kinetics of enzymatic modification of the protamines and a ... fileproposal for their binding to...
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 247, No. 24, Issue of December 25, pp. 7962-7968, 197
Printed in U.S.A.
Kinetics of Enzymatic Modification of the Protamines and a Proposal for Their Binding to Chromatin*
(Received for publication, April 25, 1972)
ANDREW J. LOUIEJAND GORDON H. DIXON~
From the Department of Biochemistry, The University of British Columbia, Vancouver 8, B.C., Canada
SUMMARY
Trout testis maturation is characterized by the complete replacement of the histones by the protamines. During this process, the protamines are extensively modified by enzy- matic phosphorylation of their seryl residues. In addition, a methionyl residue is transiently incorporated at the NH2 terminus of newly synthesized protamine. However, these modifying groups are removed by the time that spermatid maturation is complete. To study the kinetics of these trans- formations, trout testis cells were incubated with [3H]arginine, inorganic [32P]phosphate, and [%]methionine for varying times and the labeled protamines were separated by starch gel electrophoresis. At least six modified bands (one methionyl-(PoM), three phosphoryl-(Pr, P2, P3), and two phospho methionyl-(PIM, P2M) derivatives of unmodified protamine (PO)) were observed. Analysis of radioactivity showed that within 5 to 10 min, newly synthesized protamine (PoM) is phosphorylated to PrM and P2M. Methionine is then removed to form Pi and PZ. Phosphorylation of Pr and Pz continues, and labeled arginine is found in P3 after 5 to 10 hours. Very little arginine label was seen in unmodified protamine (PO) in these in uifro studies. However, 5 to 10 days after injection of [3H]arginine, label was found in unsub- stituted protamine (PO) derived from the sequential dephos- phorylation of Pz. These data suggest that pools of each modified species of protamine exist; molecules within a given pool are modified at random by the appropriate enzyme and become part of another pool. The phosphorylation and de- phosphorylation of protamine appears to be unidirectional. The long lapse (5 to 10 days) between appearances of 13H]- arginine in newly synthesized protamine (PoM) and in fully mature, unmodified protamine may be a consequence of the label having to pass through the various pools of protamine. The controlled phosphorylation of protamine may be impor- tant in the correct binding of protamine to DNA, while the dephosphorylation of protamine is related to the controlled condensation of the spermatid chromatin.
* This research was supported by the Medical Research Council, National Research Council, and National Cancer Institute of Canada.
$ Recipient of a National Research Council of Canada Cen- tennial Science Scholarship.
Q Present address, Department of Biochemistry, University of Sussex, Sussex, England.
During spermatogenesis in trout testis, the histones, arginine- lysine-rich structural proteins of chromatin, are replaced (I, 2) by the protamines, a class of sperm-specific, small proteins (mol wt 5000) very rich in arginine. The complete sequences of three components of rainbow trout (Salmo irideus) protamine have been determined (3), and, of the 32 to 33 amino acid resi- dues, 21 to 22 are arginyl and 3 to 4 are seryl residues.
Protamine is synthesized in the cytoplasm (4) by the usual mechanisms of protein synthesis (1, 4) and rapidly transported (half-time approximately 1 to 2 min) into the nucleus (4). Ingles and Dixon (5) noted previously that 75$& of the protamine seryl residues in early protamine stage testis were phosphorylated, while the protamine of mature sperm had only 5% of its seryl residues phosphorylated. Experiments involving parallel up- take of [i*C]serine and inorganic [32P]phosphate (5) indicated that all of the seryl residues newly incorporated into protamine were phosphorylated. Coupled with evidence that protamine was phosphorylated in the cytoplasm (6), it was postulated that phosphorylation of protamine might be involved in the removal of nascent protamine from the ribosome or the transport of protamine into the nucleus (6, 7).
Ingles and Dixon (5) characterized some of the tryptic phos- pho-peptides from protamine including a tripeptide, Val-Ser(P)- Arg. More recently, Sanders and Dixon (8) have shown the existence of an Arg-Ser(P)-Ser(P)-Ser(P)-Arg-Pro-Val-Arg pep- tide (presumably from the protamine components with four serines) and an Arg-Ser(P)-Ser(P)-Arg-Pro-Val-Arg peptide (presumably from the protamine component with three serines). This evidence supported the possibility that phospho-protamines existed with all of their seryl residues phosphorylated.
In addition, it was demonstrated (9) that methionine was in- corporated at the NH2 terminus of nascent protamine and later removed. These observations indicated that methionine was involved in the initiation of protein synthesis in eucaryotic cells.
By separating trout testis cells on serum albumin gradients and analyzing their basic nuclear proteins on starch gels, we have recently shown (10, 11) that protamine is synthesized and phosphorylated in middle and late stage spermatids. Asso- ciated with the appearance of protamine was the loss of histone (10, 11). On urea starch gels, at least eight bands of protamine were observed (11) ; two were unmodified “mature” protamines as found in mature sperm while the others were phosphorylated. This prompted us to examine the kinetics of incorporation of [3H]arginine into the various protamine bands in order to char- acterize them and to explore the possible biological function(s) of protamine phosphorylation and dephosphorylation.
