isolation and characterization of new cross-linking amino acid “allodesmosine” from hydrolysate...
TRANSCRIPT

Vol. 170, No. 2, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
July 31, 1990 Pages 713-718
ISOLATION AND CHARACTERIZATION OF NEW CROSS-LINKING AMINO
ACID “ALLODESMOSINE” FROM HYDROLYSATE OF ELASTIN
Kyozo Suyama and Fumihiko Nakamura
Molecular Technology of Animal Products, Faculty of
Agriculture, Tohoku University, Sendai, 981 JAPAN
Received June 16, 1990
SUMMARY: A new pentafunctional cross-linking amino acid, termed allo-
desmosine, was isolated from bovine ligamenturn nuchae elastin. This
compound was a very hygroscopic, white amorphous solid with a faint yellow
tinge, soluble in aqueous solvents but not dry methanol: it was characte-
rized by UV, FAB mass and NMR spectroscopy. The compound was shown by UV
and 1 H-NMR to have a pyridinium ring structure similar to desmosine. Mass
spectral analysis indicated a parent compound with a mass of 655. We
postulated that it arose by condensation of a reduced aldol condensation
product of all ysine, all ysine and lysine. s1990 AcademK Press, Inc.
Elastin is a connective tissue protein found in virtually every tissue
and organs. The highest concentration is found in the aorta and pulmonary
vessels(l.2). The degradation of elastin is accelerated in several
diseases such as atherosclerosis and diabetes mellitus(3). Also aberrations
in elastin structure and metabolism have been detected in a number of
heritable and acquired diseases. Elastin is secreted from the cell as a
soluble polypeptide (tropoelastin). As tropoelastin comes in contact with
the extracellular matrix, it becomes insoluble and fibrillar in structure.
This occurs through a series of intra- and intermolecular crosslinks. It is
known that elastin is cross-linked by unique pol yfunctional amino acids
which have a quaternary pyridinium skeleton; desmosine(4), isodesmosine(4),
neodesmosine(5) and pentasine(6). It is demonstrated that a-amino adipic
acid a-semialdehyde(allysine) and lysine are embodied of these cross-
linking amino acids(7,8), however the route of formation is still
uncertain. To gain more understanding about the nature and function of
elastin, we have attempted to isolate hitherto undescribed cross-linking
amino acids. Here, we show that isolation and characterization of such a
new amino acid derived from the cross-linkage in elastin is proposed.
0006-291X/90 $1.50
713 Copyright 0 1990 by Academic Press, Inc.
All rights of reproduction in any form reserved.

Vol. 170, No. 2, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
EXPERIMENTAL
After acid hydrolysis (GN,HCl; at 110” C for 48h) of purified elastin
from bovine 1 igamentum nuchae, the hydrol ysate was chromatographed on
ion pair HPLC by a SHIMADZU LC-GA pump and SPD-6AV UV detector attached
to a LiChrospher 100 RP-18 125-4tMerck) reverse phase column using a pH
O.lM phosphate buffer/acetonitrile 5:l v/v containing 20mM SDS as a solvent
(final pH 4.0). Desmosine, isodesmosine and other unknown amino acids were
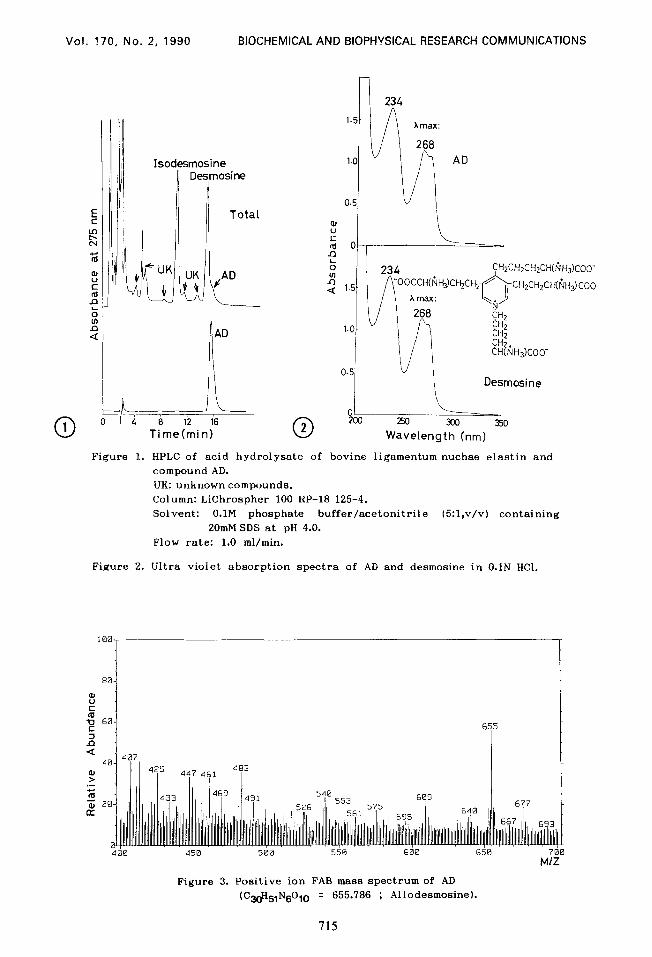
separated and detected at 275nm as shown in Figure 1.
