le regime alimentaire de la chouette effraie,
TRANSCRIPT

LE REGIME ALIMENTAIRE DE LA CHOUETTE EFFRAIE, TYTO ALBA, DANS LES PYRENEES-ORIENT ALES.
ETUDE DES V ARIA TI ONS ECOGEOGRAPHIQUES
Roland M. LIBOIS *, Roger FoNs * * e t Marie-Charlotte SAINT GIR ONS * * *
I l peut paraître surp renant qu€ l'on s'a ttache une fois d e plus à l'examen du régime alimentaire de la Chouette effraie (Tyto alba) .
L'analyse d e pelot€s de réj ection, méthode à l a fois élégante e t efficace qui permet la récolte rapide d'un matériel abondant sans porter préj udice aux animaux étudiés, a d'abord largement été utilisée par l€s ornithologues désireux de connaître en déta il le régime alimentaire de certaines espèces, notamment les rapaces. Il fallut attendre Husson (1949) pour voir la techniqu€ adoptée par des mammalogistes .
Les pelotes de rapaces ont permis de mettre en évidence des variations de la densité des populations de proi€s en fonction des saisons (Saint Girons, 1968 ; Fairley et Clark, 1972 ; 'Vebster, 1973 ; Brown, 1 981) ou d'année en année (Bovet, 1 963 ; Martin, 1973 ; Sa int Girons et Thouy, 1 978) et certains auteurs ont œmarqué que Je régime pouvait être considérablement affecté par les conditions météorologiques (Glue et Nuttall, 1971 ; Sans Coma et al., 1976) . L'inadéquation du régime de l'oiseau à la densité réelle de ses proies (vo i r Saint Giro ns et Spitz, 1966) explique sans doute que peu d'auteurs se soient intéressés à cet aspect du problème de manière approfondie . Tinbergen (1 933) estime cependant que les variations locales ou temporaires du régime du Hibou moyen-duc ( Asio otus) sont touj ours causées par des variations co ncordantes
* Laboratoi J·c d'éth o l o g i e d e l 'Un iversité de L i ège, q u a i V a n Beneden, 22,B 4020 Liège . Cet auteur 1·e mercie l e F.N.R.S . pour I 'oct1·o i des crédits (réf. V3f5-MB 287 OE ct V3f5-LC 20842) qu i l u i ont permis de couvrir une partie des fra i s de m i s s i o n .
* * Centl"c d ' é c o l o g i e méditeiT anéenne . Laboratoire Arago, F 66650 Banyu l ssur-M er.
* * * Lab01·a t o i r e d ' E Y o l u t i o n des Eh·es Organisés , 1 05 , boul evard Rasp a i l , F 7 5006 P a r i s .
Rev. Ecu / . ( Terre Y i e ) , v o l . . 7 , 1983

des réserves de nourriture disponibles. Pour Blondel et Frochot (1967) , même la spécialisation individuelle ne serait que la traduction de l'opportunisme habituel du rapace : une réponse aux conditions de milieu (voir par exemple Cruz et Garcia Rodriguez, 1969) . De Bruij n (1979) a observé, aux Pays-Bas, d'énormes changements dans la composition du régime alimentaire de l'Effraie selon que les pelotes étaient récoltées lors d'une année où le Campagnol des champs, Microtus arvalis, était abondant ou au contraire très rare. Il constate aussi des différences saisonnières accentuées e t les met en relation avec l 'importance des ressources en campagnols, moindres en hiver et au printemps.
L'Effraie traduirait donc au niveau de la composition de son menu celle des faunes locales de petits mammifères à un moment donné. Ainsi, certains ont abordé l'étude du milieu de chasse de l'oiseau (Von Knorre, 1973) , celle de l'influence des mo difications de paysage sur les communautés de micromammifères (Zelenka et Pricam, 1964 ; Rybar, 1969) ou encore celle des variations régionales ou géographiques du régime de cette espèce, en relation avec les populations de proies. A cet égard, Schmidt (1 973 a) e t Cheylan (1976) comparent des régions trop différentes sur le plan faunistique pour leur permettre d'envisager une approche écologique . Herrera (1974 a) regroupe ses échantillons en cinq ensembles correspondant chacun à un grand type de milieu . D 'autres se réfèrent à des découpages régionaux préexistants reposant sur des critères définis p ar des géographes (régions n aturelles : Saint Girons et Martin, 1 973 ; Gorner, 1979 ; d ivisions administratives : Glue, 1974) ou par des botanistes (districts phytosociologiques : De Bruij n, 1 979) . Les différences locales ou régionales y sont masquées puisque les variations du régi me alimentaire y sont généralement établies sur des moyennes régionales. Si tel n'est p as J e cas, j amais les auteurs n e s e sont a ttachés à la description précise des conditions locales e t leurs conclusions, par ailleurs souvent j udicieuses, reposent essentiellement sur leur bon sens et sur la connaissance générale qu'ils ont de l'écologie des petits mammifères proies (Balat, 1956 ; Tyrner et Barta, 1971 ; Chaline et al . , 1974 ; Ruprecht, 1979) .
Quelques travaux seulement tentent une approche moins empirique et mettent directement en relation les fluctuations du régime avec des paramètres écologiques : fertilité des sols (CabonRaczyns'ka et Ruprecht, 1977) , altitude, d i s tance des poinl s de récolte par rapport à la mer (Martin e t Vericad, 1977) , variables climatiques (Saint Girons et Vesco, 1 974 ; Fayard et Erome, 1 977) , importance de la couverture boisée (Lovari et al . , 1 976) .
D ans les Pyrénées-Orientales, no us nous sommes aussi servis du régime de l'Effraie pour mener une recherche similaire (Fons et al . , 1980) . Comme celles de Fayard e t Erome, nos conclusions englobent un cadre assez limité : nous n'avons p as essayé non plus
- 188 -

d'exp l i q u e r l e s v a ri a t ions q u a n t i t a t ives d u regi m e de l 'Effraie q ue nous obse rvion s d a n s cet te région . C'es t ce poin t que nous n o u s p roposon s d 'aborder ic i e n nous efforçan t de c e rn e r avec J e p l u s d e f inesse poss i b l e l e s d i v e rses i nf luences loca les .
NIA TERIEL E T ME THODES
To u t e s l e s égl ises d e s Pyrénées- O rien ta les (en v i ro n 200) , l es r u ines , v i e i l l e s to u rs e t bà t im e n ts abandonnés rencont rés au cours d e n o s d é p lace me n ts o n t é t é sys témat iquement v i si tés , y co mpris dans les zones o ù nous n e pe nsions pas renco n t re r de Choue t t e eff r a i e . D ' a p rès Sch n e i d e r ( 1 964) , c e t o i seau n e n i c h e ra i t g u è re a u-dessus d e 700 1 11 . N o u s avons cep end a n t réco l t é des p e lotes p l u s h a u t ( 1 000 m ) d a n s u n g î t e t e mporaire .
D i x- n e u f lo ts d e p e l o t e s de Tyto alba comp re n a n t au m i n i m u m 5 0 Ve rtébrés o n t é t é co l le c tés . D a ns l a mesure d u poss i b le, nou s avons essayé d e ré d u i re l 'i m p a c t d 'éven t uelles f l u c tu a t i ons pér io -
+ + + + + + L i m ite Q ,u /"( /1 ' i!tx
A li 1 ( (, l'
./ . · 'c •
'
, ...,_ '
r .\ /' A C N
F i g u re l . - Ca r t e d e s Py r(· n (· e s -O r i c n t a l e s p o rt a n t i n d i c :l l i o n d e s s t a t i o n s et d e p a ra m èt re s é c o c l i m a t i f ] u e s .
1 . Ca n e t s . T h u i r 1 5 . M o n t b o l o 2 . C l a i ra n . S t . F e l i u d ' A v a l ! Hi. M o s set : J . C o t· n e i l l a del V e r c o l 1 o . L a t o u r d e F ra n c e 1 7 . P ra t s d e M o l l o 4 . A r ge l è s- s u r-i\l c r I l . J I l e-su r-Têt 1 8 . H a b o u i l l ct rl . E s p i ra d e l ' A g l y 1 2 . C a u d i è s d e F e n . 1 9 . Co l d ' A u s s i è 1 · e s ( i . Péz i .J l a l a R i v i èr e 1 3 . O m s 2 0 . A y g u a t e b i a 7 . S o t· è d c d e s A l bè r e s H. S o u r n i a 2 1 . S a n sa
- 1 89

TABLEA Régime alimentaire de la Chouette effraie (Tyto alb :
L e s numéros des s tat ions correspondent it ceux de l a figure
Station No 1 2 3 4 5 6 7 8
AUitude (rn) 9 1 0 I l 1 6 28 67 80 9 1
n % n % n % n % n % n % n % n INSECTIVORES
T11/ptl &I'OfHIM 2 0,2 2
Sonx «llfWtlftiS»
Sox minuhu
Neomy1 odkru 1 0, 1
NeomyJ anomDhu
Crot:ldul'tl ru81Uitl 326 19.6 42 19.8 41 I l 42 16 8,7 299 8! 34 28,3 6 1
CroclduN $UIIIIf!Oietu 66 4,0 6 1 .6 6 0,7 5 4,2 4
SunCVJ ftNICill 16 1 ,0 3 1 ,4 3 0,8 3 2,1 2 1 , 1 4 0,5 4 3,3
CHIROPTERES 1 0,1 1 0,5 1 0,3 1 0,6
RONGEURS 1 0,6
Glù tUt 1 0,6
Eliomy1 querdnus 2 0.1 1 0,7 3 0,4
Ckthrlonomy1 11t1rrohu
Mlcrotu1 •rwIU Mlcrotu1 tlfTntù 34 2.0 5 2,4 3 1 8,3 21 14.5 3 1 ,6 89 1 0,7 6 5,0 63
Mlcrotul IIIWIIU An>koM IIIPltbll 10 0,6 4 1 ,9 1 0.3 2 0,2 1 0,8 2
Pitymy11 duodedmcol/111111 35 2,1 16 7,6 79 2 1 .4 s 2,1 3 1 3,7 1 0,8
P1tymy1 py
Microtldile in<UtenniMs 3 1 .4 2 o.s 3 0,4
Apodemlll sp. 89 5,4 12 5,7 58 t s . s 32 22.1 4 2.2 1 03 1 2,4 45 !1..:!_ 4 1
Micromyl mlnutul I S 0,9 3 1 .4 1 3 1 ,6 1 0,8
Mus mucubu 64 3,8 10 4,7 9 2.3 6 4,2 1 2 1 ,5 1
Mru •''" 1 4 1 8,5 14 6,6 30 8,0 30 20.7 4 2.2 78 9,4 9
Mus sp. 65 3,9 4 1.9 33 8,8 2 1 .4 3 1 .6 I l l 13,3 1 9 1 5,8 30
&mu sp. 8 o.s 24 1 1 .3 2 1 . 1 7 0,8 2
Murldile ir\Mterminb 2 1 , 7
OISEAUX 25 I ,S 67 11.L 75 20. 1 4 2,8 142 77.6 58 7,0 2 1 ,7 I l S -REYTII.ES
AMPHffiiENS 684 !.L.l. 2 0,9 4 1 . 1 2 1 .4 8 1 ,0 2
TOT AL VERTEBRES 1 5 8 1 2 1 0 373 145 183 830 1 20 3 5 2
INVERTEBRES 82 S,O 2 0,9 1 0,3 3 0,4 1
TOT AL GENERAL 1663 2 1 2 374 145 183 833 1 20 353
diques dans la composition du régime. A i nsi, nous avons d'une part cherché à obtenir des l o ts volumineux (plus de 500 proies) et d'autre part prélevé dans un même gîtP des pelotes d'inégale fraîcheur et déposées en des endroits différents pour que l'échantillon comprenne d·es pelotes de plusieurs années.
Nous ne reviendrons pas sur les méthodes d'analyse des p elo tes de réj ection. Elles sont classiques et bien connues. Des clefs dichotomi ques permet tent la détermination des proies au niveau de l'espèce pour les mammifères (Chaline et al . , 1 974) . Ceux-ci ont donc été distingués spécifiquement sauf en ce qui concerne les genres Apodemus et Rattus. Nous renvoyons à l 'article précédent (Fons et al. , 1980) pour la discussion des problèmes relatifs à la
- 190 -
9
97
% n
0,6 16 '
2
17.3 1984 1 , 1 48 1
32
4
10 1
17,9 255
0,6 29
43
lOS
1 1 ,6 1566 2
41
0,3 89
2.5 I l 1
8,5 925 1:
0,6 ISO
78
J8,1 614 I l
0,6 30 ' 6032
0,3 I S ' 6047

