents) in the promotion of
biotrophic fungal differentiation.
In conclusion, we provide evidence for an increased disease
resistance phenotype of irg1/palm1 plants possibly due to al-
tered abaxial leaf surface signals that inhibit differentiation of
fungal preinfection structure in M. truncatula. Although both
developmental and environmental cues affect wax biosynthesis,
only a very few transcription factors that regulate wax biosyn-
thesis have been isolated (Samuels et al., 2008; Kunst and
Samuels, 2009; Seo et al., 2011). Overexpression of the tran-
scription factor WXP1 causes increased accumulation of acyl-
reduction pathway products in M. sativa leaves (Zhang et al.,
2005). Recently, a homeodomain-Leu zipper IV family transcrip-
tion factor was shown to regulate genes involved in cuticle
biosynthesis (Javelle et al., 2010). Furthermore, overexpression of
wax inducer/SHINE family in Arabidopsis and APETALA2 (AP2)/
ethylene-responsive element binding protein–type transcription
factors, WXP1 and WXP2, in Medicago positively regulate wax
biosynthesis (Aharoni et al., 2004; Broun et al., 2004; Zhang et al.,
2005, 2007). Interestingly, Cys(2)His(2) zinc finger transcription
factor (IRG1/PALM1) contains an ERF-associated amphiphilic
repression domain at the C-terminal region and is conserved in
the class II ERF transcriptional repressors of the AP2/ERF
domain proteins (Chen et al., 2010). How a zinc transcription
factor with an ERF transcriptional repressor domain regulates
wax biosynthesis, and asymmetric distribution of epicuticular
wax is a subject for future research. The irg1/palm1 mutant
identified in this study confers altered leaf morphology and wax
deposition on only one side of the leaf surface. However, it is
important to note that the epicuticular waxes are also shown to
regulate nonstomatal water loss and are important in protecting
plants against water loss (Riederer and Schreiber, 2001). Our
preliminary results suggested that irg1 mutants were not more
susceptible to water loss than the wild type, but the effect of loss
of waxes on the abaxial side of the leaf on drought susceptibility
warrants further systematic study. Further characterization of
irg1/palm1 mutants may help to improve our understanding of
asymmetric epicuticular wax loading on leaf surfaces and may
provide us with approaches to specifically engineer adaxial or
abaxial leaf surface waxes to improve fungal resistance both by
regulating fungal differentiation and by improving the penetration
ability of pesticides in agricultural spray applications.
METHODS
Plant Materials
Seeds ofMedicago truncatula cv Jemalong A17, R108, and Tnt1 insertion
lines ofM. truncatulamutant collection, NF0227 (irg1-1/palm1-5), NF1271
(irg1-2/palm1-4), NF1432 (irg1-3), NF4045 (irg1-4), and NF5022 (irg1-5/
palm1-6) in R108 background, and M469 (irg1-6/palm1-1) in A17 back-
ground from the deletion mutant collection ofM. truncatulawere scarified
for 8 min using concentrated sulfuric acid, washed thrice with distilled
water, and germinated on moist filter papers. Two days after germination
in darkness at 248C, 12 seedlings from each Tnt1 line were transferred to
soil (one seedling per cell in 6 3 12 celled trays). Following 3 weeks
incubation in the greenhouse, the plants were transferred to growth
chambers located in a USDA–Animal and Plant Health Inspection
Service–approved BSL2+ facility to conduct soybean rust or switchgrass
(Panicum virgatum) rust inoculation assays.
Screening ofM. truncatula Tnt1 Insertion Population
To identify Tnt1mutants with altered resistance to switchgrass rust, we set
up a forward genetic screen using a detached leaf assay. An isolate of the
switchgrass rust causative agent Puccinia emaculata collected from
Oklahoma (PE-OK1) wasmaintained on a susceptible lowland switchgrass
(P. virgatum cv Summer). Fresh urediniospores of PE-OK1 were collected
using a gelatin capsule spore collector designed by the Cereal Disease
Laboratory, St. Paul, MN, and suspended in distilled water with 0.001%
Tween 20. The abaxial side of the detached leaves were spray inoculated
with 105 spores/mL (0.001% Tween 20) using an artist airbrush (Paasche
Airbrush) set at 2 p.s.i. with a portable air pump (Gast Manufacturing) for
uniform spore deposition. The inoculated leaves were maintained onmoist
filter papers and incubated overnight in dark and then maintained at 248C
with 16-h-light/8-h-dark cycle. It is important to note that we used an R0 or
R1 segregating Tnt1 population for the forward genetic screen, and one
detached leaf from each of 12 plants representing one Tnt1 line was spray
inoculatedwithP. emaculata spores as described above. Tnt1 population is
shown to containmultiple copies of the Tnt1 tag in each line, andmost of the
phenotypes are suggested to segregate in the R1 progeny (Tadege et al.,
Host Surface Signaling in Rust Infection 365

2008). Therefore, we used a sufficiently large segregating R1 population (12
plants per each line) with a possibility of recovering at least one Tnt1 line with
homozygous insertion in a given gene that also confers altered NHR
response to P. emaculata.
Asian Soybean Rust Maintenance and Inoculation Procedures
An isolate of Asian soybean rust pathogen, Phakopsora pachyrhizi, from
Illinois was maintained on the susceptible soybean cultivar (Glycine max
cv Williams) grown in a growth chamber at 228C/198C with a 12-h-light/
12-h-dark cycle (1000 mmol·m–2·s–1). P. pachyrhizi urediniospores col-
lected and prepared as described above for switchgrass rust were used
to inoculate detached leaves or whole plants grown in 72-cell trays. The
inoculated plants were maintained in a dew chamber for 24 h with 100%
humidity maintained at 198C with a 0-h-light/24-h-dark cycle. The plants
were then transferred to a growth chamber (228C/198C with a 12-h-light/
12-h-dark cycle) and incubated further to allow symptom development.
