journal of biological 263, no. 25, biology, u. s. a ... journal of biological chemistry 0 1988 by...
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1988 by The American Society for Biochemistry and Molecular Biology, Inc
Vol. 263, No. 3, Issue of January 25, pp. 1157-1165,1988 Printed in U. S. A.
Comparative Study of the Mucin-type Sugar Chains of Human Chorionic Gonadotropin Present in the Urine of Patients with Trophoblastic Diseases and Healthy Pregnant Women*
(Received for publication, April 13, 1987)
Junko Amano, Ryuichiro NishimuraS, Matsuto MochizukiS, and Akira Kobata From the Department of Biochemistry, the Institute of Medical Science, the University of Tokyo, Minuto-ku, Tokyo 108, Japan and the $Department of Obstetrics and Gynecology, Kobe University School of Medicine, Chw-ku, Kobe 650, Japan
Human chorionic gonadotropins (hCGs) highly puri- fied from the urine of patients with trophoblastic dis- eases and of healthy pregnant women contain approx- imately four mucin-type sugar chains in one molecule. The structures of these sugar chains were studied com- paratively by using a new sensitive method to obtain mucin-type sugar chains quantitatively as radioactive oligosaccharides from a small amount of glycoproteins. The mucin-type sugar chains of all hCGs include sia- lylated and nonsialylated GalS1+3GalNAc and G~1+4GlcNAc~1~6(Ga~l+3)GalNAc. In the case of normal hCG and hydatidiform mole hCG, oligosac- charides containing the tetrasaccharide core occupy approximately 10% of the total mucin-type sugar chains. The ratio of the tetrasaccharide containing oligosaccharides is increased prominently to approxi- mately 60% in choriocarcinoma hCG. The proportion in invasive mole hCG was also increased, but less than the proportion of choriocarcinoma hCG.
Human chorionic gonadotropin (hCG)’ consists of two gly- coprotein subunits, (Y and p. Both subunits contain two as- paragine-linked sugar chains (1,2). In addition, the @-subunit contains four mucin-type sugar chains at Ser’”, SerlZ6, S e P , and Ser13’. The structure of the mucin-type sugar chain of hCG was elucidated to be NeuAccu2+3Gal@l--*3(NeuAc(~2+ 6)GalNAc by Kessler et al. (3).
Although hCG normally is produced by trophoblasts of placenta at the early stage of pregnancy, it has also been detected in the sera and in the urine of patients with various trophoblastic diseases including choriocarcinoma. Because of this, urinary hCG has been used as an important marker for the diagnosis and prognosis of trophoblastic disease as well as pregnancy. So far, no attempt to discriminate hCGs from different diseases has been performed. However, any device that can discriminate the hormones originated from different
*This work has been supported in part by Grants-in-Aid for Special Project Research (Cancer-Bioscience) from the Ministry of Education, Science ahd Culture of Japan. This paper is a part of the dissertation submitted by J. A. to Faculty of Pharmaceutical Science, the University of Tokyo as a requirement for the degree of Doctor of Pharmaceutical Sciences. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
The abbreviations used are: hCG, human chorionic gonadotropin; Sia, sialic acid; NeuNGc, N-glycolylneuraminic acid Fuc, fucose; GalNAc, N-acetylgalactosamine; AraNAc, N-acetylarabinosamine; ThrNAc, N-acetylthreosamine; NDV, New Castle disease virus. Sub- scripts OT and OH are used to indicate NaB3H,- and NaBH,-reduced oligosaccharide, respectively.
trophoblastic diseases will no doubt be beneficial for the accurate diagnosis of each disease.
Nishimura et al. (4) reported that no difference is found in the amino acid composition and in the reactivity with anti- hCG antisera of the hCGs purified from the urine of patients with various trophoblastic diseases. We have reported previ- ously that the structures of the asparagine-linked sugar chains of choriocarcinoma hCG are quite different from those of hCG purified from the urine of normal pregnant women ( 5 ) . Since it remained unclear whether structural change is also induced in the mucin-type sugar chains of choriocarcinoma hCG or not, we have performed this study. In order to analyze the structures of mucin-type sugar chains with limited amounts of glycoprotein samples, a new method has been developed to convert mucin-type sugar chains quantitatively to radioactive oligosaccharides. The results described in this paper indicate that the method is quite useful for the structural study of mucin-type sugar chains in glycoproteins.
EXPERIMENTAL PROCEDURES
[l-3H]Acetic anhydride (50 mCi/mmol) and NaB3H4 (341 mCi/ mmol) were purchased from Du Pont-New England Nuclear. Bio-Gel P-4 (under 400 mesh) was purchased from Bio-Rad. Sialidase (Ar- throbacter ureafaciens) was obtained from Nakarai Chemicals, Ltd., Kyoto. Another sialidase of New Castle disease virus was purified by the method of Paulson et al. (6). @-Galactosidase and @-N-acetylhex- osaminidase were purified from jack bean meal by the method of Li and Li (7). Diplococcal @-galactosidase was purified by the method of Glasgow et al. (8).
Standard oligosaccharides, Siaa2+3Gal@1+3(Siaa2+ 6)GalNAcoT, Siaa2+3Gal@l~3Ga1NAcoT, and Gal@l+3GalNAcoT were isolated from bovine K-casein by treatment with alkaline boro- hydride containing NaB3H,. Siaa2+6GalNAcoT and GalNAcal- 3(Fuca1+2)Ga1@1+3(Siaa2+6)GalNAcoT were prepared from sub- maxillary mucin purified from pig of blood type A. The porcine submaxillary mucin was kindly provided by Dr. Naohisa Kochibe, Gunma University.
Gal@l+4GlcNAcoT was prepared as follows. One hundred fifty microliters of 100 mM sodium cacodylate buffer, pH 7.3, containing 40 nmol of UDP-galactose, 250 nmol of N-acetylglucosamine, 1 pmol of MnCl,, and 2.5 milliunits of bovine milk galactosyltransferase (Sigma) were incubated at 37 “C for 5 min. The mixture was passed through a column containing 1 ml of AG 1-X3 (C1- form), and the product in the effluent was reduced with NaB3H4.
Radioactive 2-deoxy-2-acetamidoarabinitol (AraNAcoT), 2-deoxy- 2-acetamidothreitol (ThrNAcoT), and 2-acetamidopropandiol were prepared by partial oxidation of N-acetyl[l-3H]galactosaminitol (10 nmol) with 40 nmol of sodium metaperiodate and purified by paper chromatography using solvent I as reported by Takasaki et al. (9). Galp1+3ThrNAcoT was prepared by complete periodate oxidation of the Siaa2~3Gal@1+3GalNAco~. The values of these stand- ard samples on the paper chromatogram were: AraNAcoT, 1.52; ThrNAcoT, 1.79; 2-acetamidopropandiol, 2.21; and Gal@1+ 3ThrNAcoT, 0.79.
