los restos directos de dinosaurios ... - … · dinosaurios y otros reptiles mesozoicos de españa,...
TRANSCRIPT

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
1
LOS RESTOS DIRECTOS DE DINOSAURIOS TERÓPODOS
(EXCLUYENDO AVES) EN ESPAÑA
CANUDO1, J. I. y RUIZ-OMEÑACA1,2 J. I.
1 Departamento de Ciencias de la Tierra (Área de Paleontología) y Museo Paleontológico.Universidad de Zaragoza. 50009 Zaragoza. [email protected] Paleoymás, S. L. L. Nuestra Señora del Salz, 4, local, 50017 Zaragoza. [email protected]
RESUMEN
La mayoría de los restos fósiles de dinosaurios terópodos de España son dientes aislados yescasos restos postcraneales. La única excepción es el ornitomimosaurio Pelecanimimus polyodon,del Barremiense de Las Hoyas (Cuenca). Hay registro de terópodos en el Jurásico superior(Oxfordiense superior-Tithónico inferior), en el tránsito Jurásico-Cretácico (Tithónico superior-Berriasiense inferior) y en todos los pisos del Cretácico inferior, con excepción del Valanginiense. Enel Cretácico superior únicamente hay restos en el Campaniense y Maastrichtiense. La mayor parte delas determinaciones son demasiado generales, lo que impide conocer algunas de las familias queposiblemente estén representadas. Se han reconocido: Neoceratosauria, Baryonychidae,Ornithomimosauria, Dromaeosauridae, además de terópodos indeterminados, y celurosauriosindeterminados (dientes pequeños sin dentículos). La mayoría de los restos son deManiraptoriformes, siendo especialmente abundantes los dromeosáuridos. Las únicas excepcionesson por el momento, el posible Ceratosauria del Jurásico superior de Asturias, los barionícidos delHauteriviense-Barremiense de Burgos, Teruel y La Rioja, el posible carcharodontosáurido delAptiense inferior de Morella y el posible abelisáurido del Campaniense de Laño. Además hay algunosterópodos incertae sedis, como los "paronicodóntidos" (entre los que se incluye Euronychodon), yRichardoestesia.
Palabras clave. Theropoda, Jurásico, Cretácico, España
Most fossil remains of theropod dinosaurs from the Iberian Peninsula are isolated teeth and rare post-cranial bones. The only exception is the ornithomimosaur Pelecanimimus polyodon from theBarremian of Las Hoyas (Cuenca). There are records of theropods in the Upper Jurassic (UpperOxfordian –Lower Tithonian), the Jurassic-Cretaceous transition (Upper Tithonian-Lower Berriasian)and in all the stages of the Lower Cretaceous, with the exception of the Valanginian. In the UpperCretaceous they are only found in the Campanian and Maastrichtian. Most of the findings are notdiagnostic, which makes it difficult identify all the families that may be represented. The groups includeNeoceratosauria, Baryonychidae, Ornithomimosauria, Dromaeosauridae, in addition to unidentifiedtheropods and coelurosaurs (represented by small-unserrated teeth). Most of the remains areManiraptoriformes, especially dromaeosaurids. The only exceptions at the moment are the possibleceratosaur from the Upper Jurassic (from Asturias), the baryonychids from the Hauterivian-Barremian(Burgos, Teruel and La Rioja), the possible carcharodontosaurid from the Lower Aptian (Morella) andthe possible abelisaurid from the Campanian of Laño. There are also some theropods incertae sedis,like the “paronichodontids”, among which we include Euronychodon, and Richardoestesia.
Key words. Theropoda, Jurassic, Cretaceous, Spain

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
2
INTRODUCCIÓN
Los terópodos son dinosaurios generalmente carnívoros, bípedos, y de tamaño
variable. Los dinosaurios más típicos de este grupo poseen los miembros anteriores
más pequeños que los posteriores, y las manos presentan adaptaciones para
mejorar su capacidad en la caza. El pie tiene tres dedos funcionales, con el dedo I
generalmente colocado en la parte posterior y por encima del resto de los dedos, y el
dedo V ausente o muy reducido. Tienen tendencia a poseer articulaciones
intracraneales y neumaticidad en los huesos. La columna vertebral suele ser
horizontal, con excepción del cuello que tiene una característica S. Poseen dientes
comprimidos lateromedialmente, curvados hacia atrás y con dentículos en los bordes
anterior y posterior. En formas derivadas la morfología de los dientes puede variar,
por ejemplo perdiéndose los dentículos. También hay taxones que carecen de
dientes, y otros con una morfología dental que indica una dieta omnívora o incluso
herbívora. Los terópodos tienen una amplia distribución, ya que se encuentran
desde el Triásico hasta el final del Cretácico (Gauthier, 1986; Sereno, 1997; Holtz,
1998; Sanz, 2000). Se han propuesto diferentes filogenias que han permitido
avanzar en los últimos años sobre la diferenciación de los grandes clados, que son
los usados en este trabajo (véase Holtz, 1998).
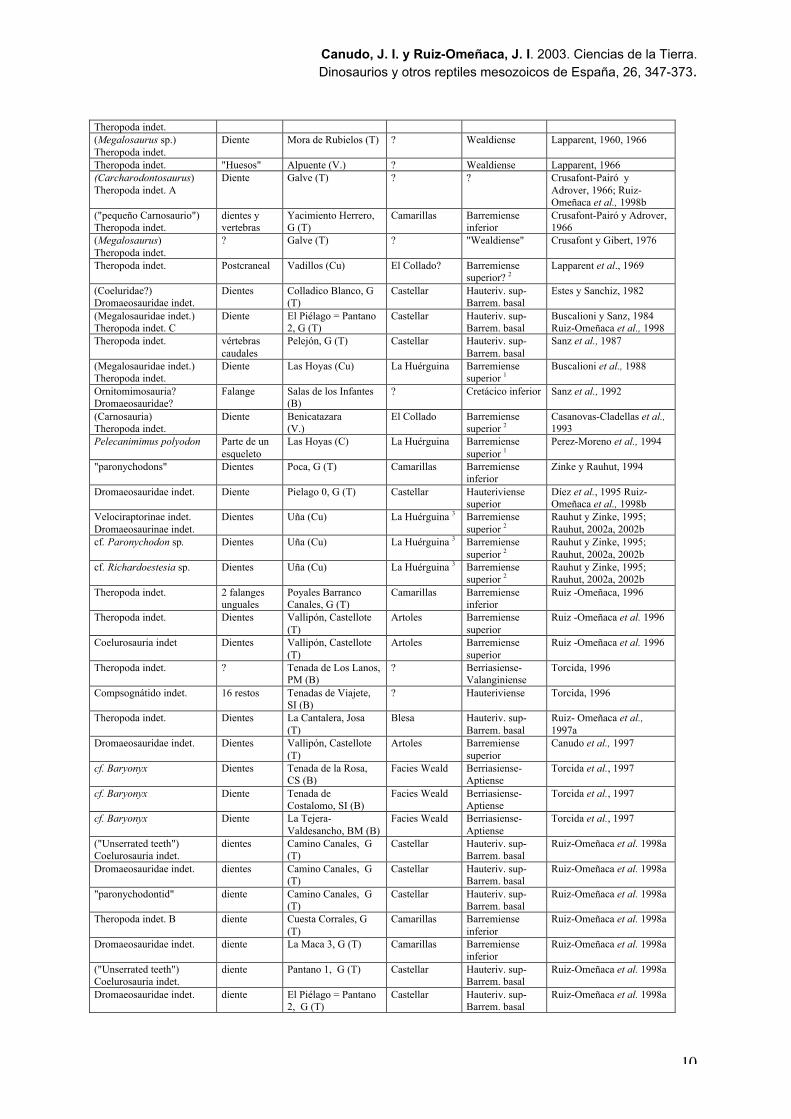
Taxón Material Yacimiento Formación Edad * Referencia(Megalosaurus sp.)Theropoda indet.
diente Ruedes (A) ? Lastres Kimmeridgiense sup-Tithónico inf. ?
Egozcue,1873
?Dromaeosauridaeindet.
dientes Entre Gijón yRibadesella (A)
Lastres Kimmeridgiense sup-Tithónico inf.
Martínez et al.,2000a, 2001
Theropoda indet. diente Entre Gijón yRibadesella (A)
Vega Oxfordiense sup-Kimmeridgiense inf.
Martínez et al.,2000a.
(Theropoda indet.)?Ceratosauria
vértebracaudal
playa de la Vega,Ribadesella (A)
Vega Oxfordiense sup-Kimmeridgiense inf.
Martínez et al.,2000b
Tabla 1. Restos directos de terópodos (excluyendo Avialae) en el Jurásico Superior de España.Abreviaturas: A: Asturias. Referencias en la bibliografía. Entre paréntesis están las determinacionesoriginales de los taxones que se han modificado en este trabajo. *: las dataciones de las Formacionesson de Schudack y Schudack (2002), y de Suarez-Vega (1974) para el diente de Ruedes.
Los restos directos de dinosaurios terópodos en España son relativamente
abundantes, pero hasta el momento son fragmentarios, con alguna notable
excepción. Los dientes aislados son los más abundantes, ya que los terópodos
solían tener más de 50 dientes en sus mandíbulas y además, estos dientes, como

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
3
los de otros dinosaurios, eran continuamente reemplazados durante la vida del
animal (Currie et al., 1990).
Figura 1. Terópodos del Jurásico Superior de España. A: diente de Theropoda indet. (JRVH-20),Formación Vega (Asturias) en vista labial/lingual. B: diente de Theropoda indet. (JRLH-180),Formación Lastres (Asturias) en vista labial/lingual. C-D: vértebra caudal anterior de Ceratosauria?indet., playa de La Vega (Ribadesella, Asturias), en vistas posterior (C) y lateral izquierda (D).Tomados de Martínez et al., 2000a (A-B) y Martínez et al., 2000b (C-D).
El objetivo de este trabajo es poner al día los conocimientos sobre los restos
directos de los terópodos españoles (excluyendo Avialae), incluyendo únicamente
los datos publicados en revistas científicas. Se han recogido todas las citas y se han
situado en el actual contexto cronológico y sistemático. Las tablas se han ordenado
por la fecha de publicación. En el caso de restos citados en varios trabajos se ha
puesto la referencia más moderna. En las Tablas se ha mantenido la determinación
original, pero dado que algunas son manifiestamente obsoletas, se discuten en el
texto, proponiéndose en su caso otras determinaciones. Se han publicado restos de
la provincias de Álava (Condado de Treviño), Asturias, Burgos, Castellón, Cuenca,
Huesca, Lérida, La Rioja y Segovia, en yacimientos del Jurásico superior, Cretácico
inferior y el final del Cretácico superior. Para una mejor comprensión, se divide el
registro en cinco intervalos: Jurásico superior, tránsito Jurásico-Cretácico,
"Neocomiense" (Berriasiense-Barremiese), Aptiense-Albiense, Campaniense y
Maastrichtiense.
Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
4
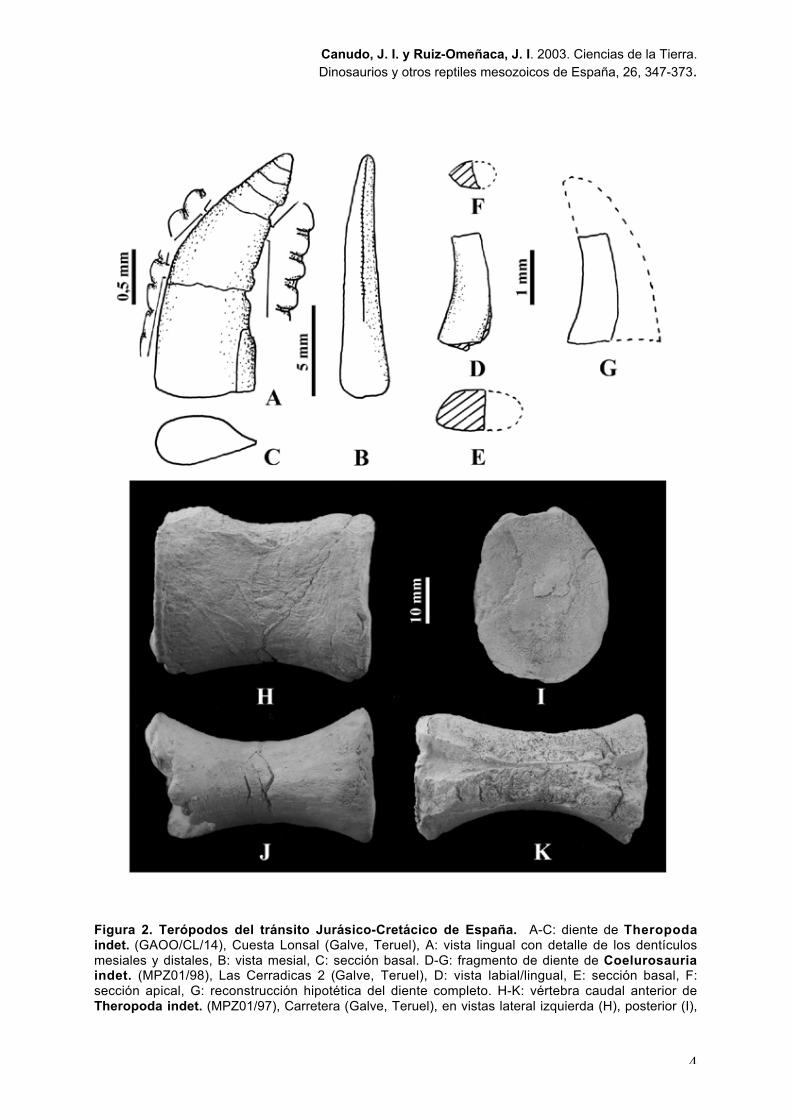
Figura 2. Terópodos del tránsito Jurásico-Cretácico de España. A-C: diente de Theropodaindet. (GAOO/CL/14), Cuesta Lonsal (Galve, Teruel), A: vista lingual con detalle de los dentículosmesiales y distales, B: vista mesial, C: sección basal. D-G: fragmento de diente de Coelurosauriaindet. (MPZ01/98), Las Cerradicas 2 (Galve, Teruel), D: vista labial/lingual, E: sección basal, F:sección apical, G: reconstrucción hipotética del diente completo. H-K: vértebra caudal anterior deTheropoda indet. (MPZ01/97), Carretera (Galve, Teruel), en vistas lateral izquierda (H), posterior (I),