7962
by guest on March 17, 2019
http://ww
w.jbc.org/
Dow
nloaded from

7963
We have used hormonally induced (12) rather than naturally maturing trout testis because the accelerated development of hormonally induced testis makes it possible to obtain testis with a larger proportion of modified protamine.
EXPERIMENTAL PROCEDURE
Materials-nL$H]Arginine (specific activity 12 Ci per mmole) and [35S]methionine (specific activity 430 to 575 Ci per mmole) and NCS solubilizer were obtained from Amersham-Searle; car- rier-free inorganic [32P]phosphate from Atomic Energy of Can- ada; penicillin and streptomycin from Baltimore Biologicals; Nonidet P-40 from Shell Oil; glass fiber filters (AP20-025-00) from Millipore Filter Corp., Bedford, Mass.; variable thickness starch gel slicer from Richmond Scientific Co., Richmond, B.C., Canada.
Trout Testis-Testes at the prot,amine stage of development were excised from rainbow trotit in which spermatogenesis had been induced by injection of salmon pituitary extracts (1, 12). Maximum testis wet weight and onset of protamine biosynthesis was reached about 50 to 60 days after start of the twice-weekly injections (1).
Cell Incubations-Cell suspensions were prepared as previously described (10) by mincing tissue with scissors in 3 volumes of Medium A-0.1 % glucose (50 InM Tris-HCl, pH 7.5, 5 my MgC&, 25 mM KCl, 0.25 M sucrose, and 0.1% glucose) and gently hand homogenizing (three strokes up and down) in a Potter-Elvehjem homogenizer with a Teflon pestle. The cell suspension was filtered through four layers of cheesecloth.
The incubation mixtures (1 ml total volume) consisted of 3 to 4 X lo8 cells, radioactivity (one or two of the following: 25 to 200 PCi of [3H]arginine, 200 PCi of [3%]methionine, or 300 &i of inorganic [azP]phosphate adjusted to pH 7.2 with NaOH and Tris buffer), 0.2 ml of Waymouth’s medium (13) (minus arginine or methionine as required and 10 mM Tris buffer, pH 7.2, instead of phosphate buffer), phenol red, 100 units of peni- cillin and streptomycin, and Medium A-0.1% glucose to a final volume of 1.0 ml. All incubations were carried out at 15-16” on a gyratory water bath with occasional mixing to resuspend sedimented cells. Control studies showed that incorporation of labeled arginine or inorganic [32P]phosphate was linear for at least 12 hours with a slight tapering off at 24 hours. In studies using inhibitors either of protein synthesis or of energy me- tabolism, cells were incubated with label as above for 10 min before addition of cycloheximide (2 X lop4 M final concentration) or 2,4-dinitrophenol (2 X 10F4 M final).
At. various times, 100 ~1 (3 to 4 X lo8 cells) were removed and diluted with 1 ml of cold Medium A. Cells were immediately fixed with 3 to 4 volumes of 95% et,hanol and filtered onto glass fiber filters over an area slightly smaller than the size of the starch gel sample slot (6 X 10 mm) with the aid of a Teflon template. The fixed cells were washed with 95% ethanol and the filters were dried.
In Viva Labeling-Rainbow trout (weight about 100 g) at the early protamine stage (55 to 60 days after initiation of hormonal injection) were injected intraperitoneally with 300 PCi of [3H]- arginine in 0.4 ml of 0.9% NaCl solution. At various times, fish were killed. The testes were excised and cell suspensions were prepared essentially as above. Cells (2.5 x 108) were filtered onto glass fiber discs (10, ll), washed with 2 X 5 ml of dedium A, then 2 X 5 ml of 0.5% Nonidet P-40 in Medium A,
and fixed with 95% ethanol and dried. The Nonidet P-40 wash stripped the cells of any cytoplasm and hence ribosomes (11, 14). Control studies showed that nuclear histones and protamines
were not removed by this wash. For electrophoresis, a 6 X 10 mm2 rectangle (containing ~4 X 10’ cells) was cut from the filter.
Starch Gel Electrophoresis-Starch gels were prepared as pre- viously described (15). Basic proteins were extracted and gel electrophoresis was conducted as in Reference 10. After elec- trophoresis, the gels were trisected horizontally. The bottom slab was stained by the sensitive cobalt-Amid0 black 10B pro- cedure and destained with sulfuric acid (15).
The protamine fractions were well separated from the histones and any ribosomal proteins (10, 11). For analysis of radioac- tivity incorporated into the protamines, the middle slab of the gel was cut with a slab-gel slicer into 2-mm slices which were then incubated with 0.5 ml of NCS solubilizer for 8 to 12 hours at room temperature (11). By the end of this period, the opaque gel had become transparent. Five milliliters of toluene scintilla- tion fluid (0.01% 1,4-bis[2-(5-phenyloxazolyl)]benzene (POPOP), 0.4% 2,5-diphenyloxazole (PPO) per liter) were then added, and the capped vials were incubated for a further 3 hours at 45” before counting in a Nuclear Chicago-Unilux II counter. Trit- ium counts were corrected for 30% ?S counts in the tritium channel.