ns le département des Pyrénées-0 rientales. 1 l e s les stations 1 à 1 9 sont p r i s es (' Il compte dans cet a rtic lc .
1 0 I l 1 2 1 3 1 4 1 5 1 6 1 7 1 8 1 9 2 0 2 1
1 0 1 1 4 9 309 S I S " ' 576 600 735 900 1000 1 365 1488
% % % % % % % % % % % %
0,:! 0.1
25 5 . 1 0,3 85 6.4 523 1 7 ,4 1 27 1 2 , 1 1 07 35,1 0.3 1 ,0
0.9 1 0 2.0 0. 1 60 4.5 391 1 3,0 38 3.6 66 19,9 0. 1
0, 1
0, 1
8 1 3,6 50 1 1 9 30.8 228 203 22.2 277 20,8 582 19,4 474 112. I l 3.3 0,9
0.8 0.9 0.8 1 0.1 82 6.2 2 0.2
1 .4 1 . 7 0.3 0.6 3 0.3 I l 0.8 0.4
0. 1 0. 1
0. 1 0,2
0.5 0.4
1 0 2,0 17 1 .3 0. 1 27 2.6 0,9 0.3 0.5
19 1 .4 0.3 28 2,7 2 1 6,3 0.6 3.6
1 .3 1 9 1 7 , 1 2 3 6.0 JO 6.1 70 7.6 1 98 14,9 783 26.1 73 7.0 22 6,7 15 1 ,5 1 . 0
1 . 1 0,2 0.6 0.5 0.1
0.5
0.2 0.8 1 . 5
0.8 23 1 ,7 0.2 0,2 94 9.1 14 7,2
27.6 23 19.0 34 30,6 203 52.6 160 32,3 604
449 lllJ. 676 22,5 257 24,5 87 26,3 1 3 1.3 2 1 ,0
0. 1
0,2 0,9 0,2 0. 1 43 3.2 0,3 0.1
6,0 8,5 2.7 18 4,6 19 3,8 1 0 1 . 1
3,8 3,4 12 3.1 0.2 0.5 44 3.4
0,7 1 ,0 0.1 1 0,1
0,2 0,3 1 .0
2.2 20 33,9 1 .8 1 .0 0,4 0.3 0.2 0.2 0.1 50 4,8 1 ,5
0. 1
59 I l l 386 494 9 1 6 1325 2998 1049 331 195 33
0.2 5 0,4 0.2 840 !Id- 1 6 1
59 I l l 386 495 9 1 6 IJJO 3005 1049 3) 1 1 035 1 94
détermination des rats e t mulots de l a Catalogne française. Nous devons l'iden tif ica ti o n précise des Mus à l 'aimable collaboration de Ph. O rsini .
RESULTA TS
A . - GÉNÉRALITÉS
La figure 1 situe avec précision nos localités de récolte tandis que le tableau 1 donne le détail des résultats des analyses depelotes .
- 191 -

Les amphibiens ne représentent qu'un pourcentage très faible des p roies de la Chouette, sauf à C anet (Station 1 ) où l'oiseau exploite visiblement les denses populations d'anoures de l'endroit . Son gîte est établi dans une ruine située au bord du grand étang côtier. Spécialisation ou simple opportunisme ? Touj ours est-il que les alentours de Canet sont bien connus pour leurs « pluies de grenouilles » : Petit et Lomont (1958) estiment à 450 000 le nombre de cadavres de Pelobates cultripes écrasés entre Canet-village et Canet-plage, lors d'une de ces p luies, en 1 956.
En règle générale, les oiseaux ne constituent j amais une part impo rtante du régime de l'Effraie (Uttendôrfer, 1939 ; Asselberg, 1 971 ; Saint Girons et Martin, 1 973 ; Glue, 1 974 ; De Bruij n, 1 979 . . . ) . C'est c e que nous constatons e n dehors d e l a p laine (St . 4, 7 , 10, 1 2, 13, 14, 15, 16, 1 7, 18 et 1 9) . Leur contribution au menu de Tyto alba y dépasse rarement 2 % du total des proies (cfr. Fig. 3a) . En revanche, dans la plaine du Roussillon, i ls p euvent constituer la base de son alimentation (St . 2 e t 8) , dépassant plus des trois quarts des proies à Espira de l ' Agly (St . 5) ! Comme l'indiquent de nombreux auteurs (Jackel, 1 891 in Schneider, 1 964 ; Uttendôrfer, 1 939 et 1952 ; Schmidt, 1 973 a et 1 973 b ; Glue, 1974 ; Herrera, 1 974 ; Saint Girons e t Thouy, 1 978 ; De Bruij n, 1979 ; Ruprecht, 1979) , les petits passereaux granivores e t notamment les moineaux domestiques, Passer domesticus, sont les oiseaux les mieux représentés dans le régime de l 'Effraie (Tableau Il) .
TABLEAU II
Les oiseaux dans le régime de l'Effraie (stations comme à la figure 1 ) .
Loca l i té s
Pro i es
ApM f.> p .
PaM vù6oJune..6
Pet i ts g ra n i vores ( * )
Pet i ts i n s e c t i vores
H,(r.undùùdae
Emb vùz-i.dae
TWtdM f.> p .
S.tWtrtM f.> p .
I ndéterm i nés
TOTAL
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2 1 3 1 4 1 5 1 6 1 7 1 8 1 9
1 - 5 3 - 2 - 2 - - - - - - - - -
5 6 5 48 - 1 4 58 - 1 33 4 7 1 1 7 2 0 - - - - -
9 - 4 - - - 33 2 - - 2 - - 3 -
3 - 8 - 3 - - - 1 6 1 1 - - 9 - 1 - 1 - - - -
- - - - - - - - - - - - -
9 1 - - 2 - 98 2 3 .. 4
25 66 75 4 1 42 58 2 1 35 6 1 4 2 1 2 0 2 4 2 3 3 6 1 -
( * ) p r i n c i pa l ement des Moi neaux ( Pllf.J f.! f.>p . )
- 192 -
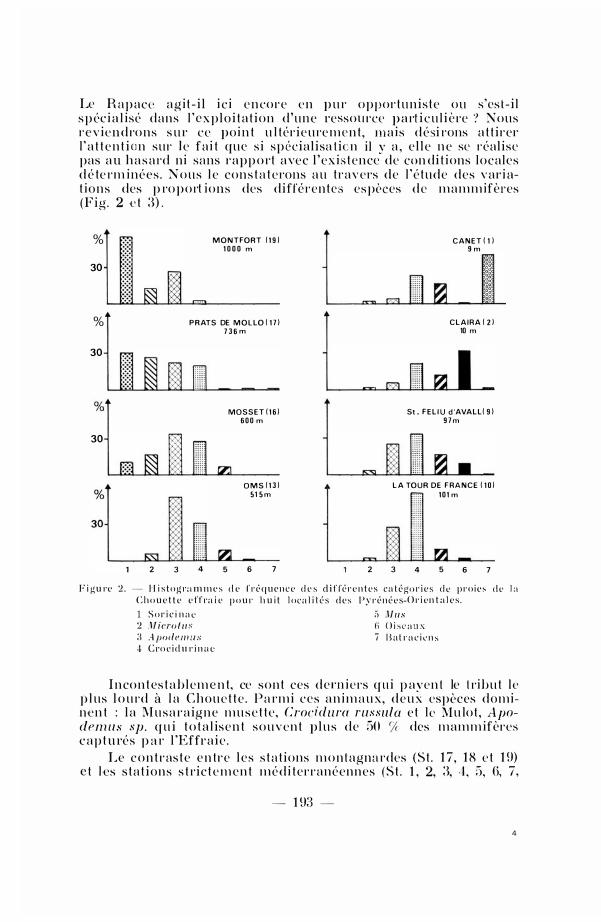
Le R a p ac-e a g i t - i l i c i e n core e n p u r oppor tu nis te ou s 'es t - i l s p éc i a l i sé dans l ' e x p l o i t a t ion d 'une ressou rce p a rt i c u l i è re ? Nous r e v i e n d ro n s s u r ce po i n t u l tér ie u re m e n t , m a i s d ési rons a t t i rer l ' a t t e n t i o n sur l e f a i t que s i sp écia l isa t ic n i l y a , e l l e n e sc réal ise pas a u h a s a rd ni sans rap p o rt a vec l ' ex i s tence de con di t i ons loca les d é te rm i nées . N o u s l e con s t a t e rons a u t ravers de l ' é tude des var i at i o n s des p ro p o rt i ons des diffé rentes es pèces d e m a m m ifères (Fig . 2 c t 3) .
%
30
%
30
%
30
3:! &'SJ 1 2 3
M O N T FO R T ( 1 9 )
1 0 0 0 m
P R AT S DE M O L L0 ( 1 7 )
7 3 6 m
. . . . 4
M O S S E T ( 1 6 )
6 0 0 m
O M S ( 1 3 )
5 1 5 m
5 6 7
l c::> 2
C A N E T ( 1 ) 9 m
o' o' o' o' o' o' o' o' ,o ,o ,g
C L A I R A ( 2 ) 10 m
St . F E L I U d "AVA L L ( 9 )
9 7 m
L A TO U R D E F R A N C E ( 1 0 ) 1 0 1 m
1 -3 4 5 6 7
F i g u re 2 . - H i s t o g r:l ln m c s C h o u e t t e c ff ra i e
d e f r é q u e n c e d e s d i f fé r e n t e s cn t é go r i c s d e p r o i e ,; p o u r h u i t l oc a l i t é s d e s Py r é n é c s-ü r i e n t n l e s .
d e 1 :1
S o l" i c i n a 10 2 ,\l iera / u s 3 A pode m u s C t·o c i d u r i n n c
5 Mus 6 O i s t• n u s 7 B a t r a c i è n s
I n co n t es t a b l e m e n t, ce sont ces d e rn i e rs q u i p ayen t le t ri b u t l e p l us l o u rd à l a C h o u e t t e . P a rmi ces a n i m a u x, d e u x espèces dominen t : l a M u s a r a ig n e m u s e t te , Crocidura russula e t l e Mu lot , Apodem u sp . q u i t o t a l isen t s o u v e n t p l u s de 50 % des mamm ifères c a p t u rés p a r l 'Effra ie .
Le c o n t r as t e e n tre l e s s t a t ions m o n t agna rdes (St . 1 7 , 1 8 e t 1 9) e t les s ta t ions s t ric t e m e n t m é d i terranéennes (S t . 1 , 2, 3, -l , 5, 6, 7,
- 1 93 -
4

A Il 1 E G E
/,-· · - - , _ "'\ . . . . _ .. / · · ·- · · · ·
.r - - · ,--1
( )._ ,
E S P A G N E
Figu re 3. - V a r i a t i o n s d u régi m e d e l 'Eff r a i e d a n s l e s Pyré n é e s- O r i e n t a l e s .
a . Pource ntage (en n o i r ) des O i s e a u x sur l 'en sembl e d e s p r o i e s .
E S P A G N E
Figu re 3h. - P o u r c e n t a g e ( e n n o i r ) d e s Crncir lu ra sur l 'e n s e m b l e Sorex + Crocidura.
- 194 -