Analytical Methods-Radioactivity was determined by Aloka liquid
1157

1158 Mucin-type Sugar Chains of Various hCGs scintillation spectrometer (LSC-700). Radioactivity on paper was detected by Packard radiochromatogram scanner model 7201. Bio- Gel P-4 column chromatography was performed as reported previ- ously (10). High voltage paper electrophoresis was performed by using pyridine/acetate buffer, pH 5.4, a t 72 volts/cm for 1.5 h. Descending paper chromatography was performed by using butanol-l/ethanol/ water, 4:l:l by volume (solvent I), or ethyl acetate/pyridine/acetic acid/water, 5:5:1:3 by volume (solvent 11). Methylation analysis of oligosaccharides was performed on the samples obtained by alkaline- NaB2H4 degradation of hCG as described previously (11). Periodate oxidation study was performed according to the method reported previously (12).
hCG Samples-hCG samples were purified from the urine of four patients with complete hydatidiform mole, YAM, ITO, AMA, and HAG; two patients with invasive mole, SHI and KAM; three patients with choriocarcinoma, MAT, MIT, and KUN, and two normal preg- nant women, YNA and YNB, as well as from the pooled urine of normal pregnant women. The case histories of the patients YAM, ITO, AMA, and MAT were described in detail in a previous paper (13), and those of the patients SHI and KAM in another paper (14). Patient HAG was confirmed to be a typical hydatidiform mole, and MIT and KUN were gestational choriocarcinomas. In this paper, these samples will be called mole hCG, invasive mole hCG, chorio- carcinoma hCG, and normal hCG, respectively. Extraction and puri- fication of these hCGs were performed as described previously (4).
Release of Mucin-type Sugar Chains by Alkaline Borohydride Treat- ment-Mucin-type sugar chains were released from hCG samples by the method of Carlson (15). hCG sample (1-2 mg) was dissolved in 200 p1 of 0.05 N NaOH containing 1 M NaBHl and kept at 48 "C for 16 h. After adjusting the pH to 5.0 with 4 N acetic acid, the mixture was passed through a column (3 ml) of AG 50-X8 (H' form) resin. The column was washed with water to recover the oligosaccharides completely. The effluent was evaporated with methanol to remove borate. In order to remove peptides, the residue was passed through a column of Bond Elut C18 (Analytichem International, Inc., Harbor City, CA). The column was washed with water and the effluent was lyophilized.
Labeling of Oligosaccharides-In order to determine the optimal condition to remove the N-acetyl groups from N-acetylamino sugar residues completely, N-acetyl[l-3H]galacto~aminitol (2 nmol, 50,000 cpm) was dissolved in 100 pl of anhydrous hydrazine and heated at 106 "C for 8,16, and 24 h. After removal of hydrazine by coevaporation with toluene, the residues were dissolved in 50 pl of water, spotted on a sheet of Whatman No. 1 paper, and subjected to high voltage paper electrophoresis. Radioactive N-acetylgalactosaminitol (-3 cm) and galactosaminitol (-19 cm) were located by radiochromatogram scan- ner, and their radioactivities were determined by liquid scintillation method. This experiment revealed that 75, 85, and 100% of N- acetylgalactosaminitol were converted to galactosaminitol by hydra- zinolysis for 8, 16, and 24 h, respectively. Accordingly, we decided to adopt 24-h hydrazinolysis for de-N-acetylation of oligosaccharides.
The oligosaccharide fraction obtained by alkaline borohydride treatment was dried thoroughly overnight in vacuo on P205 and suspended in 0.2 ml of anhydrous hydrazine. The mixture was incu- bated at 106 "C for 24 h. Hydrazine was removed from the reaction mixture by repeated evaporation with toluene. The residue was then subjected to paper chromatography with use of solvent I for 18 h. The portion of paper (0-10 cm from origin) which included de-N-acety- lated sugars larger than galactosaminitol was cut out, and the de-N- acetylated sugars were recovered by extraction with 1 N NH,OH. The fraction was neutralized with HC1 and evaporated to dryness. The residue was then N-acetylated with [3H]acetic anhydride (5 pmol, 250 pCi) in 100 pl of 0.1 M Na,CO,/NaHCO, buffer, pH 10.7, at room temperature for 1 h. Excess amount of acetic anhydride (30 pl) and 500 p1 of the buffer were added, and the mixture was kept for another hour in order to complete N-acetylation. The mixture was applied to a column (1 ml) of AG 50(H+), and the column was washed with water. The effluent was evaporated and subjected to paper chroma- tography by using solvent I for 16 h. The radioactive peaks, which moved slower than N-acetylgalactosaminitol, were extracted from the paper with water.
Alkaline Degradation Utilizing Tritiated Borohydride-The reac- tion mixture (200 pl ) containing 1-2 mg of hCG sample and 2.5 mCi of NaB3H4 in 1 M NaBH4 and 0.05 N NaOH was kept a t 48 "C for 16 h. In order to stop the reaction, 4 N acetic acid was added. The mixture was passed through a column of AG 50(H+) and the column was washed with water. The effluent was evaporated with methyl alcohol and subjected to paper chromatography using solvent I for 16
h. The radioactive peaks were extracted from the paper. Exoglycosidase Digestion-Sugar samples (2-10 nmol) were incu-
bated with one of the following mixtures (60 pl) at 37 "C for 18 h 1) A. ureajaciem sialidase digestion, enzyme (0.1 unit) in 0.2 M sodium acetate buffer, pH 5.0; 2) jack bean @-galactosidase digestion, enzyme (0.75 unit) in 0.1 M sodium citrate buffer, pH 4.0; 3) jack bean P-N- acetylhexosaminidase digestion, enzyme (0.6 unit) in 0.1 M sodium citrate buffer, pH 4.0; 4) diplococcal @-galactosidase digestion, enzyme (2 milliunits) in 0.1 M sodium citrate buffer, pH 6.0. One drop of toluene was added to each reaction mixture to inhibit bacterial growth. The reaction was terminated by heating the reaction mixture in a boiling water bath for 3 min.
New Castle Disease Virus (NDV) Sialidase Digestion-Acidic oli- gosaccharides were incubated with NDV sialidase (0.8 unit) in 50 pl of 0.05 M cacodylate buffer, pH 6.5, at 37 "C for 0-2 h. The reaction mixture was passed through a filter of mole cut (Nihon Millipore Kogyo Kabushiki Kaisha, Tokyo), and the oligosaccharides were recovered in the filtrate. The filtrate was evaporated to dryness, dissolved in 40 pl of water, and subjected to anion exchange column chromatography by using the fast protein liquid chromatography system equipped with a Mono-Q column (Pharmacia LKB Biotech- nology Inc.). After being applied to the column, the oligosaccharides were eluted by sodium acetate buffer, pH 4.0; isocratically 5 mM from 0 to 10 min and then on a linear gradient (5-300 mM) from 10 to 40 min, a t a flow rate of 1 ml/min.