RESULTS
To facilitate discussion of the results, Fig. 1 shows the starch gel electrophoresis pattern and incorporation profiles of inorganic [32P]phosphate and [35S]methionine into protamine. The various bands are labeled according to the 32P and 35S incorporation. The two fastest bands (PoA and POB) represent the two major species of unphosphorylated protamine, while the slower bands P1, Pz, and PS represent protamine with one, two, and three seryl phosphates respective1y.l Wigle and Dixon (9) showed that methionine was incorporated at the NH2 terminus of newly synthesized protamine and that the NH2 terminus of the methi- onyl residue was not blocked. When 35S-labeled protamine is subjected to electrophoresis on starch gels, the 35S pattern is con- sistent neither with protein staining nor with 32P incorporation. For example, a [%]methionine-labeled protamine species mi- grates slightly behind unmodified protamine (PO) ; this we inter- pret as newly synthesized protamine with a methionyl residue at its NH2 terminus (i.e. PcM). It is not clear why the presence of the extra methionyl residue decreases the mobility although methionyl-protamine would have molecular weight approxi- mately 3% greater than PO. Since the NH2 terminus is not blocked, the net charge is the same and the increase in molecular
1 Six phosphorylated species of protamine have been separated by chromatography on CM-cellulose. Amino acid analysis, phos- phate determinations, and starch gel electrophoresis have shown that three of these phosphorylated species correspond to PI, PZ, and P) of the A component of protamine, while the other three correspond to a similar series of the B component. Although PoA and PoB are nartiallv resolved in these gels. it is not clear why the phosphorylated components of A and B are not similarly separated. It is possible that random phosphorylation of the seryl residues gives rise to phosphoprotamines of the same net charge but slightly different conformations; phospho A and B
might then run together as a single broad band. It has also been shown that alkaline phosphatase treatment converts these deriv- atives into dephosphoprotamine (A. J. Louie and G. H. Dixon, manuscript in preparation). These analyses, which show the presence of one, two, and three phosphoryl residues per molecule are the basis for the assignment of the labels PI, P,, and P, in Fig. 1. Although all four serines can be phosphorylated (8), it should be noted that the most phosphorylated component, PB, contains three phosphoryl residues, and not four as previously suggested (II).
by guest on March 17, 2019
http://ww
w.jbc.org/
Dow
nloaded from

7964
FIG. 1. Nomenclature of protamine bands obtained by starch gel electrophoresis. A sample of whole protamine labeled with inorganic [azP]phosphate (250 uCi per ml. 14 hours) and another labeiedwiih [S%]meihionine (56pC;perml;S hours) were subjected to electrophoresis on starch gels in the presence of urea (15). The gel was trisected horizontally and the bottom slab was stained (photograph). The protamine region of the middle slab was sliced into 2-mm slices and each slice was solubilized and counted for radioactivitv. The s2P and 3% counts from the two samoles are superimposed. Electrophoresis is from left to right. 6; more phosphorylated components migrate more slowly than un- modified protamine. The various protamine bands are labeled as follows. “P” represents the protamine peptide chain; the subscript numerals represent the number of ser.yl residues esteri- fied by phosphate and the suffix “M” indicates the presence or ab- sence of NHS-terminal methionine. PO is unmodified motamine and is the major form of protamine fouhd in mature sperm. Two components of PO (PoA and PoB) are partially resolved by starch gel electrophoresis.
weight would reduce the mobility somewhat. There are cor- responding species, PIM and PzM, which migrate slightly slower than monophospho-(PI) and diphospho-(Pz) protamine. Tri- phospho protarnine (Pa) is the slowest migrating species and there appears to be no more than a trace of this species with methionine at the NH2 terminus.
Incorporation of [8H]Arginine and Inorganti [a2P]Phosphate into Protamine and Effect of Metabolic Inhibitors-To examine the incorporation of [aH]arginine and inorganic [a2P]phosphate into protamine and the effects of an inhibitor of protein synthesis and an uncoupler of oxidative phosphorylation, cells were incu- bated with [aH]arginine and [a2P]phosphate. After 10 min, the incubation mixture was divided into three portions, cyclohexi- mide was added to one, 2,4-dinitrophenol to another, and the third served as control. Incubation was continued in each case for another 5% hours. Cells were removed and processed as described under “Experimental Procedure.” Fig. 2 shows the protein stain and radioactivity profiles of the protamine region from these incubations. The fastest peak of [aH]arginine not associated with any protein band is free arginine. After 10 min of labeling (Fig. 2A) three sharp peaks of [aH]arginine which do not correspond exactly to the protein bands in the protamine region are seen. These three peaks of aH label are probably methionyl derivatives of PO, PI, and PZ protamine. However, very little [a2P]phosphate has been incorporated. The incorpora- tion of [a2P]phosphate into whole protamine is linear with time and after 546 hours (Fig. 2B) significant incorporation of [“PI- phosphate is observed. In addition, the profile of aH label has changed and shows a broad central region of incorporation with small peaks of arginine label in the slowest (Pa) and fastest (PO) regions of protamine. It is apparent from Fig. 2, A and B, that
FIG. 2. Incorporation of [sH]arginine and inorganic [a2P]phos- nhate into nrotamine and the effect of a nrotein svnthetic inhibitor and an uncoupler of oxidative phosphorylation. Cells were la- beled with 200 #i per ml of [SHJarginine and 250 pCi per ml of inorganic [3*P]phosphate. After 10 min of incubation, the mix- ture was divided into three portions and cycloheximide (2 X lo-” Y final concentration) was added to one, 2,4-dinitrophenol (2 X 10-d M final) was added to another, while the third served as control. At 10 min and 5j/2 hours of incubation, 3 X lo7 cells were removed, fixed with ethanol, and collected on glass fiber filters. Acid-sol- uble proteins were extracted and sepirated in starch gels. After electrophoresis, the gel was trisected horizontally, the bottom slab was stained (only the protamine region of the gel is shown in the photograph), the middle slab was sliced into 2-mm slices, and each slice was solubilized and counted for radioactivity. A, 10 min of labeling; B, 5x-hour control; C, 5W-hour cycloheximide (CHX) inhibited; D, 5x-hour 2,4-dinitrophenol (DNP) inhibited.
