lecture 3 implications of theory. mass & energy balance the standard deb model specifies fluxes...
Post on 20-Dec-2015
215 views
TRANSCRIPT

Lecture 3
Implications of theory

Mass & energy balance
The standard DEB model specifies fluxes of 4 organic compounds food, faeces, stucture (growth), reserve (including reproduction)
The fluxes of 4 mineral compouds (CO2, H2O, O2, NH3) follow from conservation of chemical elements C, H, O, N and strong homeostasis
The standard DEB model assumes that only food is limiting
Dissipating heat follows from conservation of energy and strong homeostasis (constant chemical potentials)

Method of indirect calorimetry
Empirical origin (multiple regression): Lavoisier 1780
Heat production = wC CO2-production + wO O2-consumption + wN N-waste production
DEB-explanation:Mass and heat fluxes = wA assimilation + wD dissipation + wG growthApplies to CO2, O2, N-waste, heat, food, faeces, …
For V1-morphs: dissipation maintenance

Mass fluxes
dioxidecarbon 2 CJ
water2 HJ
dioxygen2 OJ
ammonia10 NJ
foodXJ
structure40 VJ
reserve)(10
REE JJ faeces
PJ
llength scaled
f
lux
f
lux
bl pl
notice small dent due to transition
maturation reproductionAt abundant food: growth ceases at l = 1
allocation toreproduction
use of reservenot balanced by
feeding in embryo
bl pl
0 1
10

Methanotrophy
Yield coefficients Y and chemical indices n depend on (variable) specific growth rate rNWOWHW nnnWX3NX2OX2CX4 NOCH Y NH Y O Y CO Y CH
AC Assim (catabolic) -1 1 2 -2 0 0 0
AA Assim (anabolic) -1 0 1 0
M Maintenance 0 1 -1 0
GC Growth (catabolic) 0 1 -1 0
GA Growth (anabolic) 0 0 -1 1
C Carbon 1 1 0 0 0 1 1
H Hydrogen 4 0 2 0 3
O Oxygen 0 2 1 2 0
N Nitrogen 0 0 0 0 1
2/2/2/
2/32/2/
2/2/1
2/2/3
2/2/
2/2/32
From
GHEOVOE
GOE
GNEHVHE
GHE
NVNEG
NE
MHEOE
MOE
HENEM
HE
OEA
HXA
OX
HEA
NXA
HX
NEA
NX
YnnY
YnnY
nnY
YnY
nnY
nYY
nYY
nY
nY0
AHXY A
OXY ANXY
MHEY
GHEY
MHEY
MOEYM
OEYG
OEY GNEY
NEn
NEn
HEn
OEn
NEn
HVn
OVn
NVn
sym
bol
proc
ess
X: m
etha
ne
C: c
arbo
n di
oxid
e
H: w
ater
O: d
ioxy
gen
N: a
mm
onia
E: r
eser
ve
V: s
truc
ture
EAXE jy )1(
EAj
EGVE jy )1(
EGVE jy
EMj
EVE
EMEEVV
EVEG
MEVEM
EAmEA
ym
jkmM
dt
dMr
ryj
kyjXK
Xjj
1
For reserve density mE = ME/MV (ratio of amounts of reserve and structure), the macroscopic transformation can be decomposed into 5 microscopic ones with fixed coefficients
rate
Yie
ld c
oeff
icie
ntsT
Che
mic
al in
dice
s

Methanotrophy
spec growth rate, h-1 spec growth rate, h-1
X/O
N/O
C/O
flux
rat
io, m
ol.m
ol-1
spec
flu
x, m
ol.m
ol-1.h
-1
CE
N
X
O
X: methaneC: carbon dioxideO: dioxygenN: ammoniaE: reserve
jEAm = 1.2 mol.mol-1.h-1
yEX = 0.8yVE = 0.8kM = 0.01 h-1
kE = 2 h-1
nHE = 1.8nOE = 0.3nNE = 0.3
nHV = 1.8nOV = 0.3nNV = 0.3
chemical indices
Kooijman, Andersen &Kooi 2004. Ecology, to appear

Biomass compositionData Esener et al 1982, 1983; Kleibsiella on glycerol at 35°C
nHW
nOW
nNW
O2
CO2Spec growth rate, h-1
Spec growth rate
Spec growth rate, h-1
Rel
ativ
e ab
unda
nce
Spe
c pr
od, m
ol.m
ol-1.h
-1
Wei
ght y
ield
, mol
.mol
-1
nHE 1.66 nOE 0.422 nNE 0.312nHV 1.64 nOV 0.379 nNV 0.189
kE 2.11 h-1 kM 0.021 h-1
yEV 1.135 yXE 1.490rm 1.05 h-1 g = 1
•μE-1 pA pM pG
JC 0.14 1.00 -0.49
JH 1.15 0.36 -0.42
JO -0.35 -0.97 0.63
JN -0.31 0.31 0.02
Entropy J/C-mol.K Glycerol 69.7 Reserve 74.9 Structure 52.0
Sousa et al 2004Interface, subm