Light, Confocal, and Scanning Electron Microscopy
Initial interactions of P. pachyrhizi or P. emaculatawithM. truncatulawere
recorded by direct observations of inoculated leaves using an Olympus
stereomicroscope (SZX19) or compound microscopes (BX 41) equipped
with fluorescence attachment. For fluorescence microscopy, fungal
mycelia were stained with wheat germ agglutinin (WGA), coupled to the
green fluorescent dye Alexa Fluor 488 (WGA-Alexa Fluor 488; Invitrogen)
as described previously (Uppalapati et al., 2009). Inoculated leaves were
stained with 10 mg/mLWGA-Alexa Fluor 488 by a brief vacuum infiltration
in PBS followed by a 20-min incubation at room temperature. For
microscopy observations, after washing with PBS, whole leaves or
sections of the leaf were placed on a glass slide and mounted using a
cover glass with Dow Corning high vacuum grease for microscopy.
Fluorescence microscopy to document the infection process was done
using an Olympus epifluorescence microscope (BX 41) or a Leica TCS
SP2 AOBS confocal laser scanning microscope (Leica Microsystems)
equipped with 320 (numerical aperture of 0.70) and 363 (numerical
aperture of 1.2, water immersion) objectives using appropriate laser and
excitation filter settings (WGA-Alexa Fluor 488 to 488 nm). Chloroplast
autofluorescence was captured by exciting with the 647-nm line of the
argon-krypton laser and emission detected at 680 nm. A series of optical
sections (z series) were acquired by scanning multiple sections, and the
z-series projections were done with the software provided with the Leica
TCS SP2 AOBS confocal laser scanning microscope.
Scanning electron microscopy analysis of the air-dried leaves was
performed as described previously (Zhang et al., 2005). Briefly, leaves
from the top two internodes were harvested and air-dried at room
temperature in a Petri dish, since the conventional scanning electron
microscopy sample preparation would wash the surface waxes away.
Air-dried leaves were mounted on stubs and coated with;20 nm of 60/
40 Gold-Palladium particles using a Hummer VI sputtering system
(Anatech). Coated surfaces were viewed using a JEOL JSM-840A scan-
ning electron microscope at 15 kV.
Surface Hydrophobicity Measurements
Leaves were fixed to glass slides with double-sided tape and 10-mL
droplets of distilled water were dropped using a micropipette. The
photographs of the droplets were taken, and the contact angle (uC) was
measured using the angle tool incorporated in the image J version 1.44p
software. A total of 12 independent measurements per abaxial or adaxial
leaf surfaces were performed for wild-type and two mutant alleles. Two
water droplets per leaf surface were evaluated on three plants grown in
independent pots from three different experiments. The average of all the
12 replicates was used for contact angle measurements.
Preinfection Structure Formation and Penetration Assays
Approximately 100 spores of P. emaculata or P. pachyrhizi in 10-mL
aliquots were placed on the adaxial or abaxial surface of the detached
leaves from 4-week-old M. truncatula wild-type R108 or irg1 mutant
plants and incubated in dark overnight and then transferred to a growth
chamber (228C/198C with 12-h-light/12-h-dark cycle). To capture the
early stages of preinfection structure formation, 24 HAI, the inoculated
leaves were washed two to three times in a Petri dish with PBS to remove
the free-floating spores and were stained by floating in a PBS solution
supplementedwith 0.05%Tween 20 and 10mg/mLWGA-Alexa Fluor 488
to visualize the fungal germ tubes and appressoria. Urediniospore-
forming germ tubes >60 mm in length were counted as long germ tubes,
whereas germ tubes#50mm in lengthwere counted as short germ tubes.
The number ofP. pachyrhizi andP. emaculata spores that germinated and
formed germ tubes >10 mm in length were evaluated at 24 HAI and were
counted as the percentage of germination. The subsequent develop-
ments were followed 72 HAI, and the germinated tubes forming differen-
tiated appressoria were counted as appressoria, and the differentiated
germ tubes without appressoria that grew on the surface were also
counted from 20 random fields on three independent leaves. On M.
truncatula, the urediniospores of P. emaculata germinated and formed
long germ tubes but failed to form appressoria (on the stomata) and
penetrate. Therefore, only the percentage of germination and the differ-
entiated germ tubes without appressoria were evaluated for P. emaculata
on M. truncatula. The number of dead autofluorescing epidermal cells
resulting fromdirect penetration ofP. pachyrhiziwas counted 72HAI from
20 random fields per each inoculated site and is used to calculate the
percentage of penetration. It is important to note that the percentage of
germination reflects the total percentage of urediniospores that germi-
nated and is a representation of differentiated germ tubes without
appressoria + differentiated appressoria + percentage of penetration.
Fungal Differentiation on Artificial Surfaces and Leaf Surfaces
Manipulated to Remove the Surface Wax Coating
The abaxial and adaxial leaf surface epicuticular waxes from M. truncatula
wild-type and irg1/palm1 mutants, soybean, or switchgrass were isolated
using a polymer film of gum arabic as described (Gniwotta et al., 2005). The
polymer films peeled from two to three leaflets were pooled and extracted
with hexane, evaporated, and resuspended by sonication in fresh hexane.
The hexane solution of waxes from wild-type R108 or irg1 mutants was
adjusted to a final concentration of 0.05mL/cm2 leaf area.Uncoated, frosted
glass slides (253 75mm; VWR International) were coated (three times) with
a hexane solution to cover the whole surface. Hexane was allowed to
completely evaporate between each application as described (Podila et al.,
1993). The C30 alcohol 1-triacontanol (Sigma-Aldrich) was dissolved in
hexane to a final concentration of 73 1023mol/L (;5mg/cm2) and73 1024
mol/L (;0.5 mg/cm2) and was applied to the glass as described above for
the leaf waxes. The approximate coverage per cm2was estimated based on
the assumptions made by Hansjakob et al. (2010). Uncoated glass slides
were coated with same amount of hexane, the solvent used for wax
preparations, and were referred as mock coated/control. To measure the
effect of surface hydrophobicity or surfaces waxes on appressorium forma-
tion, a spore suspension of P. pachyrhizi was applied on the mock control
and glass slides coated with leaf waxes or C30 alcohol.