the kinetic relationship of the different. protamine species is rather complex.
Fig. 2C shows the effect of cycloheximide on the patterns of [aH]arginine and [a’JP]phosphate incorporation. The inhibitor has drastically decreased the synthesis of protamine (namely the difference in ordinate scales between Fig. 2, B and C), yet phos- phorylation of arginine-labeled protamine has continued as judged by the appearance of slower running peaks of label and the relatively unchanged incorporation of [a2P]phosphate. We can conclude that phosphorylation of protamine occurs inde- pendently of protein synthesis as previously shown (5), since the phosphorylation of newly synthesized protamine can continue in the presence of cycloheximide. We have previously calculated (10) that the approximate rate of protamine biosynthesis is 1 molecule per min per ribosome. Because of the rapid transport
by guest on March 17, 2019
http://ww
w.jbc.org/
Dow
nloaded from

7966
With increasing time, the proportion of arginine label increases in Pp, Pz, and P1 with a corresponding decrease in PzM and PIM. After 5?6 hours, the proportion of arginine label in P1 and PZ is about equal (Fig. 3F). However, this too changes, with the proportion of 3H label in P1 decreasing and PZ increasing with time (Fig. 3, G to 1). The distribution of ]35S]methionine is essentially unchanged from the beginning of the incubation (although the total %S content increases) and serves as a useful
FIG. 4. In vivo incorporation of [aH]arginine into protamine. Rainbow trout at the early protamine stage (55 days hormonal injection) were injected intraperitoneally with 350 PC1 of [3Hl- arginine in 0.4 ml of 0.9% NaCl solution. At the indicated times, a fish was killed, testes excised, and a cell suspension prepared. Cells (25 X 107) were filtered onto a glass fiber filter, washed with Medium A, and then Medium A-0.5’% Nonidet P-40 to strip the cells of cytoplasm and hence remove ribosomes. The nuclei were then fixed with ethanol. Acid-soluble proteins were extracted from 4 X lo7 nuclei, separated on starch gels, and processed as in Fig. 2. A, 1.0 day post injection; B, 4.0 days; C, 7.0 days; D, 10.0 days.
reference for the position of the arginine label in the various protamine bands.
After the initial 30 min of labeling, and even after 24 hours of labeling, only a small proportion of the labeled arginine is found in the unmodified protamine region. This implies that the phos- phorylation of protamine is unidirectional and follows a definite sequence. Thus P, labeled with arginine is phosphorylated to PZ and not dephosphorylated to PO (Fig. 31). Two questions were then posed. (a) How long does it take newly synthesized protamine to be enzymatically phosphorylated and completely dephosphorylated?; and (b) what is the biological significance of phosphorylation and dephosphorylation of protamine?
In Viva Incorporation of [aH]Arginine into Profamine-In order to observe the passage of labeled arginine into completely un- modified protamine, live trout were labeled with [3H]arginine in viva. Fig. 4, A to D, shows the labeled arginine profiles of the protamine region from these in viva experiments. After 1 or 2 days of incubation, no additional arginine-labeled protamine is synthesized (no significant labeling in the PoM, PIM, and PzM regions). This indicates that the injected label was rapidly utilized or completely diluted by endogenous unlabeled arginine.
Fig. 4 shows that even after 1 and 4 days of labeling, essen- tially no [3H]arginine is found in the unmodified (PO) protamine, while after 7 and 10 days of incubation, a significant proportion of label is found in PO. The proportion of label in P1 decreases from days 1 to 4 and increases from days 4 to 7 and 10. Pre- sumably these changing levels reflect the continued phosphoryla- tion of Pr protamine to PZ and Pa on one hand and the dephos- phorylation of Pa and Pz to P1 on the other hand. Thus it takes about 5 to 10 days for newly synthesized protamine to go through the series of phosphorylations and dephosphorylations.