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
5
ventral (J) y dorsal (K). Modificados de Barco y Ruiz-Omeñaca, 2001a (A-G) y Barco y Ruiz-Omeñaca, 2001b (H-K).
TEROPODOS DEL JURÁSICO SUPERIOR (Tabla 1, Figura 1)
Los restos de terópodos del Jurásico superior de España se han recuperado en
los afloramientos que constituyen la "Costa de los Dinosaurios" en Asturias
(Martínez et al., 2000a, 2000b, 2001). Es precisamente en este área donde se
encontró el primer resto español de dinosaurio publicado. Schulz (1858) cita un
diente aislado del Lías de Ruedes que asigna al tiburón Squalus , y que
posteriormente es determinado por Egozcue (1873) como del terópodo
Megalosaurus (Sanz, 1984; Sanz et al., 1990). A partir del trabajo de Schulz, la edad
del Jurásico detrítico en los alrededores de Lastres se consideró tradicionalmente
como Lías, hasta comienzos del siglo XX. Sin embargo los invertebrados de los
mismos niveles del diente de Megalosaurus indican una edad Jurásico superior
(Suárez Vega, 1974), y el diente de Ruedes posiblemente se encontró en la
Formación Lastres (Piñuela, com. personal), recientemente datada como
Kimmeridgiense superior-Tithónico inferior (Schudack y Schudack, 2002). Como se
discute más adelante, durante un tiempo hubo una tendencia a considerar como
Megalosaurus los dientes de grandes terópodos, por lo que sería necesaria una
futura revisión del material (actualmente en paradero desconocido) para confirmar
esta presencia en el Jurásico de España, y sólo puede asignarse a Theropoda indet.
Martínez et al. (2000a) describen un diente de Theropoda indet. en la formación
Vega, datada mediante ostrácodos como Oxfordiense superior-Kinmeridgiense
inferior (Schudack y Schudack, 2002). Este diente posee una morfología primitiva,
que se caracteriza por poseer dentículos de tamaño similar en las carenas mesial y
distal.
En la Formación Lastres (Kimmeridgiense superior-Tithónico inferior, según
Schudack y Schudack, 2002), Martínez et al. (2000a) citan un diente también con
morfología primitiva, similar al descrito en la formación Vega. Además, Martínez et
al. (2000a) describen dos dientes de esta formación, similares a los de
Dromaeosaurus, que asignan a Theropoda indet., aunque posteriormente, Martínez

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
6
et al. (2001) dicen que, por tener constricción en la base de la corona y dentículos
mesiales sólo en el tercio apical, podrían tener afinidad con Dromaeosauridae. La
presencia de dromeosáuridos en el Jurásico Superior de España no es extraña,
pues se han encontrado dientes de dromeosáuridos en el Jurásico superior de
Estados Unidos y Portugal (Chure, 1995; Zinke, 1998).
Taxón Material Yacimiento Formación Edad * ReferenciaTheropoda indet. diente Cuesta Lonsal,
Galve (T)Villar delArzobispo
Tithónico-Berriasiense
Barco y Ruiz-Omeñaca, 2001a
Coelurosauria indet. diente Cerradicas 2, Galve(T)
Villar delArzobispo
Tithónico-Berriasiense
Barco y Ruiz-Omeñaca, 2001a
Theropoda indet. vértebracaudal
Carretera, Galve(T)
Villar delArzobispo
Tithónico-Berriasiense
Barco y Ruiz-Omeñaca, 2001b
Tabla 2. Restos directos de terópodos (excluyendo Avialae) en el tránsito Jurásico-Cretácicode España. Abreviaturas: T: Teruel. Referencias en la bibliografía. *: la datación de la Fm. Villar delArzobispo en Galve es de Díaz-Molina et al. (1985) y Pérez-Lorente et al. (1997).
Respecto a material postcraneal, Martínez et al., (2000b) describen en la
formación Vega una vértebra caudal anterior de un terópodo de gran tamaño,
proveniente de los acantilados de la playa de la Vega (Ribadesella). El centro
presenta un surco ventral longitudinal bien desarrollado. Martínez et al. (2000b)
apuntan que la vértebra tiene algunos caracteres que podrían acercarle a
Ceratosaurus (Ceratosauria) y otros a Allosaurus (Tetanurae: Avetheropoda), pero
dado lo fragmentario del material prefieren considerarla como Theropoda indet. La
presencia de surco ventral en las vértebras caudales anteriores es un carácter
diagnóstico para Ceratosauria (carácter 193.1 de Holtz, 1998), por lo que esta
vértebra podría indicar la presencia de este clado en el Jurásico superior de España.
TEROPODOS DEL TRÁNSITO JURÁSICO - CRETÁCICO (Tabla 2, Figura 2)
El tránsito Jurásico-Cretácico está bien representado en la Cordillera Ibérica, pero
únicamente se han encontrado restos fragmentarios de terópodos en la Formación
Villar del Arzobispo, datada como Tithónico-Berriasiense (Díaz-Molina et al., 1985,
Pérez-Lorente et al., 1997). En Galve (Teruel) hay dos dientes de dos taxones
distintos. Uno de ellos, recuperado en el yacimiento de Cuesta Lonsal y asignado a

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
7
Theropoda indet., posee dentículos mesiales y distales (Barco y Ruiz-Omeñaca,
2001a). El segundo es un pequeño fragmento de diente del yacimiento Las
Cerradicas 2 sin dentículos en la mitad distal (la única conservada), asignado a
Coelurosauria indet. (Barco y Ruiz-Omeñaca, 2001a). El único resto postcraneal se
ha recuperado en el yacimiento Carretera, también en Galve, y es un centro
vertebral caudal anficélico. Algunos caracteres, como la ausencia de surco ventral
parecen indicar que pertenece a Tetanurae, pero dado lo fragmentario del material
se ha clasificado como Theropoda indet. (Barco y Ruiz-Omeñaca, 2001b).
TEROPODOS DEL "NEOCOMIENSE" (Tabla 3, Figuras 3 y 4)
Los restos de terópodos son relativamente abundantes en el Hauteriviense-
Barremiense de la Península Ibérica, pero generalmente son dientes aislados o
restos postcraneales fragmentarios, por lo que muchas de las determinaciones son
generales. A pesar de esta dificultad, se puede observar que hay una significativa
variedad de pequeños y grandes terópodos. La mayoría de los restos identificados
se incluyen en Maniraptoriformes, con excepción de los barionícidos.
Los dientes aislados de espinosauroideos barionícidos cercanos a Baryonyx son
relativamente frecuentes en el Hauteriviense superior y el Barremiense de España.
Se han determinado como Baryonychidae indet. y cf. Baryonyx sp. (Ruiz-Omeñaca
et al., 1998b; Torcida et al., 1997). También se han descrito elementos craneales y
postcraneales de la Formación Pinilla de los Moros en Salas de los Infantes
(Fuentes Vidarte et al., 2001). Los dientes de los barionícidos se caracterizan por su
forma cónica, una ornamentación longitudinal muy parecida a la de los dientes de
algunos cocodrilos y un reducido desarrollo de los dentículos (Martill y Hutt, 1996), y
se diferencian fácilmente de los típicos dientes de terópodos, comprimidos
lateralmente, curvados distalmente y, en general, sin ornamentación (Currie et al.,
1990). Esta similitud morfológica con los dientes de los cocodrilos, así como su
cráneo alargado y el descubrimiento de supuestos contenidos estomacales ha hecho
que se considere, al menos algunos de sus representantes, como piscívoros (Charig
y Milner, 1997). Aunque algunos dientes aislados se han asignado a Baryonyx, la

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
8
variedad morfológica de los dientes parece indicar la presencia de varios taxones
(Ruiz-Omeñaca et al., 1998b). Los dientes del holotipo de Baryonyx walkeri tienen
alrededor de 7 dentículos/mm (Charig y Milner, 1997), y en el Barremiense de Isla de
Wight (Inglaterra) se han descrito dientes aislados de barionícidos (cf. Baryonyx) que
se caracterizan por tener 7-8 dentículos/mm (Martill y Hutt, 1996). En el Barremiense
de Aragón se han descrito dientes con una morfología similar a estos barionícidos,
con 9 dentículos/mm, pero que se diferencian de Baryonyx por carecer de dentículos
en la carena mesial, por lo que se han atribuido a Baryonychidae indet. (Ruiz-
Omeñaca et al., 1998b).
Figura 3. Dientes de terópodos del Neocomiense de España. A: diente lateral deVelociraptorinae indet. (MPZ97/521), Camino Canales (Galve, Teruel), en vista labial/lingual ydetalle de los dentículos distales inclinados. B: diente premaxilar de Dromaeosauridae indet.(MPZ96/78),Vallipón (Castellote, Teruel), en vista lingual y detalle de los dentículos distales rectos. C:diente lateral de Coelurosauria indet. (MPZ96/65), Vallipón (Castellote, Teruel). D: diente de cf.Richardoestesia sp., Uña (Cuenca), en vista labial/lingual. E-G: dientes de cf. Paronychodon sp.,Uña (Cuenca), en vistas labial (E), apical (F) y lingual (G). B y C modificados de Ruiz-Omeñaca et al.,1996; D-G tomados de Rauhut, 2000b.
Los Maniraptoriformes están representados por dientes aislados, generalmente de
pequeño tamaño obtenidos por las técnicas de lavado-tamizado (Estes y Sanchiz,