For removal of epicuticular waxes, detached leaf surfaces of switch-
grass and soybean were gently rubbed with a cotton swab saturated with
a solution containing bentonite (0.02% [w/v]; Sigma-Aldrich) and celite
(1% [w/v]; Sigma-Aldrich) as described (Xia et al., 2009). The uredinio-
spores of P. emaculata and P. pachyrhizi were inoculated on the respec-
tive host leaf surfaces by spotting 10 mL aliquots of 104 spores/mL in
distillated water with 0.001% Tween 20 on the native and surfaces
manipulated to remove wax. The percentage of preinfection structures
366 The Plant Cell

was evaluated as described above in the preinfection structure formation
and penetration assays section.
Infection Assays with Colletotrichum trifolii
C. trifolii race 1 was maintained on potato dextrose agar media (Becton
Dickson and Company). Conidia from 10- to 14-d-old cultures were
harvested, washed in water, and resuspended in sterile distilled water.
Leaves from 4-week-oldwild-typeR108 and irg1mutantswere harvested
and spot inoculated with 10 mL of suspension or spray inoculated with a
suspension containing 1 3 106 spores/mL in 0.005% Tween 20. The
fungal structures were stained with lactophenol trypan blue, and the
percentage of spores forming different preinfection structures (bottom
panel) was evaluated, 72 HAI, by counting 20 random fields. The data
represent the mean of three independent experiments.
Complementation and Stable Transformation
The irg1-6/palm1-6 complementation lines were previously described
(Chen et al., 2010). In this study, we developed stable transgenic lines of
irg1-1 for complementation studies using pCAMBIA3330 carrying a
genomic fragment, including 2.718-kb 59-flanking sequence, 0.756-kb
open reading frame, and 1.028-kb 39-downstream sequence of PALM1
as described (Chen et al., 2010).
Gas Chromatography–Mass Spectrometry Analysis of Cuticular
Wax Content
Total leaf cuticularwax extractionand analysiswasconducted asdescribed
previously (Zhang et al., 2005). Leaf sampleswere collected fromone leaflet
of the top two expanded trifoliates/pentafoliates excised from the major
stems of well-watered wild-type R108 and irg1 mutant alleles. The two
leaflets from the sameplantwere combinedas one leaf sample to represent
one biological replicate. Each sample was added to 10 mL of gas chroma-
tography–mass spectrometry (GC-MS)–grade hexane (Sigma-Aldrich). Tis-
sues were agitated for 2 min, and the solvent was decanted into new glass
tubes. The same amount of hexane was used to rinse the tissues and tubes
for 10 s and was pooled to the sample tube. Hexane was evaporated to
;1mLunder a nitrogen stream, transferred to 2-mL autosampler vials, and
then evaporated completely. The abaxial and adaxial leaf waxes were
extracted as described (Gniwotta et al., 2005). The polymer films from two
leaflets were pooled to represent one biological replicate, extracted with
hexane, and evaporated to 1 mL as described above for the whole leaves.
Six replicates for the whole plant leaf extracts represent six biological
replicates from plants grown in different pots from the same experiment.
The abaxial or adaxial leaf waxes represent the average of four replicates
biological replicates from plants grown in different pots from the same
experiment. These experiments were repeated three independent times,
and data from a representative experiment are presented.
The dried extracts were derivatized using N-methyl-N-trimethylsilyl tri-
fluoroacetamide (+1% trimethylchlorosilane; Pierce Biotechnology) and
were run on an Agilent 7890A gas chromatograph using a splitless injection
as described (Zhang et al., 2005). Quantification was done using the area of
the ions with mass-to-charge ratios of M-15, 117, 57, and 218 for fatty
alcohols, fatty acids, alkanes, and sterols respectively. The amount of total
wax composition and eachcuticularwaxconstituentwas expressedper unit
of leaf area. Leaf areas were determined using a leaf area meter (LI-3000;
Li-Cor). All values represent averages of four biological replicates 6 SD.
Total Protein Isolation and Germination Inhibition Assay
Total protein was extracted from wild-type R108, irg1-1, and irg1-2 M.
truncatula leaves. One gram of leaf tissue was homogenized in buffer (1:2
[w/v]) containing 50mMTris-Cl, pH 7.4, 100mMNaCl, 10%glycerol, 1 mM
phenylmethylsulfonyl fluoride, 10 mM MgCl2, 1 mM EDTA, and complete
miniprotease inhibitor cocktail according to themanufacturer’s instructions
(Roche Diagnostics). Homogenates were centrifuged at 12,000g for 5 min
at 48C to remove debris, and protein concentrations in the homogenates
were determined by the Bradford assay (Bio-Rad). One milliliter of the
urediniospores (23 106 spores/mL) ofP. emaculata and P. pachyrhiziwere
mixed with an equal volume of total protein isolated fromM. truncatulawild
type or irg1mutants at a final concentration of 20mgofprotein/mL, then the
urediniospores were incubated in 6-well cell plastic culture plates (BD
Falcon) for 12 h under dark condition. After incubation, the numbers of P.
emaculata and P. pachyrhizi spores that germinated and germ tubes with
and without differentiated appressoria were counted from five random
fields. The average is used to calculate the percentage of germination and
spores with and without an appressorium.