Over-all Kinetics of Methionine Removal and Phosphorylation and Dephosphoylation of Protamine-Wigle and Dixon (9) showed that methionine was removed from the NH2 terminus of newly synthesized protamine, but, in their a-hour chase, only 50 y. of the label had been removed. In a situation in which one label (methionine) is being removed at a constant rate from mole- cules while the other (arginine) is stable, it would be expected that the ratio of [%]methionine to [“H]arginine would decrease linearly with time. Fig. 5 shows the ratio of 35S:3H for the PoM
L ’ * ’ “‘0
’ ’ ’ ’ 1 2 4 6 6 IO 12 14 16
HOURS
FIG. 5. Specific activity of [3%]methionine from the incubation of Fig. 3. a% and 3H counts were integrated over the PoM and whole protamine regions of Fig. 3, A to H. The ratio of %:3H was plotted at the various times. O--O, whole protamine; a---o, PoM.
by guest on March 17, 2019
http://ww
w.jbc.org/
Dow
nloaded from

of protamine into the nucleus (half-time 1 to 2 min, Reference 4) most of the phosphorylation of protamine must take place inside the nucleus and not in the cytoplasm as previously suggested (6).
The effect of an uncoupler of oxidative phosphorylation, 2,4- dinitrophenol, is shown in Fig. 20. The [3H]arginine profile changes only slightly after the addition of inhibitor, there being no formation of Pa and much less Pz. In line with this, no more [“PIphosphate is incorporated into protamine. The observed increase of P1 at the expense of PO in Fig. 20 probably represents phosphorylation that has taken place before 2,4-dinitrophenol has caused the exhaustion of the ATP pool. Thus 2,4-dinitro- phenol has abolished both protein synthesis and the conversion of labeled protamine into phosphorylated derivatives.
Incorporation of Labeled Methionine and Arginine into Prota- mine as a Function oj Time-To study the kinetic relationship of the different protamine bands, cells were doubly labeled with [35S]methionine and [3H]arginine. At various times, aliquots were removed and acid-soluble proteins were subjected to elec- trophoresis on starch gels. Fig. 3 shows the staining pattern obtained and the radioactivity profiles of the protamine region after starch gel electrophoresis. The separation is slightly different from that in Fig. 2 because of variations in starch gels prepared from different lots of starch. The fastest peak of
7965
labeled methionine not associated with protein staining is free methionine which migrates slightly slower than free arginine.
The profiles of 13H]arginine after different periods of labeling (Fig. 3, A to 1) show that the band which is first labeled with [3H]arginine is migrating in the position of PoM (Figs. 1 and 3A) and represents nascent protamine with methionine at its NH2 terminus. Thereafter, the pattern of arginine labeling changes rapidly and after 10 min label appears in two other bands, P,M and P2M. Thus phosphorylation occurs very shortly after protamine synthesis. At 30 min, the % and 3H profiles of la- beled protamine coincide; however after 135 to 3 hours (Fig. 3, D and E) they no longer coincide, and it is apparent that the arginine label formerly in PIM and P2M is migrating slightly faster in the PI and Pz regions, respectively. This must be due to the removal of methionine from the NH2 terminus of PIM and P2M in the nucleus to form P1 and Pz, respectively. There is a small proportion of [%S]methionine label in the P3 region of Fig. 3, B to G and this may represent P3M.
The profiles of labeled arginine in protamine separated by gel electrophoresis change most dramatically during the first 30 min of labeling (Fig. 3, A to C) and thereafter the changes are less pronounced (Fig. 3, D to I). Only after 3 hours does a significant proportion of arginine label appear in the P3 band of protamine
FIG. 3. Incorporation of [aH]arginine and [%]methionine into protamine as a function of time. A testis cell suspension was incubated with [aH]arginine (50 pCi per ml) and [3SS]methionine (200 pCi per ml). At various times, 3.2 X 10’ cells were removed and processed as in Fig. 2. A, 3 min labeling; B, 10 min; C, 30
min; D, lx-hours; E, 3.0 hours; F, 5?4 hours; G, 9.0 hours; H, 14.0 hours; I, 24.0 hours. -, 3H counts; - --, 8% counts. The intensely stained protein band at the far left of the photograph corresponds to histone T, which has been characterized by Wigle and Dixon (16).