1 Il 1 fC (; f
f 1 , • . / (; .Y f
F i gu re 3 c . - P o u r c e n t a ge ( e n n o i r ) de .ll u s spre l u s su r l ' c n se m h ! c M u s spre t u s + M . 111 / t s c u / u s
A li 1 E G E
' ' - \
,... _ _ , -- -- ..... - , / ' ....... ' / ........ _ _ _.,.
E S P A G N F
F i gu re 3 d . - P o u t·c c n t a ge ( e n n o i t · ) d ' A p o d e m u s su t· l ' e n s e m b l e M u s + A p o de m u s .
- 1 95 -

8, 9, 10, 1 1 et 13) mérite d'être signalé . Les p remières sont caractérisées par la présence de Microtus arualis, de Clethrionomys glareolus et l'abondance des Musaraignes du genre Sore:x: (Fig. 3b) ; les secondes, par la présence de Pitymys duodecimcostatus et surtout de Mus spretus (Fig. 3c) . Les stations 12, 14, 1 5 e t 16, situées aux confins du biome méditerranéen se singularisent par leur faune mixte : les premières Sorex apparaissent à Caudiès (12) e t à Montbolo (15) . A Sournia (14) , Mus spretus est touj ours présente m ais les Sorex gagnent en importance, alors que Clethrionomys glareolus et Pitymys pyrenaicus (1 ) font leur premièreapparition. A Mosset (St . 16) , Mus spretus a disparu, Microtus arualis apparaît e t l'élément médi terranéen est rappelé p ar Pitymys duodecimcostafus, espèce sans doute favorisée par les importantes surfaces cultivées aux alentours de ce village.
Nous retrouvons également cette opposition dans la figure 3d : dans la plaine, les souris sont nettement prépondérantes par rapport aux . mulots (St . 1 , 2, 3, 4, 5, 6, 8 e t 9) tandis qu'elles sont beaucoup moins nombreuses dans les co llines boisées (St . 7, 10, 1 1 , 12,13, 14, 1 5 et 16) et quasi absentes des localités montagnardes (St . 1 7, 18 e t 19) .
Nous ne nous attarderons p as à l 'examen des variations de la proportion des différentes catégories de proies d'une localité à l'autre. I l n'est d'ailleurs pas facile de les relier immédiatement à l'influence de facteurs écologiques donnés : ceux-ci sont généralement imbriqués et saisir la complexité de leurs interactions nécessite le recours à des procédés qui permettent de les considérer simultanément. L'analyse multidimensi onnelle s'avère être en pareil cas un outil très précieux .
B . - ANALYSE STATISTIQUE
1 . MÉTHODOLOGIE
A l'exception du lot d'Ille-sur-Têt (59 proies seulement) les échantillons de pelotes de réj ection d'Effraie ont fait l'obj e t d'un traitement statistique en deux étapes ; une analyse des correspondances portant sur 45 variables a été complétée p ar une recherche des corrélations entre variables « animales » e t variables du milieu suivant des procédés plus classiques .
Le tableau III énumère toutes ces variables e t indique dans quel(s) programme(s) elles sont intervenues.
En plus de nos relevés de terrain, divers documents ont été consultés afin d'obtenir les renseignements relatifs aux vari ables en question :
(1 ) Nous avons préféré con server la d é n o m i nat ion P. pyrenaicu. plutôt que d'uti l i ser P. gerbii, comme l e propose Spitz ( 1 978) pour des ra isons de pure commodité (concordance avec l 'art ic le p•·écédent) .
-- 196 -

TABLEAU III
Variables utilisées dans les analyses statistiques .
Prog ramme s Vari a b l e s Tai pa. eww pa.ea * SoJtex " aJu:trte.u6 " SoJtex m-i.l'lU.tuo Neomy6 6 p . CJtoudWta Jt.U.,66u1a CJtoudWta 6 uaveolen6 Sunc.M e..tluw c.M œ.M gw EUomy6 que1tu11M Clethlt-i.onomy6 glaJteolM M-i.c.Jto.t:tu. aJtvai-i.6 M-i.c.JtotM agJtUW M-i.c.Jto.t:tu. l'!-i.vai-i.6 AJtv-i.c.ola 6ap-i.dU6 P-i.tymy6 duodec.-i.mc.o6tatu:6 P-i.tymlj6 pyJtena-i.c.M ApodemM 6 p . M-i.c.Jtomy6 m-i.l'lU.tuo MM mM c.ulM MM 6 pJtetu:6 MM mM c.ulM + M . 6 pJtetM MM / ApodemM + MM RattM 6 p .
O i seaux Amph i b i en s Sé r i e phyto soc i o l og i q ue de :
QueJtc.M -<.lex QueJtc.M pub u c. en6 ( s t . no n édaph i ques ) FagM 6-i.lva.tic.a PinM 6 lflvutlti6 Alnu6 glut-i.no6a ( bord des eaux )
AJtu:ndo do nax Forêt Ma qu i s Vergers V i g nes Champs et prés Ha i es L i s i ères F l euve Etang Ma ra i s Terra s s e s d e c u l tu re Superf i c i e b o i sée Superfi c i e cu l t i vée Ru i nes L i s i ères + ha i e s Rel i ef A l t i tude Ha b i tat huma i n ** C l i mat *** * : P/A = Présence ou Ab s e n c e** : voi r § 1 . 2 . 4 . *** : vo i r Tab l ea u 4
MULTM
P/A P/A P/A P/A
> ou < 2 5 % P/A P/A P IA P /A P/A P/A
> ou < 1 0 % P/A PIA P/A P/A
> ou < 1 5 % P/A P/A P/A
> ou < 10 %
P/A > ou < 5 %
PIA
P/A P/A P/A P/A P/A P/A P/A P/A P/A P/A P/A P/A
PIA P/A P/A P/A
P/A
Pl a i nes /col l i ne s > ou < 500 m . > ou < 1 0 sec : humide
- 197 -
BMDP2R
+ + +
+
+
+
+ +
+
+ + + + + + +
+ +
+
+ +

les cartes I .G .N. au 1/25 000 ; la carte de la végétation de France, feuille de Perpignan (Ga ussen, 1 972) ; l 'atlas de la Catalogne Nord (Becat, 1 977) ; les annales climatologiques des Pyrénées-Orientales de 1 966 à 1977 ; le dictionnaire des communes de France (Anonyme, 1 973) .
1 . 1 . L'analyse des correspondances
Le programme M.U.L.T.M. (Lebm·t et al., Hl7 ï ) n e traite que des variab les catégorisées. Compte tenu du nombre trop faible de n o s relevés , nous n'avons pu affectet· plus de deux modalités à chacune des va riables retenues . Cette s impl ification a certes entraîné une perle considérable d' informat ion . N ous pensons toutefo i s l 'avoir atténuée par l e second volet du traitement .
1 .2. La recherche des corrélations
Afin d'ê tre à même d'employer les habituelles méthodes de calcul, nous devions préciser pour chacune des stations la valeur de chaque descripteur du milieu que nous avions choisi . Nous décrivons brièvement dans les lignes qui suivent la manière dont nous avons procédé pour obtenir ces valeurs.
1 .2 . 1 . La surface occupée par la forêt, le maquis, les vergers, la vigne et les champs
D'après Hosking et Newberry (1945) , les Effraies ne chassent généralement pas à plus de 1 500-2 000 rn de leur aire. En revanche, Geroudet (1965) estime à 100-200 h a (soit un rayon de env. 600-800 rn) l 'é tendue de leur domaine de chasse. Selon Glutz Von Blotzheim et Bauer (1 980) , le rayon d'action de l 'oiseau serait de 800-1 500 rn dans les conditions habituelles. Nous avons donc choisi de mesurer ces paramètres dans une zone d'étendue intermédiaire entre ces valeurs, à savoir sur une surface d'environ 710 ha (rayon = 1 500 m) centrée sur l e point de récolte des pelotes, endroi t correspondant le plus souvent à la situation de l'aire ou du gîte principal de la Chouette.
Sur carte I .G.N. , nous avons évalué la surface occupée p ar chaque type de couvert végétal à l 'aide d'une grille dont l a maille, relativement serrée (5 X 5 mm) a p e rmis des estimations précises.
1 .2.2. La longueur des /isieres e t des haies
Ces longueurs ont été mesurées dans la même zone de 1 500 m de rayon au moyen d'un curvimètre. Par lisières, nous entendons limites entre étendues boisées (maquis, forêt) e t cultures (vignes, vergers, p âtures e t champs proprement dits) .
Parfois, il était souhaitable de regrouper deux de ces variables à l'intérieur d'une troisième. Nous avons ainsi créé trois nouvelles variables dont la signification écologique n'est pas moins évidente que celle des autres :
- 1 98 -

superficie boisée (maquis + forêt) ; ensemble des haies et lisières ; surfaces cultivées : champs, prairies e t vergers (1) .
1 .2 .3 . L'altitude
L'altitude exacte du point de récolte des pelotes a été lue sur les cartes I . G.N.
1 .2.4. L'habitat humain
Pour évaluer l a concentration de l'habitat humain, un indice exprimant le rapport entre le nombre de maisons de la commune et le nombre de carrés de notre grille (§ 1 .2.1 . ) occupés par des habitations a été calculé .
1 .2 .5 . Le climat
Pour chaque localité de récolte des pelotes, le coefficient ombra-thermique d'Emberger (1942) (2) a été calculé à partir des données des stations météorologiques les plus proches ou en extrapolant les courbes isohyètes lorsque nos points de récolte étaient situés dans des régions soumises à un autre régime des pluies que celui de la s tation retenue. Le tableau IV indique les résultats de ces calculs de même que le nom des stations de référence.
Ces données ont été soumises au programme BMDP2R de corrélation p as à pas . Ce type de programme présente un net avantage p ar rapport à d'autres où les variables explicatives doivent être déterminées a priori puisqu'à chacune des étapes du calcul, la variable explicative choisie est la plus corrélée avec la variable dépendante, abstraction faite de l'influence des variables déj à sélectionnées lors d'une étape précédente.
2 . RÉSULTATS
2 .1 . L'analyse des correspondances
Les trois premiers axes factoriels extraits p ar l 'analyse ont des valeurs propres respectives de 0,394, 0,102 et 0,078. I l est mal aisé de tester leur signification. Lehart et al. (1977) ont calculé des abaques par simulation d'après lesquelles seul notre premier axe aurait une valeur propre significative. Toutefois, comme ces auteurs se plaisent à le répéter, ces valeurs donnent une idée très pessimiste de la part d'information que contiennent les facteurs extraits. Benzecri et coll . (1980) partagent cet avis . Ils écrivent même qu'en analyse multidimensionnelle, tout ce qui est interpré-
(1 ) Dans l a p la ine , en effet, i l a iTive souvent que le sol des vergers soit a ffecté à la culture maraîchè1·c. Com m e ce n'est pa s le cas d es vignes, nous ne les avons pas inclues dans ce regroupement.
(2) Q = 1 00 P jM'-m', où P i ndique la pluvio sité en mm, M, l a moyenne max. du mois l e plus chaud et m, l a m oyenne mini . du mois l e p lus froid.
- 199 -