Paulson et al. (16) reported that NDV sialidase cleaves the SiaaZ+ 3Gal linkage but not the Siaa24Gal/GalNAc linkage of glycopro- teins, the sialic acid moieties of which were added by well character- ized pure sialyltransferases. In order to clarify the extent and the limit of this specificity, we investigated the susceptibility of various mucin-type oligosaccharides to NDV sialidase digestion. As shown in Fig. 1, NeuAca2+3Gal@l+3GalNAcoT was readily converted to Gal@l+3GalNAcoT, while both sialic acid residues of NeuAcaZ+ 6GalNAcoT and NeuNGca24GalNAcoT were not hydrolyzed at all. Interesting and unexpected results were obtained by the study of the NeuAca24(Gal@l-+3)GalNAcoT and GalNAcal+3(Fucal+ 2)Gal@1+3(Siaa24)GalNAcoT (NeuAc/NeuNGc ratio = 15). Al- though these oligosaccharides contain only the Siaa24GalNAcoT linkages, the sialic acid residue was slowly but no doubt cleaved by NDV sialidase. Therefore, the Siaa2+6GalNAcoT linkage becomes susceptible to the sialidase digestion when the C-3 position of the GalNAcoT residue is substituted by sugars. In support of this rule, 1 sialic acid residue of the NeuAca2+3Gal@l-+3(NeuAca2+ 6)GalNAcoT was cleaved totally within 2 h, and the other residue was cleaved by much slower rate comparable to that of Siaa24(Gal@l+ 3)GalNAco~ (data not shown).
RESULTS
Paper Electrophoresis of Oligosaccharides Liberated from a Normal and a Choriocarcinoma hCG by Alkuline-NaB3H4 Treatment-When the radioactive oligosaccharide mixture obtained from a normal hCG sample by alkaline-NaB3H4 treatment was subjected to high voltage paper electrophoresis,
0 60 120 Time f min 1
FIG. 1. Action of NDV sialidase on various mucin-type oli- gosaccharides. Enzymatic reactions and analysis of products were performed as described under "Experimental Procedures." 0, NeuAca2+3Gal@l+3GalNAco~; 0, NeuAca2+6(Gal@l+ 3)GalNAcoT; A, a mixture of GalNAcal+3(Fuca1+2)Gal@l~ 3(NeuAca2~6)GalNAcoT and GalNAcal+3(Fucal+2)Gal~l+ 3(NeuNGca2*6)GalNAcoT in a molar ratio of 15; 0, a mixture of NeuAca24GalNAcoT and NeuNGca24GalNAco~ in a molar ratio of 1:l.

Mucin-type Sugar Chains of Various hCGs 1159
it was separated into five fractions, N, A l , A2, A3, and A4 (Fig. 2 A ) . All acidic fractions (Al-A4) were converted com- pletely to neutral components by A. ureafaciens sialidase digestion. These neutral fractions were named as AlN, A2N, A3N, and A4N, respectively. The results of limited sialidase digestion revealed that fractions A1 and A2 contained mon- osialyl oligosaccharides, while fractions A3 and A4 contained disialyl oligosaccharides (data not shown).
Alkaline-NaB3H4 treatment of a choriocarcinoma hCG also gave the five 3H-labeled fractions separated by paper electro- phoresis (Fig. 2B). As is apparent from the two electrophore- tograms in Fig. 2, A and B, the molar ratio of each oligosac- charide from choriocarcinoma hCG is quite different from those of normal hCG. In order to elucidate the difference in more detail, structural study of oligosaccharides in the five fractions was indispensable. However, low yield of radioactiv- ity incorporated into the oligosaccharide fraction made it hard to investigate the structure of each oligosaccharide thor- oughly. This is because extensive dilution of NaB3H4 was necessary to obtain high concentration of NaBH4 in order to protect the released oligosaccharides from peeling reaction. To break the bottleneck, we tried to adopt the N-3H-acetyla- tion method developed by Higashi and Basu (17) for the labeling of the sugar moieties of gangliosides.
Labeling of Oligosaccharides by N-3H-Acetylation Method-Oligosaccharide fraction obtained from the normal hCG was de-N-acetylated by hydrazinolysis and re-N-acety- lated with [3H]acetic anhydride as described under “Experi- mental Procedures.” The radioactivities incorporated into the oligosaccharide fraction by this method and by alkaline- NaB3H4 method are summarized in Table I. The data indi- cated that approximately six times more radioactivity was incorporated by using one-tenth amount of the labeling re- agent. As the result, the specific activity of the radioactive oligosaccharide fraction obtained by the N-3H-acetylation method is approximately eight times higher than that by the alkaline-NaB3H, method. When the radioactive oligosaccha- ride fraction was subjected to paper electrophoresis, it gave
N _.
A1 A2 h3 A 4
I II J 0 10 2 0 30
Dlstance f rom Orlgm (cm)
FIG. 2. Paper electrophoresis of the radioactive oligosac- charides obtained from hCG samples. Arrows indicate the mobil- ities of authentic oligosaccharides: Z, Siaa2-3Gal/31-3GalNAcoT; ZZ, Siaa2+3Ga1j31-3(Siaa2+6)GalNAcoT. Radioactive oligosaccha- rides obtained from normal hCG ( A and C ) and choriocarcinoma hCG ( B and D) by alkaline-NaB3H4 method ( A and B ) and N-3H-acetyla- tion method (C and D). Chromatograms A and B were obtained by scanning the papers at 10 times higher sensitivity than chromato- grams C and D.
TABLE I Amounts of radioactive reagents used i n the two labeling methods and
yields of radioactive mucin-type oligosaccharides N-3H-Acetylation Alkaline-NaB3H,
method method
Dry weight of hCG (mg) 1.1 1.5 Radioisotope used (mCi) 0.25 2.5 Total radioactive oligosac- 1.04 X lo6 1.82 X lo5
Radioactive oligosaccha- 9.45 x lo5 1.21 X lo5 charides obtained (cpm)
rides obtained from 1 mg of hCG (cpm)
tive oligosaccharides re-
hCG (mol) leased from 1 mol of
Total amount of radioac- 3.92“ 4.07b
a The value was derived from Table IV. The value was calculated from the total radioactivity incorporated
into oligosaccharides and the specific activity determined from the radioactivity incorporated into 100 nmol of authentic N-acetylgalac- tosamine under the same condition.
TABLE I1 Comparison of the percent molar ratio of mucin-type oligosaccharides
obtained f rom mrmal hCG by the two labeling methods Molar ratio
Fractions N-[3H]Acetylation Alkaline-NaB3H, method” method
% %
N 11.8 10.4 A1 4.8 3.3 A2 40.8 40.0 A3 8.4 10.8 A4 34.2 35.5
The values were derived from Table IV.
the five radioactive peaks as shown in Fig. 2C. Total amounts of oligosaccharides obtained from 1 mol of hCG by N-3H- acetylation method and by alkaline-NaB3H4 method were 3.92 and 4.07 mol, respectively. This result indicated that mucin- type oligosaccharides of hCG were recovered quantitatively as radioactive oligosaccharides by both methods.