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
9
1982; Cuenca-Bescós et al., 1999), siendo los restos postcraneales escasos y
fragmentarios. Como ejemplo, en el Hauteriviense-Barremiense de Galve (Teruel)
hay seis morfotipos distintos de dientes de tipo "dromeosáurido" (Ruiz-Omeñaca et
al., 1998a; Ruiz-Omeñaca et al., 1999) y en el Barremiense superior de Vallipón se
han descrito hasta diez morfotipos (Ruiz-Omeñaca et al., 1996; Canudo et al.,
1997). Los dromeosáuridos son una familia de terópodos de pequeño o mediano
tamaño que se caracteriza por el gran desarrollo de la segunda falange del pie,
formando una garra distintivamente curvada y mucho más larga que cualquier otra
falange (Ostrom, 1990). Independientemente del número de taxones válidos, estos
datos nos están indicando que los Maniraptoriformes de pequeño tamaño eran
diversos al final del Neocomiense en la Península Ibérica.
El dinosaurio más completo descrito es España es el Ornithomimosauria
Pelecanimimus polyodon Pérez-Moreno et al., 1994. Los ornitomimosaurios son un
grupo de terópodos característicos del Cretácico superior de Asia y Norteamérica.
Estos terópodos "imitadores de aves" se caracterizan por la reducción o ausencia de
dientes (Barsbold y Osmolska, 1990). El holotipo de Pelecanimimus procede del
yacimiento de Las Hoyas (Cuenca), que se encuentra en la formación La Huérguina,
datada con carofitas como Barremiense superior (Martín-Closas y Diéguez, 1998), y
consiste en la parte anterior de un individuo (incluido el cráneo) que conserva las
impresiones de los tejidos blandos (Briggs et al., 1997). Pelecanimimus se
caracteriza por la presencia de pequeños y numerosos dientes, alrededor de 220,
siendo los maxilares mayores que los mandibulares. Todos los dientes carecen de
dentículos marginales, pero presentan carenas mesial y distal, y tienen una
constricción entre la corona y la raíz. Los miembros posteriores son largos y con
dedos cortos, y la mano está modificada, ya que tiene tres dedos de tamaño similar
y subparalelos (Pérez-Moreno et al., 1994; Pérez-Moreno y Sanz, 1995, 1997). Este
terópodo tiene un gran interés filogenético al ser grupo hermano de los
Ornithomimidae del final del Cretácico inferior y del Cretácico superior de
Norteamérica y Asia (Holtz, 1998).
Taxón Material Yacimiento Formación Edad Referencia(Megalosaurus sp.)Dromaeosauridae indet.
Diente Benageber (V) ? Wealdiense Royo-Gómez 1926;Ruiz-Omeñaca y Pereda-Suberbiola 1999
(Megalosaurus cf. dunkeri)Theropoda indet.
Restos indet. Benageber (V) ? Wealdiense Royo-Gómez, 1927
Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
10
Theropoda indet.(Megalosaurus sp.)Theropoda indet.
Diente Mora de Rubielos (T) ? Wealdiense Lapparent, 1960, 1966
Theropoda indet. "Huesos" Alpuente (V.) ? Wealdiense Lapparent, 1966(Carcharodontosaurus)Theropoda indet. A
Diente Galve (T) ? ? Crusafont-Pairó yAdrover, 1966; Ruiz-Omeñaca et al., 1998b
("pequeño Carnosaurio")Theropoda indet.
dientes yvertebras
Yacimiento Herrero,G (T)
Camarillas Barremienseinferior
Crusafont-Pairó y Adrover,1966
(Megalosaurus)Theropoda indet.
? Galve (T) ? "Wealdiense" Crusafont y Gibert, 1976
Theropoda indet. Postcraneal Vadillos (Cu) El Collado? Barremiensesuperior? 2
Lapparent et al., 1969
(Coeluridae?)Dromaeosauridae indet.
Dientes Colladico Blanco, G(T)
Castellar Hauteriv. sup-Barrem. basal
Estes y Sanchiz, 1982
(Megalosauridae indet.)Theropoda indet. C
Diente El Piélago = Pantano2, G (T)
Castellar Hauteriv. sup-Barrem. basal
Buscalioni y Sanz, 1984Ruiz-Omeñaca et al., 1998
Theropoda indet. vértebrascaudales
Pelejón, G (T) Castellar Hauteriv. sup-Barrem. basal
Sanz et al., 1987
(Megalosauridae indet.)Theropoda indet.
Diente Las Hoyas (Cu) La Huérguina Barremiensesuperior 1
Buscalioni et al., 1988
Ornitomimosauria?Dromaeosauridae?
Falange Salas de los Infantes(B)
? Cretácico inferior Sanz et al., 1992
(Carnosauria)Theropoda indet.
Diente Benicatazara(V.)
El Collado Barremiensesuperior 2
Casanovas-Cladellas et al.,1993
Pelecanimimus polyodon Parte de unesqueleto
Las Hoyas (C) La Huérguina Barremiensesuperior 1
Perez-Moreno et al., 1994
"paronychodons" Dientes Poca, G (T) Camarillas Barremienseinferior
Zinke y Rauhut, 1994
Dromaeosauridae indet. Diente Pielago 0, G (T) Castellar Hauteriviensesuperior
Díez et al., 1995 Ruiz-Omeñaca et al., 1998b
Velociraptorinae indet.Dromaeosaurinae indet.
Dientes Uña (Cu) La Huérguina 3 Barremiensesuperior 2
Rauhut y Zinke, 1995;Rauhut, 2002a, 2002b
cf. Paronychodon sp. Dientes Uña (Cu) La Huérguina 3 Barremiensesuperior 2
Rauhut y Zinke, 1995;Rauhut, 2002a, 2002b
cf. Richardoestesia sp. Dientes Uña (Cu) La Huérguina 3 Barremiensesuperior 2
Rauhut y Zinke, 1995;Rauhut, 2002a, 2002b
Theropoda indet. 2 falangesunguales
Poyales BarrancoCanales, G (T)
Camarillas Barremienseinferior
Ruiz -Omeñaca, 1996
Theropoda indet. Dientes Vallipón, Castellote(T)
Artoles Barremiensesuperior
Ruiz -Omeñaca et al. 1996
Coelurosauria indet Dientes Vallipón, Castellote(T)
Artoles Barremiensesuperior
Ruiz -Omeñaca et al. 1996
Theropoda indet. ? Tenada de Los Lanos,PM (B)
? Berriasiense-Valanginiense
Torcida, 1996
Compsognátido indet. 16 restos Tenadas de Viajete,SI (B)
? Hauteriviense Torcida, 1996
Theropoda indet. Dientes La Cantalera, Josa(T)
Blesa Hauteriv. sup-Barrem. basal
Ruiz- Omeñaca et al.,1997a
Dromaeosauridae indet. Dientes Vallipón, Castellote(T)
Artoles Barremiensesuperior
Canudo et al., 1997
cf. Baryonyx Dientes Tenada de la Rosa,CS (B)
Facies Weald Berriasiense-Aptiense
Torcida et al., 1997
cf. Baryonyx Diente Tenada deCostalomo, SI (B)
Facies Weald Berriasiense-Aptiense
Torcida et al., 1997
cf. Baryonyx Diente La Tejera-Valdesancho, BM (B)
Facies Weald Berriasiense-Aptiense
Torcida et al., 1997
("Unserrated teeth")Coelurosauria indet.
dientes Camino Canales, G(T)
Castellar Hauteriv. sup-Barrem. basal
Ruiz-Omeñaca et al. 1998a
Dromaeosauridae indet. dientes Camino Canales, G(T)
Castellar Hauteriv. sup-Barrem. basal
Ruiz-Omeñaca et al. 1998a
"paronychodontid" diente Camino Canales, G(T)
Castellar Hauteriv. sup-Barrem. basal
Ruiz-Omeñaca et al. 1998a
Theropoda indet. B diente Cuesta Corrales, G(T)
Camarillas Barremienseinferior
Ruiz-Omeñaca et al. 1998a
Dromaeosauridae indet. diente La Maca 3, G (T) Camarillas Barremienseinferior
Ruiz-Omeñaca et al. 1998a
("Unserrated teeth")Coelurosauria indet.
diente Pantano 1, G (T) Castellar Hauteriv. sup-Barrem. basal
Ruiz-Omeñaca et al. 1998a
Dromaeosauridae indet. diente El Piélago = Pantano2, G (T)
Castellar Hauteriv. sup-Barrem. basal
Ruiz-Omeñaca et al. 1998a

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
11
Theropoda indet. B dientes Pajar Julian Paricio 2,G (T)
Camarillas Barremienseinferior
Ruiz-Omeñaca et al. 1998a
Dromaeosauridae indet. dientes Pajar Julian Paricio 2,G (T)
Camarillas Barremienseinferior
Ruiz-Omeñaca et al. 1998a
Baryonychidae indet. Dientes Vallipón, Castellote(T)
Artoles Barremiensesuperior
Ruiz -Omeñaca et al.,1998b
Baryonychidae indet. Dientes Poca, G (T) Camarillas Barremienseinferior
Ruiz-Omeñaca et al.,1998b
Baryonychidae indet. Diente La Cantalera, Josa(T)
Blesa Hauteriv. sup-Barrem. basal
Ruiz-Omeñaca et al.,1998b
Dromaeosauridae indet. Dientes La Cantalera, Josa(T)
Blesa Hauteriv. sup-Barrem. basal
Ruiz-Omeñaca y Canudo,2001
Baryonyx sp. cranealPostcraneal
El Juguete, SI (B) Pinilla de losMoros
Hauteriv. sup-Barrem. inf.
Fuentes-Vidarte et al.,2001
Tabla 3. Restos directos de Terópodos (excluyendo Avialae) en el Neocomiense de España.Abreviaturas: BM: Barbadillo del Mercado. CS: Cabezón de la Sierra. G: Galve. PM: Pinilla de losMoros. SI: Salas de los Infantes. B: Burgos. C: Castellón. Cu: Cuenca. T: Teruel. V: Valencia.Referencias en la bibliografía. Entre paréntesis están las determinaciones originales de los taxonesque se han modificado en este trabajo. 1: datación de Martín-Closas y Diéguez, 1998. 2: datación deGómez et al. (2001). 3: El yacimiento de Uña se encuentra según Rauhut (2000b) en la FormaciónUña, que debe ser considerada como sinónima de la Formación La Huérguina.
Los restos postcraneales aislados de Maniraptoriformes descritos son escasos.
Se puede citar una falange de Salas de los Infantes, que Sanz et al. (1992)
relacionan con Dromaeosauridae o con Ornithomimosauria. También se ha citado un
celurosaurio compsognátido en el Hauteriviense de Salas de los Infantes (yacimiento
Tenadas de Viajete; Torcida Fernández-Balbor, 1996), pero dada la singularidad de
este hallazgo, y puesto que los restos permanecen sin describir ni figurar, debe
tomarse como un taxón dudoso (?Compsognathidae en Tablas 3 y 7).
Se han descrito dientes de posibles "paronicodóntidos" en el Cretácico inferior,
asignados a cf. Euronychodon sp. y a "paronychodons". Se han encontrado en el
tránsito Hauteriviense -Barremiense de Cuesta de los Corrales (Galve, Teruel; Ruiz-
Omeñaca et al., 1998a), en el Barremiense inferior de Poca, también en Galve
(Zinke y Rauhut, 1994) y en el Barremiense superior de Uña (Cuenca; Rauhut y
Zinke, 1995; Rauhut, 2000b). Este grupo enigmático se caracteriza por tener dientes
ornamentados con surcos longitudinales. Los dientes de Uña, descritos como cf.
Paronychodon sp., carecen de dentículos en ambas carenas (Rauhut y Zinke, 1995),
mientras que los de Galve, descritos como "paronichodons", sí presentan dentículos
(Zinke y Rauhut, 1994; Ruiz-Omeñaca et al., 1998a). Algunos de los dientes de
Galve tienen un patrón de ornamentación más complicado, con surcos y crestas que
se anastomosan (Zinke y Rauhut, 1994). Los surcos y crestas longitudinales de los

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
12
dientes de Paronychodon lacustris, del Cretácico superior de Norteamérica, han sido
interpretados como consecuencia de crecimiento anormal (Currie et al., 1990).
Figura 4. Dientes de terópodo del Hauteriviense superior-Barremiense basal de La Cantalera(Josa, Teruel). A, B, G: Baryonychidae indet., A-B: vista labial/lingual, G: detalle de los dentículosdistales. C, D: Theropoda indet., vista labial/lingual. E, F, H: Dromaeosauridae indet., E-F: vistalabial/lingual, H: detalles de los dentículos distales. A-F: modificado de Ruiz-Omeñaca y Canudo,2002.

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
13
Sin embargo, otros autores apuntan que los "paronicodóntidos" son terópodos
relacionados con Maniraptoriformes primitivos, cercanos a los troodóntidos y
ornitomimosaurios (Rauhut y Zinke, 1995; Csiki y Grigorescu, 1998), o incluso que
podrían ser aves (Avialae según Rauhut, 2002a).
Otro pequeño terópodo representado en el Neocomiense de la Península Ibérica
es una forma relacionada con Richardodestesia, del Campaniense de Norteamérica.
Rauhut y Zinke (1995) describen un veintena de dientes que se caracterizan por
tener unos dentículos pequeños en relación con el tamaño del diente. Además
poseen un surco longitudinal que desaparece hacia la punta del diente. La
combinación de estos caracteres solo se ha descrito en los dientes identificados en
el género americano, y por esa razón Rauhut y Zinke (1995) los asignan a cf.
Richardoestesia sp., apuntando que la referencia al género es orientativa.
Aunque se han citado troodóntidos en el Barremiense superior de Vallipón y Uña
(Canudo et al., 1996, Rauhut, 1996) haciendo referencia a dientes con dentículos
inclinados, éstos se han asignado correctamente a dromeosáuridos velocirraptorinos
(Ruiz-Omeñaca et al., 1996; Rauhut, 2002b). De igual modo, los dientes descritos
por Estes y Sanchiz (1982) en el Hauteriviense Superior-Barremiense basal de
Galve como Coeluridae? (conservados en el Institut de Paleontologia "Miquel
Crusafont" de Sabadell) que han sido asignados a Troodontidae indet por Rauhut
(2000), se determinan como dromeosáuridos.
Los grandes terópodos están presentes en el Neocomiense (y en el resto del
Cretácico) de la Península Ibérica. Hasta la década de los 90 se utilizaron términos
como Megalosauridae o Megalosaurus para identificar a estos dinosaurios, (véanse
las citas de Royo y Gómez en las Tablas 3 y 4). Desde que Buckland (1824)
describió el terópodo Megalosaurus, la identificación de los grandes terópodos como
megalosáuridos ha sido bastante común, y de hecho los tetanuros basales han sido
informalmente clasificados como "megalosáuridos" (Gauthier, 1986). Sin embargo,
se ha demostrado que los megalosáuridos considerados de esta manera tan amplia
carecen de caracteres diagnósticos (Holtz, 1994; Sereno, 1997). El taxón
Megalosaurus representa a un grupo de Tetanurae basal con presencia únicamente
en el Jurásico medio (nodo H de Holtz, 1998). Por esta razón se ha preferido