by guest on March 17, 2019
http://ww
w.jbc.org/
Dow
nloaded from
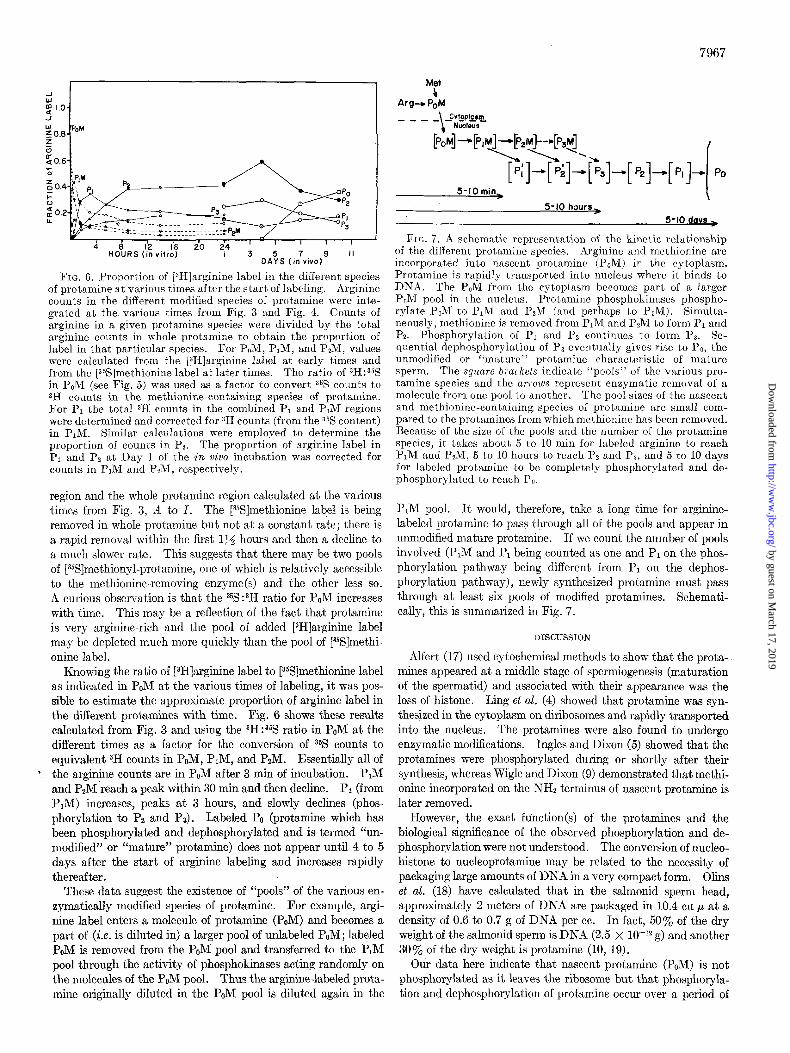
7967
Met h
Arg+ PoM
SO.6 t z 00.4 G :: E 0.2 Lo.
HOURS (in vitro) I3 5 7 9 H DAYS (in viva)
FIG. 6. Proportion of [zH]arginine label in the different species of protamine at various times after the start. of labeling. Arginine counts in the different modified species of protamine were inte- grated at the various times from Fig. 3 and Fig. 4. Counts of arginine in a given protamine species were divided by the total arginine counts in whole protamine to obtain the proportion of label in that particular species. For PoM, PIM, and P2M, values were calculated from the [zH]arginine label at early times and from the [3%]methionine label at later times. The ratio of 3H:35S in PoM (see Fig. 5) was used as a factor to convert % counts to SH counts in the methionine-containing species of protamine. For PI the total 3H counts in the combined PI and P,M regions were determined and corrected for 3H counts (from the 3% content) in PIM. Similar calculations were employed t,o determine the proportion of counts in Pz. The proportion of arginine label in PI and Pz at Day 1 of the in viva incubation was corrected for counts in PIM and PM, respectively.
region and the whole protamine region calculated at the various times from Fig. 3, B to 1. The [YSJmethionine label is being removed in whole protamine but not at a constant rate; there is a rapid removal within the first 134 hours and then a decline to a much slower rate. This suggests that there may be two pools of [%S]methionyl-protamine, one of which is relatively accessible to the methionine-removing enzyme(s) and the other less so. A curious observation is that the 35S:3H ratio for PoM increases with time. This may be a reflection of the fact that protamine is very arginine-rich and the pool of added [3H]arginine label may be depleted much more quickly than the pool of [%S]methi- onine label.
Knowing the ratio of [3H]arginine label to [%]methionine label as indicated in PoM at the various times of labeling, it was pos- sible to estimate the approximate proportion of arginine label in the different protamines with time. Fig. 6 shows these results calculated from Fig. 3 and using the 3H:35S ratio in PoM at the different times as a factor for the conversion of 35S counts to equivalent 3H counts in PoM, PIM, and PzM. Essentially all of the arginine counts are in PoM after 3 min of incubation. PIM and P2M reach a peak within 30 min and then decline. P1 (from PIM) increases, peaks at 3 hours, and slowly declines (phos- phorylation to Pz and Ps). Labeled PO (protamine which has been phosphorylated and dephosphorylated and is termed “un- modified” or “mature” protamine) does not appear until 4 to 5 days after the start of arginine labeling and increases rapidly thereafter.
These data suggest the existence of “pools” of the various en- zymatically modified species of protamine. For example, argi- nine label enters a molecule of protamine (PoM) and becomes a part of (i.e. is diluted in) a larger pool of unlabeled POM; labeled POM is removed from the PoM pool and transferred to the PIM pool through the activity of phosphokinases acting randomly on the molecules of the PoM pool. Thus the arginine-labeled prota- mine originally diluted in the P&I pool is diluted again in the
5-10 min I
5-10 hours,
5-10 davr >
FIG. 7. A schematic representation of the kinetic relationship of the different protamine species. Arginine and methionine are incorpora,ted into nascent protamine (P,M) in the cytoplasm. Protamine is rapidly transported into nucleus where it binds to DNA. The PoM from the cytoplasm becomes part of a larger P&I pool in the nucleus. Protamine phosphokinases phospho- rylate PoM to PIM and PzM (and perhaps to P3M). Simulta- neously, met.hionine is removed from PIM and PzM to form PI and Pz. Phosphorylation of PI and PZ continues to form Pa. Se- quential dephosphorylation of Pa eventually gives rise to PO, the unmodified or “mature” protamine characteristic of mature sperm. The square brackets indicate “pools” of the various pro- tamine species and the arrozOS represent enzymatic removal of a molecule from one pool to another. The pool sizes of the nascent and methionine-containing species of protamine are small com- pared to the protamines from which methionine has been removed. Because of the size of the pools and the number of the protamine species, it takes about 5 to 10 min for labeled arginine to reach PIM and PsM, 5 to 10 hours to reach PZ and Pa, and 5 to 10 days for labeled protamine to be completely phosphorylated and de- phosphorylated to reach P,.