As will be described later, similar amounts of radioactivity were incorporated into all amino sugar residues of oligosac- charides including sialic acid residues. Structural study of the oligosaccharides described later indicated that oligosaccha- rides in fractions Al, A2, A3, and A4 contain 3, 2, 4, and 3 amino sugar residues, respectively. Therefore, molar ratio of the acidic oligosaccharides could be calculated by dividing the radioactivity in each acidic peak by the number of amino sugars. The values were listed in Table I1 together with the molar ratio of each acidic peak calculated from the radioac- tivity obtained by alkaline-NaB3H4 method. Both values ac- cord well, warranting the quantitative nature of the N-3H- acetylation method. Accordingly, the structures of oligosac- charides in fractions N, Al, A2, A3, and A4 obtained by the N-3H-acetylation method from hCG sample from the pooled urine of normal pregnant women were investigated.
Structural Studies of Oligosaccharides in the Neutral Frac- tion and in the Desialylated Acidic Fractions-Fractions N, AlN, A2N, A3N, and A4N were subjected to descending paper chromatography with use of solvent I1 for 8 h. As shown in Fig. 3, two different radioactive components were detected. The faster moving component a was obtained from fractions N, A2N, and A4N, and the component b from fractions N, AlN, and A3N. Fractions N and A1N contained a small amount of radioactive components which remained at the origin (Fig. 3, A and B) . Structural studies of these compo-

1160 Mucin-type Sugar Chains of Various hCGs
b a "
0 10 20
Distance from Origin (cm)
FIG. 3. Paper chromatography of the neutral oligosaccha- ride fractions with use of solvent I1 for 8 h. Arrows indicate the mobilities of standard oligosaccharides: Z, Galj31+3GalNAcoT; ZZ, GalNAcoT.
nents revealed that they were asparagine-linked oligosaccha- rides (data not shown). The broad radioactive peak, which was detected between fractions N and A1 in Fig. 2C, was also found to contain asparagine-linked oligosaccharides. The structures of these asparagine-linked oligosaccharides have been reported previously (18).
The structure of component a was elucidated to be Gal@l+ 3GalNAcoH from the following data. The mobility of compo- nent a in Fig. 3 and in a Bio-Gel P-4 column (Fig. 4 0 ) was the same as authentic Gal@1+3GalNAcoT. Methylation analysis of component a gave 2,3,4,6-tetra-O-methyl galactitol and 1,4,5,6-tetra-O-methyl 2-N-methylacetamido-2-deoxyga- lactitol in approximately 1:l molar ratio (Table 111).
When component b was applied to a column of Bio-Gel P- 4, a single radioactive peak with a mobility of 6.3 glucose units was obtained (Fig. 4A). The oligosaccharide was con- verted to a radioactive component with a mobility of 5.3 glucose units by incubation with diplococcal @-galactosidase, indicating that 1 galactose residue was removed (Fig. 4B). Although jack bean @-N-acetylhexosaminidase could not re- lease any N-acetylhexosamine residue from the radioactive component b, the enzyme released 1 mol of 3H-labeled N- acetylglucosamine from the degalactosylated radioactive com- ponent in Fig. 4B (Fig. 4C). The results indicated that com- ponent b contains 1 Gal/31--*4GlcNAc/31-+ residue as a non- reducing terminal, because diplococcal @-galactosidase is known to cleave the Galpl4GlcNAc linkage but not the Galfi1+3GlcNAc and the Galpl4GlcNAc linkages (19). The radioactive component which eluted at 3.5 glucose units in Fig. 4C was confirmed to be Gal@l+3GalNAcoH by the same analysis as already described for component a (data not shown). Therefore, component b should have the structure of either Gal~14GlcNAc/31+Gal@1+3GalNAcoH or Gal@l+ 4GlcNAc/31+(Gal@l+3)GalNAcoH. Methylation analysis of component b gave 2,3,4,6-tetra-O-methyl galactitol as the only 0-methylated galactose derivative (Table 111), indicating that the Gal@l4GlcNAc/31+ group is not linked to the galactose residue of the Gal@l+3GalNAcoT. Detection of 1,4,5-tri-O- methyl 2-N-methylacetamido-2-deoxygalactitol (Table 111) indicated that the structure of component b is Gal@l+ 4GlcNAcfi1+6(Gal/31+3)GalNAcoH.
TABLE I11 Methylation analysis of the total oligosaccharide fraction released
from a normal hCG before and after sialidase digestion and components a and b
Molar ratio
Total oligosaccharide Components Methylated sugars fraction"
-Sialidase +Sialidase ab b'
Galactitol 2,3,4,6-Tetra-O- 6.0 11.0 1.0 2.0
methyl (1,5-di-0- acetyl)
(1,2,5-tri-O-acetyl)
(1,3,5-tri-O-acetyl)
(1,5,6-tri-O-acetyl) 2-N-Methylacetamino-
2-deoxyglucitol
(1,4,5-tri-O-acetyl)
(1,3,5-tri-O-acetyl) 2-N-Methylacetamido-
2-deoxygalactitol
methyl (3-mono-0- acetyl)
(3,6-di-O-acetyl)
3,4,6-Tri-O-methyl 0 0 0 0
2,4,6-Tri-O-methyl 5.0 0 0 0
2,3,4-Tri-O-methyl 0 0 0 0
3,6-Di-O-methyl 1.0 1.0 0 0.8
4,6-Di-O-methyl 0 0 0 0
1,4,5,6-Tetra-O- 4.5 7.3 0.7 0
1,4,5-Tri-O-methyl 3.5 0.8 0 0.7
a Numbers were calculated by taking the italicized values as 1.0. * Numbers were calculated by taking the italicized values as 1.0. e Numbers were calculated by taking the italicized values as 2.0.
t
7 6 5 4 3 2 V l l l v v
A I I I ? ?
i\
D A
210 240 270
Elution Volume ( m l )
FIG. 4. Exoglycosidase digestion of component8 a and b in Fig. 3. The oligosaccharides and radioactive digestion products were analyzed by Bio-Gel P-4 column chromatography. Black arrow-heads indicate the elution positions of glucose oligomers (numbers indicate the glucose units), and white arrows indicate those of standard oli- gosaccharides: I , Gal@l+3GalNAco~; ZZ, N-acetylglucosamine. A, component b in Fig. 3; B, the radioactive peak in A incubated with diplococcal &galactosidase; C, the radioactive peak in B incubated with jack bean P-N-acetylhexosaminidase; D, component a in Fig. 3.