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
14
cambiar la asignación de los restos del Cretácico de España identificados como
megalosáuridos a Theropoda indet. (Tabla 3), hasta que revisiones del material
permitan clarificar su filogenia.
La mayoría de los restos de grandes terópodos neocomienses son fragmentarios,
fundamentalmente dientes. Crusafont-Pairó y Adrover (1966) identifican un diente
aislado de gran tamaño del Cretácico inferior de Galve (yacimiento desconocido)
como "Carcharodontosaurus". Esta determinación parece poco adecuada: como han
puesto de manifiesto Ruiz-Omeñaca et al. (1997), el diente de Galve, conservado en
el Institut de Paleontologia "Miquel Crusafont" de Sabadell, no tiene pliegues
oblicuos a ambos lados de los márgenes (Santafé-Llopis y Casanovas-Cladellas,
1993: fig. 40), carácter diagnóstico de Carcharodontosaurus del Albiense-
Cenomaniense del Norte de África (Molnar et al., 1990; Sereno et al., 1996), por lo
que pertenecería a otro género de gran talla. También en Galve, Buscalioni y Sanz
(1984) describen un diente grande de la Formación Castellar (tránsito Hauteriviense-
Barremiense) que incluyen en Megalosauridae, pero anotando las dificultades de
esta familia que, como hemos visto anteriormente, estaba formada por taxones con
diferentes historias evolutivas. Ruiz-Omeñaca et al. (1998a) asignan los dientes de
Crusafont-Pairó y Adrover y Buscalioni y Sanz a dos tipos distintos de Theropoda
indet y aún reconocen un tercer tipo de Theropoda indet. en Galve, representado por
4 dientes de las Formaciones Castellar y Camarillas.
En Benicatazara (Aras de Alpuente, Valencia), Casanovas-Cladellas (1992) y
Casanovas-Cladellas et al., (1993) citan la parte superior de un diente que atribuyen
a un carnosaurio (Carnosauria indet.). Tiene desarrollo de dentículos tanto en el
margen distal como mesial, estando los posteriores ligeramente más desarrollados.
Dado lo fragmentario del fósil, parece mejor considerarlo como Theropoda indet.
Entre los restos postcraneales, Lapparent et al. (1969) citan dos vértebras dorsales,
una vértebra caudal y un tibia del yacimiento barremiense de Vadillos (Cuenca) que
asignan a un "Théropode carnivore". Dado que no se describe el material, parece
correcto considerarlo como Theropoda indet. Sanz et al. (1987) describen tres
centros vertebrales de Theropoda indet. en Pelejón (Galve, Formación Castellar,
tránsito Hauteriviense-Barremiense), que relacionan con los "megalosáuridos"

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
15
aunque, dada la ausencia de caracteres diagnósticos, prefieren considerar como
Theropoda indet.
La posición sistemática de estos supuestos megalosáuridos y otros grandes
terópodos del Cretácico inferior de la Península Ibérica es incierta, pero podrían
pertenecer a Carnosauria (en el sentido de Holtz, 1998). Este grupo tiene
representantes en Gondwana y Laurasia en el Neocomiense, entre los cuales están
algunos de los mayores terópodos conocidos (Coria y Salgado, 1995; Sereno et al.,
1996; Harris, 1998a, 1998b).
Taxón Material Yacimiento Formación Edad Referencia(Iguanodon)Theropoda indet.
Tibia Utrillas (T) Escucha Albiense Vilanova, 1873;Royo-Gómez, 1927
(Megalosaurus)Theropoda indet.
fémur Morella (C) Morella? Aptiense inf.? Royo-Gómez, 1925;Ruiz-Omeñaca y Pereda-Suberbiola, 1999
(Megalosaurus bucklandi)Theropoda indet.
Dien te ypostcraneal
Morella (C) Morella? Aptiense inf.? Royo-Gómez , 1921 ,1927a, 1927b
(Coeluridae indet.)Theropoda indet
D i e n t e /vértebra
Mas RomeuMorella (C)
Morella Aptiense inf. Santafé et al., 1982
(Megalosauridae indet.)Theropoda indet.
ungual El BeltránMorella (C)
Morella Aptiense inf. Santafé et al., 1978; 1982
(Megalosauridae indet.)?Carcharodontosauridae
Diente El BeltránMorella (C)
Morella Aptiense inf. Santafé et al., 1978; 1982
(Baryonyx walkeri)cf. Baryonyx sp.
maxilar sindientes
Peña CarcenaIgea (LR)
Grupo Enciso Barremiensesup.-Aptiense
Viera y Torres, 1995
Baryonychidae indet. Dientes Morella (C) Morella Aptiense inf. Ruiz-Omeñaca et al.,1998b
Tabla 4. Restos directos de Terópodos (excluyendo Avialae) en el Aptiense-Albiense deEspaña. Abreviaturas: C. Castellón. LR: La Rioja. T. Teruel. Referencias en la bibliografía. Entreparéntesis están las determinaciones originales de los taxones que se han modificado en este trabajo.
TEROPODOS DEL APTIENSE - ALBIENSE (Tabla 4)
Los terópodos del Aptiense-Albiense se han encontrado en Castellón, La Rioja y
Teruel. Al menos están representados un barionícido, un posible dromaeosáurido,
un ?carcharodontosáurido y un terópodo indeterminado. En las Capas Rojas de
Morella (Aptiense inferior) se han citado dientes y restos postcraneales identificados
como un pequeño Coeluridae indet. y un gran Megalosauridae indet. (Santafé-Llopis
et al., 1979; Santafé et al., 1982). El pequeño diente de Coeluridae, del yacimiento
de Mas Romeu ("Carnosauria ind." en Santafé et al., 1979) tiene una morfología
general cercana a Dromaeosauridae (Santafé et al., 1982), lo que podría indicar la
presencia de esta familia en el Aptiense, pero por el momento se determina como

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
16
Theropoda indet. Santafé et al. (1982) también adscribieron a Coeluridae indet. un
fragmento de centro vertebral del mismo yacimiento que el diente, que también se
determina aquí como Theropoda indet.
El diente grande, procedente del yacimiento El Beltrán está clasificado como
Megalosauridae indet., pero presenta una morfología peculiar (Santafé et al., 1982).
Es un diente premaxilar o maxilar anterior con unas crenulaciones muy finas que
además posee "pliegues oblicuos sobre las crestas" similares a los de
Carcharodontosaurus saharicus. Estos pliegues en el esmalte, además de
Carcharodontosaurus del Albiense-Cenomaniense de África (Sereno et al., 1996),
los presenta otro gran terópodo del Cretácico inferior, Giganotosaurus del Albiense-
Cenomaniense de Sudamérica (Coria y Salgado, 1995). Los últimos estudios
cladísticos sitúan a ambos en un mismo nodo sin nombrar (nodo S de Holtz, 1998,
que incluye además a Acrocanthosaurus, del Aptiense-Albiense de Norteamérica,
que no presenta pliegues en los dientes). Sereno et al. (1996) consideran a estos
tres taxones (Carcharodontosaurus, Giganotosaurus y Acrocanthosaurus) miembros
de la familia Carcharodontosauridae, aunque Currie y Carpenter (2000) consideran
que Acrocanthosaurus es un Allosauridae. Dientes con pliegues oblicuos en el
esmalte se han encontrado también en el Aptiense de Argentina (Vickers-Rich et al.,
1999), el Cenomaniense-Turoniense de Japón (Chure et al., 1999) y el Cretácico
superior de Brasil (Kellner y Campos, 2000). La figuración original de Santafé et al.
(1982) dificulta realizar comparaciones con estos carnosaurios, pero la presencia de
los pliegues oblicuos en el diente de Morella indica la presencia de un carnosaurio
cercano a los Carcharodontosauridae, lo que supone la primera evidencia en
Europa. La falange ungual de El Beltrán, descrita por Santafé et al. (1982) como
Megalosauridae indet., sólo puede determinarse como Theropoda indet.
De los mismos niveles estratigráficos (Formación Morella) provienen los restos
identificados por Royo y Gómez como Megalosaurus en el "Weald" de Morella
(Tabla 4). El más destacable es un gran fémur expuesto en la exposición
permanente del Museo Nacional de Ciencias Naturales de Madrid, que necesita de
una revisión para clarificar su posición taxonómica (Ruiz-Omeñaca y Pereda-
Suberbiola, 1999).

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
17
En Igea (La Rioja) se ha citado un fragmento de maxilar sin dientes, que se ha
atribuido a Baryonyx walkeri (Viera y Torres, 1995; Charig y Milner, 1997). Los
restos provienen del grupo Enciso, datado con carofitas como Barremiense -
Aptiense (Martín-Closas y Alonso-Millán, 1998). Dado lo fragmentario del material, y
puesto que hay otros barionícidos descritos en el Aptiense (Angaturama limai Kellner
y Campos,1996 e Irritator challengeri Martill et al., 1996 en Brasil; y Cristatusaurus
lapparenti Taquet y Russell, 1998 y Suchomimus tenerensis Sereno et al., 1998 en
Níger) es más prudente considerar este resto como cf. Baryonyx sp.
El único resto directo conocido en España de un terópodo albiense proviene de
Utrillas (Teruel). Tiene un notable interés histórico, ya que fue uno de los primeros
restos de dinosaurios identificados como tales en España (Vilanova y Piera, 1873).
Este autor cita dos huesos largos que clasifica como Iguanodon, aunque no los
describe, ni figura. A principios de siglo Royo y Gómez (1920, 1921) los cita
manteniendo su asignación a Iguanodon. Posteriormente Royo y Gómez (1926) los
menciona como "dos fragmentos de un fémur de un dinosaurio pequeño que por
error (Vilanova) determinó como de Iguanodon", y sin descartar que pertenezcan a
un ornitópodo los asigna "a una forma pequeña y quizás nueva". Definitivamente
Royo y Gómez (1927) asigna los restos a un terópodo: "Tengo además algunas
piezas de vertebrados de diferentes yacimientos, entre los cuales citaría una tibia de
terópodo de Utrillas (Teruel) de aproximadamente 25 cm de longitud, que puede
mostrar la existencia en aquellos tiempos de una forma de tamaño muy pequeño"
(Ruiz-Omeñaca y Pereda-Suberbiola, 1999).
Aunque no lo dice explícitamente, se puede asumir que es el mismo resto de
Vilanova y Piera y, por tanto, los dos huesos largos eran en realidad el extremo
proximal y distal de dicha tibia. En este momento únicamente quedan dibujos de
esta pieza en el Museo Nacional de Ciencias Naturales, desconociéndose donde se
encuentra este fósil de gran interés histórico. Dado que la tibia se recogió en los
lignitos de Utrillas (Vilanova, 1873), hoy incluidos en Formación Escucha, se puede
datar como Albiense inferior-medio (Querol et al., 1992).