Statistical Analysis
Significant differences (P# 0.05) within the data sets were tested by one-
way analysis of variance followedDuncan’smultiple range test using SAS
9.2 (SAS Institute). We fitted a general linear model to our factorial data to
estimate the significance of both main effects and interactions for the
cuticular wax content analyses performed using GC-MS (Figure 5).
Similarly, the statistical significances in urediniospore differentiation
were determined using analysis of variance followed by Duncan’s mul-
tiple range test (Figures 2 and 7). Student’s paired t test was used to
analyze the significant differences between wild-type and mutant alleles
or mock and wax treatments (Figures 1, 6, and 8).
Microarray Analyses
To represent different alleles of the irg1 mutant, two independent sam-
pling of three alleles, irg1-1, irg1-2, and irg1-5, totaling six biological
replicates, were used in themicroarray analyses in comparison with three
biological replicates of wild-type plants. All plants were planted in the
greenhouse in separate pots in a single experiment. Total RNA was
purified from leaf tissues of 4-week-old M. truncatula R108 and irg1
mutants using TRIzol reagent (Invitrogen) according to the manufac-
turer’s instructions. Three plants were used for each biological replicate.
Total RNA was extracted from individual plants, and following isolation,
was pooled to represent one biological replicate. The integrity of the RNA
was checked on an Agilent Bioanalyzer 2100, and 10 mg of total RNA was
used as a template for amplification. Probe labeling, chip hybridization,
and scanning were performed according to the manufacturer’s instruc-
tions (Affymetrix). All the raw data were imported into Robust Multichip
Average software and normalized as described (Irizarry et al., 2003). The
presence/absence call for each probe set was obtained from dCHIP
(Fiebig et al., 2000), and differentially expressed genes between sample
pairs were selected using associative analyses, as described (Dozmorov
and Centola, 2003). Genes that showed significant differences in tran-
script levels (twofold or greater and P value < 8.15954E-07) between the
wild-type and mutant plants were selected for further analyses. Type I
family-wise error rate was reduced using the Bonferroni-corrected P
value threshold of 0.05/N, where N represents the number of genes
present on the chip, which is 61,278 forM. truncatula. In this method, the
background noise presented among replicates and technical noise
during microarray experiments was measured by the residual presented
among a group of genes whose residuals are homoscedastic. Genes
whose residuals between the compared sample pairs that are signifi-
cantly higher than themeasured background noise level were considered
to be differentially expressed. Due to the large gene number in the
background measurement, this method enabled Bonferroni correction
without being overly stringent. The data and completeMIAME information
are deposited at ArrayExpress (http://www.ebi.ac.uk/microarray-as/ae/)
under accession number E-MEXP-3190.
Host Surface Signaling in Rust Infection 367

Real-Time Quantitative RT-PCR and RNA Gel Blot Analyses
Real-time qRT-PCR was performed as described (Uppalapati et al., 2009)
using thegene-specificprimers (seeSupplemental Table 2online). TotalRNA
was treatedwith TurboDNase (Ambion) to eliminate genomicDNA, and 5mg
of DNase-treated RNA was reverse transcribed using Superscript III reverse
transcriptase (Invitrogen) with oligo(dT)20 primers. The cDNA (1:10) was then
used for qRT-PCR. The qRT-PCRwas performed using Power SYBRGreen
PCRmastermix (AppliedBiosystems) in anoptical 384-well platewith anABI
Prism7900HTsequencedetection system (AppliedBiosystems).Melt-curve
analysis was performed to monitor primer-dimer formation and to check
amplification of gene-specific products. The average threshold cycles (CT)
values calculated from triplicate biological samples were used to determine
the fold expression relative to the controls. Primers specific for ubiquitin were
used to normalize small differences in template amounts. For RNA gel blot
analysis, 10 mg of total RNA was denatured, fractionated by electrophoresis
on a 1% agarose gel, and then blotted onto nylon membranes (Amersham
Biosciences). Digoxigenin-labeled DNA probes were made with DIG PCR
labelingmix (BoehringerMannheim) using gene-specific forward and reverse
primer sets described earlier (Uppalapati et al., 2009). Hybridization was
performed at 658Covernight in a solution containing 50mMChurch buffer
(pH 7.2; Church and Gilbert 1984), 53 SSC (13 SSC is 0.15 M NaCl and
0.015 M sodium citrate), 0.1% laurylsarcosine, 7% SDS (w/v), and 2%
blocking solution (Boehringer Mannheim). Further washes were done at
658C in a solution containing 0.53 SSC and 0.1% SDS. Hybridized
mRNAs were detected with anti-DIG antibody conjugated with alkaline
phosphatase (Boehringer Mannheim) and its chemiluminescent sub-
strate, CDP-Star (Boehringer Mannheim).
Accession Numbers
Sequence data from this article for M. truncatula genes can be found in the
Dana Farber Cancer InstituteMedicago gene index, J. Craig Venter Institute–
Medicago Genome Resource, or GenBank/EMBL databases under the
following TC numbers or locus number: CER1-1 (TC130292), CER2
(TC115187), CER3 (TC125004), CER4-1 (TC122585), CER4-2 (TC160800),
CER6-1 (TC116151), CER6-2 (TC125487), CER6-3 (TC121408), CER6-4
(TC113573), CER8-1 (TC140235), CER10 (TC125186), PAS2 (GE348322),
KCR1 (TC113588), KCR2 (TC145787),WSD1 (TC145247), FATB (TC148248),
MYB30 (TC142663),MYB96 (Medtr4g135250/Medtr8g112130), IRG1/PALM1
(GI:298201167), and LTP (TC97568).
Supplemental Data
The following materials are available in the online version of this article.
Supplemental Figure 1. Expression Profiles for Selected Genes in the
Phenylpropanoid Pathway (PAL, CHS, CHR, CHI, IFS, and IFR) and
Pathogenesis-Related Genes (PR3 and PR10) in the Wild Type (R108)
at 0, 8, 24, and 48 h after Inoculation with P. emaculata Urediniospores.