Product Formation
throughput rate, h-1
glyc
erol
, eth
anol
, g/l
pyru
vate
, mg/
l
glycerol
ethanol
pyru
vate
Glucose-limited growth of SaccharomycesData from Schatzmann, 1975
According to Dynamic Energy Budget theory:
Product formation rate = wA . Assimilation rate + wM . Maintenance rate + wG . Growth rate
For pyruvate: wG<0

1 Reserve – 1 Structure

2 Reserves – 1 Structure

Reserve Capacity & Growth
low turnover rate: large reserve capacity
high turnover rate: small reserve capacity

Multivariate extensionsanimal heterotroph phototroph
symbiosis plant

Interactions of substrates

Photosynthesis
2 H2O + 4 h O2 + 4 H+ + 4 e-
CO2 + 4 H+ + 4 e- CH2O + H2O
CO2 + H2O + light CH2O + O2
3222
32
NHOOHCONOCH
NOOCH
ENOEHECEnnn
ENEC
HNEOEHE
OH
yyyy
yy

Simultaneous nutrient limitation
Specific growth rate of Pavlova lutheri as function of intracellular phosphorus and vitamine B12 at 20 ºC
Data from Droop 1974Note the absence of high contents for both compounds
due to damming up of reserves, andlow contents in structure (at zero growth)

Reserve interactions
Spec growth rate, d-1 Spec growth rate, d-1 Spec growth rate, d-1
P-c
onte
nt, f
mol
.cel
l-1P
-con
c, μ
M
B12
-con
c, p
M
B12
-con
t., 1
0-21 .m
ol.c
ell-1
P Vitamin B12
kE 1.19 1.22 d-1
yXV 0.39 10-15 2.35 mol.cell-1
jEAm 4.91 10-21 76.6 10-15 mol.cell-1. d-1
κE 0.69 0.96
kM 0.0079 0.135 d-1
K 0.017 0.12 pM, μM
Data from Droop 1974 on Pavlova lutheri
P(μM) B12(pM)
1.44 68
14.4 6.8
1.44 20.4
1.44 6.8

Steps in foodGrowth of Daphnia magna at 2 constant food levels
time, d time, d time, d time, d
0 d 7 d 14 d 21 dle
ngth
, mm
leng
th, m
m
Only curves at 0 d are fittedNotice • slow response• gut content in down steps
Steps up
Steps down

Growth on reserve
Opt
ical
Den
sity
at 5
40 n
m
Con
c. p
otas
sium
, mM
Potassium limited growth of E. coli at 30 °CData Mulder 1988; DEB model fitted
OD increases by factor 4 during nutrient starvationinternal reserve fuels 9 hours of growth
time, h

Growth on reserve
Growth in starved Mytilus edulis at 21.8 °CData Strömgren & Cary 1984; DEB model fitted
internal reserve fuels 5 days of growth
time, d
grow
th r
ate,
mm
.d-1

Protein synthesis
spec growth rate, h-1 scaled spec growth rate
RN
A/d
ry w
eigh
t, μg
.μg-1
scal
ed e
long
atio
n ra
te
Data from Koch 1970Data from Bremer & Dennis 1987
RNA = wRV MV + wRE ME
dry weight = wdV MV + wdE ME

Scales of life
Life span
10log aVolume
10log m3earth
whale
bacterium
water molecule
life on earth
whale
bacteriumATP

Inter-species body size scaling• parameter values tend to co-vary across species• parameters are either intensive or extensive• ratios of extensive parameters are intensive• maximum body length is allocation fraction to growth + maint. (intensive) volume-specific maintenance power (intensive) surface area-specific assimilation power (extensive)• conclusion : (so are all extensive parameters)• write physiological property as function of parameters (including maximum body weight)• evaluate this property as function of max body weight
]/[}{ MAm ppL
}{ Ap
][ Mp
mA Lp }{
Kooijman 1986 Energy budgets can explain body size scaling relationsJ. Theor. Biol. 121: 269-282

Primary scaling relationships
assimilation {JEAm} max surface-specific assim rate Lm
feeding {b} surface- specific searching rate
digestion yEX yield of reserve on food
growth yVE yield of structure on reserve
mobilization v energy conductance
heating,osmosis {JET} surface-specific somatic maint. costs
turnover,activity [JEM] volume-specific somatic maint. costs
regulation,defence kJ maturity maintenance rate coefficient
allocation partitioning fraction
egg formation R reproduction efficiency
life cycle [MHb] volume-specific maturity at birth
life cycle [MHp] volume-specific maturity at puberty
aging ha aging acceleration
maximum length Lm = {JEAm} / [JEM] Kooijman 1986J. Theor. Biol. 121: 269-282

Follows from:1. maturity at birth equals a given value2. reserve density at birth equals that of mother
State variables:
Parameters:
Problem: Given parameter values, find
Initial reserve of an egg
Theory in Kooy2008