PIM pool. It would, therefore, take a long time for arginine- labeled protamine to pass through all of the pools and appear in unmodified mature protamine. If we count the number of pools involved (P,M and Pr being counted as one and P1 on the phos- phorylation pathway being different from PI on the dephos- phorylation pathway), newly synthesized protamine must pass through at least six pools of modified protamines. Schemati- cally, this is summarized in Fig. 7.
DISCUSSION
Alfert (17) used cytochemical methods to show that the prota- mines appeared at a middle stage of spermiogenesis (maturation of the spermatid) and associated with their appearance was the loss of histone. Ling et al. (4) showed that protamine was syn- thesized in the cytoplasm on diribosomes and rapidly transported into the nucleus. The protamines were also found to undergo enzymatic modifications. Ingles and Dixon (5) showed that the protamines were phosphorylated during or shortly after their synthesis, whereas Wigle and Dixon (9) demonstrated that methi- onine incorporated on the NH2 terminus of nascent protamine is later removed.
However, the exact funct.ion(s) of the protamines and the biological significance of the observed phosphorylation and de- phosphorylation were not understood. The conversion of nucleo- histone to nucleoprotamine may be related to the necessity of packaging large amounts of DNA in a very compact form. Olins et al. (18) have calculated that in the salmonid sperm head, approximately 2 meters of DNA are packaged in 10.4 cu p at a density of 0.6 to 0.7 g of DNA per cc. In fact, 50% of the dry weight of the salmonid sperm is DNA (2.5 X lo+ g) and another 30% of the dry weight is protamine (10, 19).
Our data here indicate that nascent protamine (PaM) is not phosphorylated as it leaves the ribosome but that phosphoryla- tion and dephosphorylation of protamine occur over a period of
by guest on March 17, 2019
http://ww
w.jbc.org/
Dow
nloaded from

7968
several days (Fig. 7). Thus, protamine phosphorylation appears not to be required for the removal of protamine from the ribosome or transport into the nucleus (7) ; in fact, most of the phosphoryla- tion must occur in the nucleus. A supply of ATP is required, both for the phosphorylation of protamine (Fig. 2), and its transport into the nucleus (7), as indicated by the inhibition of these processes by 2,4-dinitrophenol.
The existence of “pools” of the various modified species of protamine and their large number may account for the extremely long time (5 to 10 days) it takes for nascent protamine (POM) to be completely phosphorylated and dephosphorylated and become “mature” protamine (PC). The PoM, PiM, and PzM pools are part of two larger pools of methionine-containing protamine; in one pool, methionine removal occurs relatively rapidly, while in the other this process appears to be relatively slow. The phos- phorylation and dephosphorylation of protamine appears to be a closely regulated unidirection process. Thus, phosphorylation and dephosphorylation of newly synthesized protamine is an obligatory process and molecules in the phosphorylation pathway are metabolically distinct from those in the dephosphorylation pathway, even though they may have the same number of phos- phates (e.g. Pi and PI in Fig. 7).
The sequence and unidirectionality of these events together with the differing metabolic behavior of the different protamine pools suggest to us that the sequential phosphorylation of prota- mine is involved in some way in the proper binding of newly synthesized protamine to chromatin.
In fact, we have shown by cell separation techniques (10) that in the 1.5 S, spermatidsz which have just begun to synthesize protamine, the protamines present are exclusively in the phos- phorylated state. Associated with the transition of the 1.5 S, spermatid to the smaller 1.0 X, spermatid is the appearance of un- modified “mature” protamine. It takes about 1 week for 1.5 S, spermatids to be transformed into the smaller 1.0 S, spermatids (10). This time interval correlates rather well with the 5 to 10 days required for newly synthesized protamine to be completely phosphorylated and dephosphorylated. Similarly, the transi- tion of 1.0 X, spermatids to 0.6 S, mature sperm is associated with the dephosphorylation of residual phosphorylated prota- mine. These data suggest that it is the dephosphorylation of phosphorylated protamine that may cause the contraction and condensation of the spermatid chromatin.
The concept of enzymatic modifications being involved in the correct binding of basic proteins to nucleic acids is a relatively new one.3 A priori, the probability of a highly positively charged molecule such as protamine entering the nucleus and binding cor- rectly through ionic linkages to the negatively charged molecule of DNA would seem to be slight. Hence, some mechanism for the regulation of the binding of such highly charged polycationic molecules would seem to be required, assuming that the final, correct binding is highly specific and ordered.