To prepare the unknown amino acid, 50 gram of an acid hydrolysate
of elastin was charged on a large scale activated charcoal column(65 x 180
mm). Major 1 ysine derived cross-links were fractionated from water
followed by elution with 20% v/v aqueous methanol solution. Lysine derived
crosslinking amino acids thus obtained were charged on a preparative HPLC
silica gel column (10 x 240 mm; Lobar LiChroprep Si60, Merck) using ethyl
acetate/acetic acid/water 1.5:1:1 v/v as a solvent. The Fractions were
collected. Confirmation was by HPLC analysis, which indicated the presence
of another amino acid, designated as AD, eluting just after desmosine as
shown in Figure 1. Fractions containing this amino acid were pooled and
evaporated to dryness. Partially purified AD thus obtained was then
charged on the same preparative HPLC column but using a solvent system of
n-propanol/water/25%ammonia 700:330:28, v/v(9). The fractions containing AD
were collected. The UV spectrum of AD dissolved in O.lN HCl was
performed on a JASCO UVIDEC-500 spectrophotometer. Both 1 H- and ! 1 C-NMR
spectra were performed on a JEOL JNM GSX-400 instrument and FAB mass
spectrometry was done by JEOL HX-105 instrument using glycerol as a
matrix.
RESULTS AND DISCUSSION
The purity of the unknown amino acid, AD, was confirmed by HPLC and
TLC analysis . On elution systems of HPLC that will resolve desmosine and
isodesmosine, AD eluted just after desmosine and fractionated as a single
peak (Figure 1). The amino acid AD gave a heavy ninhydrin single spot on
silica gel TLC using a solvent system of ethyl acetate/acetic acid/water
l:l:l, v/v (Rf:0.098; both Rfs of desmosine and isodesmosine were 0.166).
Compound AD was a very hygroscopic, white solid with a faint yellow tinge,
soluble in aqueous solvents but not dry methanol; attempts to induce
crystallization were unsuccessful. It was thought that the hygroscopic
character might be due to the presence of the quaternary pyridinium
skeleton.
The UV spectrum of AD is shown in Figure 2 and is compared with
that of desmosine. Compound AD showed UV absorption maximum at 234
and 268 nm inflections at 274 nm which were completely identical to those
of desmosine. From the results, it is deduced that AD has the same
714
Vol. 170, No. 2, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Time(min)
234 234
1.5. 1.5.
10 10 AD AD
0.5. 0.5.
a, a,
: : 2 2 0 0
b b In In
2 2 1.5, 1.5,
1.0. 1.0.
0.5. 0.5. Desmosine Desmosine
0 2 0 200 250 300 350
Wavelength (nm)
Figure I. HPLC of acid hydrolysate of bovine ligamentum nuchae elastin and
compound AD.
UK: unknown compounds.
Column: LiChrospher 100 RP-18 125-4.
Solvent: O.lM phosphate buffer/acetonitrile (5:l,v/v) containing
20mM SDS at pH 4.0.
Flow rate: 1.0 ml/min.
Figure 2. Ultra violet absorption spectra of AD and desmosine in O.lN HCI.
400 450 500 550 600 650 700 M/Z
Figure 3. Positive ion FAB mass spectrum of AD (C.3&IN6010 = 655.786 ; Allodesmosine).
715
Vol. 170, No. 2, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
substitution pattern on the quaternary pyridinium skeleton as desmosine.