Supplemental Figure 2. A Schematic Showing the Sequence of
Events for Conducting the Forward Genetic Screens to Identify M.
truncatula Genes Involved in Nonhost Resistance against P. emaculata.
Supplemental Figure 3. Expression Profiles for Selected Defense-
Related Genes and Antimicrobial Activity of Protein Extracts on
Fungal Differentiation.
Supplemental Figure 4. Rust Germ Tube Differentiation on Multiple
Mutant Alleles of irg1.
Supplemental Figure 5. Leaf Phenotype of 4-Week-Old Wild-Type
R108 and irg1-1 Plants of M. truncatula.
Supplemental Figure 6. A Simplified Wax Biosynthesis Pathway and
Some CER Genes Implicated in Wax Biosynthesis in Arabidopsis
(Modified from Kunst and Samuels, 2003; Samuels et al., 2008).
Supplemental Figure 7. Composition of Alcohols and Alkanes in
Wild-Type and irg1 Mutant Alleles of Medicago truncatula.
Supplemental Table 1. Fold Changes in Expression of Correspond-
ing M. truncatula Orthologs of Arabidopsis Wax Biosynthesis-Related
Genes Using qRT-PCR and Microarray Analysis.
Supplemental Table 2. List of Genes and Corresponding Primers
Used for qRT-PCR.
Supplemental Data Set 1. A List of Differentially Regulated Genes
(Minimum Twofold Up- or Downregulated) in Three Different Alleles of
irg1 (irg1-1/palm1-4, irg1-2 /palm1-5, and irg1-5/palm1-6).
ACKNOWEDGMENTS
We thank Markus Frank for helpful discussions, Yongfeng Zhang for
assistance with mutant screening, and Kelly Craven, Clemencia Rojas,
and Jackie Kelley for critical reading of the manuscript. We also thank
Azhaguvel Perumal and Muhammet Sakiroglu for guidance with statis-
tical analyses and Janie Gallaway and Colleen Elles for plant mainte-
nance in the greenhouse. This work was supported through a grant to
K.S.M. and S.R.U. from BASF Plant Sciences, Germany, and National
Science Foundation–Experimental Program to Stimulate Competitive
Research (EPS-0814361) to K.S.M. The insertion mutant lines of M.
truncatula were created through research support, in part, from the
National Science Foundation (DBI-0703285).
AUTHOR CONTRIBUTIONS
S.R.U. identified the mutant, designed research, analyzed data, and
wrote the article. Y.I. designed research, performed research, and
analyzed data. K.S.M. designed research and wrote the article. S.R.U.
and Y.I. performed the bulk of the experiments, and V.D. and S.M. helped
them with genetic screens. J.N. helped with scanning electron micros-
copy. M.B. performed wax analysis using GC-MS. Y.T. analyzed micro-
array data. J.C., M.T., P.R., R.C., and H.S. contributed reagents and
provided seed stocks for Tnt1 mutant populations or PALM1 comple-
mented lines.
Received October 24, 2011; revised December 16, 2011; accepted
December 31, 2011; published January 31, 2012.
REFERENCES
Aarts, M.G., Keijzer, C.J., Stiekema, W.J., and Pereira, A. (1995).
Molecular characterization of the CER1 gene of Arabidopsis involved
in epicuticular wax biosynthesis and pollen fertility. Plant Cell 7: 2115–
2127.
Aharoni, A., Dixit, S., Jetter, R., Thoenes, E., van Arkel, G., and
Pereira, A. (2004). The SHINE clade of AP2 domain transcription
factors activates wax biosynthesis, alters cuticle properties, and
confers drought tolerance when overexpressed in Arabidopsis. Plant
Cell 16: 2463–2480.
Ayliffe, M., Singh, R., and Lagudah, E. (2008). Durable resistance to
wheat stem rust needed. Curr. Opin. Plant Biol. 11: 187–192.
Bessire, M., Chassot, C., Jacquat, A.C., Humphry, M., Borel, S.,
Petetot, J.M., Metraux, J.P., and Nawrath, C. (2007). A permeable
cuticle in Arabidopsis leads to a strong resistance to Botrytis cinerea.
EMBO J. 26: 2158–2168.
Bourdenx, B., Bernard, A., Domergue, F., Pascal, S., Leger, A., Roby,
D., Pervent, M., Vile, D., Haslam, R.P., Napier, J.A., Lessire, R., and
Joubes, J. (2011). Overexpression of Arabidopsis ECERIFERUM1
368 The Plant Cell

promotes wax very-long-chain alkane biosynthesis and influences
plant response to biotic and abiotic stresses. Plant Physiol. 156:
29–45.
Bouton, J.H. (2007). Molecular breeding of switchgrass for use as a
biofuel crop. Curr. Opin. Genet. Dev. 17: 553–558.
Broun, P., Poindexter, P., Osborne, E., Jiang, C.Z., and Riechmann,
J.L. (2004). WIN1, a transcriptional activator of epidermal wax accu-
mulation in Arabidopsis. Proc. Natl. Acad. Sci. USA 101: 4706–4711.
Carver, T.L.W., Thomas, B.J., Ingerson-Morris, S.M., and Roderick,
H.W. (1990). The role of the abaxial leaf surface waxes of Lolium spp.
in resistance to Erysiphe graminis. Plant Pathol. 39: 573–583.
Chassot, C., Nawrath, C., and Metraux, J.P. (2007). Cuticular defects
lead to full immunity to a major plant pathogen. Plant J. 49: 972–980.
Chassot, C., Nawrath, C., and Metraux, J.P. (2008). The cuticle: Not
only a barrier for plant defence: A novel defence syndrome in plants
with cuticular defects. Plant Signal. Behav. 3: 142–144.