Sialic Acid Linkages Included in the Acidic Oligosacchu- rides-The results so far described indicated that the acidic mucin-type sugar chains of hCG are mono- and disialyl de- rivatives of Gal@l+3GalNAc and Galpl+4GlcNAc@l--t 6(Gal@1+3)GalNAc. Since all the sialic acids released from normal hCGs as well as choriocarcinoma and mole hCGs by sialidase treatment were N-acetylneuraminic acid (13), the acidic mucin-type sugar chains of these hCGs should contain only N-acetylneuraminic acid. In order to find out what kinds of sialyl linkages are included in these oligosaccharides, the
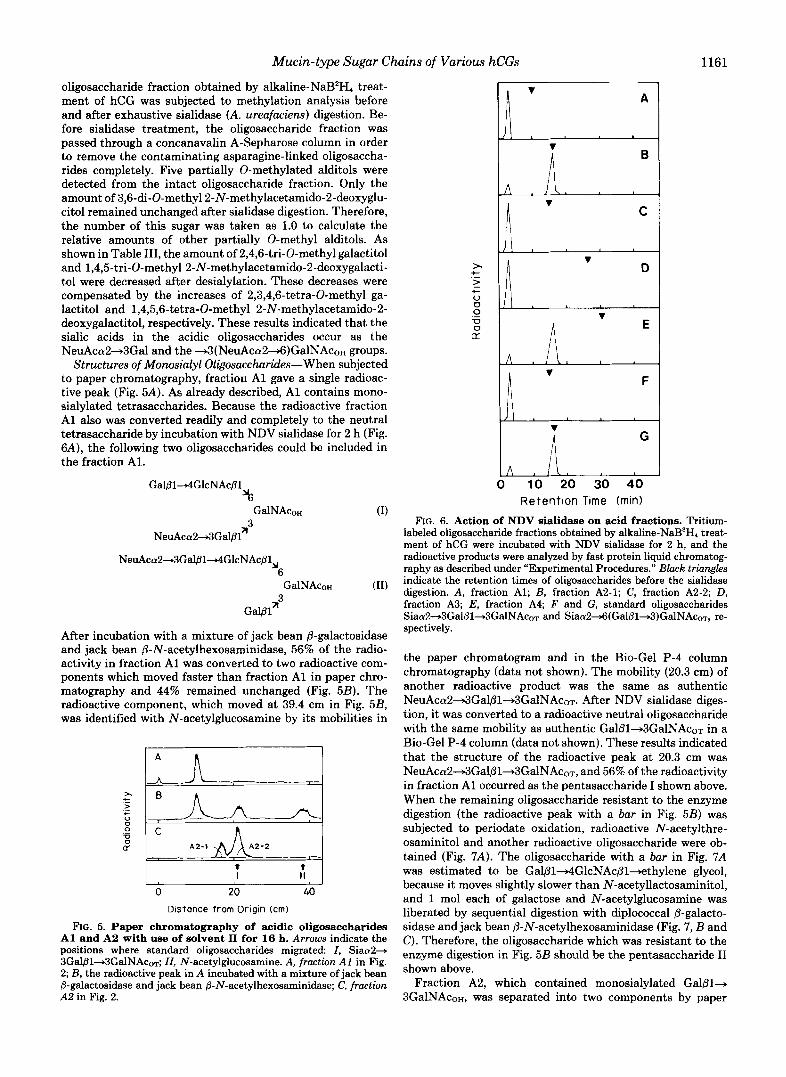
Mucin-type Sugar Chains of Various hCGs 1161
oligosaccharide fraction obtained by alkaline-NaB2H, treat- ment of hCG was subjected to methylation analysis before and after exhaustive sialidase (A. ureafacierzs) digestion. Be- fore sialidase treatment, the oligosaccharide fraction was passed through a concanavalin A-Sepharose column in order to remove the contaminating asparagine-linked oligosaccha- rides completely. Five partially 0-methylated alditols were detected from the intact oligosaccharide fraction. Only the amount of 3,6-di-O-methyl2-N-methylacetamido-2-deoxyglu- citol remained unchanged after sialidase digestion. Therefore, the number of this sugar was taken as 1.0 to calculate the relative amounts of other partially 0-methyl alditols. As shown in Table 111, the amount of 2,4,6-tri-O-methyl galactitol and 1,4,5-tri-O-methyl 2-N-methylacetamido-2-deoxygalacti- to1 were decreased after desialylation. These decreases were compensated by the increases of 2,3,4,6-tetra-O-methyl ga- lactitol and 1,4,5,6-tetra-O-methyl 2-N-methylacetamido-2- deoxygalactitol, respectively. These results indicated that the sialic acids in the acidic oligosaccharides occur as the NeuAca2+3Gal and the +3(NeuAca2+6)GalNAc0~ groups.
Structures of Monosialyl Oligosaccharides-When subjected to paper chromatography, fraction A1 gave a single radioac- tive peak (Fig. 5A). As already described, A1 contains mono- sialylated tetrasaccharides. Because the radioactive fraction A1 also was converted readily and completely to the neutral tetrasaccharide by incubation with NDV sialidase for 2 h (Fig. 6A), the following two oligosaccharides could be included in the fraction Al.
Gal@l4GlcNAc@l %
GalNAcoH (1) n
NeuAccu2+3Gal@l
NeuAca2+3Gal@14GlcNAc@l,,
a'
6
3 GalNAcoH (11)
Gal@ln
After incubation with a mixture of jack bean @-galactosidase and jack bean 0-N-acetylhexosaminidase, 56% of the radio- activity in fraction A1 was converted to two radioactive com- ponents which moved faster than fraction A1 in paper chro- matography and 44% remained unchanged (Fig. 5B). The radioactive component, which moved at 39.4 cm in Fig. 5B, was identified with N-acetylglucosamine by its mobilities in
L
0 20 40 Distonce f rom Origin (cm)
FIG. 5. Paper chromatography of acidic oligosaccharides A1 and A2 with use of solvent I1 for 16 h. Arrows indicate the positions where standard oligosaccharides migrated I, Siaa2+ 3Gal@1+3GalNAcoT; ZZ, N-acetylglucosamine. A, fraction A1 in Fig. 2; B, the radioactive peak in A incubated with a mixture of jack bean @-galactosidase and jack bean @-N-acetylhexosaminidase; C, fraction A2 in Fig. 2.
c x > t V 0 0 V
LT 0
"
.-
A
C
n v D
L
il v F
i G
A , I \ , 10 20 30 40
Retention Time (rnin) FIG. 6. Action of NDV sialidase on acid fractions. Tritium-
labeled oligosaccharide fractions obtained by alkaline-NaB3H, treat- ment of hCG were incubated with NDV sialidase for 2 h, and the radioactive products were analyzed by fast protein liquid chromatog- raphy as described under "Experimental Procedures." Black triangles indicate the retention times of oligosaccharides before the sialidase digestion. A, fraction Al; B, fraction A2-1; C , fraction A2-2; D, fraction A3; E, fraction A4; F and G, standard oligosaccharides Siaa2+3Gal@l+3GalNAcoT and Siaa2~(Ga1@1+3)GalNAcoT, re- spectively.
the paper chromatogram and in the Bio-Gel P-4 column chromatography (data not shown). The mobility (20.3 cm) of another radioactive product was the same as authentic NeuAca2+3Galfil+3GalNAc0~. After NDV sialidase diges- tion, it was converted to a radioactive neutral oligosaccharide with the same mobility as authentic Gal/31+3GalNAcoT in a Bio-Gel P-4 column (data not shown). These results indicated that the structure of the radioactive peak at 20.3 cm was NeuAca2+3Gal@l~3GalNAcoT, and 56% of the radioactivity in fraction A1 occurred as the pentasaccharide I shown above. When the remaining oligosaccharide resistant to the enzyme digestion (the radioactive peak with a bar in Fig. 5B) was subjected to periodate oxidation, radioactive N-acetylthre- osaminitol and another radioactive oligosaccharide were ob- tained (Fig. 7A) . The oligosaccharide with a bar in Fig. 7A was estimated to be Gal@l4GlcNAc~l+ethylene glycol, because it moves slightly slower than N-acetyllactosaminitol, and 1 mol each of galactose and N-acetylglucosamine was liberated by sequential digestion with diplococcal P-galacto- sidase and jack bean 8-N-acetylhexosaminidase (Fig. 7 , B and C). Therefore, the oligosaccharide which was resistant to the enzyme digestion in Fig. 5B should be the pentasaccharide I1 shown above.