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
18
Taxón Material Yacimiento Formación Edad Referencia(Megalosauridae indet.)Theropoda indet.
Radio Vilamitjana 1 =Els Nerets, (L)
Tremp Campaniense sup. 1 Casanovas-Cladellas et al.,1988
Teropoda indet. Dientes Armuña (S) "Garumniense" Campaniense sup -Maastricthiense inf.
Sanz et al., 1992
Teropoda indet. Ungual Armuña (S) "Garumniense" Campaniense sup -Maastricthiense inf.
Buscalioni, 2000
Theropoda indet. Dientes Chera (V) SierraPerenchiza
Campaniense sup -Maastricthiense inf.
Company et al., 1999
Neoceratosauria indet. Fémures L1A y L1 B,Laño (B)
Vitoria Campaniense sup. 2 Pereda-Suberbiola, 1999
Dromaeosauridae indet. Dientes L1A, Laño (B) Vitoria Campaniense sup. 2 Astibia et al., 1999cf. Euronychodon sp. Dientes L1A, Laño (B) Vitoria Campaniense sup. 2 Astibia et al., 1999Coelurosauria indet. Dientes L1A, Laño (B) Vitoria Campaniense sup. 2 Astibia et al., 1999?Ornithomimosauria ? L1A, Laño (B) Vitoria Campaniense sup. 2 Pereda-Suberbiola et al., 2000Richardoestesia-like Diente L' Abeller (L) Arén Campaniense sup. Prieto-Márquez et al. 2000Theropoda indet. Diente Montrebei (L) Tremp Campaniense final 3 Torices Hernández, en prensaCoelurosauria indet. Dientes Montrebei (L) Tremp Campaniense final 3 Torices Hernández, en prensaDromaeosauridae indet. Dientes Montrebei (L) Tremp Campaniense final 3 Torices Hernández, en prensaCoelurosauria indet. Diente Vicari 4 (L) Tremp Campaniense final 3 Torices Hernández, en prensaDromaeosauridae indet. Dientes Vicari 4 (L) Tremp Campaniense final 3 Torices Hernández, en prensa
Tabla 5. Restos directos de Terópodos (excluyendo Avialae) en el Campaniense y Campano-maastrichtiense de España. Abreviaturas: B: Burgos (condado de Treviño, Alava). L: Lérida. S:Segovia. V: Valencia. Referencias en la bibliografía. Entre paréntesis están las determinacionesoriginales de los taxones que se han modificado en este trabajo. 1: datación de López-Martínez et al.(2001), 2: datación de Núñez-Betelu (1999), 3: datación de Vicens et al. (en prensa). Vilamitjana 1 y ElNerets son el mismo yacimiento.
TERÓPODOS DEL CAMPANIENSE (Tabla 5, Figura 5)
En el Cretácico superior de España se han encontrado restos de terópodos en el
Campaniense y el Maastrichtiense. Tradicionalmente se han asignado los
yacimientos de dinosaurios del Cretácico superior español al Maastrichtiense, sin
embargo cuando se dispone de dataciones más precisas se observa que algunos de
ellos son del Campaniense (véase discusión en López-Martínez et al., 2001). En
otros casos no se han realizado todavía estudios bioestratigráficos que permitan
situarlos en el Campaniense o al Maastrichtiense. En este caso se han incluido en el
Campaniense a los yacimientos con restos del ornitópodo Rhabdodon ,
considerándose como fauna "Campaniense", aunque este dinosaurio puede
encontrarse hasta el Maastrichtiense inferior, y por eso en la Tabla 5 aparecen con
esta edad. Considerando los yacimientos, de esta manera, se han encontrado
terópodos del Campaniense en Burgos (Condado de Treviño), Lérida, Segovia y
Valencia. Recientemente se han puesto al día las faunas de dinosaurios del
Cretácico superior de la Península Ibérica (Pereda-Suberbiola, 1999); a partir de
este trabajo, y los nuevos descubrimientos se conoce que en el Campaniense hay

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
19
una amplia diversidad de terópodos compuesta por Neoceratosauria indet. y
Maniraptoriformes, entre los que hay dromeosáuridos. También están presentes
formas con afinidades inciertas, como los “paronicodóntidos” y cf. Richardoestesia.
Figura 5. Terópodos del Campaniense y Campano-maastrichtiense de España. A-C: Fémurizquierdo de Neoceratosauria indet., Laño (condado de Treviño, Burgos, Álava), en vistas anterior(A), lateral (B) y medial (C). D-F: radio izquierdo de Theropoda indet., Els Nerets (Vilamitjana,Lérida), en vistas posterior (D), proximal (E) y distal (F). G-I: diente lateral de "Richardoestesia-like",L´Abeller (Lérida), en vistas lateral con detalle de los dentículos (G), distal (H) y basal (I). J: dientelateral de Dromaeosauridae indet. (VIR-4-T7), Vicari-4 (Lérida), en vista labial/lingual. K: dientelateral de Coelurosauria indet. (MON-T6), Montrebei (Lérida), en vista lingual.A-C: tomados de Le Loeuff, 1992 (como cf. Tarascosaurus sp.); D-F: tomados de Casanovas-Cladellas et al., 1988 (como Megalosauridae indet.): G-I: tomados de Prieto-Márquez et al., 2000. J-K:tomado de Torices Hernández (en prensa).

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
20
El yacimiento más importante del Campaniense de la Península Ibérica es del
Laño (condado de Treviño, Burgos, Álava). Un equipo de la Universidad del País
Vasco ha realizado varias campañas de excavación en la que han recuperado una
diversa fauna de vertebrados (véase la lista faunística en Astibia et al., 1999). Este
yacimiento está datado con polen como Campaniense superior (Núñez-Betelu,
1999). Los terópodos de Laño están representados fundamentalmente por dientes
aislados y algunos restos postcraneales. Por el momento no se han descrito en
profundidad, pero los estudios preliminares indican una diversidad destacable.
Los neoceratosaurios están representados por dos fémures determinados como
Abelisauridae? (Astibia et al., 1990, 1999) y cf. Tarascosaurus sp. (Le Loeuff y
Buffetaut, 1991; Le Loeuff, 1992), un género descrito en el Cretácico superior de
Francia. Los abelisáuridos son un grupo característico de Gondwana, y su presencia
en Laño y en otros yacimientos europeos (Buffetaut et al., 1988; Le Loeuff y
Buffetaut, 1991) ha sido considerada como una prueba de un puente intercontinental
entre Gondwana y Laurasia en el Cretácico superior, que permitió un intercambio de
faunas de dinosaurios (Astibia et al., 1990; Le Loeuff, 1997). Sin embargo hay
autores que dudan de la presencia de abelisáuridos en Europa (Sampson et al.,
1998; Lamanna et al., 2002). Dada la ausencia de material diagnóstico en Laño,
Pereda-Suberbiola (1999) y Pereda-Suberbiola et al. (2000) prefieren atribuirlo de
manera provisional a Neoceratosauria indet. (en el sentido de Novas, 1992).
Los terópodos más diversos en Laño son los Tetanurae, entre los que se han
reconocido, mediante dientes aislados, Dromaeosauridae indet. y Coelurosauria
indet., y el terópodo de afinidades inciertas cf. Euronychodon sp. (Le Loeuff, 1988;
Astibia et al., 1999). Además, Pereda-Suberbiola et al. (2000) mencionan un
ornitomimosaurio en Laño (citado como Bullatosauria (Troodontidae +
Ornithomimosauria) en Pereda-Suberbiola, 1999), que por el momento no ha sido
descrito, por lo que en la Tabla 5 aparece como ?Ornithomimosauria.
En los Pirineos, Casanovas-Cladellas et al. (1988) describen un radio de El Nerets
(=Vilatmitjana 1), que consideran tiene afinidades con Allosauridae, pudiendo incluso
pertenecer a esta familia, pero que asignan a Megalosauridae indet. Hasta que no se
revise el material, es más correcto referir este radio a Theropoda indet. En L' Abeller

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
21
(Campaniense superior, Formación Arén) se ha descrito un diente con sección basal
subcuadrada y dentículos posteriores pequeños y numerosos (Prieto-Márquez et al.,
2000). Estos autores lo relacionan con Richardoestesia, lo que supone la primera
cita de este terópodo en el Cretácico superior de Europa. Otros dientes de terópodo
del Campaniense final (Formación Tremp) de los Pirineos de Lérida se han asignado
a Theropoda indet., Coelurosauria indet y Dromaeosauridae indet. (Torices
Hernández, en prensa).
TEROPODOS DEL MAASTRICHTIENSE (Tabla 6, Figura 6)
Los terópodos maastrichtienses están exclusivamente representados por dientes
aislados, que han aparecido en yacimientos de Burgos, Huesca y Lérida. Los más
interesantes son los del Maastrichtiense superior de la Formación Tremp en los
Pirineos de Huesca (yacimientos de Blasi, en Arén). Su importancia radica en que
son los terópodos más modernos que se conocen en Europa, un poco antes del
evento del límite Cretácico- Terciario (López-Martínez et. al., 2001). Los
dromeosáuridos son el grupo de terópodos más común, y también están
representados los "paronicodóntidos" y celurosaurios indeterminados. Hasta el
momento no hay evidencias de otros grupos de terópodos.
Dada la variedad de morfologías en los dientes descritos, los dromeosáuridos del
Maastrichtiense deben estar representados por varias especies (Pol et al., 1992;
López-Martínez et al., 2001; Canudo et al., 2001; Torices Hernández, 2002, en
prensa). Su principal característica es que los dentículos de la carena mesial,
cuando los hay, son siempre de menor tamaño que los de la carena distal. En Blasi
(Arén, Huesca) pueden estar representadas las dos subfamilias de dromeosáuridos
conocidas: velocirraptorinos y dromeosaurinos (López-Martínez et al., 2001). En
Blasi 3 hay un diente de dromaeosáurido de tamaño relativamente grande, que
conserva dentículos mesiales, y tiene los dentículos distales inclinados, por lo que
podría incluirse en Velociraptorinae (en sentido de Rauhut y Werner, 1995). En Blasi
2 hay dientes de otro dromaeosáurido más pequeño, que únicamente tiene

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
22
dentículos distales y éstos no están inclinados, lo que los acerca a Dromaeosaurinae
(en sentido de Currie et al., 1990).
Figura 6. Terópodos del Maastrichtiense de España. A: diente lateral de Dromaeosauridae indet.,Quintanilla del Coco (Burgos), en vista labial/lingual. B: diente lateral de cf. Euronychodon sp.,Quintanilla del Coco (Burgos), en vista labial/lingual. C: diente premaxilar de Dromaeosauridaeindet. (FONT-6-T1), Fontllonga 6 (Lérida), en vista lingual. C: diente lateral de Dromaeosauridaeindet. (FONT-6-T2), Fontllonga 6 (Lérida), en vista labial/lingual. E: diente lateral deDromaeosauridae indet. (MPZ98/72), Blasi-2 (Arén, Huesca), en vista labial/lingual con detalle delos dentículos distales. F-G: diente lateral de cf. Euronychodon sp. (MPZ98/76-77), Blasi-2 (Arén,Huesca), en vista lingual. H: diente lateral de Coelurosauria indet. (MPZ98/82), Blasi-2 (Arén,Huesca), en vista labial/lingual. A-B: tomados de Pol et al., 1992 (como ?Dromaeosauridae y cf.Paronychodon sp.); C-D: tomados de Torices Hernández (en prensa); E-H: tomados de López-Martínez et al., 2001.

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
23
En Quintanilla del Coco (Burgos) y Blasi 2 se han descrito unos pequeños dientes
de "paronicodóntidos", con bordes afilados pero sin dentículos, y con amplios surcos
(Pol et al., 1992; López-Martínez et al., 2001), morfológicamente parecidos a
Euronychodon del Cretácico superior de Portugal (Antunes y Sigogneau-Russell,
1991), un género posiblemente relacionado con Paronychodon, un enigmático
terópodo del Cretácico superior de Norteamérica (véase la discusión en los
terópodos del Neocomiense). Los dientes de Quintanilla del Coco, descritos como cf.
Paronychodon sp. (Pol et al., 1992), representan seguramente el mismo taxón que
los de Blasi 2 (cf. Euronychodon sp.) y se asignan en este trabajo a cf.
Euronychodon sp.
Taxón Material Yacimiento Formación Edad Referencia(cf. Paronychodon sp.)cf. Euronychodon sp.
Dientes Quintanilla delCoco (B)
Cal izas deLychnus
Maastricthiense Pol et al., 1992
¿Dromaeosauridae? Dientes Quintanilla delCoco (B)
Cal izas deLychnus
Maastricthiense Pol et al., 1992
Theropoda indet. ? Fontllonga-6(L)
Tremp Maastrichtiense inf. 1 Álvarez-Sierra et al., 1994
Theropoda indet. Dientes Blasi 2 y 3Arén (H)
Tremp Maastrichtiense sup. López-Martínez et al., 2001
Dromaeosauridae indet.-Velociraptorinae?
Diente Blasi 3Arén (H)
Tremp Maastrichtiense sup. López-Martínez et al., 2001
Dromaeosauridae indet.-Dromaeosaurinae?
Dientes Blasi 2Arén (H)
Tremp Maastrichtiense sup. López-Martínez et al., 2001
cf. Euronychodon sp. Dientes Blasi 2Arén (H)
Tremp Maastrichtiense sup. López-Martínez et al., 2001
Coelurosauria indet. Dientes Blasi 2Arén (H)
Tremp Maastrichtiense sup. López-Martínez et al., 2001
Dromaeosauridae indet. Dientes Fontllonga 6(L)
Tremp Maastrichtiense inf. 1 Torices Hernández, en prensa
Dromaeosauridae indet. Diente Figuerola-2,Figuerola deMeià (L)
Tremp Maastrichtiense inf. 2 Torices Hernández, 2002
Tabla 6. Restos directos de Terópodos (excluyendo Avialae) en el Maastrichtiense de España.Abreviaturas: B: Burgos. H: Huesca. L: Lérida. S: Segovia. Referencias en la bibliografía. Entreparéntesis están las determinaciones originales de los taxones que se han modificado en este trabajo.1: datación de López-Martínez et al. (1998), 2: datación de Vicens et al. (en prensa). La edad deArmuña es indeterminada, y podría ser también Campaniense.
También en Blasi 2 se ha encontrado varios dientes sin dentículos que se han
identificado como Coelurosauria indet. Podrían pertenecer a alguno de los grupos
anteriores, ya que se desconoce la ontogenia de estos terópodos, o bien ser un
taxón distinto (López-Martínez et al., 2001).