The model which we propose is that nascent protamine is syn-
2 8, represents the sedimentation velocity constant of cells sedimenting at unit gravity in serum albumin gradients. Larger cells have a larger S, value and hence sediment faster (Stoke’s Law) than cells with a smaller 8, constant (20, 21). In our case, the early 1.5 S, spermatids are larger than the 1.0 S, late spermatid which are in turn larger than the 0.6 S, mature sperm.
3 We have reported elsewhere (22) that the phosphorylation and acetylation of some histones (IIbl and IV, respectively) may be involved with the proper binding of these histones to DNA.
thesized in the cytoplasm and rapidly transported into the nucleus where it binds in a nonspecific fashion to DNA through ionic linkages. Protamine phosphokinases (23) bound loosely to chromatin might then function as “editing” enzymes, which recognize the improperly bound protamine and phosphorylate it. Phosphorylation (by the same or different protamine kinases) continues until the phosphorylated protamine assumes the correct conformation with respect to DNA. Phosphorylation of prota- mine would decrease its net positive charge, thus decreasing the strength of its interaction with DNA and perhaps allowing the protamine more freedom to find its correct conformation. It was shown previously by Marushige et al. (6) that [32P]protamine was bound less tightly to chromatia than unmodified protamine as judged by its elution at lower ionic strengths. Simultane- ously, histone is removed in some as yet unknown manner al- though it is possible that phosphorylation of protamine may have some direct role in the removal of histone. Finally, protamine phosphatases, which also act as “editing” enzymes, recognize those phosphorylated protamine molecules bound in the correct conformation and remove the protamine seryl phosphates, thus increasing the net positive charge on protamine and “locking” it to DNA. Thus, the progressive removal of seryl phosphates from protamine would gradually contract the spermatid chro- matin into a highly compact state characteristic of mature sperm (Fig. 7).
1.
2.
3.
4.
5.
6.
7.
8.
9. 10.
11.
12.
13. 14.
15. 16.
17. 18.
19.
20.
21.
22.
23.
REFERENCES INGLES, C. J., TREVITHICK, J. R., AND DIXON, G. H. (1966)
Biochem. Biophys. Res. Commun. 22, 627-634 MARUSHIGE, K., AND DIXON, G. H. (1969) Develop. Biol. 19,
397-414 ANDO, T., AND WATANABE, S. (1969) Int. J. Protein Res. 1,
221-224 LING, V., TREVITHICK, J. R., AND DIXON, G. H. (1969) Can. J.
Biochem. 47, 51-60 INGLES, C. J., AND DIXON, G. H. (1967) Proc. Nut. Acad. Sci.
U. S. A. 58, 1011-1018 MARUSHIGE, K., LING, V., AND DIXON, G. H. (1969) J. Biol.
Chem. 244, 5953-5958 DIXON, G. H., INGLES, C. J., JERGIL, B., LING, V., AND MARU-
SHIGE, K. (1968) Can. Cancer Conf. 8, 76-102 SANDERS, M. M., AND DIXON, G. H. (1972) J. Biol. Chem. 247,
851-855 WIGLE, D. T., AND DIXON, G. H. (1970) Nature 227,676-680 LOUIE, A. J., AND DIXON, G. H. (1972) J. Biol. Chem. 247,
5490-5497 LOUIE, A. J., AND DIXON, G. H. (1972) J. Biol. Chem. 247,5498-
5505 SCHMIDT, P. J., MITCHELL, B. S., SMITH, M., AND TSUYUKI, H.
(1965) Gen. Comp. Endocrinol. 5, 197-206 WAYMOUTH, C. (1956) J. Nat. Cancer Inst. 17,315-327 BORUN, T. W., SCHARFF, M. D., AND ROBBINS, E. (1967) Bio-
chim. Biophys. Acta 149,302304 SUNG, M., AND SMITHIES, 0. (1969) Biopolymers 7,39-58 WIGLE, D. T., AND DIXON, G. H. (1971) J. Biol. Chem. 246,
5636-5644 ALFERT, M. (1956) J. Biophys. Biochem. Cytol. 2, 109-114 OLINS, D. E., OLINS, A. L., AND VON HIPPEL, P. H. (1968) J.
Mol. BioZ. 33, 265-281 POLLISTER, A. W., AND MIRSKY, A. E. (1946) J. Gen. Physiol.
30, 101-116 MILLER, R. G., AND PHILLIPS, R. A. (1969) J. Cell. Physiol.
73, 191-201 LAM, D. M. K., FURRER, R., AND BRUCE, W. R. (1970) Proc.
Nat. Acad. Sci. U. S. A. 65, 192-199 LOUIE, A., CANDIDO, P., AND DIXON, G. H. (1972) Fed. Proc.
31, 1121 JERGIL, B., AND DIXON, G. H. (1970) J. Biol. Chem. 246,425-
434
by guest on March 17, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Andrew J. Louie and Gordon H. DixonBinding to Chromatin
Kinetics of Enzymatic Modification of the Protamines and a Proposal for Their
1972, 247:7962-7968.J. Biol. Chem.
http://www.jbc.org/content/247/24/7962Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/247/24/7962.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on March 17, 2019
http://ww
w.jbc.org/
Dow
nloaded from