TABLEAU IV
Calcul du coefficient d'Emberger (Q) .
Local i té s
1 . Canet *
2 . C l a i ra +
3 . Cornei l l a del Vercol +
4 . Arge l è s s u r Mer *
5 . E s p i ra de 1 ' Ag l y +
6 . Péz i l l a l a R i v i ère *
7 . Sorède *
8 . Thu i r *
9 . St . Fe l i u d ' Av a l l *
1 0 . La to u r de France *
1 2 . Ca ud i ès de Fen . 0
1 3 . 0ms o
1 4 . So u rn i a
1 5 . Montbo l o
1 6 . Mo s s e t
1 7 . Prats d e Mol l o
1 8 . Rabou i l l e t
1 9 . Co l d ' Au s s i ères
Sta t i on météoro l o g i q u e pl u v i omètre t h e rmomè tre
Cane t - p l age
St . L a u rent de la S a l .
V i l l e neuve de l a Raho
Arge l ès ( Va l my , C a p Bea r )
R i v e s a l te s
Le So l e r
Arge l ès ( Va l my )
Ste Col ombe
L e Sol er L a tour de France
Caud i è s ( + i s ohyète s )
L l a u ro
Sourn i a
i sohyètes
Mosset
Prats de t4ol l o
i s ohyètes
i s ohyè tes
C a ne t - p l a g e
S t . L a u r e n t de l a S .
C a n o h è s
P o r t - V e n d r e s
R i ve s a l tes
L e So l e r
L e . Bou l ou
C a n o h è s
Le So l e r
M a u ry
M a u ry
Rodès
S o u rn i a
Rodès
N o h è d e s
L a L l a u
Nohèdes
N o h è d e s
rn t 0 moyenne du moi s l e p l u s fro i d .
Q rn
7 2 , 6 3 , 2
6 3 , 1 4 , 3
6 1 , 0 3 , 8
1 1 4 , 9 3 , 8
5 4 , 9 4 , 3
89 , 2 3 , 1
1 0 1 , 0 2 , 5
86 , 0 3 , 8
89 , 2 3 , 1
8 2 , 2 3 , 1
1 1 7 , 9 3 , 1
9 1 ' 1 0 , 8
1 2 1 , 9 0 , 7
1 2 0 , 5 0 , 8 1 1 1 , 6 0 , 2 1 8 5 , 4 - 1 , 2 1 2 7 , 4 0 , 2 1 4 3 , 6 0 , 2
+ *
c l i ma t de type sem i - a r i de
c l imat subhum i de à h i ver tempéré S t a t i on s regroupées p o u r l ' a n a l ys e des c o r r e s p o n d a n c e s s o u s la dénom i na t i on :
. . c l i ma t s e c . . .
c l i ma t s u b h um i de à h i ve r fra i s
c l i mat hum i de S ta t i on s regroupées s o u s l a dénom i n a t i on :
" c l i ma t h um i de " .
table est valide (p. 479) et font appel au bon sens des utilisateurs de la méthode (p . 227) : « Pour l'honnête homme qui ne se pl ie pas au rictus des techniques, cet o util ne demeurera-t-il p as touj ours le premier ? »
Nous tenterons dès lors l'interprétation de ces t rois axes.
2 .1 . 1 . Le plan des facteurs 1 et 2 (Fig. 4) .
Si l'on proj ette dans ce premier plan la position de chaque station de récolte, il apparaît qu'à l'exception de Caudiès (St . 12) , elles s'alignent grossièrement le long d'une p arabole. C'est la figure que l'on obtient habituellement avec des suites ordonnées (effe t Guttman, voir Benzecri et coll . , 1980) . Nos s tations se succèdent donc le long d'un gradient matérialisé p ar le premier axe. Quelle est l a nature de celui-ci ? En fait , il est construi t à partir de plusieurs variables qui, par ordre d'importance de leur contribution
- 200 -

au calcul de cet axe son t : climat, altitude, présence de la Canne de Provence ( A rundo donax) , des musaraignes pygmée et carrelet (Sorex minutus et S. « arane us » ) de la série du Chêne pubescent (Quercus pubescens) , de l 'abondance des souris, de la présence de la série du Pin sylvestre (Pinus syluestris) , du Camp agnol roussâtre (Clethrionomys glareolus) , de la vigueur du relief, de la concentration de l'habitat hum ain, de la présence de vignes, du Campagnol des champs (Microtus arualis) , de la Souris sauvage (Mus spretus) , du Campagnol souterrain des Pyrénées (Pitymys pyrenaicus) et enfin, de la présence des étages du Hêtre (Fagus siluatica) e t du Chêne vert (Quercus ilex) .
Aux deux extrémités de la parabole se situent donc les sta tions les plus différentes. Le contraste mis en évidence ici n'est au tre
t I:J,. \1:JY't
* MUS ffiJ::C ;Ll! • • • • • • • •
Til1J-.'ISS!:S • • • -0• •
.· r . ouontt • . * COIJ.J'I.;
Q. PlJBJ:<'CtNS • 0· .·/:. t 10
> ':A:n m . .
CL!Yo/\1' 1 1\ IM I OE * llJS
P . PYiNA IC IJS
.
t:. '. :•:-•a.s
S . ARAMU:;. • s . H J Nurus
. !'!NUS • c GLAil(OLUS
A SI lJIJS
e r.t. t s
• M. N : VAL I S+- A Q . l U:X
M . ARVAL I S i MUS SPiifTUS ,/ .. 1'"'-lli
!l'•s•ATRI\( J ' tiS
"""' MH.S l> Y.;/ * TALPA O I SlA!JX
*
RlJ (Nffi •
*lo:A11l'S /::l. r · :DNF.
.APOO(MUS
l l l OMY *
m c Rt..vs <§>
( RUSSULA *
C . SUAVEOl ENS *
M . AGRCST J S • ! NEOHYS
• !
' .. I C . AGRES T J S rf IWVI.lS "'"""
• • • • • • ACIIAMPS
'0 . . !!rotu;s SUNCUS .
C II:USSULA
M .m!t. !Silo: C.l i S *
(Î> HUS S':j'TU; '1> FACl.t;:;M.AU S •
• A . SAP I OUS
C . Gl.f,R[Ol11S •_xnus MI CROHYS . () . !LEX * • • · . ARANEUS
·. * El ! OMVS .
• 'JI. S .M I NUTUS VI
• I!OIS A *
P . r tRD!o\ [ CUS
R.AiTUS <î> • • <!>
H'JS HUSC.ULUS : J>IO. A
.:'J.!"l.IBESC!NS :4w3J1s • nM
IWJVE 0
1\:N.JS•
• :. $"' . OUOOEC . :POOEMUS * : PlAINE A
BATRAC I ENS •
O I SlAUX •
F igure 4. - Représen tation des p 1·oj ect ions des observations sur l es facteurs 1 et 2 ( F 1 et F 2 ) de l 'analyse des correspondances . Les stations sont dé signées par
• *
e spèce présente espèce a b sente
leur n • < F ig. 1 ) .
• : descripteur présent/1 : descripteur absent (voir Ta b . I I I ) .
Des traits sont placés sur les axes à l a valeur 0 ,5 .
- 201 -

que celui que nous signalions avant d'aborder la partie statistique . D'une part, nous avons les stations de la plaine méditerranéenne (étage du Chêne vert) (climat chaud, h abitations agglomérées en villages-noyaux, vignobles étendus) où le régime de l'Effraie se singularise par le rôle qu'y j o uent les Souris (essentiellement Mus spretus) ; d'autre part des localités au climat plus frais, d'altitude plus élevée et au relief plus tourmenté : moyenne montagne où l'on rencontre les séries du Chêne pubescent, du Hêtre et du Pin sylvestre et où l'Effraie se nourrit des musaraignes du genre Sorex, de campagnols des champs, de campagnols roussàtres et de campagnols souterrains des Pyrénées.
Sur le plan faunistique, nos observations, tou t à fait semblables à celles de Heim de Balsac et de Beaufort (1966) , de Saint Girons et Vesco (1 974) et de Fayard et Erome (1977) , confirment nos conclusions antérieures (Fons et al, 1 980) : l e biome méditerranéen est caractérisé par la présence de Mus spretus, l'absence de Clethrionomys glareolus, de Microtus arvalis e t des Sorex.
Nous pouvons cependant apporter des nuances à ce portraitrobot : certaines variables, malgré leur intervention limitée dans la constitution de l' axe, n'en réagissent pas moins à l a gradation qu'il traduit. L'abondance des oiseaux dans le régime de l'Effraie de même que la présence du Rat des moissons (Micromys minutus) et du Campagnol provençal (Pitymys duodecimcostatus) , sont des traits communs aux stations plus méditerranéennes, tandis que la présence de terrasses de culture, de ruines i solées e t l 'abondance des mulots, sont plutôt typiques des localités situées dans les collines subméditerranéennes plus boisées, qui font la transition entre la plaine et l'étage montagnard.
Le second axe exprime également l 'opposition entre le p aysage des collines couvertes de maquis ou cultivées en terrasses et celui de la plaine avec ses vergers, ses haies e t ses cours d'eau bordés d'aulnes (A /nus glutinosa) .
Selon le schéma explicatif de l'effet Guttman, les points situés au voisinage du sommet de la parabole occupent une position moyenne . Nous qualifions donc les s tations 10, 13 , 14 et 15 d'intermédiaires . Mais elles ne doivent p as cette qualité aux mêmes raisons, aussi leurs proj ections ne se font-ell es pas au même droit dans le plan F1-F2. Latour de France (St . 10) est située au pied des premières collines et c 'est plutôt le type de p aysage qui influence la posi tion de sa proj ection : la faune et l a végétation y sont strictement méditerranéennes. La faune d'Oms (St. 13) est méditerranéenne mais cette station j ouit de conditions écoclimatiques qui la rapprochent de l 'étage subméditerranéen ; le cortège du Chêne pubescent y fait d'ailleurs son apparition . La tour de France e t Oms occupent donc une position intermédiaire beaucoup plus en relation avec leur p aysage qu'avec leur faune en petits mammifères, si ce n'est peut-être par l'abondance des
- 202 -

mulots e t des Crocidurinae dans les pelotes, point sur lequel nous reviendrons . Il n'en est p as de même pour Sournia (St. 14) et Montbolo (St . 1 5) où, nous l' avons vu, les éléments faunistiques méditerranéens rencontrent les montagnards. La position de Montbolo à l'intérieur de la p arabole semble indiquer que cette localité réunit plus d'éléments contradictoires que Sourni a.
Nous avions également considéré Caudiès (St . 12) et Mosset (St. 16) comme stations de transition (§ A) . On peut dès lors s'étonner de leur proj ection sur le plan des deux premiers axes factoriels. En fait d'éléments méditerranéens, i l n'y a, à Mosset, qu'un peu de végétation de la série du Chêne vert et le Campagnol p rovençal. Ces espèces débordent toutefois largement l 'étage méditerranéen strict ; elles ne peuvent donc être retenues comme critères discriminants absolus de ce dernier (Saint Girons et Vesco, 1 974 ; Fayard et Erome, 1 977 ; Fons et al , 1980) . Tous les autres éléments font de Mosset une station méditerranéenne extrême, voire plutô t montagnarde. Les influences euméditerranéennes y resten t p erceptibles : les étages de végétation s'y succèdent très rapidement dans l 'espace : i l s'agit d'une vallée assez étroite où le versant en soulane contraste très fortement, notamment par ses pelouses de la série de Chêne vert, avec le versant en ombrée au sommet duquel p oussent hêtres et pins sylvestres. Caudiès doit enfin sa p osition à la présence d'éléments très contradictoires qui attirent sa p roj ection vers chacune des extrémités de la parabole .
D ans cet échantillon, on retrouve en effet des représentants de la région méditerranéenne (Mus spretus et Quercus ile.x, absence de Microtus arvalis, de Clethrionomys glareolus , de Sorex « araneus » ) ou des caractéristiques de la plaine (altitude faible, pas de terrasses de culture) et des traits de l'étage montagnard (Sorex minutus , Pinus sylvestris) ou des collines (pas de Micromys minutus ni de Pitymys duodecimcostatus, faible proportion de Mus, abondance des Apodemus, climat moins sec) . Ne nous étendons pas sur ce cas. I l est très probable que la position de Caudiès eut été modifiée si nous avions pu disposer d'un échantillon de pelotes plus important . La présence de certaines espèces se serait sans doute révélée à la faveur d'une analyse portant sur un plus grand nombre de proies .
I l reste à éclaircir le cas des Crocidurinae e t de Microtus agrestis dont on remarque l'opposition sur F2. Les premiers occupent une posi tion semblable à celles des locali tés des collines sèches cultivées en terrasses. Fons (1 975) souligne l' affection de ces musaraignes pour les murets de soutènement des terrasses. Les stations 1 0, 14, 1 8, 7, 13, 16 et 17 où existent pareils murets en p ierres sèches non cimentées se distinguent par le pourcentage élevé des Crocidurinae d ans le régime de l'Effraie. Crocidura russula est la proie principale du rapace à Rabouillet (St. 18) où C. suaveolens et Suncus e truscus sont encore présentes. Ces faits
- 203 -

ne sont certainement pas sans rapport avec l 'abondance des murets à cet endroit. Bien que l 'altitude y soit plus faible qu'à Rabouillet et qu'il y ait aussi des murets à Prats-de-Mollo (St . 1 7) , C. russula y est le seul Crocidurinae et n'y j ou e pas un rôle prépondérant dans le régime de l'oiseau. Le climat de cette localité est vraisemblablement trop humide pour les deux autres espèces.
Le Campagnol agreste se trouve proj eté au voisinage de Neomys et de Talpa. On sait que Neomys vit exclusivement dans des milieux très humides ou au bord des eaux et on connaît l 'affinité de la Taupe pour les sols profonds. Le second facteur exprimerait donc aussi le contraste entre milieux secs au sol peu profond des collines et milieux hum des des prairies de moyenne alti tude, des fonds de vallée ou de la plaine (fleuve, canaux d'irrigation, sols alluviaux récents) . La position des points « Batraciens » , « A lnus » et « Fleuve » viendrait à l 'appui de cette hypothèse . Il se peut que ces espèces se voient de la sorte proj etées entre les deux branches de la parabole parce qu'elles répondent à la fois à certaines variables qui s'expriment dans la p laine e t à d'autres plus typiques de la montagne.
2.1 .2. Le plan des facteurs 1 e t 3 (Fig. 5)
Le troisième axe doit s'interpréter en termes de qualité de l'échantillonnage. En effet, selon que le nombre de proies d'un lot de pelotes dépasse ou non 600, la proj ection de la localité de récolte se fait en-deçà ou au-delà de l 'origine de F3. D eux exceptions : Montbolo (St. 15) e t Claira (St . 2) dont les coordonnées sur F3 demeurent cependant fort proches de zéro (respectivement 0,038 et - 0,01 6) .
Il est intéressant de constater qu'il existe une corrélation très significative (r = 0,799 ; p < 0,01) entre l a taille de chaque échantillon (log. du nombre de proies) e t sa position sur F3. On peut également voir que le nombre d'espèces d 'un prélèvement dépend, bien entendu de la richesse spécifique de l 'endroit de récolte, mais également de la taille de cet échanti l lon. Cela s·c traduit par des courbes de saturation telles que celles obtenues pour C anet (St . 1 ) et pour Olmi-Capella (Haute-Corse, voir Libois et Vranken, 1981) . Comme l'indique la figure 6, moins la r : chesse spécifique de l'endroit est importante - la Co rse ne compte p as plus de huit espèces de petits mammifères terrestres - plus vite on arrive à saturation : ainsi, 566 proies sont nécessaires pour l'attei ndre à Canet, contre 140 à Olmi-Capella . La diversité pourrait également j ouer un certain rôle.
En ce qui concerne les Pyrénées-Orientales, en dessous d'un total variant de 300 à 600 proies, i l y a de fortes chances que l'échantillon ne soit pas suffisant pour révéler la présence de toutes les espèces « rares » , rareté due à la faible densité de l'espèce sur le terrain (cas d'Eliomys quercinus et des. Neomys) , à sa stricte localisation à un milieu p articulier d'étendue res-
- 204 -

treinte (cas de MicroltLS nivalis pour les éboulis ensoleillés, de Micromys minutus pour les roselières et de Mus musculus pour les abords des habitations) à son mode de vie la mettant à l'abri de l'Effraie (arboricole : Glis glis , hypogé : Ta/pa europaea) , ou résultant du dédain de l'oiseau à son égard : Arvicola sapidus . et Rattus sp. dont la trop grande taille décourage sans doute l'Effraie.
Nous ne sero ns donc p as surpris de vérifier que ce sont précisément ces espèces (hormis la Taupe et le Loir) qui interviennent le plus dans l 'édification de F3 et que la proj ection du point représentant leur présence a une abcisse négative sur cet axe.
2.2. Les corrélations
Le tableau V reprend les résultats des calculs effectués par le programme BMDP2R pour les variables figurant au tableau III . Pour chaque variable animale, nous y avons fait figurer l'équation de la droite d 'aj ustement calculée, son coefficient de corrélation multiple (rm) ainsi que le stade et le seuil de signification correspondant à l 'entrée dans l'équation des différentes variables du milieu .
. S . H I NUTUS • Pms
Hus'\uscuLus
su*
us
A . S
I DUS
P . OUOOEC . •
OOU.INES
0 tJ,. I < i O > 500 M
• P . PYRENA I CUS
A . OCW\X
• lJ. VlœES S . ARANEUS
• C . GLAREOLUS
......, à o. nn
-- PRETUS H . ARVAL J S
• MI'QJIS
lJ. IWES
. H . N I VAL I S
!
0 * (. . RUSSULA
E L I OHVS *
*'t1 J t ROMYS
15
NEOMYS •
il- rH VAl !S NEID1YS * /'.0
II/I.JES • '-.;/
c . SUAVEOLENS
J\1J<5 •
scus M . AGREST ! S
•• HUS MUSCULUS
C . RUSSULA
VEIŒRS .
"'ï's
r>It>U> l:J. * S . M I NUTUS ll f l
CLIMAT SEC P . OUODEC • lJ. PIArNE
./.\. Q.PUB 'V' /::.. * APOOEMUS
Mus e BATRAC I E NSe'V'
• A SAYI DUS
TALPA . 0
H I OHYS •
l:r. acns
RUINES ô
I I CROMYS .
Figure 5 . - Repré sentat ion des proj ection s des observat ions sur J e facteurs 1 ct 3 (Fl ct F 3 ) de l 'analyse des corre spondances. Légende comme à la figure 4 .
- 205 --

Notons toutefois deux restrictions importantes : Ces équations sont calculées à p artir d'un (trop) peti t nombre d'échantillons des Pyrénées-Orientales . I l serait imprudent de les utiliser pour toute autre p artie de la région m éditerranéenne et a fortiori de France, où l 'abondance des espèces est conditionnée par d'autres facteurs. I l serait contraire à la déontologie statis tique d'utiliser ces modèles à des fins prédictives, y compris dans les PyrénéesOrientales, puisque certaines des variables ne répondent pas à l a condition de normalité .
"· •P· "
100 500
O L M I Y : 1,8 +3,4 l o g X
Y : O,O II + 5,JZ l o 1 1
1000 n. pro ie•
Figure 6. - Courbes de saturation de deux échant i l lons e n pron•nance de régi o n s d e richesse spécifique diffé1·ente : R ouss i l lon et Haute-Corse ( v o i r texte ) .
Les valeurs observées sont représentées par d e s points .
Etant donné ces observations, il nous p araît préférable d'accorder plus d'attention au signe des coefficients apparaissant dans les équations qu'à leur valeur précise . Néanmoins, ces équations ont le mérite de mettre en relief l 'intensité de la l iaison en tre la composition du régime de l'Effraie et certains p aramètres du milieu de chasse de l'oiseau.
Crocidura russula serait d'autant plus abondante d ans le régime de l 'Effraie que la superficie des forêts est moindre et que la longueur des lisières entre milieux boisés e t milieux cultivés est importante .
La première observation - corrobore les résultats de Fons (1975 a e t b) qui a réalisé 31 % des prises de cette espèce d ans les maquis à Banyuls. Il peut cependant p araître surprenant qu'il ai t également trouvé un pourcentage élevé (23 %) de cette espèce dans l a forêt de chênes-lièges (Quercus su ber) . Une comparaison
- 206 -

TABLEAU v Résultats des calculs effect ués par le programme BMDP2R pour les variables figurant au tableau Ill .
SURrACES LONGUEURS L A
du. wp.teuJW F v c ,If v i l H 0 i h e x. .t c a h
du a " H i L i l b u Il 9 a x, q x, è
+ i .i.. rtl EQUATION de la VRO lTE
é 9 a x, 0 .t n rn u i " t a d ' AJUSTEoiiENT .t + i + e G u m " e p
a i" e e d a m 4 .t t Yl 4 4 4 e M pècu. x, x, x, x._ x, x, x, x, x, x , . x , 1
Croc i d u r a Y, 2 1 ' l n ( Y , / 1 - Y , ) = 0 , 188 X, - 0 , 1 9 6 l n ( X , / 1 - X , ) - 2 , 9 7 5 0 , 7 23
r u s s u l a 5% 1 %
C r oc i d u ra Y, A u c u n e corré l a t i on s i g n i f i c a t i ve
s u aveo 1 e n s
S u n c u s Y, 1 2 3 l n [ ( Y3 +2 ) / ( 1 0 0 - Y, + 2 ) l = 0 , 005 X, + 0 , 0 3 6 x , e t r u s c u s 1 % 5% 5% - 0 , 2 1 6 x, - 3 , 5 38 O , d2 9
M i c r o t u s Y, 4 3 2 1 l n ( Y, / 1 - Y. ) = 0 , 02 X 1 0 + 0 , 1 3 3 x, + 0 , 0 2 3 x , a g re s t i s 5% 6% 1 % 1 % + 0 , 0 1 2 x, - 5 , 858 0 , 9 0 1
P i tymys Y, 3 2 1 l n [ ( Y, +0 , 02 ) / ( 1 - Y, + 0 , 0 2 ) 1 = -1 , 303 x, duodec i mc . 2 , 5% 2 , 5% 0 , 1 % + 0 , 0 2 7 x, + 0 , 0 1 7 x. - 1 , 938 0 , 90 1
Apodemus Y, 2 1 l n ( Y, / 1 - Y, ) = 0 , 1 4 3 X1 + 0 , 0 1 4 X , - 2 . 764 0 , 863
2 , 5% <0 , 1%
tus Y, 4 2 1 3 ' l n[ ( Y, +0 , 02 ) / ( 1 - Y7 +0 , 0 2 ) l = -0 . 760 x, + 0 , 106 x,
1 % 0 , 1 % < 0 , 1 % 5% - 0 , 0 1 1 x . , + 0 , 2 1 5 l n ( X, / 1 - X, ) - 0 , 200 0 , 96 3 Y, /Y, +Y, Y, 1 2 l n[ ( Y8 +0 , 02 ) / ( 1 - Y8 +0 , 0 2 ) 1 = - 0 , 030 x.
<0 , 1 % 1 % - 0 , 0 1 7 X o o + 1 , 8 5 9 0 , 9 5 2
O i s e a u x Y , 1 3 2 l n[ ( Y, +0 , 02 ) / ( 1 - Y, +0 , 0 2 ) ] = - 0 , 0 1 6 x ,
<0 , 1 % 2% 1 % + o , 06 5 x" - o , 0 1 6 x , . - 0 , 5 19 0 , 908
N . B . 1 . L U> 4utr.ôacu. 6 o nt exptr.im u. en % de la 6 utr.ôace .totale 6 <W. Ô dart6 lu qua.tiort6 ptr.cédé.u. d ' un ' où ellu. 6 e .ttr.ouvent6 0114 la ôotr.me d ' une ôtr.ac.tion de l ' unité .
2 . La ptr.opotr..tio n du u. pèc.U> dart6 le 1t9ime ( Y ) u..t exptr.imée en % poutr. S u n c u s e t r u s c u s , en ôtr.ac.tion de l ' unité poutr. lu au.ttr.u. u.pèc.u. . Lu lon9ueutr.6 6 o nt exptr.iméu. en c.entainu. de mè.ttr.u. ( hm . ) .
3 . L u .ttr.aYl6 ôotr.ma.tiorl6 lo9aJti.thmiquu. ont poutr. but d ' vi.tetr. la ptr.écii.c.tion de valeutr.6 né9a.tiv u. .

demeure néanmoins bien difficile p uisqu'il n 'a intensément piégé que dans les collines de la Côte Vermeille toutes couvertes de murets. Il est probable que seules les forêts méditerrané·ennes installées sur d'anciennes terrasses de culture (Q. suber p . ex.) conviennent spécialement à la Crocidure. Que celte Musaraigne soit favorisée par les lisières n'a non plus rien d'étonnant. Fons (1 975 a et b) souligne leur importance pour Suncus etruscus. On sait que les frontières entre milieux (écotones) sont riches, tant en espèces qu'en individus. Ce fait est une fois de plus mis en évidence ici, puisque dans chaque équation proposée, hormis celle du Campagnol provençal, on retrouve un élément exprimant en quelque sorte l'intensité du découpage du p aysage végétal : lisières, haies ou les deux à la fois. Souvent, les h aies e t les lisières servent de refuges : lors des travaux des champs, les anim aux dérangés ne peuvent subsister qu'à la condition de pouvoir utiliser de pareils abris.
En ce qui concerne Crocidura sua veolens, nous n'avons trouvé aucune corrélation significative. Cette espèce n'est j amais très abondante, de plus sa présence irrégulière d ans les échantillons rend assez vaine toute tentative d'explication. Fons (1 975 a et b) l 'a principalement capturée dans les maquis de la Côte Vermeille, mais son interprétation reste prudente vu le faible nombre de ses captures . Sans aucun doute, les affinités de la Musaraigne des j ardins sont méditerranéennes (voir analys·e des correspondances) ma is pourquoi ne la trouve-t-on pas en Catalogne espagnole (Claramunt et al , 1975) alors qu'elle atteint la Picardie vers le Nord (Saint Girons, 1973 a) et occupe certaines îles de la Manche ?
Suncus etruscus manifeste ses affinités méditerranéennes (corrélation négative avec l'altitude) ; elle serait favorisée dans les paysages variés (influence des lisières, cfr. Fons, 1 975 a e t b) ainsi que dans les vignobles. Fons (1 975 a et b) constate sa préférence pour les terrasses abandonnées et explique son absence des vignes cultivées p ar l'entretien régulier apporté à ces cultures et tout particulièrement par l'emploi massif d'herbicides e t d'autres p roduits chimiques qui éliminent les arthropodes. I l ne nous a pas été possible de distinguer sous deux rubriques séparées, surface du vignoble en exploitation e t surface des vignes abandonnées : les cartes I . G.N. ne le permettent pas . . . On ne peut donc conclure que ces résultats contredisent les précédents, au contraire .
Microtus agrestis subit l'influence prépondérante du climat (Tableau IV) et est avantagé p ar les h aies, les cultures et dans une moindre mesure par les forêts . Ces variables sont ant agonistes deux à deux : les champs sont plus vastes dans la p laine e t les haies plus longues, alors qu'en altitude le climat est plus frais et les forêts plus étendues. Nous touchons ici à l 'explication de la position « entre deux chaises » de ce Campagnol d ans le plan F1-F2 de l'analyse des correspondances.
- 208 -

Pour s a p art, comme l'indiquent Sans Coma et al. (1976) , Pitymys duodecimcostalus montre son appartenance à la plaine (forte corrélation négative avec l'altitude) et ses préférences pour les zones cultivées (champs proprement dits e t vergers) , voire même boisées. Il es t vrai qu'en plaine, les lambeaux de terrains incultes et les bosquets doivent lui fournir un refuge au moment des travaux des champs, notamment des labours.
A podemus ne dément p as sa réputation d'animal plutôt forestier p ar opposition aux souris qui s'avèrent plus abondantes dans les milieux franchement o uverts. Les variations de la proportion de Mus reflètent en réalité celles de Mus spretus, espèce qui représente dans le biome méditerranéen 85 à 90 % des souris capturées par l'Effraie. La corrélation fortement négative avec l'altitude (déj à constatée p ar Martin et Vericad, 1977) et avec l'indice d'Emberger dénote la préférence de la Souris sauvage pour la sécheresse. Ces observations concordent avec celles de Fayard et Erome (1976) qui trouvent cette espèce dans des biotopes secs avec une strate herbacée abondante et avec celles d'Orsini (1979) qui constate que les collines sèches avec leurs murets et leu r végétation « maigre » lui conviennent bien.
Enfin, le pourcentage d'oiseaux est influencé p ar trois facteurs principaux : la longueur des lisières, la concentration de l'habitat humain et le climat. Ces facteurs rendent bien compte des observations de différents auteurs : Schmidt (1973 b) trouve que les moineaux sont plus abondants dans le régime des effraies des zones les plus arides de la Hongrie (plaines du Sud-Est) et dans les agglomérations ; pour Ruprecht (1979) , c'est surtout dans les milieux u rb anisés. C'est également en milieu urbain que Brosset (1 956) , Saint Girons et Thouy (1978) les rencontrent en grand nombre au Maroc. On connaît l'habileté des effraies à capturer les moineaux dans leurs dortoirs (Sage, 1962) e t l 'on sait que ces dortoirs sont de préférence établis dans les agglomérations de quelque importance. Pourtant, dès qu'elles en ont l'occasion, les effraies semblent se détourner de ces proies faciles en faveur des micromammifères . Sinon, comment expliquer les résultats de Saint Girons (1 973 b) e t de Saint Girons et Thouy (1978) . Comment expliquer aussi que dans certaines grandes villes le pourcentage d'oiseaux dans leur régime soit relativement faible : 8,6 % seulement contre 60,4 % de Crocidura russula à Liège, ville de 350 000 habitants où moineaux e t étournaux abondent ? (Libois, 1977) .
O tteni et coll. (1972) ont observé que même en présence de ressources alimentaires illimitées mais constituées par des oiseaux, les effraies se reproduisaient avec moins de succès que lorsque les rongeurs étaien t abondants. Ils estiment que la Chouette cesse de nicher dès que la proportion d'oiseaux dans son régime dépasse 32 % des proies . Les oiseaux sont donc bel et bien des proies de remplacement suppléant occasionnellement la pauvreté du milieu
- 209 -
5

en petits mammifères . Gusev (1952) de même que Sans Coma (1 974) ont constaté que l'Effraie ne prenait un nombre élevé d'oiseauxque pendant et après un enneigement prolongé ; Doumeret e t Saint Girons (1977) ont fait la même observation pendant la période d'exceptionnelle sécheresse de 1 976.
Ceci est tout à fait compa tible avec la théorie de la stratégie alimentaire optimale (Curio, 1 976 ; Krebs, 1 978 ; H ainsworth e t Wolf, 1 979) : un prédateur ne capture des p roies de m auvaise qualité que si la densité de ses proies préférées est trop faible, la densité relative de la proie de remplacement n'ayant par co ntre qu'une très faible incidence sur le régime . Sans doute les régions arides, uniformes (peu de lisières) sont-elles nettement moins propices au maintien d'une faune dense e t diversifiée de petits mammifères que les pays plus humides et plus variés . Herrera (1 974 b) constate d'ailleurs un élargissement de la niche trcphique de l'Effraie en région méditerranéenne (où elle prend plus d'oiseaux) e t l'interprète comme une réponse du prédate ur à un appauvrissement de son éventail de mammifères-proies .
DISCUSSION E T CONCLUSIONS
« L'examen des proies capturées p ar la Chouette effraie fournit presque touj ours des renseignements u tiles ou simplement imprévus concernant la faunistique des m ammifères e t des oiseaux, sans oublier celle des reptiles, des amphibiens et même des arthropodes. Le seul reproche que l'on pourrait adresser à de telles investigations réside dans l'impatience de certains chercheurs à publier des notules vraiment insignifiantes, soit qu'elles évoquent un m atériel insuffisant, soit qu'elles concernent des régions par trop connues » (Heim de Balsac, 1965) .
Quels que soient les défauts de cette méthode, sans le concours de l'Effraie il serait inimaginable d'entreprendre une étude faunistique importante sur les micromammifères . Ce rapace dont l'éventail des proies va de la Musaraigne étrm:que aux rats, s'avère être le meilleur échantillonneur qui soit puisque les pièges ne p ermettent pas la capture de toutes les espèces présentes dans un milieu. Toutefois, les piégeages restent irremplaçables pour l'acquisition de renseignements précis sur le micro-habitat d'une espèce donnée.
Comme nous l'avons vu, de nombreux chercheurs ont tenté de mettre en relation les variations géographiques du régime de l'Effraie avec différents facteurs du milieu sans réaliser une approche globale du problème : chaque p aramètre était envisagé séparément. En fait, seule l 'utilisation d'une technique de description multidimensionnelle peut offrir cet avantage.
- 210 -

Par la diversi té de ses paysages, véritable carrefour de la flore et de la faune, le département des Pyrénées-Orientales est un des plus v ariés de toute la France . La carte de la végétation (feuille de Perpignan) , précieuse toile de fond pour l'écologiste, offre sur une distance de moins de 100 km, une p alette d'une remarquable richesse . Les principaux types de paysage y sont réunis, de la côte schisteuse aux hautes montagnes. L'extrême diversité des milieux naturels de ce département permettait donc l 'étude de zones très différentes sur une étendue très réduite.
En ce qui concerne notre travail, les traitements mathématiques effectués permettent de confirmer nos conclusions antérieures (Fons et al., 1980) , à savoir :
- que l 'étage euméditerranéen se caractérise par la présence de Mus spretus et p ar l'absence de Microtus arvalis, Pitym!IS pyrenaicus, Clethrionomys glareolus, Sorex « araneus » et Sorex min utus, espèces à vaste rép artition européenne mais ne descendant p as dans les Pyrénées-Orientales en dessous de l'étage montagnard ; qu'aucune espèce n'est inféodée à l'étage subméditerranéen (Chêne pubescent) mais que celui-ci est une zone de transi tion entre la faune méditerranéenne et la faune « montagnarde » (présence simultanée de Sorex et de Mus spretus p . ex.) ; que Pitymys duodecimcostatus, Suncus etruscus et Crocidura waveolens, bien que manifestant des affinités méditerranéennes ne peuvent être considérés que comme des indicateurs médiocres de cet étage. Pitymys duodecimcostatus est favorisé p ar la présence de champs cultivés au sol profond et profi te de ce trait du paysage pour dépasser les limites du biome méditerranéen. L'abondance des Crocidurinae , notamment celle de C. russula est p areillement favorisée par la présence de murets en m açonnerie sèche qui lui offrent des conditions micrcclimatiques particulières nécessaires à leur survie dans des zones plutôt défavorables (Fons, 1975 ; Genoud et Hausser, 1979) . Rabouillet est à cet égard un échantillon très révélateur p uisque en dépit de son altitude élevée (900 m) on y trouve encore les trois espèces de Crocidurinae. Notons du reste que Crocidura russula vit encore bien plus haut en altitude comme en témoigne sa présence dans les pelotes de Faucon crécerelle d'Aygua tebia (1 350 m) . Ceci ne nous étonne guère dans la mesure où en 1 978, nous avons découvert une population dense de cette espèce à Vielle Aure (800 m, Hautes-Pyrénées) dans une prairie entourée de murets et où nous avons capturé un e xemplaire de cette même espèce au pied d'un muret, à proximité de la berge du lac d'Oredon (1 850 rn, Hautes-Pyrénées) .
Nos conclusions rej oignent les observations de Heim de Balsac et de Beaufort (1966) , de Saint Girons et Vesco (1974) et de Fayard et Erome (1977) pour la vallée du Rhône. Elles peuvent également
- 21 1 -

être étendues au versant espagnol des Pyrénées ; si l 'on s'en réfère aux indications de Gallego (1970) , de Claramunt et al. (1975) , de Gosalbez (1976) , de Gosalbez e t Sans Coma (1976) et de Sans Coma (1 979) .
Enfin, ces traitements mathématiques établissant des concordances très étroites entre le régime de l'Effraie et les variables écogéographiques nous autorisent à penser que :
l a Chouette effraie est une p arfaite opportuniste : il est en effet peu probable que ces concordances eussent été aussi bonnes si nous avions eu à faire à un prédateur spécialisé. I l se peut néanmoins que la surabondance locale d'une proie induise chez le rapace la formation d'une image de recherche entraînant une prédation préférentielle . D ans ce cas, le développement de l'image de recherche ne s·erait-il tout de même pas l a traduction de l'opportunisme fondamental de l'Effraie, puisque en fin de compte, i l s 'agit d'un mécanisme d'adaptation à des conditions locales ? En cela nous rej oignons Tinbergen (1933) et Blondel e t Frochot (1967) . les travaux de reconstitution p aléoécologiques réalisés à l 'aide de l'étude des couches à rongeurs (pelotes de réj ection fossilisées) (cfr Chaline, 1970 ; Chaline et al., 1 974 ; Cordy, 1 975 ; Jeannet, 1979, etc.) peuvent trouver ici, contrairement à ce que pense notamment Von Knorre (1973) , une base écologique certaine (principe des causes actuelles) .
RESUME
Au moyen d'une méthode de description multid imensionnelle (analyse des correspondances) , les auteurs étudient les variations écogéographiques du régime de la Chouette effraie (Tyto alba') dans le département des Pyrénées-Orientales.
Les résultats reposant sur l 'examen de 19 échantillons de pelotes de réj ection (soit 18 561 proies) permettent : - de caractériser l'étage euméditerranéen p ar la présence de
Mus spretus et l 'absence de Microtus arvalis , Pitymys pyrenaicus, Clethrionomys glareolus, Sorex « araneus » et de S. minutus ; de constater que l'étage subméditerranéen fait l a transition entre l 'étage euméditerranéen et l'étage montagnard : on n'y trouve aucune espèce particulière mais un mélange d'éléments faunistiques appartenant aux étages voisins ; de considérer Pitymys duodecimcostalus, Suncus etruscus e t Crocidura suaveolens comme de médiocres indicateurs du biome méditerranéen. I ls profitent en effet de circonstances locales (champs cultivés pour le Campagnol, murets en pierres sèches pour les deux musaraignes) pour dépasser les limites de ce biome.
- 21 2 -

En outre, les variations quantitatives du régime étant étro itement liées à des facteurs écoclimatiques, ces résultats nous permettent de penser que Tyto alba agit en opportuniste. Enfin, ils nous p ermettent de j ustifier les travaux de paléoécologie du Quaternaire reposant sur l 'examen de « couches à rongeurs » .
SUMMARY
Correspondence analysis is used to study the variations of the diet of the Barn Owl (Tyto alba) in the eastern part of the Pyrénées, and their relationship with the various vegetation zones of the area. Nineteen samples of owl pellets (18561 prey items) were collected, together with information on the altitude, climate, vegetation and land-use of the sites where pellets were gathered.
ln the eumediterranean zone, the Barn Owl preys mainly on Mus spretus, Pitymys' pyrenaicus, Clethrionomys glareollls, Sorex « araneus » and S. minutus. Microtus arvalis is conspicuous by its absence .
Mus spretus i s a typical eumediterranean species . Pitymys pyrenaicus, Microtus arvalis, Clethrionomys glareolus, Sorex « araneus » and S. min utus are conspicuously absent.
Pitymys duodecimcostatus, Suncus etruscus and Crocidura suaveolens cannat be considered as « indicator species » of the m editerranean biome. These small mammals take advantage of sorne man-made environmental changes to enter areas weil outside the limits of the mediterranean bioclimate : e.g. cultivated fields with deep soils for Pitymys and dry-stone walls for Crocidurine shrews .
The strong correlation existing between ecoclimatic fa ctors and prey selection emphasizes the opportunistic nature of Barn Owl's predatory behaviour. The use of fossil pellets as an indicator of past environmental conditions of a given area is amply j ustified .
REMERCIEMENTS
I l nous e st ag1·éable de remercier Monseigneur Henry l 'Heureux, évêque de Perpignan, qui nous a fac i l ité l 'accès aux clochers de son d iocèse a ins i que les se1·vants d e s égl i s e s et l e s fonct io nnaires mun icipaux qui nous ont permis de les v i s iter.
Ph. Ors i n i nous a aidés dans l ' identification spécifique des c 1·ânes de souri s . A F ayard nous a prodigué avis et conse i l s, J .M . Cm·dy et G . Lennes ont accepté d e rel ire et de cr it iquer le manuscrit : qu'i l s eu soient remerciés . Que G. Mersch du département de statist ique de l 'Université de Liège, G. Laplanche et M. Nihon du centre de calcul rie la même université t1·ouvent i c i l 'expression de notre rec o n n a i ssance . San s leurs i ntervention s efficaces i l nous eut été impossible de mener à bien le traitement de nos données .
- 213

BIBLIO GRAPHIE
ANONYME ( 1 97 3 ) . - D ictionnaire des comm unes de France . 34• éd. , Berger-Lcv1·ault , Pa1·i s, 1 026 pp .
AssELDE RG, R . ( 1 9 7 1 ) . - De verspre i d i n g van de k le ine zoogdieren i n Bdgië aan de hand van braakballenanalyse . Bull . Inst . roy. Sc. nat . Belg . , 47 : 1 -60 .
BALAT, F. ( 1 9·5 6 ) . - Beitrag zur Erniihrung der Sch l e i ereule i n Südmahren und in der Slowakei. Zoo/ . List•y, 5 : 237-258 .
BECAT, J . ( 1 9 7 7 ) . - A tlas de Catalun y a Nord. Terra nosh·a, Prades , 220 cartes .
BENZECRI, J . P . et col l . ( 1 980 ) . - L'analyse des données . I l : L'analyse des correspondances. 3• éd. , Dunod, Par is , 632 pp.
BLONDEL, J. , F nocHOT, B. ( 1 967 ) . - Réflexions su1· l e s rapports en tre prt-dateu rs et proies chez l e s Rapaces. Terre et Vie, 2 1 : 5-62 .
BovET, J . ( 1 963 ) . - Etude par l 'analyse du contenu de pe lotes de Ch ouette e ffra i e ( Tyto alba) d e fluctuations d a n s l e s populat ion s d e micromam m i fères. Rev. suisse Zoo/., 70 : 244-249 .
B ROSSET, A . ( 1 956) . - Le régime de l 'Effra i e au Maroc m·iental . A la u da, 24 : 303-305.
BnowN, D.J. ( 1 981 ) . - Seasonal variations i n the prey of s omc b a 1·n owl s in Gwynedd. Bird Study, 28 : 1 39-1 46 .
Bnu iJN, O. DE ( 1 9 7 9 ) . - Voedseloccol ogie van de kCI·ku i l , Tyto alba , i n Nedel' land . Limo•sa, 52 : 9 1 - 1 54.
CABON-RACZYNSKA, 1\ . , HUPRECHT, A . ( 1 9 7 7 ) . - Estimation o f population dens ity of the common vole in Poland : an ana lys i s of owl pellets . A c ta theriol . , 22 : 349-354 .
CHALINE, J . ( 1 970) . - La signification des rongeurs dans l e s dépôts quaternaires . Bu ll. Ass. fr . Et. Quai., 25 : 229-241 .
CHALINE, J. , BAUDVIN, H., JAMMOT, D. , SAINT GmoNs, M . C . ( 1 9 74 ) . - Les proies des rapaces. Doin, Ed., Pa1·is, 141 pp.
CHEYLAN, G . ( 1 976 ) . - Le régime a l imentaire de l a Chouette l'ffra ie , Tyto alba, en Europe méditeiTanécnne. Terre e t Vie, 3 0 : 565-5 7 9 .
CLARAMUNT, T ., GosALBEZ, J . , S A N S CoMA, V . 0 975) . - Notes sobre la b iol(cogJ·afia de l s micromammifers a Catalnfiya. Bu/. Insl . Cala/ . Hist . nat. (Sec. Zoo/ . ) , 3 9 : 2 7-40 .
ConnY, J .M. ( 1 975 ) . - Bio ct cl1 1·on ostratigmph i e d e s dépôts tardiglac i a i res c t holocènes de la grotte d u Coléoptère à Bomal-sur-Oui·the . A n n . S o c . géol . Belg . , 98 : 2 91 -296.
Cnuz, F . , GARCIA RoDRIGUEZ, P. (1 969) . - Tyto alba a l imcntandose p i' Încipa lmen lc de Sturnus u n icolor. Ardeola, 1 5 : 1 46 .
CuRIO, E . ( 1 9 7 6 ) . - The etho logy of predation . Sp 1· in ger-Vedag, Berl i n , Heidelberg, 250 pp .
DouME RET, A., SAINT GmoNs , M.C . ( 1 97 7 ) . - Hemarques sut· J e régime de l 'Effi·a ic, Tyto alba, en Charente-Ma ritime. Ann. Soc. Sc . . Vat . Charen te-Maritime, 6 : 267-273.
EMBERGEH, L. ( 1 942 ) . - Le p1·ojet d'une cl a ssif ication des cl imats du point de vue phytogéographique. Bu ll. Soc. Hist. nat . Toulouse , 77 : 97-1 24 .
FAIHLEY, J . , CLARK, F.L. ( 1 972) . - Food of Barn owls ovcr one ycar at a roo st in Co. Galway. Irish Nat . , 17 : 2 1 9-222 .
FAYARD, A., E ROME, G . ( 1 9 7 6 ) . 4 0 : 689-690.
Mus m usculus spre tus en Ardèche . Mam maliu,
FAYARD, A., E noME, G . ( 1 9 7 7 ) . Les mict·omamm ifère s de la bordure orienta l e d u Massif centra l . Mam malia, 4 1 : 301 -3 1 9 .
214

FoNs, R . ( 1 9 7 5 a) . - Contribu tion à la connaissance de la Musaraigne é trusque Suncu s etruscus (Savi , 1 822) . Mamm ifère Soricidae. Thèse Doctorat, Université P .-et-M.-Curie , Paris, 1 92 pp.
FoNs, R . ( 1 97 5 b ) . - Premières données sur l 'écologie de la Pn chyure étrusque, Suncus e truscus (Savi, 1 822) ct comparaison avec deu x autres Crocidurinae : Crocidura rnssula (Hermann, 1 780) et Crocidtzra suaveolens (Pallas, 1 8 1 1 ) , ( l n sectivora, Soricidae) . Vie ,Uilieu, 25 : 3 1 5-360.
FONS, R ., LIBOIS, R .M. , SAINT GIRONS, M.C. (1 !180 ) . - Les micromammifères dans le département des Py1·énées-Orientales. Répartition altitudinale et l ia ison avec les étages de végétat ion . Vie Milieu , 30 : 285-299 .
GALLEGO, L . ( 1 9 7 0 ) . - D i stribuci o n de micromamiferos en Navarra . Pirineos, 98 : 4 1 -52 .
GAUSSEN, H . ( 1 97 2 ) . - Carte de végé tation de la France. Feu ille de Perpignan, 2• éd .
GENOUD, M. , HAUSSER , J . ( 1 9 7 9) . - Eco l ogie d'une population de Crocidura ntsst.tla en m i l i eu rural montagnard (Insectivora, Sol"icidac) . Terre e t l'ie, 33 : 5 3 9-554 .
GEROUDET, P. ( 1 965 ) . - Les Rapaces diurnes e t nocturnes d'Europe. Delachaux et Niestlé, Neuchâtel , 4 2 6 pp .
GLUE, D . ( 1 974) . - Food of the Barn owl in Br i ta in und Irelanrl . Bi rd Study, 21 : 200-21 o .
GLuE, D ., NUTTALL, J . ( 1 9 7 1 ) . - A dverse climntic condit ions affecting the diet of a Bam owl in Lancashire . Bird Study, 18 : 33-34 .
GLuTz von BLOTZHEIM, U.N. , BAUEH, KM. ( 1 980) . - Handbuch der Vogel Millele u ropas Bd.!!, S trigiform es . Al>ademische Verlagsgesellschaft, 'Viesbaden, pp . 235-27 6 .
G ô R N E R , M . (1 97 9 ) . - Zur Verbrei tung d e r IOcins iiuger i m Sudwcstern d e r D D R a u f der Grundlage von Gewôl lanalyscn d e r Schlciereu le ( Tyto alba) . Zoo/ . Jb . Sys t., 1 0 6 : 429-470 .
GosALBEZ, J . ( 1 976 ) . - Sorex m in u tus en las montaiias de Prades (Catalufia, Espafia ) . Mise. zoo/. , 3 : 2 1 3-2 1 6 .
GosALBEZ, J . , S A N S COMA, V . ( 1 976 ) . - Sobre c l lopil lo rojo, Cle lhrionomys glareolus (Schreber, 1 78 0 ) en la region del Mon tscny (Catalufia, Espaiia ) . Süugetierkdl Mill . , 24 : 1 2- 1 8 .
G usEv, V . M . (1 95 2 ) . - S u r l ' importance d e l a profondeur de l a couche d e neige pour les o i seaux se n ourrissant de rongeurs mul"in s (en russe ) . J. zoo/. U.R .S.S., 3 1 : 4 7 1 -4 7 3 .
HAIN SWOHTH, F .R. , WOLF, L . L . (1 9 7 9 ) . -- Feedi ng, an ecologica! a pproach. Adv. S tudy Behav. , 9 : 5 3-96 .
HEm DE BALSAC, H . ( 1 965 ) . - Quelques renseignements d'ordre faunistique tirés de l 'étude du régime a l i mentaire de Tyto alba dans l 'ouest de l 'Afrique. Alauda, 33 : 309-322 .
HEU! DE BALSAC, H . , DE BEAUFORT, F . ( 1 966) . -- Régime a l i m en taire de l 'Effraie dans l e Bas-Dauphiné. Applications à l 'étude des Vertébt·és . A lauda, 34 : 309-324 .
H E R RERA, C.M. ( 1 9 7 4 a) . - Regimen al imenticio de Tyto alba en Espafia sudoccidenta l . Ardeola, 1 9 : 359-:!94 .
H E R REHA, C.M . ( 1 9 7 4 b) . - Trophic diversity of the Barn owl, Tyto alba, in continen ta l ·western Europe . Orn is scandinavica, 5 : 1 8 1 -1 9 1 .
H o sKING, E.J . , NEWBE RHY, C .M. ( 1 945) . - Bircls o f the n igh t . Collins, London, 1 28 pp .
H u ssoN, A . M . ( 1 949) . - Gewûlle-Analysen und der Verbreitung der Kleinsiiuger von Luxembourg. Bull. L igue l!!x. Prof. Ois., Sér. 3, 28 : 82-92.
JEANNET, M . ( 1 9 7 9) . - Les Rongeurs ct l 'environnement au mont des Es1>é lugues (Lourdes, Hautes-Pyrénées) . Bnll. Soc. Rist. nat. 1'o !l louse, 1 1 5 : 5 3-64.
- 215 -

KNORRE Von, D. ( 1 9 7 3 ) . - Jagdgebiet und Ui.gl icher Nahrungsbedm·f der Schle iereule (Tyto alba Scopol i ) . Zool . Jb . Syst . , 100 : 301 -320 .
KREBS , J .R . ( 1 97 8 ) . - Optimal foraging : decis ion rul e s for predators. In Behavioural ecology : an evolu tionary approach. Krebs, J . R . Davies , N .B. eds . Blackwell . Oxford, pp. 23-63 .
LEBA RT, L. , MORINEAU, A. , TABARD, N . (1 97 7 ) . - Techniques de la description s tatis tique . Dunod, Paris, 351 pp .
Lmms, R .M. ( 1 977 ) . - Etude des mammifères e t réserves n aturelles . Bull. Rés . nat . ornithol . Belg. , 25 : 24-29.
Lmms, R .M . , VRAN KEN, M. ( 1 981 ) . - Myo tis b echsteini en Corse . Mammal ia, 45 : 380-38 1 .
LovA RI, S. , RENZONI, A . , .FoNDT, R. ( 1 9 7 6 ) . - T h e predatory habits of the B a r n o w l (Tyto alba Scopo l i ) i n relation to t h e vegetation cover. B u l l . Zoo l., 43 : 1 73-1 9 1 .
MARTIN, C . D . ( 1973) . - Ten years o f Barn o w l prey data from a Colora d o nest s ite . Wilson Bull. , 85 : 85-86.
MARTIN, J. , VERICAD, J.R. (1 977 ) . - Datos sobre l a a l i mcntacion d e l a lechuza ( Tyto alba) en Valencia . Mediterranea, 2 : 35-4 7 .
ORSINI , P . 0 97 9) . - Notes sur les souri s de Provence. Ann . S o c . S c . n a t . arch . Toufon e t V ar, 31 : 1 68-1 73 .
0TTENI, L.C . , BoLEN, E . G . , COTTAM, C.W. ( 1 972 ) . - Predator-p1·cy relat ionships and reproduction of the Barn owl i n Southern Texas . Wilson Bull . , 84 : 4 34-448.
PETIT, G. , LoMONT, H. (1 958) . - Nouvel l e pul lulat ion d e pé lobates dans la région de Canet (P .-0 . ) . l' ie Milieu, 9 : 1 31 -1 32 .
RuPRECHT, A . (1 979) . - F o o d of the B a r n o w l , T y t o a l b a g u l tata, from Kujawy. Acta o rnithol . , 16 : 493-5 1 1 .
RYBAR, P . (1 969 ) . - D i e Zweifarbige F ledermaus ( Vespertilio m urin us) und ande1·e kleine Wirbeltiere i n der Nahrung der Schle iereule (1'yto alba g u t ta ta Breh m ) i n Ca stolovice . Zool. L isty, 18 : 239·246 .
SAGE, B.L. ( 1 962 ) . - Barn owls catching sparrows at roost . British Birds, 5 5 : 237-238 .
SAINT GIRONS, M.C. (1968 ) . - Analyse des fluctuat ions du régi m e de l 'Effra ie , Tyto alba, dans l e département de l a Somme pendant u n e pul lu lat ion de Micro fus arvalis. A cta. Soc. zoo/ . Bohe mos/ . , 32 : 1 85-1 98 .
SAINT GIRONS, M .C. ( 1 9 7 3 a ) . - Les Mam m ifères de France e t du Benelux. Doin , Pari s, 481 pp.
SAINT GIHON S, M.-C. ( 1 973 b ) . - Le rég ime de l 'Effraie, Tylo afba, SU I" la côte atlant ique du Maroc. Bull . Soc . Sc. n a t. phys. Maroc, 5 3 : 1 93-1 98.
SAINT GIRONS, M.C., MARTIN, C. ( 1 9 7 3) . - A daptation du régime de quelques rapaces n octurnes au payage rnral . Les proies de l 'Effra i e et e lu Moyen-duc dans le département de la Somme. Bu ll. Eco/ . , 4 : 95-1 20 .
SAINT GIRONS, M.C., SPITZ, F . ( 1 966 ) . - A propos de l 'étude des micromammifères par l 'analyse des pelotes de Rapaces. Intérêt et l i m ites de l a méthode. Terre e t Yie, 20 : 3-1 8 .
SAINT GIRONS, M .C . , THOUY, P. (1 978) . - Fluctuation s dans l e s popula t i o n s de Souris, Mus spretus (Lataste, 1 88 3 ) , en région méditerranéenne. Bull . Ecol. , 9 : 21 1 -2 1 8 .
SAINT GIRONS, M .C., VEsco, J.P. ( 1 974) . - N ote s u r l e s Mamm ifères de France. XIII : Répartition et densité des petits mammifères dans l e couloir séquano-rhodanien . Mammalia, 38 : 244-264 .
SANS CmiA, V . (1 974) . - Sobre l a al imentacion de Ty to alba en l a region continenta l catalana. Mise. zool., 3 : 1 63-1 69 .
216 -

S A N S CoMA, V. ( 1 9 7 9 ) . - Beitrag zur Kenntn i s der Waldspitzmaus, Sorex arane u; L. (1 7 5 8 ) , i n Katalonien, Spanien . Süugetierkdl. Mitt. , 2 7 : 96-1 06 .
S A N S CoMA, V., CLARAMUNT, T . , GoSALBEZ, J. ( 1 9 7 6 ) . - Die M ittelmeer-Kleinwühlmaus Pitym ys d u odecimcostatus (de Selys Longchamps, 1 8 39 ) , i n der Ema hrung der Sehle iereule ( Ty t o albaj Katalonien (Spanien ) . Süugetierkdl. Mitt . , 24 : 7 7- 7 9 .
ScHMIDT, E. ( 1 9 7 3 a ) . - D i e Nahru n g der Schleiereule in Eump a . Z. f . angw. Zoo[. , 60 : 43-7 0 .
ScHMIDT, E . ( 1 9 7 3 b ) . - Quantitative Daten d e s Haussperl i ngs (Passer domesticus) aus ungari schen Schi·e iereulengewol len . Zoo/ . A bh . (Dresden) , 32 : 1 7 2-1 74 .
ScHNEIDER, W. ( 1 964 ) . - D ie Sch leiere u le . Neue B I·eh m Bücherei, He ft 340, Wittenberg, 1 04 p p .
SPITZ, F . ( 1 9 7 8 ) . - Etude c i·an i ométrique d u geni·e Pitymys . Mam malia, 4 2 : 267-304 .
T INBERGEN, N . ( 1 933 ) . - D i e ernahrungsokologischen Beziehungcn zwischen Asio o . o t u s u n d i l u·en Beutetieren, insbesonderc den Micro lus-ai·tcn. Eco/ .Monogr., 3 : 443-492.
TYRNER, P. , BART A, Z. ( 1 9 7 1 ) . - Kleinsauger ais Nahrung de i· Schleiereule ( Ty to alba g u / ta ta ) in Nordwestbohmen. Zoo/. A bh . ( Dresdeu ! , 32 : 5-1 6 .
UTTENDORFER, O . ( 1 939) . - Die Ernührung der deutsch en R a u bvogel und Eu len und ihre Bedeu tung in der heimischen Natur. Neudam, Berl in , 4 1 2 pp.
UTTE N D O HFER, O. ( 1 95 2 ) . - Neue Erge bnisse über die Erniihmng der Greifvogeln und E u /en . Eugen U l mer, Stuttgart, 230 pp .
WEBSTEH, J . A . ( 1 9 7 3 ) . - Seasonal variation i n mammal contents of Bai·n owl cast ings . Bird study, 2 0 : 1 85-1 96 .
ZELE N I<A, G., P HICA M , R . ( 1 964) . - Variati o n s d'effectifs des populati o n s de petits mammifères révél ées par le régi me a l i menta i i·e d'un rapace nocturne. Terre e t Vie, 1 1 1 : 1 7 8-1 84.
- 217 -