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
24
IMPLICACIONES PALEOBIOGEOGRÁFICAS
Los taxones del Jurásico, muy escasos, no permiten hacer interpretaciones de
tipo paleobiogeográfico, pero al menos pueden indicar la presencia de los dos
grandes clados de terópodos, ceratosaurios y tetanuros (Martínez et al., 2000b;
Barco y Ruiz-Omeñaca, 2001a), que ya estaban diferenciados como tales en el
Triásico Superior (Sereno et al., 1997).
La mayoría de los restos de dinosaurios del Cretácico se pueden incluir en
Tetanurae, y con excepción de los barionícidos (Ruiz-Omeñaca et al., 1998b) y el
posible "carcharodontosáurido" del Cretácico Inferior (Santafé et al., 1982) son todos
Maniraptoriformes. Esto significa que están representados clados modernos de
origen laurasiático. Los barionícidos y "carcharodontosáuridos", según las filogenias
actuales, tienen su origen en el Jurásico, y por tanto anterior a la ruptura de la
Pangea (Holtz, 1998).
El único taxón no laurasiático es el Neoceratosauria del Campaniense superior de
Laño (Pereda-Suberbiola et al., 2000), incluido en Abelisauridae (cf. Tarascosaurus
sp.) por Le Loeuff y Buffetaut (1991). La presencia de abelisáuridos en Europa (Laño
y sur de Francia), una familia característica de Gondwana, puede explicarse
únicamente por una unión intercontinental a lo largo del Cretácico Superior entre
Gondwana y Laurasia, como proponen Buffetaut et al. (1988) y Le Loeuff y Buffetaut
(1991), aunque otra posibilidad es que Tarascosaurus no sea un abelisáurido
inmigrante sino un superviviente de un grupo de neoceratosaurios laurasiático. Esta
segunda hipótesis se ve reforzada por los recientes trabajos de Sampson et al.
(1998) y Lamanna et al. (2002), quienes opinan que no hay abelisáuridos fuera de
Gondwana.
Jur.Sup.
Jur.-Cret.
Cret. Inf.pre-Apt.
Apt-Alb
Camp-Maast
Maast. sup.
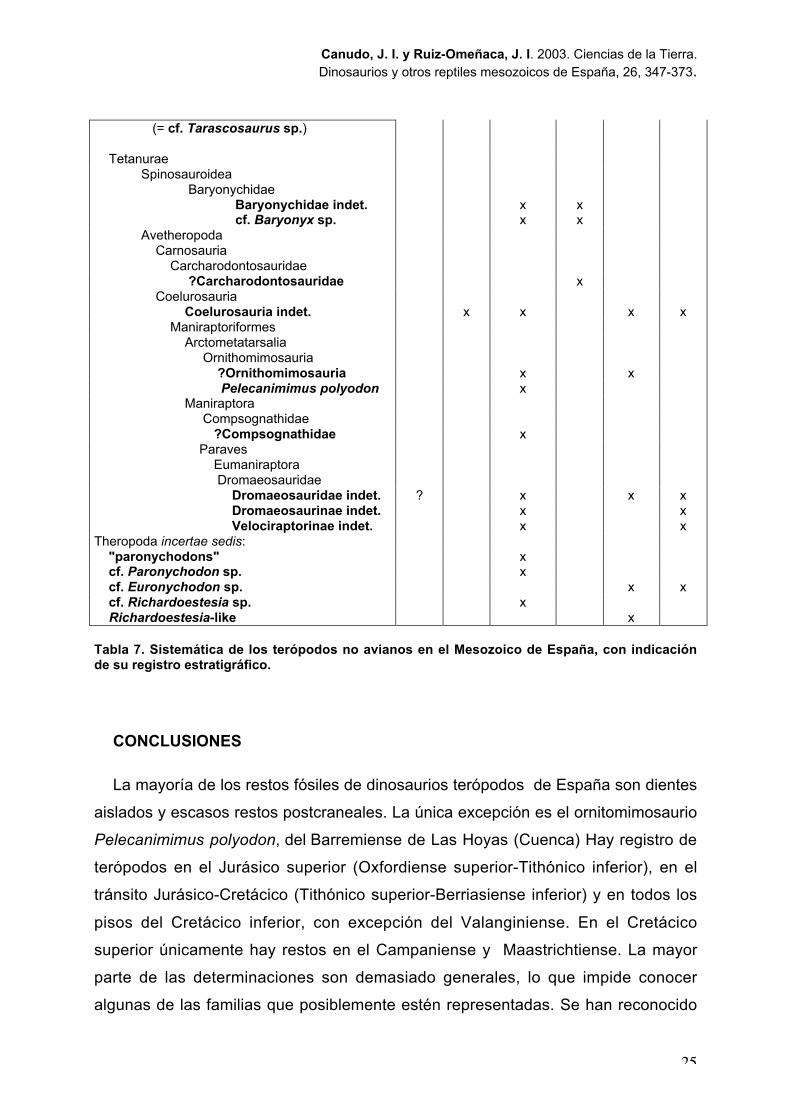
Theropoda Theropoda indet. x x x x x x Ceratosauria
?Ceratosauria xNeoceratosauria
Neoceratosauria indet. x
Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
25
(= cf. Tarascosaurus sp.)
TetanuraeSpinosauroidea
BaryonychidaeBaryonychidae indet. x xcf. Baryonyx sp. x x
Avetheropoda Carnosauria Carcharodontosauridae
?Carcharodontosauridae x Coelurosauria Coelurosauria indet. x x x x Maniraptoriformes Arctometatarsalia
Ornithomimosauria ?Ornithomimosauria x x Pelecanimimus polyodon x
Maniraptora Compsognathidae
?Compsognathidae x Paraves Eumaniraptora
Dromaeosauridae Dromaeosauridae indet. ? x x x Dromaeosaurinae indet. x x Velociraptorinae indet. x x
Theropoda incertae sedis: "paronychodons" x cf. Paronychodon sp. x cf. Euronychodon sp. x x cf. Richardoestesia sp. x Richardoestesia-like x
Tabla 7. Sistemática de los terópodos no avianos en el Mesozoico de España, con indicaciónde su registro estratigráfico.
CONCLUSIONES
La mayoría de los restos fósiles de dinosaurios terópodos de España son dientes
aislados y escasos restos postcraneales. La única excepción es el ornitomimosaurio
Pelecanimimus polyodon, del Barremiense de Las Hoyas (Cuenca) Hay registro de
terópodos en el Jurásico superior (Oxfordiense superior-Tithónico inferior), en el
tránsito Jurásico-Cretácico (Tithónico superior-Berriasiense inferior) y en todos los
pisos del Cretácico inferior, con excepción del Valanginiense. En el Cretácico
superior únicamente hay restos en el Campaniense y Maastrichtiense. La mayor
parte de las determinaciones son demasiado generales, lo que impide conocer
algunas de las familias que posiblemente estén representadas. Se han reconocido

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
26
(Tabla 7): Neoceratosauria, Baryonychidae, Ornithomimosauria, Dromaeosauridae,
además de terópodos indeterminados, y celurosaurios indeterminados (dientes
pequeños sin dentículos). La mayoría de los restos son de Maniraptoriformes, siendo
especialmente abundantes los dromeosáuridos. Las únicas excepciones son por el
momento, el posible Ceratosauria del Jurásico superior de Asturias, los barionícidos
del Hauteriviense-Barremiense de Burgos, Teruel y La Rioja, el posible
carcharodontosáurido del Aptiense inferior de Morella y los posibles abelisáuridos
del Campaniense de Laño. Además hay algunos terópodos incertae sedis, como los
"paronicodóntidos" (entre los que se incluye Euronychodon) y Richardoestesia.
AGRADECIMIENTOS
En primer lugar queremos agradecer al Doctor Félix Pérez-Lorente por su amable invitación parapresentar la ponencia Terópodos de España en el "I Congreso Internacional sobre dinosaurios y otrosreptiles mesozoicos de España". Este trabajo se enmarca en el proyecto VECOBA (VertebradosContinentales del Barremiense) subvencionado por el Ministerio de Ciencia y Tecnología (BTE 2002-1746), la Fundación Conjunto Paleontológico de Teruel y el Departamento de Cultura y Turismo de laDiputación General de Aragón. Un parte importante de los fondos bibliográficos del equipo deDinosaurios de la Universidad de Zaragoza se debe a la donación del Profesor René Lavocat.
REFERENCIAS
Álvarez-Sierra, M. A., Arribas, M. E., Ardèvol, L., Civis, J., Daams, R., Krauss, S., López-Martínez, N.,de la Peña, A., Soler, R., Vianey-Liaud, M., Lacasa, A., Marandat, B., Peláez-Campomanes, P.,Sevilla, P., Sigé, B. 1994. El limite Cretácico-Terciario en la sección de Fontllonga (Cuenca deAger, provincia de Lérida)- II Congreso del Grupo Español de Terciario. Jaca, Comunicaciones,23-26.
Antunes, M. T. y Sigogneau-Russell, D. 1991. Nouvelles données sur les Dinosaures du Crétacésupérieur du Portugal. C.R. Acad. Sci. Paris, 313, II, 113-119.
Astibia, H., Buffetaut, E., Buscalioni, A.D., Cappetta, H., Corral, C., Estes, R., Garcia-Garmilla, F.,Jaeger, J.J., Jimenez-Fuentes, E., Le Loeuff, J. Le, Mazin, J.M. Orue-Etxebarria, X., Pereda-Suberbiola, J., Powell, J.E., Rage, J.C., Rodríguez-Lazaro, J., Sanz, J.L. y Tong, H. 1990. Thefossil vertebrates from Laño (Basque Country, Spain); new evidence on the composition andaffinities of the Late Cretaceous continental faunas of Europe. Terra Nova, 2, 460-466.
Astibia, H., Murelaga, X., Pereda-Suberbiola, X., Elorza, J.J. y Gomez-Alday, J.J. 1999. Taphonomyand palaeoecology of the Upper Cretaceous continental vertebrate-bearing beds of the LañoQuarry (Iberian Peninsula). Est. Mus. Cienc. Nat. de Alava, 14 (Núm. Espec. 1), 43-104.
Barco, J.L. y Ruiz-Omeñaca, J.I. 2001a. Primeros dientes de terópodo (Dinosauria, Saurischia) en laFormación Villar del Arzobispo (Tithónico-Berriasiense): yacimientos Cuesta Lonsal y LasCerradicas 2 (Galve, Teruel). En: XVII Jornadas de la Sociedad Española de Paleontología. Losfósiles y la Paleogeografía. Albarracín, 1, 239-246.