Effects of nutrition
scaled res density at birth
scaled res density at birth
scaled res density at birth
scal
ed le
ngth
at b
irth
scal
ed in
itial
res
erve
scal
ed a
ge a
t birt
h
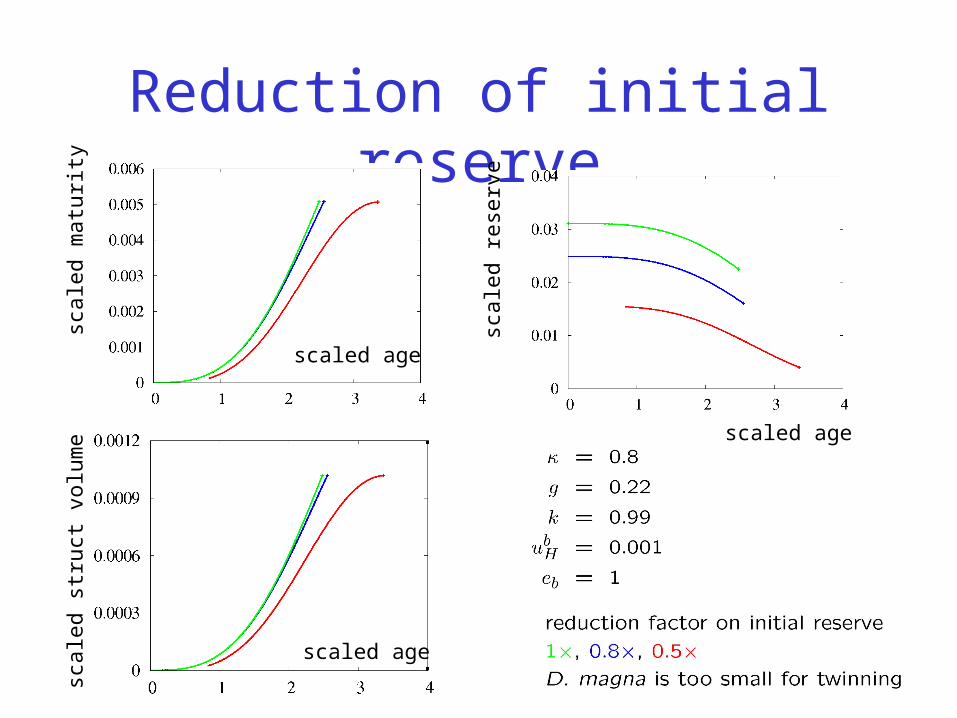
Reduction of initial reserve
1
0.8
0.5scaled age
scaled age
scaled age
scal
ed m
atur
itysc
aled
str
uct v
olum
e
scal
ed r
eser
ve

Scaling relationships
log zoom factor, z
log zoom factor, z
log zoom factor, z
log
scal
ed in
itial
res
erve
log
scal
ed a
ge a
t birt
h
log
scal
ed le
ngth
at b
irth
approximate slope at large zoom factor

Length at puberty
L, cm
Lp,
cm
Clupea• Brevoortia° Sprattus Sardinops Sardina
Sardinella+ Engraulis* Centengraulis Stolephorus
Data from Blaxter & Hunter 1982
Clupoid fishes
Length at first reproduction Lp ultimate length L

Body weight
Body weight has contributions from structure and reserveIf reserve allocated to reproduction hardly contributes:
13/4
13/100
11
1
)(][
][
W
EEmV
EEmV
f
EEV
L
μwEd
μwEdVμwEVdW
VVV/VVW
][ m
E
E
V
Eμwd
WLE
V
W
V
Wintra-spec body weightinter-spec body weightintra-spec structural volumeInter-spec structural volumereserve energycompound length-parameter
specific density for structuremolecular weight for reservechemical potential of reservemaximum reserve energy density

Feeding rateslope = 1
poikilothermic tetrapodsData: Farlow 1976
Inter-species: JXm VIntra-species: JXm V2/3
Mytilus edulisData: Winter 1973
Length, cm
Filt
ratio
n ra
te, l
/h

Scaling of metabolic rate
intra-species inter-species
maintenance
growth
weight
nrespiratio3
32
dl
llls
43
32
ldld
lll
EV
h
structure
reserve
32 vll
l0l
0
3lllh
Respiration: contributions from growth and maintenanceWeight: contributions from structure and reserveStructure ; = length; endotherms 3l l
3lllh
0hl

Metabolic rate
Log weight, g
Log metabolic rate,
w
endotherms
ectotherms
unicellulars
slope = 1
slope = 2/3
Length, cm
O2 consum
ption,
l/h
Inter-speciesIntra-species
0.0226 L2 + 0.0185 L3
0.0516 L2.44
2 curves fitted:
(Daphnia pulex)

At 25 °C : maint rate coeff kM = 400 a-1
energy conductance v = 0.3 m a-1
25 °CTA = 7 kK
10log ultimate length, mm 10log ultimate length, mm
10lo
g vo
n B
ert
grow
th r
ate
, a-1
a↑0
Von Bertalanffy growth rate