Chen, J., et al. (2010). Control of dissected leaf morphology by a Cys(2)
His(2) zinc finger transcription factor in the model legume Medicago
truncatula. Proc. Natl. Acad. Sci. USA 107: 10754–10759.
Chen, X., Goodwin, S.M., Liu, X., Chen, X., Bressan, R.A., and Jenks,
M.A. (2005). Mutation of the RESURRECTION1 locus of Arabidopsis
reveals an association of cuticular wax with embryo development.
Plant Physiol. 139: 909–919.
Church, G.M., and Gilbert, W. (1984). Genomic sequencing. Proc. Natl.
Acad. Sci. USA 81: 1991–1995.
Curvers, K., Seifi, H., Mouille, G., de Rycke, R., Asselbergh, B., Van
Hecke, A., Vanderschaeghe, D., Hofte, H., Callewaert, N., Van
Breusegem, F., and Hofte, M. (2010). Abscisic acid deficiency
causes changes in cuticle permeability and pectin composition that
influence tomato resistance to Botrytis cinerea. Plant Physiol. 154:
847–860.
Debono, A., Yeats, T.H., Rose, J.K., Bird, D., Jetter, R., Kunst, L.,
and Samuels, L. (2009). Arabidopsis LTPG is a glycosylphosphati-
dylinositol-anchored lipid transfer protein required for export of lipids
to the plant surface. Plant Cell 21: 1230–1238.
d’Erfurth, I., Cosson, V., Eschstruth, A., Lucas, H., Kondorosi, A.,
and Ratet, P. (2003). Efficient transposition of the Tnt1 tobacco
retrotransposon in the model legume Medicago truncatula. Plant J.
34: 95–106.
Dozmorov, I., and Centola, M. (2003). An associative analysis of gene
expression array data. Bioinformatics 19: 204–211.
Fiebig, A., Mayfield, J.A., Miley, N.L., Chau, S., Fischer, R.L., and
Preuss, D. (2000). Alterations in CER6, a gene identical to CUT1,
differentially affect long-chain lipid content on the surface of pollen
and stems. Plant Cell 12: 2001–2008.
Garcia, A., Calvo, E.S., de Souza Kiihl, R.A., Harada, A., Hiromoto,
D.M., and Vieira, L.G.E. (2008). Molecular mapping of soybean rust
(Phakopsora pachyrhizi) resistance genes: Discovery of a novel locus
and alleles. Theor. Appl. Genet. 117: 545–553.
Gniwotta, F., Vogg, G., Gartmann, V., Carver, T.L.W., Riederer, M.,
and Jetter, R. (2005). What do microbes encounter at the plant
surface? Chemical composition of pea leaf cuticular waxes. Plant
Physiol. 139: 519–530.
Goellner, K., Loehrer, M., Langenbach, C., Conrath, U., Koch, E.,
and Schaffrath, U. (2010). Phakopsora pachyrhizi, the causal agent of
Asian soybean rust. Mol. Plant Pathol. 11: 169–177.
Hansjakob, A., Bischof, S., Bringmann, G., Riederer, M., and
Hildebrandt, U. (2010). Very-long-chain aldehydes promote in vitro
prepenetration processes of Blumeria graminis in a dose- and chain
length-dependent manner. New Phytol. 188: 1039–1054.
Hansjakob, A., Riederer, M., and Hildebrandt, U. (2011). Wax matters:
Absence of very-long-chain aldehydes from the leaf cuticular wax of
the glossy11 mutant of maize compromises the prepenetration pro-
cesses of Blumeria graminis. Plant Pathol. 60: 1151–1161.
Heath, M.C. (1977). A comparative study of non-host interactions with
rust fungi. Physiol. Plant Pathol. 1: 73–76.
Heath, M.C. (2000). Nonhost resistance and nonspecific plant defenses.
Curr. Opin. Plant Biol. 3: 315–319.
Hoch, H.C., Staples, R.C., Whitehead, B., Comeau, J., and Wolf, E.D.
(1987). Signaling for growth orientation and cell differentiation by
surface topography in uromyces. Science 235: 1659–1662.
Hyten, D.L., Hartman, G.L., Nelson, R.L., Frederick, R.D., Concibido,
V.C., Narvel, J.M., and Cregan, P.B. (2007). Map location of the
locus that confers resistance to soybean rust in soybean. Crop Sci.
47: 837–838.
Irizarry, R.A., Bolstad, B.M., Collin, F., Cope, L.M., Hobbs, B., and
Speed, T.P. (2003). Summaries of Affymetrix GeneChip probe level
data. Nucleic Acids Res. 31: e15.
Javelle, M., Vernoud, V., Depege-Fargeix, N., Arnould, C., Oursel, D.,
Domergue, F., Sarda, X., and Rogowsky, P.M. (2010). Overexpres-
sion of the epidermis-specific homeodomain-leucine zipper IV tran-
scription factor Outer Cell Layer1 in maize identifies target genes
involved in lipid metabolism and cuticle biosynthesis. Plant Physiol.
154: 273–286.
Jenks, M.A., Tuttle, H.A., Eigenbrode, S.D., and Feldmann, K.A.
(1995). Leaf epicuticular waxes of the eceriferum mutants in Arabi-
dopsis. Plant Physiol. 108: 369–377.
Kader, J.C. (1996). Lipid-transfer proteins in plants. Annu. Rev. Plant
Physiol. Plant Mol. Biol. 47: 627–654.
Koch, E., and Hoppe, H.H. (1988). Development of infection structures
by the direct-penetrating soybean rust fungus (Phakopsora pachyrhizi
Syd.) on artificial membranes. J. Phytopathol. 122: 232–244.
Kolattukudy, P.E., Rogers, L.M., Li, D., Hwang, C.S., and Flaishman,
M.A. (1995). Surface signaling in pathogenesis. Proc. Natl. Acad. Sci.