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
27
Barco, J.L. y Ruiz-Omeñaca, J.I. 2001b. Primeros restos postcraneales de terópodo (Dinosauria,Saurischia) en la Formación Villar del Arzobispo (Tithónico-Berriasiense): Un centro vertebralcaudal del yacimiento Carretera (Galve, Teruel). En: XVII Jornadas de la Sociedad Española dePaleontología. Los fósiles y la Paleogeografía. Albarracín, 1, 247-254.
Barsbold, R. y Osmólska, H. 1990. Ornithomimosauria. In Dinosauria, Ed. D.B. Weishampel, P.Dodson, H. Osmólska. University of California Press, 225-244.
Briggs, D. E. G., Wilby, P. R., Perez, B., Sanz, J. L. y Fregenal, M.M. 1997. The mineralization ofdinosaur soft tissue in the Lower Cretaceous of Las Hoyas, Spain. Journal of the GeologicalSociety of London, 154, 4, 587-588.
Buckland, W. 1824. Notice on Megalosaurus or great fossil lizard of Stonesfield.Trans Geol. Soc.London, 21, 390-397.
Buffetaut, E., Mechin, P. y Mechin-Salessy, A. 1988. Un dinosaure théropode d'affinitésgondwaniennes dans le Crétacé supérieur de Provence. C.R. Acad. Sci. Paris, Série II, 306, 153-158.
Buscalioni, A. D. 2000. Armuña. En Sanz, J. L.: Dinosaurios. Los señores del pasado. Ed. MartínezRoca, 295-296.
Buscalioni, A.D., Barbadillo, L.J. y Sanz, J.L. 1988. Los reptiles diapsidos de las Hoyas (CretácicoInferior, Cuenca). En: IV Jornadas de Paleontología, Salamanca, 27.
Buscalioni, A.D. y Sanz, J.L. 1984. Los Arcosaurios (Reptilia) del Jurásico Superior - Cretácico Inferiorde Galve (Teruel, España). Teruel, 71, 9-28.
Canudo, J.I., Cuenca-Bescós, G., Ruíz-Omeñaca, J.I. y Soria, A.R. 1996. Estratigrafía yPaleoecología de los vertebrados del Barremiense superior (Cretácico inferior) de Vallipón(Castellote, Teruel). Mas de las Matas, 9 - 34.
Canudo, J. I., Cuenca-Bescós, G. y Ruiz-Omeñaca, J.I. 1997. Dinosaurios dromeosaúridos(Saurischia: Theropoda) en el Barremiense Superior (Cretácico Inferior) de Castellote. Teruel.Geogaceta. 22, 39-42.
Canudo, J.I., López-Martínez, N. y Ruiz-Omeñaca, J.I. 2001. Los dinosaurios del Maastrichtiensesuperior (Cretácico superior) del Pirineo de Huesca (España). En: Actas de las I Jornadasinternacionales sobre Paleontología de Dinosaurios y su entorno. 319-328.
Casanovas-Cladellas, L. 1992. Novedades en el registro fósil de dinosaurios del Levante español.Zubia. 10, 139-151.
Casanovas-Cladellas, L. M., Santafé-Llopis. V. y Sanz-García, J.L. 1988. La primera resta fòssil d'unTeròpode (Saurischia, Dinosauria) en el Cretaci superior de la Conca de Tremp (Lleida, Espanya).Paleontologia i Evolució, 22, 77-81.
Casanovas-Cladellas, M.L., Santafe-Llopis, J.V. y Santisteban-Bove, C. 1993. First dinosaur teethfrom the Lower Cretaceous of Benicatazara (Aras de Alpuente, Valencia). Revue de Paléobiologie,Vol. spec. 7, 37-44.
Charig A. J. y Milner, A. C. 1997. Baryonyx walkeri, a fish-eating dinosaur from the Wealden of Surrey.Bulletin of the Natural History Museum of London (Geology Series), 53, 1, 11-70.
Chure, D. J. 1995. The teeth of small theropods from the Morrison Formation (Upper Jurassic:Kimmeridgian), Ut. Journal of Vertebrate Paleontology, 15 (Supplement to number 3): 23A.
Chure, D.J., Manabe, M., Tanimoto, M. y Tomida, Y. 1999. An unusual theropod tooth from the MifuneGroup (Late Cenomanian to early Turonian), Kumanoto, Japan. Proceedings of the SecondGondwanan Dinosaur Symposium. National Science Museum Monographs, 15, 291-296.
Company, J., Murelaga, X., Pereda-Suberbiola, X. y Ruiz-Omeñaca, J.I. 1999. The vertebrate faunafrom the new Late Cretaceous Chera locality (Valencia Province, Spain). En: IV Europeanworkshop on Vertebrate Paleontology, Albarracín, 37-38.
Coria, R. y Salgado, L. 1995. A new giant carnivorous dinosaur from the Cretaceous of Patagonia.Nature, 377; 6546, 224-226.

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
28
Crusafont-Pairó, M. y Adrover, R. 1966. El primer representante de la clase mamíferos hallado en elMesozoico de España. Teruel, 35, 139-143.
Crusafont, M. y Gibert, R. 1976. Los primeros multituberculados de España. Nota preliminar. ActaGeológica Hispánica, 11(3), 57-64.
Csiki, Z. y Grigorescus, D. 1998. Small theropods from the Late Cretaceous of the Hateg basin(Western Romania) - an unexpected diversity at the top of the food chain, Oryctos, 1, 87-104.
Currie, P.J. y Carpenter, K. 2000. A new specimen of Acrocanthosaurus atokensis (Theropoda,Dinosauria) from the Lower Cretaceous Antlers Formation (Lower Cretaceous, Aptian) ofOklahoma, USA. Geodiversitas, 22, 2, 207 - 246.
Currie, P. J., Rigby, J. K. y Sloan, R. E. 1990. Theropod teeth from the Judith River Formation ofsouthern Alberta, Canada. In: Dinosaurs systematics. Approaches and Perspectives, 282-298.
Diaz-Molina, M., Yébenes, A., Goy, A. y Sanz, J. L.1985. Le Jurassique Superieur detritique-carbonaté du synclinal de Galve (Teruel).Strata, Série 2, 2,155-182.
Díez, J.B., Pons, D., Canudo, J.I., Cuenca, G. y Ferrer, J., 1995. Primeros datos palinológicos delCretácico inferior continental de Pielago-0, Galve (Teruel, España). XI Jornadas de Paleontología,Tremp, 79-80.
Egozcue, J. 1873. Noticia sobre la existencia en España de restos fósiles de Megalosaurus y deHyaena spelaea y brunnea. Actas Real Sociedad Española de Historia Natural, 2, 29-30.
Estes, R. y Sanchiz, F. 1982. Early Cretaceous lower vertebrates from Galve (Teruel), Spain. Journalof Vertebrate Paleontology, 2, 21-39.
Fuentes Vidarte, C., Meijide Calvo, M., Izquierdo, L.A., Molinero, P., Montero, D., Pérez, G., Urien, V.,Meijide Fuentes, F. y Meijide Fuentes, M. 1999. Restos de Baryonyx (Dinosauria, Theropoda) en elweald de Salas de los Infantes (Burgos, España). I Jornadas Internacionales sobre Paleontologíade dinosaurios y su entorno, 25-26.
Fuentes Vidarte, C., Meijide Calvo, M., Izquierdo, L.A., Montero, D., Pérex, G., Torcida, F., Urién, V.,Meijide Fuentes, F. y Meijide Fuentes, M. 2001. Restos fósiles de Baryonyx (Dinosauria,Theropoda) en el Cretácico inferior de Salas de los Infantes (Burgos, España). En: Actas de las IJornadas internacionales sobre Paleontología de Dinosaurios y su entorno. 349 - 359.
Gauthier, J.A. 1986. Saurischian monophyly and the origin of birds.In:The origin of Birds and theEvolution of Flight. Memoirs of the California Academy of Sciences, 8, 1-55.
Gómez, B., Martín-Closas, C., Méon, H., Thévenard, F. y Barale, G. 2001. Plant taphonomy andpalaeoecology in the lacustrine Uña delta (Late Barremian, Iberian ranges, Spain).Palaeogeography, Palaeoclimatology, Palaeoecology, 170, 133-148.
Harris, J. D. 1998a. Large, Early Cretaceous theropods in North America. Lower and MiddleCretaceous Terrestrial Ecosystems. New Mexico Museum of Natural History and Science, 14, 225-228.
Harris, J. D.1998b. A reanalysis of Acrocanthosaurus atokensis, its phylogenetic status, andpaleobiogeographic impliations, based on a new specimen from Texas. Bulletin of the New MexicoMuseum of Natural History and Science, 13, 1-75.
Holtz, T. R. Jr. 1994. The phylogenetic position of the Tyrannosauridae: implications for theropodsystematics. Journal of Paleontology, 68, 5, 1100-1117.
Holtz, T. R. Jr. 1996. Phylogenetic taxonomy of the Coelurosauria (Dinosauria: Theropoda). Journal ofPaleontology, 70, 3, 536-538.
Holtz, T.R. Jr. 1998. A new phylogeny of the carnivorous dinosaurs. Gaia, 15, 5 - 61.
Kellner, A. W. A. y Campos, D. A. 1996. First early Cretaceous theropod dinosaur from Brazil, withcomments on Spinosauridae. Neues Jahrbuch Geologie Paläontologie Monatshefte, 199, 2, 151-166.
Kellner, A. W. A. y Campos, D. A. 2000. Brief review of dinosaur studies and perspectives in Brazil.An. Acad. Bras. Ci., 72, 4, 509-538.

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
29
Lamanna, M.C., Martínez, R.B. y Smith, J.B. 2002. A definitive abelisaurid theropod dinosaur dinosaurfrom the early Late Cretaceous of Patagonia. Journal of Vertebrate Paleontology, 22, 1, 58 - 69.
Lapparent, A.F. de, 1966. Nouveaux gisements de reptiles mesozoiques en Espagne. Notas yComunicaciones del Instituto Geológico y Minero de España, 84, 103-110.
Lapparent, A.F., Curnelle, R., Defaut, B., De Miroschedji, A. y Pallard, B. 1969 Nouveaux gisementsde Dinosaures en Espagne centrale. Estudios geológicos, XXV, 311-315.
Le Loeuff, J. 1997. Biogeography. En: Encyclopedia of Dinosaurs, Eds. Ph. J. Currie y K. Padian.Academic Press, 51-56.
Le Loeuff, J. 1988. Etude du gisement a vertebres continentaux maastrichtiens de Laño (PaysBasque, Espagne). Essai de quantification des relations paleobiogeographiques mondiales auCretace superieur. Memoire de D.E.A. de Paléontologie des Vertébrés et de Paléontologiehumaine. Université Pierre et Marie Curie, Paris VI, 50 pp. (inédita).
Le Loeuff, J. 1992. Les vértebrés continentaux du Crétacé supérieur d´Europe: Paléoécologie,Biostratigraphie et Paléobiogéographie. Mémoires des Sciences de la Terre de l´Université Pierreet Marie Curie, Paris VI, 92-93, 273 pp. (tesis doctoral, inédita).
Le Loeuff, J. y Buffetaut, E. 1991. Tarascosaurus salluvicus nov. gen., nov. sp. dinosaure théropodedu Crétacé supérieur du sud de la France. Geobios, 25, 5, 585-594.
López-Martínez, N., Ardèvol, L., Arribas, M. E., Civis, J., y Gonzalez Delgado, A. 1998. The geologicalrecord in non-marine enviroments around the K/T boundary (Tremp Formation, Spain). Bulletin dela Societe Geologique de France 169, 11-20.
López-Martínez, N., Canudo, J.I., Ardèvol, L., Pereda-Suberbiola, X., Orue-Etxebarria, X., Cuenca-Bescós, G., Ruíz-Omeñaca, J.I., Murelaga, X. y Feist, M. 2001. New dinosaur sites correlated withUpper Maastrichtian pelagic deposits in the Spanish Pyrenees: implications for the dinosaurextinction pattern in Europe. Cretaceous Research, 22, 41-61.
Martill, D. M., Cruickshank, A. R. I., Frey, E., Small, P. G., y Clarke, M. 1996. A new crestedmaniraptoran dinosaur from the Santana Formation (Lower Cretaceous) of Brazil. J. Geol. Soc.London 153(1), 5-8
Martill, D.M. y Hutt, S.1996. Possible baryonychid dinosaur teeth from the Wessex Formation (LowerCretaceous, Barremian) of the isle of Wight, England. Proceedings of the Geologists' Association,107, 81-84.
Martín-Closas, C. y Alonso-Millán, A. 1998. Estratigrafía y bioestratigrafía (Charophyta) del Cretácicoinferior en el sector occidental de la Cuenca de Cameros (Cordillera Ibérica). Revista de laSociedad Geológica de España, 11, 253 -269.
Martín-Closas, C. y Diéguez, C. 1998. Charophytes from the Lower Cretaceous of the Iberian ranges(Spain). Palaeontology, 41, 6, 1132-1152.
Martínez, R.D., García-Ramos, J.C., Piñuela, L., Lires, J. y Luna, M. 2001. Dinosaur remains from theprincipality of Asturias, Spain. Journal of Vertebrate Paleontology, 21(Supplement to number 3),78A.
Martínez, R., García-Ramos, J.C., Piñuela, L., Lires, J., Luna, M. y Veigas, D. 2000a. Dientes deTheropoda (Dinosauria: Saurischia) del Jurásico superior de Asturias, España. En: I CongressoIbérico de Paleontologia / XVI Jornadas de la Sociedad Española de Paleontología, 109-110.
Martínez, R., García-Ramos, J.C., Piñuela, L., Lires, J., Luna, M. y Veigas, D .2000b. Vértebrascaudales de Sauropoda y Theropoda (Dinosauria: Saurischia) del Jurásico superior de Asturias,España. En: I Congresso Ibérico de Paleontologia / XVI Jornadas de la Sociedad Española dePaleontología, 113-114.
Molnar, R. E., Kurzanov, S. My Dong, Z.-M. 1990. Carnosauria. In Dinosauria, Ed. D.B. Weishampel,P. Dodson, H. Osmólska. University of California Press, 169-209.
Novas, F. E. 1992. La evolución de los dinosaurios carnívoros.In: Los dinosaurios y su entornobiótico. Actas II Curso de Paleontología, 125-164.

