lect 24 and 25: cell signal transduction cells respond to molecular cues they receive: from...
TRANSCRIPT
LECT 24 and 25: CELL SIGNAL TRANSDUCTION
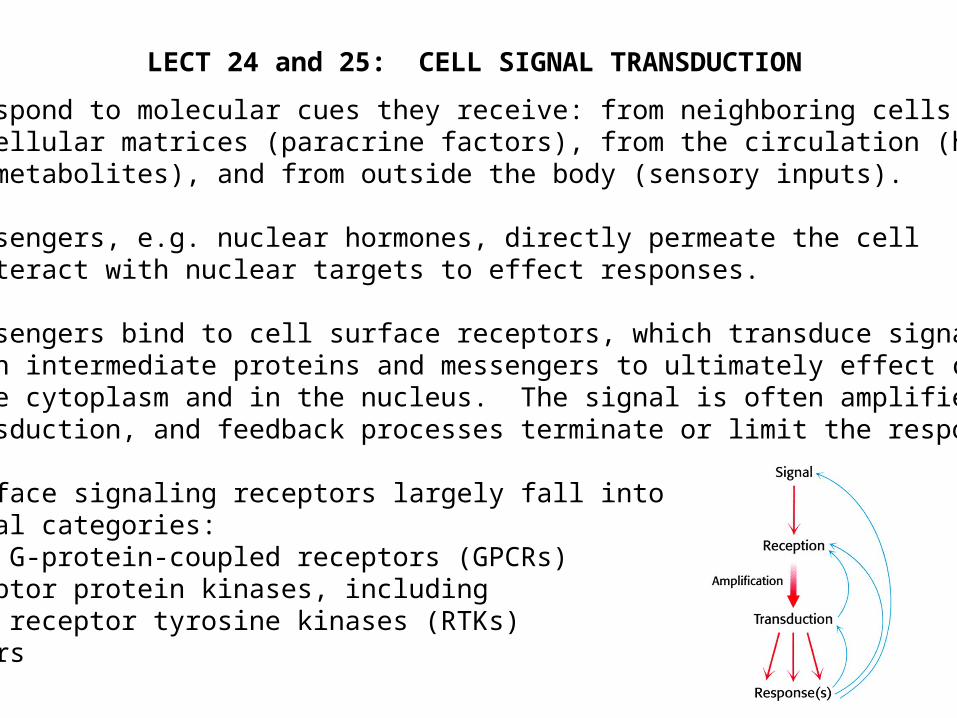
Cells respond to molecular cues they receive: from neighboring cells or extracellular matrices (paracrine factors), from the circulation (hormones and metabolites), and from outside the body (sensory inputs).
Some messengers, e.g. nuclear hormones, directly permeate the cell and interact with nuclear targets to effect responses.
Most messengers bind to cell surface receptors, which transduce signals through intermediate proteins and messengers to ultimately effect changes in the cytoplasm and in the nucleus. The signal is often amplified during transduction, and feedback processes terminate or limit the response.
Cell surface signaling receptors largely fall into several categories: …7-TM G-protein-coupled receptors (GPCRs) …Receptor protein kinases, including receptor tyrosine kinases (RTKs) …Others

Second Messengers Are Synthesized or Mobilized During Signaling

MANY SECOND MESSENGERS ACTIVATE SERINE/THREONINE PROTEIN KINASES
Each S/T-protein kinase phosphorylates exposed serine or threonine residueswithin a consensus sequence. E.g.
------Arg-Arg-Xxx-Ser-Xxx------- ------Arg-Arg-Xxx-Ser-Xxx-------
HO-P=O
O-
-
O-
ATP ADP
PKA

7-TM GPCRs Are Numerous and Mediate Many Biological Functions

7-TM GPCRs Are Numerous and Mediate Many Biological Functions
GPCRs couple to heterotrimeric G proteins to mediate signaling
Activated -adrenergic receptor activates Gas, which in turn activates adenylate cyclase (AC), which synthesizes 2o messenger cAMP, which activates protein kinase A

The Heterotrimeric G Protein Activation/Inactivation Cycle

Phospholipid Hydrolysis by PhospholipaseC Generates Two Second Messengers
There are several classes of PLC. One class is regulated by GPCRs

PLC-Derived Second Messengers Activate Protein Kinase C

ODORANT RECEPTOR STIMULATION OPENS cAMP-GATED CATION CHANNEL

Light-Activated Rhodopsin Turns Off a Dark Current ThroughcGMP-gated Cation Channel
Rhodopsin is GPCR using light-activated retinal is ligand. Couples through G protein transducin, which activates cGMP phosphodiesterase.