Fraction A2, which contained monosialylated Gal@1+ 3GalNAcoH, was separated into two components by paper

1162 Mucin-type Sugar Chains of Various hCGs
I 4 4 4 4 1 I I , II Ill iv I 0 10 20 30
Distance f rom Origln (cm)
FIG. 7. Periodate oxidation of acidic fractions. The radioac- tive products were analyzed by paper chromatography using solvent I for 20 h. Arrows at the bottom indicate the positions where standard oligosaccharides migrated: Z, Galpl+4GlcNAcoT; ZI, Gal81+ 3ThrNAcoT; IIZ, N-acetylglucosamine; ZV, ThrNAcm. A, the pro- ducts from the radioactive peak with a bur in Fig. 5B, B, the radio- active peak with a bar in A incubated with jack bean 8-galactosidase; C, the radioactive peak in B incubated with jack bean 8-N-acetylhex- osaminidase; D, the product from fraction A2-1 in Fig. 5C; E, the product from fraction A2-2 in Fig. 5C; F, the products from fraction A3 in Fig. 2 A ; G, the product from fraction A4 in Fig. 2 A .
chromatography (Fig. 5C). These two components were named A2-1 and A2-2, respectively. The mobility of A2-2 was the same as authentic NeuAca2+3Gal~l+3GalNAcoT. When digested with NDV sialidase for 2 h, it was converted com- pletely to a radioactive neutral oligosaccharide (Fig. 6C) in- dicating that all sialic acid residues occur as the NeuAca2+ 3Gal group. By periodate oxidation, Ga181+3ThrNAcoH was obtained as the only radioactive product from A2-2 (Fig. 7E). These results indicated that A2-2 is NeuAca2-+3Gal@l+ 3GalNAcoH.
When incubated with NDV sialidase, component A2-1 was converted to radioactive neutral oligosaccharide at an ex- tremely slow rate (Fig. 6B). The rate of hydrolysis was the same as that of authentic NeuAca24(Gal~l+3)GalNAcoT (Fig. 6G). By periodate oxidation, N-acetylthreosaminitol was obtained as the only radioactive product from A2-1 (Fig. 70). These results indicated that A2-1 is NeuAca2+6(Gal@l+ 3)GalNAcoH.
Structures of Disialyl Oligosaccharides-Experimental re- sults in the previous section indicated that fraction A3 con- tains disialylated tetrasaccharide. Since A3 was completely resistant to digestion with jack bean 8-galactosidase (data not shown), it was assumed that the 2 sialic acid residues are linked to the 2 galactose residues of the tetrasaccharide. Because all fraction A3 was converted readily to neutral oligosaccharide by NDV sialidase treatment (Fig. 6D), the 2 sialic acid residues should occur as the NeuAca2+3Gal group. By periodate oxidation, two radioactive components corre- sponding to Gal8l+3ThrNAcoH and Gal@l+4GlcNAc81+ ethylene glycol were obtained (Fig. 7F). These results indi-
cated that fraction A3 contains only the hexasaccharide shown in Table IV.
Fraction A4 contains disialylated Gal81+3GalNAco~. By NDV sialidase digestion for 2 h, an acidic radioactive product with the same mobility as authentic Gal/31+3(NeuAca2+ 6)GalNAcoT on high pressure liquid chromatography was obtained from fraction A4 (Fig. 6E). A small amount of neutral oligosaccharide is to be derived from the acidic trisac- charide by the action of NDV sialidase. When fraction A4 was subjected to periodate oxidation, Gal/31+3ThrNAcoH was obtained as the only radioactive product (Fig. 7G). These results indicated that fraction A4 contains only NeuAca2- 3Gal~l+3(NeuAca24)GalNAcoT.
Structural Study of the Radioactive Oligosaccharides Ob- tained from Choriocarcinoma hCG by N-3H-Acetylatwn Method-Oligosaccharide fraction obtained from the chorio- carcinoma hCG was labeled by N-3H-acetylation method. Paper electrophoresis of the radioactive oligosaccharides gave the fractionation pattern as shown in Fig. 20 . Detailed struc- tural studies of the oligosaccharides in fractions N, Al, A2, A3, and A4 revealed that no qualitative difference was found among the oligosaccharides included in the five fractions obtained from the normal and the choriocarcinoma hCGs (data not shown). In order to determine the molar ratio of each oligosaccharide in the two hCG samples, distribution of radioactivity in each amino sugar residue of all oligosaccha- rides was calculated as follows, and the values are included in Table IV. In the case of sialylated disaccharides, the radioac- tivities in sialic acid residues and in N-acetylgalactosaminitol were determined by measuring the radioactivities of sialic acid residues and of the neutral disaccharide, separated by paper electrophoresis after sialidase digestion. The radioactivities of sialic acid residues were divided by the number of sialic acid residues included in each acidic oligosaccharide. In the case of sialylated tetrasaccharides, the radioactivities in the sialic acid residues and the neutral tetrasaccharide moiety were determined in the same manner. The radioactivities in the N-acetylglucosamine residue and in the N-acetylgalac- tosaminitol were determined after 8-galactosidase and 8-N- acetylhexosaminidase digestion as described in Fig. 4C. Dis- tribution of radioactivities in each amino sugar of oligosac- charides obtained from the two hCG samples was almost the same as shown in Table IV. The data also indicated that the radioactivities incorporated in N-acetylgalactosaminitol, N- acetylglucosamine, and N-acetylneuraminic acid were 1.0:1.00.8. Based on these data, the molar ratio of each oligosaccharide from the two hCG samples was calculated from the radioactivities of the seven oligosaccharides as sum- marized in Table IV. The numbers of mucin-type sugar chains in one molecule of hCG were approximately 4 in both hCG samples. The data indicated that choriocarcinoma hCG is more enriched in the oligosaccharides with the GalPl+ 4GlcNAc~1+6(Gal~l-t3)GalNAc structure than the normal hCG.