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
30
Núñez-Betelu, K. 1999. Preliminary palynological assessment of the vertebrate-rich Laño beds: ageand paleoenvironment. Est. Mus. Cienc. Nat. de Alava, 14 (Núm. Espec. 1), 7-12.
Ostrom, J. H. 1990. Dromaeosauridae. In Dinosauria, Ed. D.B. Weishampel, P. Dodson, H. Osmólska.University of California Press, 269-279.
Pereda-Suberbiola, X. 1999. Las faunas finicretácicas de dinosaurios ibéricos. Zubia, 17, 259-279.
Pereda-Suberbiola, X., Astibia, H., Murelaga, X., Elorza, J.J. y Gómez-Alday, J.J. 2000. Taphonomyof the Late Cretaceous dinosaur-bearing beds of the Laño Quarry (Iberian Peninsula).Palaeogeogr., Palaeoclimatol., Palaeoecol., 157, 3-4, 247-275.
Pérez-Lorente, F., Cuenca-Bescós, G., Aurell, M., Canudo, J.I., Soria, A. R. y Ruiz-Omeñaca, J.I.1997. Las Cerradicas tracksite (Berriasian, Galve, Spain): Growing evidence for quadrupedalornithopods. Ichnos, 5, 109-120.
Pérez-Moreno, B. P. y Sanz, J.L. 1995. The hand of Pelecanimimus polyodon. A preliminary report. In:II International Symposium on Lithographic Limestones, Extended abstracts, 115-117.
Pérez-Moreno, B. y Sanz, J. L. 1997. Las Hoyas. In: Encyclopedia of Dinosaurs. Eds. Ph. J. Currie yK. Padian. Academic Press, 398-402.
Perez-Moreno, B. P., Sanz, J. L., Buscalioni, A. D., Moratalla, J. J., Ortega, F. y Rasskin-Gutman, D.1994. A unique multitoothed ornithomimosaur dinosaur from the Lower Cretaceous of Spain.Nature, 370, 363-367.
Pol, C., Buscalioni, A., Carballeira, J., Francés, V., López-Martínez, N., Marandat, B., Moratalla, J.J.,Sanz, J.L., Sigé , B. y Villatte, J. 1992. Reptiles and mammals from the Late Cretaceous newlocality Quintanilla del Coco (Burgos Province, Spain). N. Jb. Geol. Paläont. Mh.,184, 3, 279-314.
Prieto-Márquez, A., Gaete, R., Gallobart, A. y Ardèvol, L. 2000. A Richardoestesia-like theropod toothfrom the Late Cretaceous foredeep, south-central Pyrenees, Spain. Eclogae geol. Helv., 93, 497-501.
Querol, X., Salas, R., Pardo, G. y Ardèvol, L. 1992. Albian coal-bearing deposits of the Iberian Rangein northeastern Spain. Geological Society of America. Special Paper, 267, 193-208.
Rauhut, O. W. M. 1996. Dinosaur remains from the lignite coals of Guimarota (Kimmeridgian,Portugal) and Uña (Barremian, Spain) and paleogeographical implications. Journal of VertebratePaleontology, 16(Supplement to number 3), 59A.
Rauhut, O. W. M. 2000 The interrelationships and evolution of basal theropods (Dinosauria,Saurischia). Tesis Doctoral, Universidad de Bristol, 440 pp.
Rauhut, O. W. M. 2002a. Microrestos de dinosaurios del Cretácico inferior de Uña, España.Resumenes de las XVIII Jornadas Argentinas de Paleontología de Vertebrados, Bahia Blanca,Argentina, 36.
Rauhut, O. W. M. 2002b. Dinosaur teeth from the Barremian of Uña, Province of Cuenca, Spain.Cretaceous Research 23, 255–263.
Rauhut, O. W. M. y Zinke, J. 1995. A description of the Barremian dinosaur fauna from Uña with acomparison to that of Las Hoyas. En: II International Symposium on Lithographic limestones,Lleida-Cuenca, 123-126.
Rauhut, O. W. M. y Werner, C. 1995. First record of the family Dromaeosauridae (Dinosauria:Theropoda) in the Cretaceous of Gondwana (Wadu Milk Formation, northern Sudan).Paläontologische Zeitschrift, 69, 3/4, 475-489.
Royo y Gómez, J. 1920. Los yacimientos weáldicos del Maestrazgo. Boletín de la Real SociedadEspañola de Historia Natural, 20, 261-267.
Royo y Gómez, J. 1921. Los Facies continental en el Cretácico inferior Ibérico. En: AsociaciónEspañola para el Progreso de las Ciencias. Congreso de Oporto, IV, 221-235.
Royo y Gómez, J. 1925. Sobre los restos de reptiles weáldicos de Benageber (Valencia). Bol. R. Soc.Esp. Hist. Nat., XXV, 252.

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
31
Royo y Gómez, J. 1926. Los vertebrados del Cretácico español de facies weáldica. Boletín delInstituto Geológico de España, 47, 169-176.
Royo y Gómez, J. 1927. Sur la facies wealdien d'Espagne. Compte Rendu sommaire des Séances dela Société géologique de France, 1927, (Séance du 13 juin, communications orales), 11, 125-128.
Ruiz-Omeñaca, J. I. 1996. Los dinosaurios hipsilofodóntidos (Reptilia: Ornithischia) del Cretácicoinferior de Galve (Teruel). Tesis de Licenciatura. Universidad de Zaragoza, 1-338 p.
Ruiz-Omeñaca, J.I. y Canudo, J.I. 2001. Vallipón y La Cantalera: dos yacimientos paleontológicosexcepcionales. Naturaleza Aragonesa, 8, 8 -17.
Ruiz-Omeñaca, J. I., Canudo, J.I. y Cuenca-Bescós, G. 1996. Dientes de dinosaurios (Ornitischia,Saurischia) del Barremiense Superior (Cretácico Inferior) de Vallipón (Castellote, Teruel). Mas delas Matas,15, 59-103.
Ruiz-Omeñaca, J. I., Canudo, J.I. y Cuenca-Bescós, G. 1997. Primera evidencia de un área dealimentación de dinosaurios herbívoros en el Cretácico Inferior de España (Teruel). Monografíasde la Academia de Ciencias de Zaragoza, 10, 1-48.
Ruiz-Omeñaca, J. I., Canudo, J.I., Cuenca-Bescós, G. y Amo, O. 1998a. Theropod teeth from theLower Cretaceous of Galve (Teruel, Spain). Third European Workshop on VertebratePaleontology, Maastricht, 62-63.
Ruiz-Omeñaca, J. I., Canudo, J.I. y Cuenca-Bescós, G. 1998b. Primera cita de dinosauriosbarionícidos (Saurischia: Theropoda) en el Barremiense Superior (Cretácico Inferior) de Vallipón(Castellote, Teruel). Mas de las Matas, 17, 201-223.
Ruiz-Omeñaca, J. I., Cuenca Bescós, G. y Canudo, J.I. 1999. Dinosaur remains in the Barremian(Lower Cretaceous) of the Teruel province (Iberian Chain, NE Spain). IV European workshop onVertebrate Paleontology, Albarracín , 81-82.
Ruiz-Omeñaca, J. I. y Pereda-Suberbiola, X. 1999. Un documento inédito de Royo y Gómez sobre losdinosaurios del Levante. Temas Geológico-Mineros ITGE. Actas XV Jornadas de Paleontología.,26, 111-112.
Sampson, S.D., Witmer, L.M., Forster, C.A., Krause, D.W., O'Connor, P.M., Dodson, P. y Ravoavy, F.1998. Predatory Dinosaur Remains from Madagascar: Implications for the CretaceousBiogeography of Gondwana.Science. 280, 1048-1051.
Santafé, J.V., Casanovas, M.L., Sanz, J.L. y Calzada, S. 1982. Geología y paleontología(Dinosaurios) de las Capas Rojas de Morella (Castellón, España). Ed. Diputación Provincial deCastellón y Diputación de Barcelona, 1-169.
Santafé-Llopis, J. V.y Casanovas-Cladellas, M. L. 1993. Dinosaurios en la comunidad valenciana.Generalitat valenciana, 205 pp.
Santafé-Llopis, J. V., Casanovas-Cladellas, M. L., Sanz-García, J. L. y Calzada-Badía, S. 1979. Losdinosaurios de Morella (Nota preliminar). Acta geológica Hispánica, 13(5), 149-154.
Sanz, J.L. 1984. Las faunas españolas de Dinosaurios. En: I Congreso Español de Geología, tomo I,497-506.
Sanz, J. L. 2000. Dinosaurios. Los señores del pasado. Ed. Martínez Roca, 1-299.
Sanz, J.L., Buscalioni, A.D., Casanovas, M.L. y Santafé, J.V. 1987. Dinosaurios del Cretácico Inferiorde Galve (Teruel, España). Estudios geológicos, vol. extr. Galve - Tremp, 45-64.
Sanz, J. L., Buscalioni, A.D., Moratalla, J.J., Francés, V. y Antón, M. 1990. Los reptiles mesozoicosdel registro español. Monografías del Museo Nacional de Ciencias Naturales, 2, 1-79.
Sanz, J.L., Buscalioni, A.D., Pérez-Moreno, B., Moratalla, J. y Jiménez García, S. 1992. Losdinosaurios de Castilla y Leon. En: Vertebrados fósiles de Castilla y León, Museo de Salamanca,47-57.
Schudack, U. y Schudack, M. 2002. New biostratigraphical data for the Upper Jurassic of Asturias(Northern Spain) based on Ostracoda. Revista Española de Micropaleontología, 34, 1, 1-18.

Canudo, J. I. y Ruiz-Omeñaca, J. I. 2003. Ciencias de la Tierra.Dinosaurios y otros reptiles mesozoicos de España, 26, 347-373.
32
Schulz, G. 1858. Descripción geológica de la provincia de Oviedo. 4ª edición (1930) Obras completasde G. Schulz. Graficas Reunidas, Madrid, 1-138.
Sereno, P. 1997. The origin and evolution of dinosaurs. Annual Review of Earth and PlanetarySciences, 25, 435-489.
Sereno, P. C. Beck, A. L., Dutheil, D. B., Gado, B., Larsson, H. C. E., Lyon, G. H., Marcot, J. D,Rauhut, O. W. M., Sadleir, R. W., Sidor, C. A., Varricchio, D. J., Wilson y G. P. Wilson, J. A. 1998.A long-snouted predatory dinosaur from Africa and the evolution of spinosaurids. Science, 282:1298-1302.
Sereno, P. C., Dutheil, D. B., Iarochene, M., Larsson, H. C. E. , Lyon, G. H., Magwene, P. M., Sidor,C. A, Varricchio, D. J. y Wilson, J. A. 1996. Predatory dinosaurs from the Sahara and lateCretaceous faunal differentiation. Science, 271, 986-991.
Suarez Vega, L.C. 1974. Estratigrafía del Jurásico en Asturias. Cuadernos de geología ibérica, 3, 1,1-304.
Taquet, P. y Russell, D. A. 1998. New data on spinosaurid dinosaurs from the early Cretaceous of theSahara. C.R. Acd. Sci. Paris. 327: 347-353.
Torcida Fernández-Baldor, F. 1996. Registro de dinosaurios en el sureste de la provincia deBurgos.Zubía, 14, 89-104.
Torcida, F., Fuentes, C., Izquierdo, L A., Montero, D. y Urién, V. 1997. Dientes de DinosauriosTerópodos (cf. Baryonyx) en el Weald de Burgos (España). Stvdia Geologica Salmanticensia, 33,4, 59-65.
Torices Hernández, A. 2002. Los dinosaurios terópodos del Cretácico Superior de la Cuenca deTremp (Pineos sur-centrales, Lleida). Tesis de Licenciatura, Universidad Complutense de Madrid,58 pp (inédita).
Torices Hernández, A. (en prensa). Los dinosaurios teropodos del Cretácico Superior de la Cuencade Tremp, (Pirineos Sur-Centrales, Lleida). Coloquios de Paleontologia, 53.
Vicens, E., Ardèvol, L., López-Martínez, N. y Arribas, M. E. (en prensa). Rudist biostratigraphy in theCampanian-Maastrichtian of the South-Central Pyrenees, Spain. Courier Forsch. Seckenberg.
Vickers-Rich, P., Rich, T. H., Lanus, D. R., Rich, L. S. V. y Vacca, R. 1999. "Big Tooth" from the EarlyCretaceous of Chubut Province, Patagonia; a possible carcharodontosaurid. Proceedings of theSecond Gondwanan Dinosaur Symposium, National Science Museum Monographs, 15, 85-88.
Viera, L. I. y Torres, J.A. 1995. Presencia de Baryonyx walkeri (Saurischia, Theropoda) en el Wealdde La Rioja (España). Nota previa. Munibe (Ciencias Naturales), 47, 57-61.
Vilanova y Piera, J. 1873. Restos de Iguanodon de los lignitos de Utrillas y otro de Morella. Actas dela Sociedad Española de Historia Natural, 2, 8.
Zinke, J. 1998. Small theropod teeth from the Upper Jurassic coal mine of Guimarota (Portugal).Paläont. Z., 72 (1/2): 179-189.
Zinke, J. y Rauhut, O.W.M. 1994. Small theropods (Dinosauria, Saurischia) from the Upper Jurassicand Lower Cretaceous of the Iberian Peninsula. Berliner geowiss. Abh., E 13, 163-177.