Insulin Receptor is Receptor Tyrosine Kinase Activated byInsulin-Induced Receptor Autophosphorylation
Autophosphorylation is really receptor subunit transphosphorylation.
Autophosphorylation stimulates the receptor’s kinase activity AND creates receptor recruitment sites for signaling target proteins.

IR Kinase Domain Phosphorylation Induces Conformational Change
When unphosphorylated, the “activation loop” of the kinase tends to fold in such a way as to block the kinase’s ATP binding site. Phosphorylation prevents this inhibition.
How does insulin induce IR phosphorylation? By two steps:
1. By changing the positioning of the two subunits w.r.t. each other
2. Weak activity overcome by proximity of first target (other subunit)

Tyrosine Phosphorylation Creates Target Recruitment & Activation Sites
IR phosphorylation in juxtamembrane region creates IRS-1 recruitment site.Recruited IRS-1 is itself tyrosine phosphorylated by IR, creating a set of recruitment sites.PI3K is recruited to and activated by phosphorylated IRS-1.PIP2 is phosphorylated by PI3K, which then activates protein kinase cascade.
Activated AKT mediatesActivated AKT mediatesmany insulin responsesmany insulin responses

SH2 and PTB Domains of Target Protein Recognizes Phosphotyrosine in aSequence-Specific Context
The SH2 domain of PI3K recognizes the sequence -pY-x-x-M- on IRS-1.The PTB domain of IRS-1 recognizes -L-Y-A-S-S-N-P-A-pY- on IR.
IRS-1 PTB bound to phospho-IR peptide

Epidermal Growth Factor Activates Its Receptor by InducingDimerization and Autophosphorylation
Monomeric EGFR is dimerized by EGF, by stabilizing and intrinsic receptor dimerization site.
Dimerization brings kinase domains close together, enabling auto(trans)phosphorylation, creating substrate recruitment sites.
Ligand-mediatedLigand-mediatedreceptor clustering receptor clustering
is a commonis a commontheme in signal activationtheme in signal activation

Epidermal Growth Factor Receptor Signals to Activate RAS Protein
Oncogenic RAS mutations stabilize theOncogenic RAS mutations stabilize the GTP-bound state in absence of growthGTP-bound state in absence of growth factor, causing unregulated signalingfactor, causing unregulated signaling

Notch Receptor Signaling Controls Local Cell Fate Decisions
-- Notch (a receptor) and its ligands Delta and Serrate are conserved in all vertebrates as well as complex invertebrates (flies, worms, etc).-- Notch controls cell fate decisions at many times and places in development.-- The most classic type of decision process mediated by Notch is Lateral InhibitionLateral Inhibition: a group of equipotent cells selects some to assume a specific fate, while others of the group are inhibited. Inhibition requires Notch.Inhibition requires Notch.
UncommitedPrecursors
DifferentiatedCells
NormalDevelopment
PrecursorAblation
Notch-
Mutant
-- Ligand for Notch produced in cell assuming “black”“black” fate, acting to inhibit neighbors from assuming same fate. -- Notch signaling thereby induces the “red”“red” differentiated state

Canonical Notch Signaling Pathway
Transmembrane Notch binds a membrane-bound ligand (Delta or Serrate) on neighboring cell.
Induces Notch proteolysis, freeing cytoplasmic NICD.
NICD goes to nucleus, acting as cofactor for activation transcription.
Signaling is terminated by NICD phosphorylation, ubiquitination, and proteasomal degradation.

The Logic of Notch-Mediated Lateral Inhibition
Equipotent precursors may have randomrandom small differences in Notch and Delta expression levels.
Notch signaling mediates a pathway through E(spl) and AS-C that enhances Notch expression and decreases Delta expression
Loss of Notch signaling decreases Notch expression and enhances Delta expression.
The AS-C transcription factor also controls adoption of a particular differentiated state, so that Notch signaling blocks this differentiated state.

Fringe-Mediated Notch Glycosylation Alters Ligand Binding to Notch
Development of adult appendages (e.g. wings) in insects occurs in imaginalimaginal discsdiscs that are patternedpatterned during larval stages. A key step in patterning is
activation of Notch in a 2-cell-width stripe along dorsal/ventral midline of disc.Fringe is a cell-bound Notch glycosidaseglycosidase
that shifts Notch’s ligand preference from Serrate to Delta
At early stage of disc patterning, Notch is expressed everywhere,
Delta is not expressed, and Serrate and Fringe are only
in dorsal half of disc.
Serrate can only activate unmodified Notch at
ventral margin.
Serrate also induces Deltaexpression in ventral margin.
Delta can then activatemodified Notch at
dorsal margin.