USA 92: 4080–4087.
Kunst, L., and Samuels, A.L. (2003). Biosynthesis and secretion of
plant cuticular wax. Prog. Lipid Res. 42: 51–80.
Kunst, L., and Samuels, L. (2009). Plant cuticles shine: Advances in
wax biosynthesis and export. Curr. Opin. Plant Biol. 12: 721–727.
Lee, S.B., Go, Y.S., Bae, H.-J., Park, J.H., Cho, S.H., Cho, H.J., Lee,
D.S., Park, O.K., Hwang, I., and Suh, M.C. (2009). Disruption of
glycosylphosphatidylinositol-anchored lipid transfer protein gene al-
tered cuticular lipid composition, increased plastoglobules, and en-
hanced susceptibility to infection by the fungal pathogen Alternaria
brassicicola. Plant Physiol. 150: 42–54.
Lee, Y.-H., and Dean, R.A. (1994). Hydrophobicity of contact surface
induces appressorium formation in Magnaporthe grisea. FEMS Micro-
biol. Lett. 115: 71–75.
Li, Y., Beisson, F., Koo, A.J., Molina, I., Pollard, M., and Ohlrogge, J.
(2007). Identification of acyltransferases required for cutin biosynthe-
sis and production of cutin with suberin-like monomers. Proc. Natl.
Acad. Sci. USA 104: 18339–18344.
Lu, S., Song, T., Kosma, D.K., Parsons, E.P., Rowland, O., and
Jenks, M.A. (2009). Arabidopsis CER8 encodes LONG-CHAIN ACYL-
COA SYNTHETASE 1 (LACS1) that has overlapping functions with
LACS2 in plant wax and cutin synthesis. Plant J. 59: 553–564.
Mang, H.G., et al. (2009). The Arabidopsis RESURRECTION1 gene
regulates a novel antagonistic interaction in plant defense to biotrophs
and necrotrophs. Plant Physiol. 151: 290–305.
Mao, B., et al. (2012). Wax crystal-sparse leaf2, a rice homologue of
WAX2/GL1, is involved in synthesis of leaf cuticular wax. Planta 235:
39–52.
Millar, A.A., Clemens, S., Zachgo, S., Giblin, E.M., Taylor, D.C., and
Kunst, L. (1999). CUT1, an Arabidopsis gene required for cuticular
Host Surface Signaling in Rust Infection 369

wax biosynthesis and pollen fertility, encodes a very-long-chain fatty
acid condensing enzyme. Plant Cell 11: 825–838.
Monteros, M., Ha, B.-K., Phillips, D., and Boerma, H. (2010). SNP
assay to detect the ‘Hyuuga’ red-brown lesion resistance gene for
Asian soybean rust. Theor. Appl. Genet. 121: 1023–1032.
Muller, C., and Riederer, M. (2005). Plant surface properties in chem-
ical ecology. J. Chem. Ecol. 31: 2621–2651.
Mysore, K.S., and Ryu, C.M. (2004). Nonhost resistance: How much do
we know? Trends Plant Sci. 9: 97–104.
Pighin, J.A., Zheng, H., Balakshin, L.J., Goodman, I.P., Western,
T.L., Jetter, R., Kunst, L., and Samuels, A.L. (2004). Plant cuticular
lipid export requires an ABC transporter. Science 306: 702–704.
Podila, G.K., Rogers, L.M., and Kolattukudy, P.E. (1993). Chemical
signals from avocado surface wax trigger germination and appresso-
rium formation in Colletotrichum gloeosporioides. Plant Physiol. 103:
267–272.
Posada-Buitrago, M.L., and Frederick, R.D. (2005). Expressed se-
quence tag analysis of the soybean rust pathogen Phakopsora
pachyrhizi. Fungal Genet. Biol. 42: 949–962.
Reisige, K., Gorzelanny, C., Daniels, U., and Moerschbacher, B.M.
(2006). The C28 aldehyde octacosanal is a morphogenetically active
component involved in host plant recognition and infection structure
differentiation in the wheat stem rust fungus. Physiol. Mol. Plant
Pathol. 68: 33–40.
Riederer, M., and Schreiber, L. (2001). Protecting against water loss:
Analysis of the barrier properties of plant cuticles. J. Exp. Bot. 52:
2023–2032.
Ringelmann, A., Riedel, M., Riederer, M., and Hildebrandt, U. (2009).
Two sides of a leaf blade: Blumeria graminis needs chemical cues in
cuticular waxes of Lolium perenne for germination and differentiation.
Planta 230: 95–105.
Rowland, O., Lee, R., Franke, R., Schreiber, L., and Kunst, L. (2007).
The CER3 wax biosynthetic gene from Arabidopsis thaliana is allelic to
WAX2/YRE/FLP1. FEBS Lett. 581: 3538–3544.
Rowland, O., Zheng, H., Hepworth, S.R., Lam, P., Jetter, R., and Kunst,
L. (2006). CER4 encodes an alcohol-forming fatty acyl-coenzyme A
reductase involved in cuticular wax production in Arabidopsis. Plant
Physiol. 142: 866–877.
Samuels, L., Kunst, L., and Jetter, R. (2008). Sealing plant surfaces:
Cuticular wax formation by epidermal cells. Annu. Rev. Plant Biol. 59:
683–707.
Segura, A., Moreno, M., and Garcıa-Olmedo, F. (1993). Purification
and antipathogenic activity of lipid transfer proteins (LTPs) from the
leaves of Arabidopsis and spinach. FEBS Lett. 332: 243–246.
Seo, P.J., Lee, S.B., Suh, M.C., Park, M.J., Go, Y.S., and Park, C.M.
(2011). The MYB96 transcription factor regulates cuticular wax bio-
synthesis under drought conditions in Arabidopsis. Plant Cell 23:
1138–1152.