The FAB mass spectrum of AD gave a molecular weight of 655, consistent
with an elemental composition of C~eHslN~Oln as shown in Figure 3, whereas
desmosine gave a molecular weight of 526. For comparison and to enable us
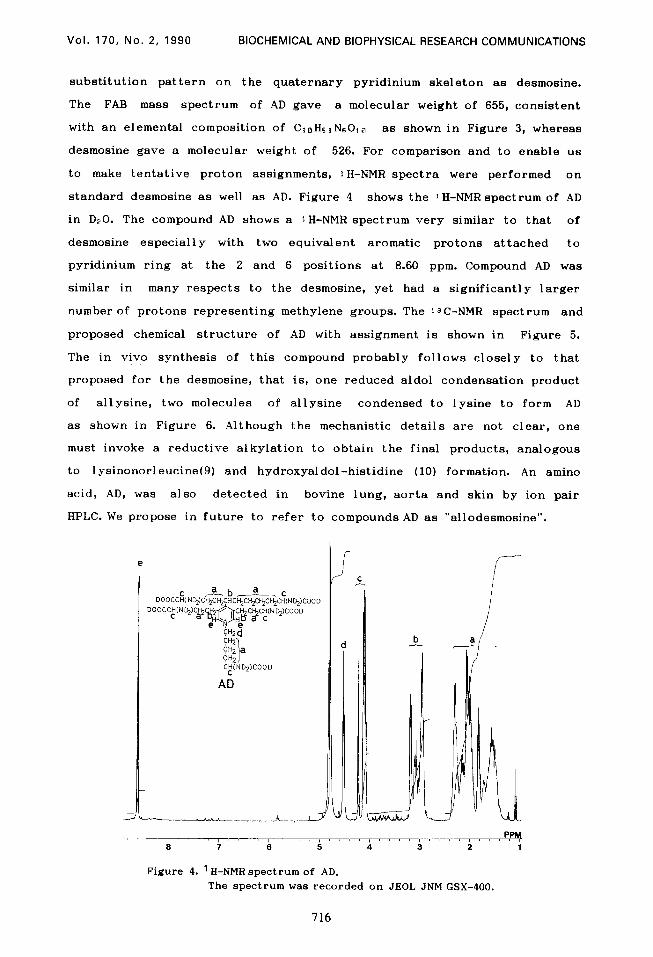
to make tentative proton assignments, 1 H-NMR spectra were performed on
standard desmosine as well as AD. Figure 4 shows the 1 H-NMR spectrum of AD
in D20. The compound AD shows a 1 H-NMR spectrum very similar to that of
desmosine especially with two equivalent aromatic protons attached to
pyridinium ring at the 2 and 6 positions at 8.60 ppm. Compound AD was
similar in many respects to the desmosine, yet had a significantly larger
number of protons representing methylene groups. The 13C-NMR spectrum and
proposed chemical structure of AD with assignment is shown in Figure 5.
The in vivo synthesis of this compound probably follows closely to that
proposed for the desmosine, that is, one reduced aldol condensation product
of allysine, two molecules of al 1 ysine condensed to lysine to form AD
as shown in Figure 6. Although the mechanistic details are not clear, one
must invoke a reductive alkylation to obtain the final products, analogous
to lysinonorleucine(9) and hydroxyaldol-histidine (10) formation. An amino
acid, AD, was also detected in bovine lung, aorta and skin by ion pair
HPLC. We propose in future to refer to compounds AD as “allodesmosine”.
CCH(ND2)CooD AD
PPM I ‘, ” I ” I ” ” I ” ” I ” ” I ” ” r
6 7 6 5 4 3 2 1
Figure 4. ' H-NMR spectrum of AD. The spectrum was recorded on JEOL JNM GSX-400.
716

Vol. 170, No. 2, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
170 160 150 140 130 120 110 100 90 80 70 60 50 40 30 20
DEPT b % t C”,CH3 t c -
PPM
Figure 5. 13C-NMR spectra of AD.
The spectra were recorded on JEOL JNM GSX-400 with proton noise
decoupled (A) and DEPT(distortionless enhancement by polarization
transfer) method (B).
-OOCCH(t?H,)CH,CH,CHCH,CH,CH,CH,CH(ljH,)COO- bH0
OOCCH(t?H,)CH$H~H, 1
yH,CH,CH,CH( NH$OO- OHC CHO >
cc? 2
%H,KOO-
Allodesmosi ne
Figure 6. Postulated mechanism of formation of allodesmosine.
717

Vol. 170, No. 2, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
ACKNOWLEDGMENT
This work was supported in part by Grants-in-Aid for Scientific
Research (No. 01560303) from the Ministry of Education, Science and Culture
of Japan.
REFERENCES
1. Desai, R., Wigglesworth, J.S. and Aber, V. (1988) Early Hum. Dev.
16,61-71.
2. John, R and Thomas, J. (1972) Biochem. J. 127,261-269.
3. Rosenbloom, J. (1982) Cons. Tiss. Res. 10,73-91.
4. Thomas, J., Elsden, D.F. and Partridge, S. (1963) Nature
200,651-652.
5. Nagai, Y. (1983) Corm. Tiss. 14,112-113.
6. Starcher, B.C., Cook, G., Gallop, P., Hensen, E. and
Shoulders, B. (1987) Bonn. Tissue Res. 16,15-25.
7. Davis, N.R. and Anwar, R.A. (1970) J. Am. Chern. Sec.
92,3778-3782
8. Francis, G., John, R. and Thomas, J. (1973) Siochem. J.
136,45-55(1973).
9. Nakamura, F. and Suyama, K. (1990) J. chrornatogr. SC i. in press.
10. Franzblau, C., Faris, B. and Papaioannou, R. (1969) Biochemistry
8,2833-2835.
11. Housley, T., Tanzer, M.L., Henson, E. and Gallop,P.M. (1975)
Biochem. Biophys. Res. Commun. 67,824-829.
718