Comparative Study of the Mucin-type Sugar Chains of hCG Samples Purified from the Urine of Pregnant Women and Patients with Trophoblastic Diseases-Having been stimu- lated with these interesting results, we then studied the ratio of the disaccharide and the tetrasaccharide cores included in the hCGs purified from the urine of various trophoblastic diseases as well as normal pregnant individuals. Radioactive oligosaccharide fractions, obtained by @elimination and N- 3H-acetylation method of hCGs from four patients with hy- datidiform mole, two with invasive mole, two others with choriocarcinoma, and two normal pregnant women were treated with A. ureafaciens sialidase. The radioactive neutral

Mucin-type Sugar Chains of Various hCGs 1163
TABLE IV Structures and amounts of mucin-type sugar ch ins of normal and choriocarcinoma hCG
Fractions Structures Normal Choriocarcinoma
N
A1
A2-1
A2-2
A3
A4
0.97" (0.98) GalB14GlcNAcpl
'6 GalNAc
Gal01
Galpl-3GalNAc
A3
1.0 (0.99) Gal/314GlcNAcpl
0.80 (0.80) '6 NeuAca2+3 I GalNAc
I Gal01 A3
0.79 (0.78) NeuAccY2
'6 GalNAc
Gala1 n3 0.74(0.75)
NeuAcc~2+3Gal~l+3GalNAc
0.78(0.78) 1.0 (1.0) NeuAcc~2+3Gal~l+4GlcNAc~l
'6
NeuAcc~2+3Gal01 213
GalNAc
0.78 (0.78)
0.81 (0.80) NeuAcaZ
6
rnol/mol of hCG
0.10' (2.6)' 0.37 (10.8)
0.36 (9.2) 0.13 (3.8)
0.19 (4.8)
0.37 (9.4)
1.23 (31.4)
0.33 (8.4)
0.69 (20.2)
0.20 (5.8)
0.66 (19.3)
1.07 (31.3)
GalNAc 1.34 (34.2) 0.30 (8.8) 3
NeuAca2+3Galpl3 0.81 (0.80)
Total 3.92 (100) 3.42 (100) The number indicates the relative radioactivity incorporated into each amino sugar of oligosaccharides from
normal hCG calculated by taking the value of N-acetylgalactosamine as 1.0. The number in the parentheses indicates the relative radioactivity from choriocarcinoma hCG.
*The value was calculated from the radioactivity incorporated into N-acetylgalactosamine residue of each oligosaccharide and the specific activity of N-acetyl group determined from the radioactivity incorporated into 100 nmol of authentic galactosaminitol.
e The number in the parentheses indicates the percent molar ratio of each oligosaccharide.
oligosaccharide fractions thus obtained were analyzed by pa- per chromatography with use of solvent 11, and the molar ratio of the two core oligosaccharides was calculated from their radioactivities and summarized in Fig. 8. The amounts of tetrasaccharide were obtained by dividing their radioactiv- ities by 2, because they contain two amino sugars.
Hydatidiform mole hCGs gave very similar molar ratios of the two neutral cores to that of normal hCGs. In contrast, the hCG from other choriocarcinoma patients gave much higher tetrasaccharide content as in the case of a choriocarcinoma hCG studied already. Other important and interesting evi- dence is that both of the two invasive mole hCGs gave intermediary values in the ratio of the tetrasaccharide core.
DISCUSSION
Since having been established by Carlson (15), alkaline borohydride degradation of glycoproteins has been used to
release the mucin-type sugar chains as oligosaccharides for structural study. Because the oligosaccharides obtained by this method are oligosaccharide alcohols, they cannot be labeled by NaB3H, reduction. Later on, alkaline-NaB3H4 treatment was devised by Aminoff et al. (20) in order to study the mucin-type sugar chains of glycoproteins obtained in small amounts. Since the method was simple and reliable as used in part of this study, it has been applied to studies of mucin-type sugar chains of glycoproteins obtained in small quantities. A drawback of this method is that an extremely high concentration of NaB3H4 is required in order to protect the released sugar chains from peeling reaction. Production of large amounts of 3H2 gas has also been hampering the frequent use of the method for the study of mucin-type sugar chains.
N-14C-Acetylation was used successfully to label glycolipids (17) and oligosaccharides released from al-fetoprotein by hydrazinolysis (21). Since all mucin-type sugar chains contain

Mucin-type Sugar Chains of Various hCGs
jl NA . . Yb
normal
:3 IA HAG .... ,
hydotidiform mole invosive mole choriocarcinoma
FIG. 8. The Occurrence of disaccharide and tetrasaccharide cores in the mucin-type sugar chains of hCGs purified from the urine of patients with various trophoblastic diseases. The percent molar ratio of the two oligosaccharides was calculated as described in the text.
at least 1 N-acetylhexosamine residue and the total amount of amino sugars in an oligosaccharide sample can be estimated by analysis of its sugar composition (22), the re-N-acetylation method can be used for the radioactive labeling of the mucin- type sugar chains without producing too much radioactive waste. The condition of hydrazinolysis for the de-N-acetyla- tion of oligosaccharides is much stronger than that used for the release of asparagine-linked sugar chains as oligosaccha- rides (23). As shown in Table IV, re-N-acetylation under the conditions described in this paper will label quantitatively all amino sugar residues of an oligosaccharide.
The radioactive oligosaccharides obtained by this procedure can be used as samples for study by the microprocedures developed for the structural analyses of asparagine-linked sugar chains (24). That the labeling occurs at several mono- saccharide residues of an oligosaccharide is somewhat incon- venient for the sequencing study by exoglycosidase digestion, because the radioactivity is not limited at the reducing ter- minal. On the other hand, this situation is beneficial because an oligosaccharide is more highly labeled than NaB3H4 reduc- tion. Furthermore, the N-acetylamino sugars released by 8- N-acetylhexosaminidase digestion can be identified to afford more precise structural information by sequential exoglyco- sidase digestion.
The results described in this paper confirmed the report of Cole et al. (25) that hCG contains not only sialylated Gal@1+ 3GalNAc but also small amounts of sialylated GalPl-, 4GlcNAc@?1+6(Gal@l+3)GalNAc. They reported that the oligosaccharides with the neutral tetrasaccharide core were 13% of total mucin-type sugar chains in tryptic @-CTP (amino acid residues 123-145 of the @-subunit) of the hCG purified from the urine of normal pregnant women. This value is approximately the same as the value (12.9%) obtained in our study. Oligosaccharides with the tetrasaccharide core occur rather rarely, while oligosaccharides with the neutral disac- charide core have been found widely in many glycoproteins.
An important finding of our study is that the proportion of oligosaccharides with the tetrasaccharide core prominently rose in choriocarcinoma hCG. Actually, in the three chorio- carcinoma hCG samples investigated, the oligosaccharides with the tetrasaccharide core were major components, occu- pying approximately 60% of total mucin-type sugar chains. Another interesting finding is that the proportion of the oligosaccharides with the tetrasaccharide core structure in- creased significantly in invasive mole hCGs, although hyda-
GalNAc-Ser/ThI
t Gal@lf3GalNAc-Ser/Thr - jGalNAc-Ser/Thr
IV S l a a h 6
GalB1'
GalB1-4GlcNAcBh6
~ ~ 1 ~ 1 " GalNAc-SerIThr
FIG. 9. The biosynthetic pathway of mucin-type sugar chains.
tidiform mole hCGs contain a similar proportion of the tetra- saccharide to normal hCG.