Silva, D.C., Yamanaka, N., Brogin, R.L., Arias, C.A., Nepomuceno,
A.L., Di Mauro, A.O., Pereira, S.S., Nogueira, L.M., Passianotto, A.
L., and Abdelnoor, R.V. (2008). Molecular mapping of two loci that
confer resistance to Asian rust in soybean. Theor. Appl. Genet. 117:
57–63.
Tadege, M., Wen, J., He, J., Tu, H., Kwak, Y., Eschstruth, A., Cayrel,
A., Endre, G., Zhao, P.X., Chabaud, M., Ratet, P., and Mysore, K.S.
(2008). Large-scale insertional mutagenesis using the Tnt1 retrotrans-
poson in the model legume Medicago truncatula. Plant J. 54: 335–347.
Tang, D., Simonich, M.T., and Innes, R.W. (2007). Mutations in LACS2,
a long-chain acyl-coenzyme A synthetase, enhance susceptibility to
avirulent Pseudomonas syringae but confer resistance to Botrytis
cinerea in Arabidopsis. Plant Physiol. 144: 1093–1103.
Uppalapati, S.R., and Fujita, Y. (2000). Carbohydrate regulation of
attachment, encystment, and appressorium formation by Pythium
porphyrae (Oomycota) zoospores on Porphyra yezoensis (Rhodophyta).
J. Phycol. 36: 359–366.
Uppalapati, S.R., Marek, S.M., Lee, H.K., Nakashima, J., Tang, Y.,
Sledge, M.K., Dixon, R.A., and Mysore, K.S. (2009). Global gene
expression profiling during Medicago truncatula-Phymatotrichopsis
omnivora interaction reveals a role for jasmonic acid, ethylene, and
the flavonoid pathway in disease development. Mol. Plant Microbe
Interact. 22: 7–17.
van de Mortel, M., Recknor, J.C., Graham, M.A., Nettleton, D.,
Dittman, J.D., Nelson, R.T., Godoy, C.V., Abdelnoor, R.V., Almeida,
A.M.R., Baum, T.J., and Whitham, S.A. (2007). Distinct biphasic
mRNA changes in response to Asian soybean rust infection. Mol. Plant
Microbe Interact. 20: 887–899.
Xia, Y., Gao, Q.M., Yu, K., Lapchyk, L., Navarre, D., Hildebrand, D.,
Kachroo, A., and Kachroo, P. (2009). An intact cuticle in distal
tissues is essential for the induction of systemic acquired resistance in
plants. Cell Host Microbe 5: 151–165.
Xia, Y., Nikolau, B.J., and Schnable, P.S. (1997). Developmental and
hormonal regulation of the Arabidopsis CER2 gene that codes for a
nuclear-localized protein required for the normal accumulation of
cuticular waxes. Plant Physiol. 115: 925–937.
Xia, Y., Yu, K., Navarre, D., Seebold, K., Kachroo, A., and Kachroo,
P. (2010). The glabra1 mutation affects cuticle formation and plant
responses to microbes. Plant Physiol. 154: 833–846.
Xiao, F., Goodwin, S.M., Xiao, Y., Sun, Z., Baker, D., Tang, X., Jenks,
M.A., and Zhou, J.M. (2004). Arabidopsis CYP86A2 represses Pseu-
domonas syringae type III genes and is required for cuticle develop-
ment. EMBO J. 23: 2903–2913.
Yeats, T.H., Howe, K.J., Matas, A.J., Buda, G.J., Thannhauser, T.W.,
and Rose, J.K. (2010). Mining the surface proteome of tomato
(Solanum lycopersicum) fruit for proteins associated with cuticle
biogenesis. J. Exp. Bot. 61: 3759–3771.
Young, N.D., and Udvardi, M. (2009). Translating Medicago truncatula
genomics to crop legumes. Curr. Opin. Plant Biol. 12: 193–201.
Zabka, V., Stangl, M., Bringmann, G., Vogg, G., Riederer, M., and
Hildebrandt, U. (2008). Host surface properties affect prepenetration
processes in the barley powdery mildew fungus. New Phytol. 177:
251–263.
Zhang, J.Y., Broeckling, C.D., Blancaflor, E.B., Sledge, M.K., Sumner,
L.W., and Wang, Z.Y. (2005). Overexpression of WXP1, a putative
Medicago truncatula AP2 domain-containing transcription factor gene,
increases cuticular wax accumulation and enhances drought tolerance
in transgenic alfalfa (Medicago sativa). Plant J. 42: 689–707.
Zhang, J.Y., Broeckling, C.D., Sumner, L.W., and Wang, Z.Y. (2007).
Heterologous expression of two Medicago truncatula putative ERF
transcription factor genes, WXP1 and WXP2, in Arabidopsis led to
increased leaf wax accumulation and improved drought tolerance, but
differential response in freezing tolerance. Plant Mol. Biol. 64: 265–278.
370 The Plant Cell

DOI 10.1105/tpc.111.093104; originally published online January 31, 2012; 2012;24;353-370Plant Cell
Schultheiss and Kirankumar S. MysoreJianghua Chen, Jin Nakashima, Yuhong Tang, Million Tadege, Pascal Ratet, Rujin Chen, Holger
Srinivasa Rao Uppalapati, Yasuhiro Ishiga, Vanthana Doraiswamy, Mohamed Bedair, Shipra Mittal,Reduced Spore Differentiation of Anthracnose and Nonhost Rust Pathogens
Mutants Results inMedicago truncatula irg1/palm1Loss of Abaxial Leaf Epicuticular Wax in
This information is current as of January 24, 2019
Supplemental Data /content/suppl/2012/01/13/tpc.111.093104.DC1.html
References /content/24/1/353.full.html#ref-list-1
This article cites 71 articles, 27 of which can be accessed free at:
Permissions https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298X
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists