kinetoplast dna minicircles are inherited from both parents in genetic crosses of trypanosoma...
TRANSCRIPT

ORIGINAL PAPER
Wendy Gibson á Mandy Crow á Julia Kearns
Kinetoplast DNA minicircles are inherited from both parentsin genetic crosses of Trypanosoma brucei
Received: 26 October 1996 /Accepted: 12 December 1996
Abstract In the order Kinetoplastida, genetic exchangehas been demonstrated only in the genus Trypanosoma.Analysis of kinetoplast DNA (kDNA) in genetic crossesof T. brucei has shown that whereas maxicircles are in-herited uniparentally, minicircles are inherited fromboth parents. This result was con®rmed for a new crossof T. b. brucei and T. b. rhodesiense by restriction en-zyme digestion and Southern analysis of puri®ed kDNA.By hybridisation with small minicircle-derived probes,we could demonstrate the presence of particular parentalminicircles in the kDNA of hybrid progeny clones. Allhybrid clones had inherited two minicircles from oneparent despite two of the four clones having maxicirclesfrom the other parent. The results suggest that ratherthan small-scale exchange of minicircles between pa-rental networks, gross breakdown and reassembly of theminicircle network occurs during genetic exchange.
Introduction
The order Kinetoplastida is characterised by possessionof a unique organelle, the kinetoplast, which containsthe tightly packaged mitochondrial DNA. Amongkinetoplastids, genetic exchange has been demonstratedonly in the genus Trypanosoma; Jenni et al. (1986)showed that hybrids were produced when di�erentclones of T. brucei were transmitted through tsetse ¯ies(genus Glossina). Extensive comparisons of parental andhybrid clones by a number of genetic techniques havesince elucidated many aspects of the mechanism ofgenetic exchange in trypanosomes. The process is not anobligatory part of the life cycle, since both parental and
hybrid clones are represented among metacyclics pro-duced by an individual ¯y (Schweizer et al. 1988).Analysis of isoenzymes and restriction-fragment-lengthpolymorphisms (RFLPs) indicates that a meiotic divi-sion probably occurs during genetic exchange; segrega-tion and reassortment of genetic markers is observed(Jenni et al. 1986; Paindavoine et al. 1986; Wells et al.1987; Gibson 1989; Sternberg et al. 1989; Turner et al.1990; Gibson and Garside 1991; Gibson et al. 1992,1995; Gibson and Whittington 1993; Gibson and Bailey1994; Schweizer et al. 1994; Degen et al. 1995), as is ahigh frequency of chromosomal recombination in hy-brids (Gibson et al. 1992, 1995; Gibson and Bailey1994). However, no haploid gamete stage has been de-tected (Shapiro et al. 1984; Kooy et al. 1989; Tait et al.1989), and genetic exchange appears to involve fusion ofparental trypanosomes (Gibson 1995).
Given this view of events, either parental cell mightprovide the kinetoplast DNA (kDNA) of a hybridprogeny clone and, indeed, initial results suggested thatinheritance of kDNA was uniparental (Sternberg et al.1988, 1989; Gibson 1989). However, kDNA has acomplex structure, consisting of two types of circularDNA molecules intercalated into a dense network (re-viewed in Shapiro and Englund 1995). There are 50±100maxicircles per network, which are homogeneous in se-quence and carry the genes for mitochondrial biogenesis.The 5,000 or so minicircles of T. brucei ssp. are hetero-geneous in sequence and encode guide RNA molecules,which direct the editing of maxicircle transcripts. Whenthese two components were considered separately, it wasshown that whereas maxicircles were inherited unipa-rentally, minicircles were inherited from both parents(Gibson and Garside 1990).
How this result is brought about is unclear. Onesuggestion is that the two parental kDNA networks mix,creating a single hybrid network. Subsequent mitoticdivisions reduce the maxicircle component to homoge-neity by random partitioning of the relatively smallnumber of molecules. Birky (1983) discussed this processwith reference to yeast mitochondrial DNA, for which
Parasitol Res (1997) 83: 483±488 Ó Springer-Verlag 1997
W. Gibson (&) á J. KearnsDepartment of Genetics, University of Leicester,Leicester LE1 7RH, UKFax:+44 (0116) 252 3378; e-mail: [email protected]
M. CrowDepartment of Biochemistry, University of Cambridge,Tennis Court Road, Cambridge CB2 1QW, UK

homogenisation was achieved in about 20 generations.The minicircles would remain heterogeneous by virtue oftheir greater number and sequence diversity. The ob-servation of putative intermediate hybrid clones withmixed maxicircles during the early stages of vegetativegrowth supports this proposal (Turner et al. 1996).However, considering the complexities of replication(Shapiro and Englund 1995), it is not easy to envisagethe fusion of kDNA networks at the molecular level. Analternative proposal is that the two parental kDNAnetworks swap minicircles while in contact during theprocess of genetic exchange but retain their maxicircles(Shapiro and Englund 1995). It is signi®cant that Sha-piro (1993) showed by selective removal of minicirclesthat the maxicircles form an independent network in-terlocked to the minicircle network.
The biparental inheritance of kDNA underpins ourcurrent model of genetic exchange in trypanosomes(Gibson 1995), since the results imply that mitochondrialfusion and, hence, cell fusion must occur. In an attemptto resolve the two proposed mechanisms for the di�er-ential inheritance of maxi- and minicircles, we present®ndings on the inheritance of individual minicircles.
Materials and methods
Trypanosomes
The parental trypanosomes (058H and KP2N) and hybrid clones(P4, P5, P11, P12) used in the present study have been describedelsewhere (Gibson and Bailey 1994). The four hybrid clones werederived from a single ¯y and were identical in all respects except thetype of kDNA maxicircles (Gibson and Bailey 1994). The two otherTrypanosoma brucei stocks used were the T. b. brucei TSW 196(MSUS/78/CI/TSW 196 CLONE A) and T. b. rhodesiense 058
(MHOM/ZM/74/58 CLONE B) clones described by Gibson (1989).Procyclic trypanosomes were cultured in Cunningham's medium(Cunningham 1977) with gentamycin at 20 lg ml)1. Bloodstream-form trypanosomes were grown in mice and puri®ed from bloodcells by anion-exchange chromatography (Lanham and Godfrey1970). KDNA was puri®ed by centrifugation of sheared lysates oftotal DNA (Simpson and Berliner 1974; Gibson et al. 1985).
Electrophoresis and hybridisation
KDNA was restricted and size-fractionated using 2±2.5% agarosegels. Gels were blotted in the standard way (Southern 1975) andnitrocellulose ®lters were hybridised as previously described (Gib-son et al. 1988) with the oligo-labeled DNA probes A±E indicatedin Fig. 1. Probe A was a 203-bp HinfI fragment containing aconserved guide RNA (Shu and Stuart 1993) from the ampli®edinsert of T. evansi clone pTK420 (Masiga and Gibson 1990). ProbesB±E were from two clones containing HincII/PstI minicircle frag-ments from T. b. brucei TSW 196 (Gibson and Garside 1990). Allprobes except C contained 40±50 bp of plasmid DNA from themultiple cloning site, since they were ampli®ed using SP6 or T7primers. All post-hybridisational washes were done to 0.1 ´ SSC(1 ´ SSC � 0.15 M NaCl, 0.015 M trisodium citrate) at 65 °Cunless indicated otherwise in the ®gure legends. End-labeledprimers p2, p3 and p4 were also used for hybridisation followingthe protocol of Mason and Williams (1985). The primers used wereas follows 5¢±3¢: p2 AAGCAGCCACTACCATC, p3 TCAGCC-CGAAAATCACC and p4 CTTTCCATATTTCTTGTTG. Blotswere reused several times, with probes being removed by washing in0.1 M NaOH 0.1% sodium dodecyl sulfate (SDS) for 10 min atroom temperature followed by several changes of 3 ´ SSC.
DNA sequence analysis
The inserts from clones pTK196-9 and pTK196-14 were sequencedby polymerase chain reaction (PCR) cycle sequencing using anautomated sequencer. The sequences were searched for the 12-meruniversal minicircle sequence GGGGTTGGTGTA to exclude thisand its conserved ¯anking region from the probes used for hy-bridisation (Fig. 1).
Fig. 1 Sketch maps of 3 kDNAminicircle clones to showpositions of probes and primersused for hybridisation. Thinlines represent plasmidsequences; thick lines,trypanosome sequences; and ahatched box indicates theposition of the conserved regionwith invariant 12-mer sequenceas shown. Primers used forampli®cation and labeling areindicated by arrows
484
Results and discussion
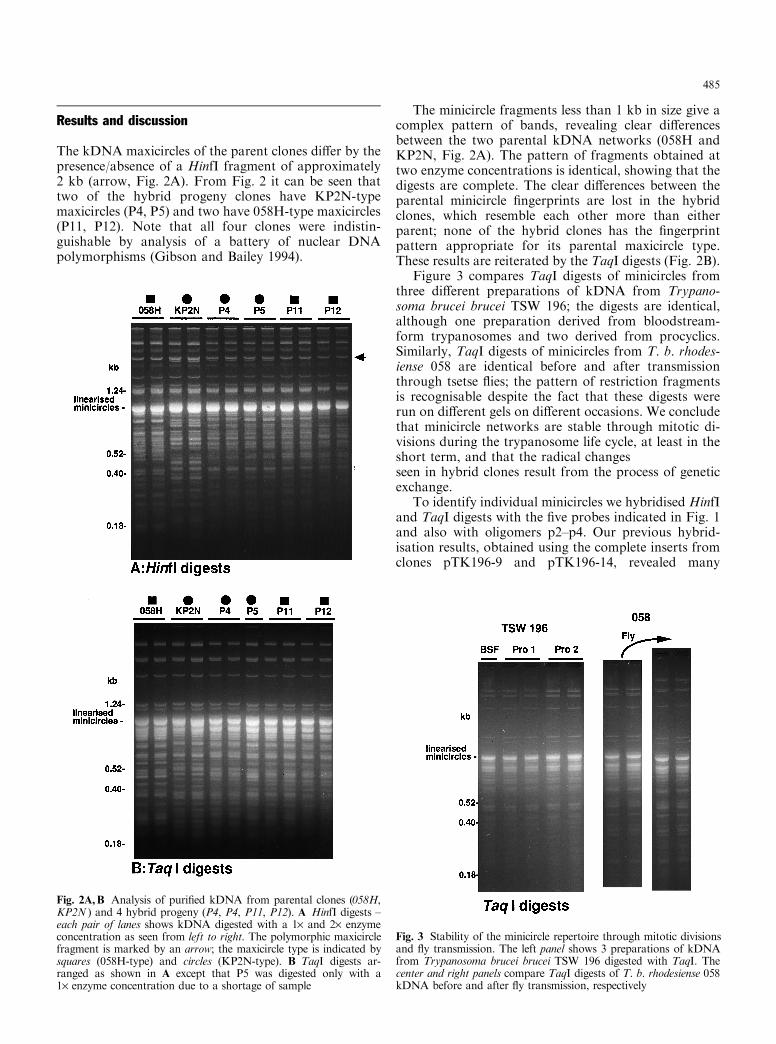
The kDNA maxicircles of the parent clones di�er by thepresence/absence of a HinfI fragment of approximately2 kb (arrow, Fig. 2A). From Fig. 2 it can be seen thattwo of the hybrid progeny clones have KP2N-typemaxicircles (P4, P5) and two have 058H-type maxicircles(P11, P12). Note that all four clones were indistin-guishable by analysis of a battery of nuclear DNApolymorphisms (Gibson and Bailey 1994).
The minicircle fragments less than 1 kb in size give acomplex pattern of bands, revealing clear di�erencesbetween the two parental kDNA networks (058H andKP2N, Fig. 2A). The pattern of fragments obtained attwo enzyme concentrations is identical, showing that thedigests are complete. The clear di�erences between theparental minicircle ®ngerprints are lost in the hybridclones, which resemble each other more than eitherparent; none of the hybrid clones has the ®ngerprintpattern appropriate for its parental maxicircle type.These results are reiterated by the TaqI digests (Fig. 2B).
Figure 3 compares TaqI digests of minicircles fromthree di�erent preparations of kDNA from Trypano-soma brucei brucei TSW 196; the digests are identical,although one preparation derived from bloodstream-form trypanosomes and two derived from procyclics.Similarly, TaqI digests of minicircles from T. b. rhodes-iense 058 are identical before and after transmissionthrough tsetse ¯ies; the pattern of restriction fragmentsis recognisable despite the fact that these digests wererun on di�erent gels on di�erent occasions. We concludethat minicircle networks are stable through mitotic di-visions during the trypanosome life cycle, at least in theshort term, and that the radical changesseen in hybrid clones result from the process of geneticexchange.
To identify individual minicircles we hybridised HinfIand TaqI digests with the ®ve probes indicated in Fig. 1and also with oligomers p2±p4. Our previous hybrid-isation results, obtained using the complete inserts fromclones pTK196-9 and pTK196-14, revealed many
Fig. 2A,B Analysis of puri®ed kDNA from parental clones (058H,KP2N ) and 4 hybrid progeny (P4, P4, P11, P12). A HinfI digests ±each pair of lanes shows kDNA digested with a 1´ and 2´ enzymeconcentration as seen from left to right. The polymorphic maxicirclefragment is marked by an arrow; the maxicircle type is indicated bysquares (058H-type) and circles (KP2N-type). B TaqI digests ar-ranged as shown in A except that P5 was digested only with a1´ enzyme concentration due to a shortage of sample
Fig. 3 Stability of the minicircle repertoire through mitotic divisionsand ¯y transmission. The left panel shows 3 preparations of kDNAfrom Trypanosoma brucei brucei TSW 196 digested with TaqI. Thecenter and right panels compare TaqI digests of T. b. rhodesiense 058kDNA before and after ¯y transmission, respectively
485

homologous minicircle fragments in each clone (Gibsonand Garside 1990). Sequence analysis enabled us tochoose shorter probes, avoiding the minicircle conservedregion contained within both inserts (Fig. 1). Probe Acontained a guide RNA (gCR4) conserved betweenT. brucei, T. evansi and T. equiperdum (Shu and Stuart1993), which we anticipated might be more widely con-served within T. brucei.
Probes A and B identi®ed minicircles that were de-tectable in parent 058H, but not KP2N, and were alsopresent in various amounts in all four hybrid clonesindependently of the maxicircle type (Fig. 4A±C). Thus,if the kDNA networks of clones P4 and P5 derived fromKP2N, then they gained a minimum of two new mini-circles from 058H during genetic exchange. In contrast,probe E hybridised strongly with several minicirclefragments from parent KP2N, and some or all of thesefragments also occurred in the four hybrid clones(Fig. 4D). From this we can conclude that if the kDNAnetworks of clones P11 and P12 derived from 058H,they gained several KP2N minicircles during geneticexchange.
Probes C and D hybridised with many minicirclebands in all digests and were therefore not informative(not shown); none of the oligomers produced a hybrid-isation signal under the conditions used. The di�erencein hybridisation pattern observed among probes A±Eshows that the contribution of plasmid sequences to thesignal was negligible.
In conclusion, we have veri®ed our original ®ndingthat minicircle networks are hybrid (Gibson and Garside1990), and this has now been shown in three indepen-dent crosses. The pattern of inheritance of individualminicircles observed in the present study and previouslysuggests that mixing of parental networks occurs on agross scale. We envisage that a major breakdown of theparental networks occurs in the heterokaryon and isfollowed by recatenation. Although we favour the hy-pothesis that the maxicircles remain attached to the coreof each parental network from considerations of net-work structure (Shapiro 1993; Shapiro and Englund1995), this would be hard to prove.
The four hybrid clones analysed herein were identicalin terms of nuclear DNA polymorphisms as judged from
B: Probe B
Fig. 4 Analysis of puri®edkDNA from parental clones(058H, KP2N) and 4 hybridprogeny (P4, P4, P11, P12).A, B Blots of HinfI digestsfrom Fig. 2A hybridised withprobes A and B; respectively.C, D Blots of TaqI digests fromFig. 2B hybridised with probesB and E, respectively (seeFig. 1). Post-hybridisationalwashes were done to lowstringency for probe A(3 ´ SSC, 0.1% SDS at 60 °C)
486

results of various ®ngerprinting techniques applicable totrypanosomes (pulsed-®eld gradient-gel electrophoresis,RFLP analysis of variant surface-glycoprotein genefamilies, random ampli®ed polymorphic DNA analysis).The four clones di�ered in kDNA maxicircle type, andthere were also small di�erences in minicircle repertoire.In our original hypothesis we assumed that such clonesprobably derived from a single heterokaryon, since thegeneration of identical sets of nuclear DNA polymor-phisms following meiosis is extremely unlikely (Gibson1989; Gibson and Garside 1990). On the basis of the ideathat parental kDNA networks remain separate but swapminicircles, clones P4 and P5, which have KP2N-typemaxicircles, should have more similar minicircle reper-toires than clones P11 and P12, which have 058H-typemaxicircles. This is not the case; all four clones appear tohave much the same minicircle network as determinedfrom both gel electrophoresis and hybridisation results.We previously obtained a similar result for two clonesfrom another cross, which also di�ered only in maxi-circle type (Gibson and Garside 1990). We thereforeconclude that minicircle mixing occurs on a gross scale.This has important consequences for hybrid viability,since correct maxicircle editing depends on a function-ally complete set of minicircles. Random exchange ofminicircles between networks would bring togethervarious combinations of minicircles, some of whichmight provide more e�cient editing than others. Highsequence redundancy among guide RNAs would alle-viate this problem.
Considering the implications of these results in termsof trypanosome cell biology, the heterokaryon resultingfrom the fusion of two parental trypanosomes wouldpresumably possess two kinetoplasts and two ¯agella.Robinson and Gull (1991) have shown that the kine-toplast and basal body of the ¯agellum are intimatelylinked during cell division. Hence, if the heterokaryonproceeded through cell division with normal replicationof these structures, the resultant trypanosomes wouldpossess two ¯agella, and a further doubling of thesestructures would occur after every subsequent mating.This is clearly not the case, so how has the trypanosomeresolved this problem? One possibility is that cell ®ssionwithout replication directly follows genetic exchange,perhaps triggered by the presence of two kinetoplast/basal body complexes in the heterokaryon; this modelrequires the heterokaryon to have either two diploidnuclei or a single tetraploid nucleus to produce twodiploid daughter cells. Alternatively, one or severalrounds of replication could be followed by multiple ®s-sion or budding of daughter cells. We await the identi-®cation of intermediate stages of genetic exchange todecide between these models.
Acknowledgements This work was supported by the Medical Re-search Council. We would like to thank many colleagues for dis-cussion of these results, particularly Paul Englund, Keith Gull,Michael Miles and Ken Stuart, and Mick Bailey and Mark Car-rington for comments on the manuscript.
References
Birky CW (1983) Relaxed cellular controls and organelle heredity.Science 222: 468±475
Cunningham I (1977) New culture medium for maintenance oftsetse tissues and growth of trypanosomatids. J Protozool 24:325±329
Degen R, Popischal H, Enyaru J, Jenni L (1995) Sexual compati-bility among Trypanosoma brucei isolates from an epidemicarea in southeastern Uganda. Parasitol Res 81: 253±257
Gibson WC (1989) Analysis of a genetic cross between Trypano-soma brucei rhodesiense and T. b. brucei. Parasitology 99: 391±402
Gibson WC (1995) The signi®cance of genetic exchange in try-panosomes. Parasitol Today 11: 465±468
Gibson W, Bailey M (1994) Genetic exchange in Trypanosomabrucei: evidence for meiosis from analysis of a cross betweendrug resistant transformants. Mol Biochem Parasitol 64: 241±252
Gibson W, Garside L (1990) Kinetoplast DNA minicircles are in-herited from both parents in genetic hybrids of Trypanosomabrucei. Mol Biochem Parasitol 42: 45±54
Gibson WC, Garside LH (1991) Genetic exchange in Trypanosomabrucei brucei: variable chromosomal location of housekeepinggenes in di�erent trypanosome stocks. Mol Biochem Parasitol45: 77±90
Gibson W, Whittington H (1993) Genetic exchange in Trypanoso-ma brucei: selection of hybrid trypanosomes by introduction ofgenes conferring drug resistance. Mol Biochem Parasitol 60: 19±26
Gibson W, Borst P, Fase-Fowler F (1985) Further analysis of in-traspeci®c variation in Trypanosoma brucei using restriction sitepolymorphisms in the maxicircle of kinetoplast DNA. MolBiochem Parasitol 15: 21±36
Gibson WC, Dukes P, Gashumba JK (1988) Species speci®c DNAprobes for the identi®cation of trypanosomes in tsetse. Para-sitology 97: 63±73
Gibson W, Garside L, Bailey M (1992) Trisomy and chromosomesize changes in hybrid trypanosomes from a genetic cross bet-ween Trypanosoma brucei rhodesiense and T. b. brucei. MolBiochem Parasitol 52: 189±200
Gibson W, Kanmogne G, Bailey M (1995) A successful backcrossin Trypanosoma brucei. Mol Biochem Parasitol 69: 101±110
Jenni L, Marti S, Schweizer J, Betschart B, Lepage RWF, WellsJM, Tait A, Paindavoine P, Pays E, Steinert M (1986) Hybridformation between African trypanosomes during cyclicaltransmission. Nature 322: 173±175
Kooy RF, Hirumi H, Moloo SK, Nantulya, VM, Dukes P, Van derLinden PM, Duijndam WAL, Janse CJ, Overdulve JP (1989)Evidence for diploidy in metacyclic forms of African trypano-somes. Proc Natl Acad Sci USA 86: 5469±5472
Lanham SM, Godfrey DG (1970) Isolation of salivarian trypano-somes from man and other mammals using DEAE-cellulose.Exp Parasitol 28: 521±534
Masiga DK, Gibson WC (1990) Speci®c probes for Trypanosomaevansi based on kinetoplast DNA minicircles. Mol BiochemParasitol 40: 279±284
Mason PJ, Williams JG (1985) Hybridisation in the analysis ofrecombinant DNA. In: Hames BD, Higgins SJ (eds) Nucleicacid hybridisation, a practical approach. IRL Press, Oxford, pp22±23
Paindavoine P, Zampetti-Bosseler F, Pays E, Schweizer J, GuyauxM, Jenni L, Steinert M (1986) Trypanosome hybrids generatedin tsetse ¯ies by nuclear fusion. EMBO J 5: 3631±3636
Robinson DR, Gull K (1991) Basal body movements as a mecha-nism for mitochondrial genome segregation in the trypanosomecell cycle. Nature 352: 731±733
Schweizer J, Tait A, Jenni L (1988) The timing and frequency ofhybrid formation in African trypanosomes during cyclicaltransmission. Parasitol Res 75: 98±101
487

Schweizer J, Pospichal H, Hide G, Buchanan N, Tait A, Jenni L(1994) Analysis of a new genetic cross between two East AfricanTrypanosoma brucei clones. Parasitology 109: 83±93
Shapiro SZ, Naessens J, Liesegang B,Moloo SK,Magondu J (1984)Analysis by ¯ow cytometry of DNA synthesis during the life-cycle of African trypanosomes. Acta Trop (Basel) 41: 313±323
Shapiro TA (1993) Kinetoplast DNA maxicircles: networks withinnetworks. Proc Natl Acad Sci USA 90: 7809±7813
Shapiro TA, Englund P (1995) The structure and replication ofkinetoplast DNA. Annu Rev Microbiol 49: 117±143
Shu H-H, Stuart K (1993) A Trypanosoma brucei minicircle en-codes the same gRNAs as do minicircles of T. equiperdumATCC 30019 and T. evansi type A minicircles. Nucleic AcidsRes 21: 2951
Simpson L, Berliner J (1974) Isolation of kDNA from Leishmaniatarentolae in the form of a network. J Protozool 21: 382±393
Southern EM (1975) Detection of speci®c sequences among DNAfragments separated by gel electrophoresis. J Mol Biol 98: 503±517
Sternberg J, Tait A, Haley S, Wells JM, Lepage RWF, Schweizer J,Jenni L (1988) Gene exchange in African trypanosomes: char-
acterisation of a new hybrid genotype. Mol Biochem Parasitol27: 191±200
Sternberg J, Turner CMR, Wells JM, Ranford-Cartwright LC,Lepage RWF, Tait A (1989) Gene exchange in African try-panosomes: frequency and allelic segregation. Mol BiochemParasitol 34: 269±280
Tait A, Turner CMR, Le Page RWF, Wells JM (1989) Geneticevidence that metacyclic forms of Trypanosoma brucei are dip-loid. Mol Biochem Parasitol 37: 247±256
Turner CMR, Sternberg J, Buchanan N, Smith E, Hide G, Tait A(1990) Evidence that the mechanism of gene exchange in Try-panosoma brucei involves meiosis and syngamy. Parasitology101: 377±386
Turner CMR, Hide G, Buchanan N, Tait A (1996) Trypanosomabrucei: inheritance of kinetoplast DNA maxicircles in a geneticcross and their segregation during vegetative growth. Exp Pa-rasitol 80: 234±241
Wells JM, Prospero TD, Jenni L, Le Page RWF (1987) DNAcontents and molecular karyotypes of hybrid Trypanosomabrucei. Mol Biochem Parasitol 24: 103±116
488