It has been reported already that the tetrasaccharide core is synthesized by the pathway shown in Fig. 9 (26). Therefore, the structural change should be the result of either an en- hanced production of the N-acetylglucosaminyltransferase which catalyzes step I1 in Fig. 9 or suppression of a sialyltrans- ferase that competes with the N-acetylglucosaminyltransfer- ase for a common acceptor Gal@1+3GalNAcal+Ser(Thr) in choriocarcinoma cells. The latter is less likely because the total amount of sialylated mucin-type oligosaccharides in normal and choriocarcinoma hCGs are almost the same. Fur- thermore, it is noteworthy that the phenomenon emerges even at the stage of invasive mole, which is considered to be a precancerous state.
The structural changes of the asparagine-linked sugar chains of choriocarcinoma hCG have been elucidated in our laboratory (5). A part but not all of the changes found in choriocarcinoma hCG was also detected in invasive mole hCGs, while hCGs purified from the urine of patients with hydatidiform mole contain exactly the same asparagine- linked sugar chains as normal hCGs (13, 14). Therefore, structural change of hCG produced in choriocarcinoma cells is induced in both asparagine-linked and mucin-type sugar chains. These changes would serve as useful markers for the diagnosis of various trophoblastic diseases.
The 8-CTP region of hCG, which contains four mucin-type sugar chains, is unique for this hormone and not found in the @-subunit of hLH closely related with hCG. Whether this region serves for the receptor binding and for the expression of biological activity of hCG or not is not known. It was reported, however, that the region does not seem to be re- quired for the binding of the @- to the a-subunit of hCG (27). The region may partially modify the activity of hCG, since some differences in binding activity and biological potency are found between hCG and luteinizing hormone (28, 29). It is expected that choriocarcinoma hCG, in which the structures of mucin-type sugar chains are modified extensively, may serve as useful material to study the functional role of the @- CTP region.
Kawakita (Department of Pure and Applied Science, College of Acknowledgments-We wish to express our gratitude to Drs. M.
General Education, The University of Tokyo) and H. Shibuta (De- partment of Viral Infection, The Institute of Medical Science, The University of Tokyo) for their kind gift of New Castle disease virus. Thanks are also due to Yumiko Kimizuka for her skilled secretarial assistance.
REFERENCES 1. Bellisario, R., Carlsen, R. B., and Bahl, 0. P. (1973) J. BWl.
Chem. 248,6796-6809

Mucin-type Sugar Chains of Various hCGs 1165
2. Carlsen, R. B., Bahl, 0. P., and Swaminathan, N. (1973) J. Biol. 16. Paulson, J. C., Weinstein, J., Dorland, L., van Halbeek, H., and Chem. 248,6810-6825 Vliegenthart, J. F. G. (1982) J. Biol. Chem. 257 , 12734-12738
3. Kessler, M. J., Mise, T., Ghai, R. D., and Bahl, 0. P. (1979) J. 17. Higashi, H., and Basu, S. (1982) Anal. Biochem. 120, 159-164 Biol. Chem. 254,7909-7914 18. Endo, Y., Yamashita, K., Tachibana, Y., Tojo, S., and Kobata,
4. Nishimura, R., Endo, Y., Tanabe, K., Ashitaka, Y., and Tojo, S. A. (1979) J. Biochem. (Tokyo) 8 5 , 669-679 (1981) J. Endocrim!. fnuest. 4,349-358 19. Paulson, J. C., Prieels, J.-P., Glasgow, L. R., and Hill, R. L.
5. Mizuochi, T., Nishimura, R., Derappe, C., Taniguchi, T., Hama- (1978) J. Biol. Chem. 253,5617-5624 mob, T., Mochizuki, M., and Kobata, A. (1983) J. Biol. Chem. 20. Aminoff, D., Gathmann, W. D., McLean, C. M., and Yadomae, 2 5 8 , 14126-14129 T. (1980) Anal. Biochem. 101 , 44-53
6. Paulson, J. C., Sadler, J. E., and Hill, R. L. (1979) J. BWl. Chem. 21. Bayard, B., and Kerckaert, J.-p. (1981) Eur. J. Biochem. 113,
7. Li, Y.-T., and ~ i , S.-C. (1972) Methods Enzymol, 28 , 702-713 22. Takeuchi, M., Takasaki, s., h u e , N., and Kobata, A. (1987) J. 8. Glasgow, L. R., Paulson, J. C., and Hill, R. L. (1977) J. Bwl. Chromutogr. 400,207-213
Chem. 252,8615-8623 23. Takasaki, S., Mizuochi, T., and Kobata, A. (1982) Methods En- 9. Takasaki, s., Yamashita, K., and Kobata, A. (1978) J. Biol. Chem. zymol. 83,263-268
24. Kobata, A. (1984) in Biology of Carbohydrates (Ginsburg, V., and 10. Yamashita, K., Mizuochi, T., and Kobata, A. (1982) Methods Robbins, R. W., eds) Vol. 2, pp. 87-161, John Wiley and Sons,
Enzymol. 83. 105-126 New York
11. Yamashita, K., Tachibana, Y., and Kobata, A. (1977) J. Biol. 25. Cole, L. A., Birken, S., and Perini, F. (1985) Biochem. Biophys.
Chem. 262,5408-5411 Res. Commun. 126,333-339
12. Amano, J., and Kobata, A. (1986) J. Biochem. (Tokyo) 99,1645- 26. Williams, D., Longmore, G., Matta, K. L., and Schachter, H.
1654 (1980) J. Biol. Chem. 255, 11253-11261
27. Hussa, R. O., Fein, H. G., Pattillo, R. A., Nagelberg, S. B., Rosen, 13. Mizuochi, T., Nishimura, R., Taniguchi, T., Utsunomiya, T., S. W., Weintraub, B. D., Perini, F., Ruddon, R. W., and Cole,
Mochizuki, M., Derappe, C., and Kobata, A. (1985) Jpn. J. L. A. (1986) Cancer Res. 46 , 1948-1954 Cancer Res. (Gann) 76,752-759 28. Huhtaniemi, I. T., and Catt, K. J. (1981) Endocrinology 108 ,
bata, A. Cancer Res. 47,5242-5245 29. Cornbarnous, Y., Guillou, F., and Martinat, N. (1986) J. Biol.
264,2120-2124 405-414
263,6086-6091
14. Endo, T., Nishimura, R., Kawano, T., Mochizuki, M., and Ko- 1931-1938
15. Carlson, D. M. (1968) J. Biol. Chem. 243,616-626 Chem. 